何光明, 邓兴旺*, 北京大学现代农学院与生命科学学院/蛋白质与植物基因研究国家重点实验室/北大-清华生命科学联合中心, 北京 100871 HeGuangming, DengXingwang*, Peking-Tsinghua Center for Life Sciences/State Key Laboratory of Protein and Plant Gene Research/School of Advanced Agricultural Sciences and School of Life Sciences, Peking University, Beijing 100871, China 引用本文 何光明, 邓兴旺. 死亡信号传递: 叶绿体与线粒体间信号交流调控植物程序性细胞死亡. 植物学报, 2018, 53(4): 441-444
Abstract: Programmed cell death (PCD) is an active and genetically controlled process that results in cell death in a multicellular organism. PCD plays important roles in plant development and defense and can be induced by increased reactive oxygen species (ROS) in mitochondria. Previous studies by Jiayang Li’s group in the Institute of Genetics and Developmental Biology, Chinese Academy of Sciences identified a cell death mutant, mod1, in Arabidopsis. Although the authors suggested that signal communication between chloroplast and mitochondrion contributes to PCD in mod1, the underlying mechanisms remain elusive. Recently, by screening the suppressors of mod1, they cloned three new suppressor genes, plNAD-MDH, DiT1 and mMDH1, which encode a plastid-localized NAD-dependent malate dehydrogenase, a chloroplastic dicarboxylate transporter, and a mitochondrial malate dehydrogenase, respectively. Mutations in these three genes each can suppress ROS accumulation and PCD in mod1. Functional analyses of these genes demonstrated that malate is transported from chloroplasts to mitochondria and triggers ROS generation and PCD in plant. This study expands our knowledge of intracellular communications between organelles and provides new understanding of molecular mechanisms of PCD in plants, which is an important progress in this field.
Key words:chloroplast ; mitochondrion ; malate ; reactive oxygen species ; programmed cell death
程序性细胞死亡(programmed cell death, PCD)是多细胞生物体中一种受遗传调控且自主而有序的细胞死亡现象。植物在正常的生长发育和响应外界环境刺激过程中都会发生PCD, 其对植物器官的发生与分化以及抵抗生物和非生物胁迫均起重要作用(Dickman et al., 2017)。例如, 玉米(Zea mays)雄花和雌花的分化就分别是未成熟花序中雌性和雄性性原基细胞发生PCD后退化的结果(Beers, 1997); 植物在抵抗活体营养型病原物侵袭过程中, 受感染部位发生的超敏反应(hypersensitive response, HR)也是一种典型的PCD (用于限制病原物的入侵和扩散) (Coll et al., 2011)。目前的研究表明, 活性氧(reactive oxygen species, ROS)在植物PCD中发挥重要作用(Van Breusegem and Dat, 2006)。ROS是生物体内一类含氧自由基或含氧分子的总称, 包括单线态氧(1O2)、超氧阴离子自由基(O2-)和过氧化氢(H2O2)等, 它们性质活跃, 具有高反应活性, 可通过电子传递或其它代谢途径产生。植物在遭受各种生物或非生物胁迫时, 体内ROS增加, 可作为信号分子诱导细胞发生程序性死亡(Van Breusegem and Dat, 2006; Van Aken and Van Breusegem, 2015)。
ROS可在叶绿体、线粒体和过氧化物酶体等多种细胞器中产生。在光下, 特别是高强度光照条件下, 叶绿体是植物产生ROS的主要来源, 并且与PCD相关。然而, 近年来的研究表明, 线粒体在PCD中起核心作用。在正常条件下, 植物的线粒体电子传递链(mitochondrial electron transport chain, mETC)通过其组分的可逆氧化还原传递电子并最终导致ATP产生。而在胁迫条件下, mETC组分中电子载体过度还原, 从而引发ROS的生成, 所产生的ROS作为信号分子增加线粒体膜的通透性, 诱导细胞发生程序性死亡(Van Aken and Van Breusegem, 2015; Gorlach et al., 2015)。一个有趣的问题是, 叶绿体中导致ROS产生的信号是否可传递到线粒体中激发ROS产生并最终诱导PCD? 近年来, 中国科学院遗传与发育生物学研究所李家洋研究组针对该问题进行了一系列研究。
李家洋研究组首先通过对拟南芥(Arabidopsis thaliana) Col-0生态型进行甲基磺酸乙酯(ethyl meth- ane sulphonate, EMS)诱变, 筛选出1个细胞死亡突变体mod1 (mosaic death 1), 并通过图位克隆鉴定出MOD1基因。该基因编码脂肪酸合酶复合体中的一个亚基——烯酰基酰基载体蛋白还原酶(enoyl-acyl carrier protein reductase, enoyl-ACP reductase), 其参与催化叶绿体中脂肪酸的合成, 负调控植物PCD (Mou et al., 2000)。随后, 该研究组发现, mod1突变体中存在明显的ROS积累, 暗示ROS的过量积累与该突变体的细胞死亡表型相关。为探明其中的具体作用机制, 他们针对mod1突变体构建了其T-DNA插入突变体库, 从中筛选出能够抑制mod1细胞死亡和ROS积累表型的抑制突变体soms (suppressors of mod1), 并克隆了这些抑制突变体对应的抑制基因, 发现它们编码线粒体电子传递链复合体I (mETC复合体I)的亚基, 或影响mETC复合体I的组装及活性, 表明完整的mETC复合体I在mod1突变体的PCD中起关键作用(Wu et al., 2015)。mod1是叶绿体中脂肪酸合酶的突变体, 其PCD表型却能被线粒体中ETC复合体I的功能缺失所恢复, 由此暗示在叶绿体中确实产生了某种信号并传递到了线粒体, 线粒体接受该信号后通过ETC产生ROS并引发PCD。那么, 这个信号是什么? 它又是如何从叶绿体传递到线粒体的? 最近, 李家洋研究组在Cell Research杂志上发表的研究论文对该问题进行了精彩且完整的解答(Zhao et al., 2018)。
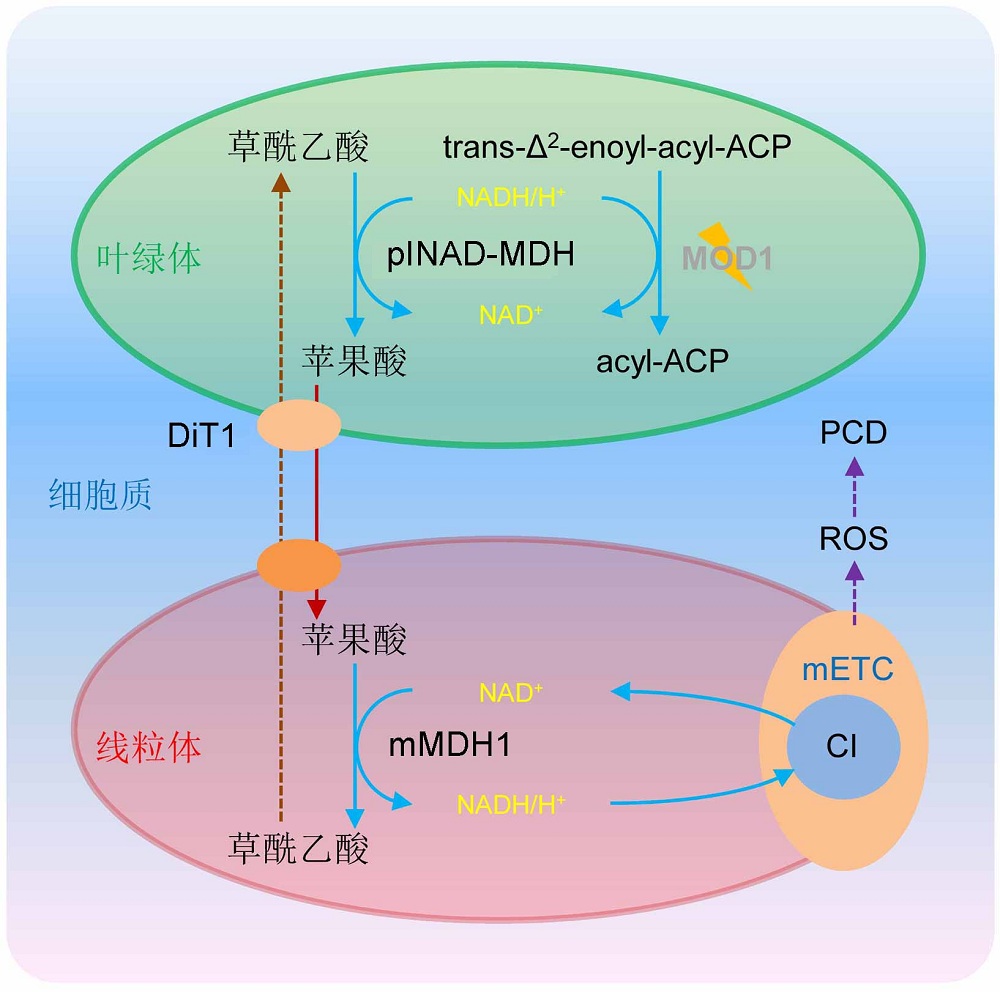
图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-441/img_1.jpg<b>图1</b> 叶绿体与线粒体间信号交流调控植物程序性细胞死亡(PCD)的工作模型(改自<xref ref-type="bibr" rid="b13-1674-3466-53-4-441">Zhao et al., 2018</xref>)<br/>植物<i>MOD1</i>基因编码烯酰基酰基载体蛋白还原酶, 该酶以NADH作为共同底物参与催化叶绿体中脂肪酸的合成。在<i>mod1</i>突变体中, MOD1的功能缺失或表达量降低导致NADH/H<sup>+</sup>在叶绿体中大量积累; 过量的NADH/H<sup>+</sup>使叶绿体中的草酰乙酸在plNAD-MDH的作用下被还原为苹果酸; 携带还原力的苹果酸通过定位于叶绿体内膜上的DiT1转运蛋白转运到细胞质中, 并进一步通过定位于线粒体膜上的一种未知转运蛋白转移到线粒体; 在线粒体中, mMDH1将携带还原力的苹果酸脱氢转化为草酰乙酸, 同时生成NADH/H<sup>+</sup>。线粒体中积累的NADH/H<sup>+</sup>为线粒体膜上的mETC复合体I (CI)提供电子, 产生活性氧(ROS), 最终导致<i>mod1</i>突变体细胞发生PCD。<br/><b>Figure 1</b> A proposed model explaining the programmed cell death in plants regulated by chloroplast-to-mitochondrion communication (modified from <xref ref-type="bibr" rid="b13-1674-3466-53-4-441">Zhao et al., 2018</xref>)<br/>In plants, <i>MOD1</i> encodes an enoyl-acyl carrier protein (ACP) reductase, which is involved in catalyzing the de novo biosynthesis of fatty acids in plastids using NADH as cosubstrate. The deficiency of MOD1 in <i>mod1</i> mutant leads to an increased level of NADH/H<sup>+</sup> in the chloroplast, which drives oxaloacetate to be converted to malate by plNAD-MDH. Malate carrying the reducing equivalents is transported out of the chloroplast into the cytosol by DiT1, which is localized in the chloroplast inner envelope membrane, and then is trans- ported into the mitochondrion by an unidentified transporter localized in the mitochondrion membrane. In the mitochondrion, malate is converted to oxaloacetate by mMDH1, and simultaneously NADH/H<sup>+</sup> is generated to provide electrons for mETC complex I (CI) to induce the ROS formation and initiate the PCD process in the <i>mod1</i> cells. Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-441/img_1.jpg<b>图1</b> 叶绿体与线粒体间信号交流调控植物程序性细胞死亡(PCD)的工作模型(改自<xref ref-type="bibr" rid="b13-1674-3466-53-4-441">Zhao et al., 2018</xref>)<br/>植物<i>MOD1</i>基因编码烯酰基酰基载体蛋白还原酶, 该酶以NADH作为共同底物参与催化叶绿体中脂肪酸的合成。在<i>mod1</i>突变体中, MOD1的功能缺失或表达量降低导致NADH/H<sup>+</sup>在叶绿体中大量积累; 过量的NADH/H<sup>+</sup>使叶绿体中的草酰乙酸在plNAD-MDH的作用下被还原为苹果酸; 携带还原力的苹果酸通过定位于叶绿体内膜上的DiT1转运蛋白转运到细胞质中, 并进一步通过定位于线粒体膜上的一种未知转运蛋白转移到线粒体; 在线粒体中, mMDH1将携带还原力的苹果酸脱氢转化为草酰乙酸, 同时生成NADH/H<sup>+</sup>。线粒体中积累的NADH/H<sup>+</sup>为线粒体膜上的mETC复合体I (CI)提供电子, 产生活性氧(ROS), 最终导致<i>mod1</i>突变体细胞发生PCD。<br/><b>Figure 1</b> A proposed model explaining the programmed cell death in plants regulated by chloroplast-to-mitochondrion communication (modified from <xref ref-type="bibr" rid="b13-1674-3466-53-4-441">Zhao et al., 2018</xref>)<br/>In plants, <i>MOD1</i> encodes an enoyl-acyl carrier protein (ACP) reductase, which is involved in catalyzing the de novo biosynthesis of fatty acids in plastids using NADH as cosubstrate. The deficiency of MOD1 in <i>mod1</i> mutant leads to an increased level of NADH/H<sup>+</sup> in the chloroplast, which drives oxaloacetate to be converted to malate by plNAD-MDH. Malate carrying the reducing equivalents is transported out of the chloroplast into the cytosol by DiT1, which is localized in the chloroplast inner envelope membrane, and then is trans- ported into the mitochondrion by an unidentified transporter localized in the mitochondrion membrane. In the mitochondrion, malate is converted to oxaloacetate by mMDH1, and simultaneously NADH/H<sup>+</sup> is generated to provide electrons for mETC complex I (CI) to induce the ROS formation and initiate the PCD process in the <i>mod1</i> cells.
图1 叶绿体与线粒体间信号交流调控植物程序性细胞死亡(PCD)的工作模型(改自Zhao et al., 2018) 植物MOD1基因编码烯酰基酰基载体蛋白还原酶, 该酶以NADH作为共同底物参与催化叶绿体中脂肪酸的合成。在mod1突变体中, MOD1的功能缺失或表达量降低导致NADH/H+在叶绿体中大量积累; 过量的NADH/H+使叶绿体中的草酰乙酸在plNAD-MDH的作用下被还原为苹果酸; 携带还原力的苹果酸通过定位于叶绿体内膜上的DiT1转运蛋白转运到细胞质中, 并进一步通过定位于线粒体膜上的一种未知转运蛋白转移到线粒体; 在线粒体中, mMDH1将携带还原力的苹果酸脱氢转化为草酰乙酸, 同时生成NADH/H+。线粒体中积累的NADH/H+为线粒体膜上的mETC复合体I (CI)提供电子, 产生活性氧(ROS), 最终导致mod1突变体细胞发生PCD。 Figure 1 A proposed model explaining the programmed cell death in plants regulated by chloroplast-to-mitochondrion communication (modified from Zhao et al., 2018) In plants, MOD1 encodes an enoyl-acyl carrier protein (ACP) reductase, which is involved in catalyzing the de novo biosynthesis of fatty acids in plastids using NADH as cosubstrate. The deficiency of MOD1 in mod1 mutant leads to an increased level of NADH/H+ in the chloroplast, which drives oxaloacetate to be converted to malate by plNAD-MDH. Malate carrying the reducing equivalents is transported out of the chloroplast into the cytosol by DiT1, which is localized in the chloroplast inner envelope membrane, and then is trans- ported into the mitochondrion by an unidentified transporter localized in the mitochondrion membrane. In the mitochondrion, malate is converted to oxaloacetate by mMDH1, and simultaneously NADH/H+ is generated to provide electrons for mETC complex I (CI) to induce the ROS formation and initiate the PCD process in the mod1 cells.
真核生物细胞中的各亚细胞单位或细胞器之间会发生物质(信息)的交流, 以协同调控细胞的各种生命活动(Schuldiner and Guo, 2015)。近年来的研究显示, 植物中存在叶绿体和线粒体之间的信息交流, 并诱导PCD的发生, 但其中的作用机制未得到详尽的解析。该研究证实了苹果酸穿梭对叶绿体到线粒体间的信息传递及线粒体中ROS的产生起重要作用, 更新了我们对细胞器之间信号交流和植物PCD发生机制的认识, 对推动植物PCD领域的研究具有重要意义。
The authors have declared that no competing interests exist.
ZhaoYN, LuoLL, XuJS, XinPY, GuoHY, WuJ, BaiL, WangGD, ChuJF, ZuoJR, YuH, HuangX, LiJY (2018). Malate transported from chloroplast to mitochondrion triggers production of ROS and PCD in Arabidopsis thaliana. Cell Res 28, 448-461. [本文引用: 3]
1 1997
... 程序性细胞死亡(programmed cell death, PCD)是多细胞生物体中一种受遗传调控且自主而有序的细胞死亡现象.植物在正常的生长发育和响应外界环境刺激过程中都会发生PCD, 其对植物器官的发生与分化以及抵抗生物和非生物胁迫均起重要作用(Dickman et al., 2017).例如, 玉米(Zea mays)雄花和雌花的分化就分别是未成熟花序中雌性和雄性性原基细胞发生PCD后退化的结果(Beers, 1997); 植物在抵抗活体营养型病原物侵袭过程中, 受感染部位发生的超敏反应(hypersensitive response, HR)也是一种典型的PCD (用于限制病原物的入侵和扩散) (Coll et al., 2011).目前的研究表明, 活性氧(reactive oxygen species, ROS)在植物PCD中发挥重要作用(Van Breusegem and Dat, 2006).ROS是生物体内一类含氧自由基或含氧分子的总称, 包括单线态氧(1O2)、超氧阴离子自由基(O2-)和过氧化氢(H2O2)等, 它们性质活跃, 具有高反应活性, 可通过电子传递或其它代谢途径产生.植物在遭受各种生物或非生物胁迫时, 体内ROS增加, 可作为信号分子诱导细胞发生程序性死亡(Van Breusegem and Dat, 2006; Van Aken and Van Breusegem, 2015). ...
... 程序性细胞死亡(programmed cell death, PCD)是多细胞生物体中一种受遗传调控且自主而有序的细胞死亡现象.植物在正常的生长发育和响应外界环境刺激过程中都会发生PCD, 其对植物器官的发生与分化以及抵抗生物和非生物胁迫均起重要作用(Dickman et al., 2017).例如, 玉米(Zea mays)雄花和雌花的分化就分别是未成熟花序中雌性和雄性性原基细胞发生PCD后退化的结果(Beers, 1997); 植物在抵抗活体营养型病原物侵袭过程中, 受感染部位发生的超敏反应(hypersensitive response, HR)也是一种典型的PCD (用于限制病原物的入侵和扩散) (Coll et al., 2011).目前的研究表明, 活性氧(reactive oxygen species, ROS)在植物PCD中发挥重要作用(Van Breusegem and Dat, 2006).ROS是生物体内一类含氧自由基或含氧分子的总称, 包括单线态氧(1O2)、超氧阴离子自由基(O2-)和过氧化氢(H2O2)等, 它们性质活跃, 具有高反应活性, 可通过电子传递或其它代谢途径产生.植物在遭受各种生物或非生物胁迫时, 体内ROS增加, 可作为信号分子诱导细胞发生程序性死亡(Van Breusegem and Dat, 2006; Van Aken and Van Breusegem, 2015). ... ... ROS可在叶绿体、线粒体和过氧化物酶体等多种细胞器中产生.在光下, 特别是高强度光照条件下, 叶绿体是植物产生ROS的主要来源, 并且与PCD相关.然而, 近年来的研究表明, 线粒体在PCD中起核心作用.在正常条件下, 植物的线粒体电子传递链(mitochondrial electron transport chain, mETC)通过其组分的可逆氧化还原传递电子并最终导致ATP产生.而在胁迫条件下, mETC组分中电子载体过度还原, 从而引发ROS的生成, 所产生的ROS作为信号分子增加线粒体膜的通透性, 诱导细胞发生程序性死亡(Van Aken and Van Breusegem, 2015; Gorlach et al., 2015).一个有趣的问题是, 叶绿体中导致ROS产生的信号是否可传递到线粒体中激发ROS产生并最终诱导PCD? 近年来, 中国科学院遗传与发育生物学研究所李家洋研究组针对该问题进行了一系列研究. ...
2 2006
... 程序性细胞死亡(programmed cell death, PCD)是多细胞生物体中一种受遗传调控且自主而有序的细胞死亡现象.植物在正常的生长发育和响应外界环境刺激过程中都会发生PCD, 其对植物器官的发生与分化以及抵抗生物和非生物胁迫均起重要作用(Dickman et al., 2017).例如, 玉米(Zea mays)雄花和雌花的分化就分别是未成熟花序中雌性和雄性性原基细胞发生PCD后退化的结果(Beers, 1997); 植物在抵抗活体营养型病原物侵袭过程中, 受感染部位发生的超敏反应(hypersensitive response, HR)也是一种典型的PCD (用于限制病原物的入侵和扩散) (Coll et al., 2011).目前的研究表明, 活性氧(reactive oxygen species, ROS)在植物PCD中发挥重要作用(Van Breusegem and Dat, 2006).ROS是生物体内一类含氧自由基或含氧分子的总称, 包括单线态氧(1O2)、超氧阴离子自由基(O2-)和过氧化氢(H2O2)等, 它们性质活跃, 具有高反应活性, 可通过电子传递或其它代谢途径产生.植物在遭受各种生物或非生物胁迫时, 体内ROS增加, 可作为信号分子诱导细胞发生程序性死亡(Van Breusegem and Dat, 2006; Van Aken and Van Breusegem, 2015). ... ... )等, 它们性质活跃, 具有高反应活性, 可通过电子传递或其它代谢途径产生.植物在遭受各种生物或非生物胁迫时, 体内ROS增加, 可作为信号分子诱导细胞发生程序性死亡(Van Breusegem and Dat, 2006; Van Aken and Van Breusegem, 2015). ...
1 2015
... 李家洋研究组首先通过对拟南芥(Arabidopsis thaliana) Col-0生态型进行甲基磺酸乙酯(ethyl meth- ane sulphonate, EMS)诱变, 筛选出1个细胞死亡突变体mod1 (mosaic death 1), 并通过图位克隆鉴定出MOD1基因.该基因编码脂肪酸合酶复合体中的一个亚基——烯酰基酰基载体蛋白还原酶(enoyl-acyl carrier protein reductase, enoyl-ACP reductase), 其参与催化叶绿体中脂肪酸的合成, 负调控植物PCD (Mou et al., 2000).随后, 该研究组发现, mod1突变体中存在明显的ROS积累, 暗示ROS的过量积累与该突变体的细胞死亡表型相关.为探明其中的具体作用机制, 他们针对mod1突变体构建了其T-DNA插入突变体库, 从中筛选出能够抑制mod1细胞死亡和ROS积累表型的抑制突变体soms (suppressors of mod1), 并克隆了这些抑制突变体对应的抑制基因, 发现它们编码线粒体电子传递链复合体I (mETC复合体I)的亚基, 或影响mETC复合体I的组装及活性, 表明完整的mETC复合体I在mod1突变体的PCD中起关键作用(Wu et al., 2015).mod1是叶绿体中脂肪酸合酶的突变体, 其PCD表型却能被线粒体中ETC复合体I的功能缺失所恢复, 由此暗示在叶绿体中确实产生了某种信号并传递到了线粒体, 线粒体接受该信号后通过ETC产生ROS并引发PCD.那么, 这个信号是什么? 它又是如何从叶绿体传递到线粒体的? 最近, 李家洋研究组在Cell Research杂志上发表的研究论文对该问题进行了精彩且完整的解答(Zhao et al., 2018). ...
3 2018
... 李家洋研究组首先通过对拟南芥(Arabidopsis thaliana) Col-0生态型进行甲基磺酸乙酯(ethyl meth- ane sulphonate, EMS)诱变, 筛选出1个细胞死亡突变体mod1 (mosaic death 1), 并通过图位克隆鉴定出MOD1基因.该基因编码脂肪酸合酶复合体中的一个亚基——烯酰基酰基载体蛋白还原酶(enoyl-acyl carrier protein reductase, enoyl-ACP reductase), 其参与催化叶绿体中脂肪酸的合成, 负调控植物PCD (Mou et al., 2000).随后, 该研究组发现, mod1突变体中存在明显的ROS积累, 暗示ROS的过量积累与该突变体的细胞死亡表型相关.为探明其中的具体作用机制, 他们针对mod1突变体构建了其T-DNA插入突变体库, 从中筛选出能够抑制mod1细胞死亡和ROS积累表型的抑制突变体soms (suppressors of mod1), 并克隆了这些抑制突变体对应的抑制基因, 发现它们编码线粒体电子传递链复合体I (mETC复合体I)的亚基, 或影响mETC复合体I的组装及活性, 表明完整的mETC复合体I在mod1突变体的PCD中起关键作用(Wu et al., 2015).mod1是叶绿体中脂肪酸合酶的突变体, 其PCD表型却能被线粒体中ETC复合体I的功能缺失所恢复, 由此暗示在叶绿体中确实产生了某种信号并传递到了线粒体, 线粒体接受该信号后通过ETC产生ROS并引发PCD.那么, 这个信号是什么? 它又是如何从叶绿体传递到线粒体的? 最近, 李家洋研究组在Cell Research杂志上发表的研究论文对该问题进行了精彩且完整的解答(Zhao et al., 2018). ... ... 叶绿体与线粒体间信号交流调控植物程序性细胞死亡(PCD)的工作模型(改自Zhao et al., 2018) 植物MOD1基因编码烯酰基酰基载体蛋白还原酶, 该酶以NADH作为共同底物参与催化叶绿体中脂肪酸的合成.在mod1突变体中, MOD1的功能缺失或表达量降低导致NADH/H+在叶绿体中大量积累; 过量的NADH/H+使叶绿体中的草酰乙酸在plNAD-MDH的作用下被还原为苹果酸; 携带还原力的苹果酸通过定位于叶绿体内膜上的DiT1转运蛋白转运到细胞质中, 并进一步通过定位于线粒体膜上的一种未知转运蛋白转移到线粒体; 在线粒体中, mMDH1将携带还原力的苹果酸脱氢转化为草酰乙酸, 同时生成NADH/H+.线粒体中积累的NADH/H+为线粒体膜上的mETC复合体I (CI)提供电子, 产生活性氧(ROS), 最终导致mod1突变体细胞发生PCD. ... ... A proposed model explaining the programmed cell death in plants regulated by chloroplast-to-mitochondrion communication (modified from Zhao et al., 2018) In plants, MOD1 encodes an enoyl-acyl carrier protein (ACP) reductase, which is involved in catalyzing the de novo biosynthesis of fatty acids in plastids using NADH as cosubstrate. The deficiency of MOD1 in mod1 mutant leads to an increased level of NADH/H+ in the chloroplast, which drives oxaloacetate to be converted to malate by plNAD-MDH. Malate carrying the reducing equivalents is transported out of the chloroplast into the cytosol by DiT1, which is localized in the chloroplast inner envelope membrane, and then is trans- ported into the mitochondrion by an unidentified transporter localized in the mitochondrion membrane. In the mitochondrion, malate is converted to oxaloacetate by mMDH1, and simultaneously NADH/H+ is generated to provide electrons for mETC complex I (CI) to induce the ROS formation and initiate the PCD process in the mod1 cells. ...


{kind=link}
{kind=link}