Abstract: We explored the different photosynthetic characteristics of three Pseudosasa cultivars: P. japonica, P. japonica f. akebonosuji, and P. japonica f. akebono. The differences in photosystem activity and photosynthetic characteristics of different leaf colors were revealed by the changes of chloroplast ultrastructure and fluorescence kinetics curves. The results showed that the photosynthetic pigment content of the three species was significantly different. Except for the intact thylakoid layer structure in the white part of the chloroplast, the green streak and the radix were significantly less than the radix. The chloroplast developmental maturity is inconsistent; the OJIP curve and parameters indicate that the open reduction of the flowering green leaf and the saplings of the PSII reaction center is lower than that of the yam, the capture energy that is used for the electron transfer share to be smaller, and the PSII activity is weaker; The redox balance of the electron transport chain of bamboo leaves P700 to QA is biased towards the reducing side, and it is presumed that the P700 reaction center P700 to PSII primary electron acceptor QA electron transport chain is damaged. Therefore, the chloroplast development caused by changes in PSII activity in the photosystem is immature, which may be the direct cause of the difference in leaf color of the species.
1 材料与方法1.1 实验材料实验于2016年6月下旬在浙江省临安市浙江农林大学翠竹园进行, 选择竹高、叶位和长势一致的矢竹(Pseu- dosasa japonica (Siebold & Zuccarini ex Steudel Makino ex Nakai))、花叶矢竹(P. japonica f. akebonosuji)和曙筋矢竹(P. japonica f. akebono)当年生新竹, 抽枝展叶成熟后, 选取花叶矢竹条纹叶片, 包括白色部分(albino sector in leaf with strips, SA)和绿色部分(green sector in leaf with strips, SG), 曙筋矢竹淡绿色叶片(virescent leaf, VL)和矢竹深绿色叶片(green leaf, GL)为实验材料。
2 结果与讨论2.1 矢竹类3个叶色叶片表型及光合色素水平差异由图1可知, 3种类型的矢竹叶片颜色存在差异。原种矢竹深绿色, 曙筋矢竹叶片淡绿色, 且颜色浅于花叶矢竹条纹叶的绿色部分, 花叶矢竹叶片具有不规则白绿相间条纹。由表1可知, 3个竹种不同颜色叶片叶绿素a (Chla)、叶绿素b (Chlb)及总叶绿素含量(Chla+b)均存在显著差异。矢竹叶片不同光合色素水平均最高, 其中Chla+b达35.86。曙筋矢竹Chla+b为矢竹的63.55%, 未能达到原种矢竹的绿色程度, 但叶绿素a/b值(Chla/b)与矢竹无显著差异; 其类胡萝卜素(Car)含量是矢竹的83.37%, 而Car含量的增加对曙筋矢竹光合能量耗散有利。花叶矢竹白色部分虽然含有叶绿素及类胡萝卜素, 但含量极少, 仅为绿色部分的1%左右, 其Chla/b仅为曙筋矢竹和花叶矢竹绿色部分的1/3左右。因此, 花叶矢竹白色部分叶绿素含量极少, 基本不存在光合作用, 在进行光合测定分析时仅用绿色部分作为实验材料。 表1 Table 1 表1 表1 不同种类矢竹叶片光合色素含量及比值(平均值±标准误) Table 1 Photosynthetic pigments content and relative ratio of different cultivars of Pseudosasa japonica leaves (means±SE)
Photosynthetic pigments
GL
SA
SG
VL
Chla (mg·g-1 FW)
27.19±1.17 a
0.34±0.17 c
25.39±2.41 a
17.09±0.52 b
Chlb (mg·g-1 FW)
8.66±0.33 a
0.16±0.07 c
8.27±0.69 a
5.70±0.17 b
Car (mg·g-1 FW)
4.89±0.28 a
0.32±0.08 c
5.52±0.55 a
4.07±0.13 b
Chla/b
3.14±0.02 a
1.99±0.09 b
3.07±0.03 a
3.00±0.00 a
Chla+b (mg·g-1 FW)
35.86±1.51 a
0.51±0.25 c
33.66±3.11 a
22.79±0.70 b
SA: 花叶矢竹条纹叶片白色部分; SG: 花叶矢竹条纹叶片绿色部分; VL: 曙筋矢竹淡绿色叶片; GL: 矢竹深绿色叶片。不同小写字母表示差异显著(P<0.05), 横向多重比较。 SA: Albino sector in leaf with strips of P. japonica f. akebonosuji; SG: Green sector in leaf with strips of P. japonica f. akebonosuji; VL: Virescent leaf of P. japonica f. akebono; GL: Green leaf of P. japonica. Different lowercase letters indicate significant differences at P<0.05 level, lateral multiple comparison.
表1 不同种类矢竹叶片光合色素含量及比值(平均值±标准误) Table 1 Photosynthetic pigments content and relative ratio of different cultivars of Pseudosasa japonica leaves (means±SE)
图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-509/img_1.png<b>图1</b> 矢竹类3个叶色叶片表型<br/>SG+SA: 花叶矢竹条纹叶片绿色部分及白色部分; VL: 曙筋矢竹淡绿色叶片; GL: 矢竹深绿色叶片<br/><b>Figure 1</b> Three kinds of <i>Pseudosasa japonica</i> leaves<br/>SA+SG: Albino and green sector in leaf of <i>P. japonica </i>f. <i>akebonosuji </i>with strips; VL: Virescent leaf of <i>P. japonica </i>f. <i>akebono</i>. GL: Green leaf of <i>P. japonica</i> Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-509/img_1.png<b>图1</b> 矢竹类3个叶色叶片表型<br/>SG+SA: 花叶矢竹条纹叶片绿色部分及白色部分; VL: 曙筋矢竹淡绿色叶片; GL: 矢竹深绿色叶片<br/><b>Figure 1</b> Three kinds of <i>Pseudosasa japonica</i> leaves<br/>SA+SG: Albino and green sector in leaf of <i>P. japonica </i>f. <i>akebonosuji </i>with strips; VL: Virescent leaf of <i>P. japonica </i>f. <i>akebono</i>. GL: Green leaf of <i>P. japonica</i>
图1 矢竹类3个叶色叶片表型 SG+SA: 花叶矢竹条纹叶片绿色部分及白色部分; VL: 曙筋矢竹淡绿色叶片; GL: 矢竹深绿色叶片 Figure 1 Three kinds of Pseudosasa japonica leaves SA+SG: Albino and green sector in leaf of P. japonica f. akebonosuji with strips; VL: Virescent leaf of P. japonica f. akebono. GL: Green leaf of P. japonica
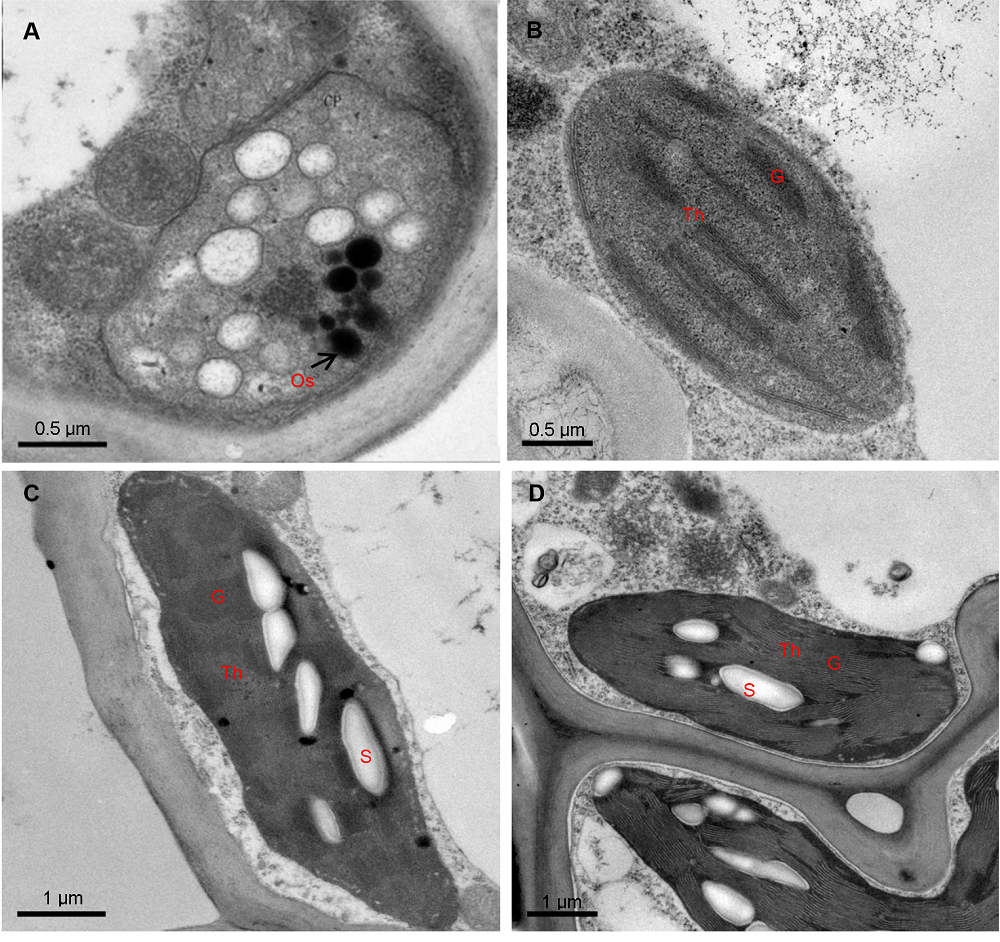
2.2 叶片叶绿体超微结构特征3种矢竹叶片叶绿体超微结构观察结果(图2A-D)表明, 不同矢竹之间叶肉细胞叶绿体结构存在较大差异。其中, 花叶矢竹白色部分的平均叶绿体数为1.43个, 叶绿体近圆形, 叶绿体外被膜良好, 内膜解体、呈囊泡状结构, 无完整类囊体片层结构, 有若干嗜锇小体分散(图2A)。花叶矢竹绿色部分平均叶绿体数为2.76个, 叶绿体椭圆形, 膜结构较完整, 基粒均匀排列, 基质片层沿着叶绿体长轴方向排列, 但无淀粉粒(图2B)。曙筋矢竹叶绿体结构较完整, 叶绿体紧贴细胞壁分布在细胞中。曙筋矢竹的叶绿体片层清晰、垛叠紧密, 平均叶绿体数为2.18个, 有较多淀粉粒(图2C)。矢竹的叶绿体类囊体片层充满整个叶绿体, 基粒片层清晰, 有较多且较大的淀粉粒。叶绿体的平均叶绿体数为4.37个, 比花叶矢竹绿色部分多36%, 且垛叠更紧密(图2D)。 图2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-509/img_2.png<b>图2</b> 3个竹种叶片叶绿体结构<br/>(A) 花叶矢竹白区叶肉细胞; (B) 花叶矢竹绿区叶肉细胞; (C) 曙筋矢竹叶肉细胞; (D) 矢竹叶肉细胞。G: 基粒; Os: 嗜锇小体; S: 淀粉粒; Th: 类囊体膜<br/><b>Figure 2</b> Plate chloroplast ultrastructure of three cultivars of <i>Pseudosasa japonica</i> leaves<br/>(A) Mesophyll cells in white zones of zebra leaf of <i>P. japonica </i>f. <i>akebonosuji</i>; (B) Mesophyll cells in green zones of <i>P. japonica </i>f. <i>akebonosuji</i>; (C) Mesophyll cells in the leaf of <i>P. japonica </i>f. <i>akebono</i>; (D) Mesophyll cells in the leaf of <i>P. japonica. </i>G: Granum; Os: Osmiophile globule; S: Starch grain; Th: Thylakoid membranes Figure 2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-509/img_2.png<b>图2</b> 3个竹种叶片叶绿体结构<br/>(A) 花叶矢竹白区叶肉细胞; (B) 花叶矢竹绿区叶肉细胞; (C) 曙筋矢竹叶肉细胞; (D) 矢竹叶肉细胞。G: 基粒; Os: 嗜锇小体; S: 淀粉粒; Th: 类囊体膜<br/><b>Figure 2</b> Plate chloroplast ultrastructure of three cultivars of <i>Pseudosasa japonica</i> leaves<br/>(A) Mesophyll cells in white zones of zebra leaf of <i>P. japonica </i>f. <i>akebonosuji</i>; (B) Mesophyll cells in green zones of <i>P. japonica </i>f. <i>akebonosuji</i>; (C) Mesophyll cells in the leaf of <i>P. japonica </i>f. <i>akebono</i>; (D) Mesophyll cells in the leaf of <i>P. japonica. </i>G: Granum; Os: Osmiophile globule; S: Starch grain; Th: Thylakoid membranes
图2 3个竹种叶片叶绿体结构 (A) 花叶矢竹白区叶肉细胞; (B) 花叶矢竹绿区叶肉细胞; (C) 曙筋矢竹叶肉细胞; (D) 矢竹叶肉细胞。G: 基粒; Os: 嗜锇小体; S: 淀粉粒; Th: 类囊体膜 Figure 2 Plate chloroplast ultrastructure of three cultivars of Pseudosasa japonica leaves (A) Mesophyll cells in white zones of zebra leaf of P. japonica f. akebonosuji; (B) Mesophyll cells in green zones of P. japonica f. akebonosuji; (C) Mesophyll cells in the leaf of P. japonica f. akebono; (D) Mesophyll cells in the leaf of P. japonica. G: Granum; Os: Osmiophile globule; S: Starch grain; Th: Thylakoid membranes
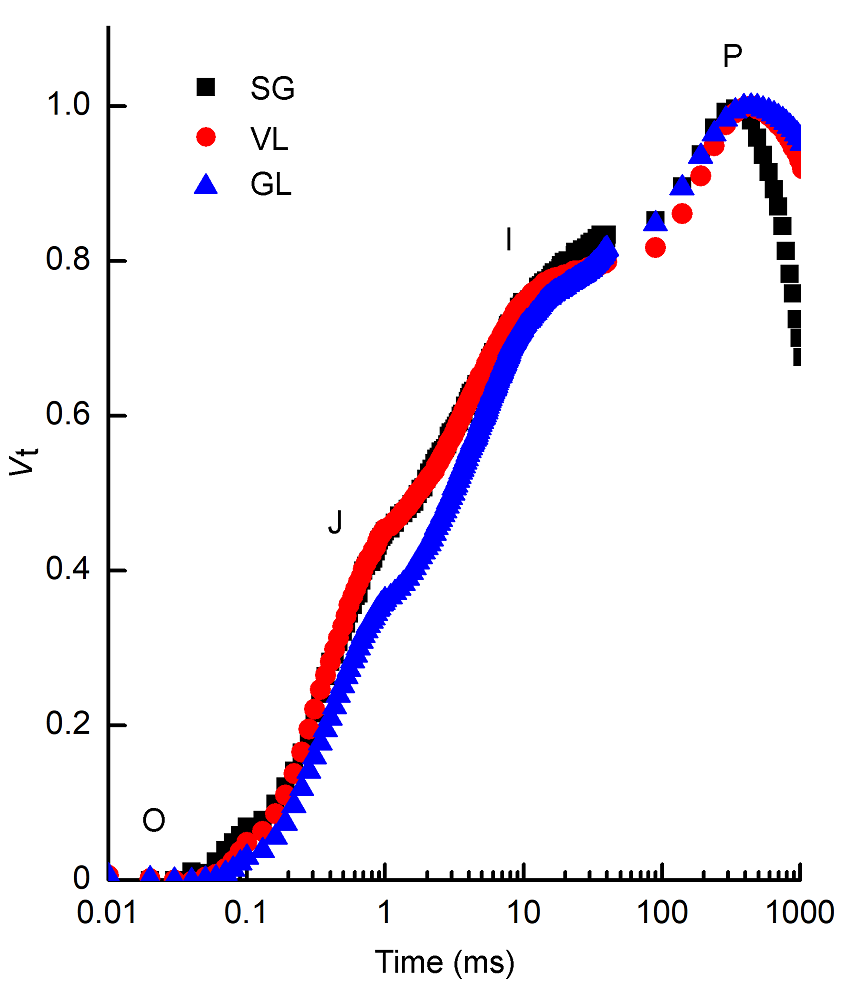
2.3 快速荧光分析2.3.1 叶绿素荧光诱导曲线及拐点的变化 非调制连续激发式荧光仪通过极短时间内照光后荧光信号的瞬时变化, 反映出暗反应活化前PSII的光化学变化(李鹏民, 2006)。从暗适应后的照光瞬间得到初始荧光F0开始到最大荧光Fp。不同种类矢竹叶片快速荧光诱导动力学曲线(图3)包含O、J、I和P等相(Strasser et al., 2004)。由图3可知, 将OJIP曲线双重归一化后各阶段都存在, 说明光合电子链仍然能够有效运转。VL和SG的OJIP曲线各相差异不大, 与GL相比, 其O-I阶段近乎相同, O-I相越高说明PSII还原侧电子从初级电子受体QA向次级电子受体QB的传递越强, 表明GL从QA向QB电子传递的能力高。P点的荧光大小趋势为GL>VL>SG, 表明曙筋矢竹和花叶矢竹的PSII活性均弱于矢竹。 图3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-509/img_3.png<b>图3</b> 不同种类矢竹叶片相对可变荧光强度(<i>V</i>t)随时间的变化<br/>SG、VL和GL同<xref ref-type="table" rid="T1-1674-3466-53-4-509">表1</xref>。<br/><b>Figure 3</b> Relative variable fluorescence (<i>V</i>t) with the time change of different cultivars of <i>Pseudosasa japonica</i> leaves<br/>SG, VL and GL see <xref ref-type="table" rid="T1-1674-3466-53-4-509">Table 1</xref>. Figure 3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-509/img_3.png<b>图3</b> 不同种类矢竹叶片相对可变荧光强度(<i>V</i>t)随时间的变化<br/>SG、VL和GL同<xref ref-type="table" rid="T1-1674-3466-53-4-509">表1</xref>。<br/><b>Figure 3</b> Relative variable fluorescence (<i>V</i>t) with the time change of different cultivars of <i>Pseudosasa japonica</i> leaves<br/>SG, VL and GL see <xref ref-type="table" rid="T1-1674-3466-53-4-509">Table 1</xref>.
图3 不同种类矢竹叶片相对可变荧光强度(Vt)随时间的变化 SG、VL和GL同表1。 Figure 3 Relative variable fluorescence (Vt) with the time change of different cultivars of Pseudosasa japonica leaves SG, VL and GL see Table 1.
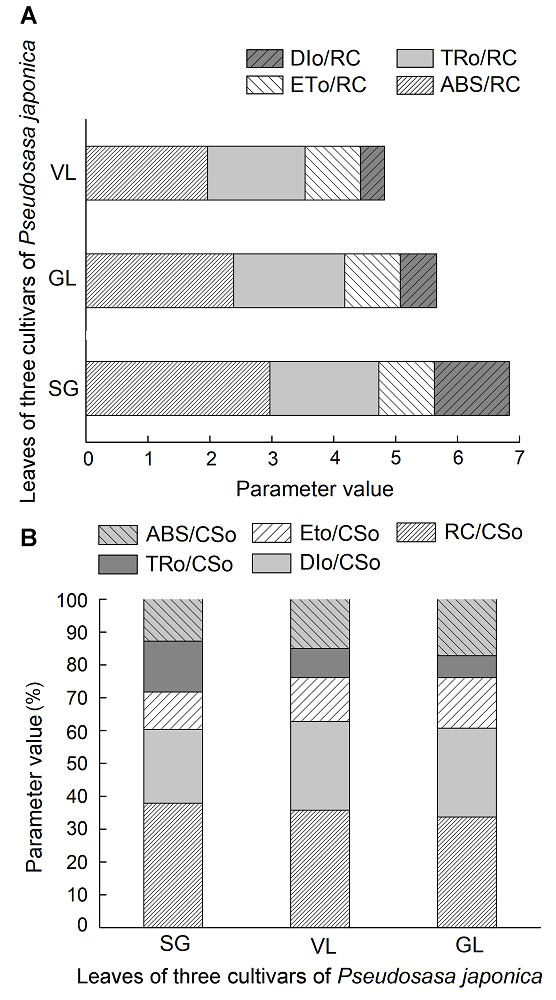
2.3.2 比活性参数的变化 叶绿素荧光曲线还可以反映光合机构的比活性, 即单位面积内活跃的反应中心的各种量子效率。图4A和B分别为单位反应中心及单位截面上吸收的光能和用于还原QA的能量, 即用于热耗散的能量。单位反应中心及单位截面积吸收的光能大小为SG>GL>VL, 其中SG用于电子传递的能量最高。Rc/Cso代表t=0时单位面积内反应中心的数量, 它反映植物光合机构的状态(Heerden et al., 2010)。不同类型叶片中, SG和VL中单位面积的光合机构含有反应中心数目Rc/Cso低于GL。以上结果表明, 矢竹反应中心数量最大, 花叶中反应中心数量最少; 而曙筋矢竹反应中心数量有所增加, 但并没有完全恢复到矢竹的水平。 图4https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-509/img_4.png<b>图4</b> 不同种类矢竹叶片比活性参数<br/>VL、GL和SG同<xref ref-type="table" rid="T1-1674-3466-53-4-509">表1</xref>。ABS/RC: 单位反应中心吸收的光能; TRo/ RC: PSII的最大捕获量; ETo/RC: 单位反应中心用于电子传递的能量; DIo/RC: 单位反应中心的热耗散; ABS/CSo: 单位面积吸收的光能; TRo/CSo: 单位面积捕获的光能; ETo/CSo: 单位面积的电子传递量子产额; DIo/CSo: 单位面积的热耗散; RC/CSo: 单位面积反应中心的数量<br/><b>Figure 4</b> Activity parameters for unit reaction center of different cultivars of <i>Pseudosasa japonica</i> leaves<br/>VL, GL and SG see <xref ref-type="table" rid="T1-1674-3466-53-4-509">Table 1</xref>. ABS/RC: The amount of light absorbed by the unit reaction center; TRo/RC: The large amount of PSII; ETo/RC: The energy of the unit reaction center for electron transfer; DIo/RC: The heat dissipation of the unit reaction center; ABS/CSo: Absorption flux per unit area; TRo/CSo: Trapped energy flux per unit area; ETo/CSo: Electron transport flux per unit area; DIo/CSo: Dissipated energy flux perunit area; RC/CSo: Number of active reaction centers per unit area Figure 4https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-509/img_4.png<b>图4</b> 不同种类矢竹叶片比活性参数<br/>VL、GL和SG同<xref ref-type="table" rid="T1-1674-3466-53-4-509">表1</xref>。ABS/RC: 单位反应中心吸收的光能; TRo/ RC: PSII的最大捕获量; ETo/RC: 单位反应中心用于电子传递的能量; DIo/RC: 单位反应中心的热耗散; ABS/CSo: 单位面积吸收的光能; TRo/CSo: 单位面积捕获的光能; ETo/CSo: 单位面积的电子传递量子产额; DIo/CSo: 单位面积的热耗散; RC/CSo: 单位面积反应中心的数量<br/><b>Figure 4</b> Activity parameters for unit reaction center of different cultivars of <i>Pseudosasa japonica</i> leaves<br/>VL, GL and SG see <xref ref-type="table" rid="T1-1674-3466-53-4-509">Table 1</xref>. ABS/RC: The amount of light absorbed by the unit reaction center; TRo/RC: The large amount of PSII; ETo/RC: The energy of the unit reaction center for electron transfer; DIo/RC: The heat dissipation of the unit reaction center; ABS/CSo: Absorption flux per unit area; TRo/CSo: Trapped energy flux per unit area; ETo/CSo: Electron transport flux per unit area; DIo/CSo: Dissipated energy flux perunit area; RC/CSo: Number of active reaction centers per unit area
图4 不同种类矢竹叶片比活性参数 VL、GL和SG同表1。ABS/RC: 单位反应中心吸收的光能; TRo/ RC: PSII的最大捕获量; ETo/RC: 单位反应中心用于电子传递的能量; DIo/RC: 单位反应中心的热耗散; ABS/CSo: 单位面积吸收的光能; TRo/CSo: 单位面积捕获的光能; ETo/CSo: 单位面积的电子传递量子产额; DIo/CSo: 单位面积的热耗散; RC/CSo: 单位面积反应中心的数量 Figure 4 Activity parameters for unit reaction center of different cultivars of Pseudosasa japonica leaves VL, GL and SG see Table 1. ABS/RC: The amount of light absorbed by the unit reaction center; TRo/RC: The large amount of PSII; ETo/RC: The energy of the unit reaction center for electron transfer; DIo/RC: The heat dissipation of the unit reaction center; ABS/CSo: Absorption flux per unit area; TRo/CSo: Trapped energy flux per unit area; ETo/CSo: Electron transport flux per unit area; DIo/CSo: Dissipated energy flux perunit area; RC/CSo: Number of active reaction centers per unit area
2.4 调制荧光结合远红光PAM2500与是否加远红光的区别仅在于F0′的有无, 而当用光化光(AL)将叶绿素荧光诱导到光稳态时, PSII-PSI-碳同化达到一个高速运转的动态平衡状态; 再用远红光(FR)激发PSI, 通过2个光系统PSI和 PSII间的线性传递, 迅速消耗掉在PQ累积的电子, 使PSII原初电子受体QA快速还原, PSII反应中心在光适应条件下完全关闭, 同时铁氧还蛋白-ADP+还原酶活性和碳同化也暂时停止。当AL再次打开时, 叶绿素荧光从F0′上升到Fs, PSII-PSI-碳同化再次被激活, 并回到Fs的动态平衡状态(钟传飞, 2008)。因此F0′-Fs是一个稳态过程, 并在300秒左右达到稳定, 这主要是由于卡尔文循环的各种酶被激活(Gilmore et al., 1997), 质子梯度逐渐稳定(Schreiber, 2004), 光暗反应逐渐达到动态平衡的结果(张阿宏等, 2008)。 2.4.1 调制荧光参数的差异分析 F0是暗适应状态下PSII反应中心完全开放时的荧光产量, 代表反应中心处于开放状态时的荧光产量, 其大小与捕光天线系统及PSII反应中心状态均有密切关系(许大全, 2001)。花叶矢竹和曙筋矢竹的F0均与矢竹差异显著。Fv/Fm表示PSII的最大光化学量子产量, 代表反应中心的活性(桂仁意等, 2010)。SG的Fv/Fm最小, 为GL的84%, 其最大光化学效率低于矢竹与曙筋矢竹。Fv/F0代表PSII光反应中心的潜在活性(林世青等, 1992)。花叶矢竹PSII光反应中心潜在活性高于曙筋矢竹而低于矢竹, 说明花叶矢竹绿叶部分PSII光反应中心潜在活性没有恢复到矢竹的水平。3个竹种间反映热耗散能力的光化学淬灭系数qP差异不显著, 表明它们用于电子传递的光能差异不显著(表2)。 表2 Table 2 表2 表2 不同种类矢竹叶片荧光参数分析 Table 2 Analysis of fluorescence parameters of different cultivars of Pseudosasa japonica leaves
GL
VL
SG
F0
0.40±0.01 b
0.44±0.03 a
0.39±0.04 ab
Fm
1.64±0.13 a
1.35±0.16 a
1.29±0.17 a
Fv/Fm
0.75±0.02 a
0.67±0.02 b
0.63±0.01 b
Fv/F0
3.11±0.36 a
2.01±0.19 b
2.26±0.14 b
Y(II)
0.38±0.04 b
0.27±0.03 a
0.31±0.04 a
NPQ
1.27±0.12 b
1.71±0.21 a
1.47±0.18 ab
qP
0.77±0.05 a
0.77±0.03 a
0.79±0.04 a
ETR
23.00±2.94 a
16.22±1.64 a
19.00±2.83 a
F0: 初始荧光; Fm: 最大荧光产量; Fv/Fm: PSII最大光化学量子产量; Fv/F0: PSII潜在光活性; Y(II): 实际量子产额; NPQ: 非光化学淬灭系数: qP: 光化学淬灭系数; ETR: 表观电子传递效率。GL、VL和SG同表1。采用LSD法进行多重比较, 同一行中不同小写字母表示差异显著(α=0.05)。 F0: Minimal fluorescence; Fm: Maximal fluorescence; Fv/Fm: Maximal quantum yield of PSII; Fv/F0: PSII potential optical activity; Y(II): The actual amount of quantum yield; NPQ: Coefficient of non-photochemical quenching; qP: Coefficient of photochemical quenching; ETR: Apparent electron transfer efficiency. GL, VL and SG see Table 1. Different lowercase letters in the same row indicate significant differences at α=0.05 according to LSD test.
表2 不同种类矢竹叶片荧光参数分析 Table 2 Analysis of fluorescence parameters of different cultivars of Pseudosasa japonica leaves
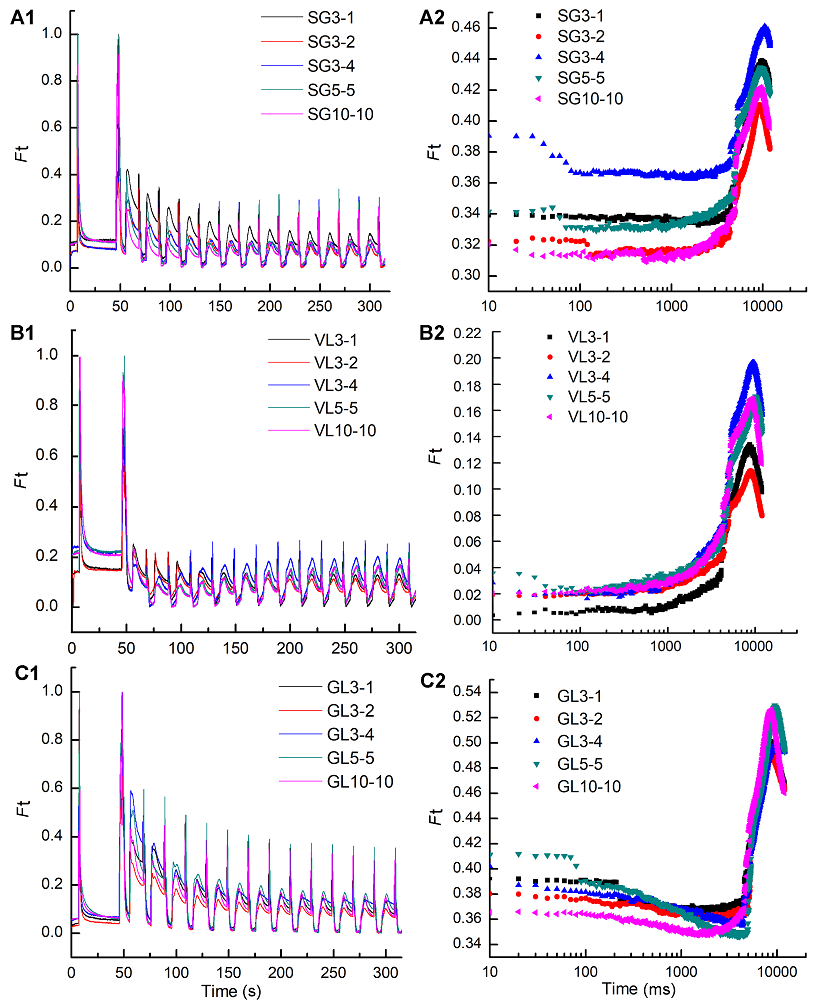
2.4.2 不同远红光组合对稳态荧光动力学的影响 光稳态时, 在每个周期中光化光转变为远红光后, 仅P700被激发而导致PSI之前的电子传递体直至QA被重氧化(钟传飞, 2008)。由图5可知, 不同远红光(FR)组合对3种矢竹叶片F0′影响差异不显著, 而对稳态荧光曲线的变化影响差异较大。FR打开时, 实时荧光曲线Ft迅速从暗适应后最大荧光Fm下降到光适应下最小荧光F0′, 而且从F0′到稳态荧光Fs过程都经历低-高-低的过程。SG从F0′到Fs过程中, Ft随着远红光组合的增强而先减小后增加。在10-10处理下的荧光上升速度最大, 当远红光组合为3-1、3-2和10-10处理时均在70 ms时(298秒)达到最低值F0′, 荧光开始上升的时间最早的是10-10处理。不同FR处理间VL对稳态荧光影响不大, 随着FR增强, 荧光上升的速度变化幅度较小, 但叶绿素荧光诱导曲线斜率呈上升趋势。 图5https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-509/img_5.png<b>图5</b> 3个竹种在不同远红光强度及时间下荧光动力学曲线变化<br/>(A1), (A2) 不同远红光强度及时间下花叶矢竹条纹叶片绿色部分荧光动力学曲线(A1)和稳态荧光(A2); (B1), (B2) 不同远红光强度及时间下曙筋矢竹淡绿色叶片荧光动力学曲线(B1)和稳态荧光(B2); (C1), (C2) 不同远红光强度及时间下矢竹深绿色叶片荧光动力学曲线(C1)和稳态荧光(C2)。SG、VL和GL同<xref ref-type="table" rid="T1-1674-3466-53-4-509">表1</xref>; <i>F</i>t: 实时荧光曲线。<br/><b>Figure 5</b> The change of fluorescence transients under different intensity and time of three cultivars of <i>Pseudosasa japonica</i> leaves under far-red light treatments<br/>(A1), (A2) The change of fluorescence transients (A1) and steady-state fluorescence (A2) of green sector in leaf with strips of <i>P. japonica</i> f. <i>akebonosuji</i> under different intensity and time of far-red light treatments; (B1), (B2) The change of fluorescence transients (B1) and steady-state fluorescence (B2) of the virescent leaves of <i>P. japonica </i>f. <i>akebono</i> under different intensity and time of far-red light treatments; (C1), (C2) The change of fluorescence transients (C1) and steady-state fluorescence (C2) of the green leaves of <i>P. japonica </i>under different intensity and time of far-red light treatments. SG, VL and GL see <xref ref-type="table" rid="T1-1674-3466-53-4-509">Table 1</xref>; <i>F</i>t: Real-time fluorescence curve. Figure 5https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-509/img_5.png<b>图5</b> 3个竹种在不同远红光强度及时间下荧光动力学曲线变化<br/>(A1), (A2) 不同远红光强度及时间下花叶矢竹条纹叶片绿色部分荧光动力学曲线(A1)和稳态荧光(A2); (B1), (B2) 不同远红光强度及时间下曙筋矢竹淡绿色叶片荧光动力学曲线(B1)和稳态荧光(B2); (C1), (C2) 不同远红光强度及时间下矢竹深绿色叶片荧光动力学曲线(C1)和稳态荧光(C2)。SG、VL和GL同<xref ref-type="table" rid="T1-1674-3466-53-4-509">表1</xref>; <i>F</i>t: 实时荧光曲线。<br/><b>Figure 5</b> The change of fluorescence transients under different intensity and time of three cultivars of <i>Pseudosasa japonica</i> leaves under far-red light treatments<br/>(A1), (A2) The change of fluorescence transients (A1) and steady-state fluorescence (A2) of green sector in leaf with strips of <i>P. japonica</i> f. <i>akebonosuji</i> under different intensity and time of far-red light treatments; (B1), (B2) The change of fluorescence transients (B1) and steady-state fluorescence (B2) of the virescent leaves of <i>P. japonica </i>f. <i>akebono</i> under different intensity and time of far-red light treatments; (C1), (C2) The change of fluorescence transients (C1) and steady-state fluorescence (C2) of the green leaves of <i>P. japonica </i>under different intensity and time of far-red light treatments. SG, VL and GL see <xref ref-type="table" rid="T1-1674-3466-53-4-509">Table 1</xref>; <i>F</i>t: Real-time fluorescence curve.
图5 3个竹种在不同远红光强度及时间下荧光动力学曲线变化 (A1), (A2) 不同远红光强度及时间下花叶矢竹条纹叶片绿色部分荧光动力学曲线(A1)和稳态荧光(A2); (B1), (B2) 不同远红光强度及时间下曙筋矢竹淡绿色叶片荧光动力学曲线(B1)和稳态荧光(B2); (C1), (C2) 不同远红光强度及时间下矢竹深绿色叶片荧光动力学曲线(C1)和稳态荧光(C2)。SG、VL和GL同表1; Ft: 实时荧光曲线。 Figure 5 The change of fluorescence transients under different intensity and time of three cultivars of Pseudosasa japonica leaves under far-red light treatments (A1), (A2) The change of fluorescence transients (A1) and steady-state fluorescence (A2) of green sector in leaf with strips of P. japonica f. akebonosuji under different intensity and time of far-red light treatments; (B1), (B2) The change of fluorescence transients (B1) and steady-state fluorescence (B2) of the virescent leaves of P. japonica f. akebono under different intensity and time of far-red light treatments; (C1), (C2) The change of fluorescence transients (C1) and steady-state fluorescence (C2) of the green leaves of P. japonica under different intensity and time of far-red light treatments. SG, VL and GL see Table 1; Ft: Real-time fluorescence curve.
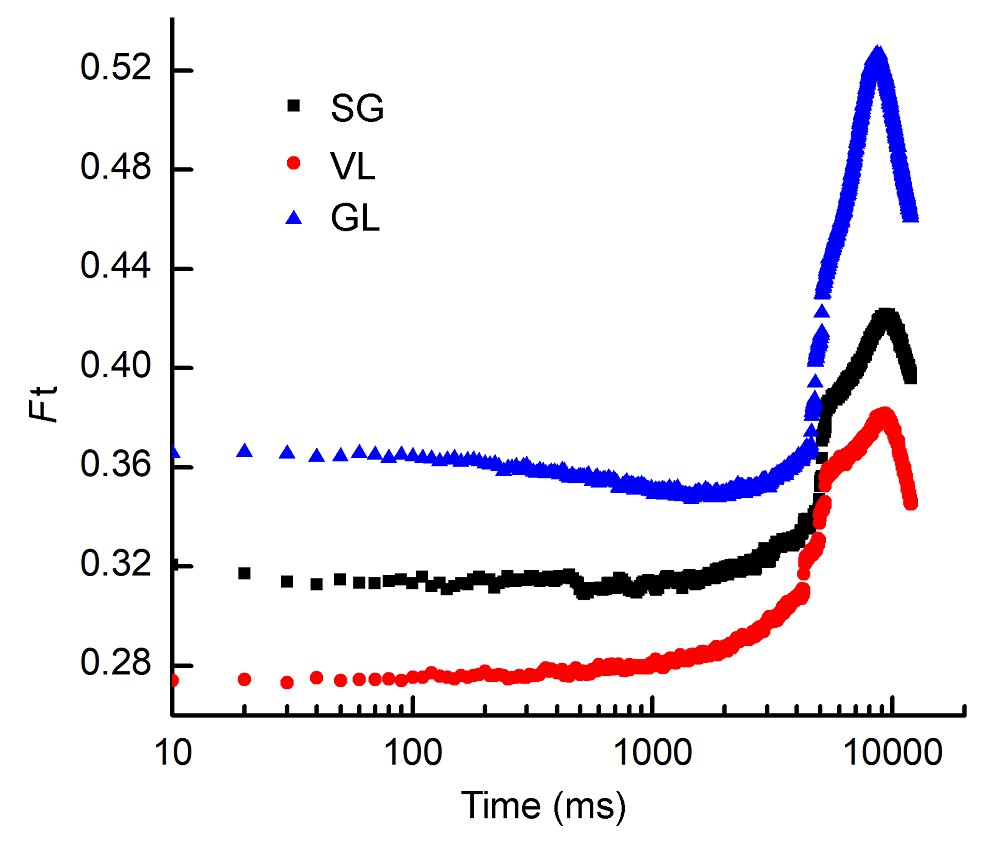
GL在FR组合10-10时荧光到达最低点的时间最早, 而对荧光上升速度影响不大。远红光组合10-10处理, 最能反映3个竹种稳态荧光动力学影响。 在10-10远红光处理下将稳态荧光诱导曲线放大之后(图6), 发现Ft很粗糙, 表明电子链不通畅。从图6中还可以看出, SG比VL和GL的动力学曲线更粗糙, F0′到Fs的上升幅度也更小, 表明花叶矢竹PSII到PSI光合电子传递链阻塞更加严重, 还未完全恢复正常。由图6的曲线斜率变化可知, 与VL相比, SG的Fm′到F0′下降速度较快, 降幅也较大, 但是它们的降幅都不如GL; 而从F0′到Fs的变化过程来看, 虽然Ft上升的幅度有所增加, 但是上升速度较慢, 且上升过程中曲线仍较粗糙, 表明光合电子传递链没有完全畅通, 可能远红光占用时间过长, 电子虽然排空, 碳代谢进行少, 可能有更多的电子用于环式电子流中。 图6https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-509/img_6.png<b>图6</b> 3个竹种在远红光10-10 (强度-时间)组合下稳态荧光动力学曲线变化<br/>SG、VL和GL同<xref ref-type="table" rid="T1-1674-3466-53-4-509">表1</xref>; <i>F</i>t同<xref ref-type="fig" rid="F5-1674-3466-53-4-509">图5</xref>。<br/><b>Figure 6</b> The change of steady-state fluorescence of 10-10 (intensity-time) of three cultivars of <i>Pseudosasa japonica</i> leaves under far-red light treatments<br/>SG, VL and GL see <xref ref-type="table" rid="T1-1674-3466-53-4-509">Table 1</xref>; <i>F</i>t see <xref ref-type="fig" rid="F5-1674-3466-53-4-509">Figure 5</xref>. Figure 6https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-509/img_6.png<b>图6</b> 3个竹种在远红光10-10 (强度-时间)组合下稳态荧光动力学曲线变化<br/>SG、VL和GL同<xref ref-type="table" rid="T1-1674-3466-53-4-509">表1</xref>; <i>F</i>t同<xref ref-type="fig" rid="F5-1674-3466-53-4-509">图5</xref>。<br/><b>Figure 6</b> The change of steady-state fluorescence of 10-10 (intensity-time) of three cultivars of <i>Pseudosasa japonica</i> leaves under far-red light treatments<br/>SG, VL and GL see <xref ref-type="table" rid="T1-1674-3466-53-4-509">Table 1</xref>; <i>F</i>t see <xref ref-type="fig" rid="F5-1674-3466-53-4-509">Figure 5</xref>.
图6 3个竹种在远红光10-10 (强度-时间)组合下稳态荧光动力学曲线变化 SG、VL和GL同表1; Ft同图5。 Figure 6 The change of steady-state fluorescence of 10-10 (intensity-time) of three cultivars of Pseudosasa japonica leaves under far-red light treatments SG, VL and GL see Table 1; Ft see Figure 5.
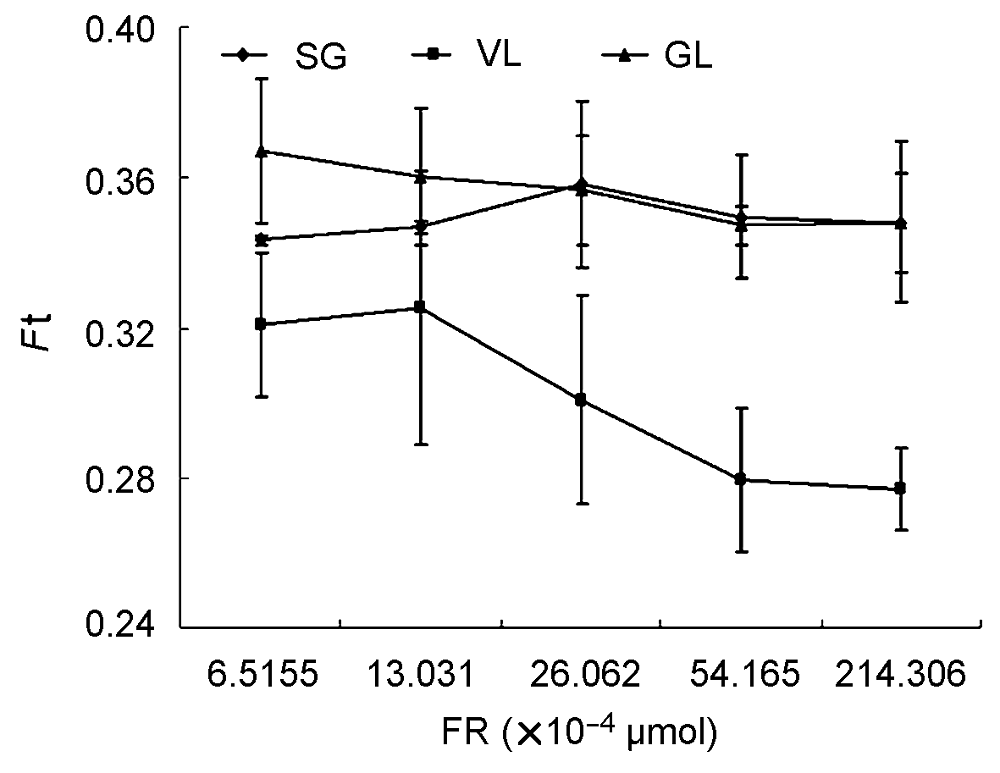
2.4.3 不同远红光强度对叶片荧光参数F0′的影响 由图7可知, 不同远红光对3种叶片的F0′影响不同。不同远红光组合使花叶矢竹的F0′在0.35上下浮动, 随着远红光加强F0′先增强后降低, 但各个处理之间差异不显著; 矢竹和曙筋矢竹的F0′随着远红光的加强而减小。矢竹的F0′差异不显著, 但曙筋矢竹前期3.03× 10-4 μmol处理下的F0′与远红光54.17×10-4 μmol处理下差异显著, 而远红光54.17×10-4 μmol处理下的F0′与远红光214.31×10-4 μmol处理下的F0′差异不显著, 即曙筋矢竹的F0′随远红光增强到一定程度差异不显著。矢竹在不同远红光处理下的电子传递变化与花叶矢竹的电子传递变化一致。曙筋矢竹的F0′随着远红光增强而持续减小, 表明PSI的P700至PSII的QA的电子链被氧化的程度持续增加。 图7https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-509/img_7.png<b>图7</b> 不同远红光强度对3个竹种<i>F</i><sub>0</sub>′ (光下最小荧光)的影响<br/>SG、VL和GL同<xref ref-type="table" rid="T1-1674-3466-53-4-509">表1</xref>; <i>F</i>t同<xref ref-type="fig" rid="F5-1674-3466-53-4-509">图5</xref>; FR: 远红光强度。<br/><b>Figure 7</b> The change of <i>F</i><sub>0</sub>′ (minimum fluorescence under the light) of three cultivars of <i>Pseudosasa japonica </i>leaves under different intensity of far-red light treatments<br/>SG, VL and GL see <xref ref-type="table" rid="T1-1674-3466-53-4-509">Table 1</xref>; <i>F</i>t see <xref ref-type="fig" rid="F5-1674-3466-53-4-509">Figure 5</xref>; FR: The intensity of far red light. Figure 7https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-509/img_7.png<b>图7</b> 不同远红光强度对3个竹种<i>F</i><sub>0</sub>′ (光下最小荧光)的影响<br/>SG、VL和GL同<xref ref-type="table" rid="T1-1674-3466-53-4-509">表1</xref>; <i>F</i>t同<xref ref-type="fig" rid="F5-1674-3466-53-4-509">图5</xref>; FR: 远红光强度。<br/><b>Figure 7</b> The change of <i>F</i><sub>0</sub>′ (minimum fluorescence under the light) of three cultivars of <i>Pseudosasa japonica </i>leaves under different intensity of far-red light treatments<br/>SG, VL and GL see <xref ref-type="table" rid="T1-1674-3466-53-4-509">Table 1</xref>; <i>F</i>t see <xref ref-type="fig" rid="F5-1674-3466-53-4-509">Figure 5</xref>; FR: The intensity of far red light.
图7 不同远红光强度对3个竹种F0′ (光下最小荧光)的影响 SG、VL和GL同表1; Ft同图5; FR: 远红光强度。 Figure 7 The change of F0′ (minimum fluorescence under the light) of three cultivars of Pseudosasa japonica leaves under different intensity of far-red light treatments SG, VL and GL see Table 1; Ft see Figure 5; FR: The intensity of far red light.
钟传飞 (2008). 稳态叶绿素荧光动力学理论构建和常绿阔叶植物越冬光合生理生态研究. 博士论文. 北京: 北京林业大学. pp. 47-49.
[本文引用: 3]
[16]
DeellJR, MurrDP, WileyL (2003). 1-Methylcyclopropene (1-MCP) increases CO2 injury in apples.Acta Horticul 600, 277-280. [本文引用: 1]
[17]
GilmoreAM, HazlettTL, Govindjee (1997). Xanthophyll cycle-dependent quenching of photosystem II chlorophyll a fluorescence: formation of a quenching complex with a short fluorescence life time.Proc Natl Acad Sci USA 92, 2273-2277. [本文引用: 1]
[18]
HeerdenPDRV, StrasserRJ, KrügerGHJ (2010). Reduction of dark chilling stress in N2-fixing soybean by nitrate as indicated by chlorophyll a fluorescence kinetics. Physiol Plantarum 121, 239-249. [本文引用: 1]
[19]
LichtenthalerHK (1987). Chlorophylls and carotenoids: pigments of photosynthetic biomembranes.Methods Enzymol 148, 350-382. [本文引用: 1]
[20]
MurchieEH, LawsonT (2013). Chlorophyll fluorescence analysis: a guide to good practice and understanding so- me new applications.J Exp Bot 64, 3983-3998. [本文引用: 1]
[21]
PapageorgiouGC, TsimillimichaelM, StamatakisK (2007). The fast and slow kinetics of chlorophyll a fluorescence induction in plants, algae and cyanobacteria: a viewpoint.Photosynth Res 94, 275-290. [本文引用: 1]
[22]
SchreiberU (2004). Pulse-amplitude-modulation (PAM) fluo- rometry and saturation pulse method: an overview. In: Papageorgiou GC, Govindjee, eds. Chlorophyll a Fluorescence. Heidelberg: Springer. pp. 279-319. [本文引用: 1]
[23]
StrasserRJ, Tsimilli-michaelM, SrivastavaA (2004). Analysis of the chlorophyll a fluorescence transient. In: Papageorgiou GC, Govindjee, eds. Chlorophyll a Fluorescence: A Signature of Photosynthesis. Dordrecht: Springer. pp. 321-362.
[24]
Tsimilli-MichaelM, StrasserRJ (2008). In vivo assessment of stress impact on Plant’s Vitality: Applications in Detecting and Evaluating the Beneficial Role of Mycorrhization on Host Plants. In: Varma A, eds. Heidelberg: Springer. pp. 679-703. [本文引用: 1]
[25]
WangQ, SullivanRW, KightA, HenryRL, HuangJR, JonesAM, KorthKL (2004). Deletion of the chloroplast- localized Thylakoid formation1 gene product in Arabidopsis leads to deficient thylakoid formation and variegated leaves. Plant Physiol 136, 3594-3604. [本文引用: 1]
[26]
WatersMT, LangdaleJA (2009). The making of a chloroplast.EMBO J 28, 2861-2873. [本文引用: 1]
[27]
WuJX, ZhangZG, ZhangQ, HanX, GuXF, LuTG (2015). The molecular cloning and clarification of a photorespiratory mutant, oscdm1, using enhancer trapping. Front Ge- net 6, 226. [本文引用: 1]
[28]
ZhuXG, Govindjee, BakerNR, deSturlerE, OrtDR, LongSP (2005). Chlorophyll a fluorescence induction kinetics in leaves predicted from a model describing each discrete step of excitation energy and electron transfer associated with Photosystem II.Planta 223, 114-133. [本文引用: 1]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}