Abstract: Plants have evolved a continuously co-evolutionary and sophisticated immune surveillance system to help themselves adapt to environmental changes. Disease resistance (R) proteins play an important role in activating potent defense responses by recognizing specific effectors to protect against microbial infection. In this review, we outline recent progress in plant R proteins, focusing on the structure and function, pathogen recognition, homeostasis control mechanisms, and potential activation mechanisms in R-mediated defense responses. These studies provide valuable information for developing transgenic plants with improved pathogen resistance.
图2 抗病蛋白稳定性调控 (A) 抗病基因的转录后调控; (B) 抗病蛋白的装配与成熟; (C) 抗病蛋白的降解 Figure 2 The homeostasis regulation of R proteins (A) Post-transcriptional regulation of R genes; (B) Chaperone-mediated R protein complex assembly; (C) R protein degradation
4.2 抗病蛋白的稳定性调控抗病蛋白的稳定性受蛋白折叠装配及降解的影响。抗病蛋白的装配过程需要分子伴侣的协助(图2B)。热激蛋白HSP90 (Heat Shock Protein 90)与分子共伴侣蛋白SGT1 (Suppressor of G2 Allele of Skp1)和RAR1 (Required for Mla12 Resistance 1)共同调控抗病蛋白的正确折叠与稳定。hsp90突变体中抗病蛋白RPM1和RPS5表达量降低。rar1和sgt1突变体中RPS5介导的病原菌抗性减弱。生化实验证明, HSP90的ATPase结构域能够与SGT1蛋白CS (Present in CHP and SGT1 Proteins)结构域互作, 而RAR1蛋白的2个锌结合结构域CHORD-I (Cysteine and Histidine Rich Domain)和CHORD-II分别与HSP90和SGT1蛋白结合, 形成复合体, 共同调控抗病蛋白的成熟(Azevedo et al., 2002; Hubert et al., 2003; Huang et al., 2014)。之前的研究证明, 磷酸酶PP5 (Protein Phosphatase 5)能够与HSP90相互作用, 调控抗病蛋白I-2介导的免疫响应(De La Fuente Bentem van, 2005)。而最新研究表明, 磷酸酶ibr5 (indole 3 butyric acid response 5)缺失导致抗病蛋白CHS3 (Chilling Sensitive 3)、RPM1和RPS4表达量降低, 同时IBR5能够与HSP90-RAR1-SGT1复合体相互作用, 证明磷酸酶能够作为分子共伴侣蛋白与HSP90复合体一起调节抗病蛋白的稳定性(Liu et al., 2015)。 抗病蛋白的降解受到泛素化蛋白酶体介导的降解途径调控(图2C)。研究表明, F-box蛋白cpr1 (constitutive expresser of PR genes 1)功能缺失突变体中抗病蛋白SNC1和RPS2 (Resistance to Pseu- domonas syringae 2)积累, 表明SCFCPR1E3复合体调控抗病蛋白SNC1和RPS2的降解(Cheng et al., 2011)。snc1增强子筛选实验证明, E4连接酶MUSE3 (Mutants snc1 Enhancing 3)、TRAR蛋白MUS13和MUSE14能够与SCFCPR1E3复合体互作, 共同调控抗病蛋白SNC1和RPS2的降解(Huang et al., 2014, 2016)。MUSE7编码1个功能未知的激酶底物蛋白, 负调控SNC1、RPS2和RPM1蛋白的积累(Johnson et al., 2016)。与HSP90复合体作为分子伴侣调控抗病蛋白的装配与成熟不同, HSP90和SGT1也能够与SKP1 (S Phase Kinase Associated Protein 1)-CUL- LIN-F-box-E3泛素复合体相互作用, 调控抗病蛋白的降解(Willhoft et al., 2017)。遗传分析表明, hsp90功能获得突变体中抗病蛋白SNC1积累(Huang et al., 2014)。sgt1b缺失突变能够抑制rar1突变体中RPS5的积累(Holt et al., 2005)。生化实验证明, HSP90和SGT1b能够与SKP1和CULLIN1互作, 促进蛋白降解; RAR1也能够结合COP9 (Constitutive Photo- morpho-genesis Complex 9)蛋白, 抑制SCF复合体的活性, 进而调控抗病蛋白的降解(Azevedo et al., 2002)。另外, SGT1a和SGT1b能够与SRFR1 (Suppressor of rps4-RLD)蛋白互作, 负调控抗病蛋白SNC1、RPS2和RPS4的积累(Li et al., 2010a)。E1泛素连接酶UBA1 (Ubiquitin Activating 1)与26S蛋白酶体亚基RPN1a (Proteasome Regulatory Subunint S2 1A)、RPN8a (RP Non-ATPase Subunit 8A)和RPT2a (Regulatory Particle AAA-ATPase 2A)参与调控抗病蛋白的降解(Goritschnig et al., 2007; Yao et al., 2012)。
4.3 环境因素对抗病蛋白稳定性的影响除内在调控方式外, 温度、湿度和昼夜节律等环境因素也能够影响抗病蛋白的稳定性。遗传筛选实验表明, 许多免疫自激活突变体均表现出温度依赖的表型, 如chs1/2/3、snc1-1、bon1-1和mkp1等。chs1/2/3在16 °C下表现为生长缺陷, 而22°C下生长正常; snc1-1和bon1-1等突变体在22°C下植株矮小、叶片卷曲, 而这种缺陷表型可以被高温恢复或部分恢复(Hua et al., 2001; Yang and Hua, 2004; Yang et al., 2010; Wang et al., 2013; Zbierzak et al., 2013)。高温影响抗病蛋白与病原菌间的相互作用, 并且高温能够抑制抗病蛋白从稳定状态向激活状态转变。此外, 高温下植物体内水杨酸(salicylic acid, SA)和脱落酸(absci- sic acid, ABA)等激素水平下降, 增强了植物的抗菌性(Mang et al., 2012)。环境湿度和昼夜周期节律变化也能够调控抗病蛋白的稳定性。例如, 高湿度能够抑制抗病蛋白SSI (Suppressor of Salicylic Acid Insensitive)与SLH1 (an allele of RRS1)依赖的免疫激活(Zhou et al., 2004; Noutoshi et al., 2005); 昼夜节律可以影响抗病基因RPP4 (Resistant to Peronospora parasitica 4)的转录(Wang et al., 2011)。
5 抗病蛋白介导的信号转导在植物免疫响应过程中, 抗病蛋白的激活导致下游免疫信号被激活, 从而调控植物对病原菌的抗性。研究表明许多抗病蛋白能够进入细胞核激活免疫响应(图3)。对SNC1蛋白的研究表明, 当其进入细胞核后, SNC1的TIR结构域与转录抑制因子TPR1 (Topless Related 1)结合, 抑制免疫负调控因子DND1 (Defense No Death 1)和DND2的转录(Zhu et al., 2010); SNC1还能够与bHLH84 (Helix Loop Helix 84)转录因子相互作用, 激活抗病相关基因的转录(Xu et al., 2014)。通过对snc1抑制子的筛选发现, MOS3和MOS7能够调控SNC1在细胞核中的积累。MOS3是一个核孔蛋白, MOS7为果蝇与人类(Homo sapiens)核孔蛋白Nup88的同源蛋白。在mos3和mos7缺失突变体中, 定位于细胞核的SNC1减少, 不能激活转录因子的表达, 从而抑制SNC1依赖的免疫响应。该研究结果表明, 核孔复合体组分MOS3和MOS7能够调控抗病蛋白的细胞核定位, 影响SNC1介导的下游转录信号激活(Zhang and Li, 2005; Cheng et al., 2009), 但其具体调控机制还不清楚。而最新研究表明, 在抗病蛋白RPS2激活过程中, 与核孔复合体结合的跨膜核孔蛋白CPR5 (Constitutive Expresser of PR Genes 5)构象改变, 降低其与CKIs (Cyclin-dep- endent Kinase Inhibitors)的亲和性, 触发Rb (Retinoblastoma)和E2F介导的细胞周期途径, 调控抗病基因的表达, 引发细胞程序性死亡, 从而激活ETI免疫响应(Gu et al., 2016)。除此以外, 某些含有DNA结合结构域的抗病蛋白进入细胞核后, 能够直接与转录因子结合, 激活抗病相关基因的转录。例如, 抗病蛋白MLA10入核后, 其N端结构域与WRKY1/2转录因子相互作用, 解除其对MYB6 (MYB Domain Protein 6)的抑制, 启动植物抗病反应(Chang et al., 2013); 而Pb1 (Panicle blast 1)与WRKY45转录因子结合后, 抑制WRKY45被蛋白酶体降解, 激活SA依赖的抗病信号通路(Inoue et al., 2013)。 图3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-2-250/img_3.png<b>图3</b> 抗病蛋白介导的植物免疫响应过程<br/>抗病蛋白激活触发的信号转导过程, 包括抗病信号激活、转录因子调控以及水杨酸(SA)信号通路激活。<br/><b>Figure 3</b> R protein mediated plant immune response pathway<br/>The regulation mechanisms of R protein involved signaling components, including the activation of R-related proteins, transcription regulators and salicylic acid (SA) dependent signaling. Figure 3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-2-250/img_3.png<b>图3</b> 抗病蛋白介导的植物免疫响应过程<br/>抗病蛋白激活触发的信号转导过程, 包括抗病信号激活、转录因子调控以及水杨酸(SA)信号通路激活。<br/><b>Figure 3</b> R protein mediated plant immune response pathway<br/>The regulation mechanisms of R protein involved signaling components, including the activation of R-related proteins, transcription regulators and salicylic acid (SA) dependent signaling.
图3 抗病蛋白介导的植物免疫响应过程 抗病蛋白激活触发的信号转导过程, 包括抗病信号激活、转录因子调控以及水杨酸(SA)信号通路激活。 Figure 3 R protein mediated plant immune response pathway The regulation mechanisms of R protein involved signaling components, including the activation of R-related proteins, transcription regulators and salicylic acid (SA) dependent signaling.
目前, 对于抗病基因下游信号通路的研究表明, EDS1 (Enhanced Disease Susceptibility 1)和NDR1 (Non Race Specific Disease Resistance 1)是抗病蛋白下游2个关键的调节因子(图3)。TNL抗病蛋白介导的免疫响应依赖于EDS1, 效应蛋白与EDS1结合后抑制PTI, 随后EDS1与抗病蛋白间的互作改变, 激活抗病蛋白依赖的免疫反应(Heidrich et al., 2011)。抗病蛋白-EDS1-SRFR1通过相互作用形成复合体, 无效应蛋白时, 细胞质中的EDS1与SRFR1结合后被固定在内质网上, 形成稳定复合体后不发挥作用; 效应蛋白入侵后与EDS1结合, 抗病蛋白-EDS1从SRFR1上释放, 并进一步激活下游防御反应基因的表达, 触发免疫响应(Kim et al., 2009, 2010)。另外, SRFR1又能够与TCP (Teosinte Branched1 Cycloidea PCF)相互作用, 拮抗调控RPS4介导的免疫响应(Kim et al., 2014)。EDS1也能够与PAD4 (Phytoalexin Deficient 4)和SAG101 (Senescence Associated Gene 101)相互作用, 影响EDS1的定位, 共同调控植物免疫响应。SAG101可以将EDS1定位于细胞核中, 而PAD4则使得EDS1定位于细胞质中, PAD4和SAG101调控EDS1在细胞核与细胞质间的动态平衡, 而细胞核中积累的EDS1可引起免疫自激活, 但SAG101- EDS1-PAD4复合物具体的生化功能尚不清楚(Feys et al., 2001; Chen et al., 2015)。与TNL抗病蛋白不同, CNL抗病蛋白介导的抗病性依赖于NDR1, 其作为病原菌的靶蛋白与效应蛋白相互作用, 激活抗病蛋白, 触发免疫响应。例如, NDR1能够负调控RIN4与RPM1的互作, 监控效应蛋白AvrRpt2, 调控RPM1依赖的免疫响应(Belkhadir et al., 2004; Day et al., 2006)。 植物免疫响应还与激素信号通路密切相关。遗传分析证明, SA相关突变体sid2 (salicylic acid induction deficient 2)和eds1能够部分恢复抗病蛋白HRT (Hypersensitive Response to TCV)和RPP8 (Recognition of Peronospora parasitic 8)激活后造成的免疫激活表型(Chandrashekara et al., 2004; Wiermer et al., 2005), 表明抗病蛋白的激活能够引起SA的积累。SA与细胞质内多聚的NPR1 (Non-expresser of PR Genes 1)结合后, NPR1解聚形成单体进入细胞核, 结合到转录因子的TGA区域, 激活PR1 (Pathogenesis Related Gene 1)以及植物防御反应相关转录因子WRKY的转录(图2); 随后, NPR1同源蛋白NPR3和NPR4作为接头蛋白连接NPR1与Cullin3 E3连接酶, NPR1被降解(Fu et al., 2012; Wu et al., 2012; Paj- erowska-Mukhtar et al., 2013)。此外, 越来越多的研究表明, 多种激素信号均参与调控植物免疫响应, 但具体的调控机制还不清楚(De Bruyne et al., 2014)。
AxtellMJ, StaskawiczBJ (2003). Initiation of RPS2-speci- fied disease resistance in Arabidopsis is coupled to the AvrRpt2-directed elimination of RIN4.Cell 112, 369-377. [本文引用: 1]
[3]
AzevedoC, SadanandomA, KitagawaK, FreialdenhovenA, ShirasuK, Schulze-LefertP (2002). The RAR1 inter- actor SGT1, an essential component of R gene-triggered disease resistance.Science 295, 2073-2076. [本文引用: 2]
[4]
BartelsS, AndersonJC, BesteiroMAG, CarreriA, HirtH, BuchalaA, MétrauxJP, PeckSC, UlmR (2009). MAP kinase phosphatase1 and protein tyrosine phosphatase1 are repressors of salicylic acid synthesis and SNC1- mediated responses in Arabidopsis.Plant Cell 21, 2884-2897. [本文引用: 1]
[5]
BelkhadirY, NimchukZ, HubertDA, MackeyD, DanglJL (2004). Arabidopsis RIN4 negatively regulates disease resistance mediated by RPS2 and RPM1 downstream or independent of the NDR1 signal modulator and is not required for the virulence functions of bacterial type III effectors AvrRpt2 or AvrRpm1.Plant Cell 16, 2822-2835. [本文引用: 1]
[6]
BernouxM, VeT, WilliamsS, WarrenC, HattersD, ValkovE, ZhangXX, EllisJG, KobeB, DoddsPN (2011). Structural and functional analysis of a plant resistance protein TIR domain reveals interfaces for self-association, signaling, and autoregulation.Cell Host Microbe 9, 200-211. [本文引用: 1]
[7]
CésariS, KanzakiH, FujiwaraT, BernouxM, ChalvonV, KawanoY, ShimamotoK, DoddsP, TerauchiR, KrojT (2014). The NB-LRR proteins RGA4 and RGA5 interact functionally and physically to confer disease resistance.EMBO J 33, 1941-1959. [本文引用: 1]
[8]
Chandra-ShekaraAC, NavarreD, KachrooA, KangHG, KlessigD, KachrooP (2004). Signaling requirements and role of salicylic acid in HRT- and rrt-mediated resistance to turnip crinkle virus in Arabidopsis.Plant J 40, 647-659. [本文引用: 1]
ChengYT, GermainH, WiermerM, BiDL, XuF, GarcíaAV, WirthmuellerL, DesprésC, ParkerJE, ZhangYL, LiX (2009). Nuclear pore complex component MOS7/ Nup88 is required for innate immunity and nuclear accu- mulation of defense regulators in Arabidopsis.Plant Cell 21, 2503-2516. [本文引用: 1]
[12]
ChengYT, LiYZ, HuangS, HuangY, DongXN, ZhangYL, LiX (2011). Stability of plant immune-receptor resistance proteins is controlled by SKP1-CULLIN1-F-box (SCF)- mediated protein degradation.Proc Natl Acad Sci USA 108, 14694-14699. [本文引用: 1]
[13]
ChungEH, da CunhaL, WuAJ, GaoZY, CherkisK, AfzalAJ, MackeyD, DanglJL (2011). Specific threonine phos- phorylation of a host target by two unrelated type III effectors activates a host innate immune receptor in plants.Cell Host Microbe 9, 125-136. [本文引用: 1]
[14]
DanglJL, HorvathDM, StaskawiczBJ (2013). Pivoting the plant immune system from dissection to deployment.Sci- ence 341, 746-751. [本文引用: 1]
[15]
DayB, DahlbeckD, StaskawiczBJ (2006). NDR1 interac- tion with RIN4 mediates the differential activation of multiple disease resistance pathways in Arabidopsis.Plant Cell 18, 2782-2791. [本文引用: 1]
[16]
De BruyneL, HöfteM, De VleesschauwerD (2014). Con- necting growth and defense: the emerging roles of brassinosteroids and gibberellins in plant innate immunity.Mol Plant 7, 943-959. [本文引用: 1]
[17]
De La Fuente Bentem van S, VossenJH, de VriesKJ, van WeesS, TamelingWIL, DekkerHL, de KosterCG, HaringMA, TakkenFLW, CornelissenBJC (2005). Heat shock protein 90 and its co-chaperone protein phospha- tase 5 interact with distinct regions of the tomato I-2 disease resistance protein.Plant J 43, 284-298. [本文引用: 1]
DeslandesL, OlivierJ, PeetersN, FengDX, KhounlothamM, BoucherC, SomssichI, GeninS, MarcoY (2003). Physical interaction between RRS1-R, a protein conferring resistance to bacterial wilt, and PopP2, a type III effector targeted to the plant nucleus.Proc Natl Acad Sci USA 100, 8024-8029. [本文引用: 1]
[20]
DoddsPN, LawrenceGJ, CatanzaritiAM, TehT, WangCIA, AyliffeMA, KobeB, EllisJG (2006). Direct protein interaction underlies gene-for-gene specificity and coevo- lution of the flax resistance genes and flax rust avirulence genes.Proc Natl Acad Sci USA 103, 8888-8893. [本文引用: 1]
[21]
DuXR, MiaoM, MaXR, LiuYS, KuhlJC, MartinGB, XiaoFM (2012). Plant programmed cell death caused by an autoactive form of Prf is suppressed by co-expression of the Prf LRR domain.Mol Plant 5, 1058-1067. [本文引用: 1]
[22]
FeiQL, XiaR, MeyersBC (2013). Phased, secondary, small interfering RNAs in posttranscriptional regulatory net- works.Plant Cell 25, 2400-2415. [本文引用: 1]
[23]
FeysBJ, MoisanLJ, NewmanMA, ParkerJE (2001). Direct interaction between the Arabidopsis disease resistance signaling proteins, EDS1 and PAD4.EMBO J 20, 5400-5411. [本文引用: 1]
[24]
FuZQ, YanSP, SalehA, WangW, RubleJ, OkaN, MohanR, SpoelSH, TadaY, ZhengN, DongXN (2012). NPR3 and NPR4 are receptors for the immune signal salicylic acid in plants.Nature 486, 228-232. [本文引用: 1]
[25]
GenotB, LangJ, BerririS, GarmierM, GilardF, PateyronS, HaustraeteK, Van Der StraetenD, HirtH, ColcombetJ (2017). Constitutively active Arabidopsis MAP kinase 3 triggers defense responses involving salicylic acid and SUMM2 resistance protein.Plant Physiol 174, 1238-1249. [本文引用: 1]
[26]
GoritschnigS, ZhangYL, LiX (2007). The ubiquitin path- way is required for innate immunity in Arabidopsis.Plant J 49, 540-551. [本文引用: 1]
[27]
GuYN, ZebellSG, LiangZZ, WangS, KangBH, DongXN (2016). Nuclear pore permeabilization is a convergent signaling event in effector-triggered immunity.Cell 166, 1526-1538. [本文引用: 1]
GutierrezJR, BalmuthAL, NtoukakisV, MucynTS, Gimenez-IbanezS, JonesAME, RathjenJP (2010). Prf immune complexes of tomato are oligomeric and contain multiple Pto-like kinases that diversify effector recognition.Plant J 61, 507-518. [本文引用: 1]
[30]
HaoW, CollierSM, MoffettP, ChaiJ (2013). Structural basis for the interaction between the potato virus X resistance protein (Rx) and its cofactor Ran GTPase- activating protein 2 (RanGAP2).J Biol Chem 288, 35868-35876. [本文引用: 1]
Holt IIIBF, BelkhadirY, DanglJL (2005). Antagonistic control of disease resistance protein stability in the plant immune system.Science 309, 929-932. [本文引用: 1]
[33]
HowdenAJM, HuitemaE (2012). Effector-triggered post- translational modifications and their role in suppression of plant immunity.Front Plant Sci 3, 160. [本文引用: 1]
[34]
HuaJ, GrisafiP, ChengSH, FinkGR (2001). Plant growth homeostasis is controlled by the Arabidopsis BON1 and BAP1 genes.Genes Dev 15, 2263-2272. [本文引用: 1]
[35]
HuangCL, HwangSY, ChiangYC, LinTP (2008). Mole- cular evolution of the Pi-ta gene resistant to rice blast in wild rice(Oryza rufipogon).Genet 179, 1527-1538. [本文引用: 1]
JonesJDG, VanceRE, DanglJL (2016). Intracellular innate immune surveillance devices in plants and animals. Sci- ence 354, aaf6395. [本文引用: 3]
[45]
KimSH, GaoF, BhattacharjeeS, AdiasorJA, NamJC, GassmannW (2010). The Arabidopsis resistance-like gene SNC1 is activated by mutations in SRFR1 and con- tributes to resistance to the bacterial effector AvrRps4.PLoS Pathog 6, e1001172. [本文引用: 1]
[46]
KimSH, KwonSI, SahaD, AnyanwuNC, GassmannW (2009) Resistance to the Pseudomonas syringae Effector HopA1 is governed by the TIR-NBS-LRR protein RPS6 and is enhanced by mutations in SRFR1.Plant Physiol 150, 1723-1732. [本文引用: 1]
[47]
KimSH, QiD, AshfieldT, HelmM, InnesRW (2016). Using decoys to expand the recognition specificity of a plant disease resistance protein.Science 351, 684-687. [本文引用: 1]
[48]
KimSH, SonGH, BhattacharjeeS, KimHJ, NamJC, NguyenPDT, HongJC, GassmannW (2014). The Arabidopsis immune adaptor SRFR1 interacts with TCP transcription factors that redundantly contribute to effector- triggered immunity.Plant J 78, 978-989. [本文引用: 1]
[49]
KongQ, QuN, GaoMH, ZhangZB, DingXJ, YangF, LiYZ, DongOX, ChenS, LiX, ZhangYL (2012). The MEKK1-MKK1/MKK2-MPK4 kinase cascade negatively regulates immunity mediated by a mitogen-activated pro- tein kinase kinase kinase in Arabidopsis.Plant Cell 24, 2225-2236. [本文引用: 1]
[50]
KrasilevaKV, DahlbeckD, StaskawiczBJ (2010). Activa- tion of an Arabidopsis resistance protein is specified by the in planta association of its leucine-rich repeat domain with the cognate oomycete effector.Plant Cell 22, 2444-2458. [本文引用: 1]
[51]
Le RouxC, HuetG, JauneauA, CambordeL, Trém- ousaygueD, KrautA, ZhouBB, LevaillantM, AdachiH, YoshiokaH, RaffaeleS, BerthoméR, CoutéY, ParkerJE, DeslandesL (2015). A receptor pair with an integrated decoy converts pathogen disabling of transcription factors to immunity.Cell 161, 1074-1088. [本文引用: 1]
LiYZ, TessaroMJ, LiX, ZhangYL (2010).b Regulation of the expression of plant Resistance gene SNC1 by a protein with a conserved BAT2 domain.Plant Physiol 153, 1425-1434. [本文引用: 1]
[56]
LiuJ, ElmoreJM, LinZJD, CoakerG (2011). A receptor-like cytoplasmic kinase phosphorylates the host target RIN4, leading to the activation of a plant innate immune receptor.Cell Host Microbe 9, 137-146. [本文引用: 1]
[57]
LiuJY, YangHB, BaoF, AoK, ZhangXY, ZhangYL, YangSH (2015). IBR5 modulates temperature-dependent, R protein CHS3-mediated defense responses in Arabidopsis.PLoS Genet 11, e1005784. [本文引用: 1]
[58]
LiuN, HakeK, WangW, ZhaoT, RomeisT, TangDZ (2017). Calcium-dependent protein kinase 5 associates with the truncated NLR protein TIR-NBS2 to contribute to exo70B1-mediated immunity.Plant Cell 29, 746-759. [本文引用: 1]
[59]
Lukasik-ShreepaathyE, SlootwegE, RichterH, GoverseA, CornelissenBJC, TakkenFLW (2012). Dual regula- tory roles of the extended N terminus for activation of the tomato Mi-1.2 resistance protein.Mol Plant Microbe In- teract 25, 1045-1057. [本文引用: 1]
[60]
MackeyD, BelkhadirY, AlonsoJM, EckerJR, DanglJL (2003). Arabidopsis RIN4 is a target of the type III vir- ulence effector AvrRpt2 and modulates RPS2-mediated resistance.Cell 112, 379-389. [本文引用: 1]
[61]
MackeyD, Holt IIIBF, WiigA, DanglJL (2002). RIN4 interacts with Pseudomonas syringae type III effector molecules and is required for RPM1-mediated resistance in Arabidopsis.Cell 108, 743-754. [本文引用: 1]
MangHG, QianWQ, ZhuY, QianJ, KangHG, KlessigDF, HuaJ (2012). Abscisic acid deficiency antagonizes high- temperature inhibition of disease resistance through en- hancing nuclear accumulation of resistance proteins SNC1 and RPS4 in Arabidopsis. Plant Cell 24, 1271-1284. [本文引用: 1]
[64]
MonaghanJ, ZipfelC (2012). Plant pattern recognition receptor complexes at the plasma membrane.Curr Opin Plant Biol 15, 349-357. [本文引用: 1]
[65]
MucynTS, ClementeA, AndriotisVME, BalmuthAL, OldroydGED, StaskawiczBJ, RathjenJP (2006). The tomato NBARC-LRR protein Prf interacts with Pto kinase in vivo to regulate specific plant immunity.Plant Cell 18, 2792-2806. [本文引用: 1]
[66]
NarusakaM, ShirasuK, NoutoshiY, KuboY, ShiraishiT, IwabuchiM, NarusakaY (2009). RRS1 and RPS4 provide a dual Resistance-gene system against fungal and bacterial pathogens. Plant J 60, 218-226. [本文引用: 2]
[67]
NoutoshiY, ItoT, SekiM, NakashitaH, YoshidaS, MarcoY, ShirasuK, ShinozakiK (2005). A single amino acid insertion in the WRKY domain of the Arabidopsis TIR- NBS-LRR-WRKY-type disease resistance protein SLH1 (sensitive to low humidity 1) causes activation of defense responses and hypersensitive cell death.Plant J 43, 873-888. [本文引用: 1]
[68]
NtoukakisV, BalmuthAL, MucynTS, GutierrezJR, JonesAME, RathjenJP (2013). The tomato Prf complex is a molecular trap for bacterial effectors based on Pto transphosphorylation.PLoS Pathog 9, e1003123. [本文引用: 2]
[69]
Pajerowska-MukhtarKM, EmerineDK, MukhtarMS (2013). Tell me more: roles of NPRs in plant immunity.Trends Plant Sci 18, 402-411. [本文引用: 1]
[70]
QiD, DeYoungBJ, InnesRW (2012). Structure-function analysis of the coiled-coil and leucine-rich repeat domains of the RPS5 disease resistance protein.Plant Physiol 158, 1819-1832. [本文引用: 2]
[71]
QiD, DubiellaU, KimSH, SlossDI, DowenRH, DixonJE, InnesRW (2014). Recognition of the protein kinase AVRPPHB SUSCEPTIBLE1 by the disease resistance protein RESISTANCE TO PSEUDOMONAS SYRINGAE5 is dependent on s-acylation and an exposed loop in AVRPPHB SUSCEPTIBLE1.Plant Physiol 164, 340-351. [本文引用: 1]
[72]
RairdanGJ, MoffettP (2006). Distinct domains in the ARC region of the potato resistance protein Rx mediate LRR binding and inhibition of activation.Plant Cell 18, 2082-2093. [本文引用: 1]
[73]
RouxME, RasmussenMW, PalmaK, LolleS, ReguéÀM, BethkeG, GlazebrookJ, ZhangWP, SieburthL, LarsenMR, MundyJ, PetersenM (2015). The mRNA decay factor PAT1 functions in a pathway including MAP kinase 4 and immune receptor SUMM2.EMBO J 34, 593-608. [本文引用: 1]
SelaH, SpiridonLN, PetrescuAJ, AkermanM, Mandel- GutfreundY, NevoE, LoutreC, KellerB, SchulmanAH, FahimaT (2012). Ancient diversity of splicing motifs and protein surfaces in the wild emmer wheat (Triticum dicoc- coides) LR10 coiled coil (CC) and leucine-rich repeat (LRR) domains.Mol Plant Pathol 13, 276-287.
[78]
SlootwegEJ, SpiridonLN, RoosienJ, ButterbachP, PompR, WesterhofL, WilbersR, BakkerE, BakkerJ, PetrescuAJ, SmantG, GoverseA (2013). Structural determinants at the interface of the ARC2 and Leucine-rich repeat domains control the activation of the plant immune receptors Rx1 and Gpa2.Plant Physiol 162, 1510-1528. [本文引用: 1]
[79]
SohnKH, SegonzacC, RallapalliG, SarrisPF, WooJY, WilliamsSJ, NewmanTE, PaekKH, KobeB, JonesJDG (2014). The nuclear immune receptor RPS4 is required for RRS1SLH1-dependent constitutive defense activation in Arabidopsis thaliana.PLoS Genet 10, e1004655. [本文引用: 1]
[80]
SwiderskiMR, BirkerD, JonesJDG (2009). The TIR domain of TIR-NB-LRR resistance proteins is a signaling domain involved in cell death induction.Mol Plant Microbe Interact 22, 157-165. [本文引用: 1]
[81]
TamelingWIL, BaulcombeDC (2007). Physical association of the NB-LRR resistance protein Rx with a Ran GTPase- activating protein is required for extreme resistance to Potato virus X.Plant Cell 19, 1682-1694. [本文引用: 1]
[82]
TsudaK, KatagiriF (2010). Comparing signaling mecha- nisms engaged in pattern-triggered and effector-triggered immunity.Curr Opin Plant Biol 13, 459-465. [本文引用: 1]
[83]
Van Der BiezenEA, JonesJDG (1998). Plant disease- resistance proteins and the gene-for-gene concept.Trends Biochem Sci 23, 454-456. [本文引用: 1]
[84]
WangGF, JiJB, El-KasmiF, DanglJL, JohalG, Balint- KurtiPJ (2015). Molecular and functional analyses of a maize autoactive NB-LRR protein identify precise struc- tural requirements for activity.PLoS Pathog 11, e1004674. [本文引用: 1]
[85]
WangW, BarnabyJY, TadaY, LiHR, TörM, CaldelariD, LeeDU, FuXD, DongXN (2011). Timing of plant immune responses by a central circadian regulator.Nature 470, 110-114. [本文引用: 1]
[86]
WangYC, ZhangY, WangZ, ZhangXY, YangSH (2013). A missense mutation in CHS1, a TIR-NB protein, induces chilling sensitivity in Arabidopsis. Plant J 75, 553-565. [本文引用: 1]
WillhoftO, KerrR, PatelD, ZhangWJ, Al-JassarC, DaviterT, MillsonSH, ThalassinosK, VaughanCK (2017). The crystal structure of the Sgt1-Skp1 complex: the link between Hsp90 and both SCF E3 ubiquitin ligases and kinetochores.Sci Rep 7, 41626. [本文引用: 1]
[89]
WilliamsSJ, SohnKH, WanL, BernouxM, SarrisPF, SegonzacC, VeT, MaY, SaucetSB, EricssonDJ, CaseyLW, LonhienneT, WinzorDJ, ZhangXX, CoerdtA, ParkerJE, DoddsPN, KobeB, JonesJDG (2014). Structural basis for assembly and function of a hetero- dimeric plant immune receptor.Science 344, 299-303. [本文引用: 1]
[90]
WilliamsSJ, SornarajP, Decourcy-IrelandE, MenzRI, KobeB, EllisJG, DoddsPN, AndersonPA (2011). An autoactive mutant of the M flax rust resistance protein has a preference for binding ATP, whereas wild-type M protein binds ADP.Mol Plant Microbe Interact 24, 897-906. [本文引用: 1]
[91]
WuCH, BelhajK, BozkurtTO, BirkMS, KamounS (2016). Helper NLR proteins NRC2a/b and NRC3 but not NRC1 are required for Pto-mediated cell death and resistance in Nicotiana benthamiana.New Phytol 209, 1344-1352. [本文引用: 1]
[92]
WuY, ZhangD, ChuJY, BoyleP, WangY, BrindleID, De LucaV, DesprésC (2012). The Arabidopsis NPR1 protein is a receptor for the plant defense hormone salicylic acid.Cell Rep 1, 639-647. [本文引用: 1]
[93]
XiaST, ChengYT, HuangS, WinJ, SoardsA, JinnTL, JonesJDG, KamounS, ChenS, ZhangYL, LiX (2013). Regulation of transcription of nucleotide-binding leucine- rich repeat-encoding genes SNC1 and RPP4 via H3K4 trimethylation.Plant Physiol 162, 1694-1705. [本文引用: 1]
[94]
XuF, KaposP, ChengYT, LiM, ZhangYL, LiX (2014). NLR-associating transcription factor bHLH84 and its para- logs function redundantly in plant immunity.PLoS Pathog 10, e1004312. [本文引用: 1]
[95]
XuF, XuSH, WiermerM, ZhangYL, LiX (2012). The cyclin L homolog MOS12 and the MOS4-associated complex are required for the proper splicing of plant resistance genes.Plant J 70, 916-928. [本文引用: 1]
XuSH, ZhangZB, JingBB, GannonP, DingJM, XuF, LiX, ZhangYL (2011). Transportin-SR is required for proper splicing of resistance genes and plant immunity.PLoS Genet 7, e1002159. [本文引用: 1]
[98]
YangHB, ShiYT, LiuJY, GuoL, ZhangXY, YangSH (2010). A mutant CHS3 protein with TIR-NB-LRR-LIM domains modulates growth, cell death and freezing toler ance in a temperature-dependent manner in Arabidopsis.Plant J 63, 283-296. [本文引用: 1]
[99]
YangSH, HuaJ (2004). A haplotype-specific resistance gene regulated by BONZAI1 mediates temperature-depe- ndent growth control in Arabidopsis.Plant Cell 16, 1060-1071. [本文引用: 2]
[100]
YaoCP, WuYY, NieHZ, TangDZ (2012). RPN1a, a 26S proteasome subunit, is required for innate immunity in Arabidopsis.Plant J 71, 1015-1028. [本文引用: 1]
[101]
ZbierzakAM, PorfirovaS, GriebelT, MelzerM, ParkerJE, DörmannP (2013). A TIR-NBS protein encoded by Ara- bidopsis Chilling Sensitive 1 (CHS1) limits chloroplast da- mage and cell death at low temperature.Plant J 75, 539-552. [本文引用: 1]
[102]
ZhangYL, LiX (2005). A putative nucleoporin 96 is required for both basal defense and constitutive resistance res- ponses mediated by suppressor of npr1-1, constitutive 1.Plant Cell 17, 1306-1316. [本文引用: 1]
[103]
ZhangZB, LiuYN, HuangH, GaoMH, WuD, KongQ, ZhangYL (2017). The NLR protein SUMM2 senses the disruption of an immune signaling MAP kinase cascade via CRCK3.EMBO Rep 18, 292-302. [本文引用: 1]
[104]
ZhangZB, WuYL, GaoMH, ZhangJ, KongQ, LiuYN, BaHP, ZhouJM, ZhangYL (2012). Disruption of PAMP- induced MAP Kinase cascade by a Pseudomonas sy- ringae effector activates plant immunity mediated by the NB-LRR protein SUMM2.Cell Host Microbe 11, 253-263. [本文引用: 1]
[105]
ZhouFS, MenkeFLH, YoshiokaK, ModerW, ShiranoY, KlessigDF (2004). High humidity suppresses ssi4-media- ted cell death and disease resistance upstream of MAP kinase activation, H2O2 production and defense gene ex- pression.Plant J 39, 920-932. [本文引用: 1]
[106]
ZhuZH, XuF, ZhangYX, ChengYT, WiermerM, LiX, ZhangYL (2010). Arabidopsis resistance protein SNC1 activates immune responses through association with a transcriptional corepressor.Proc Natl Acad Sci USA 107, 13960-13965. [本文引用: 1]
[107]
ZouBH, YangDL, ShiZY, DongHS, HuaJ (2014). Monou- biquitination of Histone 2B at the disease resistance gene locus regulates its expression and impacts immune res- ponses in Arabidopsis.Plant Physiol 165, 309-318. [本文引用: 1]
1 2011
... 除上述识别模式外, 还存在一些新的抗病蛋白激活模式.植物初级免疫PTI激活后, MAPK (Mitogen Activated Protein Kinase)信号级联通路被激活, 而微生物分泌的效应蛋白能够结合到MAPK通路相关蛋白, 抑制PTI免疫激活.遗传筛选发现, summ1 (suppressor of mkk1mkk2 1)和summ2能够抑制mpk4 (map kinase 4)突变体中免疫激活的表型, SUMM1为蛋白激酶MEKK2, 而SUMM2为抗病蛋白.该结果表明, 抗病蛋白能够监控MEKK1-MKK1/MKK2-MPK4信号级联通路.生化实验证明, MPK4能够磷酸化MEKK2, 但不能与SUMM2直接互作.遗传分析表明, MEKK2位于SUMM2上游, 但MEKK2是如何激活SUMM2的还不清楚(Kong et al., 2012; Zhang et al., 2012).随后的研究表明, MPK4能够磷酸化PAT1 (Arabidopsis Homolog of Yeast PAT1), 抑制SUMM2激活(Roux et al., 2015).最新研究表明, 蛋白激酶CRCK3 (Cal- modulin Binding Receptor like Cytoplasmic Kinase 3)也能被MPK4磷酸化, 共同调控SUMM2依赖的免疫响应(Zhang et al., 2017).无效应蛋白时, MPK4磷酸化底物蛋白, 使得SUMM2处于稳定状态; 而效应蛋白HopAI1与MPK4结合后, 抑制MPK4激酶活性, 使其不能将底物蛋白磷酸化, 导致SUMM2被激活, 触发植物免疫响应.此外, SUMM2还能够监控MPK3与MPK6引起的免疫响应(Genot et al., 2017).除上述激酶以外, 磷酸酶mkp1 (map kinase phosphat- ase1)和ptp1 (protein tyrosine phosphatase1)突变体也表现出SNC1依赖的免疫自激活表型.生化分析结果表明, MKP1能够与MPK3、MPK4和MPK6相互作用, 去磷酸化MPK6, 抑制其激酶活性, 从而拮抗调控抗病蛋白依赖的免疫激活(Schweighofer et al., 2007; Bartels et al., 2009; Anderson et al., 2011).蛋白磷酸酶AP2C1 (Arabidopsis Phosphatase 2C 1)也能够与MPK4和MPK6互作, 通过去磷酸化修饰抑制MPK4和MPK6激酶活性, 调控植物免疫响应.除MAPK依赖的途径外, 钙离子依赖的蛋白激酶CPK1 (Calcium-dependent Protein Kinase 1)、CPK2、CP- K4、CPK5、CPK6和CPK11也能够调控抗病蛋白介导的免疫响应.最新研究表明, CPK5能够与抗病蛋白TN2 (TIR-NBS2)的NBS结构域相互作用, 调控TN2依赖的免疫响应(Liu et al., 2017).综上所述, 抗病蛋白通过不同的识别模式对效应蛋白进行监控, 并且该过程多种多样. ...
1 2003
... 抗病蛋白通过保卫效应蛋白对靶蛋白的攻击激活免疫响应, 抑制病原菌的扩散.保卫模式很好地解释了RPM1 (Resistant to Pseudomonas syringae pv. maculicola 1)、RPS2 (Resistant to Pseudomonas syringae 2)以及RPS5等抗病蛋白的激活方式.目前, 对于RIN4 (RPM1 Interacting Protein 4)介导的抗病蛋白激活模式研究得较为清楚.效应蛋白AvrRpm1 (Avirulence Resistant to Pseudomonas syringae pv. maculicola 1)和AvrB (Avirulence Pseudomonas syringae B)与RIN4蛋白结合后, 蛋白激酶RIPK (RPM1 Induced Protein Kinase)对RIN4蛋白21位苏氨酸、160位丝氨酸和166位苏氨酸进行磷酸化修饰, 导致RPM1被激活, 从而触发植物免疫响应(Mackey et al., 2002; Chung et al., 2011; Liu et al., 2011).除此之外, 效应蛋白诱导的RIN4裂解也能够激活抗病蛋白.效应蛋白AvrRpt2 (Avirulence Resistant to Pseudomonas syringae pv. tomato 2)具有蛋白酶体活性, 能够裂解RIN4, 从而激活RPS2依赖的免疫响应(Axtell and Staskawicz, 2003; Mackey et al., 2003).研究表明, 拟南芥CNL类抗病蛋白RPS5也具有类似的识别方式.效应蛋白AvrPphB (Avirulence Pseudomonas phaseolicola B)具有半胱氨酸蛋白酶活性, 裂解与RPS5相结合的蛋白激酶PBS1 (Protein Kinase AvrPphB Susceptible1).在无效应蛋白时, PBS1激酶与RPS5蛋白CC结构域相互作用, 使RPS5保持失活状态; 当效应蛋白存在时, AvrPphB裂解PBS1导致RPS5蛋白构象改变并被激活, 触 发植物免疫响应(Qi et al., 2012, 2014; Kim et al., 2016). ...
2 2002
... 抗病蛋白的稳定性受蛋白折叠装配及降解的影响.抗病蛋白的装配过程需要分子伴侣的协助(图2B).热激蛋白HSP90 (Heat Shock Protein 90)与分子共伴侣蛋白SGT1 (Suppressor of G2 Allele of Skp1)和RAR1 (Required for Mla12 Resistance 1)共同调控抗病蛋白的正确折叠与稳定.hsp90突变体中抗病蛋白RPM1和RPS5表达量降低.rar1和sgt1突变体中RPS5介导的病原菌抗性减弱.生化实验证明, HSP90的ATPase结构域能够与SGT1蛋白CS (Present in CHP and SGT1 Proteins)结构域互作, 而RAR1蛋白的2个锌结合结构域CHORD-I (Cysteine and Histidine Rich Domain)和CHORD-II分别与HSP90和SGT1蛋白结合, 形成复合体, 共同调控抗病蛋白的成熟(Azevedo et al., 2002; Hubert et al., 2003; Huang et al., 2014).之前的研究证明, 磷酸酶PP5 (Protein Phosphatase 5)能够与HSP90相互作用, 调控抗病蛋白I-2介导的免疫响应(De La Fuente Bentem van, 2005).而最新研究表明, 磷酸酶ibr5 (indole 3 butyric acid response 5)缺失导致抗病蛋白CHS3 (Chilling Sensitive 3)、RPM1和RPS4表达量降低, 同时IBR5能够与HSP90-RAR1-SGT1复合体相互作用, 证明磷酸酶能够作为分子共伴侣蛋白与HSP90复合体一起调节抗病蛋白的稳定性(Liu et al., 2015). ... ... 抗病蛋白的降解受到泛素化蛋白酶体介导的降解途径调控(图2C).研究表明, F-box蛋白cpr1 (constitutive expresser of PR genes 1)功能缺失突变体中抗病蛋白SNC1和RPS2 (Resistance to Pseu- domonas syringae 2)积累, 表明SCFCPR1E3复合体调控抗病蛋白SNC1和RPS2的降解(Cheng et al., 2011).snc1增强子筛选实验证明, E4连接酶MUSE3 (Mutants snc1 Enhancing 3)、TRAR蛋白MUS13和MUSE14能够与SCFCPR1E3复合体互作, 共同调控抗病蛋白SNC1和RPS2的降解(Huang et al., 2014, 2016).MUSE7编码1个功能未知的激酶底物蛋白, 负调控SNC1、RPS2和RPM1蛋白的积累(Johnson et al., 2016).与HSP90复合体作为分子伴侣调控抗病蛋白的装配与成熟不同, HSP90和SGT1也能够与SKP1 (S Phase Kinase Associated Protein 1)-CUL- LIN-F-box-E3泛素复合体相互作用, 调控抗病蛋白的降解(Willhoft et al., 2017).遗传分析表明, hsp90功能获得突变体中抗病蛋白SNC1积累(Huang et al., 2014).sgt1b缺失突变能够抑制rar1突变体中RPS5的积累(Holt et al., 2005).生化实验证明, HSP90和SGT1b能够与SKP1和CULLIN1互作, 促进蛋白降解; RAR1也能够结合COP9 (Constitutive Photo- morpho-genesis Complex 9)蛋白, 抑制SCF复合体的活性, 进而调控抗病蛋白的降解(Azevedo et al., 2002).另外, SGT1a和SGT1b能够与SRFR1 (Suppressor of rps4-RLD)蛋白互作, 负调控抗病蛋白SNC1、RPS2和RPS4的积累(Li et al., 2010a).E1泛素连接酶UBA1 (Ubiquitin Activating 1)与26S蛋白酶体亚基RPN1a (Proteasome Regulatory Subunint S2 1A)、RPN8a (RP Non-ATPase Subunit 8A)和RPT2a (Regulatory Particle AAA-ATPase 2A)参与调控抗病蛋白的降解(Goritschnig et al., 2007; Yao et al., 2012). ...
1 2009
... 除上述识别模式外, 还存在一些新的抗病蛋白激活模式.植物初级免疫PTI激活后, MAPK (Mitogen Activated Protein Kinase)信号级联通路被激活, 而微生物分泌的效应蛋白能够结合到MAPK通路相关蛋白, 抑制PTI免疫激活.遗传筛选发现, summ1 (suppressor of mkk1mkk2 1)和summ2能够抑制mpk4 (map kinase 4)突变体中免疫激活的表型, SUMM1为蛋白激酶MEKK2, 而SUMM2为抗病蛋白.该结果表明, 抗病蛋白能够监控MEKK1-MKK1/MKK2-MPK4信号级联通路.生化实验证明, MPK4能够磷酸化MEKK2, 但不能与SUMM2直接互作.遗传分析表明, MEKK2位于SUMM2上游, 但MEKK2是如何激活SUMM2的还不清楚(Kong et al., 2012; Zhang et al., 2012).随后的研究表明, MPK4能够磷酸化PAT1 (Arabidopsis Homolog of Yeast PAT1), 抑制SUMM2激活(Roux et al., 2015).最新研究表明, 蛋白激酶CRCK3 (Cal- modulin Binding Receptor like Cytoplasmic Kinase 3)也能被MPK4磷酸化, 共同调控SUMM2依赖的免疫响应(Zhang et al., 2017).无效应蛋白时, MPK4磷酸化底物蛋白, 使得SUMM2处于稳定状态; 而效应蛋白HopAI1与MPK4结合后, 抑制MPK4激酶活性, 使其不能将底物蛋白磷酸化, 导致SUMM2被激活, 触发植物免疫响应.此外, SUMM2还能够监控MPK3与MPK6引起的免疫响应(Genot et al., 2017).除上述激酶以外, 磷酸酶mkp1 (map kinase phosphat- ase1)和ptp1 (protein tyrosine phosphatase1)突变体也表现出SNC1依赖的免疫自激活表型.生化分析结果表明, MKP1能够与MPK3、MPK4和MPK6相互作用, 去磷酸化MPK6, 抑制其激酶活性, 从而拮抗调控抗病蛋白依赖的免疫激活(Schweighofer et al., 2007; Bartels et al., 2009; Anderson et al., 2011).蛋白磷酸酶AP2C1 (Arabidopsis Phosphatase 2C 1)也能够与MPK4和MPK6互作, 通过去磷酸化修饰抑制MPK4和MPK6激酶活性, 调控植物免疫响应.除MAPK依赖的途径外, 钙离子依赖的蛋白激酶CPK1 (Calcium-dependent Protein Kinase 1)、CPK2、CP- K4、CPK5、CPK6和CPK11也能够调控抗病蛋白介导的免疫响应.最新研究表明, CPK5能够与抗病蛋白TN2 (TIR-NBS2)的NBS结构域相互作用, 调控TN2依赖的免疫响应(Liu et al., 2017).综上所述, 抗病蛋白通过不同的识别模式对效应蛋白进行监控, 并且该过程多种多样. ...
1 2004
... 目前, 对于抗病基因下游信号通路的研究表明, EDS1 (Enhanced Disease Susceptibility 1)和NDR1 (Non Race Specific Disease Resistance 1)是抗病蛋白下游2个关键的调节因子(图3).TNL抗病蛋白介导的免疫响应依赖于EDS1, 效应蛋白与EDS1结合后抑制PTI, 随后EDS1与抗病蛋白间的互作改变, 激活抗病蛋白依赖的免疫反应(Heidrich et al., 2011).抗病蛋白-EDS1-SRFR1通过相互作用形成复合体, 无效应蛋白时, 细胞质中的EDS1与SRFR1结合后被固定在内质网上, 形成稳定复合体后不发挥作用; 效应蛋白入侵后与EDS1结合, 抗病蛋白-EDS1从SRFR1上释放, 并进一步激活下游防御反应基因的表达, 触发免疫响应(Kim et al., 2009, 2010).另外, SRFR1又能够与TCP (Teosinte Branched1 Cycloidea PCF)相互作用, 拮抗调控RPS4介导的免疫响应(Kim et al., 2014).EDS1也能够与PAD4 (Phytoalexin Deficient 4)和SAG101 (Senescence Associated Gene 101)相互作用, 影响EDS1的定位, 共同调控植物免疫响应.SAG101可以将EDS1定位于细胞核中, 而PAD4则使得EDS1定位于细胞质中, PAD4和SAG101调控EDS1在细胞核与细胞质间的动态平衡, 而细胞核中积累的EDS1可引起免疫自激活, 但SAG101- EDS1-PAD4复合物具体的生化功能尚不清楚(Feys et al., 2001; Chen et al., 2015).与TNL抗病蛋白不同, CNL抗病蛋白介导的抗病性依赖于NDR1, 其作为病原菌的靶蛋白与效应蛋白相互作用, 激活抗病蛋白, 触发免疫响应.例如, NDR1能够负调控RIN4与RPM1的互作, 监控效应蛋白AvrRpt2, 调控RPM1依赖的免疫响应(Belkhadir et al., 2004; Day et al., 2006). ...
1 2011
... NB-LRR抗病蛋白作为一类免疫监测因子, 在无效应蛋白激发时, 通过不同结构域间的互作保持抗病蛋白处于失活状态.不同结构域间的互作方式主要有4种.(1) 某些抗病蛋白LRR结构域能够与效应因子直接互作, 该类蛋白NBS结构域与LRR中效应因子结合区域互作, 抑制LRR结构域的激活, 如L6 (L locus 6)抗病蛋白(Bernoux et al., 2011).(2) 抗病蛋白N端区域与NBS结构域共同介导免疫激活, 该类蛋白的LRR结构域与ARC2区域互作, 抑制N端激活域或NBS结构域的功能, 使抗病蛋白处于稳定状态, 如RPS5 (Resistant to Pseudomonas syringae 5)和RPP1 (Resistant to Pseudomonas parasitica 1)等(Rairdan and Moffett, 2006; Qi et al., 2012; Sloot- weg and Goverse, 2013).(3) 抗病蛋白的CC结构域作为其功能结构域, 其NBS结构域与CC结构域结合, 抑制抗病蛋白自激活, 如Rp1 (Resistant to Puccinia sorghi 1)抗病蛋白(Wang et al., 2015).(4) 许多抗病蛋白N端与其它蛋白互作, 共同对效应因子进行监控, LRR结构域与N端区域互作, 使抗病蛋白保持失活状态, 如Prf (Du et al., 2012; Lukasik-shreepaathy et al., 2012). ...
1 2014
... 许多抗病蛋白需要与其它抗病蛋白形成复合体, 共同调控植物免疫响应.例如, 拟南芥TNL类抗病蛋白RPS4/RRS1调控效应蛋白AvrRps4 (Pst. DC3000 Avr Rps4)和PopP2 (R. Solanacearum-PopP2)诱导的免疫反应.无效应蛋白时, RPS4的TIR结构域与RR- S1的TIR结构域相互作用, 使抗病蛋白RPS4/RRS1处于失活状态; 当效应蛋白存在时, RRS1羧基端包含的与转录因子WRKY相似的DNA结合结构域能够作为诱饵与效应蛋白PopP2结合.AvrRps4与RRS1的WRKY结构域结合后启动抗病反应; 而PopP2具有乙酰基转移酶活性, 能够将RRS1 WRKYGQK结构中第1 221位赖氨酸乙酰化, 导致其失去DNA结合能力, 激活RPS4/RRS1复合体, 触发植物免疫响应(Narusaka et al., 2009; Sohn et al., 2014; Sarris et al., 2015; Le Roux, 2015).类似地, 水稻CNL类抗病蛋白RGA4 (R Gene Analog 4)和RGA5 (R Gene Analog 5)同样以复合体的形式发挥作用.RGA5通过CC结构域与RGA4蛋白互作, 在无效应蛋白刺激时抑制后者的激活(Césari et al., 2014).此外, Prf/Pto复合体介导的抗病反应也需要抗病蛋白NRC2a/b和NRC3的参与(Jones et al., 2016; Wu et al., 2016).但不同抗病蛋白间是如何互作的还有待进一步研究. ...
1 2004
... 植物免疫响应还与激素信号通路密切相关.遗传分析证明, SA相关突变体sid2 (salicylic acid induction deficient 2)和eds1能够部分恢复抗病蛋白HRT (Hypersensitive Response to TCV)和RPP8 (Recognition of Peronospora parasitic 8)激活后造成的免疫激活表型(Chandrashekara et al., 2004; Wiermer et al., 2005), 表明抗病蛋白的激活能够引起SA的积累.SA与细胞质内多聚的NPR1 (Non-expresser of PR Genes 1)结合后, NPR1解聚形成单体进入细胞核, 结合到转录因子的TGA区域, 激活PR1 (Pathogenesis Related Gene 1)以及植物防御反应相关转录因子WRKY的转录(图2); 随后, NPR1同源蛋白NPR3和NPR4作为接头蛋白连接NPR1与Cullin3 E3连接酶, NPR1被降解(Fu et al., 2012; Wu et al., 2012; Paj- erowska-Mukhtar et al., 2013).此外, 越来越多的研究表明, 多种激素信号均参与调控植物免疫响应, 但具体的调控机制还不清楚(De Bruyne et al., 2014). ...
1 2013
... 在植物免疫响应过程中, 抗病蛋白的激活导致下游免疫信号被激活, 从而调控植物对病原菌的抗性.研究表明许多抗病蛋白能够进入细胞核激活免疫响应(图3).对SNC1蛋白的研究表明, 当其进入细胞核后, SNC1的TIR结构域与转录抑制因子TPR1 (Topless Related 1)结合, 抑制免疫负调控因子DND1 (Defense No Death 1)和DND2的转录(Zhu et al., 2010); SNC1还能够与bHLH84 (Helix Loop Helix 84)转录因子相互作用, 激活抗病相关基因的转录(Xu et al., 2014).通过对snc1抑制子的筛选发现, MOS3和MOS7能够调控SNC1在细胞核中的积累.MOS3是一个核孔蛋白, MOS7为果蝇与人类(Homo sapiens)核孔蛋白Nup88的同源蛋白.在mos3和mos7缺失突变体中, 定位于细胞核的SNC1减少, 不能激活转录因子的表达, 从而抑制SNC1依赖的免疫响应.该研究结果表明, 核孔复合体组分MOS3和MOS7能够调控抗病蛋白的细胞核定位, 影响SNC1介导的下游转录信号激活(Zhang and Li, 2005; Cheng et al., 2009), 但其具体调控机制还不清楚.而最新研究表明, 在抗病蛋白RPS2激活过程中, 与核孔复合体结合的跨膜核孔蛋白CPR5 (Constitutive Expresser of PR Genes 5)构象改变, 降低其与CKIs (Cyclin-dep- endent Kinase Inhibitors)的亲和性, 触发Rb (Retinoblastoma)和E2F介导的细胞周期途径, 调控抗病基因的表达, 引发细胞程序性死亡, 从而激活ETI免疫响应(Gu et al., 2016).除此以外, 某些含有DNA结合结构域的抗病蛋白进入细胞核后, 能够直接与转录因子结合, 激活抗病相关基因的转录.例如, 抗病蛋白MLA10入核后, 其N端结构域与WRKY1/2转录因子相互作用, 解除其对MYB6 (MYB Domain Protein 6)的抑制, 启动植物抗病反应(Chang et al., 2013); 而Pb1 (Panicle blast 1)与WRKY45转录因子结合后, 抑制WRKY45被蛋白酶体降解, 激活SA依赖的抗病信号通路(Inoue et al., 2013). ...
1 2015
... 目前, 对于抗病基因下游信号通路的研究表明, EDS1 (Enhanced Disease Susceptibility 1)和NDR1 (Non Race Specific Disease Resistance 1)是抗病蛋白下游2个关键的调节因子(图3).TNL抗病蛋白介导的免疫响应依赖于EDS1, 效应蛋白与EDS1结合后抑制PTI, 随后EDS1与抗病蛋白间的互作改变, 激活抗病蛋白依赖的免疫反应(Heidrich et al., 2011).抗病蛋白-EDS1-SRFR1通过相互作用形成复合体, 无效应蛋白时, 细胞质中的EDS1与SRFR1结合后被固定在内质网上, 形成稳定复合体后不发挥作用; 效应蛋白入侵后与EDS1结合, 抗病蛋白-EDS1从SRFR1上释放, 并进一步激活下游防御反应基因的表达, 触发免疫响应(Kim et al., 2009, 2010).另外, SRFR1又能够与TCP (Teosinte Branched1 Cycloidea PCF)相互作用, 拮抗调控RPS4介导的免疫响应(Kim et al., 2014).EDS1也能够与PAD4 (Phytoalexin Deficient 4)和SAG101 (Senescence Associated Gene 101)相互作用, 影响EDS1的定位, 共同调控植物免疫响应.SAG101可以将EDS1定位于细胞核中, 而PAD4则使得EDS1定位于细胞质中, PAD4和SAG101调控EDS1在细胞核与细胞质间的动态平衡, 而细胞核中积累的EDS1可引起免疫自激活, 但SAG101- EDS1-PAD4复合物具体的生化功能尚不清楚(Feys et al., 2001; Chen et al., 2015).与TNL抗病蛋白不同, CNL抗病蛋白介导的抗病性依赖于NDR1, 其作为病原菌的靶蛋白与效应蛋白相互作用, 激活抗病蛋白, 触发免疫响应.例如, NDR1能够负调控RIN4与RPM1的互作, 监控效应蛋白AvrRpt2, 调控RPM1依赖的免疫响应(Belkhadir et al., 2004; Day et al., 2006). ...
1 2009
... 在植物免疫响应过程中, 抗病蛋白的激活导致下游免疫信号被激活, 从而调控植物对病原菌的抗性.研究表明许多抗病蛋白能够进入细胞核激活免疫响应(图3).对SNC1蛋白的研究表明, 当其进入细胞核后, SNC1的TIR结构域与转录抑制因子TPR1 (Topless Related 1)结合, 抑制免疫负调控因子DND1 (Defense No Death 1)和DND2的转录(Zhu et al., 2010); SNC1还能够与bHLH84 (Helix Loop Helix 84)转录因子相互作用, 激活抗病相关基因的转录(Xu et al., 2014).通过对snc1抑制子的筛选发现, MOS3和MOS7能够调控SNC1在细胞核中的积累.MOS3是一个核孔蛋白, MOS7为果蝇与人类(Homo sapiens)核孔蛋白Nup88的同源蛋白.在mos3和mos7缺失突变体中, 定位于细胞核的SNC1减少, 不能激活转录因子的表达, 从而抑制SNC1依赖的免疫响应.该研究结果表明, 核孔复合体组分MOS3和MOS7能够调控抗病蛋白的细胞核定位, 影响SNC1介导的下游转录信号激活(Zhang and Li, 2005; Cheng et al., 2009), 但其具体调控机制还不清楚.而最新研究表明, 在抗病蛋白RPS2激活过程中, 与核孔复合体结合的跨膜核孔蛋白CPR5 (Constitutive Expresser of PR Genes 5)构象改变, 降低其与CKIs (Cyclin-dep- endent Kinase Inhibitors)的亲和性, 触发Rb (Retinoblastoma)和E2F介导的细胞周期途径, 调控抗病基因的表达, 引发细胞程序性死亡, 从而激活ETI免疫响应(Gu et al., 2016).除此以外, 某些含有DNA结合结构域的抗病蛋白进入细胞核后, 能够直接与转录因子结合, 激活抗病相关基因的转录.例如, 抗病蛋白MLA10入核后, 其N端结构域与WRKY1/2转录因子相互作用, 解除其对MYB6 (MYB Domain Protein 6)的抑制, 启动植物抗病反应(Chang et al., 2013); 而Pb1 (Panicle blast 1)与WRKY45转录因子结合后, 抑制WRKY45被蛋白酶体降解, 激活SA依赖的抗病信号通路(Inoue et al., 2013). ...
1 2011
... 抗病蛋白的降解受到泛素化蛋白酶体介导的降解途径调控(图2C).研究表明, F-box蛋白cpr1 (constitutive expresser of PR genes 1)功能缺失突变体中抗病蛋白SNC1和RPS2 (Resistance to Pseu- domonas syringae 2)积累, 表明SCFCPR1E3复合体调控抗病蛋白SNC1和RPS2的降解(Cheng et al., 2011).snc1增强子筛选实验证明, E4连接酶MUSE3 (Mutants snc1 Enhancing 3)、TRAR蛋白MUS13和MUSE14能够与SCFCPR1E3复合体互作, 共同调控抗病蛋白SNC1和RPS2的降解(Huang et al., 2014, 2016).MUSE7编码1个功能未知的激酶底物蛋白, 负调控SNC1、RPS2和RPM1蛋白的积累(Johnson et al., 2016).与HSP90复合体作为分子伴侣调控抗病蛋白的装配与成熟不同, HSP90和SGT1也能够与SKP1 (S Phase Kinase Associated Protein 1)-CUL- LIN-F-box-E3泛素复合体相互作用, 调控抗病蛋白的降解(Willhoft et al., 2017).遗传分析表明, hsp90功能获得突变体中抗病蛋白SNC1积累(Huang et al., 2014).sgt1b缺失突变能够抑制rar1突变体中RPS5的积累(Holt et al., 2005).生化实验证明, HSP90和SGT1b能够与SKP1和CULLIN1互作, 促进蛋白降解; RAR1也能够结合COP9 (Constitutive Photo- morpho-genesis Complex 9)蛋白, 抑制SCF复合体的活性, 进而调控抗病蛋白的降解(Azevedo et al., 2002).另外, SGT1a和SGT1b能够与SRFR1 (Suppressor of rps4-RLD)蛋白互作, 负调控抗病蛋白SNC1、RPS2和RPS4的积累(Li et al., 2010a).E1泛素连接酶UBA1 (Ubiquitin Activating 1)与26S蛋白酶体亚基RPN1a (Proteasome Regulatory Subunint S2 1A)、RPN8a (RP Non-ATPase Subunit 8A)和RPT2a (Regulatory Particle AAA-ATPase 2A)参与调控抗病蛋白的降解(Goritschnig et al., 2007; Yao et al., 2012). ...
1 2011
... 抗病蛋白通过保卫效应蛋白对靶蛋白的攻击激活免疫响应, 抑制病原菌的扩散.保卫模式很好地解释了RPM1 (Resistant to Pseudomonas syringae pv. maculicola 1)、RPS2 (Resistant to Pseudomonas syringae 2)以及RPS5等抗病蛋白的激活方式.目前, 对于RIN4 (RPM1 Interacting Protein 4)介导的抗病蛋白激活模式研究得较为清楚.效应蛋白AvrRpm1 (Avirulence Resistant to Pseudomonas syringae pv. maculicola 1)和AvrB (Avirulence Pseudomonas syringae B)与RIN4蛋白结合后, 蛋白激酶RIPK (RPM1 Induced Protein Kinase)对RIN4蛋白21位苏氨酸、160位丝氨酸和166位苏氨酸进行磷酸化修饰, 导致RPM1被激活, 从而触发植物免疫响应(Mackey et al., 2002; Chung et al., 2011; Liu et al., 2011).除此之外, 效应蛋白诱导的RIN4裂解也能够激活抗病蛋白.效应蛋白AvrRpt2 (Avirulence Resistant to Pseudomonas syringae pv. tomato 2)具有蛋白酶体活性, 能够裂解RIN4, 从而激活RPS2依赖的免疫响应(Axtell and Staskawicz, 2003; Mackey et al., 2003).研究表明, 拟南芥CNL类抗病蛋白RPS5也具有类似的识别方式.效应蛋白AvrPphB (Avirulence Pseudomonas phaseolicola B)具有半胱氨酸蛋白酶活性, 裂解与RPS5相结合的蛋白激酶PBS1 (Protein Kinase AvrPphB Susceptible1).在无效应蛋白时, PBS1激酶与RPS5蛋白CC结构域相互作用, 使RPS5保持失活状态; 当效应蛋白存在时, AvrPphB裂解PBS1导致RPS5蛋白构象改变并被激活, 触 发植物免疫响应(Qi et al., 2012, 2014; Kim et al., 2016). ...
1 2013
... 植物在生长发育过程中为了应对复杂的环境变化, 进化形成了2个不同层次的防御反应: 初级免疫反应和次级免疫反应.宿主细胞膜上的模式识别受体(pattern recognition receptors, PRR)识别病原微生物相关分子模式(pathogen associated molecular patterns, PAMP), 激活PAMP触发的免疫反应(PAMP triggered immunity, PTI) (Monaghan and Zipfel, 2012; Schwessinger and Ronald, 2012; Dangl et al., 2013), 即初级免疫反应.为了应对植物初级免疫反应, 微生物分泌出一些毒性因子(即效应因子)注入到植物细胞内, 抑制PTI, 导致植物感病.在应对效应因子的入侵方面, 植物进化出能够特异识别效应因子的抗性蛋白(resistance protein, R), 激活效应因子触发的免疫反应(effectors triggered immu- nity, ETI), 即为次级免疫反应(Tsuda and Katagiri, 2010; Howden and Huitema, 2012).ETI比PTI更为强烈和迅速, 导致局部的程序性细胞死亡(program- med cell death, PCD), 引起植物超敏反应(hyper- sensitive response, HR), 更为有效地遏制病原菌的进一步扩散(Jones and Dangl, 2006). ...
1 2006
... 目前, 对于抗病基因下游信号通路的研究表明, EDS1 (Enhanced Disease Susceptibility 1)和NDR1 (Non Race Specific Disease Resistance 1)是抗病蛋白下游2个关键的调节因子(图3).TNL抗病蛋白介导的免疫响应依赖于EDS1, 效应蛋白与EDS1结合后抑制PTI, 随后EDS1与抗病蛋白间的互作改变, 激活抗病蛋白依赖的免疫反应(Heidrich et al., 2011).抗病蛋白-EDS1-SRFR1通过相互作用形成复合体, 无效应蛋白时, 细胞质中的EDS1与SRFR1结合后被固定在内质网上, 形成稳定复合体后不发挥作用; 效应蛋白入侵后与EDS1结合, 抗病蛋白-EDS1从SRFR1上释放, 并进一步激活下游防御反应基因的表达, 触发免疫响应(Kim et al., 2009, 2010).另外, SRFR1又能够与TCP (Teosinte Branched1 Cycloidea PCF)相互作用, 拮抗调控RPS4介导的免疫响应(Kim et al., 2014).EDS1也能够与PAD4 (Phytoalexin Deficient 4)和SAG101 (Senescence Associated Gene 101)相互作用, 影响EDS1的定位, 共同调控植物免疫响应.SAG101可以将EDS1定位于细胞核中, 而PAD4则使得EDS1定位于细胞质中, PAD4和SAG101调控EDS1在细胞核与细胞质间的动态平衡, 而细胞核中积累的EDS1可引起免疫自激活, 但SAG101- EDS1-PAD4复合物具体的生化功能尚不清楚(Feys et al., 2001; Chen et al., 2015).与TNL抗病蛋白不同, CNL抗病蛋白介导的抗病性依赖于NDR1, 其作为病原菌的靶蛋白与效应蛋白相互作用, 激活抗病蛋白, 触发免疫响应.例如, NDR1能够负调控RIN4与RPM1的互作, 监控效应蛋白AvrRpt2, 调控RPM1依赖的免疫响应(Belkhadir et al., 2004; Day et al., 2006). ...
1 2014
... 植物免疫响应还与激素信号通路密切相关.遗传分析证明, SA相关突变体sid2 (salicylic acid induction deficient 2)和eds1能够部分恢复抗病蛋白HRT (Hypersensitive Response to TCV)和RPP8 (Recognition of Peronospora parasitic 8)激活后造成的免疫激活表型(Chandrashekara et al., 2004; Wiermer et al., 2005), 表明抗病蛋白的激活能够引起SA的积累.SA与细胞质内多聚的NPR1 (Non-expresser of PR Genes 1)结合后, NPR1解聚形成单体进入细胞核, 结合到转录因子的TGA区域, 激活PR1 (Pathogenesis Related Gene 1)以及植物防御反应相关转录因子WRKY的转录(图2); 随后, NPR1同源蛋白NPR3和NPR4作为接头蛋白连接NPR1与Cullin3 E3连接酶, NPR1被降解(Fu et al., 2012; Wu et al., 2012; Paj- erowska-Mukhtar et al., 2013).此外, 越来越多的研究表明, 多种激素信号均参与调控植物免疫响应, 但具体的调控机制还不清楚(De Bruyne et al., 2014). ...
1 2005
... 抗病蛋白的稳定性受蛋白折叠装配及降解的影响.抗病蛋白的装配过程需要分子伴侣的协助(图2B).热激蛋白HSP90 (Heat Shock Protein 90)与分子共伴侣蛋白SGT1 (Suppressor of G2 Allele of Skp1)和RAR1 (Required for Mla12 Resistance 1)共同调控抗病蛋白的正确折叠与稳定.hsp90突变体中抗病蛋白RPM1和RPS5表达量降低.rar1和sgt1突变体中RPS5介导的病原菌抗性减弱.生化实验证明, HSP90的ATPase结构域能够与SGT1蛋白CS (Present in CHP and SGT1 Proteins)结构域互作, 而RAR1蛋白的2个锌结合结构域CHORD-I (Cysteine and Histidine Rich Domain)和CHORD-II分别与HSP90和SGT1蛋白结合, 形成复合体, 共同调控抗病蛋白的成熟(Azevedo et al., 2002; Hubert et al., 2003; Huang et al., 2014).之前的研究证明, 磷酸酶PP5 (Protein Phosphatase 5)能够与HSP90相互作用, 调控抗病蛋白I-2介导的免疫响应(De La Fuente Bentem van, 2005).而最新研究表明, 磷酸酶ibr5 (indole 3 butyric acid response 5)缺失导致抗病蛋白CHS3 (Chilling Sensitive 3)、RPM1和RPS4表达量降低, 同时IBR5能够与HSP90-RAR1-SGT1复合体相互作用, 证明磷酸酶能够作为分子共伴侣蛋白与HSP90复合体一起调节抗病蛋白的稳定性(Liu et al., 2015). ...
... 抗病蛋白与病原菌的靶蛋白类似物相互作用, 效应蛋白将该靶蛋白类似物误认为其真正的靶蛋白, 对其进行识别修饰, 导致免疫响应激活.例如, Prf蛋白的激活方式为2个Prf蛋白通过N端结构域相互作用形成同源二聚体, 并与Pto (Pseudomonas syringae pv. tomato)蛋白激酶结合形成复合体(Mucyn et al., 2006; Gutierrez et al., 2010).无效应蛋白AvrPto时, Pto激酶上的β-1与P+1环抑制抗病蛋白激活; 当效应蛋白AvrPto存在时, Prf/Pto复合体中的1个Pto激酶作为诱饵蛋白与AvrPto结合, 使Pto激酶结构改变, 导致Prf蛋白NBS结构域对Pto的抑制作用解除, 随后另一个Pto激酶磷酸化与效应蛋白结合的Pto激酶, 抗病蛋白Prf被激活, 引起HR反应(Narusaka et al., 2009; Ntoukakis et al., 2013). ...
1 2013
... TNL类抗病蛋白N端包含TIR结构域, 其与果蝇(Droso- phila) Toll蛋白或白介素1受体N端序列类似, 在动植物中高度保守.晶体结构分析显示, TIR结构域通常由核心的5个β-片层和外围的5个α-螺旋构成.TIR结构域能够识别效应蛋白.例如, 烟草(Nicotiana tabacum) N蛋白的TIR结构域能够特异地识别花叶病毒分泌的效应蛋白p50 (Swiderski et al., 2009).此外, 该结构域也能够介导蛋白之间的相互作用.例如, RRS1 (Resistant to Ralstonia solanacearum 1)和RPS4 (Resistant to Pseudomonas syringae 4)蛋白通过TIR结构域中保守的丝氨酸-组氨酸(ser-his, SH)基序互作, 形成结合界面, 共同介导植物对病原菌的抗性(Williams et al., 2014).CNL类抗病蛋白N端包含CC结构域, 即不规则卷曲螺旋结构.通过对CC结构域蛋白结构进行解析, 发现该结构域由7个氨基酸重复序列组成, 包含2个或多个α-螺旋, 构成一个超螺旋结构.CC结构域缺乏保守性, 其中唯一的保守基序为EDVID.但并非所有的CNL蛋白CC结构域都存在该基序.CC结构域在蛋白互作中发挥重要功能.例如, CNL抗性蛋白Rx的CC结构域能够与细胞内辅因子RanGAP2直接互作, 共同监控马铃薯(Solanum tuberosum) X病毒.晶体结构揭示, 该蛋白CC结构域具有保守的EDVID基序, 其第90位色氨酸能够与Ran- GAP2的第89位丙氨酸通过疏水作用相结合.而Rx蛋白CC结构域与RanGAP2相结合的2个α-螺旋为调控Rx蛋白入核的区域, RanGAP2通过与该区域结合阻止Rx进入细胞核, 增强植物抗病能力(Tameling and Baulcombe, 2007; Hao et al., 2013).此外, 通过解析MLA10 (Mildew Resistance Locus A10)抗病蛋白CC结构域的晶体结构, 发现该蛋白CC结构域由2个长α-螺旋单体组成, 2条链相距较远, 且内侧多为疏水性氨基酸, 2个MLA10蛋白通过CC结构域形成同源二聚体, 维持整个蛋白的稳定性(Maekawa et al., 2011). ...
1 2011
... 目前, 对于抗病基因下游信号通路的研究表明, EDS1 (Enhanced Disease Susceptibility 1)和NDR1 (Non Race Specific Disease Resistance 1)是抗病蛋白下游2个关键的调节因子(图3).TNL抗病蛋白介导的免疫响应依赖于EDS1, 效应蛋白与EDS1结合后抑制PTI, 随后EDS1与抗病蛋白间的互作改变, 激活抗病蛋白依赖的免疫反应(Heidrich et al., 2011).抗病蛋白-EDS1-SRFR1通过相互作用形成复合体, 无效应蛋白时, 细胞质中的EDS1与SRFR1结合后被固定在内质网上, 形成稳定复合体后不发挥作用; 效应蛋白入侵后与EDS1结合, 抗病蛋白-EDS1从SRFR1上释放, 并进一步激活下游防御反应基因的表达, 触发免疫响应(Kim et al., 2009, 2010).另外, SRFR1又能够与TCP (Teosinte Branched1 Cycloidea PCF)相互作用, 拮抗调控RPS4介导的免疫响应(Kim et al., 2014).EDS1也能够与PAD4 (Phytoalexin Deficient 4)和SAG101 (Senescence Associated Gene 101)相互作用, 影响EDS1的定位, 共同调控植物免疫响应.SAG101可以将EDS1定位于细胞核中, 而PAD4则使得EDS1定位于细胞质中, PAD4和SAG101调控EDS1在细胞核与细胞质间的动态平衡, 而细胞核中积累的EDS1可引起免疫自激活, 但SAG101- EDS1-PAD4复合物具体的生化功能尚不清楚(Feys et al., 2001; Chen et al., 2015).与TNL抗病蛋白不同, CNL抗病蛋白介导的抗病性依赖于NDR1, 其作为病原菌的靶蛋白与效应蛋白相互作用, 激活抗病蛋白, 触发免疫响应.例如, NDR1能够负调控RIN4与RPM1的互作, 监控效应蛋白AvrRpt2, 调控RPM1依赖的免疫响应(Belkhadir et al., 2004; Day et al., 2006). ...
1 2005
... 抗病蛋白的降解受到泛素化蛋白酶体介导的降解途径调控(图2C).研究表明, F-box蛋白cpr1 (constitutive expresser of PR genes 1)功能缺失突变体中抗病蛋白SNC1和RPS2 (Resistance to Pseu- domonas syringae 2)积累, 表明SCFCPR1E3复合体调控抗病蛋白SNC1和RPS2的降解(Cheng et al., 2011).snc1增强子筛选实验证明, E4连接酶MUSE3 (Mutants snc1 Enhancing 3)、TRAR蛋白MUS13和MUSE14能够与SCFCPR1E3复合体互作, 共同调控抗病蛋白SNC1和RPS2的降解(Huang et al., 2014, 2016).MUSE7编码1个功能未知的激酶底物蛋白, 负调控SNC1、RPS2和RPM1蛋白的积累(Johnson et al., 2016).与HSP90复合体作为分子伴侣调控抗病蛋白的装配与成熟不同, HSP90和SGT1也能够与SKP1 (S Phase Kinase Associated Protein 1)-CUL- LIN-F-box-E3泛素复合体相互作用, 调控抗病蛋白的降解(Willhoft et al., 2017).遗传分析表明, hsp90功能获得突变体中抗病蛋白SNC1积累(Huang et al., 2014).sgt1b缺失突变能够抑制rar1突变体中RPS5的积累(Holt et al., 2005).生化实验证明, HSP90和SGT1b能够与SKP1和CULLIN1互作, 促进蛋白降解; RAR1也能够结合COP9 (Constitutive Photo- morpho-genesis Complex 9)蛋白, 抑制SCF复合体的活性, 进而调控抗病蛋白的降解(Azevedo et al., 2002).另外, SGT1a和SGT1b能够与SRFR1 (Suppressor of rps4-RLD)蛋白互作, 负调控抗病蛋白SNC1、RPS2和RPS4的积累(Li et al., 2010a).E1泛素连接酶UBA1 (Ubiquitin Activating 1)与26S蛋白酶体亚基RPN1a (Proteasome Regulatory Subunint S2 1A)、RPN8a (RP Non-ATPase Subunit 8A)和RPT2a (Regulatory Particle AAA-ATPase 2A)参与调控抗病蛋白的降解(Goritschnig et al., 2007; Yao et al., 2012). ...
1 2012
... 植物在生长发育过程中为了应对复杂的环境变化, 进化形成了2个不同层次的防御反应: 初级免疫反应和次级免疫反应.宿主细胞膜上的模式识别受体(pattern recognition receptors, PRR)识别病原微生物相关分子模式(pathogen associated molecular patterns, PAMP), 激活PAMP触发的免疫反应(PAMP triggered immunity, PTI) (Monaghan and Zipfel, 2012; Schwessinger and Ronald, 2012; Dangl et al., 2013), 即初级免疫反应.为了应对植物初级免疫反应, 微生物分泌出一些毒性因子(即效应因子)注入到植物细胞内, 抑制PTI, 导致植物感病.在应对效应因子的入侵方面, 植物进化出能够特异识别效应因子的抗性蛋白(resistance protein, R), 激活效应因子触发的免疫反应(effectors triggered immu- nity, ETI), 即为次级免疫反应(Tsuda and Katagiri, 2010; Howden and Huitema, 2012).ETI比PTI更为强烈和迅速, 导致局部的程序性细胞死亡(program- med cell death, PCD), 引起植物超敏反应(hyper- sensitive response, HR), 更为有效地遏制病原菌的进一步扩散(Jones and Dangl, 2006). ...
1 2001
... 除内在调控方式外, 温度、湿度和昼夜节律等环境因素也能够影响抗病蛋白的稳定性.遗传筛选实验表明, 许多免疫自激活突变体均表现出温度依赖的表型, 如chs1/2/3、snc1-1、bon1-1和mkp1等.chs1/2/3在16 °C下表现为生长缺陷, 而22°C下生长正常; snc1-1和bon1-1等突变体在22°C下植株矮小、叶片卷曲, 而这种缺陷表型可以被高温恢复或部分恢复(Hua et al., 2001; Yang and Hua, 2004; Yang et al., 2010; Wang et al., 2013; Zbierzak et al., 2013).高温影响抗病蛋白与病原菌间的相互作用, 并且高温能够抑制抗病蛋白从稳定状态向激活状态转变.此外, 高温下植物体内水杨酸(salicylic acid, SA)和脱落酸(absci- sic acid, ABA)等激素水平下降, 增强了植物的抗菌性(Mang et al., 2012).环境湿度和昼夜周期节律变化也能够调控抗病蛋白的稳定性.例如, 高湿度能够抑制抗病蛋白SSI (Suppressor of Salicylic Acid Insensitive)与SLH1 (an allele of RRS1)依赖的免疫激活(Zhou et al., 2004; Noutoshi et al., 2005); 昼夜节律可以影响抗病基因RPP4 (Resistant to Peronospora parasitica 4)的转录(Wang et al., 2011). ...
... 许多抗病蛋白需要与其它抗病蛋白形成复合体, 共同调控植物免疫响应.例如, 拟南芥TNL类抗病蛋白RPS4/RRS1调控效应蛋白AvrRps4 (Pst. DC3000 Avr Rps4)和PopP2 (R. Solanacearum-PopP2)诱导的免疫反应.无效应蛋白时, RPS4的TIR结构域与RR- S1的TIR结构域相互作用, 使抗病蛋白RPS4/RRS1处于失活状态; 当效应蛋白存在时, RRS1羧基端包含的与转录因子WRKY相似的DNA结合结构域能够作为诱饵与效应蛋白PopP2结合.AvrRps4与RRS1的WRKY结构域结合后启动抗病反应; 而PopP2具有乙酰基转移酶活性, 能够将RRS1 WRKYGQK结构中第1 221位赖氨酸乙酰化, 导致其失去DNA结合能力, 激活RPS4/RRS1复合体, 触发植物免疫响应(Narusaka et al., 2009; Sohn et al., 2014; Sarris et al., 2015; Le Roux, 2015).类似地, 水稻CNL类抗病蛋白RGA4 (R Gene Analog 4)和RGA5 (R Gene Analog 5)同样以复合体的形式发挥作用.RGA5通过CC结构域与RGA4蛋白互作, 在无效应蛋白刺激时抑制后者的激活(Césari et al., 2014).此外, Prf/Pto复合体介导的抗病反应也需要抗病蛋白NRC2a/b和NRC3的参与(Jones et al., 2016; Wu et al., 2016).但不同抗病蛋白间是如何互作的还有待进一步研究. ...
1 2012
... 植物体内大部分的抗病基因转录水平较低, 在各种组织中广泛表达.有研究表明, 病原菌入侵后植物基因组GC和CHH位点甲基化水平降低, 抗病基因表达量升高, 表明抗病基因的转录受DNA甲基化调控.例如, H3K4甲基转移酶ATXR7 (Arabidopsis Trithorax Related 7)和H2B单泛素连接酶能够结合到抗病基因的启动子区域, 对其进行甲基化修饰, 调控抗病基因SNC1 (Suppressor of npr1 Constitutive1)和RPP4 (Resistance to Peromospora parasitica 4)的表达(Xia et al., 2013; Zou et al., 2014); MOS1 (Modifier of SNC 1)蛋白N端含有BAT2 (HLA-B Associated Transcript 2)结构域, 也能够与SNC1启动子序列结合, 对其进行甲基化修饰, 正调控SNC1的表达(Li et al., 2010b, 2011).此外, miRNAs能够与抗病基因的保守结构域结合, 导致抗病基因沉默(Fei et al., 2013).例如, miRNAs nta-miR6019 (22nt)和nta- miR6020 (21nt)能够调控烟草抗病基因N的转录.研究发现, N与nta-miR6019和nta-miR6020共表达后导致抗病蛋白N介导的TMV抗性减弱, 表明miRNA在抗病基因转录调节中具有功能性作用(Li et al., 2012), 但其具体的调控机制还不是很清楚. ...
1 2011
... 植物体内大部分的抗病基因转录水平较低, 在各种组织中广泛表达.有研究表明, 病原菌入侵后植物基因组GC和CHH位点甲基化水平降低, 抗病基因表达量升高, 表明抗病基因的转录受DNA甲基化调控.例如, H3K4甲基转移酶ATXR7 (Arabidopsis Trithorax Related 7)和H2B单泛素连接酶能够结合到抗病基因的启动子区域, 对其进行甲基化修饰, 调控抗病基因SNC1 (Suppressor of npr1 Constitutive1)和RPP4 (Resistance to Peromospora parasitica 4)的表达(Xia et al., 2013; Zou et al., 2014); MOS1 (Modifier of SNC 1)蛋白N端含有BAT2 (HLA-B Associated Transcript 2)结构域, 也能够与SNC1启动子序列结合, 对其进行甲基化修饰, 正调控SNC1的表达(Li et al., 2010b, 2011).此外, miRNAs能够与抗病基因的保守结构域结合, 导致抗病基因沉默(Fei et al., 2013).例如, miRNAs nta-miR6019 (22nt)和nta- miR6020 (21nt)能够调控烟草抗病基因N的转录.研究发现, N与nta-miR6019和nta-miR6020共表达后导致抗病蛋白N介导的TMV抗性减弱, 表明miRNA在抗病基因转录调节中具有功能性作用(Li et al., 2012), 但其具体的调控机制还不是很清楚. ...
1 2010
... 抗病蛋白的降解受到泛素化蛋白酶体介导的降解途径调控(图2C).研究表明, F-box蛋白cpr1 (constitutive expresser of PR genes 1)功能缺失突变体中抗病蛋白SNC1和RPS2 (Resistance to Pseu- domonas syringae 2)积累, 表明SCFCPR1E3复合体调控抗病蛋白SNC1和RPS2的降解(Cheng et al., 2011).snc1增强子筛选实验证明, E4连接酶MUSE3 (Mutants snc1 Enhancing 3)、TRAR蛋白MUS13和MUSE14能够与SCFCPR1E3复合体互作, 共同调控抗病蛋白SNC1和RPS2的降解(Huang et al., 2014, 2016).MUSE7编码1个功能未知的激酶底物蛋白, 负调控SNC1、RPS2和RPM1蛋白的积累(Johnson et al., 2016).与HSP90复合体作为分子伴侣调控抗病蛋白的装配与成熟不同, HSP90和SGT1也能够与SKP1 (S Phase Kinase Associated Protein 1)-CUL- LIN-F-box-E3泛素复合体相互作用, 调控抗病蛋白的降解(Willhoft et al., 2017).遗传分析表明, hsp90功能获得突变体中抗病蛋白SNC1积累(Huang et al., 2014).sgt1b缺失突变能够抑制rar1突变体中RPS5的积累(Holt et al., 2005).生化实验证明, HSP90和SGT1b能够与SKP1和CULLIN1互作, 促进蛋白降解; RAR1也能够结合COP9 (Constitutive Photo- morpho-genesis Complex 9)蛋白, 抑制SCF复合体的活性, 进而调控抗病蛋白的降解(Azevedo et al., 2002).另外, SGT1a和SGT1b能够与SRFR1 (Suppressor of rps4-RLD)蛋白互作, 负调控抗病蛋白SNC1、RPS2和RPS4的积累(Li et al., 2010a).E1泛素连接酶UBA1 (Ubiquitin Activating 1)与26S蛋白酶体亚基RPN1a (Proteasome Regulatory Subunint S2 1A)、RPN8a (RP Non-ATPase Subunit 8A)和RPT2a (Regulatory Particle AAA-ATPase 2A)参与调控抗病蛋白的降解(Goritschnig et al., 2007; Yao et al., 2012). ...
1 2010
... 植物体内大部分的抗病基因转录水平较低, 在各种组织中广泛表达.有研究表明, 病原菌入侵后植物基因组GC和CHH位点甲基化水平降低, 抗病基因表达量升高, 表明抗病基因的转录受DNA甲基化调控.例如, H3K4甲基转移酶ATXR7 (Arabidopsis Trithorax Related 7)和H2B单泛素连接酶能够结合到抗病基因的启动子区域, 对其进行甲基化修饰, 调控抗病基因SNC1 (Suppressor of npr1 Constitutive1)和RPP4 (Resistance to Peromospora parasitica 4)的表达(Xia et al., 2013; Zou et al., 2014); MOS1 (Modifier of SNC 1)蛋白N端含有BAT2 (HLA-B Associated Transcript 2)结构域, 也能够与SNC1启动子序列结合, 对其进行甲基化修饰, 正调控SNC1的表达(Li et al., 2010b, 2011).此外, miRNAs能够与抗病基因的保守结构域结合, 导致抗病基因沉默(Fei et al., 2013).例如, miRNAs nta-miR6019 (22nt)和nta- miR6020 (21nt)能够调控烟草抗病基因N的转录.研究发现, N与nta-miR6019和nta-miR6020共表达后导致抗病蛋白N介导的TMV抗性减弱, 表明miRNA在抗病基因转录调节中具有功能性作用(Li et al., 2012), 但其具体的调控机制还不是很清楚. ...
1 2011
... 抗病蛋白通过保卫效应蛋白对靶蛋白的攻击激活免疫响应, 抑制病原菌的扩散.保卫模式很好地解释了RPM1 (Resistant to Pseudomonas syringae pv. maculicola 1)、RPS2 (Resistant to Pseudomonas syringae 2)以及RPS5等抗病蛋白的激活方式.目前, 对于RIN4 (RPM1 Interacting Protein 4)介导的抗病蛋白激活模式研究得较为清楚.效应蛋白AvrRpm1 (Avirulence Resistant to Pseudomonas syringae pv. maculicola 1)和AvrB (Avirulence Pseudomonas syringae B)与RIN4蛋白结合后, 蛋白激酶RIPK (RPM1 Induced Protein Kinase)对RIN4蛋白21位苏氨酸、160位丝氨酸和166位苏氨酸进行磷酸化修饰, 导致RPM1被激活, 从而触发植物免疫响应(Mackey et al., 2002; Chung et al., 2011; Liu et al., 2011).除此之外, 效应蛋白诱导的RIN4裂解也能够激活抗病蛋白.效应蛋白AvrRpt2 (Avirulence Resistant to Pseudomonas syringae pv. tomato 2)具有蛋白酶体活性, 能够裂解RIN4, 从而激活RPS2依赖的免疫响应(Axtell and Staskawicz, 2003; Mackey et al., 2003).研究表明, 拟南芥CNL类抗病蛋白RPS5也具有类似的识别方式.效应蛋白AvrPphB (Avirulence Pseudomonas phaseolicola B)具有半胱氨酸蛋白酶活性, 裂解与RPS5相结合的蛋白激酶PBS1 (Protein Kinase AvrPphB Susceptible1).在无效应蛋白时, PBS1激酶与RPS5蛋白CC结构域相互作用, 使RPS5保持失活状态; 当效应蛋白存在时, AvrPphB裂解PBS1导致RPS5蛋白构象改变并被激活, 触 发植物免疫响应(Qi et al., 2012, 2014; Kim et al., 2016). ...
1 2015
... 抗病蛋白的稳定性受蛋白折叠装配及降解的影响.抗病蛋白的装配过程需要分子伴侣的协助(图2B).热激蛋白HSP90 (Heat Shock Protein 90)与分子共伴侣蛋白SGT1 (Suppressor of G2 Allele of Skp1)和RAR1 (Required for Mla12 Resistance 1)共同调控抗病蛋白的正确折叠与稳定.hsp90突变体中抗病蛋白RPM1和RPS5表达量降低.rar1和sgt1突变体中RPS5介导的病原菌抗性减弱.生化实验证明, HSP90的ATPase结构域能够与SGT1蛋白CS (Present in CHP and SGT1 Proteins)结构域互作, 而RAR1蛋白的2个锌结合结构域CHORD-I (Cysteine and Histidine Rich Domain)和CHORD-II分别与HSP90和SGT1蛋白结合, 形成复合体, 共同调控抗病蛋白的成熟(Azevedo et al., 2002; Hubert et al., 2003; Huang et al., 2014).之前的研究证明, 磷酸酶PP5 (Protein Phosphatase 5)能够与HSP90相互作用, 调控抗病蛋白I-2介导的免疫响应(De La Fuente Bentem van, 2005).而最新研究表明, 磷酸酶ibr5 (indole 3 butyric acid response 5)缺失导致抗病蛋白CHS3 (Chilling Sensitive 3)、RPM1和RPS4表达量降低, 同时IBR5能够与HSP90-RAR1-SGT1复合体相互作用, 证明磷酸酶能够作为分子共伴侣蛋白与HSP90复合体一起调节抗病蛋白的稳定性(Liu et al., 2015). ...
1 2017
... 除上述识别模式外, 还存在一些新的抗病蛋白激活模式.植物初级免疫PTI激活后, MAPK (Mitogen Activated Protein Kinase)信号级联通路被激活, 而微生物分泌的效应蛋白能够结合到MAPK通路相关蛋白, 抑制PTI免疫激活.遗传筛选发现, summ1 (suppressor of mkk1mkk2 1)和summ2能够抑制mpk4 (map kinase 4)突变体中免疫激活的表型, SUMM1为蛋白激酶MEKK2, 而SUMM2为抗病蛋白.该结果表明, 抗病蛋白能够监控MEKK1-MKK1/MKK2-MPK4信号级联通路.生化实验证明, MPK4能够磷酸化MEKK2, 但不能与SUMM2直接互作.遗传分析表明, MEKK2位于SUMM2上游, 但MEKK2是如何激活SUMM2的还不清楚(Kong et al., 2012; Zhang et al., 2012).随后的研究表明, MPK4能够磷酸化PAT1 (Arabidopsis Homolog of Yeast PAT1), 抑制SUMM2激活(Roux et al., 2015).最新研究表明, 蛋白激酶CRCK3 (Cal- modulin Binding Receptor like Cytoplasmic Kinase 3)也能被MPK4磷酸化, 共同调控SUMM2依赖的免疫响应(Zhang et al., 2017).无效应蛋白时, MPK4磷酸化底物蛋白, 使得SUMM2处于稳定状态; 而效应蛋白HopAI1与MPK4结合后, 抑制MPK4激酶活性, 使其不能将底物蛋白磷酸化, 导致SUMM2被激活, 触发植物免疫响应.此外, SUMM2还能够监控MPK3与MPK6引起的免疫响应(Genot et al., 2017).除上述激酶以外, 磷酸酶mkp1 (map kinase phosphat- ase1)和ptp1 (protein tyrosine phosphatase1)突变体也表现出SNC1依赖的免疫自激活表型.生化分析结果表明, MKP1能够与MPK3、MPK4和MPK6相互作用, 去磷酸化MPK6, 抑制其激酶活性, 从而拮抗调控抗病蛋白依赖的免疫激活(Schweighofer et al., 2007; Bartels et al., 2009; Anderson et al., 2011).蛋白磷酸酶AP2C1 (Arabidopsis Phosphatase 2C 1)也能够与MPK4和MPK6互作, 通过去磷酸化修饰抑制MPK4和MPK6激酶活性, 调控植物免疫响应.除MAPK依赖的途径外, 钙离子依赖的蛋白激酶CPK1 (Calcium-dependent Protein Kinase 1)、CPK2、CP- K4、CPK5、CPK6和CPK11也能够调控抗病蛋白介导的免疫响应.最新研究表明, CPK5能够与抗病蛋白TN2 (TIR-NBS2)的NBS结构域相互作用, 调控TN2依赖的免疫响应(Liu et al., 2017).综上所述, 抗病蛋白通过不同的识别模式对效应蛋白进行监控, 并且该过程多种多样. ...
1 2012
... NB-LRR抗病蛋白作为一类免疫监测因子, 在无效应蛋白激发时, 通过不同结构域间的互作保持抗病蛋白处于失活状态.不同结构域间的互作方式主要有4种.(1) 某些抗病蛋白LRR结构域能够与效应因子直接互作, 该类蛋白NBS结构域与LRR中效应因子结合区域互作, 抑制LRR结构域的激活, 如L6 (L locus 6)抗病蛋白(Bernoux et al., 2011).(2) 抗病蛋白N端区域与NBS结构域共同介导免疫激活, 该类蛋白的LRR结构域与ARC2区域互作, 抑制N端激活域或NBS结构域的功能, 使抗病蛋白处于稳定状态, 如RPS5 (Resistant to Pseudomonas syringae 5)和RPP1 (Resistant to Pseudomonas parasitica 1)等(Rairdan and Moffett, 2006; Qi et al., 2012; Sloot- weg and Goverse, 2013).(3) 抗病蛋白的CC结构域作为其功能结构域, 其NBS结构域与CC结构域结合, 抑制抗病蛋白自激活, 如Rp1 (Resistant to Puccinia sorghi 1)抗病蛋白(Wang et al., 2015).(4) 许多抗病蛋白N端与其它蛋白互作, 共同对效应因子进行监控, LRR结构域与N端区域互作, 使抗病蛋白保持失活状态, 如Prf (Du et al., 2012; Lukasik-shreepaathy et al., 2012). ...
1 2003
... 抗病蛋白通过保卫效应蛋白对靶蛋白的攻击激活免疫响应, 抑制病原菌的扩散.保卫模式很好地解释了RPM1 (Resistant to Pseudomonas syringae pv. maculicola 1)、RPS2 (Resistant to Pseudomonas syringae 2)以及RPS5等抗病蛋白的激活方式.目前, 对于RIN4 (RPM1 Interacting Protein 4)介导的抗病蛋白激活模式研究得较为清楚.效应蛋白AvrRpm1 (Avirulence Resistant to Pseudomonas syringae pv. maculicola 1)和AvrB (Avirulence Pseudomonas syringae B)与RIN4蛋白结合后, 蛋白激酶RIPK (RPM1 Induced Protein Kinase)对RIN4蛋白21位苏氨酸、160位丝氨酸和166位苏氨酸进行磷酸化修饰, 导致RPM1被激活, 从而触发植物免疫响应(Mackey et al., 2002; Chung et al., 2011; Liu et al., 2011).除此之外, 效应蛋白诱导的RIN4裂解也能够激活抗病蛋白.效应蛋白AvrRpt2 (Avirulence Resistant to Pseudomonas syringae pv. tomato 2)具有蛋白酶体活性, 能够裂解RIN4, 从而激活RPS2依赖的免疫响应(Axtell and Staskawicz, 2003; Mackey et al., 2003).研究表明, 拟南芥CNL类抗病蛋白RPS5也具有类似的识别方式.效应蛋白AvrPphB (Avirulence Pseudomonas phaseolicola B)具有半胱氨酸蛋白酶活性, 裂解与RPS5相结合的蛋白激酶PBS1 (Protein Kinase AvrPphB Susceptible1).在无效应蛋白时, PBS1激酶与RPS5蛋白CC结构域相互作用, 使RPS5保持失活状态; 当效应蛋白存在时, AvrPphB裂解PBS1导致RPS5蛋白构象改变并被激活, 触 发植物免疫响应(Qi et al., 2012, 2014; Kim et al., 2016). ...
1 2002
... 抗病蛋白通过保卫效应蛋白对靶蛋白的攻击激活免疫响应, 抑制病原菌的扩散.保卫模式很好地解释了RPM1 (Resistant to Pseudomonas syringae pv. maculicola 1)、RPS2 (Resistant to Pseudomonas syringae 2)以及RPS5等抗病蛋白的激活方式.目前, 对于RIN4 (RPM1 Interacting Protein 4)介导的抗病蛋白激活模式研究得较为清楚.效应蛋白AvrRpm1 (Avirulence Resistant to Pseudomonas syringae pv. maculicola 1)和AvrB (Avirulence Pseudomonas syringae B)与RIN4蛋白结合后, 蛋白激酶RIPK (RPM1 Induced Protein Kinase)对RIN4蛋白21位苏氨酸、160位丝氨酸和166位苏氨酸进行磷酸化修饰, 导致RPM1被激活, 从而触发植物免疫响应(Mackey et al., 2002; Chung et al., 2011; Liu et al., 2011).除此之外, 效应蛋白诱导的RIN4裂解也能够激活抗病蛋白.效应蛋白AvrRpt2 (Avirulence Resistant to Pseudomonas syringae pv. tomato 2)具有蛋白酶体活性, 能够裂解RIN4, 从而激活RPS2依赖的免疫响应(Axtell and Staskawicz, 2003; Mackey et al., 2003).研究表明, 拟南芥CNL类抗病蛋白RPS5也具有类似的识别方式.效应蛋白AvrPphB (Avirulence Pseudomonas phaseolicola B)具有半胱氨酸蛋白酶活性, 裂解与RPS5相结合的蛋白激酶PBS1 (Protein Kinase AvrPphB Susceptible1).在无效应蛋白时, PBS1激酶与RPS5蛋白CC结构域相互作用, 使RPS5保持失活状态; 当效应蛋白存在时, AvrPphB裂解PBS1导致RPS5蛋白构象改变并被激活, 触 发植物免疫响应(Qi et al., 2012, 2014; Kim et al., 2016). ...
1 2011
... TNL类抗病蛋白N端包含TIR结构域, 其与果蝇(Droso- phila) Toll蛋白或白介素1受体N端序列类似, 在动植物中高度保守.晶体结构分析显示, TIR结构域通常由核心的5个β-片层和外围的5个α-螺旋构成.TIR结构域能够识别效应蛋白.例如, 烟草(Nicotiana tabacum) N蛋白的TIR结构域能够特异地识别花叶病毒分泌的效应蛋白p50 (Swiderski et al., 2009).此外, 该结构域也能够介导蛋白之间的相互作用.例如, RRS1 (Resistant to Ralstonia solanacearum 1)和RPS4 (Resistant to Pseudomonas syringae 4)蛋白通过TIR结构域中保守的丝氨酸-组氨酸(ser-his, SH)基序互作, 形成结合界面, 共同介导植物对病原菌的抗性(Williams et al., 2014).CNL类抗病蛋白N端包含CC结构域, 即不规则卷曲螺旋结构.通过对CC结构域蛋白结构进行解析, 发现该结构域由7个氨基酸重复序列组成, 包含2个或多个α-螺旋, 构成一个超螺旋结构.CC结构域缺乏保守性, 其中唯一的保守基序为EDVID.但并非所有的CNL蛋白CC结构域都存在该基序.CC结构域在蛋白互作中发挥重要功能.例如, CNL抗性蛋白Rx的CC结构域能够与细胞内辅因子RanGAP2直接互作, 共同监控马铃薯(Solanum tuberosum) X病毒.晶体结构揭示, 该蛋白CC结构域具有保守的EDVID基序, 其第90位色氨酸能够与Ran- GAP2的第89位丙氨酸通过疏水作用相结合.而Rx蛋白CC结构域与RanGAP2相结合的2个α-螺旋为调控Rx蛋白入核的区域, RanGAP2通过与该区域结合阻止Rx进入细胞核, 增强植物抗病能力(Tameling and Baulcombe, 2007; Hao et al., 2013).此外, 通过解析MLA10 (Mildew Resistance Locus A10)抗病蛋白CC结构域的晶体结构, 发现该蛋白CC结构域由2个长α-螺旋单体组成, 2条链相距较远, 且内侧多为疏水性氨基酸, 2个MLA10蛋白通过CC结构域形成同源二聚体, 维持整个蛋白的稳定性(Maekawa et al., 2011). ...
1 2012
... 除内在调控方式外, 温度、湿度和昼夜节律等环境因素也能够影响抗病蛋白的稳定性.遗传筛选实验表明, 许多免疫自激活突变体均表现出温度依赖的表型, 如chs1/2/3、snc1-1、bon1-1和mkp1等.chs1/2/3在16 °C下表现为生长缺陷, 而22°C下生长正常; snc1-1和bon1-1等突变体在22°C下植株矮小、叶片卷曲, 而这种缺陷表型可以被高温恢复或部分恢复(Hua et al., 2001; Yang and Hua, 2004; Yang et al., 2010; Wang et al., 2013; Zbierzak et al., 2013).高温影响抗病蛋白与病原菌间的相互作用, 并且高温能够抑制抗病蛋白从稳定状态向激活状态转变.此外, 高温下植物体内水杨酸(salicylic acid, SA)和脱落酸(absci- sic acid, ABA)等激素水平下降, 增强了植物的抗菌性(Mang et al., 2012).环境湿度和昼夜周期节律变化也能够调控抗病蛋白的稳定性.例如, 高湿度能够抑制抗病蛋白SSI (Suppressor of Salicylic Acid Insensitive)与SLH1 (an allele of RRS1)依赖的免疫激活(Zhou et al., 2004; Noutoshi et al., 2005); 昼夜节律可以影响抗病基因RPP4 (Resistant to Peronospora parasitica 4)的转录(Wang et al., 2011). ...
1 2012
... 植物在生长发育过程中为了应对复杂的环境变化, 进化形成了2个不同层次的防御反应: 初级免疫反应和次级免疫反应.宿主细胞膜上的模式识别受体(pattern recognition receptors, PRR)识别病原微生物相关分子模式(pathogen associated molecular patterns, PAMP), 激活PAMP触发的免疫反应(PAMP triggered immunity, PTI) (Monaghan and Zipfel, 2012; Schwessinger and Ronald, 2012; Dangl et al., 2013), 即初级免疫反应.为了应对植物初级免疫反应, 微生物分泌出一些毒性因子(即效应因子)注入到植物细胞内, 抑制PTI, 导致植物感病.在应对效应因子的入侵方面, 植物进化出能够特异识别效应因子的抗性蛋白(resistance protein, R), 激活效应因子触发的免疫反应(effectors triggered immu- nity, ETI), 即为次级免疫反应(Tsuda and Katagiri, 2010; Howden and Huitema, 2012).ETI比PTI更为强烈和迅速, 导致局部的程序性细胞死亡(program- med cell death, PCD), 引起植物超敏反应(hyper- sensitive response, HR), 更为有效地遏制病原菌的进一步扩散(Jones and Dangl, 2006). ...
1 2006
... 抗病蛋白与病原菌的靶蛋白类似物相互作用, 效应蛋白将该靶蛋白类似物误认为其真正的靶蛋白, 对其进行识别修饰, 导致免疫响应激活.例如, Prf蛋白的激活方式为2个Prf蛋白通过N端结构域相互作用形成同源二聚体, 并与Pto (Pseudomonas syringae pv. tomato)蛋白激酶结合形成复合体(Mucyn et al., 2006; Gutierrez et al., 2010).无效应蛋白AvrPto时, Pto激酶上的β-1与P+1环抑制抗病蛋白激活; 当效应蛋白AvrPto存在时, Prf/Pto复合体中的1个Pto激酶作为诱饵蛋白与AvrPto结合, 使Pto激酶结构改变, 导致Prf蛋白NBS结构域对Pto的抑制作用解除, 随后另一个Pto激酶磷酸化与效应蛋白结合的Pto激酶, 抗病蛋白Prf被激活, 引起HR反应(Narusaka et al., 2009; Ntoukakis et al., 2013). ...
2 2009
... 抗病蛋白与病原菌的靶蛋白类似物相互作用, 效应蛋白将该靶蛋白类似物误认为其真正的靶蛋白, 对其进行识别修饰, 导致免疫响应激活.例如, Prf蛋白的激活方式为2个Prf蛋白通过N端结构域相互作用形成同源二聚体, 并与Pto (Pseudomonas syringae pv. tomato)蛋白激酶结合形成复合体(Mucyn et al., 2006; Gutierrez et al., 2010).无效应蛋白AvrPto时, Pto激酶上的β-1与P+1环抑制抗病蛋白激活; 当效应蛋白AvrPto存在时, Prf/Pto复合体中的1个Pto激酶作为诱饵蛋白与AvrPto结合, 使Pto激酶结构改变, 导致Prf蛋白NBS结构域对Pto的抑制作用解除, 随后另一个Pto激酶磷酸化与效应蛋白结合的Pto激酶, 抗病蛋白Prf被激活, 引起HR反应(Narusaka et al., 2009; Ntoukakis et al., 2013). ... ... 许多抗病蛋白需要与其它抗病蛋白形成复合体, 共同调控植物免疫响应.例如, 拟南芥TNL类抗病蛋白RPS4/RRS1调控效应蛋白AvrRps4 (Pst. DC3000 Avr Rps4)和PopP2 (R. Solanacearum-PopP2)诱导的免疫反应.无效应蛋白时, RPS4的TIR结构域与RR- S1的TIR结构域相互作用, 使抗病蛋白RPS4/RRS1处于失活状态; 当效应蛋白存在时, RRS1羧基端包含的与转录因子WRKY相似的DNA结合结构域能够作为诱饵与效应蛋白PopP2结合.AvrRps4与RRS1的WRKY结构域结合后启动抗病反应; 而PopP2具有乙酰基转移酶活性, 能够将RRS1 WRKYGQK结构中第1 221位赖氨酸乙酰化, 导致其失去DNA结合能力, 激活RPS4/RRS1复合体, 触发植物免疫响应(Narusaka et al., 2009; Sohn et al., 2014; Sarris et al., 2015; Le Roux, 2015).类似地, 水稻CNL类抗病蛋白RGA4 (R Gene Analog 4)和RGA5 (R Gene Analog 5)同样以复合体的形式发挥作用.RGA5通过CC结构域与RGA4蛋白互作, 在无效应蛋白刺激时抑制后者的激活(Césari et al., 2014).此外, Prf/Pto复合体介导的抗病反应也需要抗病蛋白NRC2a/b和NRC3的参与(Jones et al., 2016; Wu et al., 2016).但不同抗病蛋白间是如何互作的还有待进一步研究. ...
1 2005
... 除内在调控方式外, 温度、湿度和昼夜节律等环境因素也能够影响抗病蛋白的稳定性.遗传筛选实验表明, 许多免疫自激活突变体均表现出温度依赖的表型, 如chs1/2/3、snc1-1、bon1-1和mkp1等.chs1/2/3在16 °C下表现为生长缺陷, 而22°C下生长正常; snc1-1和bon1-1等突变体在22°C下植株矮小、叶片卷曲, 而这种缺陷表型可以被高温恢复或部分恢复(Hua et al., 2001; Yang and Hua, 2004; Yang et al., 2010; Wang et al., 2013; Zbierzak et al., 2013).高温影响抗病蛋白与病原菌间的相互作用, 并且高温能够抑制抗病蛋白从稳定状态向激活状态转变.此外, 高温下植物体内水杨酸(salicylic acid, SA)和脱落酸(absci- sic acid, ABA)等激素水平下降, 增强了植物的抗菌性(Mang et al., 2012).环境湿度和昼夜周期节律变化也能够调控抗病蛋白的稳定性.例如, 高湿度能够抑制抗病蛋白SSI (Suppressor of Salicylic Acid Insensitive)与SLH1 (an allele of RRS1)依赖的免疫激活(Zhou et al., 2004; Noutoshi et al., 2005); 昼夜节律可以影响抗病基因RPP4 (Resistant to Peronospora parasitica 4)的转录(Wang et al., 2011). ...
2 2013
... NBS结构域通过催化核苷酸的结合与水解改变抗病蛋白的构象, 激活植物免疫响应.在无效应蛋白时, NBS结构域与ADP结合, 抗病蛋白处于失活状态; 在效应蛋白存在时, NBS结构域结合ATP, 抗病蛋白被激活.NBS结构域包含3个保守的亚结构域: NB、ARC1 (Apaf-1, R-protein and CED-4)和ARC2.其中, ARC亚结构域序列与哺乳动物细胞蛋白激酶激活凋亡因子Apaf1 (Apoptotic protease-activating factor 1)和秀丽线虫(Caenorhabditis elegans)细胞凋亡相关蛋白CED-4高度相似.在这些保守结构域中包含8个保守基序: P-loop、RNBS-A、Kinase2、RNBS-B、RNBS-C、GLPL、RNBS-D和MHD.该保守序列中的某些氨基酸突变后, 即使无效应蛋白存在, 仍会出现持续免疫激活的现象, 而有些位点突变可导致抗病蛋白持续处于失活状态, 不能激活免疫响应.例如, 亚麻抗锈病蛋白M中MHD结构域第555位氨基酸D突变为V后, 使抗病蛋白变为持续激活状态, 引发HR反应(Williams et al., 2011); 而烟草抗病蛋白Prf (Pto Resistance and Fenthion)第1 128位氨基酸K突变为A后, Prf处于持续失活状态, 即使效应蛋白存在也无法激活抗病蛋白和触发免疫响应(Ntoukakis et al., 2013).因此, NBS结构域对于抗病蛋白活性的开闭至关重要. ... ... 抗病蛋白与病原菌的靶蛋白类似物相互作用, 效应蛋白将该靶蛋白类似物误认为其真正的靶蛋白, 对其进行识别修饰, 导致免疫响应激活.例如, Prf蛋白的激活方式为2个Prf蛋白通过N端结构域相互作用形成同源二聚体, 并与Pto (Pseudomonas syringae pv. tomato)蛋白激酶结合形成复合体(Mucyn et al., 2006; Gutierrez et al., 2010).无效应蛋白AvrPto时, Pto激酶上的β-1与P+1环抑制抗病蛋白激活; 当效应蛋白AvrPto存在时, Prf/Pto复合体中的1个Pto激酶作为诱饵蛋白与AvrPto结合, 使Pto激酶结构改变, 导致Prf蛋白NBS结构域对Pto的抑制作用解除, 随后另一个Pto激酶磷酸化与效应蛋白结合的Pto激酶, 抗病蛋白Prf被激活, 引起HR反应(Narusaka et al., 2009; Ntoukakis et al., 2013). ...
1 2013
... 植物免疫响应还与激素信号通路密切相关.遗传分析证明, SA相关突变体sid2 (salicylic acid induction deficient 2)和eds1能够部分恢复抗病蛋白HRT (Hypersensitive Response to TCV)和RPP8 (Recognition of Peronospora parasitic 8)激活后造成的免疫激活表型(Chandrashekara et al., 2004; Wiermer et al., 2005), 表明抗病蛋白的激活能够引起SA的积累.SA与细胞质内多聚的NPR1 (Non-expresser of PR Genes 1)结合后, NPR1解聚形成单体进入细胞核, 结合到转录因子的TGA区域, 激活PR1 (Pathogenesis Related Gene 1)以及植物防御反应相关转录因子WRKY的转录(图2); 随后, NPR1同源蛋白NPR3和NPR4作为接头蛋白连接NPR1与Cullin3 E3连接酶, NPR1被降解(Fu et al., 2012; Wu et al., 2012; Paj- erowska-Mukhtar et al., 2013).此外, 越来越多的研究表明, 多种激素信号均参与调控植物免疫响应, 但具体的调控机制还不清楚(De Bruyne et al., 2014). ...
2 2012
... NB-LRR抗病蛋白作为一类免疫监测因子, 在无效应蛋白激发时, 通过不同结构域间的互作保持抗病蛋白处于失活状态.不同结构域间的互作方式主要有4种.(1) 某些抗病蛋白LRR结构域能够与效应因子直接互作, 该类蛋白NBS结构域与LRR中效应因子结合区域互作, 抑制LRR结构域的激活, 如L6 (L locus 6)抗病蛋白(Bernoux et al., 2011).(2) 抗病蛋白N端区域与NBS结构域共同介导免疫激活, 该类蛋白的LRR结构域与ARC2区域互作, 抑制N端激活域或NBS结构域的功能, 使抗病蛋白处于稳定状态, 如RPS5 (Resistant to Pseudomonas syringae 5)和RPP1 (Resistant to Pseudomonas parasitica 1)等(Rairdan and Moffett, 2006; Qi et al., 2012; Sloot- weg and Goverse, 2013).(3) 抗病蛋白的CC结构域作为其功能结构域, 其NBS结构域与CC结构域结合, 抑制抗病蛋白自激活, 如Rp1 (Resistant to Puccinia sorghi 1)抗病蛋白(Wang et al., 2015).(4) 许多抗病蛋白N端与其它蛋白互作, 共同对效应因子进行监控, LRR结构域与N端区域互作, 使抗病蛋白保持失活状态, 如Prf (Du et al., 2012; Lukasik-shreepaathy et al., 2012). ... ... 抗病蛋白通过保卫效应蛋白对靶蛋白的攻击激活免疫响应, 抑制病原菌的扩散.保卫模式很好地解释了RPM1 (Resistant to Pseudomonas syringae pv. maculicola 1)、RPS2 (Resistant to Pseudomonas syringae 2)以及RPS5等抗病蛋白的激活方式.目前, 对于RIN4 (RPM1 Interacting Protein 4)介导的抗病蛋白激活模式研究得较为清楚.效应蛋白AvrRpm1 (Avirulence Resistant to Pseudomonas syringae pv. maculicola 1)和AvrB (Avirulence Pseudomonas syringae B)与RIN4蛋白结合后, 蛋白激酶RIPK (RPM1 Induced Protein Kinase)对RIN4蛋白21位苏氨酸、160位丝氨酸和166位苏氨酸进行磷酸化修饰, 导致RPM1被激活, 从而触发植物免疫响应(Mackey et al., 2002; Chung et al., 2011; Liu et al., 2011).除此之外, 效应蛋白诱导的RIN4裂解也能够激活抗病蛋白.效应蛋白AvrRpt2 (Avirulence Resistant to Pseudomonas syringae pv. tomato 2)具有蛋白酶体活性, 能够裂解RIN4, 从而激活RPS2依赖的免疫响应(Axtell and Staskawicz, 2003; Mackey et al., 2003).研究表明, 拟南芥CNL类抗病蛋白RPS5也具有类似的识别方式.效应蛋白AvrPphB (Avirulence Pseudomonas phaseolicola B)具有半胱氨酸蛋白酶活性, 裂解与RPS5相结合的蛋白激酶PBS1 (Protein Kinase AvrPphB Susceptible1).在无效应蛋白时, PBS1激酶与RPS5蛋白CC结构域相互作用, 使RPS5保持失活状态; 当效应蛋白存在时, AvrPphB裂解PBS1导致RPS5蛋白构象改变并被激活, 触 发植物免疫响应(Qi et al., 2012, 2014; Kim et al., 2016). ...
1 2014
... 抗病蛋白通过保卫效应蛋白对靶蛋白的攻击激活免疫响应, 抑制病原菌的扩散.保卫模式很好地解释了RPM1 (Resistant to Pseudomonas syringae pv. maculicola 1)、RPS2 (Resistant to Pseudomonas syringae 2)以及RPS5等抗病蛋白的激活方式.目前, 对于RIN4 (RPM1 Interacting Protein 4)介导的抗病蛋白激活模式研究得较为清楚.效应蛋白AvrRpm1 (Avirulence Resistant to Pseudomonas syringae pv. maculicola 1)和AvrB (Avirulence Pseudomonas syringae B)与RIN4蛋白结合后, 蛋白激酶RIPK (RPM1 Induced Protein Kinase)对RIN4蛋白21位苏氨酸、160位丝氨酸和166位苏氨酸进行磷酸化修饰, 导致RPM1被激活, 从而触发植物免疫响应(Mackey et al., 2002; Chung et al., 2011; Liu et al., 2011).除此之外, 效应蛋白诱导的RIN4裂解也能够激活抗病蛋白.效应蛋白AvrRpt2 (Avirulence Resistant to Pseudomonas syringae pv. tomato 2)具有蛋白酶体活性, 能够裂解RIN4, 从而激活RPS2依赖的免疫响应(Axtell and Staskawicz, 2003; Mackey et al., 2003).研究表明, 拟南芥CNL类抗病蛋白RPS5也具有类似的识别方式.效应蛋白AvrPphB (Avirulence Pseudomonas phaseolicola B)具有半胱氨酸蛋白酶活性, 裂解与RPS5相结合的蛋白激酶PBS1 (Protein Kinase AvrPphB Susceptible1).在无效应蛋白时, PBS1激酶与RPS5蛋白CC结构域相互作用, 使RPS5保持失活状态; 当效应蛋白存在时, AvrPphB裂解PBS1导致RPS5蛋白构象改变并被激活, 触 发植物免疫响应(Qi et al., 2012, 2014; Kim et al., 2016). ...
1 2006
... NB-LRR抗病蛋白作为一类免疫监测因子, 在无效应蛋白激发时, 通过不同结构域间的互作保持抗病蛋白处于失活状态.不同结构域间的互作方式主要有4种.(1) 某些抗病蛋白LRR结构域能够与效应因子直接互作, 该类蛋白NBS结构域与LRR中效应因子结合区域互作, 抑制LRR结构域的激活, 如L6 (L locus 6)抗病蛋白(Bernoux et al., 2011).(2) 抗病蛋白N端区域与NBS结构域共同介导免疫激活, 该类蛋白的LRR结构域与ARC2区域互作, 抑制N端激活域或NBS结构域的功能, 使抗病蛋白处于稳定状态, 如RPS5 (Resistant to Pseudomonas syringae 5)和RPP1 (Resistant to Pseudomonas parasitica 1)等(Rairdan and Moffett, 2006; Qi et al., 2012; Sloot- weg and Goverse, 2013).(3) 抗病蛋白的CC结构域作为其功能结构域, 其NBS结构域与CC结构域结合, 抑制抗病蛋白自激活, 如Rp1 (Resistant to Puccinia sorghi 1)抗病蛋白(Wang et al., 2015).(4) 许多抗病蛋白N端与其它蛋白互作, 共同对效应因子进行监控, LRR结构域与N端区域互作, 使抗病蛋白保持失活状态, 如Prf (Du et al., 2012; Lukasik-shreepaathy et al., 2012). ...
1 2015
... 除上述识别模式外, 还存在一些新的抗病蛋白激活模式.植物初级免疫PTI激活后, MAPK (Mitogen Activated Protein Kinase)信号级联通路被激活, 而微生物分泌的效应蛋白能够结合到MAPK通路相关蛋白, 抑制PTI免疫激活.遗传筛选发现, summ1 (suppressor of mkk1mkk2 1)和summ2能够抑制mpk4 (map kinase 4)突变体中免疫激活的表型, SUMM1为蛋白激酶MEKK2, 而SUMM2为抗病蛋白.该结果表明, 抗病蛋白能够监控MEKK1-MKK1/MKK2-MPK4信号级联通路.生化实验证明, MPK4能够磷酸化MEKK2, 但不能与SUMM2直接互作.遗传分析表明, MEKK2位于SUMM2上游, 但MEKK2是如何激活SUMM2的还不清楚(Kong et al., 2012; Zhang et al., 2012).随后的研究表明, MPK4能够磷酸化PAT1 (Arabidopsis Homolog of Yeast PAT1), 抑制SUMM2激活(Roux et al., 2015).最新研究表明, 蛋白激酶CRCK3 (Cal- modulin Binding Receptor like Cytoplasmic Kinase 3)也能被MPK4磷酸化, 共同调控SUMM2依赖的免疫响应(Zhang et al., 2017).无效应蛋白时, MPK4磷酸化底物蛋白, 使得SUMM2处于稳定状态; 而效应蛋白HopAI1与MPK4结合后, 抑制MPK4激酶活性, 使其不能将底物蛋白磷酸化, 导致SUMM2被激活, 触发植物免疫响应.此外, SUMM2还能够监控MPK3与MPK6引起的免疫响应(Genot et al., 2017).除上述激酶以外, 磷酸酶mkp1 (map kinase phosphat- ase1)和ptp1 (protein tyrosine phosphatase1)突变体也表现出SNC1依赖的免疫自激活表型.生化分析结果表明, MKP1能够与MPK3、MPK4和MPK6相互作用, 去磷酸化MPK6, 抑制其激酶活性, 从而拮抗调控抗病蛋白依赖的免疫激活(Schweighofer et al., 2007; Bartels et al., 2009; Anderson et al., 2011).蛋白磷酸酶AP2C1 (Arabidopsis Phosphatase 2C 1)也能够与MPK4和MPK6互作, 通过去磷酸化修饰抑制MPK4和MPK6激酶活性, 调控植物免疫响应.除MAPK依赖的途径外, 钙离子依赖的蛋白激酶CPK1 (Calcium-dependent Protein Kinase 1)、CPK2、CP- K4、CPK5、CPK6和CPK11也能够调控抗病蛋白介导的免疫响应.最新研究表明, CPK5能够与抗病蛋白TN2 (TIR-NBS2)的NBS结构域相互作用, 调控TN2依赖的免疫响应(Liu et al., 2017).综上所述, 抗病蛋白通过不同的识别模式对效应蛋白进行监控, 并且该过程多种多样. ...
1 2015
... 许多抗病蛋白需要与其它抗病蛋白形成复合体, 共同调控植物免疫响应.例如, 拟南芥TNL类抗病蛋白RPS4/RRS1调控效应蛋白AvrRps4 (Pst. DC3000 Avr Rps4)和PopP2 (R. Solanacearum-PopP2)诱导的免疫反应.无效应蛋白时, RPS4的TIR结构域与RR- S1的TIR结构域相互作用, 使抗病蛋白RPS4/RRS1处于失活状态; 当效应蛋白存在时, RRS1羧基端包含的与转录因子WRKY相似的DNA结合结构域能够作为诱饵与效应蛋白PopP2结合.AvrRps4与RRS1的WRKY结构域结合后启动抗病反应; 而PopP2具有乙酰基转移酶活性, 能够将RRS1 WRKYGQK结构中第1 221位赖氨酸乙酰化, 导致其失去DNA结合能力, 激活RPS4/RRS1复合体, 触发植物免疫响应(Narusaka et al., 2009; Sohn et al., 2014; Sarris et al., 2015; Le Roux, 2015).类似地, 水稻CNL类抗病蛋白RGA4 (R Gene Analog 4)和RGA5 (R Gene Analog 5)同样以复合体的形式发挥作用.RGA5通过CC结构域与RGA4蛋白互作, 在无效应蛋白刺激时抑制后者的激活(Césari et al., 2014).此外, Prf/Pto复合体介导的抗病反应也需要抗病蛋白NRC2a/b和NRC3的参与(Jones et al., 2016; Wu et al., 2016).但不同抗病蛋白间是如何互作的还有待进一步研究. ...
1 2007
... 除上述识别模式外, 还存在一些新的抗病蛋白激活模式.植物初级免疫PTI激活后, MAPK (Mitogen Activated Protein Kinase)信号级联通路被激活, 而微生物分泌的效应蛋白能够结合到MAPK通路相关蛋白, 抑制PTI免疫激活.遗传筛选发现, summ1 (suppressor of mkk1mkk2 1)和summ2能够抑制mpk4 (map kinase 4)突变体中免疫激活的表型, SUMM1为蛋白激酶MEKK2, 而SUMM2为抗病蛋白.该结果表明, 抗病蛋白能够监控MEKK1-MKK1/MKK2-MPK4信号级联通路.生化实验证明, MPK4能够磷酸化MEKK2, 但不能与SUMM2直接互作.遗传分析表明, MEKK2位于SUMM2上游, 但MEKK2是如何激活SUMM2的还不清楚(Kong et al., 2012; Zhang et al., 2012).随后的研究表明, MPK4能够磷酸化PAT1 (Arabidopsis Homolog of Yeast PAT1), 抑制SUMM2激活(Roux et al., 2015).最新研究表明, 蛋白激酶CRCK3 (Cal- modulin Binding Receptor like Cytoplasmic Kinase 3)也能被MPK4磷酸化, 共同调控SUMM2依赖的免疫响应(Zhang et al., 2017).无效应蛋白时, MPK4磷酸化底物蛋白, 使得SUMM2处于稳定状态; 而效应蛋白HopAI1与MPK4结合后, 抑制MPK4激酶活性, 使其不能将底物蛋白磷酸化, 导致SUMM2被激活, 触发植物免疫响应.此外, SUMM2还能够监控MPK3与MPK6引起的免疫响应(Genot et al., 2017).除上述激酶以外, 磷酸酶mkp1 (map kinase phosphat- ase1)和ptp1 (protein tyrosine phosphatase1)突变体也表现出SNC1依赖的免疫自激活表型.生化分析结果表明, MKP1能够与MPK3、MPK4和MPK6相互作用, 去磷酸化MPK6, 抑制其激酶活性, 从而拮抗调控抗病蛋白依赖的免疫激活(Schweighofer et al., 2007; Bartels et al., 2009; Anderson et al., 2011).蛋白磷酸酶AP2C1 (Arabidopsis Phosphatase 2C 1)也能够与MPK4和MPK6互作, 通过去磷酸化修饰抑制MPK4和MPK6激酶活性, 调控植物免疫响应.除MAPK依赖的途径外, 钙离子依赖的蛋白激酶CPK1 (Calcium-dependent Protein Kinase 1)、CPK2、CP- K4、CPK5、CPK6和CPK11也能够调控抗病蛋白介导的免疫响应.最新研究表明, CPK5能够与抗病蛋白TN2 (TIR-NBS2)的NBS结构域相互作用, 调控TN2依赖的免疫响应(Liu et al., 2017).综上所述, 抗病蛋白通过不同的识别模式对效应蛋白进行监控, 并且该过程多种多样. ...
1 2012
... 植物在生长发育过程中为了应对复杂的环境变化, 进化形成了2个不同层次的防御反应: 初级免疫反应和次级免疫反应.宿主细胞膜上的模式识别受体(pattern recognition receptors, PRR)识别病原微生物相关分子模式(pathogen associated molecular patterns, PAMP), 激活PAMP触发的免疫反应(PAMP triggered immunity, PTI) (Monaghan and Zipfel, 2012; Schwessinger and Ronald, 2012; Dangl et al., 2013), 即初级免疫反应.为了应对植物初级免疫反应, 微生物分泌出一些毒性因子(即效应因子)注入到植物细胞内, 抑制PTI, 导致植物感病.在应对效应因子的入侵方面, 植物进化出能够特异识别效应因子的抗性蛋白(resistance protein, R), 激活效应因子触发的免疫反应(effectors triggered immu- nity, ETI), 即为次级免疫反应(Tsuda and Katagiri, 2010; Howden and Huitema, 2012).ETI比PTI更为强烈和迅速, 导致局部的程序性细胞死亡(program- med cell death, PCD), 引起植物超敏反应(hyper- sensitive response, HR), 更为有效地遏制病原菌的进一步扩散(Jones and Dangl, 2006). ...
0 2012
1 2013
... NB-LRR抗病蛋白作为一类免疫监测因子, 在无效应蛋白激发时, 通过不同结构域间的互作保持抗病蛋白处于失活状态.不同结构域间的互作方式主要有4种.(1) 某些抗病蛋白LRR结构域能够与效应因子直接互作, 该类蛋白NBS结构域与LRR中效应因子结合区域互作, 抑制LRR结构域的激活, 如L6 (L locus 6)抗病蛋白(Bernoux et al., 2011).(2) 抗病蛋白N端区域与NBS结构域共同介导免疫激活, 该类蛋白的LRR结构域与ARC2区域互作, 抑制N端激活域或NBS结构域的功能, 使抗病蛋白处于稳定状态, 如RPS5 (Resistant to Pseudomonas syringae 5)和RPP1 (Resistant to Pseudomonas parasitica 1)等(Rairdan and Moffett, 2006; Qi et al., 2012; Sloot- weg and Goverse, 2013).(3) 抗病蛋白的CC结构域作为其功能结构域, 其NBS结构域与CC结构域结合, 抑制抗病蛋白自激活, 如Rp1 (Resistant to Puccinia sorghi 1)抗病蛋白(Wang et al., 2015).(4) 许多抗病蛋白N端与其它蛋白互作, 共同对效应因子进行监控, LRR结构域与N端区域互作, 使抗病蛋白保持失活状态, 如Prf (Du et al., 2012; Lukasik-shreepaathy et al., 2012). ...
1 2014
... 许多抗病蛋白需要与其它抗病蛋白形成复合体, 共同调控植物免疫响应.例如, 拟南芥TNL类抗病蛋白RPS4/RRS1调控效应蛋白AvrRps4 (Pst. DC3000 Avr Rps4)和PopP2 (R. Solanacearum-PopP2)诱导的免疫反应.无效应蛋白时, RPS4的TIR结构域与RR- S1的TIR结构域相互作用, 使抗病蛋白RPS4/RRS1处于失活状态; 当效应蛋白存在时, RRS1羧基端包含的与转录因子WRKY相似的DNA结合结构域能够作为诱饵与效应蛋白PopP2结合.AvrRps4与RRS1的WRKY结构域结合后启动抗病反应; 而PopP2具有乙酰基转移酶活性, 能够将RRS1 WRKYGQK结构中第1 221位赖氨酸乙酰化, 导致其失去DNA结合能力, 激活RPS4/RRS1复合体, 触发植物免疫响应(Narusaka et al., 2009; Sohn et al., 2014; Sarris et al., 2015; Le Roux, 2015).类似地, 水稻CNL类抗病蛋白RGA4 (R Gene Analog 4)和RGA5 (R Gene Analog 5)同样以复合体的形式发挥作用.RGA5通过CC结构域与RGA4蛋白互作, 在无效应蛋白刺激时抑制后者的激活(Césari et al., 2014).此外, Prf/Pto复合体介导的抗病反应也需要抗病蛋白NRC2a/b和NRC3的参与(Jones et al., 2016; Wu et al., 2016).但不同抗病蛋白间是如何互作的还有待进一步研究. ...
1 2009
... TNL类抗病蛋白N端包含TIR结构域, 其与果蝇(Droso- phila) Toll蛋白或白介素1受体N端序列类似, 在动植物中高度保守.晶体结构分析显示, TIR结构域通常由核心的5个β-片层和外围的5个α-螺旋构成.TIR结构域能够识别效应蛋白.例如, 烟草(Nicotiana tabacum) N蛋白的TIR结构域能够特异地识别花叶病毒分泌的效应蛋白p50 (Swiderski et al., 2009).此外, 该结构域也能够介导蛋白之间的相互作用.例如, RRS1 (Resistant to Ralstonia solanacearum 1)和RPS4 (Resistant to Pseudomonas syringae 4)蛋白通过TIR结构域中保守的丝氨酸-组氨酸(ser-his, SH)基序互作, 形成结合界面, 共同介导植物对病原菌的抗性(Williams et al., 2014).CNL类抗病蛋白N端包含CC结构域, 即不规则卷曲螺旋结构.通过对CC结构域蛋白结构进行解析, 发现该结构域由7个氨基酸重复序列组成, 包含2个或多个α-螺旋, 构成一个超螺旋结构.CC结构域缺乏保守性, 其中唯一的保守基序为EDVID.但并非所有的CNL蛋白CC结构域都存在该基序.CC结构域在蛋白互作中发挥重要功能.例如, CNL抗性蛋白Rx的CC结构域能够与细胞内辅因子RanGAP2直接互作, 共同监控马铃薯(Solanum tuberosum) X病毒.晶体结构揭示, 该蛋白CC结构域具有保守的EDVID基序, 其第90位色氨酸能够与Ran- GAP2的第89位丙氨酸通过疏水作用相结合.而Rx蛋白CC结构域与RanGAP2相结合的2个α-螺旋为调控Rx蛋白入核的区域, RanGAP2通过与该区域结合阻止Rx进入细胞核, 增强植物抗病能力(Tameling and Baulcombe, 2007; Hao et al., 2013).此外, 通过解析MLA10 (Mildew Resistance Locus A10)抗病蛋白CC结构域的晶体结构, 发现该蛋白CC结构域由2个长α-螺旋单体组成, 2条链相距较远, 且内侧多为疏水性氨基酸, 2个MLA10蛋白通过CC结构域形成同源二聚体, 维持整个蛋白的稳定性(Maekawa et al., 2011). ...
1 2007
... TNL类抗病蛋白N端包含TIR结构域, 其与果蝇(Droso- phila) Toll蛋白或白介素1受体N端序列类似, 在动植物中高度保守.晶体结构分析显示, TIR结构域通常由核心的5个β-片层和外围的5个α-螺旋构成.TIR结构域能够识别效应蛋白.例如, 烟草(Nicotiana tabacum) N蛋白的TIR结构域能够特异地识别花叶病毒分泌的效应蛋白p50 (Swiderski et al., 2009).此外, 该结构域也能够介导蛋白之间的相互作用.例如, RRS1 (Resistant to Ralstonia solanacearum 1)和RPS4 (Resistant to Pseudomonas syringae 4)蛋白通过TIR结构域中保守的丝氨酸-组氨酸(ser-his, SH)基序互作, 形成结合界面, 共同介导植物对病原菌的抗性(Williams et al., 2014).CNL类抗病蛋白N端包含CC结构域, 即不规则卷曲螺旋结构.通过对CC结构域蛋白结构进行解析, 发现该结构域由7个氨基酸重复序列组成, 包含2个或多个α-螺旋, 构成一个超螺旋结构.CC结构域缺乏保守性, 其中唯一的保守基序为EDVID.但并非所有的CNL蛋白CC结构域都存在该基序.CC结构域在蛋白互作中发挥重要功能.例如, CNL抗性蛋白Rx的CC结构域能够与细胞内辅因子RanGAP2直接互作, 共同监控马铃薯(Solanum tuberosum) X病毒.晶体结构揭示, 该蛋白CC结构域具有保守的EDVID基序, 其第90位色氨酸能够与Ran- GAP2的第89位丙氨酸通过疏水作用相结合.而Rx蛋白CC结构域与RanGAP2相结合的2个α-螺旋为调控Rx蛋白入核的区域, RanGAP2通过与该区域结合阻止Rx进入细胞核, 增强植物抗病能力(Tameling and Baulcombe, 2007; Hao et al., 2013).此外, 通过解析MLA10 (Mildew Resistance Locus A10)抗病蛋白CC结构域的晶体结构, 发现该蛋白CC结构域由2个长α-螺旋单体组成, 2条链相距较远, 且内侧多为疏水性氨基酸, 2个MLA10蛋白通过CC结构域形成同源二聚体, 维持整个蛋白的稳定性(Maekawa et al., 2011). ...
1 2010
... 植物在生长发育过程中为了应对复杂的环境变化, 进化形成了2个不同层次的防御反应: 初级免疫反应和次级免疫反应.宿主细胞膜上的模式识别受体(pattern recognition receptors, PRR)识别病原微生物相关分子模式(pathogen associated molecular patterns, PAMP), 激活PAMP触发的免疫反应(PAMP triggered immunity, PTI) (Monaghan and Zipfel, 2012; Schwessinger and Ronald, 2012; Dangl et al., 2013), 即初级免疫反应.为了应对植物初级免疫反应, 微生物分泌出一些毒性因子(即效应因子)注入到植物细胞内, 抑制PTI, 导致植物感病.在应对效应因子的入侵方面, 植物进化出能够特异识别效应因子的抗性蛋白(resistance protein, R), 激活效应因子触发的免疫反应(effectors triggered immu- nity, ETI), 即为次级免疫反应(Tsuda and Katagiri, 2010; Howden and Huitema, 2012).ETI比PTI更为强烈和迅速, 导致局部的程序性细胞死亡(program- med cell death, PCD), 引起植物超敏反应(hyper- sensitive response, HR), 更为有效地遏制病原菌的进一步扩散(Jones and Dangl, 2006). ...
1 1998
... Flor的基因对基因模型(gene-for-gene model)认为,病原菌引起的植物抗病性由与其相对应的抗病蛋白所介导(Van der Biezen and Jones, 1998).自然界中病原菌种类繁多且不断进化, 因此抗病蛋白对效应蛋白的识别方式也各不相同, 主要包括直接识别和间接识别2种. ...
1 2015
... NB-LRR抗病蛋白作为一类免疫监测因子, 在无效应蛋白激发时, 通过不同结构域间的互作保持抗病蛋白处于失活状态.不同结构域间的互作方式主要有4种.(1) 某些抗病蛋白LRR结构域能够与效应因子直接互作, 该类蛋白NBS结构域与LRR中效应因子结合区域互作, 抑制LRR结构域的激活, 如L6 (L locus 6)抗病蛋白(Bernoux et al., 2011).(2) 抗病蛋白N端区域与NBS结构域共同介导免疫激活, 该类蛋白的LRR结构域与ARC2区域互作, 抑制N端激活域或NBS结构域的功能, 使抗病蛋白处于稳定状态, 如RPS5 (Resistant to Pseudomonas syringae 5)和RPP1 (Resistant to Pseudomonas parasitica 1)等(Rairdan and Moffett, 2006; Qi et al., 2012; Sloot- weg and Goverse, 2013).(3) 抗病蛋白的CC结构域作为其功能结构域, 其NBS结构域与CC结构域结合, 抑制抗病蛋白自激活, 如Rp1 (Resistant to Puccinia sorghi 1)抗病蛋白(Wang et al., 2015).(4) 许多抗病蛋白N端与其它蛋白互作, 共同对效应因子进行监控, LRR结构域与N端区域互作, 使抗病蛋白保持失活状态, 如Prf (Du et al., 2012; Lukasik-shreepaathy et al., 2012). ...
1 2011
... 除内在调控方式外, 温度、湿度和昼夜节律等环境因素也能够影响抗病蛋白的稳定性.遗传筛选实验表明, 许多免疫自激活突变体均表现出温度依赖的表型, 如chs1/2/3、snc1-1、bon1-1和mkp1等.chs1/2/3在16 °C下表现为生长缺陷, 而22°C下生长正常; snc1-1和bon1-1等突变体在22°C下植株矮小、叶片卷曲, 而这种缺陷表型可以被高温恢复或部分恢复(Hua et al., 2001; Yang and Hua, 2004; Yang et al., 2010; Wang et al., 2013; Zbierzak et al., 2013).高温影响抗病蛋白与病原菌间的相互作用, 并且高温能够抑制抗病蛋白从稳定状态向激活状态转变.此外, 高温下植物体内水杨酸(salicylic acid, SA)和脱落酸(absci- sic acid, ABA)等激素水平下降, 增强了植物的抗菌性(Mang et al., 2012).环境湿度和昼夜周期节律变化也能够调控抗病蛋白的稳定性.例如, 高湿度能够抑制抗病蛋白SSI (Suppressor of Salicylic Acid Insensitive)与SLH1 (an allele of RRS1)依赖的免疫激活(Zhou et al., 2004; Noutoshi et al., 2005); 昼夜节律可以影响抗病基因RPP4 (Resistant to Peronospora parasitica 4)的转录(Wang et al., 2011). ...
1 2013
... 除内在调控方式外, 温度、湿度和昼夜节律等环境因素也能够影响抗病蛋白的稳定性.遗传筛选实验表明, 许多免疫自激活突变体均表现出温度依赖的表型, 如chs1/2/3、snc1-1、bon1-1和mkp1等.chs1/2/3在16 °C下表现为生长缺陷, 而22°C下生长正常; snc1-1和bon1-1等突变体在22°C下植株矮小、叶片卷曲, 而这种缺陷表型可以被高温恢复或部分恢复(Hua et al., 2001; Yang and Hua, 2004; Yang et al., 2010; Wang et al., 2013; Zbierzak et al., 2013).高温影响抗病蛋白与病原菌间的相互作用, 并且高温能够抑制抗病蛋白从稳定状态向激活状态转变.此外, 高温下植物体内水杨酸(salicylic acid, SA)和脱落酸(absci- sic acid, ABA)等激素水平下降, 增强了植物的抗菌性(Mang et al., 2012).环境湿度和昼夜周期节律变化也能够调控抗病蛋白的稳定性.例如, 高湿度能够抑制抗病蛋白SSI (Suppressor of Salicylic Acid Insensitive)与SLH1 (an allele of RRS1)依赖的免疫激活(Zhou et al., 2004; Noutoshi et al., 2005); 昼夜节律可以影响抗病基因RPP4 (Resistant to Peronospora parasitica 4)的转录(Wang et al., 2011). ...
1 2005
... 植物免疫响应还与激素信号通路密切相关.遗传分析证明, SA相关突变体sid2 (salicylic acid induction deficient 2)和eds1能够部分恢复抗病蛋白HRT (Hypersensitive Response to TCV)和RPP8 (Recognition of Peronospora parasitic 8)激活后造成的免疫激活表型(Chandrashekara et al., 2004; Wiermer et al., 2005), 表明抗病蛋白的激活能够引起SA的积累.SA与细胞质内多聚的NPR1 (Non-expresser of PR Genes 1)结合后, NPR1解聚形成单体进入细胞核, 结合到转录因子的TGA区域, 激活PR1 (Pathogenesis Related Gene 1)以及植物防御反应相关转录因子WRKY的转录(图2); 随后, NPR1同源蛋白NPR3和NPR4作为接头蛋白连接NPR1与Cullin3 E3连接酶, NPR1被降解(Fu et al., 2012; Wu et al., 2012; Paj- erowska-Mukhtar et al., 2013).此外, 越来越多的研究表明, 多种激素信号均参与调控植物免疫响应, 但具体的调控机制还不清楚(De Bruyne et al., 2014). ...
1 2017
... 抗病蛋白的降解受到泛素化蛋白酶体介导的降解途径调控(图2C).研究表明, F-box蛋白cpr1 (constitutive expresser of PR genes 1)功能缺失突变体中抗病蛋白SNC1和RPS2 (Resistance to Pseu- domonas syringae 2)积累, 表明SCFCPR1E3复合体调控抗病蛋白SNC1和RPS2的降解(Cheng et al., 2011).snc1增强子筛选实验证明, E4连接酶MUSE3 (Mutants snc1 Enhancing 3)、TRAR蛋白MUS13和MUSE14能够与SCFCPR1E3复合体互作, 共同调控抗病蛋白SNC1和RPS2的降解(Huang et al., 2014, 2016).MUSE7编码1个功能未知的激酶底物蛋白, 负调控SNC1、RPS2和RPM1蛋白的积累(Johnson et al., 2016).与HSP90复合体作为分子伴侣调控抗病蛋白的装配与成熟不同, HSP90和SGT1也能够与SKP1 (S Phase Kinase Associated Protein 1)-CUL- LIN-F-box-E3泛素复合体相互作用, 调控抗病蛋白的降解(Willhoft et al., 2017).遗传分析表明, hsp90功能获得突变体中抗病蛋白SNC1积累(Huang et al., 2014).sgt1b缺失突变能够抑制rar1突变体中RPS5的积累(Holt et al., 2005).生化实验证明, HSP90和SGT1b能够与SKP1和CULLIN1互作, 促进蛋白降解; RAR1也能够结合COP9 (Constitutive Photo- morpho-genesis Complex 9)蛋白, 抑制SCF复合体的活性, 进而调控抗病蛋白的降解(Azevedo et al., 2002).另外, SGT1a和SGT1b能够与SRFR1 (Suppressor of rps4-RLD)蛋白互作, 负调控抗病蛋白SNC1、RPS2和RPS4的积累(Li et al., 2010a).E1泛素连接酶UBA1 (Ubiquitin Activating 1)与26S蛋白酶体亚基RPN1a (Proteasome Regulatory Subunint S2 1A)、RPN8a (RP Non-ATPase Subunit 8A)和RPT2a (Regulatory Particle AAA-ATPase 2A)参与调控抗病蛋白的降解(Goritschnig et al., 2007; Yao et al., 2012). ...
1 2014
... TNL类抗病蛋白N端包含TIR结构域, 其与果蝇(Droso- phila) Toll蛋白或白介素1受体N端序列类似, 在动植物中高度保守.晶体结构分析显示, TIR结构域通常由核心的5个β-片层和外围的5个α-螺旋构成.TIR结构域能够识别效应蛋白.例如, 烟草(Nicotiana tabacum) N蛋白的TIR结构域能够特异地识别花叶病毒分泌的效应蛋白p50 (Swiderski et al., 2009).此外, 该结构域也能够介导蛋白之间的相互作用.例如, RRS1 (Resistant to Ralstonia solanacearum 1)和RPS4 (Resistant to Pseudomonas syringae 4)蛋白通过TIR结构域中保守的丝氨酸-组氨酸(ser-his, SH)基序互作, 形成结合界面, 共同介导植物对病原菌的抗性(Williams et al., 2014).CNL类抗病蛋白N端包含CC结构域, 即不规则卷曲螺旋结构.通过对CC结构域蛋白结构进行解析, 发现该结构域由7个氨基酸重复序列组成, 包含2个或多个α-螺旋, 构成一个超螺旋结构.CC结构域缺乏保守性, 其中唯一的保守基序为EDVID.但并非所有的CNL蛋白CC结构域都存在该基序.CC结构域在蛋白互作中发挥重要功能.例如, CNL抗性蛋白Rx的CC结构域能够与细胞内辅因子RanGAP2直接互作, 共同监控马铃薯(Solanum tuberosum) X病毒.晶体结构揭示, 该蛋白CC结构域具有保守的EDVID基序, 其第90位色氨酸能够与Ran- GAP2的第89位丙氨酸通过疏水作用相结合.而Rx蛋白CC结构域与RanGAP2相结合的2个α-螺旋为调控Rx蛋白入核的区域, RanGAP2通过与该区域结合阻止Rx进入细胞核, 增强植物抗病能力(Tameling and Baulcombe, 2007; Hao et al., 2013).此外, 通过解析MLA10 (Mildew Resistance Locus A10)抗病蛋白CC结构域的晶体结构, 发现该蛋白CC结构域由2个长α-螺旋单体组成, 2条链相距较远, 且内侧多为疏水性氨基酸, 2个MLA10蛋白通过CC结构域形成同源二聚体, 维持整个蛋白的稳定性(Maekawa et al., 2011). ...
... RNA转录后加工对于抗病基因的表达调控十分重要.不同生态型中抗病基因的剪接方式不同, 如SNC1内含子的剪接方式在WS生态型与Col生态型中不同(Yang and Hua, 2004).抗病基因的可变剪接受到以MOS4为核心的剪接复合体MAC (MOS4 Associated Complex)的调控, 该复合体包括MOS4、CDC5 (Cell Division Cycle 5)、PRL1 (Pleiotropic Regulatory Locus 1)、MAC3以及MAC5 (图2A).MAC复合体成员功能缺失后导致抗病蛋白剪接方式改变, 而此变化将影响后续的蛋白翻译过程, 导致形成截短的抗病蛋白, 改变其原有的功能, 最终影响免疫响应的激活.此外, mRNA剪接因子MOS12、RNA结合蛋白MOS2以及MOS14也能够影响抗病基因的剪接方式, 进而参与调控植物免疫响应(Xu et al., 2011, 2012). ...
1 2010
... 除内在调控方式外, 温度、湿度和昼夜节律等环境因素也能够影响抗病蛋白的稳定性.遗传筛选实验表明, 许多免疫自激活突变体均表现出温度依赖的表型, 如chs1/2/3、snc1-1、bon1-1和mkp1等.chs1/2/3在16 °C下表现为生长缺陷, 而22°C下生长正常; snc1-1和bon1-1等突变体在22°C下植株矮小、叶片卷曲, 而这种缺陷表型可以被高温恢复或部分恢复(Hua et al., 2001; Yang and Hua, 2004; Yang et al., 2010; Wang et al., 2013; Zbierzak et al., 2013).高温影响抗病蛋白与病原菌间的相互作用, 并且高温能够抑制抗病蛋白从稳定状态向激活状态转变.此外, 高温下植物体内水杨酸(salicylic acid, SA)和脱落酸(absci- sic acid, ABA)等激素水平下降, 增强了植物的抗菌性(Mang et al., 2012).环境湿度和昼夜周期节律变化也能够调控抗病蛋白的稳定性.例如, 高湿度能够抑制抗病蛋白SSI (Suppressor of Salicylic Acid Insensitive)与SLH1 (an allele of RRS1)依赖的免疫激活(Zhou et al., 2004; Noutoshi et al., 2005); 昼夜节律可以影响抗病基因RPP4 (Resistant to Peronospora parasitica 4)的转录(Wang et al., 2011). ...
2 2004
... RNA转录后加工对于抗病基因的表达调控十分重要.不同生态型中抗病基因的剪接方式不同, 如SNC1内含子的剪接方式在WS生态型与Col生态型中不同(Yang and Hua, 2004).抗病基因的可变剪接受到以MOS4为核心的剪接复合体MAC (MOS4 Associated Complex)的调控, 该复合体包括MOS4、CDC5 (Cell Division Cycle 5)、PRL1 (Pleiotropic Regulatory Locus 1)、MAC3以及MAC5 (图2A).MAC复合体成员功能缺失后导致抗病蛋白剪接方式改变, 而此变化将影响后续的蛋白翻译过程, 导致形成截短的抗病蛋白, 改变其原有的功能, 最终影响免疫响应的激活.此外, mRNA剪接因子MOS12、RNA结合蛋白MOS2以及MOS14也能够影响抗病基因的剪接方式, 进而参与调控植物免疫响应(Xu et al., 2011, 2012). ... ... 除内在调控方式外, 温度、湿度和昼夜节律等环境因素也能够影响抗病蛋白的稳定性.遗传筛选实验表明, 许多免疫自激活突变体均表现出温度依赖的表型, 如chs1/2/3、snc1-1、bon1-1和mkp1等.chs1/2/3在16 °C下表现为生长缺陷, 而22°C下生长正常; snc1-1和bon1-1等突变体在22°C下植株矮小、叶片卷曲, 而这种缺陷表型可以被高温恢复或部分恢复(Hua et al., 2001; Yang and Hua, 2004; Yang et al., 2010; Wang et al., 2013; Zbierzak et al., 2013).高温影响抗病蛋白与病原菌间的相互作用, 并且高温能够抑制抗病蛋白从稳定状态向激活状态转变.此外, 高温下植物体内水杨酸(salicylic acid, SA)和脱落酸(absci- sic acid, ABA)等激素水平下降, 增强了植物的抗菌性(Mang et al., 2012).环境湿度和昼夜周期节律变化也能够调控抗病蛋白的稳定性.例如, 高湿度能够抑制抗病蛋白SSI (Suppressor of Salicylic Acid Insensitive)与SLH1 (an allele of RRS1)依赖的免疫激活(Zhou et al., 2004; Noutoshi et al., 2005); 昼夜节律可以影响抗病基因RPP4 (Resistant to Peronospora parasitica 4)的转录(Wang et al., 2011). ...
1 2012
... 抗病蛋白的降解受到泛素化蛋白酶体介导的降解途径调控(图2C).研究表明, F-box蛋白cpr1 (constitutive expresser of PR genes 1)功能缺失突变体中抗病蛋白SNC1和RPS2 (Resistance to Pseu- domonas syringae 2)积累, 表明SCFCPR1E3复合体调控抗病蛋白SNC1和RPS2的降解(Cheng et al., 2011).snc1增强子筛选实验证明, E4连接酶MUSE3 (Mutants snc1 Enhancing 3)、TRAR蛋白MUS13和MUSE14能够与SCFCPR1E3复合体互作, 共同调控抗病蛋白SNC1和RPS2的降解(Huang et al., 2014, 2016).MUSE7编码1个功能未知的激酶底物蛋白, 负调控SNC1、RPS2和RPM1蛋白的积累(Johnson et al., 2016).与HSP90复合体作为分子伴侣调控抗病蛋白的装配与成熟不同, HSP90和SGT1也能够与SKP1 (S Phase Kinase Associated Protein 1)-CUL- LIN-F-box-E3泛素复合体相互作用, 调控抗病蛋白的降解(Willhoft et al., 2017).遗传分析表明, hsp90功能获得突变体中抗病蛋白SNC1积累(Huang et al., 2014).sgt1b缺失突变能够抑制rar1突变体中RPS5的积累(Holt et al., 2005).生化实验证明, HSP90和SGT1b能够与SKP1和CULLIN1互作, 促进蛋白降解; RAR1也能够结合COP9 (Constitutive Photo- morpho-genesis Complex 9)蛋白, 抑制SCF复合体的活性, 进而调控抗病蛋白的降解(Azevedo et al., 2002).另外, SGT1a和SGT1b能够与SRFR1 (Suppressor of rps4-RLD)蛋白互作, 负调控抗病蛋白SNC1、RPS2和RPS4的积累(Li et al., 2010a).E1泛素连接酶UBA1 (Ubiquitin Activating 1)与26S蛋白酶体亚基RPN1a (Proteasome Regulatory Subunint S2 1A)、RPN8a (RP Non-ATPase Subunit 8A)和RPT2a (Regulatory Particle AAA-ATPase 2A)参与调控抗病蛋白的降解(Goritschnig et al., 2007; Yao et al., 2012). ...
1 2013
... 除内在调控方式外, 温度、湿度和昼夜节律等环境因素也能够影响抗病蛋白的稳定性.遗传筛选实验表明, 许多免疫自激活突变体均表现出温度依赖的表型, 如chs1/2/3、snc1-1、bon1-1和mkp1等.chs1/2/3在16 °C下表现为生长缺陷, 而22°C下生长正常; snc1-1和bon1-1等突变体在22°C下植株矮小、叶片卷曲, 而这种缺陷表型可以被高温恢复或部分恢复(Hua et al., 2001; Yang and Hua, 2004; Yang et al., 2010; Wang et al., 2013; Zbierzak et al., 2013).高温影响抗病蛋白与病原菌间的相互作用, 并且高温能够抑制抗病蛋白从稳定状态向激活状态转变.此外, 高温下植物体内水杨酸(salicylic acid, SA)和脱落酸(absci- sic acid, ABA)等激素水平下降, 增强了植物的抗菌性(Mang et al., 2012).环境湿度和昼夜周期节律变化也能够调控抗病蛋白的稳定性.例如, 高湿度能够抑制抗病蛋白SSI (Suppressor of Salicylic Acid Insensitive)与SLH1 (an allele of RRS1)依赖的免疫激活(Zhou et al., 2004; Noutoshi et al., 2005); 昼夜节律可以影响抗病基因RPP4 (Resistant to Peronospora parasitica 4)的转录(Wang et al., 2011). ...
1 2005
... 在植物免疫响应过程中, 抗病蛋白的激活导致下游免疫信号被激活, 从而调控植物对病原菌的抗性.研究表明许多抗病蛋白能够进入细胞核激活免疫响应(图3).对SNC1蛋白的研究表明, 当其进入细胞核后, SNC1的TIR结构域与转录抑制因子TPR1 (Topless Related 1)结合, 抑制免疫负调控因子DND1 (Defense No Death 1)和DND2的转录(Zhu et al., 2010); SNC1还能够与bHLH84 (Helix Loop Helix 84)转录因子相互作用, 激活抗病相关基因的转录(Xu et al., 2014).通过对snc1抑制子的筛选发现, MOS3和MOS7能够调控SNC1在细胞核中的积累.MOS3是一个核孔蛋白, MOS7为果蝇与人类(Homo sapiens)核孔蛋白Nup88的同源蛋白.在mos3和mos7缺失突变体中, 定位于细胞核的SNC1减少, 不能激活转录因子的表达, 从而抑制SNC1依赖的免疫响应.该研究结果表明, 核孔复合体组分MOS3和MOS7能够调控抗病蛋白的细胞核定位, 影响SNC1介导的下游转录信号激活(Zhang and Li, 2005; Cheng et al., 2009), 但其具体调控机制还不清楚.而最新研究表明, 在抗病蛋白RPS2激活过程中, 与核孔复合体结合的跨膜核孔蛋白CPR5 (Constitutive Expresser of PR Genes 5)构象改变, 降低其与CKIs (Cyclin-dep- endent Kinase Inhibitors)的亲和性, 触发Rb (Retinoblastoma)和E2F介导的细胞周期途径, 调控抗病基因的表达, 引发细胞程序性死亡, 从而激活ETI免疫响应(Gu et al., 2016).除此以外, 某些含有DNA结合结构域的抗病蛋白进入细胞核后, 能够直接与转录因子结合, 激活抗病相关基因的转录.例如, 抗病蛋白MLA10入核后, 其N端结构域与WRKY1/2转录因子相互作用, 解除其对MYB6 (MYB Domain Protein 6)的抑制, 启动植物抗病反应(Chang et al., 2013); 而Pb1 (Panicle blast 1)与WRKY45转录因子结合后, 抑制WRKY45被蛋白酶体降解, 激活SA依赖的抗病信号通路(Inoue et al., 2013). ...
1 2017
... 除上述识别模式外, 还存在一些新的抗病蛋白激活模式.植物初级免疫PTI激活后, MAPK (Mitogen Activated Protein Kinase)信号级联通路被激活, 而微生物分泌的效应蛋白能够结合到MAPK通路相关蛋白, 抑制PTI免疫激活.遗传筛选发现, summ1 (suppressor of mkk1mkk2 1)和summ2能够抑制mpk4 (map kinase 4)突变体中免疫激活的表型, SUMM1为蛋白激酶MEKK2, 而SUMM2为抗病蛋白.该结果表明, 抗病蛋白能够监控MEKK1-MKK1/MKK2-MPK4信号级联通路.生化实验证明, MPK4能够磷酸化MEKK2, 但不能与SUMM2直接互作.遗传分析表明, MEKK2位于SUMM2上游, 但MEKK2是如何激活SUMM2的还不清楚(Kong et al., 2012; Zhang et al., 2012).随后的研究表明, MPK4能够磷酸化PAT1 (Arabidopsis Homolog of Yeast PAT1), 抑制SUMM2激活(Roux et al., 2015).最新研究表明, 蛋白激酶CRCK3 (Cal- modulin Binding Receptor like Cytoplasmic Kinase 3)也能被MPK4磷酸化, 共同调控SUMM2依赖的免疫响应(Zhang et al., 2017).无效应蛋白时, MPK4磷酸化底物蛋白, 使得SUMM2处于稳定状态; 而效应蛋白HopAI1与MPK4结合后, 抑制MPK4激酶活性, 使其不能将底物蛋白磷酸化, 导致SUMM2被激活, 触发植物免疫响应.此外, SUMM2还能够监控MPK3与MPK6引起的免疫响应(Genot et al., 2017).除上述激酶以外, 磷酸酶mkp1 (map kinase phosphat- ase1)和ptp1 (protein tyrosine phosphatase1)突变体也表现出SNC1依赖的免疫自激活表型.生化分析结果表明, MKP1能够与MPK3、MPK4和MPK6相互作用, 去磷酸化MPK6, 抑制其激酶活性, 从而拮抗调控抗病蛋白依赖的免疫激活(Schweighofer et al., 2007; Bartels et al., 2009; Anderson et al., 2011).蛋白磷酸酶AP2C1 (Arabidopsis Phosphatase 2C 1)也能够与MPK4和MPK6互作, 通过去磷酸化修饰抑制MPK4和MPK6激酶活性, 调控植物免疫响应.除MAPK依赖的途径外, 钙离子依赖的蛋白激酶CPK1 (Calcium-dependent Protein Kinase 1)、CPK2、CP- K4、CPK5、CPK6和CPK11也能够调控抗病蛋白介导的免疫响应.最新研究表明, CPK5能够与抗病蛋白TN2 (TIR-NBS2)的NBS结构域相互作用, 调控TN2依赖的免疫响应(Liu et al., 2017).综上所述, 抗病蛋白通过不同的识别模式对效应蛋白进行监控, 并且该过程多种多样. ...
1 2012
... 除上述识别模式外, 还存在一些新的抗病蛋白激活模式.植物初级免疫PTI激活后, MAPK (Mitogen Activated Protein Kinase)信号级联通路被激活, 而微生物分泌的效应蛋白能够结合到MAPK通路相关蛋白, 抑制PTI免疫激活.遗传筛选发现, summ1 (suppressor of mkk1mkk2 1)和summ2能够抑制mpk4 (map kinase 4)突变体中免疫激活的表型, SUMM1为蛋白激酶MEKK2, 而SUMM2为抗病蛋白.该结果表明, 抗病蛋白能够监控MEKK1-MKK1/MKK2-MPK4信号级联通路.生化实验证明, MPK4能够磷酸化MEKK2, 但不能与SUMM2直接互作.遗传分析表明, MEKK2位于SUMM2上游, 但MEKK2是如何激活SUMM2的还不清楚(Kong et al., 2012; Zhang et al., 2012).随后的研究表明, MPK4能够磷酸化PAT1 (Arabidopsis Homolog of Yeast PAT1), 抑制SUMM2激活(Roux et al., 2015).最新研究表明, 蛋白激酶CRCK3 (Cal- modulin Binding Receptor like Cytoplasmic Kinase 3)也能被MPK4磷酸化, 共同调控SUMM2依赖的免疫响应(Zhang et al., 2017).无效应蛋白时, MPK4磷酸化底物蛋白, 使得SUMM2处于稳定状态; 而效应蛋白HopAI1与MPK4结合后, 抑制MPK4激酶活性, 使其不能将底物蛋白磷酸化, 导致SUMM2被激活, 触发植物免疫响应.此外, SUMM2还能够监控MPK3与MPK6引起的免疫响应(Genot et al., 2017).除上述激酶以外, 磷酸酶mkp1 (map kinase phosphat- ase1)和ptp1 (protein tyrosine phosphatase1)突变体也表现出SNC1依赖的免疫自激活表型.生化分析结果表明, MKP1能够与MPK3、MPK4和MPK6相互作用, 去磷酸化MPK6, 抑制其激酶活性, 从而拮抗调控抗病蛋白依赖的免疫激活(Schweighofer et al., 2007; Bartels et al., 2009; Anderson et al., 2011).蛋白磷酸酶AP2C1 (Arabidopsis Phosphatase 2C 1)也能够与MPK4和MPK6互作, 通过去磷酸化修饰抑制MPK4和MPK6激酶活性, 调控植物免疫响应.除MAPK依赖的途径外, 钙离子依赖的蛋白激酶CPK1 (Calcium-dependent Protein Kinase 1)、CPK2、CP- K4、CPK5、CPK6和CPK11也能够调控抗病蛋白介导的免疫响应.最新研究表明, CPK5能够与抗病蛋白TN2 (TIR-NBS2)的NBS结构域相互作用, 调控TN2依赖的免疫响应(Liu et al., 2017).综上所述, 抗病蛋白通过不同的识别模式对效应蛋白进行监控, 并且该过程多种多样. ...
1 2004
... 除内在调控方式外, 温度、湿度和昼夜节律等环境因素也能够影响抗病蛋白的稳定性.遗传筛选实验表明, 许多免疫自激活突变体均表现出温度依赖的表型, 如chs1/2/3、snc1-1、bon1-1和mkp1等.chs1/2/3在16 °C下表现为生长缺陷, 而22°C下生长正常; snc1-1和bon1-1等突变体在22°C下植株矮小、叶片卷曲, 而这种缺陷表型可以被高温恢复或部分恢复(Hua et al., 2001; Yang and Hua, 2004; Yang et al., 2010; Wang et al., 2013; Zbierzak et al., 2013).高温影响抗病蛋白与病原菌间的相互作用, 并且高温能够抑制抗病蛋白从稳定状态向激活状态转变.此外, 高温下植物体内水杨酸(salicylic acid, SA)和脱落酸(absci- sic acid, ABA)等激素水平下降, 增强了植物的抗菌性(Mang et al., 2012).环境湿度和昼夜周期节律变化也能够调控抗病蛋白的稳定性.例如, 高湿度能够抑制抗病蛋白SSI (Suppressor of Salicylic Acid Insensitive)与SLH1 (an allele of RRS1)依赖的免疫激活(Zhou et al., 2004; Noutoshi et al., 2005); 昼夜节律可以影响抗病基因RPP4 (Resistant to Peronospora parasitica 4)的转录(Wang et al., 2011). ...
1 2010
... 在植物免疫响应过程中, 抗病蛋白的激活导致下游免疫信号被激活, 从而调控植物对病原菌的抗性.研究表明许多抗病蛋白能够进入细胞核激活免疫响应(图3).对SNC1蛋白的研究表明, 当其进入细胞核后, SNC1的TIR结构域与转录抑制因子TPR1 (Topless Related 1)结合, 抑制免疫负调控因子DND1 (Defense No Death 1)和DND2的转录(Zhu et al., 2010); SNC1还能够与bHLH84 (Helix Loop Helix 84)转录因子相互作用, 激活抗病相关基因的转录(Xu et al., 2014).通过对snc1抑制子的筛选发现, MOS3和MOS7能够调控SNC1在细胞核中的积累.MOS3是一个核孔蛋白, MOS7为果蝇与人类(Homo sapiens)核孔蛋白Nup88的同源蛋白.在mos3和mos7缺失突变体中, 定位于细胞核的SNC1减少, 不能激活转录因子的表达, 从而抑制SNC1依赖的免疫响应.该研究结果表明, 核孔复合体组分MOS3和MOS7能够调控抗病蛋白的细胞核定位, 影响SNC1介导的下游转录信号激活(Zhang and Li, 2005; Cheng et al., 2009), 但其具体调控机制还不清楚.而最新研究表明, 在抗病蛋白RPS2激活过程中, 与核孔复合体结合的跨膜核孔蛋白CPR5 (Constitutive Expresser of PR Genes 5)构象改变, 降低其与CKIs (Cyclin-dep- endent Kinase Inhibitors)的亲和性, 触发Rb (Retinoblastoma)和E2F介导的细胞周期途径, 调控抗病基因的表达, 引发细胞程序性死亡, 从而激活ETI免疫响应(Gu et al., 2016).除此以外, 某些含有DNA结合结构域的抗病蛋白进入细胞核后, 能够直接与转录因子结合, 激活抗病相关基因的转录.例如, 抗病蛋白MLA10入核后, 其N端结构域与WRKY1/2转录因子相互作用, 解除其对MYB6 (MYB Domain Protein 6)的抑制, 启动植物抗病反应(Chang et al., 2013); 而Pb1 (Panicle blast 1)与WRKY45转录因子结合后, 抑制WRKY45被蛋白酶体降解, 激活SA依赖的抗病信号通路(Inoue et al., 2013). ...
1 2014
... 植物体内大部分的抗病基因转录水平较低, 在各种组织中广泛表达.有研究表明, 病原菌入侵后植物基因组GC和CHH位点甲基化水平降低, 抗病基因表达量升高, 表明抗病基因的转录受DNA甲基化调控.例如, H3K4甲基转移酶ATXR7 (Arabidopsis Trithorax Related 7)和H2B单泛素连接酶能够结合到抗病基因的启动子区域, 对其进行甲基化修饰, 调控抗病基因SNC1 (Suppressor of npr1 Constitutive1)和RPP4 (Resistance to Peromospora parasitica 4)的表达(Xia et al., 2013; Zou et al., 2014); MOS1 (Modifier of SNC 1)蛋白N端含有BAT2 (HLA-B Associated Transcript 2)结构域, 也能够与SNC1启动子序列结合, 对其进行甲基化修饰, 正调控SNC1的表达(Li et al., 2010b, 2011).此外, miRNAs能够与抗病基因的保守结构域结合, 导致抗病基因沉默(Fei et al., 2013).例如, miRNAs nta-miR6019 (22nt)和nta- miR6020 (21nt)能够调控烟草抗病基因N的转录.研究发现, N与nta-miR6019和nta-miR6020共表达后导致抗病蛋白N介导的TMV抗性减弱, 表明miRNA在抗病基因转录调节中具有功能性作用(Li et al., 2012), 但其具体的调控机制还不是很清楚. ...

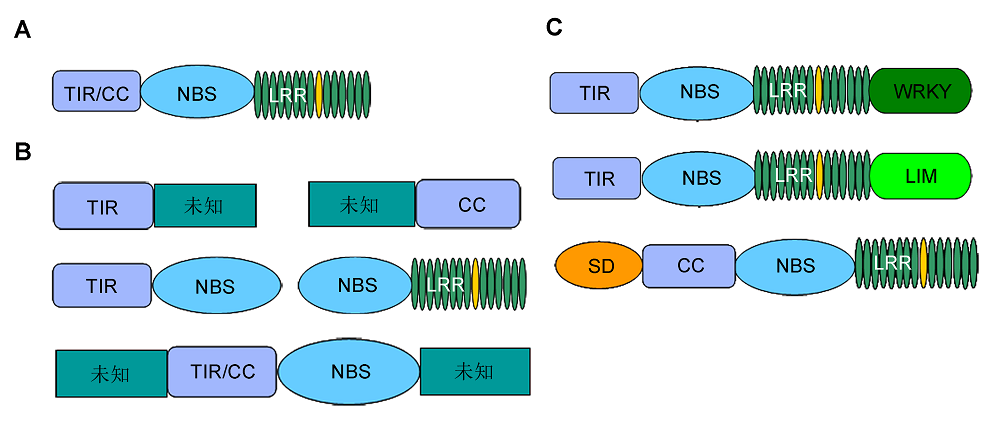
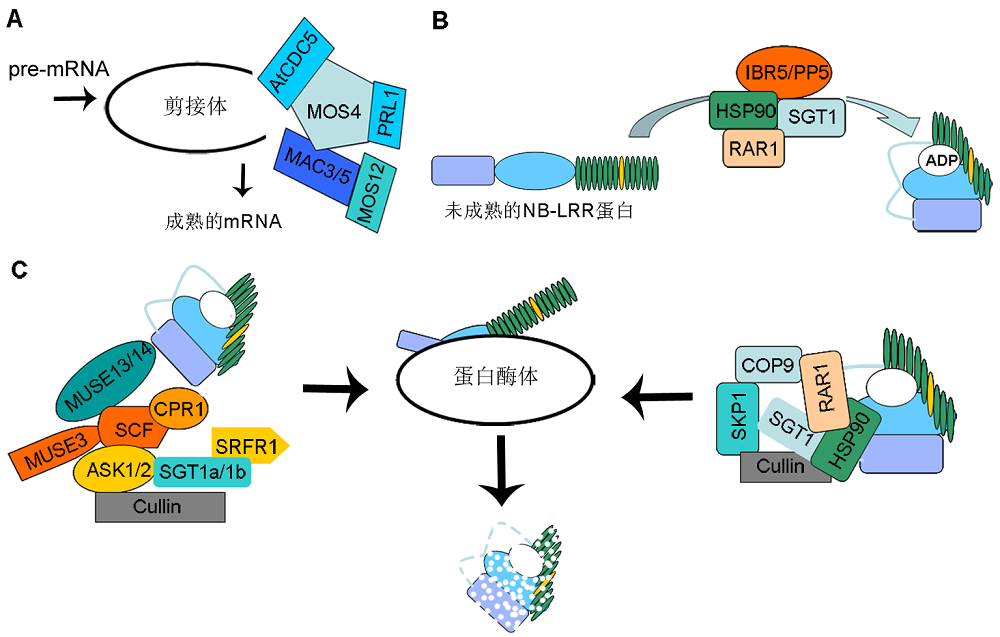
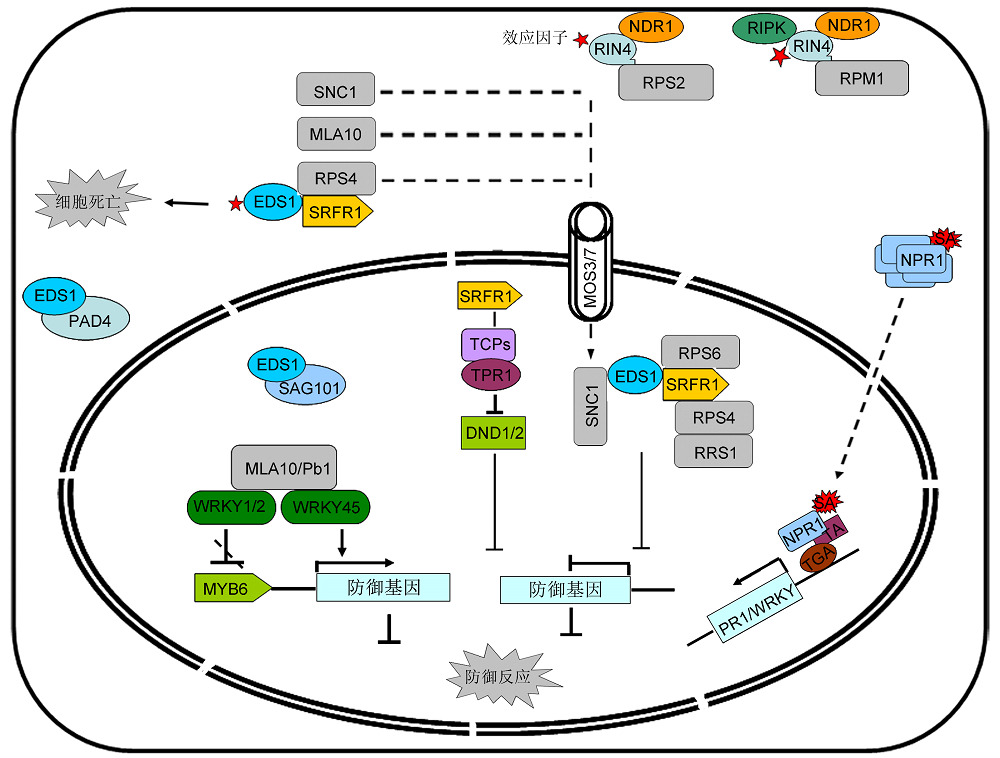

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}