Abstract: Leaf length, width, area, and angle are important components of plant architecture but also affect the efficiency of photosynthesis in maize. In this study, 620 quantitative trait loci (QTL) were used to construct an integrated map related to maize leaf shape; 22 maize QTL (mQTL) for leaf length, 22 for leaf width, 12 for leaf area and 17 for leaf angle were estimated by meta-analysis. Further bioinformatics analysis identified 44 candidate genes closely related to leaf shape within the mQTL region, with some unintegrated QTL. However, only 13 candidate genes, including NAL7-like, YABBY6-like, and GRF2, were located in the mQTL region. Most of the candidate genes, such as the cloned genes KNOTTED1, AN3/GIF1, rgd1/lbl1 and mwp1 in maize and SRL2-like, HYL1-like, and CYCB2;4-like in rice and Arabidopsis thaliana homologous genes were projected onto the interval of unintegrated QTL. The regulation mechanism of 44 candidate genes is summarized and analyzed in the development of leaf length, width and thickness, by proximal-distal, central-marginal and adaxial-abaxial. Only a few known genes revealed part of the molecular mechanism of leaf deve- lopment in maize. Further research of the mQTL/QTL and related genes will create a global view of the genetic architecture of maize leaf shape, provide useful biological information for fine mapping QTL, and identify more candidate genes to clarify the molecular mechanism of leaf morphogenesis and provide a theoretical base for ideal plant-architecture improvement of maize marker-assisted breeding.
2.2 玉米叶形相关QTL的元分析对592个叶形QTLs进行元分析, 共获得73个mQTLs (图1), 其中叶长22个、叶宽22个、叶面积12个、叶夹角17个, 每个mQTL包含至少来自2个不同群体的2-11个初始QTLs。73个mQTLs置信区间变异范围为9.5-86.9 cM, 平均为24.5 cM, 其中40%的mQTL小于20 cM, 平均每个mQTL对应于原始图谱中的3.5个初始QTL。叶长、叶宽和叶夹角mQTL在第3染色体175.3-189.5 cM存在重叠; 叶长、叶宽和叶面积mQTL在第8染色体188.3-193.5 cM存在重叠; 第1染色体150.8-160.3 cM、第10染色体309.6-327.3 cM的叶宽mQTL与叶面积重叠; 第2染色体323.7-340.0 cM和480.7-504.4 cM、第8染色体407.5-415.7 cM的叶长mQTL与叶宽重叠; 第5染色体328.4-334.7 cM和第7染色体590.9-600.1 cM的叶面积mQTL与叶长重叠; 第1染色体173.3-191.1 cM、第5染色体244.8-258.1 cM和第9染色体397.4-412.0 cM的叶夹角mQTL与叶面积重叠; 第1染色体485.9-501.9 cM的叶宽mQTL与叶夹角重叠; 第3染色体86.9- 106.4 cM和222.1-240.3 cM、第7染色体318.3- 333.3 cM、第9染色体436.9-458.9 cM、第10染色体236.8-244.7 cM的叶长mQTL与叶夹角重叠。由此可知, 上述区间可能存在控制玉米叶形性状的“一因多效”QTL, 是解释叶形性状表型变异的重要位点, 对耐密株型的分子标记辅助选择育种具有重要作用。 图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-487/img_1.png<b>图1</b> 整合图谱中叶形相关mQTL在玉米各染色体上的分布<br/>Chr: 染色体。染色体上红色区域为mQTL重叠部分所在染色体的位置; 染色体左侧为整合后mQTL的名称及位置分布; 染色体右侧为整合图谱中的标记及遗传图距(cM)。<br/><b>Figure 1</b> Distribution of leaf shape mQTL on maize chromosomes in the integrated map<br/>Chr: Chromosome. The red region is the identified location of the overlaps of mQTL on the chromosome; The position distribution and the name for mQTL are listed in the left of chromosome; Marker name and genetic distance (cM) are listed in the right of chromosome. Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-487/img_1.png<b>图1</b> 整合图谱中叶形相关mQTL在玉米各染色体上的分布<br/>Chr: 染色体。染色体上红色区域为mQTL重叠部分所在染色体的位置; 染色体左侧为整合后mQTL的名称及位置分布; 染色体右侧为整合图谱中的标记及遗传图距(cM)。<br/><b>Figure 1</b> Distribution of leaf shape mQTL on maize chromosomes in the integrated map<br/>Chr: Chromosome. The red region is the identified location of the overlaps of mQTL on the chromosome; The position distribution and the name for mQTL are listed in the left of chromosome; Marker name and genetic distance (cM) are listed in the right of chromosome.
图1 整合图谱中叶形相关mQTL在玉米各染色体上的分布 Chr: 染色体。染色体上红色区域为mQTL重叠部分所在染色体的位置; 染色体左侧为整合后mQTL的名称及位置分布; 染色体右侧为整合图谱中的标记及遗传图距(cM)。 Figure 1 Distribution of leaf shape mQTL on maize chromosomes in the integrated map Chr: Chromosome. The red region is the identified location of the overlaps of mQTL on the chromosome; The position distribution and the name for mQTL are listed in the left of chromosome; Marker name and genetic distance (cM) are listed in the right of chromosome.
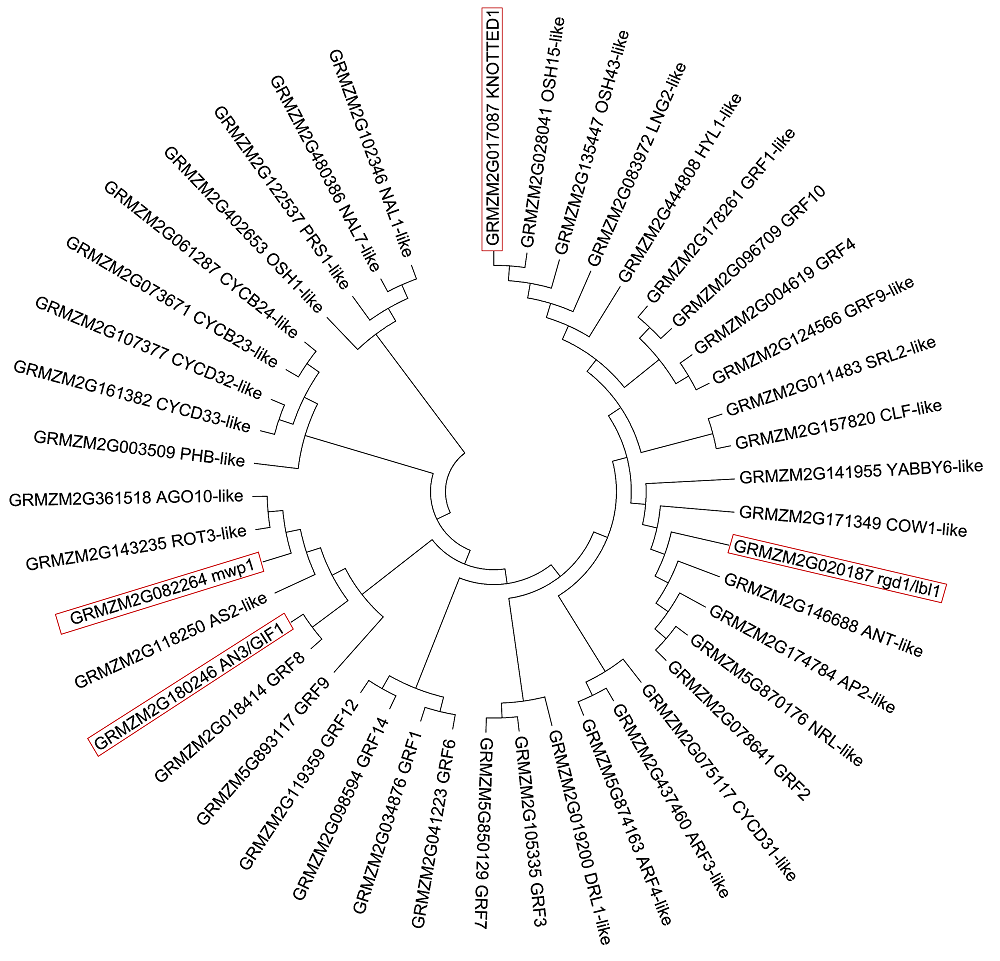
2.4 玉米叶形候选基因的聚类进化分析44个玉米叶形候选基因的系统进化关系(图2), 结果表明, 44个叶形候选基因按照亲缘关系在进化树中聚合为7大类。NAL7-like、NAL1-like、PRS1-like和OSH1-like聚为1类分支; CYCB2;4-like、CYCB2;3- like、CYCD3;2-like、CYCD3;3-like和PHB-like聚为2类分支; 功能已知(在叶片发育过程中发挥重要作用)的mwp1、AN3/GIF1与AGO10-like、ROT3-like、AS2- like、GRF8及GRF9聚为3类分支; GRF12、GRF14、GRF1和GRF6聚为4类分支; GRF7、GRF3和DRL1- like聚为5类分支; ARF3-like、ARF4-like和CYCD3; 1-like聚为6类分支; 7类分支中rgd1/lbl1和KNOT- TED1在玉米叶片发育过程中发挥相似功能, 与GRF2、NRL-like、AP2-like、COW1-like、YABBY6- like、CLF-like、SRL2-like、GRF9-like、GRF4、GRF10、GRF1-like、HYL1-like、LNG2-like、OSH43- like和OSH15-like共同聚为一大类。这些候选基因的系统进化分析为进一步克隆并研究其功能提供了参考信息。 图2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-487/img_2.png<b>图2</b> 玉米44个叶形候选基因的氨基酸序列聚类进化树<br/>方框标注的为玉米中已克隆且功能已知的叶形候选基因蛋白功能<br/><b>Figure 2</b> Phylogenetic trees of amino acid sequences of 44 candidate genes related to leaf shape of maize<br/>Function of candidate genes which have been known and cloned related to maize leaf shape are highlighted in the box Figure 2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-487/img_2.png<b>图2</b> 玉米44个叶形候选基因的氨基酸序列聚类进化树<br/>方框标注的为玉米中已克隆且功能已知的叶形候选基因蛋白功能<br/><b>Figure 2</b> Phylogenetic trees of amino acid sequences of 44 candidate genes related to leaf shape of maize<br/>Function of candidate genes which have been known and cloned related to maize leaf shape are highlighted in the box
图2 玉米44个叶形候选基因的氨基酸序列聚类进化树 方框标注的为玉米中已克隆且功能已知的叶形候选基因蛋白功能 Figure 2 Phylogenetic trees of amino acid sequences of 44 candidate genes related to leaf shape of maize Function of candidate genes which have been known and cloned related to maize leaf shape are highlighted in the box
2.5 讨论叶片是主要的侧翼器官, 其发育是一个复杂的过程, 主要由遗传因素决定(鞠培娜等, 2010)。与水稻和拟南芥相比, 玉米叶片具有较大的尺寸和线性形态, 更有利于研究叶片生长发育过程中的形态变化和分子特征, 因此玉米是研究叶形最为理想的模式作物。 2.5.1 玉米叶形相关mQTL分析 利用IBM2 2008 Neighbors图谱对玉米叶形相关QTL进行元分析, 寻找相同染色体区段或邻近位置的“一致性”QTL, 能够将不同环境条件、不同遗传背景、不同标记定位的叶型QTL信息清晰化, 缩小QTL置信区间, 提高定位的准确性。该图谱包含RFLP、SSR和RAPD等多种标记, 能够为MAS育种提供更多有效标记, 提高玉米MAS育种效率。 本研究所获得的44个叶形相关候选基因, 仅有13个基因落在mQTL内, 而且玉米中已知的叶形相关基因与mQTL位置并不重叠。其可能原因为: 一是玉米叶形的发育形成是一个复杂的过程, 由很多基因调控, 能预测到的候选基因只是其中一部分, 控制叶形相关性状的候选基因有待进一步挖掘; 二是玉米叶形相关QTL研究的局限性, 前人定位研究所用的亲本群体无法覆盖全部玉米种质, 而且大多是初级群体, 已有的定位结果并不能检测到所有叶形相关QTL, 导致两者并不完全重叠。 由于整合了多个群体的初始QTL, 我们不仅在第1、3、6、7、8和9染色体检测到与Guo等(2015)研究结果一致的mQTLW1-4、mQTLW3-1、mQTLW3-2、mQTLW6-2、mQTLW7-1、mQTLW8-2和mQTLW9, 还检测到49个新的mQTLs, 为叶形相关QTL的精细定位及候选基因克隆提供更有效的信息。其中, 叶长、叶宽、叶面积和叶夹角重叠的“一因多效” mQTL区段是控制叶形的重要区间, 是挖掘叶形候选基因的重点区域。 2.5.2 玉米叶形相关候选基因及作用路径分析 本研究通过对44个候选基因进行分析, 不仅为克隆玉米叶形性状mQTL奠定了基础, 还为研究候选基因在玉米叶片发育过程中叶原初生细胞的确定、生长轴向的形成、叶片功能区域的建立和随后的生长阶段发挥作用提供依据。 叶原初生细胞对于叶片形态建成具有重要影响。叶原初生细胞确定后, 叶片开始建立长、宽、厚3个轴向的发育, 这3个轴向包括基部(proximal)-末端(distal)、中央(central)-边缘(marginal)、远轴(adaxial)-近轴(abaxial), 分别受相关基因调控(Scanlon et al., 2000)。玉米KNOTTED1和AN3/GIF1分别通过调控顶端分生组织(shoot apical meristem, SAM)基部细胞分化和细胞伸长膨大影响叶片发育(Ramirez et al., 2009; Nelissen et al., 2015), KNOTTED1调控P0叶原基分化形成, 在基部-末端轴向形成过程中表达量逐渐降低, 甚至在发育成熟的叶片功能区并未表达, 而KNOTTED1在叶片发育过程的末端表达时, 叶片中脉发育提前终止, 叶片分叉且卷曲(Ramirez et al., 2009)。叶片发育伸展过程中, rgd1/lbl1直接或间接下调knotted1的表达, 两者相互协调建立基部-末端轴向, 并促进叶原细胞分裂, 加快叶片侧向生长(Tim- mermans et al., 1998; Ramirez et al., 2009)。此外, 水稻OsH1、OsH15和OsH43属于KNOX家族, 突变后表现与knotted1、knox5和knox1相似的叶部异常表型(Matsuoka et al., 1993; Sentoku et al., 2000; Nagasaki et al., 2001)。由此可知, knotted1、AN3/GIF1、rgd1/lbl1和OsH-like对基部-末端轴向的形成具有重要影响。 中部细胞的横向发育是影响叶片宽度的关键因素, 而玉米中央-边缘轴向的研究最为深入, ns1和ns2在该轴向的建立中发挥重要作用。研究发现ns1和ns2主要在玉米SAM的叶建成组织细胞中表达, 由于其功能缺失影响叶原基边缘区的发育, 导致成熟叶片及叶鞘变窄, ns1/ns2双突变体无叶片边缘区(Scanlon et al., 2000; Nardmann et al., 2004)。PRS-like与ns1同源, 而PRS-like与AtPRS1具有同源结构域。拟南芥Prs功能缺失导致叶片及花萼外缘横向生长被抑制, 但叶片和花萼远-近轴轴向并未发生变化(Matsumoto and Okada, 2001; Nakata et al., 2012), AtPRS1是侧生器官外缘分生组织的激活因子, 与NS1和NS2有相似的功能。因此, PRS-like可能参与中央-边缘轴向的发育。 远轴-近轴极性分化是叶片生长发育必需的, 而玉米MWP1、水稻SRL2和SLL1均作用于叶片远轴端的发育, 对叶片边缘区建立具有重要作用。OsSRL2通过影响YABBY家族基因的转录活性, 调控叶片远轴端的发育(Liu et al., 2016)。srl2和sll1表现相似的窄叶、卷曲突变表型, 但叶片的卷曲程度不同, srl2和sll1突变后均导致叶片远轴端厚壁组织细胞缺陷、叶片变窄。srl2突变体中YAB2、YAB3、YAB4、YAB5和YAB7表达量显著降低, 但SLL1、NAL1、ND1和RL14的表达并未发生变化。而前人的研究表明, SLL1参与细胞的程序性死亡, 调节叶肉细胞和厚壁组织细胞的分化及形成(Zhang et al., 2009; Liu et al., 2016)。上述结果表明, SRL2和SLL1通过不同作用路径调控叶片形态建成, 而mwp1与OsSLL1高度同源且具有KANADI家族功能域(Candela et al., 2008)。因此, SRL2-like和mwp1可能通过不同路径作用于玉米叶片远-近轴向发育。 在拟南芥中, ARF3/ARF4和PHB分别调控远轴和近轴组织的形成。生长素响应因子基因ARF3/ARF4是tasiR-ARF的靶基因, 而HD-ZIPlll类基因PHB (PHV和REV)包含miR165/166的结合位点, 上述2种小RNAs分别存在于叶原基的近轴和远轴部位, 通过限制其靶基因的表达影响叶片远轴端和近轴端细胞的分化(Pekker et al., 2005; Hunter et al., 2006; Kelley et al., 2012; Dotto et al., 2014)。ARF3/ARF4受tasiR-ARF调控的同时, 与KAN1 (KANAD1)家族基因相互协调作用于远轴组织的形成。DRL能够上调SAM中KNOX (kn1)家族基因的表达, 保持其细胞分裂活性, 而在叶片中抑制YABBY和KAN1类基因, 可促进近轴组织的分化发育(Jun et al., 2015)。AS2是HD-ZIP类转录因子, 促进叶片近轴细胞的分化, 远轴端AS2的启动子被AtKAN1结合并抑制其表达(Iwakawa et al., 2007)。另外, HYL1通过正向调控miR165/166、miR319及miR160维持远轴-近轴的正常发育调节叶片形态。ROT3和LNG2均作用于纵向极性细胞伸长膨大, 从而影响叶片变长, 但其作用机理不同(Kim et al., 1998; Mallory et al., 2004)。CLF功能的缺失导致H3K27me3的活性降低, 引起叶片卷曲变窄(Schatlowski et al., 2010)。ANT功能缺失影响叶片等侧生器官变小; 过量表达时叶细胞数目增加, 叶片变大(Mizukami and Fischer, 2000)。CYCD家族基因在叶片发育过程中调控叶细胞数目, 大多能够与ANT相互作用促进叶原基细胞分裂, 影响叶片大小(Dewitte et al., 2007)。AP2是miR172的靶基因, 同时与ARF3相互作用调控分生组织的分化, 影响叶片的形态(Würschum et al., 2006; Mlotshwa et al., 2006)。AGO10通过REV下调ARF2的表达, 负向调控生长素响应信号, 促进顶端分生组织干细胞向叶腋分生组织的分化(Zhu et al., 2011; Roodbarkelari et al., 2015)。由此可知, 上述基因的同源基因可能影响玉米叶片的发育。 植物激素的合成、代谢与叶片的发育密切相关。研究人员对水稻Nal1、Nal7和COW1进行研究时, 发现生长素的极性运输与合成异常, 导致水稻叶形发生变化。Nal1缺失突变导致生长素极性运输蛋白基因PIN1的表达显著降低, 进而引起生长素极性运输紊乱和细胞分裂发育异常, 从而影响维管束的正常分布及叶片发育, 最终导致叶片变窄(Qi et al., 2008; Kubo et al., 2017)。Nal7编码1个生长素合成的关键蛋白, 影响细胞分裂和伸长。nal7叶片窄且SAM中生长素含量明显降低, 过量表达Nal7后突变表型得到恢复(Fujino et al., 2008)。水稻YUCCA型基因COW1缺失突变引起生长素合成异常, 表现与OsYUCCA1反义植株相似的表型, 叶片卷曲且变窄(Woo et al., 2007)。由此可知, 玉米NAL1-like、NAL7-like和COW1-like可能参与生长素的合成及运输, 影响叶片发育。 不仅生长素相关基因参与调控叶宽, 细胞壁合成相关的类纤维素合成酶基因也影响叶形。除了nrl1外(Hu et al., 2010), 水稻sle1、nd1和cd1突变体均是由于编码类纤维素合成酶D4基因的突变导致叶片变窄, 而这些基因主要在分化形成不同器官的顶端分生组织中表达(Luan et al., 2011; Yoshikawa et al., 2013; Ding et al., 2015)。由此推测, 类纤维素合成酶基因NRL-like可能在玉米叶片形成过程中发挥作用。 研究表明, GRF家族是一类转录激活因子, 调控不同组织器官的发育, 影响其形态建成。玉米叶片中GRFs受miR396a调控(Candaele et al., 2014), 在不同时期和不同组织中GRFs表达量不同(Facette et al., 2013), GRF1、GRF6、GRF7、GRF12、GRF15和GRF17在分裂组织细胞中表达量很高, 但是GRF4 和GRF10在分裂细胞和伸长膨大细胞中均大量表达。GRF1过量表达时, 分裂组织细胞的分裂增殖加快, 导致叶片变大; 而过量表达GRF10时, 细胞膨大生长的速度快于分裂速度, 最终使叶片变小、变窄(Wu et al., 2014)。拟南芥grf1/grf2/grf3三突变体植株低矮、叶片窄小, 分别过量表达AtGRF1和AtGRF2, 叶片变宽、增大, 可部分恢复突变表型(Kim et al., 2003); 过量表达AtGRF5时, 细胞分裂增殖加快, 叶细胞数目增多, 叶片变大、植株增高(Horiguchi et al., 2006)。在细胞分裂和膨大生长的动态协作过程中, GRF家族基因分别通过调控细胞的分裂和膨大来调控叶片的形态大小。 GRF蛋白具有DNA结合活性, 能够结合特定的靶基因形成染色质复合物, GRF家族成员在调控不同器官发育的过程中, 通过调节GRF和GIF (GRF互作因子)的动态平衡来影响叶片的发育及不同器官的形成(Zhang et al., 2008; Facette et al., 2013; Candaele et al., 2014; Wu et al., 2014)。拟南芥AN3能够聚合SWI/SNF形成SWI/SNF-AN3 (GIF1)复合物, 进一步结合GRF3、GRF5和GRF6的启动子, 调控细胞增殖向细胞分化的过渡(Vercruyssen et al., 2014)。过量表达SWI3C时, 拟南芥叶片变大; AN3功能缺失时, 叶片细胞减少, 拟南芥叶片变窄、变小。同时, 拟南芥GRF1、GRF2、GRF3、GRF4、GRF7、GRF8和GRF9受miR396调控, miR396能够对这些生长调节因子进行转录后剪切, 从而抑制GRF的转录(Liang et al., 2014)。过量表达miR396, 拟南芥叶片变小; 过量表达miR396结合位点发生突变的GRF2、GRF7和GRF9时, 拟南芥叶片显著变大(Debernardi et al., 2014)。由此可知, miR396-GRF-GIF作用模式调控叶片发育。因此玉米中这些GRF及GRF-like基因可能与不同时期叶片形成密切相关。 上述候选基因通过不同作用路径调控叶片发育, 与候选基因的聚类进化分析相吻合, 进一步支持了以上候选基因预测的准确性, 为相应mQTL/QTL的精细定位奠定了基础。
LeeYK, KimGT, KimIJ, ParkJ, KwakSS, ChoiG, ChungWI (2006). LONGIFOLIA1 and LONGIFOLIA2, two homologous genes, regulate longitudinal cell elongation in Arabidopsis. Development 133, 4305-4314.
[43]
LiangG, HeH, LiY, WangF, YuDQ (2014). Molecular mechanism of microRNA396 mediating pistil development in Arabidopsis.Plant Physiol 164, 249-258. [本文引用: 1]
[44]
LiuXF, LiM, LiuK, TangD, SunMF, LiYF, ShenY, DuGJ, ChengZK (2016). Semi-Rolled Leaf 2 modulates rice leaf rolling by regulating abaxial side cell differentiation. J Exp Bot 67, 2139-2150. [本文引用: 2]
[45]
LiuZY, JiaLG, WangH, HeYK (2011). HYL1 regulates the balance between adaxial and abaxial identity for leaf flattening via miRNA-mediated pathways.J Exp Bot 62, 4367-6381.
[46]
LuanWJ, LiuYQ, ZhangFX, SongYL, WangZY, PengYK, SunZX (2011). OsCD1 encodes a putative member of the cellulose synthase-like D sub-family and is essential for rice plant architecture and growth. Plant Biotechnol J 9, 513-524. [本文引用: 1]
[47]
MalloryAC, ReinhartBJ, Jones-RhoadesMW, TangGL, ZamorePD, BartonMK, BartelDP (2004). MicroRNA control of PHABULOSA in leaf development: importance of pairing to the microRNA 5′ region.EMBO J 23, 3356-3364. [本文引用: 1]
[48]
MatsumotoN, OkadaK (2001). A homeobox gene, PRES- SED FLOWER, regulates lateral axis-dependent deve- lopment of Arabidopsis flowers. Genes Dev 15, 3355-3364. [本文引用: 1]
[49]
MatsuokaM, IchikawaH, SaitoA, TadaY, FujimuraT, Kano-MurakamiY (1993). Expression of a rice homeobox gene causes altered morphology of transgenic plants.Plant Cell 5, 1039-1048. [本文引用: 1]
[50]
MengesM, de JagerSM, GruissemW, MurrayJAH (2005). Global analysis of the core cell cycle regulators of Arabidopsis identifies novel genes, reveals multiple and highly specific profiles of expression and provides a coherent model for plant cell cycle control.Plant J 41, 546-566.
[51]
MickelsonSM, StuberCS, SeniorL, KaepplerSM (2002). Quantitative trait loci controlling leaf and tassel traits in a B73 × Mo17 population of maize.Crop Sci 42, 1902-1909.
[52]
MizukamiY, FischerRL (2000). Plant organ size control: AINTEGUMENTA regulates growth and cell numbers during organogenesis. Proc Natl Acad Sci USA 97, 942-947. [本文引用: 1]
[53]
MlotshwaS, YangZY, KimYJ, ChenXM (2006). Floral patterning defects induced by Arabidopsis APETALA2 and microRNA172 expression in Nicotiana benthamiana.Plant Mol Biol 61, 781-793. [本文引用: 1]
[54]
NagasakiH, SakamotoT, SatoY, MatsuokaM (2001). Functional analysis of the conserved domains of a rice KNOX homeodomain protein, OSH15.Plant Cell 13, 2085-2098. [本文引用: 1]
[55]
NakataM, MatsumotoN, TsugekiR, RikirschE, LauxT, OkadaK (2012). Roles of the middle domain-specific WUSCHEL-RELATED HOMEOBOX genes in early development of leaves in Arabidopsis. Plant Cell 24, 519-535. [本文引用: 1]
[56]
NardmannJ, JiJB, WerrW, ScanlonMJ (2004). The maize duplicate genes narrow sheath 1 and narrow sheath 2 encode a conserved homeobox gene function in a lateral domain of shoot apical meristems. Development 131, 2827-2839. [本文引用: 1]
[57]
NelissenH, EeckhoutD, DemuynckK, PersiauG, WaltonA, Van BelM, VervoortM, CandaeleJ, de BlockJ, Aes- aertS, Van LijsebettensM, GoormachtigS, VandepoeleK, Van LeeneJ, MuszynskiM, GevaertK, InzéD, De JaegerG (2015). Dynamic changes in ANGUSTIFOLIA3 complex composition reveal a growth regulatory mechanism in the maize leaf.Plant Cell 27, 1605-1619. [本文引用: 1]
[58]
PekkerI, AlvarezJP, EshedY (2005). Auxin response factors mediate Arabidopsis organ asymmetry via modulation of KANADI activity.Plant Cell 17, 2899-2910. [本文引用: 1]
[59]
QiJ, QianQ, BuQY, LiSY, ChenQ, SunJQ, LiangWX, ZhouYH, ChuCC, LiXG, RenFG, PalmeK, ZhaoBR, ChenJF, ChenMS, LiCY (2008). Mutation of the rice Narrow leaf1 gene, which encodes a novel protein, affects vein patterning and polar auxin transport. Plant Physiol 147, 1947-1959. [本文引用: 1]
[60]
RamirezJ, BolducN, LischD, HakeS (2009). Distal expression of knotted1 in maize leaves leads to reestablishment of proximal/distal patterning and leaf dissection.Plant Physiol 151, 1878-1888. [本文引用: 3]
[61]
ReymondM, MullerB, TardieuF (2004). Dealing with the genotype×environment interaction via a modelling approach: a comparison of QTLs of maize leaf length or width with QTLs of model parameters.J Exp Bot 55, 2461-2472. [本文引用: 1]
ScanlonMJ, ChenKD, McKnightCC (2000). The narrow sheath duplicate genes: sectors of dual aneuploidy reveal ancestrally conserved gene functions during maize leaf development. Genetics 155, 1379-1389. [本文引用: 2]
[64]
SchatlowskiN, StahlY, HohenstattML, GoodrichJ, SchubertD (2010). The CURLY LEAF interacting protein BLISTER controls expression of polycomb-group target genes and cellular differentiation of Arabidopsis thaliana. Plant Cell 22, 2291-2305. [本文引用: 1]
[65]
SentokuN, SatoY, MatsuokaM (2000). Overexpression of rice OSH genes induces ectopic shoots on leaf sheaths of transgenic rice plants. Dev Biol 220, 358-364. [本文引用: 1]
[66]
TianF, BradburyPJ, BrownPJ, HungH, SunQ, Flint-GarciaS, RochefordTR, McMullenMD, HollandJB, BucklerES (2011). Genome-wide association study of leaf architecture in the maize nested association mapping population.Nat Genet 43, 159-162.
[67]
TimmermansMC, SchultesNP, JankovskyJP, NelsonT (1998). Leaf bladeless 1 is required for dorsoventrality of lateral organs in maize. Development 125, 2813-2823. [本文引用: 1]
[68]
ToribaT, HaradaK, TakamuraA, NakamuraH, IchikawaH, SuzakiT, HiranoHY (2007). Molecular characterization the YABBY gene family in Oryza sativa and expression analysis of OsYABBY1. Mol Genet Genom 277, 457-468.
[69]
VercruyssenL, VerkestA, GonzalezN, HeyndrickxKS, EeckhoutD, HanSK, JéguT, ArchackiR, Van LeeneJ, AndriankajaM, De BodtS, AbeelT, CoppensF, DhondtS, De MildeL, VermeerschM, MaleuxK, GevaertK, JerzmanowskiA, BenhamedM, WagnerD, VandepoeleK, De JaegerG, InzéD (2014). ANGUSTIFOLIA3 binds to SWI/SNF chromatin remodeling complexes to regulate transcription during Arabidopsis leaf development.Plant Cell 26, 210-229. [本文引用: 1]
[70]
WassomJJ (2013). Quantitative trait loci for leaf angle, leaf width, leaf length, and plant height in a maize (Zea mays L.) B73 × Mo17 population. Maydica 58, 318-321.
[71]
WeiXM, WangXB, GuoSL, ZhouJL, ShiY, WangHT, DouDD, SongXH, LiGH, KuLX, ChenYH (2016). Epistatic and QTL × environment interaction effects on leaf area-associated traits in maize.Plant Breed 135, 671-676.
[72]
WooYM, ParkHJ, Su’udiM, YangJI, ParkJJ, BackK, ParkYM, AnG (2007). Constitutively wilted 1, a member of the rice YUCCA gene family, is required for maintaining water homeostasis and an appropriate root to shoot ratio. Plant Mol Biol 65, 125-136. [本文引用: 1]
[73]
WuL, ZhangDF, XueM, QianJJ, HeY, WangSC (2014). Overexpression of the maize GRF10, an endogenous truncated growth-regulating factor protein, leads to reduction in leaf size and plant height.J Integr Plant Biol 56, 1053-1063. [本文引用: 2]
[74]
WürschumT, Groß-HardtR, LauxT (2006). APETALA2 regulates the stem cell niche in the Arabidopsis shoot meristem. Plant Cell 18, 295-307. [本文引用: 1]
[75]
YoshikawaT, EiguchiM, HibaraKI, ItoJI, NagatoY (2013). Rice SLENDER LEAF 1 gene encodes cellulose synthase-like D4 and is specifically expressed in M-phase cells to regulate cell proliferation. J Exp Bot 64, 2049-2061. [本文引用: 1]
AgramaHAS, ZakariaAG, SaidFB, TuinstraM (1999). Identification of quantitative trait loci for nitrogen use efficiency in maize.Mol Breed 5, 187-195. [本文引用: 1]
[17]
CaiHG, ChuQ, YuanLX, LiuJC, ChenXH, ChenFJ, MiGH, ZhangFS (2012). Identification of quantitative trait loci for leaf area and chlorophyll content in maize ( Zea mays) under low nitrogen and low phosphorus supply. Mol Breed 30, 251-266.
CandelaH, JohnstonR, GerholdA, FosterT, HakeS (2008). The Milkweed pod1 gene encodes a KANADI protein that is required for abaxial/adaxial patterning in maize leaves. Plant Cell 20, 2073-2087. [本文引用: 1]
[20]
DarvasiA, SollerM (1997). A simple method to calculate resolving power and confidence interval of QTL map location.Behav Genet 27, 125-132. [本文引用: 2]
[21]
DebernardiJM, MecchiaMA, VercruyssenL, SmaczniakC, KaufmannK, InzeD, RodriguezRE, PalatnikJF (2014). Post-transcriptional control of GRF transcription factors by microRNA miR396 and GIF co-activator affects leaf size and longevity.Plant J 79, 413-426. [本文引用: 1]
[22]
DewitteW, ScofieldS, AlcasabasAA, MaughanSC, MengesM, BraunN, CollinsC, NieuwlandJ, PrinsenE, SundaresanV, MurrayJAH (2007). Arabidopsis CYCD3 D-type cyclins link cell proliferation and endocycles and are rate-limiting for cytokinin responses.Proc Natl Acad Sci USA 104, 14537-14542. [本文引用: 1]
[23]
DingZQ, LinZF, LiQ, WuH, XiangCY, WangJF (2015). DNL1, encodes cellulose synthase-like D4, is a major QTL for plant height and leaf width in rice(Oryza sativa L.). Biochem Biophys Res Commun 457, 133-140. [本文引用: 1]
[24]
DottoMC, PetschKA, AukermanMJ, BeattyM, HammellM, TimmermansMC (2014). Genome-wide analysis of leaf bladeless1-regulated and phased small RNAs underscores the importance of the TAS3 ta-siRNA pathway to maize development. PLoS Genet 10, e1004826. [本文引用: 1]
[25]
EloyNB, de Freitas LimaM, Van DammeD, VanhaerenH, GonzalezN, de MildeL, HemerlyAS, BeemsterGTS, InzéD, FerreiraPCG (2011). The APC/C subunit 10 plays an essential role in cell proliferation during leaf development. Plant J 68, 351-363.
[26]
FacetteMR, ShenZX, BjörnsdóttirFR, BriggsSP, SmithLG (2013). Parallel proteomic and phosphoproteomic ana- lyses of successive stages of maize leaf development.Plant Cell 25, 2798-2812. [本文引用: 2]
[27]
FujinoK, MatsudaY, OzawaK, NishimuraT, KoshibaT, FraaijeMW, SekiguchiH (2008). NARROW LEAF 7 controls leaf shape mediated by auxin in rice. Mol Genet Genom 279, 499-507. [本文引用: 1]
GuoSL, KuLX, QiJS, TianZQ, HanT, ZhangLK, SuHH, RenZZ, ChenYH (2015). Genetic analysis and major quantitative trait locus mapping of leaf widths at different positions in multiple populations.PLoS One 10, e0119095.
[30]
HoriguchiG, FerjaniA, FujikuraU, TsukayaH (2006). Coordination of cell proliferation and cell expansion in the control of leaf size in Arabidopsis thaliana. J Plant Res 119, 37-42. [本文引用: 1]
[31]
HuJ, ZhuL, ZengDL, GaoZY, GuoLB, FangYX, ZhangGH, DongGJ, YanMX, LiuJ, QianQ (2010). Identification and characterization of NARROW AND ROLLED LEAF 1, a novel gene regulating leaf morphology and plant architecture in rice. Plant Mol Biol 73, 283-292. [本文引用: 1]
[32]
HunterC, WillmannMR, WuG, YoshikawaM, de la Luz Gutiérrez-Nava M, PoethigSR (2006). Trans-acting siRNA-mediated repression of ETTIN and ARF4 regulates heteroblasty in Arabidopsis.Development 133, 2973-2981. [本文引用: 1]
[33]
IwakawaH, IwasakiM, KojimaS, UenoY, SomaT, TanakaH, SemiartiE, MachidaY, MachidaC (2007). Expression of the ASYMMETRIC LEAVES 2 gene in the adaxial domain of Arabidopsis leaves represses cell proliferation in this domain and is critical for the development of properly expanded leaves. Plant J 51, 173-184. [本文引用: 1]
[34]
JunSE, ChoKH, HwangJY, Abdel-FattahW, HammermeisterA, SchaffrathR, BowmanJL, KimGT (2015). Comparative analysis of the conserved functions of Arabidopsis DRL1 and yeast KTI12.Mol Cells 38, 243-250. [本文引用: 1]
[35]
KelleyDR, ArreolaA, GallagherTL, GasserCS (2012). ETTIN (ARF3) physically interacts with KANADI proteins to form a functional complex essential for integument deve- lopment and polarity determination in Arabidopsis.Deve- lopment 139, 1105-1109. [本文引用: 1]
[36]
KimGT, TsukayaH, UchimiyaH (1998). The ROTUNDIFOLIA3 gene of Arabidopsis thaliana encodes a new member of the cytochrome P450 family that is required for the regulated polar elongation of leaf cells. Genes Dev 12, 2381-2391. [本文引用: 1]
[37]
KimJH, ChoiD, KendeH (2003). The AtGRF family of putative transcription factors is involved in leaf and cotyledon growth in Arabidopsis.Plant J 36, 94-104. [本文引用: 1]
[38]
KuLX, ZhangJ, GuoSL, LiuHY, ZhaoRF, ChenYH (2012a). Integrated multiple population analysis of leaf architecture traits in maize (Zea mays L.). J Exp Bot 63, 261-274.
[39]
KuLX, ZhangJ, ZhangJC, GuoSL, LiuHY, ZhaoRF, YanQX, ChenYH (2012b). Genetic dissection of leaf area by jointing two F2:3 populations in maize (Zea mays L.). Plant Breed 131, 591-599.
[40]
KuLX, ZhaoWM, ZhangJ, WuLC, WangCL, WangPA, ZhangWQ, ChenYH (2010). Quantitative trait loci mapping of leaf angle and leaf orientation value in maize (Zea mays L.). Theor Appl Genet 121, 951-959.
KuboFC, YasuiY, KumamaruT, SatoY, HiranoHY (2017). Genetic analysis of rice mutants responsible for narrow leaf phenotype and reduced vein number.Genes Genet Syst 91, 235-240. [本文引用: 1]
0 2006
1 2014
... GRF蛋白具有DNA结合活性, 能够结合特定的靶基因形成染色质复合物, GRF家族成员在调控不同器官发育的过程中, 通过调节GRF和GIF (GRF互作因子)的动态平衡来影响叶片的发育及不同器官的形成(Zhang et al., 2008; Facette et al., 2013; Candaele et al., 2014; Wu et al., 2014).拟南芥AN3能够聚合SWI/SNF形成SWI/SNF-AN3 (GIF1)复合物, 进一步结合GRF3、GRF5和GRF6的启动子, 调控细胞增殖向细胞分化的过渡(Vercruyssen et al., 2014).过量表达SWI3C时, 拟南芥叶片变大; AN3功能缺失时, 叶片细胞减少, 拟南芥叶片变窄、变小.同时, 拟南芥GRF1、GRF2、GRF3、GRF4、GRF7、GRF8和GRF9受miR396调控, miR396能够对这些生长调节因子进行转录后剪切, 从而抑制GRF的转录(Liang et al., 2014).过量表达miR396, 拟南芥叶片变小; 过量表达miR396结合位点发生突变的GRF2、GRF7和GRF9时, 拟南芥叶片显著变大(Debernardi et al., 2014).由此可知, miR396-GRF-GIF作用模式调控叶片发育.因此玉米中这些GRF及GRF-like基因可能与不同时期叶片形成密切相关. ...
2 2016
... 远轴-近轴极性分化是叶片生长发育必需的, 而玉米MWP1、水稻SRL2和SLL1均作用于叶片远轴端的发育, 对叶片边缘区建立具有重要作用.OsSRL2通过影响YABBY家族基因的转录活性, 调控叶片远轴端的发育(Liu et al., 2016).srl2和sll1表现相似的窄叶、卷曲突变表型, 但叶片的卷曲程度不同, srl2和sll1突变后均导致叶片远轴端厚壁组织细胞缺陷、叶片变窄.srl2突变体中YAB2、YAB3、YAB4、YAB5和YAB7表达量显著降低, 但SLL1、NAL1、ND1和RL14的表达并未发生变化.而前人的研究表明, SLL1参与细胞的程序性死亡, 调节叶肉细胞和厚壁组织细胞的分化及形成(Zhang et al., 2009; Liu et al., 2016).上述结果表明, SRL2和SLL1通过不同作用路径调控叶片形态建成, 而mwp1与OsSLL1高度同源且具有KANADI家族功能域(Candela et al., 2008).因此, SRL2-like和mwp1可能通过不同路径作用于玉米叶片远-近轴向发育. ... ... ; Liu et al., 2016).上述结果表明, SRL2和SLL1通过不同作用路径调控叶片形态建成, 而mwp1与OsSLL1高度同源且具有KANADI家族功能域(Candela et al., 2008).因此, SRL2-like和mwp1可能通过不同路径作用于玉米叶片远-近轴向发育. ...
0 2011
1 2011
... 不仅生长素相关基因参与调控叶宽, 细胞壁合成相关的类纤维素合成酶基因也影响叶形.除了nrl1外(Hu et al., 2010), 水稻sle1、nd1和cd1突变体均是由于编码类纤维素合成酶D4基因的突变导致叶片变窄, 而这些基因主要在分化形成不同器官的顶端分生组织中表达(Luan et al., 2011; Yoshikawa et al., 2013; Ding et al., 2015).由此推测, 类纤维素合成酶基因NRL-like可能在玉米叶片形成过程中发挥作用. ...
1 2004
... 在拟南芥中, ARF3/ARF4和PHB分别调控远轴和近轴组织的形成.生长素响应因子基因ARF3/ARF4是tasiR-ARF的靶基因, 而HD-ZIPlll类基因PHB (PHV和REV)包含miR165/166的结合位点, 上述2种小RNAs分别存在于叶原基的近轴和远轴部位, 通过限制其靶基因的表达影响叶片远轴端和近轴端细胞的分化(Pekker et al., 2005; Hunter et al., 2006; Kelley et al., 2012; Dotto et al., 2014).ARF3/ARF4受tasiR-ARF调控的同时, 与KAN1 (KANAD1)家族基因相互协调作用于远轴组织的形成.DRL能够上调SAM中KNOX (kn1)家族基因的表达, 保持其细胞分裂活性, 而在叶片中抑制YABBY和KAN1类基因, 可促进近轴组织的分化发育(Jun et al., 2015).AS2是HD-ZIP类转录因子, 促进叶片近轴细胞的分化, 远轴端AS2的启动子被AtKAN1结合并抑制其表达(Iwakawa et al., 2007).另外, HYL1通过正向调控miR165/166、miR319及miR160维持远轴-近轴的正常发育调节叶片形态.ROT3和LNG2均作用于纵向极性细胞伸长膨大, 从而影响叶片变长, 但其作用机理不同(Kim et al., 1998; Mallory et al., 2004).CLF功能的缺失导致H3K27me3的活性降低, 引起叶片卷曲变窄(Schatlowski et al., 2010).ANT功能缺失影响叶片等侧生器官变小; 过量表达时叶细胞数目增加, 叶片变大(Mizukami and Fischer, 2000).CYCD家族基因在叶片发育过程中调控叶细胞数目, 大多能够与ANT相互作用促进叶原基细胞分裂, 影响叶片大小(Dewitte et al., 2007).AP2是miR172的靶基因, 同时与ARF3相互作用调控分生组织的分化, 影响叶片的形态(Würschum et al., 2006; Mlotshwa et al., 2006).AGO10通过REV下调ARF2的表达, 负向调控生长素响应信号, 促进顶端分生组织干细胞向叶腋分生组织的分化(Zhu et al., 2011; Roodbarkelari et al., 2015).由此可知, 上述基因的同源基因可能影响玉米叶片的发育. ...
1 2001
... 中部细胞的横向发育是影响叶片宽度的关键因素, 而玉米中央-边缘轴向的研究最为深入, ns1和ns2在该轴向的建立中发挥重要作用.研究发现ns1和ns2主要在玉米SAM的叶建成组织细胞中表达, 由于其功能缺失影响叶原基边缘区的发育, 导致成熟叶片及叶鞘变窄, ns1/ns2双突变体无叶片边缘区(Scanlon et al., 2000; Nardmann et al., 2004).PRS-like与ns1同源, 而PRS-like与AtPRS1具有同源结构域.拟南芥Prs功能缺失导致叶片及花萼外缘横向生长被抑制, 但叶片和花萼远-近轴轴向并未发生变化(Matsumoto and Okada, 2001; Nakata et al., 2012), AtPRS1是侧生器官外缘分生组织的激活因子, 与NS1和NS2有相似的功能.因此, PRS-like可能参与中央-边缘轴向的发育. ...
1 1993
... 叶原初生细胞对于叶片形态建成具有重要影响.叶原初生细胞确定后, 叶片开始建立长、宽、厚3个轴向的发育, 这3个轴向包括基部(proximal)-末端(distal)、中央(central)-边缘(marginal)、远轴(adaxial)-近轴(abaxial), 分别受相关基因调控(Scanlon et al., 2000).玉米KNOTTED1和AN3/GIF1分别通过调控顶端分生组织(shoot apical meristem, SAM)基部细胞分化和细胞伸长膨大影响叶片发育(Ramirez et al., 2009; Nelissen et al., 2015), KNOTTED1调控P0叶原基分化形成, 在基部-末端轴向形成过程中表达量逐渐降低, 甚至在发育成熟的叶片功能区并未表达, 而KNOTTED1在叶片发育过程的末端表达时, 叶片中脉发育提前终止, 叶片分叉且卷曲(Ramirez et al., 2009).叶片发育伸展过程中, rgd1/lbl1直接或间接下调knotted1的表达, 两者相互协调建立基部-末端轴向, 并促进叶原细胞分裂, 加快叶片侧向生长(Tim- mermans et al., 1998; Ramirez et al., 2009).此外, 水稻OsH1、OsH15和OsH43属于KNOX家族, 突变后表现与knotted1、knox5和knox1相似的叶部异常表型(Matsuoka et al., 1993; Sentoku et al., 2000; Nagasaki et al., 2001).由此可知, knotted1、AN3/GIF1、rgd1/lbl1和OsH-like对基部-末端轴向的形成具有重要影响. ...
0 2005
0 2002
1 2000
... 在拟南芥中, ARF3/ARF4和PHB分别调控远轴和近轴组织的形成.生长素响应因子基因ARF3/ARF4是tasiR-ARF的靶基因, 而HD-ZIPlll类基因PHB (PHV和REV)包含miR165/166的结合位点, 上述2种小RNAs分别存在于叶原基的近轴和远轴部位, 通过限制其靶基因的表达影响叶片远轴端和近轴端细胞的分化(Pekker et al., 2005; Hunter et al., 2006; Kelley et al., 2012; Dotto et al., 2014).ARF3/ARF4受tasiR-ARF调控的同时, 与KAN1 (KANAD1)家族基因相互协调作用于远轴组织的形成.DRL能够上调SAM中KNOX (kn1)家族基因的表达, 保持其细胞分裂活性, 而在叶片中抑制YABBY和KAN1类基因, 可促进近轴组织的分化发育(Jun et al., 2015).AS2是HD-ZIP类转录因子, 促进叶片近轴细胞的分化, 远轴端AS2的启动子被AtKAN1结合并抑制其表达(Iwakawa et al., 2007).另外, HYL1通过正向调控miR165/166、miR319及miR160维持远轴-近轴的正常发育调节叶片形态.ROT3和LNG2均作用于纵向极性细胞伸长膨大, 从而影响叶片变长, 但其作用机理不同(Kim et al., 1998; Mallory et al., 2004).CLF功能的缺失导致H3K27me3的活性降低, 引起叶片卷曲变窄(Schatlowski et al., 2010).ANT功能缺失影响叶片等侧生器官变小; 过量表达时叶细胞数目增加, 叶片变大(Mizukami and Fischer, 2000).CYCD家族基因在叶片发育过程中调控叶细胞数目, 大多能够与ANT相互作用促进叶原基细胞分裂, 影响叶片大小(Dewitte et al., 2007).AP2是miR172的靶基因, 同时与ARF3相互作用调控分生组织的分化, 影响叶片的形态(Würschum et al., 2006; Mlotshwa et al., 2006).AGO10通过REV下调ARF2的表达, 负向调控生长素响应信号, 促进顶端分生组织干细胞向叶腋分生组织的分化(Zhu et al., 2011; Roodbarkelari et al., 2015).由此可知, 上述基因的同源基因可能影响玉米叶片的发育. ...
1 2006
... 在拟南芥中, ARF3/ARF4和PHB分别调控远轴和近轴组织的形成.生长素响应因子基因ARF3/ARF4是tasiR-ARF的靶基因, 而HD-ZIPlll类基因PHB (PHV和REV)包含miR165/166的结合位点, 上述2种小RNAs分别存在于叶原基的近轴和远轴部位, 通过限制其靶基因的表达影响叶片远轴端和近轴端细胞的分化(Pekker et al., 2005; Hunter et al., 2006; Kelley et al., 2012; Dotto et al., 2014).ARF3/ARF4受tasiR-ARF调控的同时, 与KAN1 (KANAD1)家族基因相互协调作用于远轴组织的形成.DRL能够上调SAM中KNOX (kn1)家族基因的表达, 保持其细胞分裂活性, 而在叶片中抑制YABBY和KAN1类基因, 可促进近轴组织的分化发育(Jun et al., 2015).AS2是HD-ZIP类转录因子, 促进叶片近轴细胞的分化, 远轴端AS2的启动子被AtKAN1结合并抑制其表达(Iwakawa et al., 2007).另外, HYL1通过正向调控miR165/166、miR319及miR160维持远轴-近轴的正常发育调节叶片形态.ROT3和LNG2均作用于纵向极性细胞伸长膨大, 从而影响叶片变长, 但其作用机理不同(Kim et al., 1998; Mallory et al., 2004).CLF功能的缺失导致H3K27me3的活性降低, 引起叶片卷曲变窄(Schatlowski et al., 2010).ANT功能缺失影响叶片等侧生器官变小; 过量表达时叶细胞数目增加, 叶片变大(Mizukami and Fischer, 2000).CYCD家族基因在叶片发育过程中调控叶细胞数目, 大多能够与ANT相互作用促进叶原基细胞分裂, 影响叶片大小(Dewitte et al., 2007).AP2是miR172的靶基因, 同时与ARF3相互作用调控分生组织的分化, 影响叶片的形态(Würschum et al., 2006; Mlotshwa et al., 2006).AGO10通过REV下调ARF2的表达, 负向调控生长素响应信号, 促进顶端分生组织干细胞向叶腋分生组织的分化(Zhu et al., 2011; Roodbarkelari et al., 2015).由此可知, 上述基因的同源基因可能影响玉米叶片的发育. ...
1 2001
... 叶原初生细胞对于叶片形态建成具有重要影响.叶原初生细胞确定后, 叶片开始建立长、宽、厚3个轴向的发育, 这3个轴向包括基部(proximal)-末端(distal)、中央(central)-边缘(marginal)、远轴(adaxial)-近轴(abaxial), 分别受相关基因调控(Scanlon et al., 2000).玉米KNOTTED1和AN3/GIF1分别通过调控顶端分生组织(shoot apical meristem, SAM)基部细胞分化和细胞伸长膨大影响叶片发育(Ramirez et al., 2009; Nelissen et al., 2015), KNOTTED1调控P0叶原基分化形成, 在基部-末端轴向形成过程中表达量逐渐降低, 甚至在发育成熟的叶片功能区并未表达, 而KNOTTED1在叶片发育过程的末端表达时, 叶片中脉发育提前终止, 叶片分叉且卷曲(Ramirez et al., 2009).叶片发育伸展过程中, rgd1/lbl1直接或间接下调knotted1的表达, 两者相互协调建立基部-末端轴向, 并促进叶原细胞分裂, 加快叶片侧向生长(Tim- mermans et al., 1998; Ramirez et al., 2009).此外, 水稻OsH1、OsH15和OsH43属于KNOX家族, 突变后表现与knotted1、knox5和knox1相似的叶部异常表型(Matsuoka et al., 1993; Sentoku et al., 2000; Nagasaki et al., 2001).由此可知, knotted1、AN3/GIF1、rgd1/lbl1和OsH-like对基部-末端轴向的形成具有重要影响. ...
1 2012
... 中部细胞的横向发育是影响叶片宽度的关键因素, 而玉米中央-边缘轴向的研究最为深入, ns1和ns2在该轴向的建立中发挥重要作用.研究发现ns1和ns2主要在玉米SAM的叶建成组织细胞中表达, 由于其功能缺失影响叶原基边缘区的发育, 导致成熟叶片及叶鞘变窄, ns1/ns2双突变体无叶片边缘区(Scanlon et al., 2000; Nardmann et al., 2004).PRS-like与ns1同源, 而PRS-like与AtPRS1具有同源结构域.拟南芥Prs功能缺失导致叶片及花萼外缘横向生长被抑制, 但叶片和花萼远-近轴轴向并未发生变化(Matsumoto and Okada, 2001; Nakata et al., 2012), AtPRS1是侧生器官外缘分生组织的激活因子, 与NS1和NS2有相似的功能.因此, PRS-like可能参与中央-边缘轴向的发育. ...
1 2004
... 中部细胞的横向发育是影响叶片宽度的关键因素, 而玉米中央-边缘轴向的研究最为深入, ns1和ns2在该轴向的建立中发挥重要作用.研究发现ns1和ns2主要在玉米SAM的叶建成组织细胞中表达, 由于其功能缺失影响叶原基边缘区的发育, 导致成熟叶片及叶鞘变窄, ns1/ns2双突变体无叶片边缘区(Scanlon et al., 2000; Nardmann et al., 2004).PRS-like与ns1同源, 而PRS-like与AtPRS1具有同源结构域.拟南芥Prs功能缺失导致叶片及花萼外缘横向生长被抑制, 但叶片和花萼远-近轴轴向并未发生变化(Matsumoto and Okada, 2001; Nakata et al., 2012), AtPRS1是侧生器官外缘分生组织的激活因子, 与NS1和NS2有相似的功能.因此, PRS-like可能参与中央-边缘轴向的发育. ...
1 2015
... 叶原初生细胞对于叶片形态建成具有重要影响.叶原初生细胞确定后, 叶片开始建立长、宽、厚3个轴向的发育, 这3个轴向包括基部(proximal)-末端(distal)、中央(central)-边缘(marginal)、远轴(adaxial)-近轴(abaxial), 分别受相关基因调控(Scanlon et al., 2000).玉米KNOTTED1和AN3/GIF1分别通过调控顶端分生组织(shoot apical meristem, SAM)基部细胞分化和细胞伸长膨大影响叶片发育(Ramirez et al., 2009; Nelissen et al., 2015), KNOTTED1调控P0叶原基分化形成, 在基部-末端轴向形成过程中表达量逐渐降低, 甚至在发育成熟的叶片功能区并未表达, 而KNOTTED1在叶片发育过程的末端表达时, 叶片中脉发育提前终止, 叶片分叉且卷曲(Ramirez et al., 2009).叶片发育伸展过程中, rgd1/lbl1直接或间接下调knotted1的表达, 两者相互协调建立基部-末端轴向, 并促进叶原细胞分裂, 加快叶片侧向生长(Tim- mermans et al., 1998; Ramirez et al., 2009).此外, 水稻OsH1、OsH15和OsH43属于KNOX家族, 突变后表现与knotted1、knox5和knox1相似的叶部异常表型(Matsuoka et al., 1993; Sentoku et al., 2000; Nagasaki et al., 2001).由此可知, knotted1、AN3/GIF1、rgd1/lbl1和OsH-like对基部-末端轴向的形成具有重要影响. ...
1 2005
... 在拟南芥中, ARF3/ARF4和PHB分别调控远轴和近轴组织的形成.生长素响应因子基因ARF3/ARF4是tasiR-ARF的靶基因, 而HD-ZIPlll类基因PHB (PHV和REV)包含miR165/166的结合位点, 上述2种小RNAs分别存在于叶原基的近轴和远轴部位, 通过限制其靶基因的表达影响叶片远轴端和近轴端细胞的分化(Pekker et al., 2005; Hunter et al., 2006; Kelley et al., 2012; Dotto et al., 2014).ARF3/ARF4受tasiR-ARF调控的同时, 与KAN1 (KANAD1)家族基因相互协调作用于远轴组织的形成.DRL能够上调SAM中KNOX (kn1)家族基因的表达, 保持其细胞分裂活性, 而在叶片中抑制YABBY和KAN1类基因, 可促进近轴组织的分化发育(Jun et al., 2015).AS2是HD-ZIP类转录因子, 促进叶片近轴细胞的分化, 远轴端AS2的启动子被AtKAN1结合并抑制其表达(Iwakawa et al., 2007).另外, HYL1通过正向调控miR165/166、miR319及miR160维持远轴-近轴的正常发育调节叶片形态.ROT3和LNG2均作用于纵向极性细胞伸长膨大, 从而影响叶片变长, 但其作用机理不同(Kim et al., 1998; Mallory et al., 2004).CLF功能的缺失导致H3K27me3的活性降低, 引起叶片卷曲变窄(Schatlowski et al., 2010).ANT功能缺失影响叶片等侧生器官变小; 过量表达时叶细胞数目增加, 叶片变大(Mizukami and Fischer, 2000).CYCD家族基因在叶片发育过程中调控叶细胞数目, 大多能够与ANT相互作用促进叶原基细胞分裂, 影响叶片大小(Dewitte et al., 2007).AP2是miR172的靶基因, 同时与ARF3相互作用调控分生组织的分化, 影响叶片的形态(Würschum et al., 2006; Mlotshwa et al., 2006).AGO10通过REV下调ARF2的表达, 负向调控生长素响应信号, 促进顶端分生组织干细胞向叶腋分生组织的分化(Zhu et al., 2011; Roodbarkelari et al., 2015).由此可知, 上述基因的同源基因可能影响玉米叶片的发育. ...
1 2008
... 植物激素的合成、代谢与叶片的发育密切相关.研究人员对水稻Nal1、Nal7和COW1进行研究时, 发现生长素的极性运输与合成异常, 导致水稻叶形发生变化.Nal1缺失突变导致生长素极性运输蛋白基因PIN1的表达显著降低, 进而引起生长素极性运输紊乱和细胞分裂发育异常, 从而影响维管束的正常分布及叶片发育, 最终导致叶片变窄(Qi et al., 2008; Kubo et al., 2017).Nal7编码1个生长素合成的关键蛋白, 影响细胞分裂和伸长.nal7叶片窄且SAM中生长素含量明显降低, 过量表达Nal7后突变表型得到恢复(Fujino et al., 2008).水稻YUCCA型基因COW1缺失突变引起生长素合成异常, 表现与OsYUCCA1反义植株相似的表型, 叶片卷曲且变窄(Woo et al., 2007).由此可知, 玉米NAL1-like、NAL7-like和COW1-like可能参与生长素的合成及运输, 影响叶片发育. ...
3 2009
... 叶原初生细胞对于叶片形态建成具有重要影响.叶原初生细胞确定后, 叶片开始建立长、宽、厚3个轴向的发育, 这3个轴向包括基部(proximal)-末端(distal)、中央(central)-边缘(marginal)、远轴(adaxial)-近轴(abaxial), 分别受相关基因调控(Scanlon et al., 2000).玉米KNOTTED1和AN3/GIF1分别通过调控顶端分生组织(shoot apical meristem, SAM)基部细胞分化和细胞伸长膨大影响叶片发育(Ramirez et al., 2009; Nelissen et al., 2015), KNOTTED1调控P0叶原基分化形成, 在基部-末端轴向形成过程中表达量逐渐降低, 甚至在发育成熟的叶片功能区并未表达, 而KNOTTED1在叶片发育过程的末端表达时, 叶片中脉发育提前终止, 叶片分叉且卷曲(Ramirez et al., 2009).叶片发育伸展过程中, rgd1/lbl1直接或间接下调knotted1的表达, 两者相互协调建立基部-末端轴向, 并促进叶原细胞分裂, 加快叶片侧向生长(Tim- mermans et al., 1998; Ramirez et al., 2009).此外, 水稻OsH1、OsH15和OsH43属于KNOX家族, 突变后表现与knotted1、knox5和knox1相似的叶部异常表型(Matsuoka et al., 1993; Sentoku et al., 2000; Nagasaki et al., 2001).由此可知, knotted1、AN3/GIF1、rgd1/lbl1和OsH-like对基部-末端轴向的形成具有重要影响. ... ... 在叶片发育过程的末端表达时, 叶片中脉发育提前终止, 叶片分叉且卷曲(Ramirez et al., 2009).叶片发育伸展过程中, rgd1/lbl1直接或间接下调knotted1的表达, 两者相互协调建立基部-末端轴向, 并促进叶原细胞分裂, 加快叶片侧向生长(Tim- mermans et al., 1998; Ramirez et al., 2009).此外, 水稻OsH1、OsH15和OsH43属于KNOX家族, 突变后表现与knotted1、knox5和knox1相似的叶部异常表型(Matsuoka et al., 1993; Sentoku et al., 2000; Nagasaki et al., 2001).由此可知, knotted1、AN3/GIF1、rgd1/lbl1和OsH-like对基部-末端轴向的形成具有重要影响. ... ... ; Ramirez et al., 2009).此外, 水稻OsH1、OsH15和OsH43属于KNOX家族, 突变后表现与knotted1、knox5和knox1相似的叶部异常表型(Matsuoka et al., 1993; Sentoku et al., 2000; Nagasaki et al., 2001).由此可知, knotted1、AN3/GIF1、rgd1/lbl1和OsH-like对基部-末端轴向的形成具有重要影响. ...
 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}