Abstract: In this study, genome-wide identification of ERF family genes in ‘zhongyousihao’ peach was studied by using MEGA 6.0, MEME, GSDS, DNAMAN 6.0. We identified 102 genes, and phylogenetic analyses of the genes indicated that the transcription factors can be classified into 10 different groups (I-X). Gene structure analysis showed that 81 genes had no introns, 20 genes contained an intron, and 1 gene contained 5 introns, which had a large difference from other members. Conservative motif analysis showed that the ERF family contained 20 conserved elements: Motifs 1, 2 and 4 belonged to the APETALA2 (AP2)/ERF domain, and the same conserved motif appeared mainly in the same family; the function of most conserved motifs is unknown. qRT-PCR revealed that the expression of PpeERF068 during dormancy and germination was consistent with the developmental status of leaf buds, which suggested that it might be related to leaf bud germination. The gene expression of peach buds during each period in light culture incubation further showed the relation between PpeERF068 and the germination of leaf buds. So PpeERF068 was named PpeEBB1. We will further study the molecular mechanism of PpeEBB1 to provide theoretical guidance for the cultivation and management of peach.
Key words:peach ; ERF family ; bud germination ; PpeEBB1
2 结果与讨论2.1 桃ERF转录因子家族成员在桃中鉴定出102个ERF家族成员并对其进行编号, PpeERF基因开放式阅读框长度在417-2 115 bp之间, 蛋白长度为138-704 aa, 分子质量为15 204.7- 77 761.3 Da, 等电点为4.58-10.37。详细信息及各基因对应的表达序列标签数目见表2。 表2 Table 2 表2 表2 桃ERF家族基因及其相关信息 Table 2 List of PpeERF family genes and their related information
Group name
Gene name
Locus name
ORF length (bp)
Size (aa)
Molecular weight (Da)
PI
EST hitting
I
PpeERF#001
Prupe.6g182200
1122
373
41331.3
6.56
13
I
PpeERF#002
Prupe.3g009400
642
213
23101.2
9.79
7
I
PpeERF#003
Prupe.1g513600
1137
378
41236.8
6.37
9
I
PpeERF#004
Prupe.5g117800
933
310
34377.6
8.34
9
I
PpeERF#005
Prupe.3g157100
1386
461
51713.0
5.97
0
I
PpeERF#006
Prupe.1g008400
1230
409
45352.9
7.81
2
I
PpeERF#007
Prupe.8g198700
696
231
26301.6
9.19
0
II
PpeERF#008
Prupe.6g176400
540
179
20308.7
9.51
0
II
PpeERF#009
Prupe.1g432000
528
175
19844.9
9.39
1
II
PpeERF#010
Prupe.1g464400
453
150
17240.2
9.42
2
II
PpeERF#011
Prupe.6g231500
501
166
17849.1
8.72
0
II
PpeERF#012
Prupe.1g546100
501
166
17890.7
5.62
0
II
PpeERF#013
Prupe.1g500900
726
241
26308.4
5.61
0
II
PpeERF#014
Prupe.7g194400
648
215
23980.2
5.16
0
II
PpeERF#015
Prupe.1g001700
552
183
20044.5
7.74
0
III
PpeERF#016
Prupe.5g090700
558
185
19627.0
4.89
0
III
PpeERF#017
Prupe.5g090600
537
178
19407.7
5.67
0
III
PpeERF#018
Prupe.5g090400
543
180
19547.0
5.50
0
III
PpeERF#019
Prupe.5g090300
543
180
19510.8
5.50
0
III
PpeERF#020
Prupe.5g090800
555
184
19705.9
4.64
0
III
PpeERF#021
Prupe.2g289600
696
231
24485.1
4.83
0
III
PpeERF#022
Prupe.7g116300
609
202
21536.7
4.91
0
III
PpeERF#023
Prupe.2g190200
609
202
22713.9
5.87
1
III
PpeERF#024
Prupe.2g123200
831
276
30098.8
4.94
0
III
PpeERF#025
Prupe.5g065300
762
253
28183.5
4.91
0
III
PpeERF#026
Prupe.1g448600
771
256
27960.9
5.11
0
III
PpeERF#027
Prupe.7g222700
789
262
28536.4
4.94
0
III
PpeERF#028
Prupe.3g062800
732
243
26436.2
6.67
1
III
PpeERF#029
Prupe.7g115900
606
201
22481.2
4.95
0
III
PpeERF#030
Prupe.2g289500
690
229
24976.9
5.14
7
III
PpeERF#031
Prupe.5g090500
723
240
27217.8
7.69
0
III
PpeERF#032
Prupe.5g090100
711
236
26352.7
5.17
0
III
PpeERF#033
Prupe.5g090000
693
230
25658.8
5.13
7
III
PpeERF#034
Prupe.5g090200
699
232
26157.5
5.03
1
III
PpeERF#035
Prupe.5g089900
693
230
25521.9
7.78
0
III
PpeERF#036
Prupe.1g545700
522
173
19078.3
5.44
0
III
PpeERF#037
Prupe.1g545400
600
199
22101.0
5.54
0
IV
PpeERF#038
Prupe.2g253000
1041
346
37474.1
6.52
0
IV
PpeERF#039
Prupe.6g284400
1554
517
58370.7
4.74
0
IV
PpeERF#040
Prupe.2g256900
1167
388
42473.1
4.64
3
IV
PpeERF#041
Prupe.3g223300
591
196
21305.5
8.32
0
IV
PpeERF#042
Prupe.7g066700
858
285
32054.9
5.73
0
Group name
Gene name
Locus name
ORF length (bp)
Size (aa)
Molecular weight (Da)
PI
EST hitting
IV
PpeERF#043
Prupe.6g354000
672
223
23337.9
8.50
0
IV
PpeERF#044
Prupe.1g372100
768
255
28137.6
6.55
0
V
PpeERF#045
Prupe.3g096000
639
212
24310.2
5.94
0
V
PpeERF#046
Prupe.1g390800
588
195
22413.4
6.66
6
V
PpeERF#047
Prupe.1g480400
672
223
24759.7
9.10
0
V
PpeERF#048
Prupe.7g243600
732
243
26570.6
7.02
0
V
PpeERF#049
Prupe.3g084600
567
188
20855.3
5.97
0
V
PpeERF#050
Prupe.7g004900
945
314
34636.3
5.44
0
V
PpeERF#051
Prupe.6g004400
867
288
31688.5
6.01
0
V
PpeERF#052
Prupe.3g263000
1275
424
46904.3
4.58
0
V
PpeERF#053
Prupe.5g136200
939
312
33152.5
6.13
0
V
PpeERF#054
Prupe.1g084800
936
311
33626.0
8.92
0
V
PpeERF#055
Prupe.5g213800
675
224
25367.4
8.71
0
VI
PpeERF#056
Prupe.5g114100
1002
333
37019.0
4.98
0
VI
PpeERF#057
Prupe.2g306400
1098
365
41098.9
4.75
2
VI
PpeERF#058
Prupe.2g183200
864
287
32269.0
4.95
0
VI
PpeERF#059
Prupe.3g019900
570
189
20614.4
9.37
0
VI
PpeERF#060
Prupe.7g060700
588
195
21933.2
10.37
2
VI
PpeERF#061
Prupe.6g039700
927
308
34031.9
5.49
2
VI
PpeERF#062
Prupe.5g220700
1311
436
49386.8
6.25
1
VI
PpeERF#063
Prupe.1g310100
1020
339
38021.2
4.92
1
VII
PpeERF#064
Prupe.8g264900
966
321
35682.6
5.68
4
VII
PpeERF#065
Prupe.3g032300
1149
382
42495.9
4.81
84
VII
PpeERF#066
Prupe.1g130300
870
289
31564.0
7.66
31
VIII
PpeERF#067
Prupe.3g094700
870
289
31781.0
9.56
0
VIII
PpeERF#068
Prupe.1g245500
1275
424
46297.0
7.73
0
VIII
PpeERF#069
Prupe.1g212700
831
276
29938.4
4.94
0
VIII
PpeERF#070
Prupe.1g139600
1092
363
39938.7
5.60
0
VIII
PpeERF#071
Prupe.4g176200
687
228
24245.0
9.62
2
VIII
PpeERF#072
Prupe.4g051400
795
264
28447.6
9.47
1
VIII
PpeERF#073
Prupe.8g230600
546
181
19951.0
9.99
0
VIII
PpeERF#074
Prupe.4g051200
519
172
18303.7
9.20
3
VIII
PpeERF#075
Prupe.4g222300
696
231
25022.8
7.62
20
VIII
PpeERF#076
Prupe.3g209100
708
235
25471.3
6.58
3
IX
PpeERF#077
Prupe.2g129700
672
223
24950.8
5.41
0
IX
PpeERF#078
Prupe.2g129500
651
216
23968.9
5.99
0
IX
PpeERF#079
Prupe.2g129600
714
237
25948.7
5.47
0
IX
PpeERF#080
Prupe.2g129300
816
271
29817.0
5.17
0
IX
PpeERF#081
Prupe.2g129400
621
206
22559.1
5.97
0
IX
PpeERF#082
Prupe.5g061800
846
281
30157.0
9.09
10
IX
PpeERF#083
Prupe.6g064700
2115
704
77761.3
9.77
15
IX
PpeERF#084
Prupe.2g272300
783
260
29210.8
6.53
3
IX
PpeERF#085
Prupe.5g062000
1059
352
39322.2
5.90
5
Group name
Gene name
Locus name
ORF length (bp)
Size (aa)
Molecular weight (Da)
PI
EST hitting
IX
PpeERF#086
Prupe.2g272400
945
314
35018.8
6.17
21
IX
PpeERF#087
Prupe.2g272500
726
241
27069.2
5.92
0
IX
PpeERF#088
Prupe.8g224700
540
179
20470.7
6.08
0
IX
PpeERF#089
Prupe.1g037800
492
163
18008.7
6.85
0
IX
PpeERF#090
Prupe.8g224800
465
154
17059.7
7.79
0
IX
PpeERF#091
Prupe.1g037700
417
138
15204.7
6.84
0
IX
PpeERF#092
Prupe.4g055600
795
264
29431.3
5.61
0
IX
PpeERF#093
Prupe.4g055500
564
187
20369.6
9.51
2
IX
PpeERF#094
Prupe.6g348700
744
247
28089.7
4.92
0
IX
PpeERF#095
Prupe.8g224600
759
252
28672.0
4.99
1
IX
PpeERF#096
Prupe.1g037900
699
232
25950.8
5.54
1
X
PpeERF#097
Prupe.8g125100
816
271
30318.2
8.16
0
X
PpeERF#098
Prupe.7g134100
801
266
29255.4
7.67
0
X
PpeERF#099
Prupe.5g141200
696
231
25763.5
9.21
0
X
PpeERF#100
Prupe.6g165700
1215
404
43314.9
7.07
2
X
PpeERF#101
Prupe.5g141300
816
271
30005.0
6.16
0
X
PpeERF#102
Prupe.1g214900
741
246
27203.7
6.23
1
表2 桃ERF家族基因及其相关信息 Table 2 List of PpeERF family genes and their related information
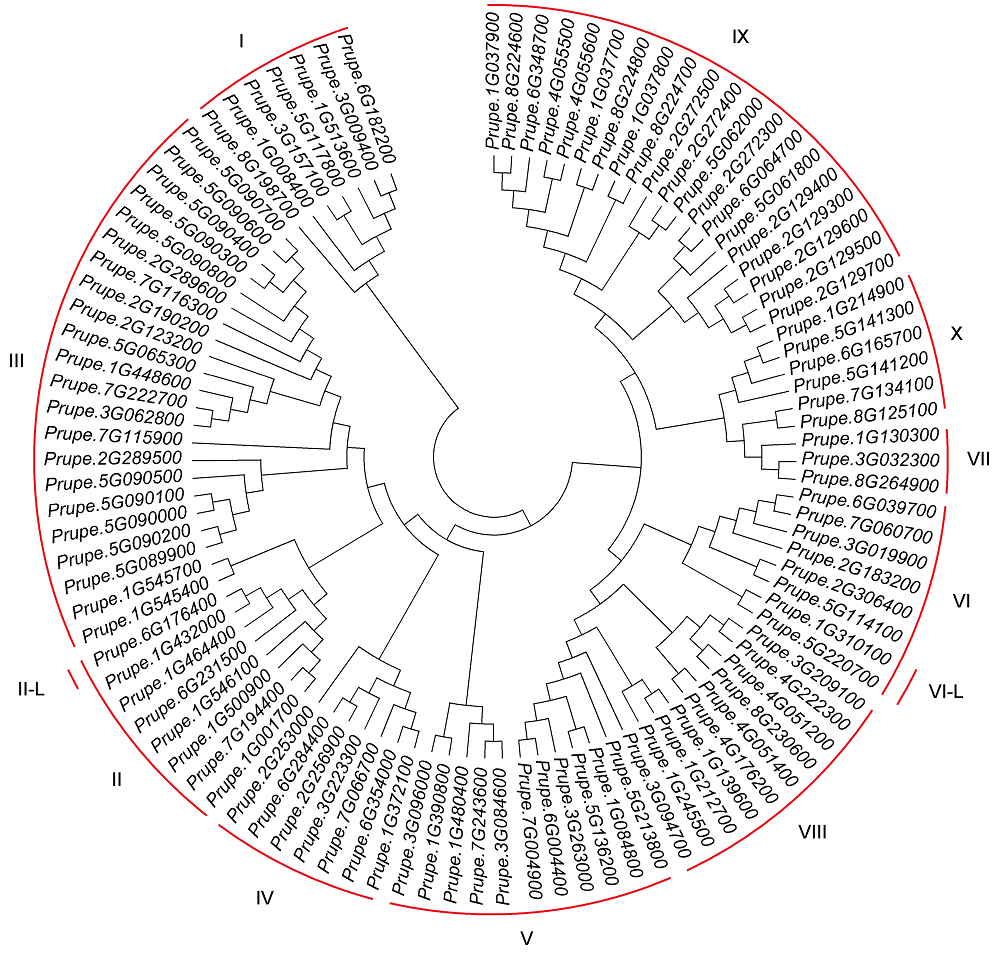
2.2 桃ERF家族成员系统进化树根据Nakano等(2006)的方法构建系统进化树, 对桃ERF家族成员的AP2/ERF结构域序列进行聚类分析, 将桃ERF家族分为10个子家族(I-X) (图1)。其中III是最大的子家族, 包含22个成员, 而在拟南芥中, III也是ERF最大的子家族, 包含23个成员; 其次为IX子家族, 含有20个成员; 另外, 通过比对ERF家族AP2/ERF结构域的蛋白序列(图2), 发现II子家族的Prupe.6g176400、VI子家族的Prupe.5g220700和Prupe.1g310100 C端的序列与ERF家族中的其它成员同源性非常低, 因此将其分别命名为II-like (II-L)和VI-like (VI-L)。 图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-612/img_1.png<b>图1</b> 桃ERF家族基因系统进化树<br/>I-X分别表示桃ERF家族的10个子家族; II-L: II-like; VI-L: VI-like<br/><b>Figure 1</b> The phylogenetic tree of peach ERF family genes<br/>I-X indicate the 10 different groups of ERF family, respectively; II-L: II-like; VI-L: VI-like Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-612/img_1.png<b>图1</b> 桃ERF家族基因系统进化树<br/>I-X分别表示桃ERF家族的10个子家族; II-L: II-like; VI-L: VI-like<br/><b>Figure 1</b> The phylogenetic tree of peach ERF family genes<br/>I-X indicate the 10 different groups of ERF family, respectively; II-L: II-like; VI-L: VI-like
图1 桃ERF家族基因系统进化树 I-X分别表示桃ERF家族的10个子家族; II-L: II-like; VI-L: VI-like Figure 1 The phylogenetic tree of peach ERF family genes I-X indicate the 10 different groups of ERF family, respectively; II-L: II-like; VI-L: VI-like
图2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-612/img_2.png<b>图2</b> 桃ERF家族基因AP2/ERF保守结构域氨基酸序列比对<br/><b>Figure 2</b> Amino acids sequence alignment of AP2/ERF domain of ERF family genes in peach Figure 2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-612/img_2.png<b>图2</b> 桃ERF家族基因AP2/ERF保守结构域氨基酸序列比对<br/><b>Figure 2</b> Amino acids sequence alignment of AP2/ERF domain of ERF family genes in peach
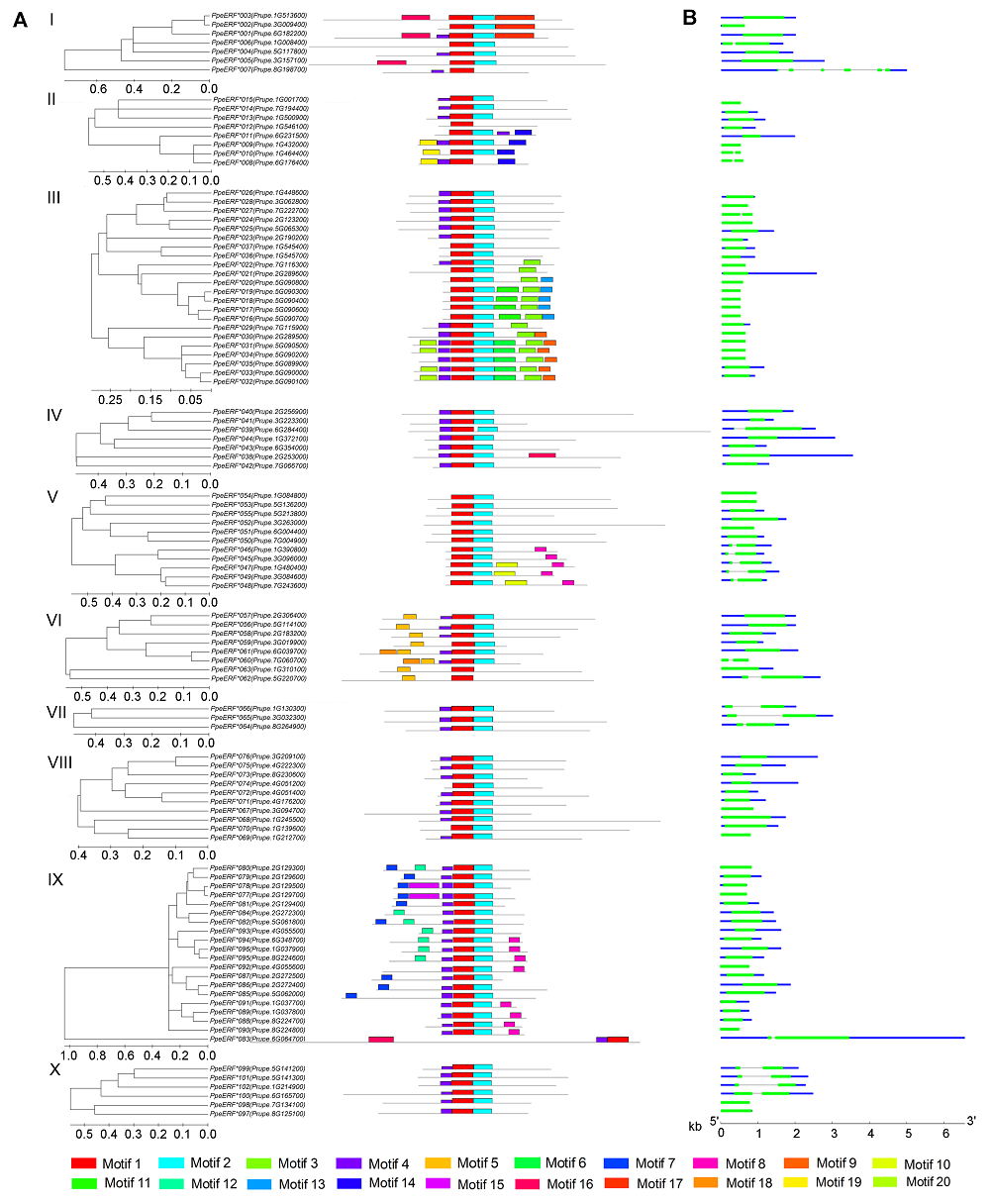
图3 桃ERF家族保守元件(A)和基因结构(B) I-X分别代表桃ERF家族的10个子家族。 Figure 3 Conserved motif (A) and structures (B) of ERF genes in peach I-X indicate the 10 different groups of ERF family, respectively.
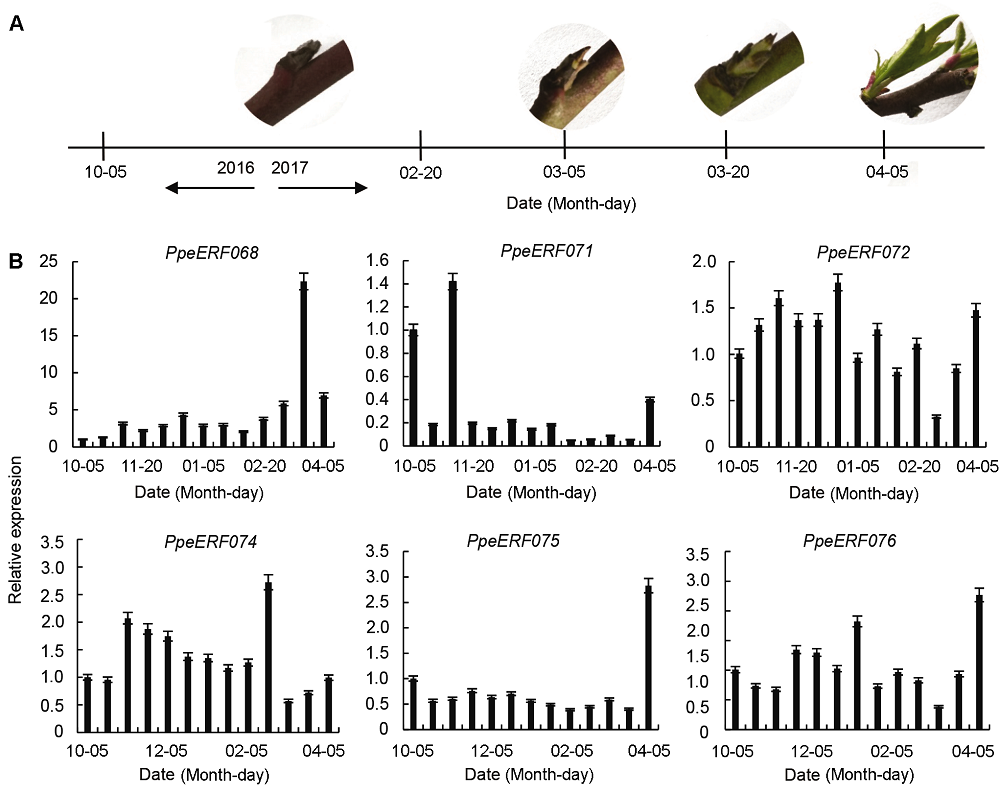
2.4 桃芽休眠及萌发过程中VIII子家族基因的表达2.4.1 桃芽发育过程 桃芽发育过程如图4A所示。在2016年10月5日至2017年2月20日期间, 桃芽被厚厚的鳞片包裹, 未见有萌发迹象; 到3月5日, 桃芽膨大, 芽背面变绿或浅红; 3月20日桃芽开始萌发; 4月5日, 长出叶片并伸长至1-2 cm。 图4https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-612/img_4.png<b>图4</b> 自然条件下桃芽发育过程(A)及VIII子家族基因的表达(B)<br/><b>Figure 4</b> Bud development process (A) and relative expression of VIII sub-family (B) in peach under natural condition Figure 4https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-612/img_4.png<b>图4</b> 自然条件下桃芽发育过程(A)及VIII子家族基因的表达(B)<br/><b>Figure 4</b> Bud development process (A) and relative expression of VIII sub-family (B) in peach under natural condition
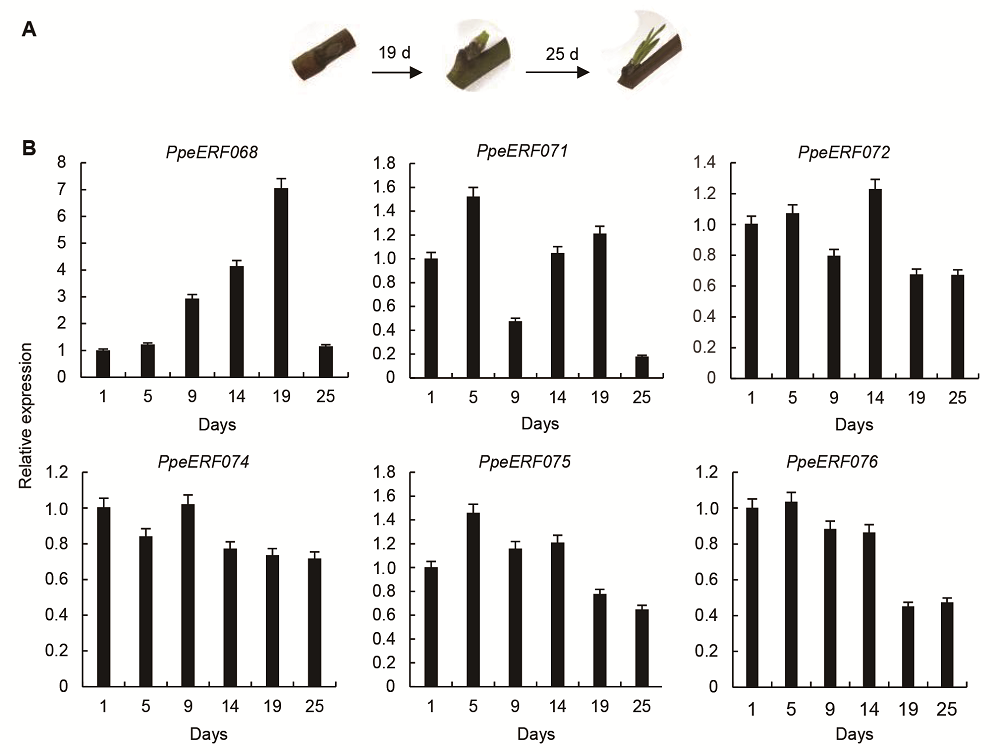
图5 光照培养箱中桃芽萌发过程(A)及VIII子家族基因的表达(B) Figure 5 Bud development process (A) and relative expression of VIII sub-family genes (B) in peach in light culture incubation
2.5 讨论在同一个基因家族中, 拥有共同保守元件的基因可能具有相似的功能, 虽然大部分保守元件的功能未知, 但它们可能在转录调控中发挥重要作用(Nakano et al., 2006)。大部分保守元件选择性地分布在同一个进化树分支上, 表明同一个子家族中的成员具有相似的结构(图3)。本研究II子家族基因中, ERF8-11 C端的Motif 14有1个DLNxxP结构。研究表明, 1个B3结构域阻遏蛋白在缺失这个保守元件后, 其阻遏作用减弱(Tsukagoshi et al., 2005)。III子家族ERF16-22与29- 35 C端的Motif 3、Motif 9和Motif 13; IX子家族N端的Motif 7和Motif 12可能与转录激活有关(Fujimoto et al., 2000)。另外, 也有研究表明, LWSY motif可能与植物抗寒性有关。现有研究表明, 拟南芥中ERF家族I子家族的WXP1和V子家族中的WIN1/SHN1是蜡质合成中的重要基因(Aharoni et al., 2004; Broun et al.,2004)。III、VI、VII和IX子家族中已获得功能鉴定的部分基因与植物抗性有关(Park et al., 2001; Magome et al., 2004; Yi et al., 2004; 翟莹等, 2013)。VIII子家族中的LEP和ESR1/DRN参与器官分化(Chandler et al., 2011), ESR1/DRN在杨树中的同源基因EBB1功能已得到证实, 其能够促进杨树提前萌发(Yordanov et al., 2014)。VIII子家族中的ERF7和X子家族中的ABR1能够响应脱落酸信号(Song et al., 2005; Pandey et al., 2005)。可见, 不同基因的功能与所在的家族并不完全对应, 具有同样功能的基因可能存在于不同的家族中, 具有不同功能的基因也可能存在于同一家族。但是, 鉴于同一家族中基因的结构较为相似, 并有相似的保守元件, 某一功能已知基因也能够在一定程度上反映该家族中其它基因的功能。 Sherif等(2013)将桃PpeERF3b (对应于本文中PpeERF076)在烟草(Nicotiana tabacum)中异位表达后, 发现该基因可以促进烟草侧芽萌发。由于PpeERF3b属于VIII子家族, 鉴于同一家族基因结构相似性较高, 我们检测了VIII子家族各基因在桃芽休眠及萌发过程中的表达量并与桃芽的发育状态进行比较分析, 发现只有PpeERF068的表达与芽的萌发状态完全一致, 因此最有可能与桃芽萌发相关。另外, 我们通过对比图4和图5中PpeERF068的表达情况以及相应的芽萌发状态, 发现桃芽在3月20日即将萌发以及在培养箱中培养至19天已经萌发时, PpeERF- 068的表达量分别是10月20日休眠时的17.5倍和7.0倍, 表明PpeERF068可能更多地参与促进芽萌发这一进程。将该基因序列与其它物种进行比对后, 我们发现该基因与杨树中已经鉴定的与萌发相关的EBB1基因同源性最高, 所以我们将其命名为PpeEBB1。Yordanov等(2014)研究表明, EBB1促进杨树萌发是由于起始了顶端分生组织和叶原基的细胞分裂, 但并未得出具体的调控途径。前人对拟南芥的研究认为, ESR1和ESR2参与拟南芥侧枝分化主要通过调控生长素的运输实现, ESR1和ESR2在拟南芥茎顶端分生组织的生长素调控网络中扮演重要角色(Chandler et al., 2011; Chandler and Werr, 2014); 而Sherif等(2013)认为桃PpeERF3b在烟草中异位表达能够促进烟草侧枝生成是通过干扰生长素生物合成、运输或信号转导实现。木本植物芽萌发与拟南芥和烟草侧枝分化可能存在差异, 因此PpeEBB1对桃芽萌发的调控是否通过生长素实现还不得而知。我们将进一步研究PpeEBB1调控芽萌发的分子机制, 以期揭示由该基因控制桃芽萌发的相关途径, 从而为桃树的栽培管理和熟期调控提供理论依据。
The authors have declared that no competing interests exist.
AharoniA, DixitS, JetterR, ThoenesE, van ArkelG, PereiraA (2004). The SHINE clade of AP2 domain transcription factors activates wax biosynthesis, alters cuticle properties, and confers drought tolerance when overexpressed in Arabidopsis.Plant Cell 16, 2463-2480. [本文引用: 1]
[7]
BrounP, PoindexterP, OsborneE, JiangCZ, RiechmannJL (2004). WIN1, a transcriptional activator of epidermal wax accumulation in Arabidopsis.Proc Natl Acad Sci USA 101, 4706-4711. [本文引用: 2]
[8]
CarlesCC, FletcherJC (2003). Shoot apical meristem maintenance: the art of a dynamic balance.Trends Plant Sci 8, 394-401. [本文引用: 1]
[9]
ChandlerJW, ColeM, JacobsB, ComelliP, WerrW (2011). Genetic integration of DORNRÖSCHEN and DORNRÖSCHEN-LIKE reveals hierarchical interactions in auxin signaling and patterning of the Arabidopsis apical embryo. Plant Mol Biol 75, 223-236. [本文引用: 2]
[10]
ChandlerJW, WerrW (2014). Arabidopsis floral phytomer development: auxin response relative to biphasic modes of organ initiation.J Exp Bot 65, 3097-3110. [本文引用: 1]
[11]
ChenM, TanQP, SunMY, LiDM, FuXL, ChenXD, XiaoW, LiL, GaoDS (2016). Genome-wide identification of WRKY family genes in peach and analysis of WRKY expression during bud dormancy. Mol Genet Genomics 291, 1319-1332. [本文引用: 1]
[12]
DubouzetJG, SakumaY, ItoY, KasugaM, DubouzetEG, MiuraS, SekiM, ShinozakiK, Yamaguchi-ShinozakiK (2003). OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in drought-, high-salt and cold-responsive gene expression. Plant J 33, 751-763. [本文引用: 1]
[13]
ElliottRC, BetznerAS, HuttnerE, OakesMP, TuckerWQ, GerentesD, PerezP, SmythDR (1996). AINTEGUMENTA, an APETALA2-like gene of Arabidopsis with pleiotropic roles in ovule development and floral organ growth. Plant Cell 8, 155-168. [本文引用: 1]
[14]
FergusonBJ, BeveridgeCA (2009). Roles for auxin, cytokinin, and strigolactone in regulating shoot branching.Plant Physiol 149, 1929-1944. [本文引用: 1]
[15]
FujimotoSY, OhtaM, UsuiA, ShinshiH, Ohme-TakagiM (2000). Arabidopsis ethylene-responsive element binding factors act as transcriptional activators or repressors of GCC box-mediated gene expression.Plant Cell 12, 393-404. [本文引用: 1]
[16]
GuYQ, WildermuthMC, ChakravarthyS, LohYT, YangCM, HeXH, HanY, MartinGB (2002). Tomato transcription factors Pti4, Pti5, and Pti6 activate defense responses when expressed in Arabidopsis.Plant Cell 14, 817-831. [本文引用: 1]
[17]
HuYX, WangYX, LiuXF, LiJY (2004). Arabidopsis RAV1 is down-regulated by brassinosteroid and may act as a negative regulator during plant development.Cell Res 14, 8-15. [本文引用: 1]
[18]
MagomeH, YamaguchiS, HanadaA, KamiyaY, OdaK (2004). dwarf and delayed-flowering 1, a novel Arabidopsis mutant deficient in gibberellin biosynthesis because of overexpression of a putative AP2 transcription factor. Plant J 37, 720-729. [本文引用: 1]
[19]
MooseSP, SiscoPH (1996). Glossy15, an APETALA2-like gene from maize that regulates leaf epidermal cell identity. Genes Dev 10, 3018-3027. [本文引用: 1]
[20]
NakanoT, SuzukiK, FujimuraT, ShinshiH (2006). Genome-wide analysis of the ERF gene family in Arabidopsis and rice.Plant Physiol 140, 411-432. [本文引用: 3]
[21]
PandeyGK, GrantJJ, CheongYH, KimBG, LiLG, LuanS (2005). ABR1, an APETALA2-domain transcription factor that functions as a repressor of ABA response in Arabidopsis.Plant Physiol 139, 1185-1193. [本文引用: 1]
[22]
ParkJM, ParkCJ, LeeSB, HamBK, ShinR, PaekKH (2001). Overexpression of the tobacco Tsi1 gene encoding an EREBP/AP2-type transcription factor enhances resistance against pathogen attack and osmotic stress in tobacco. Plant Cell 13, 1035-1046. [本文引用: 1]
[23]
Savaldi-GoldsteinS, ChoryJ (2008). Growth coordination and the shoot epidermis.Curr Opin Plant Biol 11, 42-48. [本文引用: 1]
[24]
SherifS, El-SharkawyI, PaliyathG, JayasankarS (2013). PpERF3b, a transcriptional repressor from peach, contribu- tes to disease susceptibility and side branching in EAR- dependent and -independent fashions.Plant Cell Rep 32, 1111-1124. [本文引用: 3]
[25]
SongCP, AgarwalM, OhtaM, GuoY, HalfterU, WangPC, ZhuJK (2005). Role of an Arabidopsis AP2/EREBP-type transcriptional repressor in abscisic acid and drought stress responses.Plant Cell 17, 2384-2396. [本文引用: 1]
[26]
TsukagoshiH, SaijoT, ShibataD, MorikamiA, NakamuraK (2005). Analysis of a sugar response mutant of Arabidopsis identified a novel B3 domain protein that functions as an active transcriptional repressor.Plant Physiol 138, 675-685. [本文引用: 1]
[27]
XuW, LiF, LingLZ, LiuAZ (2013). Genome-wide survey and expression profiles of the AP2/ERF family in castor bean(Ricinus communis L.). BMC Genomics 14, 785. [本文引用: 1]
[28]
XuZS, ChenM, LiLC, MaYZ (2011). Functions and application of the AP2/ERF transcription factor family in crop improvement.J Integr Plant Biol 53, 570-585. [本文引用: 1]
[29]
YiSY, KimJH, JoungYH, LeeS, KimWT, YuSH, ChoiD (2004). The pepper transcription factor CaPF1 confers pathogen and freezing tolerance in Arabidopsis.Plant Phy- siol 136, 2862-2874. [本文引用: 1]
[30]
YordanovYS, MaC, StraussSH, BusovVB (2014). EARLY BUD-BREAK 1 (EBB1) is a regulator of release from seasonal dormancy in poplar trees.Proc Natl Acad Sci USA 111, 10001-10006. [本文引用: 3]
[31]
ZhangGY, ChenM, ChenXP, XuZS, GuanS, LiLC, LiAL, GuoJM, MaoL, MaYZ (2008). Phylogeny, gene structures, and expression patterns of the ERF gene family in soybean (Glycine max L.). J Exp Bot 59, 4095-4107. [本文引用: 1]
[32]
ZhengCL, HalalyT, AcheampongAK, TakebayashiY, JikumaruY, KamiyaY, OrE (2015). Abscisic acid (ABA) regulates grape bud dormancy, and dormancy release stimuli may act through modification of ABA metabolism.J Exp Bot 66, 1527-1542. [本文引用: 1]
[33]
ZhuangJ, CaiB, PengRH, ZhuB, JinXF, XueY, GaoF, FuXY, TianYS, ZhaoW, QiaoYS, ZhangZ, XiongAS, YaoQH (2008). Genome-wide analysis of the AP2/ERF gene family in Populus trichocarpa. Biochem Biophys Res Commun 371, 468-474. [本文引用: 1]
剥鳞和激素处理对大樱桃花芽休眠解除及内源激素变化的影响 1 2004
... 休眠是多年生木本植物的重要生理特征, 是由不适宜的环境因素导致的枝条暂时停止生长.休眠芽只有达到一定的需冷量时才能够在适宜的环境条件下重新萌发.芽的萌发是新的起始, 对植物有重要影响.一方面, 促进果树的萌发能够使果实提前上市, 从而提高经济效益; 另一方面, 萌发的芽可能会因遇到冻害等恶劣天气而导致减产.因此, 了解萌发机制对于指导果树生产实践具有重要意义.已有研究表明, 植物激素的含量与萌发密切相关, 如脱落酸和独脚金内酯促进休眠(Ferguson and Beveridge, 2009; Zheng et al., 2015), 而赤霉素、细胞分裂素以及油菜素内酯等能够促进萌发(谭志一等, 1985; 段成国等, 2004; Savaldi-Goldstein and Chory, 2008). ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}