刘广超, 丁兆军*, 山东大学生命科学学院, 植物细胞工程与种质创新教育部重点实验室, 济南 250100 LiuGuangchao, DingZhaojun*, The Key Laboratory of Plant Cell Engineering and Germplasm Innovation, Ministry of Education, College of Life Sciences, Shandong University, Jinan 250100, China 引用本文 刘广超, 丁兆军. 生长素介导环境信号调控植物的生长发育. 植物学报, 2018, 53(1): 17-26
Abstract: Because plants are sessile and photo-autotrophic, they must adapt to the surrounding environment. Auxin is one of the most important plant hormones essential for plant growth and development. Recently, auxin was found to regulate plant growth by responding to endogenous developmental signals and by mediating various environmental cues. In this review, we focus on how auxin regulates plant growth by mediating various environmental cues such as light, temperature, gravity, nutrient element and metal ion signals.
Key words:auxin ; environment cues ; growth and development
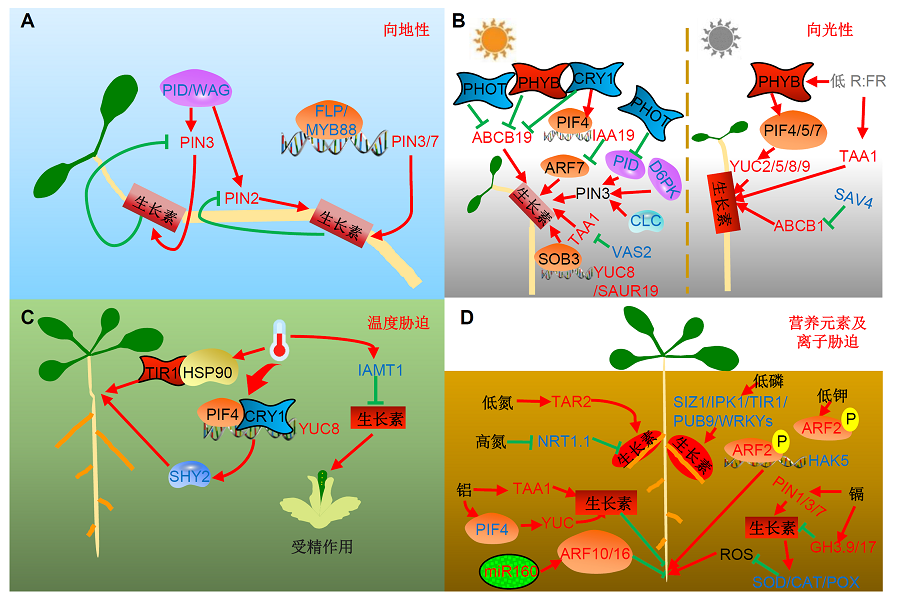
1 生长素与重力响应重力响应是植物应对外界环境的一种适应方式, 而生长素的非对称分布对于重力响应的形成具有关键的调控作用, 这种非对称分布主要通过生长素的极性运输和信号转导来实现。已有研究报道, 在侧根冠和表皮细胞表达的生长素输入载体AUX1的功能缺失性突变体具有减弱的重力反应(Swarup et al., 2001)。而生长素输出载体PIN2和PIN3的突变会分别导致植物根和下胚轴的重力反应缺失(Friml et al., 2002)。生长素信号转导对于重力响应也十分重要, 生长素受体TIR1的突变不仅使植物对生长素呈现出不敏感的响应, 也减弱了根部的重力反应。而AUX/IAA家族中的IAA3/SHY2、IAA17/AXR3、IAA14/SLR和IAA20等的功能获得性突变体均表现出根和下胚轴的重力缺失反应(Berleth et al., 2004; Sato and Yamamoto, 2008)。 植物在感知重力信号后呈现向重力性生长, 同时这一过程可被适时终止, 从而避免植物体过度弯曲生长。最近的研究结果表明, 一种生长素介导的反馈调节机制可终止这一反应。以拟南芥(Arabidopsis thali- ana)黄化苗下胚轴为材料, 通过弯曲力学实验发现, 当下胚轴感受到短时间(2-3小时)的重力刺激后, 便开始呈现出明显的重力响应, 而这种响应随着处理时间的增加逐渐减弱, 在处理30小时后几乎消失。进一步的研究证实, 生长素通过负反馈改变输出载体PIN3的极性分布参与调控这一过程(Rakusova et al., 2016)。之前的研究已发现, PIN3蛋白在质膜上对称分布, 当感应到重力刺激后, PIN3蛋白重新定位在质膜底端, 造成生长素在此端的富集, 而不断增加的生长素会重新改变PIN3的定位, 使其在质膜顶端表达, 从而缓解重力响应, 促进PIN3重新在质膜上形成均匀分布。利用蛋白酶体抑制剂MG132来抑制PIN蛋白的降解或渥曼青霉素(WM)阻止PIN蛋白向液泡的运输, 都可以明显减弱生长素介导的对PIN3极性的改变, 但并没有改变下胚轴对重力的响应, 说明液泡运输和蛋白降解途径都特异性地参与生长素介导的PIN蛋白极性的改变。另外, 通过对AGC激酶VIII亚族基因三突变体wag1/wag2/pid及过表达植株35S::PID的分析, 发现生长素调控PIN3蛋白极性的变化是依赖于PID/WAG所介导的磷酸化途径来实现(Rakusova et al., 2016) (图1A)。 另一方面, 转录因子是否直接参与生长素输出载体PIN所介导的根系重力形态建成过程还没有确切的报道。最新研究结果表明, 在感受到重力信号后, 拟南芥转录因子FLP (FOUR LIPS)和MYB88共同参与调控主根和侧根细胞中PIN3和PIN7基因的时空特异表达; 而PIN3和PIN7通过改变生长素在根两侧的定向运输和分布, 进一步影响根对重力的响应角度, 从而参与根系重力形态建成。这说明生长素运输载体PIN蛋白所参与的重力响应的确受到多个转录因子精细的动态调控, 为进一步了解植物适应重力响应所建立的信号调控网络提供了新思路(Wang et al., 2015) (图1A)。 图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-17/img_1.png<b>图1</b> 生长素介导的环境信号调控植物生长发育的工作模型<br/>(A) 在重力响应过程中, 生长素的非对称分布主要是通过改变外运载体PIN的表达模式来实现的。这一调控过程一方面依赖于PID/WAG对PIN蛋白的磷酸化, 另一方面受转录因子FLP/MYB88直接的转录调节。(B) 在光信号通路过程中, 蓝光受体向光素PHOT通过抑制PID蛋白的活性, 降低对PIN3的磷酸化修饰水平进而参与生长素在下胚轴中的非对称分布。另一蓝光受体隐花色素CRY一方面和蓝光受体向光素PHOT及红光受体PHYB共同抑制ABCB19蛋白表达水平, 另一方面则通过转录因子PIF4直接结合在<i>IAA19</i>的启动子区并抑制下游生长素响应因子ARF7的转录活性参与下胚轴向光的弯曲生长。在植物庇荫反应中, 红光受体PHYB通过PIF家族转录因子直接调控生长素合成相关基因<i>YUCCA</i>的转录水平参与茎的向光弯曲; 而<i>SAV3</i>及<i>SAV4</i>同样参与了植物的庇荫反应。(C) 在响应温度信号通路中, 生长素受体TIR1通过直接与热激蛋白HSP90互作参与植物对高温环境的适应; 另一方面蓝光受体CRY1与PIF4直接互作调控<i>YUC8</i>的转录参与高温对下胚轴的伸长。此外, 高温诱导生长素甲基转移酶IAMT1的表达, 降低植物子房内生长素信号, 进而出现显著的雄性不育表型。(D) 在响应不同离子对植物根系发育的调控网络中, 生长素合成基因<i>TAR2</i>介导了低氮对植物侧根发生的调控, 而硝态氮受体基因<i>NRT1.1</i>通过抑制生长素的极性运输促进了高氮下植物侧根的起始。在低钾处理时, ARF2可被磷酸化, 进而解除对<i>HAK5</i>转录的抑制, 增强植物对钾离子的吸收能力。生长素合成基因<i>TAA1</i>和<i>YUCCAs</i>特异性地在转换区受到铝胁迫的异位诱导表达, 造成此区生长素的积累, 而ARF7可以直接转录调控细胞分裂素合成基因<i>IPT</i>的表达, 最终抑制主根的伸长。镉胁迫则通过维持生长素的内稳态, 进而抑制下游ROS水平, 参与对根伸长的调控。<br/><b>Figure 1</b> An overview of plant growth and development in response to environment signal mediated by auxin <br/>(A) Nonsymmetrical of auxin in response to gravity due to altering expression pattern of auxin polar transporter PIN, which is regulated by phosphorylation of PIN through PID/WAG kinase or transcriptional regulation of FLP/MYB88. (B) In response to phototropism, blue light receptor PHOT reduced phosphorylation of PIN3 by inhibiting PID activity, thus mediated nonsymmetrical of auxin in hypocotyl. Another blue light receptor CRY inhibited ABCB919 expression together with PHOT and PHYB, and decreased ARF7 expression through binding to <i>IAA19</i> promoter with PIF4. In shade condition, PHYB participated in phototropism of shoot through PIF to regulate <i>YUCCA</i> expression. <i>SAV3</i> and <i>SAV4 </i>also mediate plant shade response. (C) In response to temperature signal, auxin receptor TIR1 can interact with HSP90 to participate in high temperature stress. While CRY1, together with PIF4, induced <i>YUC8</i> expression to increase auxin level in hypocotyl under high temperature condition. In addition, high temperature could induce the expression of auxin transmethylase IAMT1, reduced auxin signal in ovary and led to male sterile. (D) Auxin synthesis gene<i> TAR2</i> mediated lateral root development under low nitrogen condition, and nitrate nitrogen receptor NRT1.1 induced lateral root initiation by inhibiting auxin polar transport. ARF2 can be phosphorylated in low potassium, thus relieve the inhibition of HAK5 and enhanced the absorbing ability of phosphorus. <i>TAA1 </i>and <i>YUCCA</i> can be specific induced in the root TZ under aluminum stress, which caused excess auxin. While ARF7 directly regulated IPT expression, leading to the inhibition of root growth. Another metal ion cadmium maintained the auxin homeostasis to reduce downstream ROS level to regulate root growth. Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-17/img_1.png<b>图1</b> 生长素介导的环境信号调控植物生长发育的工作模型<br/>(A) 在重力响应过程中, 生长素的非对称分布主要是通过改变外运载体PIN的表达模式来实现的。这一调控过程一方面依赖于PID/WAG对PIN蛋白的磷酸化, 另一方面受转录因子FLP/MYB88直接的转录调节。(B) 在光信号通路过程中, 蓝光受体向光素PHOT通过抑制PID蛋白的活性, 降低对PIN3的磷酸化修饰水平进而参与生长素在下胚轴中的非对称分布。另一蓝光受体隐花色素CRY一方面和蓝光受体向光素PHOT及红光受体PHYB共同抑制ABCB19蛋白表达水平, 另一方面则通过转录因子PIF4直接结合在<i>IAA19</i>的启动子区并抑制下游生长素响应因子ARF7的转录活性参与下胚轴向光的弯曲生长。在植物庇荫反应中, 红光受体PHYB通过PIF家族转录因子直接调控生长素合成相关基因<i>YUCCA</i>的转录水平参与茎的向光弯曲; 而<i>SAV3</i>及<i>SAV4</i>同样参与了植物的庇荫反应。(C) 在响应温度信号通路中, 生长素受体TIR1通过直接与热激蛋白HSP90互作参与植物对高温环境的适应; 另一方面蓝光受体CRY1与PIF4直接互作调控<i>YUC8</i>的转录参与高温对下胚轴的伸长。此外, 高温诱导生长素甲基转移酶IAMT1的表达, 降低植物子房内生长素信号, 进而出现显著的雄性不育表型。(D) 在响应不同离子对植物根系发育的调控网络中, 生长素合成基因<i>TAR2</i>介导了低氮对植物侧根发生的调控, 而硝态氮受体基因<i>NRT1.1</i>通过抑制生长素的极性运输促进了高氮下植物侧根的起始。在低钾处理时, ARF2可被磷酸化, 进而解除对<i>HAK5</i>转录的抑制, 增强植物对钾离子的吸收能力。生长素合成基因<i>TAA1</i>和<i>YUCCAs</i>特异性地在转换区受到铝胁迫的异位诱导表达, 造成此区生长素的积累, 而ARF7可以直接转录调控细胞分裂素合成基因<i>IPT</i>的表达, 最终抑制主根的伸长。镉胁迫则通过维持生长素的内稳态, 进而抑制下游ROS水平, 参与对根伸长的调控。<br/><b>Figure 1</b> An overview of plant growth and development in response to environment signal mediated by auxin <br/>(A) Nonsymmetrical of auxin in response to gravity due to altering expression pattern of auxin polar transporter PIN, which is regulated by phosphorylation of PIN through PID/WAG kinase or transcriptional regulation of FLP/MYB88. (B) In response to phototropism, blue light receptor PHOT reduced phosphorylation of PIN3 by inhibiting PID activity, thus mediated nonsymmetrical of auxin in hypocotyl. Another blue light receptor CRY inhibited ABCB919 expression together with PHOT and PHYB, and decreased ARF7 expression through binding to <i>IAA19</i> promoter with PIF4. In shade condition, PHYB participated in phototropism of shoot through PIF to regulate <i>YUCCA</i> expression. <i>SAV3</i> and <i>SAV4 </i>also mediate plant shade response. (C) In response to temperature signal, auxin receptor TIR1 can interact with HSP90 to participate in high temperature stress. While CRY1, together with PIF4, induced <i>YUC8</i> expression to increase auxin level in hypocotyl under high temperature condition. In addition, high temperature could induce the expression of auxin transmethylase IAMT1, reduced auxin signal in ovary and led to male sterile. (D) Auxin synthesis gene<i> TAR2</i> mediated lateral root development under low nitrogen condition, and nitrate nitrogen receptor NRT1.1 induced lateral root initiation by inhibiting auxin polar transport. ARF2 can be phosphorylated in low potassium, thus relieve the inhibition of HAK5 and enhanced the absorbing ability of phosphorus. <i>TAA1 </i>and <i>YUCCA</i> can be specific induced in the root TZ under aluminum stress, which caused excess auxin. While ARF7 directly regulated IPT expression, leading to the inhibition of root growth. Another metal ion cadmium maintained the auxin homeostasis to reduce downstream ROS level to regulate root growth.
图1 生长素介导的环境信号调控植物生长发育的工作模型 (A) 在重力响应过程中, 生长素的非对称分布主要是通过改变外运载体PIN的表达模式来实现的。这一调控过程一方面依赖于PID/WAG对PIN蛋白的磷酸化, 另一方面受转录因子FLP/MYB88直接的转录调节。(B) 在光信号通路过程中, 蓝光受体向光素PHOT通过抑制PID蛋白的活性, 降低对PIN3的磷酸化修饰水平进而参与生长素在下胚轴中的非对称分布。另一蓝光受体隐花色素CRY一方面和蓝光受体向光素PHOT及红光受体PHYB共同抑制ABCB19蛋白表达水平, 另一方面则通过转录因子PIF4直接结合在IAA19的启动子区并抑制下游生长素响应因子ARF7的转录活性参与下胚轴向光的弯曲生长。在植物庇荫反应中, 红光受体PHYB通过PIF家族转录因子直接调控生长素合成相关基因YUCCA的转录水平参与茎的向光弯曲; 而SAV3及SAV4同样参与了植物的庇荫反应。(C) 在响应温度信号通路中, 生长素受体TIR1通过直接与热激蛋白HSP90互作参与植物对高温环境的适应; 另一方面蓝光受体CRY1与PIF4直接互作调控YUC8的转录参与高温对下胚轴的伸长。此外, 高温诱导生长素甲基转移酶IAMT1的表达, 降低植物子房内生长素信号, 进而出现显著的雄性不育表型。(D) 在响应不同离子对植物根系发育的调控网络中, 生长素合成基因TAR2介导了低氮对植物侧根发生的调控, 而硝态氮受体基因NRT1.1通过抑制生长素的极性运输促进了高氮下植物侧根的起始。在低钾处理时, ARF2可被磷酸化, 进而解除对HAK5转录的抑制, 增强植物对钾离子的吸收能力。生长素合成基因TAA1和YUCCAs特异性地在转换区受到铝胁迫的异位诱导表达, 造成此区生长素的积累, 而ARF7可以直接转录调控细胞分裂素合成基因IPT的表达, 最终抑制主根的伸长。镉胁迫则通过维持生长素的内稳态, 进而抑制下游ROS水平, 参与对根伸长的调控。 Figure 1 An overview of plant growth and development in response to environment signal mediated by auxin (A) Nonsymmetrical of auxin in response to gravity due to altering expression pattern of auxin polar transporter PIN, which is regulated by phosphorylation of PIN through PID/WAG kinase or transcriptional regulation of FLP/MYB88. (B) In response to phototropism, blue light receptor PHOT reduced phosphorylation of PIN3 by inhibiting PID activity, thus mediated nonsymmetrical of auxin in hypocotyl. Another blue light receptor CRY inhibited ABCB919 expression together with PHOT and PHYB, and decreased ARF7 expression through binding to IAA19 promoter with PIF4. In shade condition, PHYB participated in phototropism of shoot through PIF to regulate YUCCA expression. SAV3 and SAV4 also mediate plant shade response. (C) In response to temperature signal, auxin receptor TIR1 can interact with HSP90 to participate in high temperature stress. While CRY1, together with PIF4, induced YUC8 expression to increase auxin level in hypocotyl under high temperature condition. In addition, high temperature could induce the expression of auxin transmethylase IAMT1, reduced auxin signal in ovary and led to male sterile. (D) Auxin synthesis gene TAR2 mediated lateral root development under low nitrogen condition, and nitrate nitrogen receptor NRT1.1 induced lateral root initiation by inhibiting auxin polar transport. ARF2 can be phosphorylated in low potassium, thus relieve the inhibition of HAK5 and enhanced the absorbing ability of phosphorus. TAA1 and YUCCA can be specific induced in the root TZ under aluminum stress, which caused excess auxin. While ARF7 directly regulated IPT expression, leading to the inhibition of root growth. Another metal ion cadmium maintained the auxin homeostasis to reduce downstream ROS level to regulate root growth.
2 生长素与光信号通路光提供了植物生长发育所必需的能量, 同时光也是一种非常重要的环境信号因子, 可以调控种子萌发、幼苗形态建成以及开花等重要的植物生长发育过程。之前的研究从遗传学和生理学的角度表明, 生长素参与光信号调节的生长发育, 包括向光弯曲、下胚轴伸长和庇荫反应等。因此生长素和光信号转导途径的相互作用在这些生长发育过程的调节中发挥重要作用。 生长素的不对称分布是导致植物向光性生长的主要原因, 这种不对称分布主要由各类生长素运输载体共同调控。最近的遗传与生理分析表明, 蓝光诱导野生型下胚轴中PIN3的重定位, 引起内皮层细胞背光一侧积累更多的PIN3, 导致生长素的不对称分布和下胚轴向光弯曲生长; 在拟南芥蓝光受体突变体phot1及pin3中, 下胚轴生长素不对称分布和向光性生长均被明显抑制。此外, 在AGC激酶VIII亚族PID (PINOID)过表达株系及wag1/wag2/pid缺失突变体中, 蓝光诱导的PIN3背向定位及向光性生长也均被抑制; 进一步的生化分析证实, 蓝光显著抑制野生型植株中PID的转录, 而在phot1/phot2双突变体中这种抑制作用明显减弱, 因此PHOT介导的蓝光信号通过抑制PID的转录水平和PIN3磷酸化来促进PIN3的背向定位, 从而诱导生长素不对称分布及向光性生长(Ding et al., 2011)。另一类AGC激酶VIII亚族D6PK也参与了植物下胚轴向光性弯曲。植物下胚轴向光性弯曲主要由生长素极性转运基因PIN3、PIN4和PIN7调控, 而在拟南芥单突变体d6pk及多突变体d6pk/d6 pkl1/d6pkl2中, PIN3蛋白磷酸化水平均明显下降, 进一步的研究证实, PIN3磷酸化水平的降低最终导致下胚轴中生长素向基部的极性转运受到抑制(Willige et al., 2013)。最新研究发现, 网格蛋白CLC也通过介导PIN3的内吞与侧向定位, 进而调控由蓝光诱导的下胚轴向光面与背光面的生长素不对称分布, 说明网格蛋白也能参与调控植物的向光性生长(Zhang et al., 2017)。ABC转运器家族基因ABCB19近期也被证实参与下胚轴中生长素的极性运输。过表达ABCB19显著增加了下胚轴中生长素的含量, 无论在红光还是蓝光下都减弱了其对下胚轴伸长的抑制。而ABCB19的拟南芥功能缺失突变体对蓝光超敏感, 并能显著恢复cry1突变体下胚轴长的表型。另一方面, 在红光受体突变体phyB以及蓝光受体突变体cry1的质膜中均检测到ABCB19蛋白水平的升高, 且这类蛋白水平的升高不依赖光的调控(Wu et al., 2010)。此外, ABCB19也直接受到另一蓝光受体PHOT1的调控, PHOT1通过与ABCB19直接互作并将其磷酸化, 进而抑制其外运生长素的转运活性, 提高下胚轴中生长素的含量, 最终促进了下胚轴向光性弯曲生长(Christie et al., 2011)。因此, ABCB介导的生长素极性运输参与光信号对下胚轴生长的调控, 而且这一调控依赖于光信号受体CRY1、PHOT1和PHYB (Wu et al., 2010; Chris- tie et al., 2011) (图1B)。 除生长素运输载体外, 生长素信号通路也介导了下胚轴向光性生长反应。已有研究显示, 蓝光可以明显诱导一些阻遏蛋白AUX/IAA的上调表达, 如IAA1、IAA5和IAA18等, 而且这一调控通路依赖于蓝光受体蛋白CRY1 (Folta et al., 2003)。在蓝光下, 下胚轴背光一侧生长素的大量积累促进了IAA19的降解, 释放出有活性的ARF7, 激活与向光性生长相关基因的表达。遗传与生化证据表明, bHLH转录因子家族基因PIF4可以直接结合在IAA19启动子区的G-Box上并直接激活IAA19的表达, 抑制ARF7的活性和下胚轴的向光弯曲, 在pif4突变体中则促进了下胚轴的向光弯曲生长。因此, 生长素的积累促进了IAA19的降解从而增强ARF7的表达和向光性弯曲, 而PIF4通过直接调控IAA19的转录水平参与生长素介导的向光性生长反应(Sun et al., 2013) (图1B)。 在光下, 拟南芥中的转录因子SOB3 (SUPPRE- SSOR OF PHYTOCHROME B4-#3)以及AHL (AT-HOOK MOTIF CONTAINING NUCLEAR LOCALIZED)通过抑制下胚轴的伸长影响植株幼苗的发育。最新的研究发现生长素可能参与了这一过程。功能获得性突变体SOB3-D中YUC8 (YUCCA8)及SAU R19 (SMALL AUXIN UP-REGULATED RNA19)的表达明显下调, 相对应的, 在缺失突变体sob3-6中呈现明显的上调表达, 而超表达SAUR19可以部分恢复SOB3-D下胚轴缺陷的表型, 暗示SAUR19位于SOB3的下游。进一步的实验证实, SOB3可以直接结合在YUC8和SAUR19的启动子区, 从而调控这2个基因的转录, 而外源施加生长素极性运输抑制剂NPA也可以改变SOB3突变体下胚轴的表型, 因此, SOB3参与拟南芥下胚轴的伸长过程是通过直接调控生长素相关基因(如YUC8和SAUR19)来实现的, 这也为生长素介导光信号通路提供了新的生物学证据(Favero et al., 2016) (图1B)。
3 生长素与庇荫反应生长素介导的光信号除了参与向光性反应, 也能参与植物的庇荫反应(shade avoidance syndrome, SAS)。以拟南芥为代表的阴性植物需要一定的庇荫才能正常生长, SAS主要体现在植株形态学的变化, 如茎及叶柄的向光弯曲, 叶片的发育滞后和向下生长等(Casal, 2013; Hersch et al., 2014)。改变红光与远红光的比例(R:FR)可以有效模拟植物的庇荫反应, 因此被广泛应用于庇荫反应研究。在低R:FR条件下, 生长素相关功能缺失突变体taa1 (Tao et al., 2008)、pin3、axr1、tir1 (Keuskamp et al., 2010)或者外源施加NPA抑制生长素极性运输, 以及PEO-IAA阻断生长素信号通路均表现出不敏感的庇荫反应, 暗示生长素在植物的庇荫反应过程中发挥至关重要的作用。进一步研究发现, 低R:FR条件下, 通过在叶片中增加SAV3 (Shade Avoidance 3)/TAA1介导的生长素合成(Tao et al., 2008), 或者在下胚轴中通过PIN3蛋白参与生长素的极性转运来提高内源生长素的水平(Keuskamp et al., 2010), 进而可促进植物叶片及下胚轴中的庇荫反应(Morelli and Ruberti, 2000)。在自然环境中(R:FR较高时), 红光受体PHYB抑制植物的向光反应; 而在荫蔽条件下, PHYB通过调控3个重要的bHLH转录因子PIFs (PIF4、PIF5和PIF7)的转录活性促进了植物的向光性。进一步研究发现, 这类PIFs介导的向光反应主要依赖于一些生长素合成关键基因YUCCAs, 低R:FR可以显著诱导YUC2、YUC5、YUC8和YUC9的表达, 功能缺失突变体yuc2/yuc5/ yuc8/yuc9表现出严重的庇荫反应缺失表型。而利用子叶特异性表达的启动子FRO6构建诱导性载体YUC3i (FRO6::XVE::YUC3)并进行分析, 发现在高R:FR条件下, YUC3的诱导表达并未检测到明显的向光性缺陷, 暗示子叶中表达的YUC3对于植物向光性响应是必需的。此外, YUC3可以恢复pif4/pi5/pif7缺陷的向光性表型, 证实PIF介导的YUC基因的转录水平与植物的向光性反应有关(Goyal et al., 2016) (图1B)。 最新的研究发现, SAV4通过调控ABCB1介导的生长素极性运输参与植物下胚轴的庇荫反应。功能缺失突变体sav4具有明显的庇荫反应缺陷表型, 具体体现在庇荫条件下sav4下胚轴伸长不明显, 且生长素由顶部向基部的运输受到抑制。SAV4编码一个未知功能蛋白, 进一步的结构分析显示, 其包含一个已被证实的互作结构域(TPR), 暗示SAV4可能通过蛋白间相互作用实现其生物学功能。表达模式分析表明, SAV4主要在叶原基、根尖分生组织以及下胚轴的维管组织中表达, 而亚细胞定位发现SAV4在质膜及核中均有表达。进一步的遗传实验显示, 生长素类似物毒莠定所诱导的下胚轴伸长可以恢复sav4/taa1短下胚轴的表型, 而不能恢复sav4/dwf4短下胚轴的表型, 证实sav4对庇荫反应不敏感可能是通过影响生长素合成或转运, 而不是影响生长素信号来实现。通过共转染将SAV4及ABCB1同时转化烟草(Nicotiana tabacum), 发现SAV4可以阻断ABCB1介导的生长素外运。在功能缺失突变体sav4中可以检测到生长素由基部向顶部的运输明显增强, 进一步证实SAV4通过抑制ABCB1的转运活性参与生长素介导的植物庇荫反应(Ge et al., 2017) (图1B)。
CasalJJ (2013). Photoreceptor signaling networks in plant responses to shade.Annu Rev Plant Biol 64, 403-427. [本文引用: 1]
[5]
ChristieJM, YangH, RichterGL, SullivanS, ThomsonCE, LinJ, TitapiwatanakunB, EnnisM, KaiserliE, LeeOR, AdamecJ, PeerWA, MurphyAS (2011). phot1 inhibition of ABCB19 primes lateral auxin fluxes in the shoot apex required for phototropism.PLoS Biol 9, e1001076. [本文引用: 2]
[6]
DebS, SankaranarayananS, WewalaG, WiddupE, Sa- muelMA (2014). The S-domain receptor kinase Arabidopsis receptor kinase2 and the U box/armadillo repeat-containing E3 ubiquitin ligase9 module mediates lateral root development under phosphate starvation in Arabidopsis.Plant Physiol 165, 1647-1656. [本文引用: 1]
[7]
DevaiahBN, KarthikeyanAS, RaghothamaKG (2007). WRKY75 transcription factor is a modulator of phosphate acquisition and root development in Arabidopsis.Plant Physiol 143, 1789-1801. [本文引用: 1]
[8]
DingZJ, Galván-AmpudiaCS, DemarsyE, ŁangowskiL, Kleine-VehnJ, FanYW, MoritaMT, TasakaM, FankhauserC, OffringaR, FrimlJ (2011). Light-mediated polarization of the PIN3 auxin transporter for the phototropic response in Arabidopsis.Nat Cell Biol 13, 447-452. [本文引用: 1]
[9]
ElobeidM, GöbelC, FeussnerI, PolleA (2012). Cadmium interferes with auxin physiology and lignification in poplar.J Exp Bot 63, 1413-1421. [本文引用: 1]
[10]
FaveroDS, JacquesCN, IwaseA, LeKN, ZhaoJF, SugimotoK, NeffMM (2016). SUPPRESSOR OF PHYTOCHROME B4-#3 represses genes associated with auxin signaling to modulate hypocotyl growth.Plant Phy- siol 171, 2701-2716. [本文引用: 1]
[11]
FoltaKM, PontinMA, Karlin-NeumannG, BottiniR, SpaldingEP (2003). Genomic and physiological studies of early cryptochrome 1 action demonstrate roles for au- xin and gibberellin in the control of hypocotyl growth by blue light.Plant J 36, 203-214. [本文引用: 1]
HerschM, LorrainS, de WitM, TrevisanM, LjungK, BergmannS, FankhauserC (2014). Light intensity modu- lates the regulatory network of the shade avoidance response in Arabidopsis.Proc Natl Acad Sci USA 111, 6515-6520. [本文引用: 1]
[16]
HuYF, ZhouGY, NaXF, YangLJ, NanWB, LiuX, ZhangYQ, LiJL, BiYR (2013). Cadmium interferes with maintenance of auxin homeostasis in Arabidopsis seedlings.J Plant Physiol 170, 965-975. [本文引用: 1]
[17]
KeuskampDH, PollmannS, VoesenekLACJ, PeetersAJM, PierikR (2010). Auxin transport through PIN- FORMED 3 (PIN3) controls shade avoidance and fitness during competition.Proc Natl Acad Sci USA 107, 22740-22744. [本文引用: 2]
[18]
KollmeierM, FelleHH, HorstWJ (2000). Genotypical differences in aluminum resistance of maize are expressed in the distal part of the transition zone. Is reduced basi- petal auxin flow involved in inhibition of root elongation by aluminum?Plant Physiol 122, 945-956. [本文引用: 1]
[19]
KroukG, LacombeB, BielachA, Perrine-WalkerF, MalinskaK, MounierE, HoyerovaK, TillardP, LeonS, LjungK, ZazimalovaE, BenkovaE, NacryP, GojonA (2010). Nitrate-regulated auxin transport by NRT1.1 defines a mechanism for nutrient sensing in plants.Dev Cell 18, 927-937. [本文引用: 2]
[20]
KuoHF, ChangTY, ChiangSF, WangWD, CharngYY, ChiouTJ (2014). Arabidopsis inositol pentakisphosphate 2-kinase, AtIPK1, is required for growth and modulates phosphate homeostasis at the transcriptional level.Plant J 80, 503-515. [本文引用: 1]
[21]
LiuGC, GaoS, TianHY, WuWW, RobertHS, DingZJ (2016). Local transcriptional control of YUCCA regulates auxin promoted root-growth inhibition in response to aluminium stress in Arabidopsis.PLoS Genet 12, e1006360. [本文引用: 1]
[22]
MaDB, LiX, GuoYX, ChuJF, FangS, YanCY, NoelJP, LiuHT (2016). Cryptochrome 1 interacts with PIF4 to regulate high temperature-mediated hypocotyl elongation in response to blue light.Proc Natl Acad Sci USA 113, 224-229. [本文引用: 1]
[23]
MaWY, LiJJ, QuBY, HeX, ZhaoXQ, LiB, FuXD, TongYP (2014). Auxin biosynthetic gene TAR2 is involved in low nitrogen-mediated reprogramming of root architecture in Arabidopsis. Plant J 78, 70-79. [本文引用: 2]
[24]
MiuraK, LeeJ, GongQQ, MaSS, JinJB, YooCY, MiuraT, SatoA, BohnertHJ, HasegawaPM (2011). SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation.Plant Physiol 155, 1000-1012. [本文引用: 1]
Pérez-TorresCA, López-BucioJ, Cruz-RamírezA, Ibarra- LacletteE, DharmasiriS, EstelleM, Herrera-EstrellaL (2008). Phosphate availability alters lateral root deve- lopment in Arabidopsis by modulating auxin sensitivity via a mechanism involving the TIR1 auxin receptor.Plant Cell 20, 3258-3272. [本文引用: 1]
[27]
RakusovaH, AbbasM, HanH, SongS, RobertHS, FrimlJ (2016). Termination of shoot gravitropic responses by auxin feedback on PIN3 polarity.Curr Biol 26, 3026-3032. [本文引用: 2]
[28]
SatoA, YamamotoKT (2008). Overexpression of the non- canonical Aux/IAA genes causes auxin-related aberrant phenotypes in Arabidopsis. Physiol Plant 133, 397-405. [本文引用: 1]
[29]
SunJ, QiL, LiY, ChuJ, LiC (2012). PIF4-mediated activation of YUCCA8 expression integrates temperature into the auxin pathway in regulating Arabidopsis hypocotyl growth. PLoS Genet 8, e1002594. [本文引用: 2]
[30]
SunJQ, QiLL, LiYN, ZhaiQZ, LiCY (2013). PIF4 and PIF5 transcription factors link blue light and auxin to re- gulate the phototropic response in Arabidopsis.Plant Cell 25, 2102-2114. [本文引用: 1]
[31]
SunP, TianQY, ChenJ, ZhangWH (2010). Aluminium- induced inhibition of root elongation in Arabidopsis is mediated by ethylene and auxin.J Exp Bot 61, 347-356. [本文引用: 1]
[32]
SvistoonoffS, CreffA, ReymondM, Sigoillot-ClaudeC, RicaudL, BlanchetA, NussaumeL, DesnosT (2007). Root tip contact with low-phosphate media reprograms plant root architecture.Nat Genet 39, 792-796. [本文引用: 1]
[33]
SwarupR, FrimlJ, MarchantA, LjungK, SandbergG, PalmeK, BennettM (2001). Localization of the auxin permease AUX1 suggests two functionally distinct hormone transport pathways operate in the Arabidopsis root apex.Genes Dev 15, 2648-2653. [本文引用: 1]
[34]
TaoY, FerrerJL, LjungK, PojerF, HongF, LongJA, LiL, MorenoJE, BowmanME, IvansLJ, ChengY, LimJ, ZhaoYD, BallareCL, SandbergG, NoelJP, ChoryJ (2008). Rapid synthesis of auxin via a new tryptophan- dependent pathway is required for shade avoidance in plants.Cell 133, 164-176. [本文引用: 2]
[35]
WangHZ, YangKZ, ZouJJ, ZhuLL, XieZD, MoritaMT, TasakaM, FrimlJ, GrotewoldE, BeeckmanT, VannesteS, SackF, LeJ (2015). Transcriptional regulation of PIN genes by FOUR LIPS and MYB88 during Arabidopsis root gravitropism. Nat Commun 6, 8822. [本文引用: 1]
[36]
WangRH, ZhangY, KiefferM, YuH, KepinskiS, EstelleM (2016). HSP90 regulates temperature-dependent seed- ling growth in Arabidopsis by stabilizing the auxin co-receptor F-box protein TIR1.Nat Commun 7, 10269. [本文引用: 1]
[37]
WilligeBC, AhlersS, ZourelidouM, BarbosaICR, DemarsyE, TrevisanM, DavisPA, RoelfsemaMRG, HangarterR, FankhauserC, SchwechheimerC (2013). D6PK AGCVIII kinases are required for auxin transport and phototropic hypocotyl bending in Arabidopsis.Plant Cell 25, 1674-1688. [本文引用: 1]
[38]
WuGS, CameronJN, LjungK, SpaldingEP (2010). A role for ABCB19-mediated polar auxin transport in seedling photomorphogenesis mediated by cryptochrome 1 and phytochrome B.Plant J 62, 179-191. [本文引用: 2]
[39]
XuL, JinL, LongL, LiuLL, HeX, GaoW, ZhuLF, ZhangXL (2012). Overexpression of GbWRKY1 positively regu- lates the Pi starvation response by alteration of auxin sensitivity in Arabidopsis. Plant Cell Rep 31, 2177-2188. [本文引用: 1]
[40]
YangZB, GengXY, HeCM, ZhangF, WangR, HorstWJ, DingZJ (2014). TAA1-regulated local auxin biosynthesis in the root-apex transition zone mediates the aluminum- induced inhibition of root growth in Arabidopsis.Plant Cell 26, 2889-2904. [本文引用: 3]
[41]
YuCL, SunCD, ShenCJ, WangSK, LiuF, LiuY, ChenYL, LiCY, QianQ, AryalB, GeislerM, JiangDA, QiYH (2015). The auxin transporter, OsAUX1, is involved in primary root and root hair elongation and in Cd stress responses in rice ( Oryza sativa L.). Plant J 83, 818-830. [本文引用: 1]
[42]
YuanH, LiuD (2008). Signaling components involved in plant responses to phosphate starvation.J Integr Plant Biol 50, 849-859. [本文引用: 1]
[43]
ZhangY, YuQQ, JiangN, YanX, WangC, WangQM, LiuJZ, ZhuMY, BednarekSY, XuJ, PanJW (2017). Clathrin regulates blue light-triggered lateral auxin distribution and hypocotyl phototropism in Arabidopsis.Plant Cell Environ 40, 165-176. [本文引用: 1]
[44]
ZhaoS, ZhangML, MaTL, WangY (2016). Phosphorylation of ARF2 relieves its repression of transcription of the K+ transporter gene HAK5 in response to low potassium stress. Plant Cell 28, 3005-3019. [本文引用: 2]
... 重力响应是植物应对外界环境的一种适应方式, 而生长素的非对称分布对于重力响应的形成具有关键的调控作用, 这种非对称分布主要通过生长素的极性运输和信号转导来实现.已有研究报道, 在侧根冠和表皮细胞表达的生长素输入载体AUX1的功能缺失性突变体具有减弱的重力反应(Swarup et al., 2001).而生长素输出载体PIN2和PIN3的突变会分别导致植物根和下胚轴的重力反应缺失(Friml et al., 2002).生长素信号转导对于重力响应也十分重要, 生长素受体TIR1的突变不仅使植物对生长素呈现出不敏感的响应, 也减弱了根部的重力反应.而AUX/IAA家族中的IAA3/SHY2、IAA17/AXR3、IAA14/SLR和IAA20等的功能获得性突变体均表现出根和下胚轴的重力缺失反应(Berleth et al., 2004; Sato and Yamamoto, 2008). ...
... 重力响应是植物应对外界环境的一种适应方式, 而生长素的非对称分布对于重力响应的形成具有关键的调控作用, 这种非对称分布主要通过生长素的极性运输和信号转导来实现.已有研究报道, 在侧根冠和表皮细胞表达的生长素输入载体AUX1的功能缺失性突变体具有减弱的重力反应(Swarup et al., 2001).而生长素输出载体PIN2和PIN3的突变会分别导致植物根和下胚轴的重力反应缺失(Friml et al., 2002).生长素信号转导对于重力响应也十分重要, 生长素受体TIR1的突变不仅使植物对生长素呈现出不敏感的响应, 也减弱了根部的重力反应.而AUX/IAA家族中的IAA3/SHY2、IAA17/AXR3、IAA14/SLR和IAA20等的功能获得性突变体均表现出根和下胚轴的重力缺失反应(Berleth et al., 2004; Sato and Yamamoto, 2008). ...
... 重力响应是植物应对外界环境的一种适应方式, 而生长素的非对称分布对于重力响应的形成具有关键的调控作用, 这种非对称分布主要通过生长素的极性运输和信号转导来实现.已有研究报道, 在侧根冠和表皮细胞表达的生长素输入载体AUX1的功能缺失性突变体具有减弱的重力反应(Swarup et al., 2001).而生长素输出载体PIN2和PIN3的突变会分别导致植物根和下胚轴的重力反应缺失(Friml et al., 2002).生长素信号转导对于重力响应也十分重要, 生长素受体TIR1的突变不仅使植物对生长素呈现出不敏感的响应, 也减弱了根部的重力反应.而AUX/IAA家族中的IAA3/SHY2、IAA17/AXR3、IAA14/SLR和IAA20等的功能获得性突变体均表现出根和下胚轴的重力缺失反应(Berleth et al., 2004; Sato and Yamamoto, 2008). ...
... 铝的毒害作用被认为是酸性土壤中植物生长最主要的限制因素之一.铝主要通过抑制植物根尖的生长来产生毒害, 而生长素在植物根伸长过程中具有非常重要的调控作用.因此, 阐述铝毒害和生长素之间的相互关系一直是研究的热点.最新的研究表明, 铝胁迫可以诱导生长素合成关键基因TAA1和YUCCA在根尖转化区(TZ)特异性的异位表达, 造成生长素在此区过量积累, 从而抑制主根的伸长.进一步的研究证实, bHLH转录因子PIF4在TZ区受到铝胁迫的特异性诱导, 进而直接结合在YUC5/YUC8/YUC9的启动子区, 诱导这些基因的上调表达, 而且这一调控依赖于乙烯信号(Yang et al., 2014; Liu et al., 2016).另一方面, 铝毒害诱导大量乙烯的产生, 作为信号分子, 乙烯增强了生长素转运载体PIN2和AUX1的表达, 改变了二者所介导的根尖生长素分布, 使突变体aux1-7及pin2均表现出明显的抗铝表型(Sun et al., 2010); 而外源施加萘基酞氨酸(NPA)阻断生长素极性运输也呈明显的耐铝表型(Kollmeier et al., 2000).最新的研究证实, 生长素响应因子ARFs直接参与铝毒害对根伸长的抑制.ARF10和ARF16在根尖表皮及皮层特异性表达, 且ARF10和ARF16在铝存在的情况下受miR160的直接调控, 而双突变体arf10/arf16具有耐铝表型, 暗示了ARF10和ARF16参与介导铝毒害对根伸长的抑制依赖于miR160.此外, 通过转录组测序分析, 一些细胞壁相关基因也被证实受ARF10和ARF16的调控.以上结果表明, 铝毒害通过调控局部生长素合成、极性运输和相应信号转导共同参与植物根系的正常生长(Yang et al., 2014).另一重要的植物激素细胞分裂素最近也被证实与生长素以协同作用的方式共同参与调控植物根系对铝毒害的响应过程.铝毒害诱导细胞分裂素合成关键基因IPT (ISOPENTENYL TRANSFERASE)在TZ区特异性异位表达, ipt突变体具有明显耐铝的表型.进一步分析发现, IPT基因的启动子区具有生长素响应因子ARF的结合元件.通过染色质免疫沉淀技术及酵母单杂交实验证实, ARF7可以直接结合在IPT的启动子区并诱导IPT基因的表达, 进而介导了铝毒害下细胞分裂素在TZ区的过量积累, 从而抑制主根的伸长(Yang et al., 2017) (图1D). ...
1 2007
... 除了氮素以外, 磷和钾元素的吸收和利用也对植物的正常生长发育至关重要.植物体内存在依赖生长素的信号通路以响应低磷胁迫(Yuan and Liu, 2008).在供磷不足的条件下, 植物根尖会积累过量的生长素, 从而抑制主根的伸长和促进侧根的发生, 生长素极性运输抑制剂BFA则可以减弱低磷诱导的侧根发生, 暗示生长素的极性运输特异性参与植物对低磷的响应.进一步研究发现, 生长素特异性参与的对低磷响应是通过调控与植物主侧根发育相关基因的表达实现.其中, LPR1/2 (LOW PHOSPHATE ROOT) (Svistoonoff et al., 2007)特异性地参与生长素所介导的主根对低磷的响应, 而SIZ1 (Miura et al., 2011)、IPK1 (Kuo et al., 2014)、TIR1 (Perez-Torres et al., 2008)、PUB9 (Deb et al., 2014)及WRKYs (Devaiah et al., 2007; Xu et al., 2012)则共同调控低磷胁迫下生长素对可见侧根发生的响应(图1D). ...
1 2001
... 重力响应是植物应对外界环境的一种适应方式, 而生长素的非对称分布对于重力响应的形成具有关键的调控作用, 这种非对称分布主要通过生长素的极性运输和信号转导来实现.已有研究报道, 在侧根冠和表皮细胞表达的生长素输入载体AUX1的功能缺失性突变体具有减弱的重力反应(Swarup et al., 2001).而生长素输出载体PIN2和PIN3的突变会分别导致植物根和下胚轴的重力反应缺失(Friml et al., 2002).生长素信号转导对于重力响应也十分重要, 生长素受体TIR1的突变不仅使植物对生长素呈现出不敏感的响应, 也减弱了根部的重力反应.而AUX/IAA家族中的IAA3/SHY2、IAA17/AXR3、IAA14/SLR和IAA20等的功能获得性突变体均表现出根和下胚轴的重力缺失反应(Berleth et al., 2004; Sato and Yamamoto, 2008). ...
2 2008
... 生长素介导的光信号除了参与向光性反应, 也能参与植物的庇荫反应(shade avoidance syndrome, SAS).以拟南芥为代表的阴性植物需要一定的庇荫才能正常生长, SAS主要体现在植株形态学的变化, 如茎及叶柄的向光弯曲, 叶片的发育滞后和向下生长等(Casal, 2013; Hersch et al., 2014).改变红光与远红光的比例(R:FR)可以有效模拟植物的庇荫反应, 因此被广泛应用于庇荫反应研究.在低R:FR条件下, 生长素相关功能缺失突变体taa1 (Tao et al., 2008)、pin3、axr1、tir1 (Keuskamp et al., 2010)或者外源施加NPA抑制生长素极性运输, 以及PEO-IAA阻断生长素信号通路均表现出不敏感的庇荫反应, 暗示生长素在植物的庇荫反应过程中发挥至关重要的作用.进一步研究发现, 低R:FR条件下, 通过在叶片中增加SAV3 (Shade Avoidance 3)/TAA1介导的生长素合成(Tao et al., 2008), 或者在下胚轴中通过PIN3蛋白参与生长素的极性转运来提高内源生长素的水平(Keuskamp et al., 2010), 进而可促进植物叶片及下胚轴中的庇荫反应(Morelli and Ruberti, 2000).在自然环境中(R:FR较高时), 红光受体PHYB抑制植物的向光反应; 而在荫蔽条件下, PHYB通过调控3个重要的bHLH转录因子PIFs (PIF4、PIF5和PIF7)的转录活性促进了植物的向光性.进一步研究发现, 这类PIFs介导的向光反应主要依赖于一些生长素合成关键基因YUCCAs, 低R:FR可以显著诱导YUC2、YUC5、YUC8和YUC9的表达, 功能缺失突变体yuc2/yuc5/ yuc8/yuc9表现出严重的庇荫反应缺失表型.而利用子叶特异性表达的启动子FRO6构建诱导性载体YUC3i (FRO6::XVE::YUC3)并进行分析, 发现在高R:FR条件下, YUC3的诱导表达并未检测到明显的向光性缺陷, 暗示子叶中表达的YUC3对于植物向光性响应是必需的.此外, YUC3可以恢复pif4/pi5/pif7缺陷的向光性表型, 证实PIF介导的YUC基因的转录水平与植物的向光性反应有关(Goyal et al., 2016) (图1B). ... ... )或者外源施加NPA抑制生长素极性运输, 以及PEO-IAA阻断生长素信号通路均表现出不敏感的庇荫反应, 暗示生长素在植物的庇荫反应过程中发挥至关重要的作用.进一步研究发现, 低R:FR条件下, 通过在叶片中增加SAV3 (Shade Avoidance 3)/TAA1介导的生长素合成(Tao et al., 2008), 或者在下胚轴中通过PIN3蛋白参与生长素的极性转运来提高内源生长素的水平(Keuskamp et al., 2010), 进而可促进植物叶片及下胚轴中的庇荫反应(Morelli and Ruberti, 2000).在自然环境中(R:FR较高时), 红光受体PHYB抑制植物的向光反应; 而在荫蔽条件下, PHYB通过调控3个重要的bHLH转录因子PIFs (PIF4、PIF5和PIF7)的转录活性促进了植物的向光性.进一步研究发现, 这类PIFs介导的向光反应主要依赖于一些生长素合成关键基因YUCCAs, 低R:FR可以显著诱导YUC2、YUC5、YUC8和YUC9的表达, 功能缺失突变体yuc2/yuc5/ yuc8/yuc9表现出严重的庇荫反应缺失表型.而利用子叶特异性表达的启动子FRO6构建诱导性载体YUC3i (FRO6::XVE::YUC3)并进行分析, 发现在高R:FR条件下, YUC3的诱导表达并未检测到明显的向光性缺陷, 暗示子叶中表达的YUC3对于植物向光性响应是必需的.此外, YUC3可以恢复pif4/pi5/pif7缺陷的向光性表型, 证实PIF介导的YUC基因的转录水平与植物的向光性反应有关(Goyal et al., 2016) (图1B). ...


{kind=link}
{kind=link}