Abstract: Programmed cell death (PCD) is a fundamental biological process in plant development and is also related to plant response to environment stresses. Recently, Chinese scientists have made breakthroughs in understanding the molecular mechanisms of PCD signaling pathways in plants.
Key words:? programmed cell death ; chloroplast-to-mitochondrion communication ; reactive oxygen species ; malate
在一定的生理状态和生长发育过程中, 植物可以通过程序性细胞死亡(programmed cell death, PCD)维持体内稳定和平衡, 从而更好地适应环境并完成生命活动。PCD是一种受到遗传调控的主动性死亡过程, 通过调节细胞分化、细胞数量及超敏反应(HR)等, 在植物的生命活动和生物/非生物胁迫反应中发挥重要作用。尽管动、植物的PCD属性相似且具有共同的抑制因子(Vianello et al., 2007), 但由于植物含有叶绿体这一特有的亚细胞结构, 预示着植物的PCD信号通路还具有一定的独特性(Beers, 1997)。因此, 解析植物PCD信号通路是当前亟待解决的重要课题。最近, 中国科学家以拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)为材料, 在解析PCD发生机制及其在生产中的应用方面取得了突破性进展(Zhao et al., 2018; Heng et al., 2018)。
活性氧(reactive oxygen species, ROS)可以由叶绿体及线粒体等亚细胞产生。研究表明, ROS在植物的PCD过程中扮演了多种角色(Noctor et al., 2002)。普遍的观点认为, 光照条件下叶绿体是主要的ROS来源, 许多植物的PCD依赖光照和功能完整的叶绿体。此外, 线粒体组分参与PCD表明线粒体也能触发和执行PCD过程, ROS可以充当诱导PCD的信号分子, 改变线粒体膜通透性, 或者形成线粒体通透性转变孔(MPTP)。细胞色素C和线粒体电子传递链(mETC)崩解释放的ROS足以促使PCD的发生达到致死水平(Van Aken and Van Breusegem, 2015)。这些结果表明, 叶绿体和线粒体在执行PCD过程中起着重要作用。然而, 仍有一些根本性的问题有待解决。例如, 植物PCD过程是否需要依赖叶绿体和线粒体间的信号传递, 以及介导这种信号传递的分子是什么。解决这些问题有助于我们加深对植物PCD的认知和理解, 对于揭示动物PCD过程的分子机制也具有借鉴意义。
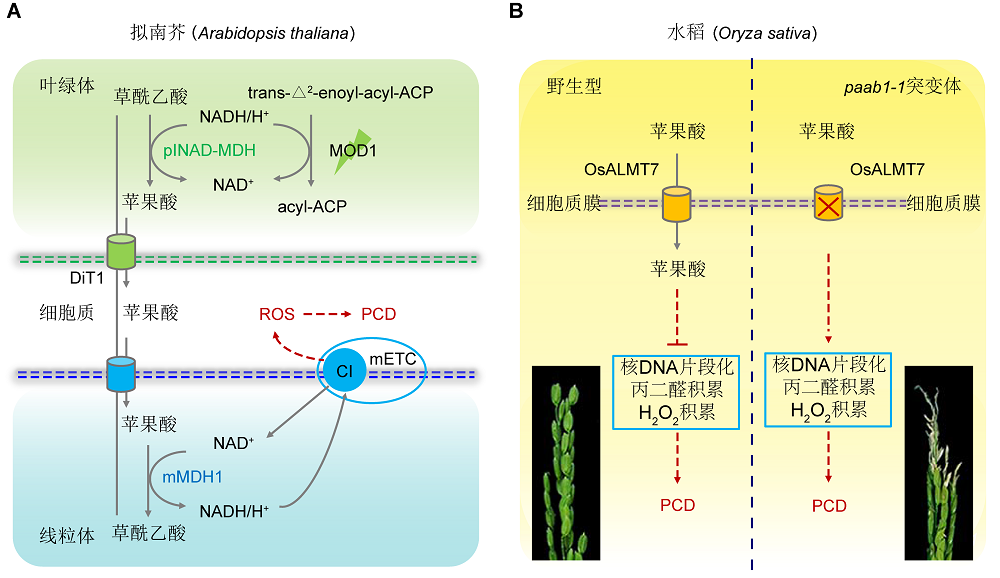
图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-447/img_1.png<b>图1</b> 苹果酸调控植物程序性细胞死亡模式图(改自<xref ref-type="bibr" rid="b8-1674-3466-53-4-447">Zhao et al., 2018</xref>; <xref ref-type="bibr" rid="b2-1674-3466-53-4-447">Heng et al., 2018</xref>)<br/>(A) 在拟南芥中, <i>MOD1</i>编码烯脂酰还原酶, 突变体<i>mod1</i>中底物NADH在叶绿体大量积累, NADH与草酰乙酸(OAA)在plNAD-MDH的催化作用下生成苹果酸, 通过DiT1运输至细胞质, 继而被某个/某些转运蛋白转运至线粒体。苹果酸在线粒体中被mMDH1催化形成草酰乙酸, 并伴随着NADH的积累, 为线粒体膜上的mETC复合体提供电子, 产生ROS, 进而导致<i>mod1</i>突变体产生程序性细胞死亡(PCD)。(B) 水稻<i>OsALMT7</i>基因编码1个定位于质膜的苹果酸转运蛋白, 可以将苹果酸转运至细胞内。苹果酸可以维持核DNA稳定, 减少H<sub>2</sub>O<sub>2</sub>和丙二醛的积累, 从而抑制PCD。在其突变体<i>paab1-1</i>中, 苹果酸积累减少, 伴随着核DNA片段化, 同时积累H<sub>2</sub>O<sub>2</sub>和丙二醛, 促进PCD。<br/><b>Figure 1</b> A proposed model of malate-regulated programmed cell death in plants (modified from <xref ref-type="bibr" rid="b8-1674-3466-53-4-447">Zhao et al., 2018</xref>; <xref ref-type="bibr" rid="b2-1674-3466-53-4-447">Heng et al., 2018</xref>)<br/>(A) In <i>Arabidopsis thaliana</i>, <i>MOD1</i> encodes an enoyl-acyl carrier protein reductase, the deficiency of MOD1 causes an increased level of NADH in the chloroplasts, which drives oxaloacetate (OAA) to be converted to malate by plNAD-MDH. Malate is transported out of the chloroplast into the cytosol by DiT1, and then be transported into the mitochondrion by an unidentified transporter or transporters. In the mitochondrion, malate is converted to OAA by mMDH1, and simultaneously NADH is generated to provide electrons for mETC to induce ROS formation and initiate programmed cell death (PCD) process in the <i>mod1</i> cells. (B) In <i>Oryza sativa</i>, <i>OsALMT7</i> encodes a putative aluminum-activated plasma membrane localized malate transporter, which could transport malate into cells. Malate is involved in nuclear DNA stability and inhibits excessive accumulation of H<sub>2</sub>O<sub>2</sub> and malondialdehyde, which protects cells from PCD. The <i>paab1-1</i> mutant harbors a mutation in <i>OsALMT7</i>, and its panicle contained less malate than wild type, particularly at the apical portions. The apical spikelets in the <i>paab1-1</i> mutant undergo PCD accompanied by nuclear DNA fragmentation and accumulation of higher levels of H<sub>2</sub>O<sub>2</sub> and malondialdehyde. Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-447/img_1.png<b>图1</b> 苹果酸调控植物程序性细胞死亡模式图(改自<xref ref-type="bibr" rid="b8-1674-3466-53-4-447">Zhao et al., 2018</xref>; <xref ref-type="bibr" rid="b2-1674-3466-53-4-447">Heng et al., 2018</xref>)<br/>(A) 在拟南芥中, <i>MOD1</i>编码烯脂酰还原酶, 突变体<i>mod1</i>中底物NADH在叶绿体大量积累, NADH与草酰乙酸(OAA)在plNAD-MDH的催化作用下生成苹果酸, 通过DiT1运输至细胞质, 继而被某个/某些转运蛋白转运至线粒体。苹果酸在线粒体中被mMDH1催化形成草酰乙酸, 并伴随着NADH的积累, 为线粒体膜上的mETC复合体提供电子, 产生ROS, 进而导致<i>mod1</i>突变体产生程序性细胞死亡(PCD)。(B) 水稻<i>OsALMT7</i>基因编码1个定位于质膜的苹果酸转运蛋白, 可以将苹果酸转运至细胞内。苹果酸可以维持核DNA稳定, 减少H<sub>2</sub>O<sub>2</sub>和丙二醛的积累, 从而抑制PCD。在其突变体<i>paab1-1</i>中, 苹果酸积累减少, 伴随着核DNA片段化, 同时积累H<sub>2</sub>O<sub>2</sub>和丙二醛, 促进PCD。<br/><b>Figure 1</b> A proposed model of malate-regulated programmed cell death in plants (modified from <xref ref-type="bibr" rid="b8-1674-3466-53-4-447">Zhao et al., 2018</xref>; <xref ref-type="bibr" rid="b2-1674-3466-53-4-447">Heng et al., 2018</xref>)<br/>(A) In <i>Arabidopsis thaliana</i>, <i>MOD1</i> encodes an enoyl-acyl carrier protein reductase, the deficiency of MOD1 causes an increased level of NADH in the chloroplasts, which drives oxaloacetate (OAA) to be converted to malate by plNAD-MDH. Malate is transported out of the chloroplast into the cytosol by DiT1, and then be transported into the mitochondrion by an unidentified transporter or transporters. In the mitochondrion, malate is converted to OAA by mMDH1, and simultaneously NADH is generated to provide electrons for mETC to induce ROS formation and initiate programmed cell death (PCD) process in the <i>mod1</i> cells. (B) In <i>Oryza sativa</i>, <i>OsALMT7</i> encodes a putative aluminum-activated plasma membrane localized malate transporter, which could transport malate into cells. Malate is involved in nuclear DNA stability and inhibits excessive accumulation of H<sub>2</sub>O<sub>2</sub> and malondialdehyde, which protects cells from PCD. The <i>paab1-1</i> mutant harbors a mutation in <i>OsALMT7</i>, and its panicle contained less malate than wild type, particularly at the apical portions. The apical spikelets in the <i>paab1-1</i> mutant undergo PCD accompanied by nuclear DNA fragmentation and accumulation of higher levels of H<sub>2</sub>O<sub>2</sub> and malondialdehyde.
图1 苹果酸调控植物程序性细胞死亡模式图(改自Zhao et al., 2018; Heng et al., 2018) (A) 在拟南芥中, MOD1编码烯脂酰还原酶, 突变体mod1中底物NADH在叶绿体大量积累, NADH与草酰乙酸(OAA)在plNAD-MDH的催化作用下生成苹果酸, 通过DiT1运输至细胞质, 继而被某个/某些转运蛋白转运至线粒体。苹果酸在线粒体中被mMDH1催化形成草酰乙酸, 并伴随着NADH的积累, 为线粒体膜上的mETC复合体提供电子, 产生ROS, 进而导致mod1突变体产生程序性细胞死亡(PCD)。(B) 水稻OsALMT7基因编码1个定位于质膜的苹果酸转运蛋白, 可以将苹果酸转运至细胞内。苹果酸可以维持核DNA稳定, 减少H2O2和丙二醛的积累, 从而抑制PCD。在其突变体paab1-1中, 苹果酸积累减少, 伴随着核DNA片段化, 同时积累H2O2和丙二醛, 促进PCD。 Figure 1 A proposed model of malate-regulated programmed cell death in plants (modified from Zhao et al., 2018; Heng et al., 2018) (A) In Arabidopsis thaliana, MOD1 encodes an enoyl-acyl carrier protein reductase, the deficiency of MOD1 causes an increased level of NADH in the chloroplasts, which drives oxaloacetate (OAA) to be converted to malate by plNAD-MDH. Malate is transported out of the chloroplast into the cytosol by DiT1, and then be transported into the mitochondrion by an unidentified transporter or transporters. In the mitochondrion, malate is converted to OAA by mMDH1, and simultaneously NADH is generated to provide electrons for mETC to induce ROS formation and initiate programmed cell death (PCD) process in the mod1 cells. (B) In Oryza sativa, OsALMT7 encodes a putative aluminum-activated plasma membrane localized malate transporter, which could transport malate into cells. Malate is involved in nuclear DNA stability and inhibits excessive accumulation of H2O2 and malondialdehyde, which protects cells from PCD. The paab1-1 mutant harbors a mutation in OsALMT7, and its panicle contained less malate than wild type, particularly at the apical portions. The apical spikelets in the paab1-1 mutant undergo PCD accompanied by nuclear DNA fragmentation and accumulation of higher levels of H2O2 and malondialdehyde.
万建民研究组的研究成果表明, 具有转运苹果酸功能的质膜蛋白OsALMT7的突变体pabb1-1苹果酸积累减少, 导致水稻穗顶部小花退化, 而外源补充苹果酸可以部分恢复突变体顶部小穗的退化表型(图1B)。他们证明了苹果酸可以抑制水稻穗顶部退化, 而穗顶部退化是发生在幼穗伸长后期的一个程序性细胞死亡过程(Heng et al., 2018)。研究结果为证明苹果酸参与程序性细胞死亡过程提供了重要证据, 也为水稻高产品种选育以及在生产上避免因穗顶部退化引起的减产提供了理论基础和解决思路。
MouZ, HeY, DaiY, LiuX, LiJ (2000). Deficiency in fatty acid synthase leads to premature cell death and dramatic alterations in plant morphology. Plant Cell 12, 405-418. [本文引用: 1]
[4]
NoctorG, Veljovic-JovanovicS, DriscollS, NovitskayaL, FoyerCH (2002). Drought and oxidative load in the leaves of C3 plants: a predominant role for photorespiration?Ann Bot 89, 841-850. [本文引用: 1]
[5]
Van AkenO, Van BreusegemF (2015). Licensed to kill: mitochondria, chloroplasts, and cell death.Trends Plant Sci 20, 754-766. [本文引用: 1]
ZhaoY, LuoL, XuJ, XinP, GuoH, WuJ, BaiL, WangG, ChuJ, ZuoJ, YuH, HuangX, LiJ (2018). Malate transported from chloroplast to mitochondrion triggers production of ROS and PCD in Arabidopsis thaliana. Cell Res 28, 448-461. [本文引用: 3]
1 1997
... 在一定的生理状态和生长发育过程中, 植物可以通过程序性细胞死亡(programmed cell death, PCD)维持体内稳定和平衡, 从而更好地适应环境并完成生命活动.PCD是一种受到遗传调控的主动性死亡过程, 通过调节细胞分化、细胞数量及超敏反应(HR)等, 在植物的生命活动和生物/非生物胁迫反应中发挥重要作用.尽管动、植物的PCD属性相似且具有共同的抑制因子(Vianello et al., 2007), 但由于植物含有叶绿体这一特有的亚细胞结构, 预示着植物的PCD信号通路还具有一定的独特性(Beers, 1997).因此, 解析植物PCD信号通路是当前亟待解决的重要课题.最近, 中国科学家以拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)为材料, 在解析PCD发生机制及其在生产中的应用方面取得了突破性进展(Zhao et al., 2018; Heng et al., 2018). ...
4 2018
... 在一定的生理状态和生长发育过程中, 植物可以通过程序性细胞死亡(programmed cell death, PCD)维持体内稳定和平衡, 从而更好地适应环境并完成生命活动.PCD是一种受到遗传调控的主动性死亡过程, 通过调节细胞分化、细胞数量及超敏反应(HR)等, 在植物的生命活动和生物/非生物胁迫反应中发挥重要作用.尽管动、植物的PCD属性相似且具有共同的抑制因子(Vianello et al., 2007), 但由于植物含有叶绿体这一特有的亚细胞结构, 预示着植物的PCD信号通路还具有一定的独特性(Beers, 1997).因此, 解析植物PCD信号通路是当前亟待解决的重要课题.最近, 中国科学家以拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)为材料, 在解析PCD发生机制及其在生产中的应用方面取得了突破性进展(Zhao et al., 2018; Heng et al., 2018). ... ... 苹果酸调控植物程序性细胞死亡模式图(改自Zhao et al., 2018; Heng et al., 2018) (A) 在拟南芥中, MOD1编码烯脂酰还原酶, 突变体mod1中底物NADH在叶绿体大量积累, NADH与草酰乙酸(OAA)在plNAD-MDH的催化作用下生成苹果酸, 通过DiT1运输至细胞质, 继而被某个/某些转运蛋白转运至线粒体.苹果酸在线粒体中被mMDH1催化形成草酰乙酸, 并伴随着NADH的积累, 为线粒体膜上的mETC复合体提供电子, 产生ROS, 进而导致mod1突变体产生程序性细胞死亡(PCD).(B) 水稻OsALMT7基因编码1个定位于质膜的苹果酸转运蛋白, 可以将苹果酸转运至细胞内.苹果酸可以维持核DNA稳定, 减少H2O2和丙二醛的积累, 从而抑制PCD.在其突变体paab1-1中, 苹果酸积累减少, 伴随着核DNA片段化, 同时积累H2O2和丙二醛, 促进PCD. ... ... A proposed model of malate-regulated programmed cell death in plants (modified from Zhao et al., 2018; Heng et al., 2018) (A) In Arabidopsis thaliana, MOD1 encodes an enoyl-acyl carrier protein reductase, the deficiency of MOD1 causes an increased level of NADH in the chloroplasts, which drives oxaloacetate (OAA) to be converted to malate by plNAD-MDH. Malate is transported out of the chloroplast into the cytosol by DiT1, and then be transported into the mitochondrion by an unidentified transporter or transporters. In the mitochondrion, malate is converted to OAA by mMDH1, and simultaneously NADH is generated to provide electrons for mETC to induce ROS formation and initiate programmed cell death (PCD) process in the mod1 cells. (B) In Oryza sativa, OsALMT7 encodes a putative aluminum-activated plasma membrane localized malate transporter, which could transport malate into cells. Malate is involved in nuclear DNA stability and inhibits excessive accumulation of H2O2 and malondialdehyde, which protects cells from PCD. The paab1-1 mutant harbors a mutation in OsALMT7, and its panicle contained less malate than wild type, particularly at the apical portions. The apical spikelets in the paab1-1 mutant undergo PCD accompanied by nuclear DNA fragmentation and accumulation of higher levels of H2O2 and malondialdehyde. ... ... 万建民研究组的研究成果表明, 具有转运苹果酸功能的质膜蛋白OsALMT7的突变体pabb1-1苹果酸积累减少, 导致水稻穗顶部小花退化, 而外源补充苹果酸可以部分恢复突变体顶部小穗的退化表型(图1B).他们证明了苹果酸可以抑制水稻穗顶部退化, 而穗顶部退化是发生在幼穗伸长后期的一个程序性细胞死亡过程(Heng et al., 2018).研究结果为证明苹果酸参与程序性细胞死亡过程提供了重要证据, 也为水稻高产品种选育以及在生产上避免因穗顶部退化引起的减产提供了理论基础和解决思路. ...
... 在一定的生理状态和生长发育过程中, 植物可以通过程序性细胞死亡(programmed cell death, PCD)维持体内稳定和平衡, 从而更好地适应环境并完成生命活动.PCD是一种受到遗传调控的主动性死亡过程, 通过调节细胞分化、细胞数量及超敏反应(HR)等, 在植物的生命活动和生物/非生物胁迫反应中发挥重要作用.尽管动、植物的PCD属性相似且具有共同的抑制因子(Vianello et al., 2007), 但由于植物含有叶绿体这一特有的亚细胞结构, 预示着植物的PCD信号通路还具有一定的独特性(Beers, 1997).因此, 解析植物PCD信号通路是当前亟待解决的重要课题.最近, 中国科学家以拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)为材料, 在解析PCD发生机制及其在生产中的应用方面取得了突破性进展(Zhao et al., 2018; Heng et al., 2018). ... ... 苹果酸调控植物程序性细胞死亡模式图(改自Zhao et al., 2018; Heng et al., 2018) (A) 在拟南芥中, MOD1编码烯脂酰还原酶, 突变体mod1中底物NADH在叶绿体大量积累, NADH与草酰乙酸(OAA)在plNAD-MDH的催化作用下生成苹果酸, 通过DiT1运输至细胞质, 继而被某个/某些转运蛋白转运至线粒体.苹果酸在线粒体中被mMDH1催化形成草酰乙酸, 并伴随着NADH的积累, 为线粒体膜上的mETC复合体提供电子, 产生ROS, 进而导致mod1突变体产生程序性细胞死亡(PCD).(B) 水稻OsALMT7基因编码1个定位于质膜的苹果酸转运蛋白, 可以将苹果酸转运至细胞内.苹果酸可以维持核DNA稳定, 减少H2O2和丙二醛的积累, 从而抑制PCD.在其突变体paab1-1中, 苹果酸积累减少, 伴随着核DNA片段化, 同时积累H2O2和丙二醛, 促进PCD. ... ... A proposed model of malate-regulated programmed cell death in plants (modified from Zhao et al., 2018; Heng et al., 2018) (A) In Arabidopsis thaliana, MOD1 encodes an enoyl-acyl carrier protein reductase, the deficiency of MOD1 causes an increased level of NADH in the chloroplasts, which drives oxaloacetate (OAA) to be converted to malate by plNAD-MDH. Malate is transported out of the chloroplast into the cytosol by DiT1, and then be transported into the mitochondrion by an unidentified transporter or transporters. In the mitochondrion, malate is converted to OAA by mMDH1, and simultaneously NADH is generated to provide electrons for mETC to induce ROS formation and initiate programmed cell death (PCD) process in the mod1 cells. (B) In Oryza sativa, OsALMT7 encodes a putative aluminum-activated plasma membrane localized malate transporter, which could transport malate into cells. Malate is involved in nuclear DNA stability and inhibits excessive accumulation of H2O2 and malondialdehyde, which protects cells from PCD. The paab1-1 mutant harbors a mutation in OsALMT7, and its panicle contained less malate than wild type, particularly at the apical portions. The apical spikelets in the paab1-1 mutant undergo PCD accompanied by nuclear DNA fragmentation and accumulation of higher levels of H2O2 and malondialdehyde. ...


{kind=link}
{kind=link}