Research Advances in the Functions of Exocyst Complex in Plants
Tonghui Li1, Xiaonan Liu2, Jing Xu2, Shipeng Li2, Su Jiang,2,*1. College of Life Sciences, Shandong Normal University, Jinan 250014, China 2. College of Life Sciences, Qilu Normal University, Jinan 250200, China
Abstract Vesicle trafficking is the biological process involving in budding, translocation, tethering and membrane fusion in all eukaryotic cells. Nine multisubunit tethering complexes (MTCs) are known to play roles in the intracellular transport, and the exocyst complex facilitates the tethering between transport vesicles and the plasma membrane (PM). The regulatory mechanism of the exocyst complex had been extensively studied in yeast and animals, whereas rapid research progress on plant exocyst has been made in recent years. Recent findings show that the plant exocyst complex has unique regulatory characteristics and is widely involved in plant growth, development and stress responses. Here we summarize research progress on the plant exocyst complex to provide a reference for future study of exocyst function in plants. Keywords:vesicle trafficking;multisubunit tethering complexes;exocyst complex
PDF (1533KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 李彤辉, 刘晓楠, 徐静, 李师鹏, 蒋苏. 胞泌复合体在植物中的功能研究进展. 植物学报, 2019, 54(5): 642-651 doi:10.11983/CBB18260 Li Tonghui, Liu Xiaonan, Xu Jing, Li Shipeng, Jiang Su. Research Advances in the Functions of Exocyst Complex in Plants. Chinese Bulletin of Botany, 2019, 54(5): 642-651 doi:10.11983/CBB18260
囊泡运输是所有真核生物中都存在的一种非常重要的细胞学过程, 负责细胞内膜系统之间及内膜系统与细胞膜之间的物质交流。囊泡运输过程主要包括4个环节: (1) 供体膜出芽形成运输囊泡; (2) 运输囊泡沿细胞骨架转运到靶膜附近; (3) 运输囊泡拴系到靶膜上; (4) SNARE (soluble N-ethyl-maleimide- sensitive fusion protein attachment protein receptor)复合体介导运输囊泡与靶膜发生融合(Bonifacino and Glick, 2004; Cai et al., 2007)。其中, 拴系过程介导运输囊泡与靶膜在空间上的最初接触, 在时空上调控下游膜融合事件的发生, 是调控极性囊泡运输的关键环节。研究表明, 一些蛋白复合体参与囊泡运输的拴系过程, 被称为拴系子(tethers)。拴系子包括两大类: 一类是卷曲螺线蛋白(coiled-coil proteins), 通常以二聚体形式存在, 被称为拴系蛋白; 另一类是多亚基的蛋白复合体(multisubunit tethering complexes, MTCs), 即拴系复合体。目前, 在真核生物中鉴定了9种拴系复合体, 分别是TRAPP I、TRAPP II、TRAPP III、COG、HOPS、CORVET、Dsl1、GARP和胞泌复合体(exocyst)。严格意义上讲, 拴系子需要同时与源于供体膜的囊泡和靶膜上的蛋白及膜脂互作, 介导两膜在物理空间上的直接接触。目前, 除了少数MTCs和卷曲螺线拴系蛋白外, 对于多数拴系子仍然缺乏直接的实验证据支持(Brunet and Sacher, 2014)。因此, 有****建议将至今还未明确拴系功能的复合体称为拴系因子(tethering factors)。与SNARE蛋白相比, MTCs的一个特点是其在细胞内具有时空特异性, 即不同MTCs作用于囊泡运输过程中不同目的膜。例如, DsI1位于内质网, COG在高尔基体, GARP作用于早期内体的返回途径, HOPS和CORVET分别在晚期内体到溶酶体以及内体之间的囊泡运输中发挥作用, TRAPP I/II/III存在于多种不同的囊泡运输途径, 胞泌复合体介导高尔基后囊泡向质膜的拴系过程(Yu and Hughson, 2010)。
胞泌复合体是介导胞吐(exocytosis)的拴系子, 了解其调控机理有助于理解细胞分泌、细胞分裂及细胞极性建立等基本生物学问题。对植物胞泌复合体的研究始于21世纪初(Elias et al., 2003), 研究显示植物胞泌复合体广泛参与植物生长发育和逆境响应。本文阐述真核细胞胞泌复合体的相关研究结果, 讨论胞泌复合体在植物中作用的特点和研究进展, 旨在为该领域研究提供参考。
目前, 对于植物胞泌复合体主要集中于遗传学研究, 细胞学研究尚处于起始阶段。已有的研究结果显示, 不同的胞泌复合体参与不同的生物学过程(表2)。在拟南芥中, 胞泌复合体成员SEC3A、SEC6、SEC8、SEC15B和EXO70A1亚基与EXO84B共定位于细胞板, EXO84B突变导致胞质分裂缺陷, 细胞质中有大量囊泡累积, 表明植物胞泌复合体介导的囊泡运输参与细胞板的形成(Fendrych et al., 2010)。玉米(Zea mays) SEC3同源物RTH1突变导致根毛变短, 显示RTH1参与根毛极性生长(Wen et al., 2005)。拟南芥SEC3A突变使花粉不能向后代传递, 表明SEC3A的功能对花粉萌发和花粉管生长非常重要(Bloch et al., 2016)。SEC5、SEC6、SEC8和SEC15A突变后同样显示出花粉萌发和花粉管生长缺陷(Hála et al., 2008)。拟南芥和水稻EXO70A1的功能与导管细胞次生壁发育有关(Li et al., 2013; Tu et al., 2015)。另据报道, 拟南芥EXO70A1突变植株生长素的运输和分布发生改变, 导致顶端优势丧失、根生长迟缓和侧根发生推迟等表型, 表明胞泌复合体可能参与PIN1和PIN2的再循环和极性定位(Drdová et al., 2013)。最近有报道显示, EXO70A1参与下胚轴-根转化区(root- hypocotyl junction)的分化, 暗培养的exo70a1幼苗在靠近转化区的下胚轴表皮细胞形成类似根毛的结构, 推测这一表型与生长素信号紊乱有关(Drdová et al., 2019)。EXO70C1在成熟花粉中高效表达, 其突变后会导致花粉管生长迟缓, 花粉的传递效率下降(Li et al., 2010)。EXO70H4在表皮毛细胞壁加厚和胼胝质沉积过程中不可或缺(Kulich et al., 2015)。大麦(Hordeum vulgare)基因沉默实验表明, EXO70F类似蛋白参与大麦抵抗真菌浸染的防御反应(Ostertag et al., 2013)。研究发现EXO70I参与丛枝菌根共生过程(Zhang et al., 2015)。BPH6蛋白可以与EXO70E1互作, 调控水稻细胞壁完整性, 抵御褐飞虱取食(Guo et al., 2018)。这些生物学过程都与胞吐关系密切, 暗示植物胞泌复合体广泛参与与胞吐有关的生物学过程, 但其具体作用机制尚需阐明。植物胞泌复合体参与囊泡拴系过程的直接证据还不充分。
AdamoJE, RossiG, BrennwaldP (1999). The Rho GTPase Rho3 has a direct role in exocytosis that is distinct from its role in actin polarity Mol Biol Cell 10, 4121-4133. [本文引用: 1]
AhmedSM, Nishida-FukudaH, LiYC, McDonaldWH, GradinaruCC, MacaraIG (2018). Exocyst dynamics during vesicle tethering and fusion Nat Commun 9, 5140. [本文引用: 3]
BlochD, PleskotR, PejcharP, PotockyM, Trpko?ováP, CwiklikL, Vuka?inoviN, SternbergH, YalovskyS, ZárskyV (2016). Exocyst SEC3 and phosphoinositides define sites of exocytosis in pollen tube initiation and growth Plant Physiol 172, 980-1002. [本文引用: 1]
BonifacinoJS, GlickBS (2004). The mechanisms of vesicle budding and fusion Cell 116, 153-166. [本文引用: 1]
BoydC, HughesT, PypaertM, NovickP (2004). Vesicles carry most exocyst subunits to exocytic sites marked by the remaining two subunits, Sec3p and Exo70p J Cell Biol 167, 889-901. [本文引用: 1]
BrunetS, SacherM (2014). Are all multisubunit tethering complexes bona fide tethers? Traffic 15, 1282-1287. [本文引用: 1]
CaiHQ, ReinischK, Ferro-NovickS (2007). Coats, tethers, Rabs, and SNAREs work together to mediate the intracellular destination of a transport vesicle Dev Cell 12, 671-682. [本文引用: 1]
ChongYT, GiddaSK, SanfordC, ParkinsonJ, MullenRT, GoringDR (2010). Characterization of the Arabidopsis thaliana exocyst complex gene families by phylogenetic, expression profiling, and subcellular localization studies New Phytol 185, 401-419. [本文引用: 1]
DingY, WangJ, LaiJHC, ChanVHL, WangXF, CaiY, TanXY, BaoYQ, XiaJ, RobinsonDG, JiangLW (2014). Exo70E2 is essential for exocyst subunit recruitment and EXPO formation in both plants and animals Mol Biol Cell 25, 412-426. [本文引用: 1]
DonovanKW, BretscherA (2015). Tracking individual secretory vesicles during exocytosis reveals an ordered and regulated process J Cell Biol 210, 181-189. [本文引用: 1]
DrdováEJ, KlejchováM, JankoK, HálaM, SoukupováH, Cvr?kováF, ?árskyV (2019). Developmental plasticity of Arabidopsis hypocotyl is dependent on exocyst complex function J Exp Bot 70, 1255-1265. [本文引用: 1]
DrdováEJ, SynekL, Pe?enkováT, HálaM, KulichI, FowlerJE, MurphyAS, ?árskyV (2013). The exocyst complex contributes to PIN auxin efflux carrier recycling and polar auxin transport in Arabidopsis Plant J 73, 709-719. [本文引用: 1]
EliasM, DrdovaE, ZiakD, BavlnkaB, HálaM, CvrckovaF, SoukupovaH, ?árskyV (2003). The exocyst complex in plants Cell Biol Int 27, 199-201. [本文引用: 3]
FendrychM, SynekL, Pe?enkováT, ToupalováH, ColeR, DrdováE, NebesárováJ, ?edinováM, HálaM, FowlerJE, ?árskyV (2010). The Arabidopsis exocyst complex is involved in cytokinesis and cell plate maturation Plant Cell 22, 3053-3065. [本文引用: 1]
FingerFP, HughesTE, NovickP (1998). Sec3p is a spatial landmark for polarized secretion in budding yeast Cell 92, 559-571. [本文引用: 1]
GrindstaffKK, YeamanC, AnandasabapathyN, HsuSC, Rodriguez-BoulanE, SchelleRH, NelsonWJ (1998). Sec6/8 complex is recruited to cell-cell contacts and specifies transport vesicle delivery to the basal-lateral membrane in epithelial cells Cell 93, 731-740. [本文引用: 1]
GuoJP, XuCX, WuD, ZhaoY, QiuYF, WangXX, OuyangYD, CaiBD, LiuX, JingSL, ShangguanXX, WangHY, MaYH, HuL, WuY, ShiSJ, WangWL, ZhuLL, XuX, ChenRZ, FengYQ, DuB, HeGC (2018). Bph6 encodes an exocyst-localized protein and confers broad resistance to planthoppers in rice Nat Genet 50, 297-306. [本文引用: 1]
GuoW, TamanoiF, NovickP (2001). Spatial regulation of the exocyst complex by Rho1 GTPase Nat Cell Biol 3, 353-360. [本文引用: 1]
HálaM, ColeR, SynekL, DrdováE, Pe?enkováT, NordheimA, LamkemeyerT, MadlungJ, HochholdingerF, FowlerJE, ?árskyV (2008). An exocyst complex functions in plant cell growth in Arabidopsis and tobacco Plant Cell 20, 1330-1345. [本文引用: 4]
KalmbachL, HématyK, De BellisD, BarberonM, FujitaS, UrsacheR, DaraspeJ, GeldnerN (2017). Transient cell-specific EXO70A1 activity in the CASP domain and casparian strip localization Nat Plants 3, 17058. [本文引用: 1]
KulichI, ColeR, DrdováE, Cvr?kováF, SoukupA, FowlerJ, ?árskyV (2010). Arabidopsis exocyst subunits SEC8 and EXO70A1 and exocyst interactor ROH1 are involved in the localized deposition of seed coat pectin New Phytol 188, 615-625. [本文引用: 1]
KulichI, Pe?enkováT, Sekere?J, SmetanaO, FendrychM, FoissnerI, H?ftbergerM, ?árskyV (2013). Arabidopsis exocyst subcomplex containing subunit EXO70B1 is involved in autophagy-related transport to the vacuole Traffic 14, 1155-1165. [本文引用: 2]
KulichI, VojtíkováZ, GlancM, OrtmannováJ, RasmannS, ?árskyV (2015). Cell wall maturation of Arabidopsis trichomes is dependent on exocyst subunit EXO70H4 and involves callose deposition Plant Physiol 168, 120-131. [本文引用: 1]
LiSP, ChenM, YuDL, RenSC, SunSF, LiuLD, KetelaarT, EmonsAMC, LiuCM (2013). EXO70A1-mediated vesicle trafficking is critical for tracheary element development in Arabidopsis Plant Cell 25, 1774-1786. [本文引用: 1]
LiSP, van OsGMA, RenSC, YuDL, KetelaarT, EmonsAMC, LiuCM (2010). Expression and functional analyses of EXO70 genes in Arabidopsis implicate their roles in regulating cell type-specific exocytosis Plant Physiol 154, 1819-1830. [本文引用: 1]
LiuDM, LiX, ShenD, NovickP (2018). Two subunits of the exocyst, Sec3p and Exo70p, can function exclusively on the plasma membrane Mol Biol Cell 29, 736-750. [本文引用: 1]
LuoGZ, ZhangJ, GuoW (2014). The role of Sec3p in secretory vesicle targeting and exocyst complex assembly Mol Biol Cell 25, 3813-3822. [本文引用: 2]
MaJ, ChenJ, WangM, RenYL, WangS, LeiCL, ChengZJ , Sodmergen (2018). Disruption of OsSEC3A increases the content of salicylic acid and induces plant defense responses in rice J Exp Bot 69, 1051-1064.
Martin-UrdirozM, DeeksMJ, HortonCG, DaweHR, JourdainI (2016). The exocyst complex in health and disease Front Cell Dev Biol 4, 24. [本文引用: 2]
MoskalenkoS, HenryDO, RosseC, MireyG, CamonisJH, WhiteMA (2002). The exocyst is a Ral effector complex Nat Cell Biol 4, 66-72. [本文引用: 1]
MunsonM, NovickP (2006). The exocyst defrocked, a framework of rods revealed Nat Struct Mol Biol 13, 577-581. [本文引用: 1]
OstertagM, StammlerJ, DouchkovD, EichmannR, HückelhovenR (2013). The conserved oligomeric golgi complex is involved in penetration resistance of barley to the barley powdery mildew fungus Mol Plant Pathol 14, 230-240. [本文引用: 1]
Pe?enkováT, HálaM, KulichI, KocourkováD, DrdováE, FendrychM, ToupalováH, ?árskyV (2011). The role for the exocyst complex subunits Exo70B2 and Exo70H1 in the plant-pathogen interaction J Exp Bot 62, 2107-2116. [本文引用: 2]
PleskotR, CwiklikL, JungwirthP, ?árskyV, PotockyM (2015). Membrane targeting of the yeast exocyst complex Biochim Biophys Acta 1848, 1481-1489. [本文引用: 1]
RawatA, Brej?kováL, HálaM, Cvr?kováF, ?árskyV (2017). The Physcomitrella patens exocyst subunit EXO70.3d has distinct roles in growth and development, and is essential for completion of the moss life cycle New Phytol 216, 438-454.
SamuelMA, ChongYT, HaasenKE, Aldea-BrydgesMG, StoneSL, GoringDR (2009). Cellular pathways regula- ting responses to compatible and self-incompatible pollen in Brassica and Arabidopsis stigmas intersect at Exo70A1, a putative component of the exocyst complex Plant Cell 21, 2655-2671.
Sekere?J, PejcharP, ?antr??ekJ, Vukasinovi?N, ?árskyV, PotockyM (2017). Analysis of exocyst subunit EXO70 family reveals distinct membrane polar domains in tobacco pollen tubes Plant Physiol 173, 1659-1675. [本文引用: 1]
ShenD, YuanH, HutagalungA, VermaA, KümmelD, WuXD, ReinischK, McNewJA, NovickP (2013). The synaptobrevin homologue Snc2p recruits the exocyst to secretory vesicles by binding to Sec6p J Cell Biol 202, 509-526. [本文引用: 1]
SivaramMVS, SaporitaJA, FurgasonMLM, BoettcherAJ, MunsonM (2005). Dimerization of the exocyst protein Sec6p and its interaction with the t-SNARE Sec9p Biochemistry 44, 6302-6311. [本文引用: 1]
SongerJA, MunsonM (2009). Sec6p anchors the assembled exocyst complex at sites of secretion Mol Biol Cell 20, 973-982. [本文引用: 1]
SynekL, SchlagerN, Eliá?M, QuentinM, HauserMT, ?árskyV (2006). AtExo70A1, a member of a family of putative exocyst subunits specifically expanded in land plants, is important for polar growth and plant development Plant J 48, 54-72.
TerBushDR, MauriceT, RothD, NovickP (1996). The exocyst is a multiprotein complex required for exocytosis in Saccharomyces cerevisiae. EMBO J 15, 6483-6494. [本文引用: 1]
TerBushDR, NovickP (1995). Sec6, Sec8, and Sec15 are components of a multisubunit complex which localizes to small bud tips in Saccharomyces cerevisiae J Cell Biol 130, 299-312. [本文引用: 1]
TingAE, HazukaCD, HsuSC, KirkMD, BeanAJ, SchellerRH (1995). rSec6 and rSec8, mammalian homologs of yeast proteins essential for secretion Proc Natl Acad Sci USA 92, 9613-9617. [本文引用: 1]
TsuboiT, RavierMA, XieH, EwartMA, GouldGW, BaldwinSA, RutterGA (2005). Mammalian exocyst complex is required for the docking step of insulin vesicle exocytosis J Biol Chem 280, 25565-25570. [本文引用: 1]
TuB, HuL, ChenWL, LiT, HuBH, ZhengL, LvZ, YouSJ, WangYP, MaBT, ChenXW, QinP, LiSG (2015). Disruption of OsEXO70A1 causes irregular vascular bundles and perturbs mineral nutrient assimilation in rice Sci Rep 5, 18609. [本文引用: 1]
VegaIE, HsuSC (2001). The exocyst complex associates with microtubules to mediate vesicle targeting and neurite outgrowth J Neuro Sci 21, 3839-3848. [本文引用: 1]
WangJ, DingY, WangJQ, HillmerS, MiaoYS, LoSW, WangXF, RobinsonDG, JiangLW (2010). EXPO, an exocyst-positive organelle distinct from multivesicular endosomes and autophagosomes, mediates cytosol to cell wall exocytosis in Arabidopsis and tobacco cells Plant Cell 22, 4009-4030. [本文引用: 2]
WangZ, LiPF, YangYJ, ChiYJ, FanBF, ChenZX (2016). Expression and functional analysis of a novel group of legume-specific WRKY and EXO70 protein variants from soybean Sci Rep 6, 32090.
WenTJ, HochholdingerF, SauerM, BruceW, SchnablePS (2005). The roothairless1 gene of maize encodes a homolog of sec3, which is involved in polar exocytosis Plant Physiol 138, 1637-1643. [本文引用: 1]
WuCY, TanL, van HoorenM, TanXY, LiuF, LiY, ZhaoYX, LiBX, RuiQC, MunnikT, BaoYQ (2017). Arabidopsis EXO70A1 recruits Patellin3 to the cell membrane independent of its role as an exocyst subunit J Integr Plant Biol 59, 851-865. [本文引用: 1]
WuH, TurnerC, GardnerJ, TempleB, BrennwaldP (2010). The Exo70 subunit of the exocyst is an effector for both Cdc42 and Rho3 function in polarized exocytosis Mol Biol Cell 21, 430-442. [本文引用: 1]
WuJD, TanXY, WuCY, CaoK, LiY, BaoYQ (2013). Regulation of cytokinesis by exocyst subunit SEC6 and KEULE in Arabidopsis thaliana Mol Plant 6, 1863-1876. [本文引用: 1]
WuSY, MehtaSQ, PichaudF, BellenHJ, QuiochoFA (2005). Sec15 interacts with Rab11 via a novel domain and affects Rab11 localization in vivo Nat Struct Mol Biol 12, 879-885. [本文引用: 1]
XiongXH, XuQW, HuangY, SinghRD, AndersonR, LeofE, HuJH, LingK (2012). An association between type Iγ PI4P 5-kinase and Exo70 directs E-cadherin clustering and epithelial polarization Mol Biol Cell 23, 87-98. [本文引用: 1]
YuIM, HughsonFM (2010). Tethering factors as organizers of intracellular vesicular traffic Annu Rev Cell Dev Biol 26, 137-156. [本文引用: 1]
?árskyV, Cvr?kováF, PotockyM, HálaM (2009). Exocytosis and cell polarity in plants exocyst and recycling domains New Phytol 183, 255-272. [本文引用: 1]
ZhangXC, PumplinN, IvanovS, HarrisonMJ (2015). EXO70I is required for development of a sub-domain of the periarbuscular membrane during arbuscular mycorrhizal symbiosis Curr Biol 25, 2189-2195. [本文引用: 1]
ZhangXY, BiEF, NovickP, DuLL, KozminskiKG, LipschutzJH, GuoW (2001). Cdc42 interacts with the exocyst and regulates polarized secretion J Biol Chem 276, 46745-46750. [本文引用: 1]
ZhangXY, OrlandoK, HeB, XiFG, ZhangJ, ZajacA, GuoW (2008). Membrane association and functional regulation of Sec3 by phospholipids and Cdc42 J Cell Biol 180, 145-158. [本文引用: 1]
ZhangXY, ZajacA, ZhangJ, WangPY, LiM, MurrayJ, TerBushD, GuoW (2005). The critical role of Exo84p in the organization and polarized localization of the exocyst complex J Biol Chem 280, 20356-20364. [本文引用: 1]
ZhangY, ImminkR, LiuCM, EmonsAM, KetelaarT (2013). The Arabidopsis exocyst subunit Sec3A is essential for embryo development and accumulates in transient puncta at the plasma membrane New Phytol 199, 74-88. [本文引用: 1]
ZhuXY, LiSD, PanSQ, XinXR, GuY (2018). CSI1, PATROL1, and exocyst complex cooperate in delivery of cellulose synthase complexes to the plasma membrane Proc Natl Acad Sci USA 115, E3578-E3587.
The Rho GTPase Rho3 has a direct role in exocytosis that is distinct from its role in actin polarity 1 1999
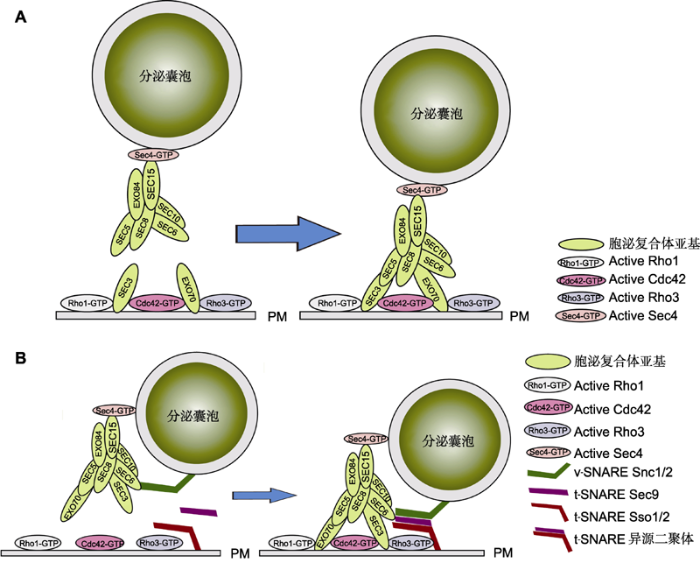
... 越来越多的实验证据表明, 胞泌复合体的功能受到Ras-GTP蛋白家族成员的调控, 并与SNARE蛋白发生关联, 调控下游的膜融合事件(Heider and Munson, 2012).质膜上的SEC3可以与小GTPase Cdc42和Rho1互作, 参与胞泌复合体在靶膜的组装(Guo et al., 2001; Zhang et al., 2001).EXO70也可与Cdc42和Rho3互作, 介导分泌囊泡与质膜的融合(Adamo et al., 1999; Wu et al., 2010).SEC3和EXO70之间的功能关系尚不明确.SEC15通过与分泌囊泡相关的Rab GTPase (SEC4)相互作用被招募到分泌囊泡(Wu et al., 2005), 同时在囊泡与质膜融合后促进马达蛋白的释放(Donovan and Bretscher, 2015).酵母双杂交实验结果显示, 在哺乳动物细胞中SEC5可以与Ras家族成员RalA-GTP相互作用, 抑制RalA-GTP的活性并影响胞泌复合体的组装, 说明RalA通过SEC5调控复合体的形成(Moskalenko et al., 2002) (图1A). ...
Exocyst dynamics during vesicle tethering and fusion 3 2018
... 最新研究表明, 哺乳动物细胞的胞泌复合体是高度动态的, 其组装方式为先形成2个四聚体亚复合物SC1 (SEC3、SEC5、SEC6和SEC8)和SC2 (SEC10、SEC15、EXO70和EXO84), 然后在胞质中组装成完整的八聚体, 并与囊泡结合共同到达质膜(Ahmed et al., 2018).有意思的是, 每个亚复合体可单独组装并与囊泡相关联, 但是两者不能单独促进囊泡融合(Ahmed et al., 2018).研究发现, 约65%的SC1和SC2可以组装成完整的八蛋白复合体, 约25%的SC1作为独立的亚复合体存在, 仅10%的亚基游离, 暗示哺乳动物细胞中的胞泌复合体处于游离亚基、亚复合体、八聚蛋白复合体的动态平衡中(Ahmed et al., 2018).SC1和SC2是否具有独立于复合体的功能值得探讨. ...
... ).有意思的是, 每个亚复合体可单独组装并与囊泡相关联, 但是两者不能单独促进囊泡融合(Ahmed et al., 2018).研究发现, 约65%的SC1和SC2可以组装成完整的八蛋白复合体, 约25%的SC1作为独立的亚复合体存在, 仅10%的亚基游离, 暗示哺乳动物细胞中的胞泌复合体处于游离亚基、亚复合体、八聚蛋白复合体的动态平衡中(Ahmed et al., 2018).SC1和SC2是否具有独立于复合体的功能值得探讨. ...
... ).研究发现, 约65%的SC1和SC2可以组装成完整的八蛋白复合体, 约25%的SC1作为独立的亚复合体存在, 仅10%的亚基游离, 暗示哺乳动物细胞中的胞泌复合体处于游离亚基、亚复合体、八聚蛋白复合体的动态平衡中(Ahmed et al., 2018).SC1和SC2是否具有独立于复合体的功能值得探讨. ...
Exocyst SEC3 and phosphoinositides define sites of exocytosis in pollen tube initiation and growth 1 2016
... 目前, 对于植物胞泌复合体主要集中于遗传学研究, 细胞学研究尚处于起始阶段.已有的研究结果显示, 不同的胞泌复合体参与不同的生物学过程(表2).在拟南芥中, 胞泌复合体成员SEC3A、SEC6、SEC8、SEC15B和EXO70A1亚基与EXO84B共定位于细胞板, EXO84B突变导致胞质分裂缺陷, 细胞质中有大量囊泡累积, 表明植物胞泌复合体介导的囊泡运输参与细胞板的形成(Fendrych et al., 2010).玉米(Zea mays) SEC3同源物RTH1突变导致根毛变短, 显示RTH1参与根毛极性生长(Wen et al., 2005).拟南芥SEC3A突变使花粉不能向后代传递, 表明SEC3A的功能对花粉萌发和花粉管生长非常重要(Bloch et al., 2016).SEC5、SEC6、SEC8和SEC15A突变后同样显示出花粉萌发和花粉管生长缺陷(Hála et al., 2008).拟南芥和水稻EXO70A1的功能与导管细胞次生壁发育有关(Li et al., 2013; Tu et al., 2015).另据报道, 拟南芥EXO70A1突变植株生长素的运输和分布发生改变, 导致顶端优势丧失、根生长迟缓和侧根发生推迟等表型, 表明胞泌复合体可能参与PIN1和PIN2的再循环和极性定位(Drdová et al., 2013).最近有报道显示, EXO70A1参与下胚轴-根转化区(root- hypocotyl junction)的分化, 暗培养的exo70a1幼苗在靠近转化区的下胚轴表皮细胞形成类似根毛的结构, 推测这一表型与生长素信号紊乱有关(Drdová et al., 2019).EXO70C1在成熟花粉中高效表达, 其突变后会导致花粉管生长迟缓, 花粉的传递效率下降(Li et al., 2010).EXO70H4在表皮毛细胞壁加厚和胼胝质沉积过程中不可或缺(Kulich et al., 2015).大麦(Hordeum vulgare)基因沉默实验表明, EXO70F类似蛋白参与大麦抵抗真菌浸染的防御反应(Ostertag et al., 2013).研究发现EXO70I参与丛枝菌根共生过程(Zhang et al., 2015).BPH6蛋白可以与EXO70E1互作, 调控水稻细胞壁完整性, 抵御褐飞虱取食(Guo et al., 2018).这些生物学过程都与胞吐关系密切, 暗示植物胞泌复合体广泛参与与胞吐有关的生物学过程, 但其具体作用机制尚需阐明.植物胞泌复合体参与囊泡拴系过程的直接证据还不充分. ...
The mechanisms of vesicle budding and fusion 1 2004
... 囊泡运输是所有真核生物中都存在的一种非常重要的细胞学过程, 负责细胞内膜系统之间及内膜系统与细胞膜之间的物质交流.囊泡运输过程主要包括4个环节: (1) 供体膜出芽形成运输囊泡; (2) 运输囊泡沿细胞骨架转运到靶膜附近; (3) 运输囊泡拴系到靶膜上; (4) SNARE (soluble N-ethyl-maleimide- sensitive fusion protein attachment protein receptor)复合体介导运输囊泡与靶膜发生融合(Bonifacino and Glick, 2004; Cai et al., 2007).其中, 拴系过程介导运输囊泡与靶膜在空间上的最初接触, 在时空上调控下游膜融合事件的发生, 是调控极性囊泡运输的关键环节.研究表明, 一些蛋白复合体参与囊泡运输的拴系过程, 被称为拴系子(tethers).拴系子包括两大类: 一类是卷曲螺线蛋白(coiled-coil proteins), 通常以二聚体形式存在, 被称为拴系蛋白; 另一类是多亚基的蛋白复合体(multisubunit tethering complexes, MTCs), 即拴系复合体.目前, 在真核生物中鉴定了9种拴系复合体, 分别是TRAPP I、TRAPP II、TRAPP III、COG、HOPS、CORVET、Dsl1、GARP和胞泌复合体(exocyst).严格意义上讲, 拴系子需要同时与源于供体膜的囊泡和靶膜上的蛋白及膜脂互作, 介导两膜在物理空间上的直接接触.目前, 除了少数MTCs和卷曲螺线拴系蛋白外, 对于多数拴系子仍然缺乏直接的实验证据支持(Brunet and Sacher, 2014).因此, 有****建议将至今还未明确拴系功能的复合体称为拴系因子(tethering factors).与SNARE蛋白相比, MTCs的一个特点是其在细胞内具有时空特异性, 即不同MTCs作用于囊泡运输过程中不同目的膜.例如, DsI1位于内质网, COG在高尔基体, GARP作用于早期内体的返回途径, HOPS和CORVET分别在晚期内体到溶酶体以及内体之间的囊泡运输中发挥作用, TRAPP I/II/III存在于多种不同的囊泡运输途径, 胞泌复合体介导高尔基后囊泡向质膜的拴系过程(Yu and Hughson, 2010). ...
Vesicles carry most exocyst subunits to exocytic sites marked by the remaining two subunits, Sec3p and Exo70p 1 2004
... 对胞泌复合体调控机制的认识主要源于酵母和动物细胞.在酵母及动物细胞中, 对于胞泌复合体在靶膜的募集机制还存在分歧.一般认为, SEC3和EXO70不依赖于微丝骨架, 两者能够通过与PIP2及小GTPase定位到胞吐位置的质膜上(Pleskot et al., 2015; Liu et al., 2018), 其它6个亚基(SEC5、SEC6、SEC8、SEC10、SEC15和EXO84)形成亚复合体(subcomplex), 在小GTPase (SEC4)参与下募集于分泌囊泡, 并依赖微丝骨架运送到胞吐位点, 在其它蛋白作用下, 与膜上的SEC3和EXO70组装成完整的复合体, 从而将囊泡拴系到质膜上(Finger et al., 1998; Boyd et al., 2004) (图1A).对酵母的研究显示, SEC3与线粒体蛋白(Tom20p)融合蛋白于线粒体异源表达时, 能够介导胞泌复合体其它亚基在线粒体中的募集, 进而导致原本运输到质膜的囊泡向线粒体转运(Luo et al., 2014), 说明SEC3蛋白在复合体组装和囊泡拴系过程中发挥关键作用. ...
Are all multisubunit tethering complexes bona fide tethers? 1 2014
... 囊泡运输是所有真核生物中都存在的一种非常重要的细胞学过程, 负责细胞内膜系统之间及内膜系统与细胞膜之间的物质交流.囊泡运输过程主要包括4个环节: (1) 供体膜出芽形成运输囊泡; (2) 运输囊泡沿细胞骨架转运到靶膜附近; (3) 运输囊泡拴系到靶膜上; (4) SNARE (soluble N-ethyl-maleimide- sensitive fusion protein attachment protein receptor)复合体介导运输囊泡与靶膜发生融合(Bonifacino and Glick, 2004; Cai et al., 2007).其中, 拴系过程介导运输囊泡与靶膜在空间上的最初接触, 在时空上调控下游膜融合事件的发生, 是调控极性囊泡运输的关键环节.研究表明, 一些蛋白复合体参与囊泡运输的拴系过程, 被称为拴系子(tethers).拴系子包括两大类: 一类是卷曲螺线蛋白(coiled-coil proteins), 通常以二聚体形式存在, 被称为拴系蛋白; 另一类是多亚基的蛋白复合体(multisubunit tethering complexes, MTCs), 即拴系复合体.目前, 在真核生物中鉴定了9种拴系复合体, 分别是TRAPP I、TRAPP II、TRAPP III、COG、HOPS、CORVET、Dsl1、GARP和胞泌复合体(exocyst).严格意义上讲, 拴系子需要同时与源于供体膜的囊泡和靶膜上的蛋白及膜脂互作, 介导两膜在物理空间上的直接接触.目前, 除了少数MTCs和卷曲螺线拴系蛋白外, 对于多数拴系子仍然缺乏直接的实验证据支持(Brunet and Sacher, 2014).因此, 有****建议将至今还未明确拴系功能的复合体称为拴系因子(tethering factors).与SNARE蛋白相比, MTCs的一个特点是其在细胞内具有时空特异性, 即不同MTCs作用于囊泡运输过程中不同目的膜.例如, DsI1位于内质网, COG在高尔基体, GARP作用于早期内体的返回途径, HOPS和CORVET分别在晚期内体到溶酶体以及内体之间的囊泡运输中发挥作用, TRAPP I/II/III存在于多种不同的囊泡运输途径, 胞泌复合体介导高尔基后囊泡向质膜的拴系过程(Yu and Hughson, 2010). ...
Coats, tethers, Rabs, and SNAREs work together to mediate the intracellular destination of a transport vesicle 1 2007
... 囊泡运输是所有真核生物中都存在的一种非常重要的细胞学过程, 负责细胞内膜系统之间及内膜系统与细胞膜之间的物质交流.囊泡运输过程主要包括4个环节: (1) 供体膜出芽形成运输囊泡; (2) 运输囊泡沿细胞骨架转运到靶膜附近; (3) 运输囊泡拴系到靶膜上; (4) SNARE (soluble N-ethyl-maleimide- sensitive fusion protein attachment protein receptor)复合体介导运输囊泡与靶膜发生融合(Bonifacino and Glick, 2004; Cai et al., 2007).其中, 拴系过程介导运输囊泡与靶膜在空间上的最初接触, 在时空上调控下游膜融合事件的发生, 是调控极性囊泡运输的关键环节.研究表明, 一些蛋白复合体参与囊泡运输的拴系过程, 被称为拴系子(tethers).拴系子包括两大类: 一类是卷曲螺线蛋白(coiled-coil proteins), 通常以二聚体形式存在, 被称为拴系蛋白; 另一类是多亚基的蛋白复合体(multisubunit tethering complexes, MTCs), 即拴系复合体.目前, 在真核生物中鉴定了9种拴系复合体, 分别是TRAPP I、TRAPP II、TRAPP III、COG、HOPS、CORVET、Dsl1、GARP和胞泌复合体(exocyst).严格意义上讲, 拴系子需要同时与源于供体膜的囊泡和靶膜上的蛋白及膜脂互作, 介导两膜在物理空间上的直接接触.目前, 除了少数MTCs和卷曲螺线拴系蛋白外, 对于多数拴系子仍然缺乏直接的实验证据支持(Brunet and Sacher, 2014).因此, 有****建议将至今还未明确拴系功能的复合体称为拴系因子(tethering factors).与SNARE蛋白相比, MTCs的一个特点是其在细胞内具有时空特异性, 即不同MTCs作用于囊泡运输过程中不同目的膜.例如, DsI1位于内质网, COG在高尔基体, GARP作用于早期内体的返回途径, HOPS和CORVET分别在晚期内体到溶酶体以及内体之间的囊泡运输中发挥作用, TRAPP I/II/III存在于多种不同的囊泡运输途径, 胞泌复合体介导高尔基后囊泡向质膜的拴系过程(Yu and Hughson, 2010). ...
Characterization of the Arabidopsis thaliana exocyst complex gene families by phylogenetic, expression profiling, and subcellular localization studies 1 2010
Exo70E2 is essential for exocyst subunit recruitment and EXPO formation in both plants and animals 1 2014
... 一般认为胞泌复合体在经典囊泡运输途径(conventional secretory pathway)中发挥作用.但也有证据显示, 复合体亚基在经典分泌途径之外的细胞学过程中行使功能(Kulich et al., 2013; Ding et al., 2014; Wu et al., 2017).姜里文研究组利用拟南芥和烟草BY-2细胞表达(X)FP标记的EXO70蛋白同源物, 并结合免疫电镜观察, 发现了一个称之为EXPO (exocyst positive organelles)的结构(Wang et al., 2010).他们利用荧光蛋白标记的EXO70E2对EXPO进行鉴定, 发现EXPO定位于细胞质、细胞膜和胞外基质, 与高尔基体、反式高尔基体网络、早期内体、多泡体和晚期内体的标记物不能共定位, 说明该结构未参与经典囊泡运输途径(Wang et al., 2010).目前, 对于EXPO的功能还不清楚, 进一步的遗传学研究将有助于明确这一结构的生物学意义. ...
Tracking individual secretory vesicles during exocytosis reveals an ordered and regulated process 1 2015
... 越来越多的实验证据表明, 胞泌复合体的功能受到Ras-GTP蛋白家族成员的调控, 并与SNARE蛋白发生关联, 调控下游的膜融合事件(Heider and Munson, 2012).质膜上的SEC3可以与小GTPase Cdc42和Rho1互作, 参与胞泌复合体在靶膜的组装(Guo et al., 2001; Zhang et al., 2001).EXO70也可与Cdc42和Rho3互作, 介导分泌囊泡与质膜的融合(Adamo et al., 1999; Wu et al., 2010).SEC3和EXO70之间的功能关系尚不明确.SEC15通过与分泌囊泡相关的Rab GTPase (SEC4)相互作用被招募到分泌囊泡(Wu et al., 2005), 同时在囊泡与质膜融合后促进马达蛋白的释放(Donovan and Bretscher, 2015).酵母双杂交实验结果显示, 在哺乳动物细胞中SEC5可以与Ras家族成员RalA-GTP相互作用, 抑制RalA-GTP的活性并影响胞泌复合体的组装, 说明RalA通过SEC5调控复合体的形成(Moskalenko et al., 2002) (图1A). ...
Developmental plasticity of Arabidopsis hypocotyl is dependent on exocyst complex function 1 2019
... 目前, 对于植物胞泌复合体主要集中于遗传学研究, 细胞学研究尚处于起始阶段.已有的研究结果显示, 不同的胞泌复合体参与不同的生物学过程(表2).在拟南芥中, 胞泌复合体成员SEC3A、SEC6、SEC8、SEC15B和EXO70A1亚基与EXO84B共定位于细胞板, EXO84B突变导致胞质分裂缺陷, 细胞质中有大量囊泡累积, 表明植物胞泌复合体介导的囊泡运输参与细胞板的形成(Fendrych et al., 2010).玉米(Zea mays) SEC3同源物RTH1突变导致根毛变短, 显示RTH1参与根毛极性生长(Wen et al., 2005).拟南芥SEC3A突变使花粉不能向后代传递, 表明SEC3A的功能对花粉萌发和花粉管生长非常重要(Bloch et al., 2016).SEC5、SEC6、SEC8和SEC15A突变后同样显示出花粉萌发和花粉管生长缺陷(Hála et al., 2008).拟南芥和水稻EXO70A1的功能与导管细胞次生壁发育有关(Li et al., 2013; Tu et al., 2015).另据报道, 拟南芥EXO70A1突变植株生长素的运输和分布发生改变, 导致顶端优势丧失、根生长迟缓和侧根发生推迟等表型, 表明胞泌复合体可能参与PIN1和PIN2的再循环和极性定位(Drdová et al., 2013).最近有报道显示, EXO70A1参与下胚轴-根转化区(root- hypocotyl junction)的分化, 暗培养的exo70a1幼苗在靠近转化区的下胚轴表皮细胞形成类似根毛的结构, 推测这一表型与生长素信号紊乱有关(Drdová et al., 2019).EXO70C1在成熟花粉中高效表达, 其突变后会导致花粉管生长迟缓, 花粉的传递效率下降(Li et al., 2010).EXO70H4在表皮毛细胞壁加厚和胼胝质沉积过程中不可或缺(Kulich et al., 2015).大麦(Hordeum vulgare)基因沉默实验表明, EXO70F类似蛋白参与大麦抵抗真菌浸染的防御反应(Ostertag et al., 2013).研究发现EXO70I参与丛枝菌根共生过程(Zhang et al., 2015).BPH6蛋白可以与EXO70E1互作, 调控水稻细胞壁完整性, 抵御褐飞虱取食(Guo et al., 2018).这些生物学过程都与胞吐关系密切, 暗示植物胞泌复合体广泛参与与胞吐有关的生物学过程, 但其具体作用机制尚需阐明.植物胞泌复合体参与囊泡拴系过程的直接证据还不充分. ...
The exocyst complex contributes to PIN auxin efflux carrier recycling and polar auxin transport in Arabidopsis 1 2013
... 目前, 对于植物胞泌复合体主要集中于遗传学研究, 细胞学研究尚处于起始阶段.已有的研究结果显示, 不同的胞泌复合体参与不同的生物学过程(表2).在拟南芥中, 胞泌复合体成员SEC3A、SEC6、SEC8、SEC15B和EXO70A1亚基与EXO84B共定位于细胞板, EXO84B突变导致胞质分裂缺陷, 细胞质中有大量囊泡累积, 表明植物胞泌复合体介导的囊泡运输参与细胞板的形成(Fendrych et al., 2010).玉米(Zea mays) SEC3同源物RTH1突变导致根毛变短, 显示RTH1参与根毛极性生长(Wen et al., 2005).拟南芥SEC3A突变使花粉不能向后代传递, 表明SEC3A的功能对花粉萌发和花粉管生长非常重要(Bloch et al., 2016).SEC5、SEC6、SEC8和SEC15A突变后同样显示出花粉萌发和花粉管生长缺陷(Hála et al., 2008).拟南芥和水稻EXO70A1的功能与导管细胞次生壁发育有关(Li et al., 2013; Tu et al., 2015).另据报道, 拟南芥EXO70A1突变植株生长素的运输和分布发生改变, 导致顶端优势丧失、根生长迟缓和侧根发生推迟等表型, 表明胞泌复合体可能参与PIN1和PIN2的再循环和极性定位(Drdová et al., 2013).最近有报道显示, EXO70A1参与下胚轴-根转化区(root- hypocotyl junction)的分化, 暗培养的exo70a1幼苗在靠近转化区的下胚轴表皮细胞形成类似根毛的结构, 推测这一表型与生长素信号紊乱有关(Drdová et al., 2019).EXO70C1在成熟花粉中高效表达, 其突变后会导致花粉管生长迟缓, 花粉的传递效率下降(Li et al., 2010).EXO70H4在表皮毛细胞壁加厚和胼胝质沉积过程中不可或缺(Kulich et al., 2015).大麦(Hordeum vulgare)基因沉默实验表明, EXO70F类似蛋白参与大麦抵抗真菌浸染的防御反应(Ostertag et al., 2013).研究发现EXO70I参与丛枝菌根共生过程(Zhang et al., 2015).BPH6蛋白可以与EXO70E1互作, 调控水稻细胞壁完整性, 抵御褐飞虱取食(Guo et al., 2018).这些生物学过程都与胞吐关系密切, 暗示植物胞泌复合体广泛参与与胞吐有关的生物学过程, 但其具体作用机制尚需阐明.植物胞泌复合体参与囊泡拴系过程的直接证据还不充分. ...
The exocyst subunit Sec6 interacts with assembled exocytic SNARE complexes 1 2015
... 拴系和膜融合事件在时序上前后紧密相连, 胞泌复合体和SNARE复合体之间存在功能关联, 二者协同作用可保证在时空上实现胞吐的精确控制.生化和遗传证据证明, 胞泌复合体能够直接帮助SNARE复合体组装并参与膜融合事件的发生(Hong and Lev, 2014).体外实验表明, 胞泌复合体亚基可与3种SNARE蛋白、SNARE复合体及其调控蛋白SEC1结合.酵母SEC3的N端结构域与t-SNARE Sso1/2结合并促进其与t-SNARE SEC9形成二聚体(Yue et al., 2017).SEC9与v-SNARE Snc1/2的互作有利于稳定囊泡与胞泌复合体的结合(Shen et al., 2013).另有研究表明, SEC6与t-SNARE (Sec9)互作促进SNARE复合体的组装、纠正错配的SNARE复合体以及激活膜融合发生(Dubuke et al., 2015).虽然目前尚不明确胞泌复合体与SNARE复合体的作用细节, 但基于现有的证据推测胞泌复合体首先与囊泡上的Rab GTPase、肌球蛋白(myosin)及v-SNARE互作, 启动胞泌复合体构象的改变, 暴露其与质膜和t-SNARE蛋白的作用位点, 从而驱动SNARE复合体的组装, 进而介导囊泡膜与靶膜的融合(图1B). ...
The exocyst complex in plants 3 2003
... 胞泌复合体是介导胞吐(exocytosis)的拴系子, 了解其调控机理有助于理解细胞分泌、细胞分裂及细胞极性建立等基本生物学问题.对植物胞泌复合体的研究始于21世纪初(Elias et al., 2003), 研究显示植物胞泌复合体广泛参与植物生长发育和逆境响应.本文阐述真核细胞胞泌复合体的相关研究结果, 讨论胞泌复合体在植物中作用的特点和研究进展, 旨在为该领域研究提供参考. ...
... 目前, 植物中编码胞泌复合体的8个亚基的同源基因已被鉴定(Elias et al., 2003).遗传学、生物化学和细胞学证据显示, 植物胞泌复合体也是以异源八聚体的形式参与胞吐相关的细胞学过程(Hála et al., 2008; Zarsky et al., 2009).但是, 目前尚未获得植物胞泌复合体作为拴系子的直接证据. ...
The Arabidopsis exocyst complex is involved in cytokinesis and cell plate maturation 1 2010
... 目前, 对于植物胞泌复合体主要集中于遗传学研究, 细胞学研究尚处于起始阶段.已有的研究结果显示, 不同的胞泌复合体参与不同的生物学过程(表2).在拟南芥中, 胞泌复合体成员SEC3A、SEC6、SEC8、SEC15B和EXO70A1亚基与EXO84B共定位于细胞板, EXO84B突变导致胞质分裂缺陷, 细胞质中有大量囊泡累积, 表明植物胞泌复合体介导的囊泡运输参与细胞板的形成(Fendrych et al., 2010).玉米(Zea mays) SEC3同源物RTH1突变导致根毛变短, 显示RTH1参与根毛极性生长(Wen et al., 2005).拟南芥SEC3A突变使花粉不能向后代传递, 表明SEC3A的功能对花粉萌发和花粉管生长非常重要(Bloch et al., 2016).SEC5、SEC6、SEC8和SEC15A突变后同样显示出花粉萌发和花粉管生长缺陷(Hála et al., 2008).拟南芥和水稻EXO70A1的功能与导管细胞次生壁发育有关(Li et al., 2013; Tu et al., 2015).另据报道, 拟南芥EXO70A1突变植株生长素的运输和分布发生改变, 导致顶端优势丧失、根生长迟缓和侧根发生推迟等表型, 表明胞泌复合体可能参与PIN1和PIN2的再循环和极性定位(Drdová et al., 2013).最近有报道显示, EXO70A1参与下胚轴-根转化区(root- hypocotyl junction)的分化, 暗培养的exo70a1幼苗在靠近转化区的下胚轴表皮细胞形成类似根毛的结构, 推测这一表型与生长素信号紊乱有关(Drdová et al., 2019).EXO70C1在成熟花粉中高效表达, 其突变后会导致花粉管生长迟缓, 花粉的传递效率下降(Li et al., 2010).EXO70H4在表皮毛细胞壁加厚和胼胝质沉积过程中不可或缺(Kulich et al., 2015).大麦(Hordeum vulgare)基因沉默实验表明, EXO70F类似蛋白参与大麦抵抗真菌浸染的防御反应(Ostertag et al., 2013).研究发现EXO70I参与丛枝菌根共生过程(Zhang et al., 2015).BPH6蛋白可以与EXO70E1互作, 调控水稻细胞壁完整性, 抵御褐飞虱取食(Guo et al., 2018).这些生物学过程都与胞吐关系密切, 暗示植物胞泌复合体广泛参与与胞吐有关的生物学过程, 但其具体作用机制尚需阐明.植物胞泌复合体参与囊泡拴系过程的直接证据还不充分. ...
Sec3p is a spatial landmark for polarized secretion in budding yeast 1 1998
... 对胞泌复合体调控机制的认识主要源于酵母和动物细胞.在酵母及动物细胞中, 对于胞泌复合体在靶膜的募集机制还存在分歧.一般认为, SEC3和EXO70不依赖于微丝骨架, 两者能够通过与PIP2及小GTPase定位到胞吐位置的质膜上(Pleskot et al., 2015; Liu et al., 2018), 其它6个亚基(SEC5、SEC6、SEC8、SEC10、SEC15和EXO84)形成亚复合体(subcomplex), 在小GTPase (SEC4)参与下募集于分泌囊泡, 并依赖微丝骨架运送到胞吐位点, 在其它蛋白作用下, 与膜上的SEC3和EXO70组装成完整的复合体, 从而将囊泡拴系到质膜上(Finger et al., 1998; Boyd et al., 2004) (图1A).对酵母的研究显示, SEC3与线粒体蛋白(Tom20p)融合蛋白于线粒体异源表达时, 能够介导胞泌复合体其它亚基在线粒体中的募集, 进而导致原本运输到质膜的囊泡向线粒体转运(Luo et al., 2014), 说明SEC3蛋白在复合体组装和囊泡拴系过程中发挥关键作用. ...
Sec6/8 complex is recruited to cell-cell contacts and specifies transport vesicle delivery to the basal-lateral membrane in epithelial cells 1 1998
... 随后, 胞泌复合体在哺乳动物大脑组织中被分离鉴定(Ting et al., 1995; Hsu et al., 1996).与酵母中的情况类似, 哺乳动物的8个亚基共沉淀于1个743 kDa的复合体(始称SEC6/SEC8, 后改为胞泌复合体)中.细胞学研究表明复合体定位于胞吐活跃的靶膜上(Grindstaff et al.,1998; Vega and Hsu, 2001; Tsuboi et al., 2005; Xiong et al., 2012), 说明胞泌复合体的功能在酵母和动物细胞中保守.除了胞吐过程, 胞泌复合体还参与多种细胞学过程, 如细胞极性建立、初级纤毛发生、胞质分裂、病原体入侵、肿瘤发生和转移(Heider and Munson, 2012; Martin-Urdiroz et al., 2016). ...
Bph6 encodes an exocyst-localized protein and confers broad resistance to planthoppers in rice 1 2018
... 目前, 对于植物胞泌复合体主要集中于遗传学研究, 细胞学研究尚处于起始阶段.已有的研究结果显示, 不同的胞泌复合体参与不同的生物学过程(表2).在拟南芥中, 胞泌复合体成员SEC3A、SEC6、SEC8、SEC15B和EXO70A1亚基与EXO84B共定位于细胞板, EXO84B突变导致胞质分裂缺陷, 细胞质中有大量囊泡累积, 表明植物胞泌复合体介导的囊泡运输参与细胞板的形成(Fendrych et al., 2010).玉米(Zea mays) SEC3同源物RTH1突变导致根毛变短, 显示RTH1参与根毛极性生长(Wen et al., 2005).拟南芥SEC3A突变使花粉不能向后代传递, 表明SEC3A的功能对花粉萌发和花粉管生长非常重要(Bloch et al., 2016).SEC5、SEC6、SEC8和SEC15A突变后同样显示出花粉萌发和花粉管生长缺陷(Hála et al., 2008).拟南芥和水稻EXO70A1的功能与导管细胞次生壁发育有关(Li et al., 2013; Tu et al., 2015).另据报道, 拟南芥EXO70A1突变植株生长素的运输和分布发生改变, 导致顶端优势丧失、根生长迟缓和侧根发生推迟等表型, 表明胞泌复合体可能参与PIN1和PIN2的再循环和极性定位(Drdová et al., 2013).最近有报道显示, EXO70A1参与下胚轴-根转化区(root- hypocotyl junction)的分化, 暗培养的exo70a1幼苗在靠近转化区的下胚轴表皮细胞形成类似根毛的结构, 推测这一表型与生长素信号紊乱有关(Drdová et al., 2019).EXO70C1在成熟花粉中高效表达, 其突变后会导致花粉管生长迟缓, 花粉的传递效率下降(Li et al., 2010).EXO70H4在表皮毛细胞壁加厚和胼胝质沉积过程中不可或缺(Kulich et al., 2015).大麦(Hordeum vulgare)基因沉默实验表明, EXO70F类似蛋白参与大麦抵抗真菌浸染的防御反应(Ostertag et al., 2013).研究发现EXO70I参与丛枝菌根共生过程(Zhang et al., 2015).BPH6蛋白可以与EXO70E1互作, 调控水稻细胞壁完整性, 抵御褐飞虱取食(Guo et al., 2018).这些生物学过程都与胞吐关系密切, 暗示植物胞泌复合体广泛参与与胞吐有关的生物学过程, 但其具体作用机制尚需阐明.植物胞泌复合体参与囊泡拴系过程的直接证据还不充分. ...
Spatial regulation of the exocyst complex by Rho1 GTPase 1 2001
... 越来越多的实验证据表明, 胞泌复合体的功能受到Ras-GTP蛋白家族成员的调控, 并与SNARE蛋白发生关联, 调控下游的膜融合事件(Heider and Munson, 2012).质膜上的SEC3可以与小GTPase Cdc42和Rho1互作, 参与胞泌复合体在靶膜的组装(Guo et al., 2001; Zhang et al., 2001).EXO70也可与Cdc42和Rho3互作, 介导分泌囊泡与质膜的融合(Adamo et al., 1999; Wu et al., 2010).SEC3和EXO70之间的功能关系尚不明确.SEC15通过与分泌囊泡相关的Rab GTPase (SEC4)相互作用被招募到分泌囊泡(Wu et al., 2005), 同时在囊泡与质膜融合后促进马达蛋白的释放(Donovan and Bretscher, 2015).酵母双杂交实验结果显示, 在哺乳动物细胞中SEC5可以与Ras家族成员RalA-GTP相互作用, 抑制RalA-GTP的活性并影响胞泌复合体的组装, 说明RalA通过SEC5调控复合体的形成(Moskalenko et al., 2002) (图1A). ...
An exocyst complex functions in plant cell growth in Arabidopsis and tobacco 4 2008
... 目前, 植物中编码胞泌复合体的8个亚基的同源基因已被鉴定(Elias et al., 2003).遗传学、生物化学和细胞学证据显示, 植物胞泌复合体也是以异源八聚体的形式参与胞吐相关的细胞学过程(Hála et al., 2008; Zarsky et al., 2009).但是, 目前尚未获得植物胞泌复合体作为拴系子的直接证据. ...
... 目前, 对于植物胞泌复合体主要集中于遗传学研究, 细胞学研究尚处于起始阶段.已有的研究结果显示, 不同的胞泌复合体参与不同的生物学过程(表2).在拟南芥中, 胞泌复合体成员SEC3A、SEC6、SEC8、SEC15B和EXO70A1亚基与EXO84B共定位于细胞板, EXO84B突变导致胞质分裂缺陷, 细胞质中有大量囊泡累积, 表明植物胞泌复合体介导的囊泡运输参与细胞板的形成(Fendrych et al., 2010).玉米(Zea mays) SEC3同源物RTH1突变导致根毛变短, 显示RTH1参与根毛极性生长(Wen et al., 2005).拟南芥SEC3A突变使花粉不能向后代传递, 表明SEC3A的功能对花粉萌发和花粉管生长非常重要(Bloch et al., 2016).SEC5、SEC6、SEC8和SEC15A突变后同样显示出花粉萌发和花粉管生长缺陷(Hála et al., 2008).拟南芥和水稻EXO70A1的功能与导管细胞次生壁发育有关(Li et al., 2013; Tu et al., 2015).另据报道, 拟南芥EXO70A1突变植株生长素的运输和分布发生改变, 导致顶端优势丧失、根生长迟缓和侧根发生推迟等表型, 表明胞泌复合体可能参与PIN1和PIN2的再循环和极性定位(Drdová et al., 2013).最近有报道显示, EXO70A1参与下胚轴-根转化区(root- hypocotyl junction)的分化, 暗培养的exo70a1幼苗在靠近转化区的下胚轴表皮细胞形成类似根毛的结构, 推测这一表型与生长素信号紊乱有关(Drdová et al., 2019).EXO70C1在成熟花粉中高效表达, 其突变后会导致花粉管生长迟缓, 花粉的传递效率下降(Li et al., 2010).EXO70H4在表皮毛细胞壁加厚和胼胝质沉积过程中不可或缺(Kulich et al., 2015).大麦(Hordeum vulgare)基因沉默实验表明, EXO70F类似蛋白参与大麦抵抗真菌浸染的防御反应(Ostertag et al., 2013).研究发现EXO70I参与丛枝菌根共生过程(Zhang et al., 2015).BPH6蛋白可以与EXO70E1互作, 调控水稻细胞壁完整性, 抵御褐飞虱取食(Guo et al., 2018).这些生物学过程都与胞吐关系密切, 暗示植物胞泌复合体广泛参与与胞吐有关的生物学过程, 但其具体作用机制尚需阐明.植物胞泌复合体参与囊泡拴系过程的直接证据还不充分. ...
... 对植物胞泌复合体各亚基之间互作关系的了解, 有助于揭示其组装机制和调控特点.目前, 通过酵母双杂交、免疫共沉淀及BiFC等手段已明确拟南芥胞泌复合体成员间的互作关系(Hála et al., 2008; Pečenková et al., 2011; Zhang et al., 2013) (图2).利用遗传学研究已对部分蛋白之间的互作进行了功能验证, 其中, SEC8和EXO70A1共同参与种子外壁中的果胶沉积(Kulich et al., 2010); EXO70B1与SEC5A和EXO84B复合物参与自噬途径(Kulich et al., 2013); EXO70B2和EXO70H1共同参与病原体的响应, 其中EXO70B2与形成抗病相关的细胞壁附着物有关(Pečenková et al., 2011).以上结果表明, 植物胞泌复合体亚基在活体中以复合体的形式发挥作用. ...
Exorcising the exocyst complex 2 2012
... 随后, 胞泌复合体在哺乳动物大脑组织中被分离鉴定(Ting et al., 1995; Hsu et al., 1996).与酵母中的情况类似, 哺乳动物的8个亚基共沉淀于1个743 kDa的复合体(始称SEC6/SEC8, 后改为胞泌复合体)中.细胞学研究表明复合体定位于胞吐活跃的靶膜上(Grindstaff et al.,1998; Vega and Hsu, 2001; Tsuboi et al., 2005; Xiong et al., 2012), 说明胞泌复合体的功能在酵母和动物细胞中保守.除了胞吐过程, 胞泌复合体还参与多种细胞学过程, 如细胞极性建立、初级纤毛发生、胞质分裂、病原体入侵、肿瘤发生和转移(Heider and Munson, 2012; Martin-Urdiroz et al., 2016). ...
... 越来越多的实验证据表明, 胞泌复合体的功能受到Ras-GTP蛋白家族成员的调控, 并与SNARE蛋白发生关联, 调控下游的膜融合事件(Heider and Munson, 2012).质膜上的SEC3可以与小GTPase Cdc42和Rho1互作, 参与胞泌复合体在靶膜的组装(Guo et al., 2001; Zhang et al., 2001).EXO70也可与Cdc42和Rho3互作, 介导分泌囊泡与质膜的融合(Adamo et al., 1999; Wu et al., 2010).SEC3和EXO70之间的功能关系尚不明确.SEC15通过与分泌囊泡相关的Rab GTPase (SEC4)相互作用被招募到分泌囊泡(Wu et al., 2005), 同时在囊泡与质膜融合后促进马达蛋白的释放(Donovan and Bretscher, 2015).酵母双杂交实验结果显示, 在哺乳动物细胞中SEC5可以与Ras家族成员RalA-GTP相互作用, 抑制RalA-GTP的活性并影响胞泌复合体的组装, 说明RalA通过SEC5调控复合体的形成(Moskalenko et al., 2002) (图1A). ...
The ROP2-RIC7 pathway negatively regulates light-induced stomatal opening by inhibiting exocyst subunit Exo70B1 in Arabidopsis 1 2016
... 酵母细胞中活化的SEC4募集于分泌囊泡膜, 并通过与SEC15互作将胞泌复合体的其它成员招募到分泌囊泡.在植物细胞中, 胞泌复合体与小GTPases直接互作的证据尚未见报道, 但研究人员发现拟南芥SEC3A与植物小G蛋白ROP GTPases之间可能通过中间蛋白ICR1发生互作(Lavy et al., 2007), 说明植物胞泌复合体可能通过植物特异的机制受到小GTPases的调控.植物ROP2 (Rho-type GTPase 2)及其下游的RIC7 (ROP-interactive Cdc42- and Rac- interactive binding motif-containing protein 7)负调控拟南芥光诱导的气孔开放.酵母双杂交实验结果表明, RIC7与EXO70B1相互作用, exo70b1和ric7/ exo70b1的敲除突变体可延缓光诱导下的气孔开放, 表明ROP2和RIC7通过抑制EXO70B1的功能来阻止气孔过度开放, 暗示EXO70B1介导的囊泡运输参与气孔开放的调控(Hong et al., 2016).进一步明确植物胞泌复合体和小GTPases的互作关系对深入理解复合体功能至关重要. ...
Tethering the assembly of SNARE complexes 1 2014
... 拴系和膜融合事件在时序上前后紧密相连, 胞泌复合体和SNARE复合体之间存在功能关联, 二者协同作用可保证在时空上实现胞吐的精确控制.生化和遗传证据证明, 胞泌复合体能够直接帮助SNARE复合体组装并参与膜融合事件的发生(Hong and Lev, 2014).体外实验表明, 胞泌复合体亚基可与3种SNARE蛋白、SNARE复合体及其调控蛋白SEC1结合.酵母SEC3的N端结构域与t-SNARE Sso1/2结合并促进其与t-SNARE SEC9形成二聚体(Yue et al., 2017).SEC9与v-SNARE Snc1/2的互作有利于稳定囊泡与胞泌复合体的结合(Shen et al., 2013).另有研究表明, SEC6与t-SNARE (Sec9)互作促进SNARE复合体的组装、纠正错配的SNARE复合体以及激活膜融合发生(Dubuke et al., 2015).虽然目前尚不明确胞泌复合体与SNARE复合体的作用细节, 但基于现有的证据推测胞泌复合体首先与囊泡上的Rab GTPase、肌球蛋白(myosin)及v-SNARE互作, 启动胞泌复合体构象的改变, 暴露其与质膜和t-SNARE蛋白的作用位点, 从而驱动SNARE复合体的组装, 进而介导囊泡膜与靶膜的融合(图1B). ...
The exocyst complex in polarized exocytosis 1 2004
... 对胞泌复合体的研究始于酵母细胞, 该复合体最早发现于温度敏感型的酿酒酵母(Saccharomyces cerevisiae)突变体中(TerBush and Novick, 1995; TerBush et al., 1996).胞泌复合体由SEC3、SEC5、SEC6、SEC8、SEC10、SEC15、EXO70和EXO84八个蛋白亚基组成, 后期研究表明该复合体广泛存在于真核生物中(Martin-Urdiroz et al., 2016).酿酒酵母增殖时, 通过胞吐将子细胞形成相关材料运输到出芽部位, 复合体中的任何一个亚基突变都会导致分泌囊泡在出芽区域细胞质内积累, 而Myo2 (V型肌动蛋白)突变导致囊泡在母细胞的细胞质中累积, 说明复合体作用于囊泡运输的下游(Hsu et al., 2004; Munson and Novick, 2006; Zhang et al., 2008; Songer and Munson, 2009).活细胞观察显示复合体的8个亚基聚集在胞吐活跃部位: 出芽初期位于出芽部位, 随后分布于正在生长的子细胞, 最后募集于母细胞与子细胞的连接部位(Zhang et al., 2005).复合体作为拴系子的直接证据来源于酵母异源表达实验, 当酵母SEC3亚基于线粒体膜异位表达时, 可观察到复合体其它亚基和囊泡在线粒体膜募集(Luo et al., 2014).该过程中SEC3蛋白CorEx基序发挥关键作用(Mei and Guo, 2018).这一作用特点表明胞泌复合体具有拴系子的功能. ...
The mammalian brain rsec6/8 complex 1 1996
... 随后, 胞泌复合体在哺乳动物大脑组织中被分离鉴定(Ting et al., 1995; Hsu et al., 1996).与酵母中的情况类似, 哺乳动物的8个亚基共沉淀于1个743 kDa的复合体(始称SEC6/SEC8, 后改为胞泌复合体)中.细胞学研究表明复合体定位于胞吐活跃的靶膜上(Grindstaff et al.,1998; Vega and Hsu, 2001; Tsuboi et al., 2005; Xiong et al., 2012), 说明胞泌复合体的功能在酵母和动物细胞中保守.除了胞吐过程, 胞泌复合体还参与多种细胞学过程, 如细胞极性建立、初级纤毛发生、胞质分裂、病原体入侵、肿瘤发生和转移(Heider and Munson, 2012; Martin-Urdiroz et al., 2016). ...
Transient cell-specific EXO70A1 activity in the CASP domain and casparian strip localization 1 2017
Arabidopsis exocyst subunits SEC8 and EXO70A1 and exocyst interactor ROH1 are involved in the localized deposition of seed coat pectin 1 2010
... 对植物胞泌复合体各亚基之间互作关系的了解, 有助于揭示其组装机制和调控特点.目前, 通过酵母双杂交、免疫共沉淀及BiFC等手段已明确拟南芥胞泌复合体成员间的互作关系(Hála et al., 2008; Pečenková et al., 2011; Zhang et al., 2013) (图2).利用遗传学研究已对部分蛋白之间的互作进行了功能验证, 其中, SEC8和EXO70A1共同参与种子外壁中的果胶沉积(Kulich et al., 2010); EXO70B1与SEC5A和EXO84B复合物参与自噬途径(Kulich et al., 2013); EXO70B2和EXO70H1共同参与病原体的响应, 其中EXO70B2与形成抗病相关的细胞壁附着物有关(Pečenková et al., 2011).以上结果表明, 植物胞泌复合体亚基在活体中以复合体的形式发挥作用. ...
Arabidopsis exocyst subcomplex containing subunit EXO70B1 is involved in autophagy-related transport to the vacuole 2 2013
... 一般认为胞泌复合体在经典囊泡运输途径(conventional secretory pathway)中发挥作用.但也有证据显示, 复合体亚基在经典分泌途径之外的细胞学过程中行使功能(Kulich et al., 2013; Ding et al., 2014; Wu et al., 2017).姜里文研究组利用拟南芥和烟草BY-2细胞表达(X)FP标记的EXO70蛋白同源物, 并结合免疫电镜观察, 发现了一个称之为EXPO (exocyst positive organelles)的结构(Wang et al., 2010).他们利用荧光蛋白标记的EXO70E2对EXPO进行鉴定, 发现EXPO定位于细胞质、细胞膜和胞外基质, 与高尔基体、反式高尔基体网络、早期内体、多泡体和晚期内体的标记物不能共定位, 说明该结构未参与经典囊泡运输途径(Wang et al., 2010).目前, 对于EXPO的功能还不清楚, 进一步的遗传学研究将有助于明确这一结构的生物学意义. ...
... 对植物胞泌复合体各亚基之间互作关系的了解, 有助于揭示其组装机制和调控特点.目前, 通过酵母双杂交、免疫共沉淀及BiFC等手段已明确拟南芥胞泌复合体成员间的互作关系(Hála et al., 2008; Pečenková et al., 2011; Zhang et al., 2013) (图2).利用遗传学研究已对部分蛋白之间的互作进行了功能验证, 其中, SEC8和EXO70A1共同参与种子外壁中的果胶沉积(Kulich et al., 2010); EXO70B1与SEC5A和EXO84B复合物参与自噬途径(Kulich et al., 2013); EXO70B2和EXO70H1共同参与病原体的响应, 其中EXO70B2与形成抗病相关的细胞壁附着物有关(Pečenková et al., 2011).以上结果表明, 植物胞泌复合体亚基在活体中以复合体的形式发挥作用. ...
Cell wall maturation of Arabidopsis trichomes is dependent on exocyst subunit EXO70H4 and involves callose deposition 1 2015
... 目前, 对于植物胞泌复合体主要集中于遗传学研究, 细胞学研究尚处于起始阶段.已有的研究结果显示, 不同的胞泌复合体参与不同的生物学过程(表2).在拟南芥中, 胞泌复合体成员SEC3A、SEC6、SEC8、SEC15B和EXO70A1亚基与EXO84B共定位于细胞板, EXO84B突变导致胞质分裂缺陷, 细胞质中有大量囊泡累积, 表明植物胞泌复合体介导的囊泡运输参与细胞板的形成(Fendrych et al., 2010).玉米(Zea mays) SEC3同源物RTH1突变导致根毛变短, 显示RTH1参与根毛极性生长(Wen et al., 2005).拟南芥SEC3A突变使花粉不能向后代传递, 表明SEC3A的功能对花粉萌发和花粉管生长非常重要(Bloch et al., 2016).SEC5、SEC6、SEC8和SEC15A突变后同样显示出花粉萌发和花粉管生长缺陷(Hála et al., 2008).拟南芥和水稻EXO70A1的功能与导管细胞次生壁发育有关(Li et al., 2013; Tu et al., 2015).另据报道, 拟南芥EXO70A1突变植株生长素的运输和分布发生改变, 导致顶端优势丧失、根生长迟缓和侧根发生推迟等表型, 表明胞泌复合体可能参与PIN1和PIN2的再循环和极性定位(Drdová et al., 2013).最近有报道显示, EXO70A1参与下胚轴-根转化区(root- hypocotyl junction)的分化, 暗培养的exo70a1幼苗在靠近转化区的下胚轴表皮细胞形成类似根毛的结构, 推测这一表型与生长素信号紊乱有关(Drdová et al., 2019).EXO70C1在成熟花粉中高效表达, 其突变后会导致花粉管生长迟缓, 花粉的传递效率下降(Li et al., 2010).EXO70H4在表皮毛细胞壁加厚和胼胝质沉积过程中不可或缺(Kulich et al., 2015).大麦(Hordeum vulgare)基因沉默实验表明, EXO70F类似蛋白参与大麦抵抗真菌浸染的防御反应(Ostertag et al., 2013).研究发现EXO70I参与丛枝菌根共生过程(Zhang et al., 2015).BPH6蛋白可以与EXO70E1互作, 调控水稻细胞壁完整性, 抵御褐飞虱取食(Guo et al., 2018).这些生物学过程都与胞吐关系密切, 暗示植物胞泌复合体广泛参与与胞吐有关的生物学过程, 但其具体作用机制尚需阐明.植物胞泌复合体参与囊泡拴系过程的直接证据还不充分. ...
A novel ROP/RAC effector links cell polarity, root-meristem maintenance, and vesicle trafficking 1 2007
... 酵母细胞中活化的SEC4募集于分泌囊泡膜, 并通过与SEC15互作将胞泌复合体的其它成员招募到分泌囊泡.在植物细胞中, 胞泌复合体与小GTPases直接互作的证据尚未见报道, 但研究人员发现拟南芥SEC3A与植物小G蛋白ROP GTPases之间可能通过中间蛋白ICR1发生互作(Lavy et al., 2007), 说明植物胞泌复合体可能通过植物特异的机制受到小GTPases的调控.植物ROP2 (Rho-type GTPase 2)及其下游的RIC7 (ROP-interactive Cdc42- and Rac- interactive binding motif-containing protein 7)负调控拟南芥光诱导的气孔开放.酵母双杂交实验结果表明, RIC7与EXO70B1相互作用, exo70b1和ric7/ exo70b1的敲除突变体可延缓光诱导下的气孔开放, 表明ROP2和RIC7通过抑制EXO70B1的功能来阻止气孔过度开放, 暗示EXO70B1介导的囊泡运输参与气孔开放的调控(Hong et al., 2016).进一步明确植物胞泌复合体和小GTPases的互作关系对深入理解复合体功能至关重要. ...
EXO70A1-mediated vesicle trafficking is critical for tracheary element development in Arabidopsis 1 2013
... 目前, 对于植物胞泌复合体主要集中于遗传学研究, 细胞学研究尚处于起始阶段.已有的研究结果显示, 不同的胞泌复合体参与不同的生物学过程(表2).在拟南芥中, 胞泌复合体成员SEC3A、SEC6、SEC8、SEC15B和EXO70A1亚基与EXO84B共定位于细胞板, EXO84B突变导致胞质分裂缺陷, 细胞质中有大量囊泡累积, 表明植物胞泌复合体介导的囊泡运输参与细胞板的形成(Fendrych et al., 2010).玉米(Zea mays) SEC3同源物RTH1突变导致根毛变短, 显示RTH1参与根毛极性生长(Wen et al., 2005).拟南芥SEC3A突变使花粉不能向后代传递, 表明SEC3A的功能对花粉萌发和花粉管生长非常重要(Bloch et al., 2016).SEC5、SEC6、SEC8和SEC15A突变后同样显示出花粉萌发和花粉管生长缺陷(Hála et al., 2008).拟南芥和水稻EXO70A1的功能与导管细胞次生壁发育有关(Li et al., 2013; Tu et al., 2015).另据报道, 拟南芥EXO70A1突变植株生长素的运输和分布发生改变, 导致顶端优势丧失、根生长迟缓和侧根发生推迟等表型, 表明胞泌复合体可能参与PIN1和PIN2的再循环和极性定位(Drdová et al., 2013).最近有报道显示, EXO70A1参与下胚轴-根转化区(root- hypocotyl junction)的分化, 暗培养的exo70a1幼苗在靠近转化区的下胚轴表皮细胞形成类似根毛的结构, 推测这一表型与生长素信号紊乱有关(Drdová et al., 2019).EXO70C1在成熟花粉中高效表达, 其突变后会导致花粉管生长迟缓, 花粉的传递效率下降(Li et al., 2010).EXO70H4在表皮毛细胞壁加厚和胼胝质沉积过程中不可或缺(Kulich et al., 2015).大麦(Hordeum vulgare)基因沉默实验表明, EXO70F类似蛋白参与大麦抵抗真菌浸染的防御反应(Ostertag et al., 2013).研究发现EXO70I参与丛枝菌根共生过程(Zhang et al., 2015).BPH6蛋白可以与EXO70E1互作, 调控水稻细胞壁完整性, 抵御褐飞虱取食(Guo et al., 2018).这些生物学过程都与胞吐关系密切, 暗示植物胞泌复合体广泛参与与胞吐有关的生物学过程, 但其具体作用机制尚需阐明.植物胞泌复合体参与囊泡拴系过程的直接证据还不充分. ...
Expression and functional analyses of EXO70 genes in Arabidopsis implicate their roles in regulating cell type-specific exocytosis 1 2010
... 目前, 对于植物胞泌复合体主要集中于遗传学研究, 细胞学研究尚处于起始阶段.已有的研究结果显示, 不同的胞泌复合体参与不同的生物学过程(表2).在拟南芥中, 胞泌复合体成员SEC3A、SEC6、SEC8、SEC15B和EXO70A1亚基与EXO84B共定位于细胞板, EXO84B突变导致胞质分裂缺陷, 细胞质中有大量囊泡累积, 表明植物胞泌复合体介导的囊泡运输参与细胞板的形成(Fendrych et al., 2010).玉米(Zea mays) SEC3同源物RTH1突变导致根毛变短, 显示RTH1参与根毛极性生长(Wen et al., 2005).拟南芥SEC3A突变使花粉不能向后代传递, 表明SEC3A的功能对花粉萌发和花粉管生长非常重要(Bloch et al., 2016).SEC5、SEC6、SEC8和SEC15A突变后同样显示出花粉萌发和花粉管生长缺陷(Hála et al., 2008).拟南芥和水稻EXO70A1的功能与导管细胞次生壁发育有关(Li et al., 2013; Tu et al., 2015).另据报道, 拟南芥EXO70A1突变植株生长素的运输和分布发生改变, 导致顶端优势丧失、根生长迟缓和侧根发生推迟等表型, 表明胞泌复合体可能参与PIN1和PIN2的再循环和极性定位(Drdová et al., 2013).最近有报道显示, EXO70A1参与下胚轴-根转化区(root- hypocotyl junction)的分化, 暗培养的exo70a1幼苗在靠近转化区的下胚轴表皮细胞形成类似根毛的结构, 推测这一表型与生长素信号紊乱有关(Drdová et al., 2019).EXO70C1在成熟花粉中高效表达, 其突变后会导致花粉管生长迟缓, 花粉的传递效率下降(Li et al., 2010).EXO70H4在表皮毛细胞壁加厚和胼胝质沉积过程中不可或缺(Kulich et al., 2015).大麦(Hordeum vulgare)基因沉默实验表明, EXO70F类似蛋白参与大麦抵抗真菌浸染的防御反应(Ostertag et al., 2013).研究发现EXO70I参与丛枝菌根共生过程(Zhang et al., 2015).BPH6蛋白可以与EXO70E1互作, 调控水稻细胞壁完整性, 抵御褐飞虱取食(Guo et al., 2018).这些生物学过程都与胞吐关系密切, 暗示植物胞泌复合体广泛参与与胞吐有关的生物学过程, 但其具体作用机制尚需阐明.植物胞泌复合体参与囊泡拴系过程的直接证据还不充分. ...
Two subunits of the exocyst, Sec3p and Exo70p, can function exclusively on the plasma membrane 1 2018
... 对胞泌复合体调控机制的认识主要源于酵母和动物细胞.在酵母及动物细胞中, 对于胞泌复合体在靶膜的募集机制还存在分歧.一般认为, SEC3和EXO70不依赖于微丝骨架, 两者能够通过与PIP2及小GTPase定位到胞吐位置的质膜上(Pleskot et al., 2015; Liu et al., 2018), 其它6个亚基(SEC5、SEC6、SEC8、SEC10、SEC15和EXO84)形成亚复合体(subcomplex), 在小GTPase (SEC4)参与下募集于分泌囊泡, 并依赖微丝骨架运送到胞吐位点, 在其它蛋白作用下, 与膜上的SEC3和EXO70组装成完整的复合体, 从而将囊泡拴系到质膜上(Finger et al., 1998; Boyd et al., 2004) (图1A).对酵母的研究显示, SEC3与线粒体蛋白(Tom20p)融合蛋白于线粒体异源表达时, 能够介导胞泌复合体其它亚基在线粒体中的募集, 进而导致原本运输到质膜的囊泡向线粒体转运(Luo et al., 2014), 说明SEC3蛋白在复合体组装和囊泡拴系过程中发挥关键作用. ...
The role of Sec3p in secretory vesicle targeting and exocyst complex assembly 2 2014
... 对胞泌复合体的研究始于酵母细胞, 该复合体最早发现于温度敏感型的酿酒酵母(Saccharomyces cerevisiae)突变体中(TerBush and Novick, 1995; TerBush et al., 1996).胞泌复合体由SEC3、SEC5、SEC6、SEC8、SEC10、SEC15、EXO70和EXO84八个蛋白亚基组成, 后期研究表明该复合体广泛存在于真核生物中(Martin-Urdiroz et al., 2016).酿酒酵母增殖时, 通过胞吐将子细胞形成相关材料运输到出芽部位, 复合体中的任何一个亚基突变都会导致分泌囊泡在出芽区域细胞质内积累, 而Myo2 (V型肌动蛋白)突变导致囊泡在母细胞的细胞质中累积, 说明复合体作用于囊泡运输的下游(Hsu et al., 2004; Munson and Novick, 2006; Zhang et al., 2008; Songer and Munson, 2009).活细胞观察显示复合体的8个亚基聚集在胞吐活跃部位: 出芽初期位于出芽部位, 随后分布于正在生长的子细胞, 最后募集于母细胞与子细胞的连接部位(Zhang et al., 2005).复合体作为拴系子的直接证据来源于酵母异源表达实验, 当酵母SEC3亚基于线粒体膜异位表达时, 可观察到复合体其它亚基和囊泡在线粒体膜募集(Luo et al., 2014).该过程中SEC3蛋白CorEx基序发挥关键作用(Mei and Guo, 2018).这一作用特点表明胞泌复合体具有拴系子的功能. ...
... 对胞泌复合体调控机制的认识主要源于酵母和动物细胞.在酵母及动物细胞中, 对于胞泌复合体在靶膜的募集机制还存在分歧.一般认为, SEC3和EXO70不依赖于微丝骨架, 两者能够通过与PIP2及小GTPase定位到胞吐位置的质膜上(Pleskot et al., 2015; Liu et al., 2018), 其它6个亚基(SEC5、SEC6、SEC8、SEC10、SEC15和EXO84)形成亚复合体(subcomplex), 在小GTPase (SEC4)参与下募集于分泌囊泡, 并依赖微丝骨架运送到胞吐位点, 在其它蛋白作用下, 与膜上的SEC3和EXO70组装成完整的复合体, 从而将囊泡拴系到质膜上(Finger et al., 1998; Boyd et al., 2004) (图1A).对酵母的研究显示, SEC3与线粒体蛋白(Tom20p)融合蛋白于线粒体异源表达时, 能够介导胞泌复合体其它亚基在线粒体中的募集, 进而导致原本运输到质膜的囊泡向线粒体转运(Luo et al., 2014), 说明SEC3蛋白在复合体组装和囊泡拴系过程中发挥关键作用. ...
Disruption of OsSEC3A increases the content of salicylic acid and induces plant defense responses in rice 0 2018
The exocyst complex in health and disease 2 2016
... 对胞泌复合体的研究始于酵母细胞, 该复合体最早发现于温度敏感型的酿酒酵母(Saccharomyces cerevisiae)突变体中(TerBush and Novick, 1995; TerBush et al., 1996).胞泌复合体由SEC3、SEC5、SEC6、SEC8、SEC10、SEC15、EXO70和EXO84八个蛋白亚基组成, 后期研究表明该复合体广泛存在于真核生物中(Martin-Urdiroz et al., 2016).酿酒酵母增殖时, 通过胞吐将子细胞形成相关材料运输到出芽部位, 复合体中的任何一个亚基突变都会导致分泌囊泡在出芽区域细胞质内积累, 而Myo2 (V型肌动蛋白)突变导致囊泡在母细胞的细胞质中累积, 说明复合体作用于囊泡运输的下游(Hsu et al., 2004; Munson and Novick, 2006; Zhang et al., 2008; Songer and Munson, 2009).活细胞观察显示复合体的8个亚基聚集在胞吐活跃部位: 出芽初期位于出芽部位, 随后分布于正在生长的子细胞, 最后募集于母细胞与子细胞的连接部位(Zhang et al., 2005).复合体作为拴系子的直接证据来源于酵母异源表达实验, 当酵母SEC3亚基于线粒体膜异位表达时, 可观察到复合体其它亚基和囊泡在线粒体膜募集(Luo et al., 2014).该过程中SEC3蛋白CorEx基序发挥关键作用(Mei and Guo, 2018).这一作用特点表明胞泌复合体具有拴系子的功能. ...
... 随后, 胞泌复合体在哺乳动物大脑组织中被分离鉴定(Ting et al., 1995; Hsu et al., 1996).与酵母中的情况类似, 哺乳动物的8个亚基共沉淀于1个743 kDa的复合体(始称SEC6/SEC8, 后改为胞泌复合体)中.细胞学研究表明复合体定位于胞吐活跃的靶膜上(Grindstaff et al.,1998; Vega and Hsu, 2001; Tsuboi et al., 2005; Xiong et al., 2012), 说明胞泌复合体的功能在酵母和动物细胞中保守.除了胞吐过程, 胞泌复合体还参与多种细胞学过程, 如细胞极性建立、初级纤毛发生、胞质分裂、病原体入侵、肿瘤发生和转移(Heider and Munson, 2012; Martin-Urdiroz et al., 2016). ...
The exocyst complex 3 2018
... 对胞泌复合体的研究始于酵母细胞, 该复合体最早发现于温度敏感型的酿酒酵母(Saccharomyces cerevisiae)突变体中(TerBush and Novick, 1995; TerBush et al., 1996).胞泌复合体由SEC3、SEC5、SEC6、SEC8、SEC10、SEC15、EXO70和EXO84八个蛋白亚基组成, 后期研究表明该复合体广泛存在于真核生物中(Martin-Urdiroz et al., 2016).酿酒酵母增殖时, 通过胞吐将子细胞形成相关材料运输到出芽部位, 复合体中的任何一个亚基突变都会导致分泌囊泡在出芽区域细胞质内积累, 而Myo2 (V型肌动蛋白)突变导致囊泡在母细胞的细胞质中累积, 说明复合体作用于囊泡运输的下游(Hsu et al., 2004; Munson and Novick, 2006; Zhang et al., 2008; Songer and Munson, 2009).活细胞观察显示复合体的8个亚基聚集在胞吐活跃部位: 出芽初期位于出芽部位, 随后分布于正在生长的子细胞, 最后募集于母细胞与子细胞的连接部位(Zhang et al., 2005).复合体作为拴系子的直接证据来源于酵母异源表达实验, 当酵母SEC3亚基于线粒体膜异位表达时, 可观察到复合体其它亚基和囊泡在线粒体膜募集(Luo et al., 2014).该过程中SEC3蛋白CorEx基序发挥关键作用(Mei and Guo, 2018).这一作用特点表明胞泌复合体具有拴系子的功能. ...
... 越来越多的实验证据表明, 胞泌复合体的功能受到Ras-GTP蛋白家族成员的调控, 并与SNARE蛋白发生关联, 调控下游的膜融合事件(Heider and Munson, 2012).质膜上的SEC3可以与小GTPase Cdc42和Rho1互作, 参与胞泌复合体在靶膜的组装(Guo et al., 2001; Zhang et al., 2001).EXO70也可与Cdc42和Rho3互作, 介导分泌囊泡与质膜的融合(Adamo et al., 1999; Wu et al., 2010).SEC3和EXO70之间的功能关系尚不明确.SEC15通过与分泌囊泡相关的Rab GTPase (SEC4)相互作用被招募到分泌囊泡(Wu et al., 2005), 同时在囊泡与质膜融合后促进马达蛋白的释放(Donovan and Bretscher, 2015).酵母双杂交实验结果显示, 在哺乳动物细胞中SEC5可以与Ras家族成员RalA-GTP相互作用, 抑制RalA-GTP的活性并影响胞泌复合体的组装, 说明RalA通过SEC5调控复合体的形成(Moskalenko et al., 2002) (图1A). ...
The exocyst defrocked, a framework of rods revealed 1 2006
... 对胞泌复合体的研究始于酵母细胞, 该复合体最早发现于温度敏感型的酿酒酵母(Saccharomyces cerevisiae)突变体中(TerBush and Novick, 1995; TerBush et al., 1996).胞泌复合体由SEC3、SEC5、SEC6、SEC8、SEC10、SEC15、EXO70和EXO84八个蛋白亚基组成, 后期研究表明该复合体广泛存在于真核生物中(Martin-Urdiroz et al., 2016).酿酒酵母增殖时, 通过胞吐将子细胞形成相关材料运输到出芽部位, 复合体中的任何一个亚基突变都会导致分泌囊泡在出芽区域细胞质内积累, 而Myo2 (V型肌动蛋白)突变导致囊泡在母细胞的细胞质中累积, 说明复合体作用于囊泡运输的下游(Hsu et al., 2004; Munson and Novick, 2006; Zhang et al., 2008; Songer and Munson, 2009).活细胞观察显示复合体的8个亚基聚集在胞吐活跃部位: 出芽初期位于出芽部位, 随后分布于正在生长的子细胞, 最后募集于母细胞与子细胞的连接部位(Zhang et al., 2005).复合体作为拴系子的直接证据来源于酵母异源表达实验, 当酵母SEC3亚基于线粒体膜异位表达时, 可观察到复合体其它亚基和囊泡在线粒体膜募集(Luo et al., 2014).该过程中SEC3蛋白CorEx基序发挥关键作用(Mei and Guo, 2018).这一作用特点表明胞泌复合体具有拴系子的功能. ...
The conserved oligomeric golgi complex is involved in penetration resistance of barley to the barley powdery mildew fungus 1 2013
... 目前, 对于植物胞泌复合体主要集中于遗传学研究, 细胞学研究尚处于起始阶段.已有的研究结果显示, 不同的胞泌复合体参与不同的生物学过程(表2).在拟南芥中, 胞泌复合体成员SEC3A、SEC6、SEC8、SEC15B和EXO70A1亚基与EXO84B共定位于细胞板, EXO84B突变导致胞质分裂缺陷, 细胞质中有大量囊泡累积, 表明植物胞泌复合体介导的囊泡运输参与细胞板的形成(Fendrych et al., 2010).玉米(Zea mays) SEC3同源物RTH1突变导致根毛变短, 显示RTH1参与根毛极性生长(Wen et al., 2005).拟南芥SEC3A突变使花粉不能向后代传递, 表明SEC3A的功能对花粉萌发和花粉管生长非常重要(Bloch et al., 2016).SEC5、SEC6、SEC8和SEC15A突变后同样显示出花粉萌发和花粉管生长缺陷(Hála et al., 2008).拟南芥和水稻EXO70A1的功能与导管细胞次生壁发育有关(Li et al., 2013; Tu et al., 2015).另据报道, 拟南芥EXO70A1突变植株生长素的运输和分布发生改变, 导致顶端优势丧失、根生长迟缓和侧根发生推迟等表型, 表明胞泌复合体可能参与PIN1和PIN2的再循环和极性定位(Drdová et al., 2013).最近有报道显示, EXO70A1参与下胚轴-根转化区(root- hypocotyl junction)的分化, 暗培养的exo70a1幼苗在靠近转化区的下胚轴表皮细胞形成类似根毛的结构, 推测这一表型与生长素信号紊乱有关(Drdová et al., 2019).EXO70C1在成熟花粉中高效表达, 其突变后会导致花粉管生长迟缓, 花粉的传递效率下降(Li et al., 2010).EXO70H4在表皮毛细胞壁加厚和胼胝质沉积过程中不可或缺(Kulich et al., 2015).大麦(Hordeum vulgare)基因沉默实验表明, EXO70F类似蛋白参与大麦抵抗真菌浸染的防御反应(Ostertag et al., 2013).研究发现EXO70I参与丛枝菌根共生过程(Zhang et al., 2015).BPH6蛋白可以与EXO70E1互作, 调控水稻细胞壁完整性, 抵御褐飞虱取食(Guo et al., 2018).这些生物学过程都与胞吐关系密切, 暗示植物胞泌复合体广泛参与与胞吐有关的生物学过程, 但其具体作用机制尚需阐明.植物胞泌复合体参与囊泡拴系过程的直接证据还不充分. ...
The role for the exocyst complex subunits Exo70B2 and Exo70H1 in the plant-pathogen interaction 2 2011
... 对植物胞泌复合体各亚基之间互作关系的了解, 有助于揭示其组装机制和调控特点.目前, 通过酵母双杂交、免疫共沉淀及BiFC等手段已明确拟南芥胞泌复合体成员间的互作关系(Hála et al., 2008; Pečenková et al., 2011; Zhang et al., 2013) (图2).利用遗传学研究已对部分蛋白之间的互作进行了功能验证, 其中, SEC8和EXO70A1共同参与种子外壁中的果胶沉积(Kulich et al., 2010); EXO70B1与SEC5A和EXO84B复合物参与自噬途径(Kulich et al., 2013); EXO70B2和EXO70H1共同参与病原体的响应, 其中EXO70B2与形成抗病相关的细胞壁附着物有关(Pečenková et al., 2011).以上结果表明, 植物胞泌复合体亚基在活体中以复合体的形式发挥作用. ...
... ); EXO70B2和EXO70H1共同参与病原体的响应, 其中EXO70B2与形成抗病相关的细胞壁附着物有关(Pečenková et al., 2011).以上结果表明, 植物胞泌复合体亚基在活体中以复合体的形式发挥作用. ...
Membrane targeting of the yeast exocyst complex 1 2015
... 对胞泌复合体调控机制的认识主要源于酵母和动物细胞.在酵母及动物细胞中, 对于胞泌复合体在靶膜的募集机制还存在分歧.一般认为, SEC3和EXO70不依赖于微丝骨架, 两者能够通过与PIP2及小GTPase定位到胞吐位置的质膜上(Pleskot et al., 2015; Liu et al., 2018), 其它6个亚基(SEC5、SEC6、SEC8、SEC10、SEC15和EXO84)形成亚复合体(subcomplex), 在小GTPase (SEC4)参与下募集于分泌囊泡, 并依赖微丝骨架运送到胞吐位点, 在其它蛋白作用下, 与膜上的SEC3和EXO70组装成完整的复合体, 从而将囊泡拴系到质膜上(Finger et al., 1998; Boyd et al., 2004) (图1A).对酵母的研究显示, SEC3与线粒体蛋白(Tom20p)融合蛋白于线粒体异源表达时, 能够介导胞泌复合体其它亚基在线粒体中的募集, 进而导致原本运输到质膜的囊泡向线粒体转运(Luo et al., 2014), 说明SEC3蛋白在复合体组装和囊泡拴系过程中发挥关键作用. ...
The Physcomitrella patens exocyst subunit EXO70.3d has distinct roles in growth and development, and is essential for completion of the moss life cycle 0 2017
Cellular pathways regula- ting responses to compatible and self-incompatible pollen in Brassica and Arabidopsis stigmas intersect at Exo70A1, a putative component of the exocyst complex 0 2009
Analysis of exocyst subunit EXO70 family reveals distinct membrane polar domains in tobacco pollen tubes 1 2017
Sec6p anchors the assembled exocyst complex at sites of secretion 1 2009
... 对胞泌复合体的研究始于酵母细胞, 该复合体最早发现于温度敏感型的酿酒酵母(Saccharomyces cerevisiae)突变体中(TerBush and Novick, 1995; TerBush et al., 1996).胞泌复合体由SEC3、SEC5、SEC6、SEC8、SEC10、SEC15、EXO70和EXO84八个蛋白亚基组成, 后期研究表明该复合体广泛存在于真核生物中(Martin-Urdiroz et al., 2016).酿酒酵母增殖时, 通过胞吐将子细胞形成相关材料运输到出芽部位, 复合体中的任何一个亚基突变都会导致分泌囊泡在出芽区域细胞质内积累, 而Myo2 (V型肌动蛋白)突变导致囊泡在母细胞的细胞质中累积, 说明复合体作用于囊泡运输的下游(Hsu et al., 2004; Munson and Novick, 2006; Zhang et al., 2008; Songer and Munson, 2009).活细胞观察显示复合体的8个亚基聚集在胞吐活跃部位: 出芽初期位于出芽部位, 随后分布于正在生长的子细胞, 最后募集于母细胞与子细胞的连接部位(Zhang et al., 2005).复合体作为拴系子的直接证据来源于酵母异源表达实验, 当酵母SEC3亚基于线粒体膜异位表达时, 可观察到复合体其它亚基和囊泡在线粒体膜募集(Luo et al., 2014).该过程中SEC3蛋白CorEx基序发挥关键作用(Mei and Guo, 2018).这一作用特点表明胞泌复合体具有拴系子的功能. ...
AtExo70A1, a member of a family of putative exocyst subunits specifically expanded in land plants, is important for polar growth and plant development 0 2006
The exocyst is a multiprotein complex required for exocytosis in 1 1996
... 对胞泌复合体的研究始于酵母细胞, 该复合体最早发现于温度敏感型的酿酒酵母(Saccharomyces cerevisiae)突变体中(TerBush and Novick, 1995; TerBush et al., 1996).胞泌复合体由SEC3、SEC5、SEC6、SEC8、SEC10、SEC15、EXO70和EXO84八个蛋白亚基组成, 后期研究表明该复合体广泛存在于真核生物中(Martin-Urdiroz et al., 2016).酿酒酵母增殖时, 通过胞吐将子细胞形成相关材料运输到出芽部位, 复合体中的任何一个亚基突变都会导致分泌囊泡在出芽区域细胞质内积累, 而Myo2 (V型肌动蛋白)突变导致囊泡在母细胞的细胞质中累积, 说明复合体作用于囊泡运输的下游(Hsu et al., 2004; Munson and Novick, 2006; Zhang et al., 2008; Songer and Munson, 2009).活细胞观察显示复合体的8个亚基聚集在胞吐活跃部位: 出芽初期位于出芽部位, 随后分布于正在生长的子细胞, 最后募集于母细胞与子细胞的连接部位(Zhang et al., 2005).复合体作为拴系子的直接证据来源于酵母异源表达实验, 当酵母SEC3亚基于线粒体膜异位表达时, 可观察到复合体其它亚基和囊泡在线粒体膜募集(Luo et al., 2014).该过程中SEC3蛋白CorEx基序发挥关键作用(Mei and Guo, 2018).这一作用特点表明胞泌复合体具有拴系子的功能. ...
Sec6, Sec8, and Sec15 are components of a multisubunit complex which localizes to small bud tips in Saccharomyces cerevisiae 1 1995
... 对胞泌复合体的研究始于酵母细胞, 该复合体最早发现于温度敏感型的酿酒酵母(Saccharomyces cerevisiae)突变体中(TerBush and Novick, 1995; TerBush et al., 1996).胞泌复合体由SEC3、SEC5、SEC6、SEC8、SEC10、SEC15、EXO70和EXO84八个蛋白亚基组成, 后期研究表明该复合体广泛存在于真核生物中(Martin-Urdiroz et al., 2016).酿酒酵母增殖时, 通过胞吐将子细胞形成相关材料运输到出芽部位, 复合体中的任何一个亚基突变都会导致分泌囊泡在出芽区域细胞质内积累, 而Myo2 (V型肌动蛋白)突变导致囊泡在母细胞的细胞质中累积, 说明复合体作用于囊泡运输的下游(Hsu et al., 2004; Munson and Novick, 2006; Zhang et al., 2008; Songer and Munson, 2009).活细胞观察显示复合体的8个亚基聚集在胞吐活跃部位: 出芽初期位于出芽部位, 随后分布于正在生长的子细胞, 最后募集于母细胞与子细胞的连接部位(Zhang et al., 2005).复合体作为拴系子的直接证据来源于酵母异源表达实验, 当酵母SEC3亚基于线粒体膜异位表达时, 可观察到复合体其它亚基和囊泡在线粒体膜募集(Luo et al., 2014).该过程中SEC3蛋白CorEx基序发挥关键作用(Mei and Guo, 2018).这一作用特点表明胞泌复合体具有拴系子的功能. ...
rSec6 and rSec8, mammalian homologs of yeast proteins essential for secretion 1 1995
... 随后, 胞泌复合体在哺乳动物大脑组织中被分离鉴定(Ting et al., 1995; Hsu et al., 1996).与酵母中的情况类似, 哺乳动物的8个亚基共沉淀于1个743 kDa的复合体(始称SEC6/SEC8, 后改为胞泌复合体)中.细胞学研究表明复合体定位于胞吐活跃的靶膜上(Grindstaff et al.,1998; Vega and Hsu, 2001; Tsuboi et al., 2005; Xiong et al., 2012), 说明胞泌复合体的功能在酵母和动物细胞中保守.除了胞吐过程, 胞泌复合体还参与多种细胞学过程, 如细胞极性建立、初级纤毛发生、胞质分裂、病原体入侵、肿瘤发生和转移(Heider and Munson, 2012; Martin-Urdiroz et al., 2016). ...
Mammalian exocyst complex is required for the docking step of insulin vesicle exocytosis 1 2005
... 随后, 胞泌复合体在哺乳动物大脑组织中被分离鉴定(Ting et al., 1995; Hsu et al., 1996).与酵母中的情况类似, 哺乳动物的8个亚基共沉淀于1个743 kDa的复合体(始称SEC6/SEC8, 后改为胞泌复合体)中.细胞学研究表明复合体定位于胞吐活跃的靶膜上(Grindstaff et al.,1998; Vega and Hsu, 2001; Tsuboi et al., 2005; Xiong et al., 2012), 说明胞泌复合体的功能在酵母和动物细胞中保守.除了胞吐过程, 胞泌复合体还参与多种细胞学过程, 如细胞极性建立、初级纤毛发生、胞质分裂、病原体入侵、肿瘤发生和转移(Heider and Munson, 2012; Martin-Urdiroz et al., 2016). ...
Disruption of OsEXO70A1 causes irregular vascular bundles and perturbs mineral nutrient assimilation in rice 1 2015
... 目前, 对于植物胞泌复合体主要集中于遗传学研究, 细胞学研究尚处于起始阶段.已有的研究结果显示, 不同的胞泌复合体参与不同的生物学过程(表2).在拟南芥中, 胞泌复合体成员SEC3A、SEC6、SEC8、SEC15B和EXO70A1亚基与EXO84B共定位于细胞板, EXO84B突变导致胞质分裂缺陷, 细胞质中有大量囊泡累积, 表明植物胞泌复合体介导的囊泡运输参与细胞板的形成(Fendrych et al., 2010).玉米(Zea mays) SEC3同源物RTH1突变导致根毛变短, 显示RTH1参与根毛极性生长(Wen et al., 2005).拟南芥SEC3A突变使花粉不能向后代传递, 表明SEC3A的功能对花粉萌发和花粉管生长非常重要(Bloch et al., 2016).SEC5、SEC6、SEC8和SEC15A突变后同样显示出花粉萌发和花粉管生长缺陷(Hála et al., 2008).拟南芥和水稻EXO70A1的功能与导管细胞次生壁发育有关(Li et al., 2013; Tu et al., 2015).另据报道, 拟南芥EXO70A1突变植株生长素的运输和分布发生改变, 导致顶端优势丧失、根生长迟缓和侧根发生推迟等表型, 表明胞泌复合体可能参与PIN1和PIN2的再循环和极性定位(Drdová et al., 2013).最近有报道显示, EXO70A1参与下胚轴-根转化区(root- hypocotyl junction)的分化, 暗培养的exo70a1幼苗在靠近转化区的下胚轴表皮细胞形成类似根毛的结构, 推测这一表型与生长素信号紊乱有关(Drdová et al., 2019).EXO70C1在成熟花粉中高效表达, 其突变后会导致花粉管生长迟缓, 花粉的传递效率下降(Li et al., 2010).EXO70H4在表皮毛细胞壁加厚和胼胝质沉积过程中不可或缺(Kulich et al., 2015).大麦(Hordeum vulgare)基因沉默实验表明, EXO70F类似蛋白参与大麦抵抗真菌浸染的防御反应(Ostertag et al., 2013).研究发现EXO70I参与丛枝菌根共生过程(Zhang et al., 2015).BPH6蛋白可以与EXO70E1互作, 调控水稻细胞壁完整性, 抵御褐飞虱取食(Guo et al., 2018).这些生物学过程都与胞吐关系密切, 暗示植物胞泌复合体广泛参与与胞吐有关的生物学过程, 但其具体作用机制尚需阐明.植物胞泌复合体参与囊泡拴系过程的直接证据还不充分. ...
The exocyst complex associates with microtubules to mediate vesicle targeting and neurite outgrowth 1 2001
... 随后, 胞泌复合体在哺乳动物大脑组织中被分离鉴定(Ting et al., 1995; Hsu et al., 1996).与酵母中的情况类似, 哺乳动物的8个亚基共沉淀于1个743 kDa的复合体(始称SEC6/SEC8, 后改为胞泌复合体)中.细胞学研究表明复合体定位于胞吐活跃的靶膜上(Grindstaff et al.,1998; Vega and Hsu, 2001; Tsuboi et al., 2005; Xiong et al., 2012), 说明胞泌复合体的功能在酵母和动物细胞中保守.除了胞吐过程, 胞泌复合体还参与多种细胞学过程, 如细胞极性建立、初级纤毛发生、胞质分裂、病原体入侵、肿瘤发生和转移(Heider and Munson, 2012; Martin-Urdiroz et al., 2016). ...
EXPO, an exocyst-positive organelle distinct from multivesicular endosomes and autophagosomes, mediates cytosol to cell wall exocytosis in Arabidopsis and tobacco cells 2 2010
... 一般认为胞泌复合体在经典囊泡运输途径(conventional secretory pathway)中发挥作用.但也有证据显示, 复合体亚基在经典分泌途径之外的细胞学过程中行使功能(Kulich et al., 2013; Ding et al., 2014; Wu et al., 2017).姜里文研究组利用拟南芥和烟草BY-2细胞表达(X)FP标记的EXO70蛋白同源物, 并结合免疫电镜观察, 发现了一个称之为EXPO (exocyst positive organelles)的结构(Wang et al., 2010).他们利用荧光蛋白标记的EXO70E2对EXPO进行鉴定, 发现EXPO定位于细胞质、细胞膜和胞外基质, 与高尔基体、反式高尔基体网络、早期内体、多泡体和晚期内体的标记物不能共定位, 说明该结构未参与经典囊泡运输途径(Wang et al., 2010).目前, 对于EXPO的功能还不清楚, 进一步的遗传学研究将有助于明确这一结构的生物学意义. ...
Expression and functional analysis of a novel group of legume-specific WRKY and EXO70 protein variants from soybean 0 2016
The roothairless1 gene of maize encodes a homolog of sec3, which is involved in polar exocytosis 1 2005
... 目前, 对于植物胞泌复合体主要集中于遗传学研究, 细胞学研究尚处于起始阶段.已有的研究结果显示, 不同的胞泌复合体参与不同的生物学过程(表2).在拟南芥中, 胞泌复合体成员SEC3A、SEC6、SEC8、SEC15B和EXO70A1亚基与EXO84B共定位于细胞板, EXO84B突变导致胞质分裂缺陷, 细胞质中有大量囊泡累积, 表明植物胞泌复合体介导的囊泡运输参与细胞板的形成(Fendrych et al., 2010).玉米(Zea mays) SEC3同源物RTH1突变导致根毛变短, 显示RTH1参与根毛极性生长(Wen et al., 2005).拟南芥SEC3A突变使花粉不能向后代传递, 表明SEC3A的功能对花粉萌发和花粉管生长非常重要(Bloch et al., 2016).SEC5、SEC6、SEC8和SEC15A突变后同样显示出花粉萌发和花粉管生长缺陷(Hála et al., 2008).拟南芥和水稻EXO70A1的功能与导管细胞次生壁发育有关(Li et al., 2013; Tu et al., 2015).另据报道, 拟南芥EXO70A1突变植株生长素的运输和分布发生改变, 导致顶端优势丧失、根生长迟缓和侧根发生推迟等表型, 表明胞泌复合体可能参与PIN1和PIN2的再循环和极性定位(Drdová et al., 2013).最近有报道显示, EXO70A1参与下胚轴-根转化区(root- hypocotyl junction)的分化, 暗培养的exo70a1幼苗在靠近转化区的下胚轴表皮细胞形成类似根毛的结构, 推测这一表型与生长素信号紊乱有关(Drdová et al., 2019).EXO70C1在成熟花粉中高效表达, 其突变后会导致花粉管生长迟缓, 花粉的传递效率下降(Li et al., 2010).EXO70H4在表皮毛细胞壁加厚和胼胝质沉积过程中不可或缺(Kulich et al., 2015).大麦(Hordeum vulgare)基因沉默实验表明, EXO70F类似蛋白参与大麦抵抗真菌浸染的防御反应(Ostertag et al., 2013).研究发现EXO70I参与丛枝菌根共生过程(Zhang et al., 2015).BPH6蛋白可以与EXO70E1互作, 调控水稻细胞壁完整性, 抵御褐飞虱取食(Guo et al., 2018).这些生物学过程都与胞吐关系密切, 暗示植物胞泌复合体广泛参与与胞吐有关的生物学过程, 但其具体作用机制尚需阐明.植物胞泌复合体参与囊泡拴系过程的直接证据还不充分. ...
Arabidopsis EXO70A1 recruits Patellin3 to the cell membrane independent of its role as an exocyst subunit 1 2017
... 一般认为胞泌复合体在经典囊泡运输途径(conventional secretory pathway)中发挥作用.但也有证据显示, 复合体亚基在经典分泌途径之外的细胞学过程中行使功能(Kulich et al., 2013; Ding et al., 2014; Wu et al., 2017).姜里文研究组利用拟南芥和烟草BY-2细胞表达(X)FP标记的EXO70蛋白同源物, 并结合免疫电镜观察, 发现了一个称之为EXPO (exocyst positive organelles)的结构(Wang et al., 2010).他们利用荧光蛋白标记的EXO70E2对EXPO进行鉴定, 发现EXPO定位于细胞质、细胞膜和胞外基质, 与高尔基体、反式高尔基体网络、早期内体、多泡体和晚期内体的标记物不能共定位, 说明该结构未参与经典囊泡运输途径(Wang et al., 2010).目前, 对于EXPO的功能还不清楚, 进一步的遗传学研究将有助于明确这一结构的生物学意义. ...
The Exo70 subunit of the exocyst is an effector for both Cdc42 and Rho3 function in polarized exocytosis 1 2010
... 越来越多的实验证据表明, 胞泌复合体的功能受到Ras-GTP蛋白家族成员的调控, 并与SNARE蛋白发生关联, 调控下游的膜融合事件(Heider and Munson, 2012).质膜上的SEC3可以与小GTPase Cdc42和Rho1互作, 参与胞泌复合体在靶膜的组装(Guo et al., 2001; Zhang et al., 2001).EXO70也可与Cdc42和Rho3互作, 介导分泌囊泡与质膜的融合(Adamo et al., 1999; Wu et al., 2010).SEC3和EXO70之间的功能关系尚不明确.SEC15通过与分泌囊泡相关的Rab GTPase (SEC4)相互作用被招募到分泌囊泡(Wu et al., 2005), 同时在囊泡与质膜融合后促进马达蛋白的释放(Donovan and Bretscher, 2015).酵母双杂交实验结果显示, 在哺乳动物细胞中SEC5可以与Ras家族成员RalA-GTP相互作用, 抑制RalA-GTP的活性并影响胞泌复合体的组装, 说明RalA通过SEC5调控复合体的形成(Moskalenko et al., 2002) (图1A). ...
Regulation of cytokinesis by exocyst subunit SEC6 and KEULE in Arabidopsis thaliana 1 2013
Sec15 interacts with Rab11 via a novel domain and affects Rab11 localization in vivo 1 2005
... 越来越多的实验证据表明, 胞泌复合体的功能受到Ras-GTP蛋白家族成员的调控, 并与SNARE蛋白发生关联, 调控下游的膜融合事件(Heider and Munson, 2012).质膜上的SEC3可以与小GTPase Cdc42和Rho1互作, 参与胞泌复合体在靶膜的组装(Guo et al., 2001; Zhang et al., 2001).EXO70也可与Cdc42和Rho3互作, 介导分泌囊泡与质膜的融合(Adamo et al., 1999; Wu et al., 2010).SEC3和EXO70之间的功能关系尚不明确.SEC15通过与分泌囊泡相关的Rab GTPase (SEC4)相互作用被招募到分泌囊泡(Wu et al., 2005), 同时在囊泡与质膜融合后促进马达蛋白的释放(Donovan and Bretscher, 2015).酵母双杂交实验结果显示, 在哺乳动物细胞中SEC5可以与Ras家族成员RalA-GTP相互作用, 抑制RalA-GTP的活性并影响胞泌复合体的组装, 说明RalA通过SEC5调控复合体的形成(Moskalenko et al., 2002) (图1A). ...
An association between type Iγ PI4P 5-kinase and Exo70 directs E-cadherin clustering and epithelial polarization 1 2012
... 随后, 胞泌复合体在哺乳动物大脑组织中被分离鉴定(Ting et al., 1995; Hsu et al., 1996).与酵母中的情况类似, 哺乳动物的8个亚基共沉淀于1个743 kDa的复合体(始称SEC6/SEC8, 后改为胞泌复合体)中.细胞学研究表明复合体定位于胞吐活跃的靶膜上(Grindstaff et al.,1998; Vega and Hsu, 2001; Tsuboi et al., 2005; Xiong et al., 2012), 说明胞泌复合体的功能在酵母和动物细胞中保守.除了胞吐过程, 胞泌复合体还参与多种细胞学过程, 如细胞极性建立、初级纤毛发生、胞质分裂、病原体入侵、肿瘤发生和转移(Heider and Munson, 2012; Martin-Urdiroz et al., 2016). ...
Tethering factors as organizers of intracellular vesicular traffic 1 2010
... 囊泡运输是所有真核生物中都存在的一种非常重要的细胞学过程, 负责细胞内膜系统之间及内膜系统与细胞膜之间的物质交流.囊泡运输过程主要包括4个环节: (1) 供体膜出芽形成运输囊泡; (2) 运输囊泡沿细胞骨架转运到靶膜附近; (3) 运输囊泡拴系到靶膜上; (4) SNARE (soluble N-ethyl-maleimide- sensitive fusion protein attachment protein receptor)复合体介导运输囊泡与靶膜发生融合(Bonifacino and Glick, 2004; Cai et al., 2007).其中, 拴系过程介导运输囊泡与靶膜在空间上的最初接触, 在时空上调控下游膜融合事件的发生, 是调控极性囊泡运输的关键环节.研究表明, 一些蛋白复合体参与囊泡运输的拴系过程, 被称为拴系子(tethers).拴系子包括两大类: 一类是卷曲螺线蛋白(coiled-coil proteins), 通常以二聚体形式存在, 被称为拴系蛋白; 另一类是多亚基的蛋白复合体(multisubunit tethering complexes, MTCs), 即拴系复合体.目前, 在真核生物中鉴定了9种拴系复合体, 分别是TRAPP I、TRAPP II、TRAPP III、COG、HOPS、CORVET、Dsl1、GARP和胞泌复合体(exocyst).严格意义上讲, 拴系子需要同时与源于供体膜的囊泡和靶膜上的蛋白及膜脂互作, 介导两膜在物理空间上的直接接触.目前, 除了少数MTCs和卷曲螺线拴系蛋白外, 对于多数拴系子仍然缺乏直接的实验证据支持(Brunet and Sacher, 2014).因此, 有****建议将至今还未明确拴系功能的复合体称为拴系因子(tethering factors).与SNARE蛋白相比, MTCs的一个特点是其在细胞内具有时空特异性, 即不同MTCs作用于囊泡运输过程中不同目的膜.例如, DsI1位于内质网, COG在高尔基体, GARP作用于早期内体的返回途径, HOPS和CORVET分别在晚期内体到溶酶体以及内体之间的囊泡运输中发挥作用, TRAPP I/II/III存在于多种不同的囊泡运输途径, 胞泌复合体介导高尔基后囊泡向质膜的拴系过程(Yu and Hughson, 2010). ...
Sec3 promotes the initial binary t-SNARE complex assembly and membrane fusion 1 2017
... 拴系和膜融合事件在时序上前后紧密相连, 胞泌复合体和SNARE复合体之间存在功能关联, 二者协同作用可保证在时空上实现胞吐的精确控制.生化和遗传证据证明, 胞泌复合体能够直接帮助SNARE复合体组装并参与膜融合事件的发生(Hong and Lev, 2014).体外实验表明, 胞泌复合体亚基可与3种SNARE蛋白、SNARE复合体及其调控蛋白SEC1结合.酵母SEC3的N端结构域与t-SNARE Sso1/2结合并促进其与t-SNARE SEC9形成二聚体(Yue et al., 2017).SEC9与v-SNARE Snc1/2的互作有利于稳定囊泡与胞泌复合体的结合(Shen et al., 2013).另有研究表明, SEC6与t-SNARE (Sec9)互作促进SNARE复合体的组装、纠正错配的SNARE复合体以及激活膜融合发生(Dubuke et al., 2015).虽然目前尚不明确胞泌复合体与SNARE复合体的作用细节, 但基于现有的证据推测胞泌复合体首先与囊泡上的Rab GTPase、肌球蛋白(myosin)及v-SNARE互作, 启动胞泌复合体构象的改变, 暴露其与质膜和t-SNARE蛋白的作用位点, 从而驱动SNARE复合体的组装, 进而介导囊泡膜与靶膜的融合(图1B). ...
Exocytosis and cell polarity in plants exocyst and recycling domains 1 2009
... 目前, 植物中编码胞泌复合体的8个亚基的同源基因已被鉴定(Elias et al., 2003).遗传学、生物化学和细胞学证据显示, 植物胞泌复合体也是以异源八聚体的形式参与胞吐相关的细胞学过程(Hála et al., 2008; Zarsky et al., 2009).但是, 目前尚未获得植物胞泌复合体作为拴系子的直接证据. ...
EXO70I is required for development of a sub-domain of the periarbuscular membrane during arbuscular mycorrhizal symbiosis 1 2015
... 目前, 对于植物胞泌复合体主要集中于遗传学研究, 细胞学研究尚处于起始阶段.已有的研究结果显示, 不同的胞泌复合体参与不同的生物学过程(表2).在拟南芥中, 胞泌复合体成员SEC3A、SEC6、SEC8、SEC15B和EXO70A1亚基与EXO84B共定位于细胞板, EXO84B突变导致胞质分裂缺陷, 细胞质中有大量囊泡累积, 表明植物胞泌复合体介导的囊泡运输参与细胞板的形成(Fendrych et al., 2010).玉米(Zea mays) SEC3同源物RTH1突变导致根毛变短, 显示RTH1参与根毛极性生长(Wen et al., 2005).拟南芥SEC3A突变使花粉不能向后代传递, 表明SEC3A的功能对花粉萌发和花粉管生长非常重要(Bloch et al., 2016).SEC5、SEC6、SEC8和SEC15A突变后同样显示出花粉萌发和花粉管生长缺陷(Hála et al., 2008).拟南芥和水稻EXO70A1的功能与导管细胞次生壁发育有关(Li et al., 2013; Tu et al., 2015).另据报道, 拟南芥EXO70A1突变植株生长素的运输和分布发生改变, 导致顶端优势丧失、根生长迟缓和侧根发生推迟等表型, 表明胞泌复合体可能参与PIN1和PIN2的再循环和极性定位(Drdová et al., 2013).最近有报道显示, EXO70A1参与下胚轴-根转化区(root- hypocotyl junction)的分化, 暗培养的exo70a1幼苗在靠近转化区的下胚轴表皮细胞形成类似根毛的结构, 推测这一表型与生长素信号紊乱有关(Drdová et al., 2019).EXO70C1在成熟花粉中高效表达, 其突变后会导致花粉管生长迟缓, 花粉的传递效率下降(Li et al., 2010).EXO70H4在表皮毛细胞壁加厚和胼胝质沉积过程中不可或缺(Kulich et al., 2015).大麦(Hordeum vulgare)基因沉默实验表明, EXO70F类似蛋白参与大麦抵抗真菌浸染的防御反应(Ostertag et al., 2013).研究发现EXO70I参与丛枝菌根共生过程(Zhang et al., 2015).BPH6蛋白可以与EXO70E1互作, 调控水稻细胞壁完整性, 抵御褐飞虱取食(Guo et al., 2018).这些生物学过程都与胞吐关系密切, 暗示植物胞泌复合体广泛参与与胞吐有关的生物学过程, 但其具体作用机制尚需阐明.植物胞泌复合体参与囊泡拴系过程的直接证据还不充分. ...
Cdc42 interacts with the exocyst and regulates polarized secretion 1 2001
... 越来越多的实验证据表明, 胞泌复合体的功能受到Ras-GTP蛋白家族成员的调控, 并与SNARE蛋白发生关联, 调控下游的膜融合事件(Heider and Munson, 2012).质膜上的SEC3可以与小GTPase Cdc42和Rho1互作, 参与胞泌复合体在靶膜的组装(Guo et al., 2001; Zhang et al., 2001).EXO70也可与Cdc42和Rho3互作, 介导分泌囊泡与质膜的融合(Adamo et al., 1999; Wu et al., 2010).SEC3和EXO70之间的功能关系尚不明确.SEC15通过与分泌囊泡相关的Rab GTPase (SEC4)相互作用被招募到分泌囊泡(Wu et al., 2005), 同时在囊泡与质膜融合后促进马达蛋白的释放(Donovan and Bretscher, 2015).酵母双杂交实验结果显示, 在哺乳动物细胞中SEC5可以与Ras家族成员RalA-GTP相互作用, 抑制RalA-GTP的活性并影响胞泌复合体的组装, 说明RalA通过SEC5调控复合体的形成(Moskalenko et al., 2002) (图1A). ...
Membrane association and functional regulation of Sec3 by phospholipids and Cdc42 1 2008
... 对胞泌复合体的研究始于酵母细胞, 该复合体最早发现于温度敏感型的酿酒酵母(Saccharomyces cerevisiae)突变体中(TerBush and Novick, 1995; TerBush et al., 1996).胞泌复合体由SEC3、SEC5、SEC6、SEC8、SEC10、SEC15、EXO70和EXO84八个蛋白亚基组成, 后期研究表明该复合体广泛存在于真核生物中(Martin-Urdiroz et al., 2016).酿酒酵母增殖时, 通过胞吐将子细胞形成相关材料运输到出芽部位, 复合体中的任何一个亚基突变都会导致分泌囊泡在出芽区域细胞质内积累, 而Myo2 (V型肌动蛋白)突变导致囊泡在母细胞的细胞质中累积, 说明复合体作用于囊泡运输的下游(Hsu et al., 2004; Munson and Novick, 2006; Zhang et al., 2008; Songer and Munson, 2009).活细胞观察显示复合体的8个亚基聚集在胞吐活跃部位: 出芽初期位于出芽部位, 随后分布于正在生长的子细胞, 最后募集于母细胞与子细胞的连接部位(Zhang et al., 2005).复合体作为拴系子的直接证据来源于酵母异源表达实验, 当酵母SEC3亚基于线粒体膜异位表达时, 可观察到复合体其它亚基和囊泡在线粒体膜募集(Luo et al., 2014).该过程中SEC3蛋白CorEx基序发挥关键作用(Mei and Guo, 2018).这一作用特点表明胞泌复合体具有拴系子的功能. ...
The critical role of Exo84p in the organization and polarized localization of the exocyst complex 1 2005
... 对胞泌复合体的研究始于酵母细胞, 该复合体最早发现于温度敏感型的酿酒酵母(Saccharomyces cerevisiae)突变体中(TerBush and Novick, 1995; TerBush et al., 1996).胞泌复合体由SEC3、SEC5、SEC6、SEC8、SEC10、SEC15、EXO70和EXO84八个蛋白亚基组成, 后期研究表明该复合体广泛存在于真核生物中(Martin-Urdiroz et al., 2016).酿酒酵母增殖时, 通过胞吐将子细胞形成相关材料运输到出芽部位, 复合体中的任何一个亚基突变都会导致分泌囊泡在出芽区域细胞质内积累, 而Myo2 (V型肌动蛋白)突变导致囊泡在母细胞的细胞质中累积, 说明复合体作用于囊泡运输的下游(Hsu et al., 2004; Munson and Novick, 2006; Zhang et al., 2008; Songer and Munson, 2009).活细胞观察显示复合体的8个亚基聚集在胞吐活跃部位: 出芽初期位于出芽部位, 随后分布于正在生长的子细胞, 最后募集于母细胞与子细胞的连接部位(Zhang et al., 2005).复合体作为拴系子的直接证据来源于酵母异源表达实验, 当酵母SEC3亚基于线粒体膜异位表达时, 可观察到复合体其它亚基和囊泡在线粒体膜募集(Luo et al., 2014).该过程中SEC3蛋白CorEx基序发挥关键作用(Mei and Guo, 2018).这一作用特点表明胞泌复合体具有拴系子的功能. ...
The Arabidopsis exocyst subunit Sec3A is essential for embryo development and accumulates in transient puncta at the plasma membrane 1 2013
... 对植物胞泌复合体各亚基之间互作关系的了解, 有助于揭示其组装机制和调控特点.目前, 通过酵母双杂交、免疫共沉淀及BiFC等手段已明确拟南芥胞泌复合体成员间的互作关系(Hála et al., 2008; Pečenková et al., 2011; Zhang et al., 2013) (图2).利用遗传学研究已对部分蛋白之间的互作进行了功能验证, 其中, SEC8和EXO70A1共同参与种子外壁中的果胶沉积(Kulich et al., 2010); EXO70B1与SEC5A和EXO84B复合物参与自噬途径(Kulich et al., 2013); EXO70B2和EXO70H1共同参与病原体的响应, 其中EXO70B2与形成抗病相关的细胞壁附着物有关(Pečenková et al., 2011).以上结果表明, 植物胞泌复合体亚基在活体中以复合体的形式发挥作用. ...
CSI1, PATROL1, and exocyst complex cooperate in delivery of cellulose synthase complexes to the plasma membrane 0 2018
 ,2,*1.
,2,*1. 
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT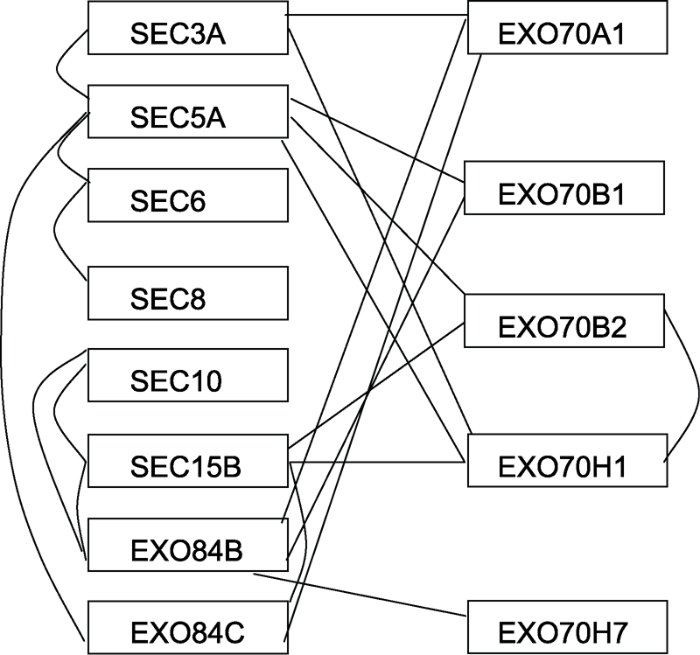 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}