王甜甜1,2, 郝怀庆1, 冯雪1, 景海春1,3,4,*, 1中国科学院植物研究所, 资源植物重点实验室, 北京 1000932东营市园林局, 东营 2570913中国科学院内蒙古草业研究中心, 呼和浩特 1501004中国科学院大学, 北京 100039 WangTiantian1,2, HaoHuaiqing1, FengXue1, JingHaichun1,3,4,*, 1Key Laboratory of Plant Resources, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China2Dongying Landscape and Greening Department, Dongying 257091, China 3Inner Mongolia Research Center for Prataculture, Chinese Academy of Sciences, Huhehaote 150100, China4University of Chinese Academy of Sciences, Beijing 100039, China 引用本文 王甜甜, 郝怀庆, 冯雪, 景海春. 植物HKT蛋白耐盐机制研究进展. 植物学报, 2018, 53(5): 710-725
Abstract: Soil salinity affects the growth of plant species, which can cause osmotic stress and ionic toxicity in the plants. Therefore, it is crucial to maintain Na+/K+ homeostasis at the cellular and whole plant levels for plant salt tolerance. The high-affinity potassium transporters (HKTs) were found to have a ability to transport and distribute Na+ and K+ in plants. Here we summarize the advance about the structure and function of the HKT proteins from glycophytes and halophytes. Further discussion is directed to genetically modified and artificially designed transport proteins for genetic engineering of salt-tolerant plants.
Key words:HKT ; ion homeostasis ; salt tolerance ; transport protein
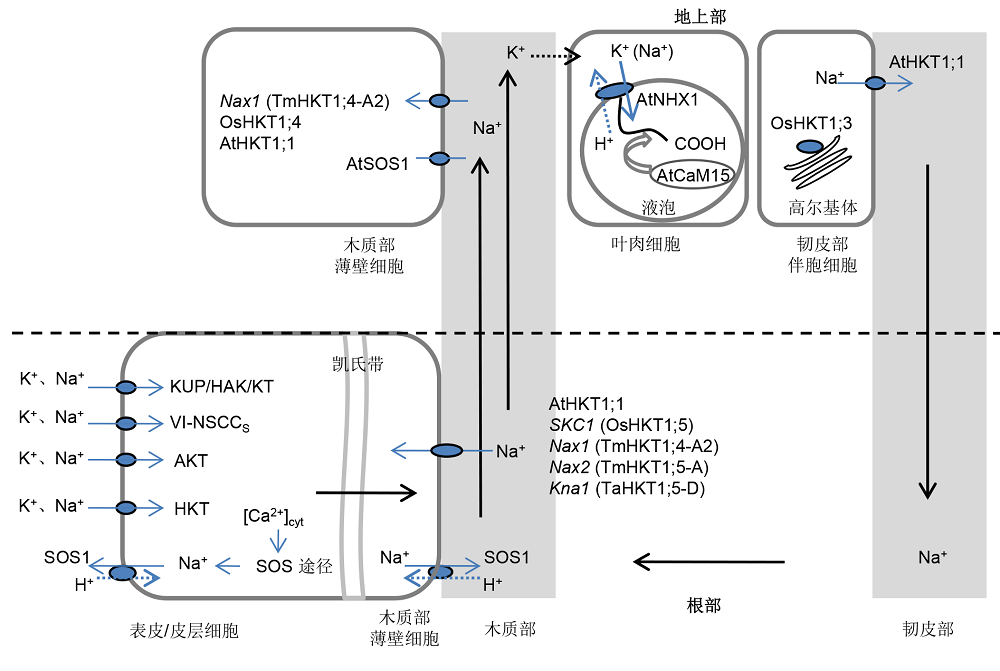
1 植物离子选择性运输的耐盐机制当外界环境NaCl含量高于40 mmol∙L-1时, 则相当于产生0.2 MPa的渗透压力, 造成植物吸水困难, 从而导致生理干旱, 影响植物正常生长(Munns and Tester, 2008)。随着外界盐离子含量的升高, Na+大量进入细胞, 同时引起K+外渗, Na+竞争执行细胞新陈代谢功能所必需酶的K+结合位点, 严重影响胞质中多种酶的活性和蛋白合成(Kronzucker and Britto, 2011)。植物通过合成渗调物质、细胞信号转导以及离子选择性运输等来响应渗透胁迫和离子毒害。同时, 植物胞质高Na+/K+比值引起胞内Ca2+浓度和pH化学势等一系列变化(Sanders et al., 1999; Tester and D- avenport, 2003), 进而影响一系列跨膜离子转运蛋白的功能。例如, HKT、质膜及液胞Na+、K+/H+反向交换体(antiporter, NHX)和KUP (K+ uptake permease)/ HAK (high-affinity K+ transporter)/KT (K+ transpor- ter)类转运蛋白活性等。图1示细胞质膜上的离子通道和转运载体在植物体内的Na+长距离运输及协同Na+/K+平衡方面的调控机制(Britto and Kronzucker, 2008; Rubio et al., 2008; Szczerba et al., 2009; Rosas- Santiago et al., 2015)。 图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-710/img_1.png<b>图1</b> 植物跨膜转运蛋白维持Na<sup>+</sup>/K<sup>+</sup>平衡的作用模型<br/>实线箭头代表钠、钾离子转运方向; 虚线箭头代表SOS1和NHX1逆向转运蛋白的H<sup>+</sup>转运方向。<br/><b>Figure 1</b> Diagram of the proposed working model for transmembrane transporters in regulating Na<sup>+</sup>/K<sup>+</sup> homeostasis<br/>The arrows in solid lines indicate the direction of Na<sup>+</sup> and K<sup>+</sup> flux, while dashed arrows indicate the direction of flux of protons (in the case of NHX1 or SOS1). Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-710/img_1.png<b>图1</b> 植物跨膜转运蛋白维持Na<sup>+</sup>/K<sup>+</sup>平衡的作用模型<br/>实线箭头代表钠、钾离子转运方向; 虚线箭头代表SOS1和NHX1逆向转运蛋白的H<sup>+</sup>转运方向。<br/><b>Figure 1</b> Diagram of the proposed working model for transmembrane transporters in regulating Na<sup>+</sup>/K<sup>+</sup> homeostasis<br/>The arrows in solid lines indicate the direction of Na<sup>+</sup> and K<sup>+</sup> flux, while dashed arrows indicate the direction of flux of protons (in the case of NHX1 or SOS1).
图1 植物跨膜转运蛋白维持Na+/K+平衡的作用模型 实线箭头代表钠、钾离子转运方向; 虚线箭头代表SOS1和NHX1逆向转运蛋白的H+转运方向。 Figure 1 Diagram of the proposed working model for transmembrane transporters in regulating Na+/K+ homeostasis The arrows in solid lines indicate the direction of Na+ and K+ flux, while dashed arrows indicate the direction of flux of protons (in the case of NHX1 or SOS1).
1.1 降低根部钠离子净积累, 提高钾离子吸收盐胁迫下, 土壤Na+随水分运输经外质体途径自由地穿过根表皮和皮层细胞的质壁间隙到达内皮层。但Na+通过质外体自由进入中柱维管组织的通路被凯氏带阻断(Enstone et al., 2002), 所以必须经过跨膜选择性运输进入共质体途径后才能被运输到木质部。植物通过吸钾排钠来维持盐胁迫下Na+/K+平衡, 因此, Na+在植物体内的累积是由被动内流和主动外流之间的平衡造成的(张宏飞和王锁民, 2007)。 非选择性的阳离子通道(nonselective cation channel, NSCC)是最初Na+内流的主要途径(Dav- enport and Tester, 2000)。钠离子通过NSCC进入细胞, 引起膜电位的去极化, 伴随着同一离子通道K+的外流。小麦LCT1是从小麦中克隆的1个可能的NSCC基因, 具有K+、Na+、Rb+、Cd2+和Ca2+的转运活性(Amtmann et al., 2001)。随后, 在电化学浓度梯度作用下, 位于膜上的离子转运载体参与钠离子的内流。然而此类离子转运蛋白一般具有较强的离子选择性。由于Na+和K+的离子半径和水合能(ion hydration)相似, Na+对于K+的吸收具有明显的竞争性抑制(Biggin et al., 2001)。 从淡土植物小麦中分离得到HKT家族的第1个成员TaHKT2;1, 以H+电化学梯度作为能量供给, 使得H+与K+同时进入细胞内。后续发现, 高亲和性钾离子转运蛋白HKT亦利用细胞膜内外的Na+电化学梯度作为K+主动转运的能量来源, 具有显著的Na+-K+协同转运特性(Rubio et al., 1995)。淡土植物高粱(Sorg- hum bicolor) SbHKT1;4蛋白在非洲爪蟾卵母细胞(Xenopuslaevis oocytes)中被证明具有显著的Na+和K+的转运特性。在拟南芥中异源表达SbHKT1;4基因, 转基因株系表现出Na+敏感特征, 但盐敏感表型可被外界升高的K+含量所缓解, 推测SbHKT1;4蛋白可在外界高钠胁迫下选择吸收K+, 与盐土植物HKT基因功能类似(Wang et al., 2014)。盐土植物盐芥可以通过TsHKT1;2在高钠胁迫下保持K+的吸收, 并且对K+的选择性高于Na+, 这是盐芥重要的耐盐机制之一(Ali et al., 2012)。盐胁迫下, 与拟南芥相比, 盐芥从根部吸收的Na+较少, 在根部组织及根部原生质体中具有较高的K+/Na+比值(Volkov et al., 2004; Wang et al., 2006)。电化学实验表明, 盐土植物赤桉(Eucalyptus camaldulensis) EcHKT1;1和EcHKT1;2蛋白对K+选择吸收能力可在外界Na+的刺激下得到加强(Liu et al., 2001)。生长至4叶期的小花碱茅(Puccinellia tenuiflora)与淡土植物小麦相比Na+单向内流速率差异显著, 小花碱茅PutHKT2;1蛋白可以在低K+和高Na+的条件下选择吸收K+, 增加K+进入共质体的数量, 从而减少Na+的净积累(Wang et al., 2004; Ardie et al., 2009)。 有研究表明, 小立碗藓(Physcomitrella patens)中Na+泵PpENA1 (Na+-ATPase, ENA)具有显著的排钠功能, 可以互补酵母(Saccharomyces cerevisiae) ena1-4突变体菌株的盐敏感表型, 从而证明低等陆生植物细胞质膜上存在Na+泵(Benito and Rodríguez- Navarro, 2003; Lunde et al., 2007)。高等植物中位于细胞膜的Na+/H+逆向转运蛋白SOS1 (Salt Overly Sensitive 1)具有重要的排钠功能。例如, 拟南芥AtSOS1基因优先在根尖表皮和木质部薄壁细胞膜表达, AtSOS1蛋白通过将Na+泵出根部细胞, 以及将Na+泵入木质部贮存, 来减少共质体途径运输至维管束的Na+含量(Shi et al., 2002)。在番茄(Solanum lycoper- slcum)中完全沉默SlSOS1基因, 则表现出根部和叶片中高Na+含量的盐敏感表型, 而特异性沉默茎部维管束中的SlSOS1基因, 则表现低Na+含量的耐盐表型, 表明SlSOS1蛋白主要在番茄根中执行排钠功能(Olías et al., 2009)。此外, SlSOS1还在根部K+跨膜吸收中具有调控保护作用(Qi and Spalding, 2004)。
1.2 控制木质部钠离子向上运输植物还可通过控制钠离子的运输及整株水平的分布来抵御钠离子的毒害。淡土植物和盐土植物均可通过排钠机制, 即控制木质部汁液中钠离子向上运输, 维持地上部较高K+/Na+比值来应答盐害胁迫。据报道,在植株整个维管束鞘都有分布的HKT转运蛋白对于Na+具有高度的选择性, 可将已进入木质部的Na+卸载在维管束薄壁细胞中, 以减少木质部汁液Na+含量, 从而降低钠离子向上运输(Mäser et al., 2002a)。例如, 拟南芥AtHKT1;1基因的作用模式是通过卸载根中木质部钠离子含量, 减少其向地上部的运输(Gong et al., 2004; Horie et al., 2009)。然而, 在近乎认定为盐土植物的大麦(Hordeum vulgare)中发现, HvHKT2;1基因过表达后, 增强了根部Na+吸收、木质部汁液及叶片Na+含量, 从而使耐盐性得到提高。这可能是由于根木质部钠离子的卸载改变了根木质部与根表皮和皮层等根外围组织之间的电位, 且这种电势的影响受外部钠或钾离子含量的调节, 尤其表现在增加外部钠离子浓度, 根部木质部与外围组织间的电化学势升高, 对根部钾离子水平的维持具有重要作用(Mian et al., 2011)。全基因组关联分析(GWAS)表明, 耐盐大麦品种中HKT1;5基因在盐胁迫下诱导表达, 通过卸载根部和叶鞘木质部Na+来减少Na+向地上部叶片运输, 从而提高大麦的耐盐性(Hazzouri et al., 2018)。有研究表明, 在木质部薄壁细胞膜表达的SOS1蛋白也具有调控木质部汁液Na+含量的功能。例如, 拟南芥AtSOS1蛋白在中等盐胁迫(如25 mmol∙L-1 NaCl)条件下, SOS1蛋白可能具有装载Na+至木质部向上运输的功能; 而在重度盐胁迫(如100 mmol∙L-1 NaCl)条件下, 在根部及地上部维管束薄壁细胞膜上表达的SOS1蛋白可能限制Na+装载入木质部, 以避免地上部Na+含量升高(Shi et al., 2002)。小花碱茅PtSOS1蛋白也被证明同样具有控制木质部汁液钠离子含量的功能(Zhang et al., 2017c)。 盐土植物也具有调控钠离子运输和分布的耐盐机制。盐土植物盐生车前(Plantago maritime)和盐敏感水车前(P. media)具有相同的根部钠离子吸收速率, 但盐生车前的耐盐性与木质部钠离子运输以及叶肉细胞液泡中贮存大量钠离子的功能有较大关系(Erdei and Kuiper, 1979)。通常情况下, 植物将已经到达地上部分的Na+转移到特定部位。植物总是优先将Na+累积在老叶片中。衰老叶片中的Na+浓度可以随着盐胁迫处理时间的延长而增加, 直至死亡, 从而保护了光合作用旺盛的新展开叶和幼嫩的部位。与成熟组织相比, 盐生植物幼嫩组织中的Na+浓度较低, 表明盐生植物和淡土植物一样, 同样需要保护代谢活跃的细胞免受Na+侵害(Flowers and Yeo, 1986)。即使是在叶片中, 钠离子的分布也有一定的规律。在同一叶片中, Na+优先积累在表皮细胞, 同时叶鞘中丰富的维管束薄壁细胞中也卸载存储了较高浓度的Na+, 以尽量减少Na+在叶肉细胞的积累, 从而保证植物的光合作用正常进行(Stelzer, 1981; Karley et al., 2000a, 2002b)。例如, 在大麦中过表达HvHKT2;1基因后其耐盐性增强, 这与Na+在叶片中特异分布的特性有关(Mian et al., 2011)。水稻OsHKT1;4基因和OsHKT1;5基因均为叶鞘维管束排钠基因, 且Os- HKT1;5蛋白还具有韧皮部排钠功能, 通过将Na+贮藏在成熟叶片来减少向幼嫩叶片的运输(Suzuki et al., 2016; Kabayashi et al., 2017)。 同时, 植物体吸收的Na+由木质部运输至地上叶片等组织中, 也有证据表明一部分Na+被韧皮部传递细胞吸收, 并分泌到韧皮部中再运回根中, 最后分泌到环境中(Berthomieu et al., 2003)。最新的研究表明, 盐胁迫下, 在拟南芥突变体tus-1地上部茎部表达的AtHKT1;1基因起主要调控作用, 通过回流茎部木质部汁液减少Na+向花器官运输, 即通过降低花器官Na+含量, 而不是降低拟南芥莲座叶片Na+含量来提高植株的耐盐性(An et al., 2017)。
1.3 植物组织耐受性机制植物组织耐受性是指将钠离子区隔化入特定组织细胞的液泡内, 从而降低离子毒害。如地上部叶片表皮细胞或维管束细胞具有发育良好的液泡, 植物体通过液泡H+-ATPase和焦磷酸酶建立起液泡内外的H+梯度, 驱动Na+/H+逆向转运蛋白, 将Na+泵到液泡贮藏, 从而减轻过量钠离子对胞质酶的伤害, 保证胞质执行正常生理功能(Blumwald, 2000)。植物(Na+, K+/H+)反向交换体(antiporter, NHX)利用质子泵产生的H+电化学梯度进行离子交换, 根据其亚细胞分布分为质膜NHX (plasma membrane NHX)、液泡NHX (vacuolar NHX)和内膜NHX (endosomal NHX)。目前, 已在包括盐土植物和淡土植物的许多物种中发现了NHX- type离子转运蛋白。盐土植物北滨藜(Atriplex gme- lini)液泡AgNHX1基因编码Na+/H+反向转运蛋白, 在盐胁迫下被诱导表达; 在水稻中异源表达AgNHX1基因可显著提高其耐盐特性, 因此推测盐胁迫下液泡NHX蛋白具有将Na+区隔化至液泡的功能(Hamada et al., 2001; Ohta et al., 2002)。OsNHX1基因编码1个液泡(Na+, K+/H+)反向交换体, 用高浓度的NaCl和KCl处理可提高OsNHX1基因在水稻茎和根中的转录水平, 过表达OsNHX1基因可显著提高水稻植株的耐盐性, 因此推测OsNHX1蛋白具有将胞质中过量积累的Na+和K+区隔化至液泡的功能(Fukuda et al., 2004)。拟南芥atnhx1突变体具有显著的盐敏感表型, 在拟南芥和油菜(Brassica napus)植株中异源表达AtNHX1基因, 可提高植株耐盐性(Apse et al., 1999; Zhang et al., 2001)。后续研究表明, 当钙调蛋白At- CaM15与液胞膜AtNHX1蛋白的C端结合, 则限制At- NHX1蛋白的Na+转运特性, 成为K+/H+反向转运蛋白; 拟南芥液泡AtNHX1蛋白可能不具有转运Na+的功能, 而在提高地上部K+含量方面发挥重要调节作用(Yamaguchi et al., 2005; Bassil et al., 2011b; Barragán et al., 2012)。例如, 在番茄中异源表达At- NHX1基因, 植株通过维持细胞K+平衡, 而非将Na+区隔化至液泡贮藏, 来提高转基因植株的耐盐性(Leidi et al., 2010)。番茄内膜LeNHX2编码K+/H+反向转运蛋白, 在番茄中过表达LeNHX2基因, 可提高盐胁迫下转基因植株对K+的吸收, 从而提高植物的耐盐性(Huertas et al., 2013)。目前, 除植物液泡NHX蛋白的功能研究较为详尽之外, 植物内膜NHX蛋白功能也有报道。拟南芥内膜AtNHX5和AtNHX6蛋白定位于高尔基体(Golgi)、反面高尔基体(TGN)和多胞体/液泡前体(MVB/PVC), 具有调节细胞pH、维持离子平衡、调控蛋白质运输及植物生长发育等方面的功能(Bassil et al., 2011a, 2012; 邱全胜, 2017)。在水稻中转化菊芋(Helianthus tuberosus) HtNHX2基因, 发现HtNHX2在转基因水稻植株中的定位类似于AtNHX5蛋白在拟南芥中的定位, 转基因水稻在盐胁迫和缺素处理下耐逆性有所提高(Zeng et al., 2018)。转化NHX基因对植物体内的哪些调控过程产生影响, 还需深入研究(Rodríguez-Rosales et al., 2009; Barragán et al., 2012; Reguera et al., 2015)。 比较盐土植物甜菜(Beta vulgaris)和淡土植物番茄(Lycopersicum esculatum)液泡膜离子通道电学特性, 发现其对Na+和K+均无特异选择性。但在甜菜中液泡膜向外正压下, 位于膜上的离子通道通透性大大降低, 防止贮存在液泡中的盐离子流入胞质, 揭示了盐土植物中位于液泡膜上离子通道的特异整流特性(Pantoja et al., 1989; Volkov, 2015)。其次, 盐生植物相对淡土植物而言, 叶片可以耐受更高的钠离子含量。盐土植物具有典型的泌盐作用, 如柽柳(Tamarix chinensis)和小花碱茅等植物叶片的盐腺, 可通过叶片泌盐的形式减少植株体内或叶片中的钠离子含量, 是存在于盐生植物中的一种特殊的耐盐机制。
2 HKT蛋白研究进展2.1 HKT蛋白种类与离子转运特性高亲和性钾离子转运蛋白是位于膜上的离子转运载体, 含有典型的TrkH保守结构域。HKT蛋白由4个跨膜MPM基序(membrane-pore-membrane motif)组成, 每个基序由2个跨膜区域和1个保守孔状P-Loop组成, 根据4个P-Loop保守位点氨基酸的不同可分为Gly-Gly-Gly-Gly和Ser-Gly-Gly-Gly两种类型。第1个MPM基序PA-Loop保守位点氨基酸决定了蛋白离子选择性。以蛋白氨基酸序列的比对为基础, HKT基因家族聚类出3个亚家族(图2)。植物HKT-type类蛋白最先被发现聚类关系较近的有亚家族1和亚家族2, 亚家族1成员属于Ser-Gly-Gly-Gly类型, 来源于双子叶植物, 具有钠离子转运特性; 亚家族2成员属于Gly- Gly-Gly-Gly类型, 来源于单子叶植物, 具有Na+-K+协同转运特性(Mäser et al., 2002b), 其中OsHKT2;1例外, 为Ser-Gly-Gly-Gly保守特征, 具有Na+转运特性; 亚家族3分支中含有原始的苔藓植物及裸子植物, 如小立碗藓和江南卷柏(Selaginella moellendorffii)的HKT-type类蛋白, 也属于Gly-Gly-Gly-Gly保守类型。但有研究表明, 有关亚家族成员的Na+、K+转运特性与PA-Loop保守位点氨基酸的分类结果不完全一致(Rodríguez-Navarro, 2000; Mäser et al., 2002b; Oomen et al., 2012)。 图2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-710/img_2.png<b>图2</b> 植物HKT蛋白家族最小进化关系聚类树(MEGA7)<br/>氨基酸序列登录号见附录1。<br/><b>Figure 2</b> An un-rooted minimum-evolution phylogenetic tree of protein sequences of HKT homologues (MEGA7)<br/>The amino acid sequences are in Appendix 1. Figure 2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-710/img_2.png<b>图2</b> 植物HKT蛋白家族最小进化关系聚类树(MEGA7)<br/>氨基酸序列登录号见附录1。<br/><b>Figure 2</b> An un-rooted minimum-evolution phylogenetic tree of protein sequences of HKT homologues (MEGA7)<br/>The amino acid sequences are in Appendix 1.
图2 植物HKT蛋白家族最小进化关系聚类树(MEGA7) 氨基酸序列登录号见附录1。 Figure 2 An un-rooted minimum-evolution phylogenetic tree of protein sequences of HKT homologues (MEGA7) The amino acid sequences are in Appendix 1.
小麦中钠离子转运相关的氨基酸功能位点已有报道, TaHKT2;1蛋白A240V、L247F、Q270L、N365S和E464Q位点氨基酸的替换, 显著影响钠离子的转运功能(Rubio et al., 1995, 1999; Diatloff et al., 1998)。TaHKT2;1蛋白519位精氨酸和AtHKT1;1蛋白487位精氨酸(Arg)突变成丙氨酸(Ala)、谷氨酰胺(Gln)、谷氨酸(Glu), 钠离子转运功能显著下降(Kato et al., 2007)。Henderson等(2017)研究表明, 小麦TaHKT1;5-D蛋白和葡萄(Vitis vinifera) VisHKT1;1蛋白的第8个跨膜区域均与钠离子选择转运特性密切相关。水稻耐盐籼稻品种Nona Bokra OsHKT1;5蛋白钠离子转运特性与395位缬氨酸(Val)密切相关, 而盐敏感品种日本晴(Nipponbare) OsHKT1;5蛋白395位为亮氨酸(Leu), 由于该位点与其它潜在的功能位点相互作用, 进而影响钠离子的选择特性(Cotsaftis et al., 2012)。 亚家族1 (图2)中AtHKT1;1与TsHKT1;2聚类关系较近, 在PA-Loop保守位点均是丝氨酸(Ser), 但TsHKT1;2具有钾离子转运特性。所以, HKT-type的离子转运蛋白的K+选择性也不只由MPM基序中的保守氨基酸决定。双子叶模式植物拟南芥仅含1个HKT基因, 拟南芥AtHKT1;1不能互补K+吸收缺陷型酵母trk1和trk2的功能。非洲爪蟾卵母细胞膜电化学实验显示, AtHKT1;1表达可引起较强的Na+流变化, 表明AtHKT1;1为Na+特异性转运载体而非K+转运载体(Uozumi et al., 2000; Kato et al., 2001)。盐芥与拟南芥亲缘关系较近, 具有较强耐盐特性, 盐芥TsHKT1;2为高亲和钾离子转运蛋白, 与盐生植物赤桉Ec- HKT1;2离子转运特性类似。TsHKT1;2蛋白K+选择特性与2nd P-Loop区域即207和238两个位点的天冬氨酸Asp (D)密切相关, 而亚家族1大多数成员(包括拟南芥)此保守位置氨基酸均是谷氨酰胺Asn (N), AtHKT1;1蛋白中N突变为D, 其钠离子转运特性转变为钠-钾协同转运特性(Ali et al., 2012; Ali and Yun, 2016)。电化学实验显示, AtHKT1;1可介导Na+内向和外向整流, 而TsHKT1;2只具有较强的内向整流特性。推测在物种进化过程中, HKT蛋白在淡土植物和盐土植物中分化出不同的功能, 从而适应物种的生理特性和生存环境。 亚家族2 (图2)成员不仅具有Na+-K+协同转运特性, 在外界环境影响下, 还可作为Na+或K+单向转运体。HKT家族中最早在小麦中分离获得TaHKT2;1基因为亚家族2成员, 在外界低钠条件下, TaHKT2;1为Na+-K+协同转运; 当处于高Na+条件下, TaHKT2;1为Na+单向转运体(Rubio et al., 1995; Gassmann et al., 1996)。水稻亚家族2成员OsHKT2;1与OsHKT2;2的氨基酸序列同源性高达91%, 然而OsHKT2;1是Na+转运蛋白, OsHKT2;2则是K+/Na+协同转运蛋白(Horieet al., 2001)。OsHKT2;1的PA-loop区域保守氨基酸是Ser, 而在OsHKT2;2中是Gly, 当两者的Gly/Ser互换后, 它们的转运活性也相应转换(Mäser et al., 2002b)。在非洲爪蟾卵母细胞电化学实验中证明, OsHKT2;4蛋白是K+单向转运体, 而不是Na+-K+协同转运, K+转运特性不需要胞外Na+的激活, 这与水稻HKT亚家族2的其它成员不同, 揭示了亚家族2另一类不同功能。同时, 当外界Na+浓度较高(>10 mmol∙L-1)时, OsHKT2;4也可作为Na+转运蛋白(Sassi et al., 2012)。
2.2 HKT蛋白的定位和表达HKT基因的表达具有组织特异性, AtHKT1;1基因主要在根部及叶片维管束组织中表达。免疫电镜及At- HKT1;1启动子-GUS报告基因表达分析表明, AtHKT- 1;1基因在叶片木质部薄壁细胞膜上具有较强的表达信号, 在韧皮部附近也有微弱的表达信号。将AtHKT1;1基因在维管束组织特异性表达后, 可以互补athkt1;1突变体盐敏感表型, 而组成型表达的转基因植株仍然表现出盐敏感表型(Mäser et al., 2002a; Møller et al., 2009)。以上表明AtHKT1;1是通过卸载木质部Na+来调节木质部与其外围的薄壁细胞组织之间的渗透平衡, 最终降低叶肉细胞中的Na+含量, 保护叶片免受盐害胁迫(Sunarpi et al., 2005)。AtHKT1;1基因是否在韧皮部发挥调控离子转运的功能? 已有研究表明, AtHKT1;1基因转录本在韧皮部特异表达, AtHKT1;1基因与韧皮部Na+含量密切相关, athkt1;1突变体韧皮部汁液中Na+含量显著降低, 推测AtHKT1;1将韧皮部伴胞细胞中的Na+运输至韧皮部, 介导Na+通过地上部韧皮部向根部回流(Berthomieu et al., 2003)。然而,利用22Na+放射性标记, 测定盐胁迫下拟南芥athkt1-1突变体植株Na+在体内的运输, 发现AtHKT1;1基因通过卸载根部木质部Na+使根中钠离子含量升高, 而不介导根部钠离子的吸收和地上部韧皮部的回流过程(Davenport et al., 2007)。 不同植物HKT基因表达部位不同, 水稻HKT成员在根、茎及叶鞘等维管束组织, 以及根表皮、皮层和叶片韧皮部等部位表达。水稻是目前已发现的HKT基因家族成员最多的植物, 在粳稻品种日本晴的基因组中已克隆出9个成员(包括2个假基因OsHKT1;2和OsHKT2;2), 其中亚家族1中有5个: OsHKT1;1、OsHKT1;2、OsHKT1;3、OsHKT1;4和OsHKT1;5; 亚家族2中有4个: OsHKT2;1、OsHKT2;2、OsHKT2;3和OsHKT2;4 (Garciadeblas et al., 2003)。 水稻HKT亚家族1成员表达方式各异, OsHKT1;1基因主要在叶片韧皮部表达, 具有Na+转运特性, oshkt1;1盐敏感突变体地上部Na+大量积累, 韧皮部汁液Na+含量降低。盐胁迫导致水稻老叶OsHKT1;1基因表达量上调, 推测是通过在老叶中积累Na+以保护幼叶正常生长(Wang et al., 2012)。后续研究表明, OsHKT1;1基因的表达还受转录因子的调节, OsMYBc转录因子调节OsHKT1;1基因的表达, 进而缓解植株遭受离子毒害的症状(Wang et al., 2015)。通过对390个水稻品种在中等盐胁迫下进行耐盐性分析, 发现籼稻(Indica rice)品种根部Na+含量较高, 而粳稻(Japonica rice)品种根部Na+含量较低。全基因组关联分析(genome-wide association, GWAS)表明, chr.4约575 kb序列(RNC4)与根部Na+含量关联度最高, 进一步鉴定出OsHKT1;1和OsHKT1;4两个候选基因。所分析的籼稻品种中存在3个高频次的OsHKT- 1;1基因非同义突变, 通过异源表达和RNA干扰实验表明, OsHKT1;1基因可调控水稻根部Na+含量, 进而使籼稻和粳稻品种根部Na+含量存在显著差异(Camp- bell et al., 2017)。 OsHKT1;3基因在叶、叶鞘及茎基部维管束中均有表达, 且在韧皮部中的表达量高于木质部。Os- HKT1;3-RNAi植株在潮霉素处理后, 具有明显的生长受抑制表型。研究表明, OsHKT1;4基因主要在地上部维管束组织表达(Suzuki et al., 2016), OsHKT1;5基因在根部和叶鞘木质部薄壁细胞膜上表达, 均具有卸载木质部Na+和减少地上部和叶片钠离子积累的作用。22Na+放射性标记实验表明, OsHKT1;5蛋白还在成熟叶片韧皮部中表达, 具有韧皮部排钠功能, 以降低钠离子向幼嫩叶片的运输(Kabayashi et al., 2017)。 水稻亚家族2中4个HKT成员在进化聚类中属于2个分支的同源基因, 其中一个分支包括OsHKT2;1与OsHKT2;2, 基因同源性较高; 另一分支为进化关系较近的OsHKT2;3与OsHKT2;4。OsHKT2;1在根的表皮和皮层细胞中表达, 水稻根部对Na+高亲和吸收的动力学特征与OsHKT2;1转运Na+的特征非常吻合(Garciadeblas et al., 2003; Kader and Lindberg, 2008)。盐胁迫下, OsHKT2;1表达量降低, 所以Os- HKT2;1是目前被认为最有可能参与根部Na+吸收的水稻HKT转运蛋白。小麦TaHKT2;1基因主要在根和叶鞘等中表达, 叶片中表达量较少。TaHKT2;1-RNAi植株根部Na+含量降低, 转基因植物耐盐性增强, 推测小麦TaHKT2;1基因具有吸收根部Na+的功能(Laurie et al., 2002)。不同的是, TaHKT2;1基因低亲和性的钠离子吸收可以被钾离子激活; OsHKT2;1钠离子转运活性则被外界高钾离子所抑制, 丧失所有的离子转运活性; 而在K+缺少的情况下转运Na+作为矿质营养素以利于植株生长(Horie et al., 2007; Yao et al., 2010)。 OsHKT2;2的全长cDNA序列是在另一个耐盐水稻品种Pokkali中分离获得。盐胁迫前, OsHKT2;2在地上部的表达部位主要为叶肉细胞; 盐胁迫后, OsHKT2;2在韧皮部的表达显著增强(Kader et al., 2006)。不同品种间水稻耐盐性差异较大, 在耐盐水稻品种(Nona Bokra)中鉴定出HKT基因新结构OsHKT2;2/1, 是由OsHKT2;2基因的5′端部分序列和OsHKT2;1基因的3′端部分序列组成, 在PA-Loop网状区域的关键位点保守氨基酸为甘氨酸(Gly)。与OsHKT2;1基因的表达不同, No-OsHKT2;2/1基因在外界高钠离子胁迫下有很高的表达量。结合在异源酵母与蛙卵细胞中的实验表明, No-OsHKT2;2/1基因在盐胁迫下对钾离子的选择吸收是水稻品种(Nona Bokra)较耐盐的重要原因(Oomen et al., 2012)。该研究表明, 在水稻不同品种进化过程中, HKT基因结构也随之发生变化。 OsHKT2;3在根、茎基部、叶和叶鞘中均有表达, 但在电生理实验中未发现其离子转运功能, 在拟南芥中异源表达OsHKT2;3基因也无任何表型特征, 由此证明OsHKT2;3可能是1个没有功能的蛋白。而OsHKT2;4基因在根表皮细胞以及茎、节间、叶鞘的维管束木质部和韧皮部中表达。前文所述OsHKT2;4具有较高的K+转运特性, 而Na+转运能力相对较低(Sassi et al., 2012)。通过将OsHKT2;4基因转化拟南芥突变体atmgt6和爪蟾卵母细胞, 发现OsHKT2;4基因还具有显著的镁离子转运功能, 在植物体内具有类似MGT-type类镁离子转运蛋白的功能(Zhang et al., 2017a)。 进一步利用比较基因组学研究方法分离和鉴定不同物种间同源基因的功能, 发现小麦亚家族2成员与水稻类似, 也具有2类分支的同源基因, 分别是TaHKT2;1基因和新分离的TaHKT2;2基因。TaHKT- 2;2位于7L染色体上, 与TaHKT2;1基因物理距离仅隔3 kb左右, 2个基因具有类似的功能。TaHKT2;1与Ta- HKT2;2表达谱不同, TaHKT2;2除了在根和叶鞘中表达, 还在叶片中表达, 这与水稻中同源基因OsHKT2;4的表达模式类似, 即均在地上部叶鞘和叶片中表达量较高, 说明HKT基因在不同物种间行驶类似的功能, 且具有不同的生物进化过程。与已有的实验结果不同, 7AL染色体上的TaHKT2;2与TaHKT2;1基因位于控制小麦地上部或叶片钠离子含量及产量的QTL位点内, 推测二者可能不参与从外界环境中吸收钠离子, 而是在Na+从根部运输至叶片或K+运输中起作用(Ariyarathna et al., 2014, 2016)。
2.3 HKT基因的QTL定位最初HKT蛋白的研究多集中在HKT分离克隆、离子选择性、表达和定位研究, 后续通过构建杂交群体, 在水稻、小麦、番茄、玉米(Zea mays)和葡萄等植物中均精确定位了耐盐主效QTL位点(含HKT转运蛋白)。 截至目前, 水稻HKT家族含有2个参与调控Na+在根-冠分配的基因, 分别是OsHKT1;5和OsHKT- 1;4。已在耐盐水稻(Nona Bokra)与粳稻品种越光(Koshihikari)杂交群体中定位到重要的耐盐QTL位点SKC1。该段序列是1个HKT类的基因OsHKT1;5, 通过膜片钳及非洲爪蟾卵母细胞的系统证明, 其编码Na+转运载体蛋白, 在根中木质部薄壁细胞膜表达, 通过卸载木质部中钠离子并限制其向地上部运输降低地上部Na+含量, 进而间接影响K+的含量, 而维持适宜的K+/Na+平衡状态(Ren et al., 2005)。粳稻品种OsHKT1;4基因在非洲爪蟾卵母细胞和酵母异源表达系统中均被证明具有显著的Na+转运能力, EGFP- OsHKT1;4融合蛋白定位在原生质体细胞膜。Os- HKT1;4在营养生长阶段主要在叶鞘中表达, 在生殖生长阶段节间II和花序梗处表达量高。生殖生长阶段OsHKT1;4-RNAi植株遭受盐胁迫, 使其地上部尤其在叶片和叶鞘中Na+过度积累, 而在营养生长阶段变化不明显。22Na+标记实验表明, OsHKT1;4在生殖生长阶段卸载茎部和叶鞘Na+, 有助于Na+从叶片排出, 而在营养生长阶段其功能不显著(Suzuki et al., 2016)。但盲目地通过增强OsHKT1;4基因在叶片的排钠功能来进行耐盐作物育种, 不仅不会提高植株的耐盐性, 还可能会造成植株根部大量吸收Na+, 因此其在育种中的应用尚需进一步研究(Oda et al., 2018)。水稻种质耐盐性存在较大差异(吴家富等, 2017), 利用耐盐水稻品种Hasawi的3个作图群体, 鉴定出Hasawi中75个耐盐QTLs, 表明水稻Hasawi品种含有的优异耐盐等位基因还有待挖掘(Bimpong et al., 2014; Bizimana et al., 2017; Rahman et al., 2017)。 在小麦中已发现3个耐盐主效QTLs, 均为亚家族1成员, Nax1包括2个基因, 命名为TmHKT1;4-A1和TmHKT1;4-A2 (Lindsay et al., 2004); Nax2命名为TmHKT1;5-A (Munns et al., 2012); Kna1命名为TaHKT1;5 (Huang et al., 2006; Byrt et al., 2007), 其与拟南芥AtHKT1;1和水稻OsHKT1;5的同源性较高且功能相似。 栽培小麦(T. aestivum)中定位的耐盐位点Kna1位于4DL染色体, 可选择性转运K+和Na+到地上部, 使叶片K+/Na+比值升高, 利于生长(Dubcovsky et al., 1996)。在硬质小麦(T. turgidum subsp. durum) Line 149品系中定位到控制叶片钠离子含量的小麦排钠基因Nax1和Nax2。Nax1位点内2个HKT1;4基因来源于T. monococcum, 位于2AL染色体上, 被证明具有卸载地上部维管束中Na+功能, 将卸载的钠离子贮藏在叶鞘, 从而降低叶片中钠离子含量; 同时将地上部Na+通过韧皮部再循环至根中(James et al., 2006)。例如, TmHKT1;4-A2基因(而非TmHKT1;4-A1基因)在根和叶鞘中表达, 表达谱与其排钠功能相吻合(Huang et al., 2006)。硬质小麦基因组(AABB)中共有5个类似HKT1;4基因, 其中2个在A基因组, 3个在B基因组, 不同基因组来源的HKT1;4具有显著的功能变异。在硬质小麦中分离出2个HKT1;4类型的cDNA, 在非洲爪蟾卵母细胞中验证TdHKT1;4-1与TdHKT1;4-2功能, 发现二者具有不同的Na+转运活性, TdHKT1;4-2对Na+转运能力较强(Ben Amar et al., 2014)。 与Nax1不同, Nax2位点位于5AL染色体, 此位点具有卸载根木质部钠离子含量、限制其往地上部分运输的功能, 进而减少地上部钠离子含量, 保持叶片高K+/Na+比值。Nax2与Kna1同源性较高且功能类似, 它们均编码HKT1;5类蛋白(Byrt et al., 2007)。Nax2位点TmHKT1;5-A基因编码1个定位在根部维管束组织外围细胞的Na+特异性转运跨膜蛋白。田间实验表明, 与缺失Nax2位点的近等基因系相比, TmHKT1;5-A可 以显著降低叶片中钠离子含量, 使硬质小麦的产量 提高25% (Munns et al., 2012)。将TmHKT1;4-A2 与TmHKT1;5-A基因转入普通栽培小麦中, 发现二者均可显著降低叶片的钠离子含量(James et al., 2011)。 栽培小麦Bread wheat控制耐盐的Kna1位点中, TaHKT1;5-D基因在限制根部钠离子向地上部运输中起重要调控作用。盐胁迫下, 沉默TaHKT1;5-D 基因的转基因株系其Na+含量显著上升(Byrt et al., 2014)。 除在水稻和小麦中通过QTL精准定位到HKT基因之外, 在玉米、番茄和葡萄中也分离得到包含HKT基因的抗盐QTL位点。在番茄中定位到控制叶片Na+/K+平衡的耐盐主效位点kc7.1, 含有HKT1;1和HKT1;2两个候选基因。栽培番茄(S. lycopersicum)和野生契斯曼尼番茄(S. cheesmaniae) 2个近等基因系, 含有纯合HKT1基因位点, 沉默ScHKT1;2和SlHKT1;2植株Na+/K+比值升高, 表现盐敏感表型; 而沉默SlHKT1;1或ScHKT1;1基因, 植株叶片Na+/K+比值变化不大(Jaime-Pérez et al., 2017)。以玉米Zheng58/Chang7-2 RIL群体为材料, 克隆了调控叶片Na+含量的主效QTL基因ZmNC1。ZmNC1编码离子转运蛋白ZmHKT1;5, 在蛙卵电化学实验中被证明具有典型的Na+转运特性, 它主要在根中柱组织表达, 通过减少木质部导管中的Na+浓度来抑制Na+从根向地上部运输, 从而促进玉米抗盐(Zhang et al., 2017b)。利用葡萄种间杂交群体图位克隆盐胁迫下控制叶片Na+含量的主效QTL位点NaE, 该位点内含有6个HKT基因, 通过表达模式分析和异源表达功能验证得出VisHKT1;1基因为控制叶片钠离子含量的功能基因(Henderson et al., 2017)。
2.4 HKT基因的分子调控截至目前, 对HKT基因的耐盐性功能已有许多报道, 然而, HKT基因所编码蛋白的调控机制仍不清楚。关于HKT蛋白分子调控的研究主要包括表观遗传研究、转录因子调节HKT基因表达以及与HKT蛋白互作等方面。 研究表明, 拟南芥AtHKT1;1基因启动子区域存在调控其表达的重复序列和调控因子。筛选获得AtHKT1;1基因2种不同生态型突变ts-1和tsu-1, 是由AtHKT1;1前约700 bp的随机重复序列的缺失造成的, 导致其根部AtHKT1;1基因表达量显著下降, 整株表现出地上部钠离子含量较高的盐敏感表型(Rus et al., 2006)。随后发现, AtHKT1;1基因启动子上游2.6 kb处存在甲基化现象, 推测该区域可能与小RNA结合, 存在DNA的高度甲基化修饰。当甲基化修饰丧失后, AtHKT1;1表达量下降, 同时发现重复序列3′端后半部分可作为增强子提高AtHKT1;1的表达(Baek et al., 2011)。Kumar等(2017)报道, 盐胁迫可诱导小麦Ta- HKT2;1与TaHKT2;3基因编码区发生胞嘧啶(cyto- sine)甲基化, 导致其在根及地上部的表达量下降。 Wang等(2015)报道, 水稻OsMYBC转录因子可与OsHKT1;1基因启动子区域结合, 调控OsHKT1;1基因表达。敲除OsMYBC后, OsHKT1;1基因在盐胁迫下的表达量降低, 植株表现出盐敏感表型。OsHKT1;3基因编码Na+转运蛋白, 在韧皮部表达(Jabnoune et al., 2009), 亚细胞定位在高尔基体。研究表明, 在水稻中发现HKT互作蛋白, 通过膜蛋白相互作用检测方法(mating-based split-ubiquitin system)及双分子荧光互补实验, 证明OsCNIH1与OsHKT1;3在内质网发生相互作用, 介导从内质网向高尔基体的物质运输(Garciadeblas et al., 2003; Rosas-Santiago et al., 2015)。 有研究表明, 盐胁迫下HKT基因参与调控植物侧根生长发育。例如, 通过全基因组关联分析发现了100个遗传位点控制拟南芥根系结构RSA (root system architecture), 仅HKT1和CYP79B2两个基因控制侧根生长。过表达AtHKT1;1基因的拟南芥, 在盐胁迫下侧根的生长和发育受到抑制, 但随着外界K+含量的升高, 侧根的生长得到恢复, 推测AtHKT1;1基因调控的Na+/K+平衡与侧根生长密切相关(Julko- wska et al., 2017)。
ApseMP, AharonGS, SneddenWA, BlumwaldE (1999). Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis.Science 285, 1256-1258. [本文引用: 1]
[9]
ArdieSW, XieLN, TakahashiR, LiuSK, TakanoT (2009). Cloning of a high-affinity K+ transporter gene PutHKT2;1 from Puccinellia tenuiflora and its functional comparison with OsHKT2;1 from rice in yeast and Arabidopsis. J Exp Bot 60, 3491-3502. [本文引用: 1]
[10]
AriyarathnaHACK, OldachKH, FranckiMG (2016). A comparative gene analysis with rice identified orthologous group II HKT genes and their association with Na+ concen- tration in bread wheat.BMC Plant Biol 16, 21. [本文引用: 1]
[11]
AriyarathnaHCCK, Ul-HaqT, ColmerTD, FranckiMG (2014). Characterization of the multigene family TaHKT 2;1 in bread wheat and the role of gene members in plant Na+ and K+ status. BMC Plant Biol 14, 159. [本文引用: 1]
[12]
BaekD, JiangJF, ChungJS, WangBS, ChenJP, XinZG, ShiHZ (2011). Regulated AtHKT1 gene expression by a distal enhancer element and DNA methylation in the promoter plays an important role in salt tolerance. Plant Cell Physiol 52, 149-161. [本文引用: 1]
[13]
BarragánV, LeidiEO, AndrésZ, RubioL, De LucaA, FernándezJA, CuberoB, PardoJM (2012). Ion exchangers NHX1 and NHX2 mediate active potassium uptake into vacuoles to regulate cell turgor and stomatal function in Arabidopsis.Plant Cell 24, 1127-1142. [本文引用: 2]
[14]
BassilE, CokuA, BlumwaldE (2012). Cellular ion homeostasis: emerging roles of intracellular NHX Na+/H+ antiporters in plant growth and development.J Exp Bot 63, 5727-5740. [本文引用: 1]
[15]
BassilE, OhtoMA, EsumiT, TajimaH, ZhuZ, CagnacO, BelmonteM, PelegZ, YamaguchiT, BlumwaldE (2011a). The Arabidopsis intracellular Na+/H+ antiporters NHX5 and NHX6 are endosome associated and necessary for plant growth and development.Plant Cell 23, 224-239. [本文引用: 1]
[16]
BassilE, TajimaH, LiangYC, OhtoMA, UshijimaK, NakanoR, EsumiT, CokuA, BelmonteM, BlumwaldE (2011b). The Arabidopsis Na+/H+ antiporters NHX1 and NHX2 control vacuolar pH and K+ homeostasis to regulate growth, flower development, and reproduction.Plant Cell 23, 3482-3497. [本文引用: 1]
[17]
Ben AmarS, BriniF, SentenacH, MasmoudiK, VéryAA (2014). Functional characterization in Xenopus oocytes of Na+ transport systems from durum wheat reveals diversity among two HKT1;4 transporters. J Exp Bot 65, 213-222. [本文引用: 1]
[18]
BenitoB, Rodríguez-NavarroA (2003). Molecular cloning and characterization of a sodium-pump ATPase of the moss Physcomitrella patens. Plant J 36, 382-389. [本文引用: 1]
[19]
BerthomieuP, ConéjéroG, NublatA, BrackenburyWJ, LambertC, SavioC, UozumiN, OikiS, YamadaK, CellierF, GostiF, SimonneauT, EssahPA, TesterM, VéryAA, SentenacH, CasseF (2003). Functional analysis of AtHKT1 in Arabidopsis shows that Na+ recirculation by the phloem is crucial for salt tolerance. EMBO J 22, 2004-2014. [本文引用: 2]
[20]
BigginPC, SmithGR, ShrivastavaI, ChoeS, SansomMSP (2001). Potassium and sodium ions in a potassium channel studied by molecular dynamics simulations.Biochim Biophys Acta 1510, 1-9. [本文引用: 1]
[21]
BimpongIK, MannehB, DiopB, GhislainK, SowA, AmoahNKA, GregorioG, SinghRK, OrtizR, WopereisM (2014). New quantitative trait loci for enhancing adaptation to salinity in rice from Hasawi, a Saudi landrace into three African cultivars at the reproductive stage.Euphytica 200, 45-60. [本文引用: 1]
[22]
BizimanaJB, Luzi-KihupiA, MuroriRW, SinghRK (2017). Identification of quantitative trait loci for salinity tolerance in rice (Oryza sativa L.) using IR29/Hasawi mapping population. J Genet 96, 571-582. [本文引用: 1]
[23]
BlumwaldE (2000). Sodium transport and salt tolerance in plants.Curr Opin Cell Biol 12, 431-434. [本文引用: 1]
[24]
BrittoDT, KronzuckerHJ (2008). Cellular mechanisms of potassium transport in plants.Physiol Plantarum 133, 637-650. [本文引用: 1]
[25]
ByrtCS, PlattenJD, SpielmeyerW, JamesRA, LagudahES, DennisES, TesterM, MunnsR (2007). HKT1;5-like cation transporters linked to Na+ exclusion loci in wheat,Nax2 and Kna1. Plant Physiol 143, 1918-1928. [本文引用: 2]
CampbellMT, BandilloN, Al ShiblawiFRA, SharmaS, LiuK, DuQ, SchmitzAJ, ZhangC, VeryAA, LorenzAJ, WaliaH (2017). Allelic variants of OsHKT1;1 underlie the divergence between indica and japonica subspecies of rice(Oryza sativa) for root sodium content. PLoS Genet 13, e1006823. [本文引用: 1]
[28]
CotsaftisO, PlettD, ShirleyN, TesterM, HrmovaM (2012). A two-staged model of Na+ exclusion in rice explained by 3D modeling of HKT transporters and alternative splicing.PLoS One 7, e39865. [本文引用: 1]
[29]
DavenportRJ, Muñoz-MayorA, JhaD, EssahPA, RusA, TesterM (2007). The Na+ transporter AtHKT1;1 controls retrieval of Na+ from the xylem in Arabidopsis.Plant Cell Environ 30, 497-507. [本文引用: 1]
DiatloffE, KumarR, SchachtmanDP (1998). Site directed mutagenesis reduces the Na+ affinity of HKT1, an Na+ energized high affinity K+ transporter.FEBS Lett 432, 31-36. [本文引用: 1]
[32]
DubcovskyJ, MaríaGS, EpsteinE, LuoMC, DvořákJ (1996). Mapping of the K+/Na+ discrimination locusKna1 in wheat. Thero Appl Genet 92, 448-454. [本文引用: 1]
[33]
EnstoneDE, PetersonCA, MaFS (2002). Root endodermis and exodermis: structure, function, and responses to the environment.J Plant Growth Regul 21, 335-351. [本文引用: 1]
[34]
ErdeiL, KuiperPJC (1979). The effect of salinity on growth, cation content, Na+-uptake and translocation in salt-sensi- tive and salt-tolerant Plantago species. Physiol Plant 47, 95-99. [本文引用: 1]
[35]
FlowersTJ, YeoAR (1986). Ion relations of plants under drought and salinity.Aust J Plant Physiol 13, 75-91. [本文引用: 1]
[36]
FukudaA, NakamuraA, TagiriA, TanakaH, MiyaoA, HirochikaH, TanakaY (2004). Function, intracellular localization and the importance in salt tolerance of a vacuolar Na+/H+ antiporter from rice.Plant Cell Physiol 45, 146-159. [本文引用: 1]
[37]
GarciadeblasB, SennME, BañuelosMA, Rodríguez- NavarroA (2003). Sodium transport and HKT transporters: the rice model.Plant J 34, 788-801. [本文引用: 4]
[38]
GassmannW, RubioF, SchroederJI (1996). Alkali cation selectivity of the wheat root high-affinity potassium transporter HKT1.Plant J 10, 869-882. [本文引用: 1]
[39]
GongJM, WanerDA, HorieT, LiSL, HorieR, AbidKB, SchroederJI (2004). Microarray-based rapid cloning of an ion accumulation deletion mutant in Arabidopsis thaliana. Proc Natl Acad Sci USA 101, 15404-15409. [本文引用: 1]
[40]
HamadaA, ShonoM, XiaT, OhtaM, HayashiY, TanakaA, HayakawaT (2001). Isolation and characterization of a Na+/H+ antiporter gene from the halophyte Atriplex gmelini. Plant Mol Biol 46, 35-42. [本文引用: 1]
[41]
HazzouriKM, KhraiweshB, AmiriKMA, PauliD, BlakeT, ShahidM, MullathSK, NelsonD, MansourAL, Salehi-AshtianiK, PuruggananM, MasmoudiK (2018). Mapping of HKT1;5 gene in barley using GWAS approach and its implication in salt tolerance mechanism. Front Plant Sci 9, 156. [本文引用: 1]
[42]
HendersonSW, DunlevyJD, WuY, BlackmoreDH, WalkerRR, EdwardsEJ, GillihamM, WalkerAR (2017). Functional differences in transport properties of natural HKT1;1 variants influence shoot Na+ exclusion in grapevine rootstocks.New Phytol 217, 1113-1127. [本文引用: 2]
[43]
HorieT, CostaA, KimTH, HanMJ, HorieR, LeungHY, MiyaoA, HirochikaH, AnG, SchroederJI (2007). Rice OsHKT2;1 transporter mediates large Na+ influx component into K+-starved roots for growth.EMBO J 26, 3003-3014. [本文引用: 1]
HorieT, YoshidaK, NakayamaH, YamadaK, OikiS, ShinmyoA (2001). Two types of HKT transporters with different properties of Na+ and K+ transport in Oryza sativa. Plant J 27, 129-138. [本文引用: 1]
[46]
HuangSB, SpielmeyerW, LagudahES, JamesRA, PlattenJD, DennisES, MunnsR (2006). A sodium transporter (HKT7) is a candidate for Nax1, a gene for salt tole- rance in durum wheat. Plant Physiol 142, 1718-1727. [本文引用: 2]
[47]
HuertasR, RubioL, CagnacO, García-SánchezMJ, AlchéJD, VenemaK, FernándezJA, RodríguezrosalesMP (2013). The K+/H+ antiporter LeNHX2 increases salt tolerance by improving K+ homeostasis in transgenic tomato.Plant Cell Environ 36, 2135-2149. [本文引用: 1]
[48]
JabnouneM, EspeoutS, MieuletD, FizamesC, VerdeilJL, ConéjéroG, Rodríguez-NavarroA, SentenacH, GuiderdoniE, AbdellyC, VéryAA (2009). Diversity in expression patterns and functional properties in the rice HKT transporter family.Plant Physiol 150, 1955-1971. [本文引用: 1]
[49]
Jaime-PérezN, PinedaB, García-SogoB, AtaresA, AthmanA, ByrtCS, OliasR, AsinsMJ, GillihamM, MorenoV, BelverA (2017). The sodium transporter encoded by the HKT1;2 gene modulates sodium/potassium homeostasis in tomato shoots under salinity. Plant Cell Environ 40, 658-671. [本文引用: 1]
[50]
JamesRA, BlakeC, ByrtCS, MunnsR (2011). Major genes for Na+ exclusion,Nax1 and Nax2(wheat HKT1;4 and HKT1;5), decrease Na+ accumulation in bread wheat lea- ves under saline and waterlogged conditions. J Exp Bot 62, 2939-2947. [本文引用: 1]
[51]
JamesRA, DavenportRJ, MunnsR (2006). Physiological characterization of two genes for Na+ exclusion in durum wheat,Nax1 and Nax2. Plant Physiol 142, 1537-1547. [本文引用: 1]
[52]
JulkowskaMM, KoevoetsIT, MolS, HoefslootH, FeronR, TesterMA, KeurentjesJJB, KorteA, HaringMA, De BoerGJ, TesterinkC (2017). Genetic components of root architecture remodeling in response to salt stress.Plant Cell 29, 3198-3213. [本文引用: 1]
[53]
KabayashiNI, YamajiN, YamamotoH, OkuboK, UenoH, CostaA, TanoiK, MatsumuraH, Fujii-KashinoM, HoriuchiT, Al NayefM, ShabalaS, AnG, MaJF, HorieT (2017). OsHKT1;5 mediates Na+ exclusion in the vasculature to protect leaf blades and reproductive tissues from salt toxicity in rice.Plant J 91, 657-670. [本文引用: 2]
[54]
KaderMA, LindbergS (2008). Cellular traits for sodium tolerance in rice (Oryza sativa L.). Plant Biotechnol 25, 247-255. [本文引用: 1]
[55]
KaderMA, SeidelT, GolldackD, LindbergS (2006). Expressions of OsHKT1, OsHKT2, and OsVHA are differentially regulated under NaCl stress in salt-sensitive and salt-tolerant rice(Oryza sativa L.) cultivars. J Exp Bot 57, 4257-4268. [本文引用: 1]
[56]
KarleyAJ, LeighRA, SandersD (2000a). Differential ion accumulation and ion fluxes in the mesophyll and epidermis of barley.Plant Physiol 122, 835-844. [本文引用: 1]
[57]
KarleyAJ, LeighRA, SandersD (2000b). Where do all the ions go? The cellular basis of differential ion accumulation in leaf cells.Trends Plant Sci 5, 465-470. [本文引用: 1]
[58]
KatoN, AkaiM, ZulkifliL, MatsudaN, KatoY, GoshimaS, HazamaA, YamagamiM, GuyHR, UozumiN (2007). Role of positively charged amino acids in the M2D transmembrane helix of Ktr/Trk/HKT type cation transporters.Channels 1, 161-171. [本文引用: 1]
[59]
KatoY, SakaguchiM, MoriY, SaitoK, NakamuraT, BakkerEP, SatoY, GoshimaS, UozumiN (2001). Evidence in support of a four transmembrane-pore-transmembrane topology model for theArabidopsis thaliana Na+/K+ translocating AtHKT1 protein, a member of the superfamily of K+ transporters. Proc Natl Acad Sci USA 98, 6488-6493. [本文引用: 1]
[60]
KronzuckerHJ, BrittoDT (2011). Sodium transport in plants: a critical review.New Phytol 189, 54-81. [本文引用: 1]
[61]
KumarS, BeenaAS, AwanaM, SinghA (2017). Salt- induced tissue-specific cytosine methylation downregulates expression of HKT genes in contrasting wheat (Triticum aestivum L.) genotypes. DNA Cell Biol 36, 283-294. [本文引用: 1]
[62]
LaurieS, FeeneyKA, MaathuisFJM, HeardPJ, BrownSJ, LeighRA (2002). A role for HKT1 in sodium uptake by wheat roots.Plant J 32, 139-149. [本文引用: 1]
[63]
LeidiEO, BarragánV, RubioL, El-HamdaouiA, RuizMT, CuberoB, FernándezJA, BressanRA, HasegawaPM, QuinteroFJ, PardoJM (2010). The AtNHX1 exchanger mediates potassium compartmentation in vacuoles of trans- genic tomato.Plant J 61, 495-506. [本文引用: 1]
[64]
LindsayMP, LagudahES, HareRA, MunnsR (2004). A locus for sodium exclusion (Nax1), a trait for salt tolerance, mapped in durum wheat. Funct Plant Biol 31, 1105-1114. [本文引用: 1]
[65]
LiuWH, FairbairnDJ, ReidRJ, SchachtmanDP (2001). Characterization of two HKT1 homologues from Eucalyptus camaldulensis that display intrinsic osmosensing capability. Plant Physiol 127, 283-294. [本文引用: 1]
[66]
LundeC, DrewPD, JacobsAK, TesterM (2007). Exclusion of Na+ via sodium ATPase (PpENA1) ensures normal growth of Physcomitrella patens under moderate salt str- ess. Plant Physiol 144, 1786-1796. [本文引用: 1]
[67]
MäserP, EckelmanB, VaidyanathanR, HorieT, FairbairnDJ, KuboM, YamagamiM, YamaguchiK, NishimuraM, UozumiN, RobertsonW, SussmanMR, SchroederJI (2002a). Altered shoot/root Na+ distribution and bifurcating salt sensitivity in Arabidopsis by genetic disruption of the Na+ transporterAtHKT1. FEBS Lett 531, 157-161. [本文引用: 2]
[68]
MäserP, HosooY, GoshimaS, HorieT, EckelmanB, YamadaK, YoshidaK, BakkerEP, ShinmyoA, OikiS, SchroederJI, UozumiN (2002b). Glycine residues in potassium channel-like selectivity filters determine potassium selectivity in four-loop-per-subunit HKT transporters from plants.Proc Natl Acad Sci USA 99, 6428-6433. [本文引用: 3]
[69]
MianA, OomenRJFJ, LsyenkovS, SentenacH, MaathuisFJM, VéryAA (2011). Over-expression of an Na+-and K+-permeable HKT transporter in barley improves salt tole- rance.Plant J 68, 468-479. [本文引用: 2]
[70]
MickelbartMV, HasegawaPM, Bailey-SerresJ (2015). Genetic mechanisms of abiotic stress tolerance that translate to crop yield stability.Nat Rev Genet 16, 237-251. [本文引用: 1]
[71]
MøllerIS, GillihamM, JhaD, MayoGM, RoySJ, CoatesJC, HaseloffJ, TesterM (2009). Shoot Na+ exclusion and increased salinity tolerance engineered by cell type- specific alteration of Na+ transport in Arabidopsis.Plant Cell 21, 2163-2178. [本文引用: 2]
[72]
MunnsR, JamesRA, XuB, AthmanA, ConnSJ, JordansC, ByrtCS, HareRA, TyermanSD, TesterM, PlettD, GillihamM (2012). Wheat grain yield on saline soils is improved by an ancestral Na+ transporter gene.Nat Biotechnol 30, 360-364. [本文引用: 3]
OdaY, KobayashiNI, TanoiK, MaJF, ItouY, KatsuharaM, ItouT, HorieT (2018). T-DNA tagging-based gain-of- function of OsHKT1;4 reinforces Na exclusion from leaves and stems but triggers Na toxicity in roots of rice under salt stress.Int J Mol Sci 19, 235. [本文引用: 1]
[75]
OhtaM, HayashiY, NakashimaA, HamadaA, TanakaA, NakamuraT, HayakawaT (2002). Introduction of a Na+/H+ antiporter gene from Atriplex gmelini confers salt tolerance to rice. FEBS Lett 532, 279-282. [本文引用: 1]
[76]
OlíasR, EljakaouiZ, LiJ, De MoralesPA, Marin-ManzanoMC, PardoJM, BelverA (2009). The plasma membrane Na+/H+ antiporter SOS1 is essential for salt tolerance in tomato and affects the partitioning of Na+ between plant organs.Plant Cell Environ 32, 904-916. [本文引用: 1]
[77]
OomenRJFJ, BenitoB, SentenacH, Rodríguez-NavarroA, TalónM, VeryAA, DomingoC (2012). HKT2;2/1, a K+-permeable transporter identified in a salt-tolerant rice cultivar through surveys of natural genetic polymorphism.Plant J 71, 750-762. [本文引用: 2]
[78]
PantojaO, DaintyJ, BlumwaldE (1989). Ion channels in vacuoles from halophytes and glycophytes.FEBS Lett 255, 92-96. [本文引用: 1]
[79]
QiZ, SpaldingEP (2004). Protection of plasma membrane K+ transport by the salt overly sensitive1 Na+-H+ antiporter during salinity stress.Plant Physiol 136, 2548-2555. [本文引用: 1]
[80]
RahmanMA, BimpongIK, BizimanaJB, PascualED, ArcetaM, SwamyBPM, DiawF, RahmanMS, SinghRK (2017). Mapping QTLs using a novel source of salinity tole- rance from Hasawi and their interaction with environments in rice.Rice 10, 47. [本文引用: 1]
[81]
RegueraM, BassilE, TajimaH, WimmerM, ChanocaA, OteguiMS, ParisN, BlumwaldE (2015). pH regulation by NHX-type antiporters is required for receptor-mediated protein trafficking to the vacuole in Arabidopsis.Plant Cell 27, 1200-1217. [本文引用: 1]
[82]
RenZH, GaoJP, LiLG, CaiXL, HuangW, ChaoDY, ZhuMZ, WangZY, LuanS, LinHX (2005). A rice quantitative trait locus for salt tolerance encodes a sodium transporter.Nat Genet 37, 1141-1146. [本文引用: 2]
[83]
Rodríguez-NavarroA (2000). Potassium transport in fungi and plants.Biochim Biophys Acta 1469, 1-30. [本文引用: 1]
RubioF, GassmannW, SchroederJI (1995). Sodium- driven potassium uptake by the plant potassium transporter HKT1 and mutations conferring salt tolerance.Science 270, 1660-1663. [本文引用: 3]
[88]
RubioF, Nieves-CordonesM, AlemánF, MartínezV (2008). Relative contribution of AtHAK5 and AtAKT1 to K+ uptake in the high-affinity range of concentrations.Physiol Plantarum 134, 598-608. [本文引用: 1]
[89]
RubioF, SchwarzM, GassmannW, SchroederJI (1999). Genetic selection of mutations in the high affinity K+ transporter HKT1 that define functions of a loop site for reduced Na+ permeability and increased Na+ tolerance.J Biol Chem 274, 6839-6847. [本文引用: 1]
[90]
RusA, BaxterI, MuthukumarB, GustinJ, LahnerB, YakubovaE, SaltDE (2006). Natural variants of AtHKT1 enhance Na+ accumulation in two wild populations of Ara- bidopsis.PLoS Genet 2, e210. [本文引用: 1]
SchroderJI, DelhaizeE, FrommerWB, GuerinotML, HarrisonMJ, Herrera-EstrellaL, HorieT, KochianLV, MunnsR, NishizawaNK, TsayYF, SandersD (2013). Using membrane transporters to improve crops for sustainable food production.Nature 497, 60-66. [本文引用: 1]
[94]
ShiHZ, QuinteroFJ, PardoJM, ZhuJK (2002). The putative plasma membrane Na+/H+ antiporter SOS1 controls long-distance Na+ transport in plants.Plant Cell 14, 465-477. [本文引用: 2]
[95]
StelzerR (1981). Ion localization in the leaves of Puccinellia peisonis. Z Pflanzenphysiol 103, 27-36. [本文引用: 1]
[96]
Sunarpi, HorieT, MotodaJ, KuboM, YangH, YodaK, HorieR, ChanWY, LeungHY, HattoriK, KonomiM, OsumiM, YamagamiM, SchroederJI, UozumiN (2005). Enhanced salt tolerance mediated by AtHKT1 transporter-induced Na+ unloading from xylem vessels to xylem parenchyma cells.Plant J 44, 928-938. [本文引用: 1]
[97]
SuzukiK, YamajiN, CostaA, OkumaE, KobayashiNI, KashiwagiT, KatsuharaM, WangC, TanoiK, MurataY, SchroederJI, MaJF, HorieT (2016). OsHKT1;4-mediated Na+ transport in stems contributes to Na+ exclusion from leaf blades of rice at the reproductive growth stage upon salt stress.BMC Plant Biol 16, 22. [本文引用: 3]
[98]
SzczerbaMW, BrittoDT, KronzuckerHJ (2009). K+ transport in plants: physiology and molecular biology.J Plant Physiol 166, 447-466. [本文引用: 1]
[99]
TesterM, DavenportR (2003). Na+ tolerance and Na+ transport in higher plants.Ann Bot 91, 503-527. [本文引用: 1]
[100]
UozumiN, KimEJ, RubioF, YamaguchiT, MutoS, TsuboiA, BakkerEP, NakamuraT, SchroederJI (2000). The Arabidopsis HKT1 gene homolog mediates inward Na+ currents in Xenopus laevis oocytes and Na+ uptake in Saccharomyces cerevisiae. Plant Physiol 122, 1249-1260. [本文引用: 1]
[101]
VolkovV (2015). Salinity tolerance in plants. quantitative approach to ion transport starting from halophytes and stepping to genetic and protein engineering for manipulating ion fluxes.Front Plant Sci 6, 873. [本文引用: 1]
[102]
VolkovV, WangB, DominyPJ, FrickeW, AmtmannA (2004). Thellungiella halophila, a salt-tolerant relative of Arabidopsis thaliana, possesses effective mechanisms to discriminate between potassium and sodium. Plant Cell Environ 27, 1-14. [本文引用: 1]
[103]
WangB, DavenportRJ, VolkovV, AmtmannA (2006). Low unidirectional sodium influx into root cells restricts net sodium accumulation in Thellungiella halophila, a salt-tol- erant relative of Arabidopsis thaliana. J Exp Bot 57, 1161-1170. [本文引用: 1]
[104]
WangH, ZhangMS, GuoR, ShiDS, LiuB, LinXY, YangCW (2012). Effects of salt stress on ion balance and nitrogen metabolism of old and young leaves in rice (Oryza sativa L.) BMC Plant Biol 12, 194. [本文引用: 1]
[105]
WangR, JingW, XiaoLY, JinYK, ShenLK, ZhangWH (2015). The rice high-affinity potassium transporter1;1 is involved in salt tolerance and regulated by an MYB-type transcription factor.Plant Physiol 168, 1076-1090. [本文引用: 2]
[106]
WangSM, ZhaoGQ, GaoYS, TangZC, ZhangCL (2004). Puccinellia tenuiflora exhibits stronger selectivity for K+ over Na+ than wheat. J Plant Nutr 27, 1841-1857. [本文引用: 1]
[107]
WangTT, RenZJ, LiuZQ, FengX, GuoRQ, LiBG, LiLG, JingHC (2014). SbHKT1;4, a member of the high-affinity potassium transporter gene family from Sorghum bicolor, functions to maintain optimal Na+/K+ balance under Na+ stress. J Integr Plant Biol 56, 315-332. [本文引用: 1]
[108]
YamaguchiT, AharonGS, SottosantoJB, BlumwaldE (2005). Vacuolar Na+/H+ antiporter cation selectivity is regu- lated by calmodulin from within the vacuole in a Ca2+-and pH-dependent manner.Proc Natl Acad Sci USA 102, 16107-16112. [本文引用: 1]
[109]
YaoX, HorieT, XueSW, LeungHY, KatsuharaM, BrodskyDE, WuY, SchroederJI (2010). Differential sodium and potassium transport selectivities of the rice OsHKT2;1 and OsHKT2;2 transporters in plant cells.Plant Physiol 152, 341-355. [本文引用: 1]
[110]
ZengY, LiQ, WangHY, ZhangJL, DuJ, FengHM, BlumwaldE, YuL, XuGH (2018). Two NHX-type transporters from Helianthus tuberosus improve the tolerance of rice to salinity and nutrient deficiency stress. Plant Biotechnol J 16, 310-321. [本文引用: 1]
ZhangHX, HodsonJN, WilliamsJP, BlumwaldE (2001). Engineering salt-tolerant Brassica plants: characterization of yield and seed oil quality in transgenic plants with increased vacuolar sodium accumulation. Proc Natl Acad Sci USA 98, 12832-12836. [本文引用: 1]
[113]
ZhangM, CaoY, WangZP, WangZQ, ShiJP, LiangXY, SongWB, ChenQJ, LaiJS, JiangCF (2017b). A retrotransposon in an HKT1 family sodium transporter causes variation of leaf Na+ exclusion and salt tolerance in maize.New Phytol 217, 1161-1176. [本文引用: 1]
[114]
ZhangWD, WangP, BaoZ, MaQ, DuanLJ, BaoAK, ZhangJL, WangSU (2017c). SOS1, HKT1;5, and NHX1 synergistically modulate Na+ homeostasis in the halophytic grass Puccinellia tenuiflora. Front Plant Sci 8, 576. [本文引用: 1]
拟南芥NHX5和NHX6: 离子平衡与蛋白质运输 1 2017
... 植物组织耐受性是指将钠离子区隔化入特定组织细胞的液泡内, 从而降低离子毒害.如地上部叶片表皮细胞或维管束细胞具有发育良好的液泡, 植物体通过液泡H+-ATPase和焦磷酸酶建立起液泡内外的H+梯度, 驱动Na+/H+逆向转运蛋白, 将Na+泵到液泡贮藏, 从而减轻过量钠离子对胞质酶的伤害, 保证胞质执行正常生理功能(Blumwald, 2000).植物(Na+, K+/H+)反向交换体(antiporter, NHX)利用质子泵产生的H+电化学梯度进行离子交换, 根据其亚细胞分布分为质膜NHX (plasma membrane NHX)、液泡NHX (vacuolar NHX)和内膜NHX (endosomal NHX).目前, 已在包括盐土植物和淡土植物的许多物种中发现了NHX- type离子转运蛋白.盐土植物北滨藜(Atriplex gme- lini)液泡AgNHX1基因编码Na+/H+反向转运蛋白, 在盐胁迫下被诱导表达; 在水稻中异源表达AgNHX1基因可显著提高其耐盐特性, 因此推测盐胁迫下液泡NHX蛋白具有将Na+区隔化至液泡的功能(Hamada et al., 2001; Ohta et al., 2002).OsNHX1基因编码1个液泡(Na+, K+/H+)反向交换体, 用高浓度的NaCl和KCl处理可提高OsNHX1基因在水稻茎和根中的转录水平, 过表达OsNHX1基因可显著提高水稻植株的耐盐性, 因此推测OsNHX1蛋白具有将胞质中过量积累的Na+和K+区隔化至液泡的功能(Fukuda et al., 2004).拟南芥atnhx1突变体具有显著的盐敏感表型, 在拟南芥和油菜(Brassica napus)植株中异源表达AtNHX1基因, 可提高植株耐盐性(Apse et al., 1999; Zhang et al., 2001).后续研究表明, 当钙调蛋白At- CaM15与液胞膜AtNHX1蛋白的C端结合, 则限制At- NHX1蛋白的Na+转运特性, 成为K+/H+反向转运蛋白; 拟南芥液泡AtNHX1蛋白可能不具有转运Na+的功能, 而在提高地上部K+含量方面发挥重要调节作用(Yamaguchi et al., 2005; Bassil et al., 2011b; Barragán et al., 2012).例如, 在番茄中异源表达At- NHX1基因, 植株通过维持细胞K+平衡, 而非将Na+区隔化至液泡贮藏, 来提高转基因植株的耐盐性(Leidi et al., 2010).番茄内膜LeNHX2编码K+/H+反向转运蛋白, 在番茄中过表达LeNHX2基因, 可提高盐胁迫下转基因植株对K+的吸收, 从而提高植物的耐盐性(Huertas et al., 2013).目前, 除植物液泡NHX蛋白的功能研究较为详尽之外, 植物内膜NHX蛋白功能也有报道.拟南芥内膜AtNHX5和AtNHX6蛋白定位于高尔基体(Golgi)、反面高尔基体(TGN)和多胞体/液泡前体(MVB/PVC), 具有调节细胞pH、维持离子平衡、调控蛋白质运输及植物生长发育等方面的功能(Bassil et al., 2011a, 2012; 邱全胜, 2017).在水稻中转化菊芋(Helianthus tuberosus) HtNHX2基因, 发现HtNHX2在转基因水稻植株中的定位类似于AtNHX5蛋白在拟南芥中的定位, 转基因水稻在盐胁迫和缺素处理下耐逆性有所提高(Zeng et al., 2018).转化NHX基因对植物体内的哪些调控过程产生影响, 还需深入研究(Rodríguez-Rosales et al., 2009; Barragán et al., 2012; Reguera et al., 2015). ...
不同水稻种质在不同生育期耐盐鉴定的差异 1 2017
... 截至目前, 水稻HKT家族含有2个参与调控Na+在根-冠分配的基因, 分别是OsHKT1;5和OsHKT- 1;4.已在耐盐水稻(Nona Bokra)与粳稻品种越光(Koshihikari)杂交群体中定位到重要的耐盐QTL位点SKC1.该段序列是1个HKT类的基因OsHKT1;5, 通过膜片钳及非洲爪蟾卵母细胞的系统证明, 其编码Na+转运载体蛋白, 在根中木质部薄壁细胞膜表达, 通过卸载木质部中钠离子并限制其向地上部运输降低地上部Na+含量, 进而间接影响K+的含量, 而维持适宜的K+/Na+平衡状态(Ren et al., 2005).粳稻品种OsHKT1;4基因在非洲爪蟾卵母细胞和酵母异源表达系统中均被证明具有显著的Na+转运能力, EGFP- OsHKT1;4融合蛋白定位在原生质体细胞膜.Os- HKT1;4在营养生长阶段主要在叶鞘中表达, 在生殖生长阶段节间II和花序梗处表达量高.生殖生长阶段OsHKT1;4-RNAi植株遭受盐胁迫, 使其地上部尤其在叶片和叶鞘中Na+过度积累, 而在营养生长阶段变化不明显.22Na+标记实验表明, OsHKT1;4在生殖生长阶段卸载茎部和叶鞘Na+, 有助于Na+从叶片排出, 而在营养生长阶段其功能不显著(Suzuki et al., 2016).但盲目地通过增强OsHKT1;4基因在叶片的排钠功能来进行耐盐作物育种, 不仅不会提高植株的耐盐性, 还可能会造成植株根部大量吸收Na+, 因此其在育种中的应用尚需进一步研究(Oda et al., 2018).水稻种质耐盐性存在较大差异(吴家富等, 2017), 利用耐盐水稻品种Hasawi的3个作图群体, 鉴定出Hasawi中75个耐盐QTLs, 表明水稻Hasawi品种含有的优异耐盐等位基因还有待挖掘(Bimpong et al., 2014; Bizimana et al., 2017; Rahman et al., 2017). ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}