Abstract: T-DNA mutants are important resources for research of gene function. High-efficiency thermal asymmetric interlaced PCR (hiTAIL-PCR) is widely used for cloning the flanking sequence near the T-DNA insertion sites. However, we found that some cloned flanking fragments in hiTAIL-PCR products corresponded not to the host genomic DNA but to the plasmid backbone DNA. In this study, with a control of the RB-S4/AC1 or LB-A4/AC1 product, we amplified PCR fragments from the plasmid backbone DNA. By excluding them from further analysis, we amplified fragments from the unknown genomic DNA more effectively. Meanwhile, by adjusting the PCR programs, the whole PCR time was greatly shortened. In cloning the flanking sequence of Arabidopsis thaliana T-DNA mutant drf1, our method with hiTAIL-PCR reduced the total 22 DNA bands required for further checking to 4 bands, which improved the efficiency by 81.8%.
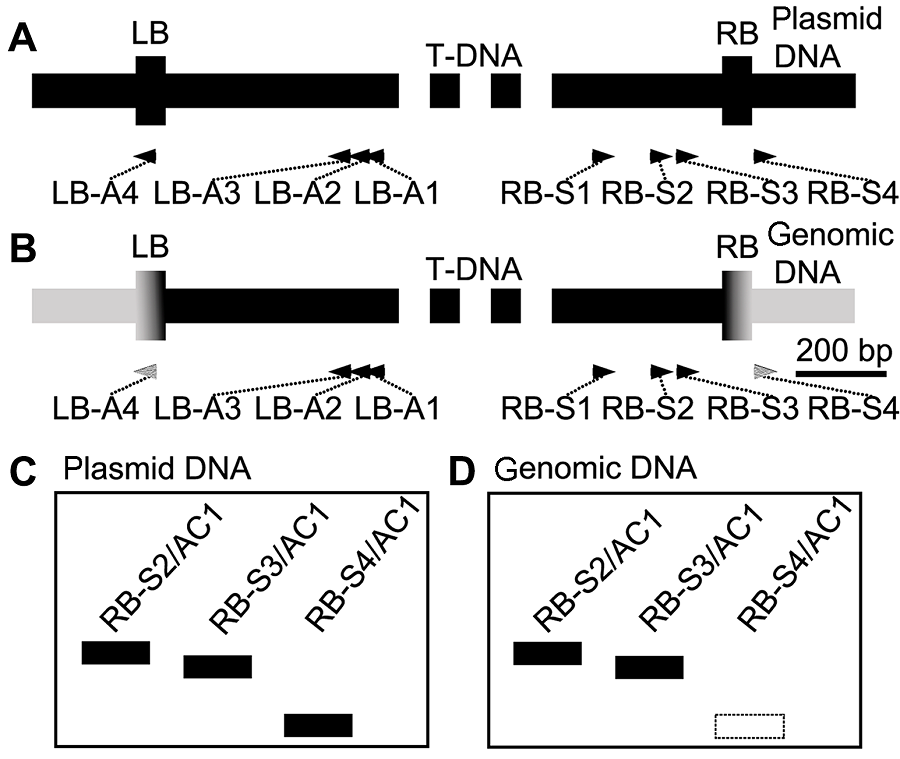
2 结果与讨论2.1 设计原理基于hiTAIL-PCR的原理, 我们增设对照RB-S4/AC1 (或LB-A4/AC1)扩增产物来鉴别侧翼序列的特异性。RB-S4结合于质粒RB的下游, LB-A4结合于质粒LB的上游(图1A)。第1轮PCR仍以RB-S1 (或者LB-A1)与多条LAD1随机引物进行反应以初次扩增侧翼序列; 第2轮反应以第1轮反应产物为模板, 分别进行RB- S2/AC1 (或LB-A2/AC1)、RB-S3/AC1 (或LB-A3/ AC1)及RB-S4/AC1 (或LB-A4/AC1) 3组反应。以RB区域为例, 当RB-S2/AC1与RB-S3/AC1扩增出的侧翼序列为质粒骨架片段时, 则RB-S4/AC1也应扩增出相应片段(图1A, C); 当RB-S2/AC1与RB-S3/AC1扩增出的侧翼序列是来自宿主染色体时, 则RB-S4/ AC1不应扩增出相应片段(图1B, D)。 图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-104/img_1.png<b>图1</b> 设计原理<br/>(A) T-DNA质粒RB和LB处区域; (B) T-DNA质粒与染色体DNA整合后RB和LB处区域; (C) 当hiTAIL-PCR扩增质粒骨架时, RB-S2/AC1、RB-S3/AC1与RB-S4/AC1都将产生相应扩增; (D) 当hiTAIL-PCR扩增基因组DNA时, RB-S4/AC1将不产生相应扩增(虚线框示未扩增)<br/><b>Figure 1</b> The principle of design<br/>(A) The regions of RB and LB of T-DNA plasmid; (B) The regions of RB and LB which integrate with chromosomal DNA; (C) When hiTAIL-PCR amplifies the backbone of plas- mid, the three PCR groups containing the RB-S2/AC1, RB-S3/AC1 and RB-S4/AC1 primer pairs, respectively, will all produce the positive bands; (D) When hiTAIL-PCR amplifies the genomic DNA, RB-S4/AC1 will not produce the positive bands (dashed frame) Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-104/img_1.png<b>图1</b> 设计原理<br/>(A) T-DNA质粒RB和LB处区域; (B) T-DNA质粒与染色体DNA整合后RB和LB处区域; (C) 当hiTAIL-PCR扩增质粒骨架时, RB-S2/AC1、RB-S3/AC1与RB-S4/AC1都将产生相应扩增; (D) 当hiTAIL-PCR扩增基因组DNA时, RB-S4/AC1将不产生相应扩增(虚线框示未扩增)<br/><b>Figure 1</b> The principle of design<br/>(A) The regions of RB and LB of T-DNA plasmid; (B) The regions of RB and LB which integrate with chromosomal DNA; (C) When hiTAIL-PCR amplifies the backbone of plas- mid, the three PCR groups containing the RB-S2/AC1, RB-S3/AC1 and RB-S4/AC1 primer pairs, respectively, will all produce the positive bands; (D) When hiTAIL-PCR amplifies the genomic DNA, RB-S4/AC1 will not produce the positive bands (dashed frame)
图1 设计原理 (A) T-DNA质粒RB和LB处区域; (B) T-DNA质粒与染色体DNA整合后RB和LB处区域; (C) 当hiTAIL-PCR扩增质粒骨架时, RB-S2/AC1、RB-S3/AC1与RB-S4/AC1都将产生相应扩增; (D) 当hiTAIL-PCR扩增基因组DNA时, RB-S4/AC1将不产生相应扩增(虚线框示未扩增) Figure 1 The principle of design (A) The regions of RB and LB of T-DNA plasmid; (B) The regions of RB and LB which integrate with chromosomal DNA; (C) When hiTAIL-PCR amplifies the backbone of plas- mid, the three PCR groups containing the RB-S2/AC1, RB-S3/AC1 and RB-S4/AC1 primer pairs, respectively, will all produce the positive bands; (D) When hiTAIL-PCR amplifies the genomic DNA, RB-S4/AC1 will not produce the positive bands (dashed frame)
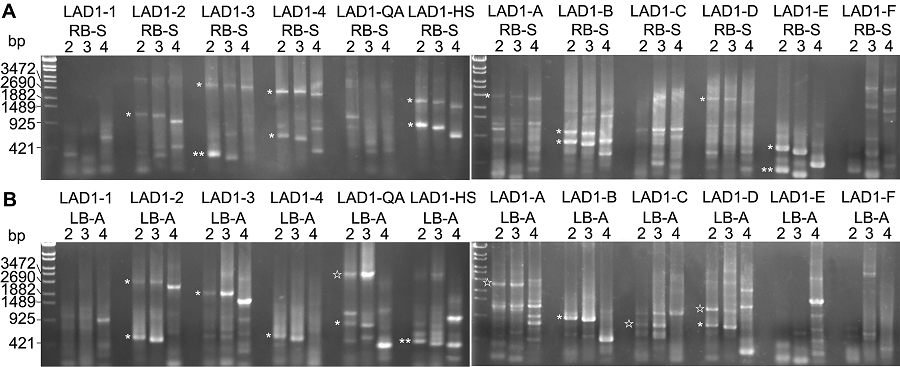
2.2 有效性验证为验证该方法的有效性, 我们选取了1株拟南芥突变体drf1进行了测试。经鉴定, 该突变体基因组至少包含1个完整质粒。我们先扩增RB处的侧翼序列。实验结果表明, 尽管扩增出11组RB-S2/AC1与RB-S3/ AC1“hiTAIL-PCR特异性片段”, 但是, 所有产物都有对应的RB-S4/AC1扩增片段(图2A, *所示), 因此, 这些扩增产物均为质粒骨架片段, 而非染色体DNA片段。我们也扩增了LB处的侧翼序列, 在11组“hiTAIL-PCR特异性片段”中, 仅4组无对应的LB-A4/AC1扩增片段, 暗示其为潜在的宿主染色体DNA特异性片段(图2B, ☆所示)。我们对这4条DNA片段(LB-A2/AC1扩增产物)进行了纯化回收, 并通过测序进一步验证其特异性。结果表明, 仅LAD1-A随机引物所对应的侧翼序列来自染色体DNA (图3)。 图2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-104/img_2.png<b>图2</b> 拟南芥突变体<i>drf1</i> T-DNA插入位点侧翼序列的克隆<br/>(A) 用RB-S系列引物扩增的第2轮PCR的结果; (B) 用LB-A系列引物扩增的第2轮PCR的结果。* 代表非特异性扩增结果; ☆代表潜在的特异性扩增结果; ** 代表来自RB之前或LB之后的T-DNA片段扩增结果。<br/><b>Figure 2</b> The cloning of flanking sequence at the T-DNA insertion site of Arabidopsis mutant <i>drf1<br/></i>(A) The second round results amplified by PCR with RB-S serial primers; (B) The second round results amplified by PCR with LB-A serial primers. * display the non-specific amplification results; ☆ represent the potential specific amplification results; ** show the amplification results from the T-DNA region before RB or after LB. Figure 2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-104/img_2.png<b>图2</b> 拟南芥突变体<i>drf1</i> T-DNA插入位点侧翼序列的克隆<br/>(A) 用RB-S系列引物扩增的第2轮PCR的结果; (B) 用LB-A系列引物扩增的第2轮PCR的结果。* 代表非特异性扩增结果; ☆代表潜在的特异性扩增结果; ** 代表来自RB之前或LB之后的T-DNA片段扩增结果。<br/><b>Figure 2</b> The cloning of flanking sequence at the T-DNA insertion site of Arabidopsis mutant <i>drf1<br/></i>(A) The second round results amplified by PCR with RB-S serial primers; (B) The second round results amplified by PCR with LB-A serial primers. * display the non-specific amplification results; ☆ represent the potential specific amplification results; ** show the amplification results from the T-DNA region before RB or after LB.
图2 拟南芥突变体drf1 T-DNA插入位点侧翼序列的克隆 (A) 用RB-S系列引物扩增的第2轮PCR的结果; (B) 用LB-A系列引物扩增的第2轮PCR的结果。* 代表非特异性扩增结果; ☆代表潜在的特异性扩增结果; ** 代表来自RB之前或LB之后的T-DNA片段扩增结果。 Figure 2 The cloning of flanking sequence at the T-DNA insertion site of Arabidopsis mutant drf1 (A) The second round results amplified by PCR with RB-S serial primers; (B) The second round results amplified by PCR with LB-A serial primers. * display the non-specific amplification results; ☆ represent the potential specific amplification results; ** show the amplification results from the T-DNA region before RB or after LB.
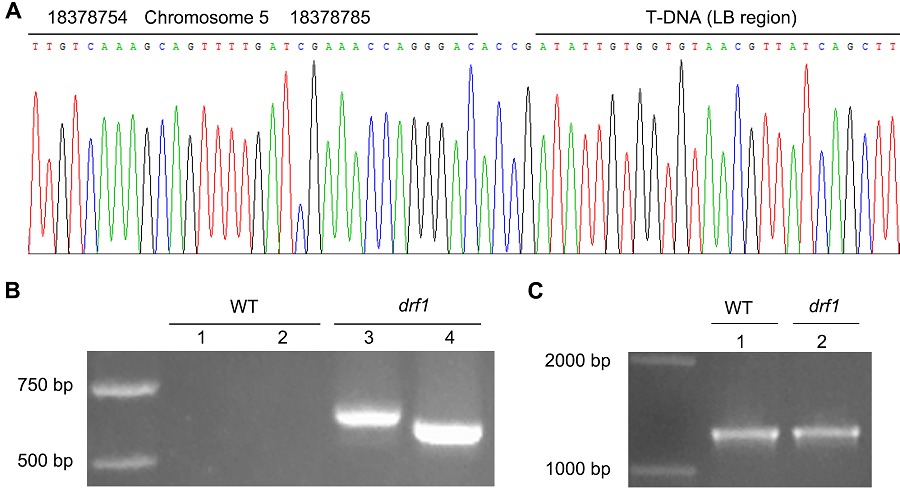
图3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-104/img_3.png<b>图3</b> 拟南芥突变体<i>drf1</i>侧翼序列的PCR验证<br/>(A) 部分测序结果; (B) 用引物对DRF1-S/LB-A2 (1,3)及DRF1-S/LB-A3 (2,4)扩增野生型(WT)与<i>drf1</i>突变体基因组的结果; (C) 用引物对QRT-S/QRT-A扩增野生型(WT)与<i>drf1</i>突变体基因组的结果<br/><b>Figure 3</b> The PCR confirmation for Arabidopsis mutant <i>drf1</i> flanking sequence<br/>(A) The partial sequencing result; (B) The results amplifying the genomic DNA from wild type (WT) and <i>drf1 </i>mutant with primer pairs of DRF1-S/LB-A2 (1,3) or DRF1-S/LB-A3 (2,4); (C) The PCR results with primer pairs of QRT-S/QRT-A to amplifying WT and <i>drf1</i> mutant genomic DNA Figure 3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-104/img_3.png<b>图3</b> 拟南芥突变体<i>drf1</i>侧翼序列的PCR验证<br/>(A) 部分测序结果; (B) 用引物对DRF1-S/LB-A2 (1,3)及DRF1-S/LB-A3 (2,4)扩增野生型(WT)与<i>drf1</i>突变体基因组的结果; (C) 用引物对QRT-S/QRT-A扩增野生型(WT)与<i>drf1</i>突变体基因组的结果<br/><b>Figure 3</b> The PCR confirmation for Arabidopsis mutant <i>drf1</i> flanking sequence<br/>(A) The partial sequencing result; (B) The results amplifying the genomic DNA from wild type (WT) and <i>drf1 </i>mutant with primer pairs of DRF1-S/LB-A2 (1,3) or DRF1-S/LB-A3 (2,4); (C) The PCR results with primer pairs of QRT-S/QRT-A to amplifying WT and <i>drf1</i> mutant genomic DNA
图3 拟南芥突变体drf1侧翼序列的PCR验证 (A) 部分测序结果; (B) 用引物对DRF1-S/LB-A2 (1,3)及DRF1-S/LB-A3 (2,4)扩增野生型(WT)与drf1突变体基因组的结果; (C) 用引物对QRT-S/QRT-A扩增野生型(WT)与drf1突变体基因组的结果 Figure 3 The PCR confirmation for Arabidopsis mutant drf1 flanking sequence (A) The partial sequencing result; (B) The results amplifying the genomic DNA from wild type (WT) and drf1 mutant with primer pairs of DRF1-S/LB-A2 (1,3) or DRF1-S/LB-A3 (2,4); (C) The PCR results with primer pairs of QRT-S/QRT-A to amplifying WT and drf1 mutant genomic DNA
CoulondreC, MillerJH (1977). Genetic studies of the lac repressor: III. Additional correlation of mutational sites with specific amino acid residues. J Mol Biol 117, 525-567. [本文引用: 1]
[4]
HellensRP, EdwardsEA, LeylandNR, BeanS, MullineauxPM (2000). pGreen: a versatile and flexible binary Ti vector for Agrobacterium-mediated plant transformation. Plant Mol Biol 42, 819-832. [本文引用: 1]
[5]
JanderG (2006). Gene identification and cloning by molecular marker mapping.Methods Mol Biol 323, 115-126. [本文引用: 1]
[6]
JanderG, NorrisSR, RounsleySD, BushDF, LevinIM, LastRL (2002). Arabidopsis map-based cloning in the post-genome era.Plant Physiol 129, 440-450. [本文引用: 1]
[7]
KleinboeltingN, HuepG, AppelhagenI, ViehoeverP, LiY, WeisshaarB (2015). The structural features of thousands of T-DNA insertion sites are consistent with a double- strand break repair-based insertion mechanism.Mol Plant 8, 1651-1664. [本文引用: 2]
[8]
LiuYG, ChenYL (2007). High-efficiency thermal asymmetric interlaced PCR for amplification of unknown flanking sequences.Biotechniques 43, 649-650. [本文引用: 5]
[9]
LiuYG, MitsukawaN, OosumiT, WhittierRF (1995). Efficient isolation and mapping of Arabidopsis thaliana T-DNA insert junctions by thermal asymmetric interlaced PCR. Plant J 8, 457-463. [本文引用: 1]
RileyJ, ButlerR, OgilvieD, FinniearR, JennerD, PowellS, AnandR, SmithJC, MarkhamAF (1990). A novel, rapid method for the isolation of terminal sequences from yeast artificial chromosome (YAC) clones.Nucleic Acids Res 18, 2887-2890. [本文引用: 1]
[15]
StachelSE, TimmermanB, ZambryskiP (1987). Activation of Agrobacterium tumefaciens vir gene expression generates multiple single-stranded T-strand molecules from the pTiA6 T-region: requirement for 5' virD gene products. EMBO J 6, 857-863. [本文引用: 1]
[16]
TrigliaT, PetersonMG, KempDJ (1988). A procedure for in vitro amplification of DNA segments that lie outside the boundaries of known sequences. Nucleic Acids Res 16, 8186. [本文引用: 1]
[17]
TzfiraT, LiJX, LacroixB, CitovskyV (2004). Agrobacterium T-DNA integration: molecules and models.Trends Ge- net 20, 375-383. [本文引用: 1]
[18]
WangHR, FangJ, LiangCZ, HeMH, LiQY, ChuCC (2011). Computation-assisted SiteFinding-PCR for isolating flanking sequence tags in rice.Biotechniques 51, 421-423. [本文引用: 1]
[19]
ZhouZW, MaHY, QuLJ, XieF, MaQW, RenZR (2012). Establishment of an improved high-efficiency thermal asym- metric interlaced PCR for identification of genomic integration sites mediated by phiC31 integrase.World J Microbiol Biotechnol 28, 1295-1299. [本文引用: 1]
1 2012
... 遗传学方法是现代分子生物学研究基因功能的重要方法.通过大规模筛选, 能够发现携带相关表型的突变体, 进而发现产生表型的基因.EMS是早期广泛使用的诱变剂, 它通过诱导染色体DNA发生单碱基改变而产生突变体(Coulondre and Miller, 1977).对于EMS突变体, 因需通过图位克隆或者深度测序技术来寻找突变基因, 致使后期的鉴定实验耗时费力或成本增大(Jander et al., 2002; Jander, 2006; Abe et al., 2012).近年来, 基于T-DNA插入获得突变体的方法被大量使用(Alonso et al., 2003).利用农杆菌(Agrobacterium)转化宿主植物, T-DNA可通过左边界(left border, LB)和右边界(right border, RB)区域随机整合至宿主染色体中(Mayerhofer et al., 1991), 从而导致插入位点的序列改变而产生突变体.与EMS突变体相比, T-DNA突变体具有插入位点易克隆的优点.此外, 由于农杆菌介导植物转化技术日渐成熟, 很多实验室都可以通过这种方法建立各自研究材料的T-DNA突变体库. ...
1 2003
... 遗传学方法是现代分子生物学研究基因功能的重要方法.通过大规模筛选, 能够发现携带相关表型的突变体, 进而发现产生表型的基因.EMS是早期广泛使用的诱变剂, 它通过诱导染色体DNA发生单碱基改变而产生突变体(Coulondre and Miller, 1977).对于EMS突变体, 因需通过图位克隆或者深度测序技术来寻找突变基因, 致使后期的鉴定实验耗时费力或成本增大(Jander et al., 2002; Jander, 2006; Abe et al., 2012).近年来, 基于T-DNA插入获得突变体的方法被大量使用(Alonso et al., 2003).利用农杆菌(Agrobacterium)转化宿主植物, T-DNA可通过左边界(left border, LB)和右边界(right border, RB)区域随机整合至宿主染色体中(Mayerhofer et al., 1991), 从而导致插入位点的序列改变而产生突变体.与EMS突变体相比, T-DNA突变体具有插入位点易克隆的优点.此外, 由于农杆菌介导植物转化技术日渐成熟, 很多实验室都可以通过这种方法建立各自研究材料的T-DNA突变体库. ...
1 1977
... 遗传学方法是现代分子生物学研究基因功能的重要方法.通过大规模筛选, 能够发现携带相关表型的突变体, 进而发现产生表型的基因.EMS是早期广泛使用的诱变剂, 它通过诱导染色体DNA发生单碱基改变而产生突变体(Coulondre and Miller, 1977).对于EMS突变体, 因需通过图位克隆或者深度测序技术来寻找突变基因, 致使后期的鉴定实验耗时费力或成本增大(Jander et al., 2002; Jander, 2006; Abe et al., 2012).近年来, 基于T-DNA插入获得突变体的方法被大量使用(Alonso et al., 2003).利用农杆菌(Agrobacterium)转化宿主植物, T-DNA可通过左边界(left border, LB)和右边界(right border, RB)区域随机整合至宿主染色体中(Mayerhofer et al., 1991), 从而导致插入位点的序列改变而产生突变体.与EMS突变体相比, T-DNA突变体具有插入位点易克隆的优点.此外, 由于农杆菌介导植物转化技术日渐成熟, 很多实验室都可以通过这种方法建立各自研究材料的T-DNA突变体库. ...
1 2000
... 拟南芥(Arabidopsis thaliana L.)突变体drf1种植于西北大学人工温室, 室内温度为23°C, 光周期为16小时光照/8小时黑暗.该突变体携带的T-DNA来自pGreenII0029质粒(Hellens et al., 2000). ...
1 2006
... 遗传学方法是现代分子生物学研究基因功能的重要方法.通过大规模筛选, 能够发现携带相关表型的突变体, 进而发现产生表型的基因.EMS是早期广泛使用的诱变剂, 它通过诱导染色体DNA发生单碱基改变而产生突变体(Coulondre and Miller, 1977).对于EMS突变体, 因需通过图位克隆或者深度测序技术来寻找突变基因, 致使后期的鉴定实验耗时费力或成本增大(Jander et al., 2002; Jander, 2006; Abe et al., 2012).近年来, 基于T-DNA插入获得突变体的方法被大量使用(Alonso et al., 2003).利用农杆菌(Agrobacterium)转化宿主植物, T-DNA可通过左边界(left border, LB)和右边界(right border, RB)区域随机整合至宿主染色体中(Mayerhofer et al., 1991), 从而导致插入位点的序列改变而产生突变体.与EMS突变体相比, T-DNA突变体具有插入位点易克隆的优点.此外, 由于农杆菌介导植物转化技术日渐成熟, 很多实验室都可以通过这种方法建立各自研究材料的T-DNA突变体库. ...
1 2002
... 遗传学方法是现代分子生物学研究基因功能的重要方法.通过大规模筛选, 能够发现携带相关表型的突变体, 进而发现产生表型的基因.EMS是早期广泛使用的诱变剂, 它通过诱导染色体DNA发生单碱基改变而产生突变体(Coulondre and Miller, 1977).对于EMS突变体, 因需通过图位克隆或者深度测序技术来寻找突变基因, 致使后期的鉴定实验耗时费力或成本增大(Jander et al., 2002; Jander, 2006; Abe et al., 2012).近年来, 基于T-DNA插入获得突变体的方法被大量使用(Alonso et al., 2003).利用农杆菌(Agrobacterium)转化宿主植物, T-DNA可通过左边界(left border, LB)和右边界(right border, RB)区域随机整合至宿主染色体中(Mayerhofer et al., 1991), 从而导致插入位点的序列改变而产生突变体.与EMS突变体相比, T-DNA突变体具有插入位点易克隆的优点.此外, 由于农杆菌介导植物转化技术日渐成熟, 很多实验室都可以通过这种方法建立各自研究材料的T-DNA突变体库. ...
2 2015
... 尽管hiTAIL-PCR在克隆侧翼序列方面显示了其高效性, 但应用到对T-DNA突变体侧翼序列的克隆时, 却经常出现一些事实上是质粒骨架片段而非染色体DNA片段的“特异性扩增结果”.通常情况下, 农杆菌仅在LB-RB之间产生可转移的单链T-DNA分子, 从而导致整合入宿主植物染色体中的仅为T-DNA区域(Tzfira et al., 2004).但是也有证据表明, T-DNA可连同质粒骨架作为一个分子整合至宿主植物的基因组中(Martineau et al., 1994; Kleinboelting et al., 2015).虽然质粒骨架随T-DNA分子一起整合至宿主植物染色体中的具体原因及机制仍不清楚, 但早期发现的“通读”现象(Stachel et al., 1987; Klein- boelting et al., 2015)可能是其原因之一.克隆这些转基因系插入位点的侧翼序列, 容易产生假阳性结果.其原因为只要随机引物LAD1能在质粒骨架上引发扩增, hiTAIL-PCR就能将部分骨架片段大量地扩增出来.然而, 由于在DNA片段测序之前, 我们无法判断这些“hiTAIL-PCR特异性片段”是来自质粒骨架还是突变体染色体DNA, 因此, 大量的后期鉴定工作将无法避免. ... ... ; Klein- boelting et al., 2015)可能是其原因之一.克隆这些转基因系插入位点的侧翼序列, 容易产生假阳性结果.其原因为只要随机引物LAD1能在质粒骨架上引发扩增, hiTAIL-PCR就能将部分骨架片段大量地扩增出来.然而, 由于在DNA片段测序之前, 我们无法判断这些“hiTAIL-PCR特异性片段”是来自质粒骨架还是突变体染色体DNA, 因此, 大量的后期鉴定工作将无法避免. ...
5 2007
... 有许多方法可用于克隆T-DNA插入位点的侧翼序列, 如质粒拯救(Nan and Walbot, 2009)、反向PCR (Triglia et al., 1988)、接头介导的PCR (Mueller and Wold, 1989; Riley et al., 1990)以及热不对称交错PCR (TAIL-PCR) (Liu et al., 1995)等.其中, TAIL- PCR由于操作简便及对突变体基因组DNA需求量少等特点, 成为大多数研究的首选方法.尤其是改进后的hiTAIL-PCR, 因具有更高效的克隆效率及操作简便而被广泛使用(Liu and Chen, 2007).该方法利用3条特异性引物(SP1、SP2和SP3)及多条随机引物(LAD1系列引物)来克隆未知侧翼序列.LAD1是简并引物, 能结合染色体DNA上的多个位点, 从而使SP1/LAD1初次扩增的侧翼序列并无较高的特异性; 在此基础上, SP2与SP3被用于第2轮及第3轮PCR扩增, 以进一步提高产物的特异性.简并引物在hiTAIL- PCR中扮演重要角色, 它的改进可进一步提高hiTAIL- PCR的效率(Wang et al., 2011).与TAIL-PCR相比, hiTAIL-PCR的随机引物LAD1的5'端都带有一段AC1序列, 这使得第2轮及第3轮PCR操作大为简化(Liu and Chen, 2007).获得的PCR产物经凝胶电泳检测, 可通过比较SP2/AC1与SP3/AC1扩增产物的分子量大小判断其特异性(Liu and Chen, 2007). ... ... ).与TAIL-PCR相比, hiTAIL-PCR的随机引物LAD1的5'端都带有一段AC1序列, 这使得第2轮及第3轮PCR操作大为简化(Liu and Chen, 2007).获得的PCR产物经凝胶电泳检测, 可通过比较SP2/AC1与SP3/AC1扩增产物的分子量大小判断其特异性(Liu and Chen, 2007). ... ... ).获得的PCR产物经凝胶电泳检测, 可通过比较SP2/AC1与SP3/AC1扩增产物的分子量大小判断其特异性(Liu and Chen, 2007). ... ... 第1轮PCR反应完全使用hiTAIL-PCR中的预扩增(pre-amplification)程序, 第2轮反应则使用略微调整的初次扩增(primary TAIL-PCR)程序(退火温度由68°C调整至65°C) (Liu and Chen, 2007).根据T-DNA序列设计的特异性引物以及随机引物如表1所示.引物RB-S2与RB-S3及RB-S3与RB-S4之间的核苷酸距离分别为51 bp和183 bp; 引物LB-A2与LB-A3及LB-A3与LB-A4之间的核苷酸距离分别为45 bp和397 bp. ... ... 根据hiTAIL-PCR的设计原理(Liu and Chen, 2007), 对照组的引入将使其前后总共需进行4轮PCR, 这无疑导致实验时间的延长.因此, 我们进一步调整了hiTAIL-PCR的程序, 将RB-S2/AC1、RB- S3/AC1和RB-S4/AC1 (或LB-A2/AC1、LB-A3/AC1和LB-A4/AC1)的扩增反应同时放在第2轮PCR中进行.这样, 就将原来的4轮PCR缩短为2轮, 从而大大节省了实验耗时.从我们已完成测试的实验来看, 迄今还未发现该程序不能有效工作的情况. ...
1 1995
... 有许多方法可用于克隆T-DNA插入位点的侧翼序列, 如质粒拯救(Nan and Walbot, 2009)、反向PCR (Triglia et al., 1988)、接头介导的PCR (Mueller and Wold, 1989; Riley et al., 1990)以及热不对称交错PCR (TAIL-PCR) (Liu et al., 1995)等.其中, TAIL- PCR由于操作简便及对突变体基因组DNA需求量少等特点, 成为大多数研究的首选方法.尤其是改进后的hiTAIL-PCR, 因具有更高效的克隆效率及操作简便而被广泛使用(Liu and Chen, 2007).该方法利用3条特异性引物(SP1、SP2和SP3)及多条随机引物(LAD1系列引物)来克隆未知侧翼序列.LAD1是简并引物, 能结合染色体DNA上的多个位点, 从而使SP1/LAD1初次扩增的侧翼序列并无较高的特异性; 在此基础上, SP2与SP3被用于第2轮及第3轮PCR扩增, 以进一步提高产物的特异性.简并引物在hiTAIL- PCR中扮演重要角色, 它的改进可进一步提高hiTAIL- PCR的效率(Wang et al., 2011).与TAIL-PCR相比, hiTAIL-PCR的随机引物LAD1的5'端都带有一段AC1序列, 这使得第2轮及第3轮PCR操作大为简化(Liu and Chen, 2007).获得的PCR产物经凝胶电泳检测, 可通过比较SP2/AC1与SP3/AC1扩增产物的分子量大小判断其特异性(Liu and Chen, 2007). ...
1 1994
... 尽管hiTAIL-PCR在克隆侧翼序列方面显示了其高效性, 但应用到对T-DNA突变体侧翼序列的克隆时, 却经常出现一些事实上是质粒骨架片段而非染色体DNA片段的“特异性扩增结果”.通常情况下, 农杆菌仅在LB-RB之间产生可转移的单链T-DNA分子, 从而导致整合入宿主植物染色体中的仅为T-DNA区域(Tzfira et al., 2004).但是也有证据表明, T-DNA可连同质粒骨架作为一个分子整合至宿主植物的基因组中(Martineau et al., 1994; Kleinboelting et al., 2015).虽然质粒骨架随T-DNA分子一起整合至宿主植物染色体中的具体原因及机制仍不清楚, 但早期发现的“通读”现象(Stachel et al., 1987; Klein- boelting et al., 2015)可能是其原因之一.克隆这些转基因系插入位点的侧翼序列, 容易产生假阳性结果.其原因为只要随机引物LAD1能在质粒骨架上引发扩增, hiTAIL-PCR就能将部分骨架片段大量地扩增出来.然而, 由于在DNA片段测序之前, 我们无法判断这些“hiTAIL-PCR特异性片段”是来自质粒骨架还是突变体染色体DNA, 因此, 大量的后期鉴定工作将无法避免. ...
1 1991
... 遗传学方法是现代分子生物学研究基因功能的重要方法.通过大规模筛选, 能够发现携带相关表型的突变体, 进而发现产生表型的基因.EMS是早期广泛使用的诱变剂, 它通过诱导染色体DNA发生单碱基改变而产生突变体(Coulondre and Miller, 1977).对于EMS突变体, 因需通过图位克隆或者深度测序技术来寻找突变基因, 致使后期的鉴定实验耗时费力或成本增大(Jander et al., 2002; Jander, 2006; Abe et al., 2012).近年来, 基于T-DNA插入获得突变体的方法被大量使用(Alonso et al., 2003).利用农杆菌(Agrobacterium)转化宿主植物, T-DNA可通过左边界(left border, LB)和右边界(right border, RB)区域随机整合至宿主染色体中(Mayerhofer et al., 1991), 从而导致插入位点的序列改变而产生突变体.与EMS突变体相比, T-DNA突变体具有插入位点易克隆的优点.此外, 由于农杆菌介导植物转化技术日渐成熟, 很多实验室都可以通过这种方法建立各自研究材料的T-DNA突变体库. ...
1 1989
... 有许多方法可用于克隆T-DNA插入位点的侧翼序列, 如质粒拯救(Nan and Walbot, 2009)、反向PCR (Triglia et al., 1988)、接头介导的PCR (Mueller and Wold, 1989; Riley et al., 1990)以及热不对称交错PCR (TAIL-PCR) (Liu et al., 1995)等.其中, TAIL- PCR由于操作简便及对突变体基因组DNA需求量少等特点, 成为大多数研究的首选方法.尤其是改进后的hiTAIL-PCR, 因具有更高效的克隆效率及操作简便而被广泛使用(Liu and Chen, 2007).该方法利用3条特异性引物(SP1、SP2和SP3)及多条随机引物(LAD1系列引物)来克隆未知侧翼序列.LAD1是简并引物, 能结合染色体DNA上的多个位点, 从而使SP1/LAD1初次扩增的侧翼序列并无较高的特异性; 在此基础上, SP2与SP3被用于第2轮及第3轮PCR扩增, 以进一步提高产物的特异性.简并引物在hiTAIL- PCR中扮演重要角色, 它的改进可进一步提高hiTAIL- PCR的效率(Wang et al., 2011).与TAIL-PCR相比, hiTAIL-PCR的随机引物LAD1的5'端都带有一段AC1序列, 这使得第2轮及第3轮PCR操作大为简化(Liu and Chen, 2007).获得的PCR产物经凝胶电泳检测, 可通过比较SP2/AC1与SP3/AC1扩增产物的分子量大小判断其特异性(Liu and Chen, 2007). ...
1 2009
... 有许多方法可用于克隆T-DNA插入位点的侧翼序列, 如质粒拯救(Nan and Walbot, 2009)、反向PCR (Triglia et al., 1988)、接头介导的PCR (Mueller and Wold, 1989; Riley et al., 1990)以及热不对称交错PCR (TAIL-PCR) (Liu et al., 1995)等.其中, TAIL- PCR由于操作简便及对突变体基因组DNA需求量少等特点, 成为大多数研究的首选方法.尤其是改进后的hiTAIL-PCR, 因具有更高效的克隆效率及操作简便而被广泛使用(Liu and Chen, 2007).该方法利用3条特异性引物(SP1、SP2和SP3)及多条随机引物(LAD1系列引物)来克隆未知侧翼序列.LAD1是简并引物, 能结合染色体DNA上的多个位点, 从而使SP1/LAD1初次扩增的侧翼序列并无较高的特异性; 在此基础上, SP2与SP3被用于第2轮及第3轮PCR扩增, 以进一步提高产物的特异性.简并引物在hiTAIL- PCR中扮演重要角色, 它的改进可进一步提高hiTAIL- PCR的效率(Wang et al., 2011).与TAIL-PCR相比, hiTAIL-PCR的随机引物LAD1的5'端都带有一段AC1序列, 这使得第2轮及第3轮PCR操作大为简化(Liu and Chen, 2007).获得的PCR产物经凝胶电泳检测, 可通过比较SP2/AC1与SP3/AC1扩增产物的分子量大小判断其特异性(Liu and Chen, 2007). ...
1 1990
... 有许多方法可用于克隆T-DNA插入位点的侧翼序列, 如质粒拯救(Nan and Walbot, 2009)、反向PCR (Triglia et al., 1988)、接头介导的PCR (Mueller and Wold, 1989; Riley et al., 1990)以及热不对称交错PCR (TAIL-PCR) (Liu et al., 1995)等.其中, TAIL- PCR由于操作简便及对突变体基因组DNA需求量少等特点, 成为大多数研究的首选方法.尤其是改进后的hiTAIL-PCR, 因具有更高效的克隆效率及操作简便而被广泛使用(Liu and Chen, 2007).该方法利用3条特异性引物(SP1、SP2和SP3)及多条随机引物(LAD1系列引物)来克隆未知侧翼序列.LAD1是简并引物, 能结合染色体DNA上的多个位点, 从而使SP1/LAD1初次扩增的侧翼序列并无较高的特异性; 在此基础上, SP2与SP3被用于第2轮及第3轮PCR扩增, 以进一步提高产物的特异性.简并引物在hiTAIL- PCR中扮演重要角色, 它的改进可进一步提高hiTAIL- PCR的效率(Wang et al., 2011).与TAIL-PCR相比, hiTAIL-PCR的随机引物LAD1的5'端都带有一段AC1序列, 这使得第2轮及第3轮PCR操作大为简化(Liu and Chen, 2007).获得的PCR产物经凝胶电泳检测, 可通过比较SP2/AC1与SP3/AC1扩增产物的分子量大小判断其特异性(Liu and Chen, 2007). ...
1 1987
... 尽管hiTAIL-PCR在克隆侧翼序列方面显示了其高效性, 但应用到对T-DNA突变体侧翼序列的克隆时, 却经常出现一些事实上是质粒骨架片段而非染色体DNA片段的“特异性扩增结果”.通常情况下, 农杆菌仅在LB-RB之间产生可转移的单链T-DNA分子, 从而导致整合入宿主植物染色体中的仅为T-DNA区域(Tzfira et al., 2004).但是也有证据表明, T-DNA可连同质粒骨架作为一个分子整合至宿主植物的基因组中(Martineau et al., 1994; Kleinboelting et al., 2015).虽然质粒骨架随T-DNA分子一起整合至宿主植物染色体中的具体原因及机制仍不清楚, 但早期发现的“通读”现象(Stachel et al., 1987; Klein- boelting et al., 2015)可能是其原因之一.克隆这些转基因系插入位点的侧翼序列, 容易产生假阳性结果.其原因为只要随机引物LAD1能在质粒骨架上引发扩增, hiTAIL-PCR就能将部分骨架片段大量地扩增出来.然而, 由于在DNA片段测序之前, 我们无法判断这些“hiTAIL-PCR特异性片段”是来自质粒骨架还是突变体染色体DNA, 因此, 大量的后期鉴定工作将无法避免. ...
1 1988
... 有许多方法可用于克隆T-DNA插入位点的侧翼序列, 如质粒拯救(Nan and Walbot, 2009)、反向PCR (Triglia et al., 1988)、接头介导的PCR (Mueller and Wold, 1989; Riley et al., 1990)以及热不对称交错PCR (TAIL-PCR) (Liu et al., 1995)等.其中, TAIL- PCR由于操作简便及对突变体基因组DNA需求量少等特点, 成为大多数研究的首选方法.尤其是改进后的hiTAIL-PCR, 因具有更高效的克隆效率及操作简便而被广泛使用(Liu and Chen, 2007).该方法利用3条特异性引物(SP1、SP2和SP3)及多条随机引物(LAD1系列引物)来克隆未知侧翼序列.LAD1是简并引物, 能结合染色体DNA上的多个位点, 从而使SP1/LAD1初次扩增的侧翼序列并无较高的特异性; 在此基础上, SP2与SP3被用于第2轮及第3轮PCR扩增, 以进一步提高产物的特异性.简并引物在hiTAIL- PCR中扮演重要角色, 它的改进可进一步提高hiTAIL- PCR的效率(Wang et al., 2011).与TAIL-PCR相比, hiTAIL-PCR的随机引物LAD1的5'端都带有一段AC1序列, 这使得第2轮及第3轮PCR操作大为简化(Liu and Chen, 2007).获得的PCR产物经凝胶电泳检测, 可通过比较SP2/AC1与SP3/AC1扩增产物的分子量大小判断其特异性(Liu and Chen, 2007). ...
1 2004
... 尽管hiTAIL-PCR在克隆侧翼序列方面显示了其高效性, 但应用到对T-DNA突变体侧翼序列的克隆时, 却经常出现一些事实上是质粒骨架片段而非染色体DNA片段的“特异性扩增结果”.通常情况下, 农杆菌仅在LB-RB之间产生可转移的单链T-DNA分子, 从而导致整合入宿主植物染色体中的仅为T-DNA区域(Tzfira et al., 2004).但是也有证据表明, T-DNA可连同质粒骨架作为一个分子整合至宿主植物的基因组中(Martineau et al., 1994; Kleinboelting et al., 2015).虽然质粒骨架随T-DNA分子一起整合至宿主植物染色体中的具体原因及机制仍不清楚, 但早期发现的“通读”现象(Stachel et al., 1987; Klein- boelting et al., 2015)可能是其原因之一.克隆这些转基因系插入位点的侧翼序列, 容易产生假阳性结果.其原因为只要随机引物LAD1能在质粒骨架上引发扩增, hiTAIL-PCR就能将部分骨架片段大量地扩增出来.然而, 由于在DNA片段测序之前, 我们无法判断这些“hiTAIL-PCR特异性片段”是来自质粒骨架还是突变体染色体DNA, 因此, 大量的后期鉴定工作将无法避免. ...
1 2011
... 有许多方法可用于克隆T-DNA插入位点的侧翼序列, 如质粒拯救(Nan and Walbot, 2009)、反向PCR (Triglia et al., 1988)、接头介导的PCR (Mueller and Wold, 1989; Riley et al., 1990)以及热不对称交错PCR (TAIL-PCR) (Liu et al., 1995)等.其中, TAIL- PCR由于操作简便及对突变体基因组DNA需求量少等特点, 成为大多数研究的首选方法.尤其是改进后的hiTAIL-PCR, 因具有更高效的克隆效率及操作简便而被广泛使用(Liu and Chen, 2007).该方法利用3条特异性引物(SP1、SP2和SP3)及多条随机引物(LAD1系列引物)来克隆未知侧翼序列.LAD1是简并引物, 能结合染色体DNA上的多个位点, 从而使SP1/LAD1初次扩增的侧翼序列并无较高的特异性; 在此基础上, SP2与SP3被用于第2轮及第3轮PCR扩增, 以进一步提高产物的特异性.简并引物在hiTAIL- PCR中扮演重要角色, 它的改进可进一步提高hiTAIL- PCR的效率(Wang et al., 2011).与TAIL-PCR相比, hiTAIL-PCR的随机引物LAD1的5'端都带有一段AC1序列, 这使得第2轮及第3轮PCR操作大为简化(Liu and Chen, 2007).获得的PCR产物经凝胶电泳检测, 可通过比较SP2/AC1与SP3/AC1扩增产物的分子量大小判断其特异性(Liu and Chen, 2007). ...


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}