 ,*, 田世平中国科学院植物研究所, 中国科学院北方资源植物重点实验室, 北京 100093
,*, 田世平中国科学院植物研究所, 中国科学院北方资源植物重点实验室, 北京 100093Advances in the Regulation on Autophagy by Reactive Oxygen Species in Plant Cells
Danying Ma, Dongchao Ji, Yong Xu, Tong Chen,*, Shiping TianKey Laboratory of Plant Resources, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China通讯作者:
收稿日期:2018-01-12接受日期:2018-03-13网络出版日期:2019-01-30
| 基金资助: |
Corresponding authors:
Received:2018-01-12Accepted:2018-03-13Online:2019-01-30

摘要
关键词:
Abstract
Keywords:
PDF (1609KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
马丹颖, 季东超, 徐勇, 陈彤, 田世平. 活性氧调控植物细胞自噬的研究进展. 植物学报, 2019, 54(1): 81-92 doi:10.11983/CBB18012
Ma Danying, Ji Dongchao, Xu Yong, Chen Tong, Tian Shiping.
自噬是真核生物中普遍存在的高度保守的胞内物质降解过程, 是生物体清除异常蛋白或受损细胞器, 以维持蛋白质代谢平衡及细胞内环境稳定的有效机制(王燕和刘玉乐, 2010; 杨小龙等, 2017)。营养缺陷能够诱导酵母细胞中自噬的发生, 以实现物质循环利用。研究表明, 自噬参与植物细胞中营养物质循环、生长发育、生物及非生物胁迫响应等过程(黄晓和李发强, 2016; Qi et al., 2017)。活性氧(reactive oxygen species, ROS)是生物进行有氧代谢的毒性副产物。正常条件下, ROS的产生与清除处于动态平衡, 而当植物衰老或受到胁迫时, ROS会在植物体内积累, 形成氧化胁迫。研究表明, ROS参与自噬的调控, 两者之间关系密切。本文结合最新的研究进展, 简要概括了植物中ROS产生和细胞自噬的机制, 阐述了ROS对自噬的调控及两者间复杂的相互作用。
1 活性氧的种类和作用
1.1 活性氧的种类及其性质
ROS是生物有氧代谢过程中产生的一类化学性质活泼且具有较高氧化活性的分子或离子的总称(Rahal et al., 2014), 主要包括超氧阴离子(O2·-)、羟基自由基(·OH)、过氧化氢(H2O2)和单线态氧(1O2)等(Dewaele et al., 2010)。其中, H2O2具有中度反应活性, 其半衰期相对较长, 是最稳定的ROS分子(景红娟等, 2012)。H2O2不带净电荷, 因此容易透过生物膜, 在细胞中进行远距离传输, 引发远距离氧化损伤(Bienert and Chaumont, 2014)。1.2 活性氧的产生和清除机制
正常或胁迫条件下, 植物细胞中质膜、叶绿体、线粒体、过氧化物酶体、内质网以及质外体等许多部位都能产生ROS (Finkel, 2011)。研究表明, 光照条件下, 绿色植物中ROS产生的主要部位是叶绿体和过氧化物酶体, 而在非绿色植物中或黑暗条件下, 线粒体和NADPH氧化酶(NADPH oxidase, NOX)是ROS的主要来源(M?ller, 2001; Foyer and Noctor, 2003)。除此之外, 定位于不同细胞区室的代谢途径(如脂肪酸β-氧化)也会产生ROS (Buchanan and Balmer, 2005)。经过长期进化, 植物已形成了酶促和非酶促两类有效的ROS清除机制(Ahmad et al., 2010)。酶促脱毒系统主要包括超氧化物歧化酶(superoxide dismutase, SOD)、过氧化氢酶(catalase, CAT)以及抗坏血酸过氧化物酶(ascorbate peroxidase, APX)等(Bla- gosklonny, 2008)。非酶促保护机制则主要包括一些直接参与ROS清除的抗氧化物, 如抗坏血酸、类胡萝卜素以及类黄酮(Apel and Hirt, 2004)。此外, 维生素等小分子物质也参与氧自由基的清除(Schafer et al., 2002)。
1.3 活性氧在植物细胞中的作用
传统观念认为, ROS是一类只会造成细胞损伤的毒性物质。但近年的研究表明, ROS作为一种信号分子还参与细胞增殖、分化及凋亡等多个生理过程, 在植物生长发育、胁迫适应和细胞程序性死亡(programmed cell death, PCD)过程中具有重要调节作用(Gechev et al., 2006)。高浓度ROS对有机体极其有害, 当ROS水平超出防御机制范围, 细胞处于氧化胁迫状态, 引发脂质过氧化、蛋白质氧化、核酸损伤和酶失活, 并激活信号通路, 最终导致PCD (van Dongen and Licausi, 2015)。但低浓度ROS能在植物细胞信号转导途径中作为第二信使, 介导植物对激素或环境胁迫的多种应答反应(Scheler et al., 2013)。例如, 高等植物和藻类中, 1O2和·OH在光合作用过程中产生并引起氧化损伤; 同时, 1O2还可通过调节特定基因的表达引发不同的信号级联反应, 从而对特定的氧化胁迫环境产生生理适应(Ledford et al., 2007)。2 植物细胞自噬
2.1 自噬的发现与分类
Ashford和Porter (1962)用电子显微镜在小鼠肝细胞中首次观察到细胞自食(self-eating)现象, 自噬(auto- phagy)这一概念则是由溶酶体的发现者De Duve在1963年提出(蔡霞等, 2016; 刘洋等, 2018)。随后, Ohsumi发现酿酒酵母(Saccharomyces cerevisiae)中存在自噬现象, 并通过遗传分析方法鉴定了自噬相关基因(autophagy-related gene, ATG), 阐明了其分子机制, 且因此荣获2016年诺贝尔生理学或医学奖(Levine and Klionsky, 2017)。目前, 研究者对自噬过程采取了不同的分类方式, 其中最经典的分类方法是基于作用机理的不同, 将自噬分为巨自噬(macro autophagy)、微自噬(microautophagy)和分子伴侣介导的自噬(chaperone-mediated autophagy) (Reumann et al., 2010; Mizushima et al., 2011)。巨自噬是最为常见和研究最多的自噬类型, 其主要特征是双层膜结构的自噬小泡(autophagosome)的形成(Ho- fius et al., 2011); 在真核生物中, 自噬小泡包裹细胞内受损的蛋白、细胞器或外源成分, 其外膜与溶酶体或液泡融合, 释放内膜包裹的自噬小体(autophagic body)。随后内含物降解, 产物得以循环利用(Liu and Bassham, 2012; 任晨霞和龚清秋, 2014 )。下文以巨自噬(简称自噬)为主, 对自噬的分子机制进行介绍。2.2 植物细胞自噬的分子机制
2.2.1 参与自噬过程的关键组分和过程自噬过程的关键基因ATGs最早在酿酒酵母中被发现(Hofius et al., 2017)。随后, 在低等真核生物、植物和动物细胞中均发现了ATGs的同源基因(Díaz-Tro- ya et al., 2011)。迄今, 酵母细胞中已有32个ATGs被鉴定(Okamoto et al., 2009); 拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)和烟草(Nicotiana benthamiana)中也分别鉴定了34、33和30个同源基因(Xia et al., 2011; Zhou et al., 2015)。ATGs的多拷贝现象在植物细胞中广泛存在, 然而具体原因尚不清楚。目前, 研究者对此主要有2种猜测, 一是ATGs存在功能冗余; 二是植物在恶劣的环境条件下采用的生存进化策略, 这些基因的功能随着时间的推移有所差别(Yoshimoto et al., 2010; Han et al., 2011)。
2.2.2 自噬发生过程
自噬的发生过程可划分为5个阶段, 即自噬诱导、自噬小泡成核、自噬小泡的扩展和闭合、自噬小泡与液泡融合、胞内成分降解及循环利用(Thompson and Vierstra, 2005)。以下我们以植物和酵母为主, 对各个步骤中所涉及的自噬组分和分子机制进行介绍。
自噬诱导: ATG1与ATG13互作在细胞自噬诱导中起关键作用。拟南芥atg13a/atg13b双突变体中不能形成自噬小泡, 说明ATG13是植物细胞自噬的必需组分(Suttangkakul et al., 2011)。ATG1激酶是自噬过程中重要的调节蛋白, 为自噬小泡前体结构形成和自噬起始所必需(Mizushima, 2010)。在植物以及酵母中, ATG1的功能主要通过雷帕霉素靶标(target of rapamycin, TOR)的磷酸化调控(Kamada et al., 2000)。当营养元素缺乏时, TOR通过形成TOR复合物作用于ATG1/ATG13复合体, 从而诱导自噬(He and Klionsky, 2009)。有研究表明, ATG9复合体也参与自噬的诱导过程(Yoshimoto et al., 2004)。
自噬小泡成核: 酵母细胞中自噬小泡的成核反应与PI3K复合体密切相关。PI3K复合体包括ATG6/ VPS30、ATG14、UVRAG、VPS15和VPS34 (Kihara et al., 2001), 其中只有ATG14尚未在植物细胞中找到同源物。VPS15和VPS34是具有催化功能的关键蛋白, ATG6则是酵母细胞自噬发生水平的关键控制因子, 为成核阶段所必需。
自噬小泡的扩展与闭合: ATG8酯化系统和AT- G12蛋白连接系统的最终产物ATG8-PE及ATG12- ATG5-ATG16对自噬小泡膜的扩展与囊泡内物质降解具有重要作用。该步骤可分为2个阶段。第1阶段为ATG12与ATG5共价结合(üstün et al., 2017)。ATG12被具有E1酶活性的ATG7活化, 首先转移到ATG10上, 然后转移到靶蛋白ATG5, 形成复合物ATG12-ATG5。研究表明, ATG12-ATG5复合体作为一种类E3酶参与ATG8的酯化作用(Chung et al., 2010)。第2阶段是ATG8-PE的共价结合反应。半胱氨酸蛋白酶ATG4切割新合成的ATG8的C末端, 使其暴露出甘氨酸残基; 之后, ATG7识别含有甘氨酸残基的ATG8, 并通过硫酯键与其结合; 活化的ATG8通过硫酯键转移到具有E2酶活性的ATG3上; 最后, ATG8通过其C末端的甘氨酸与磷脂酰乙醇胺(pho- sphatidylethanolamine, PE)之间形成酰胺键, 进而完成ATG8-PE的共价连接(Hayward and Dinesh- Kumar, 2011)。连接后的ATG8-PE定位在吞噬泡(phagophore)的内外膜上, 伴随细胞自噬发生的全过程(Minina et al., 2018)。
自噬小泡与液泡融合: 自噬小泡形成后会靶向液泡进行融合。酵母细胞中的融合过程涉及SNARE蛋白、Rab GTP酶Ypt7和C类Vps/HOPS复合物等多种因子。Vps复合物和Ypt7参与融合位点复合物的招募和结合(Wang and Klionsky, 2003)。SNARE蛋白形成的复合物是2个细胞器之间的桥梁(Ungermann and Langosch, 2005)。
降解及循环利用: 融合后, 自噬小体释放到液泡腔中进行降解。该过程依赖于液泡的酸化作用及液泡驻留水解酶的活性。自噬的主要作用是降解胞质成分, 并回收利用所产生的大分子来合成必需的组分, 以应对多种胁迫环境。
3 植物细胞中活性氧与自噬相关的实验证据
综上所述, 植物细胞中线粒体和叶绿体等多种细胞器均能产生ROS, 且胞内ROS的水平受到氧化、营养和内质网胁迫以及病原菌侵染等信号的调节。自噬作为一种大规模的胞质降解路径, 不仅能降解单个长寿蛋白和蛋白质复合体, 还能降解细胞器, 在细胞响应和适应不同胁迫环境中具关键作用。大量的研究表明, 不同来源的ROS对植物细胞自噬均具调控作用(图1)。图1
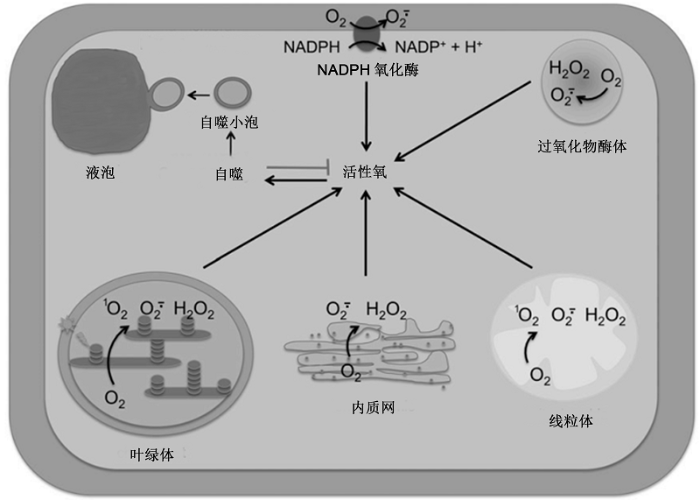 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1植物细胞中不同来源的活性氧调控自噬(改自Pérez-Pérez et al., 2012b)
定位于质膜的NADPH氧化酶, 以及叶绿体、线粒体、过氧化物酶体和内质网等细胞器均能产生活性氧。过量的活性氧会诱导细胞自噬, 以减少活性氧的产生并清除细胞受损组分。NADPH: 烟酰胺腺嘌呤二核苷酸磷酸; O2: 氧分子; O2·-: 超氧阴离子; H2O2: 过氧化氢; 1O2: 单线态氧
Figure 1Different reactive oxygen species (ROS) sources control autophagy in plant cells (modified from Pérez-Pérez et al., 2012b)
ROS can be generated by plasma membrane-localized NOX and different organelles, including chloroplast, mitochondria, peroxisome, and endoplasmic reticulum. Excess ROS then induce autophagy, which contributes to down-regulate ROS production and remove damaged cellular components. NADPH: Reduced nicotinamide adenine dinucleotide phosphate; O2: Dioxygen; O2·-: Superoxide anion; H2O2: Hydrogen peroxide; 1O2: Singlet oxygen
外源H2O2处理会对拟南芥造成严重的氧化应激, 从而诱导自噬进程(Xiong et al., 2007a)。氧化胁迫条件下, 蛋白质会发生不同类型的不可逆的氧化修饰, 包括羰基化和亚磺酸化等。这些损伤的蛋白需要被迅速有效地降解, 而自噬缺陷突变体对H2O2高度敏感, 并积累羰基化蛋白, 表明自噬降解过程为细胞适应氧化应激所必需。此外, 在拟南芥atg2以及atg5突变体中, 可以观察到H2O2的大量积累(Yoshimoto et al., 2009), 与H2O2处理结果类似。在ROS诱导剂甲基紫精(methylviologen, MV)作用下, 拟南芥野生型植株中氧化蛋白含量升高并大量积累, 且自噬活性被激活, 自噬小体数量显著增加。MV处理水稻突变体atg10b或拟南芥ATG18a下调表达的植株后, 突变体细胞内聚集的氧化蛋白显著多于野生型植株, 且对氧化胁迫更加敏感(Xiong et al., 2007b; Shin et al., 2009)。但是, MV诱导ROS产生的分子机制与外源H2O2处理不同, 它截获来自叶绿体光系统I (PSI)或植物线粒体呼吸电子传递链中的电子, 与O2反应产生O2·-并快速转化为H2O2。因此, MV处理可使植物叶绿体和/或线粒体中产生ROS, 并可能通过在不同细胞器中引起的氧化损伤来诱发自噬。
植物NADPH氧化酶(NADPH oxidase, NOX)也称作呼吸爆发氧化酶同源物(respiratory burst oxidase homologue, Rboh), 位于细胞质膜上, 以NADPH为电子供体, 从质膜胞质侧的NADPH接受电子并传递给O2, 经单电子还原在质膜的质外体侧形成O2·-, O2·-经细胞壁结合的SOD催化或自然歧化形成H2O2, H2O2再经Fenton反应或Haber-Weiss反应形成·OH (Asada, 2006; 林植芳和刘楠, 2012)。NOX通过整合ROS与其它信号通路, 在氧化还原网络中发挥核心作用(Suzuki et al., 2011), 参与不同的生物学过程中对植物细胞自噬的调控。
NOX生成的ROS在根毛生长过程中具关键作用(Swanson and Gilroy, 2010)。ATGs的缺失突变导致植株根毛变短和花粉管发育受阻等表型(Kwon et al.,2010a, 2010b)。由此可知, ROS和自噬可能参与根毛及花粉管的发育过程。但是, 两者如何共同调控根毛和花粉管发育仍然未知。盐和干旱胁迫会严重影响植物的生长发育, 在胁迫条件下离子压力和渗透胁迫使植物产生氧化损伤, 导致ROS和氧化蛋白积累(Tsu- gane et al., 1999)。Liu等(2009)研究表明, 在高盐和渗透胁迫条件下, 拟南芥AtATG18a的表达水平上调, 自噬活性被激活, 而AtATG18a-RNAi植株对上述两种胁迫均很敏感。用NOX抑制剂处理植株后, 发现高盐条件下NOX抑制剂能够抑制细胞自噬的发生, 而在干旱胁迫下细胞自噬不受NOX抑制剂影响, 仍能被诱导。此外, 他们还发现, 抑制植物细胞NOX活性可避免糖或氮饥饿诱导的自噬小泡形成。这些结果暗示, 营养和高盐胁迫过程中ROS可能作为信号分子诱导自噬; 细胞自噬对营养、高盐及干旱胁迫的响应存在NOX依赖和不依赖两种调节途径。同时, 化学抑制剂二亚苯基碘抑制NOX活性的实验也表明, 饥饿(氮或碳)或盐胁迫诱导自噬依赖于NOX活性(Pérez- Pérez et al., 2012b), 说明NOX产生的ROS为响应营养和盐胁迫的自噬激活所必需。最近有研究者发现, MdATG18a过表达显著增强苹果(Malus domestica)植株对干旱胁迫的抗性, 而且过表达植株的光合速率和抗氧化能力显著增强, 其作用机理可能是通过产生更大的自噬小体并提高自噬活性, 从而加速聚集蛋白的降解, 缓解细胞的氧化胁迫(Sun et al., 2018)。
病原菌侵染能够有效触发植物细胞自噬(Liu et al., 2005), 而自噬过程通过超敏反应诱导的细胞程序性死亡, 阻止病原菌进一步侵染(Bassham, 2007; Coll et al., 2014)。Chaouch等(2012)研究表明, 在拟南芥中, NOX (特别是AtRbohF)通过ROS将细胞氧化还原状态与水杨酸信号相关的细胞死亡下游变化相关联。另有研究表明, 在响应甘蓝链格孢菌(Alter- naria brassicicola)侵染时, 拟南芥atg5、atg10和atg18a突变体植株ROS水平升高, 坏死组织扩大且感病性增强, 说明自噬对宿主细胞有促活作用; 且上述突变体被活体营养病原菌侵染时, 会通过改变水杨酸相关信号通路来增强免疫(Lenz et al., 2011)。这些结果表明, ROS可能会诱导自噬以应对生物胁迫, 而NOX可能在病原体侵染的初始阶段参与自噬过程的激活, 但该假设仍需进一步实验验证。Henry等(2015)研究了拟南芥三磷酸甘油醛脱氢酶(glyce- raldehyde-3-phosphate dehydrogenase, GAPDH)敲除突变体, 发现其ROS水平上升且对病原菌的抗性增强, 表现出持续的自噬效应, 表明GAPDH负调控细胞自噬。另外, 对烟草的研究表明, ATG3与胞质定位的磷酸甘油醛脱氢酶(GAPCs)互作调节自噬活性。GAPC基因沉默激活了ATG3依赖的自噬活性, 同时增强了烟草中N基因介导的细胞死亡以及植株对烟草花叶病毒和丁香假单胞菌的抗性。正常条件下GPAC的结合抑制了ATG3的功能; 氧化胁迫条件下ROS水平升高则破坏了二者间的互作, 并激活了依赖ATG3的自噬活性(Han et al., 2015)。
4 内质网胁迫中活性氧对自噬活性的调节
研究表明, 细胞自噬参与内质网(endoplasmic reticulum, ER)胁迫导致的内质网降解过程。Liu等(2012b)通过内质网标记蛋白示踪, 发现降解的ER 可与自噬小体共定位; 此外, 通过透射电镜观察也发现在自噬小体中存在带有核糖体的ER碎片, 说明细胞自噬通路可响应ER胁迫, 进而参与内质网膜的循环利用。Xu等(2017)对其分子机制进行了深入研究, 发现定位于内质网的BI-1 (Bax Inhibitor-1)可与自噬过程中的关键组分ATG6互作, 从而招募自噬过程的其它重要组分, 起始内质网相关的自噬进程, 且这种互作关系对于BI-1诱导的细胞自噬和细胞死亡均非常重要。Yorimitsu等(2006)对酿酒酵母的研究表明, ER胁迫是自噬的强诱导信号; 且衣霉素和N-糖基化抑制剂等ER胁迫诱导剂也能有效激活自噬(Pérez- Pérez et al., 2010)。因此, 基于自噬和ER应激反应的高度保守性, 人们推测高等植物细胞中也存在类似的联系。ER中的蛋白质折叠是高度依赖于氧化还原状态的过程。已有多项研究报道了ER应激与ROS生成之间的关联证据(Malhotra and Kaufman, 2007; Rutkowski and Kaufman, 2007)。Santos等(2009)研究表明, 存在于ER腔中的氧化还原酶、线粒体ROS和NOX与ER应激过程中ROS的产生相关。与酵母细胞类似, 为了响应ER胁迫, 植物细胞可以诱导自噬以消除未折叠的蛋白质和有害分子(包括ROS), 或在恢复期间平衡由未折叠蛋白应答(unfolded protein response, UPR)引起的ER扩张, 从而促进细胞存活(Bernales et al., 2006)。但是, 目前还不清楚ROS产生是否是ER应激诱导自噬的主要调控机制。
5 细胞器的自噬降解
胁迫条件下产生的过量ROS常可造成细胞损伤, 这种情况下细胞必须引入更积极的机制, 方可消除受损组分并保持ROS受到控制。自噬是降解氧化分子的主要防御机制, 且在特定条件下还可能清除产生ROS的细胞器。Tolkovsky (2009)研究表明, 细胞主要通过自噬机制选择性清除受损或不需要的线粒体, 这一过程被称为线粒体自噬。酵母细胞在饥饿条件下或生长平台期会发生典型的线粒体自噬(Kissová et al., 2004; Tal et al., 2007); 而ROS清除剂N-乙酰半胱氨酸能降低酵母细胞中线粒体自噬的活性(Okamoto et al., 2009)。被氧化的过氧化物酶体聚合物同样可通过自噬过程被选择性降解(Shibata et al., 2013)。由于植物细胞中过氧化物酶体是H2O2的主要来源, 且能够产生O2·-或一氧化氮(nitric oxide, NO) (del Río et al., 2002; Foyer et al., 2009; del Río, 2011)。故过氧化物酶体可能在氧化还原信号转导及其稳态控制中起重要作用, 但是过氧化物酶体降解的生理作用及其对氧化还原信号传递的影响机制尚不明晰。叶绿体是植物和藻类中ROS的主要来源。在黑暗引起的碳饥饿条件下, 叶绿体组分可通过自噬过程被降解(Izumi et al., 2010; Wang and Liu, 2013; Wang et al., 2013)。在细胞内可以观察到包含Rubisco蛋白的亚细胞结构以及含叶绿体基质的小球形亚细胞结构(Ishida et al., 2008)。此外, 衰老植物中还可观察到整个叶绿体的降解。然而叶绿体组分存在不同的降解途径, 包括自噬和蛋白水解过程(Wang et al., 2015)。已有实验证明, Rubisco蛋白的降解在自噬缺陷植株中未受到显著影响(Wada et al., 2009)。最近有研究表明, 细胞受UV-B伤害后, 完整叶绿体通过自噬途径被运送到液泡中降解, 而在自噬相关突变体中并未观察到这种现象, 且自噬突变体表现出明显的UV-B敏感表型, 积累较多的被破坏的叶绿体。这些结果说明, 光伤害诱导的叶绿体自噬并非经RCB (Rubi- sco-containing body)途径降解(Izumi et al., 2017)。Pérez-Pérez等(2012a)研究表明, 衣藻在光氧化损伤下的自噬激活与Rubisco降解之间也未表现出直接的相关性, 暗示自噬可能并非主要途径, 而可能是通过细胞器的特异性降解来调节ROS产生。植物细胞中的这种机制有待进一步研究。
6 自噬氧化还原调节的靶标
综上所述, ROS可能通过一种高度保守的机制来调节自噬, 但是自噬如何感知并接收氧化还原信号仍然未知。迄今为止, 仅有1个靶标(ATG4)被鉴定, ATG4通过整合氧化还原信号来调节自噬, 其它调节组分(如ATG1和TOR激酶)也可能参与氧化还原信号转导。6.1 ATG4通过氧化还原信号调控自噬过程
现已知, 哺乳动物中ATG4是唯一已被证明的氧化还原调节靶标的ATG蛋白。该半胱氨酸蛋白酶在自噬小泡形成中起双重作用, ATG4通过脂质化系统将新合成的ATG8的C末端与PE结合, 并使ATG8脱脂从自噬小泡膜回收(Kirisako et al., 2000)。然而, 调节ATG4脂化和脱脂活性的分子机制目前尚不清楚。已有实验证明, 哺乳动物中H2O2可以通过氧化ATG4分子中的Cys残基使ATG4失活, 以阻止ATG8的加工; 而还原条件则激活ATG4 (Scherz-Shouval et al., 2007)。业已证明, ROS通过调节ATG4的活性来影响饥饿诱导的自噬进程。在拟南芥和玉米(Zea mays)基因组中已鉴定了ATG4的2种同源物(Chung et al., 2009), 此2种物质在拟南芥中的缺失突变导致自噬过程完全阻断(Yoshimoto et al., 2004), 说明ATG4为植物细胞中的自噬过程所必需。鉴于自噬的高度进化保守性, ATG4很可能也是植物细胞中氧化还原信号的感受器和中心整合因子。6.2 TOR-ATG1路径
TOR-ATG1是自噬的关键组分, 其催化活性为起始阶段所必需(Meijer et al., 2007)。ATG1最初在酵母中被鉴定, 该蛋白在进化过程中高度保守, 通过与ATG13等蛋白互作, 发挥调控功能(Hosokawa et al., 2009)。ATG1/ATG13复合物的活性受磷酸化/去磷酸化修饰调节, 并被TOR复合体1 (target of rapamycin complex 1, TORC1)、蛋白激酶A (protein kinase A, PKA)和腺苷酸活化蛋白激酶(adenosine 5'-mono- phosphate-activated protein kinase, AMPK)等信号通路激活(Alers et al., 2012; Xiong et al., 2013)。这些激酶不同程度地调节ATG1与ATG13间的互作强度以及ATG1的激酶活性。TORC1和PKA对ATG1和/或ATG13的磷酸化会抑制自噬, 但AMPK信号可促进自噬的发生(Nakatogawa et al., 2009; Zhang et al., 2016)。ATG1/ATG13激酶复合物在进化过程中是保守的(Robaglia et al., 2012)。酵母ATG1和ATG13的直系同源基因也存在于拟南芥基因组中, 其产物可能调节植物细胞自噬(Suttangkakul et al., 2011; Xiong and Sheen, 2014)。然而, 植物细胞中ATG1/ATG13调节的信号通路的具体作用尚未见报道。TOR和SnRK1蛋白激酶(植物细胞中AMPK的同源蛋白)在高等植物及藻类的营养与能量感知过程中具有重要作用。TORC1信号已被证明能调节藻类植物细胞中的自噬。Bassham实验室最近研究表明, 不同环境条件对植物细胞自噬活性的诱导具有特异性, 且对TOR的依赖性也存在差异。其中, 营养、盐和渗透胁迫条件下SnRK1蛋白激酶复合物通过TOR调节自噬活性; 而氧化和内质网胁迫条件下, 自噬活性的激活依赖于SnRK1, 却并非TOR (Pu et al., 2017; Soto-Burgos and Bassham, 2017)。拟南芥中AtTOR转录抑制或衣藻中雷帕霉素处理引起的TOR功能降低均会导致两种系统中的自噬活性显著增加, 与营养饥饿细胞中观察到的现象类似(Pérez-Pérez et al., 2010)。这些结果暗示, TOR在控制光合生物自噬中具有重要作用, 但不排除其它保守途径(如SnRK1信号转导)也可能参与该过程。
越来越多的研究表明, 低等和高等真核生物中, TOR是线粒体功能的关键控制器, 与氧化还原新陈代谢紧密相关(Schieke and Finkel, 2006)。AMPK等TOR的上游调节因子通过p53调节的Sestrins与氧化还原信号相关联(Budanov, 2011)。动物细胞伤害感受器(ataxia-telangiectasia mutated, ATM)能够响应胞质ROS水平的升高, 通过AMPK途径抑制mTOR C1并诱导自噬活性, 从而激活TSC/2 (Alexander et al., 2010), 表明ROS主要通过失活TOR激活自噬活性(Blagosklonny, 2008)。相比TOR, TORC1信号在高等植物和藻类细胞中更为保守(Díaz-Troya et al., 2011; Moreau et al., 2012)。但是目前仍未见关于TORC1底物的确切报道, 因而无法特异性监测这种途径的自噬活性的调节。Dames等(2005)对TOR的结构解析表明, TOR的C端FATC结构域(FRAP/ATM/ TRRAP C-terminal)的氧化还原状态可能会影响TOR的降解。但Ren等(2011)对拟南芥的研究证实, FATC结构域对TOR的功能并非必需。基于TOR在光合生物自噬过程中的作用, 以及其它系统中报道的氧化还原信号与TOR信号之间的互作, 可以推测在某些胁迫条件下, ROS可能会下调TOR的活性, 导致植物与藻类中的ATG1以及自噬活性激活。目前还未得到关于ROS通过抑制TOR活性进而调控自噬的直接实验证据。
7 展望
酵母和哺乳动物细胞中的自噬过程都已得到深入研究, 但植物细胞中对自噬的研究多集中于自噬相关基因的发现和鉴定, 具体调控机制还不清楚。尽管多种生物或非生物胁迫均能诱导ROS的产生, 但人们对ROS调控自噬过程的分子机制还了解甚少。植物细胞中多种细胞器都具有其特异的自噬过程, 包括过氧化物酶体自噬、线粒体自噬、内质网自噬和核糖体自噬等。而这些细胞器均是产生ROS的主要部位, 暗示ROS在细胞器自噬中起关键调控作用。因此, ROS如何调控细胞器自噬, ROS与自噬在植物生长发育、响应生物及非生物胁迫中的作用需要进一步研究和探讨。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.3969/j.issn.1000-6281.2016.02.013URL [本文引用: 1]

细胞自噬是真核生物对细胞内物质进行循环利用的重要途径。在植物中已发现的细胞自噬有两种形式,即微自噬和巨自噬。自噬在植物体的生长发育、免疫应答、胁迫反应、衰老等过程中具有非常重要的作用。显微镜技术是研究细胞自噬的重要手段,与生物化学和分子生物学技术相结合,荧光显微镜,激光扫描共聚焦显微镜,电子显微镜等技术的相互印证,大大地推动了对细胞自噬过程和机理的研究。本文扼要概述了显微镜技术在植物细胞自噬研究中的应用。
DOI:10.11983/CBB16011URL [本文引用: 1]
细胞自噬是真核生物中一种由液泡或溶酶体介导的,对细胞内物质进行周转的重要代谢机制.在植物中,细胞自噬作为一种重要的降解手段,参与营养物质的重新分配、受损蛋白和细胞器的清除及生物和非生物胁迫的响应等过程.此外,细胞自噬在各种程序性细胞死亡中也起着重要作用,该文主要综述了近几年来在此方面的研究进展.
DOI:10.3724/SP.J.1259.2012.00534URL [本文引用: 1]
Autophagy is a highly conserved biological progress in degradation of the cell components and has key roles in starvation, senescence and infection by pathogens. Moreover, reactive oxygen species (ROS) is a byproduct of higher plants during normal and stress conditions and plays an important role in regulation of growth and development and programmed cell death during stress response. Recent research has shown that ROS and autophagy both regulate programmed cell death in the hypersensitive reaction during infection by pathogens, growth and development and stress adaptation. We discuss the signal interaction between ROS and autophagy in plants, including species and characteristics of ROS, the molecular basis of plant autophagy, and functions of ROS in plant autophagy.
DOI:10.3724/SP.J.1259.2012.00074URL [本文引用: 1]
Reactive oxygen species (ROS) are the byproducts of plant aerobic metabolism. ROS are considered to have double functions (harmful and beneficial) in many plant processes. Oxidative damage to cells and signal transduction in the induced protection response by ROS have been investigated intensively. ROS are increasingly thought to control plant growth and development in particular. Cell growth is the important component of plant development, and control of plant development by ROS is by regulating cell growth, so ROS is an important regulator of plant growth and development. Here, we review the research progress in the control and regulation of plant growth and development by hydroxyl radicals and their precursors, superoxide radical and hydrogen peroxide, and the mechanism, the generation pathway of ROS, the methods for detecting ROS, and prospects for future study.
URL [本文引用: 1]
细胞自噬是一类依赖于溶酶体和液泡的蛋白质降解途径。在动物细胞中,靶物质通过自噬体包裹被运送到溶酶体中,由特定的水解酶降解;而植物和酵母细胞中该过程在液泡内进行。近年来,在模式植物拟南芥(Arabidopsis thaliana)中鉴定到多个关键ATG基因,它们对植物细胞自噬体的形成及自噬调控起到关键作用。该文全面综述了植物细胞自噬的调控及其在植物逆境胁迫中的生理功能。
URL [本文引用: 1]
自噬(autophagy)是真核生物细胞通过形成自噬体,回收利用胞内物质,维持细胞健康的高通量亚细胞降解途径。随着酵母和动物自噬研究的深入,植物自噬也受到越来越多的关注。近期的研究揭示了植物自噬的基本机制及其生理意义,也发现了植物特有的自噬形式与自噬相关基因。该文主要综述了自噬在植物碳、氮营养中的作用。
URL [本文引用: 1]
细胞自噬是真核生物中进化保守的对细胞内物质进行周转的重要过程,该过程中一些损坏的蛋白或细胞器被双层膜结构的自噬小泡包裹后送入溶酶体(动物)或液泡(酵母和植物)中进行降解并得以循环利用。植物中通过序列比对鉴定了诸多自噬相关基因并分离到了部分细胞自噬功能缺陷的突变体,这些研究均推进了我们对植物细胞自噬机制和功能的了解。本文主要综述了植物细胞自噬分子机制和生理功能的研究进展。
DOI:10.16420/j.issn.0513-353x.2017-0144URL [本文引用: 1]
植物在生长发育和响应环境胁迫过程中不断产生受损的细胞器和蛋白质等,这些物质必须被及时且适宜的降解。其中,细胞自噬就是一个在进化上保守的主要降解途径。自噬体通过受体蛋白选择性地识别并靶向特异的组分和蛋白复合物,然后传递至液泡中降解,使其能够循环再利用,从而调节细胞内的关键反应过程。近些年的研究揭示了自噬在植物生命活动的多个过程中均发挥重要作用。本文中主要综述了植物细胞选择性自噬的最新研究进展,包括自噬体形成过程、货物识别、分类以及在生长发育、植物免疫和响应非生物胁迫中的作用。
DOI:10.3109/07388550903524243URLPMID:20214435 [本文引用: 1]
Abstract Reactive oxygen species (ROS) are produced in plants as byproducts during many metabolic reactions, such as photosynthesis and respiration. Oxidative stress occurs when there is a serious imbalance between the production of ROS and antioxidant defense. Generation of ROS causes rapid cell damage by triggering a chain reaction. Cells have evolved an elaborate system of enzymatic and nonenzymatic antioxidants which help to scavenge these indigenously generated ROS. Various enzymes involved in ROS-scavenging have been manipulated, over expressed or downregulated to add to the present knowledge and understanding the role of the antioxidant systems. The present article reviews the manipulation of enzymatic and nonenzymatic antioxidants in plants to enhance the environmental stress tolerance and also throws light on ROS and redox signaling, calcium signaling, and ABA signaling.
DOI:10.1128/MCB.06159-11URLPMID:3255710 [本文引用: 1]
Living cells are adaptive self-sustaining systems. They strictly depend on the sufficient supply of oxygen, energy, and nutrients from the outside in order to sustain their internal organization. However, as autonomous entities they are able to monitor and appropriately adapt to any critical fluctuation in their environment. In the case of insufficient external nutrient supply or augmented energy demands, cells start to extensively digest their own interior. This process, known as macroautophagy, comprises the transport of cytosolic portions and entire organelles to the lysosomal compartment via specific double-membrane vesicles, called autophagosomes. Although extensively upregulated under nutrient restriction, a low level of basal autophagy is likewise crucial in order to sustain the cellular homeostasis. On the other hand, cells have to avoid excessive and enduring self-digestion. The delicate balance between external energy and nutrient supply and internal production and consumption is a demanding task. The complex protein network that senses and precisely reacts to environmental changes is thus mainly regulated by rapid and reversible posttranslational modifications such as phosphorylation. This review focuses on the serine/threonine protein kinases AMP-activated protein kinase, mammalian target of rapamycin (mTOR), and unc-51-like kinase 1/2 (Ulk1/2), three interconnected major junctions within the autophagy regulating signaling network.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1083/jcb.12.1.198URLPMID:1304110
De Duve and his colleagues, a few years ago, originated the concept of lysosomes based on the biochemical analysis of isolated cytoplasmic particles (1). He reported that a number of hydrolytic enzymes are concentrated in a fraction consisting of small subcellular units which sediments between the usual mitochondrial and microsomal fractions. Subsequent to this discovery, some of these properties have been demonstrated in hepatic pericanalicular dense bodies, and Essner and Novikoff (2), using the electron microscope, have shown that the enzyme product of acid phosphatase is found exclusively in these bodies. They have also found that a variety of pigment granules in human liver cells possess both
[本文引用: 1]
DOI:10.1371/journal.pbio.0040423URLPMID:1661684 [本文引用: 1]
The protein folding capacity of the endoplasmic reticulum (ER) is regulated by the unfolded protein response (UPR). The UPR senses unfolded proteins in the ER lumen and transmits that information to the cell nucleus, where it drives a transcriptional program that is tailored to re-establish homeostasis. Using thin section electron microscopy, we found that yeast cells expand their ER volume at least 5-fold under UPR-inducing conditions. Surprisingly, we discovered that ER proliferation is accompanied by the formation of autophagosome-like structures that are densely and selectively packed with membrane stacks derived from the UPR-expanded ER. In analogy to pexophagy and mitophagy, which are autophagic processes that selectively sequester and degrade peroxisomes and mitochondria, the ER-specific autophagic process described utilizes several autophagy genes: they are induced by the UPR and are essential for the survival of cells subjected to severe ER stress. Intriguingly, cell survival does not require vacuolar proteases, indicating that ER sequestration into autophagosome-like structures, rather than their degradation, is the important step. Selective ER sequestration may help cells to maintain a new steady-state level of ER abundance even in the face of continuously accumulating unfolded proteins.
DOI:10.1016/j.bbagen.2013.09.017URLPMID:24060746 [本文引用: 1]
61Aquaporins facilitate the transmembrane diffusion of hydrogen peroxide.61Aquaporins impact on the biological membrane permeability of hydrogen peroxide.61Aquaporins modulate intra- and intercellular hydrogen peroxide fluxes.61Aquaporins contribute to physiologically important transmembrane signaling pathways.
[本文引用: 2]
[本文引用: 1]
DOI:10.1089/ars.2010.3530URLPMID:20712410 [本文引用: 1]
The tumor suppressor p53 protects organisms from most types of cancer through multiple mechanisms. The p53 gene encodes a stress-activated transcriptional factor that transcriptionally regulates a large set of genes with versatile functions. These p53-activated genes mitigate consequences of stress regulating cell viability, growth, proliferation, repair, and metabolism. Recently, we described a novel antioxidant function of p53, which is important for its tumor suppressor activity. Among the many antioxidant genes activated by p53, Sestrins (Sesns) are critical for suppression of reactive oxygen species (ROS) and protection from oxidative stress, transformation, and genomic instability. Sestrins can regulate ROS through their direct effect on antioxidant peroxiredoxin proteins and through the AMP-activated protein kinase-target of rapamycin signaling pathway. The AMP-activated protein kinase-target of rapamycin axis is critical for regulation of metabolism and autophagy, two processes associated with ROS production, and deregulation of this pathway increases vulnerability of the organism to stress, aging, and age-related diseases, including cancer. Recently, we have shown that inactivation of Sestrin in fly causes accumulation of age-associated damage. Hence, Sestrins can link p53 with aging and age-related diseases.
DOI:10.1111/j.1365-313x.2011.04816.xURLPMID:21985584
This work investigated the contribution of AtRbohD and AtRbohF to regulating defence-associated metabolism during three types of interaction: (i) incompatible and (ii) compatible interaction with Pseudomonas syringae; and (iii) intracellular oxidative stress in the catalase-deficient cat2 background. In all three cases, loss of function of either gene modulated the response of defence compounds. AtRbohF gene function was necessary for rapid and full induction of salicylic acid (SA) during compatible and incompatible interactions, and for resistance to virulent bacteria. Both artrboh mutations modulated the effects of intracellular ROS in the cat2 background, although the predominant effect was mediated by atrbohF. Loss of this gene function increased lesion formation in cat2 but uncoupled this effect from cat2-triggered induction of SA and camalexin, accumulation of glutathione and disease resistance, all of which were much lower in cat2 artbohF than in cat2. A detailed comparison of GC-TOF-MS profiles produced by the three interactions revealed considerable overlap between cat2 effects and those produced by bacterial infection in the wild-type background. Analysis of the impact of the two atrboh mutations on these profiles provided further evidence that AtRbohF interacts closely with intracellular oxidative stress to tune dynamic metabolic responses during infection. Thus, AtRbohF appears to be a key player not only in HR-related cell death but also in regulating metabolomic responses and resistance. Based on the results obtained during the three types of interaction, a model is proposed of how NADPH oxidases and intracellular ROS interact to determine the outcome of pathogen defence responses.
.
DOI:10.1111/j.1365-313X.2010.04166.xURLPMID:20136727 [本文引用: 1]
Autophagic recycling of intracellular plant constituents is maintained at a basal level under normal growth conditions but can be induced in response to nutritional demand, biotic stress, and senescence. One route requires the ubiquitin-fold proteins Autophagy-related (ATG)-8 and ATG12, which become attached to the lipid phosphatidylethanolamine (PE) and the ATG5 protein, respectively, during formation of the engulfing vesicle and delivery of its cargo to the vacuole for breakdown. Here, we genetically analyzed the conjugation machinery required for ATG8/12 modification in Arabidopsis thaliana with a focus on the two loci encoding ATG12. Whereas single atg12a and atg12b mutants lack phenotypic consequences, atg12a atg12b double mutants senesce prematurely, are hypersensitive to nitrogen and fixed carbon starvation, and fail to accumulate autophagic bodies in the vacuole. By combining mutants eliminating ATG12a/b, ATG5, or the ATG10 E2 required for their condensation with a method that unequivocally detects the ATG8-PE adduct, we also show that ATG8 lipidation requires the ATG12–ATG5 conjugate. Unlike ATG8, ATG12 does not associate with autophagic bodies, implying that its role(s) during autophagy is restricted to events before the vacuolar deposition of vesicles. The expression patterns of the ATG12a and ATG12b genes and the effects of single atg12a and atg12b mutants on forming the ATG12–ATG5 conjugate reveal that the ATG12b locus is more important during basal autophagy while the ATG12a locus is more important during induced autophagy. Taken together, we conclude that the formation of the ATG12–ATG5 adduct is essential for ATG8-mediated autophagy in plants by promoting ATG8 lipidation.
[本文引用: 1]
DOI:10.1038/cdd.2014.50URLPMID:24786830 [本文引用: 1]
Abstract Autophagy is a major nutrient recycling mechanism in plants. However, its functional connection with programmed cell death (PCD) is a topic of active debate and remains not well understood. Our previous studies established the plant metacaspase AtMC1 as a positive regulator of pathogen-triggered PCD. Here, we explored the linkage between plant autophagy and AtMC1 function in the context of pathogen-triggered PCD and aging. We observed that autophagy acts as a positive regulator of pathogen-triggered PCD in a parallel pathway to AtMC1. In addition, we unveiled an additional, pro-survival homeostatic function of AtMC1 in aging plants that acts in parallel to a similar pro-survival function of autophagy. This novel pro-survival role of AtMC1 may be functionally related to its prodomain-mediated aggregate localization and potential clearance, in agreement with recent findings using the single budding yeast metacaspase YCA1. We propose a unifying model whereby autophagy and AtMC1 are part of parallel pathways, both positively regulating HR cell death in young plants, when these functions are not masked by the cumulative stresses of aging, and negatively regulating senescence in older plants.
DOI:10.1074/jbc.M501116200URLPMID:15772072
Abstract The target of rapamycin (TOR) is a highly conserved Ser/Thr kinase that plays a central role in the control of cellular growth. TOR has a characteristic multidomain structure. Only the kinase domain has catalytic function; the other domains are assumed to mediate interactions with TOR substrates and regulators. Except for the rapamycin-binding domain, there are no high-resolution structural data available for TOR. Here, we present a structural, biophysical, and mutagenesis study of the extremely conserved COOH-terminal FATC domain. The importance of this domain for TOR function has been highlighted in several publications. We show that the FATC domain, in its oxidized form, exhibits a novel structural motif consisting of an alpha-helix and a COOH-terminal disulfide-bonded loop between two completely conserved cysteine residues. Upon reduction, the flexibility of the loop region increases dramatically. The structural data, the redox potential of the disulfide bridge, and the biochemical data of a cysteine to serine mutant indicate that the intracellular redox potential can affect the cellular amount of the TOR protein via the FATC domain. Because the amount of TOR mRNA is not changed, the redox state of the FATC disulfide bond is probably influencing the degradation of TOR.
DOI:10.1016/j.abb.2010.10.022URLPMID:21055384 [本文引用: 1]
Peroxisomes are single membrane-bounded subcellular organelles with an essentially oxidative type of metabolism and are probably the major sites of intracellular H 2O 2 production. These organelles also generate superoxide radicals ( O 2 -) and besides catalase they have a complex battery of antioxidative enzymes. In recent years the existence of l-arginine-dependent nitric oxide synthase (NOS) activity and the generation of the reactive nitrogen species (RNS) nitric oxide (NO) have been demonstrated in plant peroxisomes. The inter-cellular and intracellular NO carrier S-nitrosoglutathione (GSNO) can be generated inside peroxisomes and the presence of this RNS has been demonstrated in peroxisomes from several plant species. This review analyzes the available evidence concerning the properties of the NOS activity and the generation of the RNS messengers NO and GSNO in peroxisomes in the context of the cellular function of these organelles as a source of RNS signaling molecules. The important physiological functions displayed by NO and other RNS in intra- and inter-cellular communication in different organisms indicate that more attention should be payed to the RNS signaling function of peroxisomes in human, animal and fungal cells, where it is very likely that similar mechanisms to those found in plant peroxisomes are also operative.
DOI:10.1002/(SICI)1099-0844(199709)15:3<203::AID-CBF742>3.0.CO;2-JURLPMID:11997374 [本文引用: 1]
Peroxisomes are subcellular organelles with an essentially oxidative type of metabolism. Like chloroplasts and mitochondria, plant peroxisomes also produce superoxide radicals (${\mathrm{O}}_{2}^{\cdot -}$) and there are, at least, two sites of superoxide generation: one in the organelle matrix, the generating system being xanthine oxidase, and another site in the peroxisomal membranes dependent on NAD(P)H. In peroxisomal membranes, three integral polypeptides (PMPs) with molecular masses of 18, 29 and 32 kDa have been shown to generate ${\mathrm{O}}_{2}^{\cdot -}$ radicals. Besides catalase, several antioxidative systems have been demonstrated in plant peroxisomes, including different superoxide dismutases, the ascorbate-glutathione cycle, and three NADP-dependent dehydrogenases. A CuZn-SOD and two Mn-SODs have been purified and characterized from different types of peroxisomes. The four enzymes of the ascorbate-glutathione cycle (ascorbate peroxidase, monodehydroascorbate reductase, dehydroascorbate reductase, and glutathione reductase) as well as the antioxidants glutathione and ascorbate have been found in plant peroxisomes. The recycling of NADPH from NADP+ can be carried out in peroxisomes by three dehydrogenases: glucose-6-phosphate dehydrogenase, 6-phosphogluconate dehydrogenase, and isocitrate dehydrogenase. In the last decade, different experimental evidence has suggested the existence of cellular functions for peroxisomes related to reactive oxygen species (ROS), but the recent demonstration of the presence of nitric oxide synthase (NOS) in plant peroxisomes implies that these organelles could also have a function in plant cells as a source of signal molecules like nitric oxide (NO), superoxide radicals, hydrogen peroxide, and possibly S-nitrosoglutathione (GSNO).
DOI:10.4161/auto.6.7.12113URLPMID:20505317 [本文引用: 1]
Mounting evidence suggests that reactive oxygen species (ROS) are multifaceted signalling molecules implicated in a variety of cellular programs during physiological as well as pathological conditions. Recently, ROS produced endogenously, by deranged metabolism of cancer cells, or exogenously, by ROS-generating drugs, have been shown to promote macroautophagy, a lysosomal pathway of self-degradation with essential prosurvival functions. Several molecular aspects of the modulation of autophagy pathways by ROS have been revealed in the past years and it is now clear that these processes are mutually linked and play a crucial role in cancer progression and in response to cancer therapeutics. In this review we address the molecular mechanisms underlying the activation of autophagy pathways by ROS and focus on the role of autophagy in cancer cells responding to ROS-producing agents, which are utilized as a therapeutic modality to kill cancer cells.
.
DOI:10.4161/psb.18767URLPMID:21825107 [本文引用: 2]
The target of rapamycin (TOR) kinase integrates nutritional and stress signals to coordinately control cell growth in all eukaryotes. TOR associates with highly conserved proteins to constitute two distinct signaling complexes termed TORCI and TORC2. Inactivation of TORCI by rapamycin negatively regulates protein synthesis in most eukaryotes. Here, we report that down-regulation of TOR signaling by rapamycin in the model green alga Chlamydomonas reinhardtii resulted in pronounced phosphorylation of the endoplasmic reticulum chaperone BiP. Our results indicated that Chlamydomonas TOR regulates BiP phosphorylation through the control of protein synthesis, since rapamycin and cycloheximide have similar effects on BiP modification and protein synthesis inhibition. Modification of BiP by phosphorylation was suppressed under conditions that require the chaperone activity of BiP, such as heat shock stress or tunicamycin treatment, which inhibits N-linked glycosylation of nascent proteins in the endoplasmic reticulum. A phosphopeptide localized in the substrate-binding domain of BiP was identified in Chlamydomonas cells treated with rapamycin. This peptide contains a highly conserved threonine residue that might regulate BiP function, as demonstrated by yeast functional assays. Thus, our study has revealed a regulatory mechanism of BiP in Chlamydomonas by phosphorylation/dephosphorylation events and assigns a role to the TOR pathway in the control of BiP modification.
[本文引用: 1]
DOI:10.1146/annurev.arplant.043008.091948URLPMID:19575589 [本文引用: 1]
Abstract Photorespiration is a high-flux pathway that operates alongside carbon assimilation in C(3) plants. Because most higher plant species photosynthesize using only the C(3) pathway, photorespiration has a major impact on cellular metabolism, particularly under high light, high temperatures, and CO(2) or water deficits. Although the functions of photorespiration remain controversial, it is widely accepted that this pathway influences a wide range of processes from bioenergetics, photosystem II function, and carbon metabolism to nitrogen assimilation and respiration. Crucially, the photorespiratory pathway is a major source of H(2)O(2) in photosynthetic cells. Through H(2)O(2) production and pyridine nucleotide interactions, photorespiration makes a key contribution to cellular redox homeostasis. In so doing, it influences multiple signaling pathways, particularly those that govern plant hormonal responses controlling growth, environmental and defense responses, and programmed cell death. The potential influence of photorespiration on cell physiology and fate is thus complex and wide ranging. The genes, pathways, and signaling functions of photorespiration are considered here in the context of whole plant biology, with reference to future challenges and human interventions to diminish photorespiratory flux.
[本文引用: 1]
DOI:10.1002/bies.20493URLPMID:17041898 [本文引用: 1]
Reactive oxygen species (ROS) are known as toxic metabolic products in plants and other aerobic organisms. An elaborate and highly redundant plant ROS network, composed of antioxidant enzymes, antioxidants and ROS-producing enzymes, is responsible for maintaining ROS levels under tight control. This allows ROS to serve as signaling molecules that coordinate an astonishing range of diverse plant processes. The specificity of the biological response to ROS depends on the chemical identity of ROS, intensity of the signal, sites of production, plant developmental stage, previous stresses encountered and interactions with other signaling molecules such as nitric oxide, lipid messengers and plant hormones. Although many components of the ROS signaling network have recently been identified, the challenge remains to understand how ROS-derived signals are integrated to eventually regulate such biological processes as plant growth, development, stress adaptation and programmed cell death. BioEssays 28: 1091-1101, 2006. 2006 Wiley Periodicals, Inc.
.
DOI:10.1105/tpc.114.134692URLPMID:25829441 [本文引用: 1]
Autophagy as a conserved catabolic pathway can respond to reactive oxygen species (ROS) and plays an important role in degrading oxidized proteins in plants under various stress conditions. However, how ROS regulates autophagy in response to oxidative stresses is largely unknown. Here, we show that autophagy-related protein 3 (ATG3) interacts with the cytosolic glyceraldehyde-3-phosphate dehydrogenases (GAPCs) to regulate autophagy in Nicotiana benthamiana plants. We found that oxidative stress inhibits the interaction of ATG3 with GAPCs. Silencing of GAPCs significantly activates ATG3-dependent autophagy, while overexpression of GAPCs suppresses autophagy in N. benthamiana plants. Moreover, silencing of GAPCs enhances N gene-mediated cell death and plant resistance against both incompatible pathogens Tobacco mosaic virus and Pseudomonas syringae pv tomato DC3000, as well as compatible pathogen P. syringae pv tabaci. These results indicate that GAPCs have multiple functions in the regulation of autophagy, hypersensitive response, and plant innate immunity.
DOI:10.1007/s13238-011-1104-4URLPMID:4875296 [本文引用: 1]
Autophagy is a conserved pathway for the bulk degradation of cytoplasmic components in all eukaryotes. This process plays a critical role in the adaptation of plants to drastic changing environmental stresses such as starvation, oxidative stress, drought, salt, and pathogen invasion. This paper summarizes the current knowledge about the mechanism and roles of plant autophagy in various plant stress responses.
DOI:10.1146/annurev-phyto-072910-095333URLPMID:21370973 [本文引用: 1]
Autophagy plays an established role in the execution of senescence, starvation, and stress responses in plants. More recently, an emerging role for autophagy has been discovered during the plant innate immune response. Recent papers have shown autophagy to restrict, and conversely, to also promote programmed cell death (PCD) at the site of pathogen infection. These initial studies have piqued our excitement, but they have also revealed gaps in our understanding of plant autophagy regulation, in our ability to monitor autophagy in plant cells, and in our ability to manipulate autophagic activity. In this review, we present the most pressing questions now facing the field of plant autophagy in general, with specific focus on autophagy as it occurs during a plant-pathogen interaction. To begin to answer these questions, we place recent findings in the context of studies of autophagy and immunity in other systems, and in the context of the mammalian immune response in particular.
DOI:10.1146/annurev-genet-102808-114910URLPMID:2831538 [本文引用: 1]
Abstract Autophagy is a process of self-degradation of cellular components in which double-membrane autophagosomes sequester organelles or portions of cytosol and fuse with lysosomes or vacuoles for breakdown by resident hydrolases. Autophagy is upregulated in response to extra- or intracellular stress and signals such as starvation, growth factor deprivation, ER stress, and pathogen infection. Defective autophagy plays a significant role in human pathologies, including cancer, neurodegeneration, and infectious diseases. We present our current knowledge on the key genes composing the autophagy machinery in eukaryotes from yeast to mammalian cells and the signaling pathways that sense the status of different types of stress and induce autophagy for cell survival and homeostasis. We also review the recent advances on the molecular mechanisms that regulate the autophagy machinery at various levels, from transcriptional activation to post-translational protein modification.
DOI:10.1371/journal.pgen.1005199URLPMID:25918875
Author Summary Plants can be infected by all pathogen classes, significantly impacting crop production and food security. Innate immune responses are critical to plant survival but must be tightly regulated in order to avoid negative impacts on growth and development. Here, we investigated the role of glyceraldehyde 3-phosphate dehydrogenase (GAPDH) proteins in the model plant Arabidopsis thaliana, a mustard relative. Animals have one GAPDH isoform, which has been intensely investigated and shown to exhibit diverse moonlighting, or non-traditional, activities. Plants possess multiple GAPDH isoforms that reside in distinct sub-cellular compartments. Using a combination of genetic investigation of specific GAPDH knockouts coupled with microscopy, we found that GAPDHs regulate accumulation of reactive oxygen species and cell death in response to inoculation with the bacterial pathogen Pseudomonas syringae. The GAPC1 isoform exhibits diverse sub-cellular localizations and dynamically responds to perception of bacterial flagellin. The GAPC1 and GAPA1 isoforms also negatively regulate autophagy, which is an important component of plant immune responses. Taken together, our results demonstrate that multiple GAPDH isoforms act to negatively regulate plant defense responses. Negative regulators are important for precisely regulating the duration and amplitude of immune responses.
DOI:10.1016/j.pbi.2017.04.017URLPMID:28545004 [本文引用: 1]
Autophagy is a highly conserved degradation and recycling process that controls cellular homeostasis, stress adaptation, and programmed cell death in eukaryotes. Emerging evidence indicates that autophagy is a key regulator of plant innate immunity and contributes with both pro-death and pro-survival functions to antimicrobial defences, depending on the pathogenic lifestyle. In turn, several pathogens have co-opted and evolved strategies to manipulate host autophagy pathways to the benefit of infection, while some eukaryotic microbes require their own autophagy machinery for successful pathogenesis. In this review, we present and discuss recent advances that exemplify the important role of pro- and antimicrobial autophagy in plant athogen interactions.
DOI:10.1038/cdd.2011.43URLPMID:21527936 [本文引用: 1]
Abstract Ancient autophagy pathways are emerging as key defense modules in host eukaryotic cells against microbial pathogens. Apart from actively eliminating intracellular intruders, autophagy is also responsible for cell survival, for example by reducing the deleterious effects of endoplasmic reticulum stress. At the same time, autophagy can contribute to cellular suicide. The concurrent engagement of autophagy in these processes during infection may sometimes mask its contribution to differing pro-survival and pro-death decisions. The importance of autophagy in innate immunity in mammals is well documented, but how autophagy contributes to plant innate immunity and cell death is not that clear. A few research reports have appeared recently to shed light on the roles of autophagy in plant-pathogen interactions and in disease-associated host cell death. We present a first attempt to reconcile the results of this research.
DOI:10.1091/mbc.e08-12-1248URL [本文引用: 1]
http://www.molbiolcell.org/cgi/doi/10.1091/mbc.E08-12-1248
.
DOI:10.1104/pp.108.122770URL [本文引用: 1]
During senescence and at times of stress, plants can mobilize needed nitrogen from chloroplasts in leaves to other organs. Much of the total leaf nitrogen is allocated to the most abundant plant protein, Rubisco. While bulk degradation of the cytosol and organelles in plants occurs by autophagy, the role of autophagy in the degradation of chloroplast proteins is still unclear. We have visualized the fate of Rubisco, stroma-targeted green fluorescent protein (GFP) and DsRed, and GFP-labeled Rubisco in order to investigate the involvement of autophagy in the mobilization of stromal proteins to the vacuole. Using immunoelectron microscopy, we previously demonstrated that Rubisco is released from the chloroplast into Rubisco-containing bodies (RCBs) in naturally senescent leaves. When leaves of transgenic Arabidopsis (Arabidopsis thaliana) plants expressing stroma-targeted fluorescent proteins were incubated with concanamycin A to inhibit vacuolar H鈦-ATPase activity, spherical bodies exhibiting GFP or DsRed fluorescence without chlorophyll fluorescence were observed in the vacuolar lumen. Double-labeled immunoelectron microscopy with anti-Rubisco and anti-GFP antibodies confirmed that the fluorescent bodies correspond to RCBs. RCBs could also be visualized using GFP-labeled Rubisco directly. RCBs were not observed in leaves of a T-DNA insertion mutant in ATG5, one of the essential genes for autophagy. Stroma-targeted DsRed and GFP-ATG8 fusion proteins were observed together in autophagic bodies in the vacuole. We conclude that Rubisco and stroma-targeted fluorescent proteins can be mobilized to the vacuole through an ATG gene-dependent autophagic process without prior chloroplast destruction.
DOI:10.1105/tpc.16.00637URLPMID:28123106 [本文引用: 1]
Turnover of dysfunctional organelles is vital to maintain homeostasis in eukaryotic cells. As photosynthetic organelles, plant chloroplasts can suffer sunlight-induced damage. However, the process for turnover of entire damaged chloroplasts remains unclear. Here, we demonstrate that autophagy is responsible for the elimination of sunlight-damaged, collapsed chloroplasts in Arabidopsis thaliana. We found that vacuolar transport of entire chloroplasts, termed chlorophagy, was induced by UV-B damage to the chloroplast apparatus. This transport did not occur in autophagy-defective atg mutants, which exhibited UV-B-sensitive phenotypes and accumulated collapsed chloroplasts. Use of a fluorescent protein marker of the autophagosomal membrane allowed us to image autophagosome-mediated transport of entire chloroplasts to the central vacuole. In contrast to sugar starvation, which preferentially induced distinct type of chloroplast-targeted autophagy that transports a part of stroma via the Rubisco-containing body (RCB) pathway, photooxidative damage induced chlorophagy without prior activation of RCB production. We further showed that chlorophagy is induced by chloroplast damage caused by either artificial visible light or natural sunlight. Thus, this report establishes that an autophagic process eliminates entire chloroplasts in response to light-induced damage.
DOI:10.4161/psb.6.5.14949URL [本文引用: 1]
Autophagy is an intracellular process for the vacuolar degradation of cytoplasmic components and is important for nutrient recycling during starvation. Chloroplasts can be partially mobilized to the vacuole by autophagy via spherical bodies named Rubisco-containing bodies (RCBs). Although chloroplasts contain approximately 80% of total leaf nitrogen and represent a major carbon and nitrogen source for recycling, the relationship between leaf nutrient status and RCB production remains unclear. We analyzed the effects of nutrient factors on the appearance of RCBs in Arabidopsis leaves and postulated that a close relationship exists between the autophagic degradation of chloroplasts via RCBs and leaf carbon status but not nitrogen status in autophagy. The importance of carbohydrates in RCB production during leaf senescence can be further argued. During nitrogen-limited senescence, as leaf carbohydrates were accumulated, RCB production was strongly suppressed. During the life span of leaves, RCB production increased with the progression of leaf expansion and senescence, while the production declined in late senescent leaves with a remarkable accumulation of carbohydrates, glucose and fructose. These results suggest that RCB production may be controlled by leaf carbon status during both induced and natural senescence.
[本文引用: 1]
.
DOI:10.1083/jcb.152.3.519URL [本文引用: 1]
http://www.jcb.org/cgi/doi/10.1083/jcb.152.3.519
DOI:10.1083/jcb.151.2.263URLPMID:11038174 [本文引用: 1]
Autophagy and the Cvt pathway are examples of nonclassical vesicular transport from the cytoplasm to the vacuole via double-membrane vesicles. Apg8/Aut7, which plays an important role in the formation of such vesicles, tends to bind to membranes in spite of its hydrophilic nature. We show here that the nature of the association of Apg8 with membranes changes depending on a series of modifications of the protein itself. First, the carboxy-terminal Arg residue of newly synthesized Apg8 is removed by Apg4/Aut2, a novel cysteine protease, and a Gly residue becomes the carboxy-terminal residue of the protein that is now designated Apg8FG. Subsequently, Apg8FG forms a conjugate with an unidentified molecule "X" and thereby binds tightly to membranes. This modification requires the carboxy-terminal Gly residue of Apg8FG and Apg7, a ubiquitin E1-like enzyme. Finally, the adduct Apg8FG-X is reversed to soluble or loosely membrane-bound Apg8FG by cleavage by Apg4. The mode of action of Apg4, which cleaves both newly synthesized Apg8 and modified Apg8FG, resembles that of deubiquitinating enzymes. A reaction similar to ubiquitination is probably involved in the second modification. The reversible modification of Apg8 appears to be coupled to the membrane dynamics of autophagy and the Cvt pathway.
DOI:10.1074/jbc.M406960200URLPMID:15247238 [本文引用: 1]
The absence of the outer mitochondrial membrane protein Uth1p was found to induce resistance to rapamycin treatment and starvation, two conditions that induce the autophagic process. Biochemical studies showed the onset of a fully active autophagic activity both in wild-type and uth1 strains. On the other hand, the disorganization of the mitochondrial network induced by rapamycin treatment or 15 h of nitrogen starvation was followed in cells expressing mitochondria-targeted green fluorescent protein; a rapid colocalization of green fluorescent protein fluorescence with vacuole-selective FM4-64 labeling was observed in the wild-type but not in the uth1 strain. Degradation of mitochondrial proteins, followed by Western blot analysis, did not occur in mutant strains carrying null mutations of the vacuolar protease Pep4p, the autophagy-specific protein Atg5p, and Uth1p. These data show that, although the autophagic machinery was fully functional in the absence of Uth1p, this protein is involved in the autophagic degradation of mitochondria.
DOI:10.1016/j.jneumeth.2004.07.001URLPMID:20659276 [本文引用: 1]
Abstract The tracheary elements (TEs) of the xylem serve as the water-conducting vessels of the plant vascular system. To achieve this, TEs undergo secondary cell wall thickening and cell death, during which the cell contents are completely removed. Cell death of TEs is a typical example of developmental programmed cell death that has been suggested to be autophagic. However, little evidence of autophagy in TE differentiation has been provided. The present study demonstrates that the small GTP binding protein RabG3b plays a role in TE differentiation through its function in autophagy. Differentiating wild type TE cells were found to undergo autophagy in an Arabidopsis culture system. Both autophagy and TE formation were significantly stimulated by overexpression of a constitutively active mutant (RabG3bCA), and were inhibited in transgenic plants overexpressing a dominant negative mutant (RabG3bDN) or RabG3b RNAi (RabG3bRNAi), a brassinosteroid insensitive mutant bri1-301, and an autophagy mutant atg5-1. Taken together, our results suggest that autophagy occurs during TE differentiation, and that RabG3b, as a component of autophagy, regulates TE differentiation. 2010 The Authors. Journal compilation 2010 Blackwell Publishing Ltd.
DOI:10.4161/auto.6.8.13429URLPMID:20861670 [本文引用: 1]
The vascular system of plants consists of two conducting tissues, xylem and phloem, which differentiate from procambium cells. Xylem serves as a transporting system for water and signaling molecules and is formed by sequential developmental processes, including cell division/expansion, secondary cell wall deposition, vacuole collapse and programmed cell death (PCD). PCD during xylem differentiation is accomplished by degradation of cytoplasmic constituents, and it is required for the formation of hollow vessels, known as tracheary elements (TEs). Our recent study revealed that the small GTPase RabG3b acts as a regulator of TE differentiation through its autophagic activation. By using an Arabidopsis in vitro cell culture system, we showed that autophagy is activated during TE differentiation. Overexpression of a constitutively active RabG3b (RabG3bCA) significantly enhances both autophagy and TE differentiation, which are consistently suppressed in transgenic plants overexpressing a dominant negative form (RabG3bDN) or RabG3b RNAi (RabG3bRNAi), a brassinosteroid-insensitive mutant bri1-301 and an autophagy mutant atg5-1. On the basis of our results, we propose that RabG3b functions as a component of autophagy and regulates TE differentiation by activating the process of PCD.
.
[本文引用: 1]
DOI:10.1111/j.1365-313X.2011.04546.xURLPMID:21332848 [本文引用: 1]
In plants, autophagy has been assigned ‘pro-death’ and ‘pro-survival’ roles in controlling programmed cell death associated with microbial effector-triggered immunity. The role of autophagy in basal immunity to virulent pathogens has not been addressed systematically, however. Using several autophagy-deficient (atg) genotypes, we determined the function of autophagy in basal plant immunity. Arabidopsis mutants lacking ATG5, ATG10 and ATG18a develop spreading necrosis upon infection with the necrotrophic fungal pathogen, Alternaria brassicicola, which is accompanied by the production of reactive oxygen intermediates and by enhanced hyphal growth. Likewise, treatment with the fungal toxin fumonisin B1 causes spreading lesion formation in atg mutant genotypes. We suggest that autophagy constitutes a ‘pro-survival’ mechanism that controls the containment of host tissue-destructive microbial infections. In contrast, atg plants do not show spreading necrosis, but exhibit marked resistance against the virulent biotrophic phytopathogen, Pseudomonas syringae pv. tomato. Inducible defenses associated with basal plant immunity, such as callose production or mitogen-activated protein kinase activation, were unaltered in atg genotypes. However, phytohormone analysis revealed that salicylic acid (SA) levels in non-infected and bacteria-infected atg plants were slightly higher than those in Col-0 plants, and were accompanied by elevated SA-dependent gene expression and camalexin production. This suggests that previously undetected moderate infection-induced rises in SA result in measurably enhanced bacterial resistance, and that autophagy negatively controls SA-dependent defenses and basal immunity to bacterial infection. We infer that the way in which autophagy contributes to plant immunity to different pathogens is mechanistically diverse, and thus resembles the complex role of this process in animal innate immunity.
DOI:10.1073/pnas.1619876114URLPMID:28039434 [本文引用: 1]
Abstract Autophagy is an ancient pathway in which parts of eukaryotic cells are self-digested within the lysosome or vacuole. This process has been studied for the past seven decades; however, we are only beginning to gain a molecular understanding of the key steps required for autophagy. Originally characterized as a hormonal and starvation response, we now know that autophagy has a much broader role in biology, including organellar remodeling, protein and organelle quality control, prevention of genotoxic stress, tumor suppression, pathogen elimination, regulation of immunity and inflammation, maternal DNA inheritance, metabolism, and cellular survival. Although autophagy is usually a degradative pathway, it also participates in biosynthetic and secretory processes. Given that autophagy has a fundamental role in many essential cellular functions, it is not surprising that autophagic dysfunction is associated with a wide range of human diseases. Genetic studies in various fungi, particularly Saccharomyces cerevisiae, provided the key initial breakthrough that led to an explosion of research on the basic mechanisms and the physiological connections of autophagy to health and disease. The Nobel Committee has recognized this breakthrough by the awarding of the 2016 Nobel Prize in Physiology or Medicine for research in autophagy.
DOI:10.1016/j.cell.2005.03.007URLPMID:15907470 [本文引用: 1]
The plant innate immune response includes the hypersensitive response (HR), a form of programmed cell death (PCD). PCD must be restricted to infection sites to prevent the HR from playing a pathologic rather than protective role. Here we show that plant BECLIN 1, an ortholog of the yeast and mammalian autophagy gene ATG6/VPS30/beclin 1, functions to restrict HR PCD to infection sites. Initiation of HR PCD is normal in BECLIN 1-deficient plants, but remarkably, healthy uninfected tissue adjacent to HR lesions and leaves distal to the inoculated leaf undergo unrestricted PCD. In the HR PCD response, autophagy is induced in both pathogen-infected cells and distal uninfected cells; this is reduced in BECLIN 1-deficient plants. The restriction of HR PCD also requires orthologs of other autophagy-related genes including PI3K/ VPS34, ATG3, and ATG7. Thus, the evolutionarily conserved autophagy pathway plays an essential role in plant innate immunity and negatively regulates PCD.
DOI:10.1146/annurev-arplant-042811-105441URLPMID:22242963 [本文引用: 1]
Abstract Plants have developed sophisticated mechanisms to survive when in unfavorable environments. Autophagy is a macromolecule degradation pathway that recycles damaged or unwanted cell materials upon encountering stress conditions or during specific developmental processes. Over the past decade, our molecular and physiological understanding of plant autophagy has greatly increased. Most of the essential machinery required for autophagy seems to be conserved from yeast to plants. Plant autophagy has been shown to function in various stress responses, pathogen defense, and senescence. Some of its potential upstream regulators have also been identified. Here, we describe recent advances in our understanding of autophagy in plants, discuss areas of controversy, and highlight potential future directions in autophagy research.
[本文引用: 1]
DOI:10.4161/auto.3595URLPMID:17204848 [本文引用: 1]
ATG genes encode proteins that are required for macroautophagy, the Cvt pathway and/or pexophagy. Using the published Atg protein sequences, we have screened protein and DNA databases to identify putative functional homologs (orthologs) in 21 fungal species (yeast and filamentous fungi) of which the genome sequences were available. For comparison with Atg proteins in higher eukaryotes, also the genomes of Arabidopsis thaliana and Homo sapiens were included. This analysis demonstrated that Atg proteins required for non-selective macroautophagy are conserved from yeast to man, stressing the importance of this process in cell survival and viability. Remarkably, the A. thaliana and human genomes encode multiple proteins highly similar to specific Atg proteins (paralogs), the function of which is unknown. The Atg proteins specifically involved in the Cvt pathway and/or pexophagy showed poor conservation, and were generally not present in A. thaliana and man. Furthermore, the receptor of Cvt cargo, Atg19, was only detected in S. cerevisiae. Nevertheless, Atg11, a protein that links receptor-bound cargo (peroxisomes, Cvt bodies) to the autophagic machinery was identified in all yeast species and filamentous fungi under study. This suggests that in fungi an organism-specific form of selective autophagy may occur, for which specialized Atg proteins have evolved.
DOI:10.1093/jxb/ery010URLPMID:29365132 [本文引用: 1]
Autophagy-related proteins Atg5 and Atg7 are rate-limiting components of autophagic flux in Arabidopsis. Overexpression ofATG5orATG7genes stimulates Atg8 lipidation, autophagosome formation, and autophagic flux, leading to improved plant fitness. Autophagy is a major catabolic process whereby autophagosomes deliver cytoplasmic content to the lytic compartment for recycling. Autophagosome formation requires two ubiquitin-like systems conjugating Atg12 with Atg5, and Atg8 with lipid phosphatidylethanolamine (PE), respectively. Genetic suppression of these systems causes autophagy-deficient phenotypes with reduced fitness and longevity. We show that Atg5 and the E1-like enzyme, Atg7, are rate-limiting components of Atg8 E conjugation in Arabidopsis. Overexpression ofATG5orATG7stimulates Atg8 lipidation, autophagosome formation, and autophagic flux. It also induces transcriptional changes opposite to those observed inatg5andatg7mutants, favoring stress resistance and growth. As a result,ATG5- orATG7-overexpressing plants exhibit increased resistance to necrotrophic pathogens and oxidative stress, delayed aging and enhanced growth, seed set, and seed oil content. This work provides an experimental paradigm and mechanistic insight into genetic stimulation of autophagyin plantaand shows its efficiency for improving plant productivity.
DOI:10.1016/j.ceb.2009.12.004URLPMID:20056399 [本文引用: 1]
The Atg1/ULK complex plays an essential role in the initiation of autophagy: receiving signals of cellular nutrient status, recruiting downstream Atg proteins to the autophagosome formation site, and governing autophagosome formation. Recent studies of mammalian Atg1 homologs (ULK1 and ULK2) have identified several novel interacting proteins, FIP200, mAtg13, and Atg101. FIP200 and Atg101 are not conserved in Saccharomyces cerevisiae, despite the high conservation rates of other downstream Atg proteins between the yeast and mammals. Furthermore, through studies of the Atg1/ULK1 complex, the molecular mechanism by which (m)TORC1 regulates autophagy is now being clarified in detail.
DOI:10.1146/annurev-cellbio-092910-154005URLPMID:21801009 [本文引用: 1]
Abstract Macroautophagy is mediated by a unique organelle, the autophagosome, which encloses a portion of cytoplasm for delivery to the lysosome. Autophagosome formation is dynamically regulated by starvation and other stresses and involves complicated membrane reorganization. Since the discovery of yeast Atg-related proteins, autophagosome formation has been dissected at the molecular level. In this review we describe the molecular mechanism of autophagosome formation with particular focus on the function of Atg proteins and the long-standing discussion regarding the origin of the autophagosome membrane.
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
DOI:10.1016/j.devcel.2009.06.013URLPMID:19619494 [本文引用: 2]
Mitochondria are essential organelles that produce most of the energy for a cell, but concomitantly accumulate oxidative damage. Degradation of damaged mitochondria is critical for cell homeostasis, and this process is thought to be mediated by mitophagy, an autophagy-related pathway specific for mitochondria. However, whether mitochondria are selectively degraded, and how the autophagic machinery is targeted to mitochondria, remain largely unknown. Here we demonstrate that, in post-log phase cells under respiratory conditions, a substantial fraction of mitochondria are exclusively sequestered as cargoes and transported to the vacuole, a lytic compartment in yeast, in an autophagy-dependent manner. Interestingly, we found Atg32, a mitochondria-anchored protein essential for mitophagy that is induced during respiratory growth. In addition, our data suggest that Atg32 interacts with Atg8 and Atg11, autophagy-related proteins critical for recognition of cargo receptors. We propose that Atg32 acts as a mitophagy-specific receptor and regulates selective degradation of mitochondria.
.
.
[本文引用: 2]
[本文引用: 2]
.
[本文引用: 1]
DOI:10.1105/tpc.17.00056URLPMID:28351989 [本文引用: 1]
Abstract Eukaryotic cells use autophagy to recycle cellular components. During autophagy, autophagosomes deliver cytoplasmic contents to the vacuole or lysosome for breakdown. Mammalian cells regulate the dynamics of autophagy via ubiquitin-mediated proteolysis of autophagy proteins. Here, we show that the Arabidopsis thaliana Tumor necrosis factor Receptor-Associated Factor (TRAF)-family proteins, TRAF1a and TRAF1b (previously named MUSE14 and MUSE13, respectively) help regulate autophagy via ubiquitination. Upon starvation, cytoplasmic TRAF1a and TRAF1b translocated to autophagosomes. Knockout traf1a/b lines showed reduced tolerance to nutrient deficiency, increased salicylic acid and reactive oxygen species levels, and constitutive cell death in rosettes, resembling the phenotypes of autophagy-defective mutants. Starvation-activated autophagosome accumulation decreased in traf1a/b root cells, indicating that TRAF1a and TRAF1b function redundantly in regulating autophagosome formation. TRAF1a and TRAF1b interacted in planta with ATG6 and the RING finger E3 ligases SINAT1, SINAT2, and SINAT6 (with a truncated RING-finger domain). SINAT1 and SINAT2 require the presence of TRAF1a and TRAF1b to ubiquitinate and destabilize AUTOPHAGY PROTEIN6 (ATG6) in vivo. Conversely, starvation-induced SINAT6 reduced SINAT1- and SINAT2-mediated ubiquitination and degradation of ATG6. Consistently, SINAT1/SINAT2 and SINAT6 knockout mutants exhibited increased tolerance and sensitivity, respectively, to nutrient starvation. Therefore, TRAF1a and TRAF1b function as molecular adaptors that help regulate autophagy by modulating ATG6 stability in Arabidopsis. {copyright, serif} 2017 American Society of Plant Biologists. All rights reserved.
DOI:10.1155/2014/761264URLPMID:3920909 [本文引用: 1]
Oxidative stress is a normal phenomenon in the body. Under normal conditions, the physiologically important intracellular levels of reactive oxygen species (ROS) are maintained at low levels by various enzyme systems participating in the in vivo redox homeostasis. Therefore, oxidative stress can also be viewed as an imbalance between the prooxidants and antioxidants in the body. For the last two decades, oxidative stress has been one of the most burning topics among the biological researchers all over the world. Several reasons can be assigned to justify its importance: knowledge about reactive oxygen and nitrogen species production and metabolism; identification of biomarkers for oxidative damage; evidence relating manifestation of chronic and some acute health problems to oxidative stress; identification of various dietary antioxidants present in plant foods as bioactive molecules; and so on. This review discusses the importance of oxidative stress in the body growth and development as well as proteomic and genomic evidences of its relationship with disease development, incidence of malignancies and autoimmune disorders, increased susceptibility to bacterial, viral, and parasitic diseases, and an interplay with prooxidants and antioxidants for maintaining a sound health, which would be helpful in enhancing the knowledge of any biochemist, pathophysiologist, or medical personnel regarding this important issue.
DOI:10.1104/pp.110.169045URLPMID:21266656
Target of rapamycin (TOR) is a central regulator of cell growth, cell death, nutrition, starvation, hormone, and stress responses in diverse eukaryotes. However, very little is known about TOR signaling and the associated functional domains in plants. We have taken a genetic approach to dissect TOR functions in Arabidopsis (Arabidopsis thaliana) and report here that the kinase domain is essential for the role of TOR in embryogenesis and 45S rRNA expression. Twelve new T-DNA insertion mutants, spanning 14.2 kb of TOR-encoding genomic region, have been characterized. Nine of these share expression of defective kinase domain and embryo arrest at 16 to 32 cell stage. However, three T-DNA insertion lines affecting FATC domain displayed normal embryo development, indicating that FATC domain was dispensable in Arabidopsis. Genetic complementation showed that the TOR kinase domain alone in tor-10/tor-10 mutant background can rescue early embryo lethality and restore normal development. Overexpression of full-length TOR or kinase domain in Arabidopsis displayed developmental abnormalities in meristem, leaf, root, stem, flowering time, and senescence. We further show that TOR, especially the kinase domain, plays a role in ribosome biogenesis by activating 45S rRNA production. Of the six putative nuclear localization sequences in the kinase domain, nuclear localization sequence 6 was identified to confer TOR nuclear targeting in transient expression assays. Chromatin immunoprecipitation studies revealed that the HEAT repeat domain binds to 45 S rRNA promoter and the 5' external transcribed spacer elements motif. Together, these results show that TOR controls the embryogenesis, postembryonic development, and 45S rRNA production through its kinase domain in Arabidopsis.
DOI:10.1007/s00709-010-0190-0URLPMID:20734094 [本文引用: 1]
Autophagy is an evolutionarily conserved intracellular process for the vacuolar degradation of cytoplasmic constituents. The central structures of this pathway are newly formed double-membrane vesicles (autophagosomes) that deliver excess or damaged cell components into the vacuole or lysosome for proteolytic degradation and monomer recycling. Cellular remodeling by autophagy allows organisms to survive extensive phases of nutrient starvation and exposure to abiotic and biotic stress. Autophagy was initially studied by electron microscopy in diverse organisms, followed by molecular and genetic analyses first in yeast and subsequently in mammals and plants. Experimental data demonstrate that the basic principles, mechanisms, and components characterized in yeast are conserved in mammals and plants to a large extent. However, distinct autophagy pathways appear to differ between kingdoms. Even though direct information remains scarce particularly for plants, the picture is emerging that the signal transduction cascades triggering autophagy and the mechanisms of organelle turnover evolved further in higher eukaryotes for optimization of nutrient recycling. Here, we summarize new research data on nitrogen starvation-induced signal transduction and organelle autophagy and integrate this knowledge into plant physiology.
DOI:10.1016/j.pbi.2012.01.012URLPMID:22305521 [本文引用: 1]
The perception of nutrient and energy levels inside and outside the cell is crucial to adjust growth and metabolism to available resources. The signaling pathways centered on the conserved TOR and SnRK1/Snf1/AMPK kinases have crucial and numerous roles in nutrient and energy sensing and in translating this information into metabolic and developmental adaptations. In plants evidence is mounting that, like in other eukaryotes, these signaling pathways have pivotal and antagonistic roles in connecting external or intracellular cues to many biological processes, including ribosome biogenesis, regulation of translation, cell division, accumulation of reserves and autophagy. Data on the plant TOR pathway have been hitherto rather scarce but recent findings have shed new light on its roles in plants. Moreover, the distinctive energy metabolism of photosynthetic organisms may reveal new features of these ancestral eukaryotic signaling elements.
DOI:10.1016/j.tibs.2007.09.003URLPMID:17920280 [本文引用: 1]
Cells respond to the accumulation of unfolded proteins by activating signal transduction cascades that improve protein folding. One example of such a cascade is the unfolded protein response (UPR), which senses protein folding stress in the endoplasmic reticulum (ER) and leads to improvement in the protein folding and processing capacity of the organelle. A central paradox of the UPR, and indeed of all such stress pathways, is that the response is designed to facilitate both adaptation to stress and apoptosis, depending upon the nature and severity of the stressor. Understanding how the UPR can allow for adaptation, instead of apoptosis, is of tremendous physiological importance. Recent advances have improved our understanding of ER stress and the vertebrate UPR, which suggest possible mechanisms by which cells adapt to chronic stress.
DOI:10.1515/BC.2002.069URLPMID:12033456 [本文引用: 1]
Singlet oxygen initiates lipid peroxidation via a nonfree radical mechanism by reacting directly with unsaturated lipids to form lipid hydroperoxides (LOOHs). These LOOHs can initiate free radical chain reactions leading to membrane leakage and cell death. Here we compare the ability and mechanism by which three smallmolecule membrane antioxidants (βcarotene, αtocopherol and nitric oxide) inhibit lipid peroxidation in membranes. We demonstrate that βcarotene provides protection against singlet oxygenmediated lipid peroxidation, but does not slow free radicalmediated lipid peroxidation. α Tocopherol does not protect cells from singlet oxygen, but does inhibit free radical formation in cell membranes. Nitric oxide provides no direct protection against singlet oxygen exposure, but is an exceptional chainbreaking antioxidant as evident from its ability to blunt oxygen consumption during free radical mediated lipid peroxidation. These three smallmolecule antioxidants appear to have complementary mechanisms for the protection of cell membranes from detrimental oxidations.
DOI:10.1016/j.pbi.2013.06.020URLPMID:23880111 [本文引用: 1]
Nitric oxide (NO) and reactive oxygen species (ROS) are important signaling molecules in plants. Recent progress has been made in defining their role during plant biotic interactions. Over the last decade, their function in disease resistance has been highlighted and focused a lot of investigations. Moreover, NO and ROS have recently emerged as important players of defense responses after herbivore attacks. Besides their role in plant adaptive response development, NO and ROS have been demonstrated to be involved in symbiotic interactions between plants and microorganisms. Here we review recent data concerning these three sides of NO and ROS functions in plant biotic interactions.
[本文引用: 1]
DOI:10.1515/BC.2006.170URLPMID:17081107 [本文引用: 1]
Growing evidence supports the concept that mitochondrial metabolism and reactive oxygen species (ROS) play a major role in aging and determination of an organism's life span. Cellular signaling pathways regulating mitochondrial activity, and hence the generation of ROS and retrograde signaling events originating in mitochondria, have recently moved into the spotlight in aging research. Involvement of the energy-sensing TOR pathway in both mitochondrial signaling and determination of life span has been shown in several studies. This brief review summarizes the recent progress on how mitochondrial signaling might contribute to the aging process with a particular emphasis on TOR signaling from invertebrates to humans.
DOI:10.1105/tpc.113.116947URLPMID:24368788 [本文引用: 1]
The positioning of peroxisomes in a cell is a regulated process that is closely associated with their functions. Using this feature of the peroxisomal positioning as a criterion, we identified three Arabidopsis thaliana mutants (peroxisome unusual positioning1 [peup1], peup2, and peup4) that contain aggregated peroxisomes. We found that the PEUP1, PEUP2, and PEUP4 were identical to Autophagy-related2 (ATG2), ATG18a, and ATG7, respectively, which are involved in the autophagic system. The number of peroxisomes was increased and the peroxisomal proteins were highly accumulated in the peup1 mutant, suggesting that peroxisome degradation by autophagy (pexophagy) is deficient in the peup1 mutant. These aggregated peroxisomes contained high levels of inactive catalase and were more oxidative than those of the wild type, indicating that peroxisome aggregates comprise damaged peroxisomes. In addition, peroxisome aggregation was induced in wild-type plants by exogenous application of hydrogen peroxide. The cat2 mutant also contained peroxisome aggregates. These findings demonstrate that hydrogen peroxide as a result of catalase inactivation is the inducer of peroxisome aggregation. Furthermore, an autophagosome marker, ATG8, frequently colocalized with peroxisome aggregates, indicating that peroxisomes damaged by hydrogen peroxide are selectively degraded by autophagy in the wild type. Our data provide evidence that autophagy is crucial for quality control mechanisms for peroxisomes in Arabidopsis.
[本文引用: 1]
.
[本文引用: 1]
.
DOI:10.1111/pbi.12794URLPMID:28703378 [本文引用: 1]
Autophagy is a major and conserved pathway for delivering and recycling unwanted proteins or damaged organelles to be degraded in the vacuoles. AuTophaGy elated (ATG) protein 18a has been established as one of the essential components for autophagy occurrence inArabidopsis thaliana. We previously cloned theATG18ahomolog fromMalus domestica(MdATG18a) and monitored its responsiveness to various abiotic stresses at the transcriptional level. However, it is still unclear what its function is under abiotic stress in apple. Here, we found that heterologous expression ofMdATG18ain tomato plants markedly enhanced their tolerance to drought. Overexpression (OE) of that gene in apple plants improved their drought tolerance as well. Under drought conditions, the photosynthesis rate and antioxidant capacity were significantly elevated in OE lines when compared with the untransformed wild type (WT). Transcript levels of other important appleATGgenes were more strongly up egulated in transgenicMdATG18aOE lines than in the WT. The percentage of insoluble protein in proportion to total protein was lower and less oxidized protein accumulated in the OE lines than in the WT under drought stress. This was probably due to more autophagosomes being formed in the former. These results demonstrate that overexpression ofMdATG18ain apple plants enhances their tolerance to drought stress, probably because of greater autophagosome production and a higher frequency of autophagy. Those processes help degrade protein aggregation and limit the oxidation damage, thereby suggesting that autophagy plays important roles in the drought response.
DOI:10.4161/auto.20240URLPMID:21984698 [本文引用: 2]
Autophagy is an intracellular recycling route in eukaryotes whereby organelles and cytoplasm are sequestered in vesicles, which are subsequently delivered to the vacuole for breakdown. The process is induced by various nutrient-responsive signaling cascades converging on the Autophagy-Related 1 (ATG1)/ATG13 kinase complex. Here, we describe the ATG1/13 complex in Arabidopsis thaliana and show that it is both a regulator and a target of autophagy. Plants missing ATG13 are hypersensitive to nutrient limitations and senesce prematurely similar to mutants lacking other components of the ATG system. Synthesis of the ATG12-ATG5 and ATG8-phosphatidylethanolamine adducts, which are essential for autophagy, still occurs in ATG13-deficient plants, but the biogenesis of ATG8-decorated autophagic bodies does not, indicating that the complex regulates downstream events required for autophagosome enclosure and/or vacuolar delivery. Surprisingly, levels of the ATG 1a and ATG 13a phosphoproteins drop dramatically during nutrient starvation and rise again upon nutrient addition. This turnover is abrogated by inhibition of the ATG system, indicating that the ATG 1/13 complex becomes a target of autophagy. Consistent with this mechanism, ATG 1a is delivered to the vacuole with ATG8-decorated autophagic bodies. Given its responsiveness to nutrient demands, the turnover of the ATG 1/13 kinase likely provides a dynamic mechanism to tightly connect autophagy to a plant's nutritional status.
DOI:10.1016/j.pbi.2011.07.014URLPMID:21862390 [本文引用: 1]
Highlights? Reactive oxygen species (ROS) play a key signal transduction role in cells. ? The level of ROS in cells is controlled by the ROS gene network. ? Respiratory burst oxidase homologues (RBOHs) are key signaling nodes in the ROS network. ? RBOHs regulate a large number of important biological processes in plants. ? RBOHs integrate a multitude of signal transduction pathways with ROS signaling.
[本文引用: 1]
DOI:10.1016/j.amc.2004.10.048URLPMID:17166847 [本文引用: 1]
Autophagy is a catabolic membrane-trafficking process that occurs in all eukaryotic cells and leads to the hydrolytic degradation of cytosolic material in the vacuolar or lysosomal lumen. Mitophagy, a selective form of autophagy targeting mitochondria, is poorly understood at present. Several recent reports suggest that mitophagy is a selective process that targets damaged mitochondria, whereas other studies imply a role for mitophagy in cell death processes. In a screen for protein phosphatase homologs that functionally interact with the autophagy-dedicated protein kinase Atg1p in yeast, we have identified Aup1p, encoded by Saccharomyces cerevisiae reading frame YCR079w. Aup1p is highly similar to a family of protein phosphatase homologs in animal cells that are predicted to localize to mitochondria based on sequence analysis. Interestingly, we found that Aup1p localizes to the mitochondrial intermembrane space and is required for efficient mitophagy in stationary phase cells. Viability studies demonstrate that Aup1p is required for efficient survival of cells in prolonged stationary phase cultures, implying a pro-survival role for mitophagy under our working conditions. Our data suggest that Aup1p may be part of a signal transduction mechanism that marks mitochondria for sequestration into autophagosomes.
DOI:10.1016/j.pbi.2005.01.013URLPMID:15752997 [本文引用: 1]
The autophagic engulfment of cytoplasm and organelles and their delivery to the vacuole have long been speculated to play an essential role in bulk protein turnover in plants. Until recently, however, the importance and the mechanism(s) of action of these processes have remained obscure. Aided by the discovery of numerous orthologs of the yeast AUTOPHAGY (ATG) protein system in Arabidopsis, significant advances have been now made in understanding these processes. Both reverse genetic analyses of the Arabidopsis ATG genes and the use of the encoded proteins as cytological markers have confirmed the presence of autophagy in plants and have demonstrated its importance in nutrient recycling, especially during senescence and growth under carbon- or nitrogen-limiting conditions.
[本文引用: 1]
DOI:10.1242/jcs.02561URLPMID:16129880 [本文引用: 1]
Abstract Intracellular membrane fusion occurs with exquisite coordination and specificity. Each fusion event requires three basic components: Rab-GTPases organize the fusion site; SNARE proteins act during fusion; and N-ethylmaleimide-sensitive factor (NSF) plus its cofactor alpha-SNAP are required for recycling or activation of the fusion machinery. Whereas Rab-GTPases seem to mediate the initial membrane contact, SNAREs appear to lie at the center of the fusion process. It is known that formation of complexes between SNAREs from apposed membranes is a prerequisite for lipid bilayer mixing; however, the biophysics and many details of SNARE function are still vague. Nevertheless, recent observations are shedding light on the role of SNAREs in membrane fusion. Structural studies are revealing the mechanisms by which SNARES form complexes and interact with other proteins. Furthermore, it is now apparent that the SNARE transmembrane segment not only anchors the protein but engages in SNARE-SNARE interactions and plays an active role in fusion. Recent work indicates that the fusion process itself may comprise two stages and proceed via a hemifusion intermediate.
DOI:10.3892/etm.2017.4287URLPMID:28946008 [本文引用: 1]
Abstract Autophagy is a major pathway for degradation and recycling of cytoplasmic material, including individual proteins, aggregates, and entire organelles. Autophagic processes serve mainly survival functions in cellular homeostasis, stress adaptation and immune responses but can also have death-promoting activities in different eukaryotic organisms. In plants, the role of autophagy in the regulation of programmed cell death (PCD) remained elusive and a subject of debate. More recent evidence, however, has resulted in the consensus that autophagy can either promote or restrict different forms of PCD. Here, we present latest advances in understanding the molecular mechanisms and functions of plant autophagy and discuss their implications for life and death decisions in the context of developmental and pathogen-induced PCD. Copyright 2017 Elsevier Ltd. All rights reserved.
[本文引用: 1]
DOI:10.1104/pp.108.130013URL [本文引用: 1]
Chloroplasts contain approximately 80% of total leaf nitrogen and represent a major source of recycled nitrogen during leaf senescence. While bulk degradation of the cytosol and organelles in plants is mediated by autophagy, its role in chloroplast catabolism is largely unknown. We investigated the effects of autophagy disruption on the number and size of chloroplasts during senescence. When leaves were individually darkened, senescence was promoted similarly in both wild-type Arabidopsis (Arabidopsis thaliana) and in an autophagy-defective mutant, atg4a4b-l. The number and size of chloroplasts decreased in darkened leaves of wild type, while the number remained constant and the size decrease was suppressed in atg4a4b-2. When leaves of transgenic plants expressing stroma-targeted DsRed were individually darkened, a large accumulation of fluorescence in the vacuolar lumen was observed. Chloroplasts exhibiting chlorophyll fluorescence, as well as Rubisco-containing bodies, were also observed in the vacuole. No accumulation of stroma-targeted DsRed, chloroplasts, or Rubisco-containing bodies was observed in the vacuoles of the autophagy-defective mutant. We have succeeded in demonstrating chloroplast autophagy in living cells and provide direct evidence of chloroplast transportation into the vacuole.
[本文引用: 1]
[本文引用: 2]
DOI:10.1080/15548627.2015.1113365URLPMID:4835195 [本文引用: 1]
Microtubules, the major components of cytoskeleton, are involved in various fundamental biological processes in plants. Recent studies in mammalian cells have revealed the importance of microtubule cytoskeleton in autophagy. However, little is known about the roles of microtubules in plant autophagy. Here, we found that ATG6 interacts with TUB8/-tubulin 8 and colocalizes with microtubules in Nicotiana benthamiana. Disruption of microtubules by either silencing of tubulin genes or treatment with microtubule-depolymerizing agents in N. benthamiana reduces autophagosome formation during upregulation of nocturnal or oxidation-induced macroautophagy. Furthermore, a blockage of leaf starch degradation occurred in microtubule-disrupted cells and triggered a distinct ATG6-, ATG5- and ATG7-independent autophagic pathway termed starch excess-associated chloroplast autophagy (SEX chlorophagy) for clearance of dysfunctional chloroplasts. Our findings reveal that an intact microtubule network is important for efficient macroautophagy and leaf starch degradation.
.
DOI:10.1093/dnares/dsr024URLPMID:21795261 [本文引用: 1]
Autophagy is an intracellular degradation process for recycling macromolecules and organelles. It plays important roles in plant development and in response to nutritional demand, stress, and senescence. Organisms from yeast to plants contain many autophagy-associated genes (ATG). In this study, we found that a total of 33 ATG homologues exist in the rice [Oryza sativa L. (Os)] genome, which were classified into 13 ATG subfamilies. Six of them are alternatively spliced genes. Evolutional analysis showed that expansion of 10 OsATG homologues occurred via segmental duplication events and that the occurrence of these OsATG homologues within each subfamily was asynchronous. The Ka/Ks ratios suggested purifying selection for four duplicated OsATG homologues and positive selection for two. Calculating the dates of the duplication events indicated that all duplication events might have occurred after the origin of the grasses, from 21.43 to 66.77 million years ago. Semi-quantitative RT???PCR analysis and mining the digital expression database of rice showed that all 33 OsATG homologues could be detected in at least one cell type of the various tissues under normal or stress growth conditions, but their expression was tightly regulated. The 10 duplicated genes showed expression divergence. The expression of most OsATG homologues was regulated by at least one treatment, including hormones, abiotic and biotic stresses, and nutrient limitation. The identification of OsATG homologues showing constitutive expression or responses to environmental stimuli provides new insights for in-depth characterization of selected genes of importance in rice.<br>
DOI:10.4161/auto.3847URLPMID:17312382 [本文引用: 1]
Plant cells frequently encounter oxidative stress, leading to oxidative damage and inactivation of proteins. We have recently demonstrated that oxidative stress induces autophagy in Arabidopsis seedlings in an AtATG18a-dependent manner and that RNAi-AtATG18a transgenic lines, which are defective in autophagosome formation, are hypersensitive to reactive oxygen species. Analysis of protein oxidation indicated that oxidized proteins are degraded in the vacuole after uptake by autophagy, and this degradation is impaired in RNAi-AtATG18a lines. Our results also suggest that in the absence of a functional autophagy pathway, plants are under increased oxidative stress, even under normal growth conditions.
DOI:10.1104/pp.106.092106URLPMID:17098847 [本文引用: 1]
Upon encountering oxidative stress, proteins are oxidized extensively by highly reactive .and toxic reactive oxidative species, and these damaged, oxidized proteins need to be degraded rapidly and effectively. There are two major proteolytic systems for bulk degradation in eukaryotes, the proteasome and vacuolar autophagy. In mammalian cells, the 20S proteasome and a specific type of vacuolar autophagy, chaperone-mediated autophagy, are involved in the degradation of oxidized proteins in mild oxidative stress. However, little is known about how cells remove oxidized proteins when under severe oxidative stress. Using two macroautophagy markers, monodansylcadaverine and green fluorescent protein-AtATG8e, we here show that application of hydrogen peroxide or the reactive oxidative species inducer methyl viologen can induce macroautophagy in Arabidopsis (Arabidopsis thaliana) plants. Macroautophagy-defective RNAi-AtATG18a transgenic plants are more sensitive to methyl viologen treatment than wild-type plants and accumulate a higher level of oxidized proteins due to a lower degradation rate. In the presence of a vacuolar H-ATPase inhibitor, concanamycin A, oxidized proteins were detected in the vacuole of wild-type root cells but not RNAi-AtATG18a root cells. Together, our results indicate that autophagy is involved in degrading oxidized proteins under oxidative stress conditions in Arabidopsis.
DOI:10.1038/nature12030URLPMID:23542588 [本文引用: 1]
The daily life of photosynthetic plants revolves around sugar production, transport, storage and utilization, and the complex sugar metabolic and signaling networks integrate internal regulators and environmental cues to govern and sustain plant growth and survival. Although diverse sugar signals have emerged as pivotal regulators from embryogenesis to senescence, glucose is the most ancient... [Show full abstract]
DOI:10.1104/pp.113.229948URLPMID:24385567 [本文引用: 1]
The target of rapamycin (TOR) kinase, a master regulator that is evolutionarily conserved among yeasts (Saccharomyces cerevisiae), plants, animals, and humans, integrates nutrient and energy signaling to promote cell proliferation and growth. Recent breakthroughs made possible by integrating chemical, genetic, and genomic analyses have greatly increased our understanding of the molecular functions and dynamic regulation of the TOR kinase in photosynthetic plants. TOR signaling plays fundamental roles in embryogenesis, meristem activation, root and leaf growth, flowering, senescence, and life span determination. The molecular mechanisms underlying TOR-mediated ribosomal biogenesis, translation promotion, readjustment of metabolism, and autophagy inhibition are now being uncovered. Moreover, monitoring photosynthesis-derived Glc and bioenergetics relays has revealed that TOR orchestrates unprecedented transcriptional networks that wire central metabolism and biosynthesis for energy and biomass production. In addition, these networks integrate localized stem/progenitor cell proliferation through interorgan nutrient coordination to control developmental transitions and growth.
DOI:10.1080/15548627.2017.1320633URLPMID:28537463
Abstract Autophagy is an evolutionarily conserved catabolic process and is involved in the regulation of programmed cell death during the plant immune response. However, mechanisms regulating autophagy and cell death are incompletely understood. Here, we demonstrate that plant Bax inhibitor-1 (BI-1), a highly conserved cell death regulator, interacts with ATG6, a core autophagy-related protein. Silencing of BI-1 reduced the autophagic activity induced by both N gene-mediated resistance to Tobacco mosaic virus (TMV) and methyl viologen (MV), and enhanced N gene-mediated cell death. In contrast, overexpression of plant BI-1 increased autophagic activity and surprisingly caused autophagy-dependent cell death. These results suggest that plant BI-1 has both prosurvival and prodeath effects in different physiological contexts and both depend on autophagic activity.
DOI:10.1074/jbc.M607007200URLPMID:1828866
Eukaryotic cells have evolved strategies to respond to stress conditions. For example, autophagy in yeast is primarily a response to the stress of nutrient limitation. Autophagy is a catabolic process for the degradation and recycling of cytosolic, long lived, or aggregated proteins and excess or defective organelles. In this study, we demonstrate a new pathway for the induction of autophagy. In the endoplasmic reticulum (ER), accumulation of misfolded proteins causes stress and activates the unfolded protein response to induce the expression of chaperones and proteins involved in the recovery process. ER stress stimulated the assembly of the pre-autophagosomal structure. In addition, autophagosome formation and transport to the vacuole were stimulated in an Atg protein-dependent manner. Finally, Atg1 kinase activity reflects both the nutritional status and autophagic state of the cell; starvation-induced autophagy results in increased Atg1 kinase activity. We found that Atg1 had high kinase activity during ER stress-induced autophagy. Together, these results indicate that ER stress can induce an autophagic response.
DOI:10.1105/tpc.104.025395URLPMID:15494556 [本文引用: 2]
Autophagy is an intracellular process for vacuolar degradation of cytoplasmic components. Thus far, plant autophagy has been studied primarily using morphological analyses. A recent genome-wide search revealed significant conservation among autophagy genes (ATGs) in yeast and plants. It has not been proved, however, that Arabidopsis thalianaATG genes are required for plant autophagy. To evaluate this requirement, we examined the ubiquitination-like Atg8 lipidation system, whose component genes are all found in the Arabidopsis genome. In Arabidopsis, all nine ATG8 genes and two ATG4 genes were expressed ubiquitously and were induced further by nitrogen starvation. To establish a system monitoring autophagy in whole plants, we generated transgenic Arabidopsis expressing each green fluorescent protein TG8 fusion (GFP-ATG8). In wild-type plants, GFP-ATG8s were observed as ring shapes in the cytoplasm and were delivered to vacuolar lumens under nitrogen-starved conditions. By contrast, in a T-DNA insertion double mutant of the ATG4s (atg4a4b-1), autophagosomes were not observed, and the GFP-ATG8s were not delivered to the vacuole under nitrogen-starved conditions. In addition, we detected autophagic bodies in the vacuoles of wild-type roots but not in those of atg4a4b-1 in the presence of concanamycin A, a V-ATPase inhibitor. Biochemical analyses also provided evidence that autophagy in higher plants requires ATG proteins. The phenotypic analysis of atg4a4b-1 indicated that plant autophagy contributes to the development of a root system under conditions of nutrient limitation.
[本文引用: 1]
DOI:10.1016/j.febslet.2010.01.007URLPMID:20079356 [本文引用: 1]
Plants and plant-associated microorganisms including phytopathogens have to adapt to drastic changes in environmental conditions. Because of their immobility, plants must cope with various types of environmental stresses such as starvation, oxidative stress, drought stress, and invasion by phytopathogens during their differentiation, development, and aging processes. Here we briefly describe the early studies of plant autophagy, summarize recent studies on the molecular functions of ATG genes, and speculate on the role of autophagy in plants and phytopathogens. Autophagy regulates senescence and pathogen-induced cell death in plants, and autophagy and pexophagy play critical roles in differentiation and the invasion of host cells by phytopathogenic fungi.
DOI:10.1080/15548627.2016.1185576URLPMID:27304906 [本文引用: 1]
Macroautophagy/autophagy is a conserved catabolic process that recycles cytoplasmic material during low energy conditions. BECN1/Beclin1 (Beclin 1, autophagy related) is an essential protein for function of the class 3 phosphatidylinositol 3-kinase (PtdIns3K) complexes that play a key role in autophagy nucleation and elongation. Here, we show that AMP-activated protein kinase (AMPK) regulates autophagy by phosphorylating BECN1 at Thr388. Phosphorylation of BECN1 is required for autophagy upon glucose withdrawal. BECN1T388A, a phosphorylation defective mutant, suppresses autophagy through decreasing the interaction between PIK3C3 (phosphatidylinositol 3-kinase catalytic subunit type 3) and ATG14 (autophagy-related 14). The BECN1T388A mutant has a higher affinity for BCL2 than its wild-type counterpart; the mutant is more prone to dimer formation. Conversely, a BECN1 phosphorylation mimic mutant, T388D, has stronger binding to PIK3C3 and ATG14, and promotes higher autophagy activity than the wild-type control. These findings uncover a novel mechanism of autophagy regulation.
DOI:10.1093/dnares/dsv012URLPMID:26205094 [本文引用: 1]
Autophagy is an evolutionarily conserved mechanism in both animals and plants, which has been shown to be involved in various essential developmental processes in plants. Nicotiana tabacum is considered to be an ideal model plant and has been widely used for the study of the roles of autophagy in the processes of plant development and in the response to various stresses. However, only a few autophagy-related genes (ATGs) have been identified in tobacco up to now. Here, we identified 30 ATGs belonging to 16 different groups in tobacco through a genome-wide survey. Comprehensive expression profile analysis reveals an abroad expression pattern of these ATGs, which could be detected in all tissues tested under normal growth conditions. Our series tests further reveal that majority of ATGs are sensitive and responsive to different stresses including nutrient starvation, plant hormones, heavy metal and other abiotic stresses, suggesting a central role of autophagy, likely as an effector, in plant response to various environmental cues. This work offers a detailed survey of all ATGs in tobacco and also suggests manifold functions of autophagy in both normal plant growth and plant response to environmental stresses.<br>
显微镜技术在植物细胞自噬研究中的应用
1
2016
... Ashford和Porter (1962)用电子显微镜在小鼠肝细胞中首次观察到细胞自食(self-eating)现象, 自噬(auto- phagy)这一概念则是由溶酶体的发现者De Duve在1963年提出(
细胞自噬在植物细胞程序性死亡中的作用
1
2016
... 自噬是真核生物中普遍存在的高度保守的胞内物质降解过程, 是生物体清除异常蛋白或受损细胞器, 以维持蛋白质代谢平衡及细胞内环境稳定的有效机制(
活性氧对植物自噬调控的研究进展
1
2012
... ROS是生物有氧代谢过程中产生的一类化学性质活泼且具有较高氧化活性的分子或离子的总称(
活性氧调控植物生长发育的研究进展
1
2012
... 植物NADPH氧化酶(NADPH oxidase, NOX)也称作呼吸爆发氧化酶同源物(respiratory burst oxidase homologue, Rboh), 位于细胞质膜上, 以NADPH为电子供体, 从质膜胞质侧的NADPH接受电子并传递给O2, 经单电子还原在质膜的质外体侧形成O2·-, O2·-经细胞壁结合的SOD催化或自然歧化形成H2O2, H2O2再经Fenton反应或Haber-Weiss反应形成·OH (
植物细胞自噬研究进展
1
2018
... Ashford和Porter (1962)用电子显微镜在小鼠肝细胞中首次观察到细胞自食(self-eating)现象, 自噬(auto- phagy)这一概念则是由溶酶体的发现者De Duve在1963年提出(
细胞自噬在植物碳氮营养中作用的研究进展
1
2014
... Ashford和Porter (1962)用电子显微镜在小鼠肝细胞中首次观察到细胞自食(self-eating)现象, 自噬(auto- phagy)这一概念则是由溶酶体的发现者De Duve在1963年提出(
植物细胞自噬研究进展
1
2010
... 自噬是真核生物中普遍存在的高度保守的胞内物质降解过程, 是生物体清除异常蛋白或受损细胞器, 以维持蛋白质代谢平衡及细胞内环境稳定的有效机制(
植物细胞选择性自噬研究进展
1
2017
... 自噬是真核生物中普遍存在的高度保守的胞内物质降解过程, 是生物体清除异常蛋白或受损细胞器, 以维持蛋白质代谢平衡及细胞内环境稳定的有效机制(
Roles of enzymatic and nonenzymatic antioxidants in plants during abiotic stress
1
2010
... 经过长期进化, 植物已形成了酶促和非酶促两类有效的ROS清除机制(
Role of AMPK-mTOR-Ulk1/2 in the regulation of autophagy: cross talk, shortcuts, and feedbacks
1
2012
... TOR-ATG1是自噬的关键组分, 其催化活性为起始阶段所必需(
ATM signals to TSC2 in the cytoplasm to regulate mTORC1 in response to ROS
1
2010
... 越来越多的研究表明, 低等和高等真核生物中, TOR是线粒体功能的关键控制器, 与氧化还原新陈代谢紧密相关(
Reactive oxygen species: metabolism, oxidative stress, and signal transduction
1
2004
... 经过长期进化, 植物已形成了酶促和非酶促两类有效的ROS清除机制(
Production and scavenging of reactive oxy- gen species in chloroplasts and their functions
1
2006
... 植物NADPH氧化酶(NADPH oxidase, NOX)也称作呼吸爆发氧化酶同源物(respiratory burst oxidase homologue, Rboh), 位于细胞质膜上, 以NADPH为电子供体, 从质膜胞质侧的NADPH接受电子并传递给O2, 经单电子还原在质膜的质外体侧形成O2·-, O2·-经细胞壁结合的SOD催化或自然歧化形成H2O2, H2O2再经Fenton反应或Haber-Weiss反应形成·OH (
Cytoplasmic components in hepatic cell lysosomes
0
1962
Plant autophagy—more than a starvation response
1
2007
... 病原菌侵染能够有效触发植物细胞自噬(
Autophagy counterbalances endoplasmic reticulum expansion during the unfolded protein response
1
2006
... ER中的蛋白质折叠是高度依赖于氧化还原状态的过程.已有多项研究报道了ER应激与ROS生成之间的关联证据(
Aquaporin-facilitated transmembrane diffusion of hydrogen peroxide
1
2014
... ROS是生物有氧代谢过程中产生的一类化学性质活泼且具有较高氧化活性的分子或离子的总称(
Aging: ROS or TOR
2
2008
... 经过长期进化, 植物已形成了酶促和非酶促两类有效的ROS清除机制(
... 越来越多的研究表明, 低等和高等真核生物中, TOR是线粒体功能的关键控制器, 与氧化还原新陈代谢紧密相关(
Redox regulation: a broadening horizon
1
2005
... 正常或胁迫条件下, 植物细胞中质膜、叶绿体、线粒体、过氧化物酶体、内质网以及质外体等许多部位都能产生ROS (
Stress-responsive sestrins link p53 with redox regulation and mammalian target of rapamycin signaling
1
2011
... 越来越多的研究表明, 低等和高等真核生物中, TOR是线粒体功能的关键控制器, 与氧化还原新陈代谢紧密相关(
AtRbohF is a crucial modulator of defence-associated metabolism and a key actor in the interplay between intracellular oxidative stress and pathogenesis responses in Arabidopsis
0
2012
ATG8 lipidation and ATG8-mediated autophagy in Arabidopsis require ATG12 expressed from the differentially controlled ATG- 12A and ATG12B loci
1
2010
... 自噬小泡的扩展与闭合: ATG8酯化系统和AT- G12蛋白连接系统的最终产物ATG8-PE及ATG12- ATG5-ATG16对自噬小泡膜的扩展与囊泡内物质降解具有重要作用.该步骤可分为2个阶段.第1阶段为ATG12与ATG5共价结合(
The ATG autophagic conjugation system in maize: ATG transcripts and abundance of the ATG8-lipid adduct are regulated by development and nutrient availability
1
2009
... 现已知, 哺乳动物中ATG4是唯一已被证明的氧化还原调节靶标的ATG蛋白.该半胱氨酸蛋白酶在自噬小泡形成中起双重作用, ATG4通过脂质化系统将新合成的ATG8的C末端与PE结合, 并使ATG8脱脂从自噬小泡膜回收(
The plant metacaspase AtMC1 in pathogen-trig- gered programmed cell death and aging: functional linkage with autophagy
1
2014
... 病原菌侵染能够有效触发植物细胞自噬(
The solution structure of the FATC domain of the protein kinase target of rapamycin suggests a role for redox-dependent structural and cellular stability
0
2005
Peroxisomes as a cellular source of reactive nitrogen species signal molecules
1
2011
... 胁迫条件下产生的过量ROS常可造成细胞损伤, 这种情况下细胞必须引入更积极的机制, 方可消除受损组分并保持ROS受到控制.自噬是降解氧化分子的主要防御机制, 且在特定条件下还可能清除产生ROS的细胞器.Tolkovsky (2009)研究表明, 细胞主要通过自噬机制选择性清除受损或不需要的线粒体, 这一过程被称为线粒体自噬.酵母细胞在饥饿条件下或生长平台期会发生典型的线粒体自噬(
Reactive oxygen species, antioxidant systems and nitric oxide in peroxisomes
1
2002
... 胁迫条件下产生的过量ROS常可造成细胞损伤, 这种情况下细胞必须引入更积极的机制, 方可消除受损组分并保持ROS受到控制.自噬是降解氧化分子的主要防御机制, 且在特定条件下还可能清除产生ROS的细胞器.Tolkovsky (2009)研究表明, 细胞主要通过自噬机制选择性清除受损或不需要的线粒体, 这一过程被称为线粒体自噬.酵母细胞在饥饿条件下或生长平台期会发生典型的线粒体自噬(
ROS-mediated mechanisms of autophagy stimulation and their relevance in cancer therapy
1
2010
... ROS是生物有氧代谢过程中产生的一类化学性质活泼且具有较高氧化活性的分子或离子的总称(
Inhibition of protein synthesis by TOR inactivation revealed a conserved regulatory mechanism of the BiP chaperone in Chlamydomonas
2
2011
... 自噬过程的关键基因ATGs最早在酿酒酵母中被发现(
... 越来越多的研究表明, 低等和高等真核生物中, TOR是线粒体功能的关键控制器, 与氧化还原新陈代谢紧密相关(
Signal transduction by reactive oxygen species
1
2011
... 正常或胁迫条件下, 植物细胞中质膜、叶绿体、线粒体、过氧化物酶体、内质网以及质外体等许多部位都能产生ROS (
Photorespiratory metabolism: genes, mutants, energetics, and redox signaling
1
2009
... 胁迫条件下产生的过量ROS常可造成细胞损伤, 这种情况下细胞必须引入更积极的机制, 方可消除受损组分并保持ROS受到控制.自噬是降解氧化分子的主要防御机制, 且在特定条件下还可能清除产生ROS的细胞器.Tolkovsky (2009)研究表明, 细胞主要通过自噬机制选择性清除受损或不需要的线粒体, 这一过程被称为线粒体自噬.酵母细胞在饥饿条件下或生长平台期会发生典型的线粒体自噬(
Redox sensing and signaling associated with reactive oxygen in chloroplasts, peroxi- somes and mitochondria
1
2003
... 正常或胁迫条件下, 植物细胞中质膜、叶绿体、线粒体、过氧化物酶体、内质网以及质外体等许多部位都能产生ROS (
Reactive oxygen species as signals that modulate plant stress responses and programmed cell death
1
2006
... 传统观念认为, ROS是一类只会造成细胞损伤的毒性物质.但近年的研究表明, ROS作为一种信号分子还参与细胞增殖、分化及凋亡等多个生理过程, 在植物生长发育、胁迫适应和细胞程序性死亡(programmed cell death, PCD)过程中具有重要调节作用(
Cytoplastic glyceraldehyde-3-phosph- ate dehydrogenases interact with ATG3 to negatively regulate autophagy and immunity in Nicotiana benthamia- na
1
2015
... 病原菌侵染能够有效触发植物细胞自噬(
Role of plant autophagy in stress response
1
2011
... 自噬过程的关键基因ATGs最早在酿酒酵母中被发现(
What can plant autophagy do for an innate immune response? Annu Rev Phytopathol 49, 557-576
1
2011
... 自噬小泡的扩展与闭合: ATG8酯化系统和AT- G12蛋白连接系统的最终产物ATG8-PE及ATG12- ATG5-ATG16对自噬小泡膜的扩展与囊泡内物质降解具有重要作用.该步骤可分为2个阶段.第1阶段为ATG12与ATG5共价结合(
Regulation mechanisms and signaling pathways of autophagy
1
2009
... 自噬诱导: ATG1与ATG13互作在细胞自噬诱导中起关键作用.拟南芥atg13a/atg13b双突变体中不能形成自噬小泡, 说明ATG13是植物细胞自噬的必需组分(
Beyond glycolysis: GAPDHs are multi-functional enzymes involved in regulation of ROS, autophagy, and plant immune responses
0
2015
Autophagy as an emerging arena for plant-pathogen interactions
1
2017
... 自噬过程的关键基因ATGs最早在酿酒酵母中被发现(
Role of autophagy in disease resistance and hypersensitive response-associated cell death
1
2011
... Ashford和Porter (1962)用电子显微镜在小鼠肝细胞中首次观察到细胞自食(self-eating)现象, 自噬(auto- phagy)这一概念则是由溶酶体的发现者De Duve在1963年提出(
Nutrient-dependent mTORC1 association with the ULK1-Atg13- FIP200 complex required for autophagy
1
2009
... TOR-ATG1是自噬的关键组分, 其催化活性为起始阶段所必需(
Mobilization of Rubisco and stroma-localized fluorescent proteins of chloroplasts to the vacuole by an ATG gene-dependent autophagic process
1
2008
... 叶绿体是植物和藻类中ROS的主要来源.在黑暗引起的碳饥饿条件下, 叶绿体组分可通过自噬过程被降解(
Entire photodamaged chloroplasts are transported to the central vacuole by autophagy
1
2017
... 叶绿体是植物和藻类中ROS的主要来源.在黑暗引起的碳饥饿条件下, 叶绿体组分可通过自噬过程被降解(
The autophagic degradation of chloroplasts via Rubisco-containing bodies is specifically linked to leaf carbon status but not nitrogen status in Arabidopsis
1
2010
... 叶绿体是植物和藻类中ROS的主要来源.在黑暗引起的碳饥饿条件下, 叶绿体组分可通过自噬过程被降解(
Tor-mediated induction of autophagy via an Apg1 protein kinase complex
1
2000
... 自噬诱导: ATG1与ATG13互作在细胞自噬诱导中起关键作用.拟南芥atg13a/atg13b双突变体中不能形成自噬小泡, 说明ATG13是植物细胞自噬的必需组分(
Two distinct Vps34 phosphatidylinositol 3-kinase complexes function in autophagy and carboxypeptidase Y sorting in Saccharomyces cerevisiae
1
2001
... 自噬小泡成核: 酵母细胞中自噬小泡的成核反应与PI3K复合体密切相关.PI3K复合体包括ATG6/ VPS30、ATG14、UVRAG、VPS15和VPS34 (
The reversible modi?cation regulates the membrane-binding state of Apg8/Aut7 essential for autophagy and the cytoplasm to vacuole targeting pathway
1
2000
... 现已知, 哺乳动物中ATG4是唯一已被证明的氧化还原调节靶标的ATG蛋白.该半胱氨酸蛋白酶在自噬小泡形成中起双重作用, ATG4通过脂质化系统将新合成的ATG8的C末端与PE结合, 并使ATG8脱脂从自噬小泡膜回收(
Uth1p is involved in the autophagic degradation of mitochondria
1
2004
... 胁迫条件下产生的过量ROS常可造成细胞损伤, 这种情况下细胞必须引入更积极的机制, 方可消除受损组分并保持ROS受到控制.自噬是降解氧化分子的主要防御机制, 且在特定条件下还可能清除产生ROS的细胞器.Tolkovsky (2009)研究表明, 细胞主要通过自噬机制选择性清除受损或不需要的线粒体, 这一过程被称为线粒体自噬.酵母细胞在饥饿条件下或生长平台期会发生典型的线粒体自噬(
a). The Rab GTPase RabG3b functions in auto- phagy and contributes to tracheary element differentiation in Arabidopsis
1
2010
... NOX生成的ROS在根毛生长过程中具关键作用(
b). Role of Arabidopsis RabG3b and autophagy in tracheary element differentiation
1
2010
... NOX生成的ROS在根毛生长过程中具关键作用(
Acclimation to singlet oxygen stress in Chlamydomonas reinhardtii
1
2007
... 传统观念认为, ROS是一类只会造成细胞损伤的毒性物质.但近年的研究表明, ROS作为一种信号分子还参与细胞增殖、分化及凋亡等多个生理过程, 在植物生长发育、胁迫适应和细胞程序性死亡(programmed cell death, PCD)过程中具有重要调节作用(
Autophagy differentially controls plant basal immunity to biotrophic and necrotrophic patho- gens
1
2011
... 病原菌侵染能够有效触发植物细胞自噬(
Autophagy wins the 2016 Nobel Prize in Physiology or Medicine: breakthroughs in baker’s yeast fuel advances in biomedical research
1
2017
... Ashford和Porter (1962)用电子显微镜在小鼠肝细胞中首次观察到细胞自食(self-eating)现象, 自噬(auto- phagy)这一概念则是由溶酶体的发现者De Duve在1963年提出(
Autophagy regulates programmed cell death during the plant innate immune response
1
2005
... 病原菌侵染能够有效触发植物细胞自噬(
Autophagy: pathways for self-eating in plant cells
1
2012
... Ashford和Porter (1962)用电子显微镜在小鼠肝细胞中首次观察到细胞自食(self-eating)现象, 自噬(auto- phagy)这一概念则是由溶酶体的发现者De Duve在1963年提出(
Degradation of the endoplasmic reticulum by autophagy during endoplasmic reticulum stress in Arabidopsis
0
2012
Autophagy is required for tolerance of drought and salt stress in plants
0
2009
Endoplasmic reticulum stress and oxidative stress: a vicious cycle or a double-edged sword?
1
2007
... ER中的蛋白质折叠是高度依赖于氧化还原状态的过程.已有多项研究报道了ER应激与ROS生成之间的关联证据(
ATG genes involved in non-selective autophagy are conserved from yeast to man, but the selective Cvt and pexo- phagy pathways also require organism-speci?c genes
1
2007
... TOR-ATG1是自噬的关键组分, 其催化活性为起始阶段所必需(
Transcriptional stimulation of rate-limiting components of the autophagic pathway improves plant fitness
1
2018
... 自噬小泡的扩展与闭合: ATG8酯化系统和AT- G12蛋白连接系统的最终产物ATG8-PE及ATG12- ATG5-ATG16对自噬小泡膜的扩展与囊泡内物质降解具有重要作用.该步骤可分为2个阶段.第1阶段为ATG12与ATG5共价结合(
The role of the Atg1/ULK1 complex in autophagy regulation
1
2010
... 自噬诱导: ATG1与ATG13互作在细胞自噬诱导中起关键作用.拟南芥atg13a/atg13b双突变体中不能形成自噬小泡, 说明ATG13是植物细胞自噬的必需组分(
The role of Atg proteins in autophagosome formation
1
2011
... Ashford和Porter (1962)用电子显微镜在小鼠肝细胞中首次观察到细胞自食(self-eating)现象, 自噬(auto- phagy)这一概念则是由溶酶体的发现者De Duve在1963年提出(
Plant mitochondria and oxidative stress: electron transport, NADPH turnover and metabolism of reactive oxygen species
1
2001
... 正常或胁迫条件下, 植物细胞中质膜、叶绿体、线粒体、过氧化物酶体、内质网以及质外体等许多部位都能产生ROS (
Mutations in the Arabidopsis homolog of LST8/GβL, a partner of the target of rapamycin kinase, impair plant growth, flowering, and metabolic adaptation to long days
2
2012
... 定位于质膜的NADPH氧化酶, 以及叶绿体、线粒体、过氧化物酶体和内质网等细胞器均能产生活性氧.过量的活性氧会诱导细胞自噬, 以减少活性氧的产生并清除细胞受损组分.NADPH: 烟酰胺腺嘌呤二核苷酸磷酸; O2: 氧分子; O2·-: 超氧阴离子; H2O2: 过氧化氢; 1O2: 单线态氧
... 越来越多的研究表明, 低等和高等真核生物中, TOR是线粒体功能的关键控制器, 与氧化还原新陈代谢紧密相关(
Dynamics and diversity in autophagy mechanisms: lessons from yeast
1
2009
... TOR-ATG1是自噬的关键组分, 其催化活性为起始阶段所必需(
Mitochondria-anchored receptor Atg32 mediates degradation of mitochondria via selective autophagy
2
2009
... 自噬过程的关键基因ATGs最早在酿酒酵母中被发现(
... 胁迫条件下产生的过量ROS常可造成细胞损伤, 这种情况下细胞必须引入更积极的机制, 方可消除受损组分并保持ROS受到控制.自噬是降解氧化分子的主要防御机制, 且在特定条件下还可能清除产生ROS的细胞器.Tolkovsky (2009)研究表明, 细胞主要通过自噬机制选择性清除受损或不需要的线粒体, 这一过程被称为线粒体自噬.酵母细胞在饥饿条件下或生长平台期会发生典型的线粒体自噬(
a). Carotenoid de?ciency triggers autophagy in the model green alga Chlamydomonas reinhardtii
0
2012
Inhibition of target of rapamycin signaling and stress activate autophagy in Chlamydomonas reinhardtii
2
2010
... 研究表明, 细胞自噬参与内质网(endoplasmic reticulum, ER)胁迫导致的内质网降解过程.Liu等(2012b)通过内质网标记蛋白示踪, 发现降解的ER 可与自噬小体共定位; 此外, 通过透射电镜观察也发现在自噬小体中存在带有核糖体的ER碎片, 说明细胞自噬通路可响应ER胁迫, 进而参与内质网膜的循环利用.Xu等(2017)对其分子机制进行了深入研究, 发现定位于内质网的BI-1 (Bax Inhibitor-1)可与自噬过程中的关键组分ATG6互作, 从而招募自噬过程的其它重要组分, 起始内质网相关的自噬进程, 且这种互作关系对于BI-1诱导的细胞自噬和细胞死亡均非常重要.Yorimitsu等(2006)对酿酒酵母的研究表明, ER胁迫是自噬的强诱导信号; 且衣霉素和N-糖基化抑制剂等ER胁迫诱导剂也能有效激活自噬(
... ATG1/ATG13激酶复合物在进化过程中是保守的(
b). Reactive oxygen species and autophagy in plants and algae
2
2012
... 综上所述, 植物细胞中线粒体和叶绿体等多种细胞器均能产生ROS, 且胞内ROS的水平受到氧化、营养和内质网胁迫以及病原菌侵染等信号的调节.自噬作为一种大规模的胞质降解路径, 不仅能降解单个长寿蛋白和蛋白质复合体, 还能降解细胞器, 在细胞响应和适应不同胁迫环境中具关键作用.大量的研究表明, 不同来源的ROS对植物细胞自噬均具调控作用(
... NOX生成的ROS在根毛生长过程中具关键作用(
TOR-dependent and - independent pathways regulate autophagy in Arabidopsis thaliana
1
2017
... ATG1/ATG13激酶复合物在进化过程中是保守的(
TRAF family proteins regulate autophagy dynamics by modulating AU- TOPHAGY PROTEIN6 stability in Arabidopsis
1
2017
... 自噬是真核生物中普遍存在的高度保守的胞内物质降解过程, 是生物体清除异常蛋白或受损细胞器, 以维持蛋白质代谢平衡及细胞内环境稳定的有效机制(
Oxidative stress, prooxidants, and antioxidants: the interplay
1
2014
... ROS是生物有氧代谢过程中产生的一类化学性质活泼且具有较高氧化活性的分子或离子的总称(
Target of rapamycin regulates development and ribosomal RNA expression through kinase domain in Arabidopsis
0
2011
From signal transduction to autophagy of plant cell organelles: lessons from yeast and mammals and plant-specific features
1
2010
... Ashford和Porter (1962)用电子显微镜在小鼠肝细胞中首次观察到细胞自食(self-eating)现象, 自噬(auto- phagy)这一概念则是由溶酶体的发现者De Duve在1963年提出(
Sensing nutrient and energy status by SnRK1 and TOR kinases
1
2012
... ATG1/ATG13激酶复合物在进化过程中是保守的(
That which does not kill me makes me stronger: adapting to chronic ER stress
1
2007
... ER中的蛋白质折叠是高度依赖于氧化还原状态的过程.已有多项研究报道了ER应激与ROS生成之间的关联证据(
Mechanisms and implications of reactive oxygen species generation during the unfolded protein response: roles of endoplasmic reticulum oxidoreductases, mitochondrial electron transport, and NADPH oxidase
0
2009
Comparing beta-carotene, vitamin E and nitric oxide as membrane antioxidants
1
2002
... 经过长期进化, 植物已形成了酶促和非酶促两类有效的ROS清除机制(
Nitric oxide and reactive oxygen species in plant biotic interactions
1
2013
... 传统观念认为, ROS是一类只会造成细胞损伤的毒性物质.但近年的研究表明, ROS作为一种信号分子还参与细胞增殖、分化及凋亡等多个生理过程, 在植物生长发育、胁迫适应和细胞程序性死亡(programmed cell death, PCD)过程中具有重要调节作用(
Reactive oxygen species are essential for autophagy and speci?cally regulate the activity of Atg4
1
2007
... 现已知, 哺乳动物中ATG4是唯一已被证明的氧化还原调节靶标的ATG蛋白.该半胱氨酸蛋白酶在自噬小泡形成中起双重作用, ATG4通过脂质化系统将新合成的ATG8的C末端与PE结合, 并使ATG8脱脂从自噬小泡膜回收(
Mitochondrial signaling, TOR, and life span
1
2006
... 越来越多的研究表明, 低等和高等真核生物中, TOR是线粒体功能的关键控制器, 与氧化还原新陈代谢紧密相关(
Highly oxidized peroxisomes are selectively degraded via autophagy in Arabidopsis
1
2013
... 胁迫条件下产生的过量ROS常可造成细胞损伤, 这种情况下细胞必须引入更积极的机制, 方可消除受损组分并保持ROS受到控制.自噬是降解氧化分子的主要防御机制, 且在特定条件下还可能清除产生ROS的细胞器.Tolkovsky (2009)研究表明, 细胞主要通过自噬机制选择性清除受损或不需要的线粒体, 这一过程被称为线粒体自噬.酵母细胞在饥饿条件下或生长平台期会发生典型的线粒体自噬(
OsATG10b, an autophagosome component, is needed for cell survival against oxidative stresses in rice
1
2009
... 外源H2O2处理会对拟南芥造成严重的氧化应激, 从而诱导自噬进程(
SnRK1 activates autophagy via the TOR signaling pathway in Arabidopsis thaliana
1
2017
... ATG1/ATG13激酶复合物在进化过程中是保守的(
Improvement of drought tolerance by overexpressing Md- ATG18a is mediated by modified antioxidant system and activated autophagy in transgenic apple
1
2018
... NOX生成的ROS在根毛生长过程中具关键作用(
The ATG1/ATG13 protein kinase complex is both a regulator and a target of autophagic recycling in Arabidopsis
2
2011
... 自噬诱导: ATG1与ATG13互作在细胞自噬诱导中起关键作用.拟南芥atg13a/atg13b双突变体中不能形成自噬小泡, 说明ATG13是植物细胞自噬的必需组分(
... ATG1/ATG13激酶复合物在进化过程中是保守的(
Respiratory burst oxidases: the engines of ROS signaling
1
2011
... 植物NADPH氧化酶(NADPH oxidase, NOX)也称作呼吸爆发氧化酶同源物(respiratory burst oxidase homologue, Rboh), 位于细胞质膜上, 以NADPH为电子供体, 从质膜胞质侧的NADPH接受电子并传递给O2, 经单电子还原在质膜的质外体侧形成O2·-, O2·-经细胞壁结合的SOD催化或自然歧化形成H2O2, H2O2再经Fenton反应或Haber-Weiss反应形成·OH (
ROS in plant development
1
2010
... NOX生成的ROS在根毛生长过程中具关键作用(
Aup1p, a yeast mitochondrial protein phosphatase homolog, is required for efficient stationary phase mitophagy and cell survival
1
2007
... 胁迫条件下产生的过量ROS常可造成细胞损伤, 这种情况下细胞必须引入更积极的机制, 方可消除受损组分并保持ROS受到控制.自噬是降解氧化分子的主要防御机制, 且在特定条件下还可能清除产生ROS的细胞器.Tolkovsky (2009)研究表明, 细胞主要通过自噬机制选择性清除受损或不需要的线粒体, 这一过程被称为线粒体自噬.酵母细胞在饥饿条件下或生长平台期会发生典型的线粒体自噬(
Autophagic recycling: lessons from yeast help define the process in plants
1
2005
... 自噬的发生过程可划分为5个阶段, 即自噬诱导、自噬小泡成核、自噬小泡的扩展和闭合、自噬小泡与液泡融合、胞内成分降解及循环利用(
Mitophagy
0
2009
A recessive Arabidopsis mutant that grows photoautotrophically under salt stress shows enhanced active oxygen detoxification
1
1999
... NOX生成的ROS在根毛生长过程中具关键作用(
Functions of SNAREs in intracellular membrane fusion and lipid bilayer mixing
1
2005
... 自噬小泡与液泡融合: 自噬小泡形成后会靶向液泡进行融合.酵母细胞中的融合过程涉及SNARE蛋白、Rab GTP酶Ypt7和C类Vps/HOPS复合物等多种因子.Vps复合物和Ypt7参与融合位点复合物的招募和结合(
Autophagy as a mediator of life and death in plants
1
2017
... 自噬小泡的扩展与闭合: ATG8酯化系统和AT- G12蛋白连接系统的最终产物ATG8-PE及ATG12- ATG5-ATG16对自噬小泡膜的扩展与囊泡内物质降解具有重要作用.该步骤可分为2个阶段.第1阶段为ATG12与ATG5共价结合(
Oxygen sensing and signaling
1
2015
... 传统观念认为, ROS是一类只会造成细胞损伤的毒性物质.但近年的研究表明, ROS作为一种信号分子还参与细胞增殖、分化及凋亡等多个生理过程, 在植物生长发育、胁迫适应和细胞程序性死亡(programmed cell death, PCD)过程中具有重要调节作用(
Autophagy plays a role in chloroplast degradation during senescence in individually darkened leaves
1
2009
... 叶绿体是植物和藻类中ROS的主要来源.在黑暗引起的碳饥饿条件下, 叶绿体组分可通过自噬过程被降解(
The molecular mechanism of autophagy
1
2003
... 自噬小泡与液泡融合: 自噬小泡形成后会靶向液泡进行融合.酵母细胞中的融合过程涉及SNARE蛋白、Rab GTP酶Ypt7和C类Vps/HOPS复合物等多种因子.Vps复合物和Ypt7参与融合位点复合物的招募和结合(
Autophagic degradation of leaf starch in plants
2
2013
... 叶绿体是植物和藻类中ROS的主要来源.在黑暗引起的碳饥饿条件下, 叶绿体组分可通过自噬过程被降解(
... ;
Autophagy contributes to leaf starch degradation
0
2013
Disruption of microtubules in plants suppresses macroautophagy and triggers starch excess-associated chloroplast autophagy
1
2015
... 叶绿体是植物和藻类中ROS的主要来源.在黑暗引起的碳饥饿条件下, 叶绿体组分可通过自噬过程被降解(
Genome-wide identification, classification, and expression analysis of autophagy-associated gene homologues in rice ( Oryza sativa L.)
1
2011
... 自噬过程的关键基因ATGs最早在酿酒酵母中被发现(
a). Disruption of autophagy results in constitutive oxidative stress in Ara- bidopsis
1
2007
... 外源H2O2处理会对拟南芥造成严重的氧化应激, 从而诱导自噬进程(
b). Degradation of oxidized proteins by autophagy during oxidative stress in Arabidopsis
1
2007
... 外源H2O2处理会对拟南芥造成严重的氧化应激, 从而诱导自噬进程(
Glucose-TOR signaling reprograms the transcriptome and activates meristems
1
2013
... TOR-ATG1是自噬的关键组分, 其催化活性为起始阶段所必需(
The role of target of rapamycin signaling networks in plant growth and metabolism
1
2014
... ATG1/ATG13激酶复合物在进化过程中是保守的(
Plant Bax Inhibitor-1 interacts with ATG6 to regulate autophagy and programmed cell death
0
2017
Endoplasmic reticulum stress triggers autophagy
0
2006
Processing of ATG8s, ubiquitin-like proteins, and their deconjugation by ATG4s are essential for plant autophagy
2
2004
... 自噬诱导: ATG1与ATG13互作在细胞自噬诱导中起关键作用.拟南芥atg13a/atg13b双突变体中不能形成自噬小泡, 说明ATG13是植物细胞自噬的必需组分(
... 现已知, 哺乳动物中ATG4是唯一已被证明的氧化还原调节靶标的ATG蛋白.该半胱氨酸蛋白酶在自噬小泡形成中起双重作用, ATG4通过脂质化系统将新合成的ATG8的C末端与PE结合, 并使ATG8脱脂从自噬小泡膜回收(
Autophagy negatively regulates cell death by controlling NPR1-dependent salicylic acid signaling during senescence and the innate immune response in Arabidopsis
1
2009
... 外源H2O2处理会对拟南芥造成严重的氧化应激, 从而诱导自噬进程(
Autophagy in plants and phytopathogens
1
2010
... 自噬过程的关键基因ATGs最早在酿酒酵母中被发现(
AMPK regulates autophagy by phosphorylating BECN1 at threonine 388
1
2016
... TOR-ATG1是自噬的关键组分, 其催化活性为起始阶段所必需(
A comprehensive, genome-wide analysis of autophagy-related genes identified in tobacco suggests a central role of autophagy in plant response to various environmental cues
1
2015
... 自噬过程的关键基因ATGs最早在酿酒酵母中被发现(
备案号: 京ICP备16067583号-21
版权所有 © 2021 《植物学报》编辑部
地址:北京香山南辛村20号 邮编:100093
电话:010-62836135 010-62836131 E-mail:cbb@ibcas.ac.cn
本系统由北京玛格泰克科技发展有限公司设计开发

{kind=link}
{kind=link}