 , ,
, , 引用本文
贡献者
接受日期:2018-09-2网络出版日期:2018-11-1
-->Copyright
2018《植物学报》编辑部
Contributors
History
Received:Online:
摘要:
Abstract:
Key words:
细胞骨架(cytoskeleton)系统主要包括微管(mi- crotubules, MTs)和微丝(actin filaments, F-actin)。微管遍布细胞质中, 为细胞生命活动所必需, 在细胞周期运行、细胞分裂、细胞形态维持、细胞迁移、细胞内物质运输及细胞内信号转导等过程中起重要作用。哺乳动物细胞中, 中心体(centrosomes)是主要的微管组织中心(microtubule organization center, MTOC)。高等植物细胞中没有中心体, 因此植物细胞缺乏类似动物细胞的微管组织中心来形成微管。在植物间期细胞中, 紧贴质膜、呈二维平行排列着植物特异的周质微管骨架(cortical microtubule arrays) (Ehrhardt, 2008; Wasteneys and Ambrose, 2009; Shaw, 2013)。周质微管骨架可以为细胞壁纤维素微纤丝合成酶复合体(cellulose synthase complexes)的囊泡转运提供轨道, 从而调控细胞的生长和形态建成(Paredez et al., 2006; McFarlane et al., 2014; Watanabe et al., 2015; Li et al., 2016)。另外, 微管骨架具有高度动态的特性, 不断进行活跃的重构, 从而影响植物生长发育及响应环境信号(Nick, 2013)。这种动态性包括聚合(polymerization)、解聚(depoly- merization)、切割(severing)、集束(bundling)及成 核(nucleation)等过程, 通过数百种来自不同家族 的微管结合蛋白(microtubule-associated proteins, MAPs)实现。目前, 人们对植物细胞微管骨架组装机理及其动态构象精确调控机制的认识还十分有限。最近, 中国科学家以拟南芥(Arabidopsis thaliana)为材料, 在解析微管精准切割及周质微管骨架动态重构调控机制方面取得了突破性进展(Wang et al., 2017, 2018)。
在微管阵列的动态转换过程中, 微管切割(microtubule severing)是非常重要的环节。微管切割由Katanin蛋白复合体完成, 该复合体包含1个60 kDa的具有ATPase活性的催化亚基p60, 以及1个80 kDa的包含WD-40重复基序的调节亚基p80 (McNally and Vale, 1993; Roll-Mecak and McNally, 2010; Sharp and Ross, 2012)。在体外, p60可以对单根微管的任意位点随机切割。然而在拟南芥细胞内, 微管切割仅特异性地发生在微管交叉(microtubule crossover)位点及新生微管成核(microtubule nucleation)位点, 这表明在细胞内一定存在微管切割的精确调控机制。然而, 对于Katanin在细胞内识别微管交叉和新生微管分支成核位点, 然后对微管进行切割的分子机制尚不清楚。
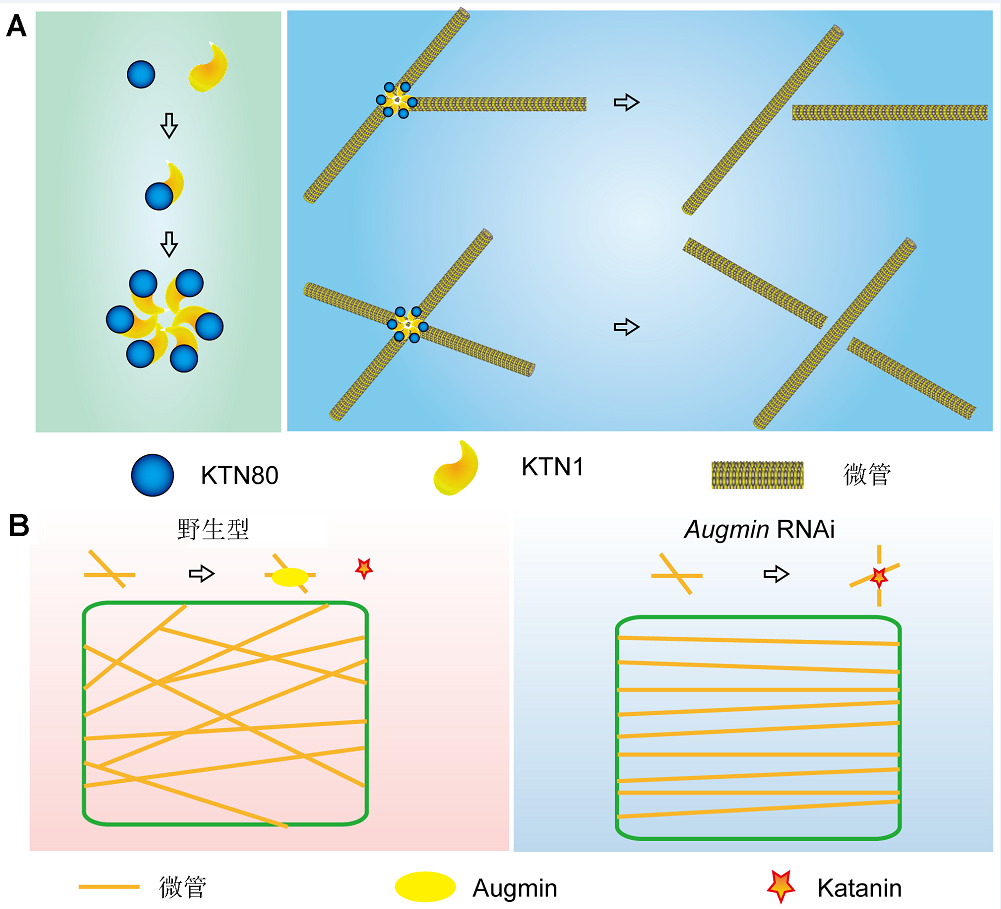
中国科学院微生物研究所孔照胜研究组综合利用遗传学、细胞生物学和生物化学等手段对Katanin蛋白复合体进行了研究, 发现拟南芥中Katanin p80亚基KTN80家族4个成员均在微管的精确切割过程中起作用, 并且功能冗余。KTN80和p60亚基KTN1以异源二聚体的形式存在于细胞质中。当发生微管切割作用时, KTN80可将KTN1-KTN80异源二聚体导向至微管切割位点; 之后, KTN1介导该异源二聚体的六聚体化, 形成十二聚体的环状超复合体, 并识别特异的微管构象进而完成切割(图1A)。
图 1
Figure 1
下载原图ZIP
生成PPT
图 1
Katanin和Augmin在植物微管(MTs)动态构象中的功能模式图(改自Wang et al., 2017, 2018)
(A) 拟南芥Katanin蛋白复合体的组装及其微管切割机制。Katanin蛋白复合体由p60亚基KTN1和p80亚基KTN80组成异源二聚体, 最终形成十二聚体的环状超复合体(左图)。KTN80可将KTN1-KTN80异源二聚体导向至微管切割位点; 而KTN1介导形成KTN1- KTN80环状超复合体, 并识别特异的微管构象、完成切割(右图)。(B) Augmin定位在微管交叉点上, 并且通过抑制Katanin的切割和稳定微管交叉构象, 来调控微管动态重组与细胞形态建成(左图)。与野生型相比, 在Augmin的基因沉默突变体中(amiR-AUG6), 抑制Augmin活性能使微管切割的频率加快并形成更多的平行微管阵列(右图)。
Figure 1
Schematic models of Katanin and Augmin functions on microtubules (MTs) organization in plants (modified from Wang et al., 2017, 2018)
(A) The formation of Katanin complexes and MTs severing in Arabidopsis cells. Katanin is composed of a p60 subunit KTN1 and a p80 subunit KTN80, which exists as a KTN1-KTN80 heterodimer in the cytosol (left panel). KTN80 determines the precise targeting of KTN1-KTN80 heterodimers to MTs crossover and branching nucleation sites. KTN1 further triggers the oligomerization of the mixed types of KTN1-KTN80 heterodimers that sense MT geometry to confer precise MT severing (right panel); (B) Augmin functions in regulating microtubule arrays by preventing katanin-mediated microtubule severing and maintaining the stability of crossovers (left panel). In the Augmin knockdown mutant (amiR-AUG6), a significantly higher microtubule severing frequency occurs and a greater proportion of aligned microtubule arrays (right panel) compared to the fine-network microtubule architectures observed in wild-type (left panel).
另外, 孔照胜研究组还对拟南芥Augmin蛋白复合体参与植物微管骨架的动态重构进行了长期深入的研究。他们发现, 在植物间期细胞中Augmin蛋白复合体能募集γ-TuRC蛋白复合体(γ-tubulin complex)到微管上, 从而产生新的成核位点, 并起始合成新的微管。该研究揭示了Augmin复合体控制植物细胞微管起始组装的机制(Liu et al., 2014)。然而, 仍有一些根本性问题有待解决。例如, 除微管成核外, Augmin是否还有其它的功能?
近期, 孔照胜研究组发现Augmin复合体倾向于定位在微管交叉位点, 一部分Augmin介导微管成核位点的形成, 另外一大部分则用于抑制Katanin的切割并维持微管交叉构象位点的稳定(图1B)。抑制Aug- min活性能够加快微管切割的频率并使平行微管阵列形成增多。最终, 该研究组通过遗传学、活细胞显微成像技术及数学模拟方法阐释了Augmin通过拮抗Katanin复合体介导的微管切割, 来精确调控周质微管骨架动态重构与细胞形态建成的新机制(Wang et al., 2018)。
他们的研究结果一方面揭示了植物细胞体内Katanin复合体在微管精准切割中的作用机制; 另一方面阐明了Augmin蛋白复合体在同一位置不仅促进新微管的形成、抑制交叉微管的切割, 还可能具有识别并修复微管交叉位点损伤的功能。该研究为系统解析植物细胞形态建成的细胞水平调控机制提供了重要的理论依据, 是植物微管研究领域的重要突破; 同时, 该研究也为微管精准切割及微管网络形成缺陷相关的疾病研究提供了新思路并为药物设计提供了新靶标。
参考文献
文献选项
原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | DOI:10.1016/j.ceb.2007.12.004URL [本文引用: 1] |
| [2] | DOI:10.1073/pnas.1613273113PMID:27647923URLCellulose, often touted as the most abundant biopolymer on Earth, is a critical component of the plant cell wall and is synthesized by plasma membrane-spanning cellulose synthase (CESA) enzymes, which in plants are organized into rosette-like CESA complexes (CSCs). Plants construct two types of cell walls, primary cell walls (PCWs)... [本文引用: 1] |
| [3] | DOI:10.1016/j.cub.2014.09.053PMID:25447999URLLiu et02al. reveal that augmin localizes to cortical MTs to initiate MT nucleation in interphase Arabidopsis cells, and impairment of augmin function severely suppresses MT nucleation. They uncover a mechanism by which the augmin complex recruits the γ-tubulin complex to cortical MTs and triggers MT-dependent MT nucleation in interphase plant cells. [本文引用: 1] |
| [4] | DOI:10.1146/annurev-arplant-050213-040240PMID:24579997URLPlant stature and development are governed by cell proliferation and directed cell growth. These parameters are determined largely by cell wall characteristics. Cellulose microfibrils, composed of hydrogen-bonded -1,4 glucans, are key components for anisotropic growth in plants. Cellulose is synthesized by plasma membrane ocalized cellulose synthase complexes. In higher plants, these complexes are assembled into hexameric rosettes in intracellular compartments and secreted to the plasma membrane. Here, the complexes typically track along cortical microtubules, which may guide cellulose synthesis, until the complexes are inactivated and/or internalized. Determining the regulatory aspects that control the behavior of cellulose synthase complexes is vital to understanding directed cell and plant growth and to tailoring cell wall content for industrial products, including paper, textiles, and fuel. In this review, we summarize and discuss cellulose synthesis and regulatory aspects of the cellulose synthase ... [本文引用: 1] |
| [5] | DOI:10.1016/j.sysconle.2010.10.008PMID:8221885URLEukaryotic cells rapidly reorganize their microtubule cytoskeleton during the cell cycle, differentiation, and cell migration. In this study, we have purified a heterodimeric protein, katanin, that severs and disassembles microtubules to tubulin dimers. The disassembled tubulin can repolymerize, indicating that it is not irreversibly modified or denatured in the reaction. Katanin is a microtubule-stimulated ATPase and requires ATP hydrolysis to sever microtubules. Katanin represents a novel type of enzyme that utilizes energy from nucleotide hydrolysis to break tubulin-tubulin bonds within a microtubule polymer, a process that may aid in disassembling complex microtubule arrays within cells. [本文引用: 1] |
| [6] | DOI:10.1111/tpj.12102PMID:23311499URLPlant microtubules, in addition to their role in cell division and axial cell expansion, convey a sensory function that is relevant for the perception of mechanical membrane stress and its derivatives, such as osmotic or cold stress. During development, sensory microtubules participate in the mechanical integration of plant architecture, including the patterning of incipient organogenesis and the alignment with gravity-dependent load. The sensory function of microtubules depends on dynamic instability, and often involves a transient elimination of cortical microtubules followed by adaptive events accompanied by subsequent formation of stable microtubule bundles. It is proposed that microtubules, because of their relative rigidity in combination with their innate nonlinear dynamics, are pre-adapted for a function as mechanosensors and, in concert with the flexible actin filaments and the anisotropic cell wall, comprise a tensegral system that allows plant cells to sense geometry and to respond to fields of mechanical strains such that the load is minimized. Microtubules are proposed as elements of a sensory hub that decodes stress-related signal signatures, with phospholipase D as an important player. [本文引用: 1] |
| [7] | DOI:10.1126/science.1126551PMID:16627697URLExpression of a functional yellow fluorescent protein fusion to cellulose synthase (CESA) in transgenic Arabidopsis plants allowed the process of cellulose deposition to be visualized in living cells. Spinning disk confocal microscopy revealed that CESA complexes in the plasma membrane moved at constant rates in linear tracks that were aligned and were coincident with cortical microtubules. Within each observed linear track, complex movement was bidirectional. Inhibition of microtubule polymerization changed the fine-scale distribution and pattern of moving CESA complexes in the membrane, indicating a relatively direct mechanism for guidance of cellulose deposition by the cytoskeleton. [本文引用: 1] |
| [8] | DOI:10.1016/j.ceb.2009.11.001URL [本文引用: 1] |
| [9] | DOI:10.1242/jcs.101139PMID:22595526URLATP-dependent severing of microtubules was first reported in Xenopus laevis egg extracts in 1991. Two years later this observation led to the purification of the first known microtubule-severing enzyme, katanin. Katanin homologs have now been identified throughout the animal kingdom and in plants. Moreover, members of two closely related enzyme subfamilies, spastin and fidgetin, have been found to sever microtubules and might act alongside katanins in some contexts (Roll-Mecak and McNally, 2010; Yu et al., 2008; Zhang et al., 2007). Over the past few years, it has become clear that microtubule-severing enzymes contribute to a wide range of cellular activities including mitosis and meiosis, morphogenesis, cilia biogenesis and disassembly, and migration. Thus, this group of enzymes is revealing itself to be among the most important of the microtubule regulators. This Commentary focuses on our growing understanding of how microtubule-severing enzymes contribute to the organization and dynamics of diverse microtubule arrays, as well as the structural and biophysical characteristics that afford them the unique capacity to catalyze the removal of tubulin from the interior microtubule lattice. Our goal is to provide a broader perspective, focusing on a limited number of particularly informative, representative and/or timely findings. [本文引用: 1] |
| [10] | DOI:10.1016/j.pbi.2013.09.006PMID:24446545URLThe interphase microtubule arrays in flowering plant cells assemble at the cell cortex into patterns that affect cellular morphogenesis. A decade of live cell imaging studies has provided significant information about the in vivo properties of the microtubule polymers. Efforts to extrapolate individual properties to larger roles in organizing or patterning the microtubule array have produced models focused on self-organization and local levels of biological control. Recent studies looking at cortical microtubule arrays as they transition from an existing pattern to a new pattern have re-emerged as a testbed for examining these models and the molecular hypotheses underpinning them. The evidence suggests that microtubule patterning is locally controlled on the scale of a cell face, using or circumventing self-organizating properties as necessary. [本文引用: 1] |
| [11] | DOI:10.15252/embj.201796823PMID:28978669URLAbstract The microtubule (MT)-severing enzyme katanin triggers dynamic reorientation of cortical MT arrays that play crucial functions during plant cell morphogenesis, such as cell elongation, cell wall biosynthesis, and hormonal signaling. MT severing specifically occurs at crossover or branching nucleation sites in living Arabidopsis cells. This differs from the random severing observed along the entire length of single MTs invitro and strongly suggests that a precise control mechanism must exist invivo . However, how katanin senses and cleaves at MT crossover and branching nucleation sites invivo has remained unknown. Here, we show that the katanin p80 subunit KTN80 confers precision to MT severing by specific targeting of the katanin p60 subunit KTN1 to MT cleavage sites and that KTN1 is required for oligomerization of functional KTN80 TN1 complexes that catalyze severing. Moreover, our findings suggest that the katanin complex in Arabidopsis is composed of a hexamer of KTN1 TN80 heterodimers that sense MT geometry to confer precise MT severing. Our findings shed light on the precise control mechanism of MT severing in plant cells, which may be relevant for other eukaryotes. [本文引用: 3] |
| [12] | DOI:10.1016/j.cub.2018.03.007PMID:29657114URLPlant cells do not possess centrosomes, which serve as the microtubule organizing center in animal cells; how plant cell microtubule arrays are established and maintain their dynamics remain poorly understood [ 1 ]. The -tubulin complex and the katanin complex play central roles in the organization of plant cortical microtubules [ 2 ; 3 ; 4 ; 5 ; 6 ]. Previously, we reported that the augmin complex recruits the -tubulin complex to preexisting microtubules and initiates microtubule nucleation [ 7 ]. Moreover, we described how an intricate interaction between the katanin p60 subunit KTN1 and the p80 subunit KTN80 confers precise microtubule severing at either microtubule branching nucleation sites or crossovers [ 8 ]. Here, we observed that augmin preferentially localizes to microtubule crossovers. Live-cell observations and analyses revealed that, whereas a small portion of crossover-localized augmin complexes act to trigger nascent microtubule nucleation, the majority function in stabilizing the architecture of microtubule crossovers. Finally, genetic analyses and computational modeling confirmed that suppression of augmin activity elevates microtubule severing frequency and facilitates the formation of aligned microtubule arrays. Combined, our findings reveal an unexpected role of augmin and demonstrate that augmin antagonizes katanin-mediated microtubule severing. Furthermore, we propose a novel mechanism for how augmin determines self-organization of plant cortical microtubules by preventing microtubule severing at crossovers in addition to triggering microtubule nucleation. [本文引用: 4] |
| [13] | DOI:10.1016/j.tcb.2008.11.004PMID:19144522URLThe shape of plant cells depends on cortical microtubules. Their freedom from central microtubule organizing centres provides a powerful experimental system to study microtubule self-organization. New ideas have emerged from live-cell imaging of microtubules, particularly in the model system Arabidopsis thaliana, revealing the importance of encounters between microtubules in driving self-organization. Encounters are modulated by intrinsic microtubule-assembly dynamics, along with polymer activities that include cortical attachment, bundling and severing. Balancing the activities of microtubule-associated proteins (such as MOR1, CLASP, MAP65s and katanins) that control these processes is crucial for fine-tuning the organization of microtubule arrays. Too much or too little of any given activity tips the balance, with often dramatic effects on array organization, cell morphogenesis and even organ chirality. [本文引用: 1] |
| [14] | DOI:10.1126/science.aac7446PMID:26450210URLCellulose biosynthesis in plant secondary cell walls forms the basis of vascular development in land plants, with xylem tissues constituting the vast majority of terrestrial biomass. We used plant lines that contained an inducible master transcription factor controlling xylem cell fate to quantitatively image fluorescently tagged cellulose synthase enzymes during cellulose deposition in living protoxylem cells. The formation of secondary cell wall thickenings was associated with a redistribution and enrichment of CESA7-containing cellulose synthase complexes (CSCs) into narrow membrane domains. The velocities of secondary cell wall-specific CSCs were faster than those of primary cell wall CSCs during abundant cellulose production. Dynamic intracellular of endomembranes, in combination with increased velocity and high density of CSCs, enables cellulose to be synthesized rapidly in secondary cell walls. [本文引用: 1] |
1
2008
... 细胞骨架(cytoskeleton)系统主要包括微管(mi- crotubules, MTs)和微丝(actin filaments, F-actin).微管遍布细胞质中, 为细胞生命活动所必需, 在细胞周期运行、细胞分裂、细胞形态维持、细胞迁移、细胞内物质运输及细胞内信号转导等过程中起重要作用.哺乳动物细胞中, 中心体(centrosomes)是主要的微管组织中心(microtubule organization center, MTOC).高等植物细胞中没有中心体, 因此植物细胞缺乏类似动物细胞的微管组织中心来形成微管.在植物间期细胞中, 紧贴质膜、呈二维平行排列着植物特异的周质微管骨架(cortical microtubule arrays) (
1
2016
... 细胞骨架(cytoskeleton)系统主要包括微管(mi- crotubules, MTs)和微丝(actin filaments, F-actin).微管遍布细胞质中, 为细胞生命活动所必需, 在细胞周期运行、细胞分裂、细胞形态维持、细胞迁移、细胞内物质运输及细胞内信号转导等过程中起重要作用.哺乳动物细胞中, 中心体(centrosomes)是主要的微管组织中心(microtubule organization center, MTOC).高等植物细胞中没有中心体, 因此植物细胞缺乏类似动物细胞的微管组织中心来形成微管.在植物间期细胞中, 紧贴质膜、呈二维平行排列着植物特异的周质微管骨架(cortical microtubule arrays) (
1
2014
... 另外, 孔照胜研究组还对拟南芥Augmin蛋白复合体参与植物微管骨架的动态重构进行了长期深入的研究.他们发现, 在植物间期细胞中Augmin蛋白复合体能募集γ-TuRC蛋白复合体(γ-tubulin complex)到微管上, 从而产生新的成核位点, 并起始合成新的微管.该研究揭示了Augmin复合体控制植物细胞微管起始组装的机制(
1
2014
... 细胞骨架(cytoskeleton)系统主要包括微管(mi- crotubules, MTs)和微丝(actin filaments, F-actin).微管遍布细胞质中, 为细胞生命活动所必需, 在细胞周期运行、细胞分裂、细胞形态维持、细胞迁移、细胞内物质运输及细胞内信号转导等过程中起重要作用.哺乳动物细胞中, 中心体(centrosomes)是主要的微管组织中心(microtubule organization center, MTOC).高等植物细胞中没有中心体, 因此植物细胞缺乏类似动物细胞的微管组织中心来形成微管.在植物间期细胞中, 紧贴质膜、呈二维平行排列着植物特异的周质微管骨架(cortical microtubule arrays) (
1
1993
... 在微管阵列的动态转换过程中, 微管切割(microtubule severing)是非常重要的环节.微管切割由Katanin蛋白复合体完成, 该复合体包含1个60 kDa的具有ATPase活性的催化亚基p60, 以及1个80 kDa的包含WD-40重复基序的调节亚基p80 (
1
2013
... 细胞骨架(cytoskeleton)系统主要包括微管(mi- crotubules, MTs)和微丝(actin filaments, F-actin).微管遍布细胞质中, 为细胞生命活动所必需, 在细胞周期运行、细胞分裂、细胞形态维持、细胞迁移、细胞内物质运输及细胞内信号转导等过程中起重要作用.哺乳动物细胞中, 中心体(centrosomes)是主要的微管组织中心(microtubule organization center, MTOC).高等植物细胞中没有中心体, 因此植物细胞缺乏类似动物细胞的微管组织中心来形成微管.在植物间期细胞中, 紧贴质膜、呈二维平行排列着植物特异的周质微管骨架(cortical microtubule arrays) (
1
2006
... 细胞骨架(cytoskeleton)系统主要包括微管(mi- crotubules, MTs)和微丝(actin filaments, F-actin).微管遍布细胞质中, 为细胞生命活动所必需, 在细胞周期运行、细胞分裂、细胞形态维持、细胞迁移、细胞内物质运输及细胞内信号转导等过程中起重要作用.哺乳动物细胞中, 中心体(centrosomes)是主要的微管组织中心(microtubule organization center, MTOC).高等植物细胞中没有中心体, 因此植物细胞缺乏类似动物细胞的微管组织中心来形成微管.在植物间期细胞中, 紧贴质膜、呈二维平行排列着植物特异的周质微管骨架(cortical microtubule arrays) (
1
2010
... 在微管阵列的动态转换过程中, 微管切割(microtubule severing)是非常重要的环节.微管切割由Katanin蛋白复合体完成, 该复合体包含1个60 kDa的具有ATPase活性的催化亚基p60, 以及1个80 kDa的包含WD-40重复基序的调节亚基p80 (
1
2012
... 在微管阵列的动态转换过程中, 微管切割(microtubule severing)是非常重要的环节.微管切割由Katanin蛋白复合体完成, 该复合体包含1个60 kDa的具有ATPase活性的催化亚基p60, 以及1个80 kDa的包含WD-40重复基序的调节亚基p80 (
1
2013
... 细胞骨架(cytoskeleton)系统主要包括微管(mi- crotubules, MTs)和微丝(actin filaments, F-actin).微管遍布细胞质中, 为细胞生命活动所必需, 在细胞周期运行、细胞分裂、细胞形态维持、细胞迁移、细胞内物质运输及细胞内信号转导等过程中起重要作用.哺乳动物细胞中, 中心体(centrosomes)是主要的微管组织中心(microtubule organization center, MTOC).高等植物细胞中没有中心体, 因此植物细胞缺乏类似动物细胞的微管组织中心来形成微管.在植物间期细胞中, 紧贴质膜、呈二维平行排列着植物特异的周质微管骨架(cortical microtubule arrays) (
3
2017
... 细胞骨架(cytoskeleton)系统主要包括微管(mi- crotubules, MTs)和微丝(actin filaments, F-actin).微管遍布细胞质中, 为细胞生命活动所必需, 在细胞周期运行、细胞分裂、细胞形态维持、细胞迁移、细胞内物质运输及细胞内信号转导等过程中起重要作用.哺乳动物细胞中, 中心体(centrosomes)是主要的微管组织中心(microtubule organization center, MTOC).高等植物细胞中没有中心体, 因此植物细胞缺乏类似动物细胞的微管组织中心来形成微管.在植物间期细胞中, 紧贴质膜、呈二维平行排列着植物特异的周质微管骨架(cortical microtubule arrays) (
... Katanin和Augmin在植物微管(MTs)动态构象中的功能模式图(
(A) 拟南芥Katanin蛋白复合体的组装及其微管切割机制.Katanin蛋白复合体由p60亚基KTN1和p80亚基KTN80组成异源二聚体, 最终形成十二聚体的环状超复合体(左图).KTN80可将KTN1-KTN80异源二聚体导向至微管切割位点; 而KTN1介导形成KTN1- KTN80环状超复合体, 并识别特异的微管构象、完成切割(右图).(B) Augmin定位在微管交叉点上, 并且通过抑制Katanin的切割和稳定微管交叉构象, 来调控微管动态重组与细胞形态建成(左图).与野生型相比, 在Augmin的基因沉默突变体中(amiR-AUG6), 抑制Augmin活性能使微管切割的频率加快并形成更多的平行微管阵列(右图). ...
... Schematic models of Katanin and Augmin functions on microtubules (MTs) organization in plants (
(A) The formation of Katanin complexes and MTs severing in Arabidopsis cells. Katanin is composed of a p60 subunit KTN1 and a p80 subunit KTN80, which exists as a KTN1-KTN80 heterodimer in the cytosol (left panel). KTN80 determines the precise targeting of KTN1-KTN80 heterodimers to MTs crossover and branching nucleation sites. KTN1 further triggers the oligomerization of the mixed types of KTN1-KTN80 heterodimers that sense MT geometry to confer precise MT severing (right panel); (B) Augmin functions in regulating microtubule arrays by preventing katanin-mediated microtubule severing and maintaining the stability of crossovers (left panel). In the Augmin knockdown mutant (amiR-AUG6), a significantly higher microtubule severing frequency occurs and a greater proportion of aligned microtubule arrays (right panel) compared to the fine-network microtubule architectures observed in wild-type (left panel). ...
4
2018
... 细胞骨架(cytoskeleton)系统主要包括微管(mi- crotubules, MTs)和微丝(actin filaments, F-actin).微管遍布细胞质中, 为细胞生命活动所必需, 在细胞周期运行、细胞分裂、细胞形态维持、细胞迁移、细胞内物质运输及细胞内信号转导等过程中起重要作用.哺乳动物细胞中, 中心体(centrosomes)是主要的微管组织中心(microtubule organization center, MTOC).高等植物细胞中没有中心体, 因此植物细胞缺乏类似动物细胞的微管组织中心来形成微管.在植物间期细胞中, 紧贴质膜、呈二维平行排列着植物特异的周质微管骨架(cortical microtubule arrays) (
... Katanin和Augmin在植物微管(MTs)动态构象中的功能模式图(
(A) 拟南芥Katanin蛋白复合体的组装及其微管切割机制.Katanin蛋白复合体由p60亚基KTN1和p80亚基KTN80组成异源二聚体, 最终形成十二聚体的环状超复合体(左图).KTN80可将KTN1-KTN80异源二聚体导向至微管切割位点; 而KTN1介导形成KTN1- KTN80环状超复合体, 并识别特异的微管构象、完成切割(右图).(B) Augmin定位在微管交叉点上, 并且通过抑制Katanin的切割和稳定微管交叉构象, 来调控微管动态重组与细胞形态建成(左图).与野生型相比, 在Augmin的基因沉默突变体中(amiR-AUG6), 抑制Augmin活性能使微管切割的频率加快并形成更多的平行微管阵列(右图). ...
... Schematic models of Katanin and Augmin functions on microtubules (MTs) organization in plants (
(A) The formation of Katanin complexes and MTs severing in Arabidopsis cells. Katanin is composed of a p60 subunit KTN1 and a p80 subunit KTN80, which exists as a KTN1-KTN80 heterodimer in the cytosol (left panel). KTN80 determines the precise targeting of KTN1-KTN80 heterodimers to MTs crossover and branching nucleation sites. KTN1 further triggers the oligomerization of the mixed types of KTN1-KTN80 heterodimers that sense MT geometry to confer precise MT severing (right panel); (B) Augmin functions in regulating microtubule arrays by preventing katanin-mediated microtubule severing and maintaining the stability of crossovers (left panel). In the Augmin knockdown mutant (amiR-AUG6), a significantly higher microtubule severing frequency occurs and a greater proportion of aligned microtubule arrays (right panel) compared to the fine-network microtubule architectures observed in wild-type (left panel). ...
... 近期, 孔照胜研究组发现Augmin复合体倾向于定位在微管交叉位点, 一部分Augmin介导微管成核位点的形成, 另外一大部分则用于抑制Katanin的切割并维持微管交叉构象位点的稳定(
1
2009
... 细胞骨架(cytoskeleton)系统主要包括微管(mi- crotubules, MTs)和微丝(actin filaments, F-actin).微管遍布细胞质中, 为细胞生命活动所必需, 在细胞周期运行、细胞分裂、细胞形态维持、细胞迁移、细胞内物质运输及细胞内信号转导等过程中起重要作用.哺乳动物细胞中, 中心体(centrosomes)是主要的微管组织中心(microtubule organization center, MTOC).高等植物细胞中没有中心体, 因此植物细胞缺乏类似动物细胞的微管组织中心来形成微管.在植物间期细胞中, 紧贴质膜、呈二维平行排列着植物特异的周质微管骨架(cortical microtubule arrays) (
1
2015
... 细胞骨架(cytoskeleton)系统主要包括微管(mi- crotubules, MTs)和微丝(actin filaments, F-actin).微管遍布细胞质中, 为细胞生命活动所必需, 在细胞周期运行、细胞分裂、细胞形态维持、细胞迁移、细胞内物质运输及细胞内信号转导等过程中起重要作用.哺乳动物细胞中, 中心体(centrosomes)是主要的微管组织中心(microtubule organization center, MTOC).高等植物细胞中没有中心体, 因此植物细胞缺乏类似动物细胞的微管组织中心来形成微管.在植物间期细胞中, 紧贴质膜、呈二维平行排列着植物特异的周质微管骨架(cortical microtubule arrays) (

{kind=link}
{kind=link}