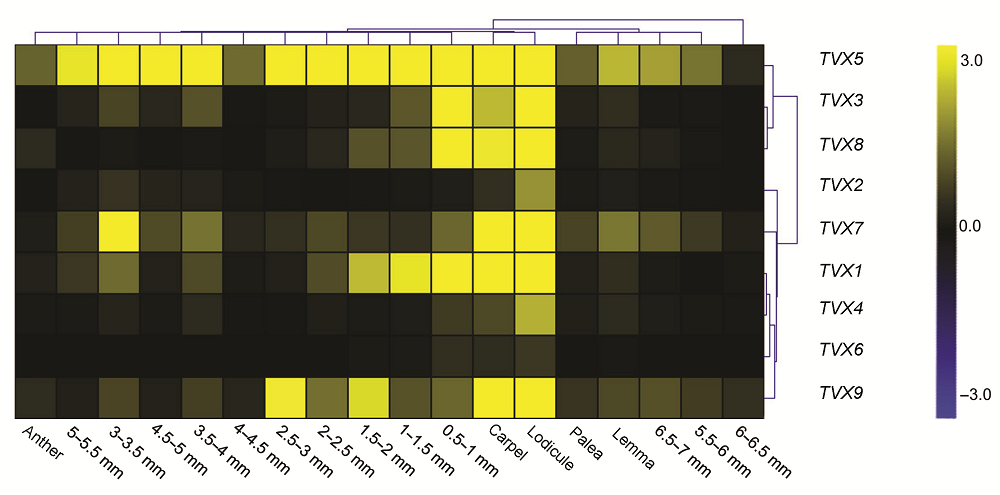
Abstract: Alternative polyadenylation (APA) is one of the important regulatory pathways of eukaryotic gene expression. By forming different lengths of 3' untranslated regions, APA affects the stability, localization and translation efficiency of mRNA and increases the complexity of transcripts. The expression of the Arabidopsis gene increased polyadenylation of BONSAI methylation 1 (IBM1) is regulated by chromatin regulatory factor enhanced downy mildew 2 (EDM2), which can further affect the CHG methylation level of the Arabidopsis thaliana genome. However, whether such regulatory mechanisms exist in other species is unknown. To answer this question, we selected OsJMJ718, an IBM1 homologous gene of rice, for research. By using bioinformatics analysis and 3'RACE experiments, we found that homologous genes of IBM1 also had APA modification. Among them, the OsJMJ718 gene may have 9 alternative polyadenylation sequences. Further sequence alignment analysis revealed that the 3' terminal sequence of OsJMJ718 in the existing Japonica genome in the NCBI database may be different from that of the other ecotype genome sequences, such as 9522 and Minghui 63. Quantitative real-time PCR showed that the 9 transcripts of OsJMJ718 present diverse dynamic expression patterns in different stages of rice reproductive development. The expression of OsJMJ718-TVX5 was higher than that of other transcripts. In summary, this work provides information for APA sequences of OsJMJ718, and the expression pattern analysis of these transcripts would also help further study of the mechanism of APA and biological function of OsJMJ718.
1.2 试剂实验所用Trizol Reagent购自Invitrogen公司。3'RA- CE Kit购自TAKARA clontech公司。SYBR Green Realtime PCR Master MIX购自天根生化科技有限公司。反转录试剂盒PrimeScript RT reagent Kit with gDNA Eraser购自TAKARA公司。本研究所需引物(表1)由上海捷瑞生物工程有限公司合成。DNA测序由华大和桑尼公司提供服务。 表1 Table 1 表1 表1 本研究所用引物 Table 1 Primers used in this research
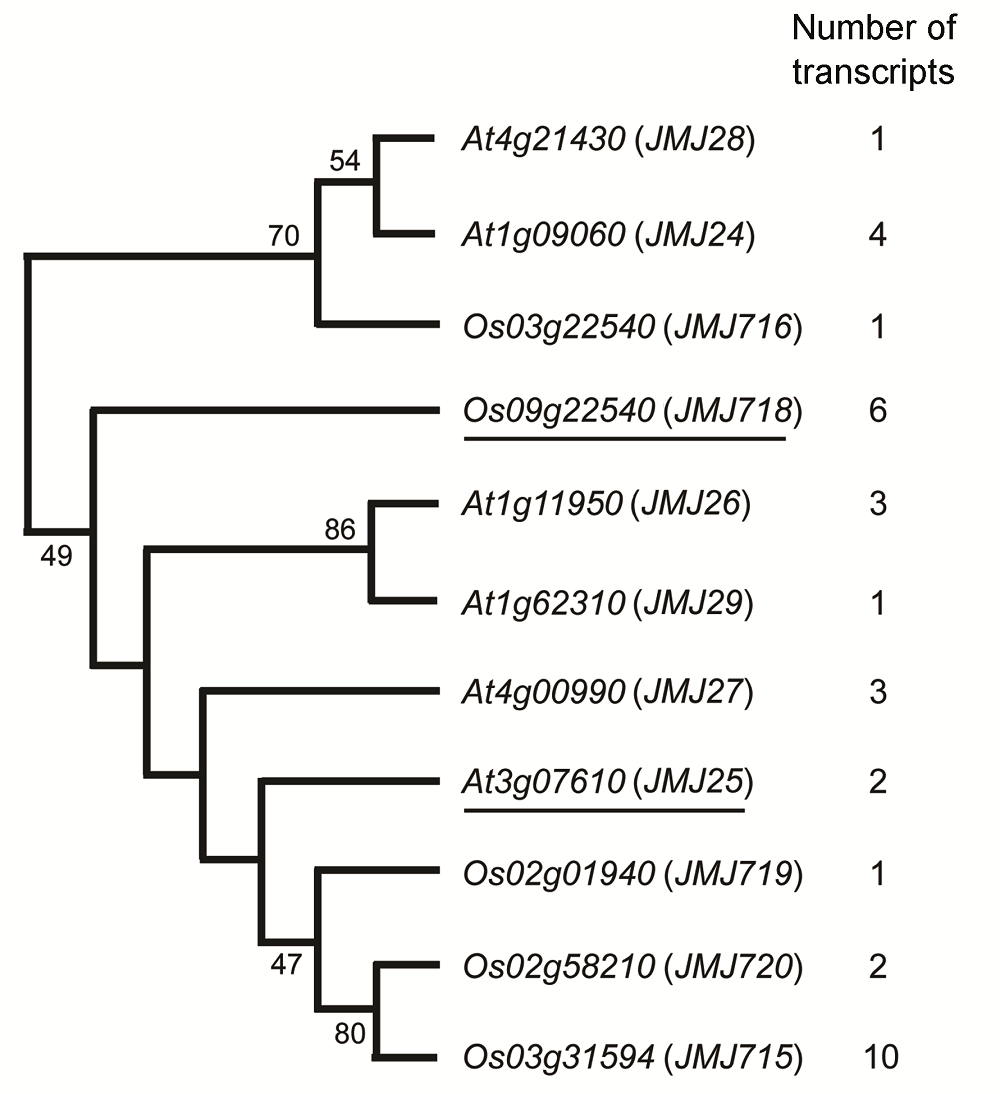
2 结果与讨论2.1 拟南芥和水稻JHDM2家族基因mRNA可选择性多聚腺苷酸化分析拟南芥IBM1 (At3g07610/JMJ25)基因属于JHDM2基因家族(Sun and Zhou, 2008)。为深入分析IBM1基因mRNA可选择性多聚腺苷酸化现象是否普遍存在, 我们首先在NCBI和TAIR (The Arabidopsis Information Resource, https://www.arabidopsis.org/index.jsp)数据库中进行转录本BLAST分析。结果表明, 拟南芥和水稻JHDM2基因家族中的其它基因也存在mRNA可选择性多聚腺苷酸化现象。例如, 拟南芥At1g09060 (JMJ24)基因有4个转录本, At1g11950基因有3个转录本, At4g00990基因有3个转录本, 多于IBM1基因转录本数目(图1); 水稻IBM1同源基因也存在类似现象, OsJMJ718基因有6个转录本, OsJMJ715基因有10个转录本(图1)。以上结果暗示, 水稻和拟南芥IBM1类基因可能存在同样的mRNA可选择性多聚腺苷酸化调控机制。 图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-594/img_1.png<b>图1</b> 拟南芥和水稻中JHDM2家族基因mRNA可选择性多聚腺苷酸化加工序列数目<br/><i>Os09g22540</i> (<i>JMJ718</i>)为研究对象<i>OsJMJ718</i>; <i>At3g07610 </i>(<i>JMJ25</i>)是拟南芥<i>IBM1</i>, <i>OsJMJ718</i>的同源基因。<br/><b>Figure 1</b> The number of alternative polyadenylation sequences of JHDM2 family genes in <i>Arabidopsis thaliana</i> and rice<br/><i>Os09g22540</i> (<i>JMJ718</i>) is <i>OsJMJ718</i>, the target sequence of this study; <i>At3g07610</i> (<i>JMJ25</i>) is <i>IBM1</i>, which is the homo- logous gene of <i>OsJMJ718 </i>in <i>Arabidopsis thaliana</i>. Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-594/img_1.png<b>图1</b> 拟南芥和水稻中JHDM2家族基因mRNA可选择性多聚腺苷酸化加工序列数目<br/><i>Os09g22540</i> (<i>JMJ718</i>)为研究对象<i>OsJMJ718</i>; <i>At3g07610 </i>(<i>JMJ25</i>)是拟南芥<i>IBM1</i>, <i>OsJMJ718</i>的同源基因。<br/><b>Figure 1</b> The number of alternative polyadenylation sequences of JHDM2 family genes in <i>Arabidopsis thaliana</i> and rice<br/><i>Os09g22540</i> (<i>JMJ718</i>) is <i>OsJMJ718</i>, the target sequence of this study; <i>At3g07610</i> (<i>JMJ25</i>) is <i>IBM1</i>, which is the homo- logous gene of <i>OsJMJ718 </i>in <i>Arabidopsis thaliana</i>.
图1 拟南芥和水稻中JHDM2家族基因mRNA可选择性多聚腺苷酸化加工序列数目 Os09g22540 (JMJ718)为研究对象OsJMJ718; At3g07610 (JMJ25)是拟南芥IBM1, OsJMJ718的同源基因。 Figure 1 The number of alternative polyadenylation sequences of JHDM2 family genes in Arabidopsis thaliana and rice Os09g22540 (JMJ718) is OsJMJ718, the target sequence of this study; At3g07610 (JMJ25) is IBM1, which is the homo- logous gene of OsJMJ718 in Arabidopsis thaliana.
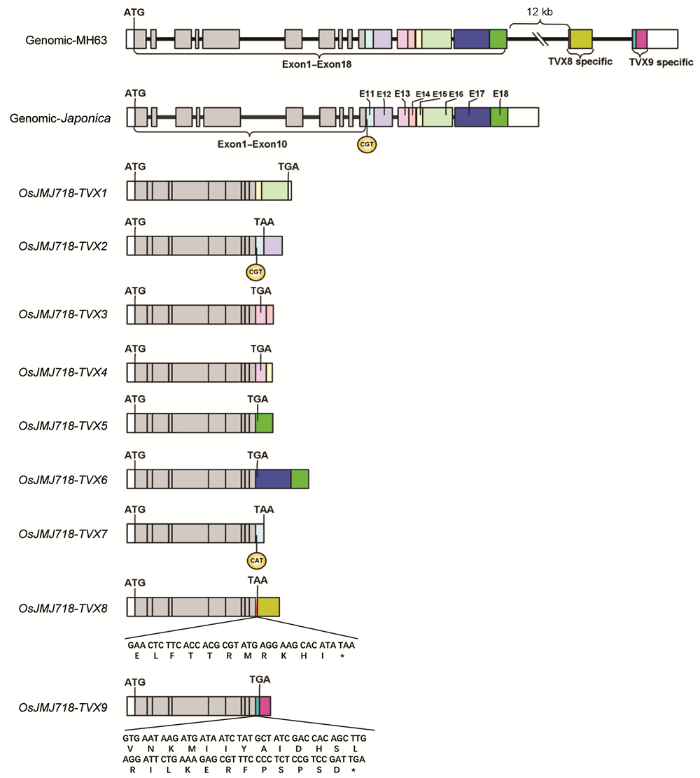
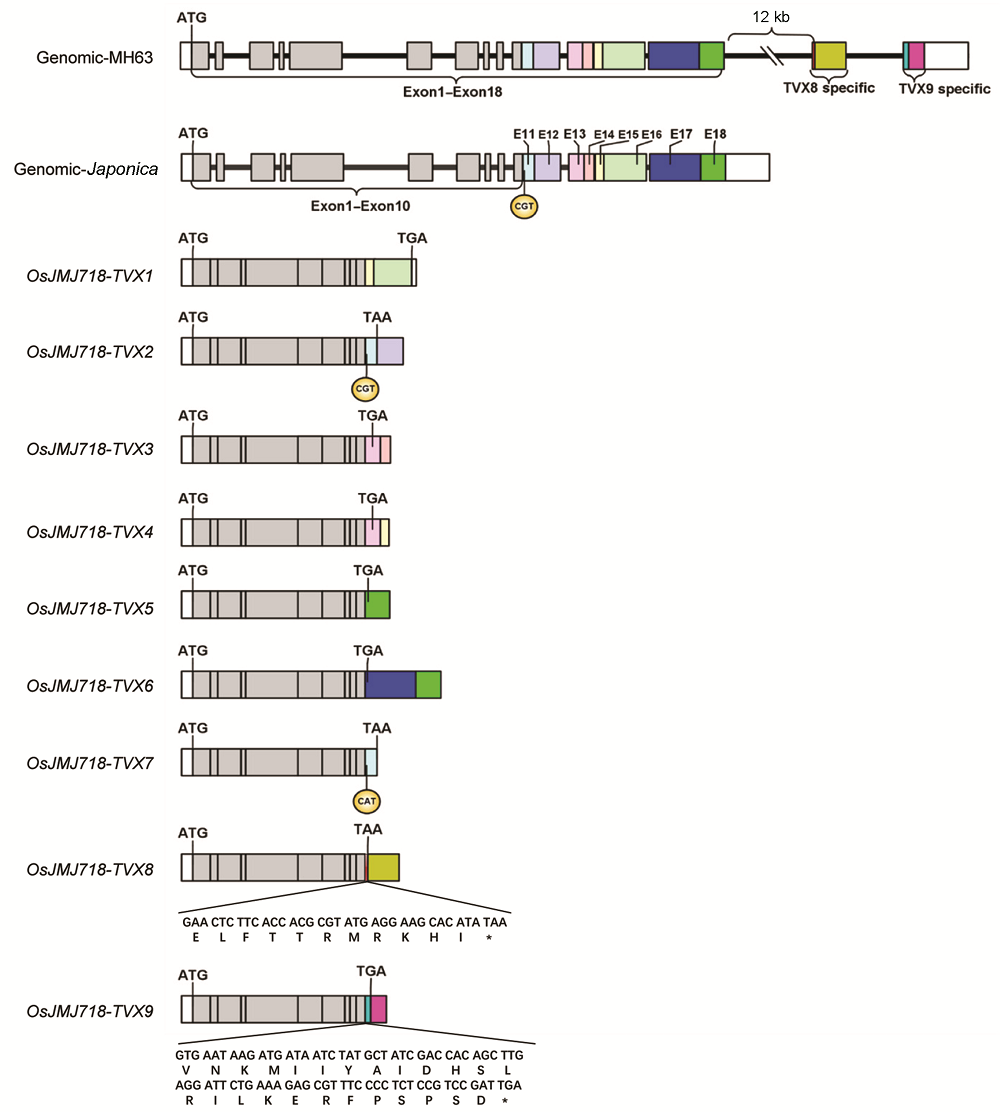
2.2 OsJMJ718转录本序列分析及3'RACE克隆为了进一步确认水稻中IBM1类基因存在mRNA可选择性多聚腺苷酸化修饰, 我们首先对从NCBI网站BLAST分析获得的6个转录本进行了序列比对。如图2所示, OsJMJ718-TVX1、TVX2、TVX3、TVX4、TVX5和TVX6六个转录本具有共同的5'UTR和前10个外显子序列, 但第11个外显子、终止密码子和3'UTR序列具有明显差异。不同转录本的第11个外显子、终止密码子和3'UTR序列分布在参考基因组日本晴基因组的不同位置, 为方便区分, 我们根据其在序列上的位置分别命名为E11-E18 (图2)。其中, TVX1转录本C端由E15和E16序列组成, 终止密码子和3'UTR均在E16序列中。TVX2转录本C端由E11及E12序列组成, 终止密码子在E11序列末端, 3'UTR在E12序列中。TVX3转录本C端由E13及E14序列组成, 终止密码子位于E13号外显子中, E13号外显子部分序列和E14号序列作为3'UTR区域。TVX4转录本C端由E13及E15序列组成, 终止密码子位于E13号外显子中, E13号外显子部分序列和E15号外显子作为3'UTR区域。值得注意的是, TVX3与TVX4转录本编码的蛋白质序列相同, 但通过可选择性多聚腺苷酸化产生了不同的3'UTR。TVX5转录本C端由E18号外显子组成, 终止密码子位于E18号外显子中, E18号外显子末端序列是3'UTR区域。TVX6转录本C端由E17及E18号外显子组成, 终止密码子位于E17号外显子中, E17号外显子部分序列及E18号外显子组成3'UTR区域。 图2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-594/img_2.png<b>图2</b> OsJMJ718-TVX转录本序列<br/>9个转录本分别命名为<i>TVX1</i>-<i>TVX9</i>; Exon1-Exon10表示各转录本共有外显子序列, Exon11-Exon18为不同颜色表示基因组上的8个外显子通过可选择性多聚腺苷酸化被特异加工至<i>TVX1</i>-<i>TVX7</i>转录本。其中, <i>TVX8</i>和<i>TVX9</i>的3'端序列仅存在于明辉63 (MH63)基因组中。* 表示终止密码子<br/><b>Figure 2</b> Sequence analysis of <i>OsJMJ718-TVX</i> transcripts<br/>Nine transcripts were named as <i>TVX1</i>-<i>TVX9</i>, respectively; Exon1-Exon10 represent common exon sequences for each transcript; Exon11-Exon18 were marked in different color, the 8 exons were processed in <i>TVX1</i>-<i>TVX7</i> by alternative polyadenylation. However, 3’ terminal sequences of <i>TVX8</i> and <i>TVX9</i> are only exist in MH63 genomic sequences. * indicates stop codon Figure 2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-594/img_2.png<b>图2</b> OsJMJ718-TVX转录本序列<br/>9个转录本分别命名为<i>TVX1</i>-<i>TVX9</i>; Exon1-Exon10表示各转录本共有外显子序列, Exon11-Exon18为不同颜色表示基因组上的8个外显子通过可选择性多聚腺苷酸化被特异加工至<i>TVX1</i>-<i>TVX7</i>转录本。其中, <i>TVX8</i>和<i>TVX9</i>的3'端序列仅存在于明辉63 (MH63)基因组中。* 表示终止密码子<br/><b>Figure 2</b> Sequence analysis of <i>OsJMJ718-TVX</i> transcripts<br/>Nine transcripts were named as <i>TVX1</i>-<i>TVX9</i>, respectively; Exon1-Exon10 represent common exon sequences for each transcript; Exon11-Exon18 were marked in different color, the 8 exons were processed in <i>TVX1</i>-<i>TVX7</i> by alternative polyadenylation. However, 3’ terminal sequences of <i>TVX8</i> and <i>TVX9</i> are only exist in MH63 genomic sequences. * indicates stop codon Figure 2 Sequence analysis of OsJMJ718-TVX transcripts Nine transcripts were named as TVX1-TVX9, respectively; Exon1-Exon10 represent common exon sequences for each transcript; Exon11-Exon18 were marked in different color, the 8 exons were processed in TVX1-TVX7 by alternative polyadenylation. However, 3’ terminal sequences of TVX8 and TVX9 are only exist in MH63 genomic sequences. * indicates stop codon'>
图2 OsJMJ718-TVX转录本序列 9个转录本分别命名为TVX1-TVX9; Exon1-Exon10表示各转录本共有外显子序列, Exon11-Exon18为不同颜色表示基因组上的8个外显子通过可选择性多聚腺苷酸化被特异加工至TVX1-TVX7转录本。其中, TVX8和TVX9的3'端序列仅存在于明辉63 (MH63)基因组中。* 表示终止密码子 Figure 2 Sequence analysis of OsJMJ718-TVX transcripts Nine transcripts were named as TVX1-TVX9, respectively; Exon1-Exon10 represent common exon sequences for each transcript; Exon11-Exon18 were marked in different color, the 8 exons were processed in TVX1-TVX7 by alternative polyadenylation. However, 3’ terminal sequences of TVX8 and TVX9 are only exist in MH63 genomic sequences. * indicates stop codon
SazeH, ShiraishiA, MiuraA, KakutaniT (2008). Control of genic DNA methylation by a jmjC domain-containing pro- tein in Arabidopsis thaliana. Science 319, 462-465. [本文引用: 3]
[12]
ShenYJ, VenuRC, NobutaK, WuXH, NotibalaV, DemirciC, MeyersBC, WangGL, JiGL, LiQQ (2011). Transcrip- tome dynamics through alternative polyadenylation in developmental and environmental responses in plants revealed by deep sequencing.Genome Res 21, 1478-1486. [本文引用: 1]
[13]
ShiYS (2012). Alternative polyadenylation: new insights from global analyses.RNA 18, 2105-2117. [本文引用: 1]
[14]
SimpsonGG, DijkwelPP, QuesadaV, HendersonI, DeanC (2003). FY is an RNA 3′ end-processing factor that inter- acts with FCA to control the Arabidopsis floral transition.Cell 113, 777-787. [本文引用: 1]
[15]
SunQ, ZhouDX (2008). Rice jmjC domain-containing gene JMJ706 encodes H3K9 demethylase required for floral organ development.Proc Natl Acad Sci USA 105, 13679-13684. [本文引用: 2]
[16]
WuXH, LiuM, DownieB, LiangC, JiGL, LiQQ, HuntAG (2011). Genome-wide landscape of polyadenylation in Arabidopsis provides evidence for extensive alternative polyadenylation.Proc Natl Acad Sci USA 108, 12533-12538. [本文引用: 1]
[17]
XingDH, LiQQ (2011). Alternative polyadenylation and gene expression regulation in plants.Wiley Interdiscip Rev RNA 2, 445-458. [本文引用: 2]
[18]
YaoP, PotdarAA, ArifA, RayPS, MukhopadhyayR, WillardB, XuYC, YanJ, SaidelGM, FoxPL (2012). Coding region polyadenylation generates a truncated tRNA synthetase that counters translation repression.Cell 149, 88-100. [本文引用: 1]
[19]
ZhangJX, AddepalliB, YunKY, HuntAG, XuRQ, RaoS, LiQQ, FalconeDL (2008). A polyadenylation factor subunit implicated in regulating oxidative signaling in Ara- bidopsis thaliana. PLoS One 3, e2410. [本文引用: 1]
LicatalosiDD, DarnellRB (2010). RNA processing and its regulation: global insights into biological networks.Nat Rev Genet 11, 75-87. [本文引用: 1]
[6]
LiuY, CuiSJ, WuF, YanS, LinXL, DuXQ, ChongK, SchillingS, TheissenG, MengZ (2013). Functional conservation of MIKC*-type MADS box genes in Ara- bidopsis and rice pollen maturation.Plant Cell 25, 1288-1303. [本文引用: 1]
[7]
MaLY, GuoC, LiQQ (2014). Role of alternative poly- adenylation in epigenetic silencing and antisilencing.Proc Natl Acad Sci USA 111, 9-10. [本文引用: 1]
[8]
MayrC, BartelDP (2009). Widespread shortening of 3'UTRs by alternative cleavage and polyadenylation activates onco- genes in cancer cells.Cell 138, 673-684. [本文引用: 2]
[9]
RigalM, KeveiZ, PélissierT, MathieuO (2012). DNA methylation in an intron of the IBM1 histone demethylase gene stabilizes chromatin modification patterns.EMBO J 31, 2981-2993. [本文引用: 1]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}