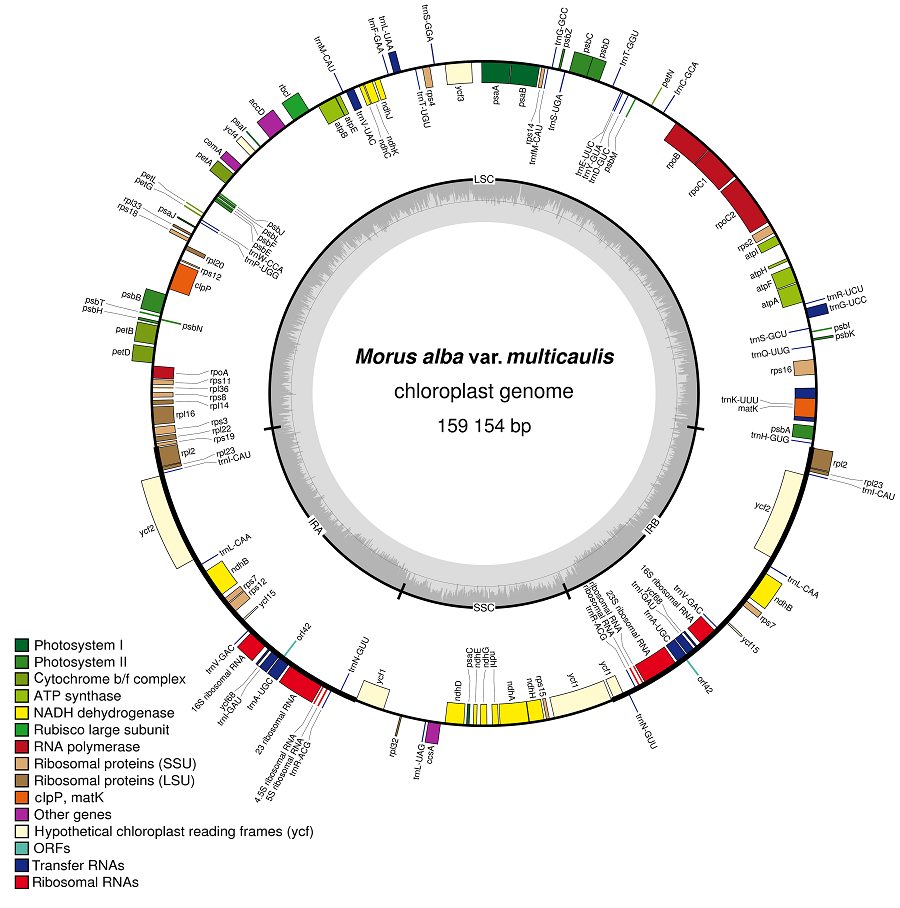
Abstract: Mulberry is an economically important crop in Asia. We determined the complete chloroplast sequence of cultivated species of Morus multicaulis. Ribenhuchen was used as experimental material. High-throughput sequencing was used to sequence the chloroplast genome and the genome structure (NCBI No.: KU355297), and we compared the chloroplast genome with those of reported sibling species (Morus mongolica, M. indica, M. notabilis). The chloroplast genome (cpDNA) of M. multicaulis with a typical quadripartite structure is 159 154 bp long. The cpDNA of M. multicaulis contains 130 genes, including 85 protein coding genes (18 genes duplicated in the inverted repeat regions), 37 transfer RNA genes and 8 ribosomal RNA genes. There are 82 simple sequence repeats, and the number of mono-, di-, tri-, tetra-, pentanucleotide repeat motifs is 63, 7, 2, 9, and 1, with no hexanucleotide repeat sequences. Mono-nucleotide repeat sequences accounted for 76.8% of the cpDNA of simple sequence repeats. MEGA 6.0 was used to construct the phylogenetic tree of 15 species and for cluster analysis of Morus plants. M. multicaulis and M. mongolica were clustered into one group. The research results have reference value for chloroplast genome research, molecular marker development and breeding of mulberry.
加粗字体表明鲁桑和蒙桑中相似的SSR位点。Bold type indicate the similar SSR in M. multicaulis and M. mongolica.
表4 鲁桑和蒙桑中简单重复序列(SSR)位点对比 Table 4 Comparison of simple sequence repeats (SSR) loci in Morus multicaulis and M. mongolica
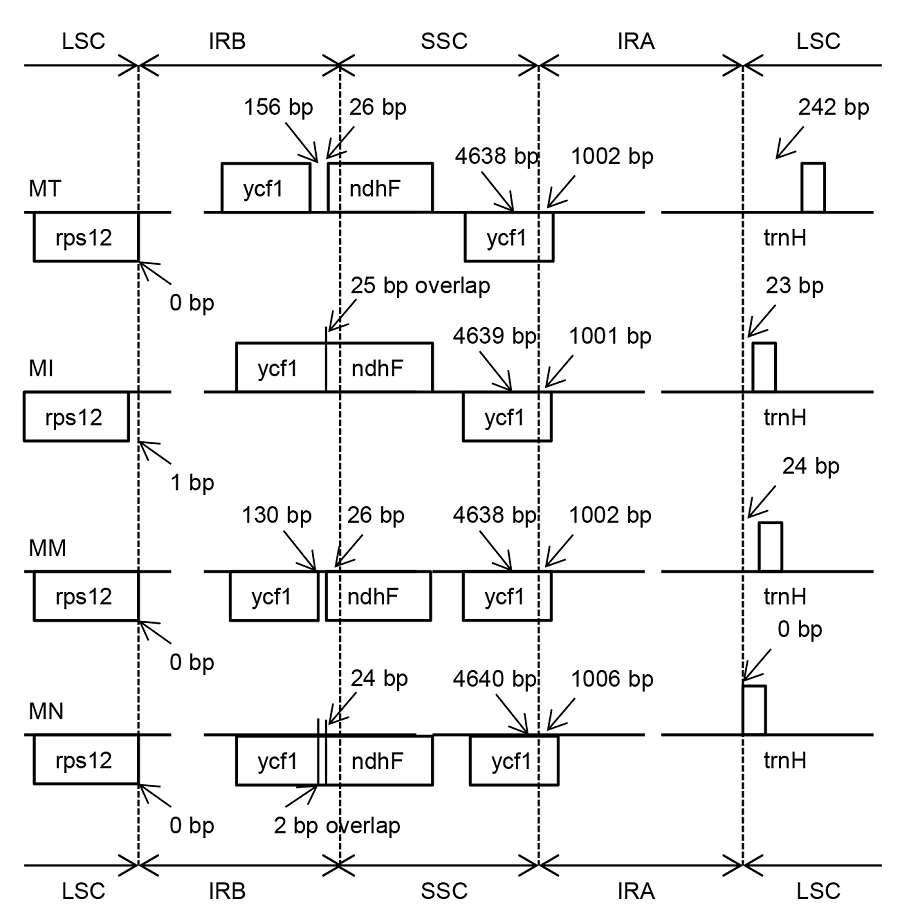
2.4 IR边界分析不同植物的叶绿体基因组大小取决于反向重复区的膨胀和收缩(Plunkett and Downie, 2000)。图2显示4个桑属物种的IR和SC边界。IR边界比较表明, 同属物种的基因组大小变化不是很明显, 表现出一定的相似性。本研究比较了4个桑属植物完整叶绿体基因组的IR/SC的边界位置, 结果显示它们具有相似的结构和基因顺序, 而在SSC/IRb交界处存在1个ycf1假基因。鲁桑的ycf1假基因与ndhF基序重叠25 bp, 这与蒙桑中的情况相同, 同时, 这种情况也出现在其它桑属植物叶绿体基因组序列中。4个桑属植物基因组的rps19基因位于非编码区的上游。印度桑中rps12基因偏离LSC/IRb边界1个碱基对, 其它3个植物rps12基因在边界处。最大的差异在LSC和IRa的边界, 鲁桑的trnH基因远离LSC/IRa边界242 bp, 而其它桑属植物中的情况不同, 只有1个很小的长度远离边界。 图2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-94/img_2.png<b>图2</b> 4个桑属物种叶绿体基因组反向重复区(IR)、大单拷贝区(LSC)和小单拷贝区(SSC)边界比对<br/>MT: 鲁桑, MI: 印度桑; MM: 蒙桑; MN: 川桑<br/><b>Figure 2</b> Comparison of the junction between inverted repeat region (IR), large single copy-region (LSC) and small single copy- region (SSC) of chloroplast genome among four <i>Morus</i> species <br/>MT: <i>Morus multicaulis</i>; MI<i>: M. indica</i>; MM: <i>M. mongolica</i>; MN: <i>M. notabilis</i> Figure 2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-94/img_2.png<b>图2</b> 4个桑属物种叶绿体基因组反向重复区(IR)、大单拷贝区(LSC)和小单拷贝区(SSC)边界比对<br/>MT: 鲁桑, MI: 印度桑; MM: 蒙桑; MN: 川桑<br/><b>Figure 2</b> Comparison of the junction between inverted repeat region (IR), large single copy-region (LSC) and small single copy- region (SSC) of chloroplast genome among four <i>Morus</i> species <br/>MT: <i>Morus multicaulis</i>; MI<i>: M. indica</i>; MM: <i>M. mongolica</i>; MN: <i>M. notabilis</i>
图2 4个桑属物种叶绿体基因组反向重复区(IR)、大单拷贝区(LSC)和小单拷贝区(SSC)边界比对 MT: 鲁桑, MI: 印度桑; MM: 蒙桑; MN: 川桑 Figure 2 Comparison of the junction between inverted repeat region (IR), large single copy-region (LSC) and small single copy- region (SSC) of chloroplast genome among four Morus species MT: Morus multicaulis; MI: M. indica; MM: M. mongolica; MN: M. notabilis
2.5 基于叶绿体基因组全序列的桑属聚类分析以烟草(Nicotiana tabacum)作为外类群, 对桑属4个物种和其它双子叶植物共15个叶绿体全基因组序列通过最大似然法(图3A)和近邻结合法(图3B)进行聚类分析, 2种方法均得出相同的结果, 即桑属的4个种聚在一起, 其中鲁桑和蒙桑聚成1个小组。 通过对鲁桑的完整叶绿体基因组的核苷酸序列和结构进行分析, 我们找到桑属植物与其它物种在序列之间的差异, 研究结果将有助于未来的进化和生态学研究。同时, 这些数据也是后续桑属种群研究指标的重要来源。此外, 完整的叶绿体基因组序列还提供了叶绿体内功能蛋白变异的数据。 图3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-94/img_3.png<b>图3</b> 基于叶绿体全基因组的桑属4个物种及其近缘种的最大似然法(ML) (A)和近邻结合法(NJ) (B)聚类结果<br/><b>Figure 3</b> Cluster analysis of four species of <i>Morus</i> using complete chloroplast genome sequence by the maximum likelihood (ML) method (A) and neighbor-joining (NL) method (B) Figure 3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-94/img_3.png<b>图3</b> 基于叶绿体全基因组的桑属4个物种及其近缘种的最大似然法(ML) (A)和近邻结合法(NJ) (B)聚类结果<br/><b>Figure 3</b> Cluster analysis of four species of <i>Morus</i> using complete chloroplast genome sequence by the maximum likelihood (ML) method (A) and neighbor-joining (NL) method (B)
图3 基于叶绿体全基因组的桑属4个物种及其近缘种的最大似然法(ML) (A)和近邻结合法(NJ) (B)聚类结果 Figure 3 Cluster analysis of four species of Morus using complete chloroplast genome sequence by the maximum likelihood (ML) method (A) and neighbor-joining (NL) method (B)
AllenderCJ, AllainguillaumeJ, LynnJ, KingGJ (2007). Simple sequence repeats reveal uneven distribution of genetic diversity in chloroplast genomes of Brassica ole- racea L. and(n=9) wild relatives. Theor Appl Genet 114, 609-618. [本文引用: 1]
[7]
ChenC, ZhouW, HuangY, WangZZ (2015). The complete chloroplast genome sequence of the mulberry Morus notabilis(Moreae). Mitochondrial DNA Part A 27, 2856-2857. [本文引用: 1]
[8]
FlanneryML, MitchellFJG, CoyneS, KavanaghTA, BurkeJI, SalaminN, DowdingP, HodkinsonTR (2006). Plastid genome characterisation in Brassica and Brassicaceae using a new set of nine SSRs. Theor Appl Genet 113, 1221-1231. [本文引用: 1]
[9]
GeorgeB, BhattBS, AwasthiM, GeorgeB, SinghAK (2015). Comparative analysis of microsatellites in chloroplast genomes of lower and higher plants.Curr Genet 61, 665-677. [本文引用: 1]
[10]
HebertPD, RatnasinghamS, de WaardJR (2003). Barcoding animal life: cytochrome c oxidase subunit 1 divergences among closely related species. Proc R Soc Lond B 270, S96-S99. [本文引用: 1]
[11]
HuangYY, MatzkeAJM, MatzkeM (2013). Complete sequence and comparative analysis of the chloroplast genome of coconut palm ( Cocos nucifera). PLoS One 8, e74736. [本文引用: 1]
JiaoY, JiaHM, LiXW, JiaHJ, ChenZ, WangGY, ChaiCY, van de WegE, GaoZS (2012). Development of simple sequence repeat (SSR) markers from a genome survey of Chinese Bayberry ( Myrica rubra). BMC Genomics 13, 201. [本文引用: 1]
[14]
KattiMV, RanjekarPK, GuptaVS (2001). Differential distribution of simple sequence repeats in eukaryotic genome sequences.Mol Biol Evol 18, 1161-1167. [本文引用: 1]
[15]
KaundunSS, MatsumotoS (2002). Heterologous nuclear and chloroplast microsatellite amplification and variation in tea, Camellia sinensis. Genome 45, 1041-1048. [本文引用: 1]
[16]
KongWQ, YangJH (2015). The complete chloroplast genome sequence of Morus mongolica and a comparative analysis within the Fabidae clade. Curr Genet 62, 165-172. [本文引用: 1]
[17]
LeighFJ, MackayI, OliveiraHR, GosmanNE, HorsnellRA, JonesH, WhiteJ, PowellW, BrownTA (2013). Using diversity of the chloroplast genome to examine evolutionary history of wheat species.Genet Resour Crop Evol 60, 1831-1842. [本文引用: 1]
[18]
LesebergCH, DuvallMR (2009). The complete chloroplast genome of Coix lacryma-jobi and a comparative molecular evolutionary analysis of plastomes in cereals. J Mol Evol 69, 311-318. [本文引用: 1]
[19]
NazarenoAG, CarlsenM, LohmannLG (2015). Complete chloroplast genome of Tanaecium tetragonolobum: the first Bignoniaceae plastome. PLoS One 10, e0129930. [本文引用: 1]
[20]
PlunkettGM, DownieSR (2000). Expansion and contraction of the chloroplast inverted repeat in Apiaceae subfamily Apioideae.Syst Bot 25, 648-667. [本文引用: 1]
[21]
RajendrakumarP, BiswalAK, BalachandranSM, SrinivasaraoK, SundaramRM (2007). Simple sequence repeats in organellar genomes of rice: frequency and distribution in genic and intergenic regions.Bioinformatics 23, 1-4. [本文引用: 1]
[22]
RaviV, KhuranaJP, TyagiAK, KhuranaP (2006). The chloroplast genome of mulberry: complete nucleotide sequence, gene organization and comparative analysis.Tree Genet Genomes 3, 49-59. [本文引用: 2]
[23]
RuhlmanTA, JansenRK (2014). The plastid genomes of flowering plants.Methods Mol Biol 1132, 3-38. [本文引用: 1]
[24]
ShawJ, LickeyEB, SchillingEE, SmallRL (2007). Comparison of whole chloroplast genome sequences to choose noncoding regions for phylogenetic studies in angiosperms: the tortoise and the hare lll.Am J Bot 94, 275-288. [本文引用: 1]
[25]
TemnykhS, DeClerckG, LukashovaA, LipovichL, CartinhourS, McCouchS (2001). Computational and experimental analysis of microsatellites in rice ( Oryza sativa L.): frequency, length variation, transposon associations, and genetic marker potential. Genome Res 11, 1441-1452. [本文引用: 1]
[26]
ZhangHY, LiC, MiaoHM, XiongSJ (2013). Insights from the complete chloroplast genome into the evolution of Se- samum indicum L. PLoS One 8, e80508. [本文引用: 1]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}