Evolution and Expression of NCED Family Genes in Vitis vinifera
Xiaolong Wang, Fengzhi Liu, Xiangbin Shi, Xiaodi Wang, Xiaohao Ji, Zhiqiang Wang, Baoliang Wang, Xiaocui Zheng, Haibo Wang,*Key Laboratory of Mineral Nutrition and Fertilizers Efficient Utilization of Deciduous Fruit Tree, Liaoning Province, Key Laboratory of Germplasm Resources Utilization of Horticultural Crops, Ministry of Agriculture, Fruit Research Institute, Chinese Academy of Agricultural Sciences, Xingcheng 125100, China
Abstract 9-cis-epoxycarotenoid dioxygenase (NCED), a key rate-limiting enzyme in ABA biosynthesis in plants, is involved in plant drought, exogenous abscisic acid (ABA) and high salt response, and can reduce the damage of environmental stress on plants. With genome-wide identification and analysis of the grape NCED gene family, we aimed to understand the species evolution relationship and study the expression patterns of various genes in different tissues and under drought, ABA and high salt (NaCl) stress treatment, to lay the foundation for further study of the biological functions of NCED genes. A total of 12 NCED genes were found in the grape genome. The amino acid residues encoded by the genes are distributed between 510 aa (VvNCED2) and 625 aa (VvNCED10). The maximum molecular weight of the VvNCED protein was 70.53 kDa (VvNCED10) and the minimum was 57.85 kDa (VvNCED2). After differentiation from the ancestral gene, the grape NCED genes had five replication events with two loss events. The NCED1/2, NCED3/4, NCED6/7 and NCED9/10 gene pairs are thought to be produced by segmental duplication. The replication time of segmental duplication ranged from 3.08 to 120.0 million years ago, which is later than the differentiation of monocotyledons. As compared with the control, VvNCED1 was significantly upregulated by 72.1% after 48 h of ABA treatment, whereas VvNCED2 was significantly downregulated by 84.0%. The expression of VvNCED6 was higher in only roots under drought treatment for 14, 21 and 28 days than in the control: 2.49, 1.05 and 1.09 times of control values, respectively. The expression of VvNCED7 was only 1.07 times higher than the control value in roots under drought treatment for 14 days. After 72 h of ABA treatment, the expression of VvNCED3 was significantly downregulated by 59.5% as compared with the control, whereas VvNCED4 was significantly upregulated by 169.9% as compared with the control. The significant peaks in expression of VvNCED3/VvNCED4 after NaCl treatment were 24 and 48 h, respectively, up by 219.2% and 114.4%. The differential conserved-domain expression patterns with different stress treatments are the basis for the functional differentiation of NCED proteins. The functional differentiation of NCED during evolution may be conducive to the occurrence of replication events. Keywords:NCED gene family;grapevine;abiotic stress;gene expression
PDF (3078KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 王小龙, 刘凤之, 史祥宾, 王孝娣, 冀晓昊, 王志强, 王宝亮, 郑晓翠, 王海波. 葡萄NCED基因家族进化及表达分析. 植物学报, 2019, 54(4): 474-485 doi:10.11983/CBB18200 Wang Xiaolong, Liu Fengzhi, Shi Xiangbin, Wang Xiaodi, Ji Xiaohao, Wang Zhiqiang, Wang Baoliang, Zheng Xiaocui, Wang Haibo. Evolution and Expression of NCED Family Genes in Vitis vinifera. Chinese Bulletin of Botany, 2019, 54(4): 474-485 doi:10.11983/CBB18200
近年来, 超过10%的耕地遭受干旱和高盐等环境胁迫, 由于土壤中含水量低导致植物光合作用及有效养分下降, 从而引起全球作物产量损失50%以上(Roychoudhury et al., 2013)。脱落酸(ABA)在植物生命周期、胁迫信号调节及内源性提示, 包括水分限制、种子发育、休眠和性别等方面发挥重要作用(Mcadam and Brodribb, 2015; Mcadam et al., 2016), 一直被认为是通用的胁迫激素(Raghavendra et al., 2010)。9-顺式-环氧类胡萝卜素双加氧酶(9-cis-epoxycarotenoid dioxygenase, NCED)能够切割质体中的C40 9-顺式-环氧类胡萝卜素, 以释放C15的中间体黄原胶毒素。这一步骤是ABA生物合成途径中的限速步骤, 在叶片与空气蒸气压差较大时, 至少1个编码9-顺式-环氧类胡萝卜素限速酶的NCED基因会快速显著上调表达(Sussmilch et al., 2017)。大多数植物都含有1个受干旱胁迫诱导的NCED基因, 它的表达水平与响应干旱而积累的ABA含量相关, 是ABA生物合成的关键基因。此外, Ahrazem等(2012)研究表明, 番红花(Crocus sativus)中CstNCED基因受盐、低温、干旱和外源ABA等非生物胁迫的诱导表达。Wang等(2018)研究表明, 梅花(Prunus mume)中PmNCED1基因的表达量与PEG胁迫相关, 并随处理时间的增加而升高。徐学中等(2018)的研究证明水稻(Oryza sativa)中OsNCED基因在ABA合成和抗旱方面发挥重要作用。在植物体中, 干旱胁迫通过大量表达NCED基因提高内源ABA水平, 促使大量ABA诱导基因表达; 而ABA则通过调节离子通道的活性以促进气孔闭合, 这样可减少叶表面的蒸腾作用进而防止水分过度流失。
葡萄、拟南芥和水稻中的NCED氨基酸序列通过MEGA 7.0软件中的Clustal W序列比对和NJ (neighbor-joining)法构建系统发育树。NJ分析时选择No. of difference、1 000次bootstrap和pairwise deletion等程序来支撑系统发育树中的每个节点。将进化树中出现的直系同源基因对(节点值≥90)定义为复制基因对。其中, 基因对位于同一条染色体上且基因对之间距离小于100 kb被定义为串联复制, 其它被定义为片段复制。通过在线网站PGDD (http://chibba.agtec.uga.edu/duplication/)计算复制基因对的Ks值, 根据公式T=Ks/2λ×10-6 Mya (葡萄的λ值为6.5×10-9)计算复制基因对复制事件的发生时间(T) (Gaut et al., 1996)。
包含内含子相位的基因结构通过在线网站GSDS (http://gsds.cbi.pku.edu.cn/)完成。VvNCED蛋白中保守结构通过在线网站MEME (http://meme-suite.org/)提取, 其相关参数the largest number of discovered and conserved motif设定为8; motif width设定在30-50个氨基酸之间; 其它参数为默认设置。
1.3 NCED基因的表达分析
通过在线网站GEO (https://www.ncbi.nlm.nih.gov/gds/)下载标准化转录表达数据(Fasoli et al., 2012), 分析NCED基因在54种不同组织中的表达特征(GSE36128)。使用本地序列BLAST, 比对候选NCED基因相应的探针, 探针序列来自Affymetrix网站(http://www.affymetrix.com)。采用MeV (v4.9.0)软件构建热图(Saeed et al., 2006)。
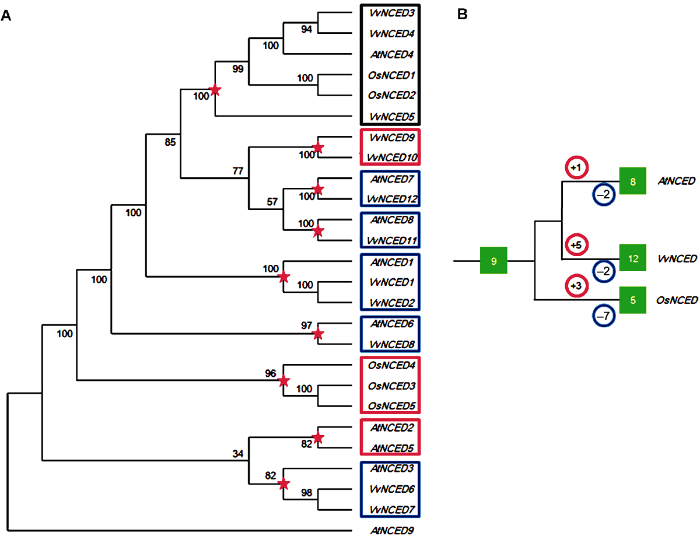
图中红色五角星代表物种分化前最近的共同祖先分化节点。红色和蓝色圆圈内的数字分别代表该物种在分化节点后发生的复制和丢失事件次数。绿色实心方框内的数字代表NCED基因的个数。 Figure 1Joint phylogenetic tree (A) and amplification model (B) of NCED gene from grape, Arabidopsis and rice
The red five-pointed stars in the figure represent the most recent common ancestral differentiation node before species differentiation. The numbers in the red and blue circles represent the number of duplication and loss events that occur after the species differentiated nodes, respectively. The numbers in the green solid box represent the number of NCED genes.
灰色条形框代表NCED蛋白全长, 其它彩色框中的数字是NCED蛋白保守结构域的随机编号。相同的数字代表相同的结构域。 Figure 3Schematic structure of conserved motifs identified in grapevine NCED proteins
The grey bars represent the full length of NCED proteins, and the numbers in the other colored boxes are random numbers for the conserved domains located at NCED protein. The same number represents the same conserved domain.
54种组织名称和基因名称分别位于热图的上方和右侧。热图上方比例尺表示基因表达量0.0-3.49。 Figure 4Expression pattern of NCED genes at different developmental stages and in some specialized tissues of grapevine
The 54 tissue names and gene names are located above and to the right of the heat map, respectively. The scale bar above the heat map indicates the geng expression level from 0.0 to 3.49.
干旱处理名称(DL1-DL5分别表示干旱处理0、7、14、21和28天叶组织; DR1-DR5分别表示干旱处理0、7、14、21和28天根组织)和基因名称分别位于热图的上方与右侧。热图上方比例尺表示基因表达量0.0-77.06。相同颜色的实心圆点代表复制基因对。 Figure 5Expression pattern of grapevine NCED genes under drought treatments
The drought treatments (DL1-DL5 indicate drought treatment for 0, 7, 14, 21 and 28 days of leaf tissue, respectively; DR1-DR5 indicate drought treatment for 0, 7, 14, 21 and 28 days of root tissue, respectively) and gene names are located above and to the right of the heat map, respectively. The scale bar above the heat map indicates the gene expression level from 0.0 to 77.06. Solid dots of the same color represent duplicated gene pairs.
AdamsKL, CronnR, PercifieldR, WendelJF ( 2003). Genes duplicated by polyploidy show unequal contributions to the transcriptome and organ-specific reciprocal silencing Proc Natl Acad Sci USA 100, 4649-4654. [本文引用: 1]
AhrazemO, Rubio-MoragaA, TraperoA, Gómez-GómezL ( 2012). Developmental and stress regulation of gene expression for a 9-cis-epoxycarotenoid dioxygenase, Cst NCED, isolated from Crocus sativus stigmas J Exp Bot 63, 681-694. [本文引用: 1]
ChawSM, ChangCC, ChenHL, LiWH ( 2004). Dating the monocot-dicot divergence and the origin of core eudicots using whole chloroplast genomes J Mol Evol 58, 424-441. [本文引用: 1]
Cohen-GihonI, SharanR, NussinovR ( 2011). Processes of fungal proteome evolution and gain of function: gene duplication and domain rearrangement Phys Biol 8, 035009. [本文引用: 1]
FasoliM, Dal SantoS, ZenoniS, TornielliGB, FarinaL, ZamboniA, PorcedduA, VenturiniL, BicegoM, MurinoV, FerrariniA, DelledonneM, PezzottiM ( 2012). The grapevine expression atlas reveals a deep transcriptome shift driving the entire plant into a maturation program Plant Cell 24, 3489-3505. [本文引用: 1]
GautBS, MortonBR, McCaigBC, CleggMT ( 1996). Substitution rate comparisons between grasses and palms: synonymous rate differences at the nuclear gene Adh parallel rate differences at the plastid gene rbcL Proc Natl Acad Sci USA 93, 10274-10279. [本文引用: 1]
GuZL, NicolaeD, LuHHS, LiWH ( 2002). Rapid divergence in expression between duplicate genes inferred from microarray data Trends Genet 18, 609-613. [本文引用: 1]
GuZL, RifkinSA, WhiteKP, LiWH ( 2004). Duplicate genes increase gene expression diversity within and between species Nat Genet 36, 577-579. [本文引用: 1]
GuoCL, GuoRR, XuXZ, GaoM, LiXQ, SongJY, ZhengY, WangXP ( 2014). Evolution and expression analysis of the grape (Vitis vinifera L.) WRKY gene family J Exp Bot 65, 1513-1528. [本文引用: 1]
MalacarneG, PerazzolliM, CestaroA, SterckL, FontanaP, Van de PeerY, ViolaR, VelascoR, SalaminiF ( 2012). Deconstruction of the (Paleo) polyploid grapevine genome based on the analysis of transposition events involving NBS resistance genes. PLoS One 7, e29762.
McAdamSAM, BrodribbTJ ( 2015). The evolution of mechanisms driving the stomatal response to vapor pressure deficit Plant Physiol 167, 833-843. [本文引用: 1]
McAdamSAM, BrodribbTJ, BanksJA, HedrichR, AtallahNM, CaiC, GeringerMA, LindC, NicholsDS, StachowskiK, GeigerD, SussmilchFC ( 2016). Abscisic acid controlled sex before transpiration in vascular plants Proc Natl Acad Sci USA 113, 12862-12867. [本文引用: 1]
RaghavendraAS, GonuguntaVK, ChristmannA, GrillE ( 2010). ABA perception and signaling Trends Plant Sci 15, 395-401. [本文引用: 1]
RoychoudhuryA, PaulS, BasuS ( 2013). Cross-talk between abscisic acid-dependent and abscisic acid-independent pathways during abiotic stress Plant Cell Rep 32, 985-1006. [本文引用: 1]
SussmilchFC, BrodribbTJ, McAdamSAM ( 2017). What are the evolutionary origins of stomatal responses to abscisic acid in land plants? J Integr Plant Biol 59, 240-260. [本文引用: 1]
TanBC, JosephLM, DengWT, LiuLJ, LiQB, ClineK, McCartyDR ( 2010). Molecular characterization of the Arabidopsis 9-cis epoxycarotenoid dioxygenase gene family Plant J 35, 44-56. [本文引用: 1]
WangRY, YangY, WangHG, ChenL, WangL, LuP, LiuMX, QiaoZJ ( 2018). Cloning of gene PmNCED1 and its response to PEG stress in common millet J Nuclear Agric Sci 32, 244-256. [本文引用: 1]
YangSH, ZhangXH, YueJX, TianDC, ChenJQ ( 2008). Recent duplications dominate NBS-encoding gene expan- sion in two woody species Mol Genet Genom 280, 187-198. [本文引用: 1]
YeX, KangBG, OsburnLD, LiY, ChengZM ( 2009). Identification of the flavin-dependent monooxygenase- encoding YUCCA gene family in Populus trichocarpa and their expression in vegetative tissues and in response to hormone and environmental stresses Plant Cell Tissue Organ Cult 97, 271-283. [本文引用: 1]
ZhangJZ ( 2003). Evolution by gene duplication: an update Trends Ecol Evol 18, 292-298. [本文引用: 1]
Genes duplicated by polyploidy show unequal contributions to the transcriptome and organ-specific reciprocal silencing 1 2003
... 基因复制是物种适应性进化的主要动力.通常, 由基因复制所产生的几个拷贝基因来维持其功能特异性, 该特异性为适应环境变化提供了更大的机会 (Cohen-Gihon et al., 2011).这种适应性的分化通常反映在复制基因的表达模式上(Gu et al., 2004).同源基因的表达可以在基因之间进行分化, 使得一个拷贝仅在某些器官中表达, 而另一个拷贝仅在其它器官中表达, 即亚功能化(Adams et al., 2003).此外, 同源基因对的表达模式变化也进一步证实了这种适应性分化.例如, VvNCED3/VvNCED4表达量均在ABA处理24小时出现显著上调, 较对照分别上调51.04%和159.32%; ABA处理72小时, VvNCED3表达量较对照显著下调59.47%, 而VvNCED4较对照显著上调169.89%.VvNCED3/VvNCED4在NaCl处理后出现的显著性峰值分别在24和48小时, 较对照上调219.19%和114.38%.由此可见, 复制基因对在响应环境胁迫时出现的差异性表达模式可能有利于进化过程中发生基因复制事件. ...
Developmental and stress regulation of gene expression for a 9-cis-epoxycarotenoid dioxygenase, Cst NCED, isolated from Crocus sativus stigmas 1 2012
... 近年来, 超过10%的耕地遭受干旱和高盐等环境胁迫, 由于土壤中含水量低导致植物光合作用及有效养分下降, 从而引起全球作物产量损失50%以上(Roychoudhury et al., 2013).脱落酸(ABA)在植物生命周期、胁迫信号调节及内源性提示, 包括水分限制、种子发育、休眠和性别等方面发挥重要作用(Mcadam and Brodribb, 2015; Mcadam et al., 2016), 一直被认为是通用的胁迫激素(Raghavendra et al., 2010).9-顺式-环氧类胡萝卜素双加氧酶(9-cis-epoxycarotenoid dioxygenase, NCED)能够切割质体中的C40 9-顺式-环氧类胡萝卜素, 以释放C15的中间体黄原胶毒素.这一步骤是ABA生物合成途径中的限速步骤, 在叶片与空气蒸气压差较大时, 至少1个编码9-顺式-环氧类胡萝卜素限速酶的NCED基因会快速显著上调表达(Sussmilch et al., 2017).大多数植物都含有1个受干旱胁迫诱导的NCED基因, 它的表达水平与响应干旱而积累的ABA含量相关, 是ABA生物合成的关键基因.此外, Ahrazem等(2012)研究表明, 番红花(Crocus sativus)中CstNCED基因受盐、低温、干旱和外源ABA等非生物胁迫的诱导表达.Wang等(2018)研究表明, 梅花(Prunus mume)中PmNCED1基因的表达量与PEG胁迫相关, 并随处理时间的增加而升高.徐学中等(2018)的研究证明水稻(Oryza sativa)中OsNCED基因在ABA合成和抗旱方面发挥重要作用.在植物体中, 干旱胁迫通过大量表达NCED基因提高内源ABA水平, 促使大量ABA诱导基因表达; 而ABA则通过调节离子通道的活性以促进气孔闭合, 这样可减少叶表面的蒸腾作用进而防止水分过度流失. ...
Dating the monocot-dicot divergence and the origin of core eudicots using whole chloroplast genomes 1 2004
... 目前葡萄单倍体基因组被认为起源于2个祖先基因组(Jaillon et al., 2007).葡萄共有19条稳定进化的染色体结构, 它们的同源三联体由2条Vc和1条Va染色体组成, 基因顺序高度保守, 只有罕见的易位才会被中断(Giulia et al., 2012).本研究发现4个(NCED1/2、NCED3/4、NCED6/7和NCED9/10)基因对被认为通过复制事件产生, 且均仅2个拷贝, 这可能与定位在Vc或Va染色体上的三联重复基因发生丢失有关.与Yang等(2008)的研究相似, 祖先同源连锁群往往分散在许多经过重排的染色体上, 期间基因组经历了大量基因丢失事件.上述4个旁系同源基因对的复制时间分布在3.08-120.0百万年前, 晚于单双子叶植物分化的时间(大约在150百万年前)(Chaw et al., 2004).这表明, NCED基因家族的扩增主要发生在单双子叶分化之后.拟南芥、水稻和葡萄联合系统发育树的结构显示, NCED祖先基因在单双子叶分化前就存在.在从祖先基因分化之后, 水稻NCED基因发生的丢失事件最多(7次), 其次是拟南芥和葡萄(均为2次); 葡萄NCED基因发生的复制事件最多(5次), 其次是水稻(3次), 最少是拟南芥(1次).本研究表明, 片段复制为NCED基因家族扩增作出主要贡献. ...
Processes of fungal proteome evolution and gain of function: gene duplication and domain rearrangement 1 2011
... 基因复制是物种适应性进化的主要动力.通常, 由基因复制所产生的几个拷贝基因来维持其功能特异性, 该特异性为适应环境变化提供了更大的机会 (Cohen-Gihon et al., 2011).这种适应性的分化通常反映在复制基因的表达模式上(Gu et al., 2004).同源基因的表达可以在基因之间进行分化, 使得一个拷贝仅在某些器官中表达, 而另一个拷贝仅在其它器官中表达, 即亚功能化(Adams et al., 2003).此外, 同源基因对的表达模式变化也进一步证实了这种适应性分化.例如, VvNCED3/VvNCED4表达量均在ABA处理24小时出现显著上调, 较对照分别上调51.04%和159.32%; ABA处理72小时, VvNCED3表达量较对照显著下调59.47%, 而VvNCED4较对照显著上调169.89%.VvNCED3/VvNCED4在NaCl处理后出现的显著性峰值分别在24和48小时, 较对照上调219.19%和114.38%.由此可见, 复制基因对在响应环境胁迫时出现的差异性表达模式可能有利于进化过程中发生基因复制事件. ...
The grapevine expression atlas reveals a deep transcriptome shift driving the entire plant into a maturation program 1 2012
... 通过在线网站GEO (https://www.ncbi.nlm.nih.gov/gds/)下载标准化转录表达数据(Fasoli et al., 2012), 分析NCED基因在54种不同组织中的表达特征(GSE36128).使用本地序列BLAST, 比对候选NCED基因相应的探针, 探针序列来自Affymetrix网站(http://www.affymetrix.com).采用MeV (v4.9.0)软件构建热图(Saeed et al., 2006). ...
Substitution rate comparisons between grasses and palms: synonymous rate differences at the nuclear gene Adh parallel rate differences at the plastid gene rbcL 1 1996
Cloning of gene PmNCED1 and its response to PEG stress in common millet 1 2018
... 近年来, 超过10%的耕地遭受干旱和高盐等环境胁迫, 由于土壤中含水量低导致植物光合作用及有效养分下降, 从而引起全球作物产量损失50%以上(Roychoudhury et al., 2013).脱落酸(ABA)在植物生命周期、胁迫信号调节及内源性提示, 包括水分限制、种子发育、休眠和性别等方面发挥重要作用(Mcadam and Brodribb, 2015; Mcadam et al., 2016), 一直被认为是通用的胁迫激素(Raghavendra et al., 2010).9-顺式-环氧类胡萝卜素双加氧酶(9-cis-epoxycarotenoid dioxygenase, NCED)能够切割质体中的C40 9-顺式-环氧类胡萝卜素, 以释放C15的中间体黄原胶毒素.这一步骤是ABA生物合成途径中的限速步骤, 在叶片与空气蒸气压差较大时, 至少1个编码9-顺式-环氧类胡萝卜素限速酶的NCED基因会快速显著上调表达(Sussmilch et al., 2017).大多数植物都含有1个受干旱胁迫诱导的NCED基因, 它的表达水平与响应干旱而积累的ABA含量相关, 是ABA生物合成的关键基因.此外, Ahrazem等(2012)研究表明, 番红花(Crocus sativus)中CstNCED基因受盐、低温、干旱和外源ABA等非生物胁迫的诱导表达.Wang等(2018)研究表明, 梅花(Prunus mume)中PmNCED1基因的表达量与PEG胁迫相关, 并随处理时间的增加而升高.徐学中等(2018)的研究证明水稻(Oryza sativa)中OsNCED基因在ABA合成和抗旱方面发挥重要作用.在植物体中, 干旱胁迫通过大量表达NCED基因提高内源ABA水平, 促使大量ABA诱导基因表达; 而ABA则通过调节离子通道的活性以促进气孔闭合, 这样可减少叶表面的蒸腾作用进而防止水分过度流失. ...
Recent duplications dominate NBS-encoding gene expan- sion in two woody species 1 2008
... 目前葡萄单倍体基因组被认为起源于2个祖先基因组(Jaillon et al., 2007).葡萄共有19条稳定进化的染色体结构, 它们的同源三联体由2条Vc和1条Va染色体组成, 基因顺序高度保守, 只有罕见的易位才会被中断(Giulia et al., 2012).本研究发现4个(NCED1/2、NCED3/4、NCED6/7和NCED9/10)基因对被认为通过复制事件产生, 且均仅2个拷贝, 这可能与定位在Vc或Va染色体上的三联重复基因发生丢失有关.与Yang等(2008)的研究相似, 祖先同源连锁群往往分散在许多经过重排的染色体上, 期间基因组经历了大量基因丢失事件.上述4个旁系同源基因对的复制时间分布在3.08-120.0百万年前, 晚于单双子叶植物分化的时间(大约在150百万年前)(Chaw et al., 2004).这表明, NCED基因家族的扩增主要发生在单双子叶分化之后.拟南芥、水稻和葡萄联合系统发育树的结构显示, NCED祖先基因在单双子叶分化前就存在.在从祖先基因分化之后, 水稻NCED基因发生的丢失事件最多(7次), 其次是拟南芥和葡萄(均为2次); 葡萄NCED基因发生的复制事件最多(5次), 其次是水稻(3次), 最少是拟南芥(1次).本研究表明, 片段复制为NCED基因家族扩增作出主要贡献. ...
Identification of the flavin-dependent monooxygenase- encoding YUCCA gene family in Populus trichocarpa and their expression in vegetative tissues and in response to hormone and environmental stresses 1 2009
... 使用北京华越洋生物科技有限公司RNA提取试剂盒提取葡萄叶片总RNA, 之后用TaKaRa试剂盒将总RNA反转录成cDNA, -20°C保存备用.利用Beacon Designer 7软件根据葡萄NCED家族基因预测的mRNA设计其特异性引物.所用引物由上海英骏生物技术有限公司(Invitrogen)合成.基因名称和引物序列见表1.qRT-PCR按照SYBR Green 试剂盒(TaKaRa公司)说明书操作, 反应在Mx3005p荧光定量PCR仪(Stratagene, 美国)上进行.每个样品设3次技术重复.用2-ΔΔCt方法计算基因相对表达量(Ye et al., 2009).使用Excel 2007和SPASS 23软件进行数据整理、图表绘制和差异显著性检验(Duncan’s). ...
 ,*中国农业科学院果树研究所, 农业部园艺作物种质资源利用重点实验室, 辽宁省落叶果树矿质营养与肥料高效利用重点实验室, 兴城 125100
,*中国农业科学院果树研究所, 农业部园艺作物种质资源利用重点实验室, 辽宁省落叶果树矿质营养与肥料高效利用重点实验室, 兴城 125100
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT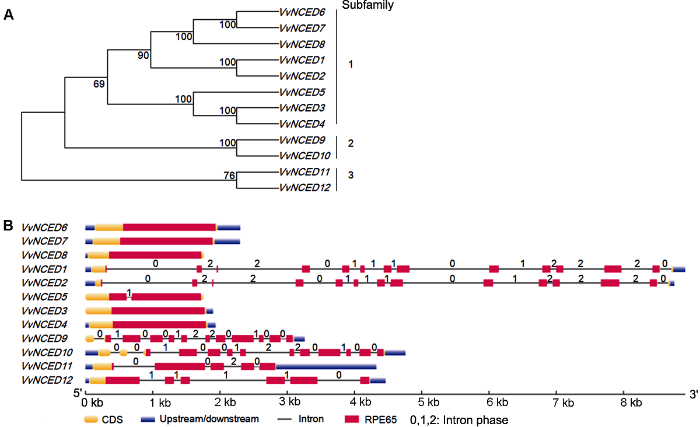 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT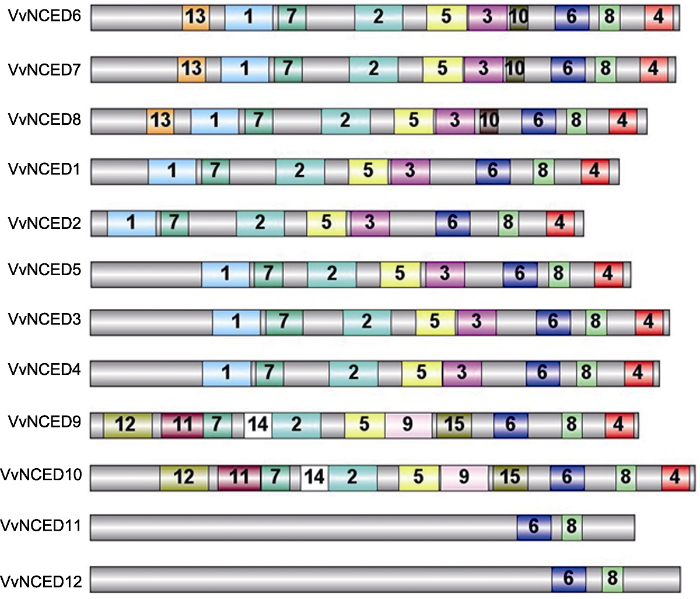 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT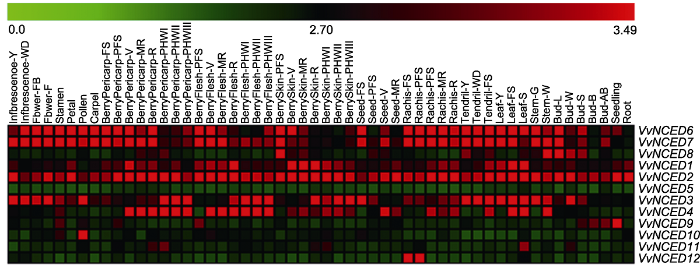 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT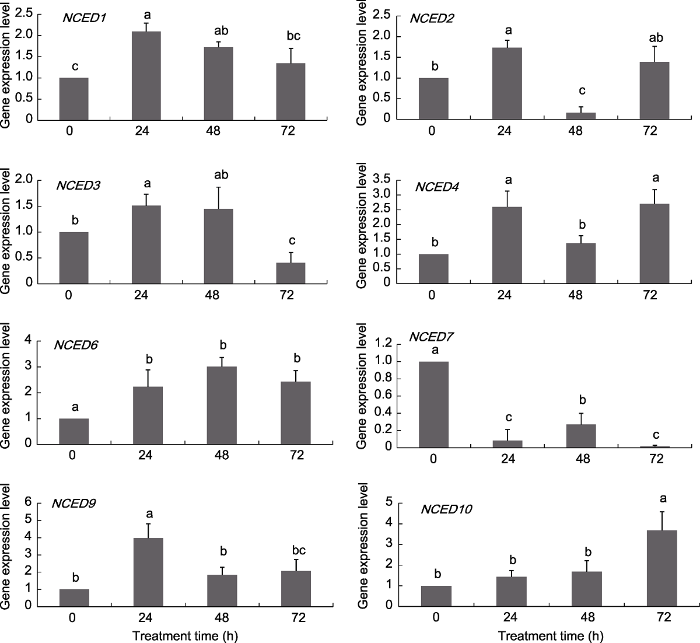 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT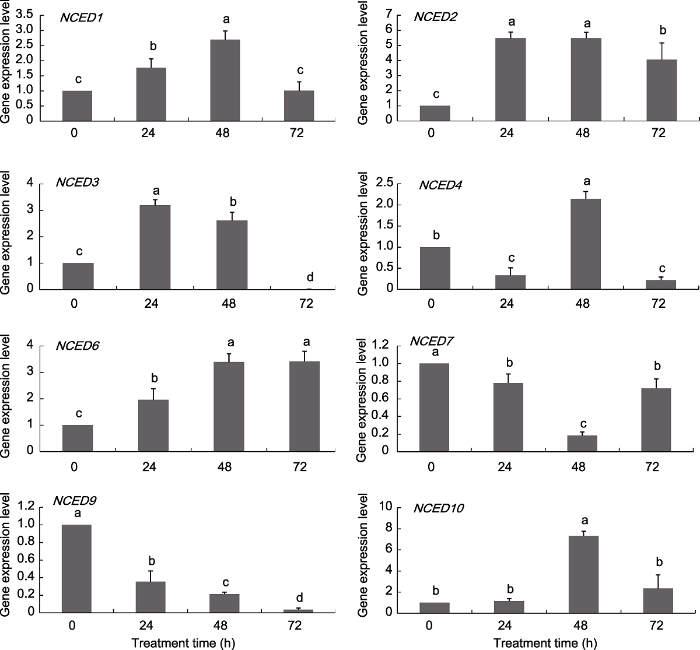 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}