吴国栋, 修宇, 王华芳*, 北京林业大学生物科学与技术学院, 林木育种国家工程实验室, 林木花卉遗传育种教育部重点实验室, 北京 100083 WuGuodong, XiuYu, WangHuafang*, Key Laboratory of Genetics and Breeding in Forest Trees and Ornamental Plants of Ministry of Education, National Engineering Laboratory for Tree Breeding, College of Biological Sciences and Biotechnology, Beijing Forestry University, Beijing 100083, China 引用本文 吴国栋, 修宇, 王华芳. 优化子叶节转化法培育大豆转基因植株. 植物学报, 2018, 53(1): 59-71
Abstract: Orthogonal factorial experiments and histochemical GUS staining were combined to optimize the genetic transformation system of Glycine max cv. ‘Dongnong 50’ and transfer the key gene MtDREB2A for drought resistance into the soybean. Sterile of soybean seeds used as explants with NaClO solution and Cl2 gas fumigation methods reached 98.67% and 93.33% germination, respectively. Histochemical staining rate of the tissues transformed with GUS by the cotyledon node method was 68.33%, significantly higher than that by the hypocotyl (14.00%) and embryo tip (0.67%) methods (P<0.05). The cotyledon node-transformed GUS gene was up to 72.00% in germinated sterile seeds for 5 days, mediated by Agrobacterium tumefaciens cultured at 25°C, OD600 0.9, and co-cultured for 5 days. The shoots were induced and differentiated with cotyledon node-transformed GUS up to 3.33% by optimal recovery culture for 5 days and were screened on culture medium containing phosphinothricin (3 mg·L-1), cefotaxime sodium (200 mg·L-1) and carbenicillin (300 mg·L-1). The transgenic efficiency was 1.11% with the optimized soybean genetic transformation system. The MtDREB2A transgenic plant roots of soybean ‘Dongnong 50’ were more dense and both taproot length and lateral root number were significantly longer and greater than those of the control (P<0.05). The study verified that the MtDREB2A gene plays a role in promoting soybean root growth, which lays a solid foundation and provides a theoretical basis for the gene using in drought resistance breeding of soybean.
Key words:soybean ; optimization of gene transformation system ; GUS ; MtDREB2A ; root enhancement
Different lowercase letters within column indicate significant differences at 0.05 level according to Duncan’s test. 同列不同小写字母表示在0.05水平差异显著(Duncan’s test)。
表3 东农50种子NaClO法表面消毒实验结果(平均值±标准差) Table 3 Effect of NaClO method on seed sterilization of Glycine max cv. ‘Dongnong 50’ (means±SD)
表4 Table 4 表4 表4 东农50种子Cl2气熏蒸法表面消毒实验结果 Table 4 Effect of Cl2 method on seed sterilization of Glycine max cv. ‘Dongnong 50’
Treatment
Time (h)
Sterilization rate (%)
Contamination rate (%)
Death rate (%)
1
1
82.00±5.29 b
13.33±5.78 a
4.66±1.16 b
2
2
82.67±6.11 b
12.00±6.00 a
5.33±3.06 b
3
4
92.00±3.46 a
2.67±2.31 b
5.33±3.06 b
4
6
93.33±1.16 a
2.67±2.31 b
4.00±2.00 b
5
8
88.00±3.46 ab
1.33±2.31 b
10.67±1.16 a
Different lowercase letters within column indicate significant differences at 0.05 level according to Duncan’s test. 同列不同小写字母表示在0.05水平差异显著(Duncan’s test)。
表4 东农50种子Cl2气熏蒸法表面消毒实验结果 Table 4 Effect of Cl2 method on seed sterilization of Glycine max cv. ‘Dongnong 50’
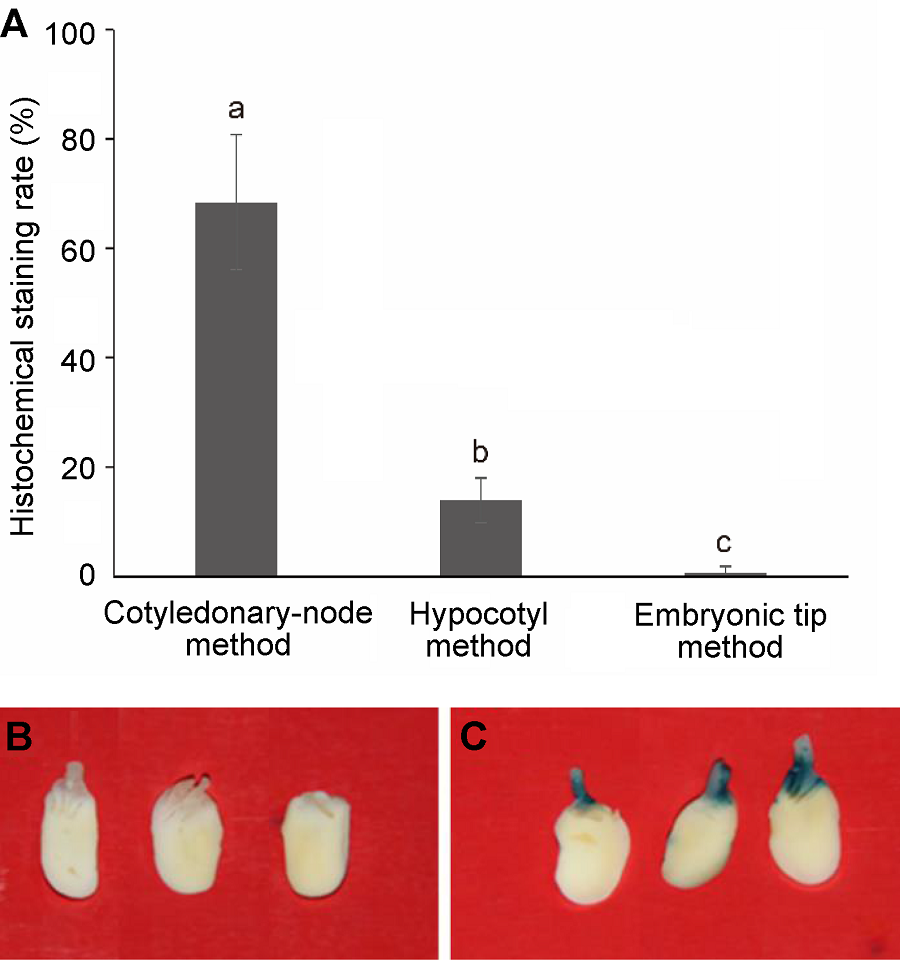
2.2.2 转化方法对GUS基因组织化学染色率的影响 大豆东农50子叶节、下胚轴和胚尖转化GUS基因的转化效率如图2A所示。子叶节法转化GUS基因的染色率达68.33%, 显著高于下胚轴法(14.00%)和胚尖法(0.67%) (P<0.05)。因此用正交因素试验对子叶节转化法进行主要影响因素的优化。 2.2.3 子叶节法遗传转化体系的优化 2.2.3.1 农杆菌侵染条件的优化 4种因素(表5)——种子萌发时间、农杆菌培养温度、浓度(OD600值)和共培养时间, L16(45)16个组合处理对子叶节GUS基因组织化学染色率(图2B, C)均影响显著(P<0.05)。极差分析表明, 4种因素的影响从大到小依次为共培养时间>农杆菌培养温度>萌发时间>OD600值。GUS基因染色率随着共培养时间的延长及农杆菌培养温度的降低而增加, 共培养5天和农杆菌培养温度为25°C时最高。此外, GUS基因染色率随着种子萌发时间的增加呈先升高后下降的趋势, 萌发5天达到最高; 随着OD600值的增大则呈先下降后升高的趋势, 在OD600值为0.9时达到最高。4种因素的优化组合为A3B1C4D4。此组合在正交试验中未出现, 以此进行的3次重复实验转GUS基因组织染色率分别为36/50、34/50和38/50, 平均为(72.00±2.00)%。 2.2.3.2 不定芽诱导条件的优化 农杆菌转化后, 4种因素——恢复培养时间、PPT浓度、Cef浓度和Cb浓度, L16(45) 16个组合对不定芽诱导率及GUS基因染色率的影响如表6所示。恢复培养时间和PPT浓度对不定芽诱导率的影响显著(P<0.05), 不定芽诱导率随恢复培养时间的延长和PPT浓度的降低而增加。4种因素的最佳组合为A4B1C4D2 (组合13), 此组合的不定芽诱导率最高, 达55.56%。 图2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-59/img_2.png<b>图2</b> <i>GUS</i>基因组织化学染色<br/>(A) 转化方法对<i>GUS</i>基因组织化学染色率的影响, 不同小写字母表示在0.05水平差异显著(Duncan’s test); (B), (C) 共培养后的子叶节<i>GUS</i>基因组织化学染色((B) 阴性对照; (C) 组织化学染色阳性)<br/><b>Figure 2</b> <i>GUS</i> gene histochemical staining<br/>(A) The effect of transgenic methods on histochemical <i>GUS</i> staining rate, different lowercase letters indicate significant differences at 0.05 level according to Duncan’s test; (B), (C) The <i>GUS</i> gene histochemical staining of cotyledonary node after co-cultivation ((B) Negative control; (C) Positive result of histochemical stain) Figure 2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-59/img_2.png<b>图2</b> <i>GUS</i>基因组织化学染色<br/>(A) 转化方法对<i>GUS</i>基因组织化学染色率的影响, 不同小写字母表示在0.05水平差异显著(Duncan’s test); (B), (C) 共培养后的子叶节<i>GUS</i>基因组织化学染色((B) 阴性对照; (C) 组织化学染色阳性)<br/><b>Figure 2</b> <i>GUS</i> gene histochemical staining<br/>(A) The effect of transgenic methods on histochemical <i>GUS</i> staining rate, different lowercase letters indicate significant differences at 0.05 level according to Duncan’s test; (B), (C) The <i>GUS</i> gene histochemical staining of cotyledonary node after co-cultivation ((B) Negative control; (C) Positive result of histochemical stain)
图2 GUS基因组织化学染色 (A) 转化方法对GUS基因组织化学染色率的影响, 不同小写字母表示在0.05水平差异显著(Duncan’s test); (B), (C) 共培养后的子叶节GUS基因组织化学染色((B) 阴性对照; (C) 组织化学染色阳性) Figure 2 GUS gene histochemical staining (A) The effect of transgenic methods on histochemical GUS staining rate, different lowercase letters indicate significant differences at 0.05 level according to Duncan’s test; (B), (C) The GUS gene histochemical staining of cotyledonary node after co-cultivation ((B) Negative control; (C) Positive result of histochemical stain)
恢复培养时间对转GUS基因不定芽组织化学染色率影响显著(P<0.05)。极差分析显示, 4种因素的影响依次为恢复培养时间>PPT浓度=Cef浓度=Cb浓度; 最佳组合为A2B2C2D3, 该组合在试验中未出现, 对其进行验证实验(3次重复, 每重复30个外植体, 染色率分别为1/30、1/30和1/30), 不定芽染色率为(3.33± 0.00)%。 表5 Table 5 表5 表5 农杆菌侵染大豆东农50的条件优化 Table 5 The results of orthogonal experiment for Agrobacterium-mediated transformation of Glycine max cv. ‘Dongnong 50’
Treatment
Factor
Histochemical staining rate of cotyledonary-node (%)
Germination time (d) (A)
Culture temperature (°C) (B)
OD600 value (C)
Co-cultivation time (d) (D)
1
3
25
0.3
2
8.00±2.00 g
2
3
26
0.5
3
31.05±10.46 def
3
3
27
0.7
4
38.27±5.46 cde
4
3
28
0.9
5
46.30±17.65 bcd
5
4
25
0.5
4
39.70±19.34 cde
6
4
26
0.3
5
59.33±3.06 ab
7
4
27
0.9
2
4.00±2.00 g
8
4
28
0.7
3
21.33±6.43 efg
9
5
25
0.7
5
68.33±12.42 a
10
5
26
0.9
4
51.06±7.65 abc
11
5
27
0.3
3
30.56±16.17 def
12
5
28
0.5
2
4.00±5.29 g
13
6
25
0.9
3
40.95±11.61 bcd
14
6
26
0.7
2
8.52±1.70 g
15
6
27
0.5
5
20.36±9.30 efg
16
6
28
0.3
4
12.67±10.26 fg
K1
30.91
39.25
27.64
6.13
K2
31.09
37.49
23.78
30.97
K3
38.49
23.30
34.12
35.43
K4
20.62
21.07
35.58
48.58
R
17.87
18.18
11.80
42.45
Kn are the average response values under the levels of n, R are the ranges of factors. Different lowercase letters with in column indicate significant differences at 0.05 level according to Duncan’s test. Kn为第n水平下的平均响应值, R为因素对染色率的效应极差。同列不同小写字母表示在0.05水平差异显著(Duncan’s test)。
表5 农杆菌侵染大豆东农50的条件优化 Table 5 The results of orthogonal experiment for Agrobacterium-mediated transformation of Glycine max cv. ‘Dongnong 50’
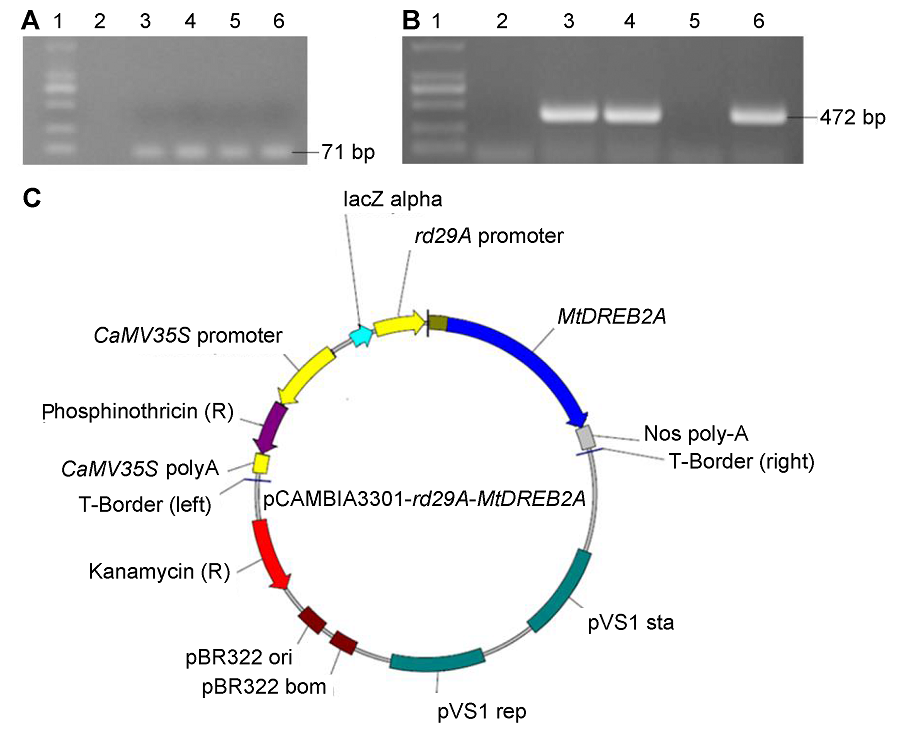
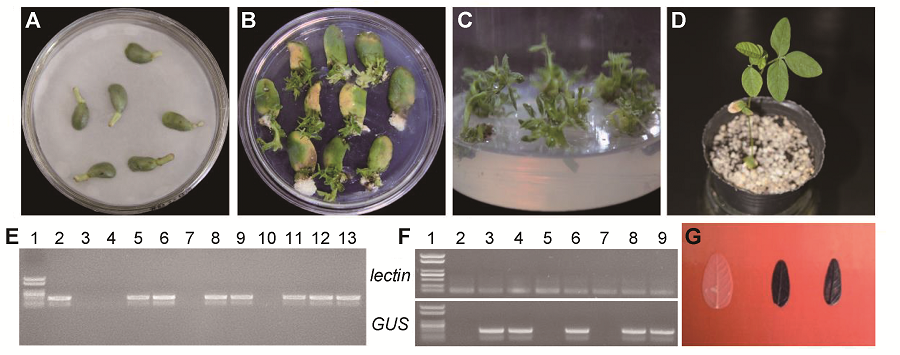
2.2.3.3 转GUS基因植株的检测 在上述优化条件下转化大豆东农50, 将再生抗性芽经诱导生根的小植株移栽到营养钵养护(图3A-D)。之后, 提取叶片基因组DNA进行GUS基因PCR检测, 显示10个抗性植株中有7个扩增出GUS基因特异片段(图3E)。对7个PCR检测呈阳性的植株的新生叶片进行GUS表达的RT-PCR检测(图3F)和组织化学染色(图3G), 结果5个植株为阳性, 表明GUS基因成功转入大豆东农50并表达; 转化效率为(1.11±0.38)% (1/ 150、2/150和2/150的统计平均值)。 图3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-59/img_3.png<b>图3</b> 优化的子叶节法转化大豆东农50<br/>(A) 农杆菌侵染后共培养; (B) 不定芽诱导; (C) 不定芽伸长; (D) 抗性芽移栽成活; (E) 抗性植株<i>GUS</i>基因PCR检测(1: Marker; 2: pCAMBIA3301-<i>35S</i>-<i>GUS</i>; 3: 对照大豆; 4-13: 抗性植株); (F) 抗性植株<i>GUS</i>基因RT-PCR检测(1: Marker; 2: 对照大豆; 3-9: 抗性植株); (G) 对照大豆(左)和抗性植株(中、右)叶片的组织化学染色<br/><b>Figure 3</b> Transformation system of <i>Glycine max</i> cv. ‘Dongnong 50’ using optimized cotyledonary-node method<br/>(A) Co-cultivation after transformation; (B) Shoot induction; (C) Shoot elongation; (D) Resistant bud transplant; (E) PCR confirmation of <i>GUS </i>gene (1: Marker; 2: pCAMBIA3301-<i>35S</i>-<i>GUS</i>; 3: Control soybean; 4-13: Resistant plant); (F) RT-PCR confirmation of <i>GUS </i>gene (1: Marker; 2: Control soybean; 3-9: Resistant plant); (G) Histochemical staining of leaves from control (left) and resistant plant (middle and right) Figure 3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-59/img_3.png<b>图3</b> 优化的子叶节法转化大豆东农50<br/>(A) 农杆菌侵染后共培养; (B) 不定芽诱导; (C) 不定芽伸长; (D) 抗性芽移栽成活; (E) 抗性植株<i>GUS</i>基因PCR检测(1: Marker; 2: pCAMBIA3301-<i>35S</i>-<i>GUS</i>; 3: 对照大豆; 4-13: 抗性植株); (F) 抗性植株<i>GUS</i>基因RT-PCR检测(1: Marker; 2: 对照大豆; 3-9: 抗性植株); (G) 对照大豆(左)和抗性植株(中、右)叶片的组织化学染色<br/><b>Figure 3</b> Transformation system of <i>Glycine max</i> cv. ‘Dongnong 50’ using optimized cotyledonary-node method<br/>(A) Co-cultivation after transformation; (B) Shoot induction; (C) Shoot elongation; (D) Resistant bud transplant; (E) PCR confirmation of <i>GUS </i>gene (1: Marker; 2: pCAMBIA3301-<i>35S</i>-<i>GUS</i>; 3: Control soybean; 4-13: Resistant plant); (F) RT-PCR confirmation of <i>GUS </i>gene (1: Marker; 2: Control soybean; 3-9: Resistant plant); (G) Histochemical staining of leaves from control (left) and resistant plant (middle and right)
图3 优化的子叶节法转化大豆东农50 (A) 农杆菌侵染后共培养; (B) 不定芽诱导; (C) 不定芽伸长; (D) 抗性芽移栽成活; (E) 抗性植株GUS基因PCR检测(1: Marker; 2: pCAMBIA3301-35S-GUS; 3: 对照大豆; 4-13: 抗性植株); (F) 抗性植株GUS基因RT-PCR检测(1: Marker; 2: 对照大豆; 3-9: 抗性植株); (G) 对照大豆(左)和抗性植株(中、右)叶片的组织化学染色 Figure 3 Transformation system of Glycine max cv. ‘Dongnong 50’ using optimized cotyledonary-node method (A) Co-cultivation after transformation; (B) Shoot induction; (C) Shoot elongation; (D) Resistant bud transplant; (E) PCR confirmation of GUS gene (1: Marker; 2: pCAMBIA3301-35S-GUS; 3: Control soybean; 4-13: Resistant plant); (F) RT-PCR confirmation of GUS gene (1: Marker; 2: Control soybean; 3-9: Resistant plant); (G) Histochemical staining of leaves from control (left) and resistant plant (middle and right)
2.3 转MtDREB2A基因再生植株根系生长以优化的大豆东农50子叶节转化法获得的转MtDRE- B2A基因再生植株叶片, PCR扩增的MtDREB2A特异片段(约484 bp)序列与MtDREB2A基因序列比对一致(结果未显示), 获得3个阳性转基因株系(图4A)。与对照相比, 转MtDREB2A基因植株根系更密集, 根的数量、长度和生长量明显增加(图4B, E)。转基因大豆可正常开花并产生种子(图4D)。收获的种子播种于营养钵中, 根系数量和长度均显著高于对照(P<0.05) (图4C, F)。 图4https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-59/img_4.png<b>图4</b> 转<i>MtDREB2A</i>基因大豆东农50鉴定和根系生长状况测定<br/>(A) <i>MtDREB2A</i>基因PCR检测(1: Marker; 2: pCAMBIA3301-<i>rd29A</i>-<i>MtDREB2A</i>; 3: 对照大豆; 4-6: 转<i>MtDREB2A</i>基因大豆); (B) 对照大豆根系(Bar=1 cm); (C) 对照和转<i>MtDREB2A</i>基因大豆主根长度; (D) 转<i>MtDREB2A</i>基因大豆荚果; (E) 转<i>MtDREB2A</i>基因大豆根系(Bar=1 cm); (F) 对照和转<i>MtDREB2A</i>基因大豆侧根数量。图(C)和(F)中不同小写字母表示在0.05水平差异显著(Duncan’s test)。<br/><b>Figure 4</b> Confirmation and root system analyses of <i>MtDREB2A</i> transgenic soybean Dongnong 50<br/>(A) PCR result of <i>MtDREB2A</i> transgenic soybean (1: Marker; 2: pCAMBIA3301-<i>rd29A</i>-<i>MtDREB2A</i>; 3: Control soybean; 4-6: <i>MtDREB2A</i> transgenic soybean); (B) The root of control soybean (Bar=1 cm); (C) The length of taproot of control and transgenic soybeans; (D) The pod of <i>MtDREB2A</i> transgenic soybean in greenhouse; (E) The root of <i>MtDREB2A</i> transgenic soybean (Bar=1 cm); (F) The number of lateral root of control and transgenic soybeans. Different lowercase letters in Figures (C) and (F) indicate significant differences at 0.05 level according to Duncan’s test. Figure 4https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-59/img_4.png<b>图4</b> 转<i>MtDREB2A</i>基因大豆东农50鉴定和根系生长状况测定<br/>(A) <i>MtDREB2A</i>基因PCR检测(1: Marker; 2: pCAMBIA3301-<i>rd29A</i>-<i>MtDREB2A</i>; 3: 对照大豆; 4-6: 转<i>MtDREB2A</i>基因大豆); (B) 对照大豆根系(Bar=1 cm); (C) 对照和转<i>MtDREB2A</i>基因大豆主根长度; (D) 转<i>MtDREB2A</i>基因大豆荚果; (E) 转<i>MtDREB2A</i>基因大豆根系(Bar=1 cm); (F) 对照和转<i>MtDREB2A</i>基因大豆侧根数量。图(C)和(F)中不同小写字母表示在0.05水平差异显著(Duncan’s test)。<br/><b>Figure 4</b> Confirmation and root system analyses of <i>MtDREB2A</i> transgenic soybean Dongnong 50<br/>(A) PCR result of <i>MtDREB2A</i> transgenic soybean (1: Marker; 2: pCAMBIA3301-<i>rd29A</i>-<i>MtDREB2A</i>; 3: Control soybean; 4-6: <i>MtDREB2A</i> transgenic soybean); (B) The root of control soybean (Bar=1 cm); (C) The length of taproot of control and transgenic soybeans; (D) The pod of <i>MtDREB2A</i> transgenic soybean in greenhouse; (E) The root of <i>MtDREB2A</i> transgenic soybean (Bar=1 cm); (F) The number of lateral root of control and transgenic soybeans. Different lowercase letters in Figures (C) and (F) indicate significant differences at 0.05 level according to Duncan’s test.
图4 转MtDREB2A基因大豆东农50鉴定和根系生长状况测定 (A) MtDREB2A基因PCR检测(1: Marker; 2: pCAMBIA3301-rd29A-MtDREB2A; 3: 对照大豆; 4-6: 转MtDREB2A基因大豆); (B) 对照大豆根系(Bar=1 cm); (C) 对照和转MtDREB2A基因大豆主根长度; (D) 转MtDREB2A基因大豆荚果; (E) 转MtDREB2A基因大豆根系(Bar=1 cm); (F) 对照和转MtDREB2A基因大豆侧根数量。图(C)和(F)中不同小写字母表示在0.05水平差异显著(Duncan’s test)。 Figure 4 Confirmation and root system analyses of MtDREB2A transgenic soybean Dongnong 50 (A) PCR result of MtDREB2A transgenic soybean (1: Marker; 2: pCAMBIA3301-rd29A-MtDREB2A; 3: Control soybean; 4-6: MtDREB2A transgenic soybean); (B) The root of control soybean (Bar=1 cm); (C) The length of taproot of control and transgenic soybeans; (D) The pod of MtDREB2A transgenic soybean in greenhouse; (E) The root of MtDREB2A transgenic soybean (Bar=1 cm); (F) The number of lateral root of control and transgenic soybeans. Different lowercase letters in Figures (C) and (F) indicate significant differences at 0.05 level according to Duncan’s test.
Histochemical staining rate of adventitious bud (%)
Recovery time (d) (A)
PPT concentration (mg·L-1) (B)
Cef concentration (mg·L-1) (C)
Cb concentration (mg·L-1) (D)
1
0
2
100
100
1.11±1.92 f
0.00±0.00
2
0
3
200
200
1.11±1.92 f
1.11±1.92
3
0
4
300
300
3.33±5.77 f
2.22±1.92
4
0
5
400
400
1.11±1.92 f
1.11±1.92
5
5
2
200
300
22.22±9.62 cd
2.22±1.92
6
5
3
100
400
7.78±3.85 ef
3.33±0.00
7
5
4
400
100
4.45±3.85 f
1.11±1.92
8
5
5
300
200
2.22±3.85 f
1.11±1.92
9
10
2
300
400
31.11±5.09 bcd
0.00±0.00
10
10
3
400
300
18.89±3.85 de
0.00±0.00
11
10
4
100
200
27.78±10.18 cd
0.00±0.00
12
10
5
200
100
21.11±8.39 cd
1.11±1.92
13
15
2
400
200
55.56±5.09 a
0.00±0.00
14
15
3
300
100
42.22±12.62 b
0.00±0.00
15
15
4
200
400
33.33±13.33 bc
0.00±0.00
16
15
5
100
300
23.33±8.82 cd
0.00±0.00
Adventitious bud induction rate
K1
1.67
27.50
15.00
17.22
K2
9.17
17.50
19.44
21.67
K3
24.72
17.22
19.72
16.94
K4
38.61
11.94
20.00
18.33
R
36.94
15.56
5.00
4.72
Histochemical staining rate
K1
1.11
0.56
0.83
0.56
K2
1.94
1.11
1.11
0.56
K3
0.28
0.83
0.83
1.11
K4
0.00
0.83
0.56
1.11
R
1.94
0.56
0.56
0.56
Different lowercase letters within column indicate significant differences at 0.05 level according to Duncan’s test. 同列不同小写字母表示在0.05水平差异显著(Duncan’s test)。
表6 不定芽诱导条件正交试验的不定芽诱导率和组织化学染色率 Table 6 The adventitious bud induction rate and histochemical staining rate of orthogonal experiment
2.4.2 大豆东农50的遗传转化体系优化 农杆菌侵染和不定芽诱导的多种因素对大豆植株再生与遗传转化效率均有明显影响(邱波等, 2011)。有关大豆遗传转化的研究多采用单因子试验方法逐一分析(林树柱等, 2005; 武小霞等, 2010; 薄路花和曹越平, 2015), 虽然简单明了但对于因素间相互作用的分析可能有局限性(林荣双等, 2003)。正交因素试验法一并分析多个因素及其相互作用的影响, 简化了实验并确定了因素的主次效应, 可获得最优组合(刘瑞江等, 2010)。该方法已用于植物分生组织、不定芽诱导和不定根发生影响因素等的研究(韩献忠等, 1990; 林荣双等, 2003; 马艳等, 2004)。本研究将正交试验方法与转GUS基因的组织化学染色技术相结合(袁鹰等, 2004; 桑庆亮等, 2014)探索多个因素对大豆子叶节法转化效率的影响, 节省了人力物力且提高了检测效率。 农杆菌培养温度显著影响大豆的转化效率。大豆遗传转化体系的农杆菌培养温度多为27-28°C (Don- aldson and Simmonds, 2000; Liu et al., 2004; Hong et al., 2007; Wang and Xu, 2008; 段莹莹等, 2010; 邱波等, 2011)。该温度最适合农杆菌生长, 此温度下细菌分裂繁殖速度较快。正交因素试验表明, 在一定范围内, 转GUS基因组织化学染色率随农杆菌培养温度的降低而升高, 当培养温度为25°C时染色率最高。对农杆菌侵染转化过程进行分析, 发现温度降低可能导致农杆菌分裂速度变慢及在溶液中的黏性增大, 影响受体细胞侵染、T-DNA插入及基因组DNA重组和在其中稳定表达。农杆菌EHA105浓度在OD600为0.3-0.5时的升高速度较慢, OD600为0.5-0.7时为对数生长期, 升高速度最快; OD600为0.9时进入平台期(杨莹, 2013); 而平台期浓度转化的组织化学染色率最高, 表明侵染液高浓度农杆菌利于大豆侵染。大豆种子萌发时间延长, GUS染色率增加, 但萌发时间达到6天的染色率反而降低(表5)。这可能与萌发阶段细胞的有丝分裂活性有关(段莹莹等, 2010)。 影响不定芽诱导的4种因素L16(45)正交试验表明, 转化材料恢复培养时间的延长或培养基PPT浓度降低均可提高不定芽的诱导率, 恢复培养时间延长至15天, PPT浓度为2 mg·L-1时不定芽的诱导率最高, 但GUS基因组织化学染色率最低(表6), 这表明GUS染色不定芽/不定芽总数的比例发生了变化, 即非转化不定芽数量的增加较快。相反, 侵染后立即在含有较高浓度PPT的诱导培养基上培养, GUS基因组织化学染色率随恢复培养时间的延长和PPT浓度的升高呈现先上升后下降趋势。这表明农杆菌介导转化的细胞对PPT抗性存在迟滞, 恢复培养时间过短易造成转化细胞死亡, 进而降低不定芽的诱导率。适度地恢复培养有利于提高转化细胞分化不定芽的比例。在本文所述条件下, 大豆子叶节法的转化效率已提高到1.11%。 2.4.3 转MtDREB2A基因大豆植株增强根系生长 DREB2基因具有促进植物根系生长和抗旱的功能(Liu et al., 1998; Shinozaki et al., 2003; Xiu et al., 2016)。本实验室前期的工作表明, MtDREB2A基因促进豆科植物蒺藜苜蓿(Medicago truncatula)根系的生长(Chen et al., 2009)。以优化的遗传转化体系将MtDREB2A基因导入大豆东农50, 转基因再生植株在温室条件下生长, 根系的长度和数量均显著高于对照(P<0.05), MtDREB2A基因促进根系生长的功能再次在大豆转化中得到了验证。后续研究需对转Mt- DREB2A基因大豆植株正常开花结实的后代的抗旱性和农艺性状进行检测, 以期为利用该基因进行大豆抗旱改良提供更丰富的基础依据。
The authors have declared that no competing interests exist.
ChenJR, LüJJ, LiuR, XiongXY, WangTX, ChenSY, GuoLB, WangHF (2010). DREB1C from Medicago truncatula enhances freezing tolerance in transgenic M. truncatula and China Rose(Rosa chinensis Jacq.). Plant Growth Regul 60, 199-211. [本文引用: 1]
[27]
ChenJR, LüJJ, WangTX, ChenSY, WangHF (2009). Activation of a DRE-binding transcription factor from Medicago truncatula by deleting a Ser/Thr-rich region. In Vitro Cell Dev Biol Plant 45, 1-11. [本文引用: 2]
[28]
DonaldsonPA, SimmondsDH (2000). Susceptibility to Agrobacterium tumefaciens and cotyledonary node trans- formation in short-season soybean. Plant Cell Rep 19, 478-484. [本文引用: 2]
[29]
DubouzetJG, SakumaY, ItoY, KasugaM, DubouzetEG, MiuraS, SekiM, ShinozakiK, Yamaguchi-ShinozakiK (2003). OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in drought-, high-salt- and cold-responsive gene expression. Plant J 33, 751-763. [本文引用: 1]
[30]
GaoMJ, AllardG, ByassL, FlanaganAM, SinghJ (2002). Regulation and characterization of four CBF transcription factors from Brassica napus. Plant Mol Biol 49, 459-471. [本文引用: 1]
HincheeMAW, Connor-WardDV, NewellCA, McDonnellRE, SatoSJ, GasserCS, FischhoffDA, ReDB, FraleyRT, HorschRB (1988). Production of transgenic soybean plants using Agrobacterium-mediated DNA transfer. Nat Biotechnol 6, 915-922. [本文引用: 1]
[33]
HongHP, ZhangHY, OlhoftP, HillS, WileyH, TorenE, HillebrandH, JonesT, ChengM (2007). Organogenic callus as the target for plant regeneration and transformation viaAgrobacterium in soybean(Glycine max 43, 558-568. [本文引用: 2]
[34]
KoTS, KorbanSS (2004). Enhancing the frequency of somatic embryogenesis followingAgrobacterium-media- ted transformation of immature cotyledons of soybean(Glycine max 40, 552-558. [本文引用: 1]
[35]
LiuHK, YangC, WeiZM (2004). Efficient Agrobacterium tumefaciens-mediated transformation of soybeans using an embryonic tip regeneration system. Planta 219, 1042-1049. [本文引用: 3]
[36]
LiuQ, KasugaM, SakumaY, AbeH, MiuraS, Yamaguchi-ShinozakiK, ShinozakiK (1998). Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis.Plant Cell 10, 1391-1406. [本文引用: 2]
OlhoftPM, FlagelLE, DonovanCM, SomersDA (2003). Efficient soybean transformation using hygromycin B selection in the cotyledonary-node method.Planta 216, 723-735. [本文引用: 3]
[39]
QinF, KakimotoM, SakumaY, MaruyamaK, OsakabeY, TranLSP, ShinozakiK, Yamaguchi-ShinozakiK (2007). Regulation and functional analysis of ZmDREB2A in response to drought and heat stresses in Zea mays L. Plant J 50, 54-69. [本文引用: 1]
[40]
SakumaY, MaruyamaK, OsakabeY, QinF, SekiM, ShinozakiK, Yamaguchi-ShinozakiK (2006). Functional analysis of an Arabidopsis transcription factor, DREB2A, involved in drought-responsive gene expression.Plant Cell 18, 1292-1309. [本文引用: 1]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}