Abstract: tRNA derived fragments (termed tRFs) are functional RNAs produced by non-random cleavage of tRNAs. The knowledge of tRNAs’ production and function is being studied but is unknown in the rice male gametophyte. Using high-throughput sequencing with 3 stages in the rice male gametophyte development, we found tRFs with a large range of length. Next, we determined four sequence-specific and one non-sequence-specific ribonuclease cleaving sites. We also summarized the distribution profile of tRFs in the three stages. Finally, a NCBI blast search was carried out to predict the tRF target gene, revealing that the major class of targets was transposable elements. This results provid new clues for rice male gametophyte research.
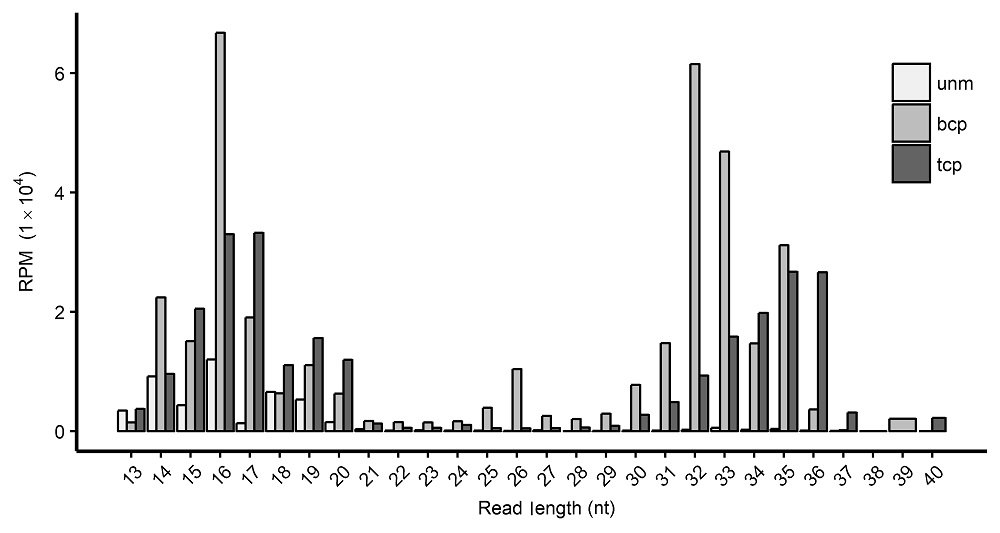
2 结果与讨论2.1 水稻雄配子三类花粉的tRF长度分布对水稻雄配子单核小孢子、二细胞花粉和三细胞花粉tRF长度进行统计(图1), 数量用“每百万条读段里的 (目标)读段数”(reads per million, RPM)来标准化, 发现它们主要分布于2个区间: 14-20 nt和31-36 nt。 图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-625/img_1.png<b>图1</b> 水稻雄配子三类花粉中的tRF长度分布 <br/>unm: 单核小孢子; bcp: 二细胞花粉; tcp: 三细胞花粉; RPM: 每百万条读段里的(目标)读段数<br/><b>Figure 1</b> Length distribution of tRFs in three types of pollens in rice male gamete<br/>unm: Uni-nucleate microspore; bcp: Bi-cellular pollen; tcp : Tri-cellular pollen; RPM: Reads per million Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-625/img_1.png<b>图1</b> 水稻雄配子三类花粉中的tRF长度分布 <br/>unm: 单核小孢子; bcp: 二细胞花粉; tcp: 三细胞花粉; RPM: 每百万条读段里的(目标)读段数<br/><b>Figure 1</b> Length distribution of tRFs in three types of pollens in rice male gamete<br/>unm: Uni-nucleate microspore; bcp: Bi-cellular pollen; tcp : Tri-cellular pollen; RPM: Reads per million
图1 水稻雄配子三类花粉中的tRF长度分布 unm: 单核小孢子; bcp: 二细胞花粉; tcp: 三细胞花粉; RPM: 每百万条读段里的(目标)读段数 Figure 1 Length distribution of tRFs in three types of pollens in rice male gamete unm: Uni-nucleate microspore; bcp: Bi-cellular pollen; tcp : Tri-cellular pollen; RPM: Reads per million
三类花粉在14-20 nt区间分布规律相似。在植物研究中, 长度在~16 nt的tRF几乎未见报道, 而在本研究中的三类花粉中均较多。~19 nt长度的tRF在多项研究中已有报道, 如水稻营养组织、小立碗藓(Phy- scomitrella patens) (Alves et al., 2016)和缺磷条件下的拟南芥根中(Hsieh et al., 2010)。 三类花粉的tRF在31-36 nt区间的表现差异很大。unm中含量很少, bcp中主要集中于~32 nt, tcp中则倾向于~35 nt (图1)。可见14-20 nt和31-36 nt两个区间tRF的形成机理可能不同。
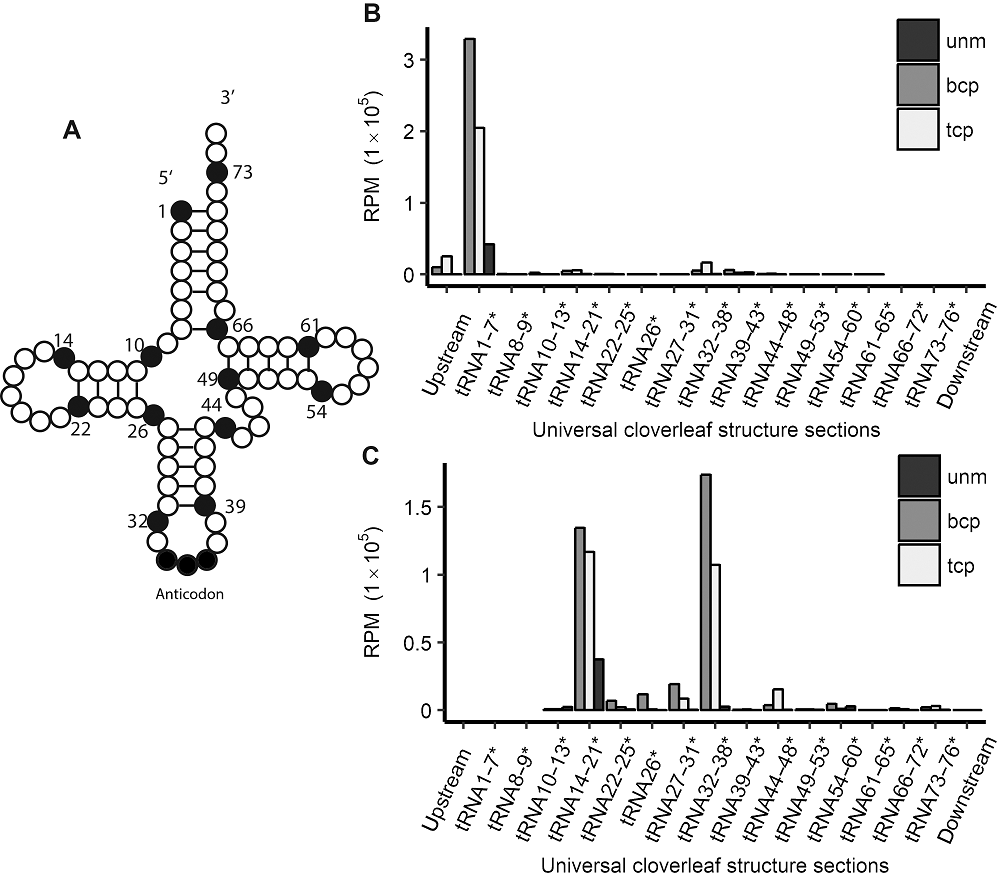
2.2 水稻雄配子三类花粉的序列特征我们发现, 水稻雄配子三类花粉tRF的起始位置主要位于tRNA的5'受体臂上, 终止位置主要在D环和反密码子环上(图2)。可见产生tRF的剪切主要发生在单链的D loop和Anticodon loop上, 这正好对应2个主要的长度类群。 图2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-625/img_2.png<b>图2</b> 水稻雄配子三类花粉的tRF起始和终止位点在四叶草结构中的分布 <br/>(A) tRNA二级结构示意图; (B) tRF起点分布; (C) tRF终点分布。*代表<xref ref-type="fig" rid="F2-1674-3466-53-5-625">图2A</xref>中数字的位置, 不代表碱基编号。unm、bcp、tcp及RPM同<xref ref-type="fig" rid="F1-1674-3466-53-5-625">图1</xref>。<br/><b>Figure 2</b> tRFs’ starting and ending site distribution in tRNAs’ universal cloverleaf structure sections in three types of pollens in rice male gamete<br/>(A) Schematic view of tRNAs’ cloverleaf-like secondary structure; (B) tRFs’ starting site positions in tRNA; (C) tRFs’ ending site positions in tRNA. * indicate that the numbers before can only be regarded as a positions represented in <xref ref-type="fig" rid="F2-1674-3466-53-5-625">Figure 2A</xref> instead of base ID. unm, bcp, tcp and RPM see <xref ref-type="fig" rid="F1-1674-3466-53-5-625">Figure 1</xref>. Figure 2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-625/img_2.png<b>图2</b> 水稻雄配子三类花粉的tRF起始和终止位点在四叶草结构中的分布 <br/>(A) tRNA二级结构示意图; (B) tRF起点分布; (C) tRF终点分布。*代表<xref ref-type="fig" rid="F2-1674-3466-53-5-625">图2A</xref>中数字的位置, 不代表碱基编号。unm、bcp、tcp及RPM同<xref ref-type="fig" rid="F1-1674-3466-53-5-625">图1</xref>。<br/><b>Figure 2</b> tRFs’ starting and ending site distribution in tRNAs’ universal cloverleaf structure sections in three types of pollens in rice male gamete<br/>(A) Schematic view of tRNAs’ cloverleaf-like secondary structure; (B) tRFs’ starting site positions in tRNA; (C) tRFs’ ending site positions in tRNA. * indicate that the numbers before can only be regarded as a positions represented in <xref ref-type="fig" rid="F2-1674-3466-53-5-625">Figure 2A</xref> instead of base ID. unm, bcp, tcp and RPM see <xref ref-type="fig" rid="F1-1674-3466-53-5-625">Figure 1</xref>.
图2 水稻雄配子三类花粉的tRF起始和终止位点在四叶草结构中的分布 (A) tRNA二级结构示意图; (B) tRF起点分布; (C) tRF终点分布。*代表图2A中数字的位置, 不代表碱基编号。unm、bcp、tcp及RPM同图1。 Figure 2 tRFs’ starting and ending site distribution in tRNAs’ universal cloverleaf structure sections in three types of pollens in rice male gamete (A) Schematic view of tRNAs’ cloverleaf-like secondary structure; (B) tRFs’ starting site positions in tRNA; (C) tRFs’ ending site positions in tRNA. * indicate that the numbers before can only be regarded as a positions represented in Figure 2A instead of base ID. unm, bcp, tcp and RPM see Figure 1.
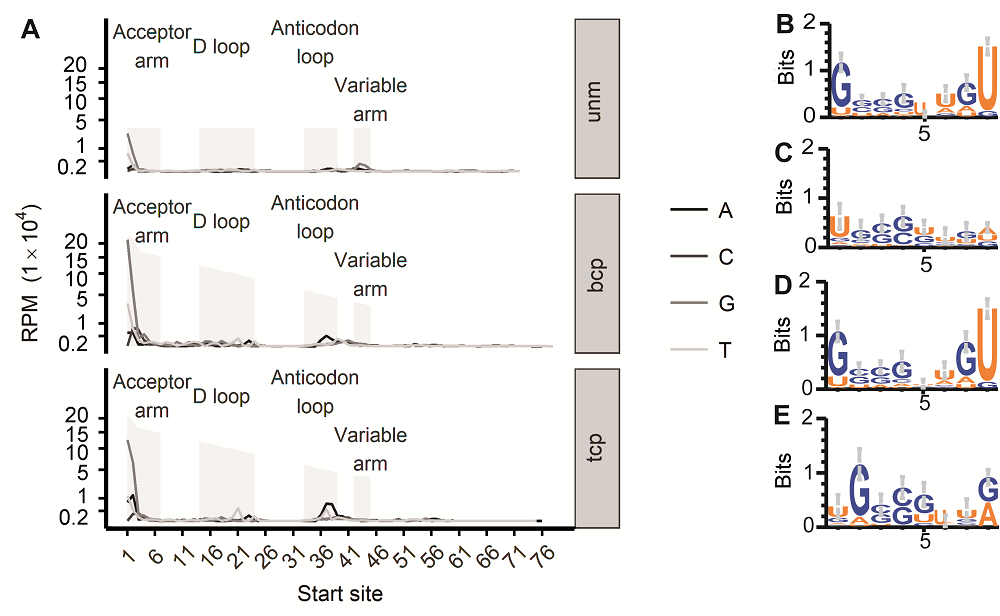
tRF5'碱基大多为G, 这与拟南芥中的碱基偏好性一致(Loss-Morais et al., 2013), 尤其是位于tRNA 5'受体臂的5'碱基(图3A, B, D)。其部分原因可以归结为tRNA在5'受体臂(尤其是前2个碱基)本就倾向于为G; 然而当tRNA第1个碱基是其它碱基的时候, tRF的起始就会延后到第2个碱基, 该现象在tcp中特别明显(图3C), 在bcp中也存在该现象, 但并不突出(图3C, E)。5'碱基倾向于G, 似乎与tRF的产生或者稳定性相关。而在反密码子环中, tRF则更多以碱基A起始(图3A), 只是其总量大不如5'受体臂起始的tRF。 图3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-625/img_3.png<b>图3</b> 水稻雄配子三类花粉的tRF 5’端碱基分布偏好性<br/>(A) 起始于tRNA各个位置的tRF起点碱基分布情况(纵坐标各点相对位置进行了1/2次幂变形处理); (B) bcp样品中tRF起始于tRNA第1位时, 对应宿主tRNA前8个碱基logo图; (C) bcp样品中tRF起始于tRNA第2位时, 对应宿主tRNA前8个碱基logo图; (D) tcp样品中tRF起始于tRNA第1位时, 对应宿主tRNA前8个碱基logo图; (E) tcp样品中tRF起始于tRNA第2位时, 对应宿主tRNA前8个碱基logo图。unm、bcp、tcp和RPM同<xref ref-type="fig" rid="F1-1674-3466-53-5-625">图1</xref>。<br/><b>Figure 3</b> tRFs’ 5’ end base bias in three types of pollens in rice male gamete<br/>(A) tRF starting site’s distribution in each position of their host tRNA (The vertical axis labels’ positions were transferred according to their square root number); (B) First 8 base logo of the tRNAs that produce tRFs from the 1<sup>st</sup> base in bcp; (C) First 8 base logo of tRNAs that produce tRFs from the 2<sup>nd</sup> base in bcp; (D) First 8 base logo of the tRNAs that produce tRFs from the 1<sup>st</sup> base in tcp; (E) First 8 base logo of tRNAs that produce tRFs from the 2<sup>nd</sup> base in tcp. unm, bcp, tcp and RPM see <xref ref-type="fig" rid="F1-1674-3466-53-5-625">Figure 1</xref>. Figure 3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-625/img_3.png<b>图3</b> 水稻雄配子三类花粉的tRF 5’端碱基分布偏好性<br/>(A) 起始于tRNA各个位置的tRF起点碱基分布情况(纵坐标各点相对位置进行了1/2次幂变形处理); (B) bcp样品中tRF起始于tRNA第1位时, 对应宿主tRNA前8个碱基logo图; (C) bcp样品中tRF起始于tRNA第2位时, 对应宿主tRNA前8个碱基logo图; (D) tcp样品中tRF起始于tRNA第1位时, 对应宿主tRNA前8个碱基logo图; (E) tcp样品中tRF起始于tRNA第2位时, 对应宿主tRNA前8个碱基logo图。unm、bcp、tcp和RPM同<xref ref-type="fig" rid="F1-1674-3466-53-5-625">图1</xref>。<br/><b>Figure 3</b> tRFs’ 5’ end base bias in three types of pollens in rice male gamete<br/>(A) tRF starting site’s distribution in each position of their host tRNA (The vertical axis labels’ positions were transferred according to their square root number); (B) First 8 base logo of the tRNAs that produce tRFs from the 1<sup>st</sup> base in bcp; (C) First 8 base logo of tRNAs that produce tRFs from the 2<sup>nd</sup> base in bcp; (D) First 8 base logo of the tRNAs that produce tRFs from the 1<sup>st</sup> base in tcp; (E) First 8 base logo of tRNAs that produce tRFs from the 2<sup>nd</sup> base in tcp. unm, bcp, tcp and RPM see <xref ref-type="fig" rid="F1-1674-3466-53-5-625">Figure 1</xref>.
图3 水稻雄配子三类花粉的tRF 5’端碱基分布偏好性 (A) 起始于tRNA各个位置的tRF起点碱基分布情况(纵坐标各点相对位置进行了1/2次幂变形处理); (B) bcp样品中tRF起始于tRNA第1位时, 对应宿主tRNA前8个碱基logo图; (C) bcp样品中tRF起始于tRNA第2位时, 对应宿主tRNA前8个碱基logo图; (D) tcp样品中tRF起始于tRNA第1位时, 对应宿主tRNA前8个碱基logo图; (E) tcp样品中tRF起始于tRNA第2位时, 对应宿主tRNA前8个碱基logo图。unm、bcp、tcp和RPM同图1。 Figure 3 tRFs’ 5’ end base bias in three types of pollens in rice male gamete (A) tRF starting site’s distribution in each position of their host tRNA (The vertical axis labels’ positions were transferred according to their square root number); (B) First 8 base logo of the tRNAs that produce tRFs from the 1st base in bcp; (C) First 8 base logo of tRNAs that produce tRFs from the 2nd base in bcp; (D) First 8 base logo of the tRNAs that produce tRFs from the 1st base in tcp; (E) First 8 base logo of tRNAs that produce tRFs from the 2nd base in tcp. unm, bcp, tcp and RPM see Figure 1.
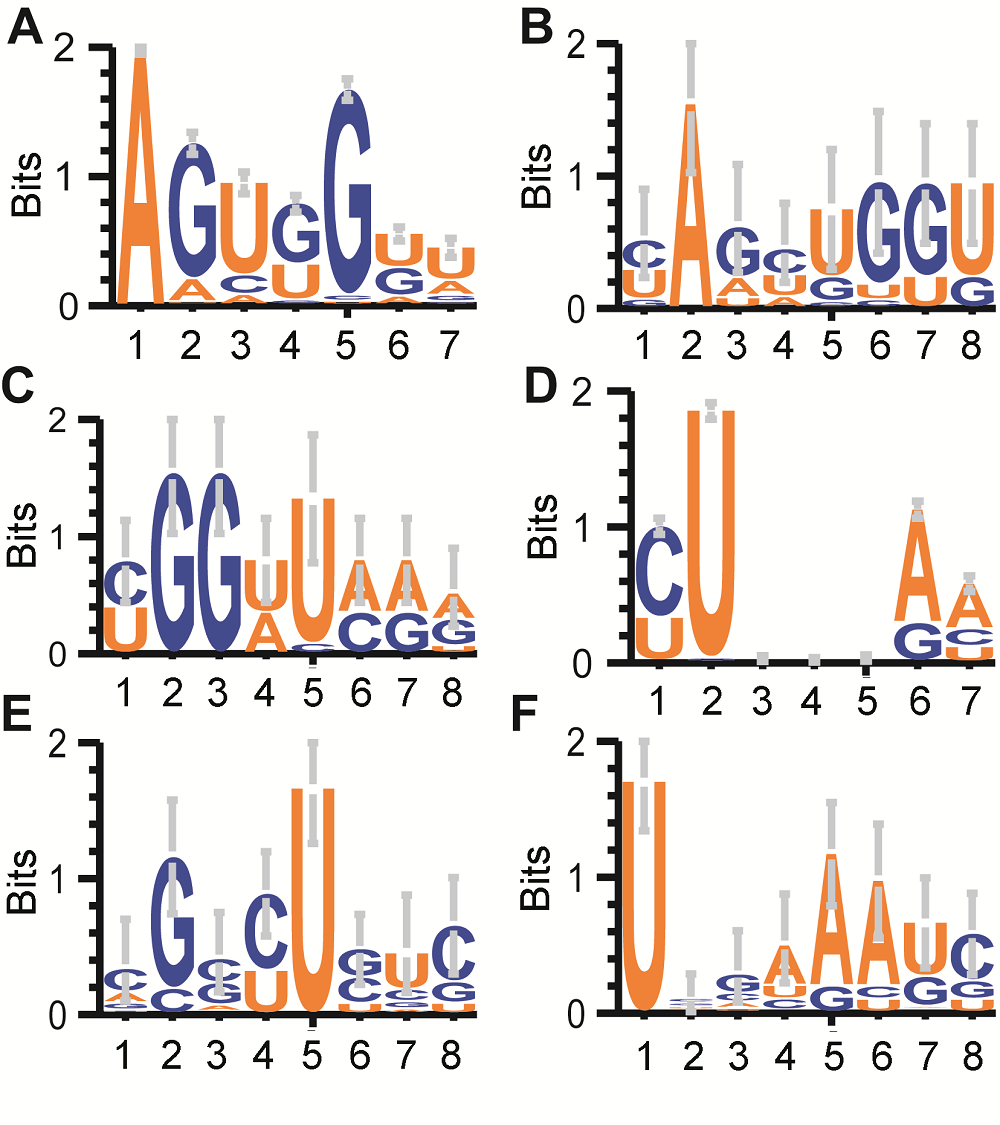
将tRF映射回tRNA。我们希望从tRF的终点(3'端)和起点(5'端)位置了解tRNA剪切位置的特征。剪切位点两侧的片段丰度多数不一致。在D环, 第3个碱基终止的tRF与在第4个碱基起始的tRF含量相距甚远; 在反密码子环, 第2个碱基终止的tRF含量也远高于第3个碱基起始的tRF (附图1)。tRNA的剪切最终得到的多是5'侧的片段(图1)。 三类花粉对应tRNA的剪切位置如附图2所示。unm几乎不在反密码子环剪切, 在D环剪切的情况也很少, 仅在距D环5'端2 nt处与后两阶段趋势一致。bcp和tcp都趋于在距D环5'端2、5 nt处, 以及距反密码子环5'端0、1、3和4 nt处剪切。有趣的是, tRNA于反密码子环的转录后修饰多发生在距5'端0、2、4 nt处(Dunin-Horkawicz et al., 2006), 剪切与修饰有可能相关。但在某些地方, bcp和tcp剪切(比如距D环3 nt的地方, bcp剪切减少, tcp剪切增加)又有不同(附图1)。对相对高频次的剪切, 都用weblogo进行了序列分析(附图2)。 在D环, bcp和tcp剪切位置富集于距GG序列1 nt的地方, 且位于U的5'侧(图4A-C; 附图2), 模型大概为“N|UGGN|UNN”(其中|表示剪切位点, N表示任意碱基)。GG序列附近的U在正常情况下会被修饰为D (二氢尿嘧啶核苷), 以维持局部茎环结构稳定, 甚至影响tRNA全局构象(Nawrot et al., 1995)。D并不会与A配对, 因此测序不太可能大量测出D。加上Dus家族的酶(二氢尿嘧啶核苷合成酶), 在bcp和tcp中的表达量也普遍低于unm (附图3)。因此此处测序得到的大量以U起始的片段很可能未经修饰, 推测是某种识别U并在其5'侧剪切的内切酶的产物。 图4https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-625/img_4.png<b>图4</b> 水稻雄配子三类花粉的tRNA剪切位置周围碱基分布<br/>(A), (D) 所有tRNA的D环和反密码子环上前7个碱基分布logo图; (B) bcp于D环第3个碱基终止的tRF对应tRNA序列logo图; (C) tcp于D环第7个碱基起始的tRF对应tRNA序列logo图; (E) bcp于反密码子环第1个碱基终止的tRF对应tRNA序列logo图; (F) tcp于反密码子环第6个碱基起始的tRF对应tRNA序列logo图。除了图A和D, 各图横轴4和5之间为“剪切发生位置”。<br/><b>Figure 4</b> Sequence motif logo for tRNA 8 mer around cleavage sites in three types of pollens in rice male gamete<br/>(A), (D) Sequence logo of all tRNA’s first 7 bases of D loop and Anticodon loop; (B) For bcp tRFs ended at the 3<sup>rd</sup> base of D loop; (C) For tcp tRFs started at the 7<sup>th</sup> base of D loop; (E) For bcp tRFs ended at the 1<sup>st</sup> base of Anticodon loop; (F) For tcp tRFs started at the 6<sup>th</sup> base of Anticodon loop. The position between 4 and 5 in X axis of Figure B, C, E and F stand for cleavage site. Figure 4https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-625/img_4.png<b>图4</b> 水稻雄配子三类花粉的tRNA剪切位置周围碱基分布<br/>(A), (D) 所有tRNA的D环和反密码子环上前7个碱基分布logo图; (B) bcp于D环第3个碱基终止的tRF对应tRNA序列logo图; (C) tcp于D环第7个碱基起始的tRF对应tRNA序列logo图; (E) bcp于反密码子环第1个碱基终止的tRF对应tRNA序列logo图; (F) tcp于反密码子环第6个碱基起始的tRF对应tRNA序列logo图。除了图A和D, 各图横轴4和5之间为“剪切发生位置”。<br/><b>Figure 4</b> Sequence motif logo for tRNA 8 mer around cleavage sites in three types of pollens in rice male gamete<br/>(A), (D) Sequence logo of all tRNA’s first 7 bases of D loop and Anticodon loop; (B) For bcp tRFs ended at the 3<sup>rd</sup> base of D loop; (C) For tcp tRFs started at the 7<sup>th</sup> base of D loop; (E) For bcp tRFs ended at the 1<sup>st</sup> base of Anticodon loop; (F) For tcp tRFs started at the 6<sup>th</sup> base of Anticodon loop. The position between 4 and 5 in X axis of Figure B, C, E and F stand for cleavage site.
图4 水稻雄配子三类花粉的tRNA剪切位置周围碱基分布 (A), (D) 所有tRNA的D环和反密码子环上前7个碱基分布logo图; (B) bcp于D环第3个碱基终止的tRF对应tRNA序列logo图; (C) tcp于D环第7个碱基起始的tRF对应tRNA序列logo图; (E) bcp于反密码子环第1个碱基终止的tRF对应tRNA序列logo图; (F) tcp于反密码子环第6个碱基起始的tRF对应tRNA序列logo图。除了图A和D, 各图横轴4和5之间为“剪切发生位置”。 Figure 4 Sequence motif logo for tRNA 8 mer around cleavage sites in three types of pollens in rice male gamete (A), (D) Sequence logo of all tRNA’s first 7 bases of D loop and Anticodon loop; (B) For bcp tRFs ended at the 3rd base of D loop; (C) For tcp tRFs started at the 7th base of D loop; (E) For bcp tRFs ended at the 1st base of Anticodon loop; (F) For tcp tRFs started at the 6th base of Anticodon loop. The position between 4 and 5 in X axis of Figure B, C, E and F stand for cleavage site.
Tcp中GG序列内部的剪切较常见(附图1, 附图2), 推测是另一种酶发挥了作用。 在反密码子环, bcp与tcp在第2碱基前后发生剪切较多, 其序列UU或CU与RNase A的作用位点(U或C的3'侧)一致(图4D, E)。有研究表明, 产生5'-tRNA halves (由反密码子环剪开产生, 长度大约为tRNA一半的tRF)的剪切留下5'羟基(Kumar et al., 2016), 且哺乳动物的angiogenin (Emara et al., 2010) (可剪切tRNA产生tRNA halves)正是属于RNase A的一种。 位于反密码子第4碱基3'侧的剪切则并未表现出碱基偏好性, 推测可能是RNase T2 (酵母中为Rny1 (Thompson and Parker, 2009))发挥作用。然而水稻含有8个RNase T2基因, 其中4种在花粉中通过基因芯片检测到, 但均无明显上调(附表1)。其剪切可能受到34位(反密码子环第3个)碱基修饰基团的调控。反密码子环第6碱基(离loop 5'边界5 nt)处往往分布着被修饰的A或者G, A的N6修饰基团阻止该碱基与第33位U通过氢键配对(Vare et al., 2017)。由此可见, 测序得到的该位置的A与正常tRNA的该位置不同; 剪切位置3'侧连续呈现2个A (图4D, F)。推测AA序列可能是酶切识别的特征所在。 表1 Table 1 表1 表1 被靶向的非TE基因和TE数的对比 Table 1 tRFs targeted loci of non-TE and TE genes
Sample
No. of non-TE genes
No. of genic TEs
No. of intergenic TEs
TE Percentage (%)
unm
317
166
506
67.95
bcp
728
403
905
64.24
tcp
529
287
736
65.91
Genic TEs: Annotated TE genes in MSU v7; Intergenic TEs: The TE loci not included in MSU v7 annotation, but represented in TIGR Oryza Repeats Database v3.3. unm, bcp and tcp see Figure 1. Genic TEs: MSU版本7中注释的TE基因; Intergenic TEs: 除genic TEs以外的在TIGR Oryza Repeats Database v3.3中出现的TE。unm、bcp和tcp同图1。
表1 被靶向的非TE基因和TE数的对比 Table 1 tRFs targeted loci of non-TE and TE genes
2.4 讨论本研究通过高通量测序及生物信息学分析, 揭示了水稻雄配子体发育过程中tRF的动态分布, 总结了水稻雄配子体tRNA剪切位点的规律, 预测了tRF靶向的TE和非TE基因。由于选取的测序片段长度范围较广, 我们发现了还几乎未见报道的长度为~16 nt的tRF, 并且其含量远超目前备受关注的19 nt tRF (Cole et al., 2009; Sobala and Hutvagner, 2013; Martinez et al., 2017)。除上述2个类群tRF, 还有~32 nt和~35 nt的tRF, 水稻雄配子体中主要有4个类群的tRF。 虽然文中只讨论了D环和反密码子环的剪切现象, 但tRF在其它位置也有一定量的剪切, 如D臂和反密码子臂交界处的G26等。剪切位置附近一般都有转录后修饰。以小鼠(Tuorto et al., 2012)和果蝇(Schaefer et al., 2010)为对象的研究表明, Dnmt2与Nsun2对tRNA的甲基化可以保护其免遭剪切成为tRNA halves, 也确实发现bcp和tcp的对应酶类有降低; 此外, 其它甲基化酶类表达量也降低了(附表1)。因此推测碱基修饰的缺失可能是tRNA剪切发生的先决条件, 然后才是序列识别。本研究得到了3个未报道过的剪切位点特征序列, 它们在水稻雄配子体3个阶段花粉中的剪切强度各不相同。 水稻雄配子体经历2次有丝分裂, 第1次是从1个细胞不均等地分裂成营养细胞和生殖细胞; 第2次则是在一层细胞膜的保护下, 生殖细胞均分成为2个精细胞。相应地, unm几乎只产生tRF-5, bcp开始就有大量tiRNA (tRNA halves)产生, 且各阶段的同类tRF都存在着剪切位点和反密码子的差别。推测此过程tRF的类群变化服务于不同的细胞环境要求。Sharma等(2016)发现, 小鼠的附睾产生大量tRF并运送给精子; 对蛋白摄入不足的小鼠来说, 其精子里还会有很多tRF-5-GlyGCC。当使用包含这部分tRF的<40 nt RNA注入到二细胞胚胎, 发现反转录转座因子MERVL靶向的基因被大量下调; 他们将人工合成的tRF-5- GlyGCC注射到二细胞胚胎, 同样发现MERVL靶向的基因表达下调, 但相应的tRF-3-GlyGCC以及tRF-2- GlyGCC则不会产生同样的效果。tRF既然广泛存在于真核生物中, 其作用机制也极可能具有保守性。tcp中的一些tRF有可能来自它的营养细胞, 并在下一代合子中发挥功能。 我们未发表的数据表明, tRF靶基因和tRF表达量的关系并没有呈现显著的负相关。Alves团队通过分析降解组数据, 也未发现被切割的tRF靶片段(Alves et al., 2016)。因此推测tRF发挥功能的方式并不依赖降解靶RNA。对水稻雄配子体各阶段的研究, 目前只停留在转录组和sRNA组层面(包括三阶段的转录组和sRNA组, 以及tcp阶段营养核和精细胞的转录组) (Wei et al., 2010, 2011; Peng et al., 2012; Anderson et al., 2013)。我们的研究也没有精确到细胞层面。因此, tRF在雄配子体发育中的具体功能还有待后续深入研究。本研究为植物tRF的来源研究以及水稻雄配子体发育相关研究提供了新线索。
附图1 水稻雄配子三类花粉的tRF在D环和反密码子环处终止或起始的位点Appendix figure 1 Distance between the tRFs’ starting or ending base and D loop’s or Anticodon loop’s first base in 3 types of pollens in rice male gamete Appendix figure 2 Sequence motif logo for tRNA 8 mer around cleavage sites Appendixtable 1 Some reference genes transcriptional level which was transferred to log2 scale, according to Peng et al., 2012. Appendix figure 3 Distribution of matched regions’ length over tRFs Appendix figure 4 Distribution of target matched tRF base positions over all tRF parts Appendix figure 5 tRF_target_GO analysis Appendix figure 6 tRFs targeted TE types in 3 types of pollens in rice male gamete
The authors have declared that no competing interests exist.
AlvesCS, VicentiniR, DuarteGT, PinotiVF, VincentzM, NogueiraFTS (2016). Genome-wide identification and characterization of tRNA-derived RNA fragments in land plants.Plant Mol Biol 93, 35-48. [本文引用: 2]
[3]
AndersonSN, JohnsonCS, JonesDS, ConradLJ, GouXP, RussellSD, SundaresanV (2013). Transcriptomes of isolated Oryza sativa gametes characterized by deep sequencing: evidence for distinct sex-dependent chromatin and epigenetic states before fertilization. Plant J 76, 729-741. [本文引用: 1]
[4]
ChenCJ, LiuQ, ZhangYC, QuLH, ChenYQ, GautheretD (2011). Genome-wide discovery and analysis of microRNAs and other small RNAs from rice embryogenic callus.RNA Biol 8, 538-547. [本文引用: 1]
[5]
ColeC, SobalaA, LuC, ThatcherSR, BowmanA, BrownJWS, GreenPJ, BartonGJ, HutvagnerG (2009). Filtering of deep sequencing data reveals the existence of abundant Dicer-dependent small RNAs derived from tRNAs.RNA 15, 2147-2160. [本文引用: 1]
[6]
CouvillionMT, SachidanandamR, CollinsK (2010). A growth-essential Tetrahymena Piwi protein carries tRNA fragment cargo. Genes Dev 24, 2742-2747. [本文引用: 1]
[7]
Dunin-HorkawiczS, CzerwoniecA, GajdaMJ, FederM, GrosjeanH, BujnickiJM (2006). MODOMICS: a database of RNA modification pathways.Nucleic Acids Res 34, D145-D149. [本文引用: 1]
GebetsbergerJ, WyssL, MleczkoAM, ReutherJ, PolacekN (2017). A tRNA-derived fragment competes with mRNA for ribosome binding and regulates translation during stress.RNA Biol 14, 1364-1373. [本文引用: 1]
[10]
GoodarziH, LiuXH, NguyenHCB, ZhangS, FishL, TavazoieSF (2015). Endogenous tRNA-derived fragments suppress breast cancer progression via YBX1 displacement.Cell 161, 790-802. [本文引用: 1]
[11]
Grant-DowntonR, Le TrionnaireG, SchmidR, Rodriguez-EnriquezJ, HafidhS, MehdiS, TwellD, DickinsonH (2009). MicroRNA and tasiRNA diversity in mature pollen of Arabidopsis thaliana. BMC Genomics 10, 643. [本文引用: 1]
[12]
HsiehLC, LinSI, KuoHF, ChiouTJ (2010). Abundance of tRNA-derived small RNAs in phosphate-starved Arabidopsis roots.Plant Signal Behav 5, 537-539. [本文引用: 1]
[13]
KaraiskosS, NaqviAS, SwansonKE, GrigorievA (2015). Age-driven modulation of tRNA-derived fragments in Drosophila and their potential targets. Biol Direct 10, 51. [本文引用: 1]
[14]
KeamSP, SobalaA, Ten HaveS, HutvagnerG (2017). tRNA-Derived RNA fragments associate with human multisynthetase complex (MSC) and modulate ribosomal protein translation.J Proteome Res 16, 413-420. [本文引用: 1]
[15]
KumarP, AnayaJ, MudunuriSB, DuttaA (2014). Meta- analysis of tRNA derived RNA fragments reveals that they are evolutionarily conserved and associate with AGO proteins to recognize specific RNA targets.BMC Biol 12, 78. [本文引用: 1]
[16]
KumarP, KuscuC, DuttaA (2016). Biogenesis and function of transfer RNA-related fragments (tRFs).Trends Biochem Sci 41, 679-689. [本文引用: 3]
[17]
Loss-MoraisG, WaterhousePM, MargisR (2013). Description of plant tRNA-derived RNA fragments (tRFs) associated with argonaute and identification of their putative targets.Biol Direct 8, 6. [本文引用: 1]
[18]
LoweTM, EddySR (1997). tRNAscan-SE: a program for improved detection of transfer RNA genes in genomic sequence.Nucleic Acids Res 25, 955-964.
[19]
MartinezG, ChoudurySG, SlotkinRK (2017). tRNA- derived small RNAs target transposable element transcripts.Nucleic Acids Res 45, 5142-5152. [本文引用: 4]
[20]
NawrotB, MalkiewiczA, SmithWS, Sierzputowska-GraczH, AgrisPF (1995). RNA modified uridines VII: chemical synthesis and initial analysis of tRNA D-Loop oligomers with tandem modified uridines.Nucleos Nucleot Nud 14, 143-165. [本文引用: 1]
[21]
PengH, ChunJ, AiTB, TongYA, ZhangR, ZhaoMM, ChenF, WangSH (2012). MicroRNA profiles and their control of male gametophyte development in rice.Plant Mol Biol 80, 85-102. [本文引用: 4]
[22]
SchaeferM, PollexT, HannaK, TuortoF, MeusburgerM, HelmM, LykoF (2010). RNA methylation by Dnmt2 protects transfer RNAs against stress-induced cleavage.Genes Dev 24, 1590-1595. [本文引用: 1]
[23]
SharmaU, ConineCC, SheaJM, BoskovicA, DerrAG, BingXY, BelleanneeC, KucukuralA, SerraRW, SunFY, SongLN, CaroneBR, RicciEP, LiXZ, FauquierL, MooreMJ, SullivanR, MelloCC, GarberM, RandoOJ (2016). Biogenesis and function of tRNA fragments during sperm maturation and fertilization in mammals.Science 351, 391-396. [本文引用: 2]
[24]
SlotkinRK, VaughnM, BorgesF, TanurdžićM, BeckerJD, FeijoJA, MartienssenRA (2009). Epigenetic reprogramming and small RNA silencing of transposable elements in pollen.Cell 136, 461-472. [本文引用: 1]
[25]
SobalaA, HutvagnerG (2013). Small RNAs derived from the 5' end of tRNA can inhibit protein translation in human cells.RNA Biol 10, 553-563. [本文引用: 1]
ThompsonDM, ParkerR (2009). The RNase Rny1p cleaves tRNAs and promotes cell death during oxidative stress in Saccharomyces cerevisiae.J Cell Biol 185, 43-50. [本文引用: 2]
[28]
TuortoF, LiebersR, MuschT, SchaeferM, HofmannS, KellnerS, FryeM, HelmM, StoecklinG, LykoF (2012). RNA cytosine methylation by Dnmt2 and NSun2 promotes tRNA stability and protein synthesis.Nat Struct Mol Biol 19, 900-905. [本文引用: 1]
[29]
VareVYP, EruysalER, NarendranA, SarachanKL, AgrisPF (2017). Chemical and conformational diversity of modified nucleosides affects tRNA structure and function.Bio- molecules 7, 29. [本文引用: 1]
[30]
WangF, JohnsonNR, CoruhC, AxtellMJ (2016a). Genome-wide analysis of single non-templated nucleotides in plant endogenous siRNAs and miRNAs.Nucleic Acids Res 44, 7395-7405.
[31]
WangQH, LiTT, XuK, ZhangW, WangXL, QuanJL, JinWB, ZhangMX, FanGJ, WangMB, ShanWX (2016b). The tRNA-derived small RNAs regulate gene expression through triggering sequence-specific degradation of target transcripts in the oomycete pathogen Phytophthora sojae. Front Plant Sci 7, 1938. [本文引用: 2]
[32]
WeiLQ, XuWY, DengZY, SuZ, XueYB, WangT (2010). Genome-scale analysis and comparison of gene expression profiles in developing and germinated pollen in Oryza sativa. BMC Genomics 11, 338. [本文引用: 1]
[33]
WeiLQ, YanLF, WangT (2011). Deep sequencing on genome-wide scale reveals the unique composition and expression patterns of microRNAs in developing pollen of Oryza sativa. Genome Biol 12, R53. [本文引用: 1]
1 2009
... 水稻的tRNA基因来自tRNADB-CE (Abe et al., 2009). ...
2 2016
... 三类花粉在14-20 nt区间分布规律相似.在植物研究中, 长度在~16 nt的tRF几乎未见报道, 而在本研究中的三类花粉中均较多.~19 nt长度的tRF在多项研究中已有报道, 如水稻营养组织、小立碗藓(Phy- scomitrella patens) (Alves et al., 2016)和缺磷条件下的拟南芥根中(Hsieh et al., 2010). ... ... 我们未发表的数据表明, tRF靶基因和tRF表达量的关系并没有呈现显著的负相关.Alves团队通过分析降解组数据, 也未发现被切割的tRF靶片段(Alves et al., 2016).因此推测tRF发挥功能的方式并不依赖降解靶RNA.对水稻雄配子体各阶段的研究, 目前只停留在转录组和sRNA组层面(包括三阶段的转录组和sRNA组, 以及tcp阶段营养核和精细胞的转录组) (Wei et al., 2010, 2011; Peng et al., 2012; Anderson et al., 2013).我们的研究也没有精确到细胞层面.因此, tRF在雄配子体发育中的具体功能还有待后续深入研究.本研究为植物tRF的来源研究以及水稻雄配子体发育相关研究提供了新线索. ...
1 2013
... 我们未发表的数据表明, tRF靶基因和tRF表达量的关系并没有呈现显著的负相关.Alves团队通过分析降解组数据, 也未发现被切割的tRF靶片段(Alves et al., 2016).因此推测tRF发挥功能的方式并不依赖降解靶RNA.对水稻雄配子体各阶段的研究, 目前只停留在转录组和sRNA组层面(包括三阶段的转录组和sRNA组, 以及tcp阶段营养核和精细胞的转录组) (Wei et al., 2010, 2011; Peng et al., 2012; Anderson et al., 2013).我们的研究也没有精确到细胞层面.因此, tRF在雄配子体发育中的具体功能还有待后续深入研究.本研究为植物tRF的来源研究以及水稻雄配子体发育相关研究提供了新线索. ...
1 2011
... tRNA为人们所熟知的功能是转运氨基酸以合成多肽, 但近年人们发现tRNA的功能似乎不止如此.tRNA产生的片段(tRNA-derived fragment, tRF)在酿酒酵母(Saccharomyces cerevisiae) (Thompson and Parker, 2009)、四膜虫(Tetrahymena sp.) (Couvillion et al., 2010)、果蝇(Drosophila melanogaster) (Karaiskos et al., 2015)、水稻(Oryza sativa) (Chen et al., 2011)、小鼠(Mus musculus) (Sharma et al., 2016)和人类(Homo sapiens) (Grant-Downton et al., 2009)等研究中已有报道.tRNA可能在反密码子环进行剪切, 产生5'和3'的tRNA halves (因最初在胁迫条件下被发现, 又称tiRNA, 后文简称5tiR和3tiR), 也可在其它位置剪切, 形成反密码子环5'侧的tRF-5和3'侧的tRF-3, 以及跨越反密码子环的tRF-2 (Kumar et al., 2016).在tRF形成过程中, 参与反密码子环剪切的酶有angiogenin (人类)和Rny1 (酵母); 参与其它位置剪切的酶尚不明确, 其中DICER是否为tRF-3和tRF-5形成所必需仍存在争议(Kumar et al., 2016).tRF有许多功能, 例如: 26 nt的tRF-5-Val被认为与mRNA相竞争地结合核糖体, 以抑制转录起始(Gebetsberger et al., 2017); 一些tRF-2被证明与某些mRNA竞争性结合蛋白YBX1, 从而抑制乳腺癌(Goodarzi et al., 2015); tRF-5-Gln可以结合MSC (Human Multisynthetase Complex)并上调核糖体蛋白的表达(Keam et al., 2017).tRF还可能结合AGO蛋白, 具有与miRNA类似的种子序列, 并能形成AGO-tRF-mRNA复合体(Kumar et al., 2014), 由此来调控其靶基因的表达(Wang et al., 2016b; Martinez et al., 2017). ...
1 2009
... 本研究通过高通量测序及生物信息学分析, 揭示了水稻雄配子体发育过程中tRF的动态分布, 总结了水稻雄配子体tRNA剪切位点的规律, 预测了tRF靶向的TE和非TE基因.由于选取的测序片段长度范围较广, 我们发现了还几乎未见报道的长度为~16 nt的tRF, 并且其含量远超目前备受关注的19 nt tRF (Cole et al., 2009; Sobala and Hutvagner, 2013; Martinez et al., 2017).除上述2个类群tRF, 还有~32 nt和~35 nt的tRF, 水稻雄配子体中主要有4个类群的tRF. ...
1 2010
... tRNA为人们所熟知的功能是转运氨基酸以合成多肽, 但近年人们发现tRNA的功能似乎不止如此.tRNA产生的片段(tRNA-derived fragment, tRF)在酿酒酵母(Saccharomyces cerevisiae) (Thompson and Parker, 2009)、四膜虫(Tetrahymena sp.) (Couvillion et al., 2010)、果蝇(Drosophila melanogaster) (Karaiskos et al., 2015)、水稻(Oryza sativa) (Chen et al., 2011)、小鼠(Mus musculus) (Sharma et al., 2016)和人类(Homo sapiens) (Grant-Downton et al., 2009)等研究中已有报道.tRNA可能在反密码子环进行剪切, 产生5'和3'的tRNA halves (因最初在胁迫条件下被发现, 又称tiRNA, 后文简称5tiR和3tiR), 也可在其它位置剪切, 形成反密码子环5'侧的tRF-5和3'侧的tRF-3, 以及跨越反密码子环的tRF-2 (Kumar et al., 2016).在tRF形成过程中, 参与反密码子环剪切的酶有angiogenin (人类)和Rny1 (酵母); 参与其它位置剪切的酶尚不明确, 其中DICER是否为tRF-3和tRF-5形成所必需仍存在争议(Kumar et al., 2016).tRF有许多功能, 例如: 26 nt的tRF-5-Val被认为与mRNA相竞争地结合核糖体, 以抑制转录起始(Gebetsberger et al., 2017); 一些tRF-2被证明与某些mRNA竞争性结合蛋白YBX1, 从而抑制乳腺癌(Goodarzi et al., 2015); tRF-5-Gln可以结合MSC (Human Multisynthetase Complex)并上调核糖体蛋白的表达(Keam et al., 2017).tRF还可能结合AGO蛋白, 具有与miRNA类似的种子序列, 并能形成AGO-tRF-mRNA复合体(Kumar et al., 2014), 由此来调控其靶基因的表达(Wang et al., 2016b; Martinez et al., 2017). ...
... 水稻是人类最重要的粮食作物之一, 其雄配子体的正常发育直接关系到水稻的结实率和产量.水稻雄配子体发育开始于早期花药药室中, 由1个二倍体小孢子母细胞经减数分裂形成4个单倍体小孢子, 随后彼此分离形成自由的单核小孢子(uni-nucleate micro- spore, unm); 小孢子体积变大并发生一次不均等有丝分裂, 形成二细胞花粉(bi-cellular pollen, bcp); 二细胞花粉中较大的营养细胞不再分裂, 但是较小的生殖细胞再经1次均等的有丝分裂形成2个精细胞, 此时花粉被称为三细胞花粉(tri-cellular pollen, tcp) (Peng et al., 2012).花粉发育过程中转座因子(TE)大量爆发, 会对基因组的完整性和稳定性产生影响; 而管控甲基化的基因DDM1 (DECREASE IN DNA METHYLATION 1)突变后, 同样导致TE大量产生(Slotkin et al., 2009).最近一项研究表明, 拟南芥(Arabidopsis thaliana)花粉和ddm1突变体大量产生tRF, 通过类似miRNA的作用方式抑制TE (Martinez et al., 2017). ... ... 我们未发表的数据表明, tRF靶基因和tRF表达量的关系并没有呈现显著的负相关.Alves团队通过分析降解组数据, 也未发现被切割的tRF靶片段(Alves et al., 2016).因此推测tRF发挥功能的方式并不依赖降解靶RNA.对水稻雄配子体各阶段的研究, 目前只停留在转录组和sRNA组层面(包括三阶段的转录组和sRNA组, 以及tcp阶段营养核和精细胞的转录组) (Wei et al., 2010, 2011; Peng et al., 2012; Anderson et al., 2013).我们的研究也没有精确到细胞层面.因此, tRF在雄配子体发育中的具体功能还有待后续深入研究.本研究为植物tRF的来源研究以及水稻雄配子体发育相关研究提供了新线索. ... ... Appendix figure 2 Sequence motif logo for tRNA 8 mer around cleavage sites <b>附表1</b> 一些基因的转录水平, 表中的值是经过log2转化后的表达量, 数据来自<xref ref-type="bibr" rid="b21-1674-3466-53-5-625">Peng等(2012)</xref>Appendixtable 1 Some reference genes transcriptional level which was transferred to log2 scale, according to Peng et al., 2012. ... ... Appendixtable 1 Some reference genes transcriptional level which was transferred to log2 scale, according to Peng et al., 2012. ...
1 2010
... 虽然文中只讨论了D环和反密码子环的剪切现象, 但tRF在其它位置也有一定量的剪切, 如D臂和反密码子臂交界处的G26等.剪切位置附近一般都有转录后修饰.以小鼠(Tuorto et al., 2012)和果蝇(Schaefer et al., 2010)为对象的研究表明, Dnmt2与Nsun2对tRNA的甲基化可以保护其免遭剪切成为tRNA halves, 也确实发现bcp和tcp的对应酶类有降低; 此外, 其它甲基化酶类表达量也降低了(附表1).因此推测碱基修饰的缺失可能是tRNA剪切发生的先决条件, 然后才是序列识别.本研究得到了3个未报道过的剪切位点特征序列, 它们在水稻雄配子体3个阶段花粉中的剪切强度各不相同. ...
2 2016
... tRNA为人们所熟知的功能是转运氨基酸以合成多肽, 但近年人们发现tRNA的功能似乎不止如此.tRNA产生的片段(tRNA-derived fragment, tRF)在酿酒酵母(Saccharomyces cerevisiae) (Thompson and Parker, 2009)、四膜虫(Tetrahymena sp.) (Couvillion et al., 2010)、果蝇(Drosophila melanogaster) (Karaiskos et al., 2015)、水稻(Oryza sativa) (Chen et al., 2011)、小鼠(Mus musculus) (Sharma et al., 2016)和人类(Homo sapiens) (Grant-Downton et al., 2009)等研究中已有报道.tRNA可能在反密码子环进行剪切, 产生5'和3'的tRNA halves (因最初在胁迫条件下被发现, 又称tiRNA, 后文简称5tiR和3tiR), 也可在其它位置剪切, 形成反密码子环5'侧的tRF-5和3'侧的tRF-3, 以及跨越反密码子环的tRF-2 (Kumar et al., 2016).在tRF形成过程中, 参与反密码子环剪切的酶有angiogenin (人类)和Rny1 (酵母); 参与其它位置剪切的酶尚不明确, 其中DICER是否为tRF-3和tRF-5形成所必需仍存在争议(Kumar et al., 2016).tRF有许多功能, 例如: 26 nt的tRF-5-Val被认为与mRNA相竞争地结合核糖体, 以抑制转录起始(Gebetsberger et al., 2017); 一些tRF-2被证明与某些mRNA竞争性结合蛋白YBX1, 从而抑制乳腺癌(Goodarzi et al., 2015); tRF-5-Gln可以结合MSC (Human Multisynthetase Complex)并上调核糖体蛋白的表达(Keam et al., 2017).tRF还可能结合AGO蛋白, 具有与miRNA类似的种子序列, 并能形成AGO-tRF-mRNA复合体(Kumar et al., 2014), 由此来调控其靶基因的表达(Wang et al., 2016b; Martinez et al., 2017). ... ... 水稻雄配子体经历2次有丝分裂, 第1次是从1个细胞不均等地分裂成营养细胞和生殖细胞; 第2次则是在一层细胞膜的保护下, 生殖细胞均分成为2个精细胞.相应地, unm几乎只产生tRF-5, bcp开始就有大量tiRNA (tRNA halves)产生, 且各阶段的同类tRF都存在着剪切位点和反密码子的差别.推测此过程tRF的类群变化服务于不同的细胞环境要求.Sharma等(2016)发现, 小鼠的附睾产生大量tRF并运送给精子; 对蛋白摄入不足的小鼠来说, 其精子里还会有很多tRF-5-GlyGCC.当使用包含这部分tRF的<40 nt RNA注入到二细胞胚胎, 发现反转录转座因子MERVL靶向的基因被大量下调; 他们将人工合成的tRF-5- GlyGCC注射到二细胞胚胎, 同样发现MERVL靶向的基因表达下调, 但相应的tRF-3-GlyGCC以及tRF-2- GlyGCC则不会产生同样的效果.tRF既然广泛存在于真核生物中, 其作用机制也极可能具有保守性.tcp中的一些tRF有可能来自它的营养细胞, 并在下一代合子中发挥功能. ...
1 2009
... 水稻是人类最重要的粮食作物之一, 其雄配子体的正常发育直接关系到水稻的结实率和产量.水稻雄配子体发育开始于早期花药药室中, 由1个二倍体小孢子母细胞经减数分裂形成4个单倍体小孢子, 随后彼此分离形成自由的单核小孢子(uni-nucleate micro- spore, unm); 小孢子体积变大并发生一次不均等有丝分裂, 形成二细胞花粉(bi-cellular pollen, bcp); 二细胞花粉中较大的营养细胞不再分裂, 但是较小的生殖细胞再经1次均等的有丝分裂形成2个精细胞, 此时花粉被称为三细胞花粉(tri-cellular pollen, tcp) (Peng et al., 2012).花粉发育过程中转座因子(TE)大量爆发, 会对基因组的完整性和稳定性产生影响; 而管控甲基化的基因DDM1 (DECREASE IN DNA METHYLATION 1)突变后, 同样导致TE大量产生(Slotkin et al., 2009).最近一项研究表明, 拟南芥(Arabidopsis thaliana)花粉和ddm1突变体大量产生tRF, 通过类似miRNA的作用方式抑制TE (Martinez et al., 2017). ...
1 2013
... 本研究通过高通量测序及生物信息学分析, 揭示了水稻雄配子体发育过程中tRF的动态分布, 总结了水稻雄配子体tRNA剪切位点的规律, 预测了tRF靶向的TE和非TE基因.由于选取的测序片段长度范围较广, 我们发现了还几乎未见报道的长度为~16 nt的tRF, 并且其含量远超目前备受关注的19 nt tRF (Cole et al., 2009; Sobala and Hutvagner, 2013; Martinez et al., 2017).除上述2个类群tRF, 还有~32 nt和~35 nt的tRF, 水稻雄配子体中主要有4个类群的tRF. ...
0 2013
2 2009
... tRNA为人们所熟知的功能是转运氨基酸以合成多肽, 但近年人们发现tRNA的功能似乎不止如此.tRNA产生的片段(tRNA-derived fragment, tRF)在酿酒酵母(Saccharomyces cerevisiae) (Thompson and Parker, 2009)、四膜虫(Tetrahymena sp.) (Couvillion et al., 2010)、果蝇(Drosophila melanogaster) (Karaiskos et al., 2015)、水稻(Oryza sativa) (Chen et al., 2011)、小鼠(Mus musculus) (Sharma et al., 2016)和人类(Homo sapiens) (Grant-Downton et al., 2009)等研究中已有报道.tRNA可能在反密码子环进行剪切, 产生5'和3'的tRNA halves (因最初在胁迫条件下被发现, 又称tiRNA, 后文简称5tiR和3tiR), 也可在其它位置剪切, 形成反密码子环5'侧的tRF-5和3'侧的tRF-3, 以及跨越反密码子环的tRF-2 (Kumar et al., 2016).在tRF形成过程中, 参与反密码子环剪切的酶有angiogenin (人类)和Rny1 (酵母); 参与其它位置剪切的酶尚不明确, 其中DICER是否为tRF-3和tRF-5形成所必需仍存在争议(Kumar et al., 2016).tRF有许多功能, 例如: 26 nt的tRF-5-Val被认为与mRNA相竞争地结合核糖体, 以抑制转录起始(Gebetsberger et al., 2017); 一些tRF-2被证明与某些mRNA竞争性结合蛋白YBX1, 从而抑制乳腺癌(Goodarzi et al., 2015); tRF-5-Gln可以结合MSC (Human Multisynthetase Complex)并上调核糖体蛋白的表达(Keam et al., 2017).tRF还可能结合AGO蛋白, 具有与miRNA类似的种子序列, 并能形成AGO-tRF-mRNA复合体(Kumar et al., 2014), 由此来调控其靶基因的表达(Wang et al., 2016b; Martinez et al., 2017). ... ... 位于反密码子第4碱基3'侧的剪切则并未表现出碱基偏好性, 推测可能是RNase T2 (酵母中为Rny1 (Thompson and Parker, 2009))发挥作用.然而水稻含有8个RNase T2基因, 其中4种在花粉中通过基因芯片检测到, 但均无明显上调(附表1).其剪切可能受到34位(反密码子环第3个)碱基修饰基团的调控.反密码子环第6碱基(离loop 5'边界5 nt)处往往分布着被修饰的A或者G, A的N6修饰基团阻止该碱基与第33位U通过氢键配对(Vare et al., 2017).由此可见, 测序得到的该位置的A与正常tRNA的该位置不同; 剪切位置3'侧连续呈现2个A (图4D, F).推测AA序列可能是酶切识别的特征所在. ...
1 2012
... 虽然文中只讨论了D环和反密码子环的剪切现象, 但tRF在其它位置也有一定量的剪切, 如D臂和反密码子臂交界处的G26等.剪切位置附近一般都有转录后修饰.以小鼠(Tuorto et al., 2012)和果蝇(Schaefer et al., 2010)为对象的研究表明, Dnmt2与Nsun2对tRNA的甲基化可以保护其免遭剪切成为tRNA halves, 也确实发现bcp和tcp的对应酶类有降低; 此外, 其它甲基化酶类表达量也降低了(附表1).因此推测碱基修饰的缺失可能是tRNA剪切发生的先决条件, 然后才是序列识别.本研究得到了3个未报道过的剪切位点特征序列, 它们在水稻雄配子体3个阶段花粉中的剪切强度各不相同. ...
... tRNA为人们所熟知的功能是转运氨基酸以合成多肽, 但近年人们发现tRNA的功能似乎不止如此.tRNA产生的片段(tRNA-derived fragment, tRF)在酿酒酵母(Saccharomyces cerevisiae) (Thompson and Parker, 2009)、四膜虫(Tetrahymena sp.) (Couvillion et al., 2010)、果蝇(Drosophila melanogaster) (Karaiskos et al., 2015)、水稻(Oryza sativa) (Chen et al., 2011)、小鼠(Mus musculus) (Sharma et al., 2016)和人类(Homo sapiens) (Grant-Downton et al., 2009)等研究中已有报道.tRNA可能在反密码子环进行剪切, 产生5'和3'的tRNA halves (因最初在胁迫条件下被发现, 又称tiRNA, 后文简称5tiR和3tiR), 也可在其它位置剪切, 形成反密码子环5'侧的tRF-5和3'侧的tRF-3, 以及跨越反密码子环的tRF-2 (Kumar et al., 2016).在tRF形成过程中, 参与反密码子环剪切的酶有angiogenin (人类)和Rny1 (酵母); 参与其它位置剪切的酶尚不明确, 其中DICER是否为tRF-3和tRF-5形成所必需仍存在争议(Kumar et al., 2016).tRF有许多功能, 例如: 26 nt的tRF-5-Val被认为与mRNA相竞争地结合核糖体, 以抑制转录起始(Gebetsberger et al., 2017); 一些tRF-2被证明与某些mRNA竞争性结合蛋白YBX1, 从而抑制乳腺癌(Goodarzi et al., 2015); tRF-5-Gln可以结合MSC (Human Multisynthetase Complex)并上调核糖体蛋白的表达(Keam et al., 2017).tRF还可能结合AGO蛋白, 具有与miRNA类似的种子序列, 并能形成AGO-tRF-mRNA复合体(Kumar et al., 2014), 由此来调控其靶基因的表达(Wang et al., 2016b; Martinez et al., 2017). ... ... 靶基因预测采用离线的NCBI Blast 2.4.0+, 由于不清楚其配对的具体形式, 我们按照Wang等(2016b)的方法, 仅规定Blast的E值不高于10.参数: blastn -task blastn-short -query [tRF fasta文件] -db [提前构建好的水稻基因组库文件] -out [输出文件] -outfmt "6 qseqid sseqid pident qcovs qlen length mismatch sstrand qstart qend sstart send evalue bitscore" -evalue 10 -ungapped.通过得到的基因组坐标, 结合已有的基因注释文件(gff3), 我们得到对应基因的相关信息. ...
1 2010
... 我们未发表的数据表明, tRF靶基因和tRF表达量的关系并没有呈现显著的负相关.Alves团队通过分析降解组数据, 也未发现被切割的tRF靶片段(Alves et al., 2016).因此推测tRF发挥功能的方式并不依赖降解靶RNA.对水稻雄配子体各阶段的研究, 目前只停留在转录组和sRNA组层面(包括三阶段的转录组和sRNA组, 以及tcp阶段营养核和精细胞的转录组) (Wei et al., 2010, 2011; Peng et al., 2012; Anderson et al., 2013).我们的研究也没有精确到细胞层面.因此, tRF在雄配子体发育中的具体功能还有待后续深入研究.本研究为植物tRF的来源研究以及水稻雄配子体发育相关研究提供了新线索. ...
1 2011
... 我们未发表的数据表明, tRF靶基因和tRF表达量的关系并没有呈现显著的负相关.Alves团队通过分析降解组数据, 也未发现被切割的tRF靶片段(Alves et al., 2016).因此推测tRF发挥功能的方式并不依赖降解靶RNA.对水稻雄配子体各阶段的研究, 目前只停留在转录组和sRNA组层面(包括三阶段的转录组和sRNA组, 以及tcp阶段营养核和精细胞的转录组) (Wei et al., 2010, 2011; Peng et al., 2012; Anderson et al., 2013).我们的研究也没有精确到细胞层面.因此, tRF在雄配子体发育中的具体功能还有待后续深入研究.本研究为植物tRF的来源研究以及水稻雄配子体发育相关研究提供了新线索. ...


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}