魏华1,2, 王岩1,2, 刘宝辉3, 王雷1,2,*, 1中国科学院植物研究所植物分子生理学重点实验室, 北京 1000932中国科学院大学, 北京 1000493广州大学生命科学学院, 广州 510006 WeiHua1,2, WangYan1,2, LiuBaohui3, WangLei1,2,*, 1Key Laboratory of Plant Molecular Physiology, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China2University of Chinese Academy of Sciences, Beijing 100049, China3School of Life Sciences, Guangzhou University, Guangzhou 510006, China 引用本文 魏华, 王岩, 刘宝辉, 王雷. 植物生物钟及其调控生长发育的研究进展. 植物学报, 2018, 53(4): 456-467
Abstract: The plant circadian system mostly includes input pathways, a core oscillator and output pathways to sense and anticipate the timing cues of the environment to optimize plant growth and fitness. As the cellular core coordinating system, the plant circadian system can sense the daily recurring light and temperature dynamics to coordinate the metabolism and multiple physiology processes, providing an adaptive advantage for plant growth and development. The core circadian oscillator regulates multiple complex downstream networks at various levels. Here, we summarize recent major research progress in deciphering the underlying mechanisms of the core oscillator and its regulatory networks. We also highlight a few fundamental questions needing to be resolved.
Key words:circadian system ; environmental adaption ; growth and development ; circadian rhythm ; stresses response
由于地球自转而产生的光-暗循环以及由该循环产生的温度昼夜周期性的变化, 是植物在生长发育过程中必须应对的昼夜节律性动态变化。在长期进化过程中产生的生物钟系统赋予了植物对光温等环境因子昼夜周期性变化的适应性。生物钟系统使植物可以追踪环境因子的昼夜周期性变化并感知和预测这种变化的动态及季节的变换, 以协调体内代谢稳态、细胞运动以及生长发育与防御反应的动态平衡, 使植物能在合适的时间完成其关键生长发育过程(Green- ham and McClung, 2015)。人们最初认为生物钟系统是一个线性过程, 即包括外界信号输入、内源的“计时员”(追踪时间的分子机制)和输出途径, 如叶片运动和基因节律性表达等(Harmer, 2009)。最新研究表明, 受生物钟调控的某些生物学过程也会反馈调节生物钟的核心振荡器, 进而形成一个复杂的互作调控网络, 使植物更好地适应生存环境(Nohales and Kay, 2016)。生物钟系统协调植物外部光温信号与内源的代谢和发育状态, 以调控生长发育与胁迫应答之间的平衡, 并决定昼夜周期性信号的输出, 从而使生物体避免资源和能量的损失以达到最优化生长。
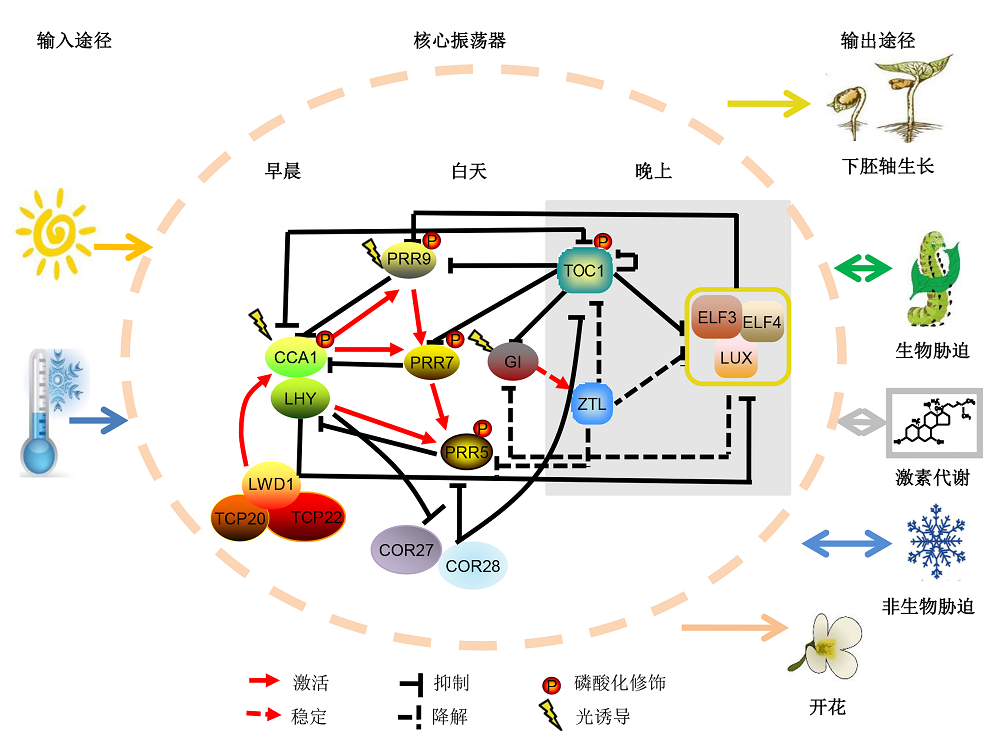
1 拟南芥生物钟核心振荡器的分子机制拟南芥(Arabidopsis thaliana)作为模式植物被广泛用于植物分子生物学和遗传学研究, 因此其生物钟分子机制也研究得较为透彻。1995年, Millar等(1995)首次采用荧光素酶作为报告基因鉴定到第1个生物钟突变体toc1 (timing of cab expression 1)。随后, Wang和Tobin (1998)利用反向遗传学方法鉴定到第1个生物钟核心振荡器的关键成员CCA1 (CIRCADIAN CLOCK ASSOCIATED 1), 该基因编码1个MYB类转录因子。超表达CCA1会使生物钟报告基因丧失昼夜节律性表达模式, 而且极度晚花, 同时也抑制内源CCA1和LHY (LATE ELONGATED HYPOCOTYL)的表达, 且该基因自身的转录水平还受到红光的瞬时诱导。基于基因表达水平和启动子活性分析, AlabadÍ等(2001)提出了拟南芥生物钟第1个分子反馈环路, 即CCA1/LHY抑制TOC1的表达, 而TOC1则促进CCA1/LHY的表达, 由此形成了生物钟核心振荡器的转录-翻译反馈环。随着技术手段的进步和计算生物学的发展, 目前认为生物钟核心振荡器由3个紧密关联的转录-翻译反馈环组成(图1)。其中, 中心反馈环由LHY和CCA1以及TOC1组成。CCA1和LHY在早晨表达, 二者存在部分功能冗余, 它们转录并翻译成熟后会相互作用进而抑制TOC1的表达。在晚上, CCA1和LHY的表达水平降低, TOC1的表达则达到峰值, 反过来抑制CCA1和LHY的表达, 进而使它们处于此起彼伏的状态(Gendron et al., 2012; Huang et al., 2012)。CCA1/LHY也和PRR (PSEUDO RESPONSE REGULATOR)基因家族的PRR9/PRR7组成早反馈环。PRRs成员中PRR9、PRR7及PRR5从早到晚每隔2-3小时依次出现表达峰值, 它们会通过与TOPL- ESS基因家族的成员互作招募组蛋白去乙酰化酶HDA6/19 (HISTONE DEACETYLASE 6/19)形成转录抑制复合体, 从而在转录水平上抑制CCA1和LHY的表达(Nakamichi et al., 2010; Wang et al., 2013)。反之, CCA1/LHY也抑制PRR9/7的表达, 从而形成早反馈环。晚反馈环包含MYB类转录因子LUX ARRHYTHMO (又命名为PHYTO-CLOCK1)以及2个核蛋白ELF3 (EARLY FLOWERING 3)和ELF4组成的EC复合体(evening complex) (Nusinow et al., 2011)和TOC1。EC复合体与CCA1/LHY相互调节, 间接促进CCA1/LHY的表达, 反过来CCA1和LHY会直接结合在EC复合体启动子上抑制其转录(Adams et al., 2015)。早反馈环、中心反馈环和晚反馈环相互连锁形成生物钟核心振荡器的抑制振荡子(repressilator)基本架构。 图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-456/img_1.png<b>图1</b> 生物钟调控拟南芥生长发育模型<br/>拟南芥生物钟系统主要包括输入途径、核心振荡器和输出途径三大部分。其中核心振荡器由多个相互联锁的转录翻译反馈环组成, 是生物钟系统的重要组成部分, 包括早循环、中心循环和晚循环, 它能够整合外界环境信号协调多种生理进程(虚线椭圆内部分)。核心振荡器接收传入的环境时间信号, 在细胞中产生内源性的昼夜节律, 并将时间信息传达到输出途径, 从而控制众多生命活动, 下胚轴伸长、病原体防御、生物和非生物胁迫响应、激素代谢和光周期调控的开花等。<br/><b>Figure 1</b> A proposed model of circadian clock regulated growth and development in Arabidopsis<br/>In higher plants, circadian clock is composed of three major parts, including input pathways, core oscillator and output pathways. The core oscillator is formed by interlocked feedback loops, such as morning loop, central loop and evening loop, indicated by the dotted oval in the figure. Circadian core oscillator regulates multiple outputs, such as hypocotyl elongation, pathogen defense, biotic and abiotic stress adaption, plant hormone signaling pathways and flowering time. Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-456/img_1.png<b>图1</b> 生物钟调控拟南芥生长发育模型<br/>拟南芥生物钟系统主要包括输入途径、核心振荡器和输出途径三大部分。其中核心振荡器由多个相互联锁的转录翻译反馈环组成, 是生物钟系统的重要组成部分, 包括早循环、中心循环和晚循环, 它能够整合外界环境信号协调多种生理进程(虚线椭圆内部分)。核心振荡器接收传入的环境时间信号, 在细胞中产生内源性的昼夜节律, 并将时间信息传达到输出途径, 从而控制众多生命活动, 下胚轴伸长、病原体防御、生物和非生物胁迫响应、激素代谢和光周期调控的开花等。<br/><b>Figure 1</b> A proposed model of circadian clock regulated growth and development in Arabidopsis<br/>In higher plants, circadian clock is composed of three major parts, including input pathways, core oscillator and output pathways. The core oscillator is formed by interlocked feedback loops, such as morning loop, central loop and evening loop, indicated by the dotted oval in the figure. Circadian core oscillator regulates multiple outputs, such as hypocotyl elongation, pathogen defense, biotic and abiotic stress adaption, plant hormone signaling pathways and flowering time.
图1 生物钟调控拟南芥生长发育模型 拟南芥生物钟系统主要包括输入途径、核心振荡器和输出途径三大部分。其中核心振荡器由多个相互联锁的转录翻译反馈环组成, 是生物钟系统的重要组成部分, 包括早循环、中心循环和晚循环, 它能够整合外界环境信号协调多种生理进程(虚线椭圆内部分)。核心振荡器接收传入的环境时间信号, 在细胞中产生内源性的昼夜节律, 并将时间信息传达到输出途径, 从而控制众多生命活动, 下胚轴伸长、病原体防御、生物和非生物胁迫响应、激素代谢和光周期调控的开花等。 Figure 1 A proposed model of circadian clock regulated growth and development in Arabidopsis In higher plants, circadian clock is composed of three major parts, including input pathways, core oscillator and output pathways. The core oscillator is formed by interlocked feedback loops, such as morning loop, central loop and evening loop, indicated by the dotted oval in the figure. Circadian core oscillator regulates multiple outputs, such as hypocotyl elongation, pathogen defense, biotic and abiotic stress adaption, plant hormone signaling pathways and flowering time.
除上述生物钟核心振荡器基本构架模型以外, 一些参与生物钟周期调控的新机制和新成员也陆续被发现和鉴定。近期研究表明, CCA1和LHY直接抑制晚间基因PRR5的表达, 进而保持PRR5在早晨处于抑制状态; 反之, 在晚间PRR5会抑制CCA1和LHY表达, 使二者在晚间处于转录抑制状态(Kamioka et al., 2016)。已有研究表明, 光能够诱导CCA1表达。利用数学模型, 人们发现TCP20 (TEOSINTE BR- ANCHED1-CYCLOIDEA-PCF20)和TCP22能够与LWD1/2 (LIGHT-REGULATED WD1/2)形成转录激活复合体, 结合在CCA1启动子区域激活CCA1表达(Wu et al., 2016)。最近研究表明, 受蓝光和温度调 控的COR27/COR28 (COLD-REGULATED GENE 27/28)同样受生物钟调控, 即其表达在晚间达到峰值, 且二者功能缺失后生物钟周期延长。进一步研究表明, COR27和COR28通过抑制核心振荡器组分PRR5和TOC1的表达而调控植物生物节律, 进而调控植物开花时间和冷胁迫响应(Li et al., 2016b)。另外, 转录组数据分析显示, COR27和COR28参与调控晚间基因TOC1、ELF4以及冷胁迫响应相关基因的表达。进一步研究表明, CCA1结合在COR27和COR- 28的启动子上抑制二者表达。以上研究表明, COR27和COR28可能是生物钟组分的新成员(Wang et al., 2017)。 生物钟核心振荡器除了存在转录-翻译反馈环的相互促进和相互抑制调控之外, 还存在复杂的转录后调控。例如, F-box类蛋白ZTL (ZEITLUPE)参与PRR5和TOC1蛋白的降解过程。PRR5可以通过PR结构域与TOC1蛋白互作, 从而促进TOC1蛋白的磷酸化及其向细胞核内的迁移过程(Wang et al., 2010)。GI (GIGANTEA)可以作为分子伴侣蛋白与HSP90 (HE- AT SHOCK PROTEIN 90)以及ZTL形成三元复合体, 进而促进ZTL成为有活性的形式, 发挥其转录后调控的功能(Cha et al., 2017)。除此之外, mRNA的成熟与加工过程也参与生物钟的周期调控, 如功能保守的含SKIP (SNW/SKI-INTERACTING PROTEIN)结构域的SKIP蛋白, 参与PRR7和PRR9 mRNA的可变剪切和成熟。当SKIP1突变以后, 植物表现出温度敏感而延长生物钟周期, 并且影响生物钟系统对光信号的敏感性, 进而影响光信号输入(Wang et al., 2012)。另外, 生物钟核心振荡器的关键组成因子也受到如磷酸化和糖基化等翻译后修饰以及转录水平的表观遗传修饰(Seo and Mas, 2014; Nolte and Staiger, 2015; Romanowski and Yanovsky, 2015), 进而构成复杂的信号网络。目前, 关于蛋白质翻译后修饰调控植物生物钟周期的分子机制研究较少, 人们对于关键因子磷酸化和糖基化的生物学意义还不是非常清楚。
2 生物钟调控拟南芥生长发育与胁迫应答生物钟通过感知和预测生存环境中光温等环境因子动态变化的时间信息调控植物生长发育的诸多过程。调控的下游过程统称为生物钟的信号输出, 主要包括开花时间决定、下胚轴细胞的节律性伸长以及生物与非生物胁迫应答等(图1)。此外, 有些生物钟输出信号也通过反馈调控生物钟的核心振荡器来参与生物钟的周期调控, 从而构成复杂的信号互作网络, 以协调生物体对外界环境变化的适应性。 2.1 开花时间调控在光周期依赖途径的成花诱导过程中, 日长的变化会诱导叶片中开花整合因子FT (FLOWERING LOCUS T)基因的表达, FT蛋白被运送到顶端分生组织诱导植物开花。生物钟对植物开花时间的调控主要体现在对CO/B-BOX1 (CONSTANS)等光周期依赖途径关键基因的表达节律和时相调控上, 这些基因感受特定日长的光周期信号并且在转录和转录后水平上受到生物钟系统多层级的调控与修饰(Song et al., 2015)。在拟南芥中, 长日照促进开花, CO蛋白在长日照下更稳定, 进而诱导FT基因表达; 但在短日照条件下, FT基因的表达水平非常低, 导致植物开花时间延迟(Kobayashi et al., 1999)。生物钟关键基因突变导致的生物钟功能紊乱会改变开花时间, 说明生物钟在感知日长信息调控光周期途径开花启动方面发挥重要作用。生物钟核心振荡器重要组分还在转录后水平调节CO蛋白的稳定性, 生物钟关键因子PRR5/7/9的突变导致植物极度晚花。最新研究表明, 它们通过与CO蛋白直接在细胞核中互作, 进而调控CO蛋白的稳定性而促进开花。由于PRR蛋白表达的时间特异性, 它们能与CO蛋白在一天之中的特定时间内互作而增强其稳定性, 并促进CO蛋白结合在FT启动子上, 从而诱导FT的表达, 促进植物开花(Hayama et al., 2017)。上述研究从分子水平部分解析了prr5/7/9突变体植物极度晚花的机制, 也为生物钟关键组分在蛋白质翻译后修饰水平调节开花时间提供了新证据。最近研究表明, 与CO蛋白家族一样含有保守B-box锌指基序, 但是不含有DNA结合结构域的BBX (B-BOX GENES)基因家族成员也可能是生物钟周期调控和开花时间调控的衔接者。例如, 研究表明BBX32 (B- BOX32)蛋白参与生物钟输出途径控制开花时间, 同时其过表达转基因株系在持续光照的条件下生物钟周期变长, 但其参与生物钟周期调控的机制还不清楚(Tripathi et al., 2017)。此外, BBX32与COL3 (CON- STANTS-LIKE 3)/BBX4相互作用抑制FT基因的表达而调控开花(Tripathi et al., 2017)。关于这类基因家族在介导开花时间与生物钟周期调控方面的研究有望成为新的研究热点, 可为更复杂层级的生物钟感知光周期途径调控开花时间的信号网络提供更多的实验证据。
2.2 下胚轴伸长的昼夜节律性调控下胚轴的伸长对于植物种子的萌发及实现从异养到自养的转变至关重要, 该过程受到光、温度和激素等信号的多层级调控。其中, PIF4 (PHYTOCHROME INTERACTING FACTOR 4)和PIF5是拟南芥中介导下胚轴伸长的关键因子, 它们属于螺旋-环-螺旋(basic helix-loop-helix, bHLH)类转录因子亚家族。该亚家族包括PIF1、PIF3、PIF4和PIF5。PIF1和PIF3蛋白在一天之中维持相对较恒定的水平, 而PIF4和PIF5的表达则呈现明显的昼夜节律性。因此认为PIF4和PIF5转录水平的表达受到生物钟更为严密的调控(Soy et al., 2016)。早在2007年, 科学家发现外部的光信号和生物钟协调互作调控下胚轴细胞的昼夜节律性伸长。研究表明, 生物钟严格调控下胚轴伸长信号整合因子PIF4和PIF5基因的昼夜节律性表达, 而光信号则在蛋白水平对蛋白稳定性和丰度进行调控, 但关于PIF4和PIF5的昼夜节律性表达的控制因子却并不十分清楚(Nozue et al., 2007)。研究表明, 由ELF3、ELF4以及转录因子LUX组成的EC复合体通过LUX直接结合在PIF4和PIF5的启动子上, 抑制PIF4和PIF5的表达, 从而调节下胚轴细胞伸长的节律性。而随着EC复合体的降解, PIF4蛋白不断积累, 使植物下胚轴的伸长速度在黎明时达到峰值(Nusinow et al., 2011)。还有研究表明, ELF3可以独立于EC复合体与PIF4通过蛋白-蛋白互作的方式调节植物生长(Nomoto et al., 2012; Nieto et al., 2015)。另外, Nakamichi等(2010, 2012)发现, PRRs基因家族中的TOC1、PRR5、PRR7及PRR9功能缺失后都会出现下胚轴伸长的表型。在短日照条件下, 下胚轴伸长速度也在黎明达到最大值。研究表明, PIF3和TOC1共同结合在一些黎明前表达的与发育相关基因的启动子上。TOC1能与PIF3和PIF4通过蛋白互作的方式抑制它们的转录激活活性, 从而抑制PIFs介导的下胚轴伸长(Soy et al., 2016; Zhu et al., 2016)。但是PRR基因家族是否会直接影响PIF基因家族的转录活性还有待深入研究。
3 生物钟调控生物胁迫和非生物胁迫应答3.1 生物胁迫应答的昼夜节律性调控早期通过转录组数据分析发现, 生物钟系统可能参与茉莉酸(jasmonate acid, JA)与水杨酸(salic acid, SA)的生物合成和信号转导途径(Covington et al., 2008), 而JA和SA被认为是调控植物响应生物胁迫的2类关键激素。MYC2是植物JA信号途径的下游关键因子, 当JA存在时, JAZ (jasmonate ZIM-domain)对MYC2的转录激活活性的阻遏效应被解除, MYC2可以激活JA信号下游效应基因的表达(Song et al., 2014)。TIC (TIME FOR COFFEE)是生物钟周期调控的关键因子, 它编码1个含有1 555个氨基酸但生化功能未知的蛋白, 其调控生物钟周期的分子机制目前还不清楚。有研究表明, TIC可以与MYC2通过蛋白互作的方式在细胞核中负调控MYC2的蛋白稳定性。因此, 在tic突变体中JA介导的免疫系统故障导致植物易感病(Shin et al., 2012)。此外, 人们也发现MYC2的表达在转录水平和蛋白水平都受到生物钟的调控(Shin et al., 2012), 其表达的峰值出现在傍晚时分, 但是转录水平上调控MYC2基因昼夜节律性表达的生物钟关键组分还有待进一步验证。最新研究显示, 由ELF4- ELF3-LUX组成的生物钟EC复合体可以在转录水平上直接介导时间维度对MYC2的转录抑制, 进而精细调节茉莉酸下游的发育事件, 如叶片衰老等(Zhang et al., 2018)。 水杨酸的生物合成和积累受到生物钟核心组分CHE (CCA1HIKING EXPEDITION)的调控(Zheng et al., 2015)。SA介导的防御反应中磷酸转移酶PHT4;1 (PHOSPHATE TRANSPORTER 4;1)的表达也受到CCA1的直接调控(Wang et al., 2014)。介导植物对真菌免疫响应的RPP4 (RECOGNITION FOR PERO- NOSPORA PARASITICA 4)基因的启动子上存在EE (evening element)元件, 该元件可被CCA1蛋白识别并结合, 从而调控RPP4基因的转录活性。CCA1对RPP4基因的转录调控使植物即使在没有病菌感染的条件下, 在侵染性真菌的孢子开始释放的黎明时分开启免疫系统, 增强植物对病原菌的免疫作用(Wang et al., 2011a)。因此, 当CCA1或者LHY超表达使生物钟节律紊乱或者LUX基因的功能缺失都会导致植物对病原菌更敏感(Bhardwaj et al., 2011; Zhang et al., 2013; Korneli et al., 2014)。虽然植物叶片表面的气孔在早晨会完全张开, 使病原菌容易入侵, 但植物会激活相关防御机制, 使其在早晨的抗病能力比夜晚更强(Zhang et al., 2013; Korneli et al., 2014)。TIC也会通过生物钟系统调控气孔开度, 进而调控植物对病原菌的敏感性(Korneli et al., 2014)。由此可见, 生物钟调节植物对生物胁迫应答存在多个节点的复杂调控, 而全面揭示这一复杂网络仍需开展更系统的研究。
3.2 生物钟调控非生物胁迫生物钟广泛调节植物对非生物胁迫的耐受性, 是协调植物生长发育与胁迫响应的重要调节者和平衡者。在冷胁迫信号应答中, 植物通过CBF/DREB1 (CREPEAT BINDING FACTOR/DEHYDRATION-RES- POSIVE-ELEMENT-BINDING)来激活COR基因的表达, 进而调控植物对冷胁迫的响应, 且CBF基因家族成员自身的表达也呈现明显的昼夜节律性(Liu et al., 1998; Fowler et al., 2005)。许多生物钟基因的突变体均表现出对冷胁迫适应性改变的表型, 如生物钟核心组分PRR5/7/9突变后, 可以间接增强CBF的表达水平使其表达丧失昼夜节律性, 从而导致植物具有更强的耐冷性(Nakamichi et al., 2009)。PRR基因家族的另一个成员TOC1的突变也会使CBF3表达水平上调, 增强植物对冷胁迫的耐受性(Keily et al., 2013), 说明PRR基因家族在植物冷胁迫的适应方面可能发挥重要作用。在农作物水稻(Oryza sativa)、小麦(Triticum aestivum)和大麦(Hordeum vulgare)中, CBF也参与调控对冷和干旱的响应, 进而影响农作物生长(Chew and Halliday, 2011)。以上结果进一步说明, 生物钟系统通过调节CBF基因家族表达的昼夜节律性来协调植物对非生物胁迫响应和自身发育所需能量分配。另一方面, 冷胁迫也可以反馈调节生物钟系统, 如CBF1结合在LUX的启动子上调控其表达(Chow et al., 2014); 低温可诱导CCA1及其它生物钟基因的可变剪切并形成不同的转录本, 从而影响生物钟的功能(Grundy et al., 2015)。核心振荡器组分还受到高温的调控, 高温主要影响生物钟组分基因的可变剪切或通过调控热激响应因子HsfB2b (HEAT SHOCK FACTOR B2B)和FBH1 (FLOW-ERING BA- SIC HELIX-LOOP-HELIX 1)分别结合在PRR7及CC- A1的启动子上来调控其表达水平(Kolmos et al., 2014; Nagel et al., 2014)。在植物干旱和盐胁迫应答方面, 生物钟系统也发挥着重要功能。研究表明, 许多与盐、脱水、渗透压调控和响应相关基因的表达均受生物钟调控(Kreps et al., 2002; Covington et al., 2008)。在拟南芥中, 参与调控盐胁迫的基因如SOS1 (SALT OVERLY SENSITIVE 1)等的表达呈现出较强的昼夜节律性(http://diurnal.mocklerlab.org/), 表明生物钟可能调控植物对盐胁迫的适应。另外, 生物钟组分GI参与调控植物在高盐环境下的Na+/H+质子泵通道的打开, 进而将Na+排出体外, 减少Na+对植物的毒害(Kim et al., 2013)。也有研究表明, PRR7可能参与调控植物响应盐胁迫(Liu et al., 2013)。但是关于生物钟系统如何调控植物对盐胁迫的响应及其相关分子机制仍需更系统地挖掘和研究, 并将有望为作物耐盐育种提供理论依据和相关遗传资源。
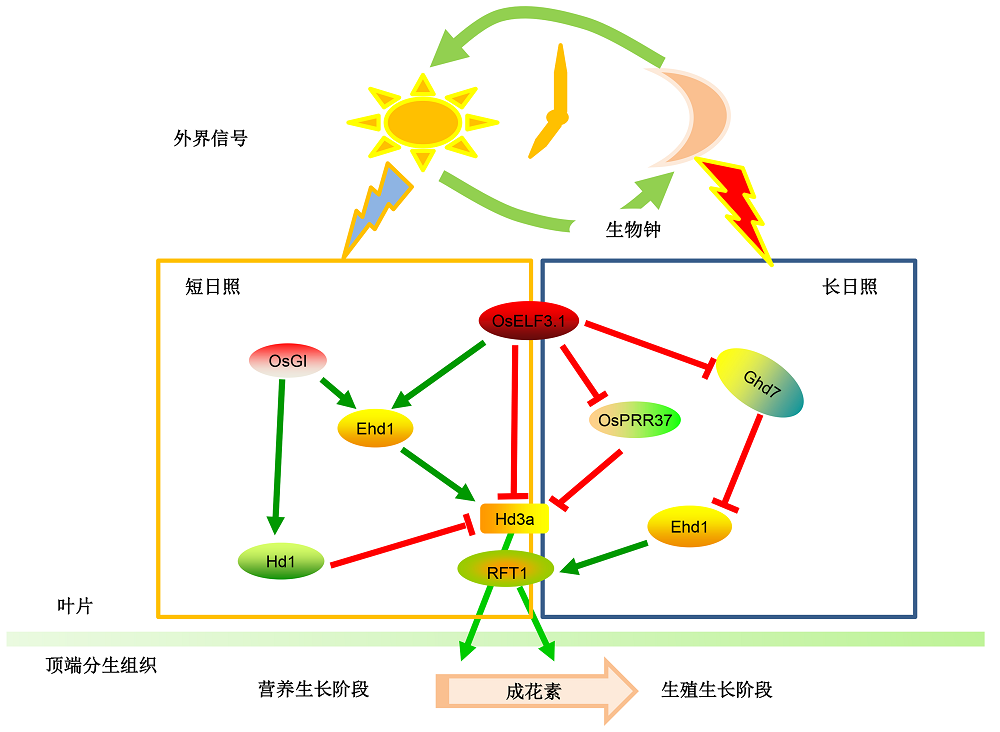
4 农作物生物钟基因挖掘及功能除了在模式植物拟南芥中发现生物钟调控生长发育过程以及应对外界环境变化和胁迫应答方面具有不可忽视的作用外, 近年来, 农作物的生物钟研究逐步展开。水稻是主要粮食作物, 生物钟如何调控水稻的生长发育(尤其是抽穗期)和非生物胁迫应答已越来越受到科学家的关注。抽穗期是水稻的重要农艺性状之一, 抽穗期的长短(从萌发到始穗)直接决定水稻品种生育期的长短, 而适宜的生育期在维持水稻高产稳产中扮演着极为关键的角色, 也在一定程度上反映水稻对种植区域和季节的适应性。研究表明, 生物钟在调控水稻抽穗期方面具有重要作用。图位克隆分析表明, OsELF3 (与拟南芥生物钟核心组分同源的ELF3)的核苷酸变异导致Nipponbare和Koshihikari品种间抽穗期的差异(Matsubara et al., 2012)。进一步研究表明, 水稻中存在2个与拟南芥ELF3同源的基因Os- ELF3.1 (LOC_Os06g05060)和OsELF3.2 (LOC_Os- 01g38530) (Zhao et al., 2012)。在长日照条件下, OsELF3.1是水稻抽穗期正调节因子, 作用于OsGI (Os-GIGANTEA)和Ghd7 (Grain number, Plant height and heading date 7)的上游, 负调控它们的表达, 因此oself3.1突变体表现出晚抽穗的表型; 但是OsELF3.2的突变并不影响水稻抽穗期。OsELF3.1突变后影响生物钟其它组分OsLHY、OsPRR1、Os- PRR37和OsPRR73的表达, 表明OsELF3.1在维持生物钟周期和调控抽穗期方面具有重要作用(Yang et al., 2013) (图2)。有趣的是, 在植物叶片衰老方面, 拟南芥ELF3通过抑制PIF4和PIF5的表达负调控叶片衰老(Sakuraba et al., 2014); 但是水稻OsELF3.1的功能恰恰相反, 它是水稻叶片衰老的正调节因子, 主要调控NAC (N-ACETYL CYSTEINE)和WRKY基因家族成员的表达(Sakuraba et al., 2016)。在拟南芥中超表达OsELF3.1则会表现出与ELF3超表达相同的表型, 即延缓叶片衰老、晚花和短周期, 表明ELF3在水稻和拟南芥中功能保守, 可能由于在不同物种中参与调控的下游靶基因完全不同, 导致其在不同植物中的功能差异(Atamian and Harmer, 2016)。水稻同源基因OsELF3.2则与E3连接酶APIP6 (AVRPIZT INTERACT PROTEIN 6)互作, 负调控水稻对稻瘟病的免疫(Ning et al., 2015)。在春大麦中ELF3通过抑制FT1的表达和产生有活性的GAs (gibberellic acid)来调控其对光周期的敏感性, 控制春大麦由营养生长阶段向生殖生长阶段的转变(Boden et al., 2014)。近期研究表明, 大麦中ELF3通过生物钟系统介导植物对温度信号的响应。受高温刺激后, CCA1和PRR73的表达水平明显降低, 而GI和PRR59的表达水平明显升高, 但是这种变化在elf3突变体中消失, 表明GI和PRR对温度响应依赖于ELF3的功能(Ford et al., 2016), 但ELF3调控大麦对温度响应的分子机制还不清楚。在另一重要作物大豆(Glycine max)中, ELF3基因突变导致其成熟时间延长, 即“长青春期”, 这能够提高大豆产量, 增强大豆在低纬度地区的区域适应性(Lu et al., 2017; Yue et al., 2017)。进一步研究表明, GmELF3结合在E1启动子上抑制其表达, 进而使下游FT2a和FT5a的表达水平降低而延长大豆青春期(Lu et al., 2017)。ELF3基因在长日照植物拟南芥中抑制开花, 而在短日照植物(如水稻和大豆)中促进开花, 这一有趣的现象是否与生物钟对日长的感知机制有关以及它们差异化调控的分子机制还有待进一步研究。 图2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-456/img_2.png<b>图2</b> 生物钟调控水稻抽穗期模式<br/>水稻中<i>OsELF3.1</i>维持生物钟周期并参与调控水稻抽穗期。在长日照条件下, <i>OsELF3.1</i>是开花负调节因子, 作用于<i>OsPRR73</i>和<i>Ghd7</i>的上游, 通过抑制<i>Hd3a</i>和<i>Ehd1</i>调控开花; 在短日照条件下, <i>OsELF3.1</i>是开花正调节因子, 通过感受蓝光信号激活<i>Ehd1</i>的表达进而促进开花。<i>OsPRR37</i>和<i>OsGI</i>也参与维持生物钟周期, 具体调控对光周期的敏感性, 控制水稻抽穗期。<br/><b>Figure 2</b> The proposed model for circadian clock regulating heading date in rice<br/>In rice, <i>OsELF3.1</i> is a key factor to maintain circadian rhythm and regulate heading date. In the long day condition, <i>OsELF3.1</i> acts as a negative factor by repressing<i> Hd3a </i>and <i>Ehd1 </i>to regulate heading date, genetically works at the upstream of<i> OsPRR73 </i>and <i>Ghd7</i>, while in short day condition, <i>OsELF3.1 </i>can work as a positive regulator by sensing the blue light signaling to promote <i>Ehd1 </i>expression. <i>OsPRR37</i> and <i>OsGI</i> are other essential components of rice circadian clock to regulate heading date in rice through photoperiod dependent pathway. Figure 2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-456/img_2.png<b>图2</b> 生物钟调控水稻抽穗期模式<br/>水稻中<i>OsELF3.1</i>维持生物钟周期并参与调控水稻抽穗期。在长日照条件下, <i>OsELF3.1</i>是开花负调节因子, 作用于<i>OsPRR73</i>和<i>Ghd7</i>的上游, 通过抑制<i>Hd3a</i>和<i>Ehd1</i>调控开花; 在短日照条件下, <i>OsELF3.1</i>是开花正调节因子, 通过感受蓝光信号激活<i>Ehd1</i>的表达进而促进开花。<i>OsPRR37</i>和<i>OsGI</i>也参与维持生物钟周期, 具体调控对光周期的敏感性, 控制水稻抽穗期。<br/><b>Figure 2</b> The proposed model for circadian clock regulating heading date in rice<br/>In rice, <i>OsELF3.1</i> is a key factor to maintain circadian rhythm and regulate heading date. In the long day condition, <i>OsELF3.1</i> acts as a negative factor by repressing<i> Hd3a </i>and <i>Ehd1 </i>to regulate heading date, genetically works at the upstream of<i> OsPRR73 </i>and <i>Ghd7</i>, while in short day condition, <i>OsELF3.1 </i>can work as a positive regulator by sensing the blue light signaling to promote <i>Ehd1 </i>expression. <i>OsPRR37</i> and <i>OsGI</i> are other essential components of rice circadian clock to regulate heading date in rice through photoperiod dependent pathway.
图2 生物钟调控水稻抽穗期模式 水稻中OsELF3.1维持生物钟周期并参与调控水稻抽穗期。在长日照条件下, OsELF3.1是开花负调节因子, 作用于OsPRR73和Ghd7的上游, 通过抑制Hd3a和Ehd1调控开花; 在短日照条件下, OsELF3.1是开花正调节因子, 通过感受蓝光信号激活Ehd1的表达进而促进开花。OsPRR37和OsGI也参与维持生物钟周期, 具体调控对光周期的敏感性, 控制水稻抽穗期。 Figure 2 The proposed model for circadian clock regulating heading date in rice In rice, OsELF3.1 is a key factor to maintain circadian rhythm and regulate heading date. In the long day condition, OsELF3.1 acts as a negative factor by repressing Hd3a and Ehd1 to regulate heading date, genetically works at the upstream of OsPRR73 and Ghd7, while in short day condition, OsELF3.1 can work as a positive regulator by sensing the blue light signaling to promote Ehd1 expression. OsPRR37 and OsGI are other essential components of rice circadian clock to regulate heading date in rice through photoperiod dependent pathway.
生物钟核心组分PRR基因家族在作物中也发挥重要功能。研究表明水稻OsPRR37可能参与调控Hd3a基因的表达, 从而调控水稻对光周期的敏感性(Koo et al., 2013), 但对于其调控的精确的分子机制尚不清楚。通过近等基因系定位到的DTH7 (Days To Heading 7), 其表达受生物钟调控, 呈现出昼夜节律, 在短日照和长日照条件下, DTH7表达量均在ZT8达到最高峰。进一步实验证明, DTH7位点即是OsPRR- 37基因的变异, 参与调控水稻开花时间和区域的适应性(Gao et al., 2014)。最新利用大数据的研究进一步证明OsPRR37和Ghd7是影响水稻开花时间、株高和每穗小穗数量等的主要数量性状位点(Liu et al., 2015)。GI是另一个在作物中发现的也具有保守功能的生物钟关键组分。Izawa等(2011)利用水稻osgi突变体在自然生长条件下, 连续多时间点取叶片进行转录组分析, 发现在检测到的27 201个基因中约75%的基因受到OsGI调控, 并且OsGI对于维持这些基因的生物节律是必需的。研究表明, OsGI在维持水稻生物钟功能中具有重要作用, 但是其具体作用机制仍然未知。最近研究表明, 在短日照条件下, OsPhyA突变后导致水稻抽穗期延迟, 主要是由于突变体中OsGI表达水平降低, 进而影响下游3个开花促进因子Hd1 (Heading Date 1)、OsMADS51和OsId1 (Oryza sativa Indeterminate 1)表达水平下调(Lee et al., 2016)。在大豆中, GI的同源基因有3个, 分别为GmGI1、GmGI2和GmGI3, 其中GmGI2的表达受光周期和生物钟调控, 可能参与调控大豆开花时间(Li et al., 2013), 但是其作用机制尚不明确。水稻生物钟系统除了参与调控抽穗期之外, 还有可能参与对非生物胁迫的响应。前期的研究表明, 水稻es1-1 (early sense-cence 1)突变体在正常和冷处理条件下其生物钟关键组分OsGI、OsLHY、OsPRR1、OsPRR37、OsPRR73和OsPRR95的表达模式与表达水平均有不同程度的改变, 表明水稻生物钟系统可能参与植物对冷胁迫的响应(徐江民等, 2016), 但是具体的分子机制仍需进一步研究。 目前, 人们对玉米(Zea mays)的生物钟系统知之甚少。Wang等(2011b)发现, ZmCCA1的表达呈现节律性, 在早晨达到峰值, 将其超表达转化拟南芥后也表现出长下胚轴和晚花的表型, 表明ZmCCA1可能是玉米生物钟系统的核心组分。ZmCCA1a和Zm- CCA1b还参与调控玉米的杂种优势, 能够较早激活体内早晨相关基因的表达, 促进其光合作用, 提高产量(Ko et al., 2016)。目前, 农作物关于生物钟组分功能的研究大多集中在开花时间调控方面, 对于其调控其它生长发育过程的研究则相对较少。近期研究表明, OsGI参与调控水稻对干旱和盐胁迫的响应。RNA-seq数据分析表明, OsSalT和一些干旱相关基因在osgi突变体中上调表达, 使水稻对于盐和干旱具有较强的耐受性(Li et al., 2016a)。以上结果表明, 生物钟基因在农作物中的功能具有多样性和保守性, 然而生物钟组分在调控水稻、玉米及大豆等生长发育和胁迫应答过程中的作用机制仍需进一步研究。解析农作物中生物钟系统如何调控生物和非生物胁迫应答以及生长发育过程, 将有望为作物分子育种和提高农作物产量提供时间生物学的理论依据及优质遗传资源。
AdamsS, ManfieldI, StockleyP, CarréIA (2015). Revised morning loops of the Arabidopsis circadian clock based on analyses of direct regulatory interactions.PLoS One 10, e0143943. [本文引用: 1]
[3]
Alabadı?D, OyamaT, YanovskyMJ, HarmonFG, MásP, KaySA (2001). Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock. Science 293, 880-883. [本文引用: 1]
[4]
AtamianHS, HarmerSL (2016). Circadian regulation of hormone signaling and plant physiology.Plant Mol Biol 91, 691-702. [本文引用: 1]
[5]
BhardwajV, MeierS, PetersenLN, IngleRA, RodenLC (2011). Defence responses of Arabidopsis thaliana to infection by Pseudomonas syringae are regulated by the circadian clock. PLoS One 6, e26968. [本文引用: 1]
[6]
BodenSA, WeissD, RossJJ, DaviesNW, TrevaskisB, ChandlerPM, SwainSM (2014). EARLY FLOWERING 3 regulates flowering in spring barley by mediating gibberellin production and FLOWERING LOCUS T expression. Plant Cell 26, 1557-1569. [本文引用: 1]
[7]
ChaJY, KimJ, KimTS, ZengQN, WangL, LeeSY, KimWY, SomersDE (2017). GIGANTEA is a co-chaperone which facilitates maturation of ZEITLUPE in the Arabidopsis circadian clock.Nat Commun 8, 3. [本文引用: 1]
[8]
ChewYH, HallidayKJ (2011). A stress-free walk from Arabidopsis to crops.Curr Opin Biotechnol 22, 281-286. [本文引用: 1]
[9]
ChowBY, SanchezSE, BretonG, Pruneda-PazJL, KroganNT, KaySA (2014). Transcriptional regulation of LUX by CBF1 mediates cold input to the circadian clock in Ara- bidopsis. Curr Biol 24, 1518-1524. [本文引用: 1]
[10]
CovingtonMF, MaloofJN, StraumeM, KaySA, HarmerSL (2008). Global transcriptome analysis reveals circadian regulation of key pathways in plant growth and development.Genome Biol 9, R130. [本文引用: 2]
[11]
FordB, DengWW, ClausenJ, OliverS, BodenS, HemmingM, TrevaskisB (2016). Barley (Hordeum vulgare) circadian clock genes can respond rapidly to temperature in an EARLY FLOWERING 3-dependent manner. J Exp Bot 67, 5517-5528. [本文引用: 1]
[12]
FowlerSG, CookD, ThomashowMF (2005). Low temperature induction of Arabidopsis CBF1, 2, and 3 is gated by the circadian clock. Plant Physiol 137, 961-968. [本文引用: 1]
[13]
GaoH, JinMN, ZhengXM, ChenJ, YuanDY, XinYY, WangMQ, HuangDY, ZhangZ, ZhouKN, ShengPK, MaJ, MaWW, DengHF, JiangL, LiuSJ, WangHY, WuCY, YuanLP, WanJM (2014). Days to heading 7, a major quantitative locus determining photoperiod sensitivity and regional adaptation in rice. Proc Natl Acad Sci USA 111, 16337-16342. [本文引用: 1]
[14]
GendronJM, Pruneda-PazJL, DohertyCJ, GrossAM, KangSE, KaySA (2012). Arabidopsis circadian clock protein, TOC1, is a DNA-binding transcription factor.Proc Natl Acad Sci USA 109, 3167-3172. [本文引用: 1]
[15]
GreenhamK, McClungCR (2015). Integrating circadian dynamics with physiological processes in plants.Nat Rev Genet 16, 598-610. [本文引用: 1]
[16]
GrundyJ, StokerC, CarréIA (2015). Circadian regulation of abiotic stress tolerance in plants.Front Plant Sci 6, 648. [本文引用: 1]
[17]
HarmerSL (2009). The circadian system in higher plants.Annu Rev Plant Biol 60, 357-377. [本文引用: 1]
[18]
HayamaR, Sarid-KrebsL, RichterR, FernándezV, JangS, CouplandG (2017). PSEUDO RESPONSE REGULATORs stabilize CONSTANS protein to promote flowering in response to day length.EMBO J 36, 904-918. [本文引用: 1]
[19]
HuangW, Peréz-GarcíaP, PokhilkoA, MillarAJ, AntoshechkinI, RiechmannJL, MasP (2012). Mapping the core of the Arabidopsis circadian clock defines the network structure of the oscillator.Science 336, 75-79. [本文引用: 1]
[20]
IzawaT, MiharaM, SuzukiY, GuptaM, ItohH, NaganoAJ, MotoyamaR, SawadaY, YanoM, HiraiMY, MakinoA, NagamuraY (2011). Os-GIGANTEA confers robust diurnal rhythms on the global transcriptome of rice in the field. Plant Cell 23, 1741-1755. [本文引用: 1]
[21]
KamiokaM, TakaoS, SuzukiT, TakiK, HigashiyamaT, KinoshitaT, NakamichiN (2016). Direct repression of evening genes by CIRCADIAN CLOCK-ASSOCIATED1 in the Arabidopsis circadian clock. Plant Cell 28, 696-711. [本文引用: 1]
[22]
KeilyJ, MacGregorDR, SmithRW, MillarAJ, HallidayKJ, PenfieldS (2013). Model selection reveals control of cold signaling by evening-phased components of the plant circadian clock.Plant J 76, 247-257. [本文引用: 1]
[23]
KimWY, AliZ, ParkHJ, ParkSJ, ChaJY, Perez- HormaecheJ, QuinteroFJ, ShinG, KimMR, QiangZ, NingL, ParkHC, LeeSY, BressanRA, PardoJM, BohnertHJ, YunDJ (2013). Release of SOS2 kinase from sequestration with GIGANTEA determines salt tole- rance in Arabidopsis.Nat Commun 4, 1352. [本文引用: 1]
[24]
KoDK, RohozinskiD, SongQX, TaylorSH, JuengerTE, HarmonFG, ChenZJ (2016). Temporal shift of circa- dian-mediated gene expression and carbon fixation contributes to biomass heterosis in maize hybrids.PLoS Genet 12, e1006197. [本文引用: 1]
[25]
KobayashiY, KayaH, GotoK, IwabuchiM, ArakiT (1999). A pair of related genes with antagonistic roles in mediating flowering signals.Science 286, 1960-1962. [本文引用: 1]
[26]
KolmosE, ChowBY, Pruneda-PazJL, KaySA (2014). HsfB2b-mediated repression of PRR7 directs abiotic stress responses of the circadian clock. Proc Natl Acad Sci USA 111, 16172-16177. [本文引用: 1]
[27]
KooBH, YooSC, ParkJW, KwonCT, LeeBD, AnG, ZhangZY, LiJJ, LiZC, PaekNC (2013). Natural variation in OsPRR37 regulates heading date and contributes to rice cultivation at a wide range of latitudes. Mol Plant 6, 1877-1888. [本文引用: 1]
[28]
KorneliC, DanismanS, StaigerD (2014). Differential control of pre-invasive and post-invasive antibacterial defense by the Arabidopsis circadian clock.Plant Cell Physiol 55, 1613-1622. [本文引用: 3]
[29]
KrepsJA, WuYJ, ChangHS, ZhuT, WangX, HarperJF (2002). Transcriptome changes for Arabidopsis in response to salt, osmotic, and cold stress.Plant Physiol 130, 2129-2141. [本文引用: 1]
[30]
LeeYS, YiJ, AnG (2016). OsPhyA modulates rice flowering time mainly through OsGI under short days and Ghd7 under long days in the absence of phytochrome B. Plant Mol Biol 91, 413-427. [本文引用: 1]
[31]
LiF, ZhangXM, HuRB, WuFQ, MaJH, MengY, FuYF (2013). Identification and molecular characterization of FKF1 and GI homologous genes in soybean. PLoS One 8, e79036. [本文引用: 1]
[32]
LiS, YueWH, WangM, QiuWM, ZhouL, ShouHX (2016a). Mutation of OsGIGANTEA leads to enhanced tolerance to polyethylene glycol-generated osmotic stress in rice. Front Plant Sci 7, 465. [本文引用: 1]
[33]
LiX, MaDB, LuSX, HuXY, HuangRF, LiangT, XuTD, TobinEM, LiuHT (2016b). Blue light- and low temperature-regulated COR27 and COR28 play roles in the Ara- bidopsis circadian clock.Plant Cell 28, 2755-2769. [本文引用: 1]
[34]
LiuC, SongGY, ZhouYH, QuXF, GuoZB, LiuZW, JiangDM, YangDC (2015). OsPRR37 and Ghd7 are the major genes for general combining ability of DTH, PH and SPP in rice. Sci Rep 5, 12803. [本文引用: 1]
[35]
LiuQ, KasugaM, SakumaY, AbeH, MiuraS, Yamaguchi-ShinozakiK, ShinozakiK (1998). Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis.Plant Cell 10, 1391-1406. [本文引用: 1]
[36]
LiuT, CarlssonJ, TakeuchiT, NewtonL, FarréEM (2013). Direct regulation of abiotic responses by the Arabidopsis circadian clock component PRR7.Plant J 76, 101-114. [本文引用: 1]
[37]
LuSJ, ZhaoXH, HuYL, LiuSL, NanHY, LiXM, FangC, CaoD, ShiXY, KongLP, SuT, ZhangFG, LiSC, WangZ, YuanXH, CoberER, WellerJL, LiuBH, HouXL, TianZX, KongFJ (2017). Natural variation at the soybean J locus improves adaptation to the tropics and enhances yield. Nat Genet 49, 773-779. [本文引用: 2]
[38]
MatsubaraK, Ogiso-TanakaE, HoriK, EbanaK, AndoT, YanoM (2012). Natural variation in Hd17, a homolog of Arabidopsis ELF3 that is involved in rice photoperiodic flowering. Plant Cell Physiol 53, 709-716. [本文引用: 1]
[39]
MillarAJ, CarreIA, StrayerCA, ChuaNH, KaySA (1995). Circadian clock mutants in Arabidopsis identified by luci- ferase imaging.Science 267, 1161-1163. [本文引用: 1]
[40]
NagelDH, Pruneda-PazJL, KaySA (2014). FBH1 affects warm temperature responses in the Arabidopsis circadian clock.Proc Natl Acad Sci USA 111, 14595-14600. [本文引用: 1]
[41]
NakamichiN, KibaT, HenriquesR, MizunoT, ChuaNH, SakakibaraH (2010). PSEUDO-RESPONSE REGULATORS 9, 7, and 5 are transcriptional repressors in the Arabidopsis circadian clock.Plant Cell 22, 594-605. [本文引用: 2]
SeoPJ, MasP (2014). Multiple layers of posttranslational regulation refine circadian clock activity in Arabidopsis.Plant Cell 26, 79-87. [本文引用: 1]
[55]
ShinJ, HeidrichK, Sanchez-VillarrealA, ParkerJE, DavisSJ (2012). TIME FOR COFFEE represses accumulation of the MYC2 transcription factor to provide time-of-day regulation of jasmonate signaling in Arabidopsis.Plant Cell 24, 2470-2482. [本文引用: 2]
[56]
SongSS, HuangH, GaoH, WangJJ, WuDW, LiuXL, YangSH, ZhaiQZ, LiCY, QiTC, XieDX (2014). Interaction between MYC2 and ETHYLENE INSENSITIVE3 mo- dulates antagonism between jasmonate and ethylene signaling in Arabidopsis.Plant Cell 26, 263-279. [本文引用: 1]
[57]
SongYH, ShimJS, Kinmonth-SchultzHA, ImaizumiT (2015). Photoperiodic flowering: time measurement mech- anisms in leaves.Annu Rev Plant Biol 66, 441-464. [本文引用: 1]
[58]
SoyJ, LeivarP, González-SchainN, MartínG, DiazC, SentandreuM, Al-SadyB, QuailPH, MonteE (2016). Molecular convergence of clock and photosensory pathways through PIF3-TOC1 interaction and co-occupancy of target promoters.Proc Natl Acad Sci USA 113, 4870-4875. [本文引用: 2]
[59]
TripathiP, CarvalloM, HamiltonEE, PreussS, KaySA (2017). Arabidopsis B-BOX32 interacts with CONSTANS- LIKE3 to regulate flowering.Proc Natl Acad Sci USA 114, 172-177. [本文引用: 2]
[60]
WangGY, ZhangC, BattleS, LuH (2014). The phosphate transporter PHT4;1 is a salicylic acid regulator likely controlled by the circadian clock protein CCA1.Front Plant Sci 5, 701. [本文引用: 1]
[61]
WangL, FujiwaraS, SomersDE (2010). PRR5 regulates phosphorylation, nuclear import and subnuclear localization of TOC1 in the Arabidopsis circadian clock.EMBO J 29, 1903-1915. [本文引用: 1]
[62]
WangL, KimJ, SomersDE (2013). Transcriptional corepressor TOPLESS complexes with pseudoresponse regulator proteins and histone deacetylases to regulate circadian transcription.Proc Natl Acad Sci USA 110, 761-766. [本文引用: 1]
WangW, BarnabyJY, TadaY, LiHr, TörM, CaldelariD, LeeDU, FuXD, DongXN (2011a). Timing of plant immune responses by a central circadian regulator.Nature 470, 110-114. [本文引用: 1]
[65]
WangXT, WuLJ, ZhangSF, WuLC, KuLX, WeiXM, XieLL, ChenYH (2011b). Robust expression and association of ZmCCA1 with circadian rhythms in maize. Plant Cell Rep 30, 1261-1272. [本文引用: 1]
[66]
WangXX, WuFM, XieQG, WangHM, WangY, YueYL, GahuraO, MaSS, LiuL, CaoY, JiaoYL, PutaF, McClungCR, XuXD, MaLG (2012). SKIP is a component of the spliceosome linking alternative splicing and the circadian clock in Arabidopsis.Plant Cell 24, 3278-3295. [本文引用: 1]
[67]
WangZY, TobinEM (1998). Constitutive expression of the CIRCADIAN CLOCK ASSOCIATED 1 (CCA1) gene disrupts circadian rhythms and suppresses its own expression. Cell 93, 1207-1217. [本文引用: 1]
YangY, PengQ, ChenGX, LiXH, WuCY (2013). OsELF3 is involved in circadian clock regulation for promoting flow- ering under long-day conditions in rice.Mol Plant 6, 202-215. [本文引用: 1]
[70]
YueYL, LiuNX, JiangBJ, LiM, WangHJ, JiangZ, PanHT, XiaQJ, MaQB, HanTF, NianH (2017). A single nucleotide deletion in J encoding GmELF3 confers long juvenility and is associated with adaption of tropic soybean. Mol Plant 10, 656-658. [本文引用: 1]
[71]
ZhangC, XieQG, AndersonRG, NgG, SeitzNC, PetersonT, McClungCR, McDowellJM, KongDD, KwakJM, LuH (2013). Crosstalk between the circadian clock and innate immunity in Arabidopsis.PLoS Pathog 9, e1003370. [本文引用: 2]
ZhaoJM, HuangX, OuyangXH, ChenWL, DuAP, ZhuL, WangSG, DengXW, LiSG (2012). OsELF3-1, an ortholog of Arabidopsis EARLY FLOWERING 3, regulates rice circadian rhythm and photoperiodic flowering. PLoS One 7, e43705. [本文引用: 1]
[74]
ZhengXY, ZhouM, YooH, Pruneda-PazJL, SpiveyNW, KaySA, DongXN (2015). Spatial and temporal regulation of biosynthesis of the plant immune signal salicylic acid.Proc Natl Acad Sci USA 112, 9166-9173. [本文引用: 1]
[75]
ZhuJY, OhE, WangTN, WangZY (2016). TOC1-PIF4 interaction mediates the circadian gating of thermores- ponsive growth in Arabidopsis.Nat Commun 7, 13692. [本文引用: 1]
水稻ES1参与生物钟基因表达调控以及逆境胁迫响应 1 2016
... 生物钟核心组分PRR基因家族在作物中也发挥重要功能.研究表明水稻OsPRR37可能参与调控Hd3a基因的表达, 从而调控水稻对光周期的敏感性(Koo et al., 2013), 但对于其调控的精确的分子机制尚不清楚.通过近等基因系定位到的DTH7 (Days To Heading 7), 其表达受生物钟调控, 呈现出昼夜节律, 在短日照和长日照条件下, DTH7表达量均在ZT8达到最高峰.进一步实验证明, DTH7位点即是OsPRR- 37基因的变异, 参与调控水稻开花时间和区域的适应性(Gao et al., 2014).最新利用大数据的研究进一步证明OsPRR37和Ghd7是影响水稻开花时间、株高和每穗小穗数量等的主要数量性状位点(Liu et al., 2015).GI是另一个在作物中发现的也具有保守功能的生物钟关键组分.Izawa等(2011)利用水稻osgi突变体在自然生长条件下, 连续多时间点取叶片进行转录组分析, 发现在检测到的27 201个基因中约75%的基因受到OsGI调控, 并且OsGI对于维持这些基因的生物节律是必需的.研究表明, OsGI在维持水稻生物钟功能中具有重要作用, 但是其具体作用机制仍然未知.最近研究表明, 在短日照条件下, OsPhyA突变后导致水稻抽穗期延迟, 主要是由于突变体中OsGI表达水平降低, 进而影响下游3个开花促进因子Hd1 (Heading Date 1)、OsMADS51和OsId1 (Oryza sativa Indeterminate 1)表达水平下调(Lee et al., 2016).在大豆中, GI的同源基因有3个, 分别为GmGI1、GmGI2和GmGI3, 其中GmGI2的表达受光周期和生物钟调控, 可能参与调控大豆开花时间(Li et al., 2013), 但是其作用机制尚不明确.水稻生物钟系统除了参与调控抽穗期之外, 还有可能参与对非生物胁迫的响应.前期的研究表明, 水稻es1-1 (early sense-cence 1)突变体在正常和冷处理条件下其生物钟关键组分OsGI、OsLHY、OsPRR1、OsPRR37、OsPRR73和OsPRR95的表达模式与表达水平均有不同程度的改变, 表明水稻生物钟系统可能参与植物对冷胁迫的响应(徐江民等, 2016), 但是具体的分子机制仍需进一步研究. ...
1 2015
... 拟南芥(Arabidopsis thaliana)作为模式植物被广泛用于植物分子生物学和遗传学研究, 因此其生物钟分子机制也研究得较为透彻.1995年, Millar等(1995)首次采用荧光素酶作为报告基因鉴定到第1个生物钟突变体toc1 (timing of cab expression 1).随后, Wang和Tobin (1998)利用反向遗传学方法鉴定到第1个生物钟核心振荡器的关键成员CCA1 (CIRCADIAN CLOCK ASSOCIATED 1), 该基因编码1个MYB类转录因子.超表达CCA1会使生物钟报告基因丧失昼夜节律性表达模式, 而且极度晚花, 同时也抑制内源CCA1和LHY (LATE ELONGATED HYPOCOTYL)的表达, 且该基因自身的转录水平还受到红光的瞬时诱导.基于基因表达水平和启动子活性分析, AlabadÍ等(2001)提出了拟南芥生物钟第1个分子反馈环路, 即CCA1/LHY抑制TOC1的表达, 而TOC1则促进CCA1/LHY的表达, 由此形成了生物钟核心振荡器的转录-翻译反馈环.随着技术手段的进步和计算生物学的发展, 目前认为生物钟核心振荡器由3个紧密关联的转录-翻译反馈环组成(图1).其中, 中心反馈环由LHY和CCA1以及TOC1组成.CCA1和LHY在早晨表达, 二者存在部分功能冗余, 它们转录并翻译成熟后会相互作用进而抑制TOC1的表达.在晚上, CCA1和LHY的表达水平降低, TOC1的表达则达到峰值, 反过来抑制CCA1和LHY的表达, 进而使它们处于此起彼伏的状态(Gendron et al., 2012; Huang et al., 2012).CCA1/LHY也和PRR (PSEUDO RESPONSE REGULATOR)基因家族的PRR9/PRR7组成早反馈环.PRRs成员中PRR9、PRR7及PRR5从早到晚每隔2-3小时依次出现表达峰值, 它们会通过与TOPL- ESS基因家族的成员互作招募组蛋白去乙酰化酶HDA6/19 (HISTONE DEACETYLASE 6/19)形成转录抑制复合体, 从而在转录水平上抑制CCA1和LHY的表达(Nakamichi et al., 2010; Wang et al., 2013).反之, CCA1/LHY也抑制PRR9/7的表达, 从而形成早反馈环.晚反馈环包含MYB类转录因子LUX ARRHYTHMO (又命名为PHYTO-CLOCK1)以及2个核蛋白ELF3 (EARLY FLOWERING 3)和ELF4组成的EC复合体(evening complex) (Nusinow et al., 2011)和TOC1.EC复合体与CCA1/LHY相互调节, 间接促进CCA1/LHY的表达, 反过来CCA1和LHY会直接结合在EC复合体启动子上抑制其转录(Adams et al., 2015).早反馈环、中心反馈环和晚反馈环相互连锁形成生物钟核心振荡器的抑制振荡子(repressilator)基本架构. ...
1 2001
... 拟南芥(Arabidopsis thaliana)作为模式植物被广泛用于植物分子生物学和遗传学研究, 因此其生物钟分子机制也研究得较为透彻.1995年, Millar等(1995)首次采用荧光素酶作为报告基因鉴定到第1个生物钟突变体toc1 (timing of cab expression 1).随后, Wang和Tobin (1998)利用反向遗传学方法鉴定到第1个生物钟核心振荡器的关键成员CCA1 (CIRCADIAN CLOCK ASSOCIATED 1), 该基因编码1个MYB类转录因子.超表达CCA1会使生物钟报告基因丧失昼夜节律性表达模式, 而且极度晚花, 同时也抑制内源CCA1和LHY (LATE ELONGATED HYPOCOTYL)的表达, 且该基因自身的转录水平还受到红光的瞬时诱导.基于基因表达水平和启动子活性分析, AlabadÍ等(2001)提出了拟南芥生物钟第1个分子反馈环路, 即CCA1/LHY抑制TOC1的表达, 而TOC1则促进CCA1/LHY的表达, 由此形成了生物钟核心振荡器的转录-翻译反馈环.随着技术手段的进步和计算生物学的发展, 目前认为生物钟核心振荡器由3个紧密关联的转录-翻译反馈环组成(图1).其中, 中心反馈环由LHY和CCA1以及TOC1组成.CCA1和LHY在早晨表达, 二者存在部分功能冗余, 它们转录并翻译成熟后会相互作用进而抑制TOC1的表达.在晚上, CCA1和LHY的表达水平降低, TOC1的表达则达到峰值, 反过来抑制CCA1和LHY的表达, 进而使它们处于此起彼伏的状态(Gendron et al., 2012; Huang et al., 2012).CCA1/LHY也和PRR (PSEUDO RESPONSE REGULATOR)基因家族的PRR9/PRR7组成早反馈环.PRRs成员中PRR9、PRR7及PRR5从早到晚每隔2-3小时依次出现表达峰值, 它们会通过与TOPL- ESS基因家族的成员互作招募组蛋白去乙酰化酶HDA6/19 (HISTONE DEACETYLASE 6/19)形成转录抑制复合体, 从而在转录水平上抑制CCA1和LHY的表达(Nakamichi et al., 2010; Wang et al., 2013).反之, CCA1/LHY也抑制PRR9/7的表达, 从而形成早反馈环.晚反馈环包含MYB类转录因子LUX ARRHYTHMO (又命名为PHYTO-CLOCK1)以及2个核蛋白ELF3 (EARLY FLOWERING 3)和ELF4组成的EC复合体(evening complex) (Nusinow et al., 2011)和TOC1.EC复合体与CCA1/LHY相互调节, 间接促进CCA1/LHY的表达, 反过来CCA1和LHY会直接结合在EC复合体启动子上抑制其转录(Adams et al., 2015).早反馈环、中心反馈环和晚反馈环相互连锁形成生物钟核心振荡器的抑制振荡子(repressilator)基本架构. ...
1 2016
... 除了在模式植物拟南芥中发现生物钟调控生长发育过程以及应对外界环境变化和胁迫应答方面具有不可忽视的作用外, 近年来, 农作物的生物钟研究逐步展开.水稻是主要粮食作物, 生物钟如何调控水稻的生长发育(尤其是抽穗期)和非生物胁迫应答已越来越受到科学家的关注.抽穗期是水稻的重要农艺性状之一, 抽穗期的长短(从萌发到始穗)直接决定水稻品种生育期的长短, 而适宜的生育期在维持水稻高产稳产中扮演着极为关键的角色, 也在一定程度上反映水稻对种植区域和季节的适应性.研究表明, 生物钟在调控水稻抽穗期方面具有重要作用.图位克隆分析表明, OsELF3 (与拟南芥生物钟核心组分同源的ELF3)的核苷酸变异导致Nipponbare和Koshihikari品种间抽穗期的差异(Matsubara et al., 2012).进一步研究表明, 水稻中存在2个与拟南芥ELF3同源的基因Os- ELF3.1 (LOC_Os06g05060)和OsELF3.2 (LOC_Os- 01g38530) (Zhao et al., 2012).在长日照条件下, OsELF3.1是水稻抽穗期正调节因子, 作用于OsGI (Os-GIGANTEA)和Ghd7 (Grain number, Plant height and heading date 7)的上游, 负调控它们的表达, 因此oself3.1突变体表现出晚抽穗的表型; 但是OsELF3.2的突变并不影响水稻抽穗期.OsELF3.1突变后影响生物钟其它组分OsLHY、OsPRR1、Os- PRR37和OsPRR73的表达, 表明OsELF3.1在维持生物钟周期和调控抽穗期方面具有重要作用(Yang et al., 2013) (图2).有趣的是, 在植物叶片衰老方面, 拟南芥ELF3通过抑制PIF4和PIF5的表达负调控叶片衰老(Sakuraba et al., 2014); 但是水稻OsELF3.1的功能恰恰相反, 它是水稻叶片衰老的正调节因子, 主要调控NAC (N-ACETYL CYSTEINE)和WRKY基因家族成员的表达(Sakuraba et al., 2016).在拟南芥中超表达OsELF3.1则会表现出与ELF3超表达相同的表型, 即延缓叶片衰老、晚花和短周期, 表明ELF3在水稻和拟南芥中功能保守, 可能由于在不同物种中参与调控的下游靶基因完全不同, 导致其在不同植物中的功能差异(Atamian and Harmer, 2016).水稻同源基因OsELF3.2则与E3连接酶APIP6 (AVRPIZT INTERACT PROTEIN 6)互作, 负调控水稻对稻瘟病的免疫(Ning et al., 2015).在春大麦中ELF3通过抑制FT1的表达和产生有活性的GAs (gibberellic acid)来调控其对光周期的敏感性, 控制春大麦由营养生长阶段向生殖生长阶段的转变(Boden et al., 2014).近期研究表明, 大麦中ELF3通过生物钟系统介导植物对温度信号的响应.受高温刺激后, CCA1和PRR73的表达水平明显降低, 而GI和PRR59的表达水平明显升高, 但是这种变化在elf3突变体中消失, 表明GI和PRR对温度响应依赖于ELF3的功能(Ford et al., 2016), 但ELF3调控大麦对温度响应的分子机制还不清楚.在另一重要作物大豆(Glycine max)中, ELF3基因突变导致其成熟时间延长, 即“长青春期”, 这能够提高大豆产量, 增强大豆在低纬度地区的区域适应性(Lu et al., 2017; Yue et al., 2017).进一步研究表明, GmELF3结合在E1启动子上抑制其表达, 进而使下游FT2a和FT5a的表达水平降低而延长大豆青春期(Lu et al., 2017).ELF3基因在长日照植物拟南芥中抑制开花, 而在短日照植物(如水稻和大豆)中促进开花, 这一有趣的现象是否与生物钟对日长的感知机制有关以及它们差异化调控的分子机制还有待进一步研究. ...
1 2011
... 水杨酸的生物合成和积累受到生物钟核心组分CHE (CCA1HIKING EXPEDITION)的调控(Zheng et al., 2015).SA介导的防御反应中磷酸转移酶PHT4;1 (PHOSPHATE TRANSPORTER 4;1)的表达也受到CCA1的直接调控(Wang et al., 2014).介导植物对真菌免疫响应的RPP4 (RECOGNITION FOR PERO- NOSPORA PARASITICA 4)基因的启动子上存在EE (evening element)元件, 该元件可被CCA1蛋白识别并结合, 从而调控RPP4基因的转录活性.CCA1对RPP4基因的转录调控使植物即使在没有病菌感染的条件下, 在侵染性真菌的孢子开始释放的黎明时分开启免疫系统, 增强植物对病原菌的免疫作用(Wang et al., 2011a).因此, 当CCA1或者LHY超表达使生物钟节律紊乱或者LUX基因的功能缺失都会导致植物对病原菌更敏感(Bhardwaj et al., 2011; Zhang et al., 2013; Korneli et al., 2014).虽然植物叶片表面的气孔在早晨会完全张开, 使病原菌容易入侵, 但植物会激活相关防御机制, 使其在早晨的抗病能力比夜晚更强(Zhang et al., 2013; Korneli et al., 2014).TIC也会通过生物钟系统调控气孔开度, 进而调控植物对病原菌的敏感性(Korneli et al., 2014).由此可见, 生物钟调节植物对生物胁迫应答存在多个节点的复杂调控, 而全面揭示这一复杂网络仍需开展更系统的研究. ...
1 2014
... 除了在模式植物拟南芥中发现生物钟调控生长发育过程以及应对外界环境变化和胁迫应答方面具有不可忽视的作用外, 近年来, 农作物的生物钟研究逐步展开.水稻是主要粮食作物, 生物钟如何调控水稻的生长发育(尤其是抽穗期)和非生物胁迫应答已越来越受到科学家的关注.抽穗期是水稻的重要农艺性状之一, 抽穗期的长短(从萌发到始穗)直接决定水稻品种生育期的长短, 而适宜的生育期在维持水稻高产稳产中扮演着极为关键的角色, 也在一定程度上反映水稻对种植区域和季节的适应性.研究表明, 生物钟在调控水稻抽穗期方面具有重要作用.图位克隆分析表明, OsELF3 (与拟南芥生物钟核心组分同源的ELF3)的核苷酸变异导致Nipponbare和Koshihikari品种间抽穗期的差异(Matsubara et al., 2012).进一步研究表明, 水稻中存在2个与拟南芥ELF3同源的基因Os- ELF3.1 (LOC_Os06g05060)和OsELF3.2 (LOC_Os- 01g38530) (Zhao et al., 2012).在长日照条件下, OsELF3.1是水稻抽穗期正调节因子, 作用于OsGI (Os-GIGANTEA)和Ghd7 (Grain number, Plant height and heading date 7)的上游, 负调控它们的表达, 因此oself3.1突变体表现出晚抽穗的表型; 但是OsELF3.2的突变并不影响水稻抽穗期.OsELF3.1突变后影响生物钟其它组分OsLHY、OsPRR1、Os- PRR37和OsPRR73的表达, 表明OsELF3.1在维持生物钟周期和调控抽穗期方面具有重要作用(Yang et al., 2013) (图2).有趣的是, 在植物叶片衰老方面, 拟南芥ELF3通过抑制PIF4和PIF5的表达负调控叶片衰老(Sakuraba et al., 2014); 但是水稻OsELF3.1的功能恰恰相反, 它是水稻叶片衰老的正调节因子, 主要调控NAC (N-ACETYL CYSTEINE)和WRKY基因家族成员的表达(Sakuraba et al., 2016).在拟南芥中超表达OsELF3.1则会表现出与ELF3超表达相同的表型, 即延缓叶片衰老、晚花和短周期, 表明ELF3在水稻和拟南芥中功能保守, 可能由于在不同物种中参与调控的下游靶基因完全不同, 导致其在不同植物中的功能差异(Atamian and Harmer, 2016).水稻同源基因OsELF3.2则与E3连接酶APIP6 (AVRPIZT INTERACT PROTEIN 6)互作, 负调控水稻对稻瘟病的免疫(Ning et al., 2015).在春大麦中ELF3通过抑制FT1的表达和产生有活性的GAs (gibberellic acid)来调控其对光周期的敏感性, 控制春大麦由营养生长阶段向生殖生长阶段的转变(Boden et al., 2014).近期研究表明, 大麦中ELF3通过生物钟系统介导植物对温度信号的响应.受高温刺激后, CCA1和PRR73的表达水平明显降低, 而GI和PRR59的表达水平明显升高, 但是这种变化在elf3突变体中消失, 表明GI和PRR对温度响应依赖于ELF3的功能(Ford et al., 2016), 但ELF3调控大麦对温度响应的分子机制还不清楚.在另一重要作物大豆(Glycine max)中, ELF3基因突变导致其成熟时间延长, 即“长青春期”, 这能够提高大豆产量, 增强大豆在低纬度地区的区域适应性(Lu et al., 2017; Yue et al., 2017).进一步研究表明, GmELF3结合在E1启动子上抑制其表达, 进而使下游FT2a和FT5a的表达水平降低而延长大豆青春期(Lu et al., 2017).ELF3基因在长日照植物拟南芥中抑制开花, 而在短日照植物(如水稻和大豆)中促进开花, 这一有趣的现象是否与生物钟对日长的感知机制有关以及它们差异化调控的分子机制还有待进一步研究. ...
1 2017
... 生物钟核心振荡器除了存在转录-翻译反馈环的相互促进和相互抑制调控之外, 还存在复杂的转录后调控.例如, F-box类蛋白ZTL (ZEITLUPE)参与PRR5和TOC1蛋白的降解过程.PRR5可以通过PR结构域与TOC1蛋白互作, 从而促进TOC1蛋白的磷酸化及其向细胞核内的迁移过程(Wang et al., 2010).GI (GIGANTEA)可以作为分子伴侣蛋白与HSP90 (HE- AT SHOCK PROTEIN 90)以及ZTL形成三元复合体, 进而促进ZTL成为有活性的形式, 发挥其转录后调控的功能(Cha et al., 2017).除此之外, mRNA的成熟与加工过程也参与生物钟的周期调控, 如功能保守的含SKIP (SNW/SKI-INTERACTING PROTEIN)结构域的SKIP蛋白, 参与PRR7和PRR9 mRNA的可变剪切和成熟.当SKIP1突变以后, 植物表现出温度敏感而延长生物钟周期, 并且影响生物钟系统对光信号的敏感性, 进而影响光信号输入(Wang et al., 2012).另外, 生物钟核心振荡器的关键组成因子也受到如磷酸化和糖基化等翻译后修饰以及转录水平的表观遗传修饰(Seo and Mas, 2014; Nolte and Staiger, 2015; Romanowski and Yanovsky, 2015), 进而构成复杂的信号网络.目前, 关于蛋白质翻译后修饰调控植物生物钟周期的分子机制研究较少, 人们对于关键因子磷酸化和糖基化的生物学意义还不是非常清楚. ...
1 2011
... 生物钟广泛调节植物对非生物胁迫的耐受性, 是协调植物生长发育与胁迫响应的重要调节者和平衡者.在冷胁迫信号应答中, 植物通过CBF/DREB1 (CREPEAT BINDING FACTOR/DEHYDRATION-RES- POSIVE-ELEMENT-BINDING)来激活COR基因的表达, 进而调控植物对冷胁迫的响应, 且CBF基因家族成员自身的表达也呈现明显的昼夜节律性(Liu et al., 1998; Fowler et al., 2005).许多生物钟基因的突变体均表现出对冷胁迫适应性改变的表型, 如生物钟核心组分PRR5/7/9突变后, 可以间接增强CBF的表达水平使其表达丧失昼夜节律性, 从而导致植物具有更强的耐冷性(Nakamichi et al., 2009).PRR基因家族的另一个成员TOC1的突变也会使CBF3表达水平上调, 增强植物对冷胁迫的耐受性(Keily et al., 2013), 说明PRR基因家族在植物冷胁迫的适应方面可能发挥重要作用.在农作物水稻(Oryza sativa)、小麦(Triticum aestivum)和大麦(Hordeum vulgare)中, CBF也参与调控对冷和干旱的响应, 进而影响农作物生长(Chew and Halliday, 2011).以上结果进一步说明, 生物钟系统通过调节CBF基因家族表达的昼夜节律性来协调植物对非生物胁迫响应和自身发育所需能量分配.另一方面, 冷胁迫也可以反馈调节生物钟系统, 如CBF1结合在LUX的启动子上调控其表达(Chow et al., 2014); 低温可诱导CCA1及其它生物钟基因的可变剪切并形成不同的转录本, 从而影响生物钟的功能(Grundy et al., 2015).核心振荡器组分还受到高温的调控, 高温主要影响生物钟组分基因的可变剪切或通过调控热激响应因子HsfB2b (HEAT SHOCK FACTOR B2B)和FBH1 (FLOW-ERING BA- SIC HELIX-LOOP-HELIX 1)分别结合在PRR7及CC- A1的启动子上来调控其表达水平(Kolmos et al., 2014; Nagel et al., 2014).在植物干旱和盐胁迫应答方面, 生物钟系统也发挥着重要功能.研究表明, 许多与盐、脱水、渗透压调控和响应相关基因的表达均受生物钟调控(Kreps et al., 2002; Covington et al., 2008).在拟南芥中, 参与调控盐胁迫的基因如SOS1 (SALT OVERLY SENSITIVE 1)等的表达呈现出较强的昼夜节律性(http://diurnal.mocklerlab.org/), 表明生物钟可能调控植物对盐胁迫的适应.另外, 生物钟组分GI参与调控植物在高盐环境下的Na+/H+质子泵通道的打开, 进而将Na+排出体外, 减少Na+对植物的毒害(Kim et al., 2013).也有研究表明, PRR7可能参与调控植物响应盐胁迫(Liu et al., 2013).但是关于生物钟系统如何调控植物对盐胁迫的响应及其相关分子机制仍需更系统地挖掘和研究, 并将有望为作物耐盐育种提供理论依据和相关遗传资源. ...
1 2014
... 生物钟广泛调节植物对非生物胁迫的耐受性, 是协调植物生长发育与胁迫响应的重要调节者和平衡者.在冷胁迫信号应答中, 植物通过CBF/DREB1 (CREPEAT BINDING FACTOR/DEHYDRATION-RES- POSIVE-ELEMENT-BINDING)来激活COR基因的表达, 进而调控植物对冷胁迫的响应, 且CBF基因家族成员自身的表达也呈现明显的昼夜节律性(Liu et al., 1998; Fowler et al., 2005).许多生物钟基因的突变体均表现出对冷胁迫适应性改变的表型, 如生物钟核心组分PRR5/7/9突变后, 可以间接增强CBF的表达水平使其表达丧失昼夜节律性, 从而导致植物具有更强的耐冷性(Nakamichi et al., 2009).PRR基因家族的另一个成员TOC1的突变也会使CBF3表达水平上调, 增强植物对冷胁迫的耐受性(Keily et al., 2013), 说明PRR基因家族在植物冷胁迫的适应方面可能发挥重要作用.在农作物水稻(Oryza sativa)、小麦(Triticum aestivum)和大麦(Hordeum vulgare)中, CBF也参与调控对冷和干旱的响应, 进而影响农作物生长(Chew and Halliday, 2011).以上结果进一步说明, 生物钟系统通过调节CBF基因家族表达的昼夜节律性来协调植物对非生物胁迫响应和自身发育所需能量分配.另一方面, 冷胁迫也可以反馈调节生物钟系统, 如CBF1结合在LUX的启动子上调控其表达(Chow et al., 2014); 低温可诱导CCA1及其它生物钟基因的可变剪切并形成不同的转录本, 从而影响生物钟的功能(Grundy et al., 2015).核心振荡器组分还受到高温的调控, 高温主要影响生物钟组分基因的可变剪切或通过调控热激响应因子HsfB2b (HEAT SHOCK FACTOR B2B)和FBH1 (FLOW-ERING BA- SIC HELIX-LOOP-HELIX 1)分别结合在PRR7及CC- A1的启动子上来调控其表达水平(Kolmos et al., 2014; Nagel et al., 2014).在植物干旱和盐胁迫应答方面, 生物钟系统也发挥着重要功能.研究表明, 许多与盐、脱水、渗透压调控和响应相关基因的表达均受生物钟调控(Kreps et al., 2002; Covington et al., 2008).在拟南芥中, 参与调控盐胁迫的基因如SOS1 (SALT OVERLY SENSITIVE 1)等的表达呈现出较强的昼夜节律性(http://diurnal.mocklerlab.org/), 表明生物钟可能调控植物对盐胁迫的适应.另外, 生物钟组分GI参与调控植物在高盐环境下的Na+/H+质子泵通道的打开, 进而将Na+排出体外, 减少Na+对植物的毒害(Kim et al., 2013).也有研究表明, PRR7可能参与调控植物响应盐胁迫(Liu et al., 2013).但是关于生物钟系统如何调控植物对盐胁迫的响应及其相关分子机制仍需更系统地挖掘和研究, 并将有望为作物耐盐育种提供理论依据和相关遗传资源. ...
2 2008
... 早期通过转录组数据分析发现, 生物钟系统可能参与茉莉酸(jasmonate acid, JA)与水杨酸(salic acid, SA)的生物合成和信号转导途径(Covington et al., 2008), 而JA和SA被认为是调控植物响应生物胁迫的2类关键激素.MYC2是植物JA信号途径的下游关键因子, 当JA存在时, JAZ (jasmonate ZIM-domain)对MYC2的转录激活活性的阻遏效应被解除, MYC2可以激活JA信号下游效应基因的表达(Song et al., 2014).TIC (TIME FOR COFFEE)是生物钟周期调控的关键因子, 它编码1个含有1 555个氨基酸但生化功能未知的蛋白, 其调控生物钟周期的分子机制目前还不清楚.有研究表明, TIC可以与MYC2通过蛋白互作的方式在细胞核中负调控MYC2的蛋白稳定性.因此, 在tic突变体中JA介导的免疫系统故障导致植物易感病(Shin et al., 2012).此外, 人们也发现MYC2的表达在转录水平和蛋白水平都受到生物钟的调控(Shin et al., 2012), 其表达的峰值出现在傍晚时分, 但是转录水平上调控MYC2基因昼夜节律性表达的生物钟关键组分还有待进一步验证.最新研究显示, 由ELF4- ELF3-LUX组成的生物钟EC复合体可以在转录水平上直接介导时间维度对MYC2的转录抑制, 进而精细调节茉莉酸下游的发育事件, 如叶片衰老等(Zhang et al., 2018). ... ... 生物钟广泛调节植物对非生物胁迫的耐受性, 是协调植物生长发育与胁迫响应的重要调节者和平衡者.在冷胁迫信号应答中, 植物通过CBF/DREB1 (CREPEAT BINDING FACTOR/DEHYDRATION-RES- POSIVE-ELEMENT-BINDING)来激活COR基因的表达, 进而调控植物对冷胁迫的响应, 且CBF基因家族成员自身的表达也呈现明显的昼夜节律性(Liu et al., 1998; Fowler et al., 2005).许多生物钟基因的突变体均表现出对冷胁迫适应性改变的表型, 如生物钟核心组分PRR5/7/9突变后, 可以间接增强CBF的表达水平使其表达丧失昼夜节律性, 从而导致植物具有更强的耐冷性(Nakamichi et al., 2009).PRR基因家族的另一个成员TOC1的突变也会使CBF3表达水平上调, 增强植物对冷胁迫的耐受性(Keily et al., 2013), 说明PRR基因家族在植物冷胁迫的适应方面可能发挥重要作用.在农作物水稻(Oryza sativa)、小麦(Triticum aestivum)和大麦(Hordeum vulgare)中, CBF也参与调控对冷和干旱的响应, 进而影响农作物生长(Chew and Halliday, 2011).以上结果进一步说明, 生物钟系统通过调节CBF基因家族表达的昼夜节律性来协调植物对非生物胁迫响应和自身发育所需能量分配.另一方面, 冷胁迫也可以反馈调节生物钟系统, 如CBF1结合在LUX的启动子上调控其表达(Chow et al., 2014); 低温可诱导CCA1及其它生物钟基因的可变剪切并形成不同的转录本, 从而影响生物钟的功能(Grundy et al., 2015).核心振荡器组分还受到高温的调控, 高温主要影响生物钟组分基因的可变剪切或通过调控热激响应因子HsfB2b (HEAT SHOCK FACTOR B2B)和FBH1 (FLOW-ERING BA- SIC HELIX-LOOP-HELIX 1)分别结合在PRR7及CC- A1的启动子上来调控其表达水平(Kolmos et al., 2014; Nagel et al., 2014).在植物干旱和盐胁迫应答方面, 生物钟系统也发挥着重要功能.研究表明, 许多与盐、脱水、渗透压调控和响应相关基因的表达均受生物钟调控(Kreps et al., 2002; Covington et al., 2008).在拟南芥中, 参与调控盐胁迫的基因如SOS1 (SALT OVERLY SENSITIVE 1)等的表达呈现出较强的昼夜节律性(http://diurnal.mocklerlab.org/), 表明生物钟可能调控植物对盐胁迫的适应.另外, 生物钟组分GI参与调控植物在高盐环境下的Na+/H+质子泵通道的打开, 进而将Na+排出体外, 减少Na+对植物的毒害(Kim et al., 2013).也有研究表明, PRR7可能参与调控植物响应盐胁迫(Liu et al., 2013).但是关于生物钟系统如何调控植物对盐胁迫的响应及其相关分子机制仍需更系统地挖掘和研究, 并将有望为作物耐盐育种提供理论依据和相关遗传资源. ...
1 2016
... 除了在模式植物拟南芥中发现生物钟调控生长发育过程以及应对外界环境变化和胁迫应答方面具有不可忽视的作用外, 近年来, 农作物的生物钟研究逐步展开.水稻是主要粮食作物, 生物钟如何调控水稻的生长发育(尤其是抽穗期)和非生物胁迫应答已越来越受到科学家的关注.抽穗期是水稻的重要农艺性状之一, 抽穗期的长短(从萌发到始穗)直接决定水稻品种生育期的长短, 而适宜的生育期在维持水稻高产稳产中扮演着极为关键的角色, 也在一定程度上反映水稻对种植区域和季节的适应性.研究表明, 生物钟在调控水稻抽穗期方面具有重要作用.图位克隆分析表明, OsELF3 (与拟南芥生物钟核心组分同源的ELF3)的核苷酸变异导致Nipponbare和Koshihikari品种间抽穗期的差异(Matsubara et al., 2012).进一步研究表明, 水稻中存在2个与拟南芥ELF3同源的基因Os- ELF3.1 (LOC_Os06g05060)和OsELF3.2 (LOC_Os- 01g38530) (Zhao et al., 2012).在长日照条件下, OsELF3.1是水稻抽穗期正调节因子, 作用于OsGI (Os-GIGANTEA)和Ghd7 (Grain number, Plant height and heading date 7)的上游, 负调控它们的表达, 因此oself3.1突变体表现出晚抽穗的表型; 但是OsELF3.2的突变并不影响水稻抽穗期.OsELF3.1突变后影响生物钟其它组分OsLHY、OsPRR1、Os- PRR37和OsPRR73的表达, 表明OsELF3.1在维持生物钟周期和调控抽穗期方面具有重要作用(Yang et al., 2013) (图2).有趣的是, 在植物叶片衰老方面, 拟南芥ELF3通过抑制PIF4和PIF5的表达负调控叶片衰老(Sakuraba et al., 2014); 但是水稻OsELF3.1的功能恰恰相反, 它是水稻叶片衰老的正调节因子, 主要调控NAC (N-ACETYL CYSTEINE)和WRKY基因家族成员的表达(Sakuraba et al., 2016).在拟南芥中超表达OsELF3.1则会表现出与ELF3超表达相同的表型, 即延缓叶片衰老、晚花和短周期, 表明ELF3在水稻和拟南芥中功能保守, 可能由于在不同物种中参与调控的下游靶基因完全不同, 导致其在不同植物中的功能差异(Atamian and Harmer, 2016).水稻同源基因OsELF3.2则与E3连接酶APIP6 (AVRPIZT INTERACT PROTEIN 6)互作, 负调控水稻对稻瘟病的免疫(Ning et al., 2015).在春大麦中ELF3通过抑制FT1的表达和产生有活性的GAs (gibberellic acid)来调控其对光周期的敏感性, 控制春大麦由营养生长阶段向生殖生长阶段的转变(Boden et al., 2014).近期研究表明, 大麦中ELF3通过生物钟系统介导植物对温度信号的响应.受高温刺激后, CCA1和PRR73的表达水平明显降低, 而GI和PRR59的表达水平明显升高, 但是这种变化在elf3突变体中消失, 表明GI和PRR对温度响应依赖于ELF3的功能(Ford et al., 2016), 但ELF3调控大麦对温度响应的分子机制还不清楚.在另一重要作物大豆(Glycine max)中, ELF3基因突变导致其成熟时间延长, 即“长青春期”, 这能够提高大豆产量, 增强大豆在低纬度地区的区域适应性(Lu et al., 2017; Yue et al., 2017).进一步研究表明, GmELF3结合在E1启动子上抑制其表达, 进而使下游FT2a和FT5a的表达水平降低而延长大豆青春期(Lu et al., 2017).ELF3基因在长日照植物拟南芥中抑制开花, 而在短日照植物(如水稻和大豆)中促进开花, 这一有趣的现象是否与生物钟对日长的感知机制有关以及它们差异化调控的分子机制还有待进一步研究. ...
1 2005
... 生物钟广泛调节植物对非生物胁迫的耐受性, 是协调植物生长发育与胁迫响应的重要调节者和平衡者.在冷胁迫信号应答中, 植物通过CBF/DREB1 (CREPEAT BINDING FACTOR/DEHYDRATION-RES- POSIVE-ELEMENT-BINDING)来激活COR基因的表达, 进而调控植物对冷胁迫的响应, 且CBF基因家族成员自身的表达也呈现明显的昼夜节律性(Liu et al., 1998; Fowler et al., 2005).许多生物钟基因的突变体均表现出对冷胁迫适应性改变的表型, 如生物钟核心组分PRR5/7/9突变后, 可以间接增强CBF的表达水平使其表达丧失昼夜节律性, 从而导致植物具有更强的耐冷性(Nakamichi et al., 2009).PRR基因家族的另一个成员TOC1的突变也会使CBF3表达水平上调, 增强植物对冷胁迫的耐受性(Keily et al., 2013), 说明PRR基因家族在植物冷胁迫的适应方面可能发挥重要作用.在农作物水稻(Oryza sativa)、小麦(Triticum aestivum)和大麦(Hordeum vulgare)中, CBF也参与调控对冷和干旱的响应, 进而影响农作物生长(Chew and Halliday, 2011).以上结果进一步说明, 生物钟系统通过调节CBF基因家族表达的昼夜节律性来协调植物对非生物胁迫响应和自身发育所需能量分配.另一方面, 冷胁迫也可以反馈调节生物钟系统, 如CBF1结合在LUX的启动子上调控其表达(Chow et al., 2014); 低温可诱导CCA1及其它生物钟基因的可变剪切并形成不同的转录本, 从而影响生物钟的功能(Grundy et al., 2015).核心振荡器组分还受到高温的调控, 高温主要影响生物钟组分基因的可变剪切或通过调控热激响应因子HsfB2b (HEAT SHOCK FACTOR B2B)和FBH1 (FLOW-ERING BA- SIC HELIX-LOOP-HELIX 1)分别结合在PRR7及CC- A1的启动子上来调控其表达水平(Kolmos et al., 2014; Nagel et al., 2014).在植物干旱和盐胁迫应答方面, 生物钟系统也发挥着重要功能.研究表明, 许多与盐、脱水、渗透压调控和响应相关基因的表达均受生物钟调控(Kreps et al., 2002; Covington et al., 2008).在拟南芥中, 参与调控盐胁迫的基因如SOS1 (SALT OVERLY SENSITIVE 1)等的表达呈现出较强的昼夜节律性(http://diurnal.mocklerlab.org/), 表明生物钟可能调控植物对盐胁迫的适应.另外, 生物钟组分GI参与调控植物在高盐环境下的Na+/H+质子泵通道的打开, 进而将Na+排出体外, 减少Na+对植物的毒害(Kim et al., 2013).也有研究表明, PRR7可能参与调控植物响应盐胁迫(Liu et al., 2013).但是关于生物钟系统如何调控植物对盐胁迫的响应及其相关分子机制仍需更系统地挖掘和研究, 并将有望为作物耐盐育种提供理论依据和相关遗传资源. ...
1 2014
... 生物钟核心组分PRR基因家族在作物中也发挥重要功能.研究表明水稻OsPRR37可能参与调控Hd3a基因的表达, 从而调控水稻对光周期的敏感性(Koo et al., 2013), 但对于其调控的精确的分子机制尚不清楚.通过近等基因系定位到的DTH7 (Days To Heading 7), 其表达受生物钟调控, 呈现出昼夜节律, 在短日照和长日照条件下, DTH7表达量均在ZT8达到最高峰.进一步实验证明, DTH7位点即是OsPRR- 37基因的变异, 参与调控水稻开花时间和区域的适应性(Gao et al., 2014).最新利用大数据的研究进一步证明OsPRR37和Ghd7是影响水稻开花时间、株高和每穗小穗数量等的主要数量性状位点(Liu et al., 2015).GI是另一个在作物中发现的也具有保守功能的生物钟关键组分.Izawa等(2011)利用水稻osgi突变体在自然生长条件下, 连续多时间点取叶片进行转录组分析, 发现在检测到的27 201个基因中约75%的基因受到OsGI调控, 并且OsGI对于维持这些基因的生物节律是必需的.研究表明, OsGI在维持水稻生物钟功能中具有重要作用, 但是其具体作用机制仍然未知.最近研究表明, 在短日照条件下, OsPhyA突变后导致水稻抽穗期延迟, 主要是由于突变体中OsGI表达水平降低, 进而影响下游3个开花促进因子Hd1 (Heading Date 1)、OsMADS51和OsId1 (Oryza sativa Indeterminate 1)表达水平下调(Lee et al., 2016).在大豆中, GI的同源基因有3个, 分别为GmGI1、GmGI2和GmGI3, 其中GmGI2的表达受光周期和生物钟调控, 可能参与调控大豆开花时间(Li et al., 2013), 但是其作用机制尚不明确.水稻生物钟系统除了参与调控抽穗期之外, 还有可能参与对非生物胁迫的响应.前期的研究表明, 水稻es1-1 (early sense-cence 1)突变体在正常和冷处理条件下其生物钟关键组分OsGI、OsLHY、OsPRR1、OsPRR37、OsPRR73和OsPRR95的表达模式与表达水平均有不同程度的改变, 表明水稻生物钟系统可能参与植物对冷胁迫的响应(徐江民等, 2016), 但是具体的分子机制仍需进一步研究. ...
1 2012
... 拟南芥(Arabidopsis thaliana)作为模式植物被广泛用于植物分子生物学和遗传学研究, 因此其生物钟分子机制也研究得较为透彻.1995年, Millar等(1995)首次采用荧光素酶作为报告基因鉴定到第1个生物钟突变体toc1 (timing of cab expression 1).随后, Wang和Tobin (1998)利用反向遗传学方法鉴定到第1个生物钟核心振荡器的关键成员CCA1 (CIRCADIAN CLOCK ASSOCIATED 1), 该基因编码1个MYB类转录因子.超表达CCA1会使生物钟报告基因丧失昼夜节律性表达模式, 而且极度晚花, 同时也抑制内源CCA1和LHY (LATE ELONGATED HYPOCOTYL)的表达, 且该基因自身的转录水平还受到红光的瞬时诱导.基于基因表达水平和启动子活性分析, AlabadÍ等(2001)提出了拟南芥生物钟第1个分子反馈环路, 即CCA1/LHY抑制TOC1的表达, 而TOC1则促进CCA1/LHY的表达, 由此形成了生物钟核心振荡器的转录-翻译反馈环.随着技术手段的进步和计算生物学的发展, 目前认为生物钟核心振荡器由3个紧密关联的转录-翻译反馈环组成(图1).其中, 中心反馈环由LHY和CCA1以及TOC1组成.CCA1和LHY在早晨表达, 二者存在部分功能冗余, 它们转录并翻译成熟后会相互作用进而抑制TOC1的表达.在晚上, CCA1和LHY的表达水平降低, TOC1的表达则达到峰值, 反过来抑制CCA1和LHY的表达, 进而使它们处于此起彼伏的状态(Gendron et al., 2012; Huang et al., 2012).CCA1/LHY也和PRR (PSEUDO RESPONSE REGULATOR)基因家族的PRR9/PRR7组成早反馈环.PRRs成员中PRR9、PRR7及PRR5从早到晚每隔2-3小时依次出现表达峰值, 它们会通过与TOPL- ESS基因家族的成员互作招募组蛋白去乙酰化酶HDA6/19 (HISTONE DEACETYLASE 6/19)形成转录抑制复合体, 从而在转录水平上抑制CCA1和LHY的表达(Nakamichi et al., 2010; Wang et al., 2013).反之, CCA1/LHY也抑制PRR9/7的表达, 从而形成早反馈环.晚反馈环包含MYB类转录因子LUX ARRHYTHMO (又命名为PHYTO-CLOCK1)以及2个核蛋白ELF3 (EARLY FLOWERING 3)和ELF4组成的EC复合体(evening complex) (Nusinow et al., 2011)和TOC1.EC复合体与CCA1/LHY相互调节, 间接促进CCA1/LHY的表达, 反过来CCA1和LHY会直接结合在EC复合体启动子上抑制其转录(Adams et al., 2015).早反馈环、中心反馈环和晚反馈环相互连锁形成生物钟核心振荡器的抑制振荡子(repressilator)基本架构. ...
1 2015
... 由于地球自转而产生的光-暗循环以及由该循环产生的温度昼夜周期性的变化, 是植物在生长发育过程中必须应对的昼夜节律性动态变化.在长期进化过程中产生的生物钟系统赋予了植物对光温等环境因子昼夜周期性变化的适应性.生物钟系统使植物可以追踪环境因子的昼夜周期性变化并感知和预测这种变化的动态及季节的变换, 以协调体内代谢稳态、细胞运动以及生长发育与防御反应的动态平衡, 使植物能在合适的时间完成其关键生长发育过程(Green- ham and McClung, 2015).人们最初认为生物钟系统是一个线性过程, 即包括外界信号输入、内源的“计时员”(追踪时间的分子机制)和输出途径, 如叶片运动和基因节律性表达等(Harmer, 2009).最新研究表明, 受生物钟调控的某些生物学过程也会反馈调节生物钟的核心振荡器, 进而形成一个复杂的互作调控网络, 使植物更好地适应生存环境(Nohales and Kay, 2016).生物钟系统协调植物外部光温信号与内源的代谢和发育状态, 以调控生长发育与胁迫应答之间的平衡, 并决定昼夜周期性信号的输出, 从而使生物体避免资源和能量的损失以达到最优化生长. ...
1 2015
... 生物钟广泛调节植物对非生物胁迫的耐受性, 是协调植物生长发育与胁迫响应的重要调节者和平衡者.在冷胁迫信号应答中, 植物通过CBF/DREB1 (CREPEAT BINDING FACTOR/DEHYDRATION-RES- POSIVE-ELEMENT-BINDING)来激活COR基因的表达, 进而调控植物对冷胁迫的响应, 且CBF基因家族成员自身的表达也呈现明显的昼夜节律性(Liu et al., 1998; Fowler et al., 2005).许多生物钟基因的突变体均表现出对冷胁迫适应性改变的表型, 如生物钟核心组分PRR5/7/9突变后, 可以间接增强CBF的表达水平使其表达丧失昼夜节律性, 从而导致植物具有更强的耐冷性(Nakamichi et al., 2009).PRR基因家族的另一个成员TOC1的突变也会使CBF3表达水平上调, 增强植物对冷胁迫的耐受性(Keily et al., 2013), 说明PRR基因家族在植物冷胁迫的适应方面可能发挥重要作用.在农作物水稻(Oryza sativa)、小麦(Triticum aestivum)和大麦(Hordeum vulgare)中, CBF也参与调控对冷和干旱的响应, 进而影响农作物生长(Chew and Halliday, 2011).以上结果进一步说明, 生物钟系统通过调节CBF基因家族表达的昼夜节律性来协调植物对非生物胁迫响应和自身发育所需能量分配.另一方面, 冷胁迫也可以反馈调节生物钟系统, 如CBF1结合在LUX的启动子上调控其表达(Chow et al., 2014); 低温可诱导CCA1及其它生物钟基因的可变剪切并形成不同的转录本, 从而影响生物钟的功能(Grundy et al., 2015).核心振荡器组分还受到高温的调控, 高温主要影响生物钟组分基因的可变剪切或通过调控热激响应因子HsfB2b (HEAT SHOCK FACTOR B2B)和FBH1 (FLOW-ERING BA- SIC HELIX-LOOP-HELIX 1)分别结合在PRR7及CC- A1的启动子上来调控其表达水平(Kolmos et al., 2014; Nagel et al., 2014).在植物干旱和盐胁迫应答方面, 生物钟系统也发挥着重要功能.研究表明, 许多与盐、脱水、渗透压调控和响应相关基因的表达均受生物钟调控(Kreps et al., 2002; Covington et al., 2008).在拟南芥中, 参与调控盐胁迫的基因如SOS1 (SALT OVERLY SENSITIVE 1)等的表达呈现出较强的昼夜节律性(http://diurnal.mocklerlab.org/), 表明生物钟可能调控植物对盐胁迫的适应.另外, 生物钟组分GI参与调控植物在高盐环境下的Na+/H+质子泵通道的打开, 进而将Na+排出体外, 减少Na+对植物的毒害(Kim et al., 2013).也有研究表明, PRR7可能参与调控植物响应盐胁迫(Liu et al., 2013).但是关于生物钟系统如何调控植物对盐胁迫的响应及其相关分子机制仍需更系统地挖掘和研究, 并将有望为作物耐盐育种提供理论依据和相关遗传资源. ...
1 2009
... 由于地球自转而产生的光-暗循环以及由该循环产生的温度昼夜周期性的变化, 是植物在生长发育过程中必须应对的昼夜节律性动态变化.在长期进化过程中产生的生物钟系统赋予了植物对光温等环境因子昼夜周期性变化的适应性.生物钟系统使植物可以追踪环境因子的昼夜周期性变化并感知和预测这种变化的动态及季节的变换, 以协调体内代谢稳态、细胞运动以及生长发育与防御反应的动态平衡, 使植物能在合适的时间完成其关键生长发育过程(Green- ham and McClung, 2015).人们最初认为生物钟系统是一个线性过程, 即包括外界信号输入、内源的“计时员”(追踪时间的分子机制)和输出途径, 如叶片运动和基因节律性表达等(Harmer, 2009).最新研究表明, 受生物钟调控的某些生物学过程也会反馈调节生物钟的核心振荡器, 进而形成一个复杂的互作调控网络, 使植物更好地适应生存环境(Nohales and Kay, 2016).生物钟系统协调植物外部光温信号与内源的代谢和发育状态, 以调控生长发育与胁迫应答之间的平衡, 并决定昼夜周期性信号的输出, 从而使生物体避免资源和能量的损失以达到最优化生长. ...
1 2017
... 在光周期依赖途径的成花诱导过程中, 日长的变化会诱导叶片中开花整合因子FT (FLOWERING LOCUS T)基因的表达, FT蛋白被运送到顶端分生组织诱导植物开花.生物钟对植物开花时间的调控主要体现在对CO/B-BOX1 (CONSTANS)等光周期依赖途径关键基因的表达节律和时相调控上, 这些基因感受特定日长的光周期信号并且在转录和转录后水平上受到生物钟系统多层级的调控与修饰(Song et al., 2015).在拟南芥中, 长日照促进开花, CO蛋白在长日照下更稳定, 进而诱导FT基因表达; 但在短日照条件下, FT基因的表达水平非常低, 导致植物开花时间延迟(Kobayashi et al., 1999).生物钟关键基因突变导致的生物钟功能紊乱会改变开花时间, 说明生物钟在感知日长信息调控光周期途径开花启动方面发挥重要作用.生物钟核心振荡器重要组分还在转录后水平调节CO蛋白的稳定性, 生物钟关键因子PRR5/7/9的突变导致植物极度晚花.最新研究表明, 它们通过与CO蛋白直接在细胞核中互作, 进而调控CO蛋白的稳定性而促进开花.由于PRR蛋白表达的时间特异性, 它们能与CO蛋白在一天之中的特定时间内互作而增强其稳定性, 并促进CO蛋白结合在FT启动子上, 从而诱导FT的表达, 促进植物开花(Hayama et al., 2017).上述研究从分子水平部分解析了prr5/7/9突变体植物极度晚花的机制, 也为生物钟关键组分在蛋白质翻译后修饰水平调节开花时间提供了新证据.最近研究表明, 与CO蛋白家族一样含有保守B-box锌指基序, 但是不含有DNA结合结构域的BBX (B-BOX GENES)基因家族成员也可能是生物钟周期调控和开花时间调控的衔接者.例如, 研究表明BBX32 (B- BOX32)蛋白参与生物钟输出途径控制开花时间, 同时其过表达转基因株系在持续光照的条件下生物钟周期变长, 但其参与生物钟周期调控的机制还不清楚(Tripathi et al., 2017).此外, BBX32与COL3 (CON- STANTS-LIKE 3)/BBX4相互作用抑制FT基因的表达而调控开花(Tripathi et al., 2017).关于这类基因家族在介导开花时间与生物钟周期调控方面的研究有望成为新的研究热点, 可为更复杂层级的生物钟感知光周期途径调控开花时间的信号网络提供更多的实验证据. ...
1 2012
... 拟南芥(Arabidopsis thaliana)作为模式植物被广泛用于植物分子生物学和遗传学研究, 因此其生物钟分子机制也研究得较为透彻.1995年, Millar等(1995)首次采用荧光素酶作为报告基因鉴定到第1个生物钟突变体toc1 (timing of cab expression 1).随后, Wang和Tobin (1998)利用反向遗传学方法鉴定到第1个生物钟核心振荡器的关键成员CCA1 (CIRCADIAN CLOCK ASSOCIATED 1), 该基因编码1个MYB类转录因子.超表达CCA1会使生物钟报告基因丧失昼夜节律性表达模式, 而且极度晚花, 同时也抑制内源CCA1和LHY (LATE ELONGATED HYPOCOTYL)的表达, 且该基因自身的转录水平还受到红光的瞬时诱导.基于基因表达水平和启动子活性分析, AlabadÍ等(2001)提出了拟南芥生物钟第1个分子反馈环路, 即CCA1/LHY抑制TOC1的表达, 而TOC1则促进CCA1/LHY的表达, 由此形成了生物钟核心振荡器的转录-翻译反馈环.随着技术手段的进步和计算生物学的发展, 目前认为生物钟核心振荡器由3个紧密关联的转录-翻译反馈环组成(图1).其中, 中心反馈环由LHY和CCA1以及TOC1组成.CCA1和LHY在早晨表达, 二者存在部分功能冗余, 它们转录并翻译成熟后会相互作用进而抑制TOC1的表达.在晚上, CCA1和LHY的表达水平降低, TOC1的表达则达到峰值, 反过来抑制CCA1和LHY的表达, 进而使它们处于此起彼伏的状态(Gendron et al., 2012; Huang et al., 2012).CCA1/LHY也和PRR (PSEUDO RESPONSE REGULATOR)基因家族的PRR9/PRR7组成早反馈环.PRRs成员中PRR9、PRR7及PRR5从早到晚每隔2-3小时依次出现表达峰值, 它们会通过与TOPL- ESS基因家族的成员互作招募组蛋白去乙酰化酶HDA6/19 (HISTONE DEACETYLASE 6/19)形成转录抑制复合体, 从而在转录水平上抑制CCA1和LHY的表达(Nakamichi et al., 2010; Wang et al., 2013).反之, CCA1/LHY也抑制PRR9/7的表达, 从而形成早反馈环.晚反馈环包含MYB类转录因子LUX ARRHYTHMO (又命名为PHYTO-CLOCK1)以及2个核蛋白ELF3 (EARLY FLOWERING 3)和ELF4组成的EC复合体(evening complex) (Nusinow et al., 2011)和TOC1.EC复合体与CCA1/LHY相互调节, 间接促进CCA1/LHY的表达, 反过来CCA1和LHY会直接结合在EC复合体启动子上抑制其转录(Adams et al., 2015).早反馈环、中心反馈环和晚反馈环相互连锁形成生物钟核心振荡器的抑制振荡子(repressilator)基本架构. ...
1 2011
... 生物钟核心组分PRR基因家族在作物中也发挥重要功能.研究表明水稻OsPRR37可能参与调控Hd3a基因的表达, 从而调控水稻对光周期的敏感性(Koo et al., 2013), 但对于其调控的精确的分子机制尚不清楚.通过近等基因系定位到的DTH7 (Days To Heading 7), 其表达受生物钟调控, 呈现出昼夜节律, 在短日照和长日照条件下, DTH7表达量均在ZT8达到最高峰.进一步实验证明, DTH7位点即是OsPRR- 37基因的变异, 参与调控水稻开花时间和区域的适应性(Gao et al., 2014).最新利用大数据的研究进一步证明OsPRR37和Ghd7是影响水稻开花时间、株高和每穗小穗数量等的主要数量性状位点(Liu et al., 2015).GI是另一个在作物中发现的也具有保守功能的生物钟关键组分.Izawa等(2011)利用水稻osgi突变体在自然生长条件下, 连续多时间点取叶片进行转录组分析, 发现在检测到的27 201个基因中约75%的基因受到OsGI调控, 并且OsGI对于维持这些基因的生物节律是必需的.研究表明, OsGI在维持水稻生物钟功能中具有重要作用, 但是其具体作用机制仍然未知.最近研究表明, 在短日照条件下, OsPhyA突变后导致水稻抽穗期延迟, 主要是由于突变体中OsGI表达水平降低, 进而影响下游3个开花促进因子Hd1 (Heading Date 1)、OsMADS51和OsId1 (Oryza sativa Indeterminate 1)表达水平下调(Lee et al., 2016).在大豆中, GI的同源基因有3个, 分别为GmGI1、GmGI2和GmGI3, 其中GmGI2的表达受光周期和生物钟调控, 可能参与调控大豆开花时间(Li et al., 2013), 但是其作用机制尚不明确.水稻生物钟系统除了参与调控抽穗期之外, 还有可能参与对非生物胁迫的响应.前期的研究表明, 水稻es1-1 (early sense-cence 1)突变体在正常和冷处理条件下其生物钟关键组分OsGI、OsLHY、OsPRR1、OsPRR37、OsPRR73和OsPRR95的表达模式与表达水平均有不同程度的改变, 表明水稻生物钟系统可能参与植物对冷胁迫的响应(徐江民等, 2016), 但是具体的分子机制仍需进一步研究. ...


{kind=link}
{kind=link}
{kind=link}
{kind=link}