Abstract: Plant growth and development are easily affected by environmental changes, and epigenetic mechanisms play important roles in regulating gene expression in response to environmental stimuli. In recent years, epigenetic stu- dies have achieved important progress in the response to abiotic stresses in plants, providing a good foundation for further understanding the potential molecular mechanisms. In this review, we summarize the plant epigenetic regulations, including DNA methylation, histone modification, chromatin remodeling and small RNA, in response to abiotic stresses.
Key words:epigenetic ; abiotic stress ; plant ; growth and development ; research advance
在自然状态下, 植物的整个生长周期均处于固着状态, 不像动物能够主动自发地躲避不良环境, 因此必然会受到来自外界生物与非生物胁迫的影响。正是由于植物处于这样一个极易受到环境胁迫的状态, 其自身进化出一系列自我保护和适应与抵制不良环境的机制(Boyko and Kovalchuk, 2008)。对植物生长发育的胁迫包括内部或外部因素。内部胁迫主要为植物自身基因的突变或异常细胞对机体代谢产生的不利影响; 外部胁迫则可分为生物与非生物胁迫。生物胁迫包括病原感染、食草动物和种内竞争等; 非生物胁迫由不利的环境条件引起(如不适宜的温度、水分和营养物质的利用以及光照条件等), 这两种外部胁迫可以是永久性的也可以是临时状态(Madlung and Comai, 2004)。
图1 植物响应非生物胁迫的表观遗传调控途径 Figure 1 Epigenetic regulation in abiotic stress tolerance in plant
表观遗传最早被定义为有丝分裂以及减数分裂过程中无法用DNA序列的改变来解释基因功能的可继承性改变(Russo et al., 1996)。目前, 人们一致认为表观遗传是研究没有DNA序列变化的、可遗传的基因表达改变(Wu and Morris, 2001)。表观遗传调控机制主要包括DNA甲基化、组蛋白修饰、染色质重塑以及非编码RNA等。越来越多的研究表明, 上述表观遗传机制在植物应答环境胁迫中发挥了非常重要的作用。本文简要阐述近年来表观遗传机制调节植物响应非生物胁迫的一系列研究成果。
1 DNA甲基化1.1 植物体内的DNA甲基化修饰DNA甲基化是指以DNA为受体, 在DNA甲基转移酶的作用下, 将供体S-腺苷甲硫氨酸上的1个甲基转移至胞嘧啶的第5位碳原子上, 从而形成5-甲基胞嘧啶(m5C)的过程(Sahu et al., 2013)。1925年, m5C首次在结核分枝杆菌的结核菌素的水解产物中被发现(Johnson and Coghill, 1925)。随后, 在植物中发现了较高水平的m5C甲基化修饰(Vanyushin and Belozerskii, 1959)。DNA胞嘧啶甲基化修饰主要包括不对称(mCpHpH)甲基化和对称(mCpG和mCpHpG)甲基化。例如, 在拟南芥(Arabidopsis thaliana)中, CG、CHG与CHH的甲基化水平分别为24%、6.7%和1.7%; 并且在同一植物的不同组织或同一组织的不同发育阶段, 其基因组DNA的甲基化位点和水平也不一样(Dhar et al., 2014)。 植物基因组DNA甲基化的方式主要有2种, 一种是从头甲基化, 在DNA复制后的新生链中, DNA甲基化酶在没有甲基化修饰的位点上重新催化DNA甲基化。第2种是维持甲基化, 即在半甲基化的新生DNA双链中, 新合成的DNA链以旧模板链为范本, 完成另一条新合成的DNA单链的甲基化。由于其对称特性, 在DNA复制之后, CpG和CpHpG位点的甲基化比较容易被复制, 所以该位点的甲基化通常是通过维持机制实现。相比之下, 非对称的CpHpH位点的甲基化则需要在每一次的DNA复制后重新建立(Karlsson et al., 2011)。研究发现, 3种位点的甲基化由3种特定的甲基化酶催化, 分别为MET1 (DNA methyltransfer- ase 1)、CMT3 (Chromomethylase 3)和DRM (Domains rearranged methylase)。MET1是哺乳动物Dnmt1的同源基因, 主要负责CpG位点的甲基化, 在拟南芥met1突变体中, DNA全局性的甲基化程度明显下降, 导致拟南芥表现出晚花表型(Kankel et al., 2003); CMT3是植物特有的甲基化酶, 负责着丝粒附近的重复序列以及转座子的CpHpG位点甲基化(Lindroth et al., 2001)。DRM特异性介导植物DNA上CHG与CHH位点的甲基化修饰。研究发现, DRM2负责的对称与非对称位点甲基化修饰主要通过RNA介导的DNA甲基化修饰实现, 即RdDM (RNA-directed DNA methylation)途径完成。植物基因组全局性DNA甲基化水平需要通过DNA甲基化和去甲基化的动态平衡进行调节, 然而DNA去甲基化的机制一直备受争议。人们普遍认为DNA去甲基化既存在主动去甲基化过程又存在被动去甲基化过程。被动去甲基化的发生主要是由于DNA复制后从头甲基化的过程被抑制, 或者亲本甲基化印迹不能够被维持(Kankel et al., 2003)。而主动去甲基化的过程主要由一些DNA糖基化酶和AP裂解酶调控, 如DME (DEMETER)、DML2 (DEMETER LIKE 2)、DML3以及ROS1 (REPRES- SOR OF SILENCING 1)等(Gong et al., 2002; Kino- shita et al., 2004)。
1.2 DNA甲基化调控植物响应非生物胁迫DNA甲基化修饰在调节植物响应外界环境胁迫中发挥了非常重要的作用。最近几年的研究表明, 全基因组DNA甲基化修饰的改变与植物响应非生物胁迫密切相关(Boyko et al., 2010; Karan et al., 2012; Wang et al., 2014)。许多与DNA甲基化修饰相关的基因参与了植物的非生物胁迫应答过程(表1)。在植物响应高盐、干旱、温度和重金属等非生物胁迫过程中, 植物的特定基因或整个基因组水平的DNA甲基化修饰都会发生改变, 从而提高植物对不良环境的适应性, 确保植物在逆境条件下的生长发育。通常情况下, 非生物胁迫诱导的DNA甲基化修饰的改变发生在整个基因组水平。例如, Wang等(2011)以水稻(Oryza sativa) DK151和IR64为研究材料, 发现由干旱胁迫诱导的DNA甲基化位点的改变占全基因组DNA甲基化位点的12%左右, 这些甲基化的变化有29%在胁迫解除时仍然被保留; 并且干旱诱导的DNA甲基化还表现出组织及发育时期的特异性。Ferreira等(2015)在研究盐胁迫下DNA甲基化水平的变化时, 发现盐胁迫耐受型水稻材料Pokkali和敏感型水稻材料IR29基因组全局性的DNA甲基化水平在盐胁迫下发生了明显改变, 并且这种DNA甲基化水平的改变具有基因型及组织特异性。此外, 非生物胁迫下, DNA甲基化水平的改变往往还会伴随其它调节机制。例如, Karan等(2012)在研究水稻响应盐胁迫时, 发现逆转录转座子、非生物胁迫应答基因以及染色质修饰因子在盐胁迫下都具有不同的表达模式, 同时鉴定了不同水稻品种盐胁迫诱导的DNA甲基化模式。而对于水稻而言, 在水稻根细胞内相关基因的去甲基化修饰对其响应盐胁迫也具有重要作用(Wang et al., 2011)。Zhu等(2015)在研究水稻R2R3类型的MYB转录因子OsMYB91时, 发现OsMYB91的表达受非生物胁迫诱导, 特别是盐胁迫。进一步研究发现, 该基因启动子区域的DNA甲基化水平改变是导致该基因在胁迫下表达发生变化的主要原因。除了拟南芥和水稻等常见模式植物, 人们也对其它植物响应胁迫时的DNA甲基化修饰进行了研究。例如, Bharti等(2015)在研究盐胁迫下, 类黄酮生物合成和抗氧化途径的酶基因是否受到表观遗传调控时, 发现这些基因启动子区域甲基化状态改变使得相关基因的表达水平升高, 从而导致过表达AtROS1的转基因烟草(Nicotiana tabacum)表现出对盐胁迫的耐受性。Lu等(2017)利用Me-DIP测序技术对棉花(Gossypium hirsutum)在盐胁迫下全基因组的DNA甲基化水平进行了研究, 为理解棉花盐胁迫反应的表观遗传修饰变化提供了有价值的信息。同样, 也有研究发现, 苜蓿(Medicago spp.)在盐胁迫下, 全基因组水平的DNA甲基化程度增加, 特别是在高盐胁迫下尤为明显(Al-Lawati et al., 2016)。目前, 有关DNA甲基化修饰水平影响植物响应非生物胁迫的具体机制还有待深入研究。 表1 Table 1 表1 表1 植物中受DNA甲基化修饰调节的非生物胁迫应答相关基因 Table 1 Abiotic stress responsive genes regulated by DNA-methylation in plants
基因
物种
胁迫类型
参考文献
Asr1, Asr2
番茄(Solanum lycopersicum)
干旱胁迫
González et al., 2011; 2013
NtGPDL
烟草(Nicotiana tabaccum cv. ‘Xanthi nc’)
低温胁迫
Choi and Sano, 2007
OsMYB91
水稻(Oryza sativa)
盐胁迫
Zhu et al., 2015
Glyma11g02400 Glyma16g27950 Glyma20g30840
大豆(Glycine max)
盐胁迫
Song et al., 2012
SPCH, FAMA
拟南芥(Arabidopsis thaliana)
湿度胁迫
Tricker et al., 2012
ZmMI1, Ac/Ds
玉米(Zea mays)
低温胁迫
Steward et al., 2002
Tam3
金鱼草(Antirrhinum majus)
低温胁迫
Hashida et al., 2006
NRPD2
拟南芥(A. thaliana)
高温胁迫
Popova et al., 2013
表1 植物中受DNA甲基化修饰调节的非生物胁迫应答相关基因 Table 1 Abiotic stress responsive genes regulated by DNA-methylation in plants
2 组蛋白修饰2.1 植物体内的组蛋白修饰组蛋白修饰是表观遗传调控研究领域的热点。核小体是染色质的基本组成单位, 由约146 bp的DNA缠绕各两分子的核心组蛋白H2A、H2B、H3和H4形成的八聚体构成(Luger et al., 1997)。伸出核小体外的核心组蛋白的N端可被共价修饰, 这些修饰主要包括乙酰化、甲基化、磷酸化、泛素化、生物素化、ADP核糖基化以及类泛素化等(Strahl and Allis, 2000)。核心组蛋白N端赖氨酸可被组蛋白乙酰化酶HATs (histone acetyltransferase)和组蛋白去乙酰化酶HDACs (histone deacetylation transferase)进行乙酰化和去乙酰化修饰。在拟南芥中, HATs分为4个不同的家族, 研究表明HATs在植物发育(Bertrand et al., 2003; Long et al., 2006)和胁迫应答(Pavangadkar et al., 2010)中具有重要作用。真核生物的HDACs包含RP- D3 (Reduced Potassium Deficiency 3)、SIR2 (Silent Information Regulator 2)和HD2 (TypeIIHDAC) 3个主要蛋白家族, 其中HD2是植物特有的HDACs (Pan- dey et al., 2002)。在拟南芥中含有10个RPD3/HDA1去乙酰化酶家族蛋白(Alinsug et al., 2009), 其中HDA6能影响转基因的表达(Murfett et al., 2001)、DNA甲基化(Aufsatz et al., 2002)以及rRNA基因的调控(Probst et al., 2004)。同时研究发现, HDA19能与WRKY转录因子结合, 从而调控植物的基本防御反应(Kim et al., 2008)。另外, HDA6和HDA19在拟南芥种子萌发、盐胁迫以及ABA诱导的基因表达方面都发挥十分重要的作用(Chen et al., 2010)。 组蛋白的甲基化和去甲基化通常发生在精氨酸和赖氨酸残基上, 赖氨酸残基的甲基化主要由含有SET结构域的赖氨酸甲基转移酶HKMTs催化形成(Pontvianne et al., 2010)。SET结构域蛋白可分为TrxG (Trithorax Group)、E(Z) (Enhancer of Zeste)、SU (VAR)以及ASH1 (Absent, Small, or Homeotic dis- cs) 4个家族, 分别负责组蛋白H3K4、H3K9、H3K27和H3K36的甲基化修饰。在拟南芥中, TrxG家族蛋白SDG25 (Berr et al., 2009)、E(Z)家族蛋白CLF (Chanvivattana et al., 2004)以及ASH1家族蛋白SDG8和SDG26 (Xu et al., 2008)都参与开花时间的调控。组蛋白去甲基化酶HDMs包括KDM1/LSD1家族和JmjC结构域蛋白家族两类(Liu et al., 2010)。这两类蛋白质都可通过氧化反应直接去除组蛋白上的甲基化修饰。KDM1/LSD1以黄素为辅助因子, 只能对一、二甲基化修饰进行去甲基, 并不能对三甲基化去甲基; JmjC结构域蛋白需要二价铁和α酮戊二酸作辅助因子, 可以对一、二、三甲基化修饰去甲基。截至目前, 拟南芥中报道了4个KDM1/LSD1家族以及21个JMJ家族组蛋白去甲基化酶, 它们主要参与拟南芥的开花调控(Saze et al., 2008; Liu et al., 2010)。组蛋白的乙酰化、磷酸化和泛素化修饰一般与基因转录激活有关, SUMO化和生物素化与转录抑制相关。已有的研究显示, 与胁迫相关的基因激活主要由H3K4、H3K36甲基化以及H3K9乙酰化调控, 而H3K9、H3K27甲基化和H3去乙酰化主要参与相关基因的沉默过程(Qiao and Fan, 2011)。
2.2 组蛋白修饰与植物响应非生物胁迫除DNA甲基化修饰外, 组蛋白修饰也是植物响应非生物胁迫的重要表观调控途径之一。近年来, 有许多关于组蛋白甲基化、乙酰化和磷酸化等修饰参与植物非生物胁迫应答过程的报道(表2)。在模式植物拟南芥中, SKB1可以改变胁迫相关基因的H4R3sme2水平, 其缺失突变会导致拟南芥产生对盐胁迫超敏感的表型(Zhang et al., 2011b)。类似地, 拟南芥组蛋白乙酰转移酶GCN5通过影响HSFA3 (Heat Stress Transcription Factors 3)和UVH6 (UV-HYPERSENSI- TIVE 6)基因启动子区域的H3K9和H3K14的乙酰化水平来调节应答基因的转录。因此, 缺失GCN5的拟南芥突变体表现出对热胁迫的耐受性(Hu et al., 2015)。在水稻中, 组蛋白乙酰转移酶基因OsHATs的转录水平和组蛋白H3和H4赖氨酸残基乙酰化水平的变化都参与了水稻的干旱胁迫调控(Fang et al., 2014)。Luo等(2012)在研究植物特异的组蛋白去乙酰化酶HD2时, 发现ABA和NaCl能抑制HD2A、HD2B、HD2C以及HD2D等基因的表达水平, HD2的T-DNA插入突变体表现出对ABA和NaCl的敏感性, 同时降低了对盐胁迫的耐受性。此外, 在组蛋白去乙酰化方面, Zheng等(2016)发现拟南芥组蛋白去乙酰化酶HDA9在调节拟南芥响应盐胁迫和干旱胁迫时发挥重要作用。由此可见, 组蛋白的多种修饰类型都参与了非生物胁迫的调节过程。 表2 Table 2 表2 表2 植物中参与非生物胁迫应答的组蛋白修饰相关基因 Table 2 Histone modification related genes involved in abiotic stresses in plants
基因
物种
胁迫类型
参考文献
HDACs
拟南芥(Arabidopsis thaliana )
盐胁迫
Luo et al., 2012
SKB1
拟南芥(A. thaliana )
盐胁迫
Zhang et al., 2011b
MSI1, HDA19
拟南芥(A. thaliana )
盐胁迫
Mehdi et al., 2016
HDA9
拟南芥(A. thaliana )
盐胁迫, 干旱胁迫
Zheng et al., 2016
HDA705
水稻(Oryza sativa)
盐胁迫
Zhao et al., 2016
GCN5
拟南芥(A. thaliana )
热胁迫
Hu et al., 2015
HDT701
水稻(O. sativa)
盐胁迫
Zhao et al., 2014
AtABO1
拟南芥(A. thaliana )
干旱胁迫
Chen et al., 2006
ADH1, PDC1
水稻(O. sativa)
淹水胁迫
Tsuji et al., 2006
OsHAG702, OsHAG704 OsHAC701, OsHAC704
水稻(O. sativa)
干旱胁迫
Liu et al., 2012
OsHAG703, OsHAM701 OsHAC703, OsHAF701
水稻(O. sativa)
干旱胁迫
Liu et al., 2012; Fang et al., 2014
AtATX1
拟南芥(A. thaliana )
干旱胁迫
Ding et al., 2011
OsDREB1bc
水稻(O. sativa)
盐胁迫, 低温胁迫
Roy et al., 2014
HvTX1, HvPKDM7
大麦(Hordeum vulgare)
干旱胁迫
Papaefthimiou and Tsaftaris, 2012
MYST, ELP3, GCN5
大麦(H. vulgare)
干旱胁迫
Papaefthimiou et al., 2010
AtHD2C
拟南芥(A. thaliana )
干旱胁迫
Sridha and Wu, 2006
AtMSI1, AtCHR12
拟南芥(A. thaliana )
干旱胁迫, 高温胁迫
Alexandre et al., 2009
AtBRM
拟南芥(A. thaliana )
干旱胁迫
Berr et al., 2012
表2 植物中参与非生物胁迫应答的组蛋白修饰相关基因 Table 2 Histone modification related genes involved in abiotic stresses in plants
AlexandreC, Möller-SteinbachY, SchönrockN, GruissemW, HennigL (2009). Arabidopsis MSI1 is required for negative regulation of the response to drought stress.Mol Plant 2, 675-687. [本文引用: 1]
[2]
AlinsugMV, YuCW, WuKQ (2009). Phylogenetic analysis, subcellular localization, and expression patterns of RPD3/ HDA1 family histone deacetylases in plants.BMC Plant Biol 9, 37. [本文引用: 1]
[3]
Al-LawatiA, Al-BahryS, VictorR, AL-LawatiAH, YaishMW (2016). Salt stress alters DNA methylation levels in alfalfa (Medicago spp.). Genet Mol Res 15, 15018299. [本文引用: 1]
[4]
AufsatzW, MetteMF, Van Der WindenJ, MatzkeM, MatzkeAJM (2002). HDA6, a putative histone deacety- lase needed to enhance DNA methylation induced by dou- ble-stranded RNA.EMBO J 21, 6832-6841. [本文引用: 1]
[5]
AvramovaZ (2015). Transcriptional ‘memory’ of a stress: transient chromatin and memory (epigenetic) marks at st- ress-response genes.Plant J 83, 149-159. [本文引用: 1]
[6]
BerrA, MénardR, HeitzT, ShenWH (2012). Chromatin modification and remodelling: a regulatory landscape for the control of Arabidopsis defence responses upon patho- gen attack.Cell Microbiol 14, 829-839. [本文引用: 1]
[7]
BerrA, XuL, GaoJ, CognatV, SteinmetzA, DongAW, ShenWH (2009). SET DOMAIN GROUP25 encodes a histone methyltransferase and is involved in FLOWERING LOCUS C activation and repression of flowering. Plant Phy- siol 151, 1476-1485. [本文引用: 1]
[8]
BertrandC, BergouniouxC, DomenichiniS, DelarueM, ZhouDX (2003). Arabidopsis histone acetyltransferase AtGCN5 regulates the floral meristem activity through the WUSCHEL/AGAMOUS pathway. J Biol Chem 278, 28246-28251. [本文引用: 1]
[9]
BhartiP, MahajanM, VishwakarmaAK, BhardwajJ, YadavSK (2015). AtROS1 overexpression provides evidence for epigenetic regulation of genes encoding enzymes of flavonoid biosynthesis and antioxidant pathways during salt stress in transgenic tobacco. J Exp Bot 66, 5959-5969. [本文引用: 1]
[10]
BostockRM (2005). Signal crosstalk and induced resistance: straddling the line between cost and benefit.Annu Rev Phytopathol 43, 545-580. [本文引用: 1]
[11]
BoykoA, BlevinsT, YaoY, GolubovA, BilichakA, IlnytskyyY, HollunderJ, Meins JrF, KovalchukI (2010). Transgenerational adaptation of Arabidopsis to stress requires DNA methylation and the function of dicer-like proteins.PLoS One 5, e9514. [本文引用: 1]
[12]
BoykoA, KovalchukI (2008). Epigenetic control of plant stress response.Environ Mol Mutagen 49, 61-72. [本文引用: 1]
[13]
ChanvivattanaY, BishoppA, SchubertD, StockC, MoonYH, SungZR, GoodrichJ (2004). Interaction of polycomb-group proteins controlling flowering in Arabidopsis.Development 131, 5263-5276. [本文引用: 1]
[14]
ChenFF, HeGM, HeH, ChenW, ZhuXP, LiangMZ, ChenLB, DengXW (2010). Expression analysis of miRNAs and highly-expressed small RNAs in two rice subspecies and their reciprocal hybrids.J Integr Plant Biol 52, 971-980. [本文引用: 1]
[15]
ChenLT, WuKQ (2010). Role of histone deacetylases HDA6 and HDA19 in ABA and abiotic stress response.Plant Sig- nal Behav 5, 1318-1320. [本文引用: 1]
[16]
ChenZZ, ZhangHR, JablonowskiD, ZhouXF, RenXZ, HongXH, SchaffrathR, ZhuJK, GongZZ (2006). Mutations in ABO1/ELO2, a subunit of holo-Elongator, increase abscisic acid sensitivity and drought tolerance in Arabidopsis thaliana. Mol Cell Biol 26, 6902-6912. [本文引用: 1]
[17]
ChoiCS, SanoH (2007). Abiotic-stress induces demethyla- tion and transcriptional activation of a gene encoding a glycerophosphodiesterase-like protein in tobacco plants.Mol Genet Genomics 277, 589-600. [本文引用: 1]
DharMK, VishalP, SharmaR, KaulS (2014). Epigenetic dynamics: role of epimarks and underlying machinery in plants exposed to abiotic stress.Int J Genomics2014, 187146. [本文引用: 1]
[20]
DingY, AvramovaZ, FrommM (2011). The Arabidopsis trithorax-like factor ATX1 functions in dehydration stress responses via ABA-dependent and ABA-independent pathways.Plant J 66, 735-744. [本文引用: 2]
FangH, LiuX, ThornG, DuanJ, TianLN (2014). Expression analysis of histone acetyltransferases in rice under drought stress.Biochem Biophys Res Commun 443, 400-405. [本文引用: 2]
[23]
FengQZ, YangCW, LinXY, WangJM, OuXF, ZhangCY, LiuB (2012). Salt and alkaline stress induced transgene- rational alteration in DNA methylation of rice (Oryza sativa). Aust J Crop Sci 6, 877. [本文引用: 1]
[24]
FerreiraLJ, AzevedoV, MarocoJ, OliveiraMM, SantosAP (2015). Salt tolerant and sensitive rice varieties display differential methylome flexibility under salt stress.PLoS One 10, e0124060. [本文引用: 1]
[25]
FoltaA, SeveringEI, KrauskopfJ, Van De GeestH, VerverJ, NapJP, MlynarovaL (2014). Over-expression of Arabidopsis AtCHR23 chromatin remodeling ATPase results in increased variability of growth and gene expression. BMC Plant Biol 14, 76. [本文引用: 1]
[26]
FujitaM, FujitaY, NoutoshiY, TakahashiF, NarusakaY, YamaguchiSK, ShinozakiK (2006). Crosstalk between abiotic and biotic stress responses: a current view from the points of convergence in the stress signaling networks.Curr Opin Plant Biol 9, 436-442. [本文引用: 1]
[27]
GeimanTM, RobertsonKD (2002). Chromatin remodeling, histone modifications, and DNA methylation—how does it all fit together?J Cell Biochem 87, 117-125. [本文引用: 1]
[28]
GongZZ, Morales-RuizT, ArizaRR, Roldán-ArjonaT, DavidL, ZhuJK (2002). ROS1, a repressor of transcriptional gene silencing in Arabidopsis, encodes a DNA glycosylase/lyase. Cell 11, 803-814. [本文引用: 1]
[29]
GonzálezRM, RicardiMM, IusemND (2011). Atypical epigenetic mark in an atypical location: cytosine methylation at asymmetric (CNN) sites within the body of a non-repeti- tive tomato gene.BMC Plant Biol 11, 94. [本文引用: 1]
[30]
GonzálezRM, RicardiMM, IusemND (2013). Epigenetic marks in an adaptive water stress-responsive gene in tomato roots under normal and drought conditions.Epigenetics 8, 864-872. [本文引用: 1]
[31]
GuanQM, LuXY, ZengHT, ZhangYY, ZhuJH (2013). Heat stress induction of miR398 triggers a regulatory loop that is critical for thermotolerance in Arabidopsis. Plant J 74, 840-851. [本文引用: 1]
[32]
GuoMX, WangRC, WangJ, HuaK, WangYM, LiuXQ, YaoSG (2014). ALT1, a Snf2 family chromatin remodeling ATPase, negatively regulates alkaline tolerance through enhanced defense against oxidative stress in rice.PLoS One 9, e112515. [本文引用: 1]
HanSK, SangY, RodriguesA, WuMF, RodriguezPL, WagnerD (2012). The SWI2/SNF2 chromatin remodeling ATPase BRAHMA represses abscisic acid responses in the absence of the stress stimulus in Arabidopsis.Plant Cell 24, 4892-4906. [本文引用: 2]
[35]
HashidaSN, UchiyamaT, MartinC, KishimaY, SanoY, MikamiT (2006). The temperature-dependent change in methylation of the Antirrhinum transposon Tam3 is controlled by the activity of its transposase.Plant Cell 18, 104-118. [本文引用: 1]
[36]
HeGM, ZhuXP, EllingAA, ChenLB, WangXF, GuoL, LiangMZ, HeH, ZhangHY, ChenFF, QiYJ, ChenRS, DengXW (2010). Global epigenetic and transcriptional trends among two rice subspecies and their reciprocal hybrids.Plant Cell 22, 17-33. [本文引用: 1]
[37]
HuZR, SongN, ZhengM, LiuXY, LiuZS, XingJW, MaJH, GuoWW, YaoYY, PengHR, XinMM, ZhouDX, NiZF, SunQX (2015). Histone acetyltransferase GCN5 is essential for heat stress-responsive gene activation and thermotolerance in Arabidopsis.Plant J 84, 1178-1191. [本文引用: 2]
[38]
HuangW, XianZQ, HuGJ, ZhengGL (2016). SlAGO4A, a core factor of RNA-directed DNA methylation (RdDM) pathway, plays an important role under salt and drought stress in tomato.Mol Breed 36, 28. [本文引用: 2]
[39]
JamesA, WangYB, RajeH, RosbyR, DiMarioP (2014). Nucleolar stress with and without p53.Nucleus 5, 402-426. [本文引用: 1]
[40]
JohnsonTB, CoghillRD (1925). Researches on pyrimidines. C111. The discovery of 5-methyl-cytosine in tuberculinic acid, the nucleic acid of the tubercle bacillus.J Am Chem Soc 11, 2838-2844. [本文引用: 1]
KaranR, DeLeonT, BiradarH, SubudhiPK (2012). Salt stress induced variation in DNA methylation pattern and its influence on gene expression in contrasting rice genoty- pes.PLoS One 7, e40203. [本文引用: 2]
[44]
KarlssonM, WeberW, FusseneggerM (2011). De novo design and construction of an inducible gene expres- sion system in mammalian cells. Meth Enzymol 497, 239-253. [本文引用: 1]
[45]
KimJM, ToTK, IshidaJ, MatsuiA, KimuraH, SekiM (2012). Transition of chromatin status during the process of recovery from drought stress in Arabidopsis thaliana. Plant Cell Physiol 53, 847-856. [本文引用: 1]
[46]
KimKC, LaiZB, FanBF, ChenZX (2008). Arabidopsis WRKY38 and WRKY62 transcription factors interact with histone deacetylase 19 in basal defense.Plant Cell 20, 2357-2371. [本文引用: 1]
[47]
KinoshitaT, MiuraA, ChoiY, YukiK, CaoXF, JacobsenSE, FischerRL, KakutaniT (2004). One-way control of FWA imprinting in Arabidopsis endosperm by DNA met- hylation. Science 303, 521-523. [本文引用: 1]
[48]
KinoshitaT, SekiM (2014). Epigenetic memory for stress response and adaptation in plant.Plant Cell Physiol 55, 1859-1863. [本文引用: 2]
LiuZQ, GaoJ, DongAW, ShenWH (2009). A truncated Arabidopsis NUCLEOSOME ASSEMBLY PROTEIN 1, At- NAP1;3T, alters plant growth responses to abscisic acid and salt in the Atnap1;3-2 mutant. Mol Plant 2, 688-699. [本文引用: 1]
[55]
LongJA, OhnoC, SmithZR, MeyerowitzEM (2006). TOP- LESS regulates apical embryonic fate in Arabidopsis.Sci- ence 312, 1520-1523. [本文引用: 1]
LuXK, ShuN, WangJJ, ChenXG, WangDL, WangS, FanWL, GuoXN, GuoLX, YeWW (2017). Genome-wide analysis of salinity-stress induced DNA methylation alterations in cotton (Gossypium hirsutum L.) using the Me-DIP sequencing technology. Genet Mol Res 16, 1-16. [本文引用: 1]
[58]
LugerK, MäderAW, RichmondRK, SargentDF, RichmondTJ (1997). Crystal structure of the nucleosome core particle at 2.8 Å resolution.Nature 389, 251-260. [本文引用: 1]
[59]
LuoM, WangYY, LiuXC, YangSG, LuQ, CuiYH, WuKQ (2012). HD2C interacts with HDA6 and is involved in ABA and salt stress response in Arabidopsis.J Exp Bot 63, 3297-3306. [本文引用: 2]
[60]
MadlungA, ComaiL (2004). The effect of stress on genome regulation and structure.Ann Bot 94, 481-495. [本文引用: 1]
[61]
MalloryAC, VaucheretH (2006). Function of microRNAs and related small RNAs in plants.Nat Genet 38, 31-36. [本文引用: 1]
[62]
Mauch-ManiB, MauchF (2005). The role of abscisic acid in plant-pathogen interactions.Curr Opin Plant Biol 8, 409-414. [本文引用: 1]
[63]
MehdiS, DerkachevaM, RamströmM, KralemannL, BergquistJ, HennigL (2016). The WD40 domain protein MSI1 functions in a histone deacetylase complex to fine- tune abscisic acid signaling.Plant Cell 28, 42-54. [本文引用: 2]
[64]
MillerMJ, Barrett-WiltGA, HuaZZ, VierstraRD (2010). Proteomic analyses identify a diverse array of nuclear processes affected by small ubiquitin-like modifier conjugation in Arabidopsis.Proc Natl Acad Sci USA 107, 16512-16517. [本文引用: 1]
[65]
MlynárováL, NapJP, BisselingT (2007). The SWI/SNF chromatin-remodeling geneAtCHR12 mediates temporary growth arrest in Arabidopsis thaliana upon perceiving environmental stress. Plant J 51, 874-885. [本文引用: 2]
NaydenovM, BaevV, ApostolovaE, GospodinovaN, SablokG, GozmanovaM, YahubyanG (2015). High- temperature effect on genes engaged in DNA methylation and affected by DNA methylation in Arabidopsis.Plant Physiol Biochem 87, 102-108. [本文引用: 1]
[68]
NiZY, HuZ, JiangQY, ZhangH (2013). GmNFYA3, a target gene of miR169, is a positive regulator of plant tolerance to drought stress. Plant Mol Biol 82, 113-129. [本文引用: 1]
[69]
PandeyR, MüllerA, NapoliCA, SelingerDA, PikaardCS, RichardsEJ, BenderJ, MountDW, JorgensenRA (2002). Analysis of histone acetyltransferase and histone deacetylase families ofArabidopsis thaliana suggests functional diversification of chromatin modification among multicellular eukaryotes. Nucleic Acids Res 30, 5036-5055. [本文引用: 1]
[70]
PapaefthimiouD, LikotrafitiE, KapazoglouA, BladenopoulosK, TsaftarisA (2010). Epigenetic chromatin modifiers in barley: III. Isolation and characterization of the barley GNAT-MYST family of histone acetyltransferases and responses to exogenous ABA.Plant Physiol Biochem 48, 98-107. [本文引用: 1]
[71]
PapaefthimiouD, TsaftarisAS (2012). Significant induction by drought of HvPKDM7-1, a gene encoding a jumonji-like histone demethylase homologue in barley(H. vulgare). Ac- ta Physiol Plant 34, 1187-1198. [本文引用: 1]
[72]
PavangadkarK, ThomashowMF, TriezenbergSJ (2010). Histone dynamics and roles of histone acetyltransferases during cold-induced gene regulation in Arabidopsis.Plant Mol Biol 74, 183-200. [本文引用: 1]
[73]
Peirats-LlobetM, HanSK, Gonzalez-GuzmanM, JeongCW, RodriguezL, Belda-PalazonB, WagnerD, RodriguezPL (2015). A direct link between abscisic acid sensing and the chromatin-remodeling ATPase BRAHMA via core ABA signaling pathway components.Mol Plant 9, 136-147. [本文引用: 1]
[74]
PerrucE, KinoshitaN, Lopez-MolinaL (2007). The role of chromatin-remodeling factor PKL in balancing osmotic stress responses during Arabidopsis seed germination.Plant J 52, 927-936. [本文引用: 1]
[75]
PhillipsJR, DalmayT, BartelsD (2007). The role of small RNAs in abiotic stress.FEBS Lett 581, 3592-3597. [本文引用: 1]
PopovaOV, DinhHQ, AufsatzW, JonakC (2013). The RdDM pathway is required for basal heat tolerance in Arabidopsis.Mol Plant 6, 396-410. [本文引用: 1]
[78]
ProbstAV, FagardM, ProuxF, MourrainP, BoutetS, EarleyK, LawrenceRJ, PikaardCS, MurfettJ, FurnerL, VaucheretH, ScheidOM (2004). Arabidopsis histone deacetylase HDA6 is required for maintenance of trans- criptional gene silencing and determines nuclear organization of rDNA repeats.Plant Cell 16, 1021-1034. [本文引用: 1]
[79]
QiaoWH, FanLM (2011). Epigenetics, a mode for plants to respond to abiotic stresses.Front Biol 6, 477-481. [本文引用: 1]
[80]
RackiLR, NarlikarGJ (2008). ATP-dependent chromatin remodeling enzymes: two heads are not better, just different.Curr Opin Genet Dev 18, 137-144. [本文引用: 1]
[81]
RamachandranV, ChenXM (2008). Small RNA metabolism in Arabidopsis.Trends Plant Sci 13, 368-374. [本文引用: 1]
[82]
RoyD, PaulA, RoyA, GhoshR, GangulyP, ChaudhuriS (2014). Differential acetylation of histone H3 at the regulatory region of OsDREB1b promoter facilitates chromatin remodelling and transcription activation during cold stress. PLoS One 9, e100343. [本文引用: 2]
[83]
RussoVEA, MartienssenRA, RiggsAD (1996). Epigenetic Mechanisms of Gene Regulation. Plainview, NY: Cold Sp- ring Harbor Laboratory Press. [本文引用: 1]
[84]
SaezA, RodriguesA, SantiagoJ, RubioS, RodriguezPL (2008). HAB1-SWI3B interaction reveals a link between abscisic acid signaling and putative SWI/SNF chromatin-remodeling complexes in Arabidopsis.Plant Cell 20, 2972-2988. [本文引用: 2]
SazeH, ShiraishiA, MiuraA, KakutaniT (2008). Control of genic DNA methylation by a jmjC domain-containing protein in Arabidopsis thaliana. Science 319, 462-465. [本文引用: 1]
[87]
SongYG, JiDD, LiS, WangP, LiQ, XiangFN (2012). The dynamic changes of DNA methylation and histone modifications of salt responsive transcription factor genes in soybean.PLoS One 7, e41274. [本文引用: 1]
[88]
Sosa-ValenciaG, PalomarM, CovarrubiasAA, ReyesJ (2017). The legume miR1514a modulates a NAC trans- cription factor transcript to trigger phasiRNA formation in response to drought.J Exp Bot 68, 2013-2026. [本文引用: 1]
[89]
SridhaS, WuKQ (2006). Identification of AtHD2C as a novel regulator of abscisic acid responses in Arabidopsis. Plant J 46, 124-133. [本文引用: 1]
[90]
StewardN, ItoM, YamaguchiY, KoizumiN, SanoH (2002). Periodic DNA methylation in maize nucleosomes and demethylation by environmental stress.J Biol Chem 277, 37741-37746. [本文引用: 1]
[91]
StrahlBD, AllisCD (2000). The language of covalent histone modifications.Nature 403, 41-45. [本文引用: 1]
[92]
SudarsanamP, WinstonF (2000). The Swi/Snf family: nucleosome-remodeling complexes and transcriptional control.Trends Genet 16, 345-351. [本文引用: 1]
[93]
SunkarR, ChinnusamyV, ZhuJH, ZhuJK (2007). Small RNAs as big players in plant abiotic stress responses and nutrient deprivation.Trends Plant Sci 12, 301-309. [本文引用: 1]
[94]
TorresMA, DanglJL (2005). Functions of the respiratory burst oxidase in biotic interactions, abiotic stress and development.Curr Opin Plant Biol 8, 397-403. [本文引用: 1]
[95]
TrickerPJ, GibbingsJG, LópezCMR, HadleyP, WilkinsonMJ (2012). Low relative humidity triggers RNA-di- rected de novo DNA methylation and suppression of genes controlling stomatal development. J Exp Bot 63, 3799-3813. [本文引用: 2]
[96]
TsujiH, SaikaH, TsutsumiN, HiraiA, NakazonoM (2006). Dynamic and reversible changes in histone H3-Lys4 methylation and H3 acetylation occurring at submergence- inducible genes in rice.Plant Cell Physiol 47, 995-1003. [本文引用: 2]
[97]
VanyushinBF, BelozerskiiAN (1959). Nucleotide composition of deoxyribonucleic acid in higher plants.Dokl Akad Nauk SSSR 129, 944-946. [本文引用: 1]
[98]
VrietC, HennigL, LaloiC (2015). Stress-induced chromatin changes in plants: of memories, metabolites and crop improvement.Cell Mol Life Sci 72, 1261-1273. [本文引用: 1]
[99]
WangM, QinLM, XieC, LiW, YuanJR, KongLN, YuWL, XiaGM, LiuSW (2014). Induced and constitutive DNA methylation in a salinity-tolerant wheat introgression line.Plant Cell Physiol 55, 1354-1365. [本文引用: 1]
[100]
WangP, ZhaoL, HouHL, ZhangH, HuangY, WangYP, LiH, GaoF, YanSH, LiLJ (2015). Epigenetic changes are associated with programmed cell death induced by heat stress in seedling leaves of Zea mays. Plant Cell Physiol 56, 965-976. [本文引用: 1]
[101]
WangWS, PanYJ, ZhaoXQ, DwivediD, ZhuLH, AliJ, FuBY, LiZK (2011). Drought-induced site-specific DNA methylation and its association with drought tolerance in rice (Oryza sativa L.). J Exp Bot 62, 1951-1960. [本文引用: 2]
[102]
WangWS, ZhaoXQ, PanYJ, ZhuLH, FuBY, LiZK (2011). DNA methylation changes detected by methylation-sen- sitive amplified polymorphism in two contrasting rice geno- types under salt stress.J Genet Genomics 38, 419-424.
[103]
WengMJ, YangY, FengHY, PanZD, ShenWH, ZhuY, DongAW (2014). Histone chaperone ASF1 is involved in gene transcription activation in response to heat stress in Arabidopsis thaliana. Plant Cell Environ 37, 2128-2138. [本文引用: 2]
[104]
WuCT, MorrisJR (2001). Genes, genetics, and epigenetics: a correspondence.Science 293, 1103-1105. [本文引用: 1]
[105]
XuL, ZhaoZ, DongAW, Soubigou-TaconnatL, RenouJP, SteinmetzA, ShenWH (2008). Di- and tri- but not monomethylation on histone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsis thaliana. Mol Cell Biol 28, 1348-1360. [本文引用: 1]
[106]
YangCH, LiDY, MaoDH, LiuX, JiCJ, LiXB, ZhaoXF, ChengZK, ChenCY, ZhuLH (2013). Overexpression of microRNA319 impacts leaf morphogenesis and leads to enhanced cold tolerance in rice ( Oryza sativa L.). Plant Cell Environ 36, 2207-2218. [本文引用: 2]
ZhangJJ, LaiJB, WangFG, YangSG, HeZP, JiangJM, LiQL, WuQ, LiuYY, YuMY, DuJJ, XieQ, WuKQ, YangCW (2017). A SUMO ligase AtMMS21 regulates the stability of the chromatin remodeler BRAHMA in root deve- lopment.Plant Physiol 173, 1574-1582. [本文引用: 1]
[109]
ZhangL, PengYG, WeiXL, DaiY, YuanDW, LuYF, PanYY, ZhuZ (2014). Small RNAs as important regulators for the hybrid vigour of super-hybrid rice.J Exp Bot 65, 5989-6002. [本文引用: 1]
[110]
ZhangXH, ZouZ, GongPJ, ZhangJH, ZiafK, LiHX, XiaoFM, YeZB (2011a). Over-expression of microRNA169 confers enhanced drought tolerance to tomato.Biotechnol Lett 33, 403-409. [本文引用: 1]
[111]
ZhangZL, ZhangSP, ZhangY, WangX, LiD, LiQL, YueMH, LiQ, ZhangYE, XuYY, XueYB, ChongK, BaoSL (2011b). Arabidopsis floral initiator SKB1 confers high salt tolerance by regulating transcription and Pre-mRNA splicing through altering histone H4R3 and small nuclear ribonucleoprotein LSM4 methylation.Plant Cell 23, 396-411. [本文引用: 2]
[112]
ZhaoJH, LiMZ, GuDC, LiuXC, ZhangJX, WuKL, ZhangXH, Da SilvaJAT, DuanJ (2016). Involvement of rice histone deacetylase HDA705 in seed germination and in response to ABA and abiotic stresses.Biochem Biophys Res Commun 470, 439-444. [本文引用: 1]
[113]
ZhaoJH, ZhangJX, ZhangW, WuKL, ZhengF, TianLN, LiuXC, DuanJ (2014). Expression and functional analysis of the plant-specific histone deacetylase HDT701 in rice. Front Plant Sci 5, 764. [本文引用: 1]
[114]
ZhengY, DingY, SunX, XieSS, WangD, LiuXY, SuLF, WeiW, PanL, ZhouDX (2016). Histone deacetylase HDA9 negatively regulates salt and drought stress respon- siveness in Arabidopsis.J Exp Bot 67, 1703-1713. [本文引用: 2]
[115]
ZhouLG, LiuYH, LiuZC, KongDY, DuanM, LuoLJ (2010). Genome-wide identification and analysis of drought- responsive microRNAs in Oryza sativa.J Exp Bot 61, 4157-4168. [本文引用: 2]
[116]
ZhuN, ChengSF, LiuXY, DuH, DaiMQ, ZhouDX, YangWJ, ZhaoY (2015). The R2R3-type MYB gene OsMYB91 has a function in coordinating plant growth and salt stress tolerance in rice. Plant Sci 236, 146-156. [本文引用: 2]
1 2009
... Histone modification related genes involved in abiotic stresses in plants
... 组蛋白修饰是表观遗传调控研究领域的热点.核小体是染色质的基本组成单位, 由约146 bp的DNA缠绕各两分子的核心组蛋白H2A、H2B、H3和H4形成的八聚体构成(Luger et al., 1997).伸出核小体外的核心组蛋白的N端可被共价修饰, 这些修饰主要包括乙酰化、甲基化、磷酸化、泛素化、生物素化、ADP核糖基化以及类泛素化等(Strahl and Allis, 2000).核心组蛋白N端赖氨酸可被组蛋白乙酰化酶HATs (histone acetyltransferase)和组蛋白去乙酰化酶HDACs (histone deacetylation transferase)进行乙酰化和去乙酰化修饰.在拟南芥中, HATs分为4个不同的家族, 研究表明HATs在植物发育(Bertrand et al., 2003; Long et al., 2006)和胁迫应答(Pavangadkar et al., 2010)中具有重要作用.真核生物的HDACs包含RP- D3 (Reduced Potassium Deficiency 3)、SIR2 (Silent Information Regulator 2)和HD2 (TypeIIHDAC) 3个主要蛋白家族, 其中HD2是植物特有的HDACs (Pan- dey et al., 2002).在拟南芥中含有10个RPD3/HDA1去乙酰化酶家族蛋白(Alinsug et al., 2009), 其中HDA6能影响转基因的表达(Murfett et al., 2001)、DNA甲基化(Aufsatz et al., 2002)以及rRNA基因的调控(Probst et al., 2004).同时研究发现, HDA19能与WRKY转录因子结合, 从而调控植物的基本防御反应(Kim et al., 2008).另外, HDA6和HDA19在拟南芥种子萌发、盐胁迫以及ABA诱导的基因表达方面都发挥十分重要的作用(Chen et al., 2010). ...
1 2016
... DNA甲基化修饰在调节植物响应外界环境胁迫中发挥了非常重要的作用.最近几年的研究表明, 全基因组DNA甲基化修饰的改变与植物响应非生物胁迫密切相关(Boyko et al., 2010; Karan et al., 2012; Wang et al., 2014).许多与DNA甲基化修饰相关的基因参与了植物的非生物胁迫应答过程(表1).在植物响应高盐、干旱、温度和重金属等非生物胁迫过程中, 植物的特定基因或整个基因组水平的DNA甲基化修饰都会发生改变, 从而提高植物对不良环境的适应性, 确保植物在逆境条件下的生长发育.通常情况下, 非生物胁迫诱导的DNA甲基化修饰的改变发生在整个基因组水平.例如, Wang等(2011)以水稻(Oryza sativa) DK151和IR64为研究材料, 发现由干旱胁迫诱导的DNA甲基化位点的改变占全基因组DNA甲基化位点的12%左右, 这些甲基化的变化有29%在胁迫解除时仍然被保留; 并且干旱诱导的DNA甲基化还表现出组织及发育时期的特异性.Ferreira等(2015)在研究盐胁迫下DNA甲基化水平的变化时, 发现盐胁迫耐受型水稻材料Pokkali和敏感型水稻材料IR29基因组全局性的DNA甲基化水平在盐胁迫下发生了明显改变, 并且这种DNA甲基化水平的改变具有基因型及组织特异性.此外, 非生物胁迫下, DNA甲基化水平的改变往往还会伴随其它调节机制.例如, Karan等(2012)在研究水稻响应盐胁迫时, 发现逆转录转座子、非生物胁迫应答基因以及染色质修饰因子在盐胁迫下都具有不同的表达模式, 同时鉴定了不同水稻品种盐胁迫诱导的DNA甲基化模式.而对于水稻而言, 在水稻根细胞内相关基因的去甲基化修饰对其响应盐胁迫也具有重要作用(Wang et al., 2011).Zhu等(2015)在研究水稻R2R3类型的MYB转录因子OsMYB91时, 发现OsMYB91的表达受非生物胁迫诱导, 特别是盐胁迫.进一步研究发现, 该基因启动子区域的DNA甲基化水平改变是导致该基因在胁迫下表达发生变化的主要原因.除了拟南芥和水稻等常见模式植物, 人们也对其它植物响应胁迫时的DNA甲基化修饰进行了研究.例如, Bharti等(2015)在研究盐胁迫下, 类黄酮生物合成和抗氧化途径的酶基因是否受到表观遗传调控时, 发现这些基因启动子区域甲基化状态改变使得相关基因的表达水平升高, 从而导致过表达AtROS1的转基因烟草(Nicotiana tabacum)表现出对盐胁迫的耐受性.Lu等(2017)利用Me-DIP测序技术对棉花(Gossypium hirsutum)在盐胁迫下全基因组的DNA甲基化水平进行了研究, 为理解棉花盐胁迫反应的表观遗传修饰变化提供了有价值的信息.同样, 也有研究发现, 苜蓿(Medicago spp.)在盐胁迫下, 全基因组水平的DNA甲基化程度增加, 特别是在高盐胁迫下尤为明显(Al-Lawati et al., 2016).目前, 有关DNA甲基化修饰水平影响植物响应非生物胁迫的具体机制还有待深入研究. ...
1 2002
... 组蛋白修饰是表观遗传调控研究领域的热点.核小体是染色质的基本组成单位, 由约146 bp的DNA缠绕各两分子的核心组蛋白H2A、H2B、H3和H4形成的八聚体构成(Luger et al., 1997).伸出核小体外的核心组蛋白的N端可被共价修饰, 这些修饰主要包括乙酰化、甲基化、磷酸化、泛素化、生物素化、ADP核糖基化以及类泛素化等(Strahl and Allis, 2000).核心组蛋白N端赖氨酸可被组蛋白乙酰化酶HATs (histone acetyltransferase)和组蛋白去乙酰化酶HDACs (histone deacetylation transferase)进行乙酰化和去乙酰化修饰.在拟南芥中, HATs分为4个不同的家族, 研究表明HATs在植物发育(Bertrand et al., 2003; Long et al., 2006)和胁迫应答(Pavangadkar et al., 2010)中具有重要作用.真核生物的HDACs包含RP- D3 (Reduced Potassium Deficiency 3)、SIR2 (Silent Information Regulator 2)和HD2 (TypeIIHDAC) 3个主要蛋白家族, 其中HD2是植物特有的HDACs (Pan- dey et al., 2002).在拟南芥中含有10个RPD3/HDA1去乙酰化酶家族蛋白(Alinsug et al., 2009), 其中HDA6能影响转基因的表达(Murfett et al., 2001)、DNA甲基化(Aufsatz et al., 2002)以及rRNA基因的调控(Probst et al., 2004).同时研究发现, HDA19能与WRKY转录因子结合, 从而调控植物的基本防御反应(Kim et al., 2008).另外, HDA6和HDA19在拟南芥种子萌发、盐胁迫以及ABA诱导的基因表达方面都发挥十分重要的作用(Chen et al., 2010). ...
1 2015
... 通常情况下, 植物处于自然环境状态受到的非生物胁迫具有短暂性和循环性等特点.研究认为这种可预测的、循环的环境改变能够激发植物产生胁迫记忆.“防御启动”是目前人们最为熟知的一种胁迫记忆现象, 即植物会记忆病原体的第1次攻击, 从而产生一个更加强大的对同一病原第2次攻击的防御反应(Kinoshita and Seki, 2014).这种反应往往具有快速、有效和增强性等特点.植物在非生物胁迫响应方面也表现出同样的记忆反应, 并且这些由胁迫诱导的表观遗传改变可以通过有丝分裂或减数分裂稳定地遗传, 从而产生当代或跨代胁迫记忆.例如, Feng等(2012)发现盐碱胁迫诱导的水稻DNA甲基化的改变能通过自交遗传给下一代.近年来许多研究表明, 这种胁迫记忆参与了植物响应干旱、低温、盐、ABA以及SA等各种非生物胁迫的应答过程(Crisp et al., 2016).有研究表明, 植物产生记忆现象可能与关键信号途径代谢产物或转录因子的持续改变有关(Kinoshita and Seki, 2014; Vriet et al., 2015); 另外还可能通过改变染色质的状态(如组蛋白修饰、DNA甲基化以及RNA聚合酶滞留等)实现(Eichten et al., 2014; Avramova, 2015).但也有研究者认为, 表观遗传记忆其实是一种相对罕见的事件, 其关键的调控步骤是胁迫恢复期间表观遗传记忆是否形成, 并提出RNA代谢、转录后基因沉默以及RNA介导的DNA甲基化途径在其中发挥了关键作用(Crisp et al., 2016).目前, 表观遗传记忆的相关研究还非常少, 但其在植物抗逆驯化以及获得性抗性等方面无疑具有非常重要的理论意义和应用前景. ...
1 2012
... Histone modification related genes involved in abiotic stresses in plants
... 组蛋白的甲基化和去甲基化通常发生在精氨酸和赖氨酸残基上, 赖氨酸残基的甲基化主要由含有SET结构域的赖氨酸甲基转移酶HKMTs催化形成(Pontvianne et al., 2010).SET结构域蛋白可分为TrxG (Trithorax Group)、E(Z) (Enhancer of Zeste)、SU (VAR)以及ASH1 (Absent, Small, or Homeotic dis- cs) 4个家族, 分别负责组蛋白H3K4、H3K9、H3K27和H3K36的甲基化修饰.在拟南芥中, TrxG家族蛋白SDG25 (Berr et al., 2009)、E(Z)家族蛋白CLF (Chanvivattana et al., 2004)以及ASH1家族蛋白SDG8和SDG26 (Xu et al., 2008)都参与开花时间的调控.组蛋白去甲基化酶HDMs包括KDM1/LSD1家族和JmjC结构域蛋白家族两类(Liu et al., 2010).这两类蛋白质都可通过氧化反应直接去除组蛋白上的甲基化修饰.KDM1/LSD1以黄素为辅助因子, 只能对一、二甲基化修饰进行去甲基, 并不能对三甲基化去甲基; JmjC结构域蛋白需要二价铁和α酮戊二酸作辅助因子, 可以对一、二、三甲基化修饰去甲基.截至目前, 拟南芥中报道了4个KDM1/LSD1家族以及21个JMJ家族组蛋白去甲基化酶, 它们主要参与拟南芥的开花调控(Saze et al., 2008; Liu et al., 2010).组蛋白的乙酰化、磷酸化和泛素化修饰一般与基因转录激活有关, SUMO化和生物素化与转录抑制相关.已有的研究显示, 与胁迫相关的基因激活主要由H3K4、H3K36甲基化以及H3K9乙酰化调控, 而H3K9、H3K27甲基化和H3去乙酰化主要参与相关基因的沉默过程(Qiao and Fan, 2011). ...
1 2003
... 组蛋白修饰是表观遗传调控研究领域的热点.核小体是染色质的基本组成单位, 由约146 bp的DNA缠绕各两分子的核心组蛋白H2A、H2B、H3和H4形成的八聚体构成(Luger et al., 1997).伸出核小体外的核心组蛋白的N端可被共价修饰, 这些修饰主要包括乙酰化、甲基化、磷酸化、泛素化、生物素化、ADP核糖基化以及类泛素化等(Strahl and Allis, 2000).核心组蛋白N端赖氨酸可被组蛋白乙酰化酶HATs (histone acetyltransferase)和组蛋白去乙酰化酶HDACs (histone deacetylation transferase)进行乙酰化和去乙酰化修饰.在拟南芥中, HATs分为4个不同的家族, 研究表明HATs在植物发育(Bertrand et al., 2003; Long et al., 2006)和胁迫应答(Pavangadkar et al., 2010)中具有重要作用.真核生物的HDACs包含RP- D3 (Reduced Potassium Deficiency 3)、SIR2 (Silent Information Regulator 2)和HD2 (TypeIIHDAC) 3个主要蛋白家族, 其中HD2是植物特有的HDACs (Pan- dey et al., 2002).在拟南芥中含有10个RPD3/HDA1去乙酰化酶家族蛋白(Alinsug et al., 2009), 其中HDA6能影响转基因的表达(Murfett et al., 2001)、DNA甲基化(Aufsatz et al., 2002)以及rRNA基因的调控(Probst et al., 2004).同时研究发现, HDA19能与WRKY转录因子结合, 从而调控植物的基本防御反应(Kim et al., 2008).另外, HDA6和HDA19在拟南芥种子萌发、盐胁迫以及ABA诱导的基因表达方面都发挥十分重要的作用(Chen et al., 2010). ...
1 2015
... DNA甲基化修饰在调节植物响应外界环境胁迫中发挥了非常重要的作用.最近几年的研究表明, 全基因组DNA甲基化修饰的改变与植物响应非生物胁迫密切相关(Boyko et al., 2010; Karan et al., 2012; Wang et al., 2014).许多与DNA甲基化修饰相关的基因参与了植物的非生物胁迫应答过程(表1).在植物响应高盐、干旱、温度和重金属等非生物胁迫过程中, 植物的特定基因或整个基因组水平的DNA甲基化修饰都会发生改变, 从而提高植物对不良环境的适应性, 确保植物在逆境条件下的生长发育.通常情况下, 非生物胁迫诱导的DNA甲基化修饰的改变发生在整个基因组水平.例如, Wang等(2011)以水稻(Oryza sativa) DK151和IR64为研究材料, 发现由干旱胁迫诱导的DNA甲基化位点的改变占全基因组DNA甲基化位点的12%左右, 这些甲基化的变化有29%在胁迫解除时仍然被保留; 并且干旱诱导的DNA甲基化还表现出组织及发育时期的特异性.Ferreira等(2015)在研究盐胁迫下DNA甲基化水平的变化时, 发现盐胁迫耐受型水稻材料Pokkali和敏感型水稻材料IR29基因组全局性的DNA甲基化水平在盐胁迫下发生了明显改变, 并且这种DNA甲基化水平的改变具有基因型及组织特异性.此外, 非生物胁迫下, DNA甲基化水平的改变往往还会伴随其它调节机制.例如, Karan等(2012)在研究水稻响应盐胁迫时, 发现逆转录转座子、非生物胁迫应答基因以及染色质修饰因子在盐胁迫下都具有不同的表达模式, 同时鉴定了不同水稻品种盐胁迫诱导的DNA甲基化模式.而对于水稻而言, 在水稻根细胞内相关基因的去甲基化修饰对其响应盐胁迫也具有重要作用(Wang et al., 2011).Zhu等(2015)在研究水稻R2R3类型的MYB转录因子OsMYB91时, 发现OsMYB91的表达受非生物胁迫诱导, 特别是盐胁迫.进一步研究发现, 该基因启动子区域的DNA甲基化水平改变是导致该基因在胁迫下表达发生变化的主要原因.除了拟南芥和水稻等常见模式植物, 人们也对其它植物响应胁迫时的DNA甲基化修饰进行了研究.例如, Bharti等(2015)在研究盐胁迫下, 类黄酮生物合成和抗氧化途径的酶基因是否受到表观遗传调控时, 发现这些基因启动子区域甲基化状态改变使得相关基因的表达水平升高, 从而导致过表达AtROS1的转基因烟草(Nicotiana tabacum)表现出对盐胁迫的耐受性.Lu等(2017)利用Me-DIP测序技术对棉花(Gossypium hirsutum)在盐胁迫下全基因组的DNA甲基化水平进行了研究, 为理解棉花盐胁迫反应的表观遗传修饰变化提供了有价值的信息.同样, 也有研究发现, 苜蓿(Medicago spp.)在盐胁迫下, 全基因组水平的DNA甲基化程度增加, 特别是在高盐胁迫下尤为明显(Al-Lawati et al., 2016).目前, 有关DNA甲基化修饰水平影响植物响应非生物胁迫的具体机制还有待深入研究. ...
1 2005
... 为了能够在胁迫条件下生存, 植物进化出复杂的机制来感知外部信号, 从而对环境变化做出最佳反应(图1).植物激素(如水杨酸(SA)、茉莉酸(JA)、乙烯(ET)和脱落酸(ABA)等内源性分子)在调节植物对生物以及非生物胁迫的抵御过程中扮演着重要角色(Bostock, 2005; Lorenzo and Solano, 2005; Mauch- Mani and Mauch, 2005).这些激素信号可通过MAP激酶级联反应调控转录因子(如MYC、MYB、NAC、ZF和HSF等)参与胁迫相关基因的表达调控(Fujita et al., 2006).同时, 有研究者认为活性氧(ROS)的产生是植物抵御生物胁迫与非生物胁迫共有的关键过程(Torres and Dangl, 2005).除转录因子的调控, 通过染色质结构变化调控基因转录也是植物响应胁迫的重要方式, 是表观遗传机制调控植物应答外界胁迫的主要体现. ...
1 2010
... DNA甲基化修饰在调节植物响应外界环境胁迫中发挥了非常重要的作用.最近几年的研究表明, 全基因组DNA甲基化修饰的改变与植物响应非生物胁迫密切相关(Boyko et al., 2010; Karan et al., 2012; Wang et al., 2014).许多与DNA甲基化修饰相关的基因参与了植物的非生物胁迫应答过程(表1).在植物响应高盐、干旱、温度和重金属等非生物胁迫过程中, 植物的特定基因或整个基因组水平的DNA甲基化修饰都会发生改变, 从而提高植物对不良环境的适应性, 确保植物在逆境条件下的生长发育.通常情况下, 非生物胁迫诱导的DNA甲基化修饰的改变发生在整个基因组水平.例如, Wang等(2011)以水稻(Oryza sativa) DK151和IR64为研究材料, 发现由干旱胁迫诱导的DNA甲基化位点的改变占全基因组DNA甲基化位点的12%左右, 这些甲基化的变化有29%在胁迫解除时仍然被保留; 并且干旱诱导的DNA甲基化还表现出组织及发育时期的特异性.Ferreira等(2015)在研究盐胁迫下DNA甲基化水平的变化时, 发现盐胁迫耐受型水稻材料Pokkali和敏感型水稻材料IR29基因组全局性的DNA甲基化水平在盐胁迫下发生了明显改变, 并且这种DNA甲基化水平的改变具有基因型及组织特异性.此外, 非生物胁迫下, DNA甲基化水平的改变往往还会伴随其它调节机制.例如, Karan等(2012)在研究水稻响应盐胁迫时, 发现逆转录转座子、非生物胁迫应答基因以及染色质修饰因子在盐胁迫下都具有不同的表达模式, 同时鉴定了不同水稻品种盐胁迫诱导的DNA甲基化模式.而对于水稻而言, 在水稻根细胞内相关基因的去甲基化修饰对其响应盐胁迫也具有重要作用(Wang et al., 2011).Zhu等(2015)在研究水稻R2R3类型的MYB转录因子OsMYB91时, 发现OsMYB91的表达受非生物胁迫诱导, 特别是盐胁迫.进一步研究发现, 该基因启动子区域的DNA甲基化水平改变是导致该基因在胁迫下表达发生变化的主要原因.除了拟南芥和水稻等常见模式植物, 人们也对其它植物响应胁迫时的DNA甲基化修饰进行了研究.例如, Bharti等(2015)在研究盐胁迫下, 类黄酮生物合成和抗氧化途径的酶基因是否受到表观遗传调控时, 发现这些基因启动子区域甲基化状态改变使得相关基因的表达水平升高, 从而导致过表达AtROS1的转基因烟草(Nicotiana tabacum)表现出对盐胁迫的耐受性.Lu等(2017)利用Me-DIP测序技术对棉花(Gossypium hirsutum)在盐胁迫下全基因组的DNA甲基化水平进行了研究, 为理解棉花盐胁迫反应的表观遗传修饰变化提供了有价值的信息.同样, 也有研究发现, 苜蓿(Medicago spp.)在盐胁迫下, 全基因组水平的DNA甲基化程度增加, 特别是在高盐胁迫下尤为明显(Al-Lawati et al., 2016).目前, 有关DNA甲基化修饰水平影响植物响应非生物胁迫的具体机制还有待深入研究. ...
1 2008
... 在自然状态下, 植物的整个生长周期均处于固着状态, 不像动物能够主动自发地躲避不良环境, 因此必然会受到来自外界生物与非生物胁迫的影响.正是由于植物处于这样一个极易受到环境胁迫的状态, 其自身进化出一系列自我保护和适应与抵制不良环境的机制(Boyko and Kovalchuk, 2008).对植物生长发育的胁迫包括内部或外部因素.内部胁迫主要为植物自身基因的突变或异常细胞对机体代谢产生的不利影响; 外部胁迫则可分为生物与非生物胁迫.生物胁迫包括病原感染、食草动物和种内竞争等; 非生物胁迫由不利的环境条件引起(如不适宜的温度、水分和营养物质的利用以及光照条件等), 这两种外部胁迫可以是永久性的也可以是临时状态(Madlung and Comai, 2004). ...
1 2004
... 组蛋白的甲基化和去甲基化通常发生在精氨酸和赖氨酸残基上, 赖氨酸残基的甲基化主要由含有SET结构域的赖氨酸甲基转移酶HKMTs催化形成(Pontvianne et al., 2010).SET结构域蛋白可分为TrxG (Trithorax Group)、E(Z) (Enhancer of Zeste)、SU (VAR)以及ASH1 (Absent, Small, or Homeotic dis- cs) 4个家族, 分别负责组蛋白H3K4、H3K9、H3K27和H3K36的甲基化修饰.在拟南芥中, TrxG家族蛋白SDG25 (Berr et al., 2009)、E(Z)家族蛋白CLF (Chanvivattana et al., 2004)以及ASH1家族蛋白SDG8和SDG26 (Xu et al., 2008)都参与开花时间的调控.组蛋白去甲基化酶HDMs包括KDM1/LSD1家族和JmjC结构域蛋白家族两类(Liu et al., 2010).这两类蛋白质都可通过氧化反应直接去除组蛋白上的甲基化修饰.KDM1/LSD1以黄素为辅助因子, 只能对一、二甲基化修饰进行去甲基, 并不能对三甲基化去甲基; JmjC结构域蛋白需要二价铁和α酮戊二酸作辅助因子, 可以对一、二、三甲基化修饰去甲基.截至目前, 拟南芥中报道了4个KDM1/LSD1家族以及21个JMJ家族组蛋白去甲基化酶, 它们主要参与拟南芥的开花调控(Saze et al., 2008; Liu et al., 2010).组蛋白的乙酰化、磷酸化和泛素化修饰一般与基因转录激活有关, SUMO化和生物素化与转录抑制相关.已有的研究显示, 与胁迫相关的基因激活主要由H3K4、H3K36甲基化以及H3K9乙酰化调控, 而H3K9、H3K27甲基化和H3去乙酰化主要参与相关基因的沉默过程(Qiao and Fan, 2011). ...
1 2010
... 组蛋白修饰是表观遗传调控研究领域的热点.核小体是染色质的基本组成单位, 由约146 bp的DNA缠绕各两分子的核心组蛋白H2A、H2B、H3和H4形成的八聚体构成(Luger et al., 1997).伸出核小体外的核心组蛋白的N端可被共价修饰, 这些修饰主要包括乙酰化、甲基化、磷酸化、泛素化、生物素化、ADP核糖基化以及类泛素化等(Strahl and Allis, 2000).核心组蛋白N端赖氨酸可被组蛋白乙酰化酶HATs (histone acetyltransferase)和组蛋白去乙酰化酶HDACs (histone deacetylation transferase)进行乙酰化和去乙酰化修饰.在拟南芥中, HATs分为4个不同的家族, 研究表明HATs在植物发育(Bertrand et al., 2003; Long et al., 2006)和胁迫应答(Pavangadkar et al., 2010)中具有重要作用.真核生物的HDACs包含RP- D3 (Reduced Potassium Deficiency 3)、SIR2 (Silent Information Regulator 2)和HD2 (TypeIIHDAC) 3个主要蛋白家族, 其中HD2是植物特有的HDACs (Pan- dey et al., 2002).在拟南芥中含有10个RPD3/HDA1去乙酰化酶家族蛋白(Alinsug et al., 2009), 其中HDA6能影响转基因的表达(Murfett et al., 2001)、DNA甲基化(Aufsatz et al., 2002)以及rRNA基因的调控(Probst et al., 2004).同时研究发现, HDA19能与WRKY转录因子结合, 从而调控植物的基本防御反应(Kim et al., 2008).另外, HDA6和HDA19在拟南芥种子萌发、盐胁迫以及ABA诱导的基因表达方面都发挥十分重要的作用(Chen et al., 2010). ...
1 2010
... 在植物中, 小RNA (sRNA)是不具有编码功能的、对基因表达有调控作用的一类RNA分子, 其主要通过调节mRNA的稳定性、沉默基因转录或参与DNA甲基化等过程参与植物的生长发育(Ramachandran and Chen, 2008).因此, 小RNA也是一种重要的表观遗传调控因子.随着研究的不断深入, 对植物中sRNA的认识也有了突破性进展.以水稻为例, 研究发现, 在水稻中存在上百种sRNA, 并且在不同的生长条件下, 不同水稻品种的不同组织中sRNA的表达水平明显不同(Chen and Wu, 2010; He et al., 2010; Zhang et al., 2014).目前, 在植物中研究较多的是siRNA (small interfering RNA)和miRNA (microRNA).si- RNA是在DCL (Dicer-like protein)酶作用下由长双链RNA前体加工而来, 目前发现参与基因转录和转录后调控的内源性siRNA有nat-siRNA、ta-siRNA和hc-siRNA等(Mallory and Vaucheret, 2006).miRNA是内源基因编码的一类能与靶mRNA互补配对的一种单链小RNA, 主要参与基因的转录后调控.大量研究表明, siRNA和miRNA在植物生长发育和非生物胁迫响应过程中发挥重要作用(Phillips et al., 2007; Sunkar et al., 2007). ...
1 2006
... Histone modification related genes involved in abiotic stresses in plants
... Abiotic stress responsive genes regulated by DNA-methylation in plants
基因
物种
胁迫类型
参考文献
Asr1, Asr2
番茄(Solanum lycopersicum)
干旱胁迫
González et al., 2011; 2013
NtGPDL
烟草(Nicotiana tabaccum cv. ‘Xanthi nc’)
低温胁迫
Choi and Sano, 2007
OsMYB91
水稻(Oryza sativa)
盐胁迫
Zhu et al., 2015
Glyma11g02400 Glyma16g27950 Glyma20g30840
大豆(Glycine max)
盐胁迫
Song et al., 2012
SPCH, FAMA
拟南芥(Arabidopsis thaliana)
湿度胁迫
Tricker et al., 2012
ZmMI1, Ac/Ds
玉米(Zea mays)
低温胁迫
Steward et al., 2002
Tam3
金鱼草(Antirrhinum majus)
低温胁迫
Hashida et al., 2006
NRPD2
拟南芥(A. thaliana)
高温胁迫
Popova et al., 2013
2 组蛋白修饰2.1 植物体内的组蛋白修饰 组蛋白修饰是表观遗传调控研究领域的热点.核小体是染色质的基本组成单位, 由约146 bp的DNA缠绕各两分子的核心组蛋白H2A、H2B、H3和H4形成的八聚体构成(Luger et al., 1997).伸出核小体外的核心组蛋白的N端可被共价修饰, 这些修饰主要包括乙酰化、甲基化、磷酸化、泛素化、生物素化、ADP核糖基化以及类泛素化等(Strahl and Allis, 2000).核心组蛋白N端赖氨酸可被组蛋白乙酰化酶HATs (histone acetyltransferase)和组蛋白去乙酰化酶HDACs (histone deacetylation transferase)进行乙酰化和去乙酰化修饰.在拟南芥中, HATs分为4个不同的家族, 研究表明HATs在植物发育(Bertrand et al., 2003; Long et al., 2006)和胁迫应答(Pavangadkar et al., 2010)中具有重要作用.真核生物的HDACs包含RP- D3 (Reduced Potassium Deficiency 3)、SIR2 (Silent Information Regulator 2)和HD2 (TypeIIHDAC) 3个主要蛋白家族, 其中HD2是植物特有的HDACs (Pan- dey et al., 2002).在拟南芥中含有10个RPD3/HDA1去乙酰化酶家族蛋白(Alinsug et al., 2009), 其中HDA6能影响转基因的表达(Murfett et al., 2001)、DNA甲基化(Aufsatz et al., 2002)以及rRNA基因的调控(Probst et al., 2004).同时研究发现, HDA19能与WRKY转录因子结合, 从而调控植物的基本防御反应(Kim et al., 2008).另外, HDA6和HDA19在拟南芥种子萌发、盐胁迫以及ABA诱导的基因表达方面都发挥十分重要的作用(Chen et al., 2010). ...
2 2016
... 通常情况下, 植物处于自然环境状态受到的非生物胁迫具有短暂性和循环性等特点.研究认为这种可预测的、循环的环境改变能够激发植物产生胁迫记忆.“防御启动”是目前人们最为熟知的一种胁迫记忆现象, 即植物会记忆病原体的第1次攻击, 从而产生一个更加强大的对同一病原第2次攻击的防御反应(Kinoshita and Seki, 2014).这种反应往往具有快速、有效和增强性等特点.植物在非生物胁迫响应方面也表现出同样的记忆反应, 并且这些由胁迫诱导的表观遗传改变可以通过有丝分裂或减数分裂稳定地遗传, 从而产生当代或跨代胁迫记忆.例如, Feng等(2012)发现盐碱胁迫诱导的水稻DNA甲基化的改变能通过自交遗传给下一代.近年来许多研究表明, 这种胁迫记忆参与了植物响应干旱、低温、盐、ABA以及SA等各种非生物胁迫的应答过程(Crisp et al., 2016).有研究表明, 植物产生记忆现象可能与关键信号途径代谢产物或转录因子的持续改变有关(Kinoshita and Seki, 2014; Vriet et al., 2015); 另外还可能通过改变染色质的状态(如组蛋白修饰、DNA甲基化以及RNA聚合酶滞留等)实现(Eichten et al., 2014; Avramova, 2015).但也有研究者认为, 表观遗传记忆其实是一种相对罕见的事件, 其关键的调控步骤是胁迫恢复期间表观遗传记忆是否形成, 并提出RNA代谢、转录后基因沉默以及RNA介导的DNA甲基化途径在其中发挥了关键作用(Crisp et al., 2016).目前, 表观遗传记忆的相关研究还非常少, 但其在植物抗逆驯化以及获得性抗性等方面无疑具有非常重要的理论意义和应用前景. ... ... ).但也有研究者认为, 表观遗传记忆其实是一种相对罕见的事件, 其关键的调控步骤是胁迫恢复期间表观遗传记忆是否形成, 并提出RNA代谢、转录后基因沉默以及RNA介导的DNA甲基化途径在其中发挥了关键作用(Crisp et al., 2016).目前, 表观遗传记忆的相关研究还非常少, 但其在植物抗逆驯化以及获得性抗性等方面无疑具有非常重要的理论意义和应用前景. ...
1 2014
... DNA甲基化是指以DNA为受体, 在DNA甲基转移酶的作用下, 将供体S-腺苷甲硫氨酸上的1个甲基转移至胞嘧啶的第5位碳原子上, 从而形成5-甲基胞嘧啶(m5C)的过程(Sahu et al., 2013).1925年, m5C首次在结核分枝杆菌的结核菌素的水解产物中被发现(Johnson and Coghill, 1925).随后, 在植物中发现了较高水平的m5C甲基化修饰(Vanyushin and Belozerskii, 1959).DNA胞嘧啶甲基化修饰主要包括不对称(mCpHpH)甲基化和对称(mCpG和mCpHpG)甲基化.例如, 在拟南芥(Arabidopsis thaliana)中, CG、CHG与CHH的甲基化水平分别为24%、6.7%和1.7%; 并且在同一植物的不同组织或同一组织的不同发育阶段, 其基因组DNA的甲基化位点和水平也不一样(Dhar et al., 2014). ...
2 2011
... Histone modification related genes involved in abiotic stresses in plants
... 通常情况下, 植物处于自然环境状态受到的非生物胁迫具有短暂性和循环性等特点.研究认为这种可预测的、循环的环境改变能够激发植物产生胁迫记忆.“防御启动”是目前人们最为熟知的一种胁迫记忆现象, 即植物会记忆病原体的第1次攻击, 从而产生一个更加强大的对同一病原第2次攻击的防御反应(Kinoshita and Seki, 2014).这种反应往往具有快速、有效和增强性等特点.植物在非生物胁迫响应方面也表现出同样的记忆反应, 并且这些由胁迫诱导的表观遗传改变可以通过有丝分裂或减数分裂稳定地遗传, 从而产生当代或跨代胁迫记忆.例如, Feng等(2012)发现盐碱胁迫诱导的水稻DNA甲基化的改变能通过自交遗传给下一代.近年来许多研究表明, 这种胁迫记忆参与了植物响应干旱、低温、盐、ABA以及SA等各种非生物胁迫的应答过程(Crisp et al., 2016).有研究表明, 植物产生记忆现象可能与关键信号途径代谢产物或转录因子的持续改变有关(Kinoshita and Seki, 2014; Vriet et al., 2015); 另外还可能通过改变染色质的状态(如组蛋白修饰、DNA甲基化以及RNA聚合酶滞留等)实现(Eichten et al., 2014; Avramova, 2015).但也有研究者认为, 表观遗传记忆其实是一种相对罕见的事件, 其关键的调控步骤是胁迫恢复期间表观遗传记忆是否形成, 并提出RNA代谢、转录后基因沉默以及RNA介导的DNA甲基化途径在其中发挥了关键作用(Crisp et al., 2016).目前, 表观遗传记忆的相关研究还非常少, 但其在植物抗逆驯化以及获得性抗性等方面无疑具有非常重要的理论意义和应用前景. ...
1 2015
... DNA甲基化修饰在调节植物响应外界环境胁迫中发挥了非常重要的作用.最近几年的研究表明, 全基因组DNA甲基化修饰的改变与植物响应非生物胁迫密切相关(Boyko et al., 2010; Karan et al., 2012; Wang et al., 2014).许多与DNA甲基化修饰相关的基因参与了植物的非生物胁迫应答过程(表1).在植物响应高盐、干旱、温度和重金属等非生物胁迫过程中, 植物的特定基因或整个基因组水平的DNA甲基化修饰都会发生改变, 从而提高植物对不良环境的适应性, 确保植物在逆境条件下的生长发育.通常情况下, 非生物胁迫诱导的DNA甲基化修饰的改变发生在整个基因组水平.例如, Wang等(2011)以水稻(Oryza sativa) DK151和IR64为研究材料, 发现由干旱胁迫诱导的DNA甲基化位点的改变占全基因组DNA甲基化位点的12%左右, 这些甲基化的变化有29%在胁迫解除时仍然被保留; 并且干旱诱导的DNA甲基化还表现出组织及发育时期的特异性.Ferreira等(2015)在研究盐胁迫下DNA甲基化水平的变化时, 发现盐胁迫耐受型水稻材料Pokkali和敏感型水稻材料IR29基因组全局性的DNA甲基化水平在盐胁迫下发生了明显改变, 并且这种DNA甲基化水平的改变具有基因型及组织特异性.此外, 非生物胁迫下, DNA甲基化水平的改变往往还会伴随其它调节机制.例如, Karan等(2012)在研究水稻响应盐胁迫时, 发现逆转录转座子、非生物胁迫应答基因以及染色质修饰因子在盐胁迫下都具有不同的表达模式, 同时鉴定了不同水稻品种盐胁迫诱导的DNA甲基化模式.而对于水稻而言, 在水稻根细胞内相关基因的去甲基化修饰对其响应盐胁迫也具有重要作用(Wang et al., 2011).Zhu等(2015)在研究水稻R2R3类型的MYB转录因子OsMYB91时, 发现OsMYB91的表达受非生物胁迫诱导, 特别是盐胁迫.进一步研究发现, 该基因启动子区域的DNA甲基化水平改变是导致该基因在胁迫下表达发生变化的主要原因.除了拟南芥和水稻等常见模式植物, 人们也对其它植物响应胁迫时的DNA甲基化修饰进行了研究.例如, Bharti等(2015)在研究盐胁迫下, 类黄酮生物合成和抗氧化途径的酶基因是否受到表观遗传调控时, 发现这些基因启动子区域甲基化状态改变使得相关基因的表达水平升高, 从而导致过表达AtROS1的转基因烟草(Nicotiana tabacum)表现出对盐胁迫的耐受性.Lu等(2017)利用Me-DIP测序技术对棉花(Gossypium hirsutum)在盐胁迫下全基因组的DNA甲基化水平进行了研究, 为理解棉花盐胁迫反应的表观遗传修饰变化提供了有价值的信息.同样, 也有研究发现, 苜蓿(Medicago spp.)在盐胁迫下, 全基因组水平的DNA甲基化程度增加, 特别是在高盐胁迫下尤为明显(Al-Lawati et al., 2016).目前, 有关DNA甲基化修饰水平影响植物响应非生物胁迫的具体机制还有待深入研究. ...
1 2014
... Chromatin remodeling related genes involved in abiotic stresses in plants
基因
物种
胁迫类型
参考文献
AtCHR12
拟南芥(Arabidopsis thaliana )
干旱, 高温胁迫
Mlynárová et al., 2007
BRM
拟南芥(A. thaliana )
ABA, 干旱胁迫
Han et al., 2012
AtCHR23
拟南芥(A. thaliana )
盐胁迫
Folta et al., 2014
ATL1
水稻(Oryza sativa)
碱胁迫
Guo et al., 2014
AtASF1A/B
拟南芥(A. thaliana )
热胁迫
Weng et al., 2014
AtNAP1
拟南芥(A. thaliana )
盐胁迫
Liu et al., 2009
不依赖于ATP的染色质组装蛋白(如拟南芥组蛋白分子伴侣ASF1)也参与非生物胁迫过程.在热激条件下, AtASF1A和AtASF1B缺失突变体Atas1fab表现出对热胁迫高度敏感.进一步研究发现, 突变体内的HSPs和HsfA2等与热胁迫相关的基因表达受到抑制, 同时AtASF1A/B缺失也影响了拟南芥体内的H3K56- ac水平(Weng et al., 2014).该研究进一步揭示了在植物响应非生物逆境的过程中, 多种表观遗传调控途径可以协同发挥作用.另外, 有研究表明, 核仁染色质与胁迫引起的DNA损伤有关(James et al., 2014).染色质多重构象变化的平衡取决于染色质与特异的核仁DNA结合蛋白、组蛋白修饰、组蛋白变体以及DNA甲基化等相关因子的相互作用.染色质重塑复合体成员众多, 功能复杂, 目前仅有很少成员的功能被报道, 相信随着对更多染色质重塑因子的研究, 其在植物响应非生物胁迫中的作用也将不断得到揭示. ...
1 2006
... 为了能够在胁迫条件下生存, 植物进化出复杂的机制来感知外部信号, 从而对环境变化做出最佳反应(图1).植物激素(如水杨酸(SA)、茉莉酸(JA)、乙烯(ET)和脱落酸(ABA)等内源性分子)在调节植物对生物以及非生物胁迫的抵御过程中扮演着重要角色(Bostock, 2005; Lorenzo and Solano, 2005; Mauch- Mani and Mauch, 2005).这些激素信号可通过MAP激酶级联反应调控转录因子(如MYC、MYB、NAC、ZF和HSF等)参与胁迫相关基因的表达调控(Fujita et al., 2006).同时, 有研究者认为活性氧(ROS)的产生是植物抵御生物胁迫与非生物胁迫共有的关键过程(Torres and Dangl, 2005).除转录因子的调控, 通过染色质结构变化调控基因转录也是植物响应胁迫的重要方式, 是表观遗传机制调控植物应答外界胁迫的主要体现. ...
1 2002
... ATP依赖的染色质重塑是染色质重塑复合体利用ATP水解释放的能量来改变染色质结构, 从而调控真核生物基因表达的一种重要的表观遗传调控机制(Racki and Narlikar, 2008).ATP依赖的染色质重塑复合物主要分为4个家族: SWI/SNF家族、ISWI家族、CHD家族和INO80家族.SWI/SNF是第1个被发现的ATP依赖的染色质重塑复合物(Sudarsanam and Winston, 2000).SWI/SNF复合物通过改变染色质结构在DNA修饰、重组以及基因表达中发挥重要作用(Geiman and Robertson, 2002).研究表明, SWI/ SNF复合物也参与了环境胁迫对植物生长发育的调控.例如, 在拟南芥中, 过表达SNF2/Brahma类型的染色质重塑蛋白AtCHR12, 该过表达植株在干旱和高温等胁迫下表现出花芽以及初生茎生长停滞现象(Mlynárová et al., 2007).同时, 有研究表明, SWI/SNF和CHD复合物都参与了ABA调控的拟南芥种子萌发及生长等过程(Perruc et al., 2007; Saez et al., 2008). ...
1 2002
... 植物基因组DNA甲基化的方式主要有2种, 一种是从头甲基化, 在DNA复制后的新生链中, DNA甲基化酶在没有甲基化修饰的位点上重新催化DNA甲基化.第2种是维持甲基化, 即在半甲基化的新生DNA双链中, 新合成的DNA链以旧模板链为范本, 完成另一条新合成的DNA单链的甲基化.由于其对称特性, 在DNA复制之后, CpG和CpHpG位点的甲基化比较容易被复制, 所以该位点的甲基化通常是通过维持机制实现.相比之下, 非对称的CpHpH位点的甲基化则需要在每一次的DNA复制后重新建立(Karlsson et al., 2011).研究发现, 3种位点的甲基化由3种特定的甲基化酶催化, 分别为MET1 (DNA methyltransfer- ase 1)、CMT3 (Chromomethylase 3)和DRM (Domains rearranged methylase).MET1是哺乳动物Dnmt1的同源基因, 主要负责CpG位点的甲基化, 在拟南芥met1突变体中, DNA全局性的甲基化程度明显下降, 导致拟南芥表现出晚花表型(Kankel et al., 2003); CMT3是植物特有的甲基化酶, 负责着丝粒附近的重复序列以及转座子的CpHpG位点甲基化(Lindroth et al., 2001).DRM特异性介导植物DNA上CHG与CHH位点的甲基化修饰.研究发现, DRM2负责的对称与非对称位点甲基化修饰主要通过RNA介导的DNA甲基化修饰实现, 即RdDM (RNA-directed DNA methylation)途径完成.植物基因组全局性DNA甲基化水平需要通过DNA甲基化和去甲基化的动态平衡进行调节, 然而DNA去甲基化的机制一直备受争议.人们普遍认为DNA去甲基化既存在主动去甲基化过程又存在被动去甲基化过程.被动去甲基化的发生主要是由于DNA复制后从头甲基化的过程被抑制, 或者亲本甲基化印迹不能够被维持(Kankel et al., 2003).而主动去甲基化的过程主要由一些DNA糖基化酶和AP裂解酶调控, 如DME (DEMETER)、DML2 (DEMETER LIKE 2)、DML3以及ROS1 (REPRES- SOR OF SILENCING 1)等(Gong et al., 2002; Kino- shita et al., 2004). ...
1 2011
... Abiotic stress responsive genes regulated by DNA-methylation in plants
基因
物种
胁迫类型
参考文献
Asr1, Asr2
番茄(Solanum lycopersicum)
干旱胁迫
González et al., 2011; 2013
NtGPDL
烟草(Nicotiana tabaccum cv. ‘Xanthi nc’)
低温胁迫
Choi and Sano, 2007
OsMYB91
水稻(Oryza sativa)
盐胁迫
Zhu et al., 2015
Glyma11g02400 Glyma16g27950 Glyma20g30840
大豆(Glycine max)
盐胁迫
Song et al., 2012
SPCH, FAMA
拟南芥(Arabidopsis thaliana)
湿度胁迫
Tricker et al., 2012
ZmMI1, Ac/Ds
玉米(Zea mays)
低温胁迫
Steward et al., 2002
Tam3
金鱼草(Antirrhinum majus)
低温胁迫
Hashida et al., 2006
NRPD2
拟南芥(A. thaliana)
高温胁迫
Popova et al., 2013
2 组蛋白修饰2.1 植物体内的组蛋白修饰 组蛋白修饰是表观遗传调控研究领域的热点.核小体是染色质的基本组成单位, 由约146 bp的DNA缠绕各两分子的核心组蛋白H2A、H2B、H3和H4形成的八聚体构成(Luger et al., 1997).伸出核小体外的核心组蛋白的N端可被共价修饰, 这些修饰主要包括乙酰化、甲基化、磷酸化、泛素化、生物素化、ADP核糖基化以及类泛素化等(Strahl and Allis, 2000).核心组蛋白N端赖氨酸可被组蛋白乙酰化酶HATs (histone acetyltransferase)和组蛋白去乙酰化酶HDACs (histone deacetylation transferase)进行乙酰化和去乙酰化修饰.在拟南芥中, HATs分为4个不同的家族, 研究表明HATs在植物发育(Bertrand et al., 2003; Long et al., 2006)和胁迫应答(Pavangadkar et al., 2010)中具有重要作用.真核生物的HDACs包含RP- D3 (Reduced Potassium Deficiency 3)、SIR2 (Silent Information Regulator 2)和HD2 (TypeIIHDAC) 3个主要蛋白家族, 其中HD2是植物特有的HDACs (Pan- dey et al., 2002).在拟南芥中含有10个RPD3/HDA1去乙酰化酶家族蛋白(Alinsug et al., 2009), 其中HDA6能影响转基因的表达(Murfett et al., 2001)、DNA甲基化(Aufsatz et al., 2002)以及rRNA基因的调控(Probst et al., 2004).同时研究发现, HDA19能与WRKY转录因子结合, 从而调控植物的基本防御反应(Kim et al., 2008).另外, HDA6和HDA19在拟南芥种子萌发、盐胁迫以及ABA诱导的基因表达方面都发挥十分重要的作用(Chen et al., 2010). ...
1 2013
... Abiotic stress responsive genes regulated by DNA-methylation in plants
基因
物种
胁迫类型
参考文献
Asr1, Asr2
番茄(Solanum lycopersicum)
干旱胁迫
González et al., 2011; 2013
NtGPDL
烟草(Nicotiana tabaccum cv. ‘Xanthi nc’)
低温胁迫
Choi and Sano, 2007
OsMYB91
水稻(Oryza sativa)
盐胁迫
Zhu et al., 2015
Glyma11g02400 Glyma16g27950 Glyma20g30840
大豆(Glycine max)
盐胁迫
Song et al., 2012
SPCH, FAMA
拟南芥(Arabidopsis thaliana)
湿度胁迫
Tricker et al., 2012
ZmMI1, Ac/Ds
玉米(Zea mays)
低温胁迫
Steward et al., 2002
Tam3
金鱼草(Antirrhinum majus)
低温胁迫
Hashida et al., 2006
NRPD2
拟南芥(A. thaliana)
高温胁迫
Popova et al., 2013
2 组蛋白修饰2.1 植物体内的组蛋白修饰 组蛋白修饰是表观遗传调控研究领域的热点.核小体是染色质的基本组成单位, 由约146 bp的DNA缠绕各两分子的核心组蛋白H2A、H2B、H3和H4形成的八聚体构成(Luger et al., 1997).伸出核小体外的核心组蛋白的N端可被共价修饰, 这些修饰主要包括乙酰化、甲基化、磷酸化、泛素化、生物素化、ADP核糖基化以及类泛素化等(Strahl and Allis, 2000).核心组蛋白N端赖氨酸可被组蛋白乙酰化酶HATs (histone acetyltransferase)和组蛋白去乙酰化酶HDACs (histone deacetylation transferase)进行乙酰化和去乙酰化修饰.在拟南芥中, HATs分为4个不同的家族, 研究表明HATs在植物发育(Bertrand et al., 2003; Long et al., 2006)和胁迫应答(Pavangadkar et al., 2010)中具有重要作用.真核生物的HDACs包含RP- D3 (Reduced Potassium Deficiency 3)、SIR2 (Silent Information Regulator 2)和HD2 (TypeIIHDAC) 3个主要蛋白家族, 其中HD2是植物特有的HDACs (Pan- dey et al., 2002).在拟南芥中含有10个RPD3/HDA1去乙酰化酶家族蛋白(Alinsug et al., 2009), 其中HDA6能影响转基因的表达(Murfett et al., 2001)、DNA甲基化(Aufsatz et al., 2002)以及rRNA基因的调控(Probst et al., 2004).同时研究发现, HDA19能与WRKY转录因子结合, 从而调控植物的基本防御反应(Kim et al., 2008).另外, HDA6和HDA19在拟南芥种子萌发、盐胁迫以及ABA诱导的基因表达方面都发挥十分重要的作用(Chen et al., 2010). ...
1 2013
... sRNA and sRNA-regulated genes involved in abiotic stresses in plants
基因
物种
胁迫类型
参考文献
SlAGO4A
番茄(Solanum lycopersicum)
干旱胁迫, 盐胁迫
Huang et al., 2016
19 different miRNAs
水稻(Oryza sativa)
干旱胁迫
Zhou et al., 2010
13 different miRNAs
野生二粒小麦(Triticum turgidum ssp. dicoccoides)
干旱胁迫
Kantar et al., 2011
CSD1, CSD2, CCS
拟南芥(Arabidopsis thaliana )
高温胁迫
Guan et al., 2013
NFYA5
拟南芥(A. thaliana )
干旱胁迫
Li et al., 2008
OsPCF5, OsPCF8
水稻(O. sativa)
低温胁迫
Yang et al., 2013
GmNFYA3
大豆(Glycine max )
干旱胁迫
Ni et al., 2013
Phvul.010g120700
菜豆(Phaseolus vulgaris)
干旱胁迫
Sosa-Valencia et al., 2017
SlNF-YA1/2/3, SlMRP1
番茄(S. lycopersicum)
干旱胁迫
Zhang et al., 2011a
BHLH23
鹰嘴豆(Cicer arietinum)
干旱胁迫
Hajyzadeh et al., 2015
4 染色质重塑与非生物胁迫 ATP依赖的染色质重塑是染色质重塑复合体利用ATP水解释放的能量来改变染色质结构, 从而调控真核生物基因表达的一种重要的表观遗传调控机制(Racki and Narlikar, 2008).ATP依赖的染色质重塑复合物主要分为4个家族: SWI/SNF家族、ISWI家族、CHD家族和INO80家族.SWI/SNF是第1个被发现的ATP依赖的染色质重塑复合物(Sudarsanam and Winston, 2000).SWI/SNF复合物通过改变染色质结构在DNA修饰、重组以及基因表达中发挥重要作用(Geiman and Robertson, 2002).研究表明, SWI/ SNF复合物也参与了环境胁迫对植物生长发育的调控.例如, 在拟南芥中, 过表达SNF2/Brahma类型的染色质重塑蛋白AtCHR12, 该过表达植株在干旱和高温等胁迫下表现出花芽以及初生茎生长停滞现象(Mlynárová et al., 2007).同时, 有研究表明, SWI/SNF和CHD复合物都参与了ABA调控的拟南芥种子萌发及生长等过程(Perruc et al., 2007; Saez et al., 2008). ...
1 2014
... Chromatin remodeling related genes involved in abiotic stresses in plants
基因
物种
胁迫类型
参考文献
AtCHR12
拟南芥(Arabidopsis thaliana )
干旱, 高温胁迫
Mlynárová et al., 2007
BRM
拟南芥(A. thaliana )
ABA, 干旱胁迫
Han et al., 2012
AtCHR23
拟南芥(A. thaliana )
盐胁迫
Folta et al., 2014
ATL1
水稻(Oryza sativa)
碱胁迫
Guo et al., 2014
AtASF1A/B
拟南芥(A. thaliana )
热胁迫
Weng et al., 2014
AtNAP1
拟南芥(A. thaliana )
盐胁迫
Liu et al., 2009
不依赖于ATP的染色质组装蛋白(如拟南芥组蛋白分子伴侣ASF1)也参与非生物胁迫过程.在热激条件下, AtASF1A和AtASF1B缺失突变体Atas1fab表现出对热胁迫高度敏感.进一步研究发现, 突变体内的HSPs和HsfA2等与热胁迫相关的基因表达受到抑制, 同时AtASF1A/B缺失也影响了拟南芥体内的H3K56- ac水平(Weng et al., 2014).该研究进一步揭示了在植物响应非生物逆境的过程中, 多种表观遗传调控途径可以协同发挥作用.另外, 有研究表明, 核仁染色质与胁迫引起的DNA损伤有关(James et al., 2014).染色质多重构象变化的平衡取决于染色质与特异的核仁DNA结合蛋白、组蛋白修饰、组蛋白变体以及DNA甲基化等相关因子的相互作用.染色质重塑复合体成员众多, 功能复杂, 目前仅有很少成员的功能被报道, 相信随着对更多染色质重塑因子的研究, 其在植物响应非生物胁迫中的作用也将不断得到揭示. ...
1 2015
... sRNA and sRNA-regulated genes involved in abiotic stresses in plants
基因
物种
胁迫类型
参考文献
SlAGO4A
番茄(Solanum lycopersicum)
干旱胁迫, 盐胁迫
Huang et al., 2016
19 different miRNAs
水稻(Oryza sativa)
干旱胁迫
Zhou et al., 2010
13 different miRNAs
野生二粒小麦(Triticum turgidum ssp. dicoccoides)
干旱胁迫
Kantar et al., 2011
CSD1, CSD2, CCS
拟南芥(Arabidopsis thaliana )
高温胁迫
Guan et al., 2013
NFYA5
拟南芥(A. thaliana )
干旱胁迫
Li et al., 2008
OsPCF5, OsPCF8
水稻(O. sativa)
低温胁迫
Yang et al., 2013
GmNFYA3
大豆(Glycine max )
干旱胁迫
Ni et al., 2013
Phvul.010g120700
菜豆(Phaseolus vulgaris)
干旱胁迫
Sosa-Valencia et al., 2017
SlNF-YA1/2/3, SlMRP1
番茄(S. lycopersicum)
干旱胁迫
Zhang et al., 2011a
BHLH23
鹰嘴豆(Cicer arietinum)
干旱胁迫
Hajyzadeh et al., 2015
4 染色质重塑与非生物胁迫 ATP依赖的染色质重塑是染色质重塑复合体利用ATP水解释放的能量来改变染色质结构, 从而调控真核生物基因表达的一种重要的表观遗传调控机制(Racki and Narlikar, 2008).ATP依赖的染色质重塑复合物主要分为4个家族: SWI/SNF家族、ISWI家族、CHD家族和INO80家族.SWI/SNF是第1个被发现的ATP依赖的染色质重塑复合物(Sudarsanam and Winston, 2000).SWI/SNF复合物通过改变染色质结构在DNA修饰、重组以及基因表达中发挥重要作用(Geiman and Robertson, 2002).研究表明, SWI/ SNF复合物也参与了环境胁迫对植物生长发育的调控.例如, 在拟南芥中, 过表达SNF2/Brahma类型的染色质重塑蛋白AtCHR12, 该过表达植株在干旱和高温等胁迫下表现出花芽以及初生茎生长停滞现象(Mlynárová et al., 2007).同时, 有研究表明, SWI/SNF和CHD复合物都参与了ABA调控的拟南芥种子萌发及生长等过程(Perruc et al., 2007; Saez et al., 2008). ...
2 2012
... 染色质重塑复合物在调节非生物胁迫相关基因的表达中起重要作用(表4).对拟南芥的研究表明, SWI3B能与ABA通路重要调控因子HAB1结合, 从而参与ABA调节的植物对非生物胁迫的适应过程.HAB1是ABA信号通路中起关键作用的负调控因子, 拟南芥swi3b突变体表现出种子萌发对ABA的弱敏感性, ABA应答基因RAB18和RD29B的表达下降, 并且发现ABA能降低HAB1在RAB18和RD2B基因启动子上的富集(Saez et al., 2008; Yuan et al., 2013).在拟南芥中, 植物对ABA的响应大多通过转录因子ABI5调节.Han等(2012)发现, SWI2/SNF2家族成员BRM的功能缺失导致拟南芥对ABA超敏感, BRM能与ABI5基因结合, BRM的缺失使得核小体不稳定, 进而使ABI5的转录受到抑制, 最终使得brm突变体表现出对干旱的耐受性.而ABA信号转导途径的关键组分SnRK2和PP2CA分别能够介导BRM的磷酸化和去磷酸化, 从而快速调控BRM的活性(Peirats-Llobet et al., 2015).此外, 研究还发现, AtMMS21介导的泛素化能够提高拟南芥根发育过程中BRM的稳定性(Zhang et al., 2017).另外, 最近研究发现, 一种植物特异的H3K27去甲基化酶REF6 (RELATIVE OF EARLY FLOWERING 6)能够结合在带有CTCTGYTY基序的基因组区域, 催化H3K27me3去甲基化并促进BRM结合在该区段, 该研究揭示了组蛋白修饰与染色质重塑的协同作用(Li et al., 2016). ... ... Chromatin remodeling related genes involved in abiotic stresses in plants
基因
物种
胁迫类型
参考文献
AtCHR12
拟南芥(Arabidopsis thaliana )
干旱, 高温胁迫
Mlynárová et al., 2007
BRM
拟南芥(A. thaliana )
ABA, 干旱胁迫
Han et al., 2012
AtCHR23
拟南芥(A. thaliana )
盐胁迫
Folta et al., 2014
ATL1
水稻(Oryza sativa)
碱胁迫
Guo et al., 2014
AtASF1A/B
拟南芥(A. thaliana )
热胁迫
Weng et al., 2014
AtNAP1
拟南芥(A. thaliana )
盐胁迫
Liu et al., 2009
不依赖于ATP的染色质组装蛋白(如拟南芥组蛋白分子伴侣ASF1)也参与非生物胁迫过程.在热激条件下, AtASF1A和AtASF1B缺失突变体Atas1fab表现出对热胁迫高度敏感.进一步研究发现, 突变体内的HSPs和HsfA2等与热胁迫相关的基因表达受到抑制, 同时AtASF1A/B缺失也影响了拟南芥体内的H3K56- ac水平(Weng et al., 2014).该研究进一步揭示了在植物响应非生物逆境的过程中, 多种表观遗传调控途径可以协同发挥作用.另外, 有研究表明, 核仁染色质与胁迫引起的DNA损伤有关(James et al., 2014).染色质多重构象变化的平衡取决于染色质与特异的核仁DNA结合蛋白、组蛋白修饰、组蛋白变体以及DNA甲基化等相关因子的相互作用.染色质重塑复合体成员众多, 功能复杂, 目前仅有很少成员的功能被报道, 相信随着对更多染色质重塑因子的研究, 其在植物响应非生物胁迫中的作用也将不断得到揭示. ...
1 2006
... Abiotic stress responsive genes regulated by DNA-methylation in plants
基因
物种
胁迫类型
参考文献
Asr1, Asr2
番茄(Solanum lycopersicum)
干旱胁迫
González et al., 2011; 2013
NtGPDL
烟草(Nicotiana tabaccum cv. ‘Xanthi nc’)
低温胁迫
Choi and Sano, 2007
OsMYB91
水稻(Oryza sativa)
盐胁迫
Zhu et al., 2015
Glyma11g02400 Glyma16g27950 Glyma20g30840
大豆(Glycine max)
盐胁迫
Song et al., 2012
SPCH, FAMA
拟南芥(Arabidopsis thaliana)
湿度胁迫
Tricker et al., 2012
ZmMI1, Ac/Ds
玉米(Zea mays)
低温胁迫
Steward et al., 2002
Tam3
金鱼草(Antirrhinum majus)
低温胁迫
Hashida et al., 2006
NRPD2
拟南芥(A. thaliana)
高温胁迫
Popova et al., 2013
2 组蛋白修饰2.1 植物体内的组蛋白修饰 组蛋白修饰是表观遗传调控研究领域的热点.核小体是染色质的基本组成单位, 由约146 bp的DNA缠绕各两分子的核心组蛋白H2A、H2B、H3和H4形成的八聚体构成(Luger et al., 1997).伸出核小体外的核心组蛋白的N端可被共价修饰, 这些修饰主要包括乙酰化、甲基化、磷酸化、泛素化、生物素化、ADP核糖基化以及类泛素化等(Strahl and Allis, 2000).核心组蛋白N端赖氨酸可被组蛋白乙酰化酶HATs (histone acetyltransferase)和组蛋白去乙酰化酶HDACs (histone deacetylation transferase)进行乙酰化和去乙酰化修饰.在拟南芥中, HATs分为4个不同的家族, 研究表明HATs在植物发育(Bertrand et al., 2003; Long et al., 2006)和胁迫应答(Pavangadkar et al., 2010)中具有重要作用.真核生物的HDACs包含RP- D3 (Reduced Potassium Deficiency 3)、SIR2 (Silent Information Regulator 2)和HD2 (TypeIIHDAC) 3个主要蛋白家族, 其中HD2是植物特有的HDACs (Pan- dey et al., 2002).在拟南芥中含有10个RPD3/HDA1去乙酰化酶家族蛋白(Alinsug et al., 2009), 其中HDA6能影响转基因的表达(Murfett et al., 2001)、DNA甲基化(Aufsatz et al., 2002)以及rRNA基因的调控(Probst et al., 2004).同时研究发现, HDA19能与WRKY转录因子结合, 从而调控植物的基本防御反应(Kim et al., 2008).另外, HDA6和HDA19在拟南芥种子萌发、盐胁迫以及ABA诱导的基因表达方面都发挥十分重要的作用(Chen et al., 2010). ...
1 2010
... 在植物中, 小RNA (sRNA)是不具有编码功能的、对基因表达有调控作用的一类RNA分子, 其主要通过调节mRNA的稳定性、沉默基因转录或参与DNA甲基化等过程参与植物的生长发育(Ramachandran and Chen, 2008).因此, 小RNA也是一种重要的表观遗传调控因子.随着研究的不断深入, 对植物中sRNA的认识也有了突破性进展.以水稻为例, 研究发现, 在水稻中存在上百种sRNA, 并且在不同的生长条件下, 不同水稻品种的不同组织中sRNA的表达水平明显不同(Chen and Wu, 2010; He et al., 2010; Zhang et al., 2014).目前, 在植物中研究较多的是siRNA (small interfering RNA)和miRNA (microRNA).si- RNA是在DCL (Dicer-like protein)酶作用下由长双链RNA前体加工而来, 目前发现参与基因转录和转录后调控的内源性siRNA有nat-siRNA、ta-siRNA和hc-siRNA等(Mallory and Vaucheret, 2006).miRNA是内源基因编码的一类能与靶mRNA互补配对的一种单链小RNA, 主要参与基因的转录后调控.大量研究表明, siRNA和miRNA在植物生长发育和非生物胁迫响应过程中发挥重要作用(Phillips et al., 2007; Sunkar et al., 2007). ...
2 2015
... 除DNA甲基化修饰外, 组蛋白修饰也是植物响应非生物胁迫的重要表观调控途径之一.近年来, 有许多关于组蛋白甲基化、乙酰化和磷酸化等修饰参与植物非生物胁迫应答过程的报道(表2).在模式植物拟南芥中, SKB1可以改变胁迫相关基因的H4R3sme2水平, 其缺失突变会导致拟南芥产生对盐胁迫超敏感的表型(Zhang et al., 2011b).类似地, 拟南芥组蛋白乙酰转移酶GCN5通过影响HSFA3 (Heat Stress Transcription Factors 3)和UVH6 (UV-HYPERSENSI- TIVE 6)基因启动子区域的H3K9和H3K14的乙酰化水平来调节应答基因的转录.因此, 缺失GCN5的拟南芥突变体表现出对热胁迫的耐受性(Hu et al., 2015).在水稻中, 组蛋白乙酰转移酶基因OsHATs的转录水平和组蛋白H3和H4赖氨酸残基乙酰化水平的变化都参与了水稻的干旱胁迫调控(Fang et al., 2014).Luo等(2012)在研究植物特异的组蛋白去乙酰化酶HD2时, 发现ABA和NaCl能抑制HD2A、HD2B、HD2C以及HD2D等基因的表达水平, HD2的T-DNA插入突变体表现出对ABA和NaCl的敏感性, 同时降低了对盐胁迫的耐受性.此外, 在组蛋白去乙酰化方面, Zheng等(2016)发现拟南芥组蛋白去乙酰化酶HDA9在调节拟南芥响应盐胁迫和干旱胁迫时发挥重要作用.由此可见, 组蛋白的多种修饰类型都参与了非生物胁迫的调节过程. ... ... Histone modification related genes involved in abiotic stresses in plants
... 组蛋白修饰是表观遗传调控研究领域的热点.核小体是染色质的基本组成单位, 由约146 bp的DNA缠绕各两分子的核心组蛋白H2A、H2B、H3和H4形成的八聚体构成(Luger et al., 1997).伸出核小体外的核心组蛋白的N端可被共价修饰, 这些修饰主要包括乙酰化、甲基化、磷酸化、泛素化、生物素化、ADP核糖基化以及类泛素化等(Strahl and Allis, 2000).核心组蛋白N端赖氨酸可被组蛋白乙酰化酶HATs (histone acetyltransferase)和组蛋白去乙酰化酶HDACs (histone deacetylation transferase)进行乙酰化和去乙酰化修饰.在拟南芥中, HATs分为4个不同的家族, 研究表明HATs在植物发育(Bertrand et al., 2003; Long et al., 2006)和胁迫应答(Pavangadkar et al., 2010)中具有重要作用.真核生物的HDACs包含RP- D3 (Reduced Potassium Deficiency 3)、SIR2 (Silent Information Regulator 2)和HD2 (TypeIIHDAC) 3个主要蛋白家族, 其中HD2是植物特有的HDACs (Pan- dey et al., 2002).在拟南芥中含有10个RPD3/HDA1去乙酰化酶家族蛋白(Alinsug et al., 2009), 其中HDA6能影响转基因的表达(Murfett et al., 2001)、DNA甲基化(Aufsatz et al., 2002)以及rRNA基因的调控(Probst et al., 2004).同时研究发现, HDA19能与WRKY转录因子结合, 从而调控植物的基本防御反应(Kim et al., 2008).另外, HDA6和HDA19在拟南芥种子萌发、盐胁迫以及ABA诱导的基因表达方面都发挥十分重要的作用(Chen et al., 2010). ...
1 2004
... 植物基因组DNA甲基化的方式主要有2种, 一种是从头甲基化, 在DNA复制后的新生链中, DNA甲基化酶在没有甲基化修饰的位点上重新催化DNA甲基化.第2种是维持甲基化, 即在半甲基化的新生DNA双链中, 新合成的DNA链以旧模板链为范本, 完成另一条新合成的DNA单链的甲基化.由于其对称特性, 在DNA复制之后, CpG和CpHpG位点的甲基化比较容易被复制, 所以该位点的甲基化通常是通过维持机制实现.相比之下, 非对称的CpHpH位点的甲基化则需要在每一次的DNA复制后重新建立(Karlsson et al., 2011).研究发现, 3种位点的甲基化由3种特定的甲基化酶催化, 分别为MET1 (DNA methyltransfer- ase 1)、CMT3 (Chromomethylase 3)和DRM (Domains rearranged methylase).MET1是哺乳动物Dnmt1的同源基因, 主要负责CpG位点的甲基化, 在拟南芥met1突变体中, DNA全局性的甲基化程度明显下降, 导致拟南芥表现出晚花表型(Kankel et al., 2003); CMT3是植物特有的甲基化酶, 负责着丝粒附近的重复序列以及转座子的CpHpG位点甲基化(Lindroth et al., 2001).DRM特异性介导植物DNA上CHG与CHH位点的甲基化修饰.研究发现, DRM2负责的对称与非对称位点甲基化修饰主要通过RNA介导的DNA甲基化修饰实现, 即RdDM (RNA-directed DNA methylation)途径完成.植物基因组全局性DNA甲基化水平需要通过DNA甲基化和去甲基化的动态平衡进行调节, 然而DNA去甲基化的机制一直备受争议.人们普遍认为DNA去甲基化既存在主动去甲基化过程又存在被动去甲基化过程.被动去甲基化的发生主要是由于DNA复制后从头甲基化的过程被抑制, 或者亲本甲基化印迹不能够被维持(Kankel et al., 2003).而主动去甲基化的过程主要由一些DNA糖基化酶和AP裂解酶调控, 如DME (DEMETER)、DML2 (DEMETER LIKE 2)、DML3以及ROS1 (REPRES- SOR OF SILENCING 1)等(Gong et al., 2002; Kino- shita et al., 2004). ...
2 2014
... 通常情况下, 植物处于自然环境状态受到的非生物胁迫具有短暂性和循环性等特点.研究认为这种可预测的、循环的环境改变能够激发植物产生胁迫记忆.“防御启动”是目前人们最为熟知的一种胁迫记忆现象, 即植物会记忆病原体的第1次攻击, 从而产生一个更加强大的对同一病原第2次攻击的防御反应(Kinoshita and Seki, 2014).这种反应往往具有快速、有效和增强性等特点.植物在非生物胁迫响应方面也表现出同样的记忆反应, 并且这些由胁迫诱导的表观遗传改变可以通过有丝分裂或减数分裂稳定地遗传, 从而产生当代或跨代胁迫记忆.例如, Feng等(2012)发现盐碱胁迫诱导的水稻DNA甲基化的改变能通过自交遗传给下一代.近年来许多研究表明, 这种胁迫记忆参与了植物响应干旱、低温、盐、ABA以及SA等各种非生物胁迫的应答过程(Crisp et al., 2016).有研究表明, 植物产生记忆现象可能与关键信号途径代谢产物或转录因子的持续改变有关(Kinoshita and Seki, 2014; Vriet et al., 2015); 另外还可能通过改变染色质的状态(如组蛋白修饰、DNA甲基化以及RNA聚合酶滞留等)实现(Eichten et al., 2014; Avramova, 2015).但也有研究者认为, 表观遗传记忆其实是一种相对罕见的事件, 其关键的调控步骤是胁迫恢复期间表观遗传记忆是否形成, 并提出RNA代谢、转录后基因沉默以及RNA介导的DNA甲基化途径在其中发挥了关键作用(Crisp et al., 2016).目前, 表观遗传记忆的相关研究还非常少, 但其在植物抗逆驯化以及获得性抗性等方面无疑具有非常重要的理论意义和应用前景. ... ... ).有研究表明, 植物产生记忆现象可能与关键信号途径代谢产物或转录因子的持续改变有关(Kinoshita and Seki, 2014; Vriet et al., 2015); 另外还可能通过改变染色质的状态(如组蛋白修饰、DNA甲基化以及RNA聚合酶滞留等)实现(Eichten et al., 2014; Avramova, 2015).但也有研究者认为, 表观遗传记忆其实是一种相对罕见的事件, 其关键的调控步骤是胁迫恢复期间表观遗传记忆是否形成, 并提出RNA代谢、转录后基因沉默以及RNA介导的DNA甲基化途径在其中发挥了关键作用(Crisp et al., 2016).目前, 表观遗传记忆的相关研究还非常少, 但其在植物抗逆驯化以及获得性抗性等方面无疑具有非常重要的理论意义和应用前景. ...
1 2016
... 染色质重塑复合物在调节非生物胁迫相关基因的表达中起重要作用(表4).对拟南芥的研究表明, SWI3B能与ABA通路重要调控因子HAB1结合, 从而参与ABA调节的植物对非生物胁迫的适应过程.HAB1是ABA信号通路中起关键作用的负调控因子, 拟南芥swi3b突变体表现出种子萌发对ABA的弱敏感性, ABA应答基因RAB18和RD29B的表达下降, 并且发现ABA能降低HAB1在RAB18和RD2B基因启动子上的富集(Saez et al., 2008; Yuan et al., 2013).在拟南芥中, 植物对ABA的响应大多通过转录因子ABI5调节.Han等(2012)发现, SWI2/SNF2家族成员BRM的功能缺失导致拟南芥对ABA超敏感, BRM能与ABI5基因结合, BRM的缺失使得核小体不稳定, 进而使ABI5的转录受到抑制, 最终使得brm突变体表现出对干旱的耐受性.而ABA信号转导途径的关键组分SnRK2和PP2CA分别能够介导BRM的磷酸化和去磷酸化, 从而快速调控BRM的活性(Peirats-Llobet et al., 2015).此外, 研究还发现, AtMMS21介导的泛素化能够提高拟南芥根发育过程中BRM的稳定性(Zhang et al., 2017).另外, 最近研究发现, 一种植物特异的H3K27去甲基化酶REF6 (RELATIVE OF EARLY FLOWERING 6)能够结合在带有CTCTGYTY基序的基因组区域, 催化H3K27me3去甲基化并促进BRM结合在该区段, 该研究揭示了组蛋白修饰与染色质重塑的协同作用(Li et al., 2016). ...
2 2008
... 迄今已有许多研究证明, 小RNA在调控植物响应非生物胁迫中扮演着十分重要的角色(表3).小RNA常通过RdDM (RNA-directed DNA methylation)途径影响DNA甲基化修饰, 进而响应非生物胁迫.Huang等(2016)发现, SlAGO4A作为RNA介导的DNA甲基化途径的重要因子参与了番茄(Solanum lycopersicum)应答干旱和高盐胁迫的过程.相比野生型, SlAGO4A下调的转基因番茄植株表现出对干旱和高盐的耐受性.同样, 在研究RdDM途径的过程中, Naydenov等(2015)发现RdDM途径中的2个植物特异的RNA聚合酶PolIV和PolV参与了高温胁迫下DRM2基因的表达调控, 并且在PolIV和PolV的缺失突变体中, ROS基因的表达也受到抑制.Tricker等(2012)在研究气孔发育时, 发现siRNA参与了拟南芥气孔发育的2个关键基因SPECHLESS和FAMA在低湿度胁迫下的DNA甲基化过程, 从而调节拟南芥对湿度改变的适应.通常情况下, 当植物处于不良环境时, 体内并非仅有某一特定miRNA的表达发生变化, 而是大量的miRNAs表达都发生改变.例如, 在水稻中, 干旱诱导下有mi- R170和miR171等11个sRNA表达下调, 有miR474和miR845等8个sRNA表达上调(Zhou et al., 2010); 野生二粒小麦(Triticum turgidum ssp. dicoccoides)在干旱胁迫下有13个miRNAs会受到不同程度的调控(Kantar et al., 2011).许多研究表明, miRNA相关基因的突变都会对植物的叶片发育产生影响.拟南芥中编码NFYA5转录因子的基因在干旱胁迫下能通过ABA依赖途径被miR169靶向调控, 敲除突变体nfya5和miR169的过表达植株都表现出叶片失水加快以及对干旱胁迫十分敏感等表型.基因芯片分析发现, NFYA5对许多应答干旱胁迫的基因具调控作用(Li et al., 2008).在水稻中, miRNA319基因家族有2个成员, 即Osa-MIR319a和Osa-MIR319b.其过表达植株相比野生型都具有较宽的叶片, 并且具有较强的低温耐受性.进一步研究发现, miR319的靶基因OsPCF5和OsPCF8的RNAi转基因植物也表现出同样的低温耐受性, 表明miR319具有调控植物适应低温胁迫的重要作用(Yang et al., 2013).综上, 目前的研究表明, 在非生物胁迫过程中, 很多小RNA的表达都会发生改变, 但是哪些小RNA才是真正参与胁迫应答调控的直接分子, 尚待进一步研究. ... ... sRNA and sRNA-regulated genes involved in abiotic stresses in plants
基因
物种
胁迫类型
参考文献
SlAGO4A
番茄(Solanum lycopersicum)
干旱胁迫, 盐胁迫
Huang et al., 2016
19 different miRNAs
水稻(Oryza sativa)
干旱胁迫
Zhou et al., 2010
13 different miRNAs
野生二粒小麦(Triticum turgidum ssp. dicoccoides)
干旱胁迫
Kantar et al., 2011
CSD1, CSD2, CCS
拟南芥(Arabidopsis thaliana )
高温胁迫
Guan et al., 2013
NFYA5
拟南芥(A. thaliana )
干旱胁迫
Li et al., 2008
OsPCF5, OsPCF8
水稻(O. sativa)
低温胁迫
Yang et al., 2013
GmNFYA3
大豆(Glycine max )
干旱胁迫
Ni et al., 2013
Phvul.010g120700
菜豆(Phaseolus vulgaris)
干旱胁迫
Sosa-Valencia et al., 2017
SlNF-YA1/2/3, SlMRP1
番茄(S. lycopersicum)
干旱胁迫
Zhang et al., 2011a
BHLH23
鹰嘴豆(Cicer arietinum)
干旱胁迫
Hajyzadeh et al., 2015
4 染色质重塑与非生物胁迫 ATP依赖的染色质重塑是染色质重塑复合体利用ATP水解释放的能量来改变染色质结构, 从而调控真核生物基因表达的一种重要的表观遗传调控机制(Racki and Narlikar, 2008).ATP依赖的染色质重塑复合物主要分为4个家族: SWI/SNF家族、ISWI家族、CHD家族和INO80家族.SWI/SNF是第1个被发现的ATP依赖的染色质重塑复合物(Sudarsanam and Winston, 2000).SWI/SNF复合物通过改变染色质结构在DNA修饰、重组以及基因表达中发挥重要作用(Geiman and Robertson, 2002).研究表明, SWI/ SNF复合物也参与了环境胁迫对植物生长发育的调控.例如, 在拟南芥中, 过表达SNF2/Brahma类型的染色质重塑蛋白AtCHR12, 该过表达植株在干旱和高温等胁迫下表现出花芽以及初生茎生长停滞现象(Mlynárová et al., 2007).同时, 有研究表明, SWI/SNF和CHD复合物都参与了ABA调控的拟南芥种子萌发及生长等过程(Perruc et al., 2007; Saez et al., 2008). ...
1 2001
... 植物基因组DNA甲基化的方式主要有2种, 一种是从头甲基化, 在DNA复制后的新生链中, DNA甲基化酶在没有甲基化修饰的位点上重新催化DNA甲基化.第2种是维持甲基化, 即在半甲基化的新生DNA双链中, 新合成的DNA链以旧模板链为范本, 完成另一条新合成的DNA单链的甲基化.由于其对称特性, 在DNA复制之后, CpG和CpHpG位点的甲基化比较容易被复制, 所以该位点的甲基化通常是通过维持机制实现.相比之下, 非对称的CpHpH位点的甲基化则需要在每一次的DNA复制后重新建立(Karlsson et al., 2011).研究发现, 3种位点的甲基化由3种特定的甲基化酶催化, 分别为MET1 (DNA methyltransfer- ase 1)、CMT3 (Chromomethylase 3)和DRM (Domains rearranged methylase).MET1是哺乳动物Dnmt1的同源基因, 主要负责CpG位点的甲基化, 在拟南芥met1突变体中, DNA全局性的甲基化程度明显下降, 导致拟南芥表现出晚花表型(Kankel et al., 2003); CMT3是植物特有的甲基化酶, 负责着丝粒附近的重复序列以及转座子的CpHpG位点甲基化(Lindroth et al., 2001).DRM特异性介导植物DNA上CHG与CHH位点的甲基化修饰.研究发现, DRM2负责的对称与非对称位点甲基化修饰主要通过RNA介导的DNA甲基化修饰实现, 即RdDM (RNA-directed DNA methylation)途径完成.植物基因组全局性DNA甲基化水平需要通过DNA甲基化和去甲基化的动态平衡进行调节, 然而DNA去甲基化的机制一直备受争议.人们普遍认为DNA去甲基化既存在主动去甲基化过程又存在被动去甲基化过程.被动去甲基化的发生主要是由于DNA复制后从头甲基化的过程被抑制, 或者亲本甲基化印迹不能够被维持(Kankel et al., 2003).而主动去甲基化的过程主要由一些DNA糖基化酶和AP裂解酶调控, 如DME (DEMETER)、DML2 (DEMETER LIKE 2)、DML3以及ROS1 (REPRES- SOR OF SILENCING 1)等(Gong et al., 2002; Kino- shita et al., 2004). ...
2 2010
... 组蛋白的甲基化和去甲基化通常发生在精氨酸和赖氨酸残基上, 赖氨酸残基的甲基化主要由含有SET结构域的赖氨酸甲基转移酶HKMTs催化形成(Pontvianne et al., 2010).SET结构域蛋白可分为TrxG (Trithorax Group)、E(Z) (Enhancer of Zeste)、SU (VAR)以及ASH1 (Absent, Small, or Homeotic dis- cs) 4个家族, 分别负责组蛋白H3K4、H3K9、H3K27和H3K36的甲基化修饰.在拟南芥中, TrxG家族蛋白SDG25 (Berr et al., 2009)、E(Z)家族蛋白CLF (Chanvivattana et al., 2004)以及ASH1家族蛋白SDG8和SDG26 (Xu et al., 2008)都参与开花时间的调控.组蛋白去甲基化酶HDMs包括KDM1/LSD1家族和JmjC结构域蛋白家族两类(Liu et al., 2010).这两类蛋白质都可通过氧化反应直接去除组蛋白上的甲基化修饰.KDM1/LSD1以黄素为辅助因子, 只能对一、二甲基化修饰进行去甲基, 并不能对三甲基化去甲基; JmjC结构域蛋白需要二价铁和α酮戊二酸作辅助因子, 可以对一、二、三甲基化修饰去甲基.截至目前, 拟南芥中报道了4个KDM1/LSD1家族以及21个JMJ家族组蛋白去甲基化酶, 它们主要参与拟南芥的开花调控(Saze et al., 2008; Liu et al., 2010).组蛋白的乙酰化、磷酸化和泛素化修饰一般与基因转录激活有关, SUMO化和生物素化与转录抑制相关.已有的研究显示, 与胁迫相关的基因激活主要由H3K4、H3K36甲基化以及H3K9乙酰化调控, 而H3K9、H3K27甲基化和H3去乙酰化主要参与相关基因的沉默过程(Qiao and Fan, 2011). ... ... ; Liu et al., 2010).组蛋白的乙酰化、磷酸化和泛素化修饰一般与基因转录激活有关, SUMO化和生物素化与转录抑制相关.已有的研究显示, 与胁迫相关的基因激活主要由H3K4、H3K36甲基化以及H3K9乙酰化调控, 而H3K9、H3K27甲基化和H3去乙酰化主要参与相关基因的沉默过程(Qiao and Fan, 2011). ...
2 2012
... Histone modification related genes involved in abiotic stresses in plants
基因
物种
胁迫类型
参考文献
HDACs
拟南芥(Arabidopsis thaliana )
盐胁迫
Luo et al., 2012
SKB1
拟南芥(A. thaliana )
盐胁迫
Zhang et al., 2011b
MSI1, HDA19
拟南芥(A. thaliana )
盐胁迫
Mehdi et al., 2016
HDA9
拟南芥(A. thaliana )
盐胁迫, 干旱胁迫
Zheng et al., 2016
HDA705
水稻(Oryza sativa)
盐胁迫
Zhao et al., 2016
GCN5
拟南芥(A. thaliana )
热胁迫
Hu et al., 2015
HDT701
水稻(O. sativa)
盐胁迫
Zhao et al., 2014
AtABO1
拟南芥(A. thaliana )
干旱胁迫
Chen et al., 2006
ADH1, PDC1
水稻(O. sativa)
淹水胁迫
Tsuji et al., 2006
OsHAG702, OsHAG704 OsHAC701, OsHAC704
水稻(O. sativa)
干旱胁迫
Liu et al., 2012
OsHAG703, OsHAM701 OsHAC703, OsHAF701
水稻(O. sativa)
干旱胁迫
Liu et al., 2012; Fang et al., 2014
AtATX1
拟南芥(A. thaliana )
干旱胁迫
Ding et al., 2011
OsDREB1bc
水稻(O. sativa)
盐胁迫, 低温胁迫
Roy et al., 2014
HvTX1, HvPKDM7
大麦(Hordeum vulgare)
干旱胁迫
Papaefthimiou and Tsaftaris, 2012
MYST, ELP3, GCN5
大麦(H. vulgare)
干旱胁迫
Papaefthimiou et al., 2010
AtHD2C
拟南芥(A. thaliana )
干旱胁迫
Sridha and Wu, 2006
AtMSI1, AtCHR12
拟南芥(A. thaliana )
干旱胁迫, 高温胁迫
Alexandre et al., 2009
AtBRM
拟南芥(A. thaliana )
干旱胁迫
Berr et al., 2012
通常情况下, 植物在响应非生物胁迫时, 体内的组蛋白修饰往往会与一些植物内源激素联系在一起.例如, Ding等(2011)发现在干旱胁迫下, 组蛋白甲基转移酶ATX1能够增强ABA合成途径的关键基因NCED3启动子区域的H3K4me3修饰, 从而促进该基因的表达.后续研究发现, 在干旱胁迫下, 拟南芥atx1突变体NCED3基因上的RNA聚合酶II和H3K4me3的富集程度明显下降, 并且与ABA相关的某些基因(如RD29A和RD29B)的转录水平也明显下降.类似研究发现, MSI1-HDA19复合物也参与了ABA介导的拟南芥应答盐胁迫过程(Mehdi et al., 2016).另外, 组蛋白修饰对胁迫的调控不仅局限于单一的某一类修饰, 而是多种修饰协同作用.例如, Wang等(2015)发现玉米(Zea mays)叶片在热胁迫下, 组蛋白修饰与叶片细胞的程序性死亡过程有密切联系.玉米叶片在响应热胁迫的过程中, 体内H3K9ac和H4K5ac水平显著升高, H3K9me2水平下降, H3K4me2水平保持不变.Kim等(2012)研究发现, 拟南芥在干旱胁迫下, 组蛋白的H3K4me3和H3K9ac修饰在干旱诱导的一些基因(如RD20和RD29a)上有明显富集.水稻在淹水胁迫下, 胁迫响应基因ADH1以及PDC1编码区的H3K4me2和H3K4me3水平显著上升, 而且胁迫后期ADH1和PDC1染色质区域的H3乙酰化水平也明显升高(Tsuji et al., 2006).Roy等(2014)在研究水稻OsDREB1bc基因时, 发现该基因能被低温特异性诱导表达, 其过表达植株对高盐和低温胁迫表现出超高的耐受性.进一步研究发现, 在此类非生物胁迫下, 该基因启动子区域的H3K9ac水平大幅度升高; 同时, H3K14ac和H3K27ac水平也呈现出位点特异性增加的趋势.除了组蛋白甲基化和乙酰化修饰外, 有研究表明组蛋白SUMO化修饰也参与了植物应答非生物胁迫过程.在高温和氧化应激条件下, SUMO化修饰可调控组蛋白乙酰化和DNA甲基化, 从而影响全基因组的转录(Miller et al., 2010).由此可见, 大量研究结果表明, 组蛋白修饰在植物响应非生物胁迫过程中具有十分重要的作用.但是, 关于不同组蛋白修饰类型之间以及组蛋白修饰与其它表观遗传调控途径的协同作用还需进一步研究, 介导协同调控植物应答非生物胁迫过程中的关键分子还有待挖掘. ... ... Liu et al., 2012; Fang et al., 2014
... Chromatin remodeling related genes involved in abiotic stresses in plants
基因
物种
胁迫类型
参考文献
AtCHR12
拟南芥(Arabidopsis thaliana )
干旱, 高温胁迫
Mlynárová et al., 2007
BRM
拟南芥(A. thaliana )
ABA, 干旱胁迫
Han et al., 2012
AtCHR23
拟南芥(A. thaliana )
盐胁迫
Folta et al., 2014
ATL1
水稻(Oryza sativa)
碱胁迫
Guo et al., 2014
AtASF1A/B
拟南芥(A. thaliana )
热胁迫
Weng et al., 2014
AtNAP1
拟南芥(A. thaliana )
盐胁迫
Liu et al., 2009
不依赖于ATP的染色质组装蛋白(如拟南芥组蛋白分子伴侣ASF1)也参与非生物胁迫过程.在热激条件下, AtASF1A和AtASF1B缺失突变体Atas1fab表现出对热胁迫高度敏感.进一步研究发现, 突变体内的HSPs和HsfA2等与热胁迫相关的基因表达受到抑制, 同时AtASF1A/B缺失也影响了拟南芥体内的H3K56- ac水平(Weng et al., 2014).该研究进一步揭示了在植物响应非生物逆境的过程中, 多种表观遗传调控途径可以协同发挥作用.另外, 有研究表明, 核仁染色质与胁迫引起的DNA损伤有关(James et al., 2014).染色质多重构象变化的平衡取决于染色质与特异的核仁DNA结合蛋白、组蛋白修饰、组蛋白变体以及DNA甲基化等相关因子的相互作用.染色质重塑复合体成员众多, 功能复杂, 目前仅有很少成员的功能被报道, 相信随着对更多染色质重塑因子的研究, 其在植物响应非生物胁迫中的作用也将不断得到揭示. ...
1 2006
... 组蛋白修饰是表观遗传调控研究领域的热点.核小体是染色质的基本组成单位, 由约146 bp的DNA缠绕各两分子的核心组蛋白H2A、H2B、H3和H4形成的八聚体构成(Luger et al., 1997).伸出核小体外的核心组蛋白的N端可被共价修饰, 这些修饰主要包括乙酰化、甲基化、磷酸化、泛素化、生物素化、ADP核糖基化以及类泛素化等(Strahl and Allis, 2000).核心组蛋白N端赖氨酸可被组蛋白乙酰化酶HATs (histone acetyltransferase)和组蛋白去乙酰化酶HDACs (histone deacetylation transferase)进行乙酰化和去乙酰化修饰.在拟南芥中, HATs分为4个不同的家族, 研究表明HATs在植物发育(Bertrand et al., 2003; Long et al., 2006)和胁迫应答(Pavangadkar et al., 2010)中具有重要作用.真核生物的HDACs包含RP- D3 (Reduced Potassium Deficiency 3)、SIR2 (Silent Information Regulator 2)和HD2 (TypeIIHDAC) 3个主要蛋白家族, 其中HD2是植物特有的HDACs (Pan- dey et al., 2002).在拟南芥中含有10个RPD3/HDA1去乙酰化酶家族蛋白(Alinsug et al., 2009), 其中HDA6能影响转基因的表达(Murfett et al., 2001)、DNA甲基化(Aufsatz et al., 2002)以及rRNA基因的调控(Probst et al., 2004).同时研究发现, HDA19能与WRKY转录因子结合, 从而调控植物的基本防御反应(Kim et al., 2008).另外, HDA6和HDA19在拟南芥种子萌发、盐胁迫以及ABA诱导的基因表达方面都发挥十分重要的作用(Chen et al., 2010). ...
1 2005
... 为了能够在胁迫条件下生存, 植物进化出复杂的机制来感知外部信号, 从而对环境变化做出最佳反应(图1).植物激素(如水杨酸(SA)、茉莉酸(JA)、乙烯(ET)和脱落酸(ABA)等内源性分子)在调节植物对生物以及非生物胁迫的抵御过程中扮演着重要角色(Bostock, 2005; Lorenzo and Solano, 2005; Mauch- Mani and Mauch, 2005).这些激素信号可通过MAP激酶级联反应调控转录因子(如MYC、MYB、NAC、ZF和HSF等)参与胁迫相关基因的表达调控(Fujita et al., 2006).同时, 有研究者认为活性氧(ROS)的产生是植物抵御生物胁迫与非生物胁迫共有的关键过程(Torres and Dangl, 2005).除转录因子的调控, 通过染色质结构变化调控基因转录也是植物响应胁迫的重要方式, 是表观遗传机制调控植物应答外界胁迫的主要体现. ...
1 2017
... DNA甲基化修饰在调节植物响应外界环境胁迫中发挥了非常重要的作用.最近几年的研究表明, 全基因组DNA甲基化修饰的改变与植物响应非生物胁迫密切相关(Boyko et al., 2010; Karan et al., 2012; Wang et al., 2014).许多与DNA甲基化修饰相关的基因参与了植物的非生物胁迫应答过程(表1).在植物响应高盐、干旱、温度和重金属等非生物胁迫过程中, 植物的特定基因或整个基因组水平的DNA甲基化修饰都会发生改变, 从而提高植物对不良环境的适应性, 确保植物在逆境条件下的生长发育.通常情况下, 非生物胁迫诱导的DNA甲基化修饰的改变发生在整个基因组水平.例如, Wang等(2011)以水稻(Oryza sativa) DK151和IR64为研究材料, 发现由干旱胁迫诱导的DNA甲基化位点的改变占全基因组DNA甲基化位点的12%左右, 这些甲基化的变化有29%在胁迫解除时仍然被保留; 并且干旱诱导的DNA甲基化还表现出组织及发育时期的特异性.Ferreira等(2015)在研究盐胁迫下DNA甲基化水平的变化时, 发现盐胁迫耐受型水稻材料Pokkali和敏感型水稻材料IR29基因组全局性的DNA甲基化水平在盐胁迫下发生了明显改变, 并且这种DNA甲基化水平的改变具有基因型及组织特异性.此外, 非生物胁迫下, DNA甲基化水平的改变往往还会伴随其它调节机制.例如, Karan等(2012)在研究水稻响应盐胁迫时, 发现逆转录转座子、非生物胁迫应答基因以及染色质修饰因子在盐胁迫下都具有不同的表达模式, 同时鉴定了不同水稻品种盐胁迫诱导的DNA甲基化模式.而对于水稻而言, 在水稻根细胞内相关基因的去甲基化修饰对其响应盐胁迫也具有重要作用(Wang et al., 2011).Zhu等(2015)在研究水稻R2R3类型的MYB转录因子OsMYB91时, 发现OsMYB91的表达受非生物胁迫诱导, 特别是盐胁迫.进一步研究发现, 该基因启动子区域的DNA甲基化水平改变是导致该基因在胁迫下表达发生变化的主要原因.除了拟南芥和水稻等常见模式植物, 人们也对其它植物响应胁迫时的DNA甲基化修饰进行了研究.例如, Bharti等(2015)在研究盐胁迫下, 类黄酮生物合成和抗氧化途径的酶基因是否受到表观遗传调控时, 发现这些基因启动子区域甲基化状态改变使得相关基因的表达水平升高, 从而导致过表达AtROS1的转基因烟草(Nicotiana tabacum)表现出对盐胁迫的耐受性.Lu等(2017)利用Me-DIP测序技术对棉花(Gossypium hirsutum)在盐胁迫下全基因组的DNA甲基化水平进行了研究, 为理解棉花盐胁迫反应的表观遗传修饰变化提供了有价值的信息.同样, 也有研究发现, 苜蓿(Medicago spp.)在盐胁迫下, 全基因组水平的DNA甲基化程度增加, 特别是在高盐胁迫下尤为明显(Al-Lawati et al., 2016).目前, 有关DNA甲基化修饰水平影响植物响应非生物胁迫的具体机制还有待深入研究. ...
1 1997
... 组蛋白修饰是表观遗传调控研究领域的热点.核小体是染色质的基本组成单位, 由约146 bp的DNA缠绕各两分子的核心组蛋白H2A、H2B、H3和H4形成的八聚体构成(Luger et al., 1997).伸出核小体外的核心组蛋白的N端可被共价修饰, 这些修饰主要包括乙酰化、甲基化、磷酸化、泛素化、生物素化、ADP核糖基化以及类泛素化等(Strahl and Allis, 2000).核心组蛋白N端赖氨酸可被组蛋白乙酰化酶HATs (histone acetyltransferase)和组蛋白去乙酰化酶HDACs (histone deacetylation transferase)进行乙酰化和去乙酰化修饰.在拟南芥中, HATs分为4个不同的家族, 研究表明HATs在植物发育(Bertrand et al., 2003; Long et al., 2006)和胁迫应答(Pavangadkar et al., 2010)中具有重要作用.真核生物的HDACs包含RP- D3 (Reduced Potassium Deficiency 3)、SIR2 (Silent Information Regulator 2)和HD2 (TypeIIHDAC) 3个主要蛋白家族, 其中HD2是植物特有的HDACs (Pan- dey et al., 2002).在拟南芥中含有10个RPD3/HDA1去乙酰化酶家族蛋白(Alinsug et al., 2009), 其中HDA6能影响转基因的表达(Murfett et al., 2001)、DNA甲基化(Aufsatz et al., 2002)以及rRNA基因的调控(Probst et al., 2004).同时研究发现, HDA19能与WRKY转录因子结合, 从而调控植物的基本防御反应(Kim et al., 2008).另外, HDA6和HDA19在拟南芥种子萌发、盐胁迫以及ABA诱导的基因表达方面都发挥十分重要的作用(Chen et al., 2010). ...
2 2012
... 除DNA甲基化修饰外, 组蛋白修饰也是植物响应非生物胁迫的重要表观调控途径之一.近年来, 有许多关于组蛋白甲基化、乙酰化和磷酸化等修饰参与植物非生物胁迫应答过程的报道(表2).在模式植物拟南芥中, SKB1可以改变胁迫相关基因的H4R3sme2水平, 其缺失突变会导致拟南芥产生对盐胁迫超敏感的表型(Zhang et al., 2011b).类似地, 拟南芥组蛋白乙酰转移酶GCN5通过影响HSFA3 (Heat Stress Transcription Factors 3)和UVH6 (UV-HYPERSENSI- TIVE 6)基因启动子区域的H3K9和H3K14的乙酰化水平来调节应答基因的转录.因此, 缺失GCN5的拟南芥突变体表现出对热胁迫的耐受性(Hu et al., 2015).在水稻中, 组蛋白乙酰转移酶基因OsHATs的转录水平和组蛋白H3和H4赖氨酸残基乙酰化水平的变化都参与了水稻的干旱胁迫调控(Fang et al., 2014).Luo等(2012)在研究植物特异的组蛋白去乙酰化酶HD2时, 发现ABA和NaCl能抑制HD2A、HD2B、HD2C以及HD2D等基因的表达水平, HD2的T-DNA插入突变体表现出对ABA和NaCl的敏感性, 同时降低了对盐胁迫的耐受性.此外, 在组蛋白去乙酰化方面, Zheng等(2016)发现拟南芥组蛋白去乙酰化酶HDA9在调节拟南芥响应盐胁迫和干旱胁迫时发挥重要作用.由此可见, 组蛋白的多种修饰类型都参与了非生物胁迫的调节过程. ... ... Histone modification related genes involved in abiotic stresses in plants
... 在自然状态下, 植物的整个生长周期均处于固着状态, 不像动物能够主动自发地躲避不良环境, 因此必然会受到来自外界生物与非生物胁迫的影响.正是由于植物处于这样一个极易受到环境胁迫的状态, 其自身进化出一系列自我保护和适应与抵制不良环境的机制(Boyko and Kovalchuk, 2008).对植物生长发育的胁迫包括内部或外部因素.内部胁迫主要为植物自身基因的突变或异常细胞对机体代谢产生的不利影响; 外部胁迫则可分为生物与非生物胁迫.生物胁迫包括病原感染、食草动物和种内竞争等; 非生物胁迫由不利的环境条件引起(如不适宜的温度、水分和营养物质的利用以及光照条件等), 这两种外部胁迫可以是永久性的也可以是临时状态(Madlung and Comai, 2004). ...
1 2006
... 在植物中, 小RNA (sRNA)是不具有编码功能的、对基因表达有调控作用的一类RNA分子, 其主要通过调节mRNA的稳定性、沉默基因转录或参与DNA甲基化等过程参与植物的生长发育(Ramachandran and Chen, 2008).因此, 小RNA也是一种重要的表观遗传调控因子.随着研究的不断深入, 对植物中sRNA的认识也有了突破性进展.以水稻为例, 研究发现, 在水稻中存在上百种sRNA, 并且在不同的生长条件下, 不同水稻品种的不同组织中sRNA的表达水平明显不同(Chen and Wu, 2010; He et al., 2010; Zhang et al., 2014).目前, 在植物中研究较多的是siRNA (small interfering RNA)和miRNA (microRNA).si- RNA是在DCL (Dicer-like protein)酶作用下由长双链RNA前体加工而来, 目前发现参与基因转录和转录后调控的内源性siRNA有nat-siRNA、ta-siRNA和hc-siRNA等(Mallory and Vaucheret, 2006).miRNA是内源基因编码的一类能与靶mRNA互补配对的一种单链小RNA, 主要参与基因的转录后调控.大量研究表明, siRNA和miRNA在植物生长发育和非生物胁迫响应过程中发挥重要作用(Phillips et al., 2007; Sunkar et al., 2007). ...
1 2005
... 为了能够在胁迫条件下生存, 植物进化出复杂的机制来感知外部信号, 从而对环境变化做出最佳反应(图1).植物激素(如水杨酸(SA)、茉莉酸(JA)、乙烯(ET)和脱落酸(ABA)等内源性分子)在调节植物对生物以及非生物胁迫的抵御过程中扮演着重要角色(Bostock, 2005; Lorenzo and Solano, 2005; Mauch- Mani and Mauch, 2005).这些激素信号可通过MAP激酶级联反应调控转录因子(如MYC、MYB、NAC、ZF和HSF等)参与胁迫相关基因的表达调控(Fujita et al., 2006).同时, 有研究者认为活性氧(ROS)的产生是植物抵御生物胁迫与非生物胁迫共有的关键过程(Torres and Dangl, 2005).除转录因子的调控, 通过染色质结构变化调控基因转录也是植物响应胁迫的重要方式, 是表观遗传机制调控植物应答外界胁迫的主要体现. ...
2 2016
... Histone modification related genes involved in abiotic stresses in plants
... ATP依赖的染色质重塑是染色质重塑复合体利用ATP水解释放的能量来改变染色质结构, 从而调控真核生物基因表达的一种重要的表观遗传调控机制(Racki and Narlikar, 2008).ATP依赖的染色质重塑复合物主要分为4个家族: SWI/SNF家族、ISWI家族、CHD家族和INO80家族.SWI/SNF是第1个被发现的ATP依赖的染色质重塑复合物(Sudarsanam and Winston, 2000).SWI/SNF复合物通过改变染色质结构在DNA修饰、重组以及基因表达中发挥重要作用(Geiman and Robertson, 2002).研究表明, SWI/ SNF复合物也参与了环境胁迫对植物生长发育的调控.例如, 在拟南芥中, 过表达SNF2/Brahma类型的染色质重塑蛋白AtCHR12, 该过表达植株在干旱和高温等胁迫下表现出花芽以及初生茎生长停滞现象(Mlynárová et al., 2007).同时, 有研究表明, SWI/SNF和CHD复合物都参与了ABA调控的拟南芥种子萌发及生长等过程(Perruc et al., 2007; Saez et al., 2008). ... ... Chromatin remodeling related genes involved in abiotic stresses in plants
基因
物种
胁迫类型
参考文献
AtCHR12
拟南芥(Arabidopsis thaliana )
干旱, 高温胁迫
Mlynárová et al., 2007
BRM
拟南芥(A. thaliana )
ABA, 干旱胁迫
Han et al., 2012
AtCHR23
拟南芥(A. thaliana )
盐胁迫
Folta et al., 2014
ATL1
水稻(Oryza sativa)
碱胁迫
Guo et al., 2014
AtASF1A/B
拟南芥(A. thaliana )
热胁迫
Weng et al., 2014
AtNAP1
拟南芥(A. thaliana )
盐胁迫
Liu et al., 2009
不依赖于ATP的染色质组装蛋白(如拟南芥组蛋白分子伴侣ASF1)也参与非生物胁迫过程.在热激条件下, AtASF1A和AtASF1B缺失突变体Atas1fab表现出对热胁迫高度敏感.进一步研究发现, 突变体内的HSPs和HsfA2等与热胁迫相关的基因表达受到抑制, 同时AtASF1A/B缺失也影响了拟南芥体内的H3K56- ac水平(Weng et al., 2014).该研究进一步揭示了在植物响应非生物逆境的过程中, 多种表观遗传调控途径可以协同发挥作用.另外, 有研究表明, 核仁染色质与胁迫引起的DNA损伤有关(James et al., 2014).染色质多重构象变化的平衡取决于染色质与特异的核仁DNA结合蛋白、组蛋白修饰、组蛋白变体以及DNA甲基化等相关因子的相互作用.染色质重塑复合体成员众多, 功能复杂, 目前仅有很少成员的功能被报道, 相信随着对更多染色质重塑因子的研究, 其在植物响应非生物胁迫中的作用也将不断得到揭示. ...
1 2001
... 组蛋白修饰是表观遗传调控研究领域的热点.核小体是染色质的基本组成单位, 由约146 bp的DNA缠绕各两分子的核心组蛋白H2A、H2B、H3和H4形成的八聚体构成(Luger et al., 1997).伸出核小体外的核心组蛋白的N端可被共价修饰, 这些修饰主要包括乙酰化、甲基化、磷酸化、泛素化、生物素化、ADP核糖基化以及类泛素化等(Strahl and Allis, 2000).核心组蛋白N端赖氨酸可被组蛋白乙酰化酶HATs (histone acetyltransferase)和组蛋白去乙酰化酶HDACs (histone deacetylation transferase)进行乙酰化和去乙酰化修饰.在拟南芥中, HATs分为4个不同的家族, 研究表明HATs在植物发育(Bertrand et al., 2003; Long et al., 2006)和胁迫应答(Pavangadkar et al., 2010)中具有重要作用.真核生物的HDACs包含RP- D3 (Reduced Potassium Deficiency 3)、SIR2 (Silent Information Regulator 2)和HD2 (TypeIIHDAC) 3个主要蛋白家族, 其中HD2是植物特有的HDACs (Pan- dey et al., 2002).在拟南芥中含有10个RPD3/HDA1去乙酰化酶家族蛋白(Alinsug et al., 2009), 其中HDA6能影响转基因的表达(Murfett et al., 2001)、DNA甲基化(Aufsatz et al., 2002)以及rRNA基因的调控(Probst et al., 2004).同时研究发现, HDA19能与WRKY转录因子结合, 从而调控植物的基本防御反应(Kim et al., 2008).另外, HDA6和HDA19在拟南芥种子萌发、盐胁迫以及ABA诱导的基因表达方面都发挥十分重要的作用(Chen et al., 2010). ...
... sRNA and sRNA-regulated genes involved in abiotic stresses in plants
基因
物种
胁迫类型
参考文献
SlAGO4A
番茄(Solanum lycopersicum)
干旱胁迫, 盐胁迫
Huang et al., 2016
19 different miRNAs
水稻(Oryza sativa)
干旱胁迫
Zhou et al., 2010
13 different miRNAs
野生二粒小麦(Triticum turgidum ssp. dicoccoides)
干旱胁迫
Kantar et al., 2011
CSD1, CSD2, CCS
拟南芥(Arabidopsis thaliana )
高温胁迫
Guan et al., 2013
NFYA5
拟南芥(A. thaliana )
干旱胁迫
Li et al., 2008
OsPCF5, OsPCF8
水稻(O. sativa)
低温胁迫
Yang et al., 2013
GmNFYA3
大豆(Glycine max )
干旱胁迫
Ni et al., 2013
Phvul.010g120700
菜豆(Phaseolus vulgaris)
干旱胁迫
Sosa-Valencia et al., 2017
SlNF-YA1/2/3, SlMRP1
番茄(S. lycopersicum)
干旱胁迫
Zhang et al., 2011a
BHLH23
鹰嘴豆(Cicer arietinum)
干旱胁迫
Hajyzadeh et al., 2015
4 染色质重塑与非生物胁迫 ATP依赖的染色质重塑是染色质重塑复合体利用ATP水解释放的能量来改变染色质结构, 从而调控真核生物基因表达的一种重要的表观遗传调控机制(Racki and Narlikar, 2008).ATP依赖的染色质重塑复合物主要分为4个家族: SWI/SNF家族、ISWI家族、CHD家族和INO80家族.SWI/SNF是第1个被发现的ATP依赖的染色质重塑复合物(Sudarsanam and Winston, 2000).SWI/SNF复合物通过改变染色质结构在DNA修饰、重组以及基因表达中发挥重要作用(Geiman and Robertson, 2002).研究表明, SWI/ SNF复合物也参与了环境胁迫对植物生长发育的调控.例如, 在拟南芥中, 过表达SNF2/Brahma类型的染色质重塑蛋白AtCHR12, 该过表达植株在干旱和高温等胁迫下表现出花芽以及初生茎生长停滞现象(Mlynárová et al., 2007).同时, 有研究表明, SWI/SNF和CHD复合物都参与了ABA调控的拟南芥种子萌发及生长等过程(Perruc et al., 2007; Saez et al., 2008). ...
1 2002
... 组蛋白修饰是表观遗传调控研究领域的热点.核小体是染色质的基本组成单位, 由约146 bp的DNA缠绕各两分子的核心组蛋白H2A、H2B、H3和H4形成的八聚体构成(Luger et al., 1997).伸出核小体外的核心组蛋白的N端可被共价修饰, 这些修饰主要包括乙酰化、甲基化、磷酸化、泛素化、生物素化、ADP核糖基化以及类泛素化等(Strahl and Allis, 2000).核心组蛋白N端赖氨酸可被组蛋白乙酰化酶HATs (histone acetyltransferase)和组蛋白去乙酰化酶HDACs (histone deacetylation transferase)进行乙酰化和去乙酰化修饰.在拟南芥中, HATs分为4个不同的家族, 研究表明HATs在植物发育(Bertrand et al., 2003; Long et al., 2006)和胁迫应答(Pavangadkar et al., 2010)中具有重要作用.真核生物的HDACs包含RP- D3 (Reduced Potassium Deficiency 3)、SIR2 (Silent Information Regulator 2)和HD2 (TypeIIHDAC) 3个主要蛋白家族, 其中HD2是植物特有的HDACs (Pan- dey et al., 2002).在拟南芥中含有10个RPD3/HDA1去乙酰化酶家族蛋白(Alinsug et al., 2009), 其中HDA6能影响转基因的表达(Murfett et al., 2001)、DNA甲基化(Aufsatz et al., 2002)以及rRNA基因的调控(Probst et al., 2004).同时研究发现, HDA19能与WRKY转录因子结合, 从而调控植物的基本防御反应(Kim et al., 2008).另外, HDA6和HDA19在拟南芥种子萌发、盐胁迫以及ABA诱导的基因表达方面都发挥十分重要的作用(Chen et al., 2010). ...
1 2010
... Histone modification related genes involved in abiotic stresses in plants
... 组蛋白修饰是表观遗传调控研究领域的热点.核小体是染色质的基本组成单位, 由约146 bp的DNA缠绕各两分子的核心组蛋白H2A、H2B、H3和H4形成的八聚体构成(Luger et al., 1997).伸出核小体外的核心组蛋白的N端可被共价修饰, 这些修饰主要包括乙酰化、甲基化、磷酸化、泛素化、生物素化、ADP核糖基化以及类泛素化等(Strahl and Allis, 2000).核心组蛋白N端赖氨酸可被组蛋白乙酰化酶HATs (histone acetyltransferase)和组蛋白去乙酰化酶HDACs (histone deacetylation transferase)进行乙酰化和去乙酰化修饰.在拟南芥中, HATs分为4个不同的家族, 研究表明HATs在植物发育(Bertrand et al., 2003; Long et al., 2006)和胁迫应答(Pavangadkar et al., 2010)中具有重要作用.真核生物的HDACs包含RP- D3 (Reduced Potassium Deficiency 3)、SIR2 (Silent Information Regulator 2)和HD2 (TypeIIHDAC) 3个主要蛋白家族, 其中HD2是植物特有的HDACs (Pan- dey et al., 2002).在拟南芥中含有10个RPD3/HDA1去乙酰化酶家族蛋白(Alinsug et al., 2009), 其中HDA6能影响转基因的表达(Murfett et al., 2001)、DNA甲基化(Aufsatz et al., 2002)以及rRNA基因的调控(Probst et al., 2004).同时研究发现, HDA19能与WRKY转录因子结合, 从而调控植物的基本防御反应(Kim et al., 2008).另外, HDA6和HDA19在拟南芥种子萌发、盐胁迫以及ABA诱导的基因表达方面都发挥十分重要的作用(Chen et al., 2010). ...
1 2015
... 染色质重塑复合物在调节非生物胁迫相关基因的表达中起重要作用(表4).对拟南芥的研究表明, SWI3B能与ABA通路重要调控因子HAB1结合, 从而参与ABA调节的植物对非生物胁迫的适应过程.HAB1是ABA信号通路中起关键作用的负调控因子, 拟南芥swi3b突变体表现出种子萌发对ABA的弱敏感性, ABA应答基因RAB18和RD29B的表达下降, 并且发现ABA能降低HAB1在RAB18和RD2B基因启动子上的富集(Saez et al., 2008; Yuan et al., 2013).在拟南芥中, 植物对ABA的响应大多通过转录因子ABI5调节.Han等(2012)发现, SWI2/SNF2家族成员BRM的功能缺失导致拟南芥对ABA超敏感, BRM能与ABI5基因结合, BRM的缺失使得核小体不稳定, 进而使ABI5的转录受到抑制, 最终使得brm突变体表现出对干旱的耐受性.而ABA信号转导途径的关键组分SnRK2和PP2CA分别能够介导BRM的磷酸化和去磷酸化, 从而快速调控BRM的活性(Peirats-Llobet et al., 2015).此外, 研究还发现, AtMMS21介导的泛素化能够提高拟南芥根发育过程中BRM的稳定性(Zhang et al., 2017).另外, 最近研究发现, 一种植物特异的H3K27去甲基化酶REF6 (RELATIVE OF EARLY FLOWERING 6)能够结合在带有CTCTGYTY基序的基因组区域, 催化H3K27me3去甲基化并促进BRM结合在该区段, 该研究揭示了组蛋白修饰与染色质重塑的协同作用(Li et al., 2016). ...
1 2007
... ATP依赖的染色质重塑是染色质重塑复合体利用ATP水解释放的能量来改变染色质结构, 从而调控真核生物基因表达的一种重要的表观遗传调控机制(Racki and Narlikar, 2008).ATP依赖的染色质重塑复合物主要分为4个家族: SWI/SNF家族、ISWI家族、CHD家族和INO80家族.SWI/SNF是第1个被发现的ATP依赖的染色质重塑复合物(Sudarsanam and Winston, 2000).SWI/SNF复合物通过改变染色质结构在DNA修饰、重组以及基因表达中发挥重要作用(Geiman and Robertson, 2002).研究表明, SWI/ SNF复合物也参与了环境胁迫对植物生长发育的调控.例如, 在拟南芥中, 过表达SNF2/Brahma类型的染色质重塑蛋白AtCHR12, 该过表达植株在干旱和高温等胁迫下表现出花芽以及初生茎生长停滞现象(Mlynárová et al., 2007).同时, 有研究表明, SWI/SNF和CHD复合物都参与了ABA调控的拟南芥种子萌发及生长等过程(Perruc et al., 2007; Saez et al., 2008). ...
1 2007
... 在植物中, 小RNA (sRNA)是不具有编码功能的、对基因表达有调控作用的一类RNA分子, 其主要通过调节mRNA的稳定性、沉默基因转录或参与DNA甲基化等过程参与植物的生长发育(Ramachandran and Chen, 2008).因此, 小RNA也是一种重要的表观遗传调控因子.随着研究的不断深入, 对植物中sRNA的认识也有了突破性进展.以水稻为例, 研究发现, 在水稻中存在上百种sRNA, 并且在不同的生长条件下, 不同水稻品种的不同组织中sRNA的表达水平明显不同(Chen and Wu, 2010; He et al., 2010; Zhang et al., 2014).目前, 在植物中研究较多的是siRNA (small interfering RNA)和miRNA (microRNA).si- RNA是在DCL (Dicer-like protein)酶作用下由长双链RNA前体加工而来, 目前发现参与基因转录和转录后调控的内源性siRNA有nat-siRNA、ta-siRNA和hc-siRNA等(Mallory and Vaucheret, 2006).miRNA是内源基因编码的一类能与靶mRNA互补配对的一种单链小RNA, 主要参与基因的转录后调控.大量研究表明, siRNA和miRNA在植物生长发育和非生物胁迫响应过程中发挥重要作用(Phillips et al., 2007; Sunkar et al., 2007). ...
1 2010
... 组蛋白的甲基化和去甲基化通常发生在精氨酸和赖氨酸残基上, 赖氨酸残基的甲基化主要由含有SET结构域的赖氨酸甲基转移酶HKMTs催化形成(Pontvianne et al., 2010).SET结构域蛋白可分为TrxG (Trithorax Group)、E(Z) (Enhancer of Zeste)、SU (VAR)以及ASH1 (Absent, Small, or Homeotic dis- cs) 4个家族, 分别负责组蛋白H3K4、H3K9、H3K27和H3K36的甲基化修饰.在拟南芥中, TrxG家族蛋白SDG25 (Berr et al., 2009)、E(Z)家族蛋白CLF (Chanvivattana et al., 2004)以及ASH1家族蛋白SDG8和SDG26 (Xu et al., 2008)都参与开花时间的调控.组蛋白去甲基化酶HDMs包括KDM1/LSD1家族和JmjC结构域蛋白家族两类(Liu et al., 2010).这两类蛋白质都可通过氧化反应直接去除组蛋白上的甲基化修饰.KDM1/LSD1以黄素为辅助因子, 只能对一、二甲基化修饰进行去甲基, 并不能对三甲基化去甲基; JmjC结构域蛋白需要二价铁和α酮戊二酸作辅助因子, 可以对一、二、三甲基化修饰去甲基.截至目前, 拟南芥中报道了4个KDM1/LSD1家族以及21个JMJ家族组蛋白去甲基化酶, 它们主要参与拟南芥的开花调控(Saze et al., 2008; Liu et al., 2010).组蛋白的乙酰化、磷酸化和泛素化修饰一般与基因转录激活有关, SUMO化和生物素化与转录抑制相关.已有的研究显示, 与胁迫相关的基因激活主要由H3K4、H3K36甲基化以及H3K9乙酰化调控, 而H3K9、H3K27甲基化和H3去乙酰化主要参与相关基因的沉默过程(Qiao and Fan, 2011). ...
1 2013
... Abiotic stress responsive genes regulated by DNA-methylation in plants
基因
物种
胁迫类型
参考文献
Asr1, Asr2
番茄(Solanum lycopersicum)
干旱胁迫
González et al., 2011; 2013
NtGPDL
烟草(Nicotiana tabaccum cv. ‘Xanthi nc’)
低温胁迫
Choi and Sano, 2007
OsMYB91
水稻(Oryza sativa)
盐胁迫
Zhu et al., 2015
Glyma11g02400 Glyma16g27950 Glyma20g30840
大豆(Glycine max)
盐胁迫
Song et al., 2012
SPCH, FAMA
拟南芥(Arabidopsis thaliana)
湿度胁迫
Tricker et al., 2012
ZmMI1, Ac/Ds
玉米(Zea mays)
低温胁迫
Steward et al., 2002
Tam3
金鱼草(Antirrhinum majus)
低温胁迫
Hashida et al., 2006
NRPD2
拟南芥(A. thaliana)
高温胁迫
Popova et al., 2013
2 组蛋白修饰2.1 植物体内的组蛋白修饰 组蛋白修饰是表观遗传调控研究领域的热点.核小体是染色质的基本组成单位, 由约146 bp的DNA缠绕各两分子的核心组蛋白H2A、H2B、H3和H4形成的八聚体构成(Luger et al., 1997).伸出核小体外的核心组蛋白的N端可被共价修饰, 这些修饰主要包括乙酰化、甲基化、磷酸化、泛素化、生物素化、ADP核糖基化以及类泛素化等(Strahl and Allis, 2000).核心组蛋白N端赖氨酸可被组蛋白乙酰化酶HATs (histone acetyltransferase)和组蛋白去乙酰化酶HDACs (histone deacetylation transferase)进行乙酰化和去乙酰化修饰.在拟南芥中, HATs分为4个不同的家族, 研究表明HATs在植物发育(Bertrand et al., 2003; Long et al., 2006)和胁迫应答(Pavangadkar et al., 2010)中具有重要作用.真核生物的HDACs包含RP- D3 (Reduced Potassium Deficiency 3)、SIR2 (Silent Information Regulator 2)和HD2 (TypeIIHDAC) 3个主要蛋白家族, 其中HD2是植物特有的HDACs (Pan- dey et al., 2002).在拟南芥中含有10个RPD3/HDA1去乙酰化酶家族蛋白(Alinsug et al., 2009), 其中HDA6能影响转基因的表达(Murfett et al., 2001)、DNA甲基化(Aufsatz et al., 2002)以及rRNA基因的调控(Probst et al., 2004).同时研究发现, HDA19能与WRKY转录因子结合, 从而调控植物的基本防御反应(Kim et al., 2008).另外, HDA6和HDA19在拟南芥种子萌发、盐胁迫以及ABA诱导的基因表达方面都发挥十分重要的作用(Chen et al., 2010). ...
1 2004
... 组蛋白修饰是表观遗传调控研究领域的热点.核小体是染色质的基本组成单位, 由约146 bp的DNA缠绕各两分子的核心组蛋白H2A、H2B、H3和H4形成的八聚体构成(Luger et al., 1997).伸出核小体外的核心组蛋白的N端可被共价修饰, 这些修饰主要包括乙酰化、甲基化、磷酸化、泛素化、生物素化、ADP核糖基化以及类泛素化等(Strahl and Allis, 2000).核心组蛋白N端赖氨酸可被组蛋白乙酰化酶HATs (histone acetyltransferase)和组蛋白去乙酰化酶HDACs (histone deacetylation transferase)进行乙酰化和去乙酰化修饰.在拟南芥中, HATs分为4个不同的家族, 研究表明HATs在植物发育(Bertrand et al., 2003; Long et al., 2006)和胁迫应答(Pavangadkar et al., 2010)中具有重要作用.真核生物的HDACs包含RP- D3 (Reduced Potassium Deficiency 3)、SIR2 (Silent Information Regulator 2)和HD2 (TypeIIHDAC) 3个主要蛋白家族, 其中HD2是植物特有的HDACs (Pan- dey et al., 2002).在拟南芥中含有10个RPD3/HDA1去乙酰化酶家族蛋白(Alinsug et al., 2009), 其中HDA6能影响转基因的表达(Murfett et al., 2001)、DNA甲基化(Aufsatz et al., 2002)以及rRNA基因的调控(Probst et al., 2004).同时研究发现, HDA19能与WRKY转录因子结合, 从而调控植物的基本防御反应(Kim et al., 2008).另外, HDA6和HDA19在拟南芥种子萌发、盐胁迫以及ABA诱导的基因表达方面都发挥十分重要的作用(Chen et al., 2010). ...
1 2011
... 组蛋白的甲基化和去甲基化通常发生在精氨酸和赖氨酸残基上, 赖氨酸残基的甲基化主要由含有SET结构域的赖氨酸甲基转移酶HKMTs催化形成(Pontvianne et al., 2010).SET结构域蛋白可分为TrxG (Trithorax Group)、E(Z) (Enhancer of Zeste)、SU (VAR)以及ASH1 (Absent, Small, or Homeotic dis- cs) 4个家族, 分别负责组蛋白H3K4、H3K9、H3K27和H3K36的甲基化修饰.在拟南芥中, TrxG家族蛋白SDG25 (Berr et al., 2009)、E(Z)家族蛋白CLF (Chanvivattana et al., 2004)以及ASH1家族蛋白SDG8和SDG26 (Xu et al., 2008)都参与开花时间的调控.组蛋白去甲基化酶HDMs包括KDM1/LSD1家族和JmjC结构域蛋白家族两类(Liu et al., 2010).这两类蛋白质都可通过氧化反应直接去除组蛋白上的甲基化修饰.KDM1/LSD1以黄素为辅助因子, 只能对一、二甲基化修饰进行去甲基, 并不能对三甲基化去甲基; JmjC结构域蛋白需要二价铁和α酮戊二酸作辅助因子, 可以对一、二、三甲基化修饰去甲基.截至目前, 拟南芥中报道了4个KDM1/LSD1家族以及21个JMJ家族组蛋白去甲基化酶, 它们主要参与拟南芥的开花调控(Saze et al., 2008; Liu et al., 2010).组蛋白的乙酰化、磷酸化和泛素化修饰一般与基因转录激活有关, SUMO化和生物素化与转录抑制相关.已有的研究显示, 与胁迫相关的基因激活主要由H3K4、H3K36甲基化以及H3K9乙酰化调控, 而H3K9、H3K27甲基化和H3去乙酰化主要参与相关基因的沉默过程(Qiao and Fan, 2011). ...
1 2008
... ATP依赖的染色质重塑是染色质重塑复合体利用ATP水解释放的能量来改变染色质结构, 从而调控真核生物基因表达的一种重要的表观遗传调控机制(Racki and Narlikar, 2008).ATP依赖的染色质重塑复合物主要分为4个家族: SWI/SNF家族、ISWI家族、CHD家族和INO80家族.SWI/SNF是第1个被发现的ATP依赖的染色质重塑复合物(Sudarsanam and Winston, 2000).SWI/SNF复合物通过改变染色质结构在DNA修饰、重组以及基因表达中发挥重要作用(Geiman and Robertson, 2002).研究表明, SWI/ SNF复合物也参与了环境胁迫对植物生长发育的调控.例如, 在拟南芥中, 过表达SNF2/Brahma类型的染色质重塑蛋白AtCHR12, 该过表达植株在干旱和高温等胁迫下表现出花芽以及初生茎生长停滞现象(Mlynárová et al., 2007).同时, 有研究表明, SWI/SNF和CHD复合物都参与了ABA调控的拟南芥种子萌发及生长等过程(Perruc et al., 2007; Saez et al., 2008). ...
1 2008
... 在植物中, 小RNA (sRNA)是不具有编码功能的、对基因表达有调控作用的一类RNA分子, 其主要通过调节mRNA的稳定性、沉默基因转录或参与DNA甲基化等过程参与植物的生长发育(Ramachandran and Chen, 2008).因此, 小RNA也是一种重要的表观遗传调控因子.随着研究的不断深入, 对植物中sRNA的认识也有了突破性进展.以水稻为例, 研究发现, 在水稻中存在上百种sRNA, 并且在不同的生长条件下, 不同水稻品种的不同组织中sRNA的表达水平明显不同(Chen and Wu, 2010; He et al., 2010; Zhang et al., 2014).目前, 在植物中研究较多的是siRNA (small interfering RNA)和miRNA (microRNA).si- RNA是在DCL (Dicer-like protein)酶作用下由长双链RNA前体加工而来, 目前发现参与基因转录和转录后调控的内源性siRNA有nat-siRNA、ta-siRNA和hc-siRNA等(Mallory and Vaucheret, 2006).miRNA是内源基因编码的一类能与靶mRNA互补配对的一种单链小RNA, 主要参与基因的转录后调控.大量研究表明, siRNA和miRNA在植物生长发育和非生物胁迫响应过程中发挥重要作用(Phillips et al., 2007; Sunkar et al., 2007). ...
2 2014
... Histone modification related genes involved in abiotic stresses in plants
... 表观遗传最早被定义为有丝分裂以及减数分裂过程中无法用DNA序列的改变来解释基因功能的可继承性改变(Russo et al., 1996).目前, 人们一致认为表观遗传是研究没有DNA序列变化的、可遗传的基因表达改变(Wu and Morris, 2001).表观遗传调控机制主要包括DNA甲基化、组蛋白修饰、染色质重塑以及非编码RNA等.越来越多的研究表明, 上述表观遗传机制在植物应答环境胁迫中发挥了非常重要的作用.本文简要阐述近年来表观遗传机制调节植物响应非生物胁迫的一系列研究成果. ...
2 2008
... ATP依赖的染色质重塑是染色质重塑复合体利用ATP水解释放的能量来改变染色质结构, 从而调控真核生物基因表达的一种重要的表观遗传调控机制(Racki and Narlikar, 2008).ATP依赖的染色质重塑复合物主要分为4个家族: SWI/SNF家族、ISWI家族、CHD家族和INO80家族.SWI/SNF是第1个被发现的ATP依赖的染色质重塑复合物(Sudarsanam and Winston, 2000).SWI/SNF复合物通过改变染色质结构在DNA修饰、重组以及基因表达中发挥重要作用(Geiman and Robertson, 2002).研究表明, SWI/ SNF复合物也参与了环境胁迫对植物生长发育的调控.例如, 在拟南芥中, 过表达SNF2/Brahma类型的染色质重塑蛋白AtCHR12, 该过表达植株在干旱和高温等胁迫下表现出花芽以及初生茎生长停滞现象(Mlynárová et al., 2007).同时, 有研究表明, SWI/SNF和CHD复合物都参与了ABA调控的拟南芥种子萌发及生长等过程(Perruc et al., 2007; Saez et al., 2008). ... ... 染色质重塑复合物在调节非生物胁迫相关基因的表达中起重要作用(表4).对拟南芥的研究表明, SWI3B能与ABA通路重要调控因子HAB1结合, 从而参与ABA调节的植物对非生物胁迫的适应过程.HAB1是ABA信号通路中起关键作用的负调控因子, 拟南芥swi3b突变体表现出种子萌发对ABA的弱敏感性, ABA应答基因RAB18和RD29B的表达下降, 并且发现ABA能降低HAB1在RAB18和RD2B基因启动子上的富集(Saez et al., 2008; Yuan et al., 2013).在拟南芥中, 植物对ABA的响应大多通过转录因子ABI5调节.Han等(2012)发现, SWI2/SNF2家族成员BRM的功能缺失导致拟南芥对ABA超敏感, BRM能与ABI5基因结合, BRM的缺失使得核小体不稳定, 进而使ABI5的转录受到抑制, 最终使得brm突变体表现出对干旱的耐受性.而ABA信号转导途径的关键组分SnRK2和PP2CA分别能够介导BRM的磷酸化和去磷酸化, 从而快速调控BRM的活性(Peirats-Llobet et al., 2015).此外, 研究还发现, AtMMS21介导的泛素化能够提高拟南芥根发育过程中BRM的稳定性(Zhang et al., 2017).另外, 最近研究发现, 一种植物特异的H3K27去甲基化酶REF6 (RELATIVE OF EARLY FLOWERING 6)能够结合在带有CTCTGYTY基序的基因组区域, 催化H3K27me3去甲基化并促进BRM结合在该区段, 该研究揭示了组蛋白修饰与染色质重塑的协同作用(Li et al., 2016). ...
1 2013
... DNA甲基化是指以DNA为受体, 在DNA甲基转移酶的作用下, 将供体S-腺苷甲硫氨酸上的1个甲基转移至胞嘧啶的第5位碳原子上, 从而形成5-甲基胞嘧啶(m5C)的过程(Sahu et al., 2013).1925年, m5C首次在结核分枝杆菌的结核菌素的水解产物中被发现(Johnson and Coghill, 1925).随后, 在植物中发现了较高水平的m5C甲基化修饰(Vanyushin and Belozerskii, 1959).DNA胞嘧啶甲基化修饰主要包括不对称(mCpHpH)甲基化和对称(mCpG和mCpHpG)甲基化.例如, 在拟南芥(Arabidopsis thaliana)中, CG、CHG与CHH的甲基化水平分别为24%、6.7%和1.7%; 并且在同一植物的不同组织或同一组织的不同发育阶段, 其基因组DNA的甲基化位点和水平也不一样(Dhar et al., 2014). ...
1 2008
... 组蛋白的甲基化和去甲基化通常发生在精氨酸和赖氨酸残基上, 赖氨酸残基的甲基化主要由含有SET结构域的赖氨酸甲基转移酶HKMTs催化形成(Pontvianne et al., 2010).SET结构域蛋白可分为TrxG (Trithorax Group)、E(Z) (Enhancer of Zeste)、SU (VAR)以及ASH1 (Absent, Small, or Homeotic dis- cs) 4个家族, 分别负责组蛋白H3K4、H3K9、H3K27和H3K36的甲基化修饰.在拟南芥中, TrxG家族蛋白SDG25 (Berr et al., 2009)、E(Z)家族蛋白CLF (Chanvivattana et al., 2004)以及ASH1家族蛋白SDG8和SDG26 (Xu et al., 2008)都参与开花时间的调控.组蛋白去甲基化酶HDMs包括KDM1/LSD1家族和JmjC结构域蛋白家族两类(Liu et al., 2010).这两类蛋白质都可通过氧化反应直接去除组蛋白上的甲基化修饰.KDM1/LSD1以黄素为辅助因子, 只能对一、二甲基化修饰进行去甲基, 并不能对三甲基化去甲基; JmjC结构域蛋白需要二价铁和α酮戊二酸作辅助因子, 可以对一、二、三甲基化修饰去甲基.截至目前, 拟南芥中报道了4个KDM1/LSD1家族以及21个JMJ家族组蛋白去甲基化酶, 它们主要参与拟南芥的开花调控(Saze et al., 2008; Liu et al., 2010).组蛋白的乙酰化、磷酸化和泛素化修饰一般与基因转录激活有关, SUMO化和生物素化与转录抑制相关.已有的研究显示, 与胁迫相关的基因激活主要由H3K4、H3K36甲基化以及H3K9乙酰化调控, 而H3K9、H3K27甲基化和H3去乙酰化主要参与相关基因的沉默过程(Qiao and Fan, 2011). ...
1 2012
... Abiotic stress responsive genes regulated by DNA-methylation in plants
基因
物种
胁迫类型
参考文献
Asr1, Asr2
番茄(Solanum lycopersicum)
干旱胁迫
González et al., 2011; 2013
NtGPDL
烟草(Nicotiana tabaccum cv. ‘Xanthi nc’)
低温胁迫
Choi and Sano, 2007
OsMYB91
水稻(Oryza sativa)
盐胁迫
Zhu et al., 2015
Glyma11g02400 Glyma16g27950 Glyma20g30840
大豆(Glycine max)
盐胁迫
Song et al., 2012
SPCH, FAMA
拟南芥(Arabidopsis thaliana)
湿度胁迫
Tricker et al., 2012
ZmMI1, Ac/Ds
玉米(Zea mays)
低温胁迫
Steward et al., 2002
Tam3
金鱼草(Antirrhinum majus)
低温胁迫
Hashida et al., 2006
NRPD2
拟南芥(A. thaliana)
高温胁迫
Popova et al., 2013
2 组蛋白修饰2.1 植物体内的组蛋白修饰 组蛋白修饰是表观遗传调控研究领域的热点.核小体是染色质的基本组成单位, 由约146 bp的DNA缠绕各两分子的核心组蛋白H2A、H2B、H3和H4形成的八聚体构成(Luger et al., 1997).伸出核小体外的核心组蛋白的N端可被共价修饰, 这些修饰主要包括乙酰化、甲基化、磷酸化、泛素化、生物素化、ADP核糖基化以及类泛素化等(Strahl and Allis, 2000).核心组蛋白N端赖氨酸可被组蛋白乙酰化酶HATs (histone acetyltransferase)和组蛋白去乙酰化酶HDACs (histone deacetylation transferase)进行乙酰化和去乙酰化修饰.在拟南芥中, HATs分为4个不同的家族, 研究表明HATs在植物发育(Bertrand et al., 2003; Long et al., 2006)和胁迫应答(Pavangadkar et al., 2010)中具有重要作用.真核生物的HDACs包含RP- D3 (Reduced Potassium Deficiency 3)、SIR2 (Silent Information Regulator 2)和HD2 (TypeIIHDAC) 3个主要蛋白家族, 其中HD2是植物特有的HDACs (Pan- dey et al., 2002).在拟南芥中含有10个RPD3/HDA1去乙酰化酶家族蛋白(Alinsug et al., 2009), 其中HDA6能影响转基因的表达(Murfett et al., 2001)、DNA甲基化(Aufsatz et al., 2002)以及rRNA基因的调控(Probst et al., 2004).同时研究发现, HDA19能与WRKY转录因子结合, 从而调控植物的基本防御反应(Kim et al., 2008).另外, HDA6和HDA19在拟南芥种子萌发、盐胁迫以及ABA诱导的基因表达方面都发挥十分重要的作用(Chen et al., 2010). ...
1 2017
... sRNA and sRNA-regulated genes involved in abiotic stresses in plants
基因
物种
胁迫类型
参考文献
SlAGO4A
番茄(Solanum lycopersicum)
干旱胁迫, 盐胁迫
Huang et al., 2016
19 different miRNAs
水稻(Oryza sativa)
干旱胁迫
Zhou et al., 2010
13 different miRNAs
野生二粒小麦(Triticum turgidum ssp. dicoccoides)
干旱胁迫
Kantar et al., 2011
CSD1, CSD2, CCS
拟南芥(Arabidopsis thaliana )
高温胁迫
Guan et al., 2013
NFYA5
拟南芥(A. thaliana )
干旱胁迫
Li et al., 2008
OsPCF5, OsPCF8
水稻(O. sativa)
低温胁迫
Yang et al., 2013
GmNFYA3
大豆(Glycine max )
干旱胁迫
Ni et al., 2013
Phvul.010g120700
菜豆(Phaseolus vulgaris)
干旱胁迫
Sosa-Valencia et al., 2017
SlNF-YA1/2/3, SlMRP1
番茄(S. lycopersicum)
干旱胁迫
Zhang et al., 2011a
BHLH23
鹰嘴豆(Cicer arietinum)
干旱胁迫
Hajyzadeh et al., 2015
4 染色质重塑与非生物胁迫 ATP依赖的染色质重塑是染色质重塑复合体利用ATP水解释放的能量来改变染色质结构, 从而调控真核生物基因表达的一种重要的表观遗传调控机制(Racki and Narlikar, 2008).ATP依赖的染色质重塑复合物主要分为4个家族: SWI/SNF家族、ISWI家族、CHD家族和INO80家族.SWI/SNF是第1个被发现的ATP依赖的染色质重塑复合物(Sudarsanam and Winston, 2000).SWI/SNF复合物通过改变染色质结构在DNA修饰、重组以及基因表达中发挥重要作用(Geiman and Robertson, 2002).研究表明, SWI/ SNF复合物也参与了环境胁迫对植物生长发育的调控.例如, 在拟南芥中, 过表达SNF2/Brahma类型的染色质重塑蛋白AtCHR12, 该过表达植株在干旱和高温等胁迫下表现出花芽以及初生茎生长停滞现象(Mlynárová et al., 2007).同时, 有研究表明, SWI/SNF和CHD复合物都参与了ABA调控的拟南芥种子萌发及生长等过程(Perruc et al., 2007; Saez et al., 2008). ...
1 2006
... Histone modification related genes involved in abiotic stresses in plants
... Abiotic stress responsive genes regulated by DNA-methylation in plants
基因
物种
胁迫类型
参考文献
Asr1, Asr2
番茄(Solanum lycopersicum)
干旱胁迫
González et al., 2011; 2013
NtGPDL
烟草(Nicotiana tabaccum cv. ‘Xanthi nc’)
低温胁迫
Choi and Sano, 2007
OsMYB91
水稻(Oryza sativa)
盐胁迫
Zhu et al., 2015
Glyma11g02400 Glyma16g27950 Glyma20g30840
大豆(Glycine max)
盐胁迫
Song et al., 2012
SPCH, FAMA
拟南芥(Arabidopsis thaliana)
湿度胁迫
Tricker et al., 2012
ZmMI1, Ac/Ds
玉米(Zea mays)
低温胁迫
Steward et al., 2002
Tam3
金鱼草(Antirrhinum majus)
低温胁迫
Hashida et al., 2006
NRPD2
拟南芥(A. thaliana)
高温胁迫
Popova et al., 2013
2 组蛋白修饰2.1 植物体内的组蛋白修饰 组蛋白修饰是表观遗传调控研究领域的热点.核小体是染色质的基本组成单位, 由约146 bp的DNA缠绕各两分子的核心组蛋白H2A、H2B、H3和H4形成的八聚体构成(Luger et al., 1997).伸出核小体外的核心组蛋白的N端可被共价修饰, 这些修饰主要包括乙酰化、甲基化、磷酸化、泛素化、生物素化、ADP核糖基化以及类泛素化等(Strahl and Allis, 2000).核心组蛋白N端赖氨酸可被组蛋白乙酰化酶HATs (histone acetyltransferase)和组蛋白去乙酰化酶HDACs (histone deacetylation transferase)进行乙酰化和去乙酰化修饰.在拟南芥中, HATs分为4个不同的家族, 研究表明HATs在植物发育(Bertrand et al., 2003; Long et al., 2006)和胁迫应答(Pavangadkar et al., 2010)中具有重要作用.真核生物的HDACs包含RP- D3 (Reduced Potassium Deficiency 3)、SIR2 (Silent Information Regulator 2)和HD2 (TypeIIHDAC) 3个主要蛋白家族, 其中HD2是植物特有的HDACs (Pan- dey et al., 2002).在拟南芥中含有10个RPD3/HDA1去乙酰化酶家族蛋白(Alinsug et al., 2009), 其中HDA6能影响转基因的表达(Murfett et al., 2001)、DNA甲基化(Aufsatz et al., 2002)以及rRNA基因的调控(Probst et al., 2004).同时研究发现, HDA19能与WRKY转录因子结合, 从而调控植物的基本防御反应(Kim et al., 2008).另外, HDA6和HDA19在拟南芥种子萌发、盐胁迫以及ABA诱导的基因表达方面都发挥十分重要的作用(Chen et al., 2010). ...
1 2000
... 组蛋白修饰是表观遗传调控研究领域的热点.核小体是染色质的基本组成单位, 由约146 bp的DNA缠绕各两分子的核心组蛋白H2A、H2B、H3和H4形成的八聚体构成(Luger et al., 1997).伸出核小体外的核心组蛋白的N端可被共价修饰, 这些修饰主要包括乙酰化、甲基化、磷酸化、泛素化、生物素化、ADP核糖基化以及类泛素化等(Strahl and Allis, 2000).核心组蛋白N端赖氨酸可被组蛋白乙酰化酶HATs (histone acetyltransferase)和组蛋白去乙酰化酶HDACs (histone deacetylation transferase)进行乙酰化和去乙酰化修饰.在拟南芥中, HATs分为4个不同的家族, 研究表明HATs在植物发育(Bertrand et al., 2003; Long et al., 2006)和胁迫应答(Pavangadkar et al., 2010)中具有重要作用.真核生物的HDACs包含RP- D3 (Reduced Potassium Deficiency 3)、SIR2 (Silent Information Regulator 2)和HD2 (TypeIIHDAC) 3个主要蛋白家族, 其中HD2是植物特有的HDACs (Pan- dey et al., 2002).在拟南芥中含有10个RPD3/HDA1去乙酰化酶家族蛋白(Alinsug et al., 2009), 其中HDA6能影响转基因的表达(Murfett et al., 2001)、DNA甲基化(Aufsatz et al., 2002)以及rRNA基因的调控(Probst et al., 2004).同时研究发现, HDA19能与WRKY转录因子结合, 从而调控植物的基本防御反应(Kim et al., 2008).另外, HDA6和HDA19在拟南芥种子萌发、盐胁迫以及ABA诱导的基因表达方面都发挥十分重要的作用(Chen et al., 2010). ...
1 2000
... ATP依赖的染色质重塑是染色质重塑复合体利用ATP水解释放的能量来改变染色质结构, 从而调控真核生物基因表达的一种重要的表观遗传调控机制(Racki and Narlikar, 2008).ATP依赖的染色质重塑复合物主要分为4个家族: SWI/SNF家族、ISWI家族、CHD家族和INO80家族.SWI/SNF是第1个被发现的ATP依赖的染色质重塑复合物(Sudarsanam and Winston, 2000).SWI/SNF复合物通过改变染色质结构在DNA修饰、重组以及基因表达中发挥重要作用(Geiman and Robertson, 2002).研究表明, SWI/ SNF复合物也参与了环境胁迫对植物生长发育的调控.例如, 在拟南芥中, 过表达SNF2/Brahma类型的染色质重塑蛋白AtCHR12, 该过表达植株在干旱和高温等胁迫下表现出花芽以及初生茎生长停滞现象(Mlynárová et al., 2007).同时, 有研究表明, SWI/SNF和CHD复合物都参与了ABA调控的拟南芥种子萌发及生长等过程(Perruc et al., 2007; Saez et al., 2008). ...
1 2007
... 在植物中, 小RNA (sRNA)是不具有编码功能的、对基因表达有调控作用的一类RNA分子, 其主要通过调节mRNA的稳定性、沉默基因转录或参与DNA甲基化等过程参与植物的生长发育(Ramachandran and Chen, 2008).因此, 小RNA也是一种重要的表观遗传调控因子.随着研究的不断深入, 对植物中sRNA的认识也有了突破性进展.以水稻为例, 研究发现, 在水稻中存在上百种sRNA, 并且在不同的生长条件下, 不同水稻品种的不同组织中sRNA的表达水平明显不同(Chen and Wu, 2010; He et al., 2010; Zhang et al., 2014).目前, 在植物中研究较多的是siRNA (small interfering RNA)和miRNA (microRNA).si- RNA是在DCL (Dicer-like protein)酶作用下由长双链RNA前体加工而来, 目前发现参与基因转录和转录后调控的内源性siRNA有nat-siRNA、ta-siRNA和hc-siRNA等(Mallory and Vaucheret, 2006).miRNA是内源基因编码的一类能与靶mRNA互补配对的一种单链小RNA, 主要参与基因的转录后调控.大量研究表明, siRNA和miRNA在植物生长发育和非生物胁迫响应过程中发挥重要作用(Phillips et al., 2007; Sunkar et al., 2007). ...
1 2005
... 为了能够在胁迫条件下生存, 植物进化出复杂的机制来感知外部信号, 从而对环境变化做出最佳反应(图1).植物激素(如水杨酸(SA)、茉莉酸(JA)、乙烯(ET)和脱落酸(ABA)等内源性分子)在调节植物对生物以及非生物胁迫的抵御过程中扮演着重要角色(Bostock, 2005; Lorenzo and Solano, 2005; Mauch- Mani and Mauch, 2005).这些激素信号可通过MAP激酶级联反应调控转录因子(如MYC、MYB、NAC、ZF和HSF等)参与胁迫相关基因的表达调控(Fujita et al., 2006).同时, 有研究者认为活性氧(ROS)的产生是植物抵御生物胁迫与非生物胁迫共有的关键过程(Torres and Dangl, 2005).除转录因子的调控, 通过染色质结构变化调控基因转录也是植物响应胁迫的重要方式, 是表观遗传机制调控植物应答外界胁迫的主要体现. ...
2 2012
... Abiotic stress responsive genes regulated by DNA-methylation in plants
基因
物种
胁迫类型
参考文献
Asr1, Asr2
番茄(Solanum lycopersicum)
干旱胁迫
González et al., 2011; 2013
NtGPDL
烟草(Nicotiana tabaccum cv. ‘Xanthi nc’)
低温胁迫
Choi and Sano, 2007
OsMYB91
水稻(Oryza sativa)
盐胁迫
Zhu et al., 2015
Glyma11g02400 Glyma16g27950 Glyma20g30840
大豆(Glycine max)
盐胁迫
Song et al., 2012
SPCH, FAMA
拟南芥(Arabidopsis thaliana)
湿度胁迫
Tricker et al., 2012
ZmMI1, Ac/Ds
玉米(Zea mays)
低温胁迫
Steward et al., 2002
Tam3
金鱼草(Antirrhinum majus)
低温胁迫
Hashida et al., 2006
NRPD2
拟南芥(A. thaliana)
高温胁迫
Popova et al., 2013
2 组蛋白修饰2.1 植物体内的组蛋白修饰 组蛋白修饰是表观遗传调控研究领域的热点.核小体是染色质的基本组成单位, 由约146 bp的DNA缠绕各两分子的核心组蛋白H2A、H2B、H3和H4形成的八聚体构成(Luger et al., 1997).伸出核小体外的核心组蛋白的N端可被共价修饰, 这些修饰主要包括乙酰化、甲基化、磷酸化、泛素化、生物素化、ADP核糖基化以及类泛素化等(Strahl and Allis, 2000).核心组蛋白N端赖氨酸可被组蛋白乙酰化酶HATs (histone acetyltransferase)和组蛋白去乙酰化酶HDACs (histone deacetylation transferase)进行乙酰化和去乙酰化修饰.在拟南芥中, HATs分为4个不同的家族, 研究表明HATs在植物发育(Bertrand et al., 2003; Long et al., 2006)和胁迫应答(Pavangadkar et al., 2010)中具有重要作用.真核生物的HDACs包含RP- D3 (Reduced Potassium Deficiency 3)、SIR2 (Silent Information Regulator 2)和HD2 (TypeIIHDAC) 3个主要蛋白家族, 其中HD2是植物特有的HDACs (Pan- dey et al., 2002).在拟南芥中含有10个RPD3/HDA1去乙酰化酶家族蛋白(Alinsug et al., 2009), 其中HDA6能影响转基因的表达(Murfett et al., 2001)、DNA甲基化(Aufsatz et al., 2002)以及rRNA基因的调控(Probst et al., 2004).同时研究发现, HDA19能与WRKY转录因子结合, 从而调控植物的基本防御反应(Kim et al., 2008).另外, HDA6和HDA19在拟南芥种子萌发、盐胁迫以及ABA诱导的基因表达方面都发挥十分重要的作用(Chen et al., 2010). ... ... 迄今已有许多研究证明, 小RNA在调控植物响应非生物胁迫中扮演着十分重要的角色(表3).小RNA常通过RdDM (RNA-directed DNA methylation)途径影响DNA甲基化修饰, 进而响应非生物胁迫.Huang等(2016)发现, SlAGO4A作为RNA介导的DNA甲基化途径的重要因子参与了番茄(Solanum lycopersicum)应答干旱和高盐胁迫的过程.相比野生型, SlAGO4A下调的转基因番茄植株表现出对干旱和高盐的耐受性.同样, 在研究RdDM途径的过程中, Naydenov等(2015)发现RdDM途径中的2个植物特异的RNA聚合酶PolIV和PolV参与了高温胁迫下DRM2基因的表达调控, 并且在PolIV和PolV的缺失突变体中, ROS基因的表达也受到抑制.Tricker等(2012)在研究气孔发育时, 发现siRNA参与了拟南芥气孔发育的2个关键基因SPECHLESS和FAMA在低湿度胁迫下的DNA甲基化过程, 从而调节拟南芥对湿度改变的适应.通常情况下, 当植物处于不良环境时, 体内并非仅有某一特定miRNA的表达发生变化, 而是大量的miRNAs表达都发生改变.例如, 在水稻中, 干旱诱导下有mi- R170和miR171等11个sRNA表达下调, 有miR474和miR845等8个sRNA表达上调(Zhou et al., 2010); 野生二粒小麦(Triticum turgidum ssp. dicoccoides)在干旱胁迫下有13个miRNAs会受到不同程度的调控(Kantar et al., 2011).许多研究表明, miRNA相关基因的突变都会对植物的叶片发育产生影响.拟南芥中编码NFYA5转录因子的基因在干旱胁迫下能通过ABA依赖途径被miR169靶向调控, 敲除突变体nfya5和miR169的过表达植株都表现出叶片失水加快以及对干旱胁迫十分敏感等表型.基因芯片分析发现, NFYA5对许多应答干旱胁迫的基因具调控作用(Li et al., 2008).在水稻中, miRNA319基因家族有2个成员, 即Osa-MIR319a和Osa-MIR319b.其过表达植株相比野生型都具有较宽的叶片, 并且具有较强的低温耐受性.进一步研究发现, miR319的靶基因OsPCF5和OsPCF8的RNAi转基因植物也表现出同样的低温耐受性, 表明miR319具有调控植物适应低温胁迫的重要作用(Yang et al., 2013).综上, 目前的研究表明, 在非生物胁迫过程中, 很多小RNA的表达都会发生改变, 但是哪些小RNA才是真正参与胁迫应答调控的直接分子, 尚待进一步研究. ...
2 2006
... Histone modification related genes involved in abiotic stresses in plants
... 迄今已有许多研究证明, 小RNA在调控植物响应非生物胁迫中扮演着十分重要的角色(表3).小RNA常通过RdDM (RNA-directed DNA methylation)途径影响DNA甲基化修饰, 进而响应非生物胁迫.Huang等(2016)发现, SlAGO4A作为RNA介导的DNA甲基化途径的重要因子参与了番茄(Solanum lycopersicum)应答干旱和高盐胁迫的过程.相比野生型, SlAGO4A下调的转基因番茄植株表现出对干旱和高盐的耐受性.同样, 在研究RdDM途径的过程中, Naydenov等(2015)发现RdDM途径中的2个植物特异的RNA聚合酶PolIV和PolV参与了高温胁迫下DRM2基因的表达调控, 并且在PolIV和PolV的缺失突变体中, ROS基因的表达也受到抑制.Tricker等(2012)在研究气孔发育时, 发现siRNA参与了拟南芥气孔发育的2个关键基因SPECHLESS和FAMA在低湿度胁迫下的DNA甲基化过程, 从而调节拟南芥对湿度改变的适应.通常情况下, 当植物处于不良环境时, 体内并非仅有某一特定miRNA的表达发生变化, 而是大量的miRNAs表达都发生改变.例如, 在水稻中, 干旱诱导下有mi- R170和miR171等11个sRNA表达下调, 有miR474和miR845等8个sRNA表达上调(Zhou et al., 2010); 野生二粒小麦(Triticum turgidum ssp. dicoccoides)在干旱胁迫下有13个miRNAs会受到不同程度的调控(Kantar et al., 2011).许多研究表明, miRNA相关基因的突变都会对植物的叶片发育产生影响.拟南芥中编码NFYA5转录因子的基因在干旱胁迫下能通过ABA依赖途径被miR169靶向调控, 敲除突变体nfya5和miR169的过表达植株都表现出叶片失水加快以及对干旱胁迫十分敏感等表型.基因芯片分析发现, NFYA5对许多应答干旱胁迫的基因具调控作用(Li et al., 2008).在水稻中, miRNA319基因家族有2个成员, 即Osa-MIR319a和Osa-MIR319b.其过表达植株相比野生型都具有较宽的叶片, 并且具有较强的低温耐受性.进一步研究发现, miR319的靶基因OsPCF5和OsPCF8的RNAi转基因植物也表现出同样的低温耐受性, 表明miR319具有调控植物适应低温胁迫的重要作用(Yang et al., 2013).综上, 目前的研究表明, 在非生物胁迫过程中, 很多小RNA的表达都会发生改变, 但是哪些小RNA才是真正参与胁迫应答调控的直接分子, 尚待进一步研究. ... ... sRNA and sRNA-regulated genes involved in abiotic stresses in plants
基因
物种
胁迫类型
参考文献
SlAGO4A
番茄(Solanum lycopersicum)
干旱胁迫, 盐胁迫
Huang et al., 2016
19 different miRNAs
水稻(Oryza sativa)
干旱胁迫
Zhou et al., 2010
13 different miRNAs
野生二粒小麦(Triticum turgidum ssp. dicoccoides)
干旱胁迫
Kantar et al., 2011
CSD1, CSD2, CCS
拟南芥(Arabidopsis thaliana )
高温胁迫
Guan et al., 2013
NFYA5
拟南芥(A. thaliana )
干旱胁迫
Li et al., 2008
OsPCF5, OsPCF8
水稻(O. sativa)
低温胁迫
Yang et al., 2013
GmNFYA3
大豆(Glycine max )
干旱胁迫
Ni et al., 2013
Phvul.010g120700
菜豆(Phaseolus vulgaris)
干旱胁迫
Sosa-Valencia et al., 2017
SlNF-YA1/2/3, SlMRP1
番茄(S. lycopersicum)
干旱胁迫
Zhang et al., 2011a
BHLH23
鹰嘴豆(Cicer arietinum)
干旱胁迫
Hajyzadeh et al., 2015
4 染色质重塑与非生物胁迫 ATP依赖的染色质重塑是染色质重塑复合体利用ATP水解释放的能量来改变染色质结构, 从而调控真核生物基因表达的一种重要的表观遗传调控机制(Racki and Narlikar, 2008).ATP依赖的染色质重塑复合物主要分为4个家族: SWI/SNF家族、ISWI家族、CHD家族和INO80家族.SWI/SNF是第1个被发现的ATP依赖的染色质重塑复合物(Sudarsanam and Winston, 2000).SWI/SNF复合物通过改变染色质结构在DNA修饰、重组以及基因表达中发挥重要作用(Geiman and Robertson, 2002).研究表明, SWI/ SNF复合物也参与了环境胁迫对植物生长发育的调控.例如, 在拟南芥中, 过表达SNF2/Brahma类型的染色质重塑蛋白AtCHR12, 该过表达植株在干旱和高温等胁迫下表现出花芽以及初生茎生长停滞现象(Mlynárová et al., 2007).同时, 有研究表明, SWI/SNF和CHD复合物都参与了ABA调控的拟南芥种子萌发及生长等过程(Perruc et al., 2007; Saez et al., 2008). ...
2 2015
... DNA甲基化修饰在调节植物响应外界环境胁迫中发挥了非常重要的作用.最近几年的研究表明, 全基因组DNA甲基化修饰的改变与植物响应非生物胁迫密切相关(Boyko et al., 2010; Karan et al., 2012; Wang et al., 2014).许多与DNA甲基化修饰相关的基因参与了植物的非生物胁迫应答过程(表1).在植物响应高盐、干旱、温度和重金属等非生物胁迫过程中, 植物的特定基因或整个基因组水平的DNA甲基化修饰都会发生改变, 从而提高植物对不良环境的适应性, 确保植物在逆境条件下的生长发育.通常情况下, 非生物胁迫诱导的DNA甲基化修饰的改变发生在整个基因组水平.例如, Wang等(2011)以水稻(Oryza sativa) DK151和IR64为研究材料, 发现由干旱胁迫诱导的DNA甲基化位点的改变占全基因组DNA甲基化位点的12%左右, 这些甲基化的变化有29%在胁迫解除时仍然被保留; 并且干旱诱导的DNA甲基化还表现出组织及发育时期的特异性.Ferreira等(2015)在研究盐胁迫下DNA甲基化水平的变化时, 发现盐胁迫耐受型水稻材料Pokkali和敏感型水稻材料IR29基因组全局性的DNA甲基化水平在盐胁迫下发生了明显改变, 并且这种DNA甲基化水平的改变具有基因型及组织特异性.此外, 非生物胁迫下, DNA甲基化水平的改变往往还会伴随其它调节机制.例如, Karan等(2012)在研究水稻响应盐胁迫时, 发现逆转录转座子、非生物胁迫应答基因以及染色质修饰因子在盐胁迫下都具有不同的表达模式, 同时鉴定了不同水稻品种盐胁迫诱导的DNA甲基化模式.而对于水稻而言, 在水稻根细胞内相关基因的去甲基化修饰对其响应盐胁迫也具有重要作用(Wang et al., 2011).Zhu等(2015)在研究水稻R2R3类型的MYB转录因子OsMYB91时, 发现OsMYB91的表达受非生物胁迫诱导, 特别是盐胁迫.进一步研究发现, 该基因启动子区域的DNA甲基化水平改变是导致该基因在胁迫下表达发生变化的主要原因.除了拟南芥和水稻等常见模式植物, 人们也对其它植物响应胁迫时的DNA甲基化修饰进行了研究.例如, Bharti等(2015)在研究盐胁迫下, 类黄酮生物合成和抗氧化途径的酶基因是否受到表观遗传调控时, 发现这些基因启动子区域甲基化状态改变使得相关基因的表达水平升高, 从而导致过表达AtROS1的转基因烟草(Nicotiana tabacum)表现出对盐胁迫的耐受性.Lu等(2017)利用Me-DIP测序技术对棉花(Gossypium hirsutum)在盐胁迫下全基因组的DNA甲基化水平进行了研究, 为理解棉花盐胁迫反应的表观遗传修饰变化提供了有价值的信息.同样, 也有研究发现, 苜蓿(Medicago spp.)在盐胁迫下, 全基因组水平的DNA甲基化程度增加, 特别是在高盐胁迫下尤为明显(Al-Lawati et al., 2016).目前, 有关DNA甲基化修饰水平影响植物响应非生物胁迫的具体机制还有待深入研究. ... ... Abiotic stress responsive genes regulated by DNA-methylation in plants
基因
物种
胁迫类型
参考文献
Asr1, Asr2
番茄(Solanum lycopersicum)
干旱胁迫
González et al., 2011; 2013
NtGPDL
烟草(Nicotiana tabaccum cv. ‘Xanthi nc’)
低温胁迫
Choi and Sano, 2007
OsMYB91
水稻(Oryza sativa)
盐胁迫
Zhu et al., 2015
Glyma11g02400 Glyma16g27950 Glyma20g30840
大豆(Glycine max)
盐胁迫
Song et al., 2012
SPCH, FAMA
拟南芥(Arabidopsis thaliana)
湿度胁迫
Tricker et al., 2012
ZmMI1, Ac/Ds
玉米(Zea mays)
低温胁迫
Steward et al., 2002
Tam3
金鱼草(Antirrhinum majus)
低温胁迫
Hashida et al., 2006
NRPD2
拟南芥(A. thaliana)
高温胁迫
Popova et al., 2013
2 组蛋白修饰2.1 植物体内的组蛋白修饰 组蛋白修饰是表观遗传调控研究领域的热点.核小体是染色质的基本组成单位, 由约146 bp的DNA缠绕各两分子的核心组蛋白H2A、H2B、H3和H4形成的八聚体构成(Luger et al., 1997).伸出核小体外的核心组蛋白的N端可被共价修饰, 这些修饰主要包括乙酰化、甲基化、磷酸化、泛素化、生物素化、ADP核糖基化以及类泛素化等(Strahl and Allis, 2000).核心组蛋白N端赖氨酸可被组蛋白乙酰化酶HATs (histone acetyltransferase)和组蛋白去乙酰化酶HDACs (histone deacetylation transferase)进行乙酰化和去乙酰化修饰.在拟南芥中, HATs分为4个不同的家族, 研究表明HATs在植物发育(Bertrand et al., 2003; Long et al., 2006)和胁迫应答(Pavangadkar et al., 2010)中具有重要作用.真核生物的HDACs包含RP- D3 (Reduced Potassium Deficiency 3)、SIR2 (Silent Information Regulator 2)和HD2 (TypeIIHDAC) 3个主要蛋白家族, 其中HD2是植物特有的HDACs (Pan- dey et al., 2002).在拟南芥中含有10个RPD3/HDA1去乙酰化酶家族蛋白(Alinsug et al., 2009), 其中HDA6能影响转基因的表达(Murfett et al., 2001)、DNA甲基化(Aufsatz et al., 2002)以及rRNA基因的调控(Probst et al., 2004).同时研究发现, HDA19能与WRKY转录因子结合, 从而调控植物的基本防御反应(Kim et al., 2008).另外, HDA6和HDA19在拟南芥种子萌发、盐胁迫以及ABA诱导的基因表达方面都发挥十分重要的作用(Chen et al., 2010). ...

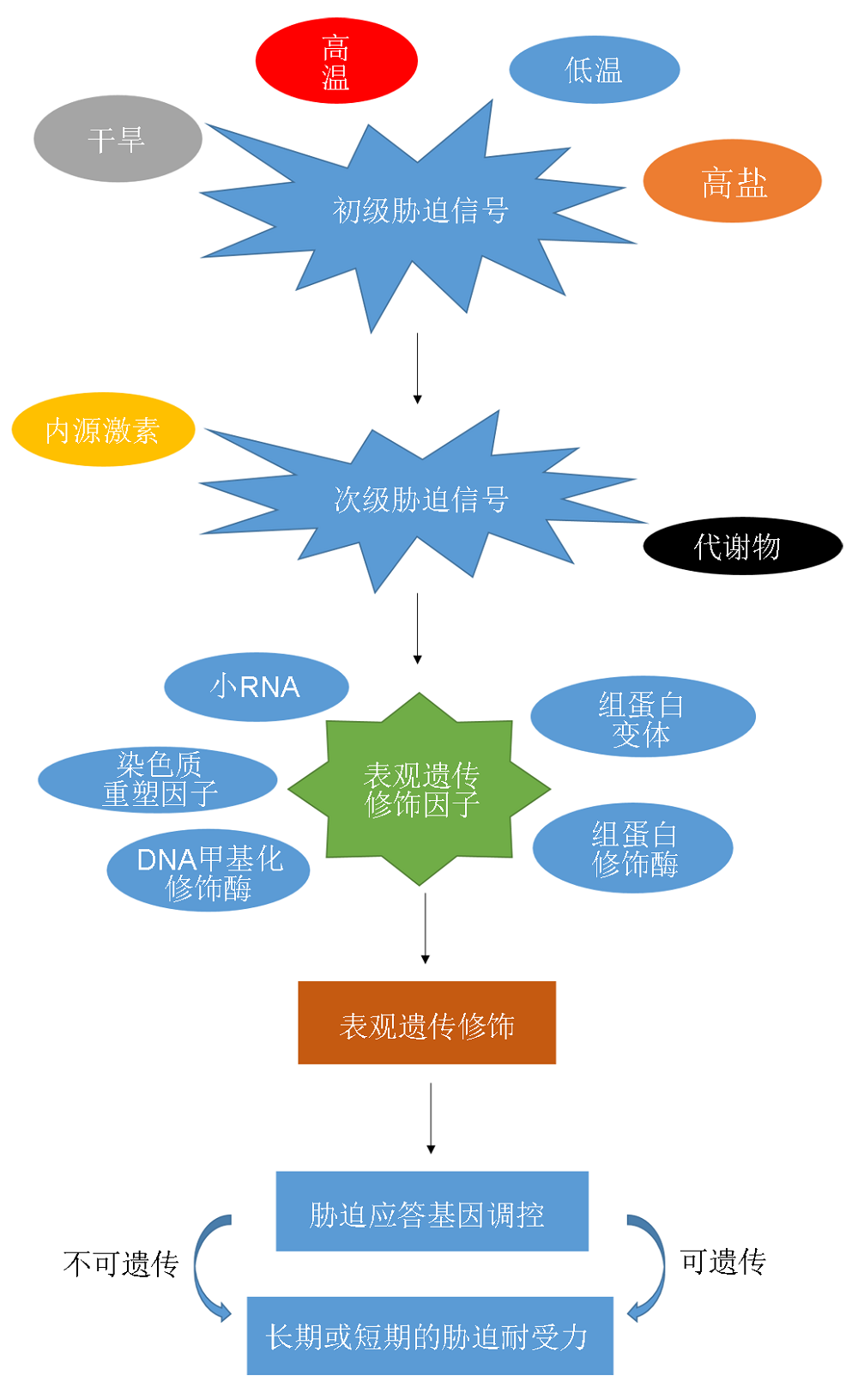

{kind=link}
{kind=link}