Advances in Plasma Membrane Ion Channels of Plant Cells
Yanqiu Tan, Shujing Sun, Jingyun Dong, Wei Xu, Lingling Wang, Yongfei Wang,*National Key Laboratory of Plant Molecular Genetics, Center for Excellence in Molecular Plant Sciences, Shanghai Institute of Plant Physiology and Ecology, Chinese Academy of Sciences, Shanghai 200032, China
Abstract Multiple ions, including cations and anions, are involved in growth, development, reproduction and stress responses as essential nutrients, osmotic components, cofactors of proteins and signaling molecules in plants. Ion channels localized in the plasma membrane and endomembrane of plant cells are essential tunnels and dynamic regulators of ion flux across biological membranes and are involved in the direct regulation of ion homeostasis in plant cells. A large number of ion channels have been identified in diverse plant species, especially in the model plant Arabidopsis, in recent years. Many ion channels are localized in the plasma membrane, and their protein structure, ion selectivity and permeability, gating patterns, activity-regulating mechanisms and the functional coordinating mechanisms between different ion channels have been analyzed. In this review, we summarize the research progress on the plasma membrane ion channels of plant cells and discuss the outstanding questions. Keywords:ions;ion flux across membrane;ion channels;plasma membrane;plant cells
PDF (5484KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 谭艳秋, 孙姝璟, 董静云, 徐伟, 王玲玲, 王永飞. 植物细胞质膜离子通道研究进展. 植物学报, 2019, 54(1): 102-118 doi:10.11983/CBB18022 Tan Yanqiu, Sun Shujing, Dong Jingyun, Xu Wei, Wang Lingling, Wang Yongfei. Advances in Plasma Membrane Ion Channels of Plant Cells. Chinese Bulletin of Botany, 2019, 54(1): 102-118 doi:10.11983/CBB18022
(A) 单个亚基拓扑结构; (B) Shaker离子通道的多种不同四聚体组成形式, 包括内向和外向质膜K+通道。S1-S6: 单亚基的第1-6个跨膜区; N/C端: 氨/羧基末端; P: 通道孔区; +: 带正电荷的电压感受器结构域; cNBD: 环核苷酸结合域 Figure 1Topological structure of a representative subunit and tetramers of Shaker family from Arabidopsis (modified from Sharma et al., 2013)
(A) Topological structure of a Shaker subunit; (B) Diverse tetramers of K+ channels composed of Shaker members, including inward and outward K+ channels. S1-S6: Transmembrane domains 1-6; N/C termini: The amino terminal and carboxyl terminal; P: Pore region; +: With positive charged voltage sensor domain; cNBD: Cyclic nucleotide binding domain.
目前, 对拟南芥Shaker家族的组织分布和功能研究得较为深入全面; 而对包括水稻(Oryza sativa)和玉米(Zea mays)在内的其它植物Shaker家族的研究相对较少。已有的研究鉴定出多个参与K+吸收、转运和气孔运动的玉米和水稻Shaker同源基因。其与拟南芥同源蛋白相比较有很多类似之处, 但也有不同。例如, 玉米KZM2通过与KZM3组成异源多聚体, 抑制K+in通道的活性和气孔运动(Gao et al., 2017)。而水稻OsKAT3通过与OsKAT2互作抑制OsKAT3的活性(Hwang et al., 2013)。这种K+通道两个同源蛋白之间的互作和调控方式与拟南芥中的AtKC1对AKT1和KAT1等的抑制作用类似(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010)。再比如, 水稻KAT2与拟南芥KAT2均作为K+in通道参与光照诱导的气孔开放, 但拟南芥气孔保卫细胞质膜K+in通道编码基因数目明显多于水稻(Pilot et al., 2001; Moon et al., 2017)。对于外向K+通道而言, 拟南芥SKOR分布于根部, 负责K+向木质部的卸载, 而GORK则主要参与拟南芥的气孔关闭, 二者的组织分布和生物学功能明显不同。而水稻的同一个Shaker蛋白同时参与气孔关闭和K+营养在根部的卸载(Nguyen et al., 2017)。因此, 加强对更多植物, 尤其是重要作物中Shaker家族的研究, 将会促进人们对植物吸收、运输和利用K+的分子机理的深入理解。
2 植物细胞质膜Ca2+通道及其活性调控机制
植物体内存在大量的钙元素。一方面钙作为细胞壁等结构组分发挥作用; 另一方面, 离子形式的钙(Ca2+)可以作为信号分子参与各种生物学反应和信号传递过程, 后者则备受关注。胞内Ca2+信号以细胞质Ca2+浓度的升高和震荡形式出现。一般认为, 胞外Ca2+内流是细胞质Ca2+信号产生所需Ca2+的主要来源, 而质膜内向Ca2+通道是胞外Ca2+内流的主要渠道。同时, Ca2+通道的活性变化通过控制胞外Ca2+内流的节奏直接调控胞内Ca2+震荡, 发挥Ca2+信号上游调控因子的作用。鉴于此, 质膜Ca2+通道受到广泛关注, 并成为动植物各信号领域的重点研究对象。科学家首先在动物细胞中发现多个Ca2+通道家族, 主要包括受环核苷酸cAMP和cGMP激活的CNG (cyclic nucleotide-gated)家族、受谷氨酸门控的GLR (glutamate receptor)家族、TRPC (transient receptor potential channel)和受胞内Ca2+库倒空激活的CRACM1/Orai1 (Zagotta and Siegelbaum, 1996; Vig and Kinet, 2007; Sobolevsky, 2015; Ambudkar et al., 2017)。通过序列比对, 多个动物细胞质膜Ca2+通道的同源基因家族先后在植物尤其是模式植物拟南芥基因组中被鉴定出来, 主要包括CNGC (cyclic nucleotide- gated channel)、GLR (glutamate receptor-like)、高渗敏感的OSCA (hyperosmolality-gated calcium- permeable channel)家族和低渗敏感的张力激活离子通道MCA (mechano-sensitive channel) (M?ser et al., 2001; Ward et al., 2009; Iida et al., 2014; Yuan et al., 2014)。
S: 跨膜区; EF: EF-手型Ca2+结合结构域; N/C端: 氨/羧基端 Figure 3Protein structure of Arabidopsis MCA1 and MCA2 (A) and a molecular mechanism of how MCA1/2 channels are activated by the tension of plasma membrane (B) (Kamano et al., 2015)
S: The transmembrane domain; EF: EF-hand Ca2+ binding domain; N/C termini: The amino terminal and carboxyl terminal
Figure 6The topological structures of a SLAC1 subunit (A) and the structure of a SLAC1 channel as a trimer (B) (Vahisalu et al., 2008; Chen et al., 2010)
除了SLAC1家族, 植物细胞质膜上还包含张力激活的阴离子通道MSL (mechano-sensitive or stret- ch-activated channels) (Crosgrove and Hedrich, 1991)。MSL为典型的张力激活的阴离子通道家族。MSL9和MSL10是最早得到电生理鉴定的MSL家族成员, 定位于细胞质膜, 其激活对质膜张力具有依赖性, 主要选择性介导阴离子的跨膜流动(Haswell et al., 2008; Maksaev and Haswell, 2012)。随后有研究表明, 拟南芥MSL10参与植物机械损伤信号反应和茉莉酸的累积(Veley et al., 2014; Zou et al., 2016)。此外, 有些拟南芥MSL成员定位在细胞器膜上, 调控细胞器的大小和形状(Veley et al., 2012)。根据氨基酸残基序列预测, MSL蛋白存在多个跨膜区, 具备形成离子通道的基本结构特点。而质膜受到拉伸时对MSL的激活是通过机械拉力直接改变MSL蛋白的结构, 使离子通过的孔被拉开。但这基本上还停留在理论推测阶段, 缺乏有力的直接实验证据支持。总体来讲, 目前对MSL家族的生物学功能还了解较少, 其组成的离子通道蛋白结构、感知生物膜张力的分子机理和生物学功能等还有待深入研究。
AcheP, BeckerD, IvashikinaN, DietrichP, RoelfsemaMRG, HedrichR ( 2000). GORK, a delayed outward rectifier expressed in guard cells of Arabidopsis thaliana, is a K +-selective, K +-sensing ion channel. FEBS Lett 486, 93-98. [本文引用: 1]
AmbudkarIS, de SouzaLB, OngHL ( 2017). TRPC1, Orai1, and STIM1 in SOCE: friends in tight spaces Cell Calcium 63, 33-39. DOI:10.1016/j.ceca.2016.12.009URLPMID:28089266 [本文引用: 1] Store-operated calcium entry (SOCE) is a ubiquitous Ca(2+) entry pathway that is activated in response to depletion of ER-Ca(2+) stores and critically controls the regulation of physiological functions in miscellaneous cell types. The transient receptor potential canonical 1 (TRPC1) is the first member of the TRPC channel subfamily to be identified as a molecular component of SOCE. While TRPC1 has been shown to contribute to SOCE and regulate various functions in many cells, none of the reported TRPC1-mediated currents resembled ICRAC, the highly Ca(2+)-selective store-dependent current first identified in lymphocytes and mast cells. Almost a decade after the cloning of TRPC1 two proteins were identified as the primary components of the CRAC channel. The first, STIM1, is an ER-Ca(2+) sensor protein involved in activating SOCE. The second, Orai1 is the pore-forming component of the CRAC channel. Co-expression of STIM1 and Orai1 generated robust ICRAC. Importantly, STIM1 was shown to also activate TRPC1 via its C-terminal polybasic domain, which is distinct from its Orai1-activating domain, SOAR. In addition, TRPC1 function critically depends on Orai1-mediated Ca(2+) entry which triggers recruitment of TRPC1 into the plasma membrane where it is then activated by STIM1. TRPC1 and Orai1 form discrete STIM1-gated channels that generate distinct Ca(2+) signals and regulate specific cellular functions. Surface expression of TRPC1 can be modulated by trafficking of the channel to and from the plasma membrane, resulting in changes to the phenotype of TRPC1-mediated current and [Ca(2+)]i signals. Thus, TRPC1 is activated downstream of Orai1 and modifies the initial [Ca(2+)]i signal generated by Orai1 following store depletion. This review will summarize the important findings that underlie the current concepts for activation and regulation of TRPC1, as well as its impact on cell function.
ArdieSW, LiuSK, TakanoT ( 2010). Expression of the AKT1-type K + channel gene from Puccinellia tenuiflora, PutAKT1, enhances salt tolerance in Arabidopsis .Plant Cell Rep 29, 865-874. [本文引用: 1]
BaiY, DoughertyL, LiMJ, FazioG, ChengLL, XuKN ( 2012). A natural mutation-led truncation in one of the two aluminum-activated malate transporter-like genes at the Ma locus is associated with low fruit acidity in apple .Mol Genet Genomics 287, 663-678. DOI:10.1007/s00438-012-0707-7URLPMID:22806345 [本文引用: 1] Abstract largely controls the variation of fruit acidity levels. The locus has recently been defined in a region of 15002kb that contains 44 predicted genes on chromosome 16 in the Golden Delicious genome. In this study, we identified two aluminum-activated malate transporter-like genes, designated and , as strong candidates of by narrowing down the locus to 65–8202kb containing 12–19 predicted genes depending on the haplotypes. The haplotypes were determined by sequencing two bacterial artificial chromosome clones from G.41 (an apple rootstock of genotype ) that cover the two distinct haplotypes at the locus. Gene expression profiling in 18 apple germplasm accessions suggested that is the major determinant at the locus controlling fruit acidity as is expressed at a much higher level than and the expression is significantly correlated with fruit titratable acidity (02=020.4543, 02=020.0021). In the coding sequences of low acidity alleles of and , sequence variations at the amino acid level between Golden Delicious and G.41 were not detected. But the alleles for high acidity vary considerably between the two genotypes. The low acidity allele of , - that targets the SNP in showed that the CAPS for low pH or high acidity in apple.
BeheraS, LongY, Schmitz-ThomI, WangXP, ZhangCX, LiH, SteinhorstL, ManishankarP, RenXL, OffenbornJN, WuWH, KudlaJ, WangY ( 2017). Two spatially and temporally distinct Ca 2+ signals convey Arabidopsis thaliana responses to K + deficiency. New Phytol 213, 739-750. [本文引用: 1]
BrandtB, BrodskyDE, XueSW, NegiJ, IbaK, Kang- asjarviJ, GhassemianM, StephanAB, HuHH, Sch- roederJI ( 2012). Reconstitution of abscisic acid activation of SLAC1 anion channel by CPK6 and OST1 kinases and branched ABI1 PP2C phosphatase action Proc Natl Acad Sci USA 109, 10593-10598. [本文引用: 2]
ChenYH, HuL, PuntaM, BruniR, HillerichB, KlossB, RostB, LoveJ, SiegelbaumSA, HendricksonWA ( 2010). Homologue structure of the SLAC1 anion channel for closing stomata in leaves Nature 467, 1074-1080. DOI:10.1038/nature09487URLPMID:20981093 [本文引用: 3] The plant SLAC1 anion channel controls turgor pressure in the aperture-defining guard cells of plant stomata, thereby regulating exchange of water vapor and photosynthetic gases in response to environmental signals such as drought or high levels of carbon dioxide. We determined the crystal structure of a bacterial homolog of SLAC1 at 1.20 resolution, and we have used structure-inspired mutagenesis to analyze the conductance properties of SLAC1 channels. SLAC1 is a symmetric trimer composed from quasi-symmetric subunits, each having ten transmembrane helices arranged from helical hairpin pairs to form a central five-helix transmembrane pore that is gated by an extremely conserved phenylalanine residue. Conformational features suggest a mechanism for control of gating by kinase activation, and electrostatic features of the pore coupled with electrophysiological characteristics suggest that selectivity among different anions is largely a function of the energetic cost of ion dehydration.
ChenZC, YokoshoK, KashinoM, ZhaoFJ, YamajiN, MaJF ( 2013). Adaptation to acidic soil is achieved by inc- reased numbers of cis-acting elements regulating ALMT1 expression in Holcus lanatus .Plant J 76, 10-23. [本文引用: 1]
CheongYH, PandeyGK, GrantJJ, BatisticO, LiLG, KimBG, LeeSC, KudlaJ, LuanS ( 2007). Two calcineurin B-like calcium sensors, interacting with protein kinase CIPK23, regulate leaf transpiration and root potassium uptake in Arabidopsis Plant J 52, 223-239. DOI:10.1111/j.1365-313X.2007.03236.xURLPMID:17922773 [本文引用: 1] Abstract Calcium signalling involves sensor proteins that decode temporal and spatial changes in cellular Ca2+ concentration. Calcineurin B-like proteins (CBLs) represent a unique family of plant calcium sensors that relay signals by interacting with a family of protein kinases, designated as CBL-interacting protein kinases (CIPKs). In a reverse genetic screen for altered drought tolerance, we identified a loss-of-function allele of CIPK23 as exhibiting a drought-tolerant phenotype. In the cipk23 mutant, reduced transpirational water loss from leaves coincides with enhanced ABA sensitivity of guard cells during opening as well as closing reactions, without noticeable alterations in ABA content in the plant. We identified the calcium sensors CBL1 and CBL9 as CIPK23-interacting proteins that targeted CIPK23 to the plasma membrane in vivo. Expression analysis of the CIPK23, CBL1 and CBL9 genes suggested that they may function together in diverse tissues, including guard cells and root hairs. In addition, expression of the CIPK23 gene was induced by low-potassium conditions, implicating a function of this gene product in potassium nutrition. Indeed, cipk23 mutants displayed severe growth impairment on media with low concentrations of potassium. This phenotype correlates with a reduced efficiency of K+ uptake into the roots. In support of the conclusion that CBL1 and CBL9 interact with and synergistically serve as upstream regulators of CIPK23, the cbl1 cbl9 double mutant, but not the cbl1 or cbl9 single mutants, exhibit altered phenotypes for stomatal responses and low-potassium sensitivity. Together with the recent identification of the potassium channel AKT1 as a target of CIPK23, these results imply that plasma membrane-localized CBL1- and CBL9-CIPK23 complexes simultaneously regulate K+ transport processes in roots and in stomatal guard cells.
ChiassonDM, HaageK, SollweckK, BrachmannA, Die- trichP, ParniskeM ( 2017). A quantitative hypermorphic CNGC allele confers ectopic calcium flux and impairs cellular development .eLife 6, e25012. DOI:10.7554/eLife.25012URLPMID:28933692 [本文引用: 1] 10.7554/eLife.25012.001The coordinated control of Ca2+ signaling is essential for development in eukaryotes. Cyclic nucleotide-gated channel (CNGC) family members mediate Ca2+ influx from cellular stores in plants (Charpentier et al., 2016; Gao et al., 2016; Frietsch et al., 2007; Urquhart et al., 2007). Here, we report the unusual genetic behavior of a quantitative gain-of-function CNGC mutation (brush) in Lotus japonicus resulting in a leaky tetrameric channel. brush resides in a cluster of redundant CNGCs encoding subunits which resemble metazoan voltage-gated potassium (Kv1-Kv4) channels in assembly and gating properties. The recessive mongenic brush mutation impaired root development and infection by nitrogen-fixing rhizobia. The brush allele exhibited quantitative behavior since overexpression of the cluster subunits was required to suppress the brush phenotype. The results reveal a mechanism by which quantitative competition between channel subunits for tetramer assembly can impact the phenotype of the mutation carrier.
ChinK, DeFalcoTA, MoederW, YoshiokaK ( 2013). The Arabidopsis cyclic nucleotide-gated ion channels AtCNGC2 and AtCNGC4 work in the same signaling pathway to regulate pathogen defense and floral transition Plant Phy-siol 163, 611-624. DOI:10.1104/pp.113.225680URL [本文引用: 1] Arabidopsis (Arabidopsis thaliana) cyclic nucleotide-gated ion channels (CNGCs) form a large family consisting of 20 members and have been implicated in Ca2+ signaling related to various physiological processes, such as pathogen defense, development, and thermotolerance. The null mutant of AtCNGC2, defense, no death (dnd1), exhibits autoimmune phenotypes, while it is impaired in mounting the hypersensitive response, which is a hallmark of effector-triggered (i.e. RESISTANCE-gene mediated) resistance. It has been suggested that AtCNGC2 is involved in defense responses and likely other aspects of physiology through its role as a Ca2+-conducting channel. However, the downstream signaling components and its relation with AtCNGC4, which is the closest paralog of AtCNGC2, remain elusive. Despite the fact that cngc4 mutants display almost identical phenotypes to those seen in cngc2 mutants, not much is known about their relationship. Here, we report the identification and characterization of the Arabidopsis mutant repressor of defense no death1 (rdd1), obtained from a suppressor screen of a transfer DNA insertion knockout mutant of AtCNGC2 in order to identify downstream components of dnd1-mediated signal transduction. rdd1 suppressed the majority of dnd1-mediated phenotypes except Ca2+ hypersensitivity. In addition, rdd1 also suppressed the dnd1-mediated late-flowering phenotype that was discovered in this study. Our genetic analysis conducted to elucidate the relationship between AtCNGC2 and AtCNGC4 indicates that RDD1 is also involved in AtCNGC4-mediated signal transduction. Lastly, bimolecular fluorescence complementation analysis suggests that AtCNGC2 and AtCNGC4 are likely part of the same channel complex.
ClarksonDT, HansonJB ( 1980). The mineral nutrition of higher plants Ann Rev Plant Physiol 31, 239-298. [本文引用: 1]
Corratgé-FaillieC, RonzierE, SanchezF, PradoK, KimJH, LancianoS, LeonhardtN, LacombeB, XiongTC ( 2017). The Arabidopsis guard cell outward potassium channel GORK is regulated by CPK33 FEBS Lett 591, 1982-1992. DOI:10.1002/1873-3468.12687URLPMID:28543075 [本文引用: 1] A complex signaling network involving voltage-gated potassium channels from the Shaker family contributes to the regulation of stomatal aperture. Several kinases and phosphatases have been shown to be crucial for ABA-dependent regulation of the ion transporters. To date, the Ca(2+) -dependent regulation of Shaker channels by Ca(2+) -dependent protein kinases (CPKs) is still elusive. A functional screen in Xenopus oocytes was launched to identify such CPKs able to regulate the three main guard cell Shaker channels KAT1, KAT2, and GORK. Seven guard cell CPKs were tested and multiple CPK/Shaker couples were identified. Further work on CPK33 indicates that GORK activity is enhanced by CPK33 and unaffected by a nonfunctional CPK33 (CPK33-K102M). Furthermore, Ca(2+) -induced stomatal closure is impaired in two cpk33 mutant plants.
CosgroveDJ, HedrichR ( 1991). Stretch-activated chloride, potassium, and calcium channels coexisting in plasma membranes of guard cells of Vicia faba L .Planta 186, 143-153. DOI:10.1007/BF00201510URLPMID:11538499 [本文引用: 1] Mechanosensitive ion channels in the plasma membrane of Vicia faba guard cell protoplasts were studied by use of the patch clamp technique. Stretch-activated (SA) channels in outside-out patches were analyzed for channel conductance, kinetics and ion selectivity. We found three distinct SA channels, permeable to Cl-, K+ and Ca2+ and distinguishable from spontaneous (non-SA) channels for these ions on the basis of conductance, kinetics, and voltage-dependence, as well as sensitivity to membrane stretch. In the attached patch configuration, light suction (2 to 10 kPa) reversibly induced channel opening with multiple amplitudes and complex kinetics. The open probability for SA channels increased nonlinearly with pipette suction. In guard cells in situ, these SA channels may mediate ion transport across the plasma membrane directly, as well as influence the activity of non-SA channels via effects on membrane voltage and cytoplasmic calcium. Through such effects, SA channels likely influence volume and turgor regulation of guard cells, and thereby control of leaf gas exchange.
Cubero-FontP, MaierhoferT, JaslanJ, RosalesMA, EsparteroJ, Diaz-RuedaP, MullerHM, HurterAL, Al- RasheidKAS, MartenI, HedrichR, Colmenero-FloresJM, GeigerD ( 2016). Silent S-type anion channel subunit SLAH1 gates SLAH3 open for chloride root-to-shoot trans- location Curr Biol 26, 2213-2220. DOI:10.1016/j.cub.2016.06.045URLPMID:27397895 [本文引用: 2] Cubero-Font, Maierhofer, et02al. report that plants adjust the ratio between Cl61and NO361in the shoot via heteromerization ofArabidopsisSLAH1/SLAH3 anion channel subunits. Differential expression of SLAH1 controls Cl61fluxes to the shoot without affecting the uptake of NO361, making SLAH1 a promising target for engineering salinity-tolerant plants.
DaramP, UrbachS, GaymardF, SentenacH, ChérelI ( 1997). Tetramerization of the AKT1 plant potassium chan- nel involves its C-terminal cytoplasmic domain EMBO J 16, 3455-3463. DOI:10.1093/emboj/16.12.3455URLPMID:9218788 [本文引用: 1] All plant channels identified so far show high conservation throughout the polypeptide sequence except in the ankyrin domain which is present only in those closely related to AKT1. In this study, the architecture of the AKT1 protein has been investigated. AKT1 polypeptides expressed in the baculovirus/Sf9 cells system were found to assemble into tetramers as observed with animal Shaker-like potassium channel subunits. The AKT1 C-terminal intracytoplasmic region (downstream from the transmembrane domain) alone formed tetrameric structures when expressed in Sf9 cells, revealing a tetramerization process different from that of Shaker channels. Tests of subfragments from this sequence in the two-hybrid system detected two kinds of interaction. The first, involving two identical segments (amino acids 371-516), would form a contact between subunits, probably via their putative cyclic nucleotide-binding domains. The second interaction was found between the last 81 amino acids of the protein and a region lying between the channel hydrophobic core and the putative cyclic nucleotide-binding domain. As the interacting regions are highly conserved in all known plant potassium channels, the structural organization of AKT1 is likely to extend to these channels. The significance of this model with respect to animal cyclic nucleotide-gated channels is also discussed.
De AngeliA, BaetzU, FranciscoR, ZhangJB, ChavesMM, RegaladoA ( 2013 a). The vacuolar channel VvALMT9 mediates malate and tartrate accumulation in berries of Vitis vinifera .Planta 238, 283-291. [本文引用: 1]
De AngeliA, ZhangJB, MeyerS, MartinoiaE ( 2013 b). AtALMT9 is a malate-activated vacuolar chloride channel required for stomatal opening in Arabidopsis Nat Commun 4, 1804. DOI:10.1038/ncomms2815URLPMID:23653216 [本文引用: 1] Abstract Water deficit strongly affects crop productivity. Plants control water loss and CO2 uptake by regulating the aperture of the stomatal pores within the leaf epidermis. Stomata aperture is regulated by the two guard cells forming the pore and changing their size in response to ion uptake and release. While our knowledge about potassium and chloride fluxes across the plasma membrane of guard cells is advanced, little is known about fluxes across the vacuolar membrane. Here we present the molecular identification of the long-sought-after vacuolar chloride channel. AtALMT9 is a chloride channel activated by physiological concentrations of cytosolic malate. Single-channel measurements demonstrate that this activation is due to a malate-dependent increase in the channel open probability. Arabidopsis thaliana atalmt9 knockout mutants exhibited impaired stomatal opening and wilt more slowly than the wild type. Our findings show that AtALMT9 is a vacuolar chloride channel having a major role in controlling stomata aperture.
DreyerI, AntunesS, HoshiT, Müller-R?berB, PalmeK, PongsO, ReintanzB, HedrichR ( 1997). Plant K + channel alpha-subunits assemble indiscriminately Biophys J 72, 2143-2150. DOI:10.1016/S0006-3495(97)78857-XURLPMID:9129816 [本文引用: 1] Abstract In plants a large diversity of inwardly rectifying K+ channels (K(in) channels) has been observed between tissues and species. However, only three different types of voltage-dependent plant K+ uptake channel subfamilies have been cloned so far; they relate either to KAT1, AKT1, or AtKC1. To explore the mechanisms underlying the channel diversity, we investigated the assembly of plant inwardly rectifying alpha-subunits. cRNA encoding five different K+ channel alpha-subunits of the three subfamilies (KAT1, KST1, AKT1, SKT1, and AtKC1) which were isolated from different tissues, species, and plant families (Arabidopsis thaliana and Solanum tuberosum) was reciprocally co-injected into Xenopus oocytes. We identified plant K+ channels as multimers. Moreover, using K+ channel mutants expressing different sensitivities to voltage, Cs+, Ca2+, and H+, we could prove heteromers on the basis of their altered voltage and modulator susceptibility. We discovered that, in contrast to animal K+ channel alpha-subunits, functional aggregates of plant K(in) channel alpha-subunits assembled indiscriminately. Interestingly, AKT-type channels from A. thaliana and S. tuberosum, which as homomers were electrically silent in oocytes after co-expression, mediated K+ currents. Our findings suggest that K+ channel diversity in plants results from nonselective heteromerization of different alpha-subunits, and thus depends on the spatial segregation of individual alpha-subunit pools and the degree of temporal overlap and kinetics of expression.
DreyerI, PoreeF, SchneiderA, Mittelst?dtJ, BertlA, SentenacH, ThibaudJB, Mueller-RoeberB ( 2004). Assembly of plant Shaker-like Kout channels requires two distinct sites of the channel α-subunit .Biophys J 87, 858-872. [本文引用: 1]
DubyG, HosyE, FizamesC, AlconC, CostaA, SentenacH, ThibaudJB ( 2008). AtKC1, a conditionally targeted Shaker-type subunit, regulates the activity of plant K + channels Plant J 53, 115-123. DOI:10.1111/j.1365-313X.2007.03324.xURLPMID:17976154 [本文引用: 1] Amongst the nine voltage-gated K + channel (Kv) subunits expressed in Arabidopsis, AtKC1 does not seem to form functional Kv channels on its own, and is therefore said to be silent. It has been proposed to be a regulatory subunit, and to significantly influence the functional properties of heteromeric channels in which it participates, along with other Kv channel subunits. The mechanisms underlying these properties of AtKC1 remain unknown. Here, the transient (co-)expression of AtKC1 , AKT1 and/or KAT1 genes was obtained in tobacco mesophyll protoplasts, which lack endogenous inward Kv channel activity. Our experimental conditions allowed both localization of expressed polypeptides (GFP-tagging) and recording of heterologously expressed Kv channel activity (untagged polypeptides). It is shown that AtKC1 remains in the endoplasmic reticulum unless it is co-expressed with AKT1. In these conditions heteromeric AtKC1-AKT1 channels are obtained, and display functional properties different from those of homomeric AKT1 channels in the same context. In particular, the activation threshold voltage of the former channels is more negative than that of the latter ones. Also, it is proposed that AtKC1-AKT1 heterodimers are preferred to AKT1-AKT1 homodimers during the process of tetramer assembly. Similar results are obtained upon co-expression of AtKC1 with KAT1 . The whole set of data provides evidence that AtKC1 is a conditionally-targeted Kv subunit, which probably downregulates the physiological activity of other Kv channel subunits in Arabidopsis.
DuttaR, RobinsonKR ( 2004). Identification and characteri- zation of stretch-activated ion channels in pollen protop- lasts Plant Physiol 135, 1398-1406. [本文引用: 1]
FanLM, WangYF, WangH, WuWH ( 2001). In vitro Arabidopsis pollen germination and characterization of the inward potassium currents in Arabidopsis pollen grain pro- toplasts J Exp Bot 52, 1603-1614. DOI:10.1093/jexbot/52.361.1603URLPMID:11479325 [本文引用: 1] The focus of this study is to investigate the regulatory role of K+ influx in Arabidopsis pollen germination and pollen tube growth. Using agar-containing media, in vitro methods for Arabidopsis pollen germination have been successfully established for the first time. The pollen germination percentage was nearly 75% and the average pollen tube length reached 135 m after a 6 h incubation. A decrease in external K+ concentration from 1 mM to 35 M resulted in 30% inhibition of pollen germination and 40% inhibition of pollen tube growth. An increase in external K+ concentration from 1 mM to 30 mM stimulated pollen tube growth but inhibited pollen germination. To study how K+ influx is associated with pollen germination and tube growth, regulation of the inward K+ channels in the pollen plasma membrane was investigated by conducting patch-clamp whole-cell recording with pollen protoplasts. K+ currents were first identified in Arabidopsis pollen protoplasts. The inward K+ currents were insensitive to changes in cytoplasmic Ca2+ but were inhibited by a high concentration of external Ca2+. A decrease of external Ca2+ concentration from 10 mM (control) to 1 mM had no significant effect on the inward K+ currents, while an increase of external Ca2+ concentration from 10 mM to 50 mM inhibited the inward K+ currents by 46%. Changes in external pH significantly affected the magnitude, conductance, voltage-independent maximal conductance, and activation kinetics of the inward K+ currents. The physiological importance of potassium influx mediated by the inward K+-channels during Arabidopsis pollen germination and tube growth is discussed.
FanLM, WangYF, WuWH ( 2003). Outward K + channels in Brassica chinensis pollen protoplasts are regulated by external and internal pH .Protoplasma 220, 143-152. DOI:10.1007/s00709-002-0037-4URLPMID:12664278 [本文引用: 1] Abstract Patch-clamp whole-cell and single-channel recording techniques were used to investigate the regulation of outward K(+) channels by external and internal protons in Brassica chinensispollen protoplasts. Outward K(+) currents and conductance were insensitive to external pH (pH(o)) except at pH 4.5. Maximal conductance (G(max)) for the outward K(+) currents was inhibited at acidic external pH. Half-activation voltage ( E(1/2)) for the outward K(+) currents shifted to more positive voltages along with the decrease in pH(o). E(1/2) can be described by a modified Henderson-Hasselbalch equation expected from a single titratable binding site. The activation kinetics of the outward K(+) channels was largely insensitive to pH(o). An internal pH (pH(i)) of 4.5 significantly increased outward K(+) currents and conductance. G(max) for the outward K(+) currents decreased with elevations in pH(i). In contrast to the effect of pH(o), E(1/2) was shifted to more positive voltages with elevations in pH(i). The outward K(+) currents, G(max) and E(1/2) can be described by the modified Henderson-Hasselbalch equation. Furthermore, acidifying pH(i) accelerated the activation of the outward K(+) currents significantly. The differences in electro-physiological properties among previously reported and currently described plant outward K(+) channels may reflect differences in the structure of these channels.
FinkaA, CuendetAFH, MaathuisFJM, SaidiY, Golou- binoffP ( 2012). Plasma membrane cyclic nucleotide gated calcium channels control land plant thermal sensing and acquired thermotolerance Plant Cell 24, 3333-3348. DOI:10.1105/tpc.112.095844URLPMID:22904147 [本文引用: 1] Typically at dawn on a hot summer day, land plants need precise molecular thermometers to sense harmless increments in the ambient temperature to induce a timely heat shock response (HSR) and accumulate protective heat shock proteins in anticipation of harmful temperatures at mid-day. Here, we found that the cyclic nucleotide gated calcium channel (CNGC) CNGCb gene from Physcomitrella patens and its Arabidopsis thaliana ortholog CNGC2, encode a component of cyclic nucleotide gated Ca0562 channels that act as the primary thermosensors of land plant cells. Disruption of CNGCb or CNGC2 produced a hyper-thermosensitive phenotype, giving rise to an HSR and acquired thermotolerance at significantly milder heat-priming treatments than in wild-type plants. In an aequorin-expressing moss, CNGCb loss-of-function caused a hyperthermoresponsive Ca0562 influx and altered Ca0562 signaling. Patch clamp recordings on moss protoplasts showed the presence of three distinct thermoresponsive Ca0562 channels in wild-type cells. Deletion of CNGCb led to a total absence of one and increased the open probability of the remaining two thermoresponsive Ca0562 channels. Thus, CNGC2 and CNGCb are expected to form heteromeric Ca0562 channels with other related CNGCs. These channels in the plasma membrane respond to increments in the ambient temperature by triggering an optimal HSR, leading to the onset of plant acquired thermotolerance.
FontechaG, Silva-NavasJ, BenitoC, MestresMA, EspinoFJ, Hernández-RiquerMV, GallegoFJ ( 2007). Candidate gene identification of an aluminum-activated organic acid transporter gene at the Alt4 locus for aluminum tolerance in rye( Secale cereale L.). Theor Appl Genet 114, 249-260. [本文引用: 1]
FuchsI, St?lzleS, IvashikinaN, HedrichR ( 2005). Rice K + uptake channel OsAKT1 is sensitive to salt stress Planta 221, 212-221. [本文引用: 1]
GaoF, HanXW, WuJH, ZhengSZ, ShangZl, SunDY, ZhouRG, LiB ( 2012). A heat-activated calcium-permeable channel—Arabidopsis cyclic nucleotide-gated ion channel 6—is involved in heat shock responses Plant J 70, 1056-1069. DOI:10.1111/j.1365-313X.2012.04969.xURLPMID:22372427 [本文引用: 1] Summary An increased concentration of cytosolic calcium ions (Ca 2+ ) is an early response by plant cells to heat shock. However, the molecular mechanism underlying the heat-induced initial Ca 2+ response in plants is unclear. In this study, we identified and characterized a heat-activated Ca 2+ -permeable channel in the plasma membrane of Arabidopsis thaliana root protoplasts using reverse genetic analysis and the whole-cell patch-clamp technique. The results indicated that A.thaliana cyclic nucleotide-gated ion channel6 (CNGC6) mediates heat-induced Ca 2+ influx and facilitates expression of heat shock protein (HSP) genes and the acquisition of thermotolerance. GUS and GFP reporter assays showed that CNGC6 expression is ubiquitous in A.thaliana , and the protein is localized to the plasma membrane of cells. Furthermore, it was found that the level of cytosolic cAMP was increased by a mild heat shock, that CNGC6 was activated by cytosolic cAMP, and that exogenous cAMP promoted the expression of HSP genes. The results reveal the role of cAMP in transduction of heat shock signals in plants. The correlation of an increased level of cytosolic cAMP in a heat-shocked plant with activation of the Ca 2+ channels and downstream expression of HSP genes sheds some light on how plants transduce a heat stimulus into a signal cascade that leads to a heat shock response.
GaoQF, FeiCF, DongJY, GuLL, WangYF ( 2014). Arabidopsis CNGC18 is a Ca 2+-permeable channel Mol Plant 7, 739-743. [本文引用: 1]
GaoQF, GuLL, WangHQ, FeiCF, FangX, HussainJ, SunSJ, DongJY, LiuHT, WangYF ( 2016). Cyclic nucleotide- gated channel 18 is an essential Ca 2+ channel in pollen tube tips for pollen tube guidance to ovules in Arabidopsis Proc Natl Acad Sci USA 113, 3096-3101. DOI:10.1073/pnas.1524629113URLPMID:26929345 [本文引用: 2] Abstract In flowering plants, pollen tubes are guided into ovules by multiple attractants from female gametophytes to release paired sperm cells for double fertilization. It has been well-established that Ca(2+) gradients in the pollen tube tips are essential for pollen tube guidance and that plasma membrane Ca(2+) channels in pollen tube tips are core components that regulate Ca(2+) gradients by mediating and regulating external Ca(2+) influx. Therefore, Ca(2+) channels are the core components for pollen tube guidance. However, there is still no genetic evidence for the identification of the putative Ca(2+) channels essential for pollen tube guidance. Here, we report that the point mutations R491Q or R578K in cyclic nucleotide-gated channel 18 (CNGC18) resulted in abnormal Ca(2+) gradients and strong pollen tube guidance defects by impairing the activation of CNGC18 in Arabidopsis. The pollen tube guidance defects of cngc18-17 (R491Q) and of the transfer DNA (T-DNA) insertion mutant cngc18-1 (+/-) were completely rescued by CNGC18. Furthermore, domain-swapping experiments showed that CNGC18's transmembrane domains are indispensable for pollen tube guidance. Additionally, we found that, among eight Ca(2+) channels (including six CNGCs and two glutamate receptor-like channels), CNGC18 was the only one essential for pollen tube guidance. Thus, CNGC18 is the long-sought essential Ca(2+) channel for pollen tube guidance in Arabidopsis.
GaoYQ, WuWH, WangY ( 2017). The K + channel KZM2 is involved in stomatal movement by modulating inward K + currents in maize guard cells Plant J 92, 662-675. DOI:10.1111/tpj.13712URLPMID:28891257 [本文引用: 1] react-text: 116 To identify mechanisms for the simultaneous release of anions and cations into the xylem sap in roots, we investigated voltage- dependent ion conductances in the plasmalemma of xylem paren- chyma cells. We applied the patch-clamp technique to protoplasts isolated from the xylem parenchyma by differential enzymic diges- tion of steles of barley roots (Hordeum vulgare 1. cv Apex). In the... /react-text react-text: 117 /react-text [Show full abstract]
GaymardF, PilotG, LacombeB, BouchezD, BruneauD, BoucherezJ, Michaux-FerrièreN, ThibaudJB, Sen- tenacH ( 1998). Identification and disruption of a plant shaker-like outward channel involved in K + release into the xylem sap Cell 94, 647-655. DOI:10.1016/S0092-8674(00)81606-2URLPMID:9741629 [本文引用: 1] Abstract SKOR, a K+ channel identified in Arabidopsis, displays the typical hydrophobic core of the Shaker channel superfamily, a cyclic nucleotide-binding domain, and an ankyrin domain. Expression in Xenopus oocytes identified SKOR as the first member of the Shaker family in plants to be endowed with outwardly rectifying properties. SKOR expression is localized in root stelar tissues. A knockout mutant shows both lower shoot K+ content and lower xylem sap K+ concentration, indicating that SKOR is involved in K+ release into the xylem sap toward the shoots. SKOR expression is strongly inhibited by the stress phytohormone abscisic acid, supporting the hypothesis that control of K+ translocation toward the shoots is part of the plant response to water stress.
GeigerD, MaierhoferT, AL-RasheidKAS, ScherzerS, MummP, LieseA, AcheP, WellmannC, MartenI, GrillE, RomeisT, HedrichR ( 2011). Stomatal closure by fast abscisic acid signaling is mediated by the guard cell anion channel SLAH3 and the receptor RCAR1 Sci Signal 4, ra32. [本文引用: 2]
GeigerD, ScherzerS, MummP, StangeA, MartenI, BauerH, AcheP, MatschiS, LieseA, Al-RasheidKAS, RomeisT, HedrichR ( 2009 b). Activity of guard cell anion channel SLAC1 is controlled by drought-stress signaling kinase- phosphatase pair Proc Natl Acad Sci USA 106, 21425-21430.
GrefenC, ChenZH, HonsbeinA, DonaldN, HillsA, BlattMR ( 2010). A novel motif essential for SNARE interaction with the K + channel KC1 and channel gating in Arabidopsis Plant Cell 22, 3076-3092. [本文引用: 1]
GruberBD, RyanPR, RichardsonAE, TyermanSD, RameshS, HebbDM, HowittSM, DelhaizeE ( 2010). HvALMT1 from barley is involved in the transport of organic anions J Exp Bot 61, 1455-1467. DOI:10.1093/jxb/erq023URLPMID:2837267 [本文引用: 1] Abstract Members of the ALMT gene family contribute to the Al(3+) resistance of several plant species by facilitating malate efflux from root cells. The first member of this family to be cloned and characterized, TaALMT1, is responsible for most of the natural variation of Al(3+) resistance in wheat. The current study describes the isolation and characterization of HvALMT1, the barley gene with the greatest sequence similarity to TaALMT1. HvALMT1 is located on chromosome 2H which has not been associated with Al(3+) resistance in barley. The relatively low levels of HvALMT1 expression detected in root and shoot tissues were independent of external aluminium or phosphorus supply. Transgenic barley plants transformed with the HvALMT1 promoter fused to the green fluorescent protein (GFP) indicated that expression of HvALMT1 was relatively high in stomatal guard cells and in root tissues containing expanding cells. GFP fused to the C-terminus of the full HvALMT1 protein localized to the plasma membrane and motile vesicles within the cytoplasm. HvALMT1 conferred both inward and outward currents when expressed in Xenopus laevis oocytes that were bathed in a range of anions including malate. Both malate uptake and efflux were confirmed in oocyte assays using [(14)C]malate as a radiotracer. It is suggested that HvALMT1 functions as an anion channel to facilitate organic anion transport in stomatal function and expanding cells.
GuLL, GaoQF, WangYF ( 2017). Cyclic nucleotide-gated channel 18 functions as an essential Ca 2+ channel for pollen germination and pollen tube growth in Arabidopsis Plant Signal Behav 12, e1197999. DOI:10.1080/15592324.2016.1197999URLPMID:5703252 [本文引用: 1] Abstract We recently revealed that cyclic nucleotide-gated channel 18 (CNGC18) functioned as the main Ca 2+ channel in pollen tube tips for pollen tube guidance to ovules by regulating external Ca 2+ influx in Arabidopsis. In this study, we found that the reduction of external Ca 2+ concentration ([Ca 2+ ] ext ) from 10 mM to 5 mM, and further to 2 mM, led to the decreases of pollen germination percentages, but led to the increases of the percentages of ruptured pollen grains and tubes, and branched pollen tubes in vitro in cngc18-17 compared with wild type. The second point mutant allele cngc18-22 showed similar phenotypes, including reduced pollen germination percentages, increased percentages of ruptured pollen tubes, but did not show obvious different percentages of ruptured pollen grains and branched pollen tubes compared with wild type. These data demonstrate that CNGC18 plays essential roles in pollen germination and tube growth as a Ca 2+ channel in Arabidopsis.
GuoKM, BabourinaO, ChristopherDA, BorsicsT, RengelZ ( 2008). The cyclic nucleotide-gated channel, AtCNGC10, influences salt tolerance in Arabidopsis Physiol Plant 134, 499-507. DOI:10.1111/j.1399-3054.2008.01157.xURLPMID:18823330 [本文引用: 1] Cyclic nucleotide-gated channels (CNGCs) in the plasma membrane transport K + and other cations; however, their roles in the response and adaptation of plants to environmental salinity are unclear. Growth, cation contents, salt tolerance and K + fluxes were assessed in wild-type and two AtCNGC10 antisense lines (A2 and A3) of Arabidopsis thaliana (L.) Heynh. Compared with the wild-type, mature plants of both antisense lines had altered K + and Na + concentrations in shoots and were more sensitive to salt stress, as assessed by biomass and Chl fluorescence. The shoots of A2 and A3 plants contained higher Na + concentrations and significantly higher Na + /K + ratios compared with wild-type, whereas roots contained higher K + concentrations and lower Na + /K + ratios. Four-day-old seedlings of both antisense lines exposed to salt stress had smaller Na + /K + ratios and longer roots than the wild-type. Under sudden salt treatment, the Na + efflux was higher and the K + efflux was smaller in the antisense lines, indicating that AtCNGC10 might function as a channel providing Na + influx and K + efflux at the root/soil interface. We conclude that the AtCNGC10 channel is involved in Na + and K + transport during cation uptake in roots and in long-distance transport, such as phloem loading and/or xylem retrieval. Mature A2 and A3 plants became more salt sensitive than wild-type plants because of impaired photosynthesis induced by a higher Na + concentration in the leaves.
GuoKM, BabourinaO, ChristopherDA, BorsicT, RengelZ ( 2010). The cyclic nucleotide-gated channel AtCNGC10 transports Ca 2+ and Mg 2+ in Arabidopsis Physiol Plant 139, 303-312. DOI:10.1111/j.1399-3054.2010.01366.xURLPMID:20210874 [本文引用: 1] The suppression of the cyclic nucleotide-gated channel (CNGC) AtCNGC10 alters K+ transport in Arabidopsis plants. Other CNGCs have been shown to transport Ca2+, K+, Li+, Cs+ and Rb+ across the plasma membrane when expressed in heterologous systems; however, the ability of the AtCNGC10 channel to transport nutrients other than K+ in plants has not been previously tested. The ion fluxes along different zones of the seedling roots, as estimated by the non-invasive ion-specific microelectrode technique, were significantly different in two AtCNGC10 antisense lines (A2 and A3) in comparison to the wild type (WT). Most notably, the influxes of H+, Ca2+ and Mg2+ in the meristem and distal elongation zones of the antisense A2 and A3 lines were significantly lower than in the WT. The lower Ca2+ influx from the external media corresponded to a lower intracellular Ca2+ activity, which was estimated by fluorescence lifetime imaging measurements (FLIM). On the other hand, the intracellular pH values in the meristem zone of the roots of A2 and A3 seedlings were significantly lower (more acidic) than that of the WT, which might indicate a feedback block of H+ influx into meristematic cells caused by low intracellular pH. Under the control conditions, mature plants from the A2 and A3 lines contained significantly higher K+ and lower Ca2+ and Mg2+ content in the shoots, indicating disturbed long-distance ion transport of these cations, possibly because of changes in xylem loading/retrieval and/or phloem loading. Exposing the plants in the flowering stage to various K+, Ca2+ and Mg2+ concentrations in the solution led to altered K+, Ca2+ and Mg2+ content in the shoots of A2 and A3 plants in comparison with the WT, suggesting a primary role of AtCNGC10 in Ca2+ (and probably Mg2+) transport in plants, which in turn regulates K+ transporters' activities.
HaswellES, PeyronnetR, Barbier-BrygooH, MeyerowitzEM, FrachisseJM ( 2008). Two MscS homologs provide mechanosensitive channel activities in the Arabidopsis root Curr Biol 18, 730-734. DOI:10.1016/j.cub.2008.04.039URLPMID:18485707 [本文引用: 1] In bacterial and animal systems, mechanosensitive (MS) ion channels are thought to mediate the perception of pressure, touch, and sound [1-3]. Although plants respond to a wide variety of mechanical stimuli, and although many mechanosensitive channel activities have been characterized in plant membranes by the patch-clamp method, the molecular nature of mechanoperception in plant systems has remained elusive [4]. Likely candidates are relatives of MscS (Mechanosensitive channel of small conductance), a well-characterized MS channel that serves to protect E. coli from osmotic shock [5]. Ten MscS-Like (MSL) proteins are found in the genome of the model flowering plant Arabidopsis thaliana[4, 6, 7]. MSL2 and MSL3, along with MSC1, a MscS family member from green algae, are implicated in the control of organelle morphology [8, 9]. Here, we characterize MSL9 and MSL10, two MSL proteins found in the plasma membrane of root cells. We use a combined genetic and electrophysiological approach to show that MSL9 and MSL10, along with three other members of the MSL family, are required for MS channel activities detected in protoplasts derived from root cells. This is the first molecular identification and characterization of MS channels in plant membranes.
HoekengaOA, MaronLG, Pi?erosMA, CancadoGMA, ShaffJ, KobayashiY, RyanPR, DongB, DelhaizeE, SasakiT, MatsumotoH, YamamotoY, KoyamaH, KochianLV ( 2006). AtALMT1, which encodes a malate transporter, is identified as one of several genes critical for aluminum tolerance in Arabidopsis Proc Natl Acad Sci USA 103, 9738-9743. DOI:10.1073/pnas.0602868103URLPMID:16740662 [本文引用: 2] Aluminum (Al) tolerance in Arabidopsis is a genetically complex trait, yet it is mediated by a single physiological mechanism based on Al-activated root malate efflux. We investigated a possible molecular determinant for Al tolerance involving a homolog of the wheat Al-activated malate transporter, ALMT1. This gene, named AtALMT1 (Atlg08430), was the best candidate from the 14-member AtALMT family to be involved with Al tolerance based on expression patterns and genomic location. Physiological analysis of a transferred DNA knockout mutant for AtALMT1 as well as electrophysiological examination of the protein expressed in Xenopus oocytes showed that AtALMT1 is critical for Arabidopsis Al tolerance and encodes the Al-activated root malate efflux transporter associated with tolerance. However, gene expression and sequence analysis of AtALMT1 alleles from tolerant Columbia (Col), sensitive Landsberg erecta (Ler), and other ecotypes that varied in Al tolerance suggested that variation observed at AtALMT1 is not correlated with the differences observed in Al tolerance among these ecotypes. Genetic complementation experiments indicated that the Ler allele of AtALMT1 is equally effective as the Col allele in conferring Al tolerance and Al-activated malate release. Finally, fine-scale mapping of a quantitative trait locus (QTL) for Al tolerance on chromosome 1 indicated that AtALMT1 is located proximal to this QTL. These results indicate that AtALMT1 is an essential factor for Al tolerance in Arabidopsis but does not represent the major Al tolerance QTL also found on chromosome 1.
HonsbeinA, SokolovskiS, GrefenC, CampanoniP, PratelliR, PanequeM, ChenZH, JohanssonI, BlattMR ( 2009). A tripartite SNARE-K + channel complex mediates in channel-dependent K + nutrition in Arabidopsis Plant Cell 21, 2859-2877. [本文引用: 1]
HosyE, VavasseurA, MoulineK, DreyerI, GaymardF, PoréeF, BoucherezJ, LebaudyA, BouchezD, VéryAA, SimonneauT, ThibaudJB, SentenacH ( 2003). The Arabidopsis outward K + channel GORK is involved in regulation of stomatal movements and plant transpiration Proc Natl Acad Sci USA 100, 5549-5554. [本文引用: 1]
HouCC, TianW, KleistT, HeK, GarciaV, BaiFL, HaoYL, LuanS, LiLG ( 2014). DUF221 proteins are a family of osmosensitive calcium-permeable cation channels conser- ved across eukaryotes Cell Res 24, 632-635. DOI:10.1038/cr.2014.14URLPMID:24503647 [本文引用: 1] Cell death and differentiation is a monthly research journal focused on the exciting field of programmed cell death and apoptosis. It provides a single accessible source of information for both scientists and clinicians, keeping them up-to-date with advances in the field. It encompasses programmed cell death, cell death induced by toxic agents, differentiation and the interrelation of these with cell proliferation.
HwangH, YoonJ, KimHY, MinMK, KimJA, ChoiEH, LanWZ, BaeYM, LuanS, ChoH, KimBG ( 2013). Unique features of two potassium channels, OsKAT2 and OsKAT3, expressed in rice guard cells PLoS One 8, e72541. DOI:10.1371/journal.pone.0072541URLPMID:23967316 [本文引用: 1] Potassium is the most abundant cation and a myriad of transporters regulate K+ homeostasis in plant. Potassium plays a role as a major osmolyte to regulate stomatal movements that control water utility of land plants. Here we report the characterization of two inward rectifying shaker-like potassium channels, OsKAT2 and OsKAT3, expressed in guard cell of rice plants. While OsKAT2 showed typical potassium channel activity, like that of Arabidopsis KAT1, OsKAT3 did not despite high sequence similarity between the two channel proteins. Interestingly, the two potassium channels physically interacted with each other and such interaction negatively regulated the OsKAT2 channel activity in CHO cell system. Furthermore, deletion of the C-terminal domain recovered the channel activity of OsKAT3, suggesting that the C-terminal region was regulatory domain that inhibited channel activity. Two homologous channels with antagonistic interaction has not been previously reported and presents new information for potassium channel regulation in plants, especially in stomatal regulation.
IidaH, FuruichiT, NakanoM, ToyotaM, SokabeM, TatsumiH ( 2014). New candidates for mechano-sensitive channels potentially involved in gravity sensing in Ara- bidopsis thaliana .Plant Biol 16, 39-42. DOI:10.1111/plb.12044URLPMID:23731064 [本文引用: 2] The mechano-sensitive channels of plants may sense increases in tension induced by mechanical stimuli, such as touch, wind and turgor pressure, and a gravitational stimulus. Recent studies have identified plant homologues of the bacterial mechano-sensitive channel MscS, which is gated by membrane tension and reduces intracellular osmolality by releasing small osmolytes from bacterial cells. However, the physiological roles of these homologues have not yet been clearly elucidated, and only two of them have been shown to be involved in the protection of osmotically stressed plastids in Arabidopsis thaliana. We identified another group of candidates for mechano-sensitive channels in Arabidopsis, named MCA1 and MCA2, whose homologues are exclusively found in plant genomes. MCA1 and MCA2 are composed of 421 and 416 amino acid residues, respectively, share 73% homology in their amino acid sequences, and are not homologous to any known ion channels or transporters. Our structural study revealed that the N-terminal region (one to 173 amino acids) of both proteins was necessary and sufficient for Ca2+ influx activity. Interestingly, this region had one putative transmembrane segment containing an Asp residue whose substitution mutation abolished this activity. Our physiological study suggested that MCA1 expressed at the root tip was required for sensing the hardness of the agar medium or soil. In addition, MCA1 and MCA2 were shown to be responsible for hypo-osmotic shock-induced increases in [Ca2+]cyt. Thus, both proteins appear to be involved in the process of sensing mechanical stresses. We discussed the possible role of both proteins in sensing mechanical and gravitational stimuli.
JanLY, JanYN ( 1997). Voltage-gated and inwardly rectifying potassium channels J Physiol 505, 267-282. DOI:10.1111/j.1469-7793.1997.267bb.xURLPMID:9423171 [本文引用: 1] This lecture is dedicated to Max Delbrück and Seymour Benzer. Max Delbrück was our graduate advisor. He introduced us to a variety of biophysical problems, and taught us ways of thinking about these problems by example. Potassium channels was one of the topics included in his journal club in the early seventies; Max also carefully considered the feasibility of purifying potassium channels then. It was in Seymour Benzer's laboratory that we began to look for Drosophila mutants that a01fect synaptic transmission at the larval neuromuscular junction. Shaker was the first behavioural mutant we tested that gave a robust phenotype, a phenotype that could be mimicked by treating wild-type preparations with a potassium channel blocker. This mutant fly has led us to our subsequent molecular studies of potassium channels.
JeangueninL, AlconC, DubyG, BoeglinM, ChérelI, GaillardI, ZimmermannS, SentenacH, VéryAA ( 2011). AtKC1 is a general modulator of Arabidopsis inward Shaker channel activity Plant J 67, 570-582. DOI:10.1111/j.1365-313X.2011.04617.xURLPMID:21518051 [本文引用: 3] A functional Shaker potassium channel requires assembly of four -subunits encoded by a single gene or various genes from the Shaker family. In Arabidopsis thaliana, AtKC1, a Shaker -subunit that is silent when expressed alone, has been shown to regulate the activity of AKT1 by forming heteromeric AtKC1 KT1 channels. Here, we investigated whether AtKC1 is a general regulator of channel activity. Co-expression in Xenopus oocytes of a dominant negative (pore-mutated) AtKC1 subunit with the inward Shaker channel subunits KAT1, KAT2 or AKT2, or the outward subunits SKOR or GORK, revealed that the three inward subunits functionally interact with AtKC1 while the outward ones cannot. Localization experiments in plant protoplasts showed that KAT2 was able to re-locate AtKC1 fused to GFP from endomembranes to the plasma membrane, indicating that heteromeric AtKC1 AT2 channels are efficiently targeted to the plasma membrane. Functional properties of heteromeric channels involving AtKC1 and KAT1, KAT2 or AKT2 were analysed by voltage clamp after co-expression of the respective subunits in Xenopus oocytes. AtKC1 behaved as a regulatory subunit within the heterotetrameric channel, reducing the macroscopic conductance and negatively shifting the channel activation potential. Expression studies showed that AtKC1 and its identified Shaker partners have overlapping expression patterns, supporting the hypothesis of a general regulation of inward channel activity by AtKC1 in planta. Lastly, AtKC1 disruption appeared to reduce plant biomass production, showing that AtKC1-mediated channel activity regulation is required for normal plant growth.
JinYK, JingW, ZhangQ, ZhangWH ( 2015). Cyclic nucleotide gated channel 10 negatively regulates salt to- lerance by mediating Na + transport in Arabidopsis J Plant Res 128, 211-220. DOI:10.1007/s10265-014-0679-2URLPMID:25416933 [本文引用: 1] A number of cyclic nucleotide gated channel ( CNGC ) genes have been identified in plant genomes, but their functions are mainly undefined. In this study, we identified the role of CNGC10 in the response of Arabidopsis thaliana to salt stress. The cngc10 T-DNA insertion mutant showed greater tolerance to salt than wild-type A. thaliana during seed germination and seedling growth. The cngc10 mutant accumulated less Na + and K + , but not less Ca 2+ , in shoots in response to salt stress. By contrast, overexpression of CNGC10 resulted in greater sensitivity to salt stress, and complementation of this gene recovered salt sensitivity. In response to salt stress, heterologous expression of CNGC10 in the Na + sensitive yeast mutant strain B31 inhibited growth due to accumulation of Na + at a rate greater than that of yeast transformed with an empty vector. Quantitative RT-PCR analysis demonstrated that CNGC10 was expressed mainly in roots and flowers. GUS analysis of a root cross section indicated that CNGC10 was expressed mainly in the endodermis and epidermis. Furthermore, the expression of CNGC10 in roots was dramatically inhibited by exposure to 200 mM NaCl for 6 h. These data suggest that CNGC10 negatively regulates salt tolerance in A. thaliana and may be involved in mediating Na + transport.
JohanssonI, WulfetangeK, PoréeF, MichardE, Gajd- anowiczP, LacombeB, SentenacH, ThibaudJB, Mueller-RoeberB, BlattMR, DreyerI ( 2006). External K + modulates the activity of the Arabidopsis potassium chan- nel SKOR via an unusual mechanism Plant J 46, 269-281. DOI:10.1111/j.1365-313X.2006.02690.xURLPMID:16623889 [本文引用: 1] Plant outward-rectifying K+ channels mediate K+ efflux from guard cells during stomatal closure and from root cells into the xylem for root-shoot allocation of potassium (K). Intriguingly, the gating of these channels depends on the extracellular K+ concentration, although the ions carrying the current are derived from inside the cell. This K+ dependence confers a sensitivity to the extracellular K+ concentration ([K+]) that ensures that the channels mediate K+ efflux only, regardless of the [K+] prevailing outside. We investigated the mechanism of K+-dependent gating of the K+ channel SKOR of Arabidopsis by site-directed mutagenesis. Mutations affecting the intrinsic K+ dependence of gating were found to cluster in the pore and within the sixth transmembrane helix (S6), identifying an 'S6 gating domain' deep within the membrane. Mapping the SKOR sequence to the crystal structure of the voltage-dependent K+ channel KvAP from Aeropyrum pernix suggested interaction between the S6 gating domain and the base of the pore helix, a prediction supported by mutations at this site. These results offer a unique insight into the molecular basis for a physiologically important K+-sensory process in plants.
KamanoS, KumeS, IidaK, LeiKJ, NakanoM, NakayamaY, IidaH ( 2015). Transmembrane topologies of Ca 2+- permeable mechanosensitive channels MCA1 and MCA2 in Arabidopsis thaliana .J Biol Chem 290, 30901-30909. [本文引用: 3]
KangJM, MehtaS, TuranoFJ ( 2004). The putative glutamate receptor 1.1 (AtGLR1.1) in Arabidopsis thaliana regulates abscisic acid biosynthesis and signaling to con- trol development and water loss .Plant Cell Physiol 45, 1380-1389. [本文引用: 1]
KangJM, TuranoFJ ( 2003). The putative glutamate receptor 1.1 (AtGLR1.1) functions as a regulator of carbon and ni- trogen metabolism in Arabidopsis thaliana .Proc Natl Acad Sci USA 100, 6872-6877. DOI:10.1073/pnas.1030961100URLPMID:12738881 [本文引用: 1] The ability to coordinate carbon (C) and nitrogen (N) metabolism enables plants to regulate development and metabolic responses to different environmental conditions. The regulator(s) or sensor(s) that monitor crosstalk between biosynthetic pathways and ultimately control the flow of C or N through them have remained elusive. We used an antisense strategy to demonstrate that the putative glutamate receptor 1.1 (AtGLR1.1) functions as a regulator of C and N metabolism in Arabidopsis. Seeds from AtGLR1.1-deficient Arabidopsis (antiAtGLR1.1) lines did not germinate in the presence of an animal ionotropic glutamate receptor (iGLR) antagonist, but germination was restored upon coincubation with an iGLR agonist or the putative ligand glutamate. In antiAtGLR1.1 lines, endogenous abscisic acid (ABA) concentrations increased with iGLR antagonist treatments and decreased with coincubation with an iGLR agonist, suggesting that germination was controlled by ABA. antiAtGLR1.1 seedlings also exhibited sensitivity to increased levels of Ca2+compared with wild type, and they exhibited a conditional phenotype that was sensitive to the C:N ratio. In the presence of C, specifically sucrose, but not glucose, mannitol, or sorbitol, antiAtGLR1.1 seeds did not germinate, but germination was restored upon coincubation with NO3-, but not NH4+. Immunoblot, isoenzyme, and RT-PCR analyses indicate that AtGLR1.1 regulates the accumulation of distinct C- and N-metabolic enzymes, hexokinase 1 (HXK1) and zeaxanthin epoxidase (ABA1), by transcriptional control. We provide a model to describe the role of AtGLR1.1 in C/N metabolism and ABA biosynthesis, which in turn controls seed germination.
KauppUB, SeifertR ( 2002). Cyclic nucleotide-gated ion channels Physiol Rev 82, 769-824. DOI:10.1152/physrev.00008.2002URLPMID:12087135 [本文引用: 1] Cyclic nucleotide-gated (CNG) channels are nonselective cation channels first identified in retinal photoreceptors and olfactory sensory neurons (OSNs). They are opened by the direct binding of cyclic nucleotides, cAMP and cGMP. Although their activity shows very little voltage dependence, CNG channels belong to the superfamily of voltage-gated ion channels. Like their cousins the voltage-gated K+ channels, CNG channels form heterotetrameric complexes consisting of two or three different types of subunits. Six different genes encoding CNG channels, four A subunits (A1 to A4) and two B subunits (B1 and B3), give rise to three different channels in rod and cone photoreceptors and in OSNs. Important functional features of these channels, i.e., ligand sensitivity and selectivity, ion permeation, and gating, are determined by the subunit composition of the respective channel complex. The function of CNG channels has been firmly established in retinal photoreceptors and in OSNs. Studies on their presence in other sensory and nonsensory cells have produced mixed results, and their purported roles in neuronal pathfinding or synaptic plasticity are not as well understood as their role in sensory neurons. Similarly, the function of invertebrate homologs found in Caenorhabditis elegans, Drosophila,and Limulus is largely unknown, except for two subunits of C. elegans that play a role in chemosensation. CNG channels are nonselective cation channels that do not discriminate well between alkali ions and even pass divalent cations, in particular Ca2+. Ca2+ entry through CNG channels is important for both excitation and adaptation of sensory cells. CNG channel activity is modulated by Ca2+/calmodulin and by phosphorylation. Other factors may also be involved in channel regulation. Mutations in CNG channel genes give rise to retinal degeneration and color blindness. In particular, mutations in the A and B subunits of the CNG channel expressed in human cones cause various forms of complete and incomplete achromatopsia.
KobayashiY, KobayashiY, SugimotoM, LakshmananV, IuchiS, KobayashiM, BaisHP, KoyamaH ( 2013). Characterization of the complex regulation of AtALMT1 expression in response to phytohormones and other indu- cers .Plant Physiol 162, 732-740. [本文引用: 1]
K?hlerC, MerkleT, NeuhausG ( 1999). Characterisation of a novel gene family of putative cyclic nucleotide- and calmodulin-regulated ion channels in Arabidopsis thaliana .Plant J 18, 97-104. DOI:10.1046/j.1365-313X.1999.00422.xURLPMID:10341447 [本文引用: 2] In plants, cyclic GMP is involved in signal transduction in response to light and gibberellic acid. For cyclic AMP, a potential role during the plant cell cycle was recently reported. However, cellular targets for cyclic nucleotides in plants are largely unknown. Here we report on the identification and characterisation of a new gene family in Arabidopsis, which share features with cyclic nucleotide-gated channels from animals and inward-rectifying K+ channels from plants. The identified gene family comprises six members (Arabidopsis thaliana cyclic nucleotide-gated channels, AtCNGC1-6) with significant homology among the deduced proteins. Hydrophobicity analysis predicted six membrane-spanning domains flanked by hydrophilic amino and carboxy termini. A putative cyclic nucleotide binding domain (CNBD) which contains several residues that are invariant in other CNBDs was located in the carboxy terminus. This domain overlaps with a predicted calmodulin (CaM) binding site, suggesting interaction between cyclic nucleotide and CaM regulation. We demonstrated interaction of the carboxy termini of AtCNGC1 and AtCNGC2 with CaM in yeast, indicating that the CaM binding sites are functional. Furthermore, it was shown that both AtCNGC1 and AtCNGC2 can partly complement the K(+)-uptake-deficient yeast mutant CY162. Therefore, we propose that the identified genes constitute a family of plant cyclic nucleotide- and CaM-regulated ion channels.
KovermannP, MeyerS, H?rtensteinerS, PiccoC, Scholz-StarkeJ, RaveraS, LeeY, MartinoiaE ( 2007). The Arabidopsis vacuolar malate channel is a member of the ALMT family Plant J 52, 1169-1180. DOI:10.1111/j.1365-313X.2007.03367.xURLPMID:18005230 [本文引用: 1] In plants, malate is a central metabolite and fulfills a large number of functions. Vacuolar malate may reach very high concentrations and fluctuate rapidly, whereas cytosolic malate is kept at a constant level allowing optimal metabolism. Recently, a vacuolar malate transporter ( Arabidopsis thaliana tonoplast dicarboxylate transporter, At tDT) was identified that did not correspond to the well-characterized vacuolar malate channel. We therefore hypothesized that a member of the aluminum-activated malate transporter (ALMT) gene family could code for a vacuolar malate channel. Using GFP fusion constructs, we could show that At ALMT9 ( A. thaliana ALMT9) is targeted to the vacuole. Promoter-GUS fusion constructs demonstrated that this gene is expressed in all organs, but is cell-type specific as GUS activity in leaves was detected nearly exclusively in mesophyll cells. Patch-clamp analysis of an Atalmt9 T-DNA insertion mutant exhibited strongly reduced vacuolar malate channel activity. In order to functionally characterize At ALMT9 as a malate channel, we heterologously expressed this gene in tobacco and in oocytes. Overexpression of At ALMT9-GFP in Nicotiana benthamiana leaves strongly enhanced the malate current densities across the mesophyll tonoplasts. Functional expression of At ALMT9 in Xenopus oocytes induced anion currents, which were clearly distinguishable from endogenous oocyte currents. Our results demonstrate that At ALMT9 is a vacuolar malate channel. Deletion mutants for At ALMT9 exhibit only slightly reduced malate content in mesophyll protoplasts and no visible phenotype, indicating that At tDT and the residual malate channel activity are sufficient to sustain the transport activity necessary to regulate the cytosolic malate homeostasis.
KwakJM, MurataY, Baizabal-AguirreVM, MerrillJ, WangM, KemperA, HawkeSD, TallmanG, SchroederJI ( 2001). Dominant negative guard cell K + channel mutants reduce inward-rectifying K + currents and light-induced stomatal opening in Arabidopsis Plant Physiol 127, 473-485. [本文引用: 1]
LaanemetsK, WangYF, LindgrenO, WuJY, NishimuraN, LeeS, CaddellD, MeriloE, BroscheM, KilkK, SoometsU, Kangasj?rviJ, SchroederJI, KollistH ( 2013). Mutations in the SLAC1 anion channel slow stomatal opening and severely reduce K + uptake channel activity via enhanced cytosolic [Ca 2+] and increased Ca 2+ sensitivity of K + uptake channels New Phytol 197, 88-98. [本文引用: 1]
LadwigF, DahlkeRI, StührwohldtN, HartmannJ, HarterK, SauterM ( 2015). Phytosulfokine regulates growth in Arabidopsis through a response module at the plasma membrane that includes CYCLIC NUCLEOTIDE-GATED CHANNEL 17, H +-ATPase, and BAK1 Plant Cell 27, 1718-1729. DOI:10.1105/tpc.15.00306URLPMID:26071421 [本文引用: 1] Phytosulfokine (PSK) is perceived by the leucine-rich repeat receptor kinase PSKR1 and promotes growth in Arabidopsis thaliana. PSKR1 is coexpressed with the CYCLIC NUCLEOTIDE-GATED CHANNEL gene CNGC17. PSK promotes protoplast expansion in the wild type but not in cngc17. Protoplast expansion is likewise promoted by cGMP in a CNGC17-dependent manner. Furthermore, PSKR1-deficient protoplasts do not expand in response to PSK but are still responsive to cGMP, suggesting that cGMP acts downstream of PSKR1. Mutating the guanylate cyclase center of PSKR1 impairs seedling growth, supporting a role for PSKR1 signaling via cGMP in planta. While PSKR1 does not interact directly with CNGC17, it interacts with the plasma membrane-localized H(+)-ATPases AHA1 and AHA2 and with the BRI-associated receptor kinase 1 (BAK1). CNGC17 likewise interacts with AHA1, AHA2, and BAK1, suggesting that PSKR1, BAK1, CNGC17, and AHA assemble in a functional complex. Roots of deetiolated bak1-3 and bak1-4 seedlings were unresponsive to PSK, and bak1-3 and bak1-4 protoplasts expanded less in response to PSK but were fully responsive to cGMP, indicating that BAK1 acts in the PSK signal pathway upstream of cGMP. We hypothesize that CNGC17 and AHAs form a functional cation-translocating unit that is activated by PSKR1/BAK1 and possibly other BAK1/RLK complexes.
LakshmananV, KittoSL, CaplanJL, HsuehYH, KearnsDB, WuYS, BaisHP ( 2012). Microbe-associated mole- cular patterns-triggered root responses mediate beneficial rhizobacterial recruitment in Arabidopsis Plant Physiol 160, 1642-1661. DOI:10.1104/pp.112.200386URL [本文引用: 1] This study demonstrated that foliar infection by Pseudomonas syringae pv tomato DC3000 induced malic acid (MA) transporter (ALUMINUM-ACTIVATED MALATE TRANSPORTER1 [ALMT1]) expression leading to increased MA titers in the rhizosphere of Arabidopsis (Arabidopsis thaliana). MA secretion in the rhizosphere increased beneficial rhizobacteria Bacillus subtilis (hereafter FB17) titers causing an induced systemic resistance response in plants against P. syringae pv tomato DC3000. Having shown that a live pathogen could induce an intraplant signal from shoot-to-root to recruit FB17 belowground, we hypothesized that pathogen-derived microbe-associated molecular patterns (MAMPs) may relay a similar response specific to FB17 recruitment. The involvement of MAMPs in triggering plant innate immune response is well studied in the plant's response against foliar pathogens. In contrast, MAMPs-elicited plant responses on the roots and the belowground microbial community are not well understood. It is known that pathogen-derived MAMPs suppress the root immune responses, which may facilitate pathogenicity. Plants subjected to known MAMPs such as a flagellar peptide, flagellin22 (flg22), and a pathogen-derived phytotoxin, coronatine (COR), induced a shoot-to-root signal regulating ALMT1 for recruitment of FB17. Micrografts using either a COR-insensitive mutant (coil) or a flagellin-insensitive mutant (fls2) as the scion and ALMT1 pro :脽-glucuronidase as the rootstock revealed that both COR and flg22 are required for a graft transmissible signal to recruit FB17 belowground. The data suggest that MAMPs-induced signaling to regulate ALMT1 is salicylic acid and JASMONIC ACID RESISTANT1 (JAR1)/JASMONATE INSENSITIVE1 (JIN1)/MYC2 independent. Interestingly, a cell culture filtrate of FB17 suppressed flg22-induced MAMPsactivated root defense responses, which are similar to suppression of COR-mediated MAMPs-activated root defense, revealing a diffusible bacterial component that may regulate plant immune responses. Further analysis showed that the biofilm formation in B. subtilis negates suppression of MAMPs-activated defense responses in roots. Moreover, B. subtilis suppression of MAMPs-activated root defense does require JAR1/JIN1/MYC2. The ability of FB17 to block the MAMPselicited signaling pathways related to antibiosis reflects a strategy adapted by FB17 for efficient root colonization. These experiments demonstrate a remarkable strategy adapted by beneficial rhizobacteria to suppress a host defense response, which may facilitate rhizobacterial colonization and host-mutualistic association.
LamHM, ChiuJ, HsiehMH, MeiselL, OliveiraIC, ShinM, CoruzziG ( 1998). Glutamate-receptor genes in plants Nature 396, 125-126. DOI:10.1038/24066URLPMID:9823891 [本文引用: 4] Glutamate receptor-like genes (GLR) have been described in a number of plant species including Arabidopsis thaliana (Lam etal., 1998; Kim etal., 2001; Lacombe etaL, 2001) and Brassica napus (Genbank accession number AF109392). These GLR genes have similarity in primary sequence and domain organization to ionotropic glutamate receptors (iGluRs) in animals. At the moment, plant genes that have... [Show full abstract]
LanWZ, LeeSC, CheYF, JiangYQ, LuanS ( 2011). Mechanistic analysis of AKT1 regulation by the CBL-CIPK- PP2CA interactions Mol Plant 4, 527-536. DOI:10.1093/mp/ssr031URLPMID:21596690 [本文引用: 1] Arabidopsis K+ transporter 1 (AKT1) participates in K+ uptake in roots, especially under low-K conditions. We recently identified a Ca2+ signaling pathway consisting of multiple calcineurin B-like calcium sensors (CBLs) and multiple target kinases (CBL-interacting protein kinases or CIPKs) that phosphorylate and activate AKT1, whereas a specific PP2C-type phosphatase inactivates CIPK-dependent AKT1 activity. In this study, we analyzed the interactions between PP2Cs and the CBL–CIPK pathway and found previously unsuspected mechanisms underlying the CBL–CIPK–PP2C signaling processes. The interaction between the CIPKs and PP2Cs involves the kinase domain of the CIPK component, in addition to the protein phosphatase interacting motif (PPI) in the regulatory domain. Furthermore, specific CBLs physically interact with and inactivate PP2C phosphatases to recover the CIPK-dependent AKT1 channel activity. These findings provide further insights into the signaling network consisting of CBL–CIPK–PP2C interactions in the activation of the AKT1 channel.
LebaudyA, HosyE, SimonneauT, SentenacH, ThibaudJB, DreyerI ( 2008). Heteromeric K + channels in plants Plant J 54, 1076-1082. DOI:10.1111/j.1365-313X.2008.03479.xURLPMID:18346194 [本文引用: 2] Voltage-gated potassium channels of plants are multimeric proteins built of four subunits. In the model plant Arabidopsis thaliana , nine genes coding for K + channel subunits have been identified. When co-expressed in heterologous expression systems, most of them display the ability to form heteromeric K + channels. Till now it was not clear whether plants use this potential of heteromerization to increase the functional diversity of potassium channels. Here, we designed an experimental approach employing different transgenic plant lines that allowed us to prove the existence of heteromeric K + channels in plants. The chosen strategy might also be useful for investigating the activity and function of other multimeric channel proteins like, for instance, cyclic-nucleotide gated channels, tandem-pore K + channels and glutamate receptor channels.
LebaudyA, PascaudF, VeryAA, AlconC, DreyerI, ThibaudJB, LacombeB ( 2010). Preferential KAT1-KAT2 heteromerization determines inward K + current properties in Arabidopsis guard cells J Biol Chem 285, 6265-6274. DOI:10.1074/jbc.M109.068445URLPMID:2825422 [本文引用: 2] Guard cells adjust their volume by changing their ion content due to intense fluxes that, for K(+), are believed to flow through inward or outward Shaker channels. Because Shaker channels can be homo- or heterotetramers and Arabidopsis guard cells express at least five genes encoding inward Shaker subunits, including the two major ones, KAT1 and KAT2, the molecular identity of inward Shaker channels operating therein is not yet completely elucidated. Here, we first addressed the properties of KAT1-KAT2 heteromers by expressing KAT1-KAT2 tandems in Xenopus oocytes. Then, computer analyses of the data suggested that coexpression of free KAT1 and KAT2 subunits resulted mainly in heteromeric channels made of two subunits of each type due to some preferential association of KAT1-KAT2 heterodimers at the first step of channel assembly. This was further supported by the analysis of KAT2 effect on KAT1 targeting in tobacco cells. Finally, patch-clamp recordings of native inward channels in wild-type and mutant genotypes strongly suggested that this preferential heteromerization occurs in planta and that Arabidopsis guard cell inward Shaker channels are mainly heteromers of KAT1 and KAT2 subunits.
LebaudyA, VavasseurA, HosyE, DreyerI, LeonhardtN, ThibaudJB, VéryAA, SimonneauT, SentenacH ( 2008). Plant adaptation to fluctuating environment and biomass production are strongly dependent on guard cell potassium channels Proc Natl Acad Sci USA 105, 5271-5276. DOI:10.1073/pnas.0709732105URLPMID:18367672 [本文引用: 3] At least four genes encoding plasma membrane inward K62 channels (${\rm K}_{{\rm in}}$ channels) are expressed in Arabidopsis guard cells. A double mutant plant was engineered by disruption of a major ${\rm K}_{{\rm in}}$ channel gene and expression of a dominant negative channel construct. Using the patch-clamp technique revealed that this mutant was totally deprived of guard cell ${\rm K}_{{\rm in}}$ channel $({\rm GCK}_{{\rm in}})$ activity, providing a model to investigate the roles of this activity in the plant. ${\rm GCK}_{{\rm in}}$ activity was found to be an essential effector of stomatal opening triggered by membrane hyperpolarization and thereby of blue light-induced stomatal opening at dawn. It improved stomatal reactivity to external or internal signals (light, CO60 availability, and evaporative demand). It protected stomatal function against detrimental effects of Na62 when plants were grown in the presence of physiological concentrations of this cation, probably by enabling guard cells to selectively and rapidly take up K62 instead of Na62 during stomatal opening, thereby preventing deleterious effects of Na62 on stomatal closure. It was also shown to be a key component of the mechanisms that underlie the circadian rhythm of stomatal opening, which is known to gate stomatal responses to extracellular and intracellular signals. Finally, in a meteorological scenario with higher light intensity during the first hours of the photophase, ${\rm GCK}_{{\rm in}}$ activity was found to allow a strong increase (35%) in plant biomass production. Thus, a large diversity of approaches indicates that ${\rm GCK}_{{\rm in}}$ activity plays pleiotropic roles that crucially contribute to plant adaptation to fluctuating and stressing natural environments.
LebaudyA, VéryAA, SentenacH ( 2007). K + channel activity in plants: genes, regulations and functions FEBS Lett 581, 2357-2366. DOI:10.1016/j.febslet.2007.03.058URLPMID:17418142 [本文引用: 2] Potassium (K +) is the most abundant cation in the cytosol, and plant growth requires that large amounts of K + are transported from the soil to the growing organs. K + uptake and fluxes within the plant are mediated by several families of transporters and channels. Here, we describe the different families of K +-selective channels that have been identified in plants, the so-called Shaker, TPK and Kir-like channels, and what is known so far on their regulations and physiological functions in the plant.
LeeSC, LanWZ, BuchananBB, LuanS ( 2009). A protein kinase-phosphatase pair interacts with an ion channel to regulate ABA signaling in plant guard cells Proc Natl Acad Sci USA 106, 21419-21424. DOI:10.1073/pnas.0910601106URLPMID:19955427 [本文引用: 2] The plant hormone abscisic acid (ABA) serves as a physiological monitor to assess the water status of plants and, under drought conditions, induces stomatal pore closure by activating specific ion channels, such as a slow-anion channel (SLAC1) that, in turn, mediate ion efflux from the guard cells. Earlier genetic analyses uncovered a protein kinase (OST1) and several 2C-type phosphatases, as respective positive and negative regulators of ABA-induced stomatal closure. Here we show that the OST1 kinase interacts with the SLAC1 anion channel, leading to its activation via phosphorylation. PP2CA, one of the PP2C phosphatase family members acts in an opposing manner and inhibits the activity of SLAC1 by two mechanisms: (1) direct interaction with SLAC1 itself, and (2) physical interaction with OSTI leading to inhibition of the kinase independently of phosphatase activity. The results suggest that ABA signaling is mediated by a physical interaction chain consisting of several components, including a PP2C member, SnRK2-type kinase (OST1), and an ion channel, SLAC1, to regulate stomatal movements. The findings are in keeping with a paradigm in which a protein kinase-phosphatase pair interacts physically with a target protein to couple a signal with a specific response.
LefoulonC, BoeglinM, MoreauB, VéryAA, SzponarskiW, DauzatM, MichardE, GaillardI, ChérelI ( 2016). The Arabidopsis AtPP2CA protein phosphatase inhibits the GORK K + efflux channel and exerts a dominant suppres- sive effect on phosphomimetic-activating mutations J Biol Chem 291, 6521-6533. [本文引用: 1]
LengQ, MercierRW, HuaBG, FrommH, BerkowitzGA ( 2002). Electrophysiological analysis of cloned cyclic nucleotide-gated ion channels Plant Physiol 128, 400-410. DOI:10.1104/pp.010832URLPMID:11842144 [本文引用: 1] Electrophysiological studies were conducted on the cloned plant cyclic nucleotide-gated ion channels AtCNGC2 and AtCNGC1 from Arabidopsis, and NtCBP4 from tobacco (Nicotiana tobacum). The nucleotide coding sequences for these proteins were expressed in Xenopus laevis oocytes or HEK 293 cells. Channel characteristics were evaluated using voltage clamp analysis of currents in the presence of cAMP. AtCNGC2 was demonstrated to conduct K+and other monovalent cations, but exclude Na+; this conductivity profile is unique for any ion channel not possessing the amino acid sequence found in the selectivity filter of K+-selective ion channels. Application of cAMP evoked currents in membrane patches of oocytes injected with AtCNGC2 cRNA. Direct activation of the channel by cyclic nucleotide, demonstrated by application of cyclic nucleotide to patches of membranes expressing such channels, is a hallmark characteristic of this ion channel family. Voltage clamp studies (two-electrode configuration) demonstrated that AtCNGC1 and NtCBP4 are also cyclic nucleotide-gated channels. Addition of a lipophilic analog of cAMP to the perfusion bath of oocytes injected with NtCBP4 and AtCNGC1 cRNAs induced inward rectified, noninactivating K+currents.
LengQ, MercierRW, YaoWZ, BerkowitzGA ( 1999). Cloning and first functional characterization of a plant cyclic nucleotide-gated cation channel Plant Physiol 121, 753-761. DOI:10.1104/pp.121.3.753URLPMID:10557223 [本文引用: 1] Cyclic nucleotide-gated (cng) non-selective cation channels have been cloned from a number of animal systems. These channels are characterized by direct gating upon cAMP or cGMP binding to the intracellular portion of the channel protein, which leads to an increase in channel conductance. Animal cng channels are involved in signal transduction systems; they translate stimulus-induced changes in cytosolic cyclic nucleotide into altered cell membrane potential and/or cation flux as part of a signal cascade pathway. Putative plant homologs of animal cng channels have been identified. However, functional characterization (i.e. demonstration of cyclic-nucleotide-dependent ion currents) of a plant cng channel has not yet been accomplished. We report the cloning and first functional characterization of a plant member of this family of ion channels. The Arabidopsis cDNA AtCNGC2 encodes a polypeptide with deduced homology to the subunit of animal channels, and facilitates cyclic nucleotide-dependent cation currents upon expression in a number of heterologous systems. AtCNGC2 expression in a yeast mutant lacking a low-affinity K+uptake system complements growth inhibition only when lipophilic cyclic nucleotides are present in the culture medium. Voltage clamp analysis indicates that Xenopus laevis oocytes injected with AtCNGC2 cRNA demonstrate cyclic-nucleotide-dependent, inward-rectifying K+currents. Human embryonic kidney cells (HEK293) transfected with AtCNGC2 cDNA demonstrate increased permeability to Ca2+only in the presence of lipophilic cyclic nucleotides. The evidence presented here supports the functional classification of AtCNGC2 as a cyclic-nucleotide-gated cation channel, and presents the first direct evidence (to our knowledge) identifying a plant member of this ion channel family.
LiJ, LongY, QiGN, LiJ, XuZJ, WuWH, WangY ( 2014). The Os-AKT1 channel is critical for K + uptake in rice roots and is modulated by the rice CBL1-CIPK23 complex Plant Cell 26, 3387-3402. DOI:10.1105/tpc.114.123455URLPMID:25096783 [本文引用: 1] Potassium (K(+)) is one of the essential nutrient elements for plant growth and development. Plants absorb K(+) ions from the environment via root cell K(+) channels and/or transporters. In this study, the Shaker K(+) channel Os-AKT1 was characterized for its function in K(+) uptake in rice (Oryza sativa) roots, and its regulation by Os-CBL1 (Calcineurin B-Like protein1) and Os-CIPK23 (CBL-Interacting Protein Kinase23) was investigated. As an inward K(+) channel, Os-AKT1 could carry out K(+) uptake and rescue the low-K(+)-sensitive phenotype of Arabidopsis thaliana akt1 mutant plants. Rice Os-akt1 mutant plants showed decreased K(+) uptake and displayed an obvious low-K(+)-sensitive phenotype. Disruption of Os-AKT1 significantly reduced the K(+) content, which resulted in inhibition of plant growth and development. Similar to the AKT1 regulation in Arabidopsis, Os-CBL1 and Os-CIPK23 were identified as the upstream regulators of Os-AKT1 in rice. The Os-CBL1-Os-CIPK23 complex could enhance Os-AKT1-mediated K(+) uptake. A phenotype test confirmed that Os-CIPK23 RNAi lines exhibited similar K(+)-deficient symptoms as the Os-akt1 mutant under low K(+) conditions. These findings demonstrate that Os-AKT1-mediated K(+) uptake in rice roots is modulated by the Os-CBL1-Os-CIPK23 complex.
LiLG, KimBG, CheongYH, PandeyGK, LuanS ( 2006). A Ca 2+ signaling pathway regulates a K + channel for low-K response in Arabidopsis Proc Natl Acad Sci USA 103, 12625-12630. [本文引用: 1]
LiangCY, Pi?erosMA, TianJ, YaoZF, SunLL, LiuJP, ShaffJ, ColuccioA, KochianLV, LiaoH ( 2013). Low pH, aluminum, and phosphorus coordinately regulate malate exudation through GmALMT1 to improve soybean adap- tation to acid soils .Plant Physiol 161, 1347-1361. DOI:10.1104/pp.112.208934URLPMID:23341359 [本文引用: 2] Low pH, aluminum (Al) toxicity, and low phosphorus (P) often coexist and are heterogeneously distributed in acid soils. To date, the underlying mechanisms of crop adaptation to these multiple factors on acid soils remain poorly understood. In this study, we found that P addition to acid soils could stimulate Al tolerance, especially for the P-efficient genotype HN89. Subsequent hydroponic studies demonstrated that solution pH, Al, and P levels coordinately altered soybean (Glycine max) root growth and malate exudation. Interestingly, HN89 released more malate under conditions mimicking acid soils (low pH, +P, and +Al), suggesting that root malate exudation might be critical for soybean adaptation to both Al toxicity and P deficiency on acid soils. GmALMT1, a soybean malate transporter gene, was cloned from the Al-treated root tips of HN89. Like root malate exudation, GmALMT1 expression was also pH dependent, being suppressed by low pH but enhanced by Al plus P addition in roots of HN89. Quantitative real-time PCR, transient expression of a GmALMT1-yellow fluorescent protein chimera in Arabidopsis protoplasts, and electrophysiological analysis of Xenopus laevis oocytes expressing GmALMT1 demonstrated that GmALMT1 encodes a root cell plasma membrane transporter that mediates malate efflux in an extracellular pH-dependent and Al-independent manner. Overexpression of GmALMT1 in transgenic Arabidopsis, as well as overexpression and knockdown of GmALMT1 in transgenic soybean hairy roots, indicated that GmALMT1-mediated root malate efflux does underlie soybean Al tolerance. Taken together, our results suggest that malate exudation is an important component of soybean adaptation to acid soils and is coordinately regulated by three factors, pH, Al, and P, through the regulation of GmALMT1 expression and GmALMT1 function.
LigabaA, KatsuharaM, RyanPR, ShibasakaM, Matsu- motoH ( 2006). The BnALMT1 and BnALMT2 genes from rape encode aluminum-activated malate transporters that enhance the aluminum resistance of plant cells .Plant Phy- siol 142, 1294-1303. DOI:10.1104/pp.106.085233URL [本文引用: 1] The release of organic anions from roots can protect plants from aluminum (Al) toxicity and help them overcome phosphorus (P) deficiency. Our previous findings showed that Al treatment induced malate and citrate efflux from rape (Brassica napus) roots, and that P deficiency did not induce the efflux. Since this response is similar to the malate efflux from wheat (Triticum aestivum) that is controlled by the TaALMT1 gene, we investigated whether homologs of TaALMT1 are present in rape and whether they are involved in the release of organic anions. We isolated two TaALMT1 homologs from rape designated BnALMT1 and BnALMT2 (B. napus Al-activated malate transporter). The expression of these genes was induced in roots, but not shoots, by Al treatment but P deficiency had no effect. Several other cations (lanthanum, ytterbium, and erbium) also increased BnALMT1 and BnALMT2 expression in the roots. The function of the BnALMT1 and BnALMT2 proteins was investigated by heterologous expression in cultured tobacco (Nicotiana tabacum) cells and in Xenopus laevis oocytes. Both transfection systems showed an enhanced capacity for malate efflux but not citrate efflux, when exposed to Al. Smaller malate fluxes were also activated by ytterbium and erbium treatment. Transgenic tobacco cells grew significantly better than control cells following an 18 h treatment with Al, indicating that the expression of BnALMT1 and BnALMT2 increased the resistance of these plant cells to Al stress. This report demonstrates that homologs of the TaALMT1 gene from wheat perform similar functions in other species.
LigabaA, MaronL, ShaffJON, KochianL, Pi?erosM ( 2012). Maize ZmALMT2 is a root anion transporter that mediates constitutive root malate efflux Plant Cell Environ 35, 1185-1200. DOI:10.1111/j.1365-3040.2011.02479.xURLPMID:22211473 [本文引用: 1] Root efflux of organic acid anions underlies a major mechanism of plant aluminium (Al) tolerance on acid soils. This efflux is mediated by transporters of the Al-activated malate transporter (ALMT) or the multi-drug and toxin extrusion (MATE) families. ZmALMT2 was previously suggested to be involved in Al tolerance based on joint association-linkage mapping for maize Al tolerance. In the current study, we functionally characterized ZmALMT2 by heterologously expressing it in Xenopus laevis oocytes and transgenic Arabidopsis. In oocytes, ZmALMT2 mediated an Al-independent electrogenic transport product of organic and inorganic anion efflux. Ectopic overexpression of ZmALMT2 in an Al-hypersensitive Arabidopsis KO/KD line lacking the Al tolerance genes, AtALMT1 and AtMATE, resulted in Al-independent constitutive root malate efflux which partially restored the Al tolerance phenotype. The lack of correlation between ZmALMT2 expression and Al tolerance (e.g., expression not localized to the root tip, not up-regulated by Al, and higher in sensitive versus tolerance maize lines) also led us to question ZmALMT2's role in Al tolerance. The functional properties of the ZmALMT2 transporter presented here, along with the gene expression data, suggest that ZmALMT2 is not involved in maize Al tolerance but, rather, may play a role in mineral nutrient acquisition and transport.
LindC, DreyerI, López-SanjurjoEJ, von MeyerK, IshizakiK, KohchiT, LangD, ZhaoY, KreuzerI, Al-RasheidKAS, RonneH, ReskiR, ZhuJK, GeigerD, HedrichR ( 2015). Stomatal guard cells co-opted an ancient ABA-dependent desiccation survival system to regulate stomatal closure Curr Biol 25, 928-935. DOI:10.1016/j.cub.2015.01.067URLPMID:25802151 [本文引用: 1] Fast drought/ABA-signaling in guard cells of higher plants involves the SnRK2 kinase OST1 activating the anion channel SLAC1. Here, Lind et al. show that during evolution of early land plants, SLAC1 co-opted the ancient ABA-signaling pathway by developing motifs at its cytosolic termini that allow ABA-activated OSTs to regulate SLAC1 activity.
LiuK, LiLG, LuanS ( 2006). Intracellular K + sensing of SKOR, a Shaker-type K + channel from Arabidopsis Plant J 46, 260-268. DOI:10.1111/j.1365-313X.2006.02689.xURLPMID:16623888 [本文引用: 2] Most K + channels in plants are structurally classified into the Shaker family named after the shaker K + channel in Drosophila . Plant K + channels function in many physiological processes including osmotic regulation and K + nutrition. An outwardly rectifying K + channel, SKOR, mediates the delivery of K + from stelar cells to the xylem in the roots, a critical step in the long-distance distribution of K + from roots to the upper parts of the plant. Here we report that SKOR channel activity is strictly dependent on intracellular K + concentrations. Activation by K + did not affect the kinetics of voltage dependence in SKOR, indicating that a voltage-independent gating mechanism underlies the K + sensing process. Further analysis showed that the C-terminal non-transmembrane region of the SKOR protein was required for this sensing process. The intracellular K + sensing mechanism couples SKOR activity to K + nutrition status in the 'source cells', thereby establishing a supply-based unloading system for the regulation of K + distribution.
LuGH, WangXP, LiuJH, YuK, GaoY, LiuHY, WangCG, WangW, WangGK, LiuM, MaoGF, LiBF, QinJY, XiaM, ZhouJL, LiuJM, JiangSQ, MoH, CuiJT, NagasawaN, SivasankarS, AlbertsenMC, SakaiH, MazurBJ, LassnerMW, BroglieRM ( 2014). Application of T-DNA activation tagging to identify glutamate receptor-like genes that enhance drought tolerance in plants Plant Cell Rep 33, 617-631. DOI:10.1007/s00299-014-1586-7URLPMID:24682459 [本文引用: 1] Key message A high-quality rice activation tagging population has been developed and screened for drought-tolerant lines using various water stress assays. One drought-tolerant line activated two...
LuM, ZhangYY, TangSK, PanJB, YuYK, HanJ, LiYY, DuXH, NanZJ, SunQP ( 2016). AtCNGC2 is involved in jasmonic acid-induced calcium mobilization J Exp Bot 67, 809-819. DOI:10.1093/jxb/erv500URLPMID:26608645 [本文引用: 1] Calcium (Ca(2+)) mobilization is a central theme in various plant signal transduction pathways. We demonstrate that Arabidopsis thaliana cyclic nucleotide-gated channel 2 (AtCNGC2) is involved in jasmonic acid (JA)-induced apoplastic Ca(2+) influx in Arabidopsis epidermal cells. Ca(2+) imaging results showed that JA can induce an elevation in the cytosolic cAMP concentration ([cAMP]cyt), reaching a maximum within 3min. Dibutyryl cAMP (db-cAMP), a cell membrane-permeable analogue of cAMP, induced an increase in the cytosolic Ca(2+) concentration ([Ca(2+)]cyt), with a peak at 4min. This [Ca(2+)]cyt increase was triggered by the JA-induced increase in [cAMP]cyt. W-7[N-(6-aminohexyl)-5-chloro-1-naphthalenesulfonamide], an antagonist of calmodulin, positively modulated the JA-induced increase in [Ca(2+)]cyt, while W-5[N-(6-aminohexyl)-1-naphthalenesulfonamide], an inactive antagonist of calmodulin, had no apparent effect. db-cAMP and JA positively induced the expression of primary (i.e. JAZ1 and MYC2) and secondary (i.e. VSP1) response genes in the JA signalling pathway in wild-type Arabidopsis thaliana, whereas they had no significant effect in the AtCNGC2 mutant 'defense, no death (dnd1) plants. These data provide evidence that JA first induces the elevation of cAMP, and cAMP, as an activating ligand, activates the AtCNGC2 channel, resulting in apoplastic Ca(2+) influx through AtCNGC2.
MaW, SmigelA, WalkerRK, MoederW, YoshiokaK, BerkowitzGA ( 2010). Leaf senescence signaling: the Ca 2+-conducting Arabidopsis cyclic nucleotide gated chan- nel 2 acts through nitric oxide to repress senescence pro- gramming Plant Physiol 154, 733-743. [本文引用: 1]
MacKinnonR ( 1991). Determination of the subunit stoi- chiometry of a voltage-activated potassium channel Na- ture 350, 232-235. DOI:10.1038/350232a0URLPMID:1706481 [本文引用: 1] The voltage-activated K+, Na+ and Ca2+ channels are responsible for the generation and propagation of electrical signals in cell membranes. The K+ channels are multimeric membrane proteins formed by the aggregation of an unknown number of independent subunits. By studying the interaction of a scorpion toxin with coexpressed wild-type and toxin-insensitive mutant Shaker K+ channels, the subunit stoichiometry can be determined. The Shaker K+ channel is found to have a tetrameric structure. This is consistent with the sequence relationship between a K+ channel and each of the four internally homologous repeats of Na+ and Ca2+ channels.
MacRobbieEAC, LettauJ ( 1980). Potassium content and aperture in “intact” stomatal and epidermal cells of Com- melina communis L .J Membr Biol 56, 249-256. [本文引用: 1]
MaierhoferT, LindC, HüttlS, ScherzerS, PapenfuβM, SimonJ, Al-RasheidKAS, AcheP, RennenbergH, HedrichR, MüllerTD, GeigerD ( 2014). A single-pore residue renders the Arabidopsis root anion channel SLAH2 highly nitrate selective Plant Cell 26, 2554-2567. DOI:10.1105/tpc.114.125849URLPMID:24938289 [本文引用: 2] In contrast to animal cells, plants use nitrate as a major source of nitrogen. Following the uptake of nitrate, this major macronutrient is fed into the vasculature for long-distance transport. The Arabidopsis thaliana shoot expresses the anion channel SLOW ANION CHANNEL1 (SLAC1) and its homolog SLAC1 HOMOLOGOUS3 (SLAH3), which prefer nitrate as substrate but cannot exclude chloride ions. By contrast, we identified SLAH2 as a nitrate-specific channel that is impermeable for chloride. To understand the molecular basis for nitrate selection in the SLAH2 channel, SLAC1 and SLAH2 were modeled to the structure of HiTehA, a distantly related bacterial member. Structure-guided site-directed mutations converted SLAC1 into a SLAH2-like nitrate-specific anion channel and vice versa. Our findings indicate that two pore-occluding phenylalanines constrict the pore. The selectivity filter of SLAC/SLAH anion channels is determined by the polarity of pore-lining residues located on alpha helix 3. Changing the polar character of a single amino acid side chain (Ser-228) to a nonpolar residue turned the nitrate-selective SLAH2 into a chloride/nitrate-permeable anion channel. Thus, the molecular basis of the anion specificity of SLAC/SLAH anion channels seems to be determined by the presence and constellation of polar side chains that act in concert with the two pore-occluding phenylalanines.
MaksaevG, HaswellES ( 2012). MscS-Like10 is a stretch- activated ion channel from Arabidopsis thaliana with a preference for anions .Proc Natl Acad Sci USA 109, 19015-19020. [本文引用: 1]
M?serP, ThomineS, SchroederJI, WardJM, HirschiK, SzeH, TalkeIN, AmtmannA, MaathuisFJM, SandersD, HarperJF, TchieuJ, GribskovM, PersansMW, SaltDE, KimSA, GuerinotML ( 2001). Phylogenetic relationships within cation transporter families of Arabidopsis Plant Phy- siol 126, 1646-1667. DOI:10.1104/pp.126.4.1646URLPMID:11500563 [本文引用: 1] Uptake and translocation of cationic nutrients play essential roles in physiological processes including plant growth, nutrition, signal transduction, and development. Approximately 5% of the Arabidopsis genome appears to encode membrane transport proteins. These proteins are classified in 46 unique families containing approximately 880 members. In addition, several hundred putative transporters have not yet been assigned to families. In this paper, we have analyzed the phylogenetic relationships of over 150 cation transport proteins. This analysis has focused on cation transporter gene families for which initial characterizations have been achieved for individual members, including potassium transporters and channels, sodium transporters, calcium antiporters, cyclic nucleotide-gated channels, cation diffusion facilitator proteins, natural resistance-associated macrophage proteins (NRAMP), and Zn-regulated transporter Fe-regulated transporter-like proteins. Phylogenetic trees of each family define the evolutionary relationships of the members to each other. These families contain numerous members, indicating diverse functions in vivo. Closely related isoforms and separate subfamilies exist within many of these gene families, indicating possible redundancies and specialized functions. To facilitate their further study, the PlantsT database (http://plantst.sdsc.edu) has been created that includes alignments of the analyzed cation transporters and their chromosomal locations.
MatulefK, ZagottaWN ( 2003). Cyclic nucleotide-gated ion channels Annu Rev Cell Dev Biol 19, 23-44. DOI:10.1146/annurev.cellbio.19.110701.154854URLPMID:14570562 [本文引用: 1] Abstract Cyclic nucleotide-gated (CNG) ion channels were first discovered in rod photoreceptors, where they are responsible for the primary electrical signal of the photoreceptor in response to light. CNG channels are highly specialized membrane proteins that open an ion-permeable pore across the membrane in response to the direct binding of intracellular cyclic nucleotides. CNG channels have been identified in a number of other tissues, including the brain, where their roles are only beginning to be appreciated. Recently, significant progress has been made in understanding the molecular mechanisms underlying their functional specializations. From these studies, a picture is beginning to emerge for how the binding of cyclic nucleotide is transduced into the opening of the pore and how this allosteric transition is modulated by various physiological effectors.
McAinshMR, PittmanJK ( 2009). Shaping the calcium signature New Phytol 181, 275-294. [本文引用: 1]
MeyerS, MummP, ImesD, EndlerA, WederB, Al-RasheidKAS, GeigerD, MartenI, MartinoiaE, HedrichR ( 2010). AtALMT12 represents an R-type anion channel required for stomatal movement in Arabidopsis guard cells Plant J 63, 1054-1062. [本文引用: 1]
MeyerS, Scholz-StarkeJ, De AngeliA, KovermannP, BurlaB, GambaleF, MartinoiaE ( 2011). Malate transport by the vacuolar AtALMT6 channel in guard cells is subject to multiple regulation Plant J 67, 247-257. DOI:10.1111/j.1365-313X.2011.04587.xURLPMID:21443686 [本文引用: 1] Gas exchange in plants is controlled by guard cells, specialized cells acting as turgor pressure-driven valves. Malate is one of the major anions accumulated inside the vacuole during stomatal opening counteracting the positive charge of potassium. AtALMT6, a member of the aluminum-activated malate transporter family, is expressed in guard cells of leaves and stems as well as in flower organs of Arabidopsis thaliana. An AtALMT6-GFP fusion protein was targeted to the vacuolar membrane both in transient and stable expression systems. Patch-clamp experiments on vacuoles isolated from AtALMT6-GFP over-expressing Arabidopsis plants revealed large inward-rectifying malate currents only in the presence of micromolar cytosolic calcium concentrations. Further analyses showed that vacuolar pH and cytosolic malate regulate the threshold of activation of AtALMT6-mediated currents. The interplay of these two factors determines the AtALMT6 function as a malate influx or efflux channel depending on the tonoplast potential. Guard cell vacuoles isolated from Atalmt6 knock-out plants displayed reduced malate currents compared with wild-type vacuoles. This reduction, however, was not accompanied by phenotypic differences in the stomatal movements in knock-out plants, probably because of functional redundancy of malate transporters in guard cell vacuoles.
MichardE, LimaPT, BorgesF, SilvaAC, PortesMT, CarvalhoJE, GillihamM, LiuLH, ObermeyerG, FeijóJA ( 2011). Glutamate receptor-like genes form Ca 2+ channels in pollen tubes and are regulated by pistil D-serine Sci- ence 332, 434-437. DOI:10.1126/science.1201101URLPMID:21415319 [本文引用: 1] Elevations in cytosolic free calcium concentration ([Ca(2+)](cyt)) constitute a fundamental signal transduction mechanism in eukaryotic cells, but the molecular identity of Ca(2+) channels initiating this signal in plants is still under debate. Here, we show by pharmacology and loss-of-function mutants that in tobacco and Arabidopsis, glutamate receptor-like channels (GLRs) facilitate Ca(2+) influx across the plasma membrane, modulate apical [Ca(2+)](cyt) gradient, and consequently affect pollen tube growth and morphogenesis. Additionally, wild-type pollen tubes grown in pistils of knock-out mutants for serine-racemase (SR1) displayed growth defects consistent with a decrease in GLR activity. Our findings reveal a novel plant signaling mechanism between male gametophyte and pistil tissue similar to amino acid-mediated communication commonly observed in animal nervous systems.
MoonSJ, KimHY, HwangH, KimJA, LeeY, MinMK, YoonIS, KwonTR, KimBG ( 2017). A dominant negative OsKAT2 mutant delays light-induced stomatal opening and improves drought tolerance without yield penalty in rice Front Plant Sci 8, 772. DOI:10.3389/fpls.2017.00772URLPMID:28553305 [本文引用: 1] Stomata are the main gateways for water and air transport between leaves and the environment. Inward-rectifying potassium channels regulate photo-induced stomatal opening. Rice contains three inward rectifying shaker-like potassium channel proteins, OsKAT1, OsKAT2, and OsKAT3. Among these, onlyOsKAT2is specifically expressed in guard cells. Here, we investigated the functions of OsKAT2 in stomatal regulation using three dominant negative mutant proteins, OsKAT2(T235R), OsKAT2(T285A) and OsKAT2(T285D), which are altered in amino acids in the channel pore and at a phosphorylation site. Yeast complementation and patch clamp assays showed that all three mutant proteins lost channel activity. However, among plants overexpressing these mutant proteins, only plants overexpressing OsKAT2(T235R) showed significantly less water loss than the control. Moreover, overexpression of this mutant protein led to delayed photo-induced stomatal opening and increased drought tolerance. Our results indicate that OsKAT2 is an inward- rectifying shaker-like potassium channel that mainly functions in stomatal opening. Interestingly, overexpression of OsKAT2(T235R) did not cause serious defects in growth or yield in rice, suggesting that OsKAT2 is a potential target for engineering plants with improved drought tolerance without yield penalty.
MoriK, RenhuN, NaitoM, NakamuraA, ShibaH, Yam- amotoT, SuzakiT, IidaH, MiuraK ( 2018). Ca 2+- permeable mechanosensitive channels MCA1 and MCA2 mediate cold-induced cytosolic Ca 2+ increase and cold tolerance in Arabidopsis Sci Rep 8, 550. DOI:10.1038/s41598-017-17483-yURL [本文引用: 1] Cold shock triggers an immediate rise in the cytosolic free calcium concentration ([Ca2+]cyt) inArabidopsis thalianaand this cold-induced elevation of [Ca2+]cytis inhibited by lanthanum or EGTA. It is suggested that intracellular calcium mainly contributes to the cold-induced [Ca2+]cytresponse by entering into the cytosol. Two calcium-permeable mechanosensitive channels, MCA1 and MCA2 (mid1-complementing activity), have been identified in Arabidopsis. Here, we demonstrate that MCA1 and MCA2 are involved in a cold-induced increase in [Ca2+]cyt. The cold-induced [Ca2+]cytincrease inmca1andmca2mutants was markedly lower than that in wild types. Themca1 mca2double mutant exhibited chilling and freezing sensitivity, compared to wild-type plants. Expression ofAt5g61820,At3g51660, andAt4g15490, which are not regulated by theCBF/DREB1stranscription factor, was down-regulated inmca1 mca2. These results suggest that MCA1 and MCA2 are involved in the cold-induced elevation of [Ca2+]cyt, cold tolerance, andCBF/DREB1-independent cold signaling.
MoulineK, VéryAA, GaymardF, BoucherezJ, PilotG, DevicM, BouchezD, ThibaudJB, SentenacH ( 2002). Pollen tube development and competitive ability are impaired by disruption of a Shaker K + channel in Arabi- dopsis Genes Dev 16, 339-350. [本文引用: 1]
NakagawaY, KatagiriT, ShinozakiK, QiZ, TatsumiH, FuruichiT, KishigamiA, SokabeM, KojimaI, SatoS, KatoT, TabataS, IidaK, TerashimaA, NakanoM, IkedaM, YamanakaT, IidaH ( 2007). Arabidopsis plasma mem- brane protein crucial for Ca 2+ influx and touch sensing in roots Proc Natl Acad Sci USA 104, 3639-3644. DOI:10.1073/pnas.0607703104URLPMID:17360695 [本文引用: 1] Plants can sense and respond to mechanical stimuli, like animals. An early mechanism of mechanosensing and response is speculated to be governed by as-yet-unidentified sensory complexes containing a Ca0562-permeable, stretch-activated (SA) channel. However, the components or regulators of such complexes are poorly understood at the molecular level in plants. Here, we report the molecular identification of a plasma membrane protein (designated Mca1) that correlates Ca0562 influx with mechanosensing in Arabidopsis thaliana. MCA1 cDNA was cloned by the functional complementation of lethality of a yeast mid1 mutant lacking a putative Ca0562-permeable SA channel component. Mca1 was localized to the yeast plasma membrane as an integral membrane protein and mediated Ca0562 influx. Mca1 also increased $[{\rm Ca}^{2+}]_{{\rm cyt}}$ upon plasma membrane distortion in Arabidopsis. The growth of MCA1-overexpressing plants was impaired in a high-calcium but not a low-calcium medium. The primary roots of mca1-null plants failed to penetrate a harder agar medium from a softer one. These observations demonstrate that Mca1 plays a crucial role in a Ca0562-permeable SA channel system that leads to mechanosensing in Arabidopsis. We anticipate our findings to be a starting point for a deeper understanding of the molecular mechanisms of mechanotransduction in plants.
NegiJ, MatsudaO, NagasawaT, ObaY, TakahashiH, Kawai-YamadaM, UchimiyaH, HashimotoM, IbaK ( 2008). CO2 regulator SLAC1 and its homologues are essential for anion homeostasis in plant cells Nature 452, 483-486. DOI:10.1038/nature06720URLPMID:18305482 [本文引用: 2] Abstract The continuing rise in atmospheric [CO2] is predicted to have diverse and dramatic effects on the productivity of agriculture, plant ecosystems and gas exchange. Stomatal pores in the epidermis provide gates for the exchange of CO2 and water between plants and the atmosphere, processes vital to plant life. Increased [CO2] has been shown to enhance anion channel activity proposed to mediate efflux of osmoregulatory anions (Cl- and malate(2-)) from guard cells during stomatal closure. However, the genes encoding anion efflux channels in plant plasma membranes remain unknown. Here we report the isolation of an Arabidopsis gene, SLAC1 (SLOW ANION CHANNEL-ASSOCIATED 1, At1g12480), which mediates CO2 sensitivity in regulation of plant gas exchange. The SLAC1 protein is a distant homologue of bacterial and fungal C4-dicarboxylate transporters, and is localized specifically to the plasma membrane of guard cells. It belongs to a protein family that in Arabidopsis consists of four structurally related members that are common in their plasma membrane localization, but show distinct tissue-specific expression patterns. The loss-of-function mutation in SLAC1 was accompanied by an over-accumulation of the osmoregulatory anions in guard cell protoplasts. Guard-cell-specific expression of SLAC1 or its family members resulted in restoration of the wild-type stomatal responses, including CO2 sensitivity, and also in the dissipation of the over-accumulated anions. These results suggest that SLAC1-family proteins have an evolutionarily conserved function that is required for the maintenance of organic/inorganic anion homeostasis on the cellular level.
NguyenTH, HuangSG, MeynardD, ChaineC, MichelR, RoelfsemaMRG, GuiderdoniE, SentenacH, VéryAA ( 2017). A dual role for the OsK5.2 ion channel in stomatal movements and K + loading into xylem sap Plant Physiol 174, 2409-2418. DOI:10.1104/pp.17.00691URLPMID:28626008 [本文引用: 1] The roles of potassium channels from the Shaker family in stomatal movements have been investigated by reverse genetics analyses in Arabidopsis (Arabidopsis thaliana), but corresponding information is lacking outside this model species. Rice (Oryza sativa) and other cereals possess stomata that are more complex than those of Arabidopsis. We examined the role of the outward Shaker K+ channel gene OsK5.2. Expression of the OsK5.2 gene (GUS reporter strategy) was observed in the whole stomatal complex (guard cells and subsidiary cells), root vasculature, and root cortex. In stomata, loss of OsK5.2 functional expression resulted in lack of time-dependent outward potassium currents in guard cells, higher rates of water loss through transpiration, and severe slowdown of stomatal closure. In line with the expression of OsK5.2 in the plant vasculature, mutant plants displayed a reduced K+ translocation from the root system toward the leaves via the xylem. The comparison between rice and Arabidopsis show that despite the strong conservation of Shaker family in plants, substantial differences can exist between the physiological roles of seemingly orthologous genes, as xylem loading depends on SKOR and stomatal closure on GORK in Arabidopsis, whereas both functions are executed by the single OsK5.2 Shaker in rice.
OoiA, Lemtiri-ChliehF, WongA, GehringC ( 2017). Direct modulation of the guard cell outward-rectifying potassium channel (GORK) by abscisic acid Mol Plant 10, 1469-1472. DOI:10.1016/j.molp.2017.08.010URLPMID:28844521 [本文引用: 1] @@
PilotG, LacombeB, GaymardF, ChérelI, BoucherezJ, ThibaudJB, SentenacH ( 2001). Guard cell inward K + channel activity in Arabidopsis involves expression of the twin channel subunits KAT1 and KAT2 J Biol Chem 276, 3215-3221. DOI:10.1074/jbc.M007303200URLPMID:11042178 [本文引用: 2] Abstract Stomatal opening, which controls gas exchanges between plants and the atmosphere, results from an increase in turgor of the two guard cells that surround the pore of the stoma. KAT1 was the only inward K(+) channel shown to be expressed in Arabidopsis guard cells, where it was proposed to mediate a K(+) influx that enables stomatal opening. We report that another Arabidopsis K(+) channel, KAT2, is expressed in guard cells. More than KAT1, KAT2 displays functional features resembling those of native inward K(+) channels in guard cells. Coexpression in Xenopus oocytes and two-hybrid experiments indicated that KAT1 and KAT2 can form heteromultimeric channels. The data indicate that KAT2 plays a crucial role in the stomatal opening machinery.
Pi?erosMA, Can?adoGMA, MaronLG, LyiSM, MenossiM, KochianLV ( 2008). Not all ALMT1-type transporters mediate aluminum-activated organic acid responses: the case of ZmALMT1—an anion-selective transporter .Plant J 53, 352-367. DOI:10.1111/j.1365-313X.2007.03344.xURLPMID:18069943 [本文引用: 1] The phytotoxic effects of aluminum (Al) on root systems of crop plants constitute a major agricultural problem in many areas of the world. Root exudation of Al-chelating molecules such as low-molecular-weight organic acids has been shown to be an important mechanism of plant Al tolerance/resistance. Differences observed in the physiology and electrophysiology of root function for two maize genotypes with contrasting Al tolerance revealed an association between rates of Al-activated root organic acid release and Al tolerance. Using these genotypes, we cloned ZmALMT1 , a maize gene homologous to the wheat ALMT1 and Arabidopsis AtALMT1 genes that have recently been described as encoding functional, Al-activated transporters that play a role in tolerance by mediating Al-activated organic acid exudation in roots. The ZmALMT1 cDNA encodes a 451 amino acid protein containing six transmembrane helices. Transient expression of a ZmALMT1 ::GFP chimera confirmed that the protein is targeted to the plant cell plasma membrane. We addressed whether ZmALMT1 might underlie the Al-resistance response (i.e. Al-activated citrate exudation) observed in the roots of the Al-tolerant genotype. The physiological, gene expression and functional data from this study confirm that ZmALMT1 is a plasma membrane transporter that is capable of mediating elective anion efflux and influx. However, gene expression data as well as biophysical transport characteristics obtained from Xenopus oocytes expressing ZmALMT1 indicate that this transporter is implicated in the selective transport of anions involved in mineral nutrition and ion homeostasis processes, rather than mediating a specific Al-activated citrate exudation response at the rhizosphere of maize roots.
PriceMB, JeleskoJ, OkumotoS ( 2012). Glutamate receptor homologs in plants: functions and evolutionary origins Front Plant Sci 3, 235. DOI:10.3389/fpls.2012.00235URL [本文引用: 3] The plant glutamate-like receptor homologs (GLRs) are homologs of mammalian ionotropic glutamate receptors (iGluRs) which were discovered more than 10 ears ago, and are hypothesized to be potential amino acid sensors in plants. Although initial progress on this gene family has been hampered by gene redundancy and technical issues such as gene toxicity; genetic, pharmacological, and electrophysiological approaches are starting to uncover the functions of this protein family. In parallel, there has been tremendous progress in elucidating the structure of animal glutamate receptors (iGluRs), which in turn will help understanding of the molecular mechanisms of plant GLR functions. In this review, we will summarize recent progress on the plant GLRs. Emerging evidence implicates plant GLRs in various biological processes in and beyond N sensing, and implies that there is some overlap in the signaling mechanisms of amino acids between plants and animals. Phylogenetic analysis using iGluRs from metazoans, plants, and bacteria showed that the plant GLRs are no more closely related to metazoan iGluRs as they are to bacterial iGluRs, indicating the separation of plant, other eukaryotic, and bacterial GLRs might have happened as early on as the last universal common ancestor. Structural similarities and differences with animal iGluRs, and the implication thereof, are also discussed.
QiGN, YaoFY, RenHM, SunSJ, TanYQ, ZhangZC, QiuBS, WangYF ( 2018). The S-type anion channel ZmSLAC1 plays essential roles in stomatal closure by mediating nitrate efflux in maize Plant Cell Physiol 59, 614-623. DOI:10.1093/pcp/pcy015URLPMID:29390155 [本文引用: 1] Abstract Diverse stimuli induce stomatal closure by triggering the efflux of osmotic anions, which is mainly mediated by the main anion channel SLAC1 in plants, and the anion permeability and selectivity of SLAC1 channels from several plant species have been reported to be variable. However, the genetic identity as well as the anion permeability and selectivity of the main S-type anion channel ZmSLAC1 in maize are still unknown. In this study, we identified GRMZM2G106921 as the gene encoding ZmSLAC1 in maize, and the maize mutants zmslac1-1 and zmslac1-2 harboring a mutator (Mu) transposon in ZmSLAC1 exhibited strong insensitive phenotypes of stomatal closure in response to diverse stimuli. We further found that ZmSLAC1 functions as a nitrate-selective anion channel without obvious permeability to chloride, sulfate and malate, clearly different from SLAC1 channels of Arabidopsis thaliana, Brassica rapa ssp. chinensis and Solanum lycopersicum L. Further experimental data show that the expression of ZmSLAC1 successfully rescued the stomatal movement phenotypes of the Arabidopsis double mutant atslac1-3atslah3-2 by mainly restoring nitrate-carried anion channel currents of guard cells. Together, these findings demonstrate that ZmSLAC1 is involved in stomatal closure mainly by mediating the efflux of nitrate in maize.
RameshSA, TyermanSD, XuB, BoseJ, KaurS, ConnV, DomingosP, UllahS, WegeS, ShabalaS, FeijóJA, RyanPR, GillhamM ( 2015). GABA signaling modulates plant growth by directly regulating the activity of plant- specific anion transporters Nat Commun 6, 7879. DOI:10.1038/ncomms8879URLPMID:26219411 [本文引用: 1] http://www.nature.com/doifinder/10.1038/ncomms8879
ReintanzB, SzyrokiA, IvashikinaN, AcheP, GoddeM, BeckerD, PalmeK, HedrichR ( 2002). AtKC1, a silent Arabidopsis potassium channel α-subunit modulates root hair K + influx Proc Natl Acad Sci USA 99, 4079-4084. DOI:10.1073/pnas.052677799URLPMID:11904452 [本文引用: 2] Ion channels in roots allow the plant to gain access to nutrients. The composition of the individual ion channels and the functional contribution of different α-subunits is largely unknown. Focusing on K+-selective ion channels, we have characterized AtKC1, a new α-subunit from the Arabidopsis shaker-like ion channel family. Promoter-β-glucuronidase (GUS) studies identified AtKC1 expression predominantly in root hairs and root endodermis. Specific antibodies recognized AtKC1 at the plasma membrane. To analyze further the abundance and the functional contribution of the different K+channels α-subunits in root cells, we performed real-time reverse transcription-PCR and patch-clamp experiments on isolated root hair protoplasts. Studying all shaker-like ion channel α-subunits, we only found the K+inward rectifier AtKC1 and AKT1 and the K+outward rectifier GORK to be expressed in this cell type. Akt1 knockout plants essentially lacked inward rectifying K+currents. In contrast, inward rectifying K+currents were present in AtKC1 knockout plants, but fundamentally altered with respect to gating and cation sensitivity. This indicates that the AtKC1 α-subunit represents an integral component of functional root hair K+uptake channels.
RenXL, QiGN, FengHQ, ZhaoS, ZhaoSS, WangY, WuWH ( 2013). Calcineurin B-like protein CBL10 directly interacts with AKT1 and modulates K + homeostasis in Ara- bidopsis Plant J 74, 258-266. DOI:10.1111/tpj.12123URLPMID:23331977 [本文引用: 1] Potassium transporters and channels play crucial roles in K(+) uptake and translocation in plant cells, which are essential for plant growth and development. AKT1 has been reported as an important K(+) channel in Arabidopsis roots involved in K(+) uptake. It is known that AKT1 is activated by a protein kinase CIPK23 interacting with two calcineurin B-like proteins CBL1/CBL9. The present study demonstrated that another CBL protein (CBL10) could also regulate AKT1 activity. The CBL10-overexpressing lines showed similarly sensitive phenotype as akt1 mutant under low K(+) conditions. In addition, the K(+) content of both CBL10-overexpressing lines and akt1 mutant plants were significantly reduced compared with wild-type plants. Moreover, CBL10 could directly interact with AKT1, which was verified in yeast two-hybrid, BiFC and Co-IP experiments. The results of electrophysiological analysis in both Xenopus oocytes and Arabidopsis root cell protoplasts demonstrated that CBL10 impairs AKT1-mediated inward K(+) currents. Furthermore, the results from yeast two-hybrid competition assay indicated that CBL10 may compete with CIPK23 for binding with AKT1 and negatively modulate AKT1 activity. The present study revealed a CIPK-independent regulatory mechanism of CBL proteins, which suggested that CBL10 directly regulates AKT1 activity and affects ion homeostasis in plant cells. 2013 The Authors. The Plant Journal 2013 Blackwell Publishing Ltd.
RonzierE, Corratgé-FaillieC, SanchezF, PradoK, BrièreC, LeonhardtN, ThibaudJB, XiongTC ( 2014). CPK13, a noncanonical Ca 2+-dependent protein kinase, specifically inhibits KAT2 and KAT1 shaker K + channels and reduces stomatal opening Plant Physiol 166, 314-326. [本文引用: 1]
RubioF, ArévaloL, CaballeroF, BotellaMA, RubioJS, García-SánchezF, MartínezV ( 2010). Systems involved in K + uptake from diluted solutions in pepper plants as revealed by the use of specific inhibitors J Plant Physiol 167, 1494-1499. DOI:10.1016/j.jplph.2010.05.022URLPMID:20691498 [本文引用: 1] Here, the contribution of the HAK1 transporter, the AKT1 channel and a putative AtCHX13 homolog to K + uptake in the high-affinity range of concentrations in pepper plants was examined. The limited development of molecular tools in pepper plants precluded a reverse genetics study in this species. By contrast, in the model plant Arabidopsis thaliana, these type of studies have shown that NH 4 + and Ba 2+ may be used as specific inhibitors of the two K + uptake systems to dissect their contribution in species in which, as in pepper, specific mutant lines are not available. By using these inhibitors together with Na + and Cs +, the relative contributions of CaHAK1, CaAKT1 and a putative AtCHX13 homolog to K + acquisition from diluted solutions under different regimens of K + supply were studied. The results showed that, in plants completely starved of K +, the gene encoding CaHAK1 was highly expressed and this system is a major contributor to K + uptake. However, K + concentrations as low as 50 M reduced CaHAK1 expression and the CaAKT1 channel came into play, participating together with CaHAK1 in K + absorption. The contribution of a putative AtCHX13 homolog seemed to be low under this low K + supply, but it cannot be ruled out that at higher K + concentrations this system participates in K + uptake. Studies of this type allow extension of the tools developed in model plants to understand nutrition in important crops.
SasakiT, MoriIC, FuruichiT, MunemasaS, ToyookaK, MatsuokaK, MurataY, YamamotoY ( 2010). Closing plant stomata requires a homolog of an aluminum-activated malate transporter Plant Cell Physiol 51, 354-365. DOI:10.1093/pcp/pcq016URLPMID:2835873 [本文引用: 1] Plant stomata limit both carbon dioxide uptake and water loss; hence, stomatal aperture is carefully set as the environment fluctuates. Aperture area is known to be regulated in part by ion transport, but few of the transporters have been characterized. Here we report thatAtALMT12(At4g17970), a homolog of the aluminum-activated malate transporter (ALMT) of wheat, is expressed in guard cells ofArabidopsis thaliana. Loss-of-function mutations inAtALMT12impair stomatal closure induced by ABA, calcium and darkness, but do not abolish either the rapidly activated or the slowly activated anion currents previously identified as being important for stomatal closure. Expressed inXenopusoocytes, AtALMT12 facilitates chloride and nitrate currents, but not those of organic solutes. Therefore, we conclude that AtALMT12 is a novel class of anion transporter involved in stomatal closure.
SasakiT, YamamotoY, EzakiB, KatsuharaM, AhnSJ, RyanPR, DelhaizeE, MatsumotoH ( 2004). A wheat gene encoding an aluminum-activated malate transporter Plant J 37, 645-653. DOI:10.1111/j.1365-313X.2003.01991.xURLPMID:14871306 [本文引用: 2] Summary The major constraint to plant growth in acid soils is the presence of toxic aluminum (Al) cations, which inhibit root elongation. The enhanced Al tolerance exhibited by some cultivars of wheat is associated with the Al-dependent efflux of malate from root apices. Malate forms a stable complex with Al that is harmless to plants and, therefore, this efflux of malate forms the basis of a hypothesis to explain Al tolerance in wheat. Here, we report on the cloning of a wheat gene, ALMT1 ( al uminum-activated m alate t ransporter), that co-segregates with Al tolerance in F 2 and F 3 populations derived from crosses between near-isogenic wheat lines that differ in Al tolerance. The ALMT1 gene encodes a membrane protein, which is constitutively expressed in the root apices of the Al-tolerant line at greater levels than in the near-isogenic but Al-sensitive line. Heterologous expression of ALMT1 in Xenopus oocytes, rice and cultured tobacco cells conferred an Al-activated malate efflux. Additionally, ALMT1 increased the tolerance of tobacco cells to Al treatment. These findings demonstrate that ALMT1 encodes an Al-activated malate transporter that is capable of conferring Al tolerance to plant cells.
SatoA, GambaleF, DreyerI, UozumiN ( 2010). Modulation of the Arabidopsis KAT1 channel by an activator of protein kinase C in Xenopus laevis oocytes .FEBS J 277, 2318-2328. [本文引用: 1]
SatoA, SatoY, FukaoY, FujiwaraM, UmezawaT, Shin- ozakiK, HibiT, TaniguchiM, MiyakeH, GotoDB, UozumiN ( 2009). Threonine at position 306 of the KAT1 potassium channel is essential for channel activity and is a target site for ABA-activated SnRK2/OST1/SnRK2.6 pro- tein kinase Biochem J 424, 439-448. DOI:10.1042/BJ20091221URLPMID:19785574 [本文引用: 1] The Arabidopsis thaliana K+ channel KAT1 has been suggested to have a key role in mediating the aperture of stomata pores on the surface of plant leaves. Although the activity of KAT1 is thought to be regulated by phosphorylation, the endogenous pathway and the primary target site for thismodification remained unknown. In the present study, we have demonstrated that the C-terminal region of KAT1 acts as a phosphorylation target for the Arabidopsis calcium-independent ABA (abscisic acid)-activated protein kinase SnRK2.6 (Snf1-related protein kinase 2.6). This was confirmed by LC-MS/MS (liquid chromatography tandem MS) analysis, which showed that Thr306 and Thr308 of KAT1 were modified by phosphorylation. The role of these specific residues was examined by single point mutations and measurement of KAT1 channel activities in Xenopus oocyte and yeast systems. Modification of Thr308 hadminimal effect onKAT1 activity. On the other hand, modification of Thr306 reduced the K+ transport uptake activity of KAT1 in both systems, indicating that Thr306 is responsible for the functional regulation of KAT1. These results suggest that negative regulation of KAT1 activity, required for stomatal closure, probably occurs by phosphorylation of KAT1 Thr306 by the stress-activated endogenous SnRK2.6 protein kinase.
SchroederJI ( 1988). K + transport properties of K + channels in the plasma membrane of Vicia faba guard cells .J Gen Physiol 92, 667-683. DOI:10.1085/jgp.92.5.667URLPMID:3235976 [本文引用: 1] Electrical properties of the plasma membrane of guard cell protoplasts isolated from stomates of Vicia faba leaves were studied by application of the whole-cell configuration of the patch-clamp technique. The two types of K+ currents that have recently been identified in guard cells may allow efflux of K+ during stomatal closing, and uptake of K+ during stomatal opening (Schroeder et al., 1987). A detailed characterization of ion transport properties of the inward-rectifying (IK+,in) and the outward-rectifying (IK+,out) K+ conductance is presented here. The permeability ratios of IK+,in and IK+,out currents for K+ over monovalent alkali metal ions were determined. The resulting permeability sequences (PK+ greater than PRb+ greater than PNa+ greater than PLi+ much greater than PCs+) corresponded closely to the ion specificity of guard cell movements in V. faba. Neither K+ currents exhibited significant inactivation when K+ channels were activated for prolonged periods (greater than 10 min). The absence of inactivation may permit long durations of K+ fluxes, which occur during guard cell movements. Activation potentials of inward K+ currents were not shifted when external K+ concentrations were changed. This differs strongly from the behavior of inward-rectifying K+ channels in animal tissue. Blue light and fusicoccin induce hyperpolarization by stimulation of an electrogenic pump. From slow-whole-cell recordings it was concluded that electrogenic pumps require cytoplasmic substrates for full activation and that the magnitude of the pump current is sufficient to drive K+ uptake through IK+,in channels. First, direct evidence was gained for the hypothesis that IK+,in channels are a molecular pathway for K+ accumulation by the finding that IK+,in was blocked by Al3+ ions, which are known to inhibit stomatal opening but not closing. The results presented in this study strongly support a prominent role for IK+,in and IK+,out channels in K+ transport across the plasma membrane of guard cells.
SchroederJI ( 1989). Quantitative analysis of outward rec- tifying K + channel currents in guard cell protoplasts from Vicia faba .J Membr Biol 107, 229-235. DOI:10.1007/BF01871938URLPMID:2716046 Abstract A quantitative analysis of the time and voltage dependence of outward-rectifying K+ currents (IK+, out) in guard cells from Vicia faba is described using the whole-cell patch-clamp technique. After step depolarizations from -75 mV to potentials positive to -40 mV, time-dependent outward currents were produced, which have recently been identified as K+ channel currents. This K+ current was characterized according to its time dependence and its steady-state activation. IK+, out could be described in terms of a Hodgkin-Huxley type conductance. Activation of the current in time was sigmoid and was well fitted by raising the activation variable to the second power. Deactivating tail currents were single exponentials, which suggests that only one conductance underlies this slow outward K+ current. Rates of channel closing were strongly dependent on the membrane potential, while rates of channel opening showed only limited voltage dependence leading to a highly asymmetric voltage dependence for channel closing and opening. The presented analysis provides a quantitative basis for the understanding of IK+, out channel gating and IK+, out channel functions in plant cells.
SchroederJI, HagiwaraS ( 1989). Cytosolic calcium regula- tes ion channels in the plasma membrane of Vicia faba guard cells .Nature 338, 427-430. DOI:10.1038/338427a0URL [本文引用: 2] THE molecular mechanisms by which Ca 2+ controls a variety of ion transport-associated cellular functions in higher plant cells 1 , including movements of stomatal pores, remain unknown. Stomatal pores regulate the gas exchange in leaves. Openings of stomatal pores are mediated by an increase in the intracellular potassium and anion content of guard cells 2,3 . Voltage-dependent K + channels 4–7 and hyperpolarizing H + pumps 8,9 have been identified as mechanisms controlling stomatal movements. But depolarizing mechanisms required for K + efflux during stomatal closing remain unknown. Indirect evidence suggests that Ca 2+ triggers stomatal closing and inhibits stomatal opening 10–12 . Using patch-clamp techniques 13 , we show here that elevated (micromolar) concentrations of cytosolic Ca 2+ in guard cells block inward rectifying K + channels. Furthermore, elevation of cytosolic Ca 2+ leads to the activation of a voltage-dependent depolarizing conductance with a permeability to anions. The Ca 2+ -induced modulation of ion channels reported here could provide a molecular basis for Ca 2+ -dependent regulation of stomatal movements in leaves.
SchroederJI, RaschkeK, NeherE ( 1987). Voltage dependence of K + channels in guard-cell protoplasts Proc Natl Acad Sci USA 84, 4108-4112. [本文引用: 1]
SchuurinkRC, ShartzerSF, FathA, JonesRL ( 1998). Characterization of a calmodulin-binding transporter from the plasma membrane of barley aleurone Proc Natl Acad Sci USA 95, 1944-1949. DOI:10.1073/pnas.95.4.1944URLPMID:9465122 [本文引用: 1] We have used Arabidopsis calmodulin (CaM) covalently coupled to horseradish peroxidase to screen a barley aleurone cDNA expression library for CaM binding proteins. The deduced amino acid sequence of one cDNA obtained by this screen was shown to be a unique protein of 702 amino acids with CaM and cyclic nucleotide binding domains at the carboxyl terminus and high similarity to olfactory and K+channels. This cDNA was designated HvCBT1 (Hordeum vulgare CaM binding transporter). Hydropathy plots of HvCBT1 showed the presence of six putative transmembrane domains, but sequence alignment indicated a pore domain that was unlike the consensus domains in K+and olfactory channels. Expression of a subclone of amino acids 482-702 in Escherichia coli generated a peptide that bound CaM. When a fusion protein of HvCBT1 and green fluorescent protein was expressed in barley aleurone protoplasts, fluorescence accumulated in the plasma membrane. Expression of HvCBT1 in the K+transport deficient Saccharomyces cerevisiae mutant CY162 showed no rescue of the mutant phenotype. However, growth of CY162 expressing HvCBT1 with its pore mutated to GYGD, the consensus sequence of K+channels, was compromised. We interpret these data as indicating that HvCBT1 acts to interfere with ion transport.
SharmaT, DreyerI, KochianL, Pi?erosMA ( 2016). The ALMT family of organic acid transporters in plants and their involvement in detoxification and nutrient security Front Plant Sci 7, 1488. DOI:10.3389/fpls.2016.01488URLPMID:5047901 [本文引用: 5] About a decade ago, members of a new protein family of anion channels were discovered on the basis of their ability to confer on plants the tolerance toward toxic aluminum ions in the soil. The efflux of Al3+-chelating malate anions through these channels is stimulated by external Al3+ions. This feature of a few proteins determined the name of the entire protein family asAluminum-activatedMalateTransporters (ALMT). Meanwhile, after several years of research, it is known that the physiological roles of ALMTs go far beyond Al-detoxification. In this review article we summarize the current knowledge on this transporter family and assess their involvement in diverse physiological processes.
SharmaT, DreyerI, RiedelsbergerJ ( 2013). The role of K + channels in uptake and redistribution of potassium in the model plant Arabidopsis thaliana .Front Plant Sci 4, 224. DOI:10.3389/fpls.2013.00224URLPMID:3694395 [本文引用: 3] Potassium (K+) is inevitable for plant growth and development. It plays a crucial role in the regulation of enzyme activities, in adjusting the electrical membrane potential and the cellular turgor, in regulating cellular homeostasis and in the stabilization of protein synthesis. Uptake of K+ from the soil and its transport to growing organs is essential for a healthy plant development. Uptake and allocation of K+ are performed by K+ channels and transporters belonging to different protein families. In this review we summarize the knowledge on the versatile physiological roles of plant K+ channels and their behavior under stress conditions in the model plant Arabidopsis thaliana.
SivaguruM, PikeS, GassmannW, BaskinTI ( 2003). Aluminum rapidly depolymerizes cortical microtubules and depolarizes the plasma membrane: evidence that these responses are mediated by a glutamate receptor Plant Cell Physiol 44, 667-675. DOI:10.1093/pcp/pcg094URLPMID:12881494 [本文引用: 1] Abstract Efforts to understand how plants respond to aluminum have focused on describing the symptoms of toxicity and elucidating mechanisms of tolerance; however, little is known about the signal transduction steps that initiate the plant's response. Here, we image cortical microtubules and quantify plasma-membrane potential in living, root cells of intact Arabidopsis seedlings. We show that aluminum depolymerizes microtubules and depolarizes the membrane, and that these responses are prevented by calcium channel blockade. Calcium influx might involve glutamate receptors, which in animals are ligand-gated cation channels and are present in the Arabidopsis genome. We show that glutamate depolymerizes microtubules and depolarizes the plasma membrane. These responses, and also the inhibition of root elongation, occur within the first few min of treatment, but are evoked more rapidly by glutamate than by aluminum. Microtubule depolymerization and membrane depolarization, induced by either glutamate or aluminum, are blocked by a specific antagonist of ionotropic glutamate receptors, 2-amino-5-phosphonopentanoate; whereas an antagonist of an aluminum-gated anion channel blocks the two responses to aluminum but not to glutamate. For growth, microtubule integrity, and membrane potential, responses to combined glutamate and aluminum were not greater than to glutamate alone. We propose that signaling in response to aluminum is initiated by efflux of a glutamate-like ligand through an anion channel and the binding of this ligand to a glutamate receptor.
SobolevskyAI ( 2015). Structure and gating of tetrameric glutamate receptors J Physiol 593, 29-38. DOI:10.1113/jphysiol.2013.264911URLPMID:25556785 [本文引用: 1] Abstract Ionotropic glutamate receptors (iGluRs) are ligand-gated ion channels that open their ion-conducting pores in response to the binding of agonist glutamate. In recent years, significant progress has been achieved in studies of iGluRs by determining numerous structures of isolated water-soluble ligand-binding and amino-terminal domains, as well as solving the first crystal structure of the full-length AMPA receptor in the closed, antagonist-bound state. These structural data combined with electrophysiological and fluorescence recordings, biochemical experiments, mutagenesis and molecular dynamics simulations have greatly improved our understanding of iGluR assembly, activation and desensitization processes. This article reviews the recent structural and functional advances in the iGluR field and summarizes them in a simplified model of full-length iGluR gating.
SukweenadhiJ, KimYJ, ChoiES, KohSC, LeeSW, KimYJ, YangDC ( 2015). Paenibacillus yonginensis DCY84 T induces changes in Arabidopsis thaliana gene expression against aluminum, drought, and salt stress .Microbiol Res 172, 7-15. [本文引用: 1]
SunSJ, QiGN, GaoQF, WangHQ, YaoFY, HussainJ, WangYF ( 2016). Protein kinase OsSAPK8 functions as an essential activator of S-type anion channel OsSLAC1, which is nitrate-selective in rice Planta 243, 489-500. DOI:10.1007/s00425-015-2418-xURLPMID:26481009 [本文引用: 1] Main conclusion OsSAPK8 is an essential activator of OsSLAC1 by phosphorylation, and OsSLAC1 is a nitrate-selective anion channel. S-type anion channel AtSLAC1 and protein kinase AtOST1 have been...
SutterJU, CampanoniP, TyrrellM, BlattMR ( 2006). Selective mobility and sensitivity to SNAREs is exhibited by the Arabidopsis KAT1 K + channel at the plasma membrane Plant Cell 18, 935-954. DOI:10.1105/tpc.105.038950URLPMID:16531497 [本文引用: 1] Recent findings indicate that proteins in the SNARE superfamily are essential for cell signaling, in addition to facilitating vesicle traffic in plant cell homeostasis, growth, and development. We previously identified SNAREs SYP121/Syr1 from tobacco (Nicotiana tabacum) and the Arabidopsis thaliana homolog SYP121 associated with abscisic acid and drought stress. Disrupting tobacco SYP121 function by expressing a dominant-negative Sp2 fragment had severe effects on growth, development, and traffic to the plasma membrane, and it blocked K62 and Cl63 channel responses to abscisic acid in guard cells. These observations raise questions about SNARE control in exocytosis and endocytosis of ion channel proteins and their organization within the plane of the membrane. We have used a dual, in vivo tagging strategy with a photoactivatable green fluorescent protein and externally exposed hemagglutinin epitopes to monitor the distribution and trafficking dynamics of the KAT1 K62 channel transiently expressed in tobacco leaves. KAT1 is localized to the plasma membrane within positionally stable microdomains of 650.5 μm in diameter; delivery of the K62 channel, but not of the PMA2 H62-ATPase, to the plasma membrane is suppressed by Sp2 fragments of tobacco and Arabidopsis SYP121, and Sp2 expression leads to profound changes in KAT1 distribution and mobility within the plane of the plasma membrane. These results offer direct evidence for SNARE-mediated traffic of the K62 channel and a role in its distribution within subdomains of the plasma membrane, and they implicate a role for SNAREs in positional anchoring of the K62 channel protein.
SutterJU, SiebenC, HartelA, EisenachC, ThielG, BlattMR ( 2007). Abscisic acid triggers the endocytosis of the Arabidopsis KAT1 K + channel and its recycling to the plasma membrane Curr Biol 17, 1396-1402. DOI:10.1016/j.cub.2007.07.020URLPMID:17683934 [本文引用: 1] Membrane vesicle traffic to and from the plasma membrane is essential for cellular homeostasis in all eukaryotes . In plants, constitutive traffic to and from the plasma membrane has been implicated in maintaining the population of integral plasma-membrane proteins and its adjustment to a variety of hormonal and environmental stimuli . However, direct evidence for evoked and selective traffic has been lacking. Here, we report that the hormone abscisic acid (ABA), which controls ion transport and transpiration in plants under water stress , triggers the selective endocytosis of the KAT1 K channel protein in epidermal and guard cells. Endocytosis of the K channel from the plasma membrane initiates in concert with changes in K channel activities evoked by ABA and leads to sequestration of the K channel within an endosomal membrane pool that recycles back to the plasma membrane over a period of hours. Selective K channel endocytosis, sequestration, and recycling demonstrates a tight and dynamic control of the population of K channels at the plasma membrane as part of a key plant signaling and response mechanism, and the observations point to a role for channel traffic in adaptive changes in the capacity for osmotic solute flux of stomatal guard cells .
SzyrokiA, IvashikinaN, DietrichP, RoelfsemaMRG, AcheP, ReintanzB, DeekenR, GoddeM, FelleH, SteinmeyerR, PalmeK, HedrichR ( 2001). KAT1 is not essential for stomatal opening Proc Natl Acad Sci USA 98, 2917-2921. DOI:10.1073/pnas.051616698URLPMID:11226341 [本文引用: 2] It is generally accepted that K+ uptake into guard cells via inward-rectifying K+ channels is required for stomatal opening. To test whether the guard cell K+ channel KAT1 is essential for stomatal opening, a knockout mutant, KAT1::En-1, was isolated from an En-1 mutagenized Arabidopsis thaliana population. Stomatal action and K+ uptake, however, were not impaired in KAT1-deficient plants. Reverse transcription-PCR experiments with isolated guard cell protoplasts showed that in addition to KAT1, the K+ channels AKT1, AKT2/3, AtKC1, and KAT2 were expressed in this cell type. In impalement measurements, intact guard cells exhibited inward-rectifying K+ currents across the plasma membrane of both wild-type and KAT1::En-1 plants. This study demonstrates that multiple K+ channel transcripts exist in guard cells and that KAT1 is not essential for stomatal action.
TakanashiK, SasakiT, KanT, SaidaY, SugiyamaA, YamamotoY, YazakiK ( 2016). A dicarboxylate transporter, LjALMT4, mainly expressed in nodules of Lotus japonicus .Mol Plant Microbe Interact 29, 584-592. DOI:10.1094/MPMI-04-16-0071-RURLPMID:27183039 [本文引用: 1] Legume plants can establish symbiosis with soil bacteria called rhizobia to obtain nitrogen as a nutrient directly from atmospheric N2 via symbiotic nitrogen fixation. Legumes and rhizobia form nodules, symbiotic organs in which fixed-nitrogen and photosynthetic products are exchanged between rhizobia and plant cells. The photosynthetic products supplied to rhizobia are thought to be dicarboxylates but little is known about the movement of dicarboxylates in the nodules. In terms of dicarboxylate transporters, an aluminum-activated malate transporter (ALMT) family is a strong candidate responsible for the membrane transport of carboxylates in nodules. Among the seven ALMT genes in the Lotus japonicus genome, only one, LjALMT4, shows a high expression in the nodules. LjALMT4 showed transport activity in a Xenopus oocyte system, with LjALMT4 mediating the efflux of dicarboxylates including malate, succinate, and fumarate, but not tricarboxylates such as citrate. LjALMT4 also mediated the influx of several inorganic anions. Organ-specific gene expression analysis showed LjALMT4 mRNA mainly in the parenchyma cells of nodule vascular bundles. These results suggest that LjALMT4 may not be involved in the direct supply of dicarboxylates to rhizobia in infected cells but is responsible for supplying malate as well as several anions necessary for symbiotic nitrogen fixation, via nodule vasculatures.
Tunc-OzdemirM, RatoC, BrownE, RogersS, Moo- neyhamA, FrietschS, MyersCT, PoulsenLR, MalhóR, HarperJF ( 2013). Cyclic nucleotide gated channels 7 and 8 are essential for male reproductive fertility PLoS One 8, e55277. DOI:10.1371/journal.pone.0055277URLPMID:23424627 [本文引用: 1] TheArabidopsis thalianagenome contains 20 CNGCs, which are proposed to encodecyclicnucleotidegated, non-selective, Ca2+-permeable ionchannels. CNGC7 and CNGC8 are the two most similar with 74% protein sequence identity, and both genes are preferentially expressed in pollen. Two independent loss-of-functionT-DNAinsertions were identified for both genes and used to generate plant lines in which only one of the two alleles was segregating (e.g.,cngc7-1+/61/cngc8-261/61andcngc7-361/61/cngc8-1+/61). While normal pollen transmission was observed for single gene mutations, pollen harboring mutations in bothcngc7and8were found to be male sterile (transmission efficiency reduced by more than 3000-fold). Pollen grains harboringT-DNAdisruptions of bothcngc7and8displayed a high frequency of bursting when germinatedin vitro.The male sterile defect could be rescued through pollen expression of aCNGC7or8transgene including a CNGC7 with an N-terminal GFP-tag. However, rescue efficiencies were reduced 6510-fold when the CNGC7 or 8 included an F to W substitution (F589W and F624W, respectively) at the junction between the putative cyclic nucleotide binding-site and the calmodulin binding-site, identifying this junction as important for proper functioning of a plant CNGC. Using confocal microscopy, GFP-CNGC7 was found to preferentially localize to the plasma membrane at the flanks of the growing tip. Together these results indicate that CNGC7 and 8 are at least partially redundant and provide an essential function at the initiation of pollen tube tip growth.
Tunc-OzdemirM, TangC, IshkaMR, BrownE, GrovesNR, MyersCT, RatoC, PoulsenLR, McDowellS, MillerG, MittlerR, HarperJF ( 2013). A cyclic nucleotide-gated channel (CNGC16) in pollen is critical for stress tolerance in pollen reproductive development Plant Physiol 161, 1010-1020. DOI:10.1104/pp.112.206888URLPMID:23370720 [本文引用: 1] Cyclic nucleotide-gated channels (CNGCs) have been implicated in diverse aspects of plant growth and development, including responses to biotic and abiotic stress, as well as pollen tube growth and fertility. Here, genetic evidence identifies CNGC16 in Arabidopsis (Arabidopsis thaliana) as critical for pollen fertility under conditions of heat stress and drought. Two independent transfer DNA disruptions of cngc16 resulted in a greater than 10-fold stress-dependent reduction in pollen fitness and seed set. This phenotype was fully rescued through pollen expression of a CNGC16 transgene, indicating that cngc16-1 and 16-2 were both loss-of-function null alleles. The most stress-sensitive period for cngc16 pollen was during germination and the initiation of pollen tube tip growth. Pollen viability assays indicate that mutant pollen are also hypersensitive to external calcium chloride, a phenomenon analogous to calcium chloride hypersensitivities observed in other cngc mutants. A heat stress was found to increase concentrations of 3',5'-cyclic guanyl monophosphate in both pollen and leaves, as detected using an antibody-binding assay. A quantitative PCR analysis indicates that cngc16 mutant pollen have attenuated expression of several heat-stress response genes, including two heat shock transcription factor genes, HsfA2 and HsfB1. Together, these results provide evidence for a heat stress response pathway in pollen that connects a cyclic nucleotide signal, a Ca2+-permeable ion channel, and a signaling network that activates a downstream transcriptional heat shock response.
UrquhartW, ChinK, UngH, MoederW, YoshiokaK ( 2011). The cyclic nucleotide-gated channels AtCNGC11 and 12 are involved in multiple Ca 2+-dependent physiological responses and act in a synergistic manner J Exp Bot 62, 3671-3682. [本文引用: 1]
UrquhartW, GunawardenaAHLAN, MoederW, AliR, BerkowitzGA, YoshiokaK ( 2007). The chimeric cyclic nucleotide-gated ion channel ATCNGC11/12 constitutively induces programmed cell death in a Ca 2+ dependent manner Plant Mol Biol 65, 747-761. DOI:10.1007/s11103-007-9239-7URLPMID:17885810 [本文引用: 1] The hypersensitive response (HR) involves programmed cell death (PCD) in response to pathogen infection. To investigate the pathogen resistance signaling pathway, we previously identified the Arabidopsis mutant cpr22 , which displays constitutive activation of multiple defense responses including HR like cell death. The cpr22 mutation has been identified as a 3 kb deletion that fuses two cyclic nucleotide-gated ion channel (CNGC)-encoding genes, ATCNGC11 and ATCNGC12 , to generate a novel chimeric gene, ATCNGC11/12 . In this study, we conducted a characterization of cell death induced by transient expression of ATCNGC11/12 in Nicotiana benthamiana. Electron microscopic analysis of this cell death showed similar characteristics to PCD, such as plasma membrane shrinkage and vesicle formation. The hallmark of animal PCD, fragmentation of nuclear DNA, was also observed in ATCNGC11/12 -induced cell death. The development of cell death was significantly suppressed by caspase-1 inhibitors, suggesting the involvement of caspases in this process. Recently, vacuolar processing enzyme (VPE) was isolated as the first plant caspase-like protein, which is involved in HR development. In VPE -silenced plants development of cell death induced by ATCNGC11/12 was much slower and weaker compared to control plants, suggesting the involvement of VPE as a caspase in ATCNGC11/12 -induced cell death. Complementation analysis using a Ca 2+ uptake deficient yeast mutant demonstrated that the ATCNGC11/12 channel is permeable to Ca 2+ . Additionally, calcium channel blockers such as GdCl 3 inhibited ATCNGC11/12 -induced HR formation, whereas potassium channel blockers did not. Taken together, these results indicate that the cell death that develops in the cpr22 mutant is indeed PCD and that the chimeric channel, ATCNGC11/12, is at the point of, or up-stream of the calcium signal necessary for the development of HR.
VahisaluT, KollistH, WangYF, NishimuraN, ChanWY, ValerioG, Lamminm?kiA, BroschéM, MoldauH, DesikanR, SchroederJI, Kangasj?rviJ ( 2008). SLAC1 is required for plant guard cell S-type anion channel function in stomatal signaling Nature 452, 487-491. DOI:10.1038/nature06608URLPMID:18305484 [本文引用: 5] Stomatal pores, formed by two surrounding guard cells in the epidermis of plant leaves, allow influx of atmospheric carbon dioxide in exchange for transpirational water loss. Stomata also restrict the entry of ozone--an important air pollutant that has an increasingly negative impact on crop yields, and thus global carbon fixation and climate change. The aperture of stomatal pores is regulated by the transport of osmotically active ions and metabolites across guard cell membranes. Despite the vital role of guard cells in controlling plant water loss, ozone sensitivity and CO2 supply, the genes encoding some of the main regulators of stomatal movements remain unknown. It has been proposed that guard cell anion channels function as important regulators of stomatal closure and are essential in mediating stomatal responses to physiological and stress stimuli. However, the genes encoding membrane proteins that mediate guard cell anion efflux have not yet been identified. Here we report the mapping and characterization of an ozone-sensitive Arabidopsis thaliana mutant, slac1. We show that SLAC1 (SLOW ANION CHANNEL-ASSOCIATED 1) is preferentially expressed in guard cells and encodes a distant homologue of fungal and bacterial dicarboxylate/malic acid transport proteins. The plasma membrane protein SLAC1 is essential for stomatal closure in response to CO2, abscisic acid, ozone, light/dark transitions, humidity change, calcium ions, hydrogen peroxide and nitric oxide. Mutations in SLAC1 impair slow (S-type) anion channel currents that are activated by cytosolic Ca2+ and abscisic acid, but do not affect rapid (R-type) anion channel currents or Ca2+ channel function. A low homology of SLAC1 to bacterial and fungal organic acid transport proteins, and the permeability of S-type anion channels to malate suggest a vital role for SLAC1 in the function of S-type anion channels.
VeleyKM, MaksaevG, FrickEM, JanuaryE, KloepperSC, HaswellES ( 2014). Arabidopsis MSL10 has a regulated cell death signaling activity that is separable from its mechanosensitive ion channel activity Plant Cell 26, 3115-3131. DOI:10.1105/tpc.114.128082URLPMID:25052715 [本文引用: 1] Members of the MscS superfamily of mechanosensitive ion channels function as osmotic safety valves, releasing osmolytes under increased membrane tension. MscS homologs exhibit diverse topology and domain structure, and it has been proposed that the more complex members of the family might have novel regulatory mechanisms or molecular functions. Here, we present a study of MscS-Like (MSL)10 from Arabidopsis thaliana that supports these ideas. High-level expression of MSL10-GFP in Arabidopsis induced small stature, hydrogen peroxide accumulation, ectopic cell death, and reactive oxygen species- and cell death-associated gene expression. Phosphomimetic mutations in the MSL10 N-terminal domain prevented these phenotypes. The phosphorylation state of MSL10 also regulated its ability to induce cell death when transiently expressed in Nicotiana benthamiana leaves but did not affect subcellular localization, assembly, or channel behavior. Finally, the N-terminal domain of MSL10 was sufficient to induce cell death in tobacco, independent of phosphorylation state. We conclude that the plant-specific N-terminal domain of MSL10 is capable of inducing cell death, this activity is regulated by phosphorylation, and MSL10 has two separable activities-one as an ion channel and one as an inducer of cell death. These findings further our understanding of the evolution and significance of mechanosensitive ion channels.
VeleyKM, MarshburnS, ClureCE, HaswellES ( 2012). Mechanosensitive channels protect plastids from hypoo- smotic stress during normal plant growth Curr Biol 22, 408-413. DOI:10.4161/psb.19991URLPMID:3472428 [本文引用: 1] In bacteria, MscS-type mechanosensitive channels serve to protect cells from lysis as they swell during extreme osmotic stress. We recently showed that two MscS homologs from Arabidopsis thaliana serve a similar purpose in the epidermal plastids of the leaf, indicating that the plant cell cytoplasm can present a dynamic osmotic challenge to the plastid. MscS homologs are predicted to be targeted to both plastids and mitochondrial envelopes and have been found in the genomes of intracellular pathogens. Here we discuss the implications of these observations, and propose that MS channels provide an essential mechanism for osmotic adaptation to both intracellular and the extracellular environments.
VigM, KinetJP ( 2007). The long and arduous road to CRAC Cell Calcium 42, 157-162. DOI:10.1016/j.ceca.2007.03.008URLPMID:17517435 [本文引用: 1] Store-operated calcium (SOC) entry is the major route of calcium influx in non-excitable cells, especially immune cells. The best characterized store-operated current, , is carried by calcium release activated calcium (CRAC) channels. The existence of the phenomenon of store-operated calcium influx was proposed almost two decades ago. However, in spite of rigorous research by many laboratories, the identity of the key molecules participating in the process has remained a mystery. In all these years, multiple different approaches have been adopted by countless researchers to identify the molecular players in this fundamental process. Along the way, many crucial discoveries have been made, some of which have been summarized here. The last couple of years have seen significant breakthroughs in the field dentification of STIM1 as the store Ca sensor and CRACM1 (Orai1) as the pore-forming subunit of the CRAC channel. The field is now actively engaged in deciphering the gating mechanism of CRAC channels. We summarize here the latest progress in this direction.
WangXP, ChenLM, LiuWX, ShenLK, WangFL, ZhouY, ZhangZD, WuWH, WangY ( 2016). AtKC1 and CIPK23 synergistically modulate AKT1-mediated low-potassium stress responses in Arabidopsis Plant Physiol 170, 2264-2277. DOI:10.1104/pp.15.01493URLPMID:26829980 [本文引用: 1] Abstract In Arabidopsis, the Shaker K+ channel AKT1 conducts K+ uptake in root cells, and its activity is regulated by CBL1/9-CIPK23 complexes, as well as by the AtKC1 channel subunit. CIPK23 and AtKC1 are both involved in the AKT1-mediated low K+ (LK) response; however, the relationship between them remains unclear. In this study, we screened suppressors of lks1 (cipk23) and isolated the sls1 (suppressor of lks1) mutant, which suppressed the leaf chlorosis phenotype of lks1 under LK conditions. Map-based cloning revealed a point mutation in AtKC1 of sls1 that led to an amino acid substitution (G322D) in the S6 region of AtKC1. The G322D substitution generated a gain-of-function mutation AtKC1D that enhanced K+ uptake capacity and LK tolerance in Arabidopsis. Structural prediction suggested that G322 is highly conserved in K+ channels and may function as the gating hinge of plant Shaker K+ channels. Electrophysiological analyses revealed that, compared with AtKC1 wild-type, AtKC1D showed enhanced inhibition of AKT1 activity and strongly reduced K+ leakage through AKT1 under LK conditions. In addition, phenotype analysis revealed distinct phenotypes of lks1 and atkc1 mutants in different LK assays, but the lks1 atkc1 double mutant always showed a LK-sensitive phenotype similar to that of akt1. This study revealed a link between CIPK-mediated activation and AtKC1-mediated modification in AKT1 regulation. CIPK23 and AtKC1 exhibit distinct effects; however, they act synergistically and balance K+ uptake/leakage to modulate AKT1-mediated LK responses in Arabidopsis. 2016 American Society of Plant Biologists. All rights reserved.
WangY, HeL, LiHD, XuJ, WuWH ( 2010). Potassium channel α-subunit AtKC1 negatively regulates AKT1-me- diated K + uptake in Arabidopsis roots under low-K + stress Cell Res 20, 826-837. DOI:10.1038/cr.2010.74URLPMID:20514083 [本文引用: 3] Potassium transporters play crucial roles in K(+) uptake and translocation in plants. However, so far little is known about the regulatory mechanism of potassium transporters. Here, we show that a Shaker-like potassium channel AtKC1, encoded by the AtLKT1 gene cloned from the Arabidopsis thaliana low-K(+) (LK)-tolerant mutant Atlkt1, significantly regulates AKT1-mediated K(+) uptake under LK conditions. Under LK conditions, the Atkc1 mutants maintained their root growth, whereas wild-type plants stopped their root growth. Lesion of AtKC1 significantly enhanced the tolerance of the Atkc1 mutants to LK stress and markedly increased K(+) uptake and K(+) accumulation in the Atkc1-mutant roots under LK conditions. Electrophysiological results showed that AtKC1 inhibited the AKT1-mediated inward K(+) currents and negatively shifted the voltage dependence of AKT1 channels. These results demonstrate that the 'silent' K(+) channel alpha-subunit AtKC1 negatively regulates the AKT1-mediated K(+) uptake in Arabidopsis roots and consequently alters the ratio of root-to-shoot under LK stress conditions.
WangY, KangY, MaCL, MiaoRY, WuCL, LongY, GeT, WuZN, HouXY, ZhangJX, QiZ ( 2017). CNGC2 is a Ca 2+ influx channel that prevents accumulation of apoplastic Ca 2+ in the leaf Plant Physiol 173, 1342-1354. DOI:10.1104/pp.16.01222URLPMID:27999084 [本文引用: 1] Abstract Ca 2+ is absorbed by roots and transported upward through the xylem to the apoplastic space of the leaf, after which it is deposited into the leaf cell. In Arabidopsis (Arabidopsis thaliana), the tonoplast-localized Ca 2+ /H + transporters CATION EXCHANGER1 (CAX1) and CAX3 sequester Ca 2+ from the cytosol into the vacuole, but it is not known what transporter mediates the initial Ca 2+ influx from the apoplast to the cytosol. Here, we report that Arabidopsis CYCLIC NUCLEOTIDE-GATED CHANNEL2 (CNGC2) encodes a protein with Ca 2+ influx channel activity and is expressed in the leaf areas surrounding the free endings of minor veins, which is the primary site for Ca 2+ unloading from the vasculature and influx into leaf cells. Under hydroponic growth conditions, with 0.1 mm Ca 2+ , both Arabidopsis cngc2 and cax1cax3 loss-of-function mutants grew normally. Increasing the Ca 2+ concentration to 10 mm induced H 2 O 2 accumulation, cell death, and leaf senescence and partially suppressed the hypersensitive response to avirulent pathogens in the mutants but not in the wild type. In vivo apoplastic Ca 2+ overaccumulation was found in the leaves of cngc2 and cax1cax3 but not the wild type under the 10 mm Ca 2+ condition, as monitored by Oregon Green BAPTA 488 5N, a low-affinity and membrane-impermeable Ca 2+ probe. Our results indicate that CNGC2 likely has no direct roles in leaf development or the hypersensitive response but, instead, that CNGC2 could mediate Ca 2+ influx into leaf cells. Finally, the in vivo extracellular Ca 2+ imaging method developed in this study provides a new tool for investigating Ca 2+ dynamics in plant cells. 2017 American Society of Plant Biologists. All Rights Reserved.
WangY, WuWH ( 2013). Potassium transport and signaling in higher plants Annu Rev Plant Biol 64, 451-476. DOI:10.1146/annurev-arplant-050312-120153URLPMID:23330792 [本文引用: 1] Abstract As one of the most important mineral nutrient elements, potassium (K(+)) participates in many plant physiological processes and determines the yield and quality of crop production. In this review, we summarize K(+) signaling processes and K(+) transport regulation in higher plants, especially in plant responses to K(+)-deficiency stress. Plants perceive external K(+) fluctuations and generate the initial K(+) signal in root cells. This signal is transduced into the cytoplasm and encoded as Ca(2+) and reactive oxygen species signaling. K(+)-deficiency-induced signals are subsequently decoded by cytoplasmic sensors, which regulate the downstream transcriptional and posttranslational responses. Eventually, plants produce a series of adaptive events in both physiological and morphological alterations that help them survive K(+) deficiency.
WardJM, M?serP, SchroederJI ( 2009). Plant ion channels: gene families, physiology, and functional genomics ana- lyses Annu Rev Physiol 71, 59-82. DOI:10.1146/annurev.physiol.010908.163204URLPMID:18842100 [本文引用: 1] Distinct potassium, anion, and calcium channels in the plasma membrane and vacuolar membrane of plant cells have been identified and characterized by patch clamping. Primarily owing to advances in Arabidopsis genetics and genomics, and yeast functional complementation, many of the corresponding genes have been identified. Recent advances in our understanding of ion channel genes that mediate signal transduction and ion transport are discussed here. Some plant ion channels, for example, ALMT and SLAC anion channel subunits, are unique. The majority of plant ion channel families exhibit homology to animal genes; such families include both hyperpolarization- and depolarization-activated Shaker-type potassium channels, CLC chloride transporters/channels, cyclic nucleotide-gated channels, and ionotropic glutamate receptor homologs. These plant ion channels offer unique opportunities to analyze the structural mechanisms and functions of ion channels. Here we review gene families of selected plant ion channel classes and discuss unique structure-function aspects and their physiological roles in plant cell signaling and transport.
WudickMM, PortesMT, MichardE, Rosas-SantiagoP, LizzioMA, NunesCO, CamposC, Sonta Cruz DamineliD, CarvalhoJC, LimaPT, PantojaO, FeijoJA ( 2018). CO- RNICHON sorting and regulation of GLR channels underlie pollen tube Ca 2+ homeostasis Science 360, 533-536. DOI:10.1126/science.aar6464URL [本文引用: 1] Compared to animals, evolution of plant calcium (Ca2+) physiology has led to a loss of proteins for influx and small ligand–operated control of cytosolic Ca2+, leaving many Ca2+ mechanisms unaccounted for. Here, we show a mechanism for sorting and activation of glutamate receptor–like channels (GLRs) by CORNICHON HOMOLOG (CNIH) proteins. Single mutants of pollen-expressed Arabidopsis thaliana GLRs (AtGLRs) showed growth and Ca2+ flux phenotypes expected for plasma membrane Ca2+ channels. However, higher-order mutants of AtGLR3.3 revealed phenotypes contradicting this assumption. These discrepancies could be explained by subcellular AtGLR localization, and we explored the implication of AtCNIHs in this sorting. We found that AtGLRs interact with AtCNIH pairs, yielding specific intracellular localizations. AtCNIHs further trigger AtGLR activity in mammalian cells without any ligand. These results reveal a regulatory mechanism underlying Ca2+ homeostasis by sorting and activation of AtGLRs by AtCNIHs.
XiclunaJ, LacombeB, DreyerI, AlconC, JeangueninL, SentenacH, ThibaudJB, ChérelI ( 2007). Increased functional diversity of plant K + channels by preferential heteromerization of the shaker-like subunits AKT2 and KAT2 J Biol Chem 282, 486-494. DOI:10.1074/jbc.M607607200URLPMID:17085433 [本文引用: 2] Abstract Assembly of plant Shaker subunits as heterotetramers, increasing channel functional diversity, has been reported. Here we focus on a new interaction, between AKT2 and KAT2 subunits. The assembly as AKT2/KAT2 heterotetramers is demonstrated by (i) a strong signal in two-hybrid tests with intracytoplasmic C-terminal regions, (ii) the effect of KAT2 on AKT2 subunit targeting in tobacco cells, (iii) the complete inhibition of AKT2 currents by co-expression with a dominant-negative KAT2 subunit in Xenopus oocytes, and reciprocally, and (iv) the appearance, upon co-expression of wild-type AKT2 and KAT2 subunits, of new channel functional properties that cannot be explained by the co-existence of two kinds of homotetrameric channels. In particular, the instantaneous current, characteristic of AKT2, displayed new functional features when compared with those of AKT2 homotetramers: activation by external acidification (instead of inhibition) and weak inhibition by calcium. Single channel current measurements in oocytes co-expressing AKT2 and KAT2 revealed a strong preference for incorporation of subunits into heteromultimers and a diversity of individual channels. In planta, these new channels, which may undergo specific regulations, are likely to be formed in guard cells and in the phloem, where they could participate in the control of membrane potential and potassium fluxes.
XuJ, LiHD, ChenLQ, WangY, LiuLL, HeL, WuWH ( 2006). A protein kinase, interacting with two calcineurin B-like proteins, regulates K + transporter AKT1 in Arabi- dopsis Cell 125, 1347-1360. [本文引用: 1]
XuJ, TianXL, Egrinya EnejiA, LiZH ( 2014). Functional characterization of GhAKT1, a novel Shaker-like K + chan- nel gene involved in K + uptake from cotton ( Gossypium hirsutum) .Gene 545, 61-71. DOI:10.1016/j.gene.2014.05.006URLPMID:24802116 [本文引用: 1] 61We characterized GhAKT1 gene from cotton.61It encodes a plasma membrane-localized protein, which can mediateK+ uptake in roots from 100 08M K+.61Overexpression of GhAKT1 in Arabidopsis resulted in an enhanced tolerance to low K+ stress.
XuMY, GruberBD, DelhaizeE, WhiteRG, JamesRA, YouJF, YangZM, RyanPR ( 2015). The barley anion channel, HvALMT1, has multiple roles in guard cell physiology and grain metabolism Physiol Plant 153, 183-193. DOI:10.1111/ppl.12234URLPMID:24853664 [本文引用: 1] The () gene HvALMT1 encodes an channel in guard cells and in certain root tissues indicating that it may perform multiple roles. The protein localizes to the and facilitates malate efflux from cells when constitutively expressed in and oocytes. This study investigated the function of HvALMT1 further by identifying its tissue-specific expression and by generating and characterizing lines with reduced HvALMT1 expression. We show that transgenic with 18-30% of wild-type HvALMT1 expression had impaired guard cell function. They maintained higher stomatal conductance in low light intensity and lost water more rapidly from excised leaves than the null segregant control . Tissue-specific expression of HvALMT1 was investigated in developing grain and during germination using transgenic lines expressing the () with the HvALMT1 promoter. We found that HvALMT1 is expressed in the nucellar projection, the aleurone layer and the scutellum of developing grain. Malate release measured from isolated aleurone layers prepared from imbibed grain was significantly lower in the compared with control . These data provide molecular and physiological evidence that HvALMT1 functions in guard cells, in grain and during germination. We propose that HvALMT1 releases malate and perhaps other from guard cells to promote stomatal closure. The likely roles of HvALMT1 during seed and grain germination are also discussed.
YuanF, YangHM, XueY, KongDD, YeR, LiCJ, ZhangJY, TheprungsirikulL, ShriftT, KrichilskyB, JohnsonDM, SwiftGB, HeYK, SiedowJN, PeiZM ( 2014). OSCA1 mediates osmotic-stress-evoked Ca 2+ increases vital for osmosensing in Arabidopsis Nature 514, 367-371. DOI:10.1038/nature13593URLPMID:25162526 [本文引用: 8] Water is crucial to plant growth and development. Environmental water deficiency triggers an osmotic stress signalling cascade, which induces short-term cellular responses to reduce water loss and long-term responses to remodel the transcriptional network and physiological and developmental processes. Several signalling components that have been identified by extensive genetic screens for altered sensitivities to osmotic stress seem to function downstream of the perception of osmotic stress. It is known that hyperosmolality and various other stimuli trigger increases in cytosolic free calcium concentration ([Ca2+]i). Considering that in bacteria and animals osmosensing Ca2+ channels serve as osmosensors, hyperosmolality-induced [Ca2+]i increases have been widely speculated to be involved in osmosensing in plants. However, the molecular nature of corresponding Ca2+ channels remain unclear. Here we describe a hyperosmolality-gated calcium-permeable channel and its function in osmosensing in plants. Using calcium-imaging-based unbiased forward genetic screens we isolated Arabidopsis mutants that exhibit low hyperosmolality-induced [Ca2+]i increases. These mutants were rescreened for their cellular, physiological and developmental responses to osmotic stress, and those with clear combined phenotypes were selected for further physical mapping. One of the mutants, reduced hyperosmolality-induced [Ca2+]i increase 1 (osca1), displays impaired osmotic Ca2+ signalling in guard cells and root cells, and attenuated water transpiration regulation and root growth in response to osmotic stress. OSCA1 is identified as a previously unknown plasma membrane protein and forms hyperosmolality-gated calcium-permeable channels, revealing that OSCA1 may be an osmosensor. OSCA1 represents a channel responsible for [Ca2+]i increases induced by a stimulus in plants, opening up new avenues for studying Ca2+ machineries for other stimuli and providing potential molecular genetic targets for engineering drought-resistant crops.
ZagottaWN, SiegelbaumSA ( 1996). Structure and function of cyclic nucleotide-gated channels Annu Rev Neurosci 19, 235-263. DOI:10.1007/BFb0033672URLPMID:8833443 [本文引用: 1] Abstract Cyclic nucleotide-gated (CNG) channels play important roles in both visual (Yau & Baylor 1989) and olfactory (Zufall et al 1994) signal transduction. The cloning of the gene coding for a rod photoreceptor channel (Kaupp et al 1989) and the subsequent cloning of related genes from olfactory neurons (Dhallan et al 1990, Ludwig et al 1990, Goulding et al 1992) has sparked much progress over the past several years in elucidating the structural bases for the function of the CNG channels. One of the surprising features to emerge from these cloning studies was that the CNG channels are structurally homologous to the voltage-gated channels (Jan & Jan 1990) despite the fact that the CNG channels are gated by the binding of a ligand-cAMP or cGMP-and not by voltage. In this review we focus on recent studies of the relationship between the structure and function of the CNG channels. Given the homology between CNG channels and voltage-gated channels, such studies are likely to provide important general information about the structure and function of a wide variety of channel types.
ZhangA, RenHM, TanYQ, QiGN, YaoFY, WuGL, YangLW, HussainJ, SunSJ, WangYF ( 2016). S-type anion channels SLAC1 and SLAH3 function as essential negative regulators of Inward K + channels and stomatal opening in Arabidopsis Plant Cell 28, 949-965. DOI:10.1105/tpc.16.01050URLPMID:27002025 [本文引用: 2] Drought stress induces stomatal closure and inhibits stomatal opening simultaneously. However, the underlying molecular mechanism is still largely unknown. Here we show that S-type anion channels SLAC1 and SLAH3 mainly inhibit inward K+ (K+in) channel KAT1 by protein-protein interaction, and consequently prevent stomatal opening in Arabidopsis. Voltage-clamp results demonstrated that SLAC1 inhibited KAT1 dramatically, but did not inhibit KAT2. SLAH3 inhibited KAT1 to a weaker degree relative to SLAC1. Both the N terminus and the C terminuses of SLAC1 inhibited KAT1, but the inhibition by the N terminus was stronger. The C terminus was essential for the inhibition of KAT1 by SLAC1. Furthermore, drought stress strongly up-regulated the expression of SLAC1 and SLAH3 in Arabidopsis guard cells, and the over-expression of wild type and truncated SLAC1 dramatically impaired K+in currents of guard cells and light-induced stomatal opening. Additionally, the inhibition of KAT1 by SLAC1 and KC1 only partially overlapped, suggesting that SLAC1 and KC1 inhibited K+in channels using different molecular mechanisms. Taken together, we discovered a novel regulatory mechanism for stomatal movement, in which singling pathways for stomatal closure and opening are directly coupled together by protein-protein interaction between SLAC1/SLAH3 and KAT1 in Arabidopsis.
ZhangHC, YinWL, XiaXL ( 2010). Shaker-like potassium channels in Populus, regulated by the CBL-CIPK signal transduction pathway, increase tolerance to low-K + stress. Plant Cell Rep 29, 1007-1012. DOI:10.1007/s00299-010-0886-9URLPMID:20582419 [本文引用: 2] Abstract Shaker-like potassium channels in plants play an important role in potassium absorption and transport. Here, we characterized 11 genes encoding shaker-like channels from Populus trichocarpa. Furthermore, two homologs from this family were isolated from Populus euphratica and named PeKC1 and PeKC2. Subcellular localization analysis of them in Nicotiana benthamiana revealed that they are located in the cell membrane. Yeast two-hybrid assays showed that they not only interacted strongly with PeCIPK24, a homolog of AtCIPK23, but also interacted with several other CIPK members, including PeCIPK10 and PeCIPK17. To further analyze their function, we over-expressed PeKC1 or PeKC2 in akt1 mutant, the results show that the transgenic plant can recover the mutant phonotype sensitive to low-K(+) stress. This means PeKC1 or PeKC2 can complement the function of AKT1 in akt1 mutant, involved in the CBL1-CIPK23 signal transduction pathway and play an important role under low-K(+) stress.
ZhangW, FanLM, WuWH ( 2007). Osmo-sensitive and stretch-activated calcium-permeable channels in Vicia faba guard cells are regulated by actin dynamics .Plant Physiol 143, 1140-1151. DOI:10.1104/pp.106.091405URLPMID:17259289 [本文引用: 1] In responses to a number of environmental stimuli, changes of cytoplasmic$[{\rm{Ca}}^{2 + } ]_{{\rm{cyt}}} $in stomatal guard cells play important roles in regulation of stomatal movements. In this study, the osmo-sensitive and stretch-activated (SA) Ca0562 channels in the plasma membrane of Vicia faba guard cells are identified, and their regulation by osmotic changes and actin dynamics are characterized. The identified Ca0562 channels were activated under hypotonic conditions at both whole-cell and single-channel levels. The channels were also activated by a stretch force directly applied to the membrane patches. The channel-mediated inward currents observed under hypotonic conditions or in the presence of a stretch force were blocked by the Ca0562 channel inhibitor Gd0662. Disruption of actin filaments activated SA Ca0562 channels, whereas stabilization of actin filaments blocked the channel activation induced by stretch or hypotonic treatment, indicating that actin dynamics may mediate the stretch activation of these channels. In addition,$[{\rm{Ca}}^{2 + } ]_{{\rm{cyt}}} $cyt imaging demonstrated that both the hypotonic treatment and disruption of actin filaments induced significant Ca0562 elevation in guard cell protoplasts, which is consistent with our electrophysiological results. It is concluded that stomatal guard cells may utilize SA Ca0562 channels as osmo sensors, by which swelling of guard cells causes elevation of$[{\rm{Ca}}^{2 + } ]_{{\rm{cyt}}} $and consequently inhibits overswelling of guard cells. This SA Ca0562 channel-mediated negative feedback mechanism may coordinate with previously hypothesized positive feedback mechanisms and regulate stomatal movement in response to environmental changes.
ZhaoLN, ShenLK, ZhangWZ, ZhangW, WangY, WuWH ( 2013). Ca 2+-dependent protein kinase11 and 24 modulate the activity of the inward rectifying K + channels in Arabi- dopsis pollen tubes Plant Cell 25, 649-661. [本文引用: 1]
ZouY, ChintamananiS, HeP, FukushigeH, YuLP, ShaoMY, ZhuLH, HildebrandDF, TangXY, ZhouJM ( 2016). A gain-of-function mutation in Msl10 triggers cell death and wound-induced hyperaccumulation of jasmonic acid in Arabidopsis .J Integr Plant Biol 58, 600-609. DOI:10.1111/jipb.12427URLPMID:26356550 [本文引用: 1] 当为创伤的关键管理者导致了发信号的小径, Jasmonates (JA ) 很快在伤害和行为以后被导致。不管多么,什么并且怎么察觉创伤信号那扳机 JA 生合成仍然保持糟糕理解。为了识别部件,在 Arabidopsis 包含了创伤和 JA 发信号小径,我们与一个酶记者的反常表示为异种屏蔽了,它在一个乙烯反应因素的一个创伤应答的倡导者的控制下面(小块土地) 抄写因素基因, RAP2.6 (与 APetala 2.6 有关) 。rea1 (在射击顶更快速的 RAP2.6 ) 异种组成地在年轻叶子表示了 RAP2.6-LUC 记者基因。与包括更短的叶柄,顶端的优势的损失, anthocyanin 颜料的累积和 JA 反应基因的组成的表示的典型 JA 显型一起, rea1 响应伤害种也显示的房间死亡和 JA 的积累的高水平。rea1 异种的显型被一个 gain-of-function 变化在 mechanosensitive 离子隧道象 MscS 一样的 C 终点引起 10 (MSL10 ) 。MSL10 在血浆膜是局部性的并且在根尖端,射击顶和脉管的纸巾主要被表示。这些结果建议 MSL10 在调整 JA 的积极反馈合成涉及被触发创伤的早信号 transduction 小径并且可能。 植物谷氨酸受体的研究进展 0 2016
植物环核苷酸门控离子通道及其功能研究进展 0 2015
GORK, a delayed outward rectifier expressed in guard cells of Arabidopsis thaliana, is a K +-selective, K +-sensing ion channel. 1 2000
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
TRPC1, Orai1, and STIM1 in SOCE: friends in tight spaces 1 2017
... 植物体内存在大量的钙元素.一方面钙作为细胞壁等结构组分发挥作用; 另一方面, 离子形式的钙(Ca2+)可以作为信号分子参与各种生物学反应和信号传递过程, 后者则备受关注.胞内Ca2+信号以细胞质Ca2+浓度的升高和震荡形式出现.一般认为, 胞外Ca2+内流是细胞质Ca2+信号产生所需Ca2+的主要来源, 而质膜内向Ca2+通道是胞外Ca2+内流的主要渠道.同时, Ca2+通道的活性变化通过控制胞外Ca2+内流的节奏直接调控胞内Ca2+震荡, 发挥Ca2+信号上游调控因子的作用.鉴于此, 质膜Ca2+通道受到广泛关注, 并成为动植物各信号领域的重点研究对象.科学家首先在动物细胞中发现多个Ca2+通道家族, 主要包括受环核苷酸cAMP和cGMP激活的CNG (cyclic nucleotide-gated)家族、受谷氨酸门控的GLR (glutamate receptor)家族、TRPC (transient receptor potential channel)和受胞内Ca2+库倒空激活的CRACM1/Orai1 (Zagotta and Siegelbaum, 1996; Vig and Kinet, 2007; Sobolevsky, 2015; Ambudkar et al., 2017).通过序列比对, 多个动物细胞质膜Ca2+通道的同源基因家族先后在植物尤其是模式植物拟南芥基因组中被鉴定出来, 主要包括CNGC (cyclic nucleotide- gated channel)、GLR (glutamate receptor-like)、高渗敏感的OSCA (hyperosmolality-gated calcium- permeable channel)家族和低渗敏感的张力激活离子通道MCA (mechano-sensitive channel) (M?ser et al., 2001; Ward et al., 2009; Iida et al., 2014; Yuan et al., 2014). ...
Expression of the AKT1-type K + channel gene from Puccinellia tenuiflora, PutAKT1, enhances salt tolerance in Arabidopsis 1 2010
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
A natural mutation-led truncation in one of the two aluminum-activated malate transporter-like genes at the Ma locus is associated with low fruit acidity in apple 1 2012
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
Two spatially and temporally distinct Ca 2+ signals convey Arabidopsis thaliana responses to K + deficiency. 1 2017
... Shaker家族离子通道的活性受多种调控因子的影响.首先, 细胞内和细胞外K+浓度对K+in和K+out通道活性均有明显影响.这种影响可能是基于K+从高浓度向低浓度方向流动的基本化学特性.酸性环境可以显著抑制拟南芥花粉原生质体质膜K+in通道活性, 而碱性条件则具有一定的激活作用(Fan et al., 2001, 2003).这种作用可能通过影响K+in通道的某些氨基酸残基电离状态和离子通道蛋白结构变化来实现.其次, K+in四聚体的亚基组成形式本身就是一种通道活性调控方式.在非洲爪蟾卵母细胞中, 以不同的排列组合共表达2个不同的拟南芥Shaker家族K+in通道基因, 可以利用电生理技术记录到具有不同电生理特性的K+in通道电流信号(Xicluna et al., 2007; Lebaudy et al., 2010).而AtKC1作为抑制亚基, 通过重组进入K+in四聚体发挥对K+in通道的抑制作用, 是K+in通道的重要负调控因子.K+in四聚体组成形式的变化可能通过影响K+in通道蛋白离子通道孔的结构而影响其离子通道活性.SNARE蛋白SYP121不仅可以影响KAT1等K+in通道的亚细胞定位, 而且可使KAT1内化离开细胞质膜, 从而抑制全细胞K+in通道的电流大小(Sutter et al., 2006, 2007).进一步研究表明, SNARE还可以通过与AtKC1互作调控K+in通道的蛋白活性(Honsbein et al., 2009; Grefen et al., 2010; Jeanguenin et al., 2011).由此可知, SNARE作为K+in通道的上游调控因子可通过对K+in四聚体组成形式的影响发挥调控作用.蛋白激酶和磷酸酶参与调控多种蛋白的功能, 包括内向和外向质膜K+通道, 比如CPK (calcium-dependent protein kinase)家族的CPK33对GORK的激活作用和PP2CA对GORK的抑制作用(Lefoulon et al., 2016; Corratgé-Faillie et al., 2017).K+in通道可以被多种蛋白激酶磷酸化所抑制, 包括CPK家族和SnRK (Rnf1-related protein kinase)家族的多个蛋白激酶(Sato et al., 2009, 2010; Ronzier et al., 2014).此外, AKT1等多个质膜K+in通道的活性还受Ca2+信号及其与CBL-CIPK组成的信号链的正调控(Li et al., 2006; Cheong et al., 2007; Zhang et al., 2010; Ren et al., 2013; Behera et al., 2017).而PP2CA对AKT1的去磷酸化可以与Ca2+-CBL-CIPK信号链协同调控AKT1的活性(Lan et al., 2011).有趣的是, ABA甚至可以直接激活GORK的离子通道活性(Ooi et al., 2017).K+通道的调控因子众多, 这些调控因子并非各自单独发挥作用, 而是彼此协同发挥作用.AtKC1和Ca2+-CBL-CIPK23信号链协同调控AKT1就是一个典型的例子(Wang et al., 2016). ...
Reconstitution of abscisic acid activation of SLAC1 anion channel by CPK6 and OST1 kinases and branched ABI1 PP2C phosphatase action 2 2012
... SLAC1家族是较晚被发现的一个植物细胞质膜阴离子通道家族, 但植物进化分析表明, SLAC1基因从低等到高等植物均很保守(Lind et al., 2015).拟南芥SLAC1家族由5个成员组成, 包括SLAC1及其同源蛋白SLAH1、2、3和4.其中, SLAC1主要定位在气孔保卫细胞质膜上, 通过介导Cl-和NO3-的外流驱动气孔关闭, 是干旱胁迫及ABA、CO2、臭氧和活性氧(reactive oxygen species, ROS)等信号传递途径的关键组分(Negi et al., 2008; Vahisalu et al., 2008).SLAH3在气孔保卫细胞中分布, 与SLAC1一起通过介导NO3-和Cl-外流驱动气孔关闭(Geiger et al., 2011), 但SLAH3在气孔关闭运动中的作用, 相对于SLAC1而言是辅助性的.拟南芥4个SLAH主要分布在根、茎和叶脉等输导组织中(Negi et al., 2008).其中, SLAH1通过调控SLAH3的活性调控Cl-从根向茎的运输(Cubero-Font et al., 2016).在离子选择性和通透性方面, 拟南芥SLAC1和SLAH3均对Cl-和NO3-有明显的通透能力, 但对NO3-的通透能力明显大于Cl- (Geiger et al., 2009; Lee et al., 2009; Brandt et al., 2012).而SLAH2则为NO3-选择性的阴离子通道, 对Cl-的通透能力微弱(Maierhofer et al., 2014).最近的研究表明, 来自单子叶植物玉米和水稻的SLAC1, 相对于Cl-而言, 对NO3-的选择性明显大于拟南芥SLAC1 (Sun et al., 2016; Qi et al., 2018); 而鸭跖草SLAC1则为NO3-激活的Cl-通透性的阴离子通道(Müller et al., 2017).因此, 来自不同高等植物的SLAC1, 其离子选择性不完全一致, 即不同的高等植物可能采用不同的阴离子通道组合来驱动气孔关闭运动.需要注意的是, 尽管在几十年前就发现苹果酸在气孔保卫细胞内大量积累, 并曾被认为是气孔关闭运动中主要的外流阴离子物质.但实际上, 在气孔关闭运动中发挥核心作用的SLAC1和SLAH3均对苹果酸没有明显的通透能力, 真正通过介导苹果酸外流而参与驱动气孔关闭运动的是来自ALMT家族的ALMT12.因此, 气孔关闭运动应该是由SLAC1、SLAH3和ALMT12等多种质膜阴离子协同作用的结果.每个SLAC1亚基具有10个跨膜区, 其C端和N端均位于质膜的胞质一侧(Vahisalu et al., 2008) (图6A).3个SLAC1亚基互相结合, 以三聚体的形式形成质膜阴离子通道, 但每个亚基独立形成1个允许阴离子通过的孔(Chen et al., 2010) (图6B).由此可见, 包括多个SLAHs在内, SLAC1家族形成的阴离子通道可能均为三聚体, 而SLAH1对SLAH3活性的调控作用, 有可能通过与SLAH3组成异源三聚体而实现.SLAC1家族的离子通道亚基组成形式与CNGC和Shaker家族不同, 但它们在不同亚基组成影响离子通道活性和选择性方面又具有类似之处.SLAC1家族离子通道的激活主要依赖蛋白激酶的磷酸化, 而失活则依赖于磷酸酶的去磷酸化(Geiger et al., 2009, 2011; Lee et al., 2009; Brandt et al., 2012; Maierhofer et al., 2014; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
... ; Brandt et al., 2012; Maierhofer et al., 2014; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
Homologue structure of the SLAC1 anion channel for closing stomata in leaves 3 2010
... SLAC1家族是较晚被发现的一个植物细胞质膜阴离子通道家族, 但植物进化分析表明, SLAC1基因从低等到高等植物均很保守(Lind et al., 2015).拟南芥SLAC1家族由5个成员组成, 包括SLAC1及其同源蛋白SLAH1、2、3和4.其中, SLAC1主要定位在气孔保卫细胞质膜上, 通过介导Cl-和NO3-的外流驱动气孔关闭, 是干旱胁迫及ABA、CO2、臭氧和活性氧(reactive oxygen species, ROS)等信号传递途径的关键组分(Negi et al., 2008; Vahisalu et al., 2008).SLAH3在气孔保卫细胞中分布, 与SLAC1一起通过介导NO3-和Cl-外流驱动气孔关闭(Geiger et al., 2011), 但SLAH3在气孔关闭运动中的作用, 相对于SLAC1而言是辅助性的.拟南芥4个SLAH主要分布在根、茎和叶脉等输导组织中(Negi et al., 2008).其中, SLAH1通过调控SLAH3的活性调控Cl-从根向茎的运输(Cubero-Font et al., 2016).在离子选择性和通透性方面, 拟南芥SLAC1和SLAH3均对Cl-和NO3-有明显的通透能力, 但对NO3-的通透能力明显大于Cl- (Geiger et al., 2009; Lee et al., 2009; Brandt et al., 2012).而SLAH2则为NO3-选择性的阴离子通道, 对Cl-的通透能力微弱(Maierhofer et al., 2014).最近的研究表明, 来自单子叶植物玉米和水稻的SLAC1, 相对于Cl-而言, 对NO3-的选择性明显大于拟南芥SLAC1 (Sun et al., 2016; Qi et al., 2018); 而鸭跖草SLAC1则为NO3-激活的Cl-通透性的阴离子通道(Müller et al., 2017).因此, 来自不同高等植物的SLAC1, 其离子选择性不完全一致, 即不同的高等植物可能采用不同的阴离子通道组合来驱动气孔关闭运动.需要注意的是, 尽管在几十年前就发现苹果酸在气孔保卫细胞内大量积累, 并曾被认为是气孔关闭运动中主要的外流阴离子物质.但实际上, 在气孔关闭运动中发挥核心作用的SLAC1和SLAH3均对苹果酸没有明显的通透能力, 真正通过介导苹果酸外流而参与驱动气孔关闭运动的是来自ALMT家族的ALMT12.因此, 气孔关闭运动应该是由SLAC1、SLAH3和ALMT12等多种质膜阴离子协同作用的结果.每个SLAC1亚基具有10个跨膜区, 其C端和N端均位于质膜的胞质一侧(Vahisalu et al., 2008) (图6A).3个SLAC1亚基互相结合, 以三聚体的形式形成质膜阴离子通道, 但每个亚基独立形成1个允许阴离子通过的孔(Chen et al., 2010) (图6B).由此可见, 包括多个SLAHs在内, SLAC1家族形成的阴离子通道可能均为三聚体, 而SLAH1对SLAH3活性的调控作用, 有可能通过与SLAH3组成异源三聚体而实现.SLAC1家族的离子通道亚基组成形式与CNGC和Shaker家族不同, 但它们在不同亚基组成影响离子通道活性和选择性方面又具有类似之处.SLAC1家族离子通道的激活主要依赖蛋白激酶的磷酸化, 而失活则依赖于磷酸酶的去磷酸化(Geiger et al., 2009, 2011; Lee et al., 2009; Brandt et al., 2012; Maierhofer et al., 2014; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
... ; Chen et al., 2010) The topological structures of a SLAC1 subunit (A) and the structure of a SLAC1 channel as a trimer (B) (<xref ref-type="bibr" rid="b139">Vahisalu et al., 2008</xref>; <xref ref-type="bibr" rid="b9">Chen et al., 2010</xref>)Figure 6 除了SLAC1家族, 植物细胞质膜上还包含张力激活的阴离子通道MSL (mechano-sensitive or stret- ch-activated channels) (Crosgrove and Hedrich, 1991).MSL为典型的张力激活的阴离子通道家族.MSL9和MSL10是最早得到电生理鉴定的MSL家族成员, 定位于细胞质膜, 其激活对质膜张力具有依赖性, 主要选择性介导阴离子的跨膜流动(Haswell et al., 2008; Maksaev and Haswell, 2012).随后有研究表明, 拟南芥MSL10参与植物机械损伤信号反应和茉莉酸的累积(Veley et al., 2014; Zou et al., 2016).此外, 有些拟南芥MSL成员定位在细胞器膜上, 调控细胞器的大小和形状(Veley et al., 2012).根据氨基酸残基序列预测, MSL蛋白存在多个跨膜区, 具备形成离子通道的基本结构特点.而质膜受到拉伸时对MSL的激活是通过机械拉力直接改变MSL蛋白的结构, 使离子通过的孔被拉开.但这基本上还停留在理论推测阶段, 缺乏有力的直接实验证据支持.总体来讲, 目前对MSL家族的生物学功能还了解较少, 其组成的离子通道蛋白结构、感知生物膜张力的分子机理和生物学功能等还有待深入研究. ...
... ; Chen et al., 2010) Figure 6 除了SLAC1家族, 植物细胞质膜上还包含张力激活的阴离子通道MSL (mechano-sensitive or stret- ch-activated channels) (Crosgrove and Hedrich, 1991).MSL为典型的张力激活的阴离子通道家族.MSL9和MSL10是最早得到电生理鉴定的MSL家族成员, 定位于细胞质膜, 其激活对质膜张力具有依赖性, 主要选择性介导阴离子的跨膜流动(Haswell et al., 2008; Maksaev and Haswell, 2012).随后有研究表明, 拟南芥MSL10参与植物机械损伤信号反应和茉莉酸的累积(Veley et al., 2014; Zou et al., 2016).此外, 有些拟南芥MSL成员定位在细胞器膜上, 调控细胞器的大小和形状(Veley et al., 2012).根据氨基酸残基序列预测, MSL蛋白存在多个跨膜区, 具备形成离子通道的基本结构特点.而质膜受到拉伸时对MSL的激活是通过机械拉力直接改变MSL蛋白的结构, 使离子通过的孔被拉开.但这基本上还停留在理论推测阶段, 缺乏有力的直接实验证据支持.总体来讲, 目前对MSL家族的生物学功能还了解较少, 其组成的离子通道蛋白结构、感知生物膜张力的分子机理和生物学功能等还有待深入研究. ...
Adaptation to acidic soil is achieved by inc- reased numbers of cis-acting elements regulating ALMT1 expression in Holcus lanatus 1 2013
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
Two calcineurin B-like calcium sensors, interacting with protein kinase CIPK23, regulate leaf transpiration and root potassium uptake in Arabidopsis 1 2007
... Shaker家族离子通道的活性受多种调控因子的影响.首先, 细胞内和细胞外K+浓度对K+in和K+out通道活性均有明显影响.这种影响可能是基于K+从高浓度向低浓度方向流动的基本化学特性.酸性环境可以显著抑制拟南芥花粉原生质体质膜K+in通道活性, 而碱性条件则具有一定的激活作用(Fan et al., 2001, 2003).这种作用可能通过影响K+in通道的某些氨基酸残基电离状态和离子通道蛋白结构变化来实现.其次, K+in四聚体的亚基组成形式本身就是一种通道活性调控方式.在非洲爪蟾卵母细胞中, 以不同的排列组合共表达2个不同的拟南芥Shaker家族K+in通道基因, 可以利用电生理技术记录到具有不同电生理特性的K+in通道电流信号(Xicluna et al., 2007; Lebaudy et al., 2010).而AtKC1作为抑制亚基, 通过重组进入K+in四聚体发挥对K+in通道的抑制作用, 是K+in通道的重要负调控因子.K+in四聚体组成形式的变化可能通过影响K+in通道蛋白离子通道孔的结构而影响其离子通道活性.SNARE蛋白SYP121不仅可以影响KAT1等K+in通道的亚细胞定位, 而且可使KAT1内化离开细胞质膜, 从而抑制全细胞K+in通道的电流大小(Sutter et al., 2006, 2007).进一步研究表明, SNARE还可以通过与AtKC1互作调控K+in通道的蛋白活性(Honsbein et al., 2009; Grefen et al., 2010; Jeanguenin et al., 2011).由此可知, SNARE作为K+in通道的上游调控因子可通过对K+in四聚体组成形式的影响发挥调控作用.蛋白激酶和磷酸酶参与调控多种蛋白的功能, 包括内向和外向质膜K+通道, 比如CPK (calcium-dependent protein kinase)家族的CPK33对GORK的激活作用和PP2CA对GORK的抑制作用(Lefoulon et al., 2016; Corratgé-Faillie et al., 2017).K+in通道可以被多种蛋白激酶磷酸化所抑制, 包括CPK家族和SnRK (Rnf1-related protein kinase)家族的多个蛋白激酶(Sato et al., 2009, 2010; Ronzier et al., 2014).此外, AKT1等多个质膜K+in通道的活性还受Ca2+信号及其与CBL-CIPK组成的信号链的正调控(Li et al., 2006; Cheong et al., 2007; Zhang et al., 2010; Ren et al., 2013; Behera et al., 2017).而PP2CA对AKT1的去磷酸化可以与Ca2+-CBL-CIPK信号链协同调控AKT1的活性(Lan et al., 2011).有趣的是, ABA甚至可以直接激活GORK的离子通道活性(Ooi et al., 2017).K+通道的调控因子众多, 这些调控因子并非各自单独发挥作用, 而是彼此协同发挥作用.AtKC1和Ca2+-CBL-CIPK23信号链协同调控AKT1就是一个典型的例子(Wang et al., 2016). ...
A quantitative hypermorphic CNGC allele confers ectopic calcium flux and impairs cellular development 1 2017
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
The Arabidopsis cyclic nucleotide-gated ion channels AtCNGC2 and AtCNGC4 work in the same signaling pathway to regulate pathogen defense and floral transition 1 2013
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
The mineral nutrition of higher plants 1 1980
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
The Arabidopsis guard cell outward potassium channel GORK is regulated by CPK33 1 2017
... Shaker家族离子通道的活性受多种调控因子的影响.首先, 细胞内和细胞外K+浓度对K+in和K+out通道活性均有明显影响.这种影响可能是基于K+从高浓度向低浓度方向流动的基本化学特性.酸性环境可以显著抑制拟南芥花粉原生质体质膜K+in通道活性, 而碱性条件则具有一定的激活作用(Fan et al., 2001, 2003).这种作用可能通过影响K+in通道的某些氨基酸残基电离状态和离子通道蛋白结构变化来实现.其次, K+in四聚体的亚基组成形式本身就是一种通道活性调控方式.在非洲爪蟾卵母细胞中, 以不同的排列组合共表达2个不同的拟南芥Shaker家族K+in通道基因, 可以利用电生理技术记录到具有不同电生理特性的K+in通道电流信号(Xicluna et al., 2007; Lebaudy et al., 2010).而AtKC1作为抑制亚基, 通过重组进入K+in四聚体发挥对K+in通道的抑制作用, 是K+in通道的重要负调控因子.K+in四聚体组成形式的变化可能通过影响K+in通道蛋白离子通道孔的结构而影响其离子通道活性.SNARE蛋白SYP121不仅可以影响KAT1等K+in通道的亚细胞定位, 而且可使KAT1内化离开细胞质膜, 从而抑制全细胞K+in通道的电流大小(Sutter et al., 2006, 2007).进一步研究表明, SNARE还可以通过与AtKC1互作调控K+in通道的蛋白活性(Honsbein et al., 2009; Grefen et al., 2010; Jeanguenin et al., 2011).由此可知, SNARE作为K+in通道的上游调控因子可通过对K+in四聚体组成形式的影响发挥调控作用.蛋白激酶和磷酸酶参与调控多种蛋白的功能, 包括内向和外向质膜K+通道, 比如CPK (calcium-dependent protein kinase)家族的CPK33对GORK的激活作用和PP2CA对GORK的抑制作用(Lefoulon et al., 2016; Corratgé-Faillie et al., 2017).K+in通道可以被多种蛋白激酶磷酸化所抑制, 包括CPK家族和SnRK (Rnf1-related protein kinase)家族的多个蛋白激酶(Sato et al., 2009, 2010; Ronzier et al., 2014).此外, AKT1等多个质膜K+in通道的活性还受Ca2+信号及其与CBL-CIPK组成的信号链的正调控(Li et al., 2006; Cheong et al., 2007; Zhang et al., 2010; Ren et al., 2013; Behera et al., 2017).而PP2CA对AKT1的去磷酸化可以与Ca2+-CBL-CIPK信号链协同调控AKT1的活性(Lan et al., 2011).有趣的是, ABA甚至可以直接激活GORK的离子通道活性(Ooi et al., 2017).K+通道的调控因子众多, 这些调控因子并非各自单独发挥作用, 而是彼此协同发挥作用.AtKC1和Ca2+-CBL-CIPK23信号链协同调控AKT1就是一个典型的例子(Wang et al., 2016). ...
Stretch-activated chloride, potassium, and calcium channels coexisting in plasma membranes of guard cells of Vicia faba L 1 1991
... 除了SLAC1家族, 植物细胞质膜上还包含张力激活的阴离子通道MSL (mechano-sensitive or stret- ch-activated channels) (Crosgrove and Hedrich, 1991).MSL为典型的张力激活的阴离子通道家族.MSL9和MSL10是最早得到电生理鉴定的MSL家族成员, 定位于细胞质膜, 其激活对质膜张力具有依赖性, 主要选择性介导阴离子的跨膜流动(Haswell et al., 2008; Maksaev and Haswell, 2012).随后有研究表明, 拟南芥MSL10参与植物机械损伤信号反应和茉莉酸的累积(Veley et al., 2014; Zou et al., 2016).此外, 有些拟南芥MSL成员定位在细胞器膜上, 调控细胞器的大小和形状(Veley et al., 2012).根据氨基酸残基序列预测, MSL蛋白存在多个跨膜区, 具备形成离子通道的基本结构特点.而质膜受到拉伸时对MSL的激活是通过机械拉力直接改变MSL蛋白的结构, 使离子通过的孔被拉开.但这基本上还停留在理论推测阶段, 缺乏有力的直接实验证据支持.总体来讲, 目前对MSL家族的生物学功能还了解较少, 其组成的离子通道蛋白结构、感知生物膜张力的分子机理和生物学功能等还有待深入研究. ...
Silent S-type anion channel subunit SLAH1 gates SLAH3 open for chloride root-to-shoot trans- location 2 2016
... SLAC1家族是较晚被发现的一个植物细胞质膜阴离子通道家族, 但植物进化分析表明, SLAC1基因从低等到高等植物均很保守(Lind et al., 2015).拟南芥SLAC1家族由5个成员组成, 包括SLAC1及其同源蛋白SLAH1、2、3和4.其中, SLAC1主要定位在气孔保卫细胞质膜上, 通过介导Cl-和NO3-的外流驱动气孔关闭, 是干旱胁迫及ABA、CO2、臭氧和活性氧(reactive oxygen species, ROS)等信号传递途径的关键组分(Negi et al., 2008; Vahisalu et al., 2008).SLAH3在气孔保卫细胞中分布, 与SLAC1一起通过介导NO3-和Cl-外流驱动气孔关闭(Geiger et al., 2011), 但SLAH3在气孔关闭运动中的作用, 相对于SLAC1而言是辅助性的.拟南芥4个SLAH主要分布在根、茎和叶脉等输导组织中(Negi et al., 2008).其中, SLAH1通过调控SLAH3的活性调控Cl-从根向茎的运输(Cubero-Font et al., 2016).在离子选择性和通透性方面, 拟南芥SLAC1和SLAH3均对Cl-和NO3-有明显的通透能力, 但对NO3-的通透能力明显大于Cl- (Geiger et al., 2009; Lee et al., 2009; Brandt et al., 2012).而SLAH2则为NO3-选择性的阴离子通道, 对Cl-的通透能力微弱(Maierhofer et al., 2014).最近的研究表明, 来自单子叶植物玉米和水稻的SLAC1, 相对于Cl-而言, 对NO3-的选择性明显大于拟南芥SLAC1 (Sun et al., 2016; Qi et al., 2018); 而鸭跖草SLAC1则为NO3-激活的Cl-通透性的阴离子通道(Müller et al., 2017).因此, 来自不同高等植物的SLAC1, 其离子选择性不完全一致, 即不同的高等植物可能采用不同的阴离子通道组合来驱动气孔关闭运动.需要注意的是, 尽管在几十年前就发现苹果酸在气孔保卫细胞内大量积累, 并曾被认为是气孔关闭运动中主要的外流阴离子物质.但实际上, 在气孔关闭运动中发挥核心作用的SLAC1和SLAH3均对苹果酸没有明显的通透能力, 真正通过介导苹果酸外流而参与驱动气孔关闭运动的是来自ALMT家族的ALMT12.因此, 气孔关闭运动应该是由SLAC1、SLAH3和ALMT12等多种质膜阴离子协同作用的结果.每个SLAC1亚基具有10个跨膜区, 其C端和N端均位于质膜的胞质一侧(Vahisalu et al., 2008) (图6A).3个SLAC1亚基互相结合, 以三聚体的形式形成质膜阴离子通道, 但每个亚基独立形成1个允许阴离子通过的孔(Chen et al., 2010) (图6B).由此可见, 包括多个SLAHs在内, SLAC1家族形成的阴离子通道可能均为三聚体, 而SLAH1对SLAH3活性的调控作用, 有可能通过与SLAH3组成异源三聚体而实现.SLAC1家族的离子通道亚基组成形式与CNGC和Shaker家族不同, 但它们在不同亚基组成影响离子通道活性和选择性方面又具有类似之处.SLAC1家族离子通道的激活主要依赖蛋白激酶的磷酸化, 而失活则依赖于磷酸酶的去磷酸化(Geiger et al., 2009, 2011; Lee et al., 2009; Brandt et al., 2012; Maierhofer et al., 2014; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
... ; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
Tetramerization of the AKT1 plant potassium chan- nel involves its C-terminal cytoplasmic domain 1 1997
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
a). The vacuolar channel VvALMT9 mediates malate and tartrate accumulation in berries of Vitis vinifera 1 2013
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
b). AtALMT9 is a malate-activated vacuolar chloride channel required for stomatal opening in Arabidopsis 1 2013
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
Plant K + channel alpha-subunits assemble indiscriminately 1 1997
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
Assembly of plant Shaker-like Kout channels requires two distinct sites of the channel α-subunit 1 2004
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
AtKC1, a conditionally targeted Shaker-type subunit, regulates the activity of plant K + channels 1 2008
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
Identification and characteri- zation of stretch-activated ion channels in pollen protop- lasts 1 2004
... 早期的研究中, 人们发现植物细胞中存在一类受质膜张力激活的Ca2+信号, 从而预测在植物细胞质膜上存在张力激活类质膜Ca2+通道(Crosgrove and Hedrich, 1991; Dutta and Robinson, 2004; Qi et al., 2004; Zhang et al., 2007).多年来, 人们一直没有鉴定到此类受机械或渗透张力激活的离子通道编码基因.近年来, 科学家找到了2类对质膜张力敏感的离子通道, 其一是受低渗条件激活, 通过介导胞外Ca2+内流发挥功能的MCA1和MCA2 (Iida et al., 2014); 其二是对高渗胁迫敏感的OSCA家族(Hou et al., 2014; Yuan et al., 2014).MCA1和MCA2蛋白只有1个跨膜区, 其N端较短且位于细胞质膜外侧, 而C端较长且位于质膜内侧(图3A).研究表明, MCA1和MCA2的N端和跨膜区为其离子通道组成及活性所必需, 而C端则对其离子通道活性发挥调控作用(Kam- ano et al., 2015).MCA1和MCA2组成的离子通道也为四聚体结构, 张力和低渗等刺激通过拉伸质膜改变该离子通道蛋白的结构, 从而使通道被激活, 引起胞外Ca2+内流和细胞质Ca2+水平升高(图3B).MCA1的拟南芥突变体根无法插入较硬的固体培养基(Nakagawa et al., 2007), 说明其在感知机械压力方面发挥作用.最近的研究表明, MCA1和MCA2还作为质膜Ca2+通道介导拟南芥的耐冷反应(Mori et al., 2018).与MCA1和MCA2相反, OSCA是对高渗敏感的质膜离子通道, 可以被细胞外的高渗处理激活(Yuan et al., 2014).拟南芥OSCA家族的蛋白结构与MCA1和MCA2大不相同, 其每个亚基均具有多达9个跨膜区, 在其第8和第9跨膜区之间具有1个孔形成所需的环状结构(Yuan et al., 2014) (图4).OSCA与MCA1和MCA2类似之处在于其亚基N端位于质膜的胞外侧, 而C端则位于质膜胞质侧(Yuan et al., 2014) (图4).OSCA1允许多种离子通过, 其对各种离子的通透能力大小排序为K+>Ba2+≈Ca2+>Na+=Mg2+=Cs+ (Yuan et al., 2014).目前, 人们对质膜张力激活的质膜离子通道的生物学功能了解较少, 但其对渗透敏感的特点暗示此类质膜离子通道可能通过对质膜张力变化做出反应.对低渗敏感的MCA1和MCA2与对高渗敏感的OSCA家族有可能作为一对质膜张力和渗透敏感元件, 协同配合发挥作用, 以共同应对渗透环境变化.一方面, 其可以通过影响胞内Ca2+信号而参与耐盐、耐旱和气孔运动等与细胞渗透和膨压调节密切相关的生物学过程; 另一方面, OSCA对K+的通透能力也表明其可能同时介导Ca2+和K+的跨膜流动而参与这些生物学过程. ...
In vitro Arabidopsis pollen germination and characterization of the inward potassium currents in Arabidopsis pollen grain pro- toplasts 1 2001
... Shaker家族离子通道的活性受多种调控因子的影响.首先, 细胞内和细胞外K+浓度对K+in和K+out通道活性均有明显影响.这种影响可能是基于K+从高浓度向低浓度方向流动的基本化学特性.酸性环境可以显著抑制拟南芥花粉原生质体质膜K+in通道活性, 而碱性条件则具有一定的激活作用(Fan et al., 2001, 2003).这种作用可能通过影响K+in通道的某些氨基酸残基电离状态和离子通道蛋白结构变化来实现.其次, K+in四聚体的亚基组成形式本身就是一种通道活性调控方式.在非洲爪蟾卵母细胞中, 以不同的排列组合共表达2个不同的拟南芥Shaker家族K+in通道基因, 可以利用电生理技术记录到具有不同电生理特性的K+in通道电流信号(Xicluna et al., 2007; Lebaudy et al., 2010).而AtKC1作为抑制亚基, 通过重组进入K+in四聚体发挥对K+in通道的抑制作用, 是K+in通道的重要负调控因子.K+in四聚体组成形式的变化可能通过影响K+in通道蛋白离子通道孔的结构而影响其离子通道活性.SNARE蛋白SYP121不仅可以影响KAT1等K+in通道的亚细胞定位, 而且可使KAT1内化离开细胞质膜, 从而抑制全细胞K+in通道的电流大小(Sutter et al., 2006, 2007).进一步研究表明, SNARE还可以通过与AtKC1互作调控K+in通道的蛋白活性(Honsbein et al., 2009; Grefen et al., 2010; Jeanguenin et al., 2011).由此可知, SNARE作为K+in通道的上游调控因子可通过对K+in四聚体组成形式的影响发挥调控作用.蛋白激酶和磷酸酶参与调控多种蛋白的功能, 包括内向和外向质膜K+通道, 比如CPK (calcium-dependent protein kinase)家族的CPK33对GORK的激活作用和PP2CA对GORK的抑制作用(Lefoulon et al., 2016; Corratgé-Faillie et al., 2017).K+in通道可以被多种蛋白激酶磷酸化所抑制, 包括CPK家族和SnRK (Rnf1-related protein kinase)家族的多个蛋白激酶(Sato et al., 2009, 2010; Ronzier et al., 2014).此外, AKT1等多个质膜K+in通道的活性还受Ca2+信号及其与CBL-CIPK组成的信号链的正调控(Li et al., 2006; Cheong et al., 2007; Zhang et al., 2010; Ren et al., 2013; Behera et al., 2017).而PP2CA对AKT1的去磷酸化可以与Ca2+-CBL-CIPK信号链协同调控AKT1的活性(Lan et al., 2011).有趣的是, ABA甚至可以直接激活GORK的离子通道活性(Ooi et al., 2017).K+通道的调控因子众多, 这些调控因子并非各自单独发挥作用, 而是彼此协同发挥作用.AtKC1和Ca2+-CBL-CIPK23信号链协同调控AKT1就是一个典型的例子(Wang et al., 2016). ...
Outward K + channels in Brassica chinensis pollen protoplasts are regulated by external and internal pH 1 2003
... Shaker家族离子通道的活性受多种调控因子的影响.首先, 细胞内和细胞外K+浓度对K+in和K+out通道活性均有明显影响.这种影响可能是基于K+从高浓度向低浓度方向流动的基本化学特性.酸性环境可以显著抑制拟南芥花粉原生质体质膜K+in通道活性, 而碱性条件则具有一定的激活作用(Fan et al., 2001, 2003).这种作用可能通过影响K+in通道的某些氨基酸残基电离状态和离子通道蛋白结构变化来实现.其次, K+in四聚体的亚基组成形式本身就是一种通道活性调控方式.在非洲爪蟾卵母细胞中, 以不同的排列组合共表达2个不同的拟南芥Shaker家族K+in通道基因, 可以利用电生理技术记录到具有不同电生理特性的K+in通道电流信号(Xicluna et al., 2007; Lebaudy et al., 2010).而AtKC1作为抑制亚基, 通过重组进入K+in四聚体发挥对K+in通道的抑制作用, 是K+in通道的重要负调控因子.K+in四聚体组成形式的变化可能通过影响K+in通道蛋白离子通道孔的结构而影响其离子通道活性.SNARE蛋白SYP121不仅可以影响KAT1等K+in通道的亚细胞定位, 而且可使KAT1内化离开细胞质膜, 从而抑制全细胞K+in通道的电流大小(Sutter et al., 2006, 2007).进一步研究表明, SNARE还可以通过与AtKC1互作调控K+in通道的蛋白活性(Honsbein et al., 2009; Grefen et al., 2010; Jeanguenin et al., 2011).由此可知, SNARE作为K+in通道的上游调控因子可通过对K+in四聚体组成形式的影响发挥调控作用.蛋白激酶和磷酸酶参与调控多种蛋白的功能, 包括内向和外向质膜K+通道, 比如CPK (calcium-dependent protein kinase)家族的CPK33对GORK的激活作用和PP2CA对GORK的抑制作用(Lefoulon et al., 2016; Corratgé-Faillie et al., 2017).K+in通道可以被多种蛋白激酶磷酸化所抑制, 包括CPK家族和SnRK (Rnf1-related protein kinase)家族的多个蛋白激酶(Sato et al., 2009, 2010; Ronzier et al., 2014).此外, AKT1等多个质膜K+in通道的活性还受Ca2+信号及其与CBL-CIPK组成的信号链的正调控(Li et al., 2006; Cheong et al., 2007; Zhang et al., 2010; Ren et al., 2013; Behera et al., 2017).而PP2CA对AKT1的去磷酸化可以与Ca2+-CBL-CIPK信号链协同调控AKT1的活性(Lan et al., 2011).有趣的是, ABA甚至可以直接激活GORK的离子通道活性(Ooi et al., 2017).K+通道的调控因子众多, 这些调控因子并非各自单独发挥作用, 而是彼此协同发挥作用.AtKC1和Ca2+-CBL-CIPK23信号链协同调控AKT1就是一个典型的例子(Wang et al., 2016). ...
Plasma membrane cyclic nucleotide gated calcium channels control land plant thermal sensing and acquired thermotolerance 1 2012
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
Candidate gene identification of an aluminum-activated organic acid transporter gene at the 1 2007
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
Rice K + uptake channel OsAKT1 is sensitive to salt stress 1 2005
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
A heat-activated calcium-permeable channel—Arabidopsis cyclic nucleotide-gated ion channel 6—is involved in heat shock responses 1 2012
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
Arabidopsis CNGC18 is a Ca 2+-permeable channel 1 2014
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
Cyclic nucleotide- gated channel 18 is an essential Ca 2+ channel in pollen tube tips for pollen tube guidance to ovules in Arabidopsis 2 2016
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
... ; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
The K + channel KZM2 is involved in stomatal movement by modulating inward K + currents in maize guard cells 1 2017
... 目前, 对拟南芥Shaker家族的组织分布和功能研究得较为深入全面; 而对包括水稻(Oryza sativa)和玉米(Zea mays)在内的其它植物Shaker家族的研究相对较少.已有的研究鉴定出多个参与K+吸收、转运和气孔运动的玉米和水稻Shaker同源基因.其与拟南芥同源蛋白相比较有很多类似之处, 但也有不同.例如, 玉米KZM2通过与KZM3组成异源多聚体, 抑制K+in通道的活性和气孔运动(Gao et al., 2017).而水稻OsKAT3通过与OsKAT2互作抑制OsKAT3的活性(Hwang et al., 2013).这种K+通道两个同源蛋白之间的互作和调控方式与拟南芥中的AtKC1对AKT1和KAT1等的抑制作用类似(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).再比如, 水稻KAT2与拟南芥KAT2均作为K+in通道参与光照诱导的气孔开放, 但拟南芥气孔保卫细胞质膜K+in通道编码基因数目明显多于水稻(Pilot et al., 2001; Moon et al., 2017).对于外向K+通道而言, 拟南芥SKOR分布于根部, 负责K+向木质部的卸载, 而GORK则主要参与拟南芥的气孔关闭, 二者的组织分布和生物学功能明显不同.而水稻的同一个Shaker蛋白同时参与气孔关闭和K+营养在根部的卸载(Nguyen et al., 2017).因此, 加强对更多植物, 尤其是重要作物中Shaker家族的研究, 将会促进人们对植物吸收、运输和利用K+的分子机理的深入理解. ...
Identification and disruption of a plant shaker-like outward channel involved in K + release into the xylem sap 1 1998
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
a). Heteromeric AtKC1·AKT1 channels in Arabidopsis roots facilitate growth under K +-limiting conditions 5 2009
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
... ; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
... 目前, 对拟南芥Shaker家族的组织分布和功能研究得较为深入全面; 而对包括水稻(Oryza sativa)和玉米(Zea mays)在内的其它植物Shaker家族的研究相对较少.已有的研究鉴定出多个参与K+吸收、转运和气孔运动的玉米和水稻Shaker同源基因.其与拟南芥同源蛋白相比较有很多类似之处, 但也有不同.例如, 玉米KZM2通过与KZM3组成异源多聚体, 抑制K+in通道的活性和气孔运动(Gao et al., 2017).而水稻OsKAT3通过与OsKAT2互作抑制OsKAT3的活性(Hwang et al., 2013).这种K+通道两个同源蛋白之间的互作和调控方式与拟南芥中的AtKC1对AKT1和KAT1等的抑制作用类似(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).再比如, 水稻KAT2与拟南芥KAT2均作为K+in通道参与光照诱导的气孔开放, 但拟南芥气孔保卫细胞质膜K+in通道编码基因数目明显多于水稻(Pilot et al., 2001; Moon et al., 2017).对于外向K+通道而言, 拟南芥SKOR分布于根部, 负责K+向木质部的卸载, 而GORK则主要参与拟南芥的气孔关闭, 二者的组织分布和生物学功能明显不同.而水稻的同一个Shaker蛋白同时参与气孔关闭和K+营养在根部的卸载(Nguyen et al., 2017).因此, 加强对更多植物, 尤其是重要作物中Shaker家族的研究, 将会促进人们对植物吸收、运输和利用K+的分子机理的深入理解. ...
... SLAC1家族是较晚被发现的一个植物细胞质膜阴离子通道家族, 但植物进化分析表明, SLAC1基因从低等到高等植物均很保守(Lind et al., 2015).拟南芥SLAC1家族由5个成员组成, 包括SLAC1及其同源蛋白SLAH1、2、3和4.其中, SLAC1主要定位在气孔保卫细胞质膜上, 通过介导Cl-和NO3-的外流驱动气孔关闭, 是干旱胁迫及ABA、CO2、臭氧和活性氧(reactive oxygen species, ROS)等信号传递途径的关键组分(Negi et al., 2008; Vahisalu et al., 2008).SLAH3在气孔保卫细胞中分布, 与SLAC1一起通过介导NO3-和Cl-外流驱动气孔关闭(Geiger et al., 2011), 但SLAH3在气孔关闭运动中的作用, 相对于SLAC1而言是辅助性的.拟南芥4个SLAH主要分布在根、茎和叶脉等输导组织中(Negi et al., 2008).其中, SLAH1通过调控SLAH3的活性调控Cl-从根向茎的运输(Cubero-Font et al., 2016).在离子选择性和通透性方面, 拟南芥SLAC1和SLAH3均对Cl-和NO3-有明显的通透能力, 但对NO3-的通透能力明显大于Cl- (Geiger et al., 2009; Lee et al., 2009; Brandt et al., 2012).而SLAH2则为NO3-选择性的阴离子通道, 对Cl-的通透能力微弱(Maierhofer et al., 2014).最近的研究表明, 来自单子叶植物玉米和水稻的SLAC1, 相对于Cl-而言, 对NO3-的选择性明显大于拟南芥SLAC1 (Sun et al., 2016; Qi et al., 2018); 而鸭跖草SLAC1则为NO3-激活的Cl-通透性的阴离子通道(Müller et al., 2017).因此, 来自不同高等植物的SLAC1, 其离子选择性不完全一致, 即不同的高等植物可能采用不同的阴离子通道组合来驱动气孔关闭运动.需要注意的是, 尽管在几十年前就发现苹果酸在气孔保卫细胞内大量积累, 并曾被认为是气孔关闭运动中主要的外流阴离子物质.但实际上, 在气孔关闭运动中发挥核心作用的SLAC1和SLAH3均对苹果酸没有明显的通透能力, 真正通过介导苹果酸外流而参与驱动气孔关闭运动的是来自ALMT家族的ALMT12.因此, 气孔关闭运动应该是由SLAC1、SLAH3和ALMT12等多种质膜阴离子协同作用的结果.每个SLAC1亚基具有10个跨膜区, 其C端和N端均位于质膜的胞质一侧(Vahisalu et al., 2008) (图6A).3个SLAC1亚基互相结合, 以三聚体的形式形成质膜阴离子通道, 但每个亚基独立形成1个允许阴离子通过的孔(Chen et al., 2010) (图6B).由此可见, 包括多个SLAHs在内, SLAC1家族形成的阴离子通道可能均为三聚体, 而SLAH1对SLAH3活性的调控作用, 有可能通过与SLAH3组成异源三聚体而实现.SLAC1家族的离子通道亚基组成形式与CNGC和Shaker家族不同, 但它们在不同亚基组成影响离子通道活性和选择性方面又具有类似之处.SLAC1家族离子通道的激活主要依赖蛋白激酶的磷酸化, 而失活则依赖于磷酸酶的去磷酸化(Geiger et al., 2009, 2011; Lee et al., 2009; Brandt et al., 2012; Maierhofer et al., 2014; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
... ).由此可见, 包括多个SLAHs在内, SLAC1家族形成的阴离子通道可能均为三聚体, 而SLAH1对SLAH3活性的调控作用, 有可能通过与SLAH3组成异源三聚体而实现.SLAC1家族的离子通道亚基组成形式与CNGC和Shaker家族不同, 但它们在不同亚基组成影响离子通道活性和选择性方面又具有类似之处.SLAC1家族离子通道的激活主要依赖蛋白激酶的磷酸化, 而失活则依赖于磷酸酶的去磷酸化(Geiger et al., 2009, 2011; Lee et al., 2009; Brandt et al., 2012; Maierhofer et al., 2014; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
Stomatal closure by fast abscisic acid signaling is mediated by the guard cell anion channel SLAH3 and the receptor RCAR1 2 2011
... SLAC1家族是较晚被发现的一个植物细胞质膜阴离子通道家族, 但植物进化分析表明, SLAC1基因从低等到高等植物均很保守(Lind et al., 2015).拟南芥SLAC1家族由5个成员组成, 包括SLAC1及其同源蛋白SLAH1、2、3和4.其中, SLAC1主要定位在气孔保卫细胞质膜上, 通过介导Cl-和NO3-的外流驱动气孔关闭, 是干旱胁迫及ABA、CO2、臭氧和活性氧(reactive oxygen species, ROS)等信号传递途径的关键组分(Negi et al., 2008; Vahisalu et al., 2008).SLAH3在气孔保卫细胞中分布, 与SLAC1一起通过介导NO3-和Cl-外流驱动气孔关闭(Geiger et al., 2011), 但SLAH3在气孔关闭运动中的作用, 相对于SLAC1而言是辅助性的.拟南芥4个SLAH主要分布在根、茎和叶脉等输导组织中(Negi et al., 2008).其中, SLAH1通过调控SLAH3的活性调控Cl-从根向茎的运输(Cubero-Font et al., 2016).在离子选择性和通透性方面, 拟南芥SLAC1和SLAH3均对Cl-和NO3-有明显的通透能力, 但对NO3-的通透能力明显大于Cl- (Geiger et al., 2009; Lee et al., 2009; Brandt et al., 2012).而SLAH2则为NO3-选择性的阴离子通道, 对Cl-的通透能力微弱(Maierhofer et al., 2014).最近的研究表明, 来自单子叶植物玉米和水稻的SLAC1, 相对于Cl-而言, 对NO3-的选择性明显大于拟南芥SLAC1 (Sun et al., 2016; Qi et al., 2018); 而鸭跖草SLAC1则为NO3-激活的Cl-通透性的阴离子通道(Müller et al., 2017).因此, 来自不同高等植物的SLAC1, 其离子选择性不完全一致, 即不同的高等植物可能采用不同的阴离子通道组合来驱动气孔关闭运动.需要注意的是, 尽管在几十年前就发现苹果酸在气孔保卫细胞内大量积累, 并曾被认为是气孔关闭运动中主要的外流阴离子物质.但实际上, 在气孔关闭运动中发挥核心作用的SLAC1和SLAH3均对苹果酸没有明显的通透能力, 真正通过介导苹果酸外流而参与驱动气孔关闭运动的是来自ALMT家族的ALMT12.因此, 气孔关闭运动应该是由SLAC1、SLAH3和ALMT12等多种质膜阴离子协同作用的结果.每个SLAC1亚基具有10个跨膜区, 其C端和N端均位于质膜的胞质一侧(Vahisalu et al., 2008) (图6A).3个SLAC1亚基互相结合, 以三聚体的形式形成质膜阴离子通道, 但每个亚基独立形成1个允许阴离子通过的孔(Chen et al., 2010) (图6B).由此可见, 包括多个SLAHs在内, SLAC1家族形成的阴离子通道可能均为三聚体, 而SLAH1对SLAH3活性的调控作用, 有可能通过与SLAH3组成异源三聚体而实现.SLAC1家族的离子通道亚基组成形式与CNGC和Shaker家族不同, 但它们在不同亚基组成影响离子通道活性和选择性方面又具有类似之处.SLAC1家族离子通道的激活主要依赖蛋白激酶的磷酸化, 而失活则依赖于磷酸酶的去磷酸化(Geiger et al., 2009, 2011; Lee et al., 2009; Brandt et al., 2012; Maierhofer et al., 2014; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
... , 2011; Lee et al., 2009; Brandt et al., 2012; Maierhofer et al., 2014; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
b). Activity of guard cell anion channel SLAC1 is controlled by drought-stress signaling kinase- phosphatase pair 0 2009
A novel motif essential for SNARE interaction with the K + channel KC1 and channel gating in Arabidopsis 1 2010
... Shaker家族离子通道的活性受多种调控因子的影响.首先, 细胞内和细胞外K+浓度对K+in和K+out通道活性均有明显影响.这种影响可能是基于K+从高浓度向低浓度方向流动的基本化学特性.酸性环境可以显著抑制拟南芥花粉原生质体质膜K+in通道活性, 而碱性条件则具有一定的激活作用(Fan et al., 2001, 2003).这种作用可能通过影响K+in通道的某些氨基酸残基电离状态和离子通道蛋白结构变化来实现.其次, K+in四聚体的亚基组成形式本身就是一种通道活性调控方式.在非洲爪蟾卵母细胞中, 以不同的排列组合共表达2个不同的拟南芥Shaker家族K+in通道基因, 可以利用电生理技术记录到具有不同电生理特性的K+in通道电流信号(Xicluna et al., 2007; Lebaudy et al., 2010).而AtKC1作为抑制亚基, 通过重组进入K+in四聚体发挥对K+in通道的抑制作用, 是K+in通道的重要负调控因子.K+in四聚体组成形式的变化可能通过影响K+in通道蛋白离子通道孔的结构而影响其离子通道活性.SNARE蛋白SYP121不仅可以影响KAT1等K+in通道的亚细胞定位, 而且可使KAT1内化离开细胞质膜, 从而抑制全细胞K+in通道的电流大小(Sutter et al., 2006, 2007).进一步研究表明, SNARE还可以通过与AtKC1互作调控K+in通道的蛋白活性(Honsbein et al., 2009; Grefen et al., 2010; Jeanguenin et al., 2011).由此可知, SNARE作为K+in通道的上游调控因子可通过对K+in四聚体组成形式的影响发挥调控作用.蛋白激酶和磷酸酶参与调控多种蛋白的功能, 包括内向和外向质膜K+通道, 比如CPK (calcium-dependent protein kinase)家族的CPK33对GORK的激活作用和PP2CA对GORK的抑制作用(Lefoulon et al., 2016; Corratgé-Faillie et al., 2017).K+in通道可以被多种蛋白激酶磷酸化所抑制, 包括CPK家族和SnRK (Rnf1-related protein kinase)家族的多个蛋白激酶(Sato et al., 2009, 2010; Ronzier et al., 2014).此外, AKT1等多个质膜K+in通道的活性还受Ca2+信号及其与CBL-CIPK组成的信号链的正调控(Li et al., 2006; Cheong et al., 2007; Zhang et al., 2010; Ren et al., 2013; Behera et al., 2017).而PP2CA对AKT1的去磷酸化可以与Ca2+-CBL-CIPK信号链协同调控AKT1的活性(Lan et al., 2011).有趣的是, ABA甚至可以直接激活GORK的离子通道活性(Ooi et al., 2017).K+通道的调控因子众多, 这些调控因子并非各自单独发挥作用, 而是彼此协同发挥作用.AtKC1和Ca2+-CBL-CIPK23信号链协同调控AKT1就是一个典型的例子(Wang et al., 2016). ...
HvALMT1 from barley is involved in the transport of organic anions 1 2010
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
Cyclic nucleotide-gated channel 18 functions as an essential Ca 2+ channel for pollen germination and pollen tube growth in Arabidopsis 1 2017
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
The cyclic nucleotide-gated channel, AtCNGC10, influences salt tolerance in Arabidopsis 1 2008
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
The cyclic nucleotide-gated channel AtCNGC10 transports Ca 2+ and Mg 2+ in Arabidopsis 1 2010
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
Two MscS homologs provide mechanosensitive channel activities in the Arabidopsis root 1 2008
... 除了SLAC1家族, 植物细胞质膜上还包含张力激活的阴离子通道MSL (mechano-sensitive or stret- ch-activated channels) (Crosgrove and Hedrich, 1991).MSL为典型的张力激活的阴离子通道家族.MSL9和MSL10是最早得到电生理鉴定的MSL家族成员, 定位于细胞质膜, 其激活对质膜张力具有依赖性, 主要选择性介导阴离子的跨膜流动(Haswell et al., 2008; Maksaev and Haswell, 2012).随后有研究表明, 拟南芥MSL10参与植物机械损伤信号反应和茉莉酸的累积(Veley et al., 2014; Zou et al., 2016).此外, 有些拟南芥MSL成员定位在细胞器膜上, 调控细胞器的大小和形状(Veley et al., 2012).根据氨基酸残基序列预测, MSL蛋白存在多个跨膜区, 具备形成离子通道的基本结构特点.而质膜受到拉伸时对MSL的激活是通过机械拉力直接改变MSL蛋白的结构, 使离子通过的孔被拉开.但这基本上还停留在理论推测阶段, 缺乏有力的直接实验证据支持.总体来讲, 目前对MSL家族的生物学功能还了解较少, 其组成的离子通道蛋白结构、感知生物膜张力的分子机理和生物学功能等还有待深入研究. ...
AtALMT1, which encodes a malate transporter, is identified as one of several genes critical for aluminum tolerance in Arabidopsis 2 2006
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
... ; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
A tripartite SNARE-K + channel complex mediates in channel-dependent K + nutrition in Arabidopsis 1 2009
... Shaker家族离子通道的活性受多种调控因子的影响.首先, 细胞内和细胞外K+浓度对K+in和K+out通道活性均有明显影响.这种影响可能是基于K+从高浓度向低浓度方向流动的基本化学特性.酸性环境可以显著抑制拟南芥花粉原生质体质膜K+in通道活性, 而碱性条件则具有一定的激活作用(Fan et al., 2001, 2003).这种作用可能通过影响K+in通道的某些氨基酸残基电离状态和离子通道蛋白结构变化来实现.其次, K+in四聚体的亚基组成形式本身就是一种通道活性调控方式.在非洲爪蟾卵母细胞中, 以不同的排列组合共表达2个不同的拟南芥Shaker家族K+in通道基因, 可以利用电生理技术记录到具有不同电生理特性的K+in通道电流信号(Xicluna et al., 2007; Lebaudy et al., 2010).而AtKC1作为抑制亚基, 通过重组进入K+in四聚体发挥对K+in通道的抑制作用, 是K+in通道的重要负调控因子.K+in四聚体组成形式的变化可能通过影响K+in通道蛋白离子通道孔的结构而影响其离子通道活性.SNARE蛋白SYP121不仅可以影响KAT1等K+in通道的亚细胞定位, 而且可使KAT1内化离开细胞质膜, 从而抑制全细胞K+in通道的电流大小(Sutter et al., 2006, 2007).进一步研究表明, SNARE还可以通过与AtKC1互作调控K+in通道的蛋白活性(Honsbein et al., 2009; Grefen et al., 2010; Jeanguenin et al., 2011).由此可知, SNARE作为K+in通道的上游调控因子可通过对K+in四聚体组成形式的影响发挥调控作用.蛋白激酶和磷酸酶参与调控多种蛋白的功能, 包括内向和外向质膜K+通道, 比如CPK (calcium-dependent protein kinase)家族的CPK33对GORK的激活作用和PP2CA对GORK的抑制作用(Lefoulon et al., 2016; Corratgé-Faillie et al., 2017).K+in通道可以被多种蛋白激酶磷酸化所抑制, 包括CPK家族和SnRK (Rnf1-related protein kinase)家族的多个蛋白激酶(Sato et al., 2009, 2010; Ronzier et al., 2014).此外, AKT1等多个质膜K+in通道的活性还受Ca2+信号及其与CBL-CIPK组成的信号链的正调控(Li et al., 2006; Cheong et al., 2007; Zhang et al., 2010; Ren et al., 2013; Behera et al., 2017).而PP2CA对AKT1的去磷酸化可以与Ca2+-CBL-CIPK信号链协同调控AKT1的活性(Lan et al., 2011).有趣的是, ABA甚至可以直接激活GORK的离子通道活性(Ooi et al., 2017).K+通道的调控因子众多, 这些调控因子并非各自单独发挥作用, 而是彼此协同发挥作用.AtKC1和Ca2+-CBL-CIPK23信号链协同调控AKT1就是一个典型的例子(Wang et al., 2016). ...
The Arabidopsis outward K + channel GORK is involved in regulation of stomatal movements and plant transpiration 1 2003
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
DUF221 proteins are a family of osmosensitive calcium-permeable cation channels conser- ved across eukaryotes 1 2014
... 早期的研究中, 人们发现植物细胞中存在一类受质膜张力激活的Ca2+信号, 从而预测在植物细胞质膜上存在张力激活类质膜Ca2+通道(Crosgrove and Hedrich, 1991; Dutta and Robinson, 2004; Qi et al., 2004; Zhang et al., 2007).多年来, 人们一直没有鉴定到此类受机械或渗透张力激活的离子通道编码基因.近年来, 科学家找到了2类对质膜张力敏感的离子通道, 其一是受低渗条件激活, 通过介导胞外Ca2+内流发挥功能的MCA1和MCA2 (Iida et al., 2014); 其二是对高渗胁迫敏感的OSCA家族(Hou et al., 2014; Yuan et al., 2014).MCA1和MCA2蛋白只有1个跨膜区, 其N端较短且位于细胞质膜外侧, 而C端较长且位于质膜内侧(图3A).研究表明, MCA1和MCA2的N端和跨膜区为其离子通道组成及活性所必需, 而C端则对其离子通道活性发挥调控作用(Kam- ano et al., 2015).MCA1和MCA2组成的离子通道也为四聚体结构, 张力和低渗等刺激通过拉伸质膜改变该离子通道蛋白的结构, 从而使通道被激活, 引起胞外Ca2+内流和细胞质Ca2+水平升高(图3B).MCA1的拟南芥突变体根无法插入较硬的固体培养基(Nakagawa et al., 2007), 说明其在感知机械压力方面发挥作用.最近的研究表明, MCA1和MCA2还作为质膜Ca2+通道介导拟南芥的耐冷反应(Mori et al., 2018).与MCA1和MCA2相反, OSCA是对高渗敏感的质膜离子通道, 可以被细胞外的高渗处理激活(Yuan et al., 2014).拟南芥OSCA家族的蛋白结构与MCA1和MCA2大不相同, 其每个亚基均具有多达9个跨膜区, 在其第8和第9跨膜区之间具有1个孔形成所需的环状结构(Yuan et al., 2014) (图4).OSCA与MCA1和MCA2类似之处在于其亚基N端位于质膜的胞外侧, 而C端则位于质膜胞质侧(Yuan et al., 2014) (图4).OSCA1允许多种离子通过, 其对各种离子的通透能力大小排序为K+>Ba2+≈Ca2+>Na+=Mg2+=Cs+ (Yuan et al., 2014).目前, 人们对质膜张力激活的质膜离子通道的生物学功能了解较少, 但其对渗透敏感的特点暗示此类质膜离子通道可能通过对质膜张力变化做出反应.对低渗敏感的MCA1和MCA2与对高渗敏感的OSCA家族有可能作为一对质膜张力和渗透敏感元件, 协同配合发挥作用, 以共同应对渗透环境变化.一方面, 其可以通过影响胞内Ca2+信号而参与耐盐、耐旱和气孔运动等与细胞渗透和膨压调节密切相关的生物学过程; 另一方面, OSCA对K+的通透能力也表明其可能同时介导Ca2+和K+的跨膜流动而参与这些生物学过程. ...
Unique features of two potassium channels, OsKAT2 and OsKAT3, expressed in rice guard cells 1 2013
... 目前, 对拟南芥Shaker家族的组织分布和功能研究得较为深入全面; 而对包括水稻(Oryza sativa)和玉米(Zea mays)在内的其它植物Shaker家族的研究相对较少.已有的研究鉴定出多个参与K+吸收、转运和气孔运动的玉米和水稻Shaker同源基因.其与拟南芥同源蛋白相比较有很多类似之处, 但也有不同.例如, 玉米KZM2通过与KZM3组成异源多聚体, 抑制K+in通道的活性和气孔运动(Gao et al., 2017).而水稻OsKAT3通过与OsKAT2互作抑制OsKAT3的活性(Hwang et al., 2013).这种K+通道两个同源蛋白之间的互作和调控方式与拟南芥中的AtKC1对AKT1和KAT1等的抑制作用类似(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).再比如, 水稻KAT2与拟南芥KAT2均作为K+in通道参与光照诱导的气孔开放, 但拟南芥气孔保卫细胞质膜K+in通道编码基因数目明显多于水稻(Pilot et al., 2001; Moon et al., 2017).对于外向K+通道而言, 拟南芥SKOR分布于根部, 负责K+向木质部的卸载, 而GORK则主要参与拟南芥的气孔关闭, 二者的组织分布和生物学功能明显不同.而水稻的同一个Shaker蛋白同时参与气孔关闭和K+营养在根部的卸载(Nguyen et al., 2017).因此, 加强对更多植物, 尤其是重要作物中Shaker家族的研究, 将会促进人们对植物吸收、运输和利用K+的分子机理的深入理解. ...
New candidates for mechano-sensitive channels potentially involved in gravity sensing in Ara- bidopsis thaliana 2 2014
... 植物体内存在大量的钙元素.一方面钙作为细胞壁等结构组分发挥作用; 另一方面, 离子形式的钙(Ca2+)可以作为信号分子参与各种生物学反应和信号传递过程, 后者则备受关注.胞内Ca2+信号以细胞质Ca2+浓度的升高和震荡形式出现.一般认为, 胞外Ca2+内流是细胞质Ca2+信号产生所需Ca2+的主要来源, 而质膜内向Ca2+通道是胞外Ca2+内流的主要渠道.同时, Ca2+通道的活性变化通过控制胞外Ca2+内流的节奏直接调控胞内Ca2+震荡, 发挥Ca2+信号上游调控因子的作用.鉴于此, 质膜Ca2+通道受到广泛关注, 并成为动植物各信号领域的重点研究对象.科学家首先在动物细胞中发现多个Ca2+通道家族, 主要包括受环核苷酸cAMP和cGMP激活的CNG (cyclic nucleotide-gated)家族、受谷氨酸门控的GLR (glutamate receptor)家族、TRPC (transient receptor potential channel)和受胞内Ca2+库倒空激活的CRACM1/Orai1 (Zagotta and Siegelbaum, 1996; Vig and Kinet, 2007; Sobolevsky, 2015; Ambudkar et al., 2017).通过序列比对, 多个动物细胞质膜Ca2+通道的同源基因家族先后在植物尤其是模式植物拟南芥基因组中被鉴定出来, 主要包括CNGC (cyclic nucleotide- gated channel)、GLR (glutamate receptor-like)、高渗敏感的OSCA (hyperosmolality-gated calcium- permeable channel)家族和低渗敏感的张力激活离子通道MCA (mechano-sensitive channel) (M?ser et al., 2001; Ward et al., 2009; Iida et al., 2014; Yuan et al., 2014). ...
... 早期的研究中, 人们发现植物细胞中存在一类受质膜张力激活的Ca2+信号, 从而预测在植物细胞质膜上存在张力激活类质膜Ca2+通道(Crosgrove and Hedrich, 1991; Dutta and Robinson, 2004; Qi et al., 2004; Zhang et al., 2007).多年来, 人们一直没有鉴定到此类受机械或渗透张力激活的离子通道编码基因.近年来, 科学家找到了2类对质膜张力敏感的离子通道, 其一是受低渗条件激活, 通过介导胞外Ca2+内流发挥功能的MCA1和MCA2 (Iida et al., 2014); 其二是对高渗胁迫敏感的OSCA家族(Hou et al., 2014; Yuan et al., 2014).MCA1和MCA2蛋白只有1个跨膜区, 其N端较短且位于细胞质膜外侧, 而C端较长且位于质膜内侧(图3A).研究表明, MCA1和MCA2的N端和跨膜区为其离子通道组成及活性所必需, 而C端则对其离子通道活性发挥调控作用(Kam- ano et al., 2015).MCA1和MCA2组成的离子通道也为四聚体结构, 张力和低渗等刺激通过拉伸质膜改变该离子通道蛋白的结构, 从而使通道被激活, 引起胞外Ca2+内流和细胞质Ca2+水平升高(图3B).MCA1的拟南芥突变体根无法插入较硬的固体培养基(Nakagawa et al., 2007), 说明其在感知机械压力方面发挥作用.最近的研究表明, MCA1和MCA2还作为质膜Ca2+通道介导拟南芥的耐冷反应(Mori et al., 2018).与MCA1和MCA2相反, OSCA是对高渗敏感的质膜离子通道, 可以被细胞外的高渗处理激活(Yuan et al., 2014).拟南芥OSCA家族的蛋白结构与MCA1和MCA2大不相同, 其每个亚基均具有多达9个跨膜区, 在其第8和第9跨膜区之间具有1个孔形成所需的环状结构(Yuan et al., 2014) (图4).OSCA与MCA1和MCA2类似之处在于其亚基N端位于质膜的胞外侧, 而C端则位于质膜胞质侧(Yuan et al., 2014) (图4).OSCA1允许多种离子通过, 其对各种离子的通透能力大小排序为K+>Ba2+≈Ca2+>Na+=Mg2+=Cs+ (Yuan et al., 2014).目前, 人们对质膜张力激活的质膜离子通道的生物学功能了解较少, 但其对渗透敏感的特点暗示此类质膜离子通道可能通过对质膜张力变化做出反应.对低渗敏感的MCA1和MCA2与对高渗敏感的OSCA家族有可能作为一对质膜张力和渗透敏感元件, 协同配合发挥作用, 以共同应对渗透环境变化.一方面, 其可以通过影响胞内Ca2+信号而参与耐盐、耐旱和气孔运动等与细胞渗透和膨压调节密切相关的生物学过程; 另一方面, OSCA对K+的通透能力也表明其可能同时介导Ca2+和K+的跨膜流动而参与这些生物学过程. ...
Voltage-gated and inwardly rectifying potassium channels 1 1997
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
AtKC1 is a general modulator of Arabidopsis inward Shaker channel activity 3 2011
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
... ; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
... Shaker家族离子通道的活性受多种调控因子的影响.首先, 细胞内和细胞外K+浓度对K+in和K+out通道活性均有明显影响.这种影响可能是基于K+从高浓度向低浓度方向流动的基本化学特性.酸性环境可以显著抑制拟南芥花粉原生质体质膜K+in通道活性, 而碱性条件则具有一定的激活作用(Fan et al., 2001, 2003).这种作用可能通过影响K+in通道的某些氨基酸残基电离状态和离子通道蛋白结构变化来实现.其次, K+in四聚体的亚基组成形式本身就是一种通道活性调控方式.在非洲爪蟾卵母细胞中, 以不同的排列组合共表达2个不同的拟南芥Shaker家族K+in通道基因, 可以利用电生理技术记录到具有不同电生理特性的K+in通道电流信号(Xicluna et al., 2007; Lebaudy et al., 2010).而AtKC1作为抑制亚基, 通过重组进入K+in四聚体发挥对K+in通道的抑制作用, 是K+in通道的重要负调控因子.K+in四聚体组成形式的变化可能通过影响K+in通道蛋白离子通道孔的结构而影响其离子通道活性.SNARE蛋白SYP121不仅可以影响KAT1等K+in通道的亚细胞定位, 而且可使KAT1内化离开细胞质膜, 从而抑制全细胞K+in通道的电流大小(Sutter et al., 2006, 2007).进一步研究表明, SNARE还可以通过与AtKC1互作调控K+in通道的蛋白活性(Honsbein et al., 2009; Grefen et al., 2010; Jeanguenin et al., 2011).由此可知, SNARE作为K+in通道的上游调控因子可通过对K+in四聚体组成形式的影响发挥调控作用.蛋白激酶和磷酸酶参与调控多种蛋白的功能, 包括内向和外向质膜K+通道, 比如CPK (calcium-dependent protein kinase)家族的CPK33对GORK的激活作用和PP2CA对GORK的抑制作用(Lefoulon et al., 2016; Corratgé-Faillie et al., 2017).K+in通道可以被多种蛋白激酶磷酸化所抑制, 包括CPK家族和SnRK (Rnf1-related protein kinase)家族的多个蛋白激酶(Sato et al., 2009, 2010; Ronzier et al., 2014).此外, AKT1等多个质膜K+in通道的活性还受Ca2+信号及其与CBL-CIPK组成的信号链的正调控(Li et al., 2006; Cheong et al., 2007; Zhang et al., 2010; Ren et al., 2013; Behera et al., 2017).而PP2CA对AKT1的去磷酸化可以与Ca2+-CBL-CIPK信号链协同调控AKT1的活性(Lan et al., 2011).有趣的是, ABA甚至可以直接激活GORK的离子通道活性(Ooi et al., 2017).K+通道的调控因子众多, 这些调控因子并非各自单独发挥作用, 而是彼此协同发挥作用.AtKC1和Ca2+-CBL-CIPK23信号链协同调控AKT1就是一个典型的例子(Wang et al., 2016). ...
Cyclic nucleotide gated channel 10 negatively regulates salt to- lerance by mediating Na + transport in Arabidopsis 1 2015
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
External K + modulates the activity of the Arabidopsis potassium chan- nel SKOR via an unusual mechanism 1 2006
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
Transmembrane topologies of Ca 2+- permeable mechanosensitive channels MCA1 and MCA2 in Arabidopsis thaliana 3 2015
... 早期的研究中, 人们发现植物细胞中存在一类受质膜张力激活的Ca2+信号, 从而预测在植物细胞质膜上存在张力激活类质膜Ca2+通道(Crosgrove and Hedrich, 1991; Dutta and Robinson, 2004; Qi et al., 2004; Zhang et al., 2007).多年来, 人们一直没有鉴定到此类受机械或渗透张力激活的离子通道编码基因.近年来, 科学家找到了2类对质膜张力敏感的离子通道, 其一是受低渗条件激活, 通过介导胞外Ca2+内流发挥功能的MCA1和MCA2 (Iida et al., 2014); 其二是对高渗胁迫敏感的OSCA家族(Hou et al., 2014; Yuan et al., 2014).MCA1和MCA2蛋白只有1个跨膜区, 其N端较短且位于细胞质膜外侧, 而C端较长且位于质膜内侧(图3A).研究表明, MCA1和MCA2的N端和跨膜区为其离子通道组成及活性所必需, 而C端则对其离子通道活性发挥调控作用(Kam- ano et al., 2015).MCA1和MCA2组成的离子通道也为四聚体结构, 张力和低渗等刺激通过拉伸质膜改变该离子通道蛋白的结构, 从而使通道被激活, 引起胞外Ca2+内流和细胞质Ca2+水平升高(图3B).MCA1的拟南芥突变体根无法插入较硬的固体培养基(Nakagawa et al., 2007), 说明其在感知机械压力方面发挥作用.最近的研究表明, MCA1和MCA2还作为质膜Ca2+通道介导拟南芥的耐冷反应(Mori et al., 2018).与MCA1和MCA2相反, OSCA是对高渗敏感的质膜离子通道, 可以被细胞外的高渗处理激活(Yuan et al., 2014).拟南芥OSCA家族的蛋白结构与MCA1和MCA2大不相同, 其每个亚基均具有多达9个跨膜区, 在其第8和第9跨膜区之间具有1个孔形成所需的环状结构(Yuan et al., 2014) (图4).OSCA与MCA1和MCA2类似之处在于其亚基N端位于质膜的胞外侧, 而C端则位于质膜胞质侧(Yuan et al., 2014) (图4).OSCA1允许多种离子通过, 其对各种离子的通透能力大小排序为K+>Ba2+≈Ca2+>Na+=Mg2+=Cs+ (Yuan et al., 2014).目前, 人们对质膜张力激活的质膜离子通道的生物学功能了解较少, 但其对渗透敏感的特点暗示此类质膜离子通道可能通过对质膜张力变化做出反应.对低渗敏感的MCA1和MCA2与对高渗敏感的OSCA家族有可能作为一对质膜张力和渗透敏感元件, 协同配合发挥作用, 以共同应对渗透环境变化.一方面, 其可以通过影响胞内Ca2+信号而参与耐盐、耐旱和气孔运动等与细胞渗透和膨压调节密切相关的生物学过程; 另一方面, OSCA对K+的通透能力也表明其可能同时介导Ca2+和K+的跨膜流动而参与这些生物学过程. ...
... S: 跨膜区; EF: EF-手型Ca2+结合结构域; N/C端: 氨/羧基端 Protein structure of Arabidopsis MCA1 and MCA2 (A) and a molecular mechanism of how MCA1/2 channels are activated by the tension of plasma membrane (B) (<xref ref-type="bibr" rid="b54">Kamano et al., 2015</xref>) S: The transmembrane domain; EF: EF-hand Ca2+ binding domain; N/C termini: The amino terminal and carboxyl terminal ...
The putative glutamate receptor 1.1 (AtGLR1.1) in Arabidopsis thaliana regulates abscisic acid biosynthesis and signaling to con- trol development and water loss 1 2004
... GLR家族是受多种氨基酸门控的离子通道, 但其通道门控特点并非如其名称仅为谷氨酸所门控, 而是受多种氨基酸影响.1998年, 第1个GLR被克隆出来(Lam et al., 1998), 人们开始对GLR进行研究.研究表明, 拟南芥GLR1和GLR2的N端位于细胞质膜外侧, C端位于细胞质膜内侧, 中间包含4个跨膜区, 但第2个跨膜区并不能完整地跨过细胞质膜(Lam et al., 1998) (图2).这种结构特点明显不同于Shaker和CNGC家族.GLR组成的质膜离子通道的亚基组成、多数GLR离子通道的离子选择性等基本特性, 以及GLR离子通道的活性调控机理目前还不甚清楚.但已经解析出动物GLR四聚体的通道蛋白结构(Price et al., 2012).据此推测, 植物GLR组成的通道蛋白结构可能与动物类似.GLR在植物体内具有多种生物学功能, 包括参与调控花粉管的极性生长(Michard et al., 2011; Wudick et al., 2018)、铝诱导的微管解聚和质膜去极化(Sivaguru et al., 2003)、离子运输(McAinsh and Pittman, 2009)、碳氮代谢(Kang and Turano, 2003)、ABA合成和耐旱反应(Kang et al., 2004; Lu et al., 2014)以及机械损伤信号的长距离传输(Mousavi et al., 2013)等.总体而言, 目前对GLR家族的研究还不够深入.关于植物GLR家族更为详细的介绍可以参阅何明杰等(2016)的综述. ...
The putative glutamate receptor 1.1 (AtGLR1.1) functions as a regulator of carbon and ni- trogen metabolism in Arabidopsis thaliana 1 2003
... GLR家族是受多种氨基酸门控的离子通道, 但其通道门控特点并非如其名称仅为谷氨酸所门控, 而是受多种氨基酸影响.1998年, 第1个GLR被克隆出来(Lam et al., 1998), 人们开始对GLR进行研究.研究表明, 拟南芥GLR1和GLR2的N端位于细胞质膜外侧, C端位于细胞质膜内侧, 中间包含4个跨膜区, 但第2个跨膜区并不能完整地跨过细胞质膜(Lam et al., 1998) (图2).这种结构特点明显不同于Shaker和CNGC家族.GLR组成的质膜离子通道的亚基组成、多数GLR离子通道的离子选择性等基本特性, 以及GLR离子通道的活性调控机理目前还不甚清楚.但已经解析出动物GLR四聚体的通道蛋白结构(Price et al., 2012).据此推测, 植物GLR组成的通道蛋白结构可能与动物类似.GLR在植物体内具有多种生物学功能, 包括参与调控花粉管的极性生长(Michard et al., 2011; Wudick et al., 2018)、铝诱导的微管解聚和质膜去极化(Sivaguru et al., 2003)、离子运输(McAinsh and Pittman, 2009)、碳氮代谢(Kang and Turano, 2003)、ABA合成和耐旱反应(Kang et al., 2004; Lu et al., 2014)以及机械损伤信号的长距离传输(Mousavi et al., 2013)等.总体而言, 目前对GLR家族的研究还不够深入.关于植物GLR家族更为详细的介绍可以参阅何明杰等(2016)的综述. ...
Cyclic nucleotide-gated ion channels 1 2002
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
Characterization of the complex regulation of AtALMT1 expression in response to phytohormones and other indu- cers 1 2013
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
Characterisation of a novel gene family of putative cyclic nucleotide- and calmodulin-regulated ion channels in Arabidopsis thaliana 2 1999
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
... 通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
The Arabidopsis vacuolar malate channel is a member of the ALMT family 1 2007
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
Dominant negative guard cell K + channel mutants reduce inward-rectifying K + currents and light-induced stomatal opening in Arabidopsis 1 2001
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
Mutations in the SLAC1 anion channel slow stomatal opening and severely reduce K + uptake channel activity via enhanced cytosolic [Ca 2+] and increased Ca 2+ sensitivity of K + uptake channels 1 2013
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
Phytosulfokine regulates growth in Arabidopsis through a response module at the plasma membrane that includes CYCLIC NUCLEOTIDE-GATED CHANNEL 17, H +-ATPase, and BAK1 1 2015
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
Glutamate-receptor genes in plants 4 1998
... GLR家族是受多种氨基酸门控的离子通道, 但其通道门控特点并非如其名称仅为谷氨酸所门控, 而是受多种氨基酸影响.1998年, 第1个GLR被克隆出来(Lam et al., 1998), 人们开始对GLR进行研究.研究表明, 拟南芥GLR1和GLR2的N端位于细胞质膜外侧, C端位于细胞质膜内侧, 中间包含4个跨膜区, 但第2个跨膜区并不能完整地跨过细胞质膜(Lam et al., 1998) (图2).这种结构特点明显不同于Shaker和CNGC家族.GLR组成的质膜离子通道的亚基组成、多数GLR离子通道的离子选择性等基本特性, 以及GLR离子通道的活性调控机理目前还不甚清楚.但已经解析出动物GLR四聚体的通道蛋白结构(Price et al., 2012).据此推测, 植物GLR组成的通道蛋白结构可能与动物类似.GLR在植物体内具有多种生物学功能, 包括参与调控花粉管的极性生长(Michard et al., 2011; Wudick et al., 2018)、铝诱导的微管解聚和质膜去极化(Sivaguru et al., 2003)、离子运输(McAinsh and Pittman, 2009)、碳氮代谢(Kang and Turano, 2003)、ABA合成和耐旱反应(Kang et al., 2004; Lu et al., 2014)以及机械损伤信号的长距离传输(Mousavi et al., 2013)等.总体而言, 目前对GLR家族的研究还不够深入.关于植物GLR家族更为详细的介绍可以参阅何明杰等(2016)的综述. ...
... ), 人们开始对GLR进行研究.研究表明, 拟南芥GLR1和GLR2的N端位于细胞质膜外侧, C端位于细胞质膜内侧, 中间包含4个跨膜区, 但第2个跨膜区并不能完整地跨过细胞质膜(Lam et al., 1998) (图2).这种结构特点明显不同于Shaker和CNGC家族.GLR组成的质膜离子通道的亚基组成、多数GLR离子通道的离子选择性等基本特性, 以及GLR离子通道的活性调控机理目前还不甚清楚.但已经解析出动物GLR四聚体的通道蛋白结构(Price et al., 2012).据此推测, 植物GLR组成的通道蛋白结构可能与动物类似.GLR在植物体内具有多种生物学功能, 包括参与调控花粉管的极性生长(Michard et al., 2011; Wudick et al., 2018)、铝诱导的微管解聚和质膜去极化(Sivaguru et al., 2003)、离子运输(McAinsh and Pittman, 2009)、碳氮代谢(Kang and Turano, 2003)、ABA合成和耐旱反应(Kang et al., 2004; Lu et al., 2014)以及机械损伤信号的长距离传输(Mousavi et al., 2013)等.总体而言, 目前对GLR家族的研究还不够深入.关于植物GLR家族更为详细的介绍可以参阅何明杰等(2016)的综述. ...
... 拟南芥GLR亚基的蛋白结构(Lam et al., 1998; Price et al., 2012) S1-S4: 第1-4跨膜区; LBD: 配体结合结构域; N/C端: 氨/羧基末端 ...
... S1-S4: 第1-4跨膜区; LBD: 配体结合结构域; N/C端: 氨/羧基末端 Putative protein structure of an Arabidopsis GLR member (<xref ref-type="bibr" rid="b65">Lam et al., 1998</xref>; <xref ref-type="bibr" rid="b106">Price et al., 2012</xref>) S1-S4: Transmembrane domains 1-4; LBD: Ligand binding domain; N/C termini: The amino terminal and carboxyl terminal ...
Mechanistic analysis of AKT1 regulation by the CBL-CIPK- PP2CA interactions 1 2011
... Shaker家族离子通道的活性受多种调控因子的影响.首先, 细胞内和细胞外K+浓度对K+in和K+out通道活性均有明显影响.这种影响可能是基于K+从高浓度向低浓度方向流动的基本化学特性.酸性环境可以显著抑制拟南芥花粉原生质体质膜K+in通道活性, 而碱性条件则具有一定的激活作用(Fan et al., 2001, 2003).这种作用可能通过影响K+in通道的某些氨基酸残基电离状态和离子通道蛋白结构变化来实现.其次, K+in四聚体的亚基组成形式本身就是一种通道活性调控方式.在非洲爪蟾卵母细胞中, 以不同的排列组合共表达2个不同的拟南芥Shaker家族K+in通道基因, 可以利用电生理技术记录到具有不同电生理特性的K+in通道电流信号(Xicluna et al., 2007; Lebaudy et al., 2010).而AtKC1作为抑制亚基, 通过重组进入K+in四聚体发挥对K+in通道的抑制作用, 是K+in通道的重要负调控因子.K+in四聚体组成形式的变化可能通过影响K+in通道蛋白离子通道孔的结构而影响其离子通道活性.SNARE蛋白SYP121不仅可以影响KAT1等K+in通道的亚细胞定位, 而且可使KAT1内化离开细胞质膜, 从而抑制全细胞K+in通道的电流大小(Sutter et al., 2006, 2007).进一步研究表明, SNARE还可以通过与AtKC1互作调控K+in通道的蛋白活性(Honsbein et al., 2009; Grefen et al., 2010; Jeanguenin et al., 2011).由此可知, SNARE作为K+in通道的上游调控因子可通过对K+in四聚体组成形式的影响发挥调控作用.蛋白激酶和磷酸酶参与调控多种蛋白的功能, 包括内向和外向质膜K+通道, 比如CPK (calcium-dependent protein kinase)家族的CPK33对GORK的激活作用和PP2CA对GORK的抑制作用(Lefoulon et al., 2016; Corratgé-Faillie et al., 2017).K+in通道可以被多种蛋白激酶磷酸化所抑制, 包括CPK家族和SnRK (Rnf1-related protein kinase)家族的多个蛋白激酶(Sato et al., 2009, 2010; Ronzier et al., 2014).此外, AKT1等多个质膜K+in通道的活性还受Ca2+信号及其与CBL-CIPK组成的信号链的正调控(Li et al., 2006; Cheong et al., 2007; Zhang et al., 2010; Ren et al., 2013; Behera et al., 2017).而PP2CA对AKT1的去磷酸化可以与Ca2+-CBL-CIPK信号链协同调控AKT1的活性(Lan et al., 2011).有趣的是, ABA甚至可以直接激活GORK的离子通道活性(Ooi et al., 2017).K+通道的调控因子众多, 这些调控因子并非各自单独发挥作用, 而是彼此协同发挥作用.AtKC1和Ca2+-CBL-CIPK23信号链协同调控AKT1就是一个典型的例子(Wang et al., 2016). ...
Heteromeric K + channels in plants 2 2008
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
... ; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
Preferential KAT1-KAT2 heteromerization determines inward K + current properties in Arabidopsis guard cells 2 2010
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
... Shaker家族离子通道的活性受多种调控因子的影响.首先, 细胞内和细胞外K+浓度对K+in和K+out通道活性均有明显影响.这种影响可能是基于K+从高浓度向低浓度方向流动的基本化学特性.酸性环境可以显著抑制拟南芥花粉原生质体质膜K+in通道活性, 而碱性条件则具有一定的激活作用(Fan et al., 2001, 2003).这种作用可能通过影响K+in通道的某些氨基酸残基电离状态和离子通道蛋白结构变化来实现.其次, K+in四聚体的亚基组成形式本身就是一种通道活性调控方式.在非洲爪蟾卵母细胞中, 以不同的排列组合共表达2个不同的拟南芥Shaker家族K+in通道基因, 可以利用电生理技术记录到具有不同电生理特性的K+in通道电流信号(Xicluna et al., 2007; Lebaudy et al., 2010).而AtKC1作为抑制亚基, 通过重组进入K+in四聚体发挥对K+in通道的抑制作用, 是K+in通道的重要负调控因子.K+in四聚体组成形式的变化可能通过影响K+in通道蛋白离子通道孔的结构而影响其离子通道活性.SNARE蛋白SYP121不仅可以影响KAT1等K+in通道的亚细胞定位, 而且可使KAT1内化离开细胞质膜, 从而抑制全细胞K+in通道的电流大小(Sutter et al., 2006, 2007).进一步研究表明, SNARE还可以通过与AtKC1互作调控K+in通道的蛋白活性(Honsbein et al., 2009; Grefen et al., 2010; Jeanguenin et al., 2011).由此可知, SNARE作为K+in通道的上游调控因子可通过对K+in四聚体组成形式的影响发挥调控作用.蛋白激酶和磷酸酶参与调控多种蛋白的功能, 包括内向和外向质膜K+通道, 比如CPK (calcium-dependent protein kinase)家族的CPK33对GORK的激活作用和PP2CA对GORK的抑制作用(Lefoulon et al., 2016; Corratgé-Faillie et al., 2017).K+in通道可以被多种蛋白激酶磷酸化所抑制, 包括CPK家族和SnRK (Rnf1-related protein kinase)家族的多个蛋白激酶(Sato et al., 2009, 2010; Ronzier et al., 2014).此外, AKT1等多个质膜K+in通道的活性还受Ca2+信号及其与CBL-CIPK组成的信号链的正调控(Li et al., 2006; Cheong et al., 2007; Zhang et al., 2010; Ren et al., 2013; Behera et al., 2017).而PP2CA对AKT1的去磷酸化可以与Ca2+-CBL-CIPK信号链协同调控AKT1的活性(Lan et al., 2011).有趣的是, ABA甚至可以直接激活GORK的离子通道活性(Ooi et al., 2017).K+通道的调控因子众多, 这些调控因子并非各自单独发挥作用, 而是彼此协同发挥作用.AtKC1和Ca2+-CBL-CIPK23信号链协同调控AKT1就是一个典型的例子(Wang et al., 2016). ...
Plant adaptation to fluctuating environment and biomass production are strongly dependent on guard cell potassium channels 3 2008
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
... ).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
... ; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
K + channel activity in plants: genes, regulations and functions 2 2007
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
... ).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
A protein kinase-phosphatase pair interacts with an ion channel to regulate ABA signaling in plant guard cells 2 2009
... SLAC1家族是较晚被发现的一个植物细胞质膜阴离子通道家族, 但植物进化分析表明, SLAC1基因从低等到高等植物均很保守(Lind et al., 2015).拟南芥SLAC1家族由5个成员组成, 包括SLAC1及其同源蛋白SLAH1、2、3和4.其中, SLAC1主要定位在气孔保卫细胞质膜上, 通过介导Cl-和NO3-的外流驱动气孔关闭, 是干旱胁迫及ABA、CO2、臭氧和活性氧(reactive oxygen species, ROS)等信号传递途径的关键组分(Negi et al., 2008; Vahisalu et al., 2008).SLAH3在气孔保卫细胞中分布, 与SLAC1一起通过介导NO3-和Cl-外流驱动气孔关闭(Geiger et al., 2011), 但SLAH3在气孔关闭运动中的作用, 相对于SLAC1而言是辅助性的.拟南芥4个SLAH主要分布在根、茎和叶脉等输导组织中(Negi et al., 2008).其中, SLAH1通过调控SLAH3的活性调控Cl-从根向茎的运输(Cubero-Font et al., 2016).在离子选择性和通透性方面, 拟南芥SLAC1和SLAH3均对Cl-和NO3-有明显的通透能力, 但对NO3-的通透能力明显大于Cl- (Geiger et al., 2009; Lee et al., 2009; Brandt et al., 2012).而SLAH2则为NO3-选择性的阴离子通道, 对Cl-的通透能力微弱(Maierhofer et al., 2014).最近的研究表明, 来自单子叶植物玉米和水稻的SLAC1, 相对于Cl-而言, 对NO3-的选择性明显大于拟南芥SLAC1 (Sun et al., 2016; Qi et al., 2018); 而鸭跖草SLAC1则为NO3-激活的Cl-通透性的阴离子通道(Müller et al., 2017).因此, 来自不同高等植物的SLAC1, 其离子选择性不完全一致, 即不同的高等植物可能采用不同的阴离子通道组合来驱动气孔关闭运动.需要注意的是, 尽管在几十年前就发现苹果酸在气孔保卫细胞内大量积累, 并曾被认为是气孔关闭运动中主要的外流阴离子物质.但实际上, 在气孔关闭运动中发挥核心作用的SLAC1和SLAH3均对苹果酸没有明显的通透能力, 真正通过介导苹果酸外流而参与驱动气孔关闭运动的是来自ALMT家族的ALMT12.因此, 气孔关闭运动应该是由SLAC1、SLAH3和ALMT12等多种质膜阴离子协同作用的结果.每个SLAC1亚基具有10个跨膜区, 其C端和N端均位于质膜的胞质一侧(Vahisalu et al., 2008) (图6A).3个SLAC1亚基互相结合, 以三聚体的形式形成质膜阴离子通道, 但每个亚基独立形成1个允许阴离子通过的孔(Chen et al., 2010) (图6B).由此可见, 包括多个SLAHs在内, SLAC1家族形成的阴离子通道可能均为三聚体, 而SLAH1对SLAH3活性的调控作用, 有可能通过与SLAH3组成异源三聚体而实现.SLAC1家族的离子通道亚基组成形式与CNGC和Shaker家族不同, 但它们在不同亚基组成影响离子通道活性和选择性方面又具有类似之处.SLAC1家族离子通道的激活主要依赖蛋白激酶的磷酸化, 而失活则依赖于磷酸酶的去磷酸化(Geiger et al., 2009, 2011; Lee et al., 2009; Brandt et al., 2012; Maierhofer et al., 2014; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
... ; Lee et al., 2009; Brandt et al., 2012; Maierhofer et al., 2014; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
The Arabidopsis AtPP2CA protein phosphatase inhibits the GORK K + efflux channel and exerts a dominant suppres- sive effect on phosphomimetic-activating mutations 1 2016
... Shaker家族离子通道的活性受多种调控因子的影响.首先, 细胞内和细胞外K+浓度对K+in和K+out通道活性均有明显影响.这种影响可能是基于K+从高浓度向低浓度方向流动的基本化学特性.酸性环境可以显著抑制拟南芥花粉原生质体质膜K+in通道活性, 而碱性条件则具有一定的激活作用(Fan et al., 2001, 2003).这种作用可能通过影响K+in通道的某些氨基酸残基电离状态和离子通道蛋白结构变化来实现.其次, K+in四聚体的亚基组成形式本身就是一种通道活性调控方式.在非洲爪蟾卵母细胞中, 以不同的排列组合共表达2个不同的拟南芥Shaker家族K+in通道基因, 可以利用电生理技术记录到具有不同电生理特性的K+in通道电流信号(Xicluna et al., 2007; Lebaudy et al., 2010).而AtKC1作为抑制亚基, 通过重组进入K+in四聚体发挥对K+in通道的抑制作用, 是K+in通道的重要负调控因子.K+in四聚体组成形式的变化可能通过影响K+in通道蛋白离子通道孔的结构而影响其离子通道活性.SNARE蛋白SYP121不仅可以影响KAT1等K+in通道的亚细胞定位, 而且可使KAT1内化离开细胞质膜, 从而抑制全细胞K+in通道的电流大小(Sutter et al., 2006, 2007).进一步研究表明, SNARE还可以通过与AtKC1互作调控K+in通道的蛋白活性(Honsbein et al., 2009; Grefen et al., 2010; Jeanguenin et al., 2011).由此可知, SNARE作为K+in通道的上游调控因子可通过对K+in四聚体组成形式的影响发挥调控作用.蛋白激酶和磷酸酶参与调控多种蛋白的功能, 包括内向和外向质膜K+通道, 比如CPK (calcium-dependent protein kinase)家族的CPK33对GORK的激活作用和PP2CA对GORK的抑制作用(Lefoulon et al., 2016; Corratgé-Faillie et al., 2017).K+in通道可以被多种蛋白激酶磷酸化所抑制, 包括CPK家族和SnRK (Rnf1-related protein kinase)家族的多个蛋白激酶(Sato et al., 2009, 2010; Ronzier et al., 2014).此外, AKT1等多个质膜K+in通道的活性还受Ca2+信号及其与CBL-CIPK组成的信号链的正调控(Li et al., 2006; Cheong et al., 2007; Zhang et al., 2010; Ren et al., 2013; Behera et al., 2017).而PP2CA对AKT1的去磷酸化可以与Ca2+-CBL-CIPK信号链协同调控AKT1的活性(Lan et al., 2011).有趣的是, ABA甚至可以直接激活GORK的离子通道活性(Ooi et al., 2017).K+通道的调控因子众多, 这些调控因子并非各自单独发挥作用, 而是彼此协同发挥作用.AtKC1和Ca2+-CBL-CIPK23信号链协同调控AKT1就是一个典型的例子(Wang et al., 2016). ...
Electrophysiological analysis of cloned cyclic nucleotide-gated ion channels 1 2002
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
Cloning and first functional characterization of a plant cyclic nucleotide-gated cation channel 1 1999
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
The Os-AKT1 channel is critical for K + uptake in rice roots and is modulated by the rice CBL1-CIPK23 complex 1 2014
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
A Ca 2+ signaling pathway regulates a K + channel for low-K response in Arabidopsis 1 2006
... Shaker家族离子通道的活性受多种调控因子的影响.首先, 细胞内和细胞外K+浓度对K+in和K+out通道活性均有明显影响.这种影响可能是基于K+从高浓度向低浓度方向流动的基本化学特性.酸性环境可以显著抑制拟南芥花粉原生质体质膜K+in通道活性, 而碱性条件则具有一定的激活作用(Fan et al., 2001, 2003).这种作用可能通过影响K+in通道的某些氨基酸残基电离状态和离子通道蛋白结构变化来实现.其次, K+in四聚体的亚基组成形式本身就是一种通道活性调控方式.在非洲爪蟾卵母细胞中, 以不同的排列组合共表达2个不同的拟南芥Shaker家族K+in通道基因, 可以利用电生理技术记录到具有不同电生理特性的K+in通道电流信号(Xicluna et al., 2007; Lebaudy et al., 2010).而AtKC1作为抑制亚基, 通过重组进入K+in四聚体发挥对K+in通道的抑制作用, 是K+in通道的重要负调控因子.K+in四聚体组成形式的变化可能通过影响K+in通道蛋白离子通道孔的结构而影响其离子通道活性.SNARE蛋白SYP121不仅可以影响KAT1等K+in通道的亚细胞定位, 而且可使KAT1内化离开细胞质膜, 从而抑制全细胞K+in通道的电流大小(Sutter et al., 2006, 2007).进一步研究表明, SNARE还可以通过与AtKC1互作调控K+in通道的蛋白活性(Honsbein et al., 2009; Grefen et al., 2010; Jeanguenin et al., 2011).由此可知, SNARE作为K+in通道的上游调控因子可通过对K+in四聚体组成形式的影响发挥调控作用.蛋白激酶和磷酸酶参与调控多种蛋白的功能, 包括内向和外向质膜K+通道, 比如CPK (calcium-dependent protein kinase)家族的CPK33对GORK的激活作用和PP2CA对GORK的抑制作用(Lefoulon et al., 2016; Corratgé-Faillie et al., 2017).K+in通道可以被多种蛋白激酶磷酸化所抑制, 包括CPK家族和SnRK (Rnf1-related protein kinase)家族的多个蛋白激酶(Sato et al., 2009, 2010; Ronzier et al., 2014).此外, AKT1等多个质膜K+in通道的活性还受Ca2+信号及其与CBL-CIPK组成的信号链的正调控(Li et al., 2006; Cheong et al., 2007; Zhang et al., 2010; Ren et al., 2013; Behera et al., 2017).而PP2CA对AKT1的去磷酸化可以与Ca2+-CBL-CIPK信号链协同调控AKT1的活性(Lan et al., 2011).有趣的是, ABA甚至可以直接激活GORK的离子通道活性(Ooi et al., 2017).K+通道的调控因子众多, 这些调控因子并非各自单独发挥作用, 而是彼此协同发挥作用.AtKC1和Ca2+-CBL-CIPK23信号链协同调控AKT1就是一个典型的例子(Wang et al., 2016). ...
Low pH, aluminum, and phosphorus coordinately regulate malate exudation through GmALMT1 to improve soybean adap- tation to acid soils 2 2013
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
... ; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
The BnALMT1 and BnALMT2 genes from rape encode aluminum-activated malate transporters that enhance the aluminum resistance of plant cells 1 2006
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
Maize ZmALMT2 is a root anion transporter that mediates constitutive root malate efflux 1 2012
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
Stomatal guard cells co-opted an ancient ABA-dependent desiccation survival system to regulate stomatal closure 1 2015
... SLAC1家族是较晚被发现的一个植物细胞质膜阴离子通道家族, 但植物进化分析表明, SLAC1基因从低等到高等植物均很保守(Lind et al., 2015).拟南芥SLAC1家族由5个成员组成, 包括SLAC1及其同源蛋白SLAH1、2、3和4.其中, SLAC1主要定位在气孔保卫细胞质膜上, 通过介导Cl-和NO3-的外流驱动气孔关闭, 是干旱胁迫及ABA、CO2、臭氧和活性氧(reactive oxygen species, ROS)等信号传递途径的关键组分(Negi et al., 2008; Vahisalu et al., 2008).SLAH3在气孔保卫细胞中分布, 与SLAC1一起通过介导NO3-和Cl-外流驱动气孔关闭(Geiger et al., 2011), 但SLAH3在气孔关闭运动中的作用, 相对于SLAC1而言是辅助性的.拟南芥4个SLAH主要分布在根、茎和叶脉等输导组织中(Negi et al., 2008).其中, SLAH1通过调控SLAH3的活性调控Cl-从根向茎的运输(Cubero-Font et al., 2016).在离子选择性和通透性方面, 拟南芥SLAC1和SLAH3均对Cl-和NO3-有明显的通透能力, 但对NO3-的通透能力明显大于Cl- (Geiger et al., 2009; Lee et al., 2009; Brandt et al., 2012).而SLAH2则为NO3-选择性的阴离子通道, 对Cl-的通透能力微弱(Maierhofer et al., 2014).最近的研究表明, 来自单子叶植物玉米和水稻的SLAC1, 相对于Cl-而言, 对NO3-的选择性明显大于拟南芥SLAC1 (Sun et al., 2016; Qi et al., 2018); 而鸭跖草SLAC1则为NO3-激活的Cl-通透性的阴离子通道(Müller et al., 2017).因此, 来自不同高等植物的SLAC1, 其离子选择性不完全一致, 即不同的高等植物可能采用不同的阴离子通道组合来驱动气孔关闭运动.需要注意的是, 尽管在几十年前就发现苹果酸在气孔保卫细胞内大量积累, 并曾被认为是气孔关闭运动中主要的外流阴离子物质.但实际上, 在气孔关闭运动中发挥核心作用的SLAC1和SLAH3均对苹果酸没有明显的通透能力, 真正通过介导苹果酸外流而参与驱动气孔关闭运动的是来自ALMT家族的ALMT12.因此, 气孔关闭运动应该是由SLAC1、SLAH3和ALMT12等多种质膜阴离子协同作用的结果.每个SLAC1亚基具有10个跨膜区, 其C端和N端均位于质膜的胞质一侧(Vahisalu et al., 2008) (图6A).3个SLAC1亚基互相结合, 以三聚体的形式形成质膜阴离子通道, 但每个亚基独立形成1个允许阴离子通过的孔(Chen et al., 2010) (图6B).由此可见, 包括多个SLAHs在内, SLAC1家族形成的阴离子通道可能均为三聚体, 而SLAH1对SLAH3活性的调控作用, 有可能通过与SLAH3组成异源三聚体而实现.SLAC1家族的离子通道亚基组成形式与CNGC和Shaker家族不同, 但它们在不同亚基组成影响离子通道活性和选择性方面又具有类似之处.SLAC1家族离子通道的激活主要依赖蛋白激酶的磷酸化, 而失活则依赖于磷酸酶的去磷酸化(Geiger et al., 2009, 2011; Lee et al., 2009; Brandt et al., 2012; Maierhofer et al., 2014; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
Intracellular K + sensing of SKOR, a Shaker-type K + channel from Arabidopsis 2 2006
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
... ; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
Application of T-DNA activation tagging to identify glutamate receptor-like genes that enhance drought tolerance in plants 1 2014
... GLR家族是受多种氨基酸门控的离子通道, 但其通道门控特点并非如其名称仅为谷氨酸所门控, 而是受多种氨基酸影响.1998年, 第1个GLR被克隆出来(Lam et al., 1998), 人们开始对GLR进行研究.研究表明, 拟南芥GLR1和GLR2的N端位于细胞质膜外侧, C端位于细胞质膜内侧, 中间包含4个跨膜区, 但第2个跨膜区并不能完整地跨过细胞质膜(Lam et al., 1998) (图2).这种结构特点明显不同于Shaker和CNGC家族.GLR组成的质膜离子通道的亚基组成、多数GLR离子通道的离子选择性等基本特性, 以及GLR离子通道的活性调控机理目前还不甚清楚.但已经解析出动物GLR四聚体的通道蛋白结构(Price et al., 2012).据此推测, 植物GLR组成的通道蛋白结构可能与动物类似.GLR在植物体内具有多种生物学功能, 包括参与调控花粉管的极性生长(Michard et al., 2011; Wudick et al., 2018)、铝诱导的微管解聚和质膜去极化(Sivaguru et al., 2003)、离子运输(McAinsh and Pittman, 2009)、碳氮代谢(Kang and Turano, 2003)、ABA合成和耐旱反应(Kang et al., 2004; Lu et al., 2014)以及机械损伤信号的长距离传输(Mousavi et al., 2013)等.总体而言, 目前对GLR家族的研究还不够深入.关于植物GLR家族更为详细的介绍可以参阅何明杰等(2016)的综述. ...
AtCNGC2 is involved in jasmonic acid-induced calcium mobilization 1 2016
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
Leaf senescence signaling: the Ca 2+-conducting Arabidopsis cyclic nucleotide gated chan- nel 2 acts through nitric oxide to repress senescence pro- gramming 1 2010
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
Determination of the subunit stoi- chiometry of a voltage-activated potassium channel 1 1991
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
Potassium content and aperture in “intact” stomatal and epidermal cells of Com- melina communis L 1 1980
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
A single-pore residue renders the Arabidopsis root anion channel SLAH2 highly nitrate selective 2 2014
... SLAC1家族是较晚被发现的一个植物细胞质膜阴离子通道家族, 但植物进化分析表明, SLAC1基因从低等到高等植物均很保守(Lind et al., 2015).拟南芥SLAC1家族由5个成员组成, 包括SLAC1及其同源蛋白SLAH1、2、3和4.其中, SLAC1主要定位在气孔保卫细胞质膜上, 通过介导Cl-和NO3-的外流驱动气孔关闭, 是干旱胁迫及ABA、CO2、臭氧和活性氧(reactive oxygen species, ROS)等信号传递途径的关键组分(Negi et al., 2008; Vahisalu et al., 2008).SLAH3在气孔保卫细胞中分布, 与SLAC1一起通过介导NO3-和Cl-外流驱动气孔关闭(Geiger et al., 2011), 但SLAH3在气孔关闭运动中的作用, 相对于SLAC1而言是辅助性的.拟南芥4个SLAH主要分布在根、茎和叶脉等输导组织中(Negi et al., 2008).其中, SLAH1通过调控SLAH3的活性调控Cl-从根向茎的运输(Cubero-Font et al., 2016).在离子选择性和通透性方面, 拟南芥SLAC1和SLAH3均对Cl-和NO3-有明显的通透能力, 但对NO3-的通透能力明显大于Cl- (Geiger et al., 2009; Lee et al., 2009; Brandt et al., 2012).而SLAH2则为NO3-选择性的阴离子通道, 对Cl-的通透能力微弱(Maierhofer et al., 2014).最近的研究表明, 来自单子叶植物玉米和水稻的SLAC1, 相对于Cl-而言, 对NO3-的选择性明显大于拟南芥SLAC1 (Sun et al., 2016; Qi et al., 2018); 而鸭跖草SLAC1则为NO3-激活的Cl-通透性的阴离子通道(Müller et al., 2017).因此, 来自不同高等植物的SLAC1, 其离子选择性不完全一致, 即不同的高等植物可能采用不同的阴离子通道组合来驱动气孔关闭运动.需要注意的是, 尽管在几十年前就发现苹果酸在气孔保卫细胞内大量积累, 并曾被认为是气孔关闭运动中主要的外流阴离子物质.但实际上, 在气孔关闭运动中发挥核心作用的SLAC1和SLAH3均对苹果酸没有明显的通透能力, 真正通过介导苹果酸外流而参与驱动气孔关闭运动的是来自ALMT家族的ALMT12.因此, 气孔关闭运动应该是由SLAC1、SLAH3和ALMT12等多种质膜阴离子协同作用的结果.每个SLAC1亚基具有10个跨膜区, 其C端和N端均位于质膜的胞质一侧(Vahisalu et al., 2008) (图6A).3个SLAC1亚基互相结合, 以三聚体的形式形成质膜阴离子通道, 但每个亚基独立形成1个允许阴离子通过的孔(Chen et al., 2010) (图6B).由此可见, 包括多个SLAHs在内, SLAC1家族形成的阴离子通道可能均为三聚体, 而SLAH1对SLAH3活性的调控作用, 有可能通过与SLAH3组成异源三聚体而实现.SLAC1家族的离子通道亚基组成形式与CNGC和Shaker家族不同, 但它们在不同亚基组成影响离子通道活性和选择性方面又具有类似之处.SLAC1家族离子通道的激活主要依赖蛋白激酶的磷酸化, 而失活则依赖于磷酸酶的去磷酸化(Geiger et al., 2009, 2011; Lee et al., 2009; Brandt et al., 2012; Maierhofer et al., 2014; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
... ; Maierhofer et al., 2014; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
MscS-Like10 is a stretch- activated ion channel from Arabidopsis thaliana with a preference for anions 1 2012
... 除了SLAC1家族, 植物细胞质膜上还包含张力激活的阴离子通道MSL (mechano-sensitive or stret- ch-activated channels) (Crosgrove and Hedrich, 1991).MSL为典型的张力激活的阴离子通道家族.MSL9和MSL10是最早得到电生理鉴定的MSL家族成员, 定位于细胞质膜, 其激活对质膜张力具有依赖性, 主要选择性介导阴离子的跨膜流动(Haswell et al., 2008; Maksaev and Haswell, 2012).随后有研究表明, 拟南芥MSL10参与植物机械损伤信号反应和茉莉酸的累积(Veley et al., 2014; Zou et al., 2016).此外, 有些拟南芥MSL成员定位在细胞器膜上, 调控细胞器的大小和形状(Veley et al., 2012).根据氨基酸残基序列预测, MSL蛋白存在多个跨膜区, 具备形成离子通道的基本结构特点.而质膜受到拉伸时对MSL的激活是通过机械拉力直接改变MSL蛋白的结构, 使离子通过的孔被拉开.但这基本上还停留在理论推测阶段, 缺乏有力的直接实验证据支持.总体来讲, 目前对MSL家族的生物学功能还了解较少, 其组成的离子通道蛋白结构、感知生物膜张力的分子机理和生物学功能等还有待深入研究. ...
Phylogenetic relationships within cation transporter families of Arabidopsis 1 2001
... 植物体内存在大量的钙元素.一方面钙作为细胞壁等结构组分发挥作用; 另一方面, 离子形式的钙(Ca2+)可以作为信号分子参与各种生物学反应和信号传递过程, 后者则备受关注.胞内Ca2+信号以细胞质Ca2+浓度的升高和震荡形式出现.一般认为, 胞外Ca2+内流是细胞质Ca2+信号产生所需Ca2+的主要来源, 而质膜内向Ca2+通道是胞外Ca2+内流的主要渠道.同时, Ca2+通道的活性变化通过控制胞外Ca2+内流的节奏直接调控胞内Ca2+震荡, 发挥Ca2+信号上游调控因子的作用.鉴于此, 质膜Ca2+通道受到广泛关注, 并成为动植物各信号领域的重点研究对象.科学家首先在动物细胞中发现多个Ca2+通道家族, 主要包括受环核苷酸cAMP和cGMP激活的CNG (cyclic nucleotide-gated)家族、受谷氨酸门控的GLR (glutamate receptor)家族、TRPC (transient receptor potential channel)和受胞内Ca2+库倒空激活的CRACM1/Orai1 (Zagotta and Siegelbaum, 1996; Vig and Kinet, 2007; Sobolevsky, 2015; Ambudkar et al., 2017).通过序列比对, 多个动物细胞质膜Ca2+通道的同源基因家族先后在植物尤其是模式植物拟南芥基因组中被鉴定出来, 主要包括CNGC (cyclic nucleotide- gated channel)、GLR (glutamate receptor-like)、高渗敏感的OSCA (hyperosmolality-gated calcium- permeable channel)家族和低渗敏感的张力激活离子通道MCA (mechano-sensitive channel) (M?ser et al., 2001; Ward et al., 2009; Iida et al., 2014; Yuan et al., 2014). ...
Cyclic nucleotide-gated ion channels 1 2003
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
Shaping the calcium signature 1 2009
... GLR家族是受多种氨基酸门控的离子通道, 但其通道门控特点并非如其名称仅为谷氨酸所门控, 而是受多种氨基酸影响.1998年, 第1个GLR被克隆出来(Lam et al., 1998), 人们开始对GLR进行研究.研究表明, 拟南芥GLR1和GLR2的N端位于细胞质膜外侧, C端位于细胞质膜内侧, 中间包含4个跨膜区, 但第2个跨膜区并不能完整地跨过细胞质膜(Lam et al., 1998) (图2).这种结构特点明显不同于Shaker和CNGC家族.GLR组成的质膜离子通道的亚基组成、多数GLR离子通道的离子选择性等基本特性, 以及GLR离子通道的活性调控机理目前还不甚清楚.但已经解析出动物GLR四聚体的通道蛋白结构(Price et al., 2012).据此推测, 植物GLR组成的通道蛋白结构可能与动物类似.GLR在植物体内具有多种生物学功能, 包括参与调控花粉管的极性生长(Michard et al., 2011; Wudick et al., 2018)、铝诱导的微管解聚和质膜去极化(Sivaguru et al., 2003)、离子运输(McAinsh and Pittman, 2009)、碳氮代谢(Kang and Turano, 2003)、ABA合成和耐旱反应(Kang et al., 2004; Lu et al., 2014)以及机械损伤信号的长距离传输(Mousavi et al., 2013)等.总体而言, 目前对GLR家族的研究还不够深入.关于植物GLR家族更为详细的介绍可以参阅何明杰等(2016)的综述. ...
AtALMT12 represents an R-type anion channel required for stomatal movement in Arabidopsis guard cells 1 2010
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
Malate transport by the vacuolar AtALMT6 channel in guard cells is subject to multiple regulation 1 2011
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
Glutamate receptor-like genes form Ca 2+ channels in pollen tubes and are regulated by pistil D-serine 1 2011
... GLR家族是受多种氨基酸门控的离子通道, 但其通道门控特点并非如其名称仅为谷氨酸所门控, 而是受多种氨基酸影响.1998年, 第1个GLR被克隆出来(Lam et al., 1998), 人们开始对GLR进行研究.研究表明, 拟南芥GLR1和GLR2的N端位于细胞质膜外侧, C端位于细胞质膜内侧, 中间包含4个跨膜区, 但第2个跨膜区并不能完整地跨过细胞质膜(Lam et al., 1998) (图2).这种结构特点明显不同于Shaker和CNGC家族.GLR组成的质膜离子通道的亚基组成、多数GLR离子通道的离子选择性等基本特性, 以及GLR离子通道的活性调控机理目前还不甚清楚.但已经解析出动物GLR四聚体的通道蛋白结构(Price et al., 2012).据此推测, 植物GLR组成的通道蛋白结构可能与动物类似.GLR在植物体内具有多种生物学功能, 包括参与调控花粉管的极性生长(Michard et al., 2011; Wudick et al., 2018)、铝诱导的微管解聚和质膜去极化(Sivaguru et al., 2003)、离子运输(McAinsh and Pittman, 2009)、碳氮代谢(Kang and Turano, 2003)、ABA合成和耐旱反应(Kang et al., 2004; Lu et al., 2014)以及机械损伤信号的长距离传输(Mousavi et al., 2013)等.总体而言, 目前对GLR家族的研究还不够深入.关于植物GLR家族更为详细的介绍可以参阅何明杰等(2016)的综述. ...
A dominant negative OsKAT2 mutant delays light-induced stomatal opening and improves drought tolerance without yield penalty in rice 1 2017
... 目前, 对拟南芥Shaker家族的组织分布和功能研究得较为深入全面; 而对包括水稻(Oryza sativa)和玉米(Zea mays)在内的其它植物Shaker家族的研究相对较少.已有的研究鉴定出多个参与K+吸收、转运和气孔运动的玉米和水稻Shaker同源基因.其与拟南芥同源蛋白相比较有很多类似之处, 但也有不同.例如, 玉米KZM2通过与KZM3组成异源多聚体, 抑制K+in通道的活性和气孔运动(Gao et al., 2017).而水稻OsKAT3通过与OsKAT2互作抑制OsKAT3的活性(Hwang et al., 2013).这种K+通道两个同源蛋白之间的互作和调控方式与拟南芥中的AtKC1对AKT1和KAT1等的抑制作用类似(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).再比如, 水稻KAT2与拟南芥KAT2均作为K+in通道参与光照诱导的气孔开放, 但拟南芥气孔保卫细胞质膜K+in通道编码基因数目明显多于水稻(Pilot et al., 2001; Moon et al., 2017).对于外向K+通道而言, 拟南芥SKOR分布于根部, 负责K+向木质部的卸载, 而GORK则主要参与拟南芥的气孔关闭, 二者的组织分布和生物学功能明显不同.而水稻的同一个Shaker蛋白同时参与气孔关闭和K+营养在根部的卸载(Nguyen et al., 2017).因此, 加强对更多植物, 尤其是重要作物中Shaker家族的研究, 将会促进人们对植物吸收、运输和利用K+的分子机理的深入理解. ...
Ca 2+- permeable mechanosensitive channels MCA1 and MCA2 mediate cold-induced cytosolic Ca 2+ increase and cold tolerance in Arabidopsis 1 2018
... 早期的研究中, 人们发现植物细胞中存在一类受质膜张力激活的Ca2+信号, 从而预测在植物细胞质膜上存在张力激活类质膜Ca2+通道(Crosgrove and Hedrich, 1991; Dutta and Robinson, 2004; Qi et al., 2004; Zhang et al., 2007).多年来, 人们一直没有鉴定到此类受机械或渗透张力激活的离子通道编码基因.近年来, 科学家找到了2类对质膜张力敏感的离子通道, 其一是受低渗条件激活, 通过介导胞外Ca2+内流发挥功能的MCA1和MCA2 (Iida et al., 2014); 其二是对高渗胁迫敏感的OSCA家族(Hou et al., 2014; Yuan et al., 2014).MCA1和MCA2蛋白只有1个跨膜区, 其N端较短且位于细胞质膜外侧, 而C端较长且位于质膜内侧(图3A).研究表明, MCA1和MCA2的N端和跨膜区为其离子通道组成及活性所必需, 而C端则对其离子通道活性发挥调控作用(Kam- ano et al., 2015).MCA1和MCA2组成的离子通道也为四聚体结构, 张力和低渗等刺激通过拉伸质膜改变该离子通道蛋白的结构, 从而使通道被激活, 引起胞外Ca2+内流和细胞质Ca2+水平升高(图3B).MCA1的拟南芥突变体根无法插入较硬的固体培养基(Nakagawa et al., 2007), 说明其在感知机械压力方面发挥作用.最近的研究表明, MCA1和MCA2还作为质膜Ca2+通道介导拟南芥的耐冷反应(Mori et al., 2018).与MCA1和MCA2相反, OSCA是对高渗敏感的质膜离子通道, 可以被细胞外的高渗处理激活(Yuan et al., 2014).拟南芥OSCA家族的蛋白结构与MCA1和MCA2大不相同, 其每个亚基均具有多达9个跨膜区, 在其第8和第9跨膜区之间具有1个孔形成所需的环状结构(Yuan et al., 2014) (图4).OSCA与MCA1和MCA2类似之处在于其亚基N端位于质膜的胞外侧, 而C端则位于质膜胞质侧(Yuan et al., 2014) (图4).OSCA1允许多种离子通过, 其对各种离子的通透能力大小排序为K+>Ba2+≈Ca2+>Na+=Mg2+=Cs+ (Yuan et al., 2014).目前, 人们对质膜张力激活的质膜离子通道的生物学功能了解较少, 但其对渗透敏感的特点暗示此类质膜离子通道可能通过对质膜张力变化做出反应.对低渗敏感的MCA1和MCA2与对高渗敏感的OSCA家族有可能作为一对质膜张力和渗透敏感元件, 协同配合发挥作用, 以共同应对渗透环境变化.一方面, 其可以通过影响胞内Ca2+信号而参与耐盐、耐旱和气孔运动等与细胞渗透和膨压调节密切相关的生物学过程; 另一方面, OSCA对K+的通透能力也表明其可能同时介导Ca2+和K+的跨膜流动而参与这些生物学过程. ...
Pollen tube development and competitive ability are impaired by disruption of a Shaker K + channel in Arabi- dopsis 1 2002
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
... (A) 单个亚基拓扑结构; (B) Shaker离子通道的多种不同四聚体组成形式, 包括内向和外向质膜K+通道.S1-S6: 单亚基的第1-6个跨膜区; N/C端: 氨/羧基末端; P: 通道孔区; +: 带正电荷的电压感受器结构域; cNBD: 环核苷酸结合域 Topological structure of a representative subunit and tetramers of Shaker family from Arabidopsis (<xref ref-type="bibr" rid="b98">modified from Sharma et al., 2013</xref>) (A) Topological structure of a Shaker subunit; (B) Diverse tetramers of K+ channels composed of Shaker members, including inward and outward K+ channels. S1-S6: Transmembrane domains 1-6; N/C termini: The amino terminal and carboxyl terminal; P: Pore region; +: With positive charged voltage sensor domain; cNBD: Cyclic nucleotide binding domain. ...
... GLR家族是受多种氨基酸门控的离子通道, 但其通道门控特点并非如其名称仅为谷氨酸所门控, 而是受多种氨基酸影响.1998年, 第1个GLR被克隆出来(Lam et al., 1998), 人们开始对GLR进行研究.研究表明, 拟南芥GLR1和GLR2的N端位于细胞质膜外侧, C端位于细胞质膜内侧, 中间包含4个跨膜区, 但第2个跨膜区并不能完整地跨过细胞质膜(Lam et al., 1998) (图2).这种结构特点明显不同于Shaker和CNGC家族.GLR组成的质膜离子通道的亚基组成、多数GLR离子通道的离子选择性等基本特性, 以及GLR离子通道的活性调控机理目前还不甚清楚.但已经解析出动物GLR四聚体的通道蛋白结构(Price et al., 2012).据此推测, 植物GLR组成的通道蛋白结构可能与动物类似.GLR在植物体内具有多种生物学功能, 包括参与调控花粉管的极性生长(Michard et al., 2011; Wudick et al., 2018)、铝诱导的微管解聚和质膜去极化(Sivaguru et al., 2003)、离子运输(McAinsh and Pittman, 2009)、碳氮代谢(Kang and Turano, 2003)、ABA合成和耐旱反应(Kang et al., 2004; Lu et al., 2014)以及机械损伤信号的长距离传输(Mousavi et al., 2013)等.总体而言, 目前对GLR家族的研究还不够深入.关于植物GLR家族更为详细的介绍可以参阅何明杰等(2016)的综述. ...
The desert plant Phoenix dactylifera closes stomata via nitrate-regulated SLAC1 anion channel 1 2017
... SLAC1家族是较晚被发现的一个植物细胞质膜阴离子通道家族, 但植物进化分析表明, SLAC1基因从低等到高等植物均很保守(Lind et al., 2015).拟南芥SLAC1家族由5个成员组成, 包括SLAC1及其同源蛋白SLAH1、2、3和4.其中, SLAC1主要定位在气孔保卫细胞质膜上, 通过介导Cl-和NO3-的外流驱动气孔关闭, 是干旱胁迫及ABA、CO2、臭氧和活性氧(reactive oxygen species, ROS)等信号传递途径的关键组分(Negi et al., 2008; Vahisalu et al., 2008).SLAH3在气孔保卫细胞中分布, 与SLAC1一起通过介导NO3-和Cl-外流驱动气孔关闭(Geiger et al., 2011), 但SLAH3在气孔关闭运动中的作用, 相对于SLAC1而言是辅助性的.拟南芥4个SLAH主要分布在根、茎和叶脉等输导组织中(Negi et al., 2008).其中, SLAH1通过调控SLAH3的活性调控Cl-从根向茎的运输(Cubero-Font et al., 2016).在离子选择性和通透性方面, 拟南芥SLAC1和SLAH3均对Cl-和NO3-有明显的通透能力, 但对NO3-的通透能力明显大于Cl- (Geiger et al., 2009; Lee et al., 2009; Brandt et al., 2012).而SLAH2则为NO3-选择性的阴离子通道, 对Cl-的通透能力微弱(Maierhofer et al., 2014).最近的研究表明, 来自单子叶植物玉米和水稻的SLAC1, 相对于Cl-而言, 对NO3-的选择性明显大于拟南芥SLAC1 (Sun et al., 2016; Qi et al., 2018); 而鸭跖草SLAC1则为NO3-激活的Cl-通透性的阴离子通道(Müller et al., 2017).因此, 来自不同高等植物的SLAC1, 其离子选择性不完全一致, 即不同的高等植物可能采用不同的阴离子通道组合来驱动气孔关闭运动.需要注意的是, 尽管在几十年前就发现苹果酸在气孔保卫细胞内大量积累, 并曾被认为是气孔关闭运动中主要的外流阴离子物质.但实际上, 在气孔关闭运动中发挥核心作用的SLAC1和SLAH3均对苹果酸没有明显的通透能力, 真正通过介导苹果酸外流而参与驱动气孔关闭运动的是来自ALMT家族的ALMT12.因此, 气孔关闭运动应该是由SLAC1、SLAH3和ALMT12等多种质膜阴离子协同作用的结果.每个SLAC1亚基具有10个跨膜区, 其C端和N端均位于质膜的胞质一侧(Vahisalu et al., 2008) (图6A).3个SLAC1亚基互相结合, 以三聚体的形式形成质膜阴离子通道, 但每个亚基独立形成1个允许阴离子通过的孔(Chen et al., 2010) (图6B).由此可见, 包括多个SLAHs在内, SLAC1家族形成的阴离子通道可能均为三聚体, 而SLAH1对SLAH3活性的调控作用, 有可能通过与SLAH3组成异源三聚体而实现.SLAC1家族的离子通道亚基组成形式与CNGC和Shaker家族不同, 但它们在不同亚基组成影响离子通道活性和选择性方面又具有类似之处.SLAC1家族离子通道的激活主要依赖蛋白激酶的磷酸化, 而失活则依赖于磷酸酶的去磷酸化(Geiger et al., 2009, 2011; Lee et al., 2009; Brandt et al., 2012; Maierhofer et al., 2014; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
Arabidopsis plasma mem- brane protein crucial for Ca 2+ influx and touch sensing in roots 1 2007
... 早期的研究中, 人们发现植物细胞中存在一类受质膜张力激活的Ca2+信号, 从而预测在植物细胞质膜上存在张力激活类质膜Ca2+通道(Crosgrove and Hedrich, 1991; Dutta and Robinson, 2004; Qi et al., 2004; Zhang et al., 2007).多年来, 人们一直没有鉴定到此类受机械或渗透张力激活的离子通道编码基因.近年来, 科学家找到了2类对质膜张力敏感的离子通道, 其一是受低渗条件激活, 通过介导胞外Ca2+内流发挥功能的MCA1和MCA2 (Iida et al., 2014); 其二是对高渗胁迫敏感的OSCA家族(Hou et al., 2014; Yuan et al., 2014).MCA1和MCA2蛋白只有1个跨膜区, 其N端较短且位于细胞质膜外侧, 而C端较长且位于质膜内侧(图3A).研究表明, MCA1和MCA2的N端和跨膜区为其离子通道组成及活性所必需, 而C端则对其离子通道活性发挥调控作用(Kam- ano et al., 2015).MCA1和MCA2组成的离子通道也为四聚体结构, 张力和低渗等刺激通过拉伸质膜改变该离子通道蛋白的结构, 从而使通道被激活, 引起胞外Ca2+内流和细胞质Ca2+水平升高(图3B).MCA1的拟南芥突变体根无法插入较硬的固体培养基(Nakagawa et al., 2007), 说明其在感知机械压力方面发挥作用.最近的研究表明, MCA1和MCA2还作为质膜Ca2+通道介导拟南芥的耐冷反应(Mori et al., 2018).与MCA1和MCA2相反, OSCA是对高渗敏感的质膜离子通道, 可以被细胞外的高渗处理激活(Yuan et al., 2014).拟南芥OSCA家族的蛋白结构与MCA1和MCA2大不相同, 其每个亚基均具有多达9个跨膜区, 在其第8和第9跨膜区之间具有1个孔形成所需的环状结构(Yuan et al., 2014) (图4).OSCA与MCA1和MCA2类似之处在于其亚基N端位于质膜的胞外侧, 而C端则位于质膜胞质侧(Yuan et al., 2014) (图4).OSCA1允许多种离子通过, 其对各种离子的通透能力大小排序为K+>Ba2+≈Ca2+>Na+=Mg2+=Cs+ (Yuan et al., 2014).目前, 人们对质膜张力激活的质膜离子通道的生物学功能了解较少, 但其对渗透敏感的特点暗示此类质膜离子通道可能通过对质膜张力变化做出反应.对低渗敏感的MCA1和MCA2与对高渗敏感的OSCA家族有可能作为一对质膜张力和渗透敏感元件, 协同配合发挥作用, 以共同应对渗透环境变化.一方面, 其可以通过影响胞内Ca2+信号而参与耐盐、耐旱和气孔运动等与细胞渗透和膨压调节密切相关的生物学过程; 另一方面, OSCA对K+的通透能力也表明其可能同时介导Ca2+和K+的跨膜流动而参与这些生物学过程. ...
CO2 regulator SLAC1 and its homologues are essential for anion homeostasis in plant cells 2 2008
... SLAC1家族是较晚被发现的一个植物细胞质膜阴离子通道家族, 但植物进化分析表明, SLAC1基因从低等到高等植物均很保守(Lind et al., 2015).拟南芥SLAC1家族由5个成员组成, 包括SLAC1及其同源蛋白SLAH1、2、3和4.其中, SLAC1主要定位在气孔保卫细胞质膜上, 通过介导Cl-和NO3-的外流驱动气孔关闭, 是干旱胁迫及ABA、CO2、臭氧和活性氧(reactive oxygen species, ROS)等信号传递途径的关键组分(Negi et al., 2008; Vahisalu et al., 2008).SLAH3在气孔保卫细胞中分布, 与SLAC1一起通过介导NO3-和Cl-外流驱动气孔关闭(Geiger et al., 2011), 但SLAH3在气孔关闭运动中的作用, 相对于SLAC1而言是辅助性的.拟南芥4个SLAH主要分布在根、茎和叶脉等输导组织中(Negi et al., 2008).其中, SLAH1通过调控SLAH3的活性调控Cl-从根向茎的运输(Cubero-Font et al., 2016).在离子选择性和通透性方面, 拟南芥SLAC1和SLAH3均对Cl-和NO3-有明显的通透能力, 但对NO3-的通透能力明显大于Cl- (Geiger et al., 2009; Lee et al., 2009; Brandt et al., 2012).而SLAH2则为NO3-选择性的阴离子通道, 对Cl-的通透能力微弱(Maierhofer et al., 2014).最近的研究表明, 来自单子叶植物玉米和水稻的SLAC1, 相对于Cl-而言, 对NO3-的选择性明显大于拟南芥SLAC1 (Sun et al., 2016; Qi et al., 2018); 而鸭跖草SLAC1则为NO3-激活的Cl-通透性的阴离子通道(Müller et al., 2017).因此, 来自不同高等植物的SLAC1, 其离子选择性不完全一致, 即不同的高等植物可能采用不同的阴离子通道组合来驱动气孔关闭运动.需要注意的是, 尽管在几十年前就发现苹果酸在气孔保卫细胞内大量积累, 并曾被认为是气孔关闭运动中主要的外流阴离子物质.但实际上, 在气孔关闭运动中发挥核心作用的SLAC1和SLAH3均对苹果酸没有明显的通透能力, 真正通过介导苹果酸外流而参与驱动气孔关闭运动的是来自ALMT家族的ALMT12.因此, 气孔关闭运动应该是由SLAC1、SLAH3和ALMT12等多种质膜阴离子协同作用的结果.每个SLAC1亚基具有10个跨膜区, 其C端和N端均位于质膜的胞质一侧(Vahisalu et al., 2008) (图6A).3个SLAC1亚基互相结合, 以三聚体的形式形成质膜阴离子通道, 但每个亚基独立形成1个允许阴离子通过的孔(Chen et al., 2010) (图6B).由此可见, 包括多个SLAHs在内, SLAC1家族形成的阴离子通道可能均为三聚体, 而SLAH1对SLAH3活性的调控作用, 有可能通过与SLAH3组成异源三聚体而实现.SLAC1家族的离子通道亚基组成形式与CNGC和Shaker家族不同, 但它们在不同亚基组成影响离子通道活性和选择性方面又具有类似之处.SLAC1家族离子通道的激活主要依赖蛋白激酶的磷酸化, 而失活则依赖于磷酸酶的去磷酸化(Geiger et al., 2009, 2011; Lee et al., 2009; Brandt et al., 2012; Maierhofer et al., 2014; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
... ), 但SLAH3在气孔关闭运动中的作用, 相对于SLAC1而言是辅助性的.拟南芥4个SLAH主要分布在根、茎和叶脉等输导组织中(Negi et al., 2008).其中, SLAH1通过调控SLAH3的活性调控Cl-从根向茎的运输(Cubero-Font et al., 2016).在离子选择性和通透性方面, 拟南芥SLAC1和SLAH3均对Cl-和NO3-有明显的通透能力, 但对NO3-的通透能力明显大于Cl- (Geiger et al., 2009; Lee et al., 2009; Brandt et al., 2012).而SLAH2则为NO3-选择性的阴离子通道, 对Cl-的通透能力微弱(Maierhofer et al., 2014).最近的研究表明, 来自单子叶植物玉米和水稻的SLAC1, 相对于Cl-而言, 对NO3-的选择性明显大于拟南芥SLAC1 (Sun et al., 2016; Qi et al., 2018); 而鸭跖草SLAC1则为NO3-激活的Cl-通透性的阴离子通道(Müller et al., 2017).因此, 来自不同高等植物的SLAC1, 其离子选择性不完全一致, 即不同的高等植物可能采用不同的阴离子通道组合来驱动气孔关闭运动.需要注意的是, 尽管在几十年前就发现苹果酸在气孔保卫细胞内大量积累, 并曾被认为是气孔关闭运动中主要的外流阴离子物质.但实际上, 在气孔关闭运动中发挥核心作用的SLAC1和SLAH3均对苹果酸没有明显的通透能力, 真正通过介导苹果酸外流而参与驱动气孔关闭运动的是来自ALMT家族的ALMT12.因此, 气孔关闭运动应该是由SLAC1、SLAH3和ALMT12等多种质膜阴离子协同作用的结果.每个SLAC1亚基具有10个跨膜区, 其C端和N端均位于质膜的胞质一侧(Vahisalu et al., 2008) (图6A).3个SLAC1亚基互相结合, 以三聚体的形式形成质膜阴离子通道, 但每个亚基独立形成1个允许阴离子通过的孔(Chen et al., 2010) (图6B).由此可见, 包括多个SLAHs在内, SLAC1家族形成的阴离子通道可能均为三聚体, 而SLAH1对SLAH3活性的调控作用, 有可能通过与SLAH3组成异源三聚体而实现.SLAC1家族的离子通道亚基组成形式与CNGC和Shaker家族不同, 但它们在不同亚基组成影响离子通道活性和选择性方面又具有类似之处.SLAC1家族离子通道的激活主要依赖蛋白激酶的磷酸化, 而失活则依赖于磷酸酶的去磷酸化(Geiger et al., 2009, 2011; Lee et al., 2009; Brandt et al., 2012; Maierhofer et al., 2014; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
A dual role for the OsK5.2 ion channel in stomatal movements and K + loading into xylem sap 1 2017
... 目前, 对拟南芥Shaker家族的组织分布和功能研究得较为深入全面; 而对包括水稻(Oryza sativa)和玉米(Zea mays)在内的其它植物Shaker家族的研究相对较少.已有的研究鉴定出多个参与K+吸收、转运和气孔运动的玉米和水稻Shaker同源基因.其与拟南芥同源蛋白相比较有很多类似之处, 但也有不同.例如, 玉米KZM2通过与KZM3组成异源多聚体, 抑制K+in通道的活性和气孔运动(Gao et al., 2017).而水稻OsKAT3通过与OsKAT2互作抑制OsKAT3的活性(Hwang et al., 2013).这种K+通道两个同源蛋白之间的互作和调控方式与拟南芥中的AtKC1对AKT1和KAT1等的抑制作用类似(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).再比如, 水稻KAT2与拟南芥KAT2均作为K+in通道参与光照诱导的气孔开放, 但拟南芥气孔保卫细胞质膜K+in通道编码基因数目明显多于水稻(Pilot et al., 2001; Moon et al., 2017).对于外向K+通道而言, 拟南芥SKOR分布于根部, 负责K+向木质部的卸载, 而GORK则主要参与拟南芥的气孔关闭, 二者的组织分布和生物学功能明显不同.而水稻的同一个Shaker蛋白同时参与气孔关闭和K+营养在根部的卸载(Nguyen et al., 2017).因此, 加强对更多植物, 尤其是重要作物中Shaker家族的研究, 将会促进人们对植物吸收、运输和利用K+的分子机理的深入理解. ...
Direct modulation of the guard cell outward-rectifying potassium channel (GORK) by abscisic acid 1 2017
... Shaker家族离子通道的活性受多种调控因子的影响.首先, 细胞内和细胞外K+浓度对K+in和K+out通道活性均有明显影响.这种影响可能是基于K+从高浓度向低浓度方向流动的基本化学特性.酸性环境可以显著抑制拟南芥花粉原生质体质膜K+in通道活性, 而碱性条件则具有一定的激活作用(Fan et al., 2001, 2003).这种作用可能通过影响K+in通道的某些氨基酸残基电离状态和离子通道蛋白结构变化来实现.其次, K+in四聚体的亚基组成形式本身就是一种通道活性调控方式.在非洲爪蟾卵母细胞中, 以不同的排列组合共表达2个不同的拟南芥Shaker家族K+in通道基因, 可以利用电生理技术记录到具有不同电生理特性的K+in通道电流信号(Xicluna et al., 2007; Lebaudy et al., 2010).而AtKC1作为抑制亚基, 通过重组进入K+in四聚体发挥对K+in通道的抑制作用, 是K+in通道的重要负调控因子.K+in四聚体组成形式的变化可能通过影响K+in通道蛋白离子通道孔的结构而影响其离子通道活性.SNARE蛋白SYP121不仅可以影响KAT1等K+in通道的亚细胞定位, 而且可使KAT1内化离开细胞质膜, 从而抑制全细胞K+in通道的电流大小(Sutter et al., 2006, 2007).进一步研究表明, SNARE还可以通过与AtKC1互作调控K+in通道的蛋白活性(Honsbein et al., 2009; Grefen et al., 2010; Jeanguenin et al., 2011).由此可知, SNARE作为K+in通道的上游调控因子可通过对K+in四聚体组成形式的影响发挥调控作用.蛋白激酶和磷酸酶参与调控多种蛋白的功能, 包括内向和外向质膜K+通道, 比如CPK (calcium-dependent protein kinase)家族的CPK33对GORK的激活作用和PP2CA对GORK的抑制作用(Lefoulon et al., 2016; Corratgé-Faillie et al., 2017).K+in通道可以被多种蛋白激酶磷酸化所抑制, 包括CPK家族和SnRK (Rnf1-related protein kinase)家族的多个蛋白激酶(Sato et al., 2009, 2010; Ronzier et al., 2014).此外, AKT1等多个质膜K+in通道的活性还受Ca2+信号及其与CBL-CIPK组成的信号链的正调控(Li et al., 2006; Cheong et al., 2007; Zhang et al., 2010; Ren et al., 2013; Behera et al., 2017).而PP2CA对AKT1的去磷酸化可以与Ca2+-CBL-CIPK信号链协同调控AKT1的活性(Lan et al., 2011).有趣的是, ABA甚至可以直接激活GORK的离子通道活性(Ooi et al., 2017).K+通道的调控因子众多, 这些调控因子并非各自单独发挥作用, 而是彼此协同发挥作用.AtKC1和Ca2+-CBL-CIPK23信号链协同调控AKT1就是一个典型的例子(Wang et al., 2016). ...
Guard cell inward K + channel activity in Arabidopsis involves expression of the twin channel subunits KAT1 and KAT2 2 2001
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
... 目前, 对拟南芥Shaker家族的组织分布和功能研究得较为深入全面; 而对包括水稻(Oryza sativa)和玉米(Zea mays)在内的其它植物Shaker家族的研究相对较少.已有的研究鉴定出多个参与K+吸收、转运和气孔运动的玉米和水稻Shaker同源基因.其与拟南芥同源蛋白相比较有很多类似之处, 但也有不同.例如, 玉米KZM2通过与KZM3组成异源多聚体, 抑制K+in通道的活性和气孔运动(Gao et al., 2017).而水稻OsKAT3通过与OsKAT2互作抑制OsKAT3的活性(Hwang et al., 2013).这种K+通道两个同源蛋白之间的互作和调控方式与拟南芥中的AtKC1对AKT1和KAT1等的抑制作用类似(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).再比如, 水稻KAT2与拟南芥KAT2均作为K+in通道参与光照诱导的气孔开放, 但拟南芥气孔保卫细胞质膜K+in通道编码基因数目明显多于水稻(Pilot et al., 2001; Moon et al., 2017).对于外向K+通道而言, 拟南芥SKOR分布于根部, 负责K+向木质部的卸载, 而GORK则主要参与拟南芥的气孔关闭, 二者的组织分布和生物学功能明显不同.而水稻的同一个Shaker蛋白同时参与气孔关闭和K+营养在根部的卸载(Nguyen et al., 2017).因此, 加强对更多植物, 尤其是重要作物中Shaker家族的研究, 将会促进人们对植物吸收、运输和利用K+的分子机理的深入理解. ...
Not all ALMT1-type transporters mediate aluminum-activated organic acid responses: the case of ZmALMT1—an anion-selective transporter 1 2008
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
Glutamate receptor homologs in plants: functions and evolutionary origins 3 2012
... GLR家族是受多种氨基酸门控的离子通道, 但其通道门控特点并非如其名称仅为谷氨酸所门控, 而是受多种氨基酸影响.1998年, 第1个GLR被克隆出来(Lam et al., 1998), 人们开始对GLR进行研究.研究表明, 拟南芥GLR1和GLR2的N端位于细胞质膜外侧, C端位于细胞质膜内侧, 中间包含4个跨膜区, 但第2个跨膜区并不能完整地跨过细胞质膜(Lam et al., 1998) (图2).这种结构特点明显不同于Shaker和CNGC家族.GLR组成的质膜离子通道的亚基组成、多数GLR离子通道的离子选择性等基本特性, 以及GLR离子通道的活性调控机理目前还不甚清楚.但已经解析出动物GLR四聚体的通道蛋白结构(Price et al., 2012).据此推测, 植物GLR组成的通道蛋白结构可能与动物类似.GLR在植物体内具有多种生物学功能, 包括参与调控花粉管的极性生长(Michard et al., 2011; Wudick et al., 2018)、铝诱导的微管解聚和质膜去极化(Sivaguru et al., 2003)、离子运输(McAinsh and Pittman, 2009)、碳氮代谢(Kang and Turano, 2003)、ABA合成和耐旱反应(Kang et al., 2004; Lu et al., 2014)以及机械损伤信号的长距离传输(Mousavi et al., 2013)等.总体而言, 目前对GLR家族的研究还不够深入.关于植物GLR家族更为详细的介绍可以参阅何明杰等(2016)的综述. ...
... S1-S4: 第1-4跨膜区; LBD: 配体结合结构域; N/C端: 氨/羧基末端 Putative protein structure of an Arabidopsis GLR member (<xref ref-type="bibr" rid="b65">Lam et al., 1998</xref>; <xref ref-type="bibr" rid="b106">Price et al., 2012</xref>) S1-S4: Transmembrane domains 1-4; LBD: Ligand binding domain; N/C termini: The amino terminal and carboxyl terminal ...
The S-type anion channel ZmSLAC1 plays essential roles in stomatal closure by mediating nitrate efflux in maize 1 2018
... SLAC1家族是较晚被发现的一个植物细胞质膜阴离子通道家族, 但植物进化分析表明, SLAC1基因从低等到高等植物均很保守(Lind et al., 2015).拟南芥SLAC1家族由5个成员组成, 包括SLAC1及其同源蛋白SLAH1、2、3和4.其中, SLAC1主要定位在气孔保卫细胞质膜上, 通过介导Cl-和NO3-的外流驱动气孔关闭, 是干旱胁迫及ABA、CO2、臭氧和活性氧(reactive oxygen species, ROS)等信号传递途径的关键组分(Negi et al., 2008; Vahisalu et al., 2008).SLAH3在气孔保卫细胞中分布, 与SLAC1一起通过介导NO3-和Cl-外流驱动气孔关闭(Geiger et al., 2011), 但SLAH3在气孔关闭运动中的作用, 相对于SLAC1而言是辅助性的.拟南芥4个SLAH主要分布在根、茎和叶脉等输导组织中(Negi et al., 2008).其中, SLAH1通过调控SLAH3的活性调控Cl-从根向茎的运输(Cubero-Font et al., 2016).在离子选择性和通透性方面, 拟南芥SLAC1和SLAH3均对Cl-和NO3-有明显的通透能力, 但对NO3-的通透能力明显大于Cl- (Geiger et al., 2009; Lee et al., 2009; Brandt et al., 2012).而SLAH2则为NO3-选择性的阴离子通道, 对Cl-的通透能力微弱(Maierhofer et al., 2014).最近的研究表明, 来自单子叶植物玉米和水稻的SLAC1, 相对于Cl-而言, 对NO3-的选择性明显大于拟南芥SLAC1 (Sun et al., 2016; Qi et al., 2018); 而鸭跖草SLAC1则为NO3-激活的Cl-通透性的阴离子通道(Müller et al., 2017).因此, 来自不同高等植物的SLAC1, 其离子选择性不完全一致, 即不同的高等植物可能采用不同的阴离子通道组合来驱动气孔关闭运动.需要注意的是, 尽管在几十年前就发现苹果酸在气孔保卫细胞内大量积累, 并曾被认为是气孔关闭运动中主要的外流阴离子物质.但实际上, 在气孔关闭运动中发挥核心作用的SLAC1和SLAH3均对苹果酸没有明显的通透能力, 真正通过介导苹果酸外流而参与驱动气孔关闭运动的是来自ALMT家族的ALMT12.因此, 气孔关闭运动应该是由SLAC1、SLAH3和ALMT12等多种质膜阴离子协同作用的结果.每个SLAC1亚基具有10个跨膜区, 其C端和N端均位于质膜的胞质一侧(Vahisalu et al., 2008) (图6A).3个SLAC1亚基互相结合, 以三聚体的形式形成质膜阴离子通道, 但每个亚基独立形成1个允许阴离子通过的孔(Chen et al., 2010) (图6B).由此可见, 包括多个SLAHs在内, SLAC1家族形成的阴离子通道可能均为三聚体, 而SLAH1对SLAH3活性的调控作用, 有可能通过与SLAH3组成异源三聚体而实现.SLAC1家族的离子通道亚基组成形式与CNGC和Shaker家族不同, 但它们在不同亚基组成影响离子通道活性和选择性方面又具有类似之处.SLAC1家族离子通道的激活主要依赖蛋白激酶的磷酸化, 而失活则依赖于磷酸酶的去磷酸化(Geiger et al., 2009, 2011; Lee et al., 2009; Brandt et al., 2012; Maierhofer et al., 2014; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
A mechanosensitive anion channel in Arabidopsis thaliana mesophyll cells 1 2004
... 早期的研究中, 人们发现植物细胞中存在一类受质膜张力激活的Ca2+信号, 从而预测在植物细胞质膜上存在张力激活类质膜Ca2+通道(Crosgrove and Hedrich, 1991; Dutta and Robinson, 2004; Qi et al., 2004; Zhang et al., 2007).多年来, 人们一直没有鉴定到此类受机械或渗透张力激活的离子通道编码基因.近年来, 科学家找到了2类对质膜张力敏感的离子通道, 其一是受低渗条件激活, 通过介导胞外Ca2+内流发挥功能的MCA1和MCA2 (Iida et al., 2014); 其二是对高渗胁迫敏感的OSCA家族(Hou et al., 2014; Yuan et al., 2014).MCA1和MCA2蛋白只有1个跨膜区, 其N端较短且位于细胞质膜外侧, 而C端较长且位于质膜内侧(图3A).研究表明, MCA1和MCA2的N端和跨膜区为其离子通道组成及活性所必需, 而C端则对其离子通道活性发挥调控作用(Kam- ano et al., 2015).MCA1和MCA2组成的离子通道也为四聚体结构, 张力和低渗等刺激通过拉伸质膜改变该离子通道蛋白的结构, 从而使通道被激活, 引起胞外Ca2+内流和细胞质Ca2+水平升高(图3B).MCA1的拟南芥突变体根无法插入较硬的固体培养基(Nakagawa et al., 2007), 说明其在感知机械压力方面发挥作用.最近的研究表明, MCA1和MCA2还作为质膜Ca2+通道介导拟南芥的耐冷反应(Mori et al., 2018).与MCA1和MCA2相反, OSCA是对高渗敏感的质膜离子通道, 可以被细胞外的高渗处理激活(Yuan et al., 2014).拟南芥OSCA家族的蛋白结构与MCA1和MCA2大不相同, 其每个亚基均具有多达9个跨膜区, 在其第8和第9跨膜区之间具有1个孔形成所需的环状结构(Yuan et al., 2014) (图4).OSCA与MCA1和MCA2类似之处在于其亚基N端位于质膜的胞外侧, 而C端则位于质膜胞质侧(Yuan et al., 2014) (图4).OSCA1允许多种离子通过, 其对各种离子的通透能力大小排序为K+>Ba2+≈Ca2+>Na+=Mg2+=Cs+ (Yuan et al., 2014).目前, 人们对质膜张力激活的质膜离子通道的生物学功能了解较少, 但其对渗透敏感的特点暗示此类质膜离子通道可能通过对质膜张力变化做出反应.对低渗敏感的MCA1和MCA2与对高渗敏感的OSCA家族有可能作为一对质膜张力和渗透敏感元件, 协同配合发挥作用, 以共同应对渗透环境变化.一方面, 其可以通过影响胞内Ca2+信号而参与耐盐、耐旱和气孔运动等与细胞渗透和膨压调节密切相关的生物学过程; 另一方面, OSCA对K+的通透能力也表明其可能同时介导Ca2+和K+的跨膜流动而参与这些生物学过程. ...
GABA signaling modulates plant growth by directly regulating the activity of plant- specific anion transporters 1 2015
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
AtKC1, a silent Arabidopsis potassium channel α-subunit modulates root hair K + influx 2 2002
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
... 目前, 对拟南芥Shaker家族的组织分布和功能研究得较为深入全面; 而对包括水稻(Oryza sativa)和玉米(Zea mays)在内的其它植物Shaker家族的研究相对较少.已有的研究鉴定出多个参与K+吸收、转运和气孔运动的玉米和水稻Shaker同源基因.其与拟南芥同源蛋白相比较有很多类似之处, 但也有不同.例如, 玉米KZM2通过与KZM3组成异源多聚体, 抑制K+in通道的活性和气孔运动(Gao et al., 2017).而水稻OsKAT3通过与OsKAT2互作抑制OsKAT3的活性(Hwang et al., 2013).这种K+通道两个同源蛋白之间的互作和调控方式与拟南芥中的AtKC1对AKT1和KAT1等的抑制作用类似(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).再比如, 水稻KAT2与拟南芥KAT2均作为K+in通道参与光照诱导的气孔开放, 但拟南芥气孔保卫细胞质膜K+in通道编码基因数目明显多于水稻(Pilot et al., 2001; Moon et al., 2017).对于外向K+通道而言, 拟南芥SKOR分布于根部, 负责K+向木质部的卸载, 而GORK则主要参与拟南芥的气孔关闭, 二者的组织分布和生物学功能明显不同.而水稻的同一个Shaker蛋白同时参与气孔关闭和K+营养在根部的卸载(Nguyen et al., 2017).因此, 加强对更多植物, 尤其是重要作物中Shaker家族的研究, 将会促进人们对植物吸收、运输和利用K+的分子机理的深入理解. ...
Calcineurin B-like protein CBL10 directly interacts with AKT1 and modulates K + homeostasis in Ara- bidopsis 1 2013
... Shaker家族离子通道的活性受多种调控因子的影响.首先, 细胞内和细胞外K+浓度对K+in和K+out通道活性均有明显影响.这种影响可能是基于K+从高浓度向低浓度方向流动的基本化学特性.酸性环境可以显著抑制拟南芥花粉原生质体质膜K+in通道活性, 而碱性条件则具有一定的激活作用(Fan et al., 2001, 2003).这种作用可能通过影响K+in通道的某些氨基酸残基电离状态和离子通道蛋白结构变化来实现.其次, K+in四聚体的亚基组成形式本身就是一种通道活性调控方式.在非洲爪蟾卵母细胞中, 以不同的排列组合共表达2个不同的拟南芥Shaker家族K+in通道基因, 可以利用电生理技术记录到具有不同电生理特性的K+in通道电流信号(Xicluna et al., 2007; Lebaudy et al., 2010).而AtKC1作为抑制亚基, 通过重组进入K+in四聚体发挥对K+in通道的抑制作用, 是K+in通道的重要负调控因子.K+in四聚体组成形式的变化可能通过影响K+in通道蛋白离子通道孔的结构而影响其离子通道活性.SNARE蛋白SYP121不仅可以影响KAT1等K+in通道的亚细胞定位, 而且可使KAT1内化离开细胞质膜, 从而抑制全细胞K+in通道的电流大小(Sutter et al., 2006, 2007).进一步研究表明, SNARE还可以通过与AtKC1互作调控K+in通道的蛋白活性(Honsbein et al., 2009; Grefen et al., 2010; Jeanguenin et al., 2011).由此可知, SNARE作为K+in通道的上游调控因子可通过对K+in四聚体组成形式的影响发挥调控作用.蛋白激酶和磷酸酶参与调控多种蛋白的功能, 包括内向和外向质膜K+通道, 比如CPK (calcium-dependent protein kinase)家族的CPK33对GORK的激活作用和PP2CA对GORK的抑制作用(Lefoulon et al., 2016; Corratgé-Faillie et al., 2017).K+in通道可以被多种蛋白激酶磷酸化所抑制, 包括CPK家族和SnRK (Rnf1-related protein kinase)家族的多个蛋白激酶(Sato et al., 2009, 2010; Ronzier et al., 2014).此外, AKT1等多个质膜K+in通道的活性还受Ca2+信号及其与CBL-CIPK组成的信号链的正调控(Li et al., 2006; Cheong et al., 2007; Zhang et al., 2010; Ren et al., 2013; Behera et al., 2017).而PP2CA对AKT1的去磷酸化可以与Ca2+-CBL-CIPK信号链协同调控AKT1的活性(Lan et al., 2011).有趣的是, ABA甚至可以直接激活GORK的离子通道活性(Ooi et al., 2017).K+通道的调控因子众多, 这些调控因子并非各自单独发挥作用, 而是彼此协同发挥作用.AtKC1和Ca2+-CBL-CIPK23信号链协同调控AKT1就是一个典型的例子(Wang et al., 2016). ...
CPK13, a noncanonical Ca 2+-dependent protein kinase, specifically inhibits KAT2 and KAT1 shaker K + channels and reduces stomatal opening 1 2014
... Shaker家族离子通道的活性受多种调控因子的影响.首先, 细胞内和细胞外K+浓度对K+in和K+out通道活性均有明显影响.这种影响可能是基于K+从高浓度向低浓度方向流动的基本化学特性.酸性环境可以显著抑制拟南芥花粉原生质体质膜K+in通道活性, 而碱性条件则具有一定的激活作用(Fan et al., 2001, 2003).这种作用可能通过影响K+in通道的某些氨基酸残基电离状态和离子通道蛋白结构变化来实现.其次, K+in四聚体的亚基组成形式本身就是一种通道活性调控方式.在非洲爪蟾卵母细胞中, 以不同的排列组合共表达2个不同的拟南芥Shaker家族K+in通道基因, 可以利用电生理技术记录到具有不同电生理特性的K+in通道电流信号(Xicluna et al., 2007; Lebaudy et al., 2010).而AtKC1作为抑制亚基, 通过重组进入K+in四聚体发挥对K+in通道的抑制作用, 是K+in通道的重要负调控因子.K+in四聚体组成形式的变化可能通过影响K+in通道蛋白离子通道孔的结构而影响其离子通道活性.SNARE蛋白SYP121不仅可以影响KAT1等K+in通道的亚细胞定位, 而且可使KAT1内化离开细胞质膜, 从而抑制全细胞K+in通道的电流大小(Sutter et al., 2006, 2007).进一步研究表明, SNARE还可以通过与AtKC1互作调控K+in通道的蛋白活性(Honsbein et al., 2009; Grefen et al., 2010; Jeanguenin et al., 2011).由此可知, SNARE作为K+in通道的上游调控因子可通过对K+in四聚体组成形式的影响发挥调控作用.蛋白激酶和磷酸酶参与调控多种蛋白的功能, 包括内向和外向质膜K+通道, 比如CPK (calcium-dependent protein kinase)家族的CPK33对GORK的激活作用和PP2CA对GORK的抑制作用(Lefoulon et al., 2016; Corratgé-Faillie et al., 2017).K+in通道可以被多种蛋白激酶磷酸化所抑制, 包括CPK家族和SnRK (Rnf1-related protein kinase)家族的多个蛋白激酶(Sato et al., 2009, 2010; Ronzier et al., 2014).此外, AKT1等多个质膜K+in通道的活性还受Ca2+信号及其与CBL-CIPK组成的信号链的正调控(Li et al., 2006; Cheong et al., 2007; Zhang et al., 2010; Ren et al., 2013; Behera et al., 2017).而PP2CA对AKT1的去磷酸化可以与Ca2+-CBL-CIPK信号链协同调控AKT1的活性(Lan et al., 2011).有趣的是, ABA甚至可以直接激活GORK的离子通道活性(Ooi et al., 2017).K+通道的调控因子众多, 这些调控因子并非各自单独发挥作用, 而是彼此协同发挥作用.AtKC1和Ca2+-CBL-CIPK23信号链协同调控AKT1就是一个典型的例子(Wang et al., 2016). ...
Systems involved in K + uptake from diluted solutions in pepper plants as revealed by the use of specific inhibitors 1 2010
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
Closing plant stomata requires a homolog of an aluminum-activated malate transporter 1 2010
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
A wheat gene encoding an aluminum-activated malate transporter 2 2004
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
... )等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
Modulation of the Arabidopsis KAT1 channel by an activator of protein kinase C in Xenopus laevis oocytes 1 2010
... Shaker家族离子通道的活性受多种调控因子的影响.首先, 细胞内和细胞外K+浓度对K+in和K+out通道活性均有明显影响.这种影响可能是基于K+从高浓度向低浓度方向流动的基本化学特性.酸性环境可以显著抑制拟南芥花粉原生质体质膜K+in通道活性, 而碱性条件则具有一定的激活作用(Fan et al., 2001, 2003).这种作用可能通过影响K+in通道的某些氨基酸残基电离状态和离子通道蛋白结构变化来实现.其次, K+in四聚体的亚基组成形式本身就是一种通道活性调控方式.在非洲爪蟾卵母细胞中, 以不同的排列组合共表达2个不同的拟南芥Shaker家族K+in通道基因, 可以利用电生理技术记录到具有不同电生理特性的K+in通道电流信号(Xicluna et al., 2007; Lebaudy et al., 2010).而AtKC1作为抑制亚基, 通过重组进入K+in四聚体发挥对K+in通道的抑制作用, 是K+in通道的重要负调控因子.K+in四聚体组成形式的变化可能通过影响K+in通道蛋白离子通道孔的结构而影响其离子通道活性.SNARE蛋白SYP121不仅可以影响KAT1等K+in通道的亚细胞定位, 而且可使KAT1内化离开细胞质膜, 从而抑制全细胞K+in通道的电流大小(Sutter et al., 2006, 2007).进一步研究表明, SNARE还可以通过与AtKC1互作调控K+in通道的蛋白活性(Honsbein et al., 2009; Grefen et al., 2010; Jeanguenin et al., 2011).由此可知, SNARE作为K+in通道的上游调控因子可通过对K+in四聚体组成形式的影响发挥调控作用.蛋白激酶和磷酸酶参与调控多种蛋白的功能, 包括内向和外向质膜K+通道, 比如CPK (calcium-dependent protein kinase)家族的CPK33对GORK的激活作用和PP2CA对GORK的抑制作用(Lefoulon et al., 2016; Corratgé-Faillie et al., 2017).K+in通道可以被多种蛋白激酶磷酸化所抑制, 包括CPK家族和SnRK (Rnf1-related protein kinase)家族的多个蛋白激酶(Sato et al., 2009, 2010; Ronzier et al., 2014).此外, AKT1等多个质膜K+in通道的活性还受Ca2+信号及其与CBL-CIPK组成的信号链的正调控(Li et al., 2006; Cheong et al., 2007; Zhang et al., 2010; Ren et al., 2013; Behera et al., 2017).而PP2CA对AKT1的去磷酸化可以与Ca2+-CBL-CIPK信号链协同调控AKT1的活性(Lan et al., 2011).有趣的是, ABA甚至可以直接激活GORK的离子通道活性(Ooi et al., 2017).K+通道的调控因子众多, 这些调控因子并非各自单独发挥作用, 而是彼此协同发挥作用.AtKC1和Ca2+-CBL-CIPK23信号链协同调控AKT1就是一个典型的例子(Wang et al., 2016). ...
Threonine at position 306 of the KAT1 potassium channel is essential for channel activity and is a target site for ABA-activated SnRK2/OST1/SnRK2.6 pro- tein kinase 1 2009
... Shaker家族离子通道的活性受多种调控因子的影响.首先, 细胞内和细胞外K+浓度对K+in和K+out通道活性均有明显影响.这种影响可能是基于K+从高浓度向低浓度方向流动的基本化学特性.酸性环境可以显著抑制拟南芥花粉原生质体质膜K+in通道活性, 而碱性条件则具有一定的激活作用(Fan et al., 2001, 2003).这种作用可能通过影响K+in通道的某些氨基酸残基电离状态和离子通道蛋白结构变化来实现.其次, K+in四聚体的亚基组成形式本身就是一种通道活性调控方式.在非洲爪蟾卵母细胞中, 以不同的排列组合共表达2个不同的拟南芥Shaker家族K+in通道基因, 可以利用电生理技术记录到具有不同电生理特性的K+in通道电流信号(Xicluna et al., 2007; Lebaudy et al., 2010).而AtKC1作为抑制亚基, 通过重组进入K+in四聚体发挥对K+in通道的抑制作用, 是K+in通道的重要负调控因子.K+in四聚体组成形式的变化可能通过影响K+in通道蛋白离子通道孔的结构而影响其离子通道活性.SNARE蛋白SYP121不仅可以影响KAT1等K+in通道的亚细胞定位, 而且可使KAT1内化离开细胞质膜, 从而抑制全细胞K+in通道的电流大小(Sutter et al., 2006, 2007).进一步研究表明, SNARE还可以通过与AtKC1互作调控K+in通道的蛋白活性(Honsbein et al., 2009; Grefen et al., 2010; Jeanguenin et al., 2011).由此可知, SNARE作为K+in通道的上游调控因子可通过对K+in四聚体组成形式的影响发挥调控作用.蛋白激酶和磷酸酶参与调控多种蛋白的功能, 包括内向和外向质膜K+通道, 比如CPK (calcium-dependent protein kinase)家族的CPK33对GORK的激活作用和PP2CA对GORK的抑制作用(Lefoulon et al., 2016; Corratgé-Faillie et al., 2017).K+in通道可以被多种蛋白激酶磷酸化所抑制, 包括CPK家族和SnRK (Rnf1-related protein kinase)家族的多个蛋白激酶(Sato et al., 2009, 2010; Ronzier et al., 2014).此外, AKT1等多个质膜K+in通道的活性还受Ca2+信号及其与CBL-CIPK组成的信号链的正调控(Li et al., 2006; Cheong et al., 2007; Zhang et al., 2010; Ren et al., 2013; Behera et al., 2017).而PP2CA对AKT1的去磷酸化可以与Ca2+-CBL-CIPK信号链协同调控AKT1的活性(Lan et al., 2011).有趣的是, ABA甚至可以直接激活GORK的离子通道活性(Ooi et al., 2017).K+通道的调控因子众多, 这些调控因子并非各自单独发挥作用, 而是彼此协同发挥作用.AtKC1和Ca2+-CBL-CIPK23信号链协同调控AKT1就是一个典型的例子(Wang et al., 2016). ...
K + transport properties of K + channels in the plasma membrane of Vicia faba guard cells 1 1988
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
Quantitative analysis of outward rec- tifying K + channel currents in guard cell protoplasts from Vicia faba 0 1989
Cytosolic calcium regula- tes ion channels in the plasma membrane of Vicia faba guard cells 2 1989
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
Voltage dependence of K + channels in guard-cell protoplasts 1 1987
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
Characterization of a calmodulin-binding transporter from the plasma membrane of barley aleurone 1 1998
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
The ALMT family of organic acid transporters in plants and their involvement in detoxification and nutrient security 5 2016
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
... 等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
... ).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
... 拟南芥ALMT家族通道蛋白结构(改自Sharma et al., 2016) The putative protein structure of an ALMT member from Arabidopsis (<xref ref-type="bibr" rid="b124">modified from Sharma et al., 2016</xref>)Figure 5 SLAC1家族是较晚被发现的一个植物细胞质膜阴离子通道家族, 但植物进化分析表明, SLAC1基因从低等到高等植物均很保守(Lind et al., 2015).拟南芥SLAC1家族由5个成员组成, 包括SLAC1及其同源蛋白SLAH1、2、3和4.其中, SLAC1主要定位在气孔保卫细胞质膜上, 通过介导Cl-和NO3-的外流驱动气孔关闭, 是干旱胁迫及ABA、CO2、臭氧和活性氧(reactive oxygen species, ROS)等信号传递途径的关键组分(Negi et al., 2008; Vahisalu et al., 2008).SLAH3在气孔保卫细胞中分布, 与SLAC1一起通过介导NO3-和Cl-外流驱动气孔关闭(Geiger et al., 2011), 但SLAH3在气孔关闭运动中的作用, 相对于SLAC1而言是辅助性的.拟南芥4个SLAH主要分布在根、茎和叶脉等输导组织中(Negi et al., 2008).其中, SLAH1通过调控SLAH3的活性调控Cl-从根向茎的运输(Cubero-Font et al., 2016).在离子选择性和通透性方面, 拟南芥SLAC1和SLAH3均对Cl-和NO3-有明显的通透能力, 但对NO3-的通透能力明显大于Cl- (Geiger et al., 2009; Lee et al., 2009; Brandt et al., 2012).而SLAH2则为NO3-选择性的阴离子通道, 对Cl-的通透能力微弱(Maierhofer et al., 2014).最近的研究表明, 来自单子叶植物玉米和水稻的SLAC1, 相对于Cl-而言, 对NO3-的选择性明显大于拟南芥SLAC1 (Sun et al., 2016; Qi et al., 2018); 而鸭跖草SLAC1则为NO3-激活的Cl-通透性的阴离子通道(Müller et al., 2017).因此, 来自不同高等植物的SLAC1, 其离子选择性不完全一致, 即不同的高等植物可能采用不同的阴离子通道组合来驱动气孔关闭运动.需要注意的是, 尽管在几十年前就发现苹果酸在气孔保卫细胞内大量积累, 并曾被认为是气孔关闭运动中主要的外流阴离子物质.但实际上, 在气孔关闭运动中发挥核心作用的SLAC1和SLAH3均对苹果酸没有明显的通透能力, 真正通过介导苹果酸外流而参与驱动气孔关闭运动的是来自ALMT家族的ALMT12.因此, 气孔关闭运动应该是由SLAC1、SLAH3和ALMT12等多种质膜阴离子协同作用的结果.每个SLAC1亚基具有10个跨膜区, 其C端和N端均位于质膜的胞质一侧(Vahisalu et al., 2008) (图6A).3个SLAC1亚基互相结合, 以三聚体的形式形成质膜阴离子通道, 但每个亚基独立形成1个允许阴离子通过的孔(Chen et al., 2010) (图6B).由此可见, 包括多个SLAHs在内, SLAC1家族形成的阴离子通道可能均为三聚体, 而SLAH1对SLAH3活性的调控作用, 有可能通过与SLAH3组成异源三聚体而实现.SLAC1家族的离子通道亚基组成形式与CNGC和Shaker家族不同, 但它们在不同亚基组成影响离子通道活性和选择性方面又具有类似之处.SLAC1家族离子通道的激活主要依赖蛋白激酶的磷酸化, 而失活则依赖于磷酸酶的去磷酸化(Geiger et al., 2009, 2011; Lee et al., 2009; Brandt et al., 2012; Maierhofer et al., 2014; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
... The putative protein structure of an ALMT member from Arabidopsis (modified from Sharma et al., 2016) Figure 5 SLAC1家族是较晚被发现的一个植物细胞质膜阴离子通道家族, 但植物进化分析表明, SLAC1基因从低等到高等植物均很保守(Lind et al., 2015).拟南芥SLAC1家族由5个成员组成, 包括SLAC1及其同源蛋白SLAH1、2、3和4.其中, SLAC1主要定位在气孔保卫细胞质膜上, 通过介导Cl-和NO3-的外流驱动气孔关闭, 是干旱胁迫及ABA、CO2、臭氧和活性氧(reactive oxygen species, ROS)等信号传递途径的关键组分(Negi et al., 2008; Vahisalu et al., 2008).SLAH3在气孔保卫细胞中分布, 与SLAC1一起通过介导NO3-和Cl-外流驱动气孔关闭(Geiger et al., 2011), 但SLAH3在气孔关闭运动中的作用, 相对于SLAC1而言是辅助性的.拟南芥4个SLAH主要分布在根、茎和叶脉等输导组织中(Negi et al., 2008).其中, SLAH1通过调控SLAH3的活性调控Cl-从根向茎的运输(Cubero-Font et al., 2016).在离子选择性和通透性方面, 拟南芥SLAC1和SLAH3均对Cl-和NO3-有明显的通透能力, 但对NO3-的通透能力明显大于Cl- (Geiger et al., 2009; Lee et al., 2009; Brandt et al., 2012).而SLAH2则为NO3-选择性的阴离子通道, 对Cl-的通透能力微弱(Maierhofer et al., 2014).最近的研究表明, 来自单子叶植物玉米和水稻的SLAC1, 相对于Cl-而言, 对NO3-的选择性明显大于拟南芥SLAC1 (Sun et al., 2016; Qi et al., 2018); 而鸭跖草SLAC1则为NO3-激活的Cl-通透性的阴离子通道(Müller et al., 2017).因此, 来自不同高等植物的SLAC1, 其离子选择性不完全一致, 即不同的高等植物可能采用不同的阴离子通道组合来驱动气孔关闭运动.需要注意的是, 尽管在几十年前就发现苹果酸在气孔保卫细胞内大量积累, 并曾被认为是气孔关闭运动中主要的外流阴离子物质.但实际上, 在气孔关闭运动中发挥核心作用的SLAC1和SLAH3均对苹果酸没有明显的通透能力, 真正通过介导苹果酸外流而参与驱动气孔关闭运动的是来自ALMT家族的ALMT12.因此, 气孔关闭运动应该是由SLAC1、SLAH3和ALMT12等多种质膜阴离子协同作用的结果.每个SLAC1亚基具有10个跨膜区, 其C端和N端均位于质膜的胞质一侧(Vahisalu et al., 2008) (图6A).3个SLAC1亚基互相结合, 以三聚体的形式形成质膜阴离子通道, 但每个亚基独立形成1个允许阴离子通过的孔(Chen et al., 2010) (图6B).由此可见, 包括多个SLAHs在内, SLAC1家族形成的阴离子通道可能均为三聚体, 而SLAH1对SLAH3活性的调控作用, 有可能通过与SLAH3组成异源三聚体而实现.SLAC1家族的离子通道亚基组成形式与CNGC和Shaker家族不同, 但它们在不同亚基组成影响离子通道活性和选择性方面又具有类似之处.SLAC1家族离子通道的激活主要依赖蛋白激酶的磷酸化, 而失活则依赖于磷酸酶的去磷酸化(Geiger et al., 2009, 2011; Lee et al., 2009; Brandt et al., 2012; Maierhofer et al., 2014; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
The role of K + channels in uptake and redistribution of potassium in the model plant Arabidopsis thaliana 3 2013
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
... ; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
The cyclic nucleotide-gated channel CNGC14 regulates root gravitropism in Arabidopsis thaliana 1 2015
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
Aluminum rapidly depolymerizes cortical microtubules and depolarizes the plasma membrane: evidence that these responses are mediated by a glutamate receptor 1 2003
... GLR家族是受多种氨基酸门控的离子通道, 但其通道门控特点并非如其名称仅为谷氨酸所门控, 而是受多种氨基酸影响.1998年, 第1个GLR被克隆出来(Lam et al., 1998), 人们开始对GLR进行研究.研究表明, 拟南芥GLR1和GLR2的N端位于细胞质膜外侧, C端位于细胞质膜内侧, 中间包含4个跨膜区, 但第2个跨膜区并不能完整地跨过细胞质膜(Lam et al., 1998) (图2).这种结构特点明显不同于Shaker和CNGC家族.GLR组成的质膜离子通道的亚基组成、多数GLR离子通道的离子选择性等基本特性, 以及GLR离子通道的活性调控机理目前还不甚清楚.但已经解析出动物GLR四聚体的通道蛋白结构(Price et al., 2012).据此推测, 植物GLR组成的通道蛋白结构可能与动物类似.GLR在植物体内具有多种生物学功能, 包括参与调控花粉管的极性生长(Michard et al., 2011; Wudick et al., 2018)、铝诱导的微管解聚和质膜去极化(Sivaguru et al., 2003)、离子运输(McAinsh and Pittman, 2009)、碳氮代谢(Kang and Turano, 2003)、ABA合成和耐旱反应(Kang et al., 2004; Lu et al., 2014)以及机械损伤信号的长距离传输(Mousavi et al., 2013)等.总体而言, 目前对GLR家族的研究还不够深入.关于植物GLR家族更为详细的介绍可以参阅何明杰等(2016)的综述. ...
Structure and gating of tetrameric glutamate receptors 1 2015
... 植物体内存在大量的钙元素.一方面钙作为细胞壁等结构组分发挥作用; 另一方面, 离子形式的钙(Ca2+)可以作为信号分子参与各种生物学反应和信号传递过程, 后者则备受关注.胞内Ca2+信号以细胞质Ca2+浓度的升高和震荡形式出现.一般认为, 胞外Ca2+内流是细胞质Ca2+信号产生所需Ca2+的主要来源, 而质膜内向Ca2+通道是胞外Ca2+内流的主要渠道.同时, Ca2+通道的活性变化通过控制胞外Ca2+内流的节奏直接调控胞内Ca2+震荡, 发挥Ca2+信号上游调控因子的作用.鉴于此, 质膜Ca2+通道受到广泛关注, 并成为动植物各信号领域的重点研究对象.科学家首先在动物细胞中发现多个Ca2+通道家族, 主要包括受环核苷酸cAMP和cGMP激活的CNG (cyclic nucleotide-gated)家族、受谷氨酸门控的GLR (glutamate receptor)家族、TRPC (transient receptor potential channel)和受胞内Ca2+库倒空激活的CRACM1/Orai1 (Zagotta and Siegelbaum, 1996; Vig and Kinet, 2007; Sobolevsky, 2015; Ambudkar et al., 2017).通过序列比对, 多个动物细胞质膜Ca2+通道的同源基因家族先后在植物尤其是模式植物拟南芥基因组中被鉴定出来, 主要包括CNGC (cyclic nucleotide- gated channel)、GLR (glutamate receptor-like)、高渗敏感的OSCA (hyperosmolality-gated calcium- permeable channel)家族和低渗敏感的张力激活离子通道MCA (mechano-sensitive channel) (M?ser et al., 2001; Ward et al., 2009; Iida et al., 2014; Yuan et al., 2014). ...
Paenibacillus yonginensis DCY84 T induces changes in Arabidopsis thaliana gene expression against aluminum, drought, and salt stress 1 2015
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
Protein kinase OsSAPK8 functions as an essential activator of S-type anion channel OsSLAC1, which is nitrate-selective in rice 1 2016
... SLAC1家族是较晚被发现的一个植物细胞质膜阴离子通道家族, 但植物进化分析表明, SLAC1基因从低等到高等植物均很保守(Lind et al., 2015).拟南芥SLAC1家族由5个成员组成, 包括SLAC1及其同源蛋白SLAH1、2、3和4.其中, SLAC1主要定位在气孔保卫细胞质膜上, 通过介导Cl-和NO3-的外流驱动气孔关闭, 是干旱胁迫及ABA、CO2、臭氧和活性氧(reactive oxygen species, ROS)等信号传递途径的关键组分(Negi et al., 2008; Vahisalu et al., 2008).SLAH3在气孔保卫细胞中分布, 与SLAC1一起通过介导NO3-和Cl-外流驱动气孔关闭(Geiger et al., 2011), 但SLAH3在气孔关闭运动中的作用, 相对于SLAC1而言是辅助性的.拟南芥4个SLAH主要分布在根、茎和叶脉等输导组织中(Negi et al., 2008).其中, SLAH1通过调控SLAH3的活性调控Cl-从根向茎的运输(Cubero-Font et al., 2016).在离子选择性和通透性方面, 拟南芥SLAC1和SLAH3均对Cl-和NO3-有明显的通透能力, 但对NO3-的通透能力明显大于Cl- (Geiger et al., 2009; Lee et al., 2009; Brandt et al., 2012).而SLAH2则为NO3-选择性的阴离子通道, 对Cl-的通透能力微弱(Maierhofer et al., 2014).最近的研究表明, 来自单子叶植物玉米和水稻的SLAC1, 相对于Cl-而言, 对NO3-的选择性明显大于拟南芥SLAC1 (Sun et al., 2016; Qi et al., 2018); 而鸭跖草SLAC1则为NO3-激活的Cl-通透性的阴离子通道(Müller et al., 2017).因此, 来自不同高等植物的SLAC1, 其离子选择性不完全一致, 即不同的高等植物可能采用不同的阴离子通道组合来驱动气孔关闭运动.需要注意的是, 尽管在几十年前就发现苹果酸在气孔保卫细胞内大量积累, 并曾被认为是气孔关闭运动中主要的外流阴离子物质.但实际上, 在气孔关闭运动中发挥核心作用的SLAC1和SLAH3均对苹果酸没有明显的通透能力, 真正通过介导苹果酸外流而参与驱动气孔关闭运动的是来自ALMT家族的ALMT12.因此, 气孔关闭运动应该是由SLAC1、SLAH3和ALMT12等多种质膜阴离子协同作用的结果.每个SLAC1亚基具有10个跨膜区, 其C端和N端均位于质膜的胞质一侧(Vahisalu et al., 2008) (图6A).3个SLAC1亚基互相结合, 以三聚体的形式形成质膜阴离子通道, 但每个亚基独立形成1个允许阴离子通过的孔(Chen et al., 2010) (图6B).由此可见, 包括多个SLAHs在内, SLAC1家族形成的阴离子通道可能均为三聚体, 而SLAH1对SLAH3活性的调控作用, 有可能通过与SLAH3组成异源三聚体而实现.SLAC1家族的离子通道亚基组成形式与CNGC和Shaker家族不同, 但它们在不同亚基组成影响离子通道活性和选择性方面又具有类似之处.SLAC1家族离子通道的激活主要依赖蛋白激酶的磷酸化, 而失活则依赖于磷酸酶的去磷酸化(Geiger et al., 2009, 2011; Lee et al., 2009; Brandt et al., 2012; Maierhofer et al., 2014; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
Selective mobility and sensitivity to SNAREs is exhibited by the Arabidopsis KAT1 K + channel at the plasma membrane 1 2006
... Shaker家族离子通道的活性受多种调控因子的影响.首先, 细胞内和细胞外K+浓度对K+in和K+out通道活性均有明显影响.这种影响可能是基于K+从高浓度向低浓度方向流动的基本化学特性.酸性环境可以显著抑制拟南芥花粉原生质体质膜K+in通道活性, 而碱性条件则具有一定的激活作用(Fan et al., 2001, 2003).这种作用可能通过影响K+in通道的某些氨基酸残基电离状态和离子通道蛋白结构变化来实现.其次, K+in四聚体的亚基组成形式本身就是一种通道活性调控方式.在非洲爪蟾卵母细胞中, 以不同的排列组合共表达2个不同的拟南芥Shaker家族K+in通道基因, 可以利用电生理技术记录到具有不同电生理特性的K+in通道电流信号(Xicluna et al., 2007; Lebaudy et al., 2010).而AtKC1作为抑制亚基, 通过重组进入K+in四聚体发挥对K+in通道的抑制作用, 是K+in通道的重要负调控因子.K+in四聚体组成形式的变化可能通过影响K+in通道蛋白离子通道孔的结构而影响其离子通道活性.SNARE蛋白SYP121不仅可以影响KAT1等K+in通道的亚细胞定位, 而且可使KAT1内化离开细胞质膜, 从而抑制全细胞K+in通道的电流大小(Sutter et al., 2006, 2007).进一步研究表明, SNARE还可以通过与AtKC1互作调控K+in通道的蛋白活性(Honsbein et al., 2009; Grefen et al., 2010; Jeanguenin et al., 2011).由此可知, SNARE作为K+in通道的上游调控因子可通过对K+in四聚体组成形式的影响发挥调控作用.蛋白激酶和磷酸酶参与调控多种蛋白的功能, 包括内向和外向质膜K+通道, 比如CPK (calcium-dependent protein kinase)家族的CPK33对GORK的激活作用和PP2CA对GORK的抑制作用(Lefoulon et al., 2016; Corratgé-Faillie et al., 2017).K+in通道可以被多种蛋白激酶磷酸化所抑制, 包括CPK家族和SnRK (Rnf1-related protein kinase)家族的多个蛋白激酶(Sato et al., 2009, 2010; Ronzier et al., 2014).此外, AKT1等多个质膜K+in通道的活性还受Ca2+信号及其与CBL-CIPK组成的信号链的正调控(Li et al., 2006; Cheong et al., 2007; Zhang et al., 2010; Ren et al., 2013; Behera et al., 2017).而PP2CA对AKT1的去磷酸化可以与Ca2+-CBL-CIPK信号链协同调控AKT1的活性(Lan et al., 2011).有趣的是, ABA甚至可以直接激活GORK的离子通道活性(Ooi et al., 2017).K+通道的调控因子众多, 这些调控因子并非各自单独发挥作用, 而是彼此协同发挥作用.AtKC1和Ca2+-CBL-CIPK23信号链协同调控AKT1就是一个典型的例子(Wang et al., 2016). ...
Abscisic acid triggers the endocytosis of the Arabidopsis KAT1 K + channel and its recycling to the plasma membrane 1 2007
... Shaker家族离子通道的活性受多种调控因子的影响.首先, 细胞内和细胞外K+浓度对K+in和K+out通道活性均有明显影响.这种影响可能是基于K+从高浓度向低浓度方向流动的基本化学特性.酸性环境可以显著抑制拟南芥花粉原生质体质膜K+in通道活性, 而碱性条件则具有一定的激活作用(Fan et al., 2001, 2003).这种作用可能通过影响K+in通道的某些氨基酸残基电离状态和离子通道蛋白结构变化来实现.其次, K+in四聚体的亚基组成形式本身就是一种通道活性调控方式.在非洲爪蟾卵母细胞中, 以不同的排列组合共表达2个不同的拟南芥Shaker家族K+in通道基因, 可以利用电生理技术记录到具有不同电生理特性的K+in通道电流信号(Xicluna et al., 2007; Lebaudy et al., 2010).而AtKC1作为抑制亚基, 通过重组进入K+in四聚体发挥对K+in通道的抑制作用, 是K+in通道的重要负调控因子.K+in四聚体组成形式的变化可能通过影响K+in通道蛋白离子通道孔的结构而影响其离子通道活性.SNARE蛋白SYP121不仅可以影响KAT1等K+in通道的亚细胞定位, 而且可使KAT1内化离开细胞质膜, 从而抑制全细胞K+in通道的电流大小(Sutter et al., 2006, 2007).进一步研究表明, SNARE还可以通过与AtKC1互作调控K+in通道的蛋白活性(Honsbein et al., 2009; Grefen et al., 2010; Jeanguenin et al., 2011).由此可知, SNARE作为K+in通道的上游调控因子可通过对K+in四聚体组成形式的影响发挥调控作用.蛋白激酶和磷酸酶参与调控多种蛋白的功能, 包括内向和外向质膜K+通道, 比如CPK (calcium-dependent protein kinase)家族的CPK33对GORK的激活作用和PP2CA对GORK的抑制作用(Lefoulon et al., 2016; Corratgé-Faillie et al., 2017).K+in通道可以被多种蛋白激酶磷酸化所抑制, 包括CPK家族和SnRK (Rnf1-related protein kinase)家族的多个蛋白激酶(Sato et al., 2009, 2010; Ronzier et al., 2014).此外, AKT1等多个质膜K+in通道的活性还受Ca2+信号及其与CBL-CIPK组成的信号链的正调控(Li et al., 2006; Cheong et al., 2007; Zhang et al., 2010; Ren et al., 2013; Behera et al., 2017).而PP2CA对AKT1的去磷酸化可以与Ca2+-CBL-CIPK信号链协同调控AKT1的活性(Lan et al., 2011).有趣的是, ABA甚至可以直接激活GORK的离子通道活性(Ooi et al., 2017).K+通道的调控因子众多, 这些调控因子并非各自单独发挥作用, 而是彼此协同发挥作用.AtKC1和Ca2+-CBL-CIPK23信号链协同调控AKT1就是一个典型的例子(Wang et al., 2016). ...
KAT1 is not essential for stomatal opening 2 2001
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
... ).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
A dicarboxylate transporter, LjALMT4, mainly expressed in nodules of Lotus japonicus 1 2016
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
Cyclic nucleotide gated channels 7 and 8 are essential for male reproductive fertility 1 2013
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
A cyclic nucleotide-gated channel (CNGC16) in pollen is critical for stress tolerance in pollen reproductive development 1 2013
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
The cyclic nucleotide-gated channels AtCNGC11 and 12 are involved in multiple Ca 2+-dependent physiological responses and act in a synergistic manner 1 2011
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
The chimeric cyclic nucleotide-gated ion channel ATCNGC11/12 constitutively induces programmed cell death in a Ca 2+ dependent manner 1 2007
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
SLAC1 is required for plant guard cell S-type anion channel function in stomatal signaling 5 2008
... SLAC1家族是较晚被发现的一个植物细胞质膜阴离子通道家族, 但植物进化分析表明, SLAC1基因从低等到高等植物均很保守(Lind et al., 2015).拟南芥SLAC1家族由5个成员组成, 包括SLAC1及其同源蛋白SLAH1、2、3和4.其中, SLAC1主要定位在气孔保卫细胞质膜上, 通过介导Cl-和NO3-的外流驱动气孔关闭, 是干旱胁迫及ABA、CO2、臭氧和活性氧(reactive oxygen species, ROS)等信号传递途径的关键组分(Negi et al., 2008; Vahisalu et al., 2008).SLAH3在气孔保卫细胞中分布, 与SLAC1一起通过介导NO3-和Cl-外流驱动气孔关闭(Geiger et al., 2011), 但SLAH3在气孔关闭运动中的作用, 相对于SLAC1而言是辅助性的.拟南芥4个SLAH主要分布在根、茎和叶脉等输导组织中(Negi et al., 2008).其中, SLAH1通过调控SLAH3的活性调控Cl-从根向茎的运输(Cubero-Font et al., 2016).在离子选择性和通透性方面, 拟南芥SLAC1和SLAH3均对Cl-和NO3-有明显的通透能力, 但对NO3-的通透能力明显大于Cl- (Geiger et al., 2009; Lee et al., 2009; Brandt et al., 2012).而SLAH2则为NO3-选择性的阴离子通道, 对Cl-的通透能力微弱(Maierhofer et al., 2014).最近的研究表明, 来自单子叶植物玉米和水稻的SLAC1, 相对于Cl-而言, 对NO3-的选择性明显大于拟南芥SLAC1 (Sun et al., 2016; Qi et al., 2018); 而鸭跖草SLAC1则为NO3-激活的Cl-通透性的阴离子通道(Müller et al., 2017).因此, 来自不同高等植物的SLAC1, 其离子选择性不完全一致, 即不同的高等植物可能采用不同的阴离子通道组合来驱动气孔关闭运动.需要注意的是, 尽管在几十年前就发现苹果酸在气孔保卫细胞内大量积累, 并曾被认为是气孔关闭运动中主要的外流阴离子物质.但实际上, 在气孔关闭运动中发挥核心作用的SLAC1和SLAH3均对苹果酸没有明显的通透能力, 真正通过介导苹果酸外流而参与驱动气孔关闭运动的是来自ALMT家族的ALMT12.因此, 气孔关闭运动应该是由SLAC1、SLAH3和ALMT12等多种质膜阴离子协同作用的结果.每个SLAC1亚基具有10个跨膜区, 其C端和N端均位于质膜的胞质一侧(Vahisalu et al., 2008) (图6A).3个SLAC1亚基互相结合, 以三聚体的形式形成质膜阴离子通道, 但每个亚基独立形成1个允许阴离子通过的孔(Chen et al., 2010) (图6B).由此可见, 包括多个SLAHs在内, SLAC1家族形成的阴离子通道可能均为三聚体, 而SLAH1对SLAH3活性的调控作用, 有可能通过与SLAH3组成异源三聚体而实现.SLAC1家族的离子通道亚基组成形式与CNGC和Shaker家族不同, 但它们在不同亚基组成影响离子通道活性和选择性方面又具有类似之处.SLAC1家族离子通道的激活主要依赖蛋白激酶的磷酸化, 而失活则依赖于磷酸酶的去磷酸化(Geiger et al., 2009, 2011; Lee et al., 2009; Brandt et al., 2012; Maierhofer et al., 2014; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
... ).因此, 来自不同高等植物的SLAC1, 其离子选择性不完全一致, 即不同的高等植物可能采用不同的阴离子通道组合来驱动气孔关闭运动.需要注意的是, 尽管在几十年前就发现苹果酸在气孔保卫细胞内大量积累, 并曾被认为是气孔关闭运动中主要的外流阴离子物质.但实际上, 在气孔关闭运动中发挥核心作用的SLAC1和SLAH3均对苹果酸没有明显的通透能力, 真正通过介导苹果酸外流而参与驱动气孔关闭运动的是来自ALMT家族的ALMT12.因此, 气孔关闭运动应该是由SLAC1、SLAH3和ALMT12等多种质膜阴离子协同作用的结果.每个SLAC1亚基具有10个跨膜区, 其C端和N端均位于质膜的胞质一侧(Vahisalu et al., 2008) (图6A).3个SLAC1亚基互相结合, 以三聚体的形式形成质膜阴离子通道, 但每个亚基独立形成1个允许阴离子通过的孔(Chen et al., 2010) (图6B).由此可见, 包括多个SLAHs在内, SLAC1家族形成的阴离子通道可能均为三聚体, 而SLAH1对SLAH3活性的调控作用, 有可能通过与SLAH3组成异源三聚体而实现.SLAC1家族的离子通道亚基组成形式与CNGC和Shaker家族不同, 但它们在不同亚基组成影响离子通道活性和选择性方面又具有类似之处.SLAC1家族离子通道的激活主要依赖蛋白激酶的磷酸化, 而失活则依赖于磷酸酶的去磷酸化(Geiger et al., 2009, 2011; Lee et al., 2009; Brandt et al., 2012; Maierhofer et al., 2014; Cubero-Font et al., 2016).其中, 参与SLAC1及其同源蛋白离子通道活性调控的蛋白激酶主要来自SnRK、CPK和CBL-CIPK (calcineurin B-like, CBL; CBL-interacting protein kinase, CIPK)等蛋白激酶家族及PP2C (protein phosphatases 2C)磷酸酶家族. ...
... SLAC1亚基(A)和离子通道蛋白结构(B) (Vahisalu et al., 2008; Chen et al., 2010) The topological structures of a SLAC1 subunit (A) and the structure of a SLAC1 channel as a trimer (B) (<xref ref-type="bibr" rid="b139">Vahisalu et al., 2008</xref>; <xref ref-type="bibr" rid="b9">Chen et al., 2010</xref>)Figure 6 除了SLAC1家族, 植物细胞质膜上还包含张力激活的阴离子通道MSL (mechano-sensitive or stret- ch-activated channels) (Crosgrove and Hedrich, 1991).MSL为典型的张力激活的阴离子通道家族.MSL9和MSL10是最早得到电生理鉴定的MSL家族成员, 定位于细胞质膜, 其激活对质膜张力具有依赖性, 主要选择性介导阴离子的跨膜流动(Haswell et al., 2008; Maksaev and Haswell, 2012).随后有研究表明, 拟南芥MSL10参与植物机械损伤信号反应和茉莉酸的累积(Veley et al., 2014; Zou et al., 2016).此外, 有些拟南芥MSL成员定位在细胞器膜上, 调控细胞器的大小和形状(Veley et al., 2012).根据氨基酸残基序列预测, MSL蛋白存在多个跨膜区, 具备形成离子通道的基本结构特点.而质膜受到拉伸时对MSL的激活是通过机械拉力直接改变MSL蛋白的结构, 使离子通过的孔被拉开.但这基本上还停留在理论推测阶段, 缺乏有力的直接实验证据支持.总体来讲, 目前对MSL家族的生物学功能还了解较少, 其组成的离子通道蛋白结构、感知生物膜张力的分子机理和生物学功能等还有待深入研究. ...
... The topological structures of a SLAC1 subunit (A) and the structure of a SLAC1 channel as a trimer (B) (Vahisalu et al., 2008; Chen et al., 2010) Figure 6 除了SLAC1家族, 植物细胞质膜上还包含张力激活的阴离子通道MSL (mechano-sensitive or stret- ch-activated channels) (Crosgrove and Hedrich, 1991).MSL为典型的张力激活的阴离子通道家族.MSL9和MSL10是最早得到电生理鉴定的MSL家族成员, 定位于细胞质膜, 其激活对质膜张力具有依赖性, 主要选择性介导阴离子的跨膜流动(Haswell et al., 2008; Maksaev and Haswell, 2012).随后有研究表明, 拟南芥MSL10参与植物机械损伤信号反应和茉莉酸的累积(Veley et al., 2014; Zou et al., 2016).此外, 有些拟南芥MSL成员定位在细胞器膜上, 调控细胞器的大小和形状(Veley et al., 2012).根据氨基酸残基序列预测, MSL蛋白存在多个跨膜区, 具备形成离子通道的基本结构特点.而质膜受到拉伸时对MSL的激活是通过机械拉力直接改变MSL蛋白的结构, 使离子通过的孔被拉开.但这基本上还停留在理论推测阶段, 缺乏有力的直接实验证据支持.总体来讲, 目前对MSL家族的生物学功能还了解较少, 其组成的离子通道蛋白结构、感知生物膜张力的分子机理和生物学功能等还有待深入研究. ...
Arabidopsis MSL10 has a regulated cell death signaling activity that is separable from its mechanosensitive ion channel activity 1 2014
... 除了SLAC1家族, 植物细胞质膜上还包含张力激活的阴离子通道MSL (mechano-sensitive or stret- ch-activated channels) (Crosgrove and Hedrich, 1991).MSL为典型的张力激活的阴离子通道家族.MSL9和MSL10是最早得到电生理鉴定的MSL家族成员, 定位于细胞质膜, 其激活对质膜张力具有依赖性, 主要选择性介导阴离子的跨膜流动(Haswell et al., 2008; Maksaev and Haswell, 2012).随后有研究表明, 拟南芥MSL10参与植物机械损伤信号反应和茉莉酸的累积(Veley et al., 2014; Zou et al., 2016).此外, 有些拟南芥MSL成员定位在细胞器膜上, 调控细胞器的大小和形状(Veley et al., 2012).根据氨基酸残基序列预测, MSL蛋白存在多个跨膜区, 具备形成离子通道的基本结构特点.而质膜受到拉伸时对MSL的激活是通过机械拉力直接改变MSL蛋白的结构, 使离子通过的孔被拉开.但这基本上还停留在理论推测阶段, 缺乏有力的直接实验证据支持.总体来讲, 目前对MSL家族的生物学功能还了解较少, 其组成的离子通道蛋白结构、感知生物膜张力的分子机理和生物学功能等还有待深入研究. ...
Mechanosensitive channels protect plastids from hypoo- smotic stress during normal plant growth 1 2012
... 除了SLAC1家族, 植物细胞质膜上还包含张力激活的阴离子通道MSL (mechano-sensitive or stret- ch-activated channels) (Crosgrove and Hedrich, 1991).MSL为典型的张力激活的阴离子通道家族.MSL9和MSL10是最早得到电生理鉴定的MSL家族成员, 定位于细胞质膜, 其激活对质膜张力具有依赖性, 主要选择性介导阴离子的跨膜流动(Haswell et al., 2008; Maksaev and Haswell, 2012).随后有研究表明, 拟南芥MSL10参与植物机械损伤信号反应和茉莉酸的累积(Veley et al., 2014; Zou et al., 2016).此外, 有些拟南芥MSL成员定位在细胞器膜上, 调控细胞器的大小和形状(Veley et al., 2012).根据氨基酸残基序列预测, MSL蛋白存在多个跨膜区, 具备形成离子通道的基本结构特点.而质膜受到拉伸时对MSL的激活是通过机械拉力直接改变MSL蛋白的结构, 使离子通过的孔被拉开.但这基本上还停留在理论推测阶段, 缺乏有力的直接实验证据支持.总体来讲, 目前对MSL家族的生物学功能还了解较少, 其组成的离子通道蛋白结构、感知生物膜张力的分子机理和生物学功能等还有待深入研究. ...
The long and arduous road to CRAC 1 2007
... 植物体内存在大量的钙元素.一方面钙作为细胞壁等结构组分发挥作用; 另一方面, 离子形式的钙(Ca2+)可以作为信号分子参与各种生物学反应和信号传递过程, 后者则备受关注.胞内Ca2+信号以细胞质Ca2+浓度的升高和震荡形式出现.一般认为, 胞外Ca2+内流是细胞质Ca2+信号产生所需Ca2+的主要来源, 而质膜内向Ca2+通道是胞外Ca2+内流的主要渠道.同时, Ca2+通道的活性变化通过控制胞外Ca2+内流的节奏直接调控胞内Ca2+震荡, 发挥Ca2+信号上游调控因子的作用.鉴于此, 质膜Ca2+通道受到广泛关注, 并成为动植物各信号领域的重点研究对象.科学家首先在动物细胞中发现多个Ca2+通道家族, 主要包括受环核苷酸cAMP和cGMP激活的CNG (cyclic nucleotide-gated)家族、受谷氨酸门控的GLR (glutamate receptor)家族、TRPC (transient receptor potential channel)和受胞内Ca2+库倒空激活的CRACM1/Orai1 (Zagotta and Siegelbaum, 1996; Vig and Kinet, 2007; Sobolevsky, 2015; Ambudkar et al., 2017).通过序列比对, 多个动物细胞质膜Ca2+通道的同源基因家族先后在植物尤其是模式植物拟南芥基因组中被鉴定出来, 主要包括CNGC (cyclic nucleotide- gated channel)、GLR (glutamate receptor-like)、高渗敏感的OSCA (hyperosmolality-gated calcium- permeable channel)家族和低渗敏感的张力激活离子通道MCA (mechano-sensitive channel) (M?ser et al., 2001; Ward et al., 2009; Iida et al., 2014; Yuan et al., 2014). ...
AtKC1 and CIPK23 synergistically modulate AKT1-mediated low-potassium stress responses in Arabidopsis 1 2016
... Shaker家族离子通道的活性受多种调控因子的影响.首先, 细胞内和细胞外K+浓度对K+in和K+out通道活性均有明显影响.这种影响可能是基于K+从高浓度向低浓度方向流动的基本化学特性.酸性环境可以显著抑制拟南芥花粉原生质体质膜K+in通道活性, 而碱性条件则具有一定的激活作用(Fan et al., 2001, 2003).这种作用可能通过影响K+in通道的某些氨基酸残基电离状态和离子通道蛋白结构变化来实现.其次, K+in四聚体的亚基组成形式本身就是一种通道活性调控方式.在非洲爪蟾卵母细胞中, 以不同的排列组合共表达2个不同的拟南芥Shaker家族K+in通道基因, 可以利用电生理技术记录到具有不同电生理特性的K+in通道电流信号(Xicluna et al., 2007; Lebaudy et al., 2010).而AtKC1作为抑制亚基, 通过重组进入K+in四聚体发挥对K+in通道的抑制作用, 是K+in通道的重要负调控因子.K+in四聚体组成形式的变化可能通过影响K+in通道蛋白离子通道孔的结构而影响其离子通道活性.SNARE蛋白SYP121不仅可以影响KAT1等K+in通道的亚细胞定位, 而且可使KAT1内化离开细胞质膜, 从而抑制全细胞K+in通道的电流大小(Sutter et al., 2006, 2007).进一步研究表明, SNARE还可以通过与AtKC1互作调控K+in通道的蛋白活性(Honsbein et al., 2009; Grefen et al., 2010; Jeanguenin et al., 2011).由此可知, SNARE作为K+in通道的上游调控因子可通过对K+in四聚体组成形式的影响发挥调控作用.蛋白激酶和磷酸酶参与调控多种蛋白的功能, 包括内向和外向质膜K+通道, 比如CPK (calcium-dependent protein kinase)家族的CPK33对GORK的激活作用和PP2CA对GORK的抑制作用(Lefoulon et al., 2016; Corratgé-Faillie et al., 2017).K+in通道可以被多种蛋白激酶磷酸化所抑制, 包括CPK家族和SnRK (Rnf1-related protein kinase)家族的多个蛋白激酶(Sato et al., 2009, 2010; Ronzier et al., 2014).此外, AKT1等多个质膜K+in通道的活性还受Ca2+信号及其与CBL-CIPK组成的信号链的正调控(Li et al., 2006; Cheong et al., 2007; Zhang et al., 2010; Ren et al., 2013; Behera et al., 2017).而PP2CA对AKT1的去磷酸化可以与Ca2+-CBL-CIPK信号链协同调控AKT1的活性(Lan et al., 2011).有趣的是, ABA甚至可以直接激活GORK的离子通道活性(Ooi et al., 2017).K+通道的调控因子众多, 这些调控因子并非各自单独发挥作用, 而是彼此协同发挥作用.AtKC1和Ca2+-CBL-CIPK23信号链协同调控AKT1就是一个典型的例子(Wang et al., 2016). ...
Potassium channel α-subunit AtKC1 negatively regulates AKT1-me- diated K + uptake in Arabidopsis roots under low-K + stress 3 2010
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
... ; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
... 目前, 对拟南芥Shaker家族的组织分布和功能研究得较为深入全面; 而对包括水稻(Oryza sativa)和玉米(Zea mays)在内的其它植物Shaker家族的研究相对较少.已有的研究鉴定出多个参与K+吸收、转运和气孔运动的玉米和水稻Shaker同源基因.其与拟南芥同源蛋白相比较有很多类似之处, 但也有不同.例如, 玉米KZM2通过与KZM3组成异源多聚体, 抑制K+in通道的活性和气孔运动(Gao et al., 2017).而水稻OsKAT3通过与OsKAT2互作抑制OsKAT3的活性(Hwang et al., 2013).这种K+通道两个同源蛋白之间的互作和调控方式与拟南芥中的AtKC1对AKT1和KAT1等的抑制作用类似(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).再比如, 水稻KAT2与拟南芥KAT2均作为K+in通道参与光照诱导的气孔开放, 但拟南芥气孔保卫细胞质膜K+in通道编码基因数目明显多于水稻(Pilot et al., 2001; Moon et al., 2017).对于外向K+通道而言, 拟南芥SKOR分布于根部, 负责K+向木质部的卸载, 而GORK则主要参与拟南芥的气孔关闭, 二者的组织分布和生物学功能明显不同.而水稻的同一个Shaker蛋白同时参与气孔关闭和K+营养在根部的卸载(Nguyen et al., 2017).因此, 加强对更多植物, 尤其是重要作物中Shaker家族的研究, 将会促进人们对植物吸收、运输和利用K+的分子机理的深入理解. ...
CNGC2 is a Ca 2+ influx channel that prevents accumulation of apoplastic Ca 2+ in the leaf 1 2017
... 拟南芥CNGC家族由20个成员组成, 其蛋白结构与动物CNG同源蛋白和植物Shaker家族成员颇为类似(图1A), 也具有6个跨膜区, 位于质膜内侧的1个N自由端和1个C自由端, 在第4与第5跨膜区之间有1个孔形成所必需的自由环状结构(K?hler et al., 1999; Kaupp and Seifert, 2002; Matulef and Zagotta, 2003).但由CNGC组成的离子通道具体包含几个亚基尚不清楚.最近的研究表明, 百脉根(Lotus corniculatus) CNGC组成的离子通道与Shaker家族类似, 也是四聚体, 由4个相同或不同的CNGC亚基共同形成1个有功能的Ca2+通道蛋白复合体, 且不同的亚基组成影响植物表型(Chiasson et al., 2017).考虑到CNGC具有较高的序列相似性, 推测包括拟南芥在内的植物CNGC组成的质膜离子通道可能都是四聚体.关于CNGC家族各成员单独形成的同源四聚体离子通道对离子的选择性和通透性, 早期的研究表明, 拟南芥CNGC1和CNGC2允许K+通过, 是K+通透性离子通道(K?hler et al., 1999; Leng et al., 1999, 2002).但最新研究表明, CNGC2也对Ca2+具有明显的通透性, 介导Ca2+跨质膜内流(Wang et al, 2017).CNGC10对Ca2+和Mg2+均有明显的通透能力, 并可能通过二价阳离子进一步调控K+的跨膜运输(Guo et al., 2010).而CNGC7、8、9、10、16和18均为Ca2+通透性的二价阳离子通道, 对单价阳离子没有明显的通透性(Gao et al., 2014, 2016).由此表明, CNGC家族各成员的离子选择性并不一致, 且有些成员对不同离子的选择性较弱.从第1个植物CNGC家族基因被克隆(Schuurink et al., 1998)至今已有20年, 关于其生物学功能也已经有很多报道.在作为质膜Ca2+通道发挥生物学功能方面, CNGC2通过调控Ca2+内流调控质外体Ca2+的动态、叶片衰老和介导茉莉酸信号传递(Ma et al., 2010; Lu et al., 2016; Wang et al., 2017); CNGC2与CNGC4共同参与植物的病原防御反应(Chin et al., 2013); CNGC7、8、9、10、16和18均在成熟花粉和花粉管中表达, 但只有CNGC7、8、16和18被确定参与花粉萌发及花粉管的极性生长与导向调控(Tunc-Ozdemir et al., 2013a, 2013b; Gao et al., 2016; Gu et al., 2017); CNGC2和CNG- C6通过介导Ca2+内流参与拟南芥的热激反应(Finka et al., 2012; Gao et al., 2012); CNGC10参与拟南芥的耐盐反应(Guo et al., 2008; Jin et al., 2015); CNGC17参与植物磺肽素信号的传递过程(Ladwig et al., 2015); CNGC11/12参与植物衰老和细胞程序性死亡过程(Urquhart et al., 2007, 2011); 而CNGC14参与拟南芥根的重力反应(Shih et al., 2015).关于拟南芥CNGC家族的更多功能可以参考刘海娇等(2015)的专题评述. ...
Potassium transport and signaling in higher plants 1 2013
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
Plant ion channels: gene families, physiology, and functional genomics ana- lyses 1 2009
... 植物体内存在大量的钙元素.一方面钙作为细胞壁等结构组分发挥作用; 另一方面, 离子形式的钙(Ca2+)可以作为信号分子参与各种生物学反应和信号传递过程, 后者则备受关注.胞内Ca2+信号以细胞质Ca2+浓度的升高和震荡形式出现.一般认为, 胞外Ca2+内流是细胞质Ca2+信号产生所需Ca2+的主要来源, 而质膜内向Ca2+通道是胞外Ca2+内流的主要渠道.同时, Ca2+通道的活性变化通过控制胞外Ca2+内流的节奏直接调控胞内Ca2+震荡, 发挥Ca2+信号上游调控因子的作用.鉴于此, 质膜Ca2+通道受到广泛关注, 并成为动植物各信号领域的重点研究对象.科学家首先在动物细胞中发现多个Ca2+通道家族, 主要包括受环核苷酸cAMP和cGMP激活的CNG (cyclic nucleotide-gated)家族、受谷氨酸门控的GLR (glutamate receptor)家族、TRPC (transient receptor potential channel)和受胞内Ca2+库倒空激活的CRACM1/Orai1 (Zagotta and Siegelbaum, 1996; Vig and Kinet, 2007; Sobolevsky, 2015; Ambudkar et al., 2017).通过序列比对, 多个动物细胞质膜Ca2+通道的同源基因家族先后在植物尤其是模式植物拟南芥基因组中被鉴定出来, 主要包括CNGC (cyclic nucleotide- gated channel)、GLR (glutamate receptor-like)、高渗敏感的OSCA (hyperosmolality-gated calcium- permeable channel)家族和低渗敏感的张力激活离子通道MCA (mechano-sensitive channel) (M?ser et al., 2001; Ward et al., 2009; Iida et al., 2014; Yuan et al., 2014). ...
CO- RNICHON sorting and regulation of GLR channels underlie pollen tube Ca 2+ homeostasis 1 2018
... GLR家族是受多种氨基酸门控的离子通道, 但其通道门控特点并非如其名称仅为谷氨酸所门控, 而是受多种氨基酸影响.1998年, 第1个GLR被克隆出来(Lam et al., 1998), 人们开始对GLR进行研究.研究表明, 拟南芥GLR1和GLR2的N端位于细胞质膜外侧, C端位于细胞质膜内侧, 中间包含4个跨膜区, 但第2个跨膜区并不能完整地跨过细胞质膜(Lam et al., 1998) (图2).这种结构特点明显不同于Shaker和CNGC家族.GLR组成的质膜离子通道的亚基组成、多数GLR离子通道的离子选择性等基本特性, 以及GLR离子通道的活性调控机理目前还不甚清楚.但已经解析出动物GLR四聚体的通道蛋白结构(Price et al., 2012).据此推测, 植物GLR组成的通道蛋白结构可能与动物类似.GLR在植物体内具有多种生物学功能, 包括参与调控花粉管的极性生长(Michard et al., 2011; Wudick et al., 2018)、铝诱导的微管解聚和质膜去极化(Sivaguru et al., 2003)、离子运输(McAinsh and Pittman, 2009)、碳氮代谢(Kang and Turano, 2003)、ABA合成和耐旱反应(Kang et al., 2004; Lu et al., 2014)以及机械损伤信号的长距离传输(Mousavi et al., 2013)等.总体而言, 目前对GLR家族的研究还不够深入.关于植物GLR家族更为详细的介绍可以参阅何明杰等(2016)的综述. ...
Increased functional diversity of plant K + channels by preferential heteromerization of the shaker-like subunits AKT2 and KAT2 2 2007
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
... Shaker家族离子通道的活性受多种调控因子的影响.首先, 细胞内和细胞外K+浓度对K+in和K+out通道活性均有明显影响.这种影响可能是基于K+从高浓度向低浓度方向流动的基本化学特性.酸性环境可以显著抑制拟南芥花粉原生质体质膜K+in通道活性, 而碱性条件则具有一定的激活作用(Fan et al., 2001, 2003).这种作用可能通过影响K+in通道的某些氨基酸残基电离状态和离子通道蛋白结构变化来实现.其次, K+in四聚体的亚基组成形式本身就是一种通道活性调控方式.在非洲爪蟾卵母细胞中, 以不同的排列组合共表达2个不同的拟南芥Shaker家族K+in通道基因, 可以利用电生理技术记录到具有不同电生理特性的K+in通道电流信号(Xicluna et al., 2007; Lebaudy et al., 2010).而AtKC1作为抑制亚基, 通过重组进入K+in四聚体发挥对K+in通道的抑制作用, 是K+in通道的重要负调控因子.K+in四聚体组成形式的变化可能通过影响K+in通道蛋白离子通道孔的结构而影响其离子通道活性.SNARE蛋白SYP121不仅可以影响KAT1等K+in通道的亚细胞定位, 而且可使KAT1内化离开细胞质膜, 从而抑制全细胞K+in通道的电流大小(Sutter et al., 2006, 2007).进一步研究表明, SNARE还可以通过与AtKC1互作调控K+in通道的蛋白活性(Honsbein et al., 2009; Grefen et al., 2010; Jeanguenin et al., 2011).由此可知, SNARE作为K+in通道的上游调控因子可通过对K+in四聚体组成形式的影响发挥调控作用.蛋白激酶和磷酸酶参与调控多种蛋白的功能, 包括内向和外向质膜K+通道, 比如CPK (calcium-dependent protein kinase)家族的CPK33对GORK的激活作用和PP2CA对GORK的抑制作用(Lefoulon et al., 2016; Corratgé-Faillie et al., 2017).K+in通道可以被多种蛋白激酶磷酸化所抑制, 包括CPK家族和SnRK (Rnf1-related protein kinase)家族的多个蛋白激酶(Sato et al., 2009, 2010; Ronzier et al., 2014).此外, AKT1等多个质膜K+in通道的活性还受Ca2+信号及其与CBL-CIPK组成的信号链的正调控(Li et al., 2006; Cheong et al., 2007; Zhang et al., 2010; Ren et al., 2013; Behera et al., 2017).而PP2CA对AKT1的去磷酸化可以与Ca2+-CBL-CIPK信号链协同调控AKT1的活性(Lan et al., 2011).有趣的是, ABA甚至可以直接激活GORK的离子通道活性(Ooi et al., 2017).K+通道的调控因子众多, 这些调控因子并非各自单独发挥作用, 而是彼此协同发挥作用.AtKC1和Ca2+-CBL-CIPK23信号链协同调控AKT1就是一个典型的例子(Wang et al., 2016). ...
A protein kinase, interacting with two calcineurin B-like proteins, regulates K + transporter AKT1 in Arabi- dopsis 1 2006
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
Functional characterization of GhAKT1, a novel Shaker-like K + chan- nel gene involved in K + uptake from cotton ( Gossypium hirsutum) 1 2014
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
The barley anion channel, HvALMT1, has multiple roles in guard cell physiology and grain metabolism 1 2015
... 关于ALMT家族, 最早被鉴定出来的2个ALMT分别是来自小麦(Triticum aestivum)和拟南芥的ALMT1, 因其均受Al诱导且对苹果酸具有明显的通透能力而得名(Sasaki et al., 2004; Hoekenga et al., 2006).ALMT虽然被命名为Al诱导的苹果酸转运子(transporters), 但在结构和功能上都是典型的离子通道(图5), 在质膜和内膜系统均有分布.ALMT蛋白结构目前尚不清楚.根据现有信息, 推测每个ALMT亚基具有6或7个跨膜区, 其C端和N端可能均位于质膜胞质侧, 其N端和跨膜区为形成有功能的离子通道所必需, 而其C端主要对离子通道的活性发挥调控作用(Sharma et al., 2016) (图5).ALMT的离子选择性也不仅限于苹果酸, 而是对苹果酸、草酸、NO3-和Cl-等多种阴离子物质均有通透能力, 且离子流动的方向也不一致(Sharma et al., 2016).离子通道对离子的选择性及对各种离子的通透能力和通透方向往往由其自身的蛋白拓扑结构及一些关键氨基酸残基决定.由于ALMT组成的离子通道蛋白的拓扑结构还不清楚, 目前尚无法从蛋白结构层面详细解释其离子选择性和对各种离子的相对通透能力(Sharma et al., 2016).ALMT在植物各种组织中广泛分布, 其生物学功能多样.其中, 分布于植物根细胞质膜的ALMT1向土壤中分泌苹果酸的作用先后在多种植物根部被观测到, 包括小麦、拟南芥、油菜(Brassica napus)、玉米、大豆(Glycine max)和绒毛草(Holcus lanatus)等(Sasaki et al., 2004; Hoekenga et al., 2006; Ligaba et al., 2006, 2012; Fontecha et al., 2007; Chen et al., 2013; Liang et al., 2013).由植物分泌并进入土壤的苹果酸有2个作用, 其一是与土壤中处于离子状态的铝结合, 将其螯合和固定, 从而使植物免受铝毒伤害; 其二是苹果酸使土壤pH值降低, 可以有效提升土壤中离子状态磷元素的浓度, 从而促进植物对磷营养的吸收.此外, 研究表明, 豆科植物ALMT1和ALMT4基因在根瘤维管束细胞表达, 并受flg22诱导, 其蛋白通过介导有机酸的跨膜流动参与植物与微生物的互作(Rudrappa et al., 2008; Lakshmanan et al., 2012; Sukweenadhi et al., 2015; Takanashi et al., 2016).有意思的是, 玉米ALMT1虽然也在根组织中分布, 但却对苹果酸等有机酸没有明显的通透能力, 而主要介导Cl-、NO3-和SO42-等阴离子营养物质的跨质膜内流和外流(Pi?eros et al., 2008).这明显不同于其它多种植物的ALMT1.在自然情况下, 分布于植物根细胞质膜的ALMT编码基因的表达可以同时受多种信号的调控, 包括土壤中的磷和铝、土壤酸化、植物-微生物互作、体内激素、活性自由基的产生以及GABA (γ-aminobutyric acid)信号的调控(Kobayashi et al., 2013; Liang et al., 2013; Ramesh et al., 2015).在蛋白水平上, Al和苹果酸可能通过直接与ALMT1通道蛋白结合, 提高ALMT1的离子通道活性.在植物地上部, 多个ALMT成员在气孔保卫细胞中分布, 并参与调控气孔运动.其中, 拟南芥ALMT6和ALMT9主要定位于气孔保卫细胞液泡膜上, 且AtALMT9的离子通道活性受细胞质苹果酸含量的调控(Kovermann et al., 2007; Meyer et al., 2011; De Angeli et al., 2013b).苹果(Malus pumila)果实细胞液泡膜定位的ALMT6通过介导苹果酸在液泡中的累积进而影响苹果的酸性物质含量和口味; 葡萄(Vitis vinifera)果实细胞液泡膜定位的ALMT9则通过介导苹果酸和酒石酸的跨液泡膜流动及在液泡中的积累来影响用其所酿酒的品质和产量(Bai et al., 2012; De Angeli et al., 2013a).而拟南芥ALMT12作为质膜阴离子通道, 通过介导苹果酸的外流参与ABA等诱导的气孔关闭运动(Meyer et al., 2010; Sasaki et al., 2010).除了这些有特定组织和亚细胞分布的ALMT外, 也有ALMT组织分布比较复杂, 同一个蛋白在不同组织中发挥不同的生物学功能.例如, 大麦(Hordeum vulgare) ALMT1同时分布于气孔保卫细胞、根和种子中, 分别参与气孔关闭运动、植物生长发育和种子萌发(Gru- ber et al., 2010; Xu et al., 2015).这些研究不仅表明ALMT家族组成的离子通道在离子选择性和方向性方面具多样化, 也表现出其组织分布、亚细胞定位和生物学功能的复杂性. ...
S-type anion channel SLAC1’s homologues inhibit inward potassium channels AKT2 and KAT2 in Arabidopsis 1 2017
... S: The transmembrane domain; EF: EF-hand Ca2+ binding domain; N/C termini: The amino terminal and carboxyl terminal Figure 3 10.11983/CBB18022.F004
拟南芥OSCA家族通道蛋白结构(<xref ref-type="bibr" rid="b154">改自Yuan et al., 2014</xref>)
The putative protein structure of an OSCA member from Arabidopsis (<xref ref-type="bibr" rid="b154">modified from Yuan et al., 2014</xref>)Figure 4 鉴于Ca2+信号的广泛存在和重要生物学功能, 寻找其上游调控因子一直是这一研究领域的重要任务.其中, 质膜Ca2+通道是最重要的一大类Ca2+信号上游调控因子.循此思路, 很多研究组直接从寻找Ca2+通道和揭示Ca2+通道候选蛋白的生物学功能入手进行研究.但迄今为止依然有很多信号传递过程中Ca2+信号和Ca2+通道之间无法实现一对一配对, 即Ca2+信号发挥重要作用的生物学过程或信号传递链中, 负责此类Ca2+信号产生的Ca2+通道编码基因往往并不清楚, 且不能阐明一些已知生物学功能的质膜Ca2+通道调控Ca2+信号的分子机制.因此, 这一领域还有诸多科学问题等待解答. ...
... The putative protein structure of an OSCA member from Arabidopsis (modified from Yuan et al., 2014) Figure 4 鉴于Ca2+信号的广泛存在和重要生物学功能, 寻找其上游调控因子一直是这一研究领域的重要任务.其中, 质膜Ca2+通道是最重要的一大类Ca2+信号上游调控因子.循此思路, 很多研究组直接从寻找Ca2+通道和揭示Ca2+通道候选蛋白的生物学功能入手进行研究.但迄今为止依然有很多信号传递过程中Ca2+信号和Ca2+通道之间无法实现一对一配对, 即Ca2+信号发挥重要作用的生物学过程或信号传递链中, 负责此类Ca2+信号产生的Ca2+通道编码基因往往并不清楚, 且不能阐明一些已知生物学功能的质膜Ca2+通道调控Ca2+信号的分子机制.因此, 这一领域还有诸多科学问题等待解答. ...
Structure and function of cyclic nucleotide-gated channels 1 1996
... 植物体内存在大量的钙元素.一方面钙作为细胞壁等结构组分发挥作用; 另一方面, 离子形式的钙(Ca2+)可以作为信号分子参与各种生物学反应和信号传递过程, 后者则备受关注.胞内Ca2+信号以细胞质Ca2+浓度的升高和震荡形式出现.一般认为, 胞外Ca2+内流是细胞质Ca2+信号产生所需Ca2+的主要来源, 而质膜内向Ca2+通道是胞外Ca2+内流的主要渠道.同时, Ca2+通道的活性变化通过控制胞外Ca2+内流的节奏直接调控胞内Ca2+震荡, 发挥Ca2+信号上游调控因子的作用.鉴于此, 质膜Ca2+通道受到广泛关注, 并成为动植物各信号领域的重点研究对象.科学家首先在动物细胞中发现多个Ca2+通道家族, 主要包括受环核苷酸cAMP和cGMP激活的CNG (cyclic nucleotide-gated)家族、受谷氨酸门控的GLR (glutamate receptor)家族、TRPC (transient receptor potential channel)和受胞内Ca2+库倒空激活的CRACM1/Orai1 (Zagotta and Siegelbaum, 1996; Vig and Kinet, 2007; Sobolevsky, 2015; Ambudkar et al., 2017).通过序列比对, 多个动物细胞质膜Ca2+通道的同源基因家族先后在植物尤其是模式植物拟南芥基因组中被鉴定出来, 主要包括CNGC (cyclic nucleotide- gated channel)、GLR (glutamate receptor-like)、高渗敏感的OSCA (hyperosmolality-gated calcium- permeable channel)家族和低渗敏感的张力激活离子通道MCA (mechano-sensitive channel) (M?ser et al., 2001; Ward et al., 2009; Iida et al., 2014; Yuan et al., 2014). ...
S-type anion channels SLAC1 and SLAH3 function as essential negative regulators of Inward K + channels and stomatal opening in Arabidopsis 2 2016
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
Shaker-like potassium channels in Populus, regulated by the CBL-CIPK signal transduction pathway, increase tolerance to low-K + stress. 2 2010
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
... Shaker家族离子通道的活性受多种调控因子的影响.首先, 细胞内和细胞外K+浓度对K+in和K+out通道活性均有明显影响.这种影响可能是基于K+从高浓度向低浓度方向流动的基本化学特性.酸性环境可以显著抑制拟南芥花粉原生质体质膜K+in通道活性, 而碱性条件则具有一定的激活作用(Fan et al., 2001, 2003).这种作用可能通过影响K+in通道的某些氨基酸残基电离状态和离子通道蛋白结构变化来实现.其次, K+in四聚体的亚基组成形式本身就是一种通道活性调控方式.在非洲爪蟾卵母细胞中, 以不同的排列组合共表达2个不同的拟南芥Shaker家族K+in通道基因, 可以利用电生理技术记录到具有不同电生理特性的K+in通道电流信号(Xicluna et al., 2007; Lebaudy et al., 2010).而AtKC1作为抑制亚基, 通过重组进入K+in四聚体发挥对K+in通道的抑制作用, 是K+in通道的重要负调控因子.K+in四聚体组成形式的变化可能通过影响K+in通道蛋白离子通道孔的结构而影响其离子通道活性.SNARE蛋白SYP121不仅可以影响KAT1等K+in通道的亚细胞定位, 而且可使KAT1内化离开细胞质膜, 从而抑制全细胞K+in通道的电流大小(Sutter et al., 2006, 2007).进一步研究表明, SNARE还可以通过与AtKC1互作调控K+in通道的蛋白活性(Honsbein et al., 2009; Grefen et al., 2010; Jeanguenin et al., 2011).由此可知, SNARE作为K+in通道的上游调控因子可通过对K+in四聚体组成形式的影响发挥调控作用.蛋白激酶和磷酸酶参与调控多种蛋白的功能, 包括内向和外向质膜K+通道, 比如CPK (calcium-dependent protein kinase)家族的CPK33对GORK的激活作用和PP2CA对GORK的抑制作用(Lefoulon et al., 2016; Corratgé-Faillie et al., 2017).K+in通道可以被多种蛋白激酶磷酸化所抑制, 包括CPK家族和SnRK (Rnf1-related protein kinase)家族的多个蛋白激酶(Sato et al., 2009, 2010; Ronzier et al., 2014).此外, AKT1等多个质膜K+in通道的活性还受Ca2+信号及其与CBL-CIPK组成的信号链的正调控(Li et al., 2006; Cheong et al., 2007; Zhang et al., 2010; Ren et al., 2013; Behera et al., 2017).而PP2CA对AKT1的去磷酸化可以与Ca2+-CBL-CIPK信号链协同调控AKT1的活性(Lan et al., 2011).有趣的是, ABA甚至可以直接激活GORK的离子通道活性(Ooi et al., 2017).K+通道的调控因子众多, 这些调控因子并非各自单独发挥作用, 而是彼此协同发挥作用.AtKC1和Ca2+-CBL-CIPK23信号链协同调控AKT1就是一个典型的例子(Wang et al., 2016). ...
Osmo-sensitive and stretch-activated calcium-permeable channels in Vicia faba guard cells are regulated by actin dynamics 1 2007
... 早期的研究中, 人们发现植物细胞中存在一类受质膜张力激活的Ca2+信号, 从而预测在植物细胞质膜上存在张力激活类质膜Ca2+通道(Crosgrove and Hedrich, 1991; Dutta and Robinson, 2004; Qi et al., 2004; Zhang et al., 2007).多年来, 人们一直没有鉴定到此类受机械或渗透张力激活的离子通道编码基因.近年来, 科学家找到了2类对质膜张力敏感的离子通道, 其一是受低渗条件激活, 通过介导胞外Ca2+内流发挥功能的MCA1和MCA2 (Iida et al., 2014); 其二是对高渗胁迫敏感的OSCA家族(Hou et al., 2014; Yuan et al., 2014).MCA1和MCA2蛋白只有1个跨膜区, 其N端较短且位于细胞质膜外侧, 而C端较长且位于质膜内侧(图3A).研究表明, MCA1和MCA2的N端和跨膜区为其离子通道组成及活性所必需, 而C端则对其离子通道活性发挥调控作用(Kam- ano et al., 2015).MCA1和MCA2组成的离子通道也为四聚体结构, 张力和低渗等刺激通过拉伸质膜改变该离子通道蛋白的结构, 从而使通道被激活, 引起胞外Ca2+内流和细胞质Ca2+水平升高(图3B).MCA1的拟南芥突变体根无法插入较硬的固体培养基(Nakagawa et al., 2007), 说明其在感知机械压力方面发挥作用.最近的研究表明, MCA1和MCA2还作为质膜Ca2+通道介导拟南芥的耐冷反应(Mori et al., 2018).与MCA1和MCA2相反, OSCA是对高渗敏感的质膜离子通道, 可以被细胞外的高渗处理激活(Yuan et al., 2014).拟南芥OSCA家族的蛋白结构与MCA1和MCA2大不相同, 其每个亚基均具有多达9个跨膜区, 在其第8和第9跨膜区之间具有1个孔形成所需的环状结构(Yuan et al., 2014) (图4).OSCA与MCA1和MCA2类似之处在于其亚基N端位于质膜的胞外侧, 而C端则位于质膜胞质侧(Yuan et al., 2014) (图4).OSCA1允许多种离子通过, 其对各种离子的通透能力大小排序为K+>Ba2+≈Ca2+>Na+=Mg2+=Cs+ (Yuan et al., 2014).目前, 人们对质膜张力激活的质膜离子通道的生物学功能了解较少, 但其对渗透敏感的特点暗示此类质膜离子通道可能通过对质膜张力变化做出反应.对低渗敏感的MCA1和MCA2与对高渗敏感的OSCA家族有可能作为一对质膜张力和渗透敏感元件, 协同配合发挥作用, 以共同应对渗透环境变化.一方面, 其可以通过影响胞内Ca2+信号而参与耐盐、耐旱和气孔运动等与细胞渗透和膨压调节密切相关的生物学过程; 另一方面, OSCA对K+的通透能力也表明其可能同时介导Ca2+和K+的跨膜流动而参与这些生物学过程. ...
Ca 2+-dependent protein kinase11 and 24 modulate the activity of the inward rectifying K + channels in Arabi- dopsis pollen tubes 1 2013
... 钾是一种重要的植物离子营养物质, 以离子形式(K+)被植物吸收, 并以离子形式在植物细胞内存在.在正常情况下, 植物细胞内K+浓度可以达到或超过100 mmol?L-1 (Clarkson and Hanson, 1980), 有些细胞和细胞器内的K+浓度甚至可以达到数百毫摩尔的水平.例如, 处于气孔开放状态的鸭跖草(Commelina communis)气孔保卫细胞内K+浓度可达300 mmol?L-1以上(MacRobbie and Lettau, 1980).因此, K+是植物细胞内最主要的一种阳离子渗透物质, 是各种生化反应得以发生的基础环境条件.植物从土壤中吸收K+, 以及K+在植物生长发育和逆境反应中被频繁地跨膜运输, 主要依赖于K+通道的介导.模式植物拟南芥(Arabidopsis thaliana)的质膜K+通道主要来自Shaker家族.Shaker家族离子通道首先在果蝇(Drosophila melanogaster)中被发现, 因早期发现的一个果蝇突变体颤动表型而得名(Jan and Jan, 1997).通过序列比对, 科学家在拟南芥中发现了9个Shaker同源基因, 并延续了Shaker的命名.拟南芥Shaker家族每个成员均具有位于质膜内侧的1个自由N端和1个自由C端、6个跨膜结构域、1个定位于第4跨膜结构域的电压感受器、1个位于第5和第6跨膜结构域之间形成离子通道孔道的结构域和1个可能结合环核苷酸的结构域, 其中孔形成结构域包含1个保守的氨基酸序列GYGD/E (Lebaudy et al., 2007; Sharma et al., 2013) (图1A).根据其功能特点, 可以将Shaker家族划分为5个组(Lebaudy et al., 2007).其中AKT1、AKT5和AKT6/SPIK为第1组; KAT1和KAT2为第2组; AKT2为第3组; AtKC1为第4组; SKOR和GORK组成第5组.除第4组的AtKC1不能单独形成有功能的质膜K+通道外, 其它各Shaker成员均可以单独形成有功能的质膜K+通道.第1和第2两个组的成员均为强内向K+ (K+in)通道, 只允许K+从胞外向胞内方向流动, 一般不允许K+反方向流动.第3组为弱内向性K+通道, 主要介导K+的跨质膜内流, 但也允许K+的缓慢外流.第4组AtKC1虽然不能独自形成有功能的K+通道, 但可以作为内向K+通道的调控亚基发挥负调控作用(Duby et al., 2008; Jeanguenin et al., 2011).第5组为典型的质膜外向K+ (K+out)通道, 只允许K+从胞内向胞外跨质膜流动, 不允许K+跨质膜内流.动物和植物细胞的Shaker家族组成的K+通道(包括内向和外向的K+通道)均为四聚体, 且既可以是同源四聚体, 也可以是异源四聚体, 共同围成一个离子通道的孔, 每个亚基的2个自由端在四聚体的形成中发挥重要作用(MacKinnon, 1991; Daram et al., 1997; Dreyer et al., 2004; Lebaudy et al., 2008a, 2008b; Sharma et al., 2013) (图1B).采用植物基因在非洲爪蟾(Xenopus laevis)卵母细胞中的瞬时表达技术, 科学家结合电生理技术对Shaker家族各成员的基本特性作了详细分析, 确定该家族组成的细胞质膜K+通道的基本特点和功能, 并发现异源四聚体的亚基组成明显影响K+通道的电生理特性(Schroeder et al., 1987; Schroeder, 1988; Schroeder and Hagiwara, 1989; Dreyer et al., 1997; Pilot et al., 2001; Lebaudy et al., 2008a; Geiger et al., 2009; Wang et al., 2010; Jeanguenin et al., 2011).而拟南芥体内实验也证实, Shaker家族成员在植物体内以异源多聚体的形式发挥作用(Lebaudy et al., 2008b).拟南芥Shaker家族广泛分布于植物的各个组织器官和细胞类型, 其生物学功能由其组织分布特点和自身的K+通道特性共同决定.AKT1在植物根中负责K+的吸收和耐盐反应(Fuchs et al., 2005; Xu et al., 2006, 2014; Ardie et al., 2010; Rubio et al., 2010; Zhang et al., 2010; Li et al., 2014).AtKC1作为1个亚基与AKT1共同组成质膜K+通道, 一方面抑制植物对胞外K+的吸收, 另一方面也可以抑制AKT1介导的K+外流和渗漏(Reintanz et al., 2002; Geiger et al., 2009; Wang et al., 2010).因此, AtKC1的缺失可以促进AKT1介导的胞外K+吸收.K+通过AKT1被植物吸收后, 通过质膜外向K+通道SKOR被装载进入木质部, 进一步被长距离运输到植物的地上部(Gaymard et al., 1998; Liu et al., 2006).SKOR的离子通道活性受胞内和胞外K+浓度的调控, 胞内高浓度K+可以有效提高SKOR的K+通道活性, 促进K+向地上部转运, 而对胞外K+的敏感性可以确保其选择性地介导K+的外流(Johansson et al., 2006; Liu et al., 2006).植物以此机制实现对K+的吸收及长距离体内运输的联动与精细调控.关于Shaker家族在植物地上部分的生物学功能, 有研究表明AKT6/SPIK在花粉管中表达, 参与花粉管的极性生长(Mouline et al., 2002; Zhao et al., 2013).另有多个Shaker成员被发现作为质膜K+通道参与气孔运动的调控.光照等通过激活由多个Shaker家族成员组成的气孔保卫细胞质膜K+in通道及其介导的胞外K+内流而诱导气孔开放; ABA和干旱胁迫等通过激活K+out通道及其介导的K+外流驱动气孔关闭.研究表明, 5个Shaker家族成员(即KAT1、KAT2、AKT1、AKT2和AtKC1)都参与拟南芥气孔保卫细胞质膜K+in通道的形成, 其中最主要的是KAT1, 由于其功能缺失导致拟南芥气孔保卫细胞的全细胞内向K+电流减小50%以上(Szyroki et al., 2001).同时, KAT1的功能缺失并不会明显影响拟南芥气孔运动.因此, KAT1曾被认为对于气孔开放运动并不重要(Szyroki et al., 2001).但随后来自多个研究组的研究均证明, Shaker家族多个成员组成的质膜K+in通道及其介导的K+内流为气孔开放运动所必需.KAT1作为最主要的K+in通道组成亚基, 在气孔开放运动中的关键作用毋庸置疑(Kwak et al., 2001; Lebaudy et al., 2008b; Laan- emets et al., 2013; Wang and Wu, 2013; Zhang et al., 2016).由于多个Shaker成员的参与, 气孔保卫细胞内向K+通道主要以异源四聚体的形式存在(Xicluna et al., 2007; Lebaudy et al., 2010).但在气孔关闭运动中发挥重要作用的质膜K+out通道的亚基组成形式则比较简单, 其原因是该K+out通道仅有GORK一个Shaker成员参与组成, 必然为同源四聚体(Ache et al., 2000; Hosy et al., 2003). ...
A gain-of-function mutation in Msl10 triggers cell death and wound-induced hyperaccumulation of jasmonic acid in Arabidopsis 1 2016
... 除了SLAC1家族, 植物细胞质膜上还包含张力激活的阴离子通道MSL (mechano-sensitive or stret- ch-activated channels) (Crosgrove and Hedrich, 1991).MSL为典型的张力激活的阴离子通道家族.MSL9和MSL10是最早得到电生理鉴定的MSL家族成员, 定位于细胞质膜, 其激活对质膜张力具有依赖性, 主要选择性介导阴离子的跨膜流动(Haswell et al., 2008; Maksaev and Haswell, 2012).随后有研究表明, 拟南芥MSL10参与植物机械损伤信号反应和茉莉酸的累积(Veley et al., 2014; Zou et al., 2016).此外, 有些拟南芥MSL成员定位在细胞器膜上, 调控细胞器的大小和形状(Veley et al., 2012).根据氨基酸残基序列预测, MSL蛋白存在多个跨膜区, 具备形成离子通道的基本结构特点.而质膜受到拉伸时对MSL的激活是通过机械拉力直接改变MSL蛋白的结构, 使离子通过的孔被拉开.但这基本上还停留在理论推测阶段, 缺乏有力的直接实验证据支持.总体来讲, 目前对MSL家族的生物学功能还了解较少, 其组成的离子通道蛋白结构、感知生物膜张力的分子机理和生物学功能等还有待深入研究. ...
 ,*中国科学院上海植物生理生态研究所, 中国科学院分子植物科学卓越创新中心, 植物分子遗传国家重点实验室, 上海 200032
,*中国科学院上海植物生理生态研究所, 中国科学院分子植物科学卓越创新中心, 植物分子遗传国家重点实验室, 上海 200032
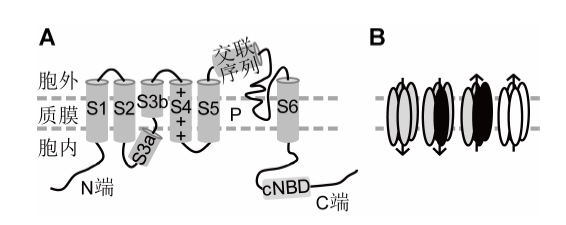 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT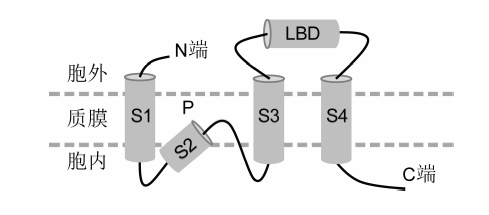 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT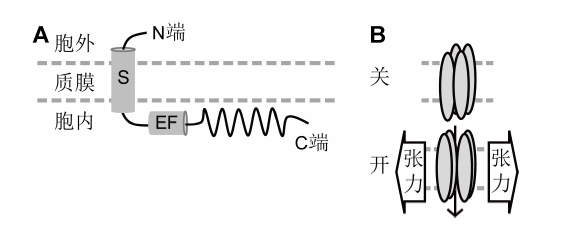 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT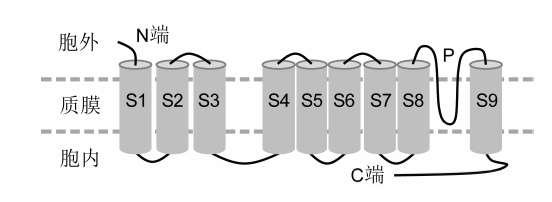 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT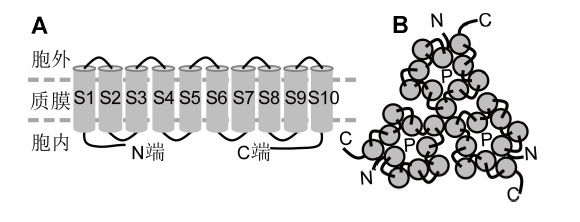 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}