Abstract: The transpirational dynamics and regulation features in leaves of Ferula krylovii grown in a desert area of Xinjiang, China were monitored, recorded and analysed systematically with a high-sensitivity humidity sensor combined with a specific leaf chamber and other types of sensors. The results were compared with those from other methods such as photosynthetic meters or weighing. Parameters associated with fast regulation (within 1-2 min) and diurnal variations in transpiration rate were clearly monitored and recorded. The parameters obtained could be used to analyse the correlations between transpiration and the effect of changes in environmental factors such as temperature, light intensity, and humidity to uncover more details on the transpirational dynamics and regulation features of a plant, details that other methods are unable to provide. Because larger samples could be measured with this method, the disadvantages of other methods could be excluded, such as errors due to the selection of the local sampling site, systematic errors due to smaller gas samples, and possible mechanical stress due to the clamp of the leaf chamber. This method, combined with other types of sensors, could yield parameters that cover more extensively the transpirational water consumption and regulation of plants under varied environmental conditions and provide a more detailed dynamic perspective of plants in their adaptation to environments, with the possibility of remote, continuous monitoring.
Key words:sensors ; temperature and humidity monitoring ; Ferula krylovii ; stomata regulation ; transpiration dynamics
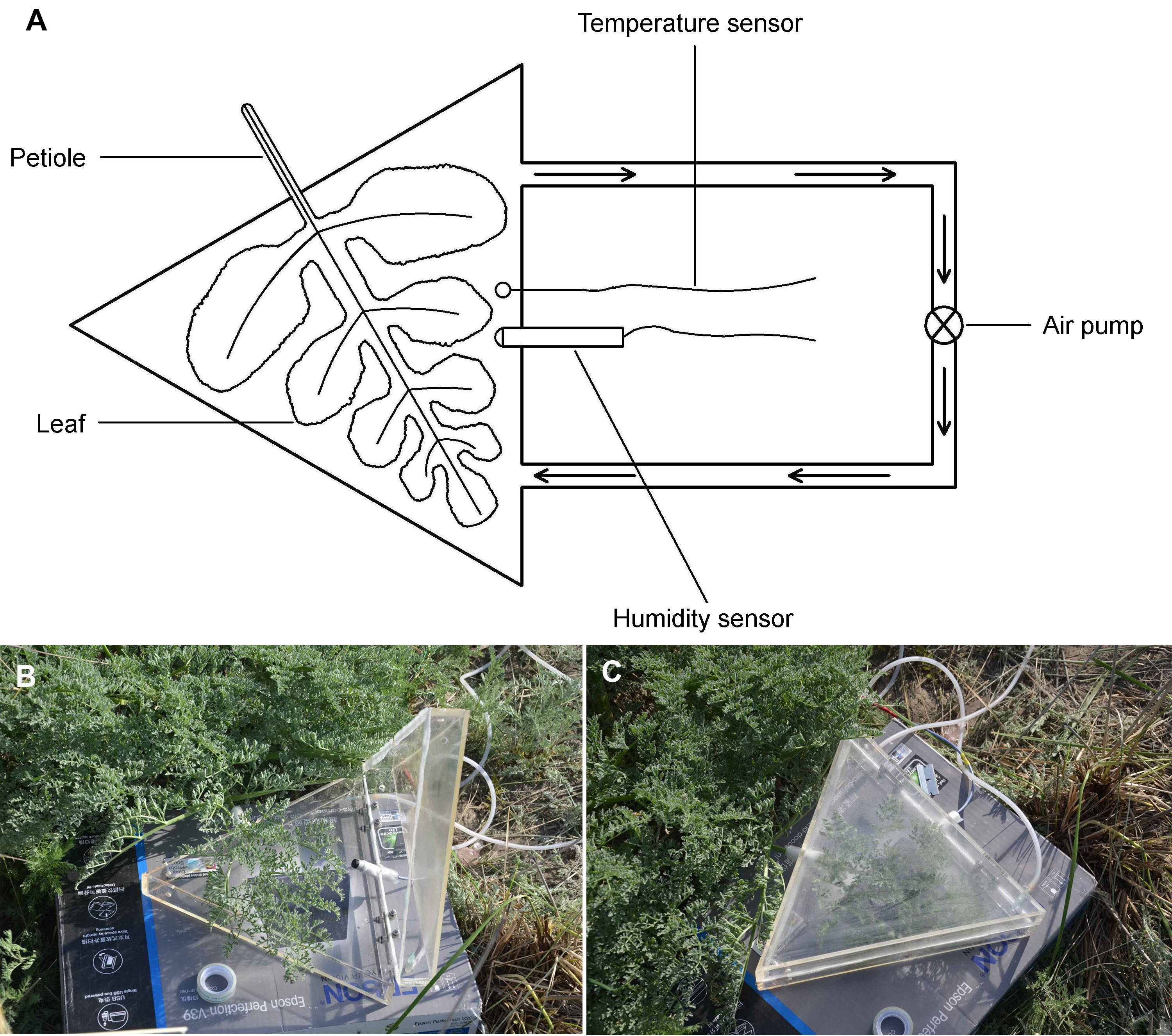
1.2 方法1.2.1 叶室结构 在为模仿阿魏大型复叶形状而特制的三角形叶室(图1A-C)内进行蒸腾测定(对于其它植物则可以根据植物或叶片形状制作其它形状的叶室)。特制叶室为等边三角形的有机玻璃盒, 叶室由上盖和底部两部分构成, 盖子和底部接合处以泡沫塑料条密封。叶室内部边长28 cm, 高7 cm (内部体积2.38 L), 能够容纳较大的整个阿魏复叶叶片。叶室上盖和下底结合部的2个侧面预先各保留1个钻孔, 以便能将叶柄和传感器分别从2个侧面伸入叶室。同时在叶室一侧的两端留有另外2个钻孔, 用于连接循环气泵, 使叶室内保持气流内循环以维持叶室内空气湿度的均匀性, 同时减少由于水蒸气扩散阻力造成的湿度传感器反应滞后。测定时, 先将叶室放置在叶片原来位置的角度和高度上, 在叶室内安装好湿度传感器, 并连接空气循环泵进出口的管子, 将整个叶片平放入叶室, 并用海绵或橡胶泥密封好叶柄在叶室侧壁上通过的地方, 立即启动空气循环泵并开始记录。循环泵气体流速为3.5 L∙min-1, 空气在直径为5 mm的循环管路进出口的速度为2.97 m·s-1, 由此带动叶室内空气形成快速的气流循环。 图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-353/img_1.png<b>图1</b> 针对托里阿魏叶片特制的叶室平面结构示意图(A)和叶室(B, C)<br/><b>Figure 1</b> A planar schematic illustration (A) and the leaf chamber (B, C) specially designed for <i>Ferula krylovii</i> Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-353/img_1.png<b>图1</b> 针对托里阿魏叶片特制的叶室平面结构示意图(A)和叶室(B, C)<br/><b>Figure 1</b> A planar schematic illustration (A) and the leaf chamber (B, C) specially designed for <i>Ferula krylovii</i>
2 结果与讨论2.1 大气温度和湿度的日变化曲线图2为大气温度和湿度的日变化曲线。图2A是大气温度从日出到日落的变化情况。由于决定水汽蒸发速率的因子不是大气湿度本身, 而是大气湿度与饱和湿度的差值, 即大气湿度越低, 大气湿度与大气饱和湿度的差值就越大, 蒸发就越强烈。图2B同时显示大气湿度的日变化和大气湿度与饱和湿度的差值(different- tial humidity)曲线。 图2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-353/img_2.png<b>图2</b> 大气温度(A)和相对湿度(B)的日变化曲线以及大气湿度与饱和湿度的差值曲线(B)<br/><b>Figure 2</b> The diurnal variations in air temperature (A), relative humidity (B), and the differential humidity (the difference between the saturation humidity and the air humidity) (B) Figure 2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-353/img_2.png<b>图2</b> 大气温度(A)和相对湿度(B)的日变化曲线以及大气湿度与饱和湿度的差值曲线(B)<br/><b>Figure 2</b> The diurnal variations in air temperature (A), relative humidity (B), and the differential humidity (the difference between the saturation humidity and the air humidity) (B)
图2 大气温度(A)和相对湿度(B)的日变化曲线以及大气湿度与饱和湿度的差值曲线(B) Figure 2 The diurnal variations in air temperature (A), relative humidity (B), and the differential humidity (the difference between the saturation humidity and the air humidity) (B)
2.2 阿魏叶片蒸腾作用的动力学分析图3为以自由水面的蒸腾模拟曲线(图3A)作为参照, 托里阿魏从9:00-13:00 (当地时间7:00-11:00)按测定时间顺序排列的阿魏叶片的蒸腾作用动力学记录曲线和指数模拟曲线(图3B-F)。从理论上来说, 如果蒸腾过程中气孔开度不变, 就等于水分蒸发表面积不变, 蒸腾过程符合指数蒸发曲线; 偏离指数蒸发曲线的蒸腾作用表明在蒸腾过程中有水分蒸发表面积的变化, 即气孔开度的调节变化。从图3B-F中可以看到上下波动或微小的震荡, 但所有曲线的动力学都接近模拟指数曲线, 表明叶片在这一时段没有表现出明显的整体性正反馈型或负反馈型气孔调节。 图3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-353/img_3.png<b>图3</b> 托里阿魏从上午9:00-13:00 (当地时间7:00-11:00)的蒸腾作用动力学记录曲线和指数模拟曲线(以自由水面的蒸发和理论模拟曲线作参照)<br/>(A) 自由水面的蒸发和理论模拟曲线; (B)-(F) 叶片的实际测定曲线和对应的理论指数模拟曲线<br/><b>Figure 3</b> The recorded dynamic curves and the corresponding theoretical simulated curve of transpiration rate in<i> Ferula krylovii</i> from 9:00 am to 13:00 pm (local time 7:00 to 11:00 am) in the morning (in comparison with the evaporation curve of free water surface)<br/>(A) The evaporation curve and the corresponding theoretical simulated curve of free water surface; (B)-(F) The actual measured curves of the leaves and the corresponding theoretical simulated curve Figure 3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-353/img_3.png<b>图3</b> 托里阿魏从上午9:00-13:00 (当地时间7:00-11:00)的蒸腾作用动力学记录曲线和指数模拟曲线(以自由水面的蒸发和理论模拟曲线作参照)<br/>(A) 自由水面的蒸发和理论模拟曲线; (B)-(F) 叶片的实际测定曲线和对应的理论指数模拟曲线<br/><b>Figure 3</b> The recorded dynamic curves and the corresponding theoretical simulated curve of transpiration rate in<i> Ferula krylovii</i> from 9:00 am to 13:00 pm (local time 7:00 to 11:00 am) in the morning (in comparison with the evaporation curve of free water surface)<br/>(A) The evaporation curve and the corresponding theoretical simulated curve of free water surface; (B)-(F) The actual measured curves of the leaves and the corresponding theoretical simulated curve
图3 托里阿魏从上午9:00-13:00 (当地时间7:00-11:00)的蒸腾作用动力学记录曲线和指数模拟曲线(以自由水面的蒸发和理论模拟曲线作参照) (A) 自由水面的蒸发和理论模拟曲线; (B)-(F) 叶片的实际测定曲线和对应的理论指数模拟曲线 Figure 3 The recorded dynamic curves and the corresponding theoretical simulated curve of transpiration rate in Ferula krylovii from 9:00 am to 13:00 pm (local time 7:00 to 11:00 am) in the morning (in comparison with the evaporation curve of free water surface) (A) The evaporation curve and the corresponding theoretical simulated curve of free water surface; (B)-(F) The actual measured curves of the leaves and the corresponding theoretical simulated curve
图4A-D为午后14:00-18:00 (当地时间12:00- 16:00)之间按测定时间顺序排列的阿魏叶片的蒸腾作用动力学记录曲线和指数模拟曲线。从图4A-D中可以看出, 当相对湿度增加到65%或更高时, 蒸腾速率出现了明显的偏离理论模拟曲线的加速现象, 即正反馈型加速。这可能是由于午后时间段蒸腾耗水量较快, 表皮细胞和保卫细胞都有轻度脱水, 影响气孔张开; 当湿度增大而蒸腾减少时, 保卫细胞出现恢复性吸水膨胀, 导致气孔开度增加, 从而使蒸腾加速。图4E和F为傍晚19:00-20:00 (当地时间17:00-18:00)阿魏叶片的蒸腾作用动力学记录曲线和指数模拟曲线。从图4E和F中可以看出, 当相对湿度增加到70%- 85%时, 蒸腾速率出现了明显的偏离理论模拟曲线和减速现象, 即负反馈型调节减速, 可能与接近傍晚光照强度下降、气孔开始加速关闭或植物生物钟性质的日变化节律有关。 图4https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-353/img_4.png<b>图4</b> 托里阿魏叶片午后的蒸腾作用动力学实际测定曲线和对应的理论指数模拟曲线<br/>(A)-(D) 14:00-18:00 (当地时间12:00-16:00); (E), (F) 19:00-20:00 (当地时间17:00-18:00)<br/><b>Figure 4</b> The recorded dynamic curves and their corresponding theoretical simulated curves of the transpiration rate in<i> Ferula krylovii</i> leaves in the afternoon<br/>(A)-(D) Curves from14:00 to 18:00 pm (local time 12:00 to 16:00 pm); (E), (F) Curves from 19:00-20:00 (local time 17:00-18:00 pm) Figure 4https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-353/img_4.png<b>图4</b> 托里阿魏叶片午后的蒸腾作用动力学实际测定曲线和对应的理论指数模拟曲线<br/>(A)-(D) 14:00-18:00 (当地时间12:00-16:00); (E), (F) 19:00-20:00 (当地时间17:00-18:00)<br/><b>Figure 4</b> The recorded dynamic curves and their corresponding theoretical simulated curves of the transpiration rate in<i> Ferula krylovii</i> leaves in the afternoon<br/>(A)-(D) Curves from14:00 to 18:00 pm (local time 12:00 to 16:00 pm); (E), (F) Curves from 19:00-20:00 (local time 17:00-18:00 pm)
图4 托里阿魏叶片午后的蒸腾作用动力学实际测定曲线和对应的理论指数模拟曲线 (A)-(D) 14:00-18:00 (当地时间12:00-16:00); (E), (F) 19:00-20:00 (当地时间17:00-18:00) Figure 4 The recorded dynamic curves and their corresponding theoretical simulated curves of the transpiration rate in Ferula krylovii leaves in the afternoon (A)-(D) Curves from14:00 to 18:00 pm (local time 12:00 to 16:00 pm); (E), (F) Curves from 19:00-20:00 (local time 17:00-18:00 pm)
2.3 不同测定方法下阿魏叶片蒸腾速率比较图5为不同方法测定得到的托里阿魏叶片蒸腾速率的日变化趋势曲线。图5A为湿度传感器的测定结果; 图5B为光合仪的测定结果; 图5C为用湿度传感器测定相对湿度标准化到65%时叶片的蒸腾速率日变化趋势。湿度传感器的测定结果(峰值152.4 g·m-2·h-1)更接近于称重测定的结果(图6A中30分钟的蒸腾值乘以2, 峰值为168.2 g·m-2·h-1); 而光合仪的测定结果(峰值118.4 g·m-2·h-1)略小, 个体差异也较大, 可能与取样面积小、误差偏大有关。同时, 光合仪的叶室在夹叶片时施加的机械压力对叶片是否有影响也不明确。从图5A, B可以看出, 托里阿魏在自然条件下的蒸腾速率日变化呈现不明显的双峰, 当叶室相对湿度都标准化到65%时(图5C), 大约在下午14:00-15:00 (当地时间12:00-13:00)叶片蒸腾作用出现了明显的变化(双峰)。这表明大气干燥或其它因素导致的蒸腾速率增大可能掩盖了叶片的气孔调节效应, 即阿魏叶片也有午休现象; 也表明用湿度传感器结合特制叶室能够对植物的蒸腾作用进行更为精细的分析。 图5https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-353/img_5.png<b>图5</b> 托里阿魏叶片蒸腾速率的日变化趋势<br/>(A) 湿度传感器测定结果; (B) TPS-2光合仪测定结果; (C) 相对湿度标准化到65%时用湿度传感器测定的结果<br/><b>Figure 5</b> The diurnal variations in the transpiration rate of <i>Ferula krylovii</i> leaves<br/>(A) The results of humidity sensors; (B) The results of a TPS-2 portable photosynthesis system; (C) The diurnal variations results of humidity sensors in the transpiration rate in <i>Ferula krylovii</i> normalized to 65% relative humidity Figure 5https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-353/img_5.png<b>图5</b> 托里阿魏叶片蒸腾速率的日变化趋势<br/>(A) 湿度传感器测定结果; (B) TPS-2光合仪测定结果; (C) 相对湿度标准化到65%时用湿度传感器测定的结果<br/><b>Figure 5</b> The diurnal variations in the transpiration rate of <i>Ferula krylovii</i> leaves<br/>(A) The results of humidity sensors; (B) The results of a TPS-2 portable photosynthesis system; (C) The diurnal variations results of humidity sensors in the transpiration rate in <i>Ferula krylovii</i> normalized to 65% relative humidity
图5 托里阿魏叶片蒸腾速率的日变化趋势 (A) 湿度传感器测定结果; (B) TPS-2光合仪测定结果; (C) 相对湿度标准化到65%时用湿度传感器测定的结果 Figure 5 The diurnal variations in the transpiration rate of Ferula krylovii leaves (A) The results of humidity sensors; (B) The results of a TPS-2 portable photosynthesis system; (C) The diurnal variations results of humidity sensors in the transpiration rate in Ferula krylovii normalized to 65% relative humidity
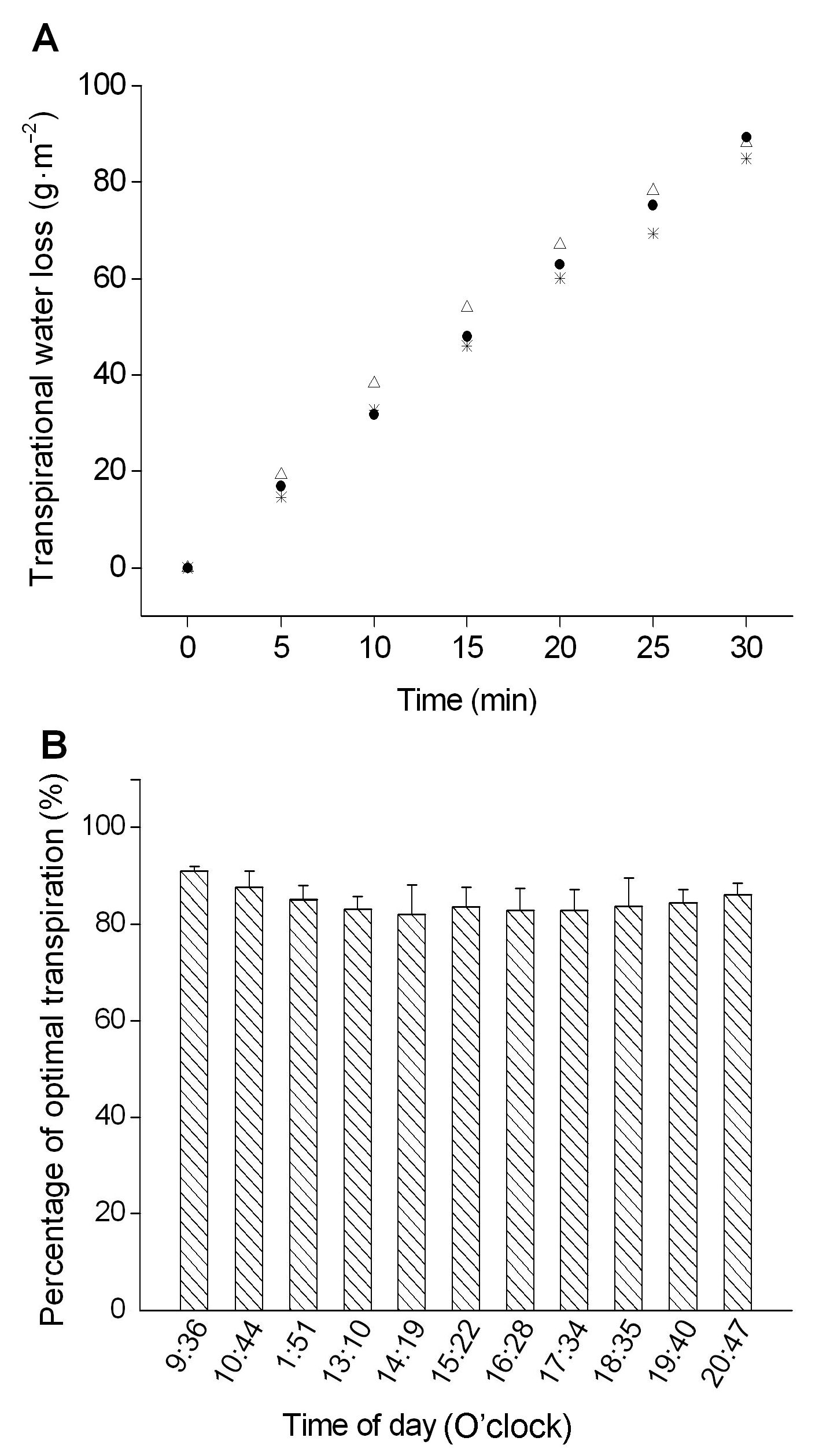
图6https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-353/img_6.png<b>图6</b> 托里阿魏原位叶片的蒸腾速率(A)与相同条件下消除了根系吸水阻力的最适蒸腾速率的百分率比值(B)<br/><b>Figure 6</b> The transpiration rate of <i>Ferula krylovii in situ</i> leaves (A) and the ratio of the optimal transpiration rate whereby the root resistance to water absorption was eliminated (B) Figure 6https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-353/img_6.png<b>图6</b> 托里阿魏原位叶片的蒸腾速率(A)与相同条件下消除了根系吸水阻力的最适蒸腾速率的百分率比值(B)<br/><b>Figure 6</b> The transpiration rate of <i>Ferula krylovii in situ</i> leaves (A) and the ratio of the optimal transpiration rate whereby the root resistance to water absorption was eliminated (B)
图6 托里阿魏原位叶片的蒸腾速率(A)与相同条件下消除了根系吸水阻力的最适蒸腾速率的百分率比值(B) Figure 6 The transpiration rate of Ferula krylovii in situ leaves (A) and the ratio of the optimal transpiration rate whereby the root resistance to water absorption was eliminated (B)
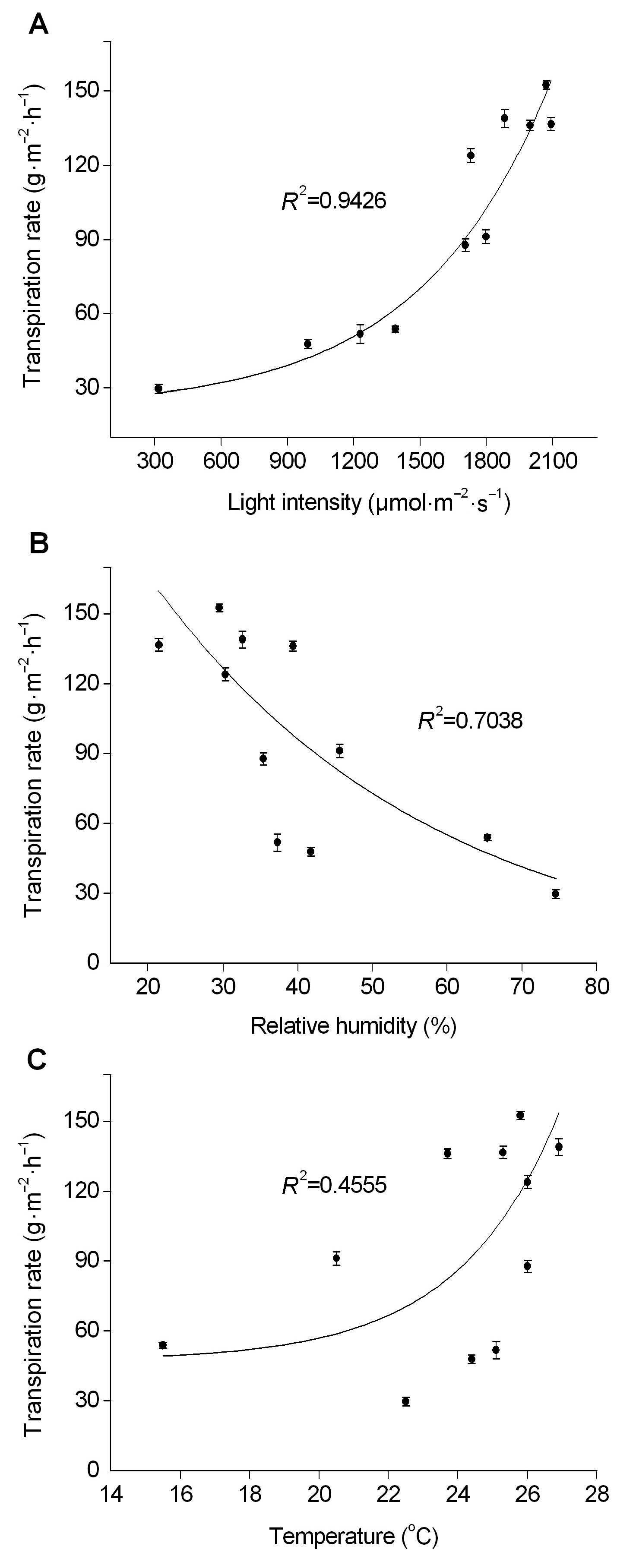
2.5 不同环境因子的变化对阿魏叶片蒸腾速率的影响图7是用湿度传感器测定的叶片初始蒸腾速率与3个环境因子之间的回归分析曲线。从图7A可以看到, 叶片蒸腾速率随着光照强度的增加升高很快, 有较高的相关性(R2=0.942 6), 比较接近指数上升曲线。叶片的蒸腾速率与湿度(图7B)也有一定的负相关关系(R2=0.703 8), 但相关性明显低于叶片蒸腾速率与光照强度的关系。从图7C可以看到, 叶片蒸腾速率与温度之间的相关性在3者之中最低(R2=0.455 3)。以上结果表明, 阿魏叶片的蒸腾作用受光照强度影响最大, 其次是大气湿度, 温度的影响最不明显。 图7https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-353/img_7.png<b>图7</b> 托里阿魏叶片蒸腾速率与光照强度(A)、湿度(B)和温度(C)之间关系的回归曲线<br/><b>Figure 7</b> The regression curves on the relationship between the transpiration rate of <i>Ferula krylovii </i>leaves to the light intensity (A), relative humidity (B) and temperature (C) Figure 7https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-353/img_7.png<b>图7</b> 托里阿魏叶片蒸腾速率与光照强度(A)、湿度(B)和温度(C)之间关系的回归曲线<br/><b>Figure 7</b> The regression curves on the relationship between the transpiration rate of <i>Ferula krylovii </i>leaves to the light intensity (A), relative humidity (B) and temperature (C)
图7 托里阿魏叶片蒸腾速率与光照强度(A)、湿度(B)和温度(C)之间关系的回归曲线 Figure 7 The regression curves on the relationship between the transpiration rate of Ferula krylovii leaves to the light intensity (A), relative humidity (B) and temperature (C)
AkhmetovaA, MukhitdinovN, YdyrysA (2015). Anatomical indicators of the leaf structure of Ferula iliensis, growing in the eastern part of Zailiyskiy Alatau (big Boguty mountains).Pak J Bot 47, 511-515. [本文引用: 1]
[9]
ChaudharyDD, NayseSP, WaghmareLM (2011). Application of wireless sensor networks for greenhouse parameter control in precision agriculture.Int J Wirel Mobile Netw 3, 1-6. [本文引用: 1]
[10]
IrmakS, KabengeI, RudnickD, KnezevicS, WoodwardD, MoravekM (2013). Evapotranspiration crop coefficients for mixed riparian plant community and transpiration crop coefficients for common reed, cottonwood and peach-leaf willow in the Platte River Basin, Nebraska-USA.J Hydrol 481, 177-190. [本文引用: 1]
[11]
LendvayB, KalaposT (2014). Population dynamics of the climate-sensitive endangered perennial Ferula sadleriana Ledeb.(Apiaceae).Plant Spec Biol 29, 138-151. [本文引用: 1]
[12]
LiuMZ, JiangGM, LiYG, NiuSL, GaoLM, DingL, PengY (2003). Leaf osmotic potentials of 104 plant species in relation to habitats and plant functional types in Hunshandak Sandland, Inner Mongolia, China.Trees 17, 554-560. [本文引用: 1]
[13]
Millan-AlmarazJR, De Jesus Romero-TroncosoR, Gue- vara-GonzalezRG, Contreras-MedinaLM, Carrillo- SerranoRV, Osornio-RiosRA, Duarte-GalvanC, Rios- AlcarazMA, Torres-PachecoI (2010). FPGA-based fused smart sensor for real-time plant-transpiration dyna- mic estimation.Sensors 10, 8316-8331. [本文引用: 1]
[14]
MontaldoN, CoronaR, AlbertsonJD (2013). On the separate effects of soil and land cover on Mediterranean ecohydrology: two contrasting case studies in Sardinia, Italy.Water Resour Res 49, 1123-1136. [本文引用: 1]
[15]
MurphyDM, KoopT (2005). Review of the vapour pressures of ice and supercooled water for atmospheric applications.Quart J Roy Meteor Soc 131, 1539-1565. [本文引用: 1]
SavageMJ (2010). Field evaluation of polymer capacitive humidity sensors for Bowen ratio energy balance flux mea- surements.Sensors 10, 7748-7771. [本文引用: 1]
[18]
TamangB, AndreuMG, RockwoodDL (2010). Microclimate patterns on the leeside of single-row tree windbreaks during different weather conditions in Florida farms: implications for improved crop production.Agrof Syst 79, 111-122. [本文引用: 1]
[19]
YaqoobU, NawchooIA (2017a). Conservation and cultivation of Ferula jaeschkeana Vatke: a species with deep complex morphophysiological dormancy.Proc Natl Acad Sci India Sect B Biol Sci 87, 315-325. [本文引用: 1]
[20]
YaqoobU, NawchooIA (2017b). Impact of habitat variability and altitude on growth dynamics and reproductive allocation in Ferula jaeschkeana Vatke.J King Saud Univ Sci 29, 19-27. [本文引用: 1]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}