Abstract: Plant root tips can sense the moisture gradient in soil and grow toward the higher water potential region. This unique response is called the hydrotropic response or hydrotropism. Hydrotropism plays a key role for plants to efficiently obtain water from soil. The root hydrotropic response has become one of the hot topics in plant biology. However, the detailed molecular mechanisms controlling the root hydrotropic response are poorly understood. Previous studies de- monstrated that MIZ1 and GNOM can positively regulate the hydrotropic response. Several phytohormones, light, ROS and Ca2+ were also thought to mediate the root hydrotropic response, but their detailed molecular mechanisms are not yet elucidated. This review highlights the research history and factors of hydrotropic response and identification and characterization of key regulators of hydrotropic response, to give a more comprehensive understanding of research progress in the plant hydrotropic response. We provide perspectives on possible future research directions.
1 向水性的研究历史及突变体筛选早期的植物生理学研究中, Porta (1658)和Knight (1811)等科学家通过大量的实验证明, 植物根系会随着生长环境中的水势梯度而改变其生长方向, 由此产生了对植物根向水性的研究。此外, 达尔文所著的《The Power of Movement in Plants》一书中也有关于向水性的描述(Darwin and Darwin, 1881)。德国植物学家Sachs (1887)将由网格布包裹的潮湿木屑斜向悬挂, 让豌豆(Pisum sativum)种子在木屑表面萌发, 当豌豆的根因向地性生长至网格布下边缘时, 会沿网格布表面弯曲生长而不会进入干燥的空气中, 据此他认为植物的根存在向水性生长(图1A) (Sachs, 1887)。另外, 研究还发现, 土壤中的水分会随着土壤深度、季节和降水等因素而分布不均, 产生水势梯度, 而且这种水势梯度会诱导根系向水性生长(Garwood and Williams,1967; Hsiao and Xu, 2000; Steudle, 2000)。然而, 由于其它植物向性的干扰, 向水性反应一直以来未被科学家认可。直到1985年, Jaffe及其同事利用豌豆突变体“Ageotropum”为实验材料才首次强有力地证明了植物根向水性不依赖于向地性而独立存在(Jaffe et al., 1985)。此豌豆突变体的根完全失去了向地性和避光性反应, 因此其部分根会向上生长出土壤(Blixt et al.,1958; Schöldéen and Burst- röm, 1960; Ekelund and Hemberg, 1966; Wilson, 1967)。在一个湿度可控的玻璃盒子中培养该突变体植物, 当盒子中空气的相对湿度(relative humidity of the air, RH)达到95%-98%时, Ageotropum的根生长出土壤后不会弯曲, 而是向着湿润的空气中继续生长(图1B); 当空气相对湿度调整为85%-90%时, 原来向上生长的Ageotropum根会迅速向下弯曲生长直至进入比空气湿润的土壤里。这一现象说明, Ageotro- pum尽管缺失了向地性和避光性, 但其根仍然能感知环境中的水势梯度, 并具有正常的向水性生长反应。除了利用失去了向地性的突变体来研究植物根的向水性外, 科学家们还用竖直旋转的转盘来固定植物材料(Hoson et al., 1997; Kraft et al., 2000)进行研究, 发现竖直方向低速旋转培养植物的根的向地性被有效抵消。用此方法对豌豆以及黄瓜(Cucumis sativus)进行了研究, 发现它们的向水性表型更加明显(Takah- ashi et al., 1996, 1999)。Takahashi及其同事发现, 在航天器失重条件下, 黄瓜的侧根也表现出更强烈地向水性反应, 而在正常培养的实验中侧根的向水性反应不易被观察到, 说明去除向地性影响后, 植物根对水势梯度的反应更明显(Takahashi et al.,1999)。 图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-2-154/img_1.png<b>图1</b> 植物根向水性反应实验筛选体系示意图<br/>(A) Sachs发现植物的根具有向水性所用的实验方法(<xref ref-type="bibr" rid="b62-1674-3466-53-2-154">Sachs, 1887</xref>); (B) Jaffe及其同事证明植物根向水性存在所用的实验方法(<xref ref-type="bibr" rid="b27-1674-3466-53-2-154">Jaffe et al., 1985</xref>); (C) 墨西哥Cassab实验室的根向水性筛选系统(<xref ref-type="bibr" rid="b15-1674-3466-53-2-154">Eapen et al., 2003</xref>; <xref ref-type="bibr" rid="b63-1674-3466-53-2-154">Saucedo et al., 2012</xref>); (D), (E) 日本东北大学Takahashi实验室的根向水性筛选系统(<xref ref-type="bibr" rid="b72-1674-3466-53-2-154">Takahashi et al., 2002</xref>; <xref ref-type="bibr" rid="b32-1674-3466-53-2-154">Kobayashi et al., 2007</xref>)。NM: 正常培养基; WSM: 水分胁迫培养基<br/><b>Figure 1</b> Methods used for demonstrating root hydrotropism and screening for mutants altering root hydrotropic response<br/>(A) The method Sachs used to demonstrate plant root hydrotropic response (<xref ref-type="bibr" rid="b62-1674-3466-53-2-154">Sachs, 1887</xref>); (B) Experiment Jaffe et al. (1985) carried out to prove the existence of the hydrotropic response using <i>Ageotropum</i> mutant; (C) Screening system for identifying mutants with altered hydrotropic response in Cassab’s lab (<xref ref-type="bibr" rid="b15-1674-3466-53-2-154">Eapen et al., 2003</xref>; <xref ref-type="bibr" rid="b63-1674-3466-53-2-154">Saucedo et al., 2012</xref>); (D), (E) Screening system used in Takahashi’s laboratory for mutants with altered hydrotropic response (<xref ref-type="bibr" rid="b72-1674-3466-53-2-154">Takahashi et al., 2002</xref>; <xref ref-type="bibr" rid="b32-1674-3466-53-2-154">Kobayashi et al., 2007</xref>). NM: Normal nutrient medium; WSM: Water stress medium Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-2-154/img_1.png<b>图1</b> 植物根向水性反应实验筛选体系示意图<br/>(A) Sachs发现植物的根具有向水性所用的实验方法(<xref ref-type="bibr" rid="b62-1674-3466-53-2-154">Sachs, 1887</xref>); (B) Jaffe及其同事证明植物根向水性存在所用的实验方法(<xref ref-type="bibr" rid="b27-1674-3466-53-2-154">Jaffe et al., 1985</xref>); (C) 墨西哥Cassab实验室的根向水性筛选系统(<xref ref-type="bibr" rid="b15-1674-3466-53-2-154">Eapen et al., 2003</xref>; <xref ref-type="bibr" rid="b63-1674-3466-53-2-154">Saucedo et al., 2012</xref>); (D), (E) 日本东北大学Takahashi实验室的根向水性筛选系统(<xref ref-type="bibr" rid="b72-1674-3466-53-2-154">Takahashi et al., 2002</xref>; <xref ref-type="bibr" rid="b32-1674-3466-53-2-154">Kobayashi et al., 2007</xref>)。NM: 正常培养基; WSM: 水分胁迫培养基<br/><b>Figure 1</b> Methods used for demonstrating root hydrotropism and screening for mutants altering root hydrotropic response<br/>(A) The method Sachs used to demonstrate plant root hydrotropic response (<xref ref-type="bibr" rid="b62-1674-3466-53-2-154">Sachs, 1887</xref>); (B) Experiment Jaffe et al. (1985) carried out to prove the existence of the hydrotropic response using <i>Ageotropum</i> mutant; (C) Screening system for identifying mutants with altered hydrotropic response in Cassab’s lab (<xref ref-type="bibr" rid="b15-1674-3466-53-2-154">Eapen et al., 2003</xref>; <xref ref-type="bibr" rid="b63-1674-3466-53-2-154">Saucedo et al., 2012</xref>); (D), (E) Screening system used in Takahashi’s laboratory for mutants with altered hydrotropic response (<xref ref-type="bibr" rid="b72-1674-3466-53-2-154">Takahashi et al., 2002</xref>; <xref ref-type="bibr" rid="b32-1674-3466-53-2-154">Kobayashi et al., 2007</xref>). NM: Normal nutrient medium; WSM: Water stress medium
图1 植物根向水性反应实验筛选体系示意图 (A) Sachs发现植物的根具有向水性所用的实验方法(Sachs, 1887); (B) Jaffe及其同事证明植物根向水性存在所用的实验方法(Jaffe et al., 1985); (C) 墨西哥Cassab实验室的根向水性筛选系统(Eapen et al., 2003; Saucedo et al., 2012); (D), (E) 日本东北大学Takahashi实验室的根向水性筛选系统(Takahashi et al., 2002; Kobayashi et al., 2007)。NM: 正常培养基; WSM: 水分胁迫培养基 Figure 1 Methods used for demonstrating root hydrotropism and screening for mutants altering root hydrotropic response (A) The method Sachs used to demonstrate plant root hydrotropic response (Sachs, 1887); (B) Experiment Jaffe et al. (1985) carried out to prove the existence of the hydrotropic response using Ageotropum mutant; (C) Screening system for identifying mutants with altered hydrotropic response in Cassab’s lab (Eapen et al., 2003; Saucedo et al., 2012); (D), (E) Screening system used in Takahashi’s laboratory for mutants with altered hydrotropic response (Takahashi et al., 2002; Kobayashi et al., 2007). NM: Normal nutrient medium; WSM: Water stress medium
植物根柱细胞中的淀粉粒在根的重力感知中起重要作用(Kiss, 2000; Perrin et al., 2005)。研究发现, 拟南芥(Arabidopsis thaliana)主根根柱细胞中的淀粉粒在水势梯度刺激后会迅速降解, 导致根的重力敏感性降低, 同时促进其向水性反应(Takahashi et al., 2003)。但根柱细胞内淀粉粒的降解不仅受到水势梯度的诱导, 而且也受到其它不同胁迫(如干旱)的调控 (Takahashi et al., 2003; Sun et al., 2008; Ding and Friml, 2010; Iyer-Pascuzzi et al., 2011)。因此, 尚不能确定向水性反应中的淀粉粒降解是由水势梯度特异诱导的还是受干旱诱导发生的。而且在一些植物中, 干旱会促进根的向地性反应(Leopold and LaFavre, 1989), 说明淀粉粒的降解可能对诱导植物根的向水性反应并非必需(Nakayama et al., 2012)。 进入21世纪后, 日本东北大学的Takahashi实验室和墨西哥国立自治大学的Cassab实验室开始利用分子遗传学手段研究根向水性反应的分子机制。Cassab实验室将培养基分成上下两部分, 其中上半部分为正常1/2MS培养基(normal nutrient medium, NM), 下半部分则是在正常培养基的基础上添加了渗透调节剂(如甘油或海藻酸)来降低培养基的水分含量(water stress medium, WSM), 进而人为地造成上下水势梯度。将萌发后的拟南芥幼苗放在上半部分离界面1 cm左右的培养基上垂直培养, 当根生长到接近两种培养基界面时, 野生型植物的根会克服向地性而向上弯曲生长, 说明其对水势梯度敏感; 而失去向水性生长的突变体会继续向下生长, 说明其对水势梯度不敏感(图1C)。利用此方法, Cassab实验室分离到两个向水性缺陷突变体, 分别命名为nhr1 (no hydrotropic response 1)和ahr1 (altered hydrotropic response 1) (Eapen et al., 2003; Saucedo et al., 2012), 这两个突变体都呈现半显性遗传的特点, 但造成二者表型的突变基因至今未被鉴定。 Takahashi实验室利用一个密闭的聚丙烯塑料盒建立了空气水分梯度(图1D)。在密闭盒子的底部放置1个盛有饱和碳酸钾溶液的器皿, 盒子的内侧上方固定垂直放置添加了琼脂糖的1/2MS培养基, 形成盒子下方水势低而上方水势高的空气水势梯度。将拟南芥幼苗垂直植入培养基, 使其根尖露出培养基边缘3 mm, 在培养过程中, 野生型的根会克服向地性朝向培养基方向弯曲生长, 而对水分梯度不敏感的突变体则不会发生弯曲, 最终根尖暴露在空气中脱水死亡。该实验室利用此系统筛选到1个具有正常向地性和生长表型, 但对水势梯度不敏感的突变体, 命名为“mizu-kussei”(日语中的向水性一词, 简称miz1) (Kobayashi et al., 2007)。研究表明, MIZ1在感应水势梯度的早期阶段扮演着重要角色, 然而其编码蛋 白在调节根向水性反应过程中的具体分子机制尚不清楚。 同时, Takahashi实验室还筛选到另一个拟南芥根向水性点突变体miz2 (miz-kussei2), MIZ2编码1个参与细胞中囊泡运输的GNOM蛋白。miz2的向地性并没有因为其向水性的改变而发生变化(Miyazawa et al., 2009)。此外, GNOM调节的囊泡运输可被囊泡运输抑制剂BFA (Brefeldin A)所抑制(Geldner et al., 2003), 而体外施加BFA也可明显抑制野生型的向水性弯曲, 说明拟南芥根的向水性反应依赖于囊泡运输(Miyazawa et al., 2009)。随后, 科学家将miz2和超表达的MIZ1 (MIZ1OE)进行杂交, 发现尽管MIZ1基因超表达可以导致拟南芥根的向水性反应显著增强, 但miz2/MIZ1OE根对水势梯度依然表现出不敏感的表型, 说明MIZ1调控的根向水性反应依赖于MIZ2的功能, 即MIZ1和MIZ2在植物根向水性反应中处于同一信号通路, 且MIZ2在MIZ1下游发挥作用(Miyazawa et al., 2012)。 在探究向水性反应中根对信号的具体感知部位时, 科学家利用Ageotropum证明了根尖在感知水势梯度时起着决定性作用, 截除根冠的豌豆突变体失去了向水性生长的功能(Jaffe et al., 1985)。然而最近的研究发现, 拟南芥根尖的分生-伸长过渡区对向水性至关重要, 而根冠对于向水性信号的感知并不重要(Dietrich et al., 2017)。目前, 尚不清楚这两个矛盾的研究结果是由于研究材料的种间差异造成的, 还是研究方法不同所致。
2.4 多种植物激素和光对植物根向性反应的影响2.4.1 多种植物激素参与植物根的向地性反应, 但其是否参与向水性反应有待研究 加拿大科学家Rood等(1987)证明, 赤霉素参与玉米茎的向地性反应。在水稻叶中, 赤霉素受生长素的诱导并参与其向地性弯曲(Cui et al., 2005)。此外, Wolbang等(2007)的研究发现, 大麦(Hordeum vul- gare)叶的向地性反应会导致弯曲部位的赤霉素和生长素不对称分布。同样, Löfke等(2013)的研究发现, 在拟南芥根的向地性反应中, 赤霉素也会在根弯曲凹面处堆积, 出现不对称分布现象。 油菜素内酯是一类广泛存在于植物体内的甾醇类激素, 它参与调节植物的生长发育。有报道指出, 油菜素内酯参与拟南芥花序的负向地性反应(Arteca and Arteca, 2011)。此外, 油菜素内酯和乙烯在参与调节拟南芥茎的负向地性反应中具相反的作用, 内源油菜素内酯信号会降低植物对乙烯信号的感知从而使拟南芥的茎失去负向地性反应(Vandenbussche et al., 2013)。虽然目前的研究确定赤霉素、油菜素内酯和乙烯都参与植物的向地性反应, 但三者是否调节植物根的向水性反应仍然未知。 2.4.2 光、ROS及钙离子参与调节植物根的向水性反应 光作为植物根的负向光性诱导因子, 也可影响植物的向水性反应。研究发现, 光影响植物的向水性反应主要通过调节植物体内MIZ1基因的表达来实现, 与生长在白光条件下的植物相比, 黑暗环境下生长的植物根中MIZ1的转录水平降低, 从而降低了其向水性反应(Moriwaki et al., 2012)。该实验还证明, 光依赖的MIZ1基因表达受光信号下游转录因子HY5的调控。表明光对植物向水性反应非常重要, 尤其是植物的根暴露在土壤表面或在极浅的土壤中时, 光可以提高植物根中MIZ1的表达, 从而使根发生向水性弯曲生长, 进入土壤深处, 免受干旱胁迫。植物的侧根发生受水势梯度的诱导, 水势梯度较高一侧产生的侧根更多(Bao et al., 2014; Robbins and Dinneny, 2018)。此外, 光还能调节MIZ1蛋白在侧根根冠处积累, 进而使侧根也具有感知水势梯度的能力(Moriwaki et al., 2012)。 有研究表明, 植物根因重力刺激发生向地性弯曲后, 弯曲根的凹面处活性氧(reactive oxygen species, ROS)积累增加, 从而抑制凹面处的生长, 但这种根中ROS的单侧积累并不会发生在向水性弯曲生长过程中(Krieger et al., 2016)。另外, 提高植物根中细胞质的钙离子浓度有助于根对水势梯度的感知, 但其作用机制并不清楚(Takano et al., 1997)。
3 展望植物根的向水性反应与植物的抗旱反应不同, 严重的干旱环境会导致植物发生脱落酸积累、气孔关闭、生长停滞或者局部死亡等一系列生理生化反应(陈善福和舒庆尧, 1999; 陈健辉等, 2011; 侯智慧, 2011; 刘晓东等, 2016); 而向水性反应是指植物针对根尖的水势梯度调整植物根系的生长方向, 对植物从环境中获取必要的水分至关重要。但是对于植物根向水性的机理研究到目前为止还停留在探索阶段。目前的研究认为, MIZ1作为植物根向水性反应的重要调节基因, 与GNOM调控的囊泡运输共同发挥作用; 植物激素参与根的向水性反应机制并不明确; 光可以通过下游转录因子HY5调控MIZ1的表达, 从而调节植物根的向水性反应(图2)。科学家们通过各种方法优化植物向水性的模拟实验体系, 以排除研究过程中重力对向水性的干扰。目前对植物向水性的研究尚有许多未解决的问题。例如, MIZ1作为调控拟南芥根向水性反应的重要基因, 其编码蛋白的功能和调控机制尚不清楚; 尽管MIZ2调控的囊泡运输在拟南芥根的向水性反应中发挥重要作用, 但其作用机制, 即囊泡具体运输了什么物质(或信号)仍有待进一步研究; 生长素在不同植物中的根向水性反应有所不同, 这种种间差异的调控机理尚未被解析; 植物根的向水性反应除了受生长素的调节, 还可能受植物体内细胞分裂素及脱落酸的影响, 但目前这2种激素参与调节向水性反应的机制仍不清楚。另外, 油菜素内酯和赤霉素等植物激素都参与调节植物根的向地性反应, 但其是否参与及如何调节植物根的向水性反应仍需进一步研究。 图2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-2-154/img_2.png<b>图2</b> 已知植物根向水性反应的调控模式<br/>MIZ1和GNOM可以调控植物根的向水性反应, 而且GNOM对MIZ1功能的发挥是必要的; 光可以通过下游转录因子HY5调控<i> MIZ1</i>的表达从而调节植物根的向水性反应; 但植物激素参与植物根向水性反应的作用机制仍有待进一步阐明。<br/><b>Figure 2</b> Current understanding of plant root hydrotropic response<br/>MIZ1 and GNOM are known proteins regulating hydrotropism. GNOM is essential for the function of MIZ1. Light can regulate the transcription of <i>MIZ1</i> via an important transcriptional factor, HY5. Additional analyses are needed to elucidate the detailed molecular mechanisms of phytohormones in controlling root hydrotropic response. Figure 2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-2-154/img_2.png<b>图2</b> 已知植物根向水性反应的调控模式<br/>MIZ1和GNOM可以调控植物根的向水性反应, 而且GNOM对MIZ1功能的发挥是必要的; 光可以通过下游转录因子HY5调控<i> MIZ1</i>的表达从而调节植物根的向水性反应; 但植物激素参与植物根向水性反应的作用机制仍有待进一步阐明。<br/><b>Figure 2</b> Current understanding of plant root hydrotropic response<br/>MIZ1 and GNOM are known proteins regulating hydrotropism. GNOM is essential for the function of MIZ1. Light can regulate the transcription of <i>MIZ1</i> via an important transcriptional factor, HY5. Additional analyses are needed to elucidate the detailed molecular mechanisms of phytohormones in controlling root hydrotropic response.
图2 已知植物根向水性反应的调控模式 MIZ1和GNOM可以调控植物根的向水性反应, 而且GNOM对MIZ1功能的发挥是必要的; 光可以通过下游转录因子HY5调控 MIZ1的表达从而调节植物根的向水性反应; 但植物激素参与植物根向水性反应的作用机制仍有待进一步阐明。 Figure 2 Current understanding of plant root hydrotropic response MIZ1 and GNOM are known proteins regulating hydrotropism. GNOM is essential for the function of MIZ1. Light can regulate the transcription of MIZ1 via an important transcriptional factor, HY5. Additional analyses are needed to elucidate the detailed molecular mechanisms of phytohormones in controlling root hydrotropic response.
MichniewiczM, BrewerPB, FrimlJ (2007). Polar auxin transport and asymmetric auxin distribution.Arabidopsis Book 5, e0108. [本文引用: 1]
[42]
MishraRC, GhoshR, BaeH (2016). Plant acoustics: in the search of a sound mechanism for sound signaling in plants.J Exp Bot 67, 4483-4494. [本文引用: 1]
[43]
MiyazawaY, TakahashiA, KobayashiA, KaneyasuT, FujiiN, TakahashiH (2009). GNOM-mediated vesicular trafficking plays an essential role in hydrotropism of Ara- bidopsis roots.Plant Physiol 149, 835-840. [本文引用: 2]
[44]
MiyazawaY, MoriwakiT, UchidaM, KobayashiA, FujiiN, TakahashiH (2012). Overexpression of MIZU-KUSSEI1 enhances the root hydrotropic response by retaining cell viability under hydrostimulated conditions in Arabidopsis thaliana.Plant Cell Physiol 53, 1926-1933. [本文引用: 2]
[45]
MizunoH, KobayashiA, FujiiN, YamashitaM, TakahashiH (2002). Hydrotropic response and expression pattern of auxin-inducible gene,CS-IAA1, in the primary roots of clinorotated cucumber seedlings.Plant Cell Physiol 43, 793-801. [本文引用: 1]
MoriwakiT, MiyazawaY, FujiiN, TakahashiH (2012). Light and abscisic acid signaling are integrated by MIZ1 gene expression and regulate hydrotropic response in roots of Arabidopsis thaliana.Plant Cell Environ 35, 1359-1368. [本文引用: 4]
[49]
MoriwakiT, MiyazawaY, KobayashiA, UchidaM, WatanabeC, FujiiN, TakahashiH (2011). Hormonal regulation of lateral root development in Arabidopsis modulated by MIZ1 and requirement of GNOM activity for MIZ1 function.Plant Physiol 157, 1209-1220. [本文引用: 1]
NagpalP, WalkerLM, YoungJC, SonawalaA, TimpteC, EstelleM, ReedJW (2000). AXR2 encodes a member of the Aux/IAA protein family. Plant Physiol 123, 563-574. [本文引用: 1]
[52]
NakajimaY, NaraY, KobayashiA, SugitaT, MiyazawaY, FujiiN, TakahashiH (2017). Auxin transport and response requirements for root hydrotropism differ between plant species.J Exp Bot 68, 3441-3456. [本文引用: 4]
[53]
NakayamaM, KanekoY, MiyazawaY, FujiiN, HigashitaniN, WadaS, IshidaH, YoshimotoK, ShirasuK, YamadaK, NishimuraM, TakahashiH (2012). A possible involvement of autophagy in amyloplast degradation in colu- mella cells during hydrotropic response of Arabidopsis roots.Planta 236, 999-1012. [本文引用: 1]
[54]
PerrinRM, YoungLS, MurthyUMN, HarrisonBR, WangY, WillJL, MassonPH (2005). Gravity signal transduction in primary roots.Ann Bot 96, 737-743. [本文引用: 1]
[55]
PonceG, RasgadoFA, CassabGI (2008). Roles of amyloplasts and water deficit in root tropisms.Plant Cell Environ 31, 205-217. [本文引用: 1]
Quiroz-FigueroaF, Rodríguez-AcostaA, Salazar-BlasA, Hernández-DomínguezE, CamposME, KitahataN, AsamiT, Galaz-AvalosRM, CassabGI (2010). Accumulation of high levels of ABA regulates the pleiotropic response of thenhr1 Arabidopsis mutant. J Plant Biol 53, 32-44. [本文引用: 1]
[58]
RashotteAM, De LongA, MudayGK (2001). Genetic and chemical reductions in protein phosphatase activity alter auxin transport, gravity response, and lateral root growth.Plant Cell 13, 1683-1697. [本文引用: 1]
[59]
Robbins IINE, DinnenyJR (2018). Growth is required for perception of water availability to pattern root branches in plants.Proc Natl Acad Sci USA 115, E822-E831. [本文引用: 1]
[60]
Rodrigo-MorenoA, BazihizinaN, AzzarelloE, MasiE, TranD, BouteauF, BaluskaF, MancusoS (2017). Root phonotropism: early signaling events following sound per- ception in Arabidopsis roots.Plant Sci 264, 9-15. [本文引用: 1]
[61]
RoodSB, KaufmanPB, AbeH, PharisRP (1987). Gibberel- lins and gravitropism in maize shoots: endogenous gibberellin-like substances and movement and metabolism of [3H] gibberellin A20.Plant Physiol 83, 645-651. [本文引用: 1]
[62]
SachsJ (1887). Lectures on the physiology of plants. Oxford: Clarendon Press. pp. 1832-1897. [本文引用: 3]
[63]
SaucedoM, PonceG, CamposME, EapenD, GarcíaE, LujánR, SánchezY, CassabGI (2012). An altered hydrotropic response (ahr1) mutant of Arabidopsis recovers root hydrotropism with cytokinin. J Exp Bot 63, 3587-3601. [本文引用: 4]
[64]
SchöldéenC, BurströmH (1960). Physiological studies of an ageotropic pea mutant.Physiol Plant 13, 831-838. [本文引用: 1]
TakahashiH (1997). Hydrotropism: the current state of our knowledge.J Plant Res 110, 163-169. [本文引用: 1]
[72]
TakahashiN, GotoN, OkadaK, TakahashiH (2002). Hydrotropism in abscisic acid, wavy, and gravitropic mutants of Arabidopsis thaliana. Planta 216, 203-211. [本文引用: 5]
[73]
TakahashiH, JaffeMJ (1990). Thigmotropism and the modulation of tropistic curvature by mechanical perturbation in cucumber hypocotyls.Physiol Plant 80, 561-567. [本文引用: 2]
[74]
TakahashiH, MizunoH, KamadaM, FujiiN, HigashitaniA, KamigaichiS, AizawaS, MukaiC, ShimazuT, FukuiK, YamashitaM (1999). A spaceflight experiment for the study of gravimorphogenesis and hydrotropism in cucumber seedlings.J Plant Res 112, 497-505. [本文引用: 2]
TakahashiH, SugeH (1991). Root hydrotropism of an agravitropic pea mutant,ageotropum.Physiol Plant 82, 24-31. [本文引用: 2]
[76]
TakahashiH, TakanoM, FujiiN, YamashitaM, SugeH (1996). Induction of hydrotropism in clinorotated seedling roots of Alaska pea,Pisum sativum L. J Plant Res 109, 335-337. [本文引用: 1]
[5]
AloniR, LanghansM, AloniE, UllrichCI (2004). Role of cytokinin in the regulation of root gravitropism.Planta 220, 177-182. [本文引用: 1]
[6]
ArtecaRN, ArtecaJM (2011). Characterization of gravitropic inflorescence bending in brassinosteroid biosynthesis and signaling Arabidopsis mutants.J Plant Physiol 168, 1200-1207. [本文引用: 1]
[77]
TakahashiN, YamazakiY, KobayashiA, HigashitaniA, TakahashiH (2003). Hydrotropism interacts with gravitropism by degrading amyloplasts in seedling roots of Ara- bidopsis and radish.Plant Physiol 132, 805-810. [本文引用: 2]
[78]
TakanoM, TakahashiH, SugeH (1997). Calcium requirement for the induction of hydrotropism and enhancement of calcium-induced curvature by water stress in primary roots of pea,Pisum sativum L. Plant Cell Physiol 38, 385-391. [本文引用: 1]
[79]
Van OverbeekJ (1939). Phototropism.Bot Rev 5, 655-681. [本文引用: 1]
[80]
VandenbusscheF, CallebertP, ZadnikovaP, BenkovaE, Van Der StraetenD (2013). Brassinosteroid control of shoot gravitropism interacts with ethylene and depends on auxin signaling components.Am J Bot 100, 215-225. [本文引用: 1]
[7]
BaoY, AggarwalP, RobbinsNE, SturrockCJ, ThompsonMC, TanHQ, ThamC, DuanLN, RodriguezPL, VernouxT, MooneySJ, BennettMJ, DinnenyJR (2014). Plant roots use a patterning mechanism to position lateral root branches toward available water.Proc Natl Acad Sci USA 111, 9319-9324. [本文引用: 1]
[8]
BlixtS, EhrenbergL, GelinO (1958). Quantitative studies of induced mutations in peas. I. Methodological investigations.Agric Hortic Genet 16, 238-250. [本文引用: 1]
[81]
WernerT, MotykaV, LaucouV, SmetsR, Van OnckelenH, SchmüllingT (2003). Cytokinin-deficient transgenic Arabidopsis plants show multiple developmental alterations indicating opposite functions of cytokinins in the re- gulation of shoot and root meristem activity.Plant Cell 15, 2532-2550. [本文引用: 1]
[82]
WernerT, NehnevajovaE, KöllmerI, NovákO, StrnadM, KrämerU, SchmüllingT (2010). Root-specific reduction of cytokinin causes enhanced root growth, drought tolerance, and leaf mineral enrichment in Arabidopsis and tobacco.Plant Cell 22, 3905-3920. [本文引用: 1]
WilsonBF (1967). Root growth around barriers.BotGaz 128, 79-82. [本文引用: 1]
[9]
CaryAJ, LiuW, HowellSH (1995). Cytokinin action is coup- led to ethylene in its effects on the inhibition of root and hypocotyl elongation inArabidopsis thaliana seedlings. Plant Physiol 107, 1075-1082. [本文引用: 1]
[10]
CorrellMJ, KissJZ (2002). Interactions between gravitropism and phototropism in plants. J Plant Growth Regul 21, 89-101. [本文引用: 1]
[85]
WolbangCM, DaviesNW, TaylorSA, RossJJ (2007). Gravistimulation leads to asymmetry of both auxin and gibberellin levels in barley pulvini.Physiol Plant 131, 140-148. [本文引用: 1]
[86]
ZhouZY, ZhangCG, WuL, ZhangCG, ChaiJ, WangM, JhaA, JiaPF, CuiSJ, YangM, ChenRJ, GuoGQ (2011). Functional characterization of the CKRC1/TAA1 gene and dissection of hormonal actions in the Arabidopsis root.Plant J 66, 516-527. [本文引用: 1]
[11]
CuiDY, NeillSJ, TangZC, CaiWM (2005). Gibberellin-regulatedXET is differentially induced by auxin in rice leaf sheath bases during gravitropic bending. J Exp Bot 56, 1327-1334. [本文引用: 1]
[12]
DarwinC, DarwinF (1881). The Power of Movement in Plants. New York: Appleton-Century. pp. 329-338. [本文引用: 1]
DingZJ, FrimlJ (2010). Auxin regulates distal stem cell differentiation in Arabidopsis roots.Proc Natl Acad Sci USA 107, 12046-12051. [本文引用: 1]
[15]
EapenD, BarrosoML, CamposME, PonceG, CorkidiG, DubrovskyJG, CassabGI (2003). A no hydrotropic response root mutant that responds positively to gravitropism in Arabidopsis.Plant Physiol 131, 536-546. [本文引用: 3]
[16]
EkelundR, HembergT (1966). A comparison between geotropism and geoelectric effect inPisum sativum and its mutant ageotropum. Physiol Plant 19, 1120-1124. [本文引用: 1]
[17]
EsmonCA, PedmaleUV, LiscumE (2005). Plant tropisms: providing the power of movement to a sessile organism.Int J Dev Biol 49, 665-674. [本文引用: 1]
[18]
Galvan-AmpudiaC, JulkowskaM, DarwishE, GandulloJ, KorverRA, BrunoudG, HaringMA, MunnikT, VernouxT, TesterinkC (2013). Halotropism is a response of plant roots to avoid a saline environment.Curr Biol 23, 2044-2050. [本文引用: 1]
[19]
GarwoodEA, WilliamsTE (1967). Soil water use and growth of a grass sward.J Agric Sci 68, 281-292. [本文引用: 1]
[20]
GehringCA, WilliamsDA, CodySH, ParishRW (1990). Phototropism and geotropism in maize coleoptiles are spatially correlated with increases in cytosolic free calcium.Nature 345, 528-530. [本文引用: 2]
HanW, RongHH, ZhangHM, WangMH (2009). Abscisic acid is a negative regulator of root gravitropism in Arabidopsis thaliana.Biochem Biophys Res Commun 378, 695-700. [本文引用: 1]
[23]
HartJW (1990). Plant Tropisms and Other Growth Movements. Netherlands: Springer. pp. 257-258. [本文引用: 1]
[24]
HosonT, KamisakaS, MasudaY, YamashitaM, BuchenB (1997). Evaluation of the three-dimensional clinostat as a simulator of weightlessness.Planta 203, S187-S197. [本文引用: 1]
[25]
HsiaoTC, XuLK (2000). Sensitivity of growth of roots versus leaves to water stress: biophysical analysis and relation to water transport.J Exp Bot 51, 1595-1616. [本文引用: 1]
[26]
Iyer-PascuzziAS, JacksonT, CuiHC, PetrickaJJ, BuschW, TsukagoshiH, BenfeyPN (2011). Cell identity regulators link development and stress responses in the Ara- bidopsis root.Dev Cell 21, 770-782. [本文引用: 1]
[27]
JaffeMJ, TakahashiH, BiroRL (1985). A pea mutant for the study of hydrotropism in roots.Science 230, 445-447. [本文引用: 3]
[28]
JonesB, GunnerasSA, PeterssonSV, TarkowskiP, GrahamN, MayS, DolezalK, SandbergG, LjungK (2010). Cytokinin regulation of auxin synthesis in Arabidopsis involves a homeostatic feedback loop regulated via auxin and cytokinin signal transduction. Plant Cell 22, 2956-2969. [本文引用: 1]
[29]
KaneyasuT, KobayashiA, NakayamaM, FujiiN, TakahashiH, MiyazawaY (2007). Auxin response, but not its polar transport, plays a role in hydrotropism of Arabidopsis roots.J Exp Bot 58, 1143-1150. [本文引用: 1]
[30]
KissJZ (2000). Mechanisms of the early phases of plant gravitropism.CRC Crit Rev Plant Sci 19, 551-573. [本文引用: 1]
[31]
KnightTA (1811). On the causes which influence the direction of the growth of roots.Philos Trans R Soc Lond B Biol Sci 101, 209-219. [本文引用: 1]
[32]
KobayashiA, TakahashiA, KakimotoY, MiyazawaY, FujiiN, HigashitaniA, TakahashiH (2007). A gene essential for hydrotropism in roots.Proc Natl Acad Sci USA 104, 4724-4729. [本文引用: 3]
[33]
KoornneefM, ReulingG, KarssenCM (1984). The isolation and characterization of abscisic acid insensitive mutants ofArabidopsis thaliana. Physiol Plant 61, 377-383. [本文引用: 1]
[34]
KoornneefM, JornaML, Brinkhorst-van der SwanDLC, KarssenCM (1982). The isolation of abscisic acid (ABA) deficient mutants by selection of induced revertants in non-germinating gibberellin sensitive lines of Arabidopsis thaliana(L.) Heynh.Theor Appl Genet 61, 385-393. [本文引用: 1]
[35]
KraftTFB, van LoonJJWA, KissJZ (2000). Plastid position in Arabidopsis columella cells is similar in microgravi- ty and on a random-positioning machine.Planta 211, 415-422. [本文引用: 1]
LeopoldAC, LaFavreAK (1989). Interactions between red light, abscisic acid, and calcium in gravitropism.Plant Physiol 89, 875-878. [本文引用: 2]
[38]
LeyserHMO, LincolnCA, TimpteC, LammerD, TurnerJ, EstelleM (1993). Arabidopsis auxin-resistance gene AXR1 encodes a protein related to ubiquitin-activating en- zyme E1.Nature 364, 161-164. [本文引用: 1]
[39]
LiscumE, AskinosieSK, LeuchtmanDL, MorrowJ, WillenburgKT, CoatsDR (2014). Phototropism: growing towards an understanding of plant movement.Plant Cell 26, 38-55. [本文引用: 1]
[40]
LöfkeC, ZwiewkaM, HeilmannI, Van MontaguMCE, TeichmannT, FrimlJ (2013). Asymmetric gibberellin signaling regulates vacuolar trafficking of PIN auxin trans- porters during root gravitropism.Proc Natl Acad Sci USA 110, 3627-3632.
1 2007
... 1991年, Takahashi在对豌豆突变体“Ageotro- pum”的向水性研究中发现, 当对Ageotropum施加生长素转运抑制剂三碘苯甲酸(TIBA)时, 其根的向水性反应被明显抑制, 说明生长素的极性运输在植物根的向水性反应过程中具有重要作用(Takahashi and Suge, 1991).然而生长素的调节效应具有种间差异.在豌豆、黄瓜和水稻(Oryza sativa)根尖, 生长素以重新分布的模式调控其根的向地性以及向水性弯曲(Takahashi and Suge, 1991; Mizuno et al., 2002; Nakajima et al., 2017); 但与豌豆同属豆科植物的百脉根(Lotus corniculatus)和十字花科的拟南芥根向水性反应却不依赖于生长素的极性运输与重新分配(Shkolnik et al., 2016; Nakajima et al., 2017).研究表明, 生长素转运抑制剂(CHPAA、NPA和TIBA)不会影响百脉根和拟南芥根的向水性反应(Kaneyasu et al., 2007; Nakajima et al., 2017), 但生长素合成抑制剂Kyn会明显减弱百脉根的向水性反应(Nakajima et al., 2017).此外, PINs是植物体内调节生长素运输方向的蛋白, 它们以极性定位的方式参与生长素在植物细胞间的极性运输(Michniewicz et al., 2007).拟南芥中, PIN2缺失突变体表现出向地性反应缺陷, 但却具有正常的向水性反应(Takahashi et al., 2002).生长素信号通路受损的拟南芥突变体axr1-3和axr2-1根的向地性反应虽然减弱(Leyser et al., 1993; Nagpal et al., 2000), 但其根对水势梯度响应的敏感性增强(Takahashi et al., 2002).研究发现, 超表达MIZ1能增强拟南芥根的向水性反应, 同时其根中生长素合成基因的表达水平明显降低(Moriwaki et al., 2011; Miyazawa et al., 2012).总之, 生长素通过极性运输和重新分布的方式参与植物根的向地性反应, 但该作用机理并不适用于所有植物(如百脉根和拟南芥)根的向水性反应, 有关生长素对向水性反应调控的种间差异及生理和分子基础有待进一步研究. ...
1 2016
... 植物在整个生命活动过程中都在被动地适应不断变化的环境.在长期的进化选择中, 植物进化出一系列特有的机制来适应周围环境的变化, 使其能够一代代繁衍生息.植物能够感知自身生长环境中各种非生物(包括光、重力、水、温度、氧气和二氧化碳等)和生物信号, 并做出相应的生理生化反应.向性反应(tropic response)作为植物适应周围环境变化的重要机制之一, 是植物实现自身趋利避害的主要途径.植物朝向环境信号生长的向性反应称为正(positive)向性反应, 反之则称为负(negative)向性反应.植物为了应对不同的环境信号进化出了一系列向性反应, 包括向光性(phototropism) (Van Overbeek,1939; Gehr- ing et al., 1990; Liscum et al., 2014)、向地性(gravi- tropism) (Gehring et al., 1990; Hart, 1990; Taka- hashi and Jaffe, 1990; Correll and Kiss, 2002)、向化性(chemotropism) (Esmon et al., 2005)、向触性(thigmotropism) (Monshausen et al., 2008)、向水性(hydrotropism) (Takahashi and Jaffe, 1990; Takaha- shi, 1997)、避盐性(halotropism) (Munns and Tes- ter, 2008; Sun et al., 2008; Shelef et al., 2010; Galvan-Ampudia et al., 2013)及向声性(phonotro pism) (Mishra et al., 2016; Rodrigo-Moreno et al., 2017)等.其中, 向地性和向光性由于较易观察和筛选等原因, 研究进展较快, 目前已对调控植物向地性和向光性的分子机理有了较为深入的认识.由于重力等因素的干扰, 根向水性的研究较为困难.目前的研究仅发现少数基因和突变体参与或改变植物的向水性反应.向水性机理研究不仅具有重要的理论意义, 而且对节水灌溉和提高植物的抗旱性等也具有指导意义. ...
2 2009
... 同时, Takahashi实验室还筛选到另一个拟南芥根向水性点突变体miz2 (miz-kussei2), MIZ2编码1个参与细胞中囊泡运输的GNOM蛋白.miz2的向地性并没有因为其向水性的改变而发生变化(Miyazawa et al., 2009).此外, GNOM调节的囊泡运输可被囊泡运输抑制剂BFA (Brefeldin A)所抑制(Geldner et al., 2003), 而体外施加BFA也可明显抑制野生型的向水性弯曲, 说明拟南芥根的向水性反应依赖于囊泡运输(Miyazawa et al., 2009).随后, 科学家将miz2和超表达的MIZ1 (MIZ1OE)进行杂交, 发现尽管MIZ1基因超表达可以导致拟南芥根的向水性反应显著增强, 但miz2/MIZ1OE根对水势梯度依然表现出不敏感的表型, 说明MIZ1调控的根向水性反应依赖于MIZ2的功能, 即MIZ1和MIZ2在植物根向水性反应中处于同一信号通路, 且MIZ2在MIZ1下游发挥作用(Miyazawa et al., 2012). ... ... ), 而体外施加BFA也可明显抑制野生型的向水性弯曲, 说明拟南芥根的向水性反应依赖于囊泡运输(Miyazawa et al., 2009).随后, 科学家将miz2和超表达的MIZ1 (MIZ1OE)进行杂交, 发现尽管MIZ1基因超表达可以导致拟南芥根的向水性反应显著增强, 但miz2/MIZ1OE根对水势梯度依然表现出不敏感的表型, 说明MIZ1调控的根向水性反应依赖于MIZ2的功能, 即MIZ1和MIZ2在植物根向水性反应中处于同一信号通路, 且MIZ2在MIZ1下游发挥作用(Miyazawa et al., 2012). ...
2 2012
... 同时, Takahashi实验室还筛选到另一个拟南芥根向水性点突变体miz2 (miz-kussei2), MIZ2编码1个参与细胞中囊泡运输的GNOM蛋白.miz2的向地性并没有因为其向水性的改变而发生变化(Miyazawa et al., 2009).此外, GNOM调节的囊泡运输可被囊泡运输抑制剂BFA (Brefeldin A)所抑制(Geldner et al., 2003), 而体外施加BFA也可明显抑制野生型的向水性弯曲, 说明拟南芥根的向水性反应依赖于囊泡运输(Miyazawa et al., 2009).随后, 科学家将miz2和超表达的MIZ1 (MIZ1OE)进行杂交, 发现尽管MIZ1基因超表达可以导致拟南芥根的向水性反应显著增强, 但miz2/MIZ1OE根对水势梯度依然表现出不敏感的表型, 说明MIZ1调控的根向水性反应依赖于MIZ2的功能, 即MIZ1和MIZ2在植物根向水性反应中处于同一信号通路, 且MIZ2在MIZ1下游发挥作用(Miyazawa et al., 2012). ... ... 1991年, Takahashi在对豌豆突变体“Ageotro- pum”的向水性研究中发现, 当对Ageotropum施加生长素转运抑制剂三碘苯甲酸(TIBA)时, 其根的向水性反应被明显抑制, 说明生长素的极性运输在植物根的向水性反应过程中具有重要作用(Takahashi and Suge, 1991).然而生长素的调节效应具有种间差异.在豌豆、黄瓜和水稻(Oryza sativa)根尖, 生长素以重新分布的模式调控其根的向地性以及向水性弯曲(Takahashi and Suge, 1991; Mizuno et al., 2002; Nakajima et al., 2017); 但与豌豆同属豆科植物的百脉根(Lotus corniculatus)和十字花科的拟南芥根向水性反应却不依赖于生长素的极性运输与重新分配(Shkolnik et al., 2016; Nakajima et al., 2017).研究表明, 生长素转运抑制剂(CHPAA、NPA和TIBA)不会影响百脉根和拟南芥根的向水性反应(Kaneyasu et al., 2007; Nakajima et al., 2017), 但生长素合成抑制剂Kyn会明显减弱百脉根的向水性反应(Nakajima et al., 2017).此外, PINs是植物体内调节生长素运输方向的蛋白, 它们以极性定位的方式参与生长素在植物细胞间的极性运输(Michniewicz et al., 2007).拟南芥中, PIN2缺失突变体表现出向地性反应缺陷, 但却具有正常的向水性反应(Takahashi et al., 2002).生长素信号通路受损的拟南芥突变体axr1-3和axr2-1根的向地性反应虽然减弱(Leyser et al., 1993; Nagpal et al., 2000), 但其根对水势梯度响应的敏感性增强(Takahashi et al., 2002).研究发现, 超表达MIZ1能增强拟南芥根的向水性反应, 同时其根中生长素合成基因的表达水平明显降低(Moriwaki et al., 2011; Miyazawa et al., 2012).总之, 生长素通过极性运输和重新分布的方式参与植物根的向地性反应, 但该作用机理并不适用于所有植物(如百脉根和拟南芥)根的向水性反应, 有关生长素对向水性反应调控的种间差异及生理和分子基础有待进一步研究. ...
1 2002
... 1991年, Takahashi在对豌豆突变体“Ageotro- pum”的向水性研究中发现, 当对Ageotropum施加生长素转运抑制剂三碘苯甲酸(TIBA)时, 其根的向水性反应被明显抑制, 说明生长素的极性运输在植物根的向水性反应过程中具有重要作用(Takahashi and Suge, 1991).然而生长素的调节效应具有种间差异.在豌豆、黄瓜和水稻(Oryza sativa)根尖, 生长素以重新分布的模式调控其根的向地性以及向水性弯曲(Takahashi and Suge, 1991; Mizuno et al., 2002; Nakajima et al., 2017); 但与豌豆同属豆科植物的百脉根(Lotus corniculatus)和十字花科的拟南芥根向水性反应却不依赖于生长素的极性运输与重新分配(Shkolnik et al., 2016; Nakajima et al., 2017).研究表明, 生长素转运抑制剂(CHPAA、NPA和TIBA)不会影响百脉根和拟南芥根的向水性反应(Kaneyasu et al., 2007; Nakajima et al., 2017), 但生长素合成抑制剂Kyn会明显减弱百脉根的向水性反应(Nakajima et al., 2017).此外, PINs是植物体内调节生长素运输方向的蛋白, 它们以极性定位的方式参与生长素在植物细胞间的极性运输(Michniewicz et al., 2007).拟南芥中, PIN2缺失突变体表现出向地性反应缺陷, 但却具有正常的向水性反应(Takahashi et al., 2002).生长素信号通路受损的拟南芥突变体axr1-3和axr2-1根的向地性反应虽然减弱(Leyser et al., 1993; Nagpal et al., 2000), 但其根对水势梯度响应的敏感性增强(Takahashi et al., 2002).研究发现, 超表达MIZ1能增强拟南芥根的向水性反应, 同时其根中生长素合成基因的表达水平明显降低(Moriwaki et al., 2011; Miyazawa et al., 2012).总之, 生长素通过极性运输和重新分布的方式参与植物根的向地性反应, 但该作用机理并不适用于所有植物(如百脉根和拟南芥)根的向水性反应, 有关生长素对向水性反应调控的种间差异及生理和分子基础有待进一步研究. ...
1 2001
... 在植物根系的生长过程中, 细胞分裂素作为重要的植物调节激素被广泛认知(Mok and Mok, 2001; Wer- ner and Schmülling, 2009).细胞分裂素不仅可以抑制植物根的伸长和分化, 还参与植物根对干旱胁迫的响应(Skoog and Miller, 1957; Cary et al., 1995; Werner et al., 2003, 2010).研究发现, 在植物根的一侧施加浓度较高的细胞分裂素会使根克服重力朝着施加位点弯曲生长(Aloni et al., 2004), 说明细胞分裂素在植物根的向地性反应中可能起着负向调节作用.此外, 有研究表明, 细胞分裂素不仅可以下调植物根中生长素的极性运输, 而且能刺激根局部区域生长素的合成, 从而参与调节植物根的向地性生长(Zhou et al., 2011). ...
1 2008
... 植物在整个生命活动过程中都在被动地适应不断变化的环境.在长期的进化选择中, 植物进化出一系列特有的机制来适应周围环境的变化, 使其能够一代代繁衍生息.植物能够感知自身生长环境中各种非生物(包括光、重力、水、温度、氧气和二氧化碳等)和生物信号, 并做出相应的生理生化反应.向性反应(tropic response)作为植物适应周围环境变化的重要机制之一, 是植物实现自身趋利避害的主要途径.植物朝向环境信号生长的向性反应称为正(positive)向性反应, 反之则称为负(negative)向性反应.植物为了应对不同的环境信号进化出了一系列向性反应, 包括向光性(phototropism) (Van Overbeek,1939; Gehr- ing et al., 1990; Liscum et al., 2014)、向地性(gravi- tropism) (Gehring et al., 1990; Hart, 1990; Taka- hashi and Jaffe, 1990; Correll and Kiss, 2002)、向化性(chemotropism) (Esmon et al., 2005)、向触性(thigmotropism) (Monshausen et al., 2008)、向水性(hydrotropism) (Takahashi and Jaffe, 1990; Takaha- shi, 1997)、避盐性(halotropism) (Munns and Tes- ter, 2008; Sun et al., 2008; Shelef et al., 2010; Galvan-Ampudia et al., 2013)及向声性(phonotro pism) (Mishra et al., 2016; Rodrigo-Moreno et al., 2017)等.其中, 向地性和向光性由于较易观察和筛选等原因, 研究进展较快, 目前已对调控植物向地性和向光性的分子机理有了较为深入的认识.由于重力等因素的干扰, 根向水性的研究较为困难.目前的研究仅发现少数基因和突变体参与或改变植物的向水性反应.向水性机理研究不仅具有重要的理论意义, 而且对节水灌溉和提高植物的抗旱性等也具有指导意义. ...
4 2012
... 日本Takahashi实验室利用新的实验体系(图1E), 发现拟南芥脱落酸合成缺失突变体aba1-1 (Koorn- neef et al., 1982)以及脱落酸不敏感突变体abi2-1 (Koornneef et al., 1984)对水势梯度不敏感, 但二者的根呈现出与野生型一致的向地性弯曲, 由此认为脱落酸能促进拟南芥根的向水性弯曲(Takahashi et al., 2002).随后的实验证明, 脱落酸可以通过调节MIZ1的表达来参与植物的向水性反应(Moriwaki et al., 2012).脱落酸信号途径正调控因子SnRK2.2及SnRK2.3的双重缺失突变体snrk2.2/snrk2.3表现出向水性反应降低的表型, 说明脱落酸正向调节拟南芥根的向水性反应.同时, 该研究还表明, 拟南芥根尖皮层细胞的分裂对向水性非常重要(Dietrich et al., 2017).然而, 脱落酸尽管可以恢复ahr1对水势梯度感知的缺陷, 但却加剧了nhr1向水性反应缺失的表型(Ponce et al., 2008; Moriwaki et al., 2012).墨西哥Cassab实验室对nhr1突变体脱落酸的定量分析显示, nhr1突变会造成植物体内脱落酸含量升高.再者, 脱落酸合成抑制剂AbamineSG会恢复nhr1向水性缺失的表型, 说明在nhr1参与的向水性反应中, 脱落酸可能起着负向调控作用(Quiroz-Figueroa et al., 2010). ... ... ; Moriwaki et al., 2012).墨西哥Cassab实验室对nhr1突变体脱落酸的定量分析显示, nhr1突变会造成植物体内脱落酸含量升高.再者, 脱落酸合成抑制剂AbamineSG会恢复nhr1向水性缺失的表型, 说明在nhr1参与的向水性反应中, 脱落酸可能起着负向调控作用(Quiroz-Figueroa et al., 2010). ... ... 光作为植物根的负向光性诱导因子, 也可影响植物的向水性反应.研究发现, 光影响植物的向水性反应主要通过调节植物体内MIZ1基因的表达来实现, 与生长在白光条件下的植物相比, 黑暗环境下生长的植物根中MIZ1的转录水平降低, 从而降低了其向水性反应(Moriwaki et al., 2012).该实验还证明, 光依赖的MIZ1基因表达受光信号下游转录因子HY5的调控.表明光对植物向水性反应非常重要, 尤其是植物的根暴露在土壤表面或在极浅的土壤中时, 光可以提高植物根中MIZ1的表达, 从而使根发生向水性弯曲生长, 进入土壤深处, 免受干旱胁迫.植物的侧根发生受水势梯度的诱导, 水势梯度较高一侧产生的侧根更多(Bao et al., 2014; Robbins and Dinneny, 2018).此外, 光还能调节MIZ1蛋白在侧根根冠处积累, 进而使侧根也具有感知水势梯度的能力(Moriwaki et al., 2012). ... ... ).此外, 光还能调节MIZ1蛋白在侧根根冠处积累, 进而使侧根也具有感知水势梯度的能力(Moriwaki et al., 2012). ...
1 2011
... 1991年, Takahashi在对豌豆突变体“Ageotro- pum”的向水性研究中发现, 当对Ageotropum施加生长素转运抑制剂三碘苯甲酸(TIBA)时, 其根的向水性反应被明显抑制, 说明生长素的极性运输在植物根的向水性反应过程中具有重要作用(Takahashi and Suge, 1991).然而生长素的调节效应具有种间差异.在豌豆、黄瓜和水稻(Oryza sativa)根尖, 生长素以重新分布的模式调控其根的向地性以及向水性弯曲(Takahashi and Suge, 1991; Mizuno et al., 2002; Nakajima et al., 2017); 但与豌豆同属豆科植物的百脉根(Lotus corniculatus)和十字花科的拟南芥根向水性反应却不依赖于生长素的极性运输与重新分配(Shkolnik et al., 2016; Nakajima et al., 2017).研究表明, 生长素转运抑制剂(CHPAA、NPA和TIBA)不会影响百脉根和拟南芥根的向水性反应(Kaneyasu et al., 2007; Nakajima et al., 2017), 但生长素合成抑制剂Kyn会明显减弱百脉根的向水性反应(Nakajima et al., 2017).此外, PINs是植物体内调节生长素运输方向的蛋白, 它们以极性定位的方式参与生长素在植物细胞间的极性运输(Michniewicz et al., 2007).拟南芥中, PIN2缺失突变体表现出向地性反应缺陷, 但却具有正常的向水性反应(Takahashi et al., 2002).生长素信号通路受损的拟南芥突变体axr1-3和axr2-1根的向地性反应虽然减弱(Leyser et al., 1993; Nagpal et al., 2000), 但其根对水势梯度响应的敏感性增强(Takahashi et al., 2002).研究发现, 超表达MIZ1能增强拟南芥根的向水性反应, 同时其根中生长素合成基因的表达水平明显降低(Moriwaki et al., 2011; Miyazawa et al., 2012).总之, 生长素通过极性运输和重新分布的方式参与植物根的向地性反应, 但该作用机理并不适用于所有植物(如百脉根和拟南芥)根的向水性反应, 有关生长素对向水性反应调控的种间差异及生理和分子基础有待进一步研究. ...
1 2008
... 植物在整个生命活动过程中都在被动地适应不断变化的环境.在长期的进化选择中, 植物进化出一系列特有的机制来适应周围环境的变化, 使其能够一代代繁衍生息.植物能够感知自身生长环境中各种非生物(包括光、重力、水、温度、氧气和二氧化碳等)和生物信号, 并做出相应的生理生化反应.向性反应(tropic response)作为植物适应周围环境变化的重要机制之一, 是植物实现自身趋利避害的主要途径.植物朝向环境信号生长的向性反应称为正(positive)向性反应, 反之则称为负(negative)向性反应.植物为了应对不同的环境信号进化出了一系列向性反应, 包括向光性(phototropism) (Van Overbeek,1939; Gehr- ing et al., 1990; Liscum et al., 2014)、向地性(gravi- tropism) (Gehring et al., 1990; Hart, 1990; Taka- hashi and Jaffe, 1990; Correll and Kiss, 2002)、向化性(chemotropism) (Esmon et al., 2005)、向触性(thigmotropism) (Monshausen et al., 2008)、向水性(hydrotropism) (Takahashi and Jaffe, 1990; Takaha- shi, 1997)、避盐性(halotropism) (Munns and Tes- ter, 2008; Sun et al., 2008; Shelef et al., 2010; Galvan-Ampudia et al., 2013)及向声性(phonotro pism) (Mishra et al., 2016; Rodrigo-Moreno et al., 2017)等.其中, 向地性和向光性由于较易观察和筛选等原因, 研究进展较快, 目前已对调控植物向地性和向光性的分子机理有了较为深入的认识.由于重力等因素的干扰, 根向水性的研究较为困难.目前的研究仅发现少数基因和突变体参与或改变植物的向水性反应.向水性机理研究不仅具有重要的理论意义, 而且对节水灌溉和提高植物的抗旱性等也具有指导意义. ...
1 2000
... 1991年, Takahashi在对豌豆突变体“Ageotro- pum”的向水性研究中发现, 当对Ageotropum施加生长素转运抑制剂三碘苯甲酸(TIBA)时, 其根的向水性反应被明显抑制, 说明生长素的极性运输在植物根的向水性反应过程中具有重要作用(Takahashi and Suge, 1991).然而生长素的调节效应具有种间差异.在豌豆、黄瓜和水稻(Oryza sativa)根尖, 生长素以重新分布的模式调控其根的向地性以及向水性弯曲(Takahashi and Suge, 1991; Mizuno et al., 2002; Nakajima et al., 2017); 但与豌豆同属豆科植物的百脉根(Lotus corniculatus)和十字花科的拟南芥根向水性反应却不依赖于生长素的极性运输与重新分配(Shkolnik et al., 2016; Nakajima et al., 2017).研究表明, 生长素转运抑制剂(CHPAA、NPA和TIBA)不会影响百脉根和拟南芥根的向水性反应(Kaneyasu et al., 2007; Nakajima et al., 2017), 但生长素合成抑制剂Kyn会明显减弱百脉根的向水性反应(Nakajima et al., 2017).此外, PINs是植物体内调节生长素运输方向的蛋白, 它们以极性定位的方式参与生长素在植物细胞间的极性运输(Michniewicz et al., 2007).拟南芥中, PIN2缺失突变体表现出向地性反应缺陷, 但却具有正常的向水性反应(Takahashi et al., 2002).生长素信号通路受损的拟南芥突变体axr1-3和axr2-1根的向地性反应虽然减弱(Leyser et al., 1993; Nagpal et al., 2000), 但其根对水势梯度响应的敏感性增强(Takahashi et al., 2002).研究发现, 超表达MIZ1能增强拟南芥根的向水性反应, 同时其根中生长素合成基因的表达水平明显降低(Moriwaki et al., 2011; Miyazawa et al., 2012).总之, 生长素通过极性运输和重新分布的方式参与植物根的向地性反应, 但该作用机理并不适用于所有植物(如百脉根和拟南芥)根的向水性反应, 有关生长素对向水性反应调控的种间差异及生理和分子基础有待进一步研究. ...
4 2017
... 1991年, Takahashi在对豌豆突变体“Ageotro- pum”的向水性研究中发现, 当对Ageotropum施加生长素转运抑制剂三碘苯甲酸(TIBA)时, 其根的向水性反应被明显抑制, 说明生长素的极性运输在植物根的向水性反应过程中具有重要作用(Takahashi and Suge, 1991).然而生长素的调节效应具有种间差异.在豌豆、黄瓜和水稻(Oryza sativa)根尖, 生长素以重新分布的模式调控其根的向地性以及向水性弯曲(Takahashi and Suge, 1991; Mizuno et al., 2002; Nakajima et al., 2017); 但与豌豆同属豆科植物的百脉根(Lotus corniculatus)和十字花科的拟南芥根向水性反应却不依赖于生长素的极性运输与重新分配(Shkolnik et al., 2016; Nakajima et al., 2017).研究表明, 生长素转运抑制剂(CHPAA、NPA和TIBA)不会影响百脉根和拟南芥根的向水性反应(Kaneyasu et al., 2007; Nakajima et al., 2017), 但生长素合成抑制剂Kyn会明显减弱百脉根的向水性反应(Nakajima et al., 2017).此外, PINs是植物体内调节生长素运输方向的蛋白, 它们以极性定位的方式参与生长素在植物细胞间的极性运输(Michniewicz et al., 2007).拟南芥中, PIN2缺失突变体表现出向地性反应缺陷, 但却具有正常的向水性反应(Takahashi et al., 2002).生长素信号通路受损的拟南芥突变体axr1-3和axr2-1根的向地性反应虽然减弱(Leyser et al., 1993; Nagpal et al., 2000), 但其根对水势梯度响应的敏感性增强(Takahashi et al., 2002).研究发现, 超表达MIZ1能增强拟南芥根的向水性反应, 同时其根中生长素合成基因的表达水平明显降低(Moriwaki et al., 2011; Miyazawa et al., 2012).总之, 生长素通过极性运输和重新分布的方式参与植物根的向地性反应, 但该作用机理并不适用于所有植物(如百脉根和拟南芥)根的向水性反应, 有关生长素对向水性反应调控的种间差异及生理和分子基础有待进一步研究. ... ... ; Nakajima et al., 2017).研究表明, 生长素转运抑制剂(CHPAA、NPA和TIBA)不会影响百脉根和拟南芥根的向水性反应(Kaneyasu et al., 2007; Nakajima et al., 2017), 但生长素合成抑制剂Kyn会明显减弱百脉根的向水性反应(Nakajima et al., 2017).此外, PINs是植物体内调节生长素运输方向的蛋白, 它们以极性定位的方式参与生长素在植物细胞间的极性运输(Michniewicz et al., 2007).拟南芥中, PIN2缺失突变体表现出向地性反应缺陷, 但却具有正常的向水性反应(Takahashi et al., 2002).生长素信号通路受损的拟南芥突变体axr1-3和axr2-1根的向地性反应虽然减弱(Leyser et al., 1993; Nagpal et al., 2000), 但其根对水势梯度响应的敏感性增强(Takahashi et al., 2002).研究发现, 超表达MIZ1能增强拟南芥根的向水性反应, 同时其根中生长素合成基因的表达水平明显降低(Moriwaki et al., 2011; Miyazawa et al., 2012).总之, 生长素通过极性运输和重新分布的方式参与植物根的向地性反应, 但该作用机理并不适用于所有植物(如百脉根和拟南芥)根的向水性反应, 有关生长素对向水性反应调控的种间差异及生理和分子基础有待进一步研究. ... ... ; Nakajima et al., 2017), 但生长素合成抑制剂Kyn会明显减弱百脉根的向水性反应(Nakajima et al., 2017).此外, PINs是植物体内调节生长素运输方向的蛋白, 它们以极性定位的方式参与生长素在植物细胞间的极性运输(Michniewicz et al., 2007).拟南芥中, PIN2缺失突变体表现出向地性反应缺陷, 但却具有正常的向水性反应(Takahashi et al., 2002).生长素信号通路受损的拟南芥突变体axr1-3和axr2-1根的向地性反应虽然减弱(Leyser et al., 1993; Nagpal et al., 2000), 但其根对水势梯度响应的敏感性增强(Takahashi et al., 2002).研究发现, 超表达MIZ1能增强拟南芥根的向水性反应, 同时其根中生长素合成基因的表达水平明显降低(Moriwaki et al., 2011; Miyazawa et al., 2012).总之, 生长素通过极性运输和重新分布的方式参与植物根的向地性反应, 但该作用机理并不适用于所有植物(如百脉根和拟南芥)根的向水性反应, 有关生长素对向水性反应调控的种间差异及生理和分子基础有待进一步研究. ... ... ), 但生长素合成抑制剂Kyn会明显减弱百脉根的向水性反应(Nakajima et al., 2017).此外, PINs是植物体内调节生长素运输方向的蛋白, 它们以极性定位的方式参与生长素在植物细胞间的极性运输(Michniewicz et al., 2007).拟南芥中, PIN2缺失突变体表现出向地性反应缺陷, 但却具有正常的向水性反应(Takahashi et al., 2002).生长素信号通路受损的拟南芥突变体axr1-3和axr2-1根的向地性反应虽然减弱(Leyser et al., 1993; Nagpal et al., 2000), 但其根对水势梯度响应的敏感性增强(Takahashi et al., 2002).研究发现, 超表达MIZ1能增强拟南芥根的向水性反应, 同时其根中生长素合成基因的表达水平明显降低(Moriwaki et al., 2011; Miyazawa et al., 2012).总之, 生长素通过极性运输和重新分布的方式参与植物根的向地性反应, 但该作用机理并不适用于所有植物(如百脉根和拟南芥)根的向水性反应, 有关生长素对向水性反应调控的种间差异及生理和分子基础有待进一步研究. ...
1 2012
... 植物根柱细胞中的淀粉粒在根的重力感知中起重要作用(Kiss, 2000; Perrin et al., 2005).研究发现, 拟南芥(Arabidopsis thaliana)主根根柱细胞中的淀粉粒在水势梯度刺激后会迅速降解, 导致根的重力敏感性降低, 同时促进其向水性反应(Takahashi et al., 2003).但根柱细胞内淀粉粒的降解不仅受到水势梯度的诱导, 而且也受到其它不同胁迫(如干旱)的调控 (Takahashi et al., 2003; Sun et al., 2008; Ding and Friml, 2010; Iyer-Pascuzzi et al., 2011).因此, 尚不能确定向水性反应中的淀粉粒降解是由水势梯度特异诱导的还是受干旱诱导发生的.而且在一些植物中, 干旱会促进根的向地性反应(Leopold and LaFavre, 1989), 说明淀粉粒的降解可能对诱导植物根的向水性反应并非必需(Nakayama et al., 2012). ...
1 2005
... 植物根柱细胞中的淀粉粒在根的重力感知中起重要作用(Kiss, 2000; Perrin et al., 2005).研究发现, 拟南芥(Arabidopsis thaliana)主根根柱细胞中的淀粉粒在水势梯度刺激后会迅速降解, 导致根的重力敏感性降低, 同时促进其向水性反应(Takahashi et al., 2003).但根柱细胞内淀粉粒的降解不仅受到水势梯度的诱导, 而且也受到其它不同胁迫(如干旱)的调控 (Takahashi et al., 2003; Sun et al., 2008; Ding and Friml, 2010; Iyer-Pascuzzi et al., 2011).因此, 尚不能确定向水性反应中的淀粉粒降解是由水势梯度特异诱导的还是受干旱诱导发生的.而且在一些植物中, 干旱会促进根的向地性反应(Leopold and LaFavre, 1989), 说明淀粉粒的降解可能对诱导植物根的向水性反应并非必需(Nakayama et al., 2012). ...
1 2008
... 日本Takahashi实验室利用新的实验体系(图1E), 发现拟南芥脱落酸合成缺失突变体aba1-1 (Koorn- neef et al., 1982)以及脱落酸不敏感突变体abi2-1 (Koornneef et al., 1984)对水势梯度不敏感, 但二者的根呈现出与野生型一致的向地性弯曲, 由此认为脱落酸能促进拟南芥根的向水性弯曲(Takahashi et al., 2002).随后的实验证明, 脱落酸可以通过调节MIZ1的表达来参与植物的向水性反应(Moriwaki et al., 2012).脱落酸信号途径正调控因子SnRK2.2及SnRK2.3的双重缺失突变体snrk2.2/snrk2.3表现出向水性反应降低的表型, 说明脱落酸正向调节拟南芥根的向水性反应.同时, 该研究还表明, 拟南芥根尖皮层细胞的分裂对向水性非常重要(Dietrich et al., 2017).然而, 脱落酸尽管可以恢复ahr1对水势梯度感知的缺陷, 但却加剧了nhr1向水性反应缺失的表型(Ponce et al., 2008; Moriwaki et al., 2012).墨西哥Cassab实验室对nhr1突变体脱落酸的定量分析显示, nhr1突变会造成植物体内脱落酸含量升高.再者, 脱落酸合成抑制剂AbamineSG会恢复nhr1向水性缺失的表型, 说明在nhr1参与的向水性反应中, 脱落酸可能起着负向调控作用(Quiroz-Figueroa et al., 2010). ...
1 1658
... 早期的植物生理学研究中, Porta (1658)和Knight (1811)等科学家通过大量的实验证明, 植物根系会随着生长环境中的水势梯度而改变其生长方向, 由此产生了对植物根向水性的研究.此外, 达尔文所著的《The Power of Movement in Plants》一书中也有关于向水性的描述(Darwin and Darwin, 1881).德国植物学家Sachs (1887)将由网格布包裹的潮湿木屑斜向悬挂, 让豌豆(Pisum sativum)种子在木屑表面萌发, 当豌豆的根因向地性生长至网格布下边缘时, 会沿网格布表面弯曲生长而不会进入干燥的空气中, 据此他认为植物的根存在向水性生长(图1A) (Sachs, 1887).另外, 研究还发现, 土壤中的水分会随着土壤深度、季节和降水等因素而分布不均, 产生水势梯度, 而且这种水势梯度会诱导根系向水性生长(Garwood and Williams,1967; Hsiao and Xu, 2000; Steudle, 2000).然而, 由于其它植物向性的干扰, 向水性反应一直以来未被科学家认可.直到1985年, Jaffe及其同事利用豌豆突变体“Ageotropum”为实验材料才首次强有力地证明了植物根向水性不依赖于向地性而独立存在(Jaffe et al., 1985).此豌豆突变体的根完全失去了向地性和避光性反应, 因此其部分根会向上生长出土壤(Blixt et al.,1958; Schöldéen and Burst- röm, 1960; Ekelund and Hemberg, 1966; Wilson, 1967).在一个湿度可控的玻璃盒子中培养该突变体植物, 当盒子中空气的相对湿度(relative humidity of the air, RH)达到95%-98%时, Ageotropum的根生长出土壤后不会弯曲, 而是向着湿润的空气中继续生长(图1B); 当空气相对湿度调整为85%-90%时, 原来向上生长的Ageotropum根会迅速向下弯曲生长直至进入比空气湿润的土壤里.这一现象说明, Ageotro- pum尽管缺失了向地性和避光性, 但其根仍然能感知环境中的水势梯度, 并具有正常的向水性生长反应.除了利用失去了向地性的突变体来研究植物根的向水性外, 科学家们还用竖直旋转的转盘来固定植物材料(Hoson et al., 1997; Kraft et al., 2000)进行研究, 发现竖直方向低速旋转培养植物的根的向地性被有效抵消.用此方法对豌豆以及黄瓜(Cucumis sativus)进行了研究, 发现它们的向水性表型更加明显(Takah- ashi et al., 1996, 1999).Takahashi及其同事发现, 在航天器失重条件下, 黄瓜的侧根也表现出更强烈地向水性反应, 而在正常培养的实验中侧根的向水性反应不易被观察到, 说明去除向地性影响后, 植物根对水势梯度的反应更明显(Takahashi et al.,1999). ...
1 2010
... 日本Takahashi实验室利用新的实验体系(图1E), 发现拟南芥脱落酸合成缺失突变体aba1-1 (Koorn- neef et al., 1982)以及脱落酸不敏感突变体abi2-1 (Koornneef et al., 1984)对水势梯度不敏感, 但二者的根呈现出与野生型一致的向地性弯曲, 由此认为脱落酸能促进拟南芥根的向水性弯曲(Takahashi et al., 2002).随后的实验证明, 脱落酸可以通过调节MIZ1的表达来参与植物的向水性反应(Moriwaki et al., 2012).脱落酸信号途径正调控因子SnRK2.2及SnRK2.3的双重缺失突变体snrk2.2/snrk2.3表现出向水性反应降低的表型, 说明脱落酸正向调节拟南芥根的向水性反应.同时, 该研究还表明, 拟南芥根尖皮层细胞的分裂对向水性非常重要(Dietrich et al., 2017).然而, 脱落酸尽管可以恢复ahr1对水势梯度感知的缺陷, 但却加剧了nhr1向水性反应缺失的表型(Ponce et al., 2008; Moriwaki et al., 2012).墨西哥Cassab实验室对nhr1突变体脱落酸的定量分析显示, nhr1突变会造成植物体内脱落酸含量升高.再者, 脱落酸合成抑制剂AbamineSG会恢复nhr1向水性缺失的表型, 说明在nhr1参与的向水性反应中, 脱落酸可能起着负向调控作用(Quiroz-Figueroa et al., 2010). ...
1 2001
... 生长素的Cholodny-Went假说认为, 植物根在受到重力刺激时, 会导致生长素在其根弯曲的凹面处大量积累, 生长素的累积会抑制该侧细胞的伸长, 而根的另一侧(弯曲的凸面)细胞仍能伸长, 从而导致根发生向地性弯曲生长.生长素的这种不对称分布是通过生长素的极性运输实现(Rashotte et al., 2001).然而, 目前研究认为该假说并不能用来解释植物根的向水性弯曲生长(Shkolnik et al., 2016; Shkolnik and Fromm, 2016). ...
1 2018
... 光作为植物根的负向光性诱导因子, 也可影响植物的向水性反应.研究发现, 光影响植物的向水性反应主要通过调节植物体内MIZ1基因的表达来实现, 与生长在白光条件下的植物相比, 黑暗环境下生长的植物根中MIZ1的转录水平降低, 从而降低了其向水性反应(Moriwaki et al., 2012).该实验还证明, 光依赖的MIZ1基因表达受光信号下游转录因子HY5的调控.表明光对植物向水性反应非常重要, 尤其是植物的根暴露在土壤表面或在极浅的土壤中时, 光可以提高植物根中MIZ1的表达, 从而使根发生向水性弯曲生长, 进入土壤深处, 免受干旱胁迫.植物的侧根发生受水势梯度的诱导, 水势梯度较高一侧产生的侧根更多(Bao et al., 2014; Robbins and Dinneny, 2018).此外, 光还能调节MIZ1蛋白在侧根根冠处积累, 进而使侧根也具有感知水势梯度的能力(Moriwaki et al., 2012). ...
1 2017
... 植物在整个生命活动过程中都在被动地适应不断变化的环境.在长期的进化选择中, 植物进化出一系列特有的机制来适应周围环境的变化, 使其能够一代代繁衍生息.植物能够感知自身生长环境中各种非生物(包括光、重力、水、温度、氧气和二氧化碳等)和生物信号, 并做出相应的生理生化反应.向性反应(tropic response)作为植物适应周围环境变化的重要机制之一, 是植物实现自身趋利避害的主要途径.植物朝向环境信号生长的向性反应称为正(positive)向性反应, 反之则称为负(negative)向性反应.植物为了应对不同的环境信号进化出了一系列向性反应, 包括向光性(phototropism) (Van Overbeek,1939; Gehr- ing et al., 1990; Liscum et al., 2014)、向地性(gravi- tropism) (Gehring et al., 1990; Hart, 1990; Taka- hashi and Jaffe, 1990; Correll and Kiss, 2002)、向化性(chemotropism) (Esmon et al., 2005)、向触性(thigmotropism) (Monshausen et al., 2008)、向水性(hydrotropism) (Takahashi and Jaffe, 1990; Takaha- shi, 1997)、避盐性(halotropism) (Munns and Tes- ter, 2008; Sun et al., 2008; Shelef et al., 2010; Galvan-Ampudia et al., 2013)及向声性(phonotro pism) (Mishra et al., 2016; Rodrigo-Moreno et al., 2017)等.其中, 向地性和向光性由于较易观察和筛选等原因, 研究进展较快, 目前已对调控植物向地性和向光性的分子机理有了较为深入的认识.由于重力等因素的干扰, 根向水性的研究较为困难.目前的研究仅发现少数基因和突变体参与或改变植物的向水性反应.向水性机理研究不仅具有重要的理论意义, 而且对节水灌溉和提高植物的抗旱性等也具有指导意义. ...
... 早期的植物生理学研究中, Porta (1658)和Knight (1811)等科学家通过大量的实验证明, 植物根系会随着生长环境中的水势梯度而改变其生长方向, 由此产生了对植物根向水性的研究.此外, 达尔文所著的《The Power of Movement in Plants》一书中也有关于向水性的描述(Darwin and Darwin, 1881).德国植物学家Sachs (1887)将由网格布包裹的潮湿木屑斜向悬挂, 让豌豆(Pisum sativum)种子在木屑表面萌发, 当豌豆的根因向地性生长至网格布下边缘时, 会沿网格布表面弯曲生长而不会进入干燥的空气中, 据此他认为植物的根存在向水性生长(图1A) (Sachs, 1887).另外, 研究还发现, 土壤中的水分会随着土壤深度、季节和降水等因素而分布不均, 产生水势梯度, 而且这种水势梯度会诱导根系向水性生长(Garwood and Williams,1967; Hsiao and Xu, 2000; Steudle, 2000).然而, 由于其它植物向性的干扰, 向水性反应一直以来未被科学家认可.直到1985年, Jaffe及其同事利用豌豆突变体“Ageotropum”为实验材料才首次强有力地证明了植物根向水性不依赖于向地性而独立存在(Jaffe et al., 1985).此豌豆突变体的根完全失去了向地性和避光性反应, 因此其部分根会向上生长出土壤(Blixt et al.,1958; Schöldéen and Burst- röm, 1960; Ekelund and Hemberg, 1966; Wilson, 1967).在一个湿度可控的玻璃盒子中培养该突变体植物, 当盒子中空气的相对湿度(relative humidity of the air, RH)达到95%-98%时, Ageotropum的根生长出土壤后不会弯曲, 而是向着湿润的空气中继续生长(图1B); 当空气相对湿度调整为85%-90%时, 原来向上生长的Ageotropum根会迅速向下弯曲生长直至进入比空气湿润的土壤里.这一现象说明, Ageotro- pum尽管缺失了向地性和避光性, 但其根仍然能感知环境中的水势梯度, 并具有正常的向水性生长反应.除了利用失去了向地性的突变体来研究植物根的向水性外, 科学家们还用竖直旋转的转盘来固定植物材料(Hoson et al., 1997; Kraft et al., 2000)进行研究, 发现竖直方向低速旋转培养植物的根的向地性被有效抵消.用此方法对豌豆以及黄瓜(Cucumis sativus)进行了研究, 发现它们的向水性表型更加明显(Takah- ashi et al., 1996, 1999).Takahashi及其同事发现, 在航天器失重条件下, 黄瓜的侧根也表现出更强烈地向水性反应, 而在正常培养的实验中侧根的向水性反应不易被观察到, 说明去除向地性影响后, 植物根对水势梯度的反应更明显(Takahashi et al.,1999). ... ... 植物根向水性反应实验筛选体系示意图 (A) Sachs发现植物的根具有向水性所用的实验方法(Sachs, 1887); (B) Jaffe及其同事证明植物根向水性存在所用的实验方法(Jaffe et al., 1985); (C) 墨西哥Cassab实验室的根向水性筛选系统(Eapen et al., 2003; Saucedo et al., 2012); (D), (E) 日本东北大学Takahashi实验室的根向水性筛选系统(Takahashi et al., 2002; Kobayashi et al., 2007).NM: 正常培养基; WSM: 水分胁迫培养基 ... ... Methods used for demonstrating root hydrotropism and screening for mutants altering root hydrotropic response (A) The method Sachs used to demonstrate plant root hydrotropic response (Sachs, 1887); (B) Experiment Jaffe et al. (1985) carried out to prove the existence of the hydrotropic response using Ageotropum mutant; (C) Screening system for identifying mutants with altered hydrotropic response in Cassab’s lab (Eapen et al., 2003; Saucedo et al., 2012); (D), (E) Screening system used in Takahashi’s laboratory for mutants with altered hydrotropic response (Takahashi et al., 2002; Kobayashi et al., 2007). NM: Normal nutrient medium; WSM: Water stress medium ...
4 2012
... 植物根向水性反应实验筛选体系示意图 (A) Sachs发现植物的根具有向水性所用的实验方法(Sachs, 1887); (B) Jaffe及其同事证明植物根向水性存在所用的实验方法(Jaffe et al., 1985); (C) 墨西哥Cassab实验室的根向水性筛选系统(Eapen et al., 2003; Saucedo et al., 2012); (D), (E) 日本东北大学Takahashi实验室的根向水性筛选系统(Takahashi et al., 2002; Kobayashi et al., 2007).NM: 正常培养基; WSM: 水分胁迫培养基 ... ... Methods used for demonstrating root hydrotropism and screening for mutants altering root hydrotropic response (A) The method Sachs used to demonstrate plant root hydrotropic response (Sachs, 1887); (B) Experiment Jaffe et al. (1985) carried out to prove the existence of the hydrotropic response using Ageotropum mutant; (C) Screening system for identifying mutants with altered hydrotropic response in Cassab’s lab (Eapen et al., 2003; Saucedo et al., 2012); (D), (E) Screening system used in Takahashi’s laboratory for mutants with altered hydrotropic response (Takahashi et al., 2002; Kobayashi et al., 2007). NM: Normal nutrient medium; WSM: Water stress medium ... ... 进入21世纪后, 日本东北大学的Takahashi实验室和墨西哥国立自治大学的Cassab实验室开始利用分子遗传学手段研究根向水性反应的分子机制.Cassab实验室将培养基分成上下两部分, 其中上半部分为正常1/2MS培养基(normal nutrient medium, NM), 下半部分则是在正常培养基的基础上添加了渗透调节剂(如甘油或海藻酸)来降低培养基的水分含量(water stress medium, WSM), 进而人为地造成上下水势梯度.将萌发后的拟南芥幼苗放在上半部分离界面1 cm左右的培养基上垂直培养, 当根生长到接近两种培养基界面时, 野生型植物的根会克服向地性而向上弯曲生长, 说明其对水势梯度敏感; 而失去向水性生长的突变体会继续向下生长, 说明其对水势梯度不敏感(图1C).利用此方法, Cassab实验室分离到两个向水性缺陷突变体, 分别命名为nhr1 (no hydrotropic response 1)和ahr1 (altered hydrotropic response 1) (Eapen et al., 2003; Saucedo et al., 2012), 这两个突变体都呈现半显性遗传的特点, 但造成二者表型的突变基因至今未被鉴定. ... ... 2012年, Cassab实验室指出细胞分裂素在植物根的向水性反应中发挥重要作用; 并且通过实验证明细胞分裂素能够恢复ahr1根的向水性表型, 且这种恢复不是通过改变其向地性来完成(Saucedo et al., 2012).此外, 细胞分裂素还可以调节生长素的合成(Jones et al., 2010).因此, Cassab实验室认为, 细胞分裂素参与植物根的向水性反应可能通过调节根中生长素的水平实现.目前, 细胞分裂素在植物根向水性反应中的作用机制尚不明确. ...
1 1960
... 早期的植物生理学研究中, Porta (1658)和Knight (1811)等科学家通过大量的实验证明, 植物根系会随着生长环境中的水势梯度而改变其生长方向, 由此产生了对植物根向水性的研究.此外, 达尔文所著的《The Power of Movement in Plants》一书中也有关于向水性的描述(Darwin and Darwin, 1881).德国植物学家Sachs (1887)将由网格布包裹的潮湿木屑斜向悬挂, 让豌豆(Pisum sativum)种子在木屑表面萌发, 当豌豆的根因向地性生长至网格布下边缘时, 会沿网格布表面弯曲生长而不会进入干燥的空气中, 据此他认为植物的根存在向水性生长(图1A) (Sachs, 1887).另外, 研究还发现, 土壤中的水分会随着土壤深度、季节和降水等因素而分布不均, 产生水势梯度, 而且这种水势梯度会诱导根系向水性生长(Garwood and Williams,1967; Hsiao and Xu, 2000; Steudle, 2000).然而, 由于其它植物向性的干扰, 向水性反应一直以来未被科学家认可.直到1985年, Jaffe及其同事利用豌豆突变体“Ageotropum”为实验材料才首次强有力地证明了植物根向水性不依赖于向地性而独立存在(Jaffe et al., 1985).此豌豆突变体的根完全失去了向地性和避光性反应, 因此其部分根会向上生长出土壤(Blixt et al.,1958; Schöldéen and Burst- röm, 1960; Ekelund and Hemberg, 1966; Wilson, 1967).在一个湿度可控的玻璃盒子中培养该突变体植物, 当盒子中空气的相对湿度(relative humidity of the air, RH)达到95%-98%时, Ageotropum的根生长出土壤后不会弯曲, 而是向着湿润的空气中继续生长(图1B); 当空气相对湿度调整为85%-90%时, 原来向上生长的Ageotropum根会迅速向下弯曲生长直至进入比空气湿润的土壤里.这一现象说明, Ageotro- pum尽管缺失了向地性和避光性, 但其根仍然能感知环境中的水势梯度, 并具有正常的向水性生长反应.除了利用失去了向地性的突变体来研究植物根的向水性外, 科学家们还用竖直旋转的转盘来固定植物材料(Hoson et al., 1997; Kraft et al., 2000)进行研究, 发现竖直方向低速旋转培养植物的根的向地性被有效抵消.用此方法对豌豆以及黄瓜(Cucumis sativus)进行了研究, 发现它们的向水性表型更加明显(Takah- ashi et al., 1996, 1999).Takahashi及其同事发现, 在航天器失重条件下, 黄瓜的侧根也表现出更强烈地向水性反应, 而在正常培养的实验中侧根的向水性反应不易被观察到, 说明去除向地性影响后, 植物根对水势梯度的反应更明显(Takahashi et al.,1999). ...
1 2010
... 植物在整个生命活动过程中都在被动地适应不断变化的环境.在长期的进化选择中, 植物进化出一系列特有的机制来适应周围环境的变化, 使其能够一代代繁衍生息.植物能够感知自身生长环境中各种非生物(包括光、重力、水、温度、氧气和二氧化碳等)和生物信号, 并做出相应的生理生化反应.向性反应(tropic response)作为植物适应周围环境变化的重要机制之一, 是植物实现自身趋利避害的主要途径.植物朝向环境信号生长的向性反应称为正(positive)向性反应, 反之则称为负(negative)向性反应.植物为了应对不同的环境信号进化出了一系列向性反应, 包括向光性(phototropism) (Van Overbeek,1939; Gehr- ing et al., 1990; Liscum et al., 2014)、向地性(gravi- tropism) (Gehring et al., 1990; Hart, 1990; Taka- hashi and Jaffe, 1990; Correll and Kiss, 2002)、向化性(chemotropism) (Esmon et al., 2005)、向触性(thigmotropism) (Monshausen et al., 2008)、向水性(hydrotropism) (Takahashi and Jaffe, 1990; Takaha- shi, 1997)、避盐性(halotropism) (Munns and Tes- ter, 2008; Sun et al., 2008; Shelef et al., 2010; Galvan-Ampudia et al., 2013)及向声性(phonotro pism) (Mishra et al., 2016; Rodrigo-Moreno et al., 2017)等.其中, 向地性和向光性由于较易观察和筛选等原因, 研究进展较快, 目前已对调控植物向地性和向光性的分子机理有了较为深入的认识.由于重力等因素的干扰, 根向水性的研究较为困难.目前的研究仅发现少数基因和突变体参与或改变植物的向水性反应.向水性机理研究不仅具有重要的理论意义, 而且对节水灌溉和提高植物的抗旱性等也具有指导意义. ...
3 2016
... 生长素的Cholodny-Went假说认为, 植物根在受到重力刺激时, 会导致生长素在其根弯曲的凹面处大量积累, 生长素的累积会抑制该侧细胞的伸长, 而根的另一侧(弯曲的凸面)细胞仍能伸长, 从而导致根发生向地性弯曲生长.生长素的这种不对称分布是通过生长素的极性运输实现(Rashotte et al., 2001).然而, 目前研究认为该假说并不能用来解释植物根的向水性弯曲生长(Shkolnik et al., 2016; Shkolnik and Fromm, 2016). ... ... ; Shkolnik and Fromm, 2016). ... ... 1991年, Takahashi在对豌豆突变体“Ageotro- pum”的向水性研究中发现, 当对Ageotropum施加生长素转运抑制剂三碘苯甲酸(TIBA)时, 其根的向水性反应被明显抑制, 说明生长素的极性运输在植物根的向水性反应过程中具有重要作用(Takahashi and Suge, 1991).然而生长素的调节效应具有种间差异.在豌豆、黄瓜和水稻(Oryza sativa)根尖, 生长素以重新分布的模式调控其根的向地性以及向水性弯曲(Takahashi and Suge, 1991; Mizuno et al., 2002; Nakajima et al., 2017); 但与豌豆同属豆科植物的百脉根(Lotus corniculatus)和十字花科的拟南芥根向水性反应却不依赖于生长素的极性运输与重新分配(Shkolnik et al., 2016; Nakajima et al., 2017).研究表明, 生长素转运抑制剂(CHPAA、NPA和TIBA)不会影响百脉根和拟南芥根的向水性反应(Kaneyasu et al., 2007; Nakajima et al., 2017), 但生长素合成抑制剂Kyn会明显减弱百脉根的向水性反应(Nakajima et al., 2017).此外, PINs是植物体内调节生长素运输方向的蛋白, 它们以极性定位的方式参与生长素在植物细胞间的极性运输(Michniewicz et al., 2007).拟南芥中, PIN2缺失突变体表现出向地性反应缺陷, 但却具有正常的向水性反应(Takahashi et al., 2002).生长素信号通路受损的拟南芥突变体axr1-3和axr2-1根的向地性反应虽然减弱(Leyser et al., 1993; Nagpal et al., 2000), 但其根对水势梯度响应的敏感性增强(Takahashi et al., 2002).研究发现, 超表达MIZ1能增强拟南芥根的向水性反应, 同时其根中生长素合成基因的表达水平明显降低(Moriwaki et al., 2011; Miyazawa et al., 2012).总之, 生长素通过极性运输和重新分布的方式参与植物根的向地性反应, 但该作用机理并不适用于所有植物(如百脉根和拟南芥)根的向水性反应, 有关生长素对向水性反应调控的种间差异及生理和分子基础有待进一步研究. ...
0 2016
1 1957
... 在植物根系的生长过程中, 细胞分裂素作为重要的植物调节激素被广泛认知(Mok and Mok, 2001; Wer- ner and Schmülling, 2009).细胞分裂素不仅可以抑制植物根的伸长和分化, 还参与植物根对干旱胁迫的响应(Skoog and Miller, 1957; Cary et al., 1995; Werner et al., 2003, 2010).研究发现, 在植物根的一侧施加浓度较高的细胞分裂素会使根克服重力朝着施加位点弯曲生长(Aloni et al., 2004), 说明细胞分裂素在植物根的向地性反应中可能起着负向调节作用.此外, 有研究表明, 细胞分裂素不仅可以下调植物根中生长素的极性运输, 而且能刺激根局部区域生长素的合成, 从而参与调节植物根的向地性生长(Zhou et al., 2011). ...
1 2000
... 早期的植物生理学研究中, Porta (1658)和Knight (1811)等科学家通过大量的实验证明, 植物根系会随着生长环境中的水势梯度而改变其生长方向, 由此产生了对植物根向水性的研究.此外, 达尔文所著的《The Power of Movement in Plants》一书中也有关于向水性的描述(Darwin and Darwin, 1881).德国植物学家Sachs (1887)将由网格布包裹的潮湿木屑斜向悬挂, 让豌豆(Pisum sativum)种子在木屑表面萌发, 当豌豆的根因向地性生长至网格布下边缘时, 会沿网格布表面弯曲生长而不会进入干燥的空气中, 据此他认为植物的根存在向水性生长(图1A) (Sachs, 1887).另外, 研究还发现, 土壤中的水分会随着土壤深度、季节和降水等因素而分布不均, 产生水势梯度, 而且这种水势梯度会诱导根系向水性生长(Garwood and Williams,1967; Hsiao and Xu, 2000; Steudle, 2000).然而, 由于其它植物向性的干扰, 向水性反应一直以来未被科学家认可.直到1985年, Jaffe及其同事利用豌豆突变体“Ageotropum”为实验材料才首次强有力地证明了植物根向水性不依赖于向地性而独立存在(Jaffe et al., 1985).此豌豆突变体的根完全失去了向地性和避光性反应, 因此其部分根会向上生长出土壤(Blixt et al.,1958; Schöldéen and Burst- röm, 1960; Ekelund and Hemberg, 1966; Wilson, 1967).在一个湿度可控的玻璃盒子中培养该突变体植物, 当盒子中空气的相对湿度(relative humidity of the air, RH)达到95%-98%时, Ageotropum的根生长出土壤后不会弯曲, 而是向着湿润的空气中继续生长(图1B); 当空气相对湿度调整为85%-90%时, 原来向上生长的Ageotropum根会迅速向下弯曲生长直至进入比空气湿润的土壤里.这一现象说明, Ageotro- pum尽管缺失了向地性和避光性, 但其根仍然能感知环境中的水势梯度, 并具有正常的向水性生长反应.除了利用失去了向地性的突变体来研究植物根的向水性外, 科学家们还用竖直旋转的转盘来固定植物材料(Hoson et al., 1997; Kraft et al., 2000)进行研究, 发现竖直方向低速旋转培养植物的根的向地性被有效抵消.用此方法对豌豆以及黄瓜(Cucumis sativus)进行了研究, 发现它们的向水性表型更加明显(Takah- ashi et al., 1996, 1999).Takahashi及其同事发现, 在航天器失重条件下, 黄瓜的侧根也表现出更强烈地向水性反应, 而在正常培养的实验中侧根的向水性反应不易被观察到, 说明去除向地性影响后, 植物根对水势梯度的反应更明显(Takahashi et al.,1999). ...
2 2008
... 植物在整个生命活动过程中都在被动地适应不断变化的环境.在长期的进化选择中, 植物进化出一系列特有的机制来适应周围环境的变化, 使其能够一代代繁衍生息.植物能够感知自身生长环境中各种非生物(包括光、重力、水、温度、氧气和二氧化碳等)和生物信号, 并做出相应的生理生化反应.向性反应(tropic response)作为植物适应周围环境变化的重要机制之一, 是植物实现自身趋利避害的主要途径.植物朝向环境信号生长的向性反应称为正(positive)向性反应, 反之则称为负(negative)向性反应.植物为了应对不同的环境信号进化出了一系列向性反应, 包括向光性(phototropism) (Van Overbeek,1939; Gehr- ing et al., 1990; Liscum et al., 2014)、向地性(gravi- tropism) (Gehring et al., 1990; Hart, 1990; Taka- hashi and Jaffe, 1990; Correll and Kiss, 2002)、向化性(chemotropism) (Esmon et al., 2005)、向触性(thigmotropism) (Monshausen et al., 2008)、向水性(hydrotropism) (Takahashi and Jaffe, 1990; Takaha- shi, 1997)、避盐性(halotropism) (Munns and Tes- ter, 2008; Sun et al., 2008; Shelef et al., 2010; Galvan-Ampudia et al., 2013)及向声性(phonotro pism) (Mishra et al., 2016; Rodrigo-Moreno et al., 2017)等.其中, 向地性和向光性由于较易观察和筛选等原因, 研究进展较快, 目前已对调控植物向地性和向光性的分子机理有了较为深入的认识.由于重力等因素的干扰, 根向水性的研究较为困难.目前的研究仅发现少数基因和突变体参与或改变植物的向水性反应.向水性机理研究不仅具有重要的理论意义, 而且对节水灌溉和提高植物的抗旱性等也具有指导意义. ... ... 植物根柱细胞中的淀粉粒在根的重力感知中起重要作用(Kiss, 2000; Perrin et al., 2005).研究发现, 拟南芥(Arabidopsis thaliana)主根根柱细胞中的淀粉粒在水势梯度刺激后会迅速降解, 导致根的重力敏感性降低, 同时促进其向水性反应(Takahashi et al., 2003).但根柱细胞内淀粉粒的降解不仅受到水势梯度的诱导, 而且也受到其它不同胁迫(如干旱)的调控 (Takahashi et al., 2003; Sun et al., 2008; Ding and Friml, 2010; Iyer-Pascuzzi et al., 2011).因此, 尚不能确定向水性反应中的淀粉粒降解是由水势梯度特异诱导的还是受干旱诱导发生的.而且在一些植物中, 干旱会促进根的向地性反应(Leopold and LaFavre, 1989), 说明淀粉粒的降解可能对诱导植物根的向水性反应并非必需(Nakayama et al., 2012). ...
... 植物在整个生命活动过程中都在被动地适应不断变化的环境.在长期的进化选择中, 植物进化出一系列特有的机制来适应周围环境的变化, 使其能够一代代繁衍生息.植物能够感知自身生长环境中各种非生物(包括光、重力、水、温度、氧气和二氧化碳等)和生物信号, 并做出相应的生理生化反应.向性反应(tropic response)作为植物适应周围环境变化的重要机制之一, 是植物实现自身趋利避害的主要途径.植物朝向环境信号生长的向性反应称为正(positive)向性反应, 反之则称为负(negative)向性反应.植物为了应对不同的环境信号进化出了一系列向性反应, 包括向光性(phototropism) (Van Overbeek,1939; Gehr- ing et al., 1990; Liscum et al., 2014)、向地性(gravi- tropism) (Gehring et al., 1990; Hart, 1990; Taka- hashi and Jaffe, 1990; Correll and Kiss, 2002)、向化性(chemotropism) (Esmon et al., 2005)、向触性(thigmotropism) (Monshausen et al., 2008)、向水性(hydrotropism) (Takahashi and Jaffe, 1990; Takaha- shi, 1997)、避盐性(halotropism) (Munns and Tes- ter, 2008; Sun et al., 2008; Shelef et al., 2010; Galvan-Ampudia et al., 2013)及向声性(phonotro pism) (Mishra et al., 2016; Rodrigo-Moreno et al., 2017)等.其中, 向地性和向光性由于较易观察和筛选等原因, 研究进展较快, 目前已对调控植物向地性和向光性的分子机理有了较为深入的认识.由于重力等因素的干扰, 根向水性的研究较为困难.目前的研究仅发现少数基因和突变体参与或改变植物的向水性反应.向水性机理研究不仅具有重要的理论意义, 而且对节水灌溉和提高植物的抗旱性等也具有指导意义. ...
5 2002
... 植物根向水性反应实验筛选体系示意图 (A) Sachs发现植物的根具有向水性所用的实验方法(Sachs, 1887); (B) Jaffe及其同事证明植物根向水性存在所用的实验方法(Jaffe et al., 1985); (C) 墨西哥Cassab实验室的根向水性筛选系统(Eapen et al., 2003; Saucedo et al., 2012); (D), (E) 日本东北大学Takahashi实验室的根向水性筛选系统(Takahashi et al., 2002; Kobayashi et al., 2007).NM: 正常培养基; WSM: 水分胁迫培养基 ... ... Methods used for demonstrating root hydrotropism and screening for mutants altering root hydrotropic response (A) The method Sachs used to demonstrate plant root hydrotropic response (Sachs, 1887); (B) Experiment Jaffe et al. (1985) carried out to prove the existence of the hydrotropic response using Ageotropum mutant; (C) Screening system for identifying mutants with altered hydrotropic response in Cassab’s lab (Eapen et al., 2003; Saucedo et al., 2012); (D), (E) Screening system used in Takahashi’s laboratory for mutants with altered hydrotropic response (Takahashi et al., 2002; Kobayashi et al., 2007). NM: Normal nutrient medium; WSM: Water stress medium ... ... 1991年, Takahashi在对豌豆突变体“Ageotro- pum”的向水性研究中发现, 当对Ageotropum施加生长素转运抑制剂三碘苯甲酸(TIBA)时, 其根的向水性反应被明显抑制, 说明生长素的极性运输在植物根的向水性反应过程中具有重要作用(Takahashi and Suge, 1991).然而生长素的调节效应具有种间差异.在豌豆、黄瓜和水稻(Oryza sativa)根尖, 生长素以重新分布的模式调控其根的向地性以及向水性弯曲(Takahashi and Suge, 1991; Mizuno et al., 2002; Nakajima et al., 2017); 但与豌豆同属豆科植物的百脉根(Lotus corniculatus)和十字花科的拟南芥根向水性反应却不依赖于生长素的极性运输与重新分配(Shkolnik et al., 2016; Nakajima et al., 2017).研究表明, 生长素转运抑制剂(CHPAA、NPA和TIBA)不会影响百脉根和拟南芥根的向水性反应(Kaneyasu et al., 2007; Nakajima et al., 2017), 但生长素合成抑制剂Kyn会明显减弱百脉根的向水性反应(Nakajima et al., 2017).此外, PINs是植物体内调节生长素运输方向的蛋白, 它们以极性定位的方式参与生长素在植物细胞间的极性运输(Michniewicz et al., 2007).拟南芥中, PIN2缺失突变体表现出向地性反应缺陷, 但却具有正常的向水性反应(Takahashi et al., 2002).生长素信号通路受损的拟南芥突变体axr1-3和axr2-1根的向地性反应虽然减弱(Leyser et al., 1993; Nagpal et al., 2000), 但其根对水势梯度响应的敏感性增强(Takahashi et al., 2002).研究发现, 超表达MIZ1能增强拟南芥根的向水性反应, 同时其根中生长素合成基因的表达水平明显降低(Moriwaki et al., 2011; Miyazawa et al., 2012).总之, 生长素通过极性运输和重新分布的方式参与植物根的向地性反应, 但该作用机理并不适用于所有植物(如百脉根和拟南芥)根的向水性反应, 有关生长素对向水性反应调控的种间差异及生理和分子基础有待进一步研究. ... ... ), 但其根对水势梯度响应的敏感性增强(Takahashi et al., 2002).研究发现, 超表达MIZ1能增强拟南芥根的向水性反应, 同时其根中生长素合成基因的表达水平明显降低(Moriwaki et al., 2011; Miyazawa et al., 2012).总之, 生长素通过极性运输和重新分布的方式参与植物根的向地性反应, 但该作用机理并不适用于所有植物(如百脉根和拟南芥)根的向水性反应, 有关生长素对向水性反应调控的种间差异及生理和分子基础有待进一步研究. ... ... 日本Takahashi实验室利用新的实验体系(图1E), 发现拟南芥脱落酸合成缺失突变体aba1-1 (Koorn- neef et al., 1982)以及脱落酸不敏感突变体abi2-1 (Koornneef et al., 1984)对水势梯度不敏感, 但二者的根呈现出与野生型一致的向地性弯曲, 由此认为脱落酸能促进拟南芥根的向水性弯曲(Takahashi et al., 2002).随后的实验证明, 脱落酸可以通过调节MIZ1的表达来参与植物的向水性反应(Moriwaki et al., 2012).脱落酸信号途径正调控因子SnRK2.2及SnRK2.3的双重缺失突变体snrk2.2/snrk2.3表现出向水性反应降低的表型, 说明脱落酸正向调节拟南芥根的向水性反应.同时, 该研究还表明, 拟南芥根尖皮层细胞的分裂对向水性非常重要(Dietrich et al., 2017).然而, 脱落酸尽管可以恢复ahr1对水势梯度感知的缺陷, 但却加剧了nhr1向水性反应缺失的表型(Ponce et al., 2008; Moriwaki et al., 2012).墨西哥Cassab实验室对nhr1突变体脱落酸的定量分析显示, nhr1突变会造成植物体内脱落酸含量升高.再者, 脱落酸合成抑制剂AbamineSG会恢复nhr1向水性缺失的表型, 说明在nhr1参与的向水性反应中, 脱落酸可能起着负向调控作用(Quiroz-Figueroa et al., 2010). ...
2 1990
... 植物在整个生命活动过程中都在被动地适应不断变化的环境.在长期的进化选择中, 植物进化出一系列特有的机制来适应周围环境的变化, 使其能够一代代繁衍生息.植物能够感知自身生长环境中各种非生物(包括光、重力、水、温度、氧气和二氧化碳等)和生物信号, 并做出相应的生理生化反应.向性反应(tropic response)作为植物适应周围环境变化的重要机制之一, 是植物实现自身趋利避害的主要途径.植物朝向环境信号生长的向性反应称为正(positive)向性反应, 反之则称为负(negative)向性反应.植物为了应对不同的环境信号进化出了一系列向性反应, 包括向光性(phototropism) (Van Overbeek,1939; Gehr- ing et al., 1990; Liscum et al., 2014)、向地性(gravi- tropism) (Gehring et al., 1990; Hart, 1990; Taka- hashi and Jaffe, 1990; Correll and Kiss, 2002)、向化性(chemotropism) (Esmon et al., 2005)、向触性(thigmotropism) (Monshausen et al., 2008)、向水性(hydrotropism) (Takahashi and Jaffe, 1990; Takaha- shi, 1997)、避盐性(halotropism) (Munns and Tes- ter, 2008; Sun et al., 2008; Shelef et al., 2010; Galvan-Ampudia et al., 2013)及向声性(phonotro pism) (Mishra et al., 2016; Rodrigo-Moreno et al., 2017)等.其中, 向地性和向光性由于较易观察和筛选等原因, 研究进展较快, 目前已对调控植物向地性和向光性的分子机理有了较为深入的认识.由于重力等因素的干扰, 根向水性的研究较为困难.目前的研究仅发现少数基因和突变体参与或改变植物的向水性反应.向水性机理研究不仅具有重要的理论意义, 而且对节水灌溉和提高植物的抗旱性等也具有指导意义. ... ... )、向水性(hydrotropism) (Takahashi and Jaffe, 1990; Takaha- shi, 1997)、避盐性(halotropism) (Munns and Tes- ter, 2008; Sun et al., 2008; Shelef et al., 2010; Galvan-Ampudia et al., 2013)及向声性(phonotro pism) (Mishra et al., 2016; Rodrigo-Moreno et al., 2017)等.其中, 向地性和向光性由于较易观察和筛选等原因, 研究进展较快, 目前已对调控植物向地性和向光性的分子机理有了较为深入的认识.由于重力等因素的干扰, 根向水性的研究较为困难.目前的研究仅发现少数基因和突变体参与或改变植物的向水性反应.向水性机理研究不仅具有重要的理论意义, 而且对节水灌溉和提高植物的抗旱性等也具有指导意义. ...
2 1999
... 早期的植物生理学研究中, Porta (1658)和Knight (1811)等科学家通过大量的实验证明, 植物根系会随着生长环境中的水势梯度而改变其生长方向, 由此产生了对植物根向水性的研究.此外, 达尔文所著的《The Power of Movement in Plants》一书中也有关于向水性的描述(Darwin and Darwin, 1881).德国植物学家Sachs (1887)将由网格布包裹的潮湿木屑斜向悬挂, 让豌豆(Pisum sativum)种子在木屑表面萌发, 当豌豆的根因向地性生长至网格布下边缘时, 会沿网格布表面弯曲生长而不会进入干燥的空气中, 据此他认为植物的根存在向水性生长(图1A) (Sachs, 1887).另外, 研究还发现, 土壤中的水分会随着土壤深度、季节和降水等因素而分布不均, 产生水势梯度, 而且这种水势梯度会诱导根系向水性生长(Garwood and Williams,1967; Hsiao and Xu, 2000; Steudle, 2000).然而, 由于其它植物向性的干扰, 向水性反应一直以来未被科学家认可.直到1985年, Jaffe及其同事利用豌豆突变体“Ageotropum”为实验材料才首次强有力地证明了植物根向水性不依赖于向地性而独立存在(Jaffe et al., 1985).此豌豆突变体的根完全失去了向地性和避光性反应, 因此其部分根会向上生长出土壤(Blixt et al.,1958; Schöldéen and Burst- röm, 1960; Ekelund and Hemberg, 1966; Wilson, 1967).在一个湿度可控的玻璃盒子中培养该突变体植物, 当盒子中空气的相对湿度(relative humidity of the air, RH)达到95%-98%时, Ageotropum的根生长出土壤后不会弯曲, 而是向着湿润的空气中继续生长(图1B); 当空气相对湿度调整为85%-90%时, 原来向上生长的Ageotropum根会迅速向下弯曲生长直至进入比空气湿润的土壤里.这一现象说明, Ageotro- pum尽管缺失了向地性和避光性, 但其根仍然能感知环境中的水势梯度, 并具有正常的向水性生长反应.除了利用失去了向地性的突变体来研究植物根的向水性外, 科学家们还用竖直旋转的转盘来固定植物材料(Hoson et al., 1997; Kraft et al., 2000)进行研究, 发现竖直方向低速旋转培养植物的根的向地性被有效抵消.用此方法对豌豆以及黄瓜(Cucumis sativus)进行了研究, 发现它们的向水性表型更加明显(Takah- ashi et al., 1996, 1999).Takahashi及其同事发现, 在航天器失重条件下, 黄瓜的侧根也表现出更强烈地向水性反应, 而在正常培养的实验中侧根的向水性反应不易被观察到, 说明去除向地性影响后, 植物根对水势梯度的反应更明显(Takahashi et al.,1999). ... ... ).Takahashi及其同事发现, 在航天器失重条件下, 黄瓜的侧根也表现出更强烈地向水性反应, 而在正常培养的实验中侧根的向水性反应不易被观察到, 说明去除向地性影响后, 植物根对水势梯度的反应更明显(Takahashi et al.,1999). ...
... 1991年, Takahashi在对豌豆突变体“Ageotro- pum”的向水性研究中发现, 当对Ageotropum施加生长素转运抑制剂三碘苯甲酸(TIBA)时, 其根的向水性反应被明显抑制, 说明生长素的极性运输在植物根的向水性反应过程中具有重要作用(Takahashi and Suge, 1991).然而生长素的调节效应具有种间差异.在豌豆、黄瓜和水稻(Oryza sativa)根尖, 生长素以重新分布的模式调控其根的向地性以及向水性弯曲(Takahashi and Suge, 1991; Mizuno et al., 2002; Nakajima et al., 2017); 但与豌豆同属豆科植物的百脉根(Lotus corniculatus)和十字花科的拟南芥根向水性反应却不依赖于生长素的极性运输与重新分配(Shkolnik et al., 2016; Nakajima et al., 2017).研究表明, 生长素转运抑制剂(CHPAA、NPA和TIBA)不会影响百脉根和拟南芥根的向水性反应(Kaneyasu et al., 2007; Nakajima et al., 2017), 但生长素合成抑制剂Kyn会明显减弱百脉根的向水性反应(Nakajima et al., 2017).此外, PINs是植物体内调节生长素运输方向的蛋白, 它们以极性定位的方式参与生长素在植物细胞间的极性运输(Michniewicz et al., 2007).拟南芥中, PIN2缺失突变体表现出向地性反应缺陷, 但却具有正常的向水性反应(Takahashi et al., 2002).生长素信号通路受损的拟南芥突变体axr1-3和axr2-1根的向地性反应虽然减弱(Leyser et al., 1993; Nagpal et al., 2000), 但其根对水势梯度响应的敏感性增强(Takahashi et al., 2002).研究发现, 超表达MIZ1能增强拟南芥根的向水性反应, 同时其根中生长素合成基因的表达水平明显降低(Moriwaki et al., 2011; Miyazawa et al., 2012).总之, 生长素通过极性运输和重新分布的方式参与植物根的向地性反应, 但该作用机理并不适用于所有植物(如百脉根和拟南芥)根的向水性反应, 有关生长素对向水性反应调控的种间差异及生理和分子基础有待进一步研究. ... ... )根尖, 生长素以重新分布的模式调控其根的向地性以及向水性弯曲(Takahashi and Suge, 1991; Mizuno et al., 2002; Nakajima et al., 2017); 但与豌豆同属豆科植物的百脉根(Lotus corniculatus)和十字花科的拟南芥根向水性反应却不依赖于生长素的极性运输与重新分配(Shkolnik et al., 2016; Nakajima et al., 2017).研究表明, 生长素转运抑制剂(CHPAA、NPA和TIBA)不会影响百脉根和拟南芥根的向水性反应(Kaneyasu et al., 2007; Nakajima et al., 2017), 但生长素合成抑制剂Kyn会明显减弱百脉根的向水性反应(Nakajima et al., 2017).此外, PINs是植物体内调节生长素运输方向的蛋白, 它们以极性定位的方式参与生长素在植物细胞间的极性运输(Michniewicz et al., 2007).拟南芥中, PIN2缺失突变体表现出向地性反应缺陷, 但却具有正常的向水性反应(Takahashi et al., 2002).生长素信号通路受损的拟南芥突变体axr1-3和axr2-1根的向地性反应虽然减弱(Leyser et al., 1993; Nagpal et al., 2000), 但其根对水势梯度响应的敏感性增强(Takahashi et al., 2002).研究发现, 超表达MIZ1能增强拟南芥根的向水性反应, 同时其根中生长素合成基因的表达水平明显降低(Moriwaki et al., 2011; Miyazawa et al., 2012).总之, 生长素通过极性运输和重新分布的方式参与植物根的向地性反应, 但该作用机理并不适用于所有植物(如百脉根和拟南芥)根的向水性反应, 有关生长素对向水性反应调控的种间差异及生理和分子基础有待进一步研究. ...
1 1996
... 早期的植物生理学研究中, Porta (1658)和Knight (1811)等科学家通过大量的实验证明, 植物根系会随着生长环境中的水势梯度而改变其生长方向, 由此产生了对植物根向水性的研究.此外, 达尔文所著的《The Power of Movement in Plants》一书中也有关于向水性的描述(Darwin and Darwin, 1881).德国植物学家Sachs (1887)将由网格布包裹的潮湿木屑斜向悬挂, 让豌豆(Pisum sativum)种子在木屑表面萌发, 当豌豆的根因向地性生长至网格布下边缘时, 会沿网格布表面弯曲生长而不会进入干燥的空气中, 据此他认为植物的根存在向水性生长(图1A) (Sachs, 1887).另外, 研究还发现, 土壤中的水分会随着土壤深度、季节和降水等因素而分布不均, 产生水势梯度, 而且这种水势梯度会诱导根系向水性生长(Garwood and Williams,1967; Hsiao and Xu, 2000; Steudle, 2000).然而, 由于其它植物向性的干扰, 向水性反应一直以来未被科学家认可.直到1985年, Jaffe及其同事利用豌豆突变体“Ageotropum”为实验材料才首次强有力地证明了植物根向水性不依赖于向地性而独立存在(Jaffe et al., 1985).此豌豆突变体的根完全失去了向地性和避光性反应, 因此其部分根会向上生长出土壤(Blixt et al.,1958; Schöldéen and Burst- röm, 1960; Ekelund and Hemberg, 1966; Wilson, 1967).在一个湿度可控的玻璃盒子中培养该突变体植物, 当盒子中空气的相对湿度(relative humidity of the air, RH)达到95%-98%时, Ageotropum的根生长出土壤后不会弯曲, 而是向着湿润的空气中继续生长(图1B); 当空气相对湿度调整为85%-90%时, 原来向上生长的Ageotropum根会迅速向下弯曲生长直至进入比空气湿润的土壤里.这一现象说明, Ageotro- pum尽管缺失了向地性和避光性, 但其根仍然能感知环境中的水势梯度, 并具有正常的向水性生长反应.除了利用失去了向地性的突变体来研究植物根的向水性外, 科学家们还用竖直旋转的转盘来固定植物材料(Hoson et al., 1997; Kraft et al., 2000)进行研究, 发现竖直方向低速旋转培养植物的根的向地性被有效抵消.用此方法对豌豆以及黄瓜(Cucumis sativus)进行了研究, 发现它们的向水性表型更加明显(Takah- ashi et al., 1996, 1999).Takahashi及其同事发现, 在航天器失重条件下, 黄瓜的侧根也表现出更强烈地向水性反应, 而在正常培养的实验中侧根的向水性反应不易被观察到, 说明去除向地性影响后, 植物根对水势梯度的反应更明显(Takahashi et al.,1999). ...
1 2004
... 在植物根系的生长过程中, 细胞分裂素作为重要的植物调节激素被广泛认知(Mok and Mok, 2001; Wer- ner and Schmülling, 2009).细胞分裂素不仅可以抑制植物根的伸长和分化, 还参与植物根对干旱胁迫的响应(Skoog and Miller, 1957; Cary et al., 1995; Werner et al., 2003, 2010).研究发现, 在植物根的一侧施加浓度较高的细胞分裂素会使根克服重力朝着施加位点弯曲生长(Aloni et al., 2004), 说明细胞分裂素在植物根的向地性反应中可能起着负向调节作用.此外, 有研究表明, 细胞分裂素不仅可以下调植物根中生长素的极性运输, 而且能刺激根局部区域生长素的合成, 从而参与调节植物根的向地性生长(Zhou et al., 2011). ...
1 2011
... 油菜素内酯是一类广泛存在于植物体内的甾醇类激素, 它参与调节植物的生长发育.有报道指出, 油菜素内酯参与拟南芥花序的负向地性反应(Arteca and Arteca, 2011).此外, 油菜素内酯和乙烯在参与调节拟南芥茎的负向地性反应中具相反的作用, 内源油菜素内酯信号会降低植物对乙烯信号的感知从而使拟南芥的茎失去负向地性反应(Vandenbussche et al., 2013).虽然目前的研究确定赤霉素、油菜素内酯和乙烯都参与植物的向地性反应, 但三者是否调节植物根的向水性反应仍然未知. ...
2 2003
... 植物根柱细胞中的淀粉粒在根的重力感知中起重要作用(Kiss, 2000; Perrin et al., 2005).研究发现, 拟南芥(Arabidopsis thaliana)主根根柱细胞中的淀粉粒在水势梯度刺激后会迅速降解, 导致根的重力敏感性降低, 同时促进其向水性反应(Takahashi et al., 2003).但根柱细胞内淀粉粒的降解不仅受到水势梯度的诱导, 而且也受到其它不同胁迫(如干旱)的调控 (Takahashi et al., 2003; Sun et al., 2008; Ding and Friml, 2010; Iyer-Pascuzzi et al., 2011).因此, 尚不能确定向水性反应中的淀粉粒降解是由水势梯度特异诱导的还是受干旱诱导发生的.而且在一些植物中, 干旱会促进根的向地性反应(Leopold and LaFavre, 1989), 说明淀粉粒的降解可能对诱导植物根的向水性反应并非必需(Nakayama et al., 2012). ... ... ).但根柱细胞内淀粉粒的降解不仅受到水势梯度的诱导, 而且也受到其它不同胁迫(如干旱)的调控 (Takahashi et al., 2003; Sun et al., 2008; Ding and Friml, 2010; Iyer-Pascuzzi et al., 2011).因此, 尚不能确定向水性反应中的淀粉粒降解是由水势梯度特异诱导的还是受干旱诱导发生的.而且在一些植物中, 干旱会促进根的向地性反应(Leopold and LaFavre, 1989), 说明淀粉粒的降解可能对诱导植物根的向水性反应并非必需(Nakayama et al., 2012). ...
1 1997
... 有研究表明, 植物根因重力刺激发生向地性弯曲后, 弯曲根的凹面处活性氧(reactive oxygen species, ROS)积累增加, 从而抑制凹面处的生长, 但这种根中ROS的单侧积累并不会发生在向水性弯曲生长过程中(Krieger et al., 2016).另外, 提高植物根中细胞质的钙离子浓度有助于根对水势梯度的感知, 但其作用机制并不清楚(Takano et al., 1997). ...
1 1939
... 植物在整个生命活动过程中都在被动地适应不断变化的环境.在长期的进化选择中, 植物进化出一系列特有的机制来适应周围环境的变化, 使其能够一代代繁衍生息.植物能够感知自身生长环境中各种非生物(包括光、重力、水、温度、氧气和二氧化碳等)和生物信号, 并做出相应的生理生化反应.向性反应(tropic response)作为植物适应周围环境变化的重要机制之一, 是植物实现自身趋利避害的主要途径.植物朝向环境信号生长的向性反应称为正(positive)向性反应, 反之则称为负(negative)向性反应.植物为了应对不同的环境信号进化出了一系列向性反应, 包括向光性(phototropism) (Van Overbeek,1939; Gehr- ing et al., 1990; Liscum et al., 2014)、向地性(gravi- tropism) (Gehring et al., 1990; Hart, 1990; Taka- hashi and Jaffe, 1990; Correll and Kiss, 2002)、向化性(chemotropism) (Esmon et al., 2005)、向触性(thigmotropism) (Monshausen et al., 2008)、向水性(hydrotropism) (Takahashi and Jaffe, 1990; Takaha- shi, 1997)、避盐性(halotropism) (Munns and Tes- ter, 2008; Sun et al., 2008; Shelef et al., 2010; Galvan-Ampudia et al., 2013)及向声性(phonotro pism) (Mishra et al., 2016; Rodrigo-Moreno et al., 2017)等.其中, 向地性和向光性由于较易观察和筛选等原因, 研究进展较快, 目前已对调控植物向地性和向光性的分子机理有了较为深入的认识.由于重力等因素的干扰, 根向水性的研究较为困难.目前的研究仅发现少数基因和突变体参与或改变植物的向水性反应.向水性机理研究不仅具有重要的理论意义, 而且对节水灌溉和提高植物的抗旱性等也具有指导意义. ...
1 2013
... 油菜素内酯是一类广泛存在于植物体内的甾醇类激素, 它参与调节植物的生长发育.有报道指出, 油菜素内酯参与拟南芥花序的负向地性反应(Arteca and Arteca, 2011).此外, 油菜素内酯和乙烯在参与调节拟南芥茎的负向地性反应中具相反的作用, 内源油菜素内酯信号会降低植物对乙烯信号的感知从而使拟南芥的茎失去负向地性反应(Vandenbussche et al., 2013).虽然目前的研究确定赤霉素、油菜素内酯和乙烯都参与植物的向地性反应, 但三者是否调节植物根的向水性反应仍然未知. ...
1 2014
... 光作为植物根的负向光性诱导因子, 也可影响植物的向水性反应.研究发现, 光影响植物的向水性反应主要通过调节植物体内MIZ1基因的表达来实现, 与生长在白光条件下的植物相比, 黑暗环境下生长的植物根中MIZ1的转录水平降低, 从而降低了其向水性反应(Moriwaki et al., 2012).该实验还证明, 光依赖的MIZ1基因表达受光信号下游转录因子HY5的调控.表明光对植物向水性反应非常重要, 尤其是植物的根暴露在土壤表面或在极浅的土壤中时, 光可以提高植物根中MIZ1的表达, 从而使根发生向水性弯曲生长, 进入土壤深处, 免受干旱胁迫.植物的侧根发生受水势梯度的诱导, 水势梯度较高一侧产生的侧根更多(Bao et al., 2014; Robbins and Dinneny, 2018).此外, 光还能调节MIZ1蛋白在侧根根冠处积累, 进而使侧根也具有感知水势梯度的能力(Moriwaki et al., 2012). ...
1 1958
... 早期的植物生理学研究中, Porta (1658)和Knight (1811)等科学家通过大量的实验证明, 植物根系会随着生长环境中的水势梯度而改变其生长方向, 由此产生了对植物根向水性的研究.此外, 达尔文所著的《The Power of Movement in Plants》一书中也有关于向水性的描述(Darwin and Darwin, 1881).德国植物学家Sachs (1887)将由网格布包裹的潮湿木屑斜向悬挂, 让豌豆(Pisum sativum)种子在木屑表面萌发, 当豌豆的根因向地性生长至网格布下边缘时, 会沿网格布表面弯曲生长而不会进入干燥的空气中, 据此他认为植物的根存在向水性生长(图1A) (Sachs, 1887).另外, 研究还发现, 土壤中的水分会随着土壤深度、季节和降水等因素而分布不均, 产生水势梯度, 而且这种水势梯度会诱导根系向水性生长(Garwood and Williams,1967; Hsiao and Xu, 2000; Steudle, 2000).然而, 由于其它植物向性的干扰, 向水性反应一直以来未被科学家认可.直到1985年, Jaffe及其同事利用豌豆突变体“Ageotropum”为实验材料才首次强有力地证明了植物根向水性不依赖于向地性而独立存在(Jaffe et al., 1985).此豌豆突变体的根完全失去了向地性和避光性反应, 因此其部分根会向上生长出土壤(Blixt et al.,1958; Schöldéen and Burst- röm, 1960; Ekelund and Hemberg, 1966; Wilson, 1967).在一个湿度可控的玻璃盒子中培养该突变体植物, 当盒子中空气的相对湿度(relative humidity of the air, RH)达到95%-98%时, Ageotropum的根生长出土壤后不会弯曲, 而是向着湿润的空气中继续生长(图1B); 当空气相对湿度调整为85%-90%时, 原来向上生长的Ageotropum根会迅速向下弯曲生长直至进入比空气湿润的土壤里.这一现象说明, Ageotro- pum尽管缺失了向地性和避光性, 但其根仍然能感知环境中的水势梯度, 并具有正常的向水性生长反应.除了利用失去了向地性的突变体来研究植物根的向水性外, 科学家们还用竖直旋转的转盘来固定植物材料(Hoson et al., 1997; Kraft et al., 2000)进行研究, 发现竖直方向低速旋转培养植物的根的向地性被有效抵消.用此方法对豌豆以及黄瓜(Cucumis sativus)进行了研究, 发现它们的向水性表型更加明显(Takah- ashi et al., 1996, 1999).Takahashi及其同事发现, 在航天器失重条件下, 黄瓜的侧根也表现出更强烈地向水性反应, 而在正常培养的实验中侧根的向水性反应不易被观察到, 说明去除向地性影响后, 植物根对水势梯度的反应更明显(Takahashi et al.,1999). ...
1 2003
... 在植物根系的生长过程中, 细胞分裂素作为重要的植物调节激素被广泛认知(Mok and Mok, 2001; Wer- ner and Schmülling, 2009).细胞分裂素不仅可以抑制植物根的伸长和分化, 还参与植物根对干旱胁迫的响应(Skoog and Miller, 1957; Cary et al., 1995; Werner et al., 2003, 2010).研究发现, 在植物根的一侧施加浓度较高的细胞分裂素会使根克服重力朝着施加位点弯曲生长(Aloni et al., 2004), 说明细胞分裂素在植物根的向地性反应中可能起着负向调节作用.此外, 有研究表明, 细胞分裂素不仅可以下调植物根中生长素的极性运输, 而且能刺激根局部区域生长素的合成, 从而参与调节植物根的向地性生长(Zhou et al., 2011). ...
1 2010
... 在植物根系的生长过程中, 细胞分裂素作为重要的植物调节激素被广泛认知(Mok and Mok, 2001; Wer- ner and Schmülling, 2009).细胞分裂素不仅可以抑制植物根的伸长和分化, 还参与植物根对干旱胁迫的响应(Skoog and Miller, 1957; Cary et al., 1995; Werner et al., 2003, 2010).研究发现, 在植物根的一侧施加浓度较高的细胞分裂素会使根克服重力朝着施加位点弯曲生长(Aloni et al., 2004), 说明细胞分裂素在植物根的向地性反应中可能起着负向调节作用.此外, 有研究表明, 细胞分裂素不仅可以下调植物根中生长素的极性运输, 而且能刺激根局部区域生长素的合成, 从而参与调节植物根的向地性生长(Zhou et al., 2011). ...
1 2009
... 在植物根系的生长过程中, 细胞分裂素作为重要的植物调节激素被广泛认知(Mok and Mok, 2001; Wer- ner and Schmülling, 2009).细胞分裂素不仅可以抑制植物根的伸长和分化, 还参与植物根对干旱胁迫的响应(Skoog and Miller, 1957; Cary et al., 1995; Werner et al., 2003, 2010).研究发现, 在植物根的一侧施加浓度较高的细胞分裂素会使根克服重力朝着施加位点弯曲生长(Aloni et al., 2004), 说明细胞分裂素在植物根的向地性反应中可能起着负向调节作用.此外, 有研究表明, 细胞分裂素不仅可以下调植物根中生长素的极性运输, 而且能刺激根局部区域生长素的合成, 从而参与调节植物根的向地性生长(Zhou et al., 2011). ...
1 1967
... 早期的植物生理学研究中, Porta (1658)和Knight (1811)等科学家通过大量的实验证明, 植物根系会随着生长环境中的水势梯度而改变其生长方向, 由此产生了对植物根向水性的研究.此外, 达尔文所著的《The Power of Movement in Plants》一书中也有关于向水性的描述(Darwin and Darwin, 1881).德国植物学家Sachs (1887)将由网格布包裹的潮湿木屑斜向悬挂, 让豌豆(Pisum sativum)种子在木屑表面萌发, 当豌豆的根因向地性生长至网格布下边缘时, 会沿网格布表面弯曲生长而不会进入干燥的空气中, 据此他认为植物的根存在向水性生长(图1A) (Sachs, 1887).另外, 研究还发现, 土壤中的水分会随着土壤深度、季节和降水等因素而分布不均, 产生水势梯度, 而且这种水势梯度会诱导根系向水性生长(Garwood and Williams,1967; Hsiao and Xu, 2000; Steudle, 2000).然而, 由于其它植物向性的干扰, 向水性反应一直以来未被科学家认可.直到1985年, Jaffe及其同事利用豌豆突变体“Ageotropum”为实验材料才首次强有力地证明了植物根向水性不依赖于向地性而独立存在(Jaffe et al., 1985).此豌豆突变体的根完全失去了向地性和避光性反应, 因此其部分根会向上生长出土壤(Blixt et al.,1958; Schöldéen and Burst- röm, 1960; Ekelund and Hemberg, 1966; Wilson, 1967).在一个湿度可控的玻璃盒子中培养该突变体植物, 当盒子中空气的相对湿度(relative humidity of the air, RH)达到95%-98%时, Ageotropum的根生长出土壤后不会弯曲, 而是向着湿润的空气中继续生长(图1B); 当空气相对湿度调整为85%-90%时, 原来向上生长的Ageotropum根会迅速向下弯曲生长直至进入比空气湿润的土壤里.这一现象说明, Ageotro- pum尽管缺失了向地性和避光性, 但其根仍然能感知环境中的水势梯度, 并具有正常的向水性生长反应.除了利用失去了向地性的突变体来研究植物根的向水性外, 科学家们还用竖直旋转的转盘来固定植物材料(Hoson et al., 1997; Kraft et al., 2000)进行研究, 发现竖直方向低速旋转培养植物的根的向地性被有效抵消.用此方法对豌豆以及黄瓜(Cucumis sativus)进行了研究, 发现它们的向水性表型更加明显(Takah- ashi et al., 1996, 1999).Takahashi及其同事发现, 在航天器失重条件下, 黄瓜的侧根也表现出更强烈地向水性反应, 而在正常培养的实验中侧根的向水性反应不易被观察到, 说明去除向地性影响后, 植物根对水势梯度的反应更明显(Takahashi et al.,1999). ...
1 1995
... 在植物根系的生长过程中, 细胞分裂素作为重要的植物调节激素被广泛认知(Mok and Mok, 2001; Wer- ner and Schmülling, 2009).细胞分裂素不仅可以抑制植物根的伸长和分化, 还参与植物根对干旱胁迫的响应(Skoog and Miller, 1957; Cary et al., 1995; Werner et al., 2003, 2010).研究发现, 在植物根的一侧施加浓度较高的细胞分裂素会使根克服重力朝着施加位点弯曲生长(Aloni et al., 2004), 说明细胞分裂素在植物根的向地性反应中可能起着负向调节作用.此外, 有研究表明, 细胞分裂素不仅可以下调植物根中生长素的极性运输, 而且能刺激根局部区域生长素的合成, 从而参与调节植物根的向地性生长(Zhou et al., 2011). ...
1 2002
... 植物在整个生命活动过程中都在被动地适应不断变化的环境.在长期的进化选择中, 植物进化出一系列特有的机制来适应周围环境的变化, 使其能够一代代繁衍生息.植物能够感知自身生长环境中各种非生物(包括光、重力、水、温度、氧气和二氧化碳等)和生物信号, 并做出相应的生理生化反应.向性反应(tropic response)作为植物适应周围环境变化的重要机制之一, 是植物实现自身趋利避害的主要途径.植物朝向环境信号生长的向性反应称为正(positive)向性反应, 反之则称为负(negative)向性反应.植物为了应对不同的环境信号进化出了一系列向性反应, 包括向光性(phototropism) (Van Overbeek,1939; Gehr- ing et al., 1990; Liscum et al., 2014)、向地性(gravi- tropism) (Gehring et al., 1990; Hart, 1990; Taka- hashi and Jaffe, 1990; Correll and Kiss, 2002)、向化性(chemotropism) (Esmon et al., 2005)、向触性(thigmotropism) (Monshausen et al., 2008)、向水性(hydrotropism) (Takahashi and Jaffe, 1990; Takaha- shi, 1997)、避盐性(halotropism) (Munns and Tes- ter, 2008; Sun et al., 2008; Shelef et al., 2010; Galvan-Ampudia et al., 2013)及向声性(phonotro pism) (Mishra et al., 2016; Rodrigo-Moreno et al., 2017)等.其中, 向地性和向光性由于较易观察和筛选等原因, 研究进展较快, 目前已对调控植物向地性和向光性的分子机理有了较为深入的认识.由于重力等因素的干扰, 根向水性的研究较为困难.目前的研究仅发现少数基因和突变体参与或改变植物的向水性反应.向水性机理研究不仅具有重要的理论意义, 而且对节水灌溉和提高植物的抗旱性等也具有指导意义. ...
... 在植物根系的生长过程中, 细胞分裂素作为重要的植物调节激素被广泛认知(Mok and Mok, 2001; Wer- ner and Schmülling, 2009).细胞分裂素不仅可以抑制植物根的伸长和分化, 还参与植物根对干旱胁迫的响应(Skoog and Miller, 1957; Cary et al., 1995; Werner et al., 2003, 2010).研究发现, 在植物根的一侧施加浓度较高的细胞分裂素会使根克服重力朝着施加位点弯曲生长(Aloni et al., 2004), 说明细胞分裂素在植物根的向地性反应中可能起着负向调节作用.此外, 有研究表明, 细胞分裂素不仅可以下调植物根中生长素的极性运输, 而且能刺激根局部区域生长素的合成, 从而参与调节植物根的向地性生长(Zhou et al., 2011). ...
... 早期的植物生理学研究中, Porta (1658)和Knight (1811)等科学家通过大量的实验证明, 植物根系会随着生长环境中的水势梯度而改变其生长方向, 由此产生了对植物根向水性的研究.此外, 达尔文所著的《The Power of Movement in Plants》一书中也有关于向水性的描述(Darwin and Darwin, 1881).德国植物学家Sachs (1887)将由网格布包裹的潮湿木屑斜向悬挂, 让豌豆(Pisum sativum)种子在木屑表面萌发, 当豌豆的根因向地性生长至网格布下边缘时, 会沿网格布表面弯曲生长而不会进入干燥的空气中, 据此他认为植物的根存在向水性生长(图1A) (Sachs, 1887).另外, 研究还发现, 土壤中的水分会随着土壤深度、季节和降水等因素而分布不均, 产生水势梯度, 而且这种水势梯度会诱导根系向水性生长(Garwood and Williams,1967; Hsiao and Xu, 2000; Steudle, 2000).然而, 由于其它植物向性的干扰, 向水性反应一直以来未被科学家认可.直到1985年, Jaffe及其同事利用豌豆突变体“Ageotropum”为实验材料才首次强有力地证明了植物根向水性不依赖于向地性而独立存在(Jaffe et al., 1985).此豌豆突变体的根完全失去了向地性和避光性反应, 因此其部分根会向上生长出土壤(Blixt et al.,1958; Schöldéen and Burst- röm, 1960; Ekelund and Hemberg, 1966; Wilson, 1967).在一个湿度可控的玻璃盒子中培养该突变体植物, 当盒子中空气的相对湿度(relative humidity of the air, RH)达到95%-98%时, Ageotropum的根生长出土壤后不会弯曲, 而是向着湿润的空气中继续生长(图1B); 当空气相对湿度调整为85%-90%时, 原来向上生长的Ageotropum根会迅速向下弯曲生长直至进入比空气湿润的土壤里.这一现象说明, Ageotro- pum尽管缺失了向地性和避光性, 但其根仍然能感知环境中的水势梯度, 并具有正常的向水性生长反应.除了利用失去了向地性的突变体来研究植物根的向水性外, 科学家们还用竖直旋转的转盘来固定植物材料(Hoson et al., 1997; Kraft et al., 2000)进行研究, 发现竖直方向低速旋转培养植物的根的向地性被有效抵消.用此方法对豌豆以及黄瓜(Cucumis sativus)进行了研究, 发现它们的向水性表型更加明显(Takah- ashi et al., 1996, 1999).Takahashi及其同事发现, 在航天器失重条件下, 黄瓜的侧根也表现出更强烈地向水性反应, 而在正常培养的实验中侧根的向水性反应不易被观察到, 说明去除向地性影响后, 植物根对水势梯度的反应更明显(Takahashi et al.,1999). ...
2 2017
... 在探究向水性反应中根对信号的具体感知部位时, 科学家利用Ageotropum证明了根尖在感知水势梯度时起着决定性作用, 截除根冠的豌豆突变体失去了向水性生长的功能(Jaffe et al., 1985).然而最近的研究发现, 拟南芥根尖的分生-伸长过渡区对向水性至关重要, 而根冠对于向水性信号的感知并不重要(Dietrich et al., 2017).目前, 尚不清楚这两个矛盾的研究结果是由于研究材料的种间差异造成的, 还是研究方法不同所致. ... ... 日本Takahashi实验室利用新的实验体系(图1E), 发现拟南芥脱落酸合成缺失突变体aba1-1 (Koorn- neef et al., 1982)以及脱落酸不敏感突变体abi2-1 (Koornneef et al., 1984)对水势梯度不敏感, 但二者的根呈现出与野生型一致的向地性弯曲, 由此认为脱落酸能促进拟南芥根的向水性弯曲(Takahashi et al., 2002).随后的实验证明, 脱落酸可以通过调节MIZ1的表达来参与植物的向水性反应(Moriwaki et al., 2012).脱落酸信号途径正调控因子SnRK2.2及SnRK2.3的双重缺失突变体snrk2.2/snrk2.3表现出向水性反应降低的表型, 说明脱落酸正向调节拟南芥根的向水性反应.同时, 该研究还表明, 拟南芥根尖皮层细胞的分裂对向水性非常重要(Dietrich et al., 2017).然而, 脱落酸尽管可以恢复ahr1对水势梯度感知的缺陷, 但却加剧了nhr1向水性反应缺失的表型(Ponce et al., 2008; Moriwaki et al., 2012).墨西哥Cassab实验室对nhr1突变体脱落酸的定量分析显示, nhr1突变会造成植物体内脱落酸含量升高.再者, 脱落酸合成抑制剂AbamineSG会恢复nhr1向水性缺失的表型, 说明在nhr1参与的向水性反应中, 脱落酸可能起着负向调控作用(Quiroz-Figueroa et al., 2010). ...
1 2010
... 植物根柱细胞中的淀粉粒在根的重力感知中起重要作用(Kiss, 2000; Perrin et al., 2005).研究发现, 拟南芥(Arabidopsis thaliana)主根根柱细胞中的淀粉粒在水势梯度刺激后会迅速降解, 导致根的重力敏感性降低, 同时促进其向水性反应(Takahashi et al., 2003).但根柱细胞内淀粉粒的降解不仅受到水势梯度的诱导, 而且也受到其它不同胁迫(如干旱)的调控 (Takahashi et al., 2003; Sun et al., 2008; Ding and Friml, 2010; Iyer-Pascuzzi et al., 2011).因此, 尚不能确定向水性反应中的淀粉粒降解是由水势梯度特异诱导的还是受干旱诱导发生的.而且在一些植物中, 干旱会促进根的向地性反应(Leopold and LaFavre, 1989), 说明淀粉粒的降解可能对诱导植物根的向水性反应并非必需(Nakayama et al., 2012). ...
3 2003
... 植物根向水性反应实验筛选体系示意图 (A) Sachs发现植物的根具有向水性所用的实验方法(Sachs, 1887); (B) Jaffe及其同事证明植物根向水性存在所用的实验方法(Jaffe et al., 1985); (C) 墨西哥Cassab实验室的根向水性筛选系统(Eapen et al., 2003; Saucedo et al., 2012); (D), (E) 日本东北大学Takahashi实验室的根向水性筛选系统(Takahashi et al., 2002; Kobayashi et al., 2007).NM: 正常培养基; WSM: 水分胁迫培养基 ... ... Methods used for demonstrating root hydrotropism and screening for mutants altering root hydrotropic response (A) The method Sachs used to demonstrate plant root hydrotropic response (Sachs, 1887); (B) Experiment Jaffe et al. (1985) carried out to prove the existence of the hydrotropic response using Ageotropum mutant; (C) Screening system for identifying mutants with altered hydrotropic response in Cassab’s lab (Eapen et al., 2003; Saucedo et al., 2012); (D), (E) Screening system used in Takahashi’s laboratory for mutants with altered hydrotropic response (Takahashi et al., 2002; Kobayashi et al., 2007). NM: Normal nutrient medium; WSM: Water stress medium ... ... 进入21世纪后, 日本东北大学的Takahashi实验室和墨西哥国立自治大学的Cassab实验室开始利用分子遗传学手段研究根向水性反应的分子机制.Cassab实验室将培养基分成上下两部分, 其中上半部分为正常1/2MS培养基(normal nutrient medium, NM), 下半部分则是在正常培养基的基础上添加了渗透调节剂(如甘油或海藻酸)来降低培养基的水分含量(water stress medium, WSM), 进而人为地造成上下水势梯度.将萌发后的拟南芥幼苗放在上半部分离界面1 cm左右的培养基上垂直培养, 当根生长到接近两种培养基界面时, 野生型植物的根会克服向地性而向上弯曲生长, 说明其对水势梯度敏感; 而失去向水性生长的突变体会继续向下生长, 说明其对水势梯度不敏感(图1C).利用此方法, Cassab实验室分离到两个向水性缺陷突变体, 分别命名为nhr1 (no hydrotropic response 1)和ahr1 (altered hydrotropic response 1) (Eapen et al., 2003; Saucedo et al., 2012), 这两个突变体都呈现半显性遗传的特点, 但造成二者表型的突变基因至今未被鉴定. ...
1 1966
... 早期的植物生理学研究中, Porta (1658)和Knight (1811)等科学家通过大量的实验证明, 植物根系会随着生长环境中的水势梯度而改变其生长方向, 由此产生了对植物根向水性的研究.此外, 达尔文所著的《The Power of Movement in Plants》一书中也有关于向水性的描述(Darwin and Darwin, 1881).德国植物学家Sachs (1887)将由网格布包裹的潮湿木屑斜向悬挂, 让豌豆(Pisum sativum)种子在木屑表面萌发, 当豌豆的根因向地性生长至网格布下边缘时, 会沿网格布表面弯曲生长而不会进入干燥的空气中, 据此他认为植物的根存在向水性生长(图1A) (Sachs, 1887).另外, 研究还发现, 土壤中的水分会随着土壤深度、季节和降水等因素而分布不均, 产生水势梯度, 而且这种水势梯度会诱导根系向水性生长(Garwood and Williams,1967; Hsiao and Xu, 2000; Steudle, 2000).然而, 由于其它植物向性的干扰, 向水性反应一直以来未被科学家认可.直到1985年, Jaffe及其同事利用豌豆突变体“Ageotropum”为实验材料才首次强有力地证明了植物根向水性不依赖于向地性而独立存在(Jaffe et al., 1985).此豌豆突变体的根完全失去了向地性和避光性反应, 因此其部分根会向上生长出土壤(Blixt et al.,1958; Schöldéen and Burst- röm, 1960; Ekelund and Hemberg, 1966; Wilson, 1967).在一个湿度可控的玻璃盒子中培养该突变体植物, 当盒子中空气的相对湿度(relative humidity of the air, RH)达到95%-98%时, Ageotropum的根生长出土壤后不会弯曲, 而是向着湿润的空气中继续生长(图1B); 当空气相对湿度调整为85%-90%时, 原来向上生长的Ageotropum根会迅速向下弯曲生长直至进入比空气湿润的土壤里.这一现象说明, Ageotro- pum尽管缺失了向地性和避光性, 但其根仍然能感知环境中的水势梯度, 并具有正常的向水性生长反应.除了利用失去了向地性的突变体来研究植物根的向水性外, 科学家们还用竖直旋转的转盘来固定植物材料(Hoson et al., 1997; Kraft et al., 2000)进行研究, 发现竖直方向低速旋转培养植物的根的向地性被有效抵消.用此方法对豌豆以及黄瓜(Cucumis sativus)进行了研究, 发现它们的向水性表型更加明显(Takah- ashi et al., 1996, 1999).Takahashi及其同事发现, 在航天器失重条件下, 黄瓜的侧根也表现出更强烈地向水性反应, 而在正常培养的实验中侧根的向水性反应不易被观察到, 说明去除向地性影响后, 植物根对水势梯度的反应更明显(Takahashi et al.,1999). ...
1 2005
... 植物在整个生命活动过程中都在被动地适应不断变化的环境.在长期的进化选择中, 植物进化出一系列特有的机制来适应周围环境的变化, 使其能够一代代繁衍生息.植物能够感知自身生长环境中各种非生物(包括光、重力、水、温度、氧气和二氧化碳等)和生物信号, 并做出相应的生理生化反应.向性反应(tropic response)作为植物适应周围环境变化的重要机制之一, 是植物实现自身趋利避害的主要途径.植物朝向环境信号生长的向性反应称为正(positive)向性反应, 反之则称为负(negative)向性反应.植物为了应对不同的环境信号进化出了一系列向性反应, 包括向光性(phototropism) (Van Overbeek,1939; Gehr- ing et al., 1990; Liscum et al., 2014)、向地性(gravi- tropism) (Gehring et al., 1990; Hart, 1990; Taka- hashi and Jaffe, 1990; Correll and Kiss, 2002)、向化性(chemotropism) (Esmon et al., 2005)、向触性(thigmotropism) (Monshausen et al., 2008)、向水性(hydrotropism) (Takahashi and Jaffe, 1990; Takaha- shi, 1997)、避盐性(halotropism) (Munns and Tes- ter, 2008; Sun et al., 2008; Shelef et al., 2010; Galvan-Ampudia et al., 2013)及向声性(phonotro pism) (Mishra et al., 2016; Rodrigo-Moreno et al., 2017)等.其中, 向地性和向光性由于较易观察和筛选等原因, 研究进展较快, 目前已对调控植物向地性和向光性的分子机理有了较为深入的认识.由于重力等因素的干扰, 根向水性的研究较为困难.目前的研究仅发现少数基因和突变体参与或改变植物的向水性反应.向水性机理研究不仅具有重要的理论意义, 而且对节水灌溉和提高植物的抗旱性等也具有指导意义. ...
1 2013
... 植物在整个生命活动过程中都在被动地适应不断变化的环境.在长期的进化选择中, 植物进化出一系列特有的机制来适应周围环境的变化, 使其能够一代代繁衍生息.植物能够感知自身生长环境中各种非生物(包括光、重力、水、温度、氧气和二氧化碳等)和生物信号, 并做出相应的生理生化反应.向性反应(tropic response)作为植物适应周围环境变化的重要机制之一, 是植物实现自身趋利避害的主要途径.植物朝向环境信号生长的向性反应称为正(positive)向性反应, 反之则称为负(negative)向性反应.植物为了应对不同的环境信号进化出了一系列向性反应, 包括向光性(phototropism) (Van Overbeek,1939; Gehr- ing et al., 1990; Liscum et al., 2014)、向地性(gravi- tropism) (Gehring et al., 1990; Hart, 1990; Taka- hashi and Jaffe, 1990; Correll and Kiss, 2002)、向化性(chemotropism) (Esmon et al., 2005)、向触性(thigmotropism) (Monshausen et al., 2008)、向水性(hydrotropism) (Takahashi and Jaffe, 1990; Takaha- shi, 1997)、避盐性(halotropism) (Munns and Tes- ter, 2008; Sun et al., 2008; Shelef et al., 2010; Galvan-Ampudia et al., 2013)及向声性(phonotro pism) (Mishra et al., 2016; Rodrigo-Moreno et al., 2017)等.其中, 向地性和向光性由于较易观察和筛选等原因, 研究进展较快, 目前已对调控植物向地性和向光性的分子机理有了较为深入的认识.由于重力等因素的干扰, 根向水性的研究较为困难.目前的研究仅发现少数基因和突变体参与或改变植物的向水性反应.向水性机理研究不仅具有重要的理论意义, 而且对节水灌溉和提高植物的抗旱性等也具有指导意义. ...
1 1967
... 早期的植物生理学研究中, Porta (1658)和Knight (1811)等科学家通过大量的实验证明, 植物根系会随着生长环境中的水势梯度而改变其生长方向, 由此产生了对植物根向水性的研究.此外, 达尔文所著的《The Power of Movement in Plants》一书中也有关于向水性的描述(Darwin and Darwin, 1881).德国植物学家Sachs (1887)将由网格布包裹的潮湿木屑斜向悬挂, 让豌豆(Pisum sativum)种子在木屑表面萌发, 当豌豆的根因向地性生长至网格布下边缘时, 会沿网格布表面弯曲生长而不会进入干燥的空气中, 据此他认为植物的根存在向水性生长(图1A) (Sachs, 1887).另外, 研究还发现, 土壤中的水分会随着土壤深度、季节和降水等因素而分布不均, 产生水势梯度, 而且这种水势梯度会诱导根系向水性生长(Garwood and Williams,1967; Hsiao and Xu, 2000; Steudle, 2000).然而, 由于其它植物向性的干扰, 向水性反应一直以来未被科学家认可.直到1985年, Jaffe及其同事利用豌豆突变体“Ageotropum”为实验材料才首次强有力地证明了植物根向水性不依赖于向地性而独立存在(Jaffe et al., 1985).此豌豆突变体的根完全失去了向地性和避光性反应, 因此其部分根会向上生长出土壤(Blixt et al.,1958; Schöldéen and Burst- röm, 1960; Ekelund and Hemberg, 1966; Wilson, 1967).在一个湿度可控的玻璃盒子中培养该突变体植物, 当盒子中空气的相对湿度(relative humidity of the air, RH)达到95%-98%时, Ageotropum的根生长出土壤后不会弯曲, 而是向着湿润的空气中继续生长(图1B); 当空气相对湿度调整为85%-90%时, 原来向上生长的Ageotropum根会迅速向下弯曲生长直至进入比空气湿润的土壤里.这一现象说明, Ageotro- pum尽管缺失了向地性和避光性, 但其根仍然能感知环境中的水势梯度, 并具有正常的向水性生长反应.除了利用失去了向地性的突变体来研究植物根的向水性外, 科学家们还用竖直旋转的转盘来固定植物材料(Hoson et al., 1997; Kraft et al., 2000)进行研究, 发现竖直方向低速旋转培养植物的根的向地性被有效抵消.用此方法对豌豆以及黄瓜(Cucumis sativus)进行了研究, 发现它们的向水性表型更加明显(Takah- ashi et al., 1996, 1999).Takahashi及其同事发现, 在航天器失重条件下, 黄瓜的侧根也表现出更强烈地向水性反应, 而在正常培养的实验中侧根的向水性反应不易被观察到, 说明去除向地性影响后, 植物根对水势梯度的反应更明显(Takahashi et al.,1999). ...
2 1990
... 植物在整个生命活动过程中都在被动地适应不断变化的环境.在长期的进化选择中, 植物进化出一系列特有的机制来适应周围环境的变化, 使其能够一代代繁衍生息.植物能够感知自身生长环境中各种非生物(包括光、重力、水、温度、氧气和二氧化碳等)和生物信号, 并做出相应的生理生化反应.向性反应(tropic response)作为植物适应周围环境变化的重要机制之一, 是植物实现自身趋利避害的主要途径.植物朝向环境信号生长的向性反应称为正(positive)向性反应, 反之则称为负(negative)向性反应.植物为了应对不同的环境信号进化出了一系列向性反应, 包括向光性(phototropism) (Van Overbeek,1939; Gehr- ing et al., 1990; Liscum et al., 2014)、向地性(gravi- tropism) (Gehring et al., 1990; Hart, 1990; Taka- hashi and Jaffe, 1990; Correll and Kiss, 2002)、向化性(chemotropism) (Esmon et al., 2005)、向触性(thigmotropism) (Monshausen et al., 2008)、向水性(hydrotropism) (Takahashi and Jaffe, 1990; Takaha- shi, 1997)、避盐性(halotropism) (Munns and Tes- ter, 2008; Sun et al., 2008; Shelef et al., 2010; Galvan-Ampudia et al., 2013)及向声性(phonotro pism) (Mishra et al., 2016; Rodrigo-Moreno et al., 2017)等.其中, 向地性和向光性由于较易观察和筛选等原因, 研究进展较快, 目前已对调控植物向地性和向光性的分子机理有了较为深入的认识.由于重力等因素的干扰, 根向水性的研究较为困难.目前的研究仅发现少数基因和突变体参与或改变植物的向水性反应.向水性机理研究不仅具有重要的理论意义, 而且对节水灌溉和提高植物的抗旱性等也具有指导意义. ... ... )、向地性(gravi- tropism) (Gehring et al., 1990; Hart, 1990; Taka- hashi and Jaffe, 1990; Correll and Kiss, 2002)、向化性(chemotropism) (Esmon et al., 2005)、向触性(thigmotropism) (Monshausen et al., 2008)、向水性(hydrotropism) (Takahashi and Jaffe, 1990; Takaha- shi, 1997)、避盐性(halotropism) (Munns and Tes- ter, 2008; Sun et al., 2008; Shelef et al., 2010; Galvan-Ampudia et al., 2013)及向声性(phonotro pism) (Mishra et al., 2016; Rodrigo-Moreno et al., 2017)等.其中, 向地性和向光性由于较易观察和筛选等原因, 研究进展较快, 目前已对调控植物向地性和向光性的分子机理有了较为深入的认识.由于重力等因素的干扰, 根向水性的研究较为困难.目前的研究仅发现少数基因和突变体参与或改变植物的向水性反应.向水性机理研究不仅具有重要的理论意义, 而且对节水灌溉和提高植物的抗旱性等也具有指导意义. ...
1 2003
... 同时, Takahashi实验室还筛选到另一个拟南芥根向水性点突变体miz2 (miz-kussei2), MIZ2编码1个参与细胞中囊泡运输的GNOM蛋白.miz2的向地性并没有因为其向水性的改变而发生变化(Miyazawa et al., 2009).此外, GNOM调节的囊泡运输可被囊泡运输抑制剂BFA (Brefeldin A)所抑制(Geldner et al., 2003), 而体外施加BFA也可明显抑制野生型的向水性弯曲, 说明拟南芥根的向水性反应依赖于囊泡运输(Miyazawa et al., 2009).随后, 科学家将miz2和超表达的MIZ1 (MIZ1OE)进行杂交, 发现尽管MIZ1基因超表达可以导致拟南芥根的向水性反应显著增强, 但miz2/MIZ1OE根对水势梯度依然表现出不敏感的表型, 说明MIZ1调控的根向水性反应依赖于MIZ2的功能, 即MIZ1和MIZ2在植物根向水性反应中处于同一信号通路, 且MIZ2在MIZ1下游发挥作用(Miyazawa et al., 2012). ...
... 植物在整个生命活动过程中都在被动地适应不断变化的环境.在长期的进化选择中, 植物进化出一系列特有的机制来适应周围环境的变化, 使其能够一代代繁衍生息.植物能够感知自身生长环境中各种非生物(包括光、重力、水、温度、氧气和二氧化碳等)和生物信号, 并做出相应的生理生化反应.向性反应(tropic response)作为植物适应周围环境变化的重要机制之一, 是植物实现自身趋利避害的主要途径.植物朝向环境信号生长的向性反应称为正(positive)向性反应, 反之则称为负(negative)向性反应.植物为了应对不同的环境信号进化出了一系列向性反应, 包括向光性(phototropism) (Van Overbeek,1939; Gehr- ing et al., 1990; Liscum et al., 2014)、向地性(gravi- tropism) (Gehring et al., 1990; Hart, 1990; Taka- hashi and Jaffe, 1990; Correll and Kiss, 2002)、向化性(chemotropism) (Esmon et al., 2005)、向触性(thigmotropism) (Monshausen et al., 2008)、向水性(hydrotropism) (Takahashi and Jaffe, 1990; Takaha- shi, 1997)、避盐性(halotropism) (Munns and Tes- ter, 2008; Sun et al., 2008; Shelef et al., 2010; Galvan-Ampudia et al., 2013)及向声性(phonotro pism) (Mishra et al., 2016; Rodrigo-Moreno et al., 2017)等.其中, 向地性和向光性由于较易观察和筛选等原因, 研究进展较快, 目前已对调控植物向地性和向光性的分子机理有了较为深入的认识.由于重力等因素的干扰, 根向水性的研究较为困难.目前的研究仅发现少数基因和突变体参与或改变植物的向水性反应.向水性机理研究不仅具有重要的理论意义, 而且对节水灌溉和提高植物的抗旱性等也具有指导意义. ...
1 1997
... 早期的植物生理学研究中, Porta (1658)和Knight (1811)等科学家通过大量的实验证明, 植物根系会随着生长环境中的水势梯度而改变其生长方向, 由此产生了对植物根向水性的研究.此外, 达尔文所著的《The Power of Movement in Plants》一书中也有关于向水性的描述(Darwin and Darwin, 1881).德国植物学家Sachs (1887)将由网格布包裹的潮湿木屑斜向悬挂, 让豌豆(Pisum sativum)种子在木屑表面萌发, 当豌豆的根因向地性生长至网格布下边缘时, 会沿网格布表面弯曲生长而不会进入干燥的空气中, 据此他认为植物的根存在向水性生长(图1A) (Sachs, 1887).另外, 研究还发现, 土壤中的水分会随着土壤深度、季节和降水等因素而分布不均, 产生水势梯度, 而且这种水势梯度会诱导根系向水性生长(Garwood and Williams,1967; Hsiao and Xu, 2000; Steudle, 2000).然而, 由于其它植物向性的干扰, 向水性反应一直以来未被科学家认可.直到1985年, Jaffe及其同事利用豌豆突变体“Ageotropum”为实验材料才首次强有力地证明了植物根向水性不依赖于向地性而独立存在(Jaffe et al., 1985).此豌豆突变体的根完全失去了向地性和避光性反应, 因此其部分根会向上生长出土壤(Blixt et al.,1958; Schöldéen and Burst- röm, 1960; Ekelund and Hemberg, 1966; Wilson, 1967).在一个湿度可控的玻璃盒子中培养该突变体植物, 当盒子中空气的相对湿度(relative humidity of the air, RH)达到95%-98%时, Ageotropum的根生长出土壤后不会弯曲, 而是向着湿润的空气中继续生长(图1B); 当空气相对湿度调整为85%-90%时, 原来向上生长的Ageotropum根会迅速向下弯曲生长直至进入比空气湿润的土壤里.这一现象说明, Ageotro- pum尽管缺失了向地性和避光性, 但其根仍然能感知环境中的水势梯度, 并具有正常的向水性生长反应.除了利用失去了向地性的突变体来研究植物根的向水性外, 科学家们还用竖直旋转的转盘来固定植物材料(Hoson et al., 1997; Kraft et al., 2000)进行研究, 发现竖直方向低速旋转培养植物的根的向地性被有效抵消.用此方法对豌豆以及黄瓜(Cucumis sativus)进行了研究, 发现它们的向水性表型更加明显(Takah- ashi et al., 1996, 1999).Takahashi及其同事发现, 在航天器失重条件下, 黄瓜的侧根也表现出更强烈地向水性反应, 而在正常培养的实验中侧根的向水性反应不易被观察到, 说明去除向地性影响后, 植物根对水势梯度的反应更明显(Takahashi et al.,1999). ...
1 2000
... 早期的植物生理学研究中, Porta (1658)和Knight (1811)等科学家通过大量的实验证明, 植物根系会随着生长环境中的水势梯度而改变其生长方向, 由此产生了对植物根向水性的研究.此外, 达尔文所著的《The Power of Movement in Plants》一书中也有关于向水性的描述(Darwin and Darwin, 1881).德国植物学家Sachs (1887)将由网格布包裹的潮湿木屑斜向悬挂, 让豌豆(Pisum sativum)种子在木屑表面萌发, 当豌豆的根因向地性生长至网格布下边缘时, 会沿网格布表面弯曲生长而不会进入干燥的空气中, 据此他认为植物的根存在向水性生长(图1A) (Sachs, 1887).另外, 研究还发现, 土壤中的水分会随着土壤深度、季节和降水等因素而分布不均, 产生水势梯度, 而且这种水势梯度会诱导根系向水性生长(Garwood and Williams,1967; Hsiao and Xu, 2000; Steudle, 2000).然而, 由于其它植物向性的干扰, 向水性反应一直以来未被科学家认可.直到1985年, Jaffe及其同事利用豌豆突变体“Ageotropum”为实验材料才首次强有力地证明了植物根向水性不依赖于向地性而独立存在(Jaffe et al., 1985).此豌豆突变体的根完全失去了向地性和避光性反应, 因此其部分根会向上生长出土壤(Blixt et al.,1958; Schöldéen and Burst- röm, 1960; Ekelund and Hemberg, 1966; Wilson, 1967).在一个湿度可控的玻璃盒子中培养该突变体植物, 当盒子中空气的相对湿度(relative humidity of the air, RH)达到95%-98%时, Ageotropum的根生长出土壤后不会弯曲, 而是向着湿润的空气中继续生长(图1B); 当空气相对湿度调整为85%-90%时, 原来向上生长的Ageotropum根会迅速向下弯曲生长直至进入比空气湿润的土壤里.这一现象说明, Ageotro- pum尽管缺失了向地性和避光性, 但其根仍然能感知环境中的水势梯度, 并具有正常的向水性生长反应.除了利用失去了向地性的突变体来研究植物根的向水性外, 科学家们还用竖直旋转的转盘来固定植物材料(Hoson et al., 1997; Kraft et al., 2000)进行研究, 发现竖直方向低速旋转培养植物的根的向地性被有效抵消.用此方法对豌豆以及黄瓜(Cucumis sativus)进行了研究, 发现它们的向水性表型更加明显(Takah- ashi et al., 1996, 1999).Takahashi及其同事发现, 在航天器失重条件下, 黄瓜的侧根也表现出更强烈地向水性反应, 而在正常培养的实验中侧根的向水性反应不易被观察到, 说明去除向地性影响后, 植物根对水势梯度的反应更明显(Takahashi et al.,1999). ...
1 2011
... 植物根柱细胞中的淀粉粒在根的重力感知中起重要作用(Kiss, 2000; Perrin et al., 2005).研究发现, 拟南芥(Arabidopsis thaliana)主根根柱细胞中的淀粉粒在水势梯度刺激后会迅速降解, 导致根的重力敏感性降低, 同时促进其向水性反应(Takahashi et al., 2003).但根柱细胞内淀粉粒的降解不仅受到水势梯度的诱导, 而且也受到其它不同胁迫(如干旱)的调控 (Takahashi et al., 2003; Sun et al., 2008; Ding and Friml, 2010; Iyer-Pascuzzi et al., 2011).因此, 尚不能确定向水性反应中的淀粉粒降解是由水势梯度特异诱导的还是受干旱诱导发生的.而且在一些植物中, 干旱会促进根的向地性反应(Leopold and LaFavre, 1989), 说明淀粉粒的降解可能对诱导植物根的向水性反应并非必需(Nakayama et al., 2012). ...
3 1985
... 早期的植物生理学研究中, Porta (1658)和Knight (1811)等科学家通过大量的实验证明, 植物根系会随着生长环境中的水势梯度而改变其生长方向, 由此产生了对植物根向水性的研究.此外, 达尔文所著的《The Power of Movement in Plants》一书中也有关于向水性的描述(Darwin and Darwin, 1881).德国植物学家Sachs (1887)将由网格布包裹的潮湿木屑斜向悬挂, 让豌豆(Pisum sativum)种子在木屑表面萌发, 当豌豆的根因向地性生长至网格布下边缘时, 会沿网格布表面弯曲生长而不会进入干燥的空气中, 据此他认为植物的根存在向水性生长(图1A) (Sachs, 1887).另外, 研究还发现, 土壤中的水分会随着土壤深度、季节和降水等因素而分布不均, 产生水势梯度, 而且这种水势梯度会诱导根系向水性生长(Garwood and Williams,1967; Hsiao and Xu, 2000; Steudle, 2000).然而, 由于其它植物向性的干扰, 向水性反应一直以来未被科学家认可.直到1985年, Jaffe及其同事利用豌豆突变体“Ageotropum”为实验材料才首次强有力地证明了植物根向水性不依赖于向地性而独立存在(Jaffe et al., 1985).此豌豆突变体的根完全失去了向地性和避光性反应, 因此其部分根会向上生长出土壤(Blixt et al.,1958; Schöldéen and Burst- röm, 1960; Ekelund and Hemberg, 1966; Wilson, 1967).在一个湿度可控的玻璃盒子中培养该突变体植物, 当盒子中空气的相对湿度(relative humidity of the air, RH)达到95%-98%时, Ageotropum的根生长出土壤后不会弯曲, 而是向着湿润的空气中继续生长(图1B); 当空气相对湿度调整为85%-90%时, 原来向上生长的Ageotropum根会迅速向下弯曲生长直至进入比空气湿润的土壤里.这一现象说明, Ageotro- pum尽管缺失了向地性和避光性, 但其根仍然能感知环境中的水势梯度, 并具有正常的向水性生长反应.除了利用失去了向地性的突变体来研究植物根的向水性外, 科学家们还用竖直旋转的转盘来固定植物材料(Hoson et al., 1997; Kraft et al., 2000)进行研究, 发现竖直方向低速旋转培养植物的根的向地性被有效抵消.用此方法对豌豆以及黄瓜(Cucumis sativus)进行了研究, 发现它们的向水性表型更加明显(Takah- ashi et al., 1996, 1999).Takahashi及其同事发现, 在航天器失重条件下, 黄瓜的侧根也表现出更强烈地向水性反应, 而在正常培养的实验中侧根的向水性反应不易被观察到, 说明去除向地性影响后, 植物根对水势梯度的反应更明显(Takahashi et al.,1999). ... ... 植物根向水性反应实验筛选体系示意图 (A) Sachs发现植物的根具有向水性所用的实验方法(Sachs, 1887); (B) Jaffe及其同事证明植物根向水性存在所用的实验方法(Jaffe et al., 1985); (C) 墨西哥Cassab实验室的根向水性筛选系统(Eapen et al., 2003; Saucedo et al., 2012); (D), (E) 日本东北大学Takahashi实验室的根向水性筛选系统(Takahashi et al., 2002; Kobayashi et al., 2007).NM: 正常培养基; WSM: 水分胁迫培养基 ... ... 在探究向水性反应中根对信号的具体感知部位时, 科学家利用Ageotropum证明了根尖在感知水势梯度时起着决定性作用, 截除根冠的豌豆突变体失去了向水性生长的功能(Jaffe et al., 1985).然而最近的研究发现, 拟南芥根尖的分生-伸长过渡区对向水性至关重要, 而根冠对于向水性信号的感知并不重要(Dietrich et al., 2017).目前, 尚不清楚这两个矛盾的研究结果是由于研究材料的种间差异造成的, 还是研究方法不同所致. ...
1 2010
... 2012年, Cassab实验室指出细胞分裂素在植物根的向水性反应中发挥重要作用; 并且通过实验证明细胞分裂素能够恢复ahr1根的向水性表型, 且这种恢复不是通过改变其向地性来完成(Saucedo et al., 2012).此外, 细胞分裂素还可以调节生长素的合成(Jones et al., 2010).因此, Cassab实验室认为, 细胞分裂素参与植物根的向水性反应可能通过调节根中生长素的水平实现.目前, 细胞分裂素在植物根向水性反应中的作用机制尚不明确. ...
1 2007
... 1991年, Takahashi在对豌豆突变体“Ageotro- pum”的向水性研究中发现, 当对Ageotropum施加生长素转运抑制剂三碘苯甲酸(TIBA)时, 其根的向水性反应被明显抑制, 说明生长素的极性运输在植物根的向水性反应过程中具有重要作用(Takahashi and Suge, 1991).然而生长素的调节效应具有种间差异.在豌豆、黄瓜和水稻(Oryza sativa)根尖, 生长素以重新分布的模式调控其根的向地性以及向水性弯曲(Takahashi and Suge, 1991; Mizuno et al., 2002; Nakajima et al., 2017); 但与豌豆同属豆科植物的百脉根(Lotus corniculatus)和十字花科的拟南芥根向水性反应却不依赖于生长素的极性运输与重新分配(Shkolnik et al., 2016; Nakajima et al., 2017).研究表明, 生长素转运抑制剂(CHPAA、NPA和TIBA)不会影响百脉根和拟南芥根的向水性反应(Kaneyasu et al., 2007; Nakajima et al., 2017), 但生长素合成抑制剂Kyn会明显减弱百脉根的向水性反应(Nakajima et al., 2017).此外, PINs是植物体内调节生长素运输方向的蛋白, 它们以极性定位的方式参与生长素在植物细胞间的极性运输(Michniewicz et al., 2007).拟南芥中, PIN2缺失突变体表现出向地性反应缺陷, 但却具有正常的向水性反应(Takahashi et al., 2002).生长素信号通路受损的拟南芥突变体axr1-3和axr2-1根的向地性反应虽然减弱(Leyser et al., 1993; Nagpal et al., 2000), 但其根对水势梯度响应的敏感性增强(Takahashi et al., 2002).研究发现, 超表达MIZ1能增强拟南芥根的向水性反应, 同时其根中生长素合成基因的表达水平明显降低(Moriwaki et al., 2011; Miyazawa et al., 2012).总之, 生长素通过极性运输和重新分布的方式参与植物根的向地性反应, 但该作用机理并不适用于所有植物(如百脉根和拟南芥)根的向水性反应, 有关生长素对向水性反应调控的种间差异及生理和分子基础有待进一步研究. ...
1 2000
... 植物根柱细胞中的淀粉粒在根的重力感知中起重要作用(Kiss, 2000; Perrin et al., 2005).研究发现, 拟南芥(Arabidopsis thaliana)主根根柱细胞中的淀粉粒在水势梯度刺激后会迅速降解, 导致根的重力敏感性降低, 同时促进其向水性反应(Takahashi et al., 2003).但根柱细胞内淀粉粒的降解不仅受到水势梯度的诱导, 而且也受到其它不同胁迫(如干旱)的调控 (Takahashi et al., 2003; Sun et al., 2008; Ding and Friml, 2010; Iyer-Pascuzzi et al., 2011).因此, 尚不能确定向水性反应中的淀粉粒降解是由水势梯度特异诱导的还是受干旱诱导发生的.而且在一些植物中, 干旱会促进根的向地性反应(Leopold and LaFavre, 1989), 说明淀粉粒的降解可能对诱导植物根的向水性反应并非必需(Nakayama et al., 2012). ...
1 1811
... 早期的植物生理学研究中, Porta (1658)和Knight (1811)等科学家通过大量的实验证明, 植物根系会随着生长环境中的水势梯度而改变其生长方向, 由此产生了对植物根向水性的研究.此外, 达尔文所著的《The Power of Movement in Plants》一书中也有关于向水性的描述(Darwin and Darwin, 1881).德国植物学家Sachs (1887)将由网格布包裹的潮湿木屑斜向悬挂, 让豌豆(Pisum sativum)种子在木屑表面萌发, 当豌豆的根因向地性生长至网格布下边缘时, 会沿网格布表面弯曲生长而不会进入干燥的空气中, 据此他认为植物的根存在向水性生长(图1A) (Sachs, 1887).另外, 研究还发现, 土壤中的水分会随着土壤深度、季节和降水等因素而分布不均, 产生水势梯度, 而且这种水势梯度会诱导根系向水性生长(Garwood and Williams,1967; Hsiao and Xu, 2000; Steudle, 2000).然而, 由于其它植物向性的干扰, 向水性反应一直以来未被科学家认可.直到1985年, Jaffe及其同事利用豌豆突变体“Ageotropum”为实验材料才首次强有力地证明了植物根向水性不依赖于向地性而独立存在(Jaffe et al., 1985).此豌豆突变体的根完全失去了向地性和避光性反应, 因此其部分根会向上生长出土壤(Blixt et al.,1958; Schöldéen and Burst- röm, 1960; Ekelund and Hemberg, 1966; Wilson, 1967).在一个湿度可控的玻璃盒子中培养该突变体植物, 当盒子中空气的相对湿度(relative humidity of the air, RH)达到95%-98%时, Ageotropum的根生长出土壤后不会弯曲, 而是向着湿润的空气中继续生长(图1B); 当空气相对湿度调整为85%-90%时, 原来向上生长的Ageotropum根会迅速向下弯曲生长直至进入比空气湿润的土壤里.这一现象说明, Ageotro- pum尽管缺失了向地性和避光性, 但其根仍然能感知环境中的水势梯度, 并具有正常的向水性生长反应.除了利用失去了向地性的突变体来研究植物根的向水性外, 科学家们还用竖直旋转的转盘来固定植物材料(Hoson et al., 1997; Kraft et al., 2000)进行研究, 发现竖直方向低速旋转培养植物的根的向地性被有效抵消.用此方法对豌豆以及黄瓜(Cucumis sativus)进行了研究, 发现它们的向水性表型更加明显(Takah- ashi et al., 1996, 1999).Takahashi及其同事发现, 在航天器失重条件下, 黄瓜的侧根也表现出更强烈地向水性反应, 而在正常培养的实验中侧根的向水性反应不易被观察到, 说明去除向地性影响后, 植物根对水势梯度的反应更明显(Takahashi et al.,1999). ...
3 2007
... 植物根向水性反应实验筛选体系示意图 (A) Sachs发现植物的根具有向水性所用的实验方法(Sachs, 1887); (B) Jaffe及其同事证明植物根向水性存在所用的实验方法(Jaffe et al., 1985); (C) 墨西哥Cassab实验室的根向水性筛选系统(Eapen et al., 2003; Saucedo et al., 2012); (D), (E) 日本东北大学Takahashi实验室的根向水性筛选系统(Takahashi et al., 2002; Kobayashi et al., 2007).NM: 正常培养基; WSM: 水分胁迫培养基 ... ... Methods used for demonstrating root hydrotropism and screening for mutants altering root hydrotropic response (A) The method Sachs used to demonstrate plant root hydrotropic response (Sachs, 1887); (B) Experiment Jaffe et al. (1985) carried out to prove the existence of the hydrotropic response using Ageotropum mutant; (C) Screening system for identifying mutants with altered hydrotropic response in Cassab’s lab (Eapen et al., 2003; Saucedo et al., 2012); (D), (E) Screening system used in Takahashi’s laboratory for mutants with altered hydrotropic response (Takahashi et al., 2002; Kobayashi et al., 2007). NM: Normal nutrient medium; WSM: Water stress medium ... ... Takahashi实验室利用一个密闭的聚丙烯塑料盒建立了空气水分梯度(图1D).在密闭盒子的底部放置1个盛有饱和碳酸钾溶液的器皿, 盒子的内侧上方固定垂直放置添加了琼脂糖的1/2MS培养基, 形成盒子下方水势低而上方水势高的空气水势梯度.将拟南芥幼苗垂直植入培养基, 使其根尖露出培养基边缘3 mm, 在培养过程中, 野生型的根会克服向地性朝向培养基方向弯曲生长, 而对水分梯度不敏感的突变体则不会发生弯曲, 最终根尖暴露在空气中脱水死亡.该实验室利用此系统筛选到1个具有正常向地性和生长表型, 但对水势梯度不敏感的突变体, 命名为“mizu-kussei”(日语中的向水性一词, 简称miz1) (Kobayashi et al., 2007).研究表明, MIZ1在感应水势梯度的早期阶段扮演着重要角色, 然而其编码蛋 白在调节根向水性反应过程中的具体分子机制尚不清楚. ...
1 1984
... 日本Takahashi实验室利用新的实验体系(图1E), 发现拟南芥脱落酸合成缺失突变体aba1-1 (Koorn- neef et al., 1982)以及脱落酸不敏感突变体abi2-1 (Koornneef et al., 1984)对水势梯度不敏感, 但二者的根呈现出与野生型一致的向地性弯曲, 由此认为脱落酸能促进拟南芥根的向水性弯曲(Takahashi et al., 2002).随后的实验证明, 脱落酸可以通过调节MIZ1的表达来参与植物的向水性反应(Moriwaki et al., 2012).脱落酸信号途径正调控因子SnRK2.2及SnRK2.3的双重缺失突变体snrk2.2/snrk2.3表现出向水性反应降低的表型, 说明脱落酸正向调节拟南芥根的向水性反应.同时, 该研究还表明, 拟南芥根尖皮层细胞的分裂对向水性非常重要(Dietrich et al., 2017).然而, 脱落酸尽管可以恢复ahr1对水势梯度感知的缺陷, 但却加剧了nhr1向水性反应缺失的表型(Ponce et al., 2008; Moriwaki et al., 2012).墨西哥Cassab实验室对nhr1突变体脱落酸的定量分析显示, nhr1突变会造成植物体内脱落酸含量升高.再者, 脱落酸合成抑制剂AbamineSG会恢复nhr1向水性缺失的表型, 说明在nhr1参与的向水性反应中, 脱落酸可能起着负向调控作用(Quiroz-Figueroa et al., 2010). ...
1 1982
... 日本Takahashi实验室利用新的实验体系(图1E), 发现拟南芥脱落酸合成缺失突变体aba1-1 (Koorn- neef et al., 1982)以及脱落酸不敏感突变体abi2-1 (Koornneef et al., 1984)对水势梯度不敏感, 但二者的根呈现出与野生型一致的向地性弯曲, 由此认为脱落酸能促进拟南芥根的向水性弯曲(Takahashi et al., 2002).随后的实验证明, 脱落酸可以通过调节MIZ1的表达来参与植物的向水性反应(Moriwaki et al., 2012).脱落酸信号途径正调控因子SnRK2.2及SnRK2.3的双重缺失突变体snrk2.2/snrk2.3表现出向水性反应降低的表型, 说明脱落酸正向调节拟南芥根的向水性反应.同时, 该研究还表明, 拟南芥根尖皮层细胞的分裂对向水性非常重要(Dietrich et al., 2017).然而, 脱落酸尽管可以恢复ahr1对水势梯度感知的缺陷, 但却加剧了nhr1向水性反应缺失的表型(Ponce et al., 2008; Moriwaki et al., 2012).墨西哥Cassab实验室对nhr1突变体脱落酸的定量分析显示, nhr1突变会造成植物体内脱落酸含量升高.再者, 脱落酸合成抑制剂AbamineSG会恢复nhr1向水性缺失的表型, 说明在nhr1参与的向水性反应中, 脱落酸可能起着负向调控作用(Quiroz-Figueroa et al., 2010). ...
1 2000
... 早期的植物生理学研究中, Porta (1658)和Knight (1811)等科学家通过大量的实验证明, 植物根系会随着生长环境中的水势梯度而改变其生长方向, 由此产生了对植物根向水性的研究.此外, 达尔文所著的《The Power of Movement in Plants》一书中也有关于向水性的描述(Darwin and Darwin, 1881).德国植物学家Sachs (1887)将由网格布包裹的潮湿木屑斜向悬挂, 让豌豆(Pisum sativum)种子在木屑表面萌发, 当豌豆的根因向地性生长至网格布下边缘时, 会沿网格布表面弯曲生长而不会进入干燥的空气中, 据此他认为植物的根存在向水性生长(图1A) (Sachs, 1887).另外, 研究还发现, 土壤中的水分会随着土壤深度、季节和降水等因素而分布不均, 产生水势梯度, 而且这种水势梯度会诱导根系向水性生长(Garwood and Williams,1967; Hsiao and Xu, 2000; Steudle, 2000).然而, 由于其它植物向性的干扰, 向水性反应一直以来未被科学家认可.直到1985年, Jaffe及其同事利用豌豆突变体“Ageotropum”为实验材料才首次强有力地证明了植物根向水性不依赖于向地性而独立存在(Jaffe et al., 1985).此豌豆突变体的根完全失去了向地性和避光性反应, 因此其部分根会向上生长出土壤(Blixt et al.,1958; Schöldéen and Burst- röm, 1960; Ekelund and Hemberg, 1966; Wilson, 1967).在一个湿度可控的玻璃盒子中培养该突变体植物, 当盒子中空气的相对湿度(relative humidity of the air, RH)达到95%-98%时, Ageotropum的根生长出土壤后不会弯曲, 而是向着湿润的空气中继续生长(图1B); 当空气相对湿度调整为85%-90%时, 原来向上生长的Ageotropum根会迅速向下弯曲生长直至进入比空气湿润的土壤里.这一现象说明, Ageotro- pum尽管缺失了向地性和避光性, 但其根仍然能感知环境中的水势梯度, 并具有正常的向水性生长反应.除了利用失去了向地性的突变体来研究植物根的向水性外, 科学家们还用竖直旋转的转盘来固定植物材料(Hoson et al., 1997; Kraft et al., 2000)进行研究, 发现竖直方向低速旋转培养植物的根的向地性被有效抵消.用此方法对豌豆以及黄瓜(Cucumis sativus)进行了研究, 发现它们的向水性表型更加明显(Takah- ashi et al., 1996, 1999).Takahashi及其同事发现, 在航天器失重条件下, 黄瓜的侧根也表现出更强烈地向水性反应, 而在正常培养的实验中侧根的向水性反应不易被观察到, 说明去除向地性影响后, 植物根对水势梯度的反应更明显(Takahashi et al.,1999). ...
1 2016
... 有研究表明, 植物根因重力刺激发生向地性弯曲后, 弯曲根的凹面处活性氧(reactive oxygen species, ROS)积累增加, 从而抑制凹面处的生长, 但这种根中ROS的单侧积累并不会发生在向水性弯曲生长过程中(Krieger et al., 2016).另外, 提高植物根中细胞质的钙离子浓度有助于根对水势梯度的感知, 但其作用机制并不清楚(Takano et al., 1997). ...
2 1989
... 植物根柱细胞中的淀粉粒在根的重力感知中起重要作用(Kiss, 2000; Perrin et al., 2005).研究发现, 拟南芥(Arabidopsis thaliana)主根根柱细胞中的淀粉粒在水势梯度刺激后会迅速降解, 导致根的重力敏感性降低, 同时促进其向水性反应(Takahashi et al., 2003).但根柱细胞内淀粉粒的降解不仅受到水势梯度的诱导, 而且也受到其它不同胁迫(如干旱)的调控 (Takahashi et al., 2003; Sun et al., 2008; Ding and Friml, 2010; Iyer-Pascuzzi et al., 2011).因此, 尚不能确定向水性反应中的淀粉粒降解是由水势梯度特异诱导的还是受干旱诱导发生的.而且在一些植物中, 干旱会促进根的向地性反应(Leopold and LaFavre, 1989), 说明淀粉粒的降解可能对诱导植物根的向水性反应并非必需(Nakayama et al., 2012). ... ... 脱落酸作为植物体内的抗逆诱导因子, 在植物干旱、高盐和低温等逆境胁迫反应中起重要作用, 因此被称为植物的“胁迫激素”.在玉米(Zea mays)根中, 脱落酸以光依赖的形式参与向地性反应, 并在干旱胁迫时作为调节因子改变玉米根的向地性反应(Leopold and LaFavre, 1989).之后, Han等(2009)的研究发现, 脱落酸可以抑制植物根的向地性, 但却不抑制生长素的极性运输. ...
1 1993
... 1991年, Takahashi在对豌豆突变体“Ageotro- pum”的向水性研究中发现, 当对Ageotropum施加生长素转运抑制剂三碘苯甲酸(TIBA)时, 其根的向水性反应被明显抑制, 说明生长素的极性运输在植物根的向水性反应过程中具有重要作用(Takahashi and Suge, 1991).然而生长素的调节效应具有种间差异.在豌豆、黄瓜和水稻(Oryza sativa)根尖, 生长素以重新分布的模式调控其根的向地性以及向水性弯曲(Takahashi and Suge, 1991; Mizuno et al., 2002; Nakajima et al., 2017); 但与豌豆同属豆科植物的百脉根(Lotus corniculatus)和十字花科的拟南芥根向水性反应却不依赖于生长素的极性运输与重新分配(Shkolnik et al., 2016; Nakajima et al., 2017).研究表明, 生长素转运抑制剂(CHPAA、NPA和TIBA)不会影响百脉根和拟南芥根的向水性反应(Kaneyasu et al., 2007; Nakajima et al., 2017), 但生长素合成抑制剂Kyn会明显减弱百脉根的向水性反应(Nakajima et al., 2017).此外, PINs是植物体内调节生长素运输方向的蛋白, 它们以极性定位的方式参与生长素在植物细胞间的极性运输(Michniewicz et al., 2007).拟南芥中, PIN2缺失突变体表现出向地性反应缺陷, 但却具有正常的向水性反应(Takahashi et al., 2002).生长素信号通路受损的拟南芥突变体axr1-3和axr2-1根的向地性反应虽然减弱(Leyser et al., 1993; Nagpal et al., 2000), 但其根对水势梯度响应的敏感性增强(Takahashi et al., 2002).研究发现, 超表达MIZ1能增强拟南芥根的向水性反应, 同时其根中生长素合成基因的表达水平明显降低(Moriwaki et al., 2011; Miyazawa et al., 2012).总之, 生长素通过极性运输和重新分布的方式参与植物根的向地性反应, 但该作用机理并不适用于所有植物(如百脉根和拟南芥)根的向水性反应, 有关生长素对向水性反应调控的种间差异及生理和分子基础有待进一步研究. ...
1 2014
... 植物在整个生命活动过程中都在被动地适应不断变化的环境.在长期的进化选择中, 植物进化出一系列特有的机制来适应周围环境的变化, 使其能够一代代繁衍生息.植物能够感知自身生长环境中各种非生物(包括光、重力、水、温度、氧气和二氧化碳等)和生物信号, 并做出相应的生理生化反应.向性反应(tropic response)作为植物适应周围环境变化的重要机制之一, 是植物实现自身趋利避害的主要途径.植物朝向环境信号生长的向性反应称为正(positive)向性反应, 反之则称为负(negative)向性反应.植物为了应对不同的环境信号进化出了一系列向性反应, 包括向光性(phototropism) (Van Overbeek,1939; Gehr- ing et al., 1990; Liscum et al., 2014)、向地性(gravi- tropism) (Gehring et al., 1990; Hart, 1990; Taka- hashi and Jaffe, 1990; Correll and Kiss, 2002)、向化性(chemotropism) (Esmon et al., 2005)、向触性(thigmotropism) (Monshausen et al., 2008)、向水性(hydrotropism) (Takahashi and Jaffe, 1990; Takaha- shi, 1997)、避盐性(halotropism) (Munns and Tes- ter, 2008; Sun et al., 2008; Shelef et al., 2010; Galvan-Ampudia et al., 2013)及向声性(phonotro pism) (Mishra et al., 2016; Rodrigo-Moreno et al., 2017)等.其中, 向地性和向光性由于较易观察和筛选等原因, 研究进展较快, 目前已对调控植物向地性和向光性的分子机理有了较为深入的认识.由于重力等因素的干扰, 根向水性的研究较为困难.目前的研究仅发现少数基因和突变体参与或改变植物的向水性反应.向水性机理研究不仅具有重要的理论意义, 而且对节水灌溉和提高植物的抗旱性等也具有指导意义. ...

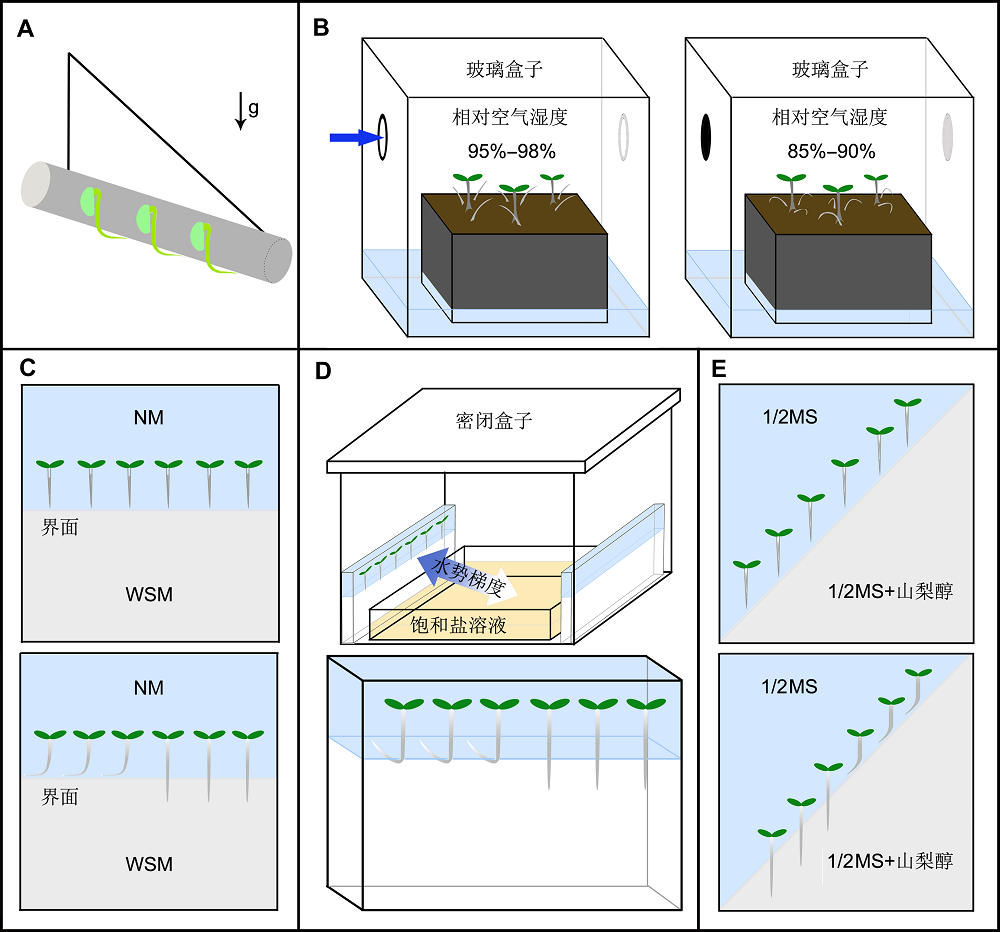
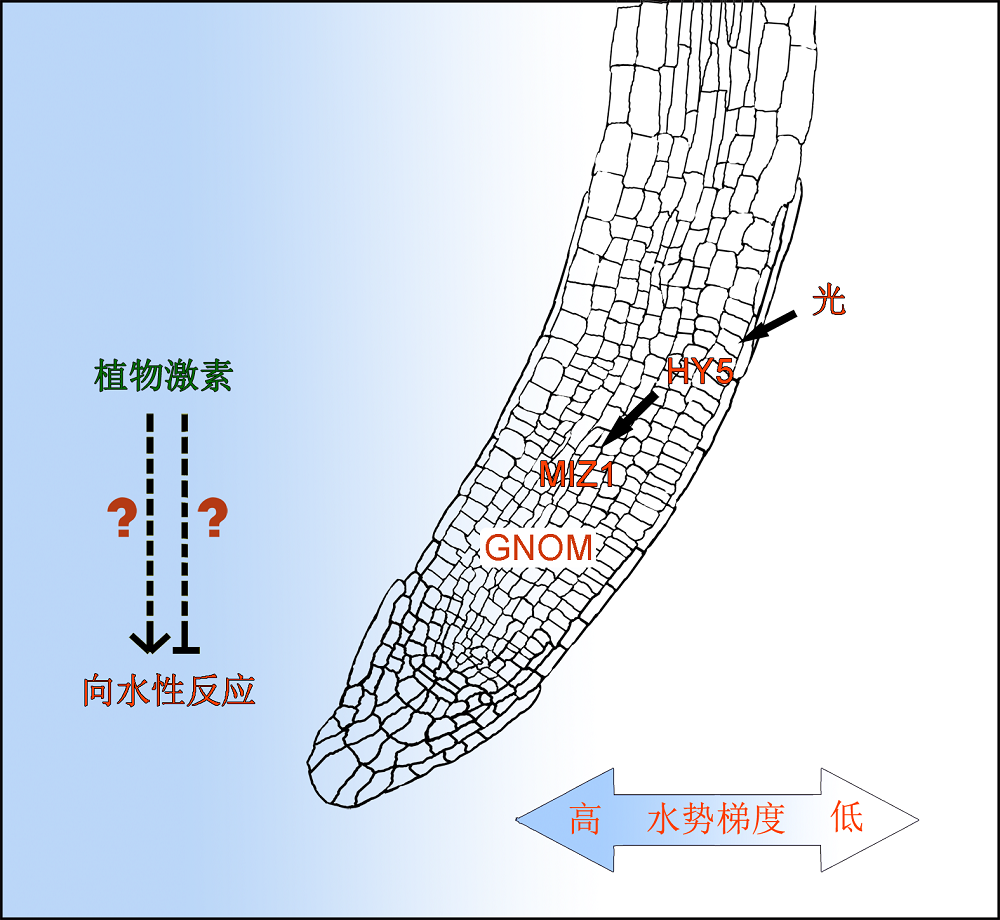

{kind=link}
{kind=link}
{kind=link}
{kind=link}