 ,1,*, 郭顺星,1,*, 田丽霞1, 严林2, 王欣11.
,1,*, 郭顺星,1,*, 田丽霞1, 严林2, 王欣11. 2.
Advances in Molecular Regulation of Sphingolipids in Plant-fungus Interactions
Tingting Shan1, Xiaomei Chen,1,*, Shunxing Guo,1,*, Lixia Tian1, Lin Yan2, Xin Wang11. 2.
通讯作者:
收稿日期:2018-04-16接受日期:2018-10-6网络出版日期:2019-07-01
| 基金资助: |
Received:2018-04-16Accepted:2018-10-6Online:2019-07-01

摘要
关键词:
Abstract
Keywords:
PDF (1324KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
单婷婷, 陈晓梅, 郭顺星, 田丽霞, 严林, 王欣. 鞘脂在植物-真菌互作中的分子调控机制研究进展. 植物学报, 2019, 54(3): 396-404 doi:10.11983/CBB18099
Shan Tingting, Chen Xiaomei, Guo Shunxing, Tian Lixia, Yan Lin, Wang Xin.
植物与真菌在长期的协同进化过程中形成了复杂的作用机制, 植物既能有效抵御病原体侵染, 又能与有益真菌共生。在植物-真菌互作过程中, 脂类如磷脂、糖脂、鞘脂(sphingolipids)和甾醇脂质参与建立两个有机体之间的膜界面(Siebers et al., 2016)。同时, 脂质还是感知病原菌、信号转导、诱导防御反应等作用的信号分子。鞘脂是在细胞膜中发现的一类生物活性脂质, 广泛存在于动物、植物和真菌中。鞘脂不仅是植物质膜、液泡膜和内膜的主要成分(Mar- kham et al., 2013), 还作为植物防御机制的第二信使, 参与多种植物应激反应。当植物被微生物侵染后, 鞘脂含量的变化和鞘脂及其磷酸化衍生物的平衡调控植物产生抗性反应(Bi et al., 2014; Yanagawa et al., 2017)。
已有许多利用遗传学方法研究细胞凋亡相关信号的报道, 涉及的信号分子主要包括Ca2+、水杨酸(salicylic acid, SA)和活性氧(reactive oxygen species, ROS)等。近几年, 研究人员通过遗传学方法对鞘脂相关性状进行了广泛分析, 相关报道主要集中在鞘脂的信号分子作用(Magnin-Robert et al., 2015), 以及鞘脂调节肿瘤、心血管、炎症以及感染性疾病等的病理过程方面(Sharma and Prakash, 2017; Ueda, 2017)。此外, 有关真菌鞘脂作为抗真菌药物的报道也较多(Healey et al., 2015)。较早有报道且数量最多的是关于动物和微生物的鞘脂研究。近20年开始出现对植物鞘脂的研究报道, 其中鞘脂在植物-真菌互作中的作用是研究热点之一。本文总结了植物和真菌鞘脂的结构及代谢特点, 综述了鞘脂参与调控植物-真菌互作的分子机制研究进展, 旨在为深入开展相关研究提供参考。
1 植物与真菌中的鞘脂化合物结构
鞘脂是含有鞘氨醇长链基团的一类结构复杂的脂类分子。典型的鞘脂结构包括3个部分: 鞘氨醇、脂肪酸和极性基团(图1)。鞘氨醇分子的2-氨基与长链脂肪酸发生酰基化反应, 1-羟基与糖和磷酸等极性基团发生醇醛缩合反应, 形成鞘脂。由于鞘氨醇和脂肪酸链长度的不同、双键和羟基取代的位置和数量差异, 以及极性头基团的种类多样, 导致鞘脂类化合物具有丰富多变的结构。图1
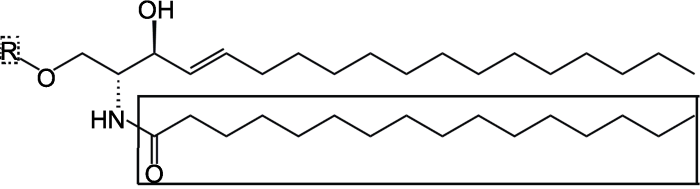 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1鞘脂的基本骨架
无框部分: 鞘氨醇; 实线框: 脂肪酸; 虚线框: 极性基团
Figure 1The structure of sphingolipids
No box: Sphingosine; Solid box: Fatty acid; Dotted box: Polar group
1.1 鞘氨醇部分
鞘脂结构中的鞘氨醇部分又被称为长链碱基(long- chain base, LCB), 链长一般为C16-C20, 以C18为主。二氢鞘氨醇(2-氨基-十八烷-1,3-二醇, dihydrosphingosine, d18:0)和4-羟基鞘氨醇(2-氨基-十八烷-1,3,4-三醇, phytosphingosine, t18:0) (图2)是植物中最常见的2种鞘氨醇类化合物(Michaelson et al., 2016)。图2
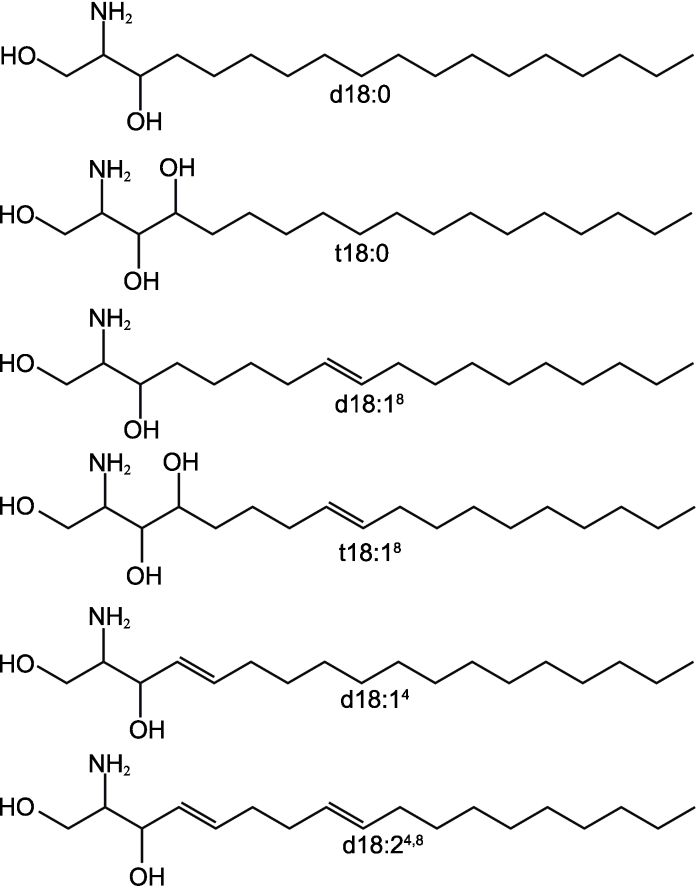 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2植物鞘脂结构中常见的鞘氨醇改自(Warnecke and Heinz, 2003)
d18:0: 二氢鞘氨醇; t18:0: 4-羟基鞘氨醇; d18:18: 2-氨基-8-十八烯-1,3-二醇; t18:18: 2-氨基-8-十八烯-1,3,4-三醇; d18:14: 2-氨基-4-十八烯-1,3-二醇; d18:24,8: 2-氨基-4,8-十八二烯-1,3-二醇
Figure 2Sphingosines in the structure of plant sphingolipids (modified from Warnecke and Heinz, 2003)
d18:0: Dihydrosphingosine; t18:0: Phytosphingosine; d18:18: Sphing-8-enine; t18:18: 4-hydroxysphing-8-enine; d18:14: Sphingosine; d18:24,8: Sphinga-4,8-dienine
植物鞘脂中LCB的结构种类较多, LCB的链长一般是C18, 在C-1、C-3和C-4易发生羟基化, 在C-4和C-8易形成双键。除了d18:0和t18:0外, 常见的植物鞘脂LCB还包括: 鞘氨醇(2-氨基-4-十八烯-1,3-二醇, sphingosine, d18:14)、2-氨基-8-十八烯-1,3-二醇(sphing-8-enine, d18:18)、2-氨基-8-十八烯-1,3,4-三醇(4-hydroxysphing-8-enine, t18:18)和2-氨基-4,8-十八二烯-1,3-二醇(鞘氨醇-4,8-二烯, sphinga-4,8- dienine, d18:24,8) (图2)。植物鞘脂中LCB的构型可变性更高, 可以在C-8顺式或反式去饱和化形成区域异构体, 主要包括(E/Z)-sphing-8-enine、(4E,8E/Z)- sphinga-4,8-dienine和(8E/Z)-4-hydroxy-8-sphingenine (Michaelson et al., 2016)。绝大多数病原真菌鞘脂的LCB具有C18长链, 在C-4和C-8形成双键, 在C-9发生甲基化(Barreto-Bergter et al., 2011)。
1.2 脂肪酸部分
鞘脂类的结构中, 鞘氨醇的2-氨基与脂肪酸发生酰基化反应生成的N-脂酰鞘氨醇类物质, 又被称为神经酰胺类物质(ceramides, Cers)。神经酰胺是鞘脂的基本结构单位, 也是最重要的中间代谢产物。植物鞘脂分子中, 脂肪酸可以酰化附着到不同的LCBs上。这些脂肪酸几乎都是α-右旋羟基化合物, 碳链长度在C14-C28之间; 脂肪酸在C-2位易发生羟基化, 在C-9位形成双键(Michaelson et al., 2016)。常见的脂肪酸包括饱和的C16、C20、C22、C24和C26的α-羟基脂肪酸。真菌鞘脂分子一般具有C16和C18链长的脂肪酸(Warnecke and Heinz, 2003)。作为真菌鞘脂研究的模式生物, 酿酒酵母(Saccharomyces cerevisiae)鞘脂的脂肪酸链长度主要在C24-C26之间, 属于超长链脂肪酸(Lindahl et al., 2017)。1.3 极性基团
极性基团是指磷酸基和糖基, 一般连接在LCB的C-1位置上。根据极性基团的种类, 可以将鞘脂分为鞘磷脂(sphingomyelins)和鞘糖脂(glycosphingolipids)两大类。前者是神经酰胺的C-1羟基连接磷酸胆碱(phosphocholine)或磷酸乙醇胺(phosphoethanolamine), 后者是神经酰胺的C-1羟基连接单糖或寡多糖。脑苷脂(cerebrosides)和糖基肌醇磷酰基神经酰胺(glycosylinositol phosphorylceramides, GIPCs)是2类主要的鞘糖脂。脑苷脂类中最常见的是葡萄糖基神经酰胺(glucosylceramides, GlcCers)和半乳糖基神经酰胺(galactosylceramides, GalCers)。GlcCers在真菌、植物和动物体内普遍存在, 而GalCers只存在于真菌和动物体内(Barreto-Bergter et al., 2011)。GlcCers和GIPCs是植物中含量最高的鞘脂类物质, 是植物细胞的质膜、液泡膜和内质网膜的重要组成部分(Warnecke and Heinz, 2003; Chen et al., 2012; Michaelson et al., 2016)。目前已经在许多真菌中发现GlcCers的合成途径, 如毕赤酵母(Pichia pastoris)、白色念珠菌(Candida albicans)、新型隐球菌(Cryptococcus neoformans)和烟曲霉(Aspergillus fumigatus) (Rittershaus et al., 2006)。除GlcCers以外, 一些真菌还可以产生GalCers, 如米曲霉(A. oryzae)、酱油曲霉(A. sojae)和细毛曲霉(A. awamori) (Tani et al., 2014)。在酿酒酵母中, 属于GIPCs类型的甘露糖肌醇磷脂酰神经酰胺(mannose inositol phosphoryl ceramide, MIPC)以及甘露糖-(肌糖磷脂酰)2-神经酰胺(mannosyl diinositol phosphoryl ceramide, M(IP)2C)是质膜中主要的鞘脂成分(Hechtberger et al., 1994)。
2 植物与真菌中的鞘脂代谢
植物和真菌体内鞘脂生物合成的第1步反应都是丝氨酸和棕榈酰辅酶A在丝氨酸棕榈酰转移酶(SPT)作用下缩合生成3-酮基二氢鞘氨醇(图3), 随后经3-酮基二氢鞘氨醇还原酶(KDHR)还原生成鞘氨醇。鞘氨醇是鞘脂代谢途径关键的中间产物之一。鞘氨醇在(二氢)神经酰胺合酶(CerS)的作用下生成二氢神经酰胺, 随后在二氢神经酰胺去饱和酶(DES)作用下合成最简单的鞘脂类化合物神经酰胺。Cer是鞘脂代谢途径的另一个关键中间产物, 其合成是生物体内各种结构复杂的鞘脂类化合物生物合成的起点(刘润华等, 2009)。从Cer开始的鞘脂生物合成途径主要有5条(图 3), 分别生成肌醇磷脂酰神经酰胺(inositol phosphoryl ceramide, IPC)、半乳糖神经酰胺、葡糖神经酰胺、鞘磷脂和神经酰胺-1-磷酸(ceramide-1-phosphate, C1P)。Cer经鞘磷脂合酶(SMS)催化生成鞘磷脂, 经神经酰胺半乳糖基转移酶(CGT)催化生成半乳糖神经酰胺, 这2条途径是真菌特有的(图3黑色加粗边框)。Cer在肌醇磷脂酰神经酰胺合酶(IPCS)的作用下生成IPC, 此途径在植物和真菌中都存在。IPC在葡萄糖肌醇磷脂酰神经酰胺醛酸基转移酶(IPUT)的作用下生成多种糖基肌醇磷脂酰神经酰胺(GIPCs)。Tartaglio等(2017)报道, 拟南芥(Arabidopsis thaliana) IPUT1可以催化IPC生成GIPCs。而在真菌中, IPC经甘露糖肌醇磷脂酰神经酰胺合酶(MIPCS)催化生成MIPC, MIPC可以在肌醇磷酸转移酶(IPT)的作用下再加入1个肌醇磷脂酰基生成M(IP)2C (Rollin-pinheiro et al., 2016)。
图3
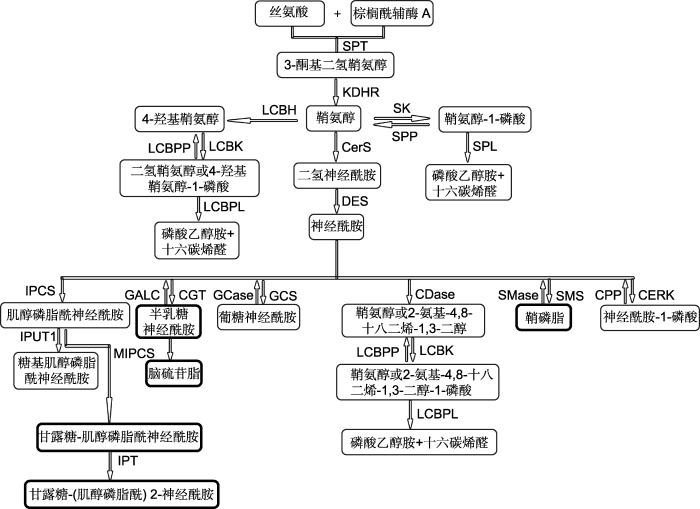 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3植物和真菌鞘脂代谢途径(改自Michaelson et al., 2016; Rollin-pinheiro et al., 2016)
黑色加粗边框为真菌特有的产物。SPT: 丝氨酸棕榈酰转移酶; KDHR: 3-酮二氢鞘氨醇还原酶; CerS: (二氢)神经酰胺合酶; DES: 二氢神经酰胺去饱和酶; SK: 鞘氨醇激酶; SPP: 鞘氨醇-1-磷酸磷酸酶; SPL: 鞘氨醇-1-磷酸裂合酶; LCBH: 鞘氨醇羟化酶; CERK: 神经酰胺激酶; CPP: 神经酰胺-1-磷酸磷酸酶; SMS: 鞘磷脂合酶; SMase: 鞘磷脂酶; LCBK: LCB激酶; LCBPP: LCB-1-磷酸磷酸酶; LCBPL: LCB-1-磷酸裂合酶; IPCS: 肌醇磷脂酰神经酰胺合酶; IPUT1: 葡萄糖肌醇磷脂酰神经酰胺醛酸基转移酶1; MIPCS: 甘露糖肌醇磷脂酰神经酰胺合酶; IPT: 肌醇磷酸转移酶; CGT: 神经酰胺半乳糖基转移酶; GALC: 半乳糖神经酰胺酶; GCS: 葡糖基神经酰胺合酶; GCase: 葡糖基神经酰胺酶; CDase: 神经鞘氨醇酶
Figure 3Sphingolipid biosynthesis in plant and fungi (modified from Michaelson et al., 2016; Rollin-pinheiro et al., 2016)
The bold border boxes are unique products of fungi. SPT: Serine palmitoyltransferase; KDHR: 3-ketodihydrosphingosine reductase; CerS: (dihydro) Ceramide synthase; DES: Dihydroceramide desaturase; SK: Sphingosine kinase; SPP: higoine- 1-phosphate phosphatases; SPL: Sphingosine-1-phosphate lyase; LCBH: LCB hydroxylase; CERK: Ceramide kinase; CPP: Ceramide-1-phosphate phosphatases; SMS: Sphingomyelin synthase; SMase: Sphingomyelinases; LCBK: LCB kinase; LCBPP: LCB-1-phosphate phosphatases; LCBPL: LCB-1-phosphate lyase; IPCS: Inositol phosphoryl ceramide synthase; IPUT1: Inositol phosphorylceramide glucuronosyl transferases 1; MIPCS: Mannosylinositol phosphorylceramide synthase; IPT: Inositol phosphoryl transferase; CGT: Galactosyltransferase; GALC: Galactosylceramidase; GCS: Glucosylceramide synthase; GCase: Glucosylceramidase; CDase: Ceramidase
3 鞘脂在植物-真菌互作中的分子调控机制
3.1 植物鞘脂
植物的鞘脂及其代谢产物是具有信号转导作用的活性脂质分子, 它们不仅能够调节细胞的生长、分化和增殖, 而且在植物防病和抗病方面发挥重要作用(Michaelson et al., 2016)。程序性细胞死亡(programmed cell death, PCD)是在一定的生理或病理条件下, 植物为了更好地适应生存环境而采取的一种由基因调控的细胞自我消亡。植物鞘脂及其代谢产物能够诱导植物细胞发生PCD。植物与病原菌的不亲和互作引起植物发生的过敏反应(hypersensitive response, HR)就是一种最常见的PCD。HR的发生依赖水杨酸信号通路, 也涉及ROS的形成(Shi et al., 2007)。鞘脂类代谢在调节植物PCD的过程中发挥重要作用。LCBs等鞘脂类信号分子诱导植物发生PCD的信号通路错综复杂, 既涉及Ca2+和ROS等信号分子, 也涉及SA信号和丝裂原活化蛋白激酶(mitogen-activated protein kinase, MAPK)通路等信号转导途径。Ca2+是细胞内信号转导的第二信使。对小麦(Triticum aestivum)根细胞的研究发现, 细胞质Ca2+浓度升高诱导线粒体通透性转换孔(permeability transition pore, PTP)开放, 使线粒体体积增大, 这是细胞发生PCD的前提(Virolainen et al., 2002)。细胞内游离钙浓度的变化是鞘脂及其代谢产物诱导细胞发生PCD的必要条件。Lachaud等(2010)研究表明, 用DHS (D-erythro-sphinganine)处理烟草(Nicotiana tabacum) BY-2细胞, 能引起细胞质和细胞核内Ca2+浓度升高, 后者在DHS诱导BY-2细胞发生PC
的过程中发挥关键作用。Townley等(2005)报道, 在C2-Ceramide (N-acetylsphingosine, N-乙酰基神经鞘氨醇)诱导拟南芥悬浮培养细胞发生PCD之前, 会在细胞内先形成钙瞬变(calcium transient), 而抑制钙瞬变的发生可以阻止细胞死亡。
ROS是需氧细胞在代谢过程中产生的一系列活性氧簇, 参与植物衰老、胁迫条件下和过敏反应中的PCD; 过氧化氢(H2O2)和超氧阴离子(O2-.)是导致细胞发生PCD的2类ROS; 但ROS究竟是细胞发生PCD的引发者, 还是仅为PCD中的一个作用因素, 目前尚不明确(林久生和王根轩, 2001)。Shi等(2007)通过研究LCBs在调节拟南芥产生活性氧中间体和发生PCD中的作用, 发现t18:0、d18:0和d18:1等LCBs处理能诱导H2O2和O2-.的积累, 进而发生PCD, 且这种PCD诱导作用具有剂量依赖性; 但这些LCBs的1-磷酸化产物均不具有PCD诱导作用; 此外, d18:0的1-磷酸化产物能特异性地阻断d18:0引起的ROS积累和后续的PCD发生, 且作用强度具有剂量依赖性, 表明维持游离LCBs与其磷酸化产物的体内平衡是细胞得以生存的决定性因素。Yanagawa等(2017)的研究也得出类似的结论: 伏马菌素B1 (fumonisin B1, FB1)是一种细胞死亡的诱导剂, 能够干扰植物的鞘脂代谢; 拟南芥中LCBP/LCB比与FB1诱导发生细胞死亡的程度相关, LCBP/LCB的体内平衡对于拟南芥的FB1耐受性以及调控细胞存亡至关重要。
RPW8是一种拟南芥抗性基因(R), 通过SA信号通路触发植物产生过敏反应来限制高氏白粉菌属(Golovinomyces)真菌引起的白粉病。Wang等(2008)报道, ERH1编码一种植物肌醇磷脂酰神经酰胺合酶(IPCS), 是RPW8潜在的转录调控因子。拟南芥erh1突变体叶片接种G. cichoracearum UCSC1后, 突变体SA含量增加, RPW8转录增强, 突变体内Cer的含量更高, RPW8-介导的细胞死亡和抗性反应增强, 证明Cer对于调节植物发生PCD有重要作用。Bi等(2014)研究表明, Cer与其磷酸化产物的平衡可以调控拟南芥发生PCD及早期抗真菌反应。ACD5编码神经酰胺激酶(CERK), 被灰霉菌(Botrytis cinerea)侵染后, 与野生型拟南芥相比, acd5突变体体内出现Cer的时间更早且积累水平更高, 胼胝质沉积减少, 线粒体H2O2含量增加; 此外, 突变体很早就表现出生长缺陷, 以限制灰霉菌孢子的萌发及生长。
MAPKs是一组能被不同种类的细胞外刺激激活的丝氨酸-苏氨酸蛋白激酶, 能够将信号从细胞表面转导至细胞核内部。Saucedo-García等(2011)通过研究拟南芥中LCBs积累与发生PCD的关系, 发现MAPK6在LCBs诱导发生PCD的过程中发挥信号转导(transducer)的作用, 从而介导PCD的下游过程。Mortimer等(2013)报道拟南芥GONST1 (golgi-localized nucleotide sugar transporter)是一种二磷酸尿苷(GDP)糖转运蛋白, 特异性地向高尔基体内膜转运GDP-Man, 用于GIPCs合成; 拟南芥gonst1突变体的鞘糖脂甘露糖基化异常, 植株对SA水平升高具有构成性的超敏反应。这表明GIPCs的糖基化修饰可以影响鞘脂功能, 进而影响植物的生长发育。
3.2 真菌鞘脂
鞘脂中的鞘糖脂是真菌细胞膜磷脂双分子层的组成成分。GlcCers是一类最简单的中性鞘糖脂, 能影响真菌的生长发育和病原真菌的致病力。植物能够识别真菌GlcCers和脑苷脂类, 并针对真菌的侵染发起特异性防御反应。一些与植物鞘脂LCBs结构类似的真菌毒素通过干扰植物鞘脂的代谢而引起植物死亡。3.2.1 真菌GlcCers调控菌丝生长, 影响真菌致病性
真菌GlcCers代谢的改变能够影响真菌的生长发育, 及其侵染宿主的能力。禾谷镰孢菌(Fusarium graminearum)的菌丝顶端富含GlcCers, 其突变体ΔFggcs1缺少GlcCer合成酶(GCS)基因, 不能合成GlcCers。与野生型相比, ΔFggcs1分生孢子短, 细胞数量少, 结构发生了很大改变; 菌丝短, 气生菌丝稀少, 细胞极性生长受到破坏; 菌丝体对细胞壁降解酶具抗性; 尽管ΔFggcs1仍能侵染小麦穗和玉米(Zea mays)须, 但菌丝体在植物组织中的延伸能力显著降低, 致病力减弱(Ramamoorthy et al., 2007)。真菌GlcCers结构具有高度的保守性, 其LCBs部分的C-9甲基对于真菌的生长发育具有重要作用。FgMT2是禾谷镰孢菌的一个鞘脂C-9甲基转移酶(C-9-MT)基因, 缺少FgMT2的禾谷镰孢菌突变体ΔFgmt2只能产生25%-35%的C-9甲基化GlcCer。ΔFgmt2表现出严重的生长缺陷, 并产生异常分生孢子, 真菌毒力降低; 被其侵染后, 小麦的病情指数(disease index)大幅度降低, 拟南芥的症状发展明显滞后(Ramamoorthy et al., 2009)。PdGcs1是柑橘绿霉菌(Penicillium digitatum)中编码GCS的关键基因, 缺失PdGcs1会导致柑橘绿霉菌的GlcCers合成量大幅度降低, 菌丝生长缓慢, 孢子萌发延迟, 并降低柑橘绿霉菌对柑橘属植物的毒力(Zhu et al., 2014)。
3.2.2 真菌GlcCers是一些植物防御素的受体
植物防御素(defensins)是一类具有抗真菌和抗细菌作用的多肽。GlcCers被认为是一些植物防御素的受体。在体外, 一些富含半胱氨酸、具有抗真菌作用的植物防御素需要在真菌GlcCers存在的条件下才能发挥作用, 如萝卜(Raphanus sativus)种子产生的植物防御素RsAFP2 (Thevissen et al., 2004)。体内不能产生GlcCers的禾谷镰孢菌突变体ΔFggcs1对苜蓿(Medicago sativa)种子产生的植物防御素MsDef1具有抗性(Ramamoorthy et al., 2007)。Ramamoorthy等(2009)研究表明, 缺少C-9-MT基因的禾谷镰孢菌突变体ΔFgmt1和ΔFgmt2以及毕赤酵母突变体ΔPpmt1均保留了对植物防御素MsDef1和RsAFP2的敏感性, 说明真菌GlcCers分子中LCB的C-9甲基对于植物防御素发挥抗真菌作用并不必要, 他们推测MsDef1和RsAFP2的作用与真菌GlcCers分子脂肪酸链的Δ3双键有关。AFP1 (Antifungal Peptide 1)是芥菜(Brassica juncea)产生的植物防御素, 能够抑制白色念珠菌生长。Oguro等(2014)报道, 真菌GlcCers分子中的C9-甲基影响AFP1的作用。大丽花(Dahlia merckii)植物防御素DmAMP1通过与酿酒酵母质膜上的糖鞘脂结合, 或者与糖鞘脂固定的糖磷脂酰肌醇锚定蛋白(GPI anchored protein)相结合来抑制真菌的生长(Thevissen et al., 2003)。Aerts等(2006)发现, DmAMP1与M(IP)2C的结合破坏了酿酒酵母细胞膜的磷脂双分子层, 导致质膜流动性变差, 物质运输效率降低, 从而抑制真菌生长。
3.2.3 真菌脑苷脂诱导植物抗性反应
真菌脑苷脂是一类能激活植物防御反应的诱导子。Umemura等(2000)报道, 田间实验中, 向水稻(Ory- za sativa)叶片喷施来自稻瘟病病原菌Magnaporthe grisea的脑苷酯, 能诱导水稻叶片合成phytocassane A、B、C和D, 以及momilactone A和B等植保素(phytoalexins, PAs), 且作用强度与脑苷脂用量呈正相关; 此外, 叶片中一些编码病程相关蛋白(pathogenesis related protein, PR protein)(如PBZ1、β-葡聚糖酶、几丁质酶和过氧化物酶)的基因被快速诱导表达。用真菌脑苷脂处理水稻悬浮培养细胞, 也能诱导细胞积累PAs和PR蛋白; 向细胞悬浮液中添加Ca2+通道抑制剂, 可以显著抑制真菌脑苷脂诱导的PAs合成, 表明Ca2+在脑苷脂诱导的抗性响应中发挥重要作用(Umemura et al., 2002)。经丝核菌属(Rhiz- octonia sp.)真菌培养物中分离的脑苷脂处理后, 可以显著提高珍珠粟(Pennisetum glaucum)对病原菌Sclerospora graminicola引起的霜霉病的抗性, 从而大幅提高珍珠粟产量(Deepak et al., 2003)。
3.2.4 真菌毒素干扰植物鞘脂合成
SAMs (sphinganine-analog mycotoxins)是真菌产生的一类与鞘氨醇结构类似的真菌毒素。SAMs与植物LCBs竞争结合CerS, 导致植物体内鞘脂合成前体物质LCBs积累, 鞘脂合成减少, 引发植物PCD或坏死(Brandwagt et al., 2002)。串珠镰孢菌(F. moniliforme)产生的FB1 (fumonisins B1)和番茄(Lycopersicon esculentum)病原菌Alternaria alternata f. sp. lycopersici产生的AAL毒素都属于SAMs。FB1是CerS的抑制剂, FB1处理可引起拟南芥体内LCBs水平升高, 进而诱导植物发生PCD (Shi et al., 2007); SPT抑制剂myriocin通过减少拟南芥LCB合成而阻断LCB的积累, 从而抑制FB1诱导的PCD反应(Saucedo-García et al., 2011)。AAL毒素是CerS抑制剂, 可引起番茄体内3-酮二氢鞘氨醇和d18:0的积累, 进而诱导番茄叶片发生PCD, 并最终导致植株坏死(Spassieva et al., 2002)。列当属(Orobanche)植物也对AAL毒素敏感; AAL毒素诱发植物体内t18:0、d18:0和d18:1等游离鞘氨醇的积累, 导致植物鞘脂代谢紊乱(De Zélicourt et al., 2009)。
4 展望
在植物-细菌和植物-真菌的相互作用中, 植物鞘脂都是主要的信号分子, 可诱导植物产生抗性反应(Peer et al., 2010; Magnin-Robert et al., 2015)。但是, 细菌鞘脂和真菌鞘脂的作用有差异。典型的革兰氏阴性细菌细胞膜含有脂多糖(lipopolysaccharides, LPSs), 鞘氨醇单胞菌(Sphingomonas)等一些厌氧细菌的细胞膜上含有鞘脂(Olsen and Jantzen, 2001)。对细菌鞘脂在动物-细菌及植物-细菌相互作用的研究表明, 动物和植物天然免疫系统识别鞘氨醇单胞菌细胞膜上鞘糖脂(GSLs)的能力弱, 导致鞘氨醇单胞菌的侵染能力增强(Arora and Porcelli, 2008)。在植物-真菌的相互作用中, 真菌鞘脂发挥的作用更为复杂, 根据其对植物的作用性质, 可以分为有利和有害2个方面。真菌鞘糖脂中的GlcCers是一些植物防御素的结合位点, 脑苷脂能诱导植物产生防卫反应, 提高植物抗病性, 这些作用有利于植物与真菌建立起互利共生的关系。SAMs多由一些病原微生物产生, 它们通过干扰植物鞘脂正常代谢, 引发植物死亡。尽管目前有关植物-病原真菌互作过程中鞘脂的作用及其机理的研究报道较多, 但鞘脂在植物-真菌互利共生关系中的作用研究还未见报道。植物与真菌的互利共生是自然界普遍存在的现象, 真菌能够提高宿主植物对水分和矿质营养元素的吸收, 通过影响宿主植物的物质代谢提高植物对资源的利用效率, 改善营养条件, 从而促进植物生长(郭顺星, 2016)。在互利共生关系中, 真菌的脂类代谢在植物和微生物的营养交换、共生信号转导及调控、共生关系的建立及维持等方面发挥着不可替代的作用(田磊等, 2016)。张海涵(2011)通过研究枸杞(Lycium chinense)根系深色有隔内生真菌(dark septate endophyte fungus, DSE)的定殖状况, 发现镜空菌丝(hyaline hyphae)和微菌核(microsclerotia)中存在大量的脂类物质, 推测丰富的脂类物质可能是DSE协助宿主植物度过干旱胁迫的能量物质。Maillet等(2011)发现菌根真菌可以产生脂质几丁寡糖(lipochitooligosac- charides), 它们作为共生信号刺激蒺藜苜蓿(M. truncatula)的根部生长, 并形成分枝状结构, 有利于菌根真菌的侵染。上述研究结果表明, 鞘脂在亲和性真菌与植物的共生关系中发挥重要作用。了解鞘脂在植物-真菌互作中的作用及其机制, 对于植物的病害防治以及利用有益真菌促进植物生长都具有重要意义。
(责任编辑: 朱亚娜)
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
URL [本文引用: 1]

在各种条件诱导的植物细胞编程性死亡 (PCD)过程中都有活性氧的参与 ,H2 O2 和O- ·2 可能是参与PCD调节的最重要的活性氧。文中概述了活性氧与植物细胞编程性死亡的关系以及活性氧的生成调节和与其它一些信号物质之间可能的相互作用。
DOI:10.3969/j.issn.1674-3466.2009.05.013URL [本文引用: 1]
众所周知,鞘脂是生物膜结构的重要组成成分,随着鞘脂在动物和酵母中的深入研究发现,鞘脂及其代谢产物是一类很重要的活性分子,它们参与调节细胞的生长、分化、衰老和细胞程序性死亡等许多重要的信号转导过程。鞘脂在植物中的研究最近几年才开始,植物鞘脂的功能还不十分清楚。最近的研究发现,鞘脂及其代谢产物在植物中也起着很重要的信号分子作用。该文详细总结了鞘脂在植物中的结构、代谢途径和主要生物学功能,并结合实验室的工作对植物鞘脂的功能研究进行了展望。
DOI:10.13343/j.cnki.wsxb.20150205URL [本文引用: 1]
丛枝菌根(AM)真菌是自然生态系统中分布最为广泛的真菌之一,在自然界物质循环和能量流动中发挥着重要作用。经过长期的协同进化,AM真菌和宿主植物之间形成了完美的互惠互利的共生关系,而真菌的脂类代谢可能是揭示共生秘密的关键所在。本文综述了AM真菌脂类代谢在共生关系建立和维持中关键作用的最新研究进展,重点探讨了AM真菌脂类代谢对共生信号调控的响应和反馈机制,主要包括:AM真菌脂类存储和释放对共生和非共生状态的响应,以及脂类代谢产物变化与共生营养传递之间的关系;脂类分解过程在共生建立初期对信号分子调控发生的响应,以及相应的物质转化和能量代谢;菌根共生互惠互利关系维持中,真菌脂类代谢与信号分子交流通道的相互渗透和影响。本文对于理解菌根共生机制,促进菌根在生产中的应用具有促进作用。
URL [本文引用: 1]
本研究采用巢式PCR-DGGE、克隆测序、实时定量PCR、BIOLOG和叶绿素荧光等技术研究了黄土高原旱区不同品种枸杞根系丛枝菌根真菌(AMF)与深色有隔内生真菌(DSE)定殖特征,不同品种枸杞根际土壤球囊霉素含量和AMF群落结构多样性季相变化特征,不同生态条件下枸杞根际土壤粒径分布与细菌和真菌群落多样性的关系,枸杞共生DSE菌株生物学特性和菌丝体对人工模拟干旱胁迫的响应,接种DSE对枸杞生长、根际细菌群落功能与遗传多样性的影响及干旱胁迫条件下接种AMF和DSE对枸杞叶片形态、光合参数和根系结构的影响。获得以下主要结果: 1.黄土高原枸杞丛枝菌根真菌(AMF)与深色有隔内生真菌(DSE)定殖特征 对柠杞1号(NQ-1)、柠杞5号(NQ-5)和柠杞菜1号(NQC-1)根系丛枝菌根真菌(AMF)与深色有隔内生真菌(DSE)定殖特征进行了调查研究,结果表明,枸杞形成P-型AM菌根。NQ-1丛枝菌根侵染率最高。不同品种枸杞的AMF侵染率不同,且呈现季节差异。同时,3种枸杞根系均存在DSE定殖。微菌核、深色(棕色)有隔菌丝主要分布在皮层细胞中。8月份DSE镜空菌丝侵染率最高,12月份最低。镜空菌丝和微菌核中存在大量脂类物质,这些脂类物质可能扮演着协助宿主渡过干旱逆境的碳源能量物质的角色。 2.枸杞根际球囊霉素含量及丛枝菌根真菌(AMF)群落结构 采用巢式PCR-DGGE和克隆测序技术研究了枸杞根际土壤AMF群落结构和球囊霉素含量,结果表明,不同品种枸杞根际土壤中易提取球囊霉素含量存在显著性差异,且呈现季节变化,8月份球囊霉素含量最高,12月份最低。总球囊霉素与易提取球囊霉素含量呈显著正相关。巢式PCR-DGGE图谱表明不同品种枸杞根际AMF群落不同,季节和品种对AMF群落丰富度指数(S)和多样性指数(H)影响显著,12月份丰富度指数显著低于8月份和10月份,但是两者交互作用不显著。冗余分析表明,枸杞根际土壤AMF群落多样性与土壤环境因子之间存在密切关系。克隆测序结果表明17条序列均属于球囊霉属(Glomus)。 3.不同生态条件下枸杞土壤粒径与微生物群落多样性 固原、同心、银川和中宁生态区枸杞根际土壤粒径最多分布在20~50μm范围之内,土壤粒径分别占总粒径的37.21±0.21%、33.25±0.21%、38.35±0.18%和42.33±0.45%。惠农地区有40.21±0.26%分布在100~200μm范围。巢式PCR-DGGE结果表明,5个生态区枸杞根际细菌群落丰富度指数为:同心中宁固原=银川惠农;真菌群落丰富度指数为:银川同心惠农中宁固原,细菌和真菌群落多样性指数之间相关性不显著。主成分分析表明,固原典型变量值的变异(离散)最小,而银川细菌典型变量值的变异(离散)最大,说明银川枸杞根际细菌群落最不稳定。固原与同心真菌群落多样性差异不显著。克隆测序结果表明枸杞根际细菌和真菌均为不可培养微生物类群。 4.枸杞DSE的分离、鉴定及其生物学特性 从枸杞根系上分离DSE菌株,对3株能与枸杞形成共生关系的菌株利用分子生物学技术(rDNA-ITS)进行鉴定。结果显示,3株DSE分别为Phoma chrysanthemicola,Cladosporium cladosporioides LBF3和C. cladosporioides LBF6。3种DSE对不同单一碳源和氮源物质利用有显著差异,P. chrysanthemicola最佳利用碳源和氮源为葡萄糖和甘氨酸,C. cladosporioides LBF3最佳利用碳源和氮源为甘露醇和脯氨酸,C. cladosporioidesLBF6最佳利用碳源和氮源为木糖和甘氨酸。模拟干旱胁迫结果发现P. chrysanthemicola的菌落形态、菌丝形态、菌丝蛋白质表达发生变化,傅里叶红外光谱分析表明模拟干旱引起菌丝中蛋白质结构发生改变。 5. DSE对枸杞根际细菌群落功能和遗传多样性的影响 为了探索接种DSE对枸杞根际微生态的影响,盆栽实验结果表明,接种3种DSE可以改变枸杞幼苗根际土壤细菌数量,接种DSE真菌60 d后枸杞根际土壤细菌代谢活性(BIOLOG)、代谢多样性均高于不接菌的对照。不同DSE菌株的作用有差异。主成分分析表明,接种深色有隔内生真菌后枸杞根际细菌群落对碳源的利用种类和程度均高于对照,细菌群落功能多样性发生了较大变化,接种DSE使得细菌群落变的更稳定;同时,巢式PCR-DGGE结果显示,DSE对细菌群落遗传多样性产生影响。 6. DSE对枸杞生长的影响 为了探索DSE与枸杞幼苗互作机制,将分离自枸杞根系的LBF-2菌株接种到枸杞幼苗根部,在实验室控制条件下研究了接种DSE对枸杞幼苗生长的影响,35天后检测结果发现深色有隔菌丝侵入根系皮层细胞,形成微菌核。首次采用巢式PCR-DGGE技术证明了DSE菌丝在植物组织中的定位和分布,结果认为DSE菌丝分布在枸杞根系、茎和叶片组织中。接种LBF-2可以提高枸杞幼苗生物量28%,接种幼苗叶绿素荧光参数显著高于不接种对照,叶绿素a和总叶绿素含量提高16%和19%。同时,接种植株黄酮物质含量比不接种高26%。结果表明LBF-2可能在枸杞育苗和栽培中发挥作用。 7.接种AMF和DSE枸杞苗叶片与根系对干旱胁迫的响应 为了探索接种AMF和DSE枸杞对干旱胁迫的响应机制,盆栽实验条件下,研究了接种AMF和DSE枸杞苗叶片与根系对干旱胁迫的响应。结果表明,接种AMF和DSE可以改变根系形态和叶片形态,显著降低叶片电解质透出率,提高叶片气孔导度和净光合速率。接种共生真菌能够有效地促进枸杞根系发育,通过提高枸杞叶片气孔导度和光合速率来减轻干旱胁迫的毒害作用。
DOI:10.1016/j.febslet.2006.02.061PMID:16527275
http://linkinghub.elsevier.com/retrieve/pii/S0014579306002638
DOI:10.1016/j.chembiol.2008.07.001URLPMID:18634999 [本文引用: 1]
In this issue of Chemistry & Biology, Kinjo et02al. (2008) propose that the addition of oligosaccharides to the02core outer membrane glycosphingolipid in Sphingomonas spp. may be an adaptation that allows bacteria to evade recognition by Natural Killer T cells, thus suggesting a remarkable process of host/pathogen coevolution.
DOI:10.3389/fmicb.2011.00239URLPMID:22164155 [本文引用: 2]
Of the ceramide monohexosides (CMHs), gluco- and galactosylceramides are the main neutral glycosphingolipids expressed in fungal cells. Their structural determination is greatly dependent on the use of mass spectrometric techniques, including fast atom bombardment-mass spectrometry (FAB-MS), electrospray ionization (ESI-MS), and energy collision-induced dissociation mass spectrometry (ESI-MS/CID-MS). Nuclear magnetic resonance (NMR) has also been used successfully. Such a combination of techniques, combined with classical analytical separation, such as HPTLC and column chromatography, has led to the structural elucidation of a great number of fungal CMHs. The structure of fungal CMH is conserved among fungal species and consists of a glucose or galactose residue attached to a ceramide moiety containing 9-methyl-4,8-sphingadienine with an amidic linkage to hydroxylated fatty acids, most commonly having 16 or 18 carbon atoms and unsaturation between C-3 and C-4. Along with their unique structural characteristics, fungal CMHs have a peculiar subcellular distribution and striking biological properties. Fungal cerebrosides were also characterized as antigenic molecules directly or indirectly involved in cell growth or differentiation in Schizophyllum commune, Cryptococcus neoformans, Pseudallescheria boydii, Candida albicans, Aspergillus nidulans, A.fumigatus and Colletotrichum gloeosporioides. Besides classical techniques for cerebroside (CMH) analysis, we now describe new approaches, combining conventional TLC and mass spectrometry, as well as emerging technologies for subcellular localization and distribution of glycosphingolipids by SIMS and imaging MALDI TOF .
DOI:10.1105/tpc.114.127050URLPMID:25149397 [本文引用: 2]
Arabidopsis thaliana plants that lack ceramide kinase, encoded by ACCELERATED CELL DEATH5 (ACD5), display spontaneous programmed cell death late in development and accumulate substrates of ACD5. Here, we compared ceramide accumulation kinetics, defense responses, ultrastructural features, and sites of reactive oxygen species (ROS) production in wild-type and acd5 plants during development and/or Botrytis cinerea infection. Quantitative sphingolipid profiling indicated that ceramide accumulation in acd5 paralleled the appearance of spontaneous cell death, and it was accompanied by autophagy and mitochondrial ROS accumulation. Plants lacking ACD5 differed significantly from the wild type in their responses to B. cinerea, showing earlier and higher increases in ceramides, greater disease, smaller cell wall appositions (papillae), reduced callose deposition and apoplastic ROS, and increased mitochondrial ROS. Together, these data show that ceramide kinase greatly affects sphingolipid metabolism and the site of ROS accumulation during development and infection, which likely explains the developmental and infection-related cell death phenotypes. The acd5 plants also showed an early defect in restricting B. cinerea germination and growth, which occurred prior to the onset of cell death. This early defect in B. cinerea restriction in acd5 points to a role for ceramide phosphate and/or the balance of ceramides in mediating early antifungal responses that are independent of cell death.
DOI:10.1016/j.quascirev.2008.01.010URLPMID:11858172 [本文引用: 1]
The sphinganine-analog mycotoxins (SAMs) fumonisin B-1 and AAL toxins are inhibitors of eukaryotic sphinganine N-acyltransferase in vitro. Treatment of eukaryotes with SAMs generally results in an accumulation of sphingoid base precursors and a depletion of complex sphingolipids. The asc,asc genotypes of tomato (Lycopersicon esculentum) and Nicotiana umbratica are sensitive to SAMs and host of the AAL toxin-producing fungus Alternaria alternata f. sp. lycopersici. Codominant insensitivity to SAMs in tomato is mediated by the Asc-1 gene, and sensitivity is associated with a frame-shift mutation present in asc-1. We investigated the function of Asc-1 in mediating insensitivity to SAMs and resistance to the fungus by overexpression of asc-1 and Asc-1. In this study, it is shown that overexpression of these genes did not lead to visual symptoms in tomato hairy roots and N. umbratica plants. Overexpression of asc-1 did not influence the (in)sensitivity to SAMs. Overexpression of Asc-1 in SAM-sensitive hairy roots and N. umbratica plants, however, mediated a high insensitivity to SAMs and resistance to plant infection by Alternaria alternata f. sp. lycopersici.
DOI:10.1111/j.1365-313X.2011.04829.xURLPMID:22023480 [本文引用: 1]
Plants contain a large diversity of sphingolipid structures, arising in part from C4 hydroxylation and Δ4 and Δ8 desaturation of the component long-chain bases (LCBs). Typically, 85–90% of sphingolipid LCBs in Arabidopsis leaves contain a cis or transΔ8 double bond produced by sphingoid LCB Δ8 desaturase (SLD). To understand the metabolic and physiological significance of Δ8 unsaturation, studies were performed using mutants of the Arabidopsis SLD genes AtSLD1 and AtSLD2. Our studies revealed that both genes are constitutively expressed, the corresponding polypeptides are ER-localized, and expression of these genes in Saccharomyces cerevisiae yields mixtures of cis/transΔ8 desaturation products, predominantly as trans isomers. Consistent in part with the higher expression of AtSLD1 in Arabidopsis plants, AtSLD1 T-DNA mutants showed large reductions in Δ8 unsaturated LCBs in all organs examined, whereas AtSLD2 mutants showed little change in LCB unsaturation. Double mutants of AtSLD1 and AtSLD2 showed no detectable LCB Δ8 unsaturation. Comprehensive analysis of sphingolipids in rosettes of these mutants revealed a 50% reduction in glucosylceramide levels and a corresponding increase in glycosylinositolphosphoceramides that were restored by complementation with a wild-type copy of AtSLD1. Double sld1 sld2 mutants lacked apparent growth phenotypes under optimal conditions, but displayed altered responses to certain stresses, including prolonged exposure to low temperatures. These results are consistent with a role for LCB Δ8 unsaturation in selective channeling of ceramides for the synthesis of complex sphingolipids and the physiological performance of Arabidopsis.
DOI:10.1007/s00425-009-1008-1URLPMID:19705146 [本文引用: 1]
Fusarium and Alternaria spp. are phytopathogenic fungi which are known to be virulent on broomrapes and to produce sphinganine-analog mycotoxins (SAMs). AAL-toxin is a SAM produced by Alternaria alternata which causes the inhibition of sphinganine N-acyltransferase, a key enzyme in sphingolipid biosynthesis, leading to accumulation of sphingoid bases. These long chain bases (LCBs) are determinant in the occurrence of programmed cell death (PCD) in susceptible plants. We showed that broomrapes are sensitive to AAL-toxin, which is not common plant behavior, and that AAL-toxin triggers cell death at the apex of the radicle as well as LCB accumulation and DNA laddering. We also demonstrated that three Lag1 homologs, encoding components of sphinganine N-acyltransferase in yeast, are present in the Orobanche cumana genome and two of them are mutated leading to an enhanced susceptibility to AAL-toxin. We therefore propose a model for the molecular mechanism governing broomrape susceptibility to the fungus Alternaria alternata.
DOI:10.1111/j.1744-7348.2003.tb00283.xURL [本文引用: 1]
Innate defence mechanisms in plants can be triggered and enhanced by certain agents, which are referred to as inducers. Inducing resistance against a broad spectrum of pathogens in otherwise susceptible plants is seen as a potentially safer alternative to other methods of chemical control of plant diseases. Cerebrosides, which are glycosphingolipids extracted from various plant pathogens, have been reported as resistance elicitors in the rice-pathogen system. In the present study, cerebroside elicited resistance against downy mildew disease (caused by Sclerospora graminicola) of pearl millet (Pennisetum glaucum) that was highly significant. The resistance was of systemic nature and the time required for the resistance to build up was from 2 days onwards. There was a significant yield enhancement due to disease suppression by cerebroside treatment. Promising results were obtained in a preliminary field trial.
DOI:10.1128/AAC.04667-14URLPMID:25824222 [本文引用: 1]
Abstract The cell wall synthesis-inhibiting echinocandins, including caspofungin and micafungin, play important roles in the treatment of candidiasis and aspergillosis. Previous studies revealed that, in the haploid yeast Candida glabrata, sphingolipid biosynthesis pathway mutations confer caspofungin reduced susceptibility (CRS) but micafungin increased susceptibility (MIS). Here we describe one Candida albicans strain (of 10 tested) that similarly yields CRS-MIS mutants at relatively high frequency. Mutants demonstrated increased levels of long chain bases (sphingolipid pathway intermediates) and, unique to this strain, loss of His104/Pro104 heterozygosity in the TSC13-encoded enoyl reductase. CRS-MIS was similarly observed in a C. albicans homozygous fen1螖 fen12螖 laboratory strain, and in diverse wild-type strains following exogenous long chain base treatment. Analogous to these results, CRS-MIS was demonstrated in an Aspergillus nidulans basA mutant encoding defective sphingolipid C4-hydroxylase, and in its wild-type parent exposed to long chain bases. Sphingolipids likely modulate echinocandin interaction with their Fks-membrane target in all susceptible fungi, with potential implications for optimizing therapy with existing antifungals and the development of novel agents. Copyright 漏 2015, American Society for Microbiology. All Rights Reserved.
DOI:10.1111/j.1432-1033.1994.00641.xURLPMID:7957179 [本文引用: 1]
In yeast, as in higher eukaryotic cells, sphingolipids are essential membrane components. The yeast, Saccharomyces cerevisiae , contains three classes of sphingolipids, inositolphosphorylceramide (Ins P Cer), mannosylinositolphosphorylceramide (ManIns P Cer) and mannosyldiinositolphosphorylceramide (Man P Ins 2 P Cer). As a prerequisite to localize these sphingolipids in subcellular membranes, authentic standards of the respective lipids were isolated and characterized using biochemical methods and electrospray ionization mass spectrometry. The complete set of yeast subcellular membranes was isolated at high purity, and sphingolipids were extracted. Ins P Cer, ManIns P Cer, and Man P Ins 2 P Cer were separated by thin-layer chromatography, stained and densitometrically scanned along with the respective standards. These methods enable a complete overview of the subcellular distribution of yeast sphingolipids to be obtained, as far as is known, for the first time. Ins P Cer was highly enriched in Golgi and vacuolar membranes, whereas the largest amounts of ManIns P Cer and ManPIns2PCer were found in the plasma membrane. The presence of inositol-containing sphingolipids in organelles of the protein-secretory pathway strongly supports the notion that protein secretion and intracellular trafficking of sphingolipids are linked processes.
DOI:10.1016/j.ceca.2009.11.011URLPMID:20022633
Studies performed in animals have highlighted the major role of sphingolipids in regulating the balance between cell proliferation and cell death. Sphingolipids have also been shown to induce cell death in plants calcium-based signalling pathways but the contribution of free cytosolic and/or nuclear calcium in the overall process has never been evaluated.Here, we show that increase in tobacco BY-2 cells of the endogenous content of Long Chain Bases (LCBs) caused by external application of --2-amino-5-phosphopentanoic acid (AP5) and [(+)-dizocilpine] (MK801), two inhibitors of animal and plant ionotropic glutamate receptors, suppress DHS-induced cell death symptoms by selectively inhibiting the variations of nuclear calcium concentration. The selective action of these compounds demonstrates the crucial role of nuclear calcium signature in controlling DHS-induced cell death in tobacco cells.
DOI:10.1038/srep41868URLPMID:28145511 [本文引用: 1]
The sustainable production of fuels and chemicals using microbial cell factories is now well established. However, many microbial production processes are still limited in scale due to inhibition from compounds that are present in the feedstock or are produced during fermentation. Some of these inhibitors interfere with cellular membranes and change the physicochemical properties of the membranes. Another group of molecules is dependent on their permeation rate through the membrane for their inhibition. We have investigated the use of membrane engineering to counteract the negative effects of inhibitors on the microorganism with focus on modulating the abundance of complex sphingolipids in the cell membrane ofSaccharomyces cerevisiae. Overexpression ofELO3, involved in fatty acid elongation, andAUR1, which catalyses the formation of complex sphingolipids, had no effect on the membrane lipid profile or on cellular physiology. Deletion of the genesORM1andORM2, encoding negative regulators of sphingolipid biosynthesis, decreased cell viability and considerably reduced phosphatidylinositol and complex sphingolipids. Additionally, combiningELO3andAUR1overexpression withorm1/2螖improved cell viability and increased fatty acyl chain length compared with onlyorm1/2螖. These findings can be used to further study the sphingolipid metabolism, as well as giving guidance in membrane engineering.
DOI:10.1104/pp.15.01126URLPMID:26378098 [本文引用: 2]
Sphingolipids are emerging as second messengers in programmed cell death and plant defense mechanisms. However, their role in plant defense is far from being understood, especially against necrotrophic pathogens. Sphingolipidomics and plant defense responses during pathogenic infection were evaluated in the mutant of long-chain base phosphate (LCB-P) lyase, encoded by the dihydrosphingosine-1-phosphate lyase1 (AtDPL1) gene and regulating long-chain base/LCB-P homeostasis. Atdpl1 mutants exhibit tolerance to the necrotrophic fungus Botrytis cinerea but susceptibility to the hemibiotrophic bacterium Pseudomonas syringae pv tomato (Pst). Here, a direct comparison of sphingolipid profiles in Arabidopsis (Arabidopsis thaliana) during infection with pathogens differing in lifestyles is described. In contrast to long-chain bases (dihydrosphingosine [d18:0] and 4,8-sphingadienine [d18:2]), hydroxyceramide and LCB-P (phytosphingosine-1-phosphate [t18:0-P] and 4-hydroxy-8-sphingenine-1-phosphate [t18:1-P]) levels are higher in Atdpl1-1 than in wild-type plants in response to B. cinerea. Following Pst infection, t18:0-P accumulates more strongly in Atdpl1-1 than in wild-type plants. Moreover, d18:0 and t18:0-P appear as key players in Pst- and B. cinerea-induced cell death and reactive oxygen species accumulation. Salicylic acid levels are similar in both types of plants, independent of the pathogen. In addition, salicylic acid-dependent gene expression is similar in both types of B. cinerea-infected plants but is repressed in Atdpl1-1 after treatment with Pst. Infection with both pathogens triggers higher jasmonic acid, jasmonoyl-isoleucine accumulation, and jasmonic acid-dependent gene expression in Atdpl1-1 mutants. Our results demonstrate that sphingolipids play an important role in plant defense, especially toward necrotrophic pathogens, and highlight a novel connection between the jasmonate signaling pathway, cell death, and sphingolipids.
DOI:10.1038/nature09622URLPMID:21209659 [本文引用: 1]
Abstract Arbuscular mycorrhiza (AM) is a root endosymbiosis between plants and glomeromycete fungi. It is the most widespread terrestrial plant symbiosis, improving plant uptake of water and mineral nutrients. Yet, despite its crucial role in land ecosystems, molecular mechanisms leading to its formation are just beginning to be unravelled. Recent evidence suggests that AM fungi produce diffusible symbiotic signals. Here we show that Glomus intraradices secretes symbiotic signals that are a mixture of sulphated and non-sulphated simple lipochitooligosaccharides (LCOs), which stimulate formation of AM in plant species of diverse families (Fabaceae, Asteraceae and Umbelliferae). In the legume Medicago truncatula these signals stimulate root growth and branching by the symbiotic DMI signalling pathway. These findings provide a better understanding of the evolution of signalling mechanisms involved in plant root endosymbioses and will greatly facilitate their molecular dissection. They also open the way to using these natural and very active molecules in agriculture.
DOI:10.1016/j.pbi.2013.02.009URLPMID:23499054 [本文引用: 1]
Plant sphingolipids are structurally diverse molecules that are important as membrane components and bioactive molecules. An appreciation of the relationship between structural diversity and functional significance of plant sphingolipids is emerging through characterization of Arabidopsis mutants coupled with advanced analytical methods. It is increasingly apparent that modifications such as hydroxylation and desaturation of the sphingolipid nonpolar long-chain bases and fatty acids influence their metabolic routing to particular complex sphingolipid classes and their functions in signaling pathways and other cellular processes, such as membrane protein targeting. Here, we review recent reports investigating some of the more prevalent sphingolipid structural modifications and their functional importance in plants.
DOI:10.1016/j.bbalip.2016.04.003URLPMID:27086144 [本文引用: 7]
Sphingolipids and their phosphorylated derivatives are ubiquitous bio-active components of cells. They are structural elements in the lipid bilayer and contribute to the dynamic nature of the membrane. They have been implicated in many cellular processes in yeast and animal cells, including aspects of signaling, apoptosis, and senescence. Although sphingolipids have a better defined role in animal systems, they have been shown to be central to many essential processes in plants including but not limited to, pollen development, signal transduction and in the response to biotic and abiotic stress. A fuller understanding of the roles of sphingolipids within plants has been facilitated by classical biochemical studies and the identification of mutants of model species. Recently the development of powerful mass spectrometry techniques hailed the advent of the emerging field of lipidomics enabling more accurate sphingolipid detection and quantitation. This review will consider plant sphingolipid biosynthesis and function in the context of these new developments. This article is part of a Special Issue entitled: Plant Lipid Biology edited by Kent D. Chapman and Ivo Feussner. 61Up-to-date review of our current understanding on plant sphingolipid biosynthesis61In depth consideration of plant-specific aspects of sphingolipid metabolism61Updates on the biosynthesis of GIPCs in plants61The sometimes overlooked role of VLCFA biosynthesis in sphingolipid biogenesis Up-to-date review of our current understanding on plant sphingolipid biosynthesis In depth consideration of plant-specific aspects of sphingolipid metabolism Updates on the biosynthesis of GIPCs in plants The sometimes overlooked role of VLCFA biosynthesis in sphingolipid biogenesis
DOI:10.1105/tpc.113.111500URLPMID:23695979 [本文引用: 1]
The Arabidopsis thaliana protein GOLGI-LOCALIZED NUCLEOTIDE SUGAR TRANSPORTER (GONST1) has been previously identified as a GDP-D-mannose transporter. It has been hypothesized that GONST1 provides precursors for the synthesis of cell wall polysaccharides, such as glucomannan. Here, we show that in vitro GONST1 can transport all four plant GDP-sugars. However, gonst1 mutants have no reduction in glucomannan quantity and show no detectable alterations in other cell wall polysaccharides. By contrast, we show that a class of glycosylated sphingolipids (glycosylinositol phosphoceramides [GIPCs]) contains Man and that this mannosylation is affected in gonst1. GONST1 therefore is a Golgi GDP-sugar transporter that specifically supplies GDP-Man to the Golgi lumen for GIPC synthesis. gonst1 plants have a dwarfed phenotype and a constitutive hypersensitive response with elevated salicylic acid levels. This suggests an unexpected role for GIPC sugar decorations in sphingolipid function and plant defense signaling. Additionally, we discuss these data in the context of substrate channeling within the Golgi.
DOI:10.1007/s00294-013-0416-8URLPMID:242532931 [本文引用: 1]
An antifungal defensin, AFP1, of Brassica juncea inhibits the growth of various microorganisms. The molecular details of this inhibition remain largely unknown. Herein, we reveal that a specific structure of fungal sphingolipid glucosylceramide (GlcCer) is critical for the sensitivity of Candida albicans cells to AFP1. Our results revealed that AFP1 induces plasma membrane permeabilization and the production of reactive oxygen species (ROS) in wild-type C. albicans cells, but not in cells lacking the ninth methyl residue of the GlcCer sphingoid base moiety, which is a characteristic feature of fungi. AFP1-induced ROS production is responsible for its antifungal activity, with a consequent loss of yeast cell viability. These findings suggest that AFP1 specifically recognizes the structural difference of GlcCer for targeting of the fungal pathogens.
DOI:10.1006/anae.2001.0376URL [本文引用: 1]
Sphingolipids constitute a distinctive group of membrane lipids characterized by a long-chain (monounsaturated), di-hydroxy amine structure (sphingosine). Sphingolipids are essential components of the plasma membrane of mammalian cells where they are typically found in the outer leaflet. They are also membrane constituents of some bacterial groups, particularly anaerobes. These groups include Bacteroides, Prevotella, Porphyromonas, Fusobacterium, Sphingomonas, Sphingobacterium, Bdellovibrio, Cystobacter, Mycoplasma, Flectobacillus, and possibly Acetobacter. Fungi in which sphingolipids have been found comprise Saccharomyces, Candida, Histoplasma, Phytophthora, Cryptococcus, Aspergillus, Neurospora, Schizosaccharomyces, Fusicoccum, Shizophyllum, Amanita, Hansenula, Lactarius, Lentinus, Penicillium, Clitocybe, Paracoccidioides, Agaricus, Sporothrix, and oomycete plant pathogens. The fact that sphingolipids are not universally present in bacteria and fungi makes them interesting as taxonomic markers. Thus in Sphingomonas spp. distinction by sphingolipid and fatty acid patterns is in agreement with phylogenetic clustering by 16 S rRNA gene sequences.
DOI:10.1016/j.febslet.2010.08.027URLPMID:20732322 [本文引用: 1]
Sphingolipids are important membrane components and also regulate cell proliferation and apoptosis. We detected a fast increase of the free sphingobase t18:0 (phytosphinganine) in Arabidopsis leaves after inoculation with an avirulent strain of the bacterial pathogen Pseudomonas syringae pathovar tomato, characterized by host cell death reactions. The induction of phytosphinganine was more transient in virulent interactions lacking cell death reactions, suggesting a positive role of t18:0 in the plants鈥 response to pathogens, e.g. the hypersensitive response. In the mutant sphingobase hydroxylase 1 ( sbh1-1), Pseudomonas induced elevated free d18:0 levels. As total t18:0 contents (after hydrolysis of ceramides) were not reduced in sbh1-1, the pathogen-triggered t18:0 increase most likely results from de novo synthesis from d18:0 which would require SBH1.
DOI:10.1111/j.1365-2958.2007.05955.xURLPMID:17908205 [本文引用: 2]
Antifungal defensins, MsDef1 and MtDef4, from Medicago spp., inhibit the growth of a fungal pathogen, Fusarium graminearum , at micromolar concentrations. However, molecular mechanisms by which they inhibit the growth of this fungus are not known. We have characterized a functional role of the fungal sphingolipid glucosylceramide in regulating sensitivity of the fungus to MsDef1 and MtDef4. A null mutation of the Fg GCS1 gene encoding glucosylceramide synthase results in a mutant lacking glucosylceramide. The 螖Fg gcs1 -null mutant becomes resistant to MsDef1, but not to MtDef4. It shows a significant change in the conidial morphology and displays dramatic polar growth defect, and its mycelia are resistant to cell wall degrading enzymes. Contrary to its essential role in the pathogenicity of a human fungal pathogen, Cryptococcus neoformans , GCS1 is not required for the pathogenicity of F. graminearum . The 螖Fg gcs1 mutant successfully colonizes wheat heads and corn silk, but its ability to spread in these tissues is significantly reduced as compared with the wild-type PH-1 strain. In contrast, it retains full virulence on tomato fruits and Arabidopsis thaliana floral and foliar tissues. Based on our findings, we conclude that glucosylceramide is essential for MsDef1-mediated growth inhibition of F. graminearum , but its role in fungal pathogenesis is host-dependent.
DOI:10.1016/S0079-6123(05)52014-4URLPMID:19028992 [本文引用: 2]
The C-9-methylated glucosylceramides (GlcCers) are sphingolipids unique to fungi. They play important roles in fungal growth and pathogenesis, and they act as receptors for some antifungal plant defensins. We have identified two genes, FgMT1 and FgMT2, that each encode a putative sphingolipid C-9 methyltransferase (C-9-MT) in the fungal pathogen Fusarium graminearum and complement a Pichia pastoris C-9-MT-null mutant. The DeltaFgmt1 mutant produced C-9-methylated GlcCer like the wild-type strain, PH-1, whereas the DeltaFgmt2 mutant produced 65 to 75% nonmethylated and 25 to 35% methylated GlcCer. No DeltaFgmt1DeltaFgmt2 double-knockout mutant producing only nonmethylated GlcCer could be recovered, suggesting that perhaps C-9-MTs are essential in this pathogen. This is in contrast to the nonessential nature of this enzyme in the unicellular fungus P. pastoris. The DeltaFgmt2 mutant exhibited severe growth defects and produced abnormal conidia, while the DeltaFgmt1 mutant grew like the wild-type strain, PH-1, under the conditions tested. The DeltaFgmt2 mutant also exhibited drastically reduced disease symptoms in wheat and much-delayed disease symptoms in Arabidopsis thaliana. Surprisingly, the DeltaFgmt2 mutant was less virulent on different host plants tested than the previously characterized DeltaFggcs1 mutant, which lacks GlcCer synthase activity and produces no GlcCer at all. Moreover, the DeltaFgmt1 and DeltaFgmt2 mutants, as well as the P. pastoris strain in which the C-9-MT gene was deleted, retained sensitivity to the antifungal plant defensins MsDef1 and RsAFP2, indicating that the C-9 methyl group is not a critical structural feature of the GlcCer receptor required for the antifungal action of plant defensins.
DOI:10.1172/JCI27890URLPMID:1466548 [本文引用: 1]
The pathogenic fungus Cryptococcus neoformans infects humans upon inhalation and causes the most common fungal meningoencephalitis in immunocompromised subjects worldwide. In the host, C. neoformans is found both intracellularly and extracellularly, but how these two components contribute to the development of the disease is largely unknown. Here we show that the glycosphingolipid glucosylceramide (GlcCer), which is present in C. neoformans, was essential for fungal growth in host extracellular environments, such as in alveolar spaces and in the bloodstream, which are characterized by a neutral/alkaline pH, but not in the host intracellular environment, such as in the phagolysosome of macrophages, which is characteristically acidic. Indeed, a C. neoformans mutant strain lacking GlcCer did not grow in vitro at a neutral/alkaline pH, yet it had no growth defect at an acidic pH. The mechanism by which GlcCer regulates alkali tolerance was by allowing the transition of C. neoformans through the cell cycle. This study establishes C. neoformans GlcCer as a key virulence factor of cryptococcal pathogenicity, with important implications for future development of new antifungal strategies.
DOI:10.4155/fmc-2016-0053URLPMID:5558548 [本文引用: 3]
Invasive fungal infections have significantly increased in the last few decades. Three classes of drugs are commonly used to treat these infections: polyenes, azoles and echinocandins. Unfortunately each of these drugs has drawbacks; polyenes are toxic, resistance against azoles is emerging and echinocandins have narrow spectrum of activity. Thus, the development of new antifungals is urgently needed. In this context, fungal sphingolipids have emerged as a potential target for new antifungals, because their biosynthesis in fungi is structurally different than in mammals. Besides, some fungal sphingolipids play an important role in the regulation of virulence in a variety of fungi. This review aims to highlight the diverse strategies that could be used to block the synthesis or/and function of fungal sphingolipids.
DOI:10.4161/psb.6.10.16981URLPMID:21534970 [本文引用: 1]
Long chain bases or sphingoid bases are building blocks of complex sphingolipids that display a signaling role in programmed cell death in plants. So far, the type of programmed cell death in which these signaling lipids have been demonstrated to participate is the cell death that occurs in plant immunity, known as the hypersensitive response. The few links that have been described in this pathway are: MPK6 activation, increased calcium concentrations, and reactive oxygen species (ROS) generation. The latter constitute one of the more elusive loops because of the chemical nature of ROS the multiple possible cell sites where they can be formed and the ways in which they influence cell structure and function.
DOI:10.3389/fimmu.2017.00378URLPMID:5372786 [本文引用: 1]
Sphingolipids are the major constituent of the mucus secreted by the cells of epithelial linings of lungs where they maintain the barrier functions and prevent microbial invasion. Sphingolipids are interconvertible, and their primary and secondary metabolites have both structural and functional roles. Out of several sphingolipid metabolites, sphingosine-1 phosphate (S1P) and ceramide are central molecules and decisive for sphingolipid signaling. These are produced by enzymatic activity of sphingosine kinase-1 (SK-1) upon the challenge with either biological or physiological stresses. S1P and ceramide rheostat are important for the progression of various pathologies, which are manifested by inflammatory cascade. S1P is a well-established secondary messenger and associated with various neuronal, metabolic, and inflammatory diseases other than respiratory infections such asChlamydia pneumoniae, Streptococcus pneumoniae, andMycobacterium tuberculosis. These pathogens are known to exploit sphingolipid metabolism for their opportunistic survival. Decreased sphingosine kinase activity/S1P content in the lung and peripheral blood of tuberculosis patients clearly indicated a dysregulation of sphingolipid metabolism during infection and suggest that sphingolipid metabolism is important for management of infection by the host. Our previous study has demonstrated that gain of SK-1 activity is important for the maturation of phagolysosomal compartment, innate activation of macrophages, and subsequent control of mycobacterial replication/growth in macrophages. Furthermore, S1P-mediated amelioration of lung pathology and disease severity in TB patients is believed to be mediated by the selective activation or rearrangement of various S1P receptors (S1PR) particularly S1PR2, which has been effective in controlling respiratory fungal pathogens. Therefore, such specificity of S1P鈥揝1PR would be paramount for triggering inflammatory events, subsequent activation, and fostering bactericidal potential in macrophages for the control of TB. In this review, we have discussed and emphasized that sphingolipids may represent effective novel, yet dual specific drug targets for controlling pulmonary infections.
DOI:10.4161/psb.4.6.8583URLPMID:18059378 [本文引用: 2]
It has long been known that fungal pathogens like Fusarium and Alternaria spp. produce toxins (mycotoxin) to kill plant cells.02 These mycotoxins have been shown to perturb the plant sphingolipid biosynthesis pathway, resulting in the necrotic cell death of plant cells.02 A recent study by Shi et al.1 revealed that an increase in the amount of cellular sphingoid bases triggers plant programmed cell death (PCD) through accumulation of reactive oxygen species (ROS).02 These studies point to the importance of sphingolipids in the regulation of plant cell in disease development as well as in defense responses.02 In the latest report,2 we showed that serine palmitoyltransferase (SPT), the key enzyme of sphingolipid biosynthesis, regulates not only plant cell death but also defense response against a non-host pathogen, soliciting further studies to elucidate the roles of sphingolipids in plant innate immunity.
DOI:10.1016/j.bbalip.2016.02.021URLPMID:26928590 [本文引用: 1]
61Plants can undergo interactions with pathogens, symbionts, and beneficial microbes.61Membrane lipids represent the natural barrier between the host and the microbial cell.61Lipids and lipid breakdown products serve as signals during plant–microbe interactions.
DOI:10.1046/j.1365-313X.2002.01444.xURLPMID:12445127 [本文引用: 1]
The nectrotrophic fungus A lternaria a lternata f.sp. l ycopersici infects tomato plants of the genotype asc / asc by utilizing a host-selective toxin, AAL-toxin, that kills the host cells by inducing programmed cell death. Asc-1 is homologous to genes found in most eukaryotes from yeast to humans, suggesting a conserved function. A yeast strain with deletions in the homologous genes LAG1 and LAC1 was functionally complemented by Asc-1 , indicating that Asc-1 functions in an analogous manner to the yeast homologues. Examination of the yeast sphingolipids, which are almost absent in the lag1螖lac1螖 mutant, showed that Asc-1 was able to restore the synthesis of sphingolipids. We therefore examined the biosynthesis of sphingolipids in tomato by labeling leaf discs with l -[3- 3 H]serine. In the absence of AAL-toxin, there was no detectable difference in sphingolipid labeling between leaf discs from Asc/Asc or asc / asc leaves. In the presence of pathologically significant concentrations of AAL-toxin however, asc / asc leaf discs showed severely reduced labeling of sphingolipids and increased label in dihydrosphingosine (DHS) and 3-ketodihydrosphingosine (3-KDHS). Leaf discs from Asc / Asc leaves responded to AAL-toxin treatment by incorporating label into different sphingolipid species. The effects of AAL-toxin on asc / asc leaflets could be partially blocked by the simultaneous application of AAL-toxin and myriocin. Leaf discs simultaneously treated with AAL-toxin and myriocin showed no incorporation of label into sphingolipids or long-chain bases as expected. These results indicate that the presence of Asc-1 is able to relieve an AAL-toxin-induced block on sphingolipid synthesis that would otherwise lead to programmed cell death.
DOI:10.1007/s10529-014-1631-1URLPMID:25129050 [本文引用: 1]
Glucosylceramide and galactosylceramide were detected in three Aspergillus species: Aspergillus oryzae, Aspergillus sojae and Aspergillus. awamori, using borate-coated TLC. The cerebrosides from A. oryzae were further purified by ion exchange and iatrobeads column chromatographies with or without borate, and determined the composition of sugar, fatty acid and sphingoid base by GC/MS, MALDI-TOF/MS and (1)H-NMR. We identified them as β-glucosylceramide and β-galactosylceramide. The ceramide moiety of both cerebrosides consisted mainly of 2-hydroxystearic acid and either 9-methyl-octadeca-4, 8-sphingadienine or octadeca-4, 8-sphingadienine. To our knowledge, this is the first study to provide evidence for the presence of β-galactosylceramide in A. oryzae.
DOI:10.1111/tpj.13382URLPMID:27643972 [本文引用: 1]
漏 2016 The Authors The Plant Journal 漏 2016 John Wiley & Sons LtdSphingolipids are a major component of plant plasma membranes and endomembranes, and mediate a diverse range of biological processes. Study of the highly glycosylated glycosyl inositol phosphorylceramide (GIPC) sphingolipids has been slow as a result of challenges associated with the extractability of GIPCs, and their functions in the plant remain poorly characterized. We recently discovered an Arabidopsis GIPC glucuronosyltransferase, INOSITOL PHOSPHORYLCERAMIDE GLUCURONOSYLTRANSFERASE聽1 (IPUT1), which is the first enzyme in the GIPC glycosylation pathway. Plants homozygous for the iput1 loss-of-function mutation were unobtainable, and so the developmental effects of reduced GIPC glucuronosylation could not be analyzed in聽planta. Using a pollen-specific rescue construct, we have here isolated homozygous iput1 mutants. The iput1 mutants show severe dwarfism, compromised pollen tube guidance, and constitutive activation of salicyclic acid-mediated defense pathways. The mutants also possess reduced GIPCs, increased ceramides, and an increased incorporation of short-chain fatty acids and dihydroxylated bases into inositol phosphorylceramides and GIPCs. The assignment of a direct role for GIPC glycan head groups in the impaired processes in iput1 mutants is complicated by the vast compensatory changes in the sphingolipidome; however, our results reveal that the glycosylation steps of GIPC biosynthesis are important regulated components of sphingolipid metabolism. This study corroborates previously suggested roles for GIPC glycans in plant growth and defense, suggests important roles for them in reproduction and demonstrates that the entire sphingolipidome is sensitive to their status.
DOI:10.1016/S0378-1097(03)00590-1URLPMID:13129623 [本文引用: 1]
DmAMP1, an antifungal plant defensin from Dahlia merckii , was shown previously to require the presence of sphingolipids for fungicidal action against Saccharomyces cerevisiae . Sphingolipids may stabilize glycosylphosphatidylinositol (GPI)-anchored proteins, which interact with DmAMP1, or they may directly serve as DmAMP1 binding sites. In the present study, we demonstrate that S. cerevisiae disruptants in GPI-anchored proteins showed small or no increased resistance towards DmAMP1 indicating no involvement of these proteins in DmAMP1 action. Further, studies using an enzyme-linked immunosorbent assay (ELISA)-based binding assay revealed that DmAMP1 interacts directly with sphingolipids isolated from S. cerevisiae and that this interaction is enhanced in the presence of equimolar concentrations of ergosterol. Therefore, DmAMP1 antifungal action involving membrane interaction with sphingolipids and ergosterol is proposed.
DOI:10.1074/jbc.M311165200URLPMID:14604982 [本文引用: 1]
Growth of the yeast species Candida albicans and Pichia pastoris is inhibited by RsAFP2, a plant defensin isolated from radish seed (Raphanus sativus), at micromolar concentrations. In contrast, gcs-deletion mutants of both yeast species are resistant toward RsAFP2. GCS genes encode UDP-glucose:ceramide glucosyltransferases, which catalyze the final step in the biosynthesis of the membrane lipid glucosylceramide. In an enzyme-linked immunosorbent assay-based binding assay, RsAFP2 was found to interact with glucosylceramides isolated from P. pastoris but not with soybean nor human glucosylceramides. Furthermore, the P. pastoris parental strain is sensitive toward RsAFP2-induced membrane permeabilization, whereas the corresponding gcs-deletion mutant is highly resistant to RsAFP2-mediated membrane permeabilization. A model for the mode of action of RsAFP2 is presented in which all of these findings are linked. Similarly to RsAFP2, heliomicin, a defensin-like peptide from the insect Heliothis virescens, is active on C. albicans and P. pastoris parental strains but displays no activity on the gcs-deletion mutants of both yeast species. Furthermore, heliomicin interacts with glucosylceramides isolated from P. pastoris and soybean but not with human glucosylceramides. These data indicate that structurally homologous anti-fungal peptides present in species from different eukaryotic kingdoms interact with the same target in the fungal plasma membrane, namely glucosylceramides, and as such support the hypothesis that defensins from plants and insects have evolved from a single precursor.
DOI:10.1515/BC.2005.020URLPMID:15843160
While the role of C2-ceramide in the induction of programmed cell death (PCD) in animal systems has been well documented, little is known of its role in plant cells. Here we show that C2-ceramide induces PCD in Arabidopsis suspension cultures, which is preceded by the generation of a calcium transient and an increase in reactive oxygen species (ROS). Inhibition of the calcium transient prevented cell death, whereas inhibition of ROS had no effect on cell survival. These observations suggest that calcium signalling plays a role in ceramide-induced PCD but is independent of the generation of ROS.
DOI:10.2174/0929867324666170112114525URLPMID:28078995 [本文引用: 1]
Sphingolipids (SLs) regulate apoptosis, proliferation, and stress response. SLs, includingceramide, glycosphingolipids(glucosylceramide, lactosylceramide, and gangliosides)and sphingosine-1-phosphate (S1P), play a role in the pathogenesis and progression of genetic(lysosomal storage disease, congenital nephrotic syndrome and polycystic kidney disease) andnon-genetic forms of chronic kidney diseases (CKDs). SLs metabolism defects promote complications(cardiovascular events, etc.) via oxidant stress in CKDs. A balancing role of apoptoticSLs and anti-apoptotic S1P is crucial in the regulation of glomerular injury and complicationsassociated with CKDs. Interaction between SLs, endothelial function and reninangiotensin-aldosterone system (RAAS) plays an important role in the regulation of glomerularinjury. SLs affect mitochondrial function that regulate the opening of mitochondrial permeabilitytransition (MPT) pore, mitochondrial outer membrane permeability (MOMP), generationof reactive oxygen species (ROS), and expression of BcL-2 family proteins, whichresult in cytochrome c release and caspase activation, leading to apoptosis, and regulateglomerular cell proliferation or renal fibrosis. This review article summarizes the current evidencesupporting a role of SLs metabolism defects in the pathogenesis and progression ofglomerular injury and discusses a role of mitochondria, including MPT pore, MOMP, ROSgeneration, BcL-2 family proteins, interaction between SLs, endothelial function and RAAS,and SLs-induced downstream signaling events in CKDs. Crosstalk between these factorsplays a role in the pathogenesis and progression of CKDs. Therapeutic strategy of targetingSLs metabolism defects for CKDs through modulation of the enzymes responsible for SLsmetabolism defects is also discussed.
DOI:10.1093/pcp/pcf090URLPMID:12154140 [本文引用: 1]
Abstract Cerebrosides, compounds categorized as glycosphingolipids, were found to occur in a wide range of phytopathogens as novel elicitors and to induce the effective disease resistance for rice plants in our previous study. Here, we showed that cerebroside elicitors lead to the accumulation of phytoalexins and pathogenesis-related (PR) protein in cell suspension cultures of rice with the structural specificity similar to that for the rice whole plants. This elicitor activity of the cerebroside was greater than jasmonic acid (JA) and chitin oligomer (which is known to be an elicitor for cell suspension cultures of rice). Treatment of cell suspension cultures with cerebroside and chitin oligomer resulted in a synergetic induction of phytoalexins, suggesting that cerebroside and carbohydrate elicitors, such as glucan and chitin elicitor, enhance the defense signals of rice in vivo. Induction of phytoalexins by the treatment with cerebroside elicitor was markedly inhibited by LaCl(3) and GdCl(3), Ca(2+ )channel blockers. It is possible that Ca(2+) may be involved in the signaling pathway of elicitor activity of cerebroside.
DOI:10.1093/pcp/41.6.676URLPMID:10945336
Abstract Cerebrosides A and C, compounds categorized as glycosphingolipids, were isolated in our previous study from the rice blast fungus (Magnaporthe grisea) as novel elicitors which induce the synthesis of rice phytoalexins. In this paper, these cerebroside elicitors showed phytoalexin-inducing activity when applied to plants by spray treatment and also induced the expression of pathogenesis-related (PR) proteins in rice leaves. This elicitor activity of the cerebrosides showed the structural specificity as that for the induction of phytoalexins. Ceramides prepared from the cerebrosides by removal of glucose also showed the elicitor activity even in lower level compared to the cerebrosides. In field experiments, the cerebroside elicitors effectively protected rice plants against the rice blast fungus, an economically devastating agent of disease of rice in Japan. The cerebrosides elicitors protected rice plants from other disease as well and were found to occur in a wide range of different phytopathogens, indicating that cerebrosides function as general elicitors in a wide variety of rice-pathogen interactions.
[本文引用: 1]
DOI:10.1105/tpc.108.060053URL [本文引用: 1]
The Arabidopsis thaliana resistance gene RPW8 triggers the hypersensitive response (HR) to restrict powdery mildew infection via the salicylic acid-dependent signalling pathway. To further understand how RPW8 signaling is regulated, we have conducted a genetic screen to identify mutations enhancing RPW8-mediated HR-like cell death (designated erh). Here, we report the isolation and characterization of the Arabidopsis erh1 mutant, in which the At2g37940 locus is knocked out by a T-DNA insertion. Loss of function of ERH1 results in salicylic acid accumulation, enhanced transcription of RPW8 and RPW8-dependent spontaneous HR-like cell death in leaf tissues, and reduction in plant stature. Sequence analysis suggests that ERH1 may encode the long-sought Arabidopsis functional homolog of yeast and protozoan inositolphosphorylceramide synthase (IPCS), which converts ceramide to inositolphosphorylceramide. Indeed, ERH1 is able to rescue the yeast aur1 mutant, which lacks the IPCS, and the erh1 mutant plants display reduced (53% of wild type) levels of leaf IPCS activity, indicating that ERH1 encodes a plant IPCS. Consistent with its biochemical function, the erh1 mutation causes ceramide accumulation in plants expressing RPW8. These data reinforce the concept that sphingolipid metabolism (specifically, ceramide accumulation) plays an important role in modulating plant programmed cell death associated with defense.
DOI:10.1007/s00018-003-2243-4URLPMID:12827281 [本文引用: 4]
Glycosphingolipids are ubiquitous membrane lipids of eukaryotic organisms and a few bacteria. Whereas inositol-containing glycosphingolipids are restricted to plants and fungi, galactosylceramide occurs only in fungi and animals. In contrast, glucosylceramide is the unique glycosphingolipid which plants, fungi and animals have in common. However, there are specific differences in the structure of the ceramide backbone of glucosylceramides from these organisms. A comparison of the structural features and the biosynthesis of glucosylceramides from plants, fungi and animals will contribute to our understanding of their functions, which so far have been analysed mainly in animals. The availability of nearly all genes involved in the biosynthesis of glucosylceramides enables the specific manipulation of glycosphingolipid metabolism by techniques of forward and reverse genetics. Application of this approach to unicellular organisms like yeasts, multicellular filamentous fungi, as well as to complex organisms like plants will reveal common and different glucosylceramide functions in these organisms. These glycolipids play a role both in intracellular processes and in cell-to-cell interactions. These interactions may occur between cells of a multicellular organism or between cells of different species, as in host-pathogen interactions.
DOI:10.1007/s10265-017-0923-7URLPMID:28303405 [本文引用: 1]
Fumonisin B 1 (FB 1 ), an inducer of cell death, disrupts sphingolipid metabolism; large accumulations of de novo synthesized free long-chain bases (LCBs) are observed. However, it remains unclear whether tolerance to FB 1 toxicity in plants is connected with preventing the accumulation of free LCBs through their phosphorylation. Here a workflow for the extraction, detection and quantification of LCB phosphates (LCBPs) in Arabidopsis thaliana was developed. We studied the effect of expression of genes for three enzymes involved in the synthesis and degradation of LCBPs, LCB kinase (LCBK1), LCBP phosphatase (SPP1) and lyase (DPL1) on FB 1 -induced cell death. As expected, large accumulations of saturated free LCBs, dihydrosphingosine and phytosphingosine, were observed in the FB 1 -treated leaves. On the other hand, a high level of sphingenine phosphate was found in the FB 1 -treated leaves even though free sphingenine was found in low amounts in these leaves. In comparison of WT and spp1 plants, the LCBP/LCB ratio is likely to be correlated with the degree of FB 1 -induced cell death determined by trypan blue staining. The FB 1 -treated leaves in dpl1 plants showed severe cell death and the elevation of free LCBs and LCBPs. LCBK1-OX and -KD plants showed resistance and sensitivity to FB 1 , respectively, whereas free LCB and LCBP levels in FB 1 -treated LCBK1-OX and -KD plants were moderately different to those in FB 1 -treated WT plants. Overall, the findings described here suggest that LCBP/LCB homeostasis is an important topic that participates in the tolerance of plant cells to FB 1 .
DOI:10.1016/j.bbrc.2014.10.142URLPMID:25449268 [本文引用: 1]
Glucosylceramides (GlcCers) are important lipid components of the membrane systems of eukaryotes. Recent studies have suggested the roles for GlcCers in regulating fungal growth and pathogenesis. In this study, we report the identification and functional characterization of PdGcs1, a gene encoding GlcCer synthase (GCS) essential for the biosynthesis of GlcCers, in Penicilliumdigitatum genome. We demonstrated that the deletion of PdGcs1 in P. digitatum resulted in the complete loss of production of GlcCer (d18:1/18:0h) and GlcCer (d18:2/18:0h), a decrease in vegetation growth and sporulation, and a delay in spore germination. The virulence of the PdGcs1 deletion mutant on citrus fruits was also impaired, as evidenced by the delayed occurrence of water soaking lesion and the formation of smaller size of lesion. These results suggest that PdGcs1 is a bona fide GCS that plays an important role in regulating cell growth, differentiation, and virulence of P. digitatum by controlling the biosynthesis of GlcCers.
1
... 植物与真菌的互利共生是自然界普遍存在的现象, 真菌能够提高宿主植物对水分和矿质营养元素的吸收, 通过影响宿主植物的物质代谢提高植物对资源的利用效率, 改善营养条件, 从而促进植物生长(
活性氧与植物细胞编程性死亡
1
2001
... ROS是需氧细胞在代谢过程中产生的一系列活性氧簇, 参与植物衰老、胁迫条件下和过敏反应中的PCD; 过氧化氢(H2O2)和超氧阴离子(O2-.)是导致细胞发生PCD的2类ROS; 但ROS究竟是细胞发生PCD的引发者, 还是仅为PCD中的一个作用因素, 目前尚不明确(
植物鞘脂的结构、代谢途径及其功能
1
2009
... 植物和真菌体内鞘脂生物合成的第1步反应都是丝氨酸和棕榈酰辅酶A在丝氨酸棕榈酰转移酶(SPT)作用下缩合生成3-酮基二氢鞘氨醇(
丛枝菌根真菌脂类代谢对共生信号调控的响应和反馈
1
2016
... 植物与真菌的互利共生是自然界普遍存在的现象, 真菌能够提高宿主植物对水分和矿质营养元素的吸收, 通过影响宿主植物的物质代谢提高植物对资源的利用效率, 改善营养条件, 从而促进植物生长(
黄土高原枸杞根际微生态特征及其共生真菌调控宿主生长与耐旱响应机制
1
2011
... 植物与真菌的互利共生是自然界普遍存在的现象, 真菌能够提高宿主植物对水分和矿质营养元素的吸收, 通过影响宿主植物的物质代谢提高植物对资源的利用效率, 改善营养条件, 从而促进植物生长(
Level of M(IP)2C sphingolipid affects plant defensin sensitivity, oxidative stress resistance and chronological life-span in yeast
0
2006
A glycan shield for bacterial sphingolipids
1
2008
... 在植物-细菌和植物-真菌的相互作用中, 植物鞘脂都是主要的信号分子, 可诱导植物产生抗性反应(
Structural analysis of fungal cerebrosides
2
2011
... 植物鞘脂中LCB的结构种类较多, LCB的链长一般是C18, 在C-1、C-3和C-4易发生羟基化, 在C-4和C-8易形成双键.除了d18:0和t18:0外, 常见的植物鞘脂LCB还包括: 鞘氨醇(2-氨基-4-十八烯-1,3-二醇, sphingosine, d18:14)、2-氨基-8-十八烯-1,3-二醇(sphing-8-enine, d18:18)、2-氨基-8-十八烯-1,3,4-三醇(4-hydroxysphing-8-enine, t18:18)和2-氨基-4,8-十八二烯-1,3-二醇(鞘氨醇-4,8-二烯, sphinga-4,8- dienine, d18:24,8) (
... 极性基团是指磷酸基和糖基, 一般连接在LCB的C-1位置上.根据极性基团的种类, 可以将鞘脂分为鞘磷脂(sphingomyelins)和鞘糖脂(glycosphingolipids)两大类.前者是神经酰胺的C-1羟基连接磷酸胆碱(phosphocholine)或磷酸乙醇胺(phosphoethanolamine), 后者是神经酰胺的C-1羟基连接单糖或寡多糖.脑苷脂(cerebrosides)和糖基肌醇磷酰基神经酰胺(glycosylinositol phosphorylceramides, GIPCs)是2类主要的鞘糖脂.脑苷脂类中最常见的是葡萄糖基神经酰胺(glucosylceramides, GlcCers)和半乳糖基神经酰胺(galactosylceramides, GalCers).GlcCers在真菌、植物和动物体内普遍存在, 而GalCers只存在于真菌和动物体内(
Loss of ceramide kinase in Arabidopsis impairs defenses and promotes ceramide accumulation and mito- chondrial H2O2 bursts
2
2014
... 植物与真菌在长期的协同进化过程中形成了复杂的作用机制, 植物既能有效抵御病原体侵染, 又能与有益真菌共生.在植物-真菌互作过程中, 脂类如磷脂、糖脂、鞘脂(sphingolipids)和甾醇脂质参与建立两个有机体之间的膜界面(
... RPW8是一种拟南芥抗性基因(R), 通过SA信号通路触发植物产生过敏反应来限制高氏白粉菌属(Golovinomyces)真菌引起的白粉病.
Overexpression of the tomato ASC-1 gene mediates high insensitivity to AAL toxins and fumonisin B1 in tomato hairy roots and confers resistance to Alternaria alternata f. sp. lycopersici in Nicotiana umbratica plants
1
2002
... SAMs (sphinganine-analog mycotoxins)是真菌产生的一类与鞘氨醇结构类似的真菌毒素.SAMs与植物LCBs竞争结合CerS, 导致植物体内鞘脂合成前体物质LCBs积累, 鞘脂合成减少, 引发植物PCD或坏死(
Sphingolipid Δ8 unsaturation is important for glucosylceramide biosynthesis and low-temperature performance in Arabidopsis
1
2012
... GlcCers和GIPCs是植物中含量最高的鞘脂类物质, 是植物细胞的质膜、液泡膜和内质网膜的重要组成部分(
Susceptibility of Phelipanche and Orobanche species to AAL-toxin
1
2009
... SAMs (sphinganine-analog mycotoxins)是真菌产生的一类与鞘氨醇结构类似的真菌毒素.SAMs与植物LCBs竞争结合CerS, 导致植物体内鞘脂合成前体物质LCBs积累, 鞘脂合成减少, 引发植物PCD或坏死(
Cerebroside as an elicitor for induced resistance against the downy mildew pathogen in pearl millet
1
2003
... 真菌脑苷脂是一类能激活植物防御反应的诱导子.Umemura等(2000)报道, 田间实验中, 向水稻(Ory- za sativa)叶片喷施来自稻瘟病病原菌Magnaporthe grisea的脑苷酯, 能诱导水稻叶片合成phytocassane A、B、C和D, 以及momilactone A和B等植保素(phytoalexins, PAs), 且作用强度与脑苷脂用量呈正相关; 此外, 叶片中一些编码病程相关蛋白(pathogenesis related protein, PR protein)(如PBZ1、β-葡聚糖酶、几丁质酶和过氧化物酶)的基因被快速诱导表达.用真菌脑苷脂处理水稻悬浮培养细胞, 也能诱导细胞积累PAs和PR蛋白; 向细胞悬浮液中添加Ca2+通道抑制剂, 可以显著抑制真菌脑苷脂诱导的PAs合成, 表明Ca2+在脑苷脂诱导的抗性响应中发挥重要作用(
Sphingolipids mediate differential echinocandin susceptibility in Candida albicans and Aspergillus nidulans
1
2015
... 已有许多利用遗传学方法研究细胞凋亡相关信号的报道, 涉及的信号分子主要包括Ca2+、水杨酸(salicylic acid, SA)和活性氧(reactive oxygen species, ROS)等.近几年, 研究人员通过遗传学方法对鞘脂相关性状进行了广泛分析, 相关报道主要集中在鞘脂的信号分子作用(
Characterization, quantification and subcellular localization of inositol-containing sphingolipids of the yeast, Saccharomyces cerevisiae
1
1994
... GlcCers和GIPCs是植物中含量最高的鞘脂类物质, 是植物细胞的质膜、液泡膜和内质网膜的重要组成部分(
Nuclear calcium controls the apoptotic-like cell death induced by D-erythro-sphinganine in tobacco cells
0
2010
Membrane engineering of S. cerevisiae targeting sphingolipid metabolism
1
2017
... 鞘脂类的结构中, 鞘氨醇的2-氨基与脂肪酸发生酰基化反应生成的N-脂酰鞘氨醇类物质, 又被称为神经酰胺类物质(ceramides, Cers).神经酰胺是鞘脂的基本结构单位, 也是最重要的中间代谢产物.植物鞘脂分子中, 脂肪酸可以酰化附着到不同的LCBs上.这些脂肪酸几乎都是α-右旋羟基化合物, 碳链长度在C14-C28之间; 脂肪酸在C-2位易发生羟基化, 在C-9位形成双键(
Modifications of sphingolipid content affect tolerance to hemibiotrophic and necrotrophic pathogens by modulating plant defense responses in Arabidopsis
2
2015
... 已有许多利用遗传学方法研究细胞凋亡相关信号的报道, 涉及的信号分子主要包括Ca2+、水杨酸(salicylic acid, SA)和活性氧(reactive oxygen species, ROS)等.近几年, 研究人员通过遗传学方法对鞘脂相关性状进行了广泛分析, 相关报道主要集中在鞘脂的信号分子作用(
... 在植物-细菌和植物-真菌的相互作用中, 植物鞘脂都是主要的信号分子, 可诱导植物产生抗性反应(
Fungal lipochitooligosaccharide symbiotic signals in arbu- scular mycorrhiza
1
2011
... 植物与真菌的互利共生是自然界普遍存在的现象, 真菌能够提高宿主植物对水分和矿质营养元素的吸收, 通过影响宿主植物的物质代谢提高植物对资源的利用效率, 改善营养条件, 从而促进植物生长(
Plant sphingolipids: function follows form
1
2013
... 植物与真菌在长期的协同进化过程中形成了复杂的作用机制, 植物既能有效抵御病原体侵染, 又能与有益真菌共生.在植物-真菌互作过程中, 脂类如磷脂、糖脂、鞘脂(sphingolipids)和甾醇脂质参与建立两个有机体之间的膜界面(
Plant sphingolipids: their importance in cellular organization and adaption
7
2016
... 鞘脂结构中的鞘氨醇部分又被称为长链碱基(long- chain base, LCB), 链长一般为C16-C20, 以C18为主.二氢鞘氨醇(2-氨基-十八烷-1,3-二醇, dihydrosphingosine, d18:0)和4-羟基鞘氨醇(2-氨基-十八烷-1,3,4-三醇, phytosphingosine, t18:0) (
... 植物鞘脂中LCB的结构种类较多, LCB的链长一般是C18, 在C-1、C-3和C-4易发生羟基化, 在C-4和C-8易形成双键.除了d18:0和t18:0外, 常见的植物鞘脂LCB还包括: 鞘氨醇(2-氨基-4-十八烯-1,3-二醇, sphingosine, d18:14)、2-氨基-8-十八烯-1,3-二醇(sphing-8-enine, d18:18)、2-氨基-8-十八烯-1,3,4-三醇(4-hydroxysphing-8-enine, t18:18)和2-氨基-4,8-十八二烯-1,3-二醇(鞘氨醇-4,8-二烯, sphinga-4,8- dienine, d18:24,8) (
... 鞘脂类的结构中, 鞘氨醇的2-氨基与脂肪酸发生酰基化反应生成的N-脂酰鞘氨醇类物质, 又被称为神经酰胺类物质(ceramides, Cers).神经酰胺是鞘脂的基本结构单位, 也是最重要的中间代谢产物.植物鞘脂分子中, 脂肪酸可以酰化附着到不同的LCBs上.这些脂肪酸几乎都是α-右旋羟基化合物, 碳链长度在C14-C28之间; 脂肪酸在C-2位易发生羟基化, 在C-9位形成双键(
... GlcCers和GIPCs是植物中含量最高的鞘脂类物质, 是植物细胞的质膜、液泡膜和内质网膜的重要组成部分(
... 从Cer开始的鞘脂生物合成途径主要有5条(
... 黑色加粗边框为真菌特有的产物.SPT: 丝氨酸棕榈酰转移酶; KDHR: 3-酮二氢鞘氨醇还原酶; CerS: (二氢)神经酰胺合酶; DES: 二氢神经酰胺去饱和酶; SK: 鞘氨醇激酶; SPP: 鞘氨醇-1-磷酸磷酸酶; SPL: 鞘氨醇-1-磷酸裂合酶; LCBH: 鞘氨醇羟化酶; CERK: 神经酰胺激酶; CPP: 神经酰胺-1-磷酸磷酸酶; SMS: 鞘磷脂合酶; SMase: 鞘磷脂酶; LCBK: LCB激酶; LCBPP: LCB-1-磷酸磷酸酶; LCBPL: LCB-1-磷酸裂合酶; IPCS: 肌醇磷脂酰神经酰胺合酶; IPUT1: 葡萄糖肌醇磷脂酰神经酰胺醛酸基转移酶1; MIPCS: 甘露糖肌醇磷脂酰神经酰胺合酶; IPT: 肌醇磷酸转移酶; CGT: 神经酰胺半乳糖基转移酶; GALC: 半乳糖神经酰胺酶; GCS: 葡糖基神经酰胺合酶; GCase: 葡糖基神经酰胺酶; CDase: 神经鞘氨醇酶
... 植物的鞘脂及其代谢产物是具有信号转导作用的活性脂质分子, 它们不仅能够调节细胞的生长、分化和增殖, 而且在植物防病和抗病方面发挥重要作用(
Abnormal glycosphingolipid mannosylation triggers salicylic acid mediated responses in Arabidopsis
1
2013
... MAPKs是一组能被不同种类的细胞外刺激激活的丝氨酸-苏氨酸蛋白激酶, 能够将信号从细胞表面转导至细胞核内部.Saucedo-García等(2011)通过研究拟南芥中LCBs积累与发生PCD的关系, 发现MAPK6在LCBs诱导发生PCD的过程中发挥信号转导(transducer)的作用, 从而介导PCD的下游过程.
Antifungal activity of plant defensin AFP1 in Brassica juncea involves the recognition of the methyl residue in glucosylceramide of target pathogen Candida albicans
1
2014
... 植物防御素(defensins)是一类具有抗真菌和抗细菌作用的多肽.GlcCers被认为是一些植物防御素的受体.在体外, 一些富含半胱氨酸、具有抗真菌作用的植物防御素需要在真菌GlcCers存在的条件下才能发挥作用, 如萝卜(Raphanus sativus)种子产生的植物防御素RsAFP2 (
Sphingolipids in bacteria and fungi
1
2001
... 在植物-细菌和植物-真菌的相互作用中, 植物鞘脂都是主要的信号分子, 可诱导植物产生抗性反应(
Pseudomonas syringae infection triggers de novo synthesis of phytosphingosine from sphinganine in Arabidopsis thaliana
1
2010
... 在植物-细菌和植物-真菌的相互作用中, 植物鞘脂都是主要的信号分子, 可诱导植物产生抗性反应(
Glucosylceramide synthase is essential for alfalfa defensin-mediated growth inhibition but not for pathogenicity of Fusarium graminearum
2
2007
... 真菌GlcCers代谢的改变能够影响真菌的生长发育, 及其侵染宿主的能力.禾谷镰孢菌(Fusarium graminearum)的菌丝顶端富含GlcCers, 其突变体ΔFggcs1缺少GlcCer合成酶(GCS)基因, 不能合成GlcCers.与野生型相比, ΔFggcs1分生孢子短, 细胞数量少, 结构发生了很大改变; 菌丝短, 气生菌丝稀少, 细胞极性生长受到破坏; 菌丝体对细胞壁降解酶具抗性; 尽管ΔFggcs1仍能侵染小麦穗和玉米(Zea mays)须, 但菌丝体在植物组织中的延伸能力显著降低, 致病力减弱(
... 植物防御素(defensins)是一类具有抗真菌和抗细菌作用的多肽.GlcCers被认为是一些植物防御素的受体.在体外, 一些富含半胱氨酸、具有抗真菌作用的植物防御素需要在真菌GlcCers存在的条件下才能发挥作用, 如萝卜(Raphanus sativus)种子产生的植物防御素RsAFP2 (
Sphingolipid C-9 methyltransferases are important for growth and virulence but not for sensitivity to antifungal plant defensins in Fusarium graminearum
2
2009
... 真菌GlcCers代谢的改变能够影响真菌的生长发育, 及其侵染宿主的能力.禾谷镰孢菌(Fusarium graminearum)的菌丝顶端富含GlcCers, 其突变体ΔFggcs1缺少GlcCer合成酶(GCS)基因, 不能合成GlcCers.与野生型相比, ΔFggcs1分生孢子短, 细胞数量少, 结构发生了很大改变; 菌丝短, 气生菌丝稀少, 细胞极性生长受到破坏; 菌丝体对细胞壁降解酶具抗性; 尽管ΔFggcs1仍能侵染小麦穗和玉米(Zea mays)须, 但菌丝体在植物组织中的延伸能力显著降低, 致病力减弱(
... 植物防御素(defensins)是一类具有抗真菌和抗细菌作用的多肽.GlcCers被认为是一些植物防御素的受体.在体外, 一些富含半胱氨酸、具有抗真菌作用的植物防御素需要在真菌GlcCers存在的条件下才能发挥作用, 如萝卜(Raphanus sativus)种子产生的植物防御素RsAFP2 (
Glucosylceramide synthase is an essential regulator of pathogenicity of Cryptococcus neoformans
1
2006
... GlcCers和GIPCs是植物中含量最高的鞘脂类物质, 是植物细胞的质膜、液泡膜和内质网膜的重要组成部分(
Sphingolipids as targets for treatment of fungal infections
3
2016
... 从Cer开始的鞘脂生物合成途径主要有5条(
... ;
... 黑色加粗边框为真菌特有的产物.SPT: 丝氨酸棕榈酰转移酶; KDHR: 3-酮二氢鞘氨醇还原酶; CerS: (二氢)神经酰胺合酶; DES: 二氢神经酰胺去饱和酶; SK: 鞘氨醇激酶; SPP: 鞘氨醇-1-磷酸磷酸酶; SPL: 鞘氨醇-1-磷酸裂合酶; LCBH: 鞘氨醇羟化酶; CERK: 神经酰胺激酶; CPP: 神经酰胺-1-磷酸磷酸酶; SMS: 鞘磷脂合酶; SMase: 鞘磷脂酶; LCBK: LCB激酶; LCBPP: LCB-1-磷酸磷酸酶; LCBPL: LCB-1-磷酸裂合酶; IPCS: 肌醇磷脂酰神经酰胺合酶; IPUT1: 葡萄糖肌醇磷脂酰神经酰胺醛酸基转移酶1; MIPCS: 甘露糖肌醇磷脂酰神经酰胺合酶; IPT: 肌醇磷酸转移酶; CGT: 神经酰胺半乳糖基转移酶; GALC: 半乳糖神经酰胺酶; GCS: 葡糖基神经酰胺合酶; GCase: 葡糖基神经酰胺酶; CDase: 神经鞘氨醇酶
MPK6, sphinganine and the LCB2a gene from serine palmitoyltransferase are required in the signaling pathway that mediates cell death induced by long chain bases in Arabidopsis
1
2011
... SAMs (sphinganine-analog mycotoxins)是真菌产生的一类与鞘氨醇结构类似的真菌毒素.SAMs与植物LCBs竞争结合CerS, 导致植物体内鞘脂合成前体物质LCBs积累, 鞘脂合成减少, 引发植物PCD或坏死(
Sphingolipids are dual specific drug targets for the management of pulmonary infections: perspective
1
2017
... 已有许多利用遗传学方法研究细胞凋亡相关信号的报道, 涉及的信号分子主要包括Ca2+、水杨酸(salicylic acid, SA)和活性氧(reactive oxygen species, ROS)等.近几年, 研究人员通过遗传学方法对鞘脂相关性状进行了广泛分析, 相关报道主要集中在鞘脂的信号分子作用(
Involvement of sphingoid bases in mediating reactive oxygen intermediate production and programmed cell death in Arabidopsis
2
2007
... 植物的鞘脂及其代谢产物是具有信号转导作用的活性脂质分子, 它们不仅能够调节细胞的生长、分化和增殖, 而且在植物防病和抗病方面发挥重要作用(
... SAMs (sphinganine-analog mycotoxins)是真菌产生的一类与鞘氨醇结构类似的真菌毒素.SAMs与植物LCBs竞争结合CerS, 导致植物体内鞘脂合成前体物质LCBs积累, 鞘脂合成减少, 引发植物PCD或坏死(
Lipids in plant-microbe interactions
1
2016
... 植物与真菌在长期的协同进化过程中形成了复杂的作用机制, 植物既能有效抵御病原体侵染, 又能与有益真菌共生.在植物-真菌互作过程中, 脂类如磷脂、糖脂、鞘脂(sphingolipids)和甾醇脂质参与建立两个有机体之间的膜界面(
The plant disease resistance gene Asc-1 prevents disruption of sphingolipid metabolism during AAL-toxin-induced programmed cell death
1
2002
... SAMs (sphinganine-analog mycotoxins)是真菌产生的一类与鞘氨醇结构类似的真菌毒素.SAMs与植物LCBs竞争结合CerS, 导致植物体内鞘脂合成前体物质LCBs积累, 鞘脂合成减少, 引发植物PCD或坏死(
Structural analysis of cerebrosides from Aspergillus fungi: the existence of galactosylceramide in A. oryzae
1
2014
... GlcCers和GIPCs是植物中含量最高的鞘脂类物质, 是植物细胞的质膜、液泡膜和内质网膜的重要组成部分(
Glycosylation of inositol phosphorylceramide sphingolipids is required for normal growth and reproduction in Arabidosis
1
2017
... 从Cer开始的鞘脂生物合成途径主要有5条(
DmAMP1, an antifungal plant defensin from dahlia (Dahlia merckii), interacts with sphingolipids from Saccharomyces cerevisiae
1
2003
... 植物防御素(defensins)是一类具有抗真菌和抗细菌作用的多肽.GlcCers被认为是一些植物防御素的受体.在体外, 一些富含半胱氨酸、具有抗真菌作用的植物防御素需要在真菌GlcCers存在的条件下才能发挥作用, 如萝卜(Raphanus sativus)种子产生的植物防御素RsAFP2 (
Defensins from insects and plants interact with fungal glucosylceramides
1
2004
... 植物防御素(defensins)是一类具有抗真菌和抗细菌作用的多肽.GlcCers被认为是一些植物防御素的受体.在体外, 一些富含半胱氨酸、具有抗真菌作用的植物防御素需要在真菌GlcCers存在的条件下才能发挥作用, 如萝卜(Raphanus sativus)种子产生的植物防御素RsAFP2 (
Ceramides induce programmed cell death in Arabidopsis cells in a calcium-dependent manner
0
2005
Sphingolipids in genetic and acquired forms of chronic kidney diseases
1
2017
... 已有许多利用遗传学方法研究细胞凋亡相关信号的报道, 涉及的信号分子主要包括Ca2+、水杨酸(salicylic acid, SA)和活性氧(reactive oxygen species, ROS)等.近几年, 研究人员通过遗传学方法对鞘脂相关性状进行了广泛分析, 相关报道主要集中在鞘脂的信号分子作用(
Elicitor activity of cerebroside, a sphingolipid elicitor, in cell suspension cultures of rice
1
2002
... 真菌脑苷脂是一类能激活植物防御反应的诱导子.Umemura等(2000)报道, 田间实验中, 向水稻(Ory- za sativa)叶片喷施来自稻瘟病病原菌Magnaporthe grisea的脑苷酯, 能诱导水稻叶片合成phytocassane A、B、C和D, 以及momilactone A和B等植保素(phytoalexins, PAs), 且作用强度与脑苷脂用量呈正相关; 此外, 叶片中一些编码病程相关蛋白(pathogenesis related protein, PR protein)(如PBZ1、β-葡聚糖酶、几丁质酶和过氧化物酶)的基因被快速诱导表达.用真菌脑苷脂处理水稻悬浮培养细胞, 也能诱导细胞积累PAs和PR蛋白; 向细胞悬浮液中添加Ca2+通道抑制剂, 可以显著抑制真菌脑苷脂诱导的PAs合成, 表明Ca2+在脑苷脂诱导的抗性响应中发挥重要作用(
Cerebroside elicitors found in diverse phytopathogens activate defense responses in rice plants
0
2000
Ca 2+-in- duced high amplitude swelling and cytochrome c release from wheat (Triticum aestivum L.) mitochondria under anoxic stress
1
2002
... Ca2+是细胞内信号转导的第二信使.对小麦(Triticum aestivum)根细胞的研究发现, 细胞质Ca2+浓度升高诱导线粒体通透性转换孔(permeability transition pore, PTP)开放, 使线粒体体积增大, 这是细胞发生PCD的前提(
An inositolphosphorylceramide synthase is involved in regulation of plant programmed cell death associated with defense in Arabidopsis
1
2008
... RPW8是一种拟南芥抗性基因(R), 通过SA信号通路触发植物产生过敏反应来限制高氏白粉菌属(Golovinomyces)真菌引起的白粉病.
Recently discovered functions of glucosylceramides in plants and fungi
4
2003
... 鞘脂结构中的鞘氨醇部分又被称为长链碱基(long- chain base, LCB), 链长一般为C16-C20, 以C18为主.二氢鞘氨醇(2-氨基-十八烷-1,3-二醇, dihydrosphingosine, d18:0)和4-羟基鞘氨醇(2-氨基-十八烷-1,3,4-三醇, phytosphingosine, t18:0) (
... d18:0: 二氢鞘氨醇; t18:0: 4-羟基鞘氨醇; d18:18: 2-氨基-8-十八烯-1,3-二醇; t18:18: 2-氨基-8-十八烯-1,3,4-三醇; d18:14: 2-氨基-4-十八烯-1,3-二醇; d18:24,8: 2-氨基-4,8-十八二烯-1,3-二醇
... 鞘脂类的结构中, 鞘氨醇的2-氨基与脂肪酸发生酰基化反应生成的N-脂酰鞘氨醇类物质, 又被称为神经酰胺类物质(ceramides, Cers).神经酰胺是鞘脂的基本结构单位, 也是最重要的中间代谢产物.植物鞘脂分子中, 脂肪酸可以酰化附着到不同的LCBs上.这些脂肪酸几乎都是α-右旋羟基化合物, 碳链长度在C14-C28之间; 脂肪酸在C-2位易发生羟基化, 在C-9位形成双键(
... GlcCers和GIPCs是植物中含量最高的鞘脂类物质, 是植物细胞的质膜、液泡膜和内质网膜的重要组成部分(
Synthesis and degradation of long-chain base phosphates affect fumonisin B1-induced cell death in Arabidopsis thaliana
1
2017
... 植物与真菌在长期的协同进化过程中形成了复杂的作用机制, 植物既能有效抵御病原体侵染, 又能与有益真菌共生.在植物-真菌互作过程中, 脂类如磷脂、糖脂、鞘脂(sphingolipids)和甾醇脂质参与建立两个有机体之间的膜界面(
Glucosylceramides are required for mycelial growth and full virulence in Penicillium digitatum
1
2014
... 真菌GlcCers代谢的改变能够影响真菌的生长发育, 及其侵染宿主的能力.禾谷镰孢菌(Fusarium graminearum)的菌丝顶端富含GlcCers, 其突变体ΔFggcs1缺少GlcCer合成酶(GCS)基因, 不能合成GlcCers.与野生型相比, ΔFggcs1分生孢子短, 细胞数量少, 结构发生了很大改变; 菌丝短, 气生菌丝稀少, 细胞极性生长受到破坏; 菌丝体对细胞壁降解酶具抗性; 尽管ΔFggcs1仍能侵染小麦穗和玉米(Zea mays)须, 但菌丝体在植物组织中的延伸能力显著降低, 致病力减弱(
备案号: 京ICP备16067583号-21
版权所有 © 2021 《植物学报》编辑部
地址:北京香山南辛村20号 邮编:100093
电话:010-62836135 010-62836131 E-mail:cbb@ibcas.ac.cn
本系统由北京玛格泰克科技发展有限公司设计开发

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}