Chloroplast Genome Structural Characteristics and Phylogenetic Relationships of Oleaceae
Yuemei Zhao1, Zhenyan Yang2, Yongping Zhao1, Xiaoling Li1, Zhixin Zhao1, Guifang Zhao,3,*1 College of Biopharmaceutical and Food Engineering, Shangluo University, Shangluo 726000, China 2 Key Laboratory for Plant Diversity and Biogeography of East Asia, Kunming Institute of Botany, Chinese Academy of Sciences, Kunming 650201, China 3 Key Laboratory of Resource Biology and Biotechnology in Western China, Ministry of Education, College of Life Sciences, Northwest University, Xi’an 710069, China
Abstract In this study, the chloroplast genomes from 19 species (11 genera) in Oleaceae were compared to reveal the general characteristics and structural variations. The chloroplast genome sizes in Oleaceae were 154-165 kb, and the differences were mainly caused by the length of large single-copy regions. The chloroplast genome sizes of 3 species from the genus Jasminum differed greatly from that for other species; in addition, the introns from the clpP and accD genes were lost in Jasminum. Synteny analyses showed several gene rearrangements in 3 Jasminum species that were probably caused by inversions. The boundary genes between IRb/small single copy (SSC) and SSC/IRa regions in 3 Jasminum species differed from others. Repeat sequences and simple sequence repeat detection demonstrated that Jasminum had significant differences in repeat number and repeat length as compared with other genera. On the basis of shared protein-coding genes among 19 species, Abeliophyllum distichum and Forsythia suspensa were the early-diverging clades in Oleaceae. Keywords:Oleaceae;chloroplast genomes;structural characteristics;phylogeny
PDF (15099KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 赵月梅, 杨振艳, 赵永平, 李筱玲, 赵志新, 赵桂仿. 木犀科植物叶绿体基因组结构特征和系统发育关系. 植物学报, 2019, 54(4): 441-454 doi:10.11983/CBB18191 Zhao Yuemei, Yang Zhenyan, Zhao Yongping, Li Xiaoling, Zhao Zhixin, Zhao Guifang. Chloroplast Genome Structural Characteristics and Phylogenetic Relationships of Oleaceae. Chinese Bulletin of Botany, 2019, 54(4): 441-454 doi:10.11983/CBB18191
叶绿体是绿色植物进行光合作用的细胞器, 具有合成蛋白质、淀粉及色素等功能, 其基因组可自主遗传, 在植物漫长的进化历史中起重要作用。在被子植物中, 多数叶绿体基因组(cpDNA)为母系遗传, 少数为双亲遗传或父系遗传(Hu et al., 2008)。叶绿体基因组结构和序列都比较保守, 大部分基因组结构为双链环状结构, 包括2个反向重复区(inverted repeats region, IRa/IRb)、1个大单拷贝区(large single copy region, LSC)和1个小单拷贝区(small single copy region, SSC) 4部分, 大小一般在120-180 kb之间(Zhang et al., 2012)。虽然与核基因组和线粒体基因组相比, cpDNA在基因组成和结构上相对保守, 但近年的研究也表明许多cpDNA发生基因变异, 如基因或内含子片段的丢失(Gao et al., 2010; 唐萍等, 2011)、反向重复区的长度变异或部分片段的插入/缺失(Chumley et al., 2006)、整个反向重复区的扩张或缺失(Chumley et al., 2006)及基因重排(Guisinger et al., 2011; Blazier et al., 2016)。由于基因组较小且结构相对保守, cpDNA已经成为进化和比较基因组研究的理想模型(Dong et al., 2013), 为揭示植物类群的系统位置和遗传发育关系提供更多有利依据。
木犀科植物多为乔木, 直立或藤状灌木, 属于被子植物中合瓣花亚纲的木犀亚目, 包含28属400余种, 广泛分布于温带和热带, 我国有10属160余种(Flora of China Editorial Committee, 1996)。该科的茉莉(Jasminum sambac)、流苏(Chionanthus retusus)和迎春花(J. nudiflorum)等均是重要的园林植物; 连翘(Forsythia suspensa)则是重要的药用植物, 其果具清热解毒、消肿散结等功效。近年来, 随着分子系统学的发展, 有关木犀科的系统发育研究也有所增加(Wallander and Albert, 2000), 但多数研究仅基于核基因ITS或叶绿体基因片段, 且这些研究仅推断出木犀科部分属(如Fraxinus、Syringa、Ligustrum和Olea) (Jeandroz et al., 1997; Li et al., 2002; Wallander, 2008; Besnard et al., 2009)或种间的系统发育关系(Besnard et al., 2002; Baali cherif and Besnard, 2005)。随着二代测序技术的不断发展, 有关木犀科叶绿体基因组方面的研究也逐渐兴起。Lee等(2007)主要分析了Jasminum和Menodora结构重排及其形成原因, 内容上更侧重于叶绿体基因组的结构变异。时至今日, 数据库中木犀科植物叶绿体基因组数据在不断更新, 但尚未见对该科叶绿体基因组结构及进化关系的系统研究。本研究在数据库共享信息不断增加的基础上, 拟通过下载整理数据库中木犀科19个物种的序列, 从叶绿体基因组的角度分析这19个物种的种间关系, 并通过比较这19个物种(11个属)的叶绿体基因组数据, 揭示该科植物叶绿体基因组的结构特征和变异情况。
Table 2 表2 表2叶绿体基因组中含内含子蛋白编码基因的编码区长度和基因全长 Table 2Length of coding region and complete gene of intron-contained protein-coding genes of chloroplast genomes
Species
petD
rps12
clpP
rpoC1
rps16
1
483/1196
372/908
591/2047
2064/2821
267/1132
2
483/1218
372/908
591/2043
2073/2834
267/1147
3
483/1230
375/911
591/2045
2052/2820
267/1134
4
483/1261
381/917
591/2043
2073/2827
267/1153
5
483/1134
372/908
591/2045
2052/2811
255/1147
6
483/1148
372/908
591/2053
2073/2830
267/1136
7
483/1217
372/908
591/2039
2073/2834
267/1142
8
483/1196
372/908
588/2045
2064/2822
267/1131
9
483/1196
381/917
660
2076/2844
267/1162
10
483/1203
387/923
786
2052/2808
267/1154
11
483/1213
387/923
786
2052/2807
267/1161
12
483/1215
381/917
591/2041
2073/2833
267/1143
13
483/1217
372/908
591/2043
2073/2830
267/1143
14
483/1215
372/908
591/2046
2073/2832
267/1141
15
483/1213
371/907
591/2043
2073/2831
267/1142
16
483/1213
372/908
591/2044
2073/2831
267/1142
17
483/1215
373/909
591/2041
2073/2833
267/1141
18
483/1203
372/908
591/2047
2073/2834
267/1147
19
483/1199
372/913
591/2041
2073/2784
237/1115
物种编号1-19同表1。表中/前数字表示编码基因长度(bp), /后数字表示基因全长(bp)。 The species number 1-19 same as Table 1. The number before / indicates the length of coding region (bp), and the number after / indicates the full length of gene (bp).
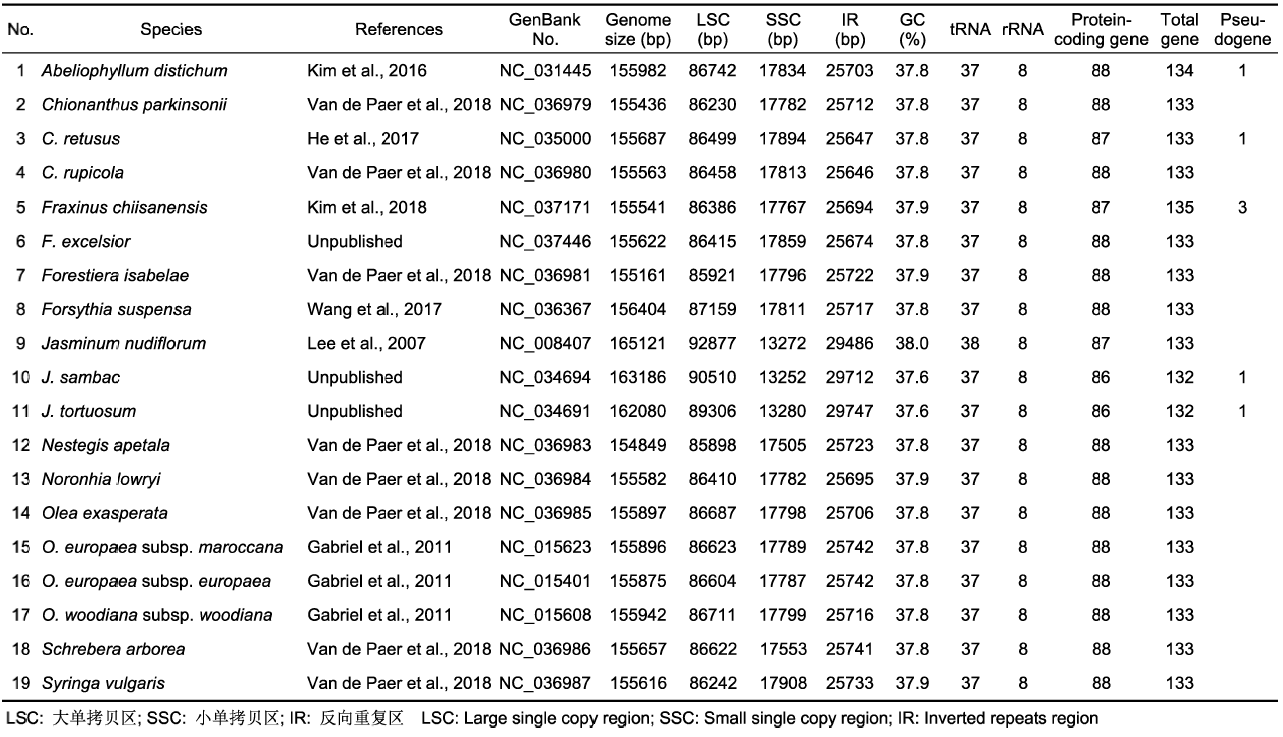
(A) 3种重复类型的数目; (B) 3种重复类型的比例; (C) SSR类型的比例; (D) SSR的数目和类型 Figure 1The type and percentage of repeated sequences in the chloroplast genomes of 19 species in Oleaceae
(A) Number of three type repeats; (B) Percentage of three type repeats; (C) Percentage of SSR types; (D) Number and types of SSRs
物种编号1-19同表1。以Abeliophyllum distichum叶绿体基因组序列为参考序列(x轴), 各物种叶绿体基因组与参考序列一致度范围为50%-100%(y轴)。箭头表示基因及转录方向。 Figure 3Graphic view of the alignment of chloroplast genomes from 19 species in Oleaceae
The species number 1-19 same as Table 1. Sequence identity varying between 50% and 100% are drawn on the y axis of the plot, the x axis corresponds to the coordinates on the Abeliophyllum distichum chloroplast genome. Arrows indicate the annotated genes and their transcriptional direction.
每个节点附近的数字分别表示ML树的自展支持率和BI树的后验概率。以Cornus controversa、Echites umbellatus和Catharanthus roseus为外类群。ML: 最大似然法; BI: 贝叶斯法 Figure 5Phylogenetic relationship of 19 species in Oleaceae inferred from ML and BI analyses based on shared protein-coding genes
The numbers near each node are bootstrap support values in ML and posterior probability in BI. The outgroups are Cornus controversa, Echites umbellatus and Catharanthus roseus. ML: Maximum likelihood; BI: Bayesian inference
3 讨论
被子植物的叶绿体基因组大小为120-180 kb, IR区长度为20-30 kb (Zhang et al., 2012)。19个(11属)木犀科物种的叶绿体基因组大小为154-165 kb, IR区长25-29 kb, 与被子植物叶绿体基因组长度相吻合。本研究表明, 19个基因组大小差异约为10 kb, LSC、SSC与IR各区域的差异为LSC (7 kb)>SSC (4.7 kb)>IR (4 kb), 可见木犀科不同种间基因组长度的差异主要是LSC的差异。研究表明, 反向互补区可增强叶绿体基因组的稳定性和保守性(Khakhlova and Bock, 2006)。本研究中木犀科11属19种植物, 除Jasminum属3个种外, 其余16个种的叶绿体基因组IR区差异均很小, 未发现IR区域过长、过短或丢失(Chumley et al., 2006; Hirao et al., 2008; Guisinger et al., 2011), 整体较保守且一致。
Baali-CherifD, BesnardG ( 2005). High genetic diversity and clonal growth in relict populations of Olea europaea subsp. laperrinei ( Oleaceae) from Hoggar, Algeria Ann Bot 96, 823-830. [本文引用: 1]
BensonG ( 1999). Tandem repeats finder: a program to analyze DNA sequences Nucleic Acids Res 27, 573-580. [本文引用: 1]
BesnardG, de CasasRR, ChristinPA, VargasP ( 2009). Phylogenetics of Olea( Oleaceae) based on plastid and nuclear ribosomal DNA sequences: tertiary climatic shifts and lineage differentiation times Ann Bot 104, 143-160. [本文引用: 1]
BesnardG, HernándezP, KhadariB, DoradoG, SavolainenV ( 2011). Genomic profiling of plastid DNA variation in the mediterranean olive tree BMC Plant Biol 11, 80.
BesnardG, KhadariB, BaradatP, BervilléA ( 2002). Olea europaea( Oleaceae) phylogeography based on chloroplast DNA polymorphism Theor Appl Genet 104, 1353-1361. [本文引用: 1]
BlazierJC, RuhlmanTA, WengML, RehmanSK, SabirJSM, JansenRK ( 2016). Divergence of RNA polymerase α subunits in angiosperm plastid genomes is mediated by genomic rearrangement Sci Rep 6, 24595. [本文引用: 1]
ChumleyTW, PalmerJD, MowerJP, FourcadeHM, CaliePJ, BooreJL, JansenRK ( 2006). The complete chloroplast genome sequence of Pelargonium x hortorum: organization and evolution of the largest and most highly rearranged chloroplast genome of land plants Mol Biol Evol 23, 2175-2190. [本文引用: 3]
DarlingACE, MauB, BlattnerFR, PernaANT ( 2004). Mauve: multiple alignment of conserved genomic sequence with rearrangements Genome Res 14, 1394-1403. [本文引用: 1]
DongWP, XuC, ChengT, LinK, ZhouSL ( 2013). Sequencing angiosperm plastid genomes made easy: a complete set of universal primers and a case study on the phylogeny of Saxifragales Genome Biol Evol 5, 989-997. [本文引用: 1]
Flora of China Editorial Committee ( 1995). Flora of China, Vol. 16. Beijing & St. Louis: Science Press & Missouri Botanical Garden Press. pp. 143-188. [本文引用: 2]
Flora of China Editorial Committee ( 1996). Flora of China, Vol. 15. Beijing & St. Louis: Science Press & Missouri Botanical Garden Press. pp. 272-319. [本文引用: 2]
FrazerKA, PachterL, PoliakovA, RubinEM, DubchakI ( 2004). Vista: computational tools for comparative genomics Nucleic Acids Res 32, W273-W279. [本文引用: 1]
GaoL, SuYJ, WangT ( 2010). Plastid genome sequencing, comparative genomics, and phylogenomics: current status and prospects J Syst Evol 48, 77-93. [本文引用: 1]
GrayBN, AhnerBA, HansonMR ( 2009). Extensive homologous recombination between introduced and native regulatory plastid DNA elements in transplastomic plants Transgenic Res 18, 559-572. [本文引用: 3]
GreinerS, WangX, RauwolfU, SilberMV, MayerK, MeurerJ, HabererG, HerrmannRG ( 2008). The complete nucleotide sequences of the five genetically distinct plastid genomes of Oenothera, subsection Oenothera. I. Sequence evaluation and plastome evolution Nucleic Acids Res 36, 2366-2378. [本文引用: 1]
GuisingerMM, KuehlJV, BooreJL, JansenRK ( 2011). Extreme reconfiguration of plastid genomes in the angiosperm family Geraniaceae: rearrangements, repeats, and codon usage Mol Biol Evol 28, 583-600.
HansenDR, DastidarSG, CaiZQ, PenaflorC, KuehlJV, BooreJL, JansenRK ( 2007). Phylogenetic and evolutionary implications of complete chloroplast genome sequences of four early-diverging angiosperms: Buxus( Buxaceae), Chloranthus 45, 547-563. [本文引用: 1]
HeYX, LiuLX, YangSH, DongMF, YuanWJ, ShangFD ( 2017). Characterization of the complete chloroplast genome of chinese fringetree (Chionanthus retusus) Conserv Genet Resour 9, 431-434. [本文引用: 1]
HiraoT, WatanabeA, KuritaM, KondoT, TakataK ( 2008). Complete nucleotide sequence of the Cryptomeria japonica D. Don. chloroplast genome and comparative chloroplast genomics: diversified genomic structure of coniferous species BMC Plant Biol 8, 70. [本文引用: 1]
HiratsukaJ, ShimadaH, WhittierR, IshibashiT, SakamotoM, MoriM, KondoC, HonjiY, SunCR, MengBY, LiYQ, KannoA, NishizawaY, HiraiA, ShinozakiK, SugiuraM ( 1989). The complete sequence of the rice (Oryza sativa) chloroplast genome: intermolecular recombination between distinct tRNA genes accounts for a major plastid DNA inversion during the evolution of the cereals Mol Gen Genet 217, 185-194. [本文引用: 1]
HuYC, ZhangQ, RaoGY , Sodmergen ( 2008). Occurrence of plastids in the sperm cells of Caprifoliaceae: biparental plastid inheritance in angiosperms is unilaterally derived from maternal inheritance Plant Cell Physiol 49, 958-968. [本文引用: 1]
JeandrozS, RoyA, BousquetJ ( 1997). Phylogeny and phylogeography of the circumpolar genus Fraxinus( Oleaceae) based on internal transcribed spacer sequences of nuclear ribosomal DNA Mol Phylogenet Evol 7, 241-251. [本文引用: 1]
KatayamaH, OgiharaY ( 1996). Phylogenetic affinities of the grasses to other monocots as revealed by molecular analysis of chloroplast DNA Curr Genet 29, 572-581.
KhakhlovaO, BockR ( 2006). Elimination of deleterious mutations in plastid genomes by gene conversion Plant J 46, 85-94.
KimC, KimHJ, DoHDK, JungJ, KimJH ( 2018). Characterization of the complete chloroplast genome of Fraxinus chiisanensis( Oleaceae), an endemic to Korea Conserv Genet Resour 11, 63-66. [本文引用: 1]
KimHW, LeeHL, LeeDK, KimKJ ( 2016). Complete plastid genome sequences of Abeliophyllum distichum Nakai (Oleaceae), a Korea endemic genus Mitochondr DNA Part B 1, 596-598. [本文引用: 1]
KnoxEB, DownieSR, PalmerJD ( 1993). Chloroplast genome rearrangements and the evolution of giant lobelias from herbaceous ancestors Mol Biol Evol 10, 414-430. [本文引用: 4]
KurtzS, ChoudhuriJV, OhlebuschE, SchleiermacherC, StoyeJ, GiegerichR ( 2001). Reputer: the manifold applications of repeat analysis on a genomic scale Nucleic Acids Res 29, 4633-4642. [本文引用: 1]
LeeHL, JansenRK, ChumleyTW, KimKJ ( 2007). Gene relocations within chloroplast genomes of Jasminum and Menodora( Oleaceae) are due to multiple, overlapping inversions Mol Biol Evol 24, 1161-1180. [本文引用: 4]
LiJH, AlexanderJH, ZhangDL ( 2002). Paraphyletic Syringa( Oleaceae): evidence from sequences of nuclear ribosomal DNA ITS and ETS regions Syst Bot 27, 592-597. [本文引用: 1]
MaierRM, NeckermannK, IgloiGL, K?selH ( 1995). Complete sequence of the maize chloroplast genome: gene content, hotspots of divergence and fine tuning of genetic information by transcript editing J Mol Biol 251, 614-628. [本文引用: 1]
OgiharaY, TerachiT, SasakumaT ( 1988). Intramolecular recombination of chloroplast genome mediated by short direct-repeat sequences in wheat species Proc Natl Acad Sci USA 85, 8573-8577. [本文引用: 1]
PalmerJD ( 1985). Comparative organization of chloroplast genomes Annu Rev Genet 19, 325-354. [本文引用: 1]
StamatakisA, HooverP, RougemontJ ( 2008). A rapid bootstrap algorithm for the raxml web servers Syst Biol 57, 758-771.
ThielT, MichalekW, VarshneyR, GranerA ( 2003). Exploiting EST databases for the development and characterization of gene-derived SSR-markers in barley (Hordeum vulgare L.) Theor Appl Genet 106, 411-422. [本文引用: 1]
Van de PaerC, BouchezO, BesnardG ( 2018). Prospects on the evolutionary mitogenomics of plants: a case study on the olive family (Oleaceae) Mol Ecol Res 18, 407-423. [本文引用: 2]
WallanderE ( 2008). Systematics of Fraxinus( Oleaceae) and evolution of dioecy Plant Syst Evol 273, 25-49.
WallanderE, AlbertVA ( 2000). Phylogeny and classification of Oleaceae based on rps16 and trnL-F sequence data Am J Bot 87, 1827-1841. [本文引用: 2]
WangWB, YuH, WangJH, LeiWJ, GaoJH, QiuXP, WangJS ( 2017). The complete chloroplast genome sequences of the medicinal plant Forsythia suspensa( Oleaceae) Int J Mol Sci 18, 2288. [本文引用: 2]
WengML, BlazierJC, GovinduM, JansenRK ( 2014). Reconstruction of the ancestral plastid genome in Geraniaceae reveals a correlation between genome rearrangements, repeats, and nucleotide substitution rates Mol Biol Evol 31, 645-659. [本文引用: 1]
ZhangTW, FangYJ, WangXM, DengX, ZhangXW, HuSN, YuJ ( 2012). The complete chloroplast and mitochondrial genome sequences of Boea hygrometrica: insights into the evolution of plant organellar genomes PLoS One 7, e30531.
ZhangX, ZhouT, YangJ, SunJJ, JuMM, ZhaoYM, ZhaoGF ( 2018). Comparative analyses of chloroplast genomes of Cucurbitaceae species: lights into selective pressures and phylogenetic relationships Molecules 23, 2165.
鲁桑叶绿体基因组序列及特征分析 1 2018
... 叶绿体基因组序列共线性分析显示, 木犀科Jasminum属3个种的叶绿体基因组部分片段有重排现象.研究表明Jasminum属部分基因位置变化具有物种特异性, 且可能与基因组结构有关(Lee et al., 2007; Greiner et al., 2008).Lee等(2007)认为J. nudiflorum中的部分基因序列重排是由2段序列的先后倒位(inversion)所致, 该研究还标注了基因重排的具体位置并估算了倒位发生的大概时间.也有研究表明重复序列可能是倒位的原因之一(Palmer, 1985; Knox et al., 1993).长且复杂的重复序列可能在基因组的重排或重组中起重要作用(Ogihara et al., 1988; Weng et al., 2014), 甚至重复序列之间的重组也可能诱导重排(Gray et al., 2009).之前牻牛儿苗科发生了基因重排的天竺葵属和牻牛儿苗属中都含有大量的大片段重复序列(Guisinger et al., 2011).本研究中出现基因重排的Jasminum属3种类型的重复序列无论数量还是长度均与其余物种有较大差异(图1A; 附表2, 附表3), 出现大量大于100 bp的重复序列, 数量也明显多, 与前人结论相符.在木犀科Jasminum属的3个物种中检测到的SSR数量与其余物种无明显差异, 但重复单元较长的3-6核苷酸重复大多分布在这3个物种中(附表4).同其它植物类似(李巧丽等, 2018; Zhang et al., 2018), 本研究检测到的大量SSR也可作为潜在的分子标记用于该类群植物的后续研究.此外, 在多个物种中存在accD基因缺失也可能与基因重排有关(Hiratsuka et al., 1989; Maier et al.,1995; Katayama and Ogihara, 1996). ...
禾本科植物叶绿体基因组结构的系统进化研究 2 2011
... 叶绿体是绿色植物进行光合作用的细胞器, 具有合成蛋白质、淀粉及色素等功能, 其基因组可自主遗传, 在植物漫长的进化历史中起重要作用.在被子植物中, 多数叶绿体基因组(cpDNA)为母系遗传, 少数为双亲遗传或父系遗传(Hu et al., 2008).叶绿体基因组结构和序列都比较保守, 大部分基因组结构为双链环状结构, 包括2个反向重复区(inverted repeats region, IRa/IRb)、1个大单拷贝区(large single copy region, LSC)和1个小单拷贝区(small single copy region, SSC) 4部分, 大小一般在120-180 kb之间(Zhang et al., 2012).虽然与核基因组和线粒体基因组相比, cpDNA在基因组成和结构上相对保守, 但近年的研究也表明许多cpDNA发生基因变异, 如基因或内含子片段的丢失(Gao et al., 2010; 唐萍等, 2011)、反向重复区的长度变异或部分片段的插入/缺失(Chumley et al., 2006)、整个反向重复区的扩张或缺失(Chumley et al., 2006)及基因重排(Guisinger et al., 2011; Blazier et al., 2016).由于基因组较小且结构相对保守, cpDNA已经成为进化和比较基因组研究的理想模型(Dong et al., 2013), 为揭示植物类群的系统位置和遗传发育关系提供更多有利依据. ...
Phylogenetics of Olea( Oleaceae) based on plastid and nuclear ribosomal DNA sequences: tertiary climatic shifts and lineage differentiation times 1 2009
... 木犀科植物多为乔木, 直立或藤状灌木, 属于被子植物中合瓣花亚纲的木犀亚目, 包含28属400余种, 广泛分布于温带和热带, 我国有10属160余种(Flora of China Editorial Committee, 1996).该科的茉莉(Jasminum sambac)、流苏(Chionanthus retusus)和迎春花(J. nudiflorum)等均是重要的园林植物; 连翘(Forsythia suspensa)则是重要的药用植物, 其果具清热解毒、消肿散结等功效.近年来, 随着分子系统学的发展, 有关木犀科的系统发育研究也有所增加(Wallander and Albert, 2000), 但多数研究仅基于核基因ITS或叶绿体基因片段, 且这些研究仅推断出木犀科部分属(如Fraxinus、Syringa、Ligustrum和Olea) (Jeandroz et al., 1997; Li et al., 2002; Wallander, 2008; Besnard et al., 2009)或种间的系统发育关系(Besnard et al., 2002; Baali cherif and Besnard, 2005).随着二代测序技术的不断发展, 有关木犀科叶绿体基因组方面的研究也逐渐兴起.Lee等(2007)主要分析了Jasminum和Menodora结构重排及其形成原因, 内容上更侧重于叶绿体基因组的结构变异.时至今日, 数据库中木犀科植物叶绿体基因组数据在不断更新, 但尚未见对该科叶绿体基因组结构及进化关系的系统研究.本研究在数据库共享信息不断增加的基础上, 拟通过下载整理数据库中木犀科19个物种的序列, 从叶绿体基因组的角度分析这19个物种的种间关系, 并通过比较这19个物种(11个属)的叶绿体基因组数据, 揭示该科植物叶绿体基因组的结构特征和变异情况. ...
Genomic profiling of plastid DNA variation in the mediterranean olive tree 0 2011
Olea europaea( Oleaceae) phylogeography based on chloroplast DNA polymorphism 1 2002
... 木犀科植物多为乔木, 直立或藤状灌木, 属于被子植物中合瓣花亚纲的木犀亚目, 包含28属400余种, 广泛分布于温带和热带, 我国有10属160余种(Flora of China Editorial Committee, 1996).该科的茉莉(Jasminum sambac)、流苏(Chionanthus retusus)和迎春花(J. nudiflorum)等均是重要的园林植物; 连翘(Forsythia suspensa)则是重要的药用植物, 其果具清热解毒、消肿散结等功效.近年来, 随着分子系统学的发展, 有关木犀科的系统发育研究也有所增加(Wallander and Albert, 2000), 但多数研究仅基于核基因ITS或叶绿体基因片段, 且这些研究仅推断出木犀科部分属(如Fraxinus、Syringa、Ligustrum和Olea) (Jeandroz et al., 1997; Li et al., 2002; Wallander, 2008; Besnard et al., 2009)或种间的系统发育关系(Besnard et al., 2002; Baali cherif and Besnard, 2005).随着二代测序技术的不断发展, 有关木犀科叶绿体基因组方面的研究也逐渐兴起.Lee等(2007)主要分析了Jasminum和Menodora结构重排及其形成原因, 内容上更侧重于叶绿体基因组的结构变异.时至今日, 数据库中木犀科植物叶绿体基因组数据在不断更新, 但尚未见对该科叶绿体基因组结构及进化关系的系统研究.本研究在数据库共享信息不断增加的基础上, 拟通过下载整理数据库中木犀科19个物种的序列, 从叶绿体基因组的角度分析这19个物种的种间关系, 并通过比较这19个物种(11个属)的叶绿体基因组数据, 揭示该科植物叶绿体基因组的结构特征和变异情况. ...
Divergence of RNA polymerase α subunits in angiosperm plastid genomes is mediated by genomic rearrangement 1 2016
... 叶绿体是绿色植物进行光合作用的细胞器, 具有合成蛋白质、淀粉及色素等功能, 其基因组可自主遗传, 在植物漫长的进化历史中起重要作用.在被子植物中, 多数叶绿体基因组(cpDNA)为母系遗传, 少数为双亲遗传或父系遗传(Hu et al., 2008).叶绿体基因组结构和序列都比较保守, 大部分基因组结构为双链环状结构, 包括2个反向重复区(inverted repeats region, IRa/IRb)、1个大单拷贝区(large single copy region, LSC)和1个小单拷贝区(small single copy region, SSC) 4部分, 大小一般在120-180 kb之间(Zhang et al., 2012).虽然与核基因组和线粒体基因组相比, cpDNA在基因组成和结构上相对保守, 但近年的研究也表明许多cpDNA发生基因变异, 如基因或内含子片段的丢失(Gao et al., 2010; 唐萍等, 2011)、反向重复区的长度变异或部分片段的插入/缺失(Chumley et al., 2006)、整个反向重复区的扩张或缺失(Chumley et al., 2006)及基因重排(Guisinger et al., 2011; Blazier et al., 2016).由于基因组较小且结构相对保守, cpDNA已经成为进化和比较基因组研究的理想模型(Dong et al., 2013), 为揭示植物类群的系统位置和遗传发育关系提供更多有利依据. ...
The complete chloroplast genome sequence of Pelargonium x hortorum: organization and evolution of the largest and most highly rearranged chloroplast genome of land plants 3 2006
... 叶绿体是绿色植物进行光合作用的细胞器, 具有合成蛋白质、淀粉及色素等功能, 其基因组可自主遗传, 在植物漫长的进化历史中起重要作用.在被子植物中, 多数叶绿体基因组(cpDNA)为母系遗传, 少数为双亲遗传或父系遗传(Hu et al., 2008).叶绿体基因组结构和序列都比较保守, 大部分基因组结构为双链环状结构, 包括2个反向重复区(inverted repeats region, IRa/IRb)、1个大单拷贝区(large single copy region, LSC)和1个小单拷贝区(small single copy region, SSC) 4部分, 大小一般在120-180 kb之间(Zhang et al., 2012).虽然与核基因组和线粒体基因组相比, cpDNA在基因组成和结构上相对保守, 但近年的研究也表明许多cpDNA发生基因变异, 如基因或内含子片段的丢失(Gao et al., 2010; 唐萍等, 2011)、反向重复区的长度变异或部分片段的插入/缺失(Chumley et al., 2006)、整个反向重复区的扩张或缺失(Chumley et al., 2006)及基因重排(Guisinger et al., 2011; Blazier et al., 2016).由于基因组较小且结构相对保守, cpDNA已经成为进化和比较基因组研究的理想模型(Dong et al., 2013), 为揭示植物类群的系统位置和遗传发育关系提供更多有利依据. ...
... )、整个反向重复区的扩张或缺失(Chumley et al., 2006)及基因重排(Guisinger et al., 2011; Blazier et al., 2016).由于基因组较小且结构相对保守, cpDNA已经成为进化和比较基因组研究的理想模型(Dong et al., 2013), 为揭示植物类群的系统位置和遗传发育关系提供更多有利依据. ...
... 被子植物的叶绿体基因组大小为120-180 kb, IR区长度为20-30 kb (Zhang et al., 2012).19个(11属)木犀科物种的叶绿体基因组大小为154-165 kb, IR区长25-29 kb, 与被子植物叶绿体基因组长度相吻合.本研究表明, 19个基因组大小差异约为10 kb, LSC、SSC与IR各区域的差异为LSC (7 kb)>SSC (4.7 kb)>IR (4 kb), 可见木犀科不同种间基因组长度的差异主要是LSC的差异.研究表明, 反向互补区可增强叶绿体基因组的稳定性和保守性(Khakhlova and Bock, 2006).本研究中木犀科11属19种植物, 除Jasminum属3个种外, 其余16个种的叶绿体基因组IR区差异均很小, 未发现IR区域过长、过短或丢失(Chumley et al., 2006; Hirao et al., 2008; Guisinger et al., 2011), 整体较保守且一致. ...
Mauve: multiple alignment of conserved genomic sequence with rearrangements 1 2004
... 叶绿体是绿色植物进行光合作用的细胞器, 具有合成蛋白质、淀粉及色素等功能, 其基因组可自主遗传, 在植物漫长的进化历史中起重要作用.在被子植物中, 多数叶绿体基因组(cpDNA)为母系遗传, 少数为双亲遗传或父系遗传(Hu et al., 2008).叶绿体基因组结构和序列都比较保守, 大部分基因组结构为双链环状结构, 包括2个反向重复区(inverted repeats region, IRa/IRb)、1个大单拷贝区(large single copy region, LSC)和1个小单拷贝区(small single copy region, SSC) 4部分, 大小一般在120-180 kb之间(Zhang et al., 2012).虽然与核基因组和线粒体基因组相比, cpDNA在基因组成和结构上相对保守, 但近年的研究也表明许多cpDNA发生基因变异, 如基因或内含子片段的丢失(Gao et al., 2010; 唐萍等, 2011)、反向重复区的长度变异或部分片段的插入/缺失(Chumley et al., 2006)、整个反向重复区的扩张或缺失(Chumley et al., 2006)及基因重排(Guisinger et al., 2011; Blazier et al., 2016).由于基因组较小且结构相对保守, cpDNA已经成为进化和比较基因组研究的理想模型(Dong et al., 2013), 为揭示植物类群的系统位置和遗传发育关系提供更多有利依据. ...
... 木犀科植物多为乔木, 直立或藤状灌木, 属于被子植物中合瓣花亚纲的木犀亚目, 包含28属400余种, 广泛分布于温带和热带, 我国有10属160余种(Flora of China Editorial Committee, 1996).该科的茉莉(Jasminum sambac)、流苏(Chionanthus retusus)和迎春花(J. nudiflorum)等均是重要的园林植物; 连翘(Forsythia suspensa)则是重要的药用植物, 其果具清热解毒、消肿散结等功效.近年来, 随着分子系统学的发展, 有关木犀科的系统发育研究也有所增加(Wallander and Albert, 2000), 但多数研究仅基于核基因ITS或叶绿体基因片段, 且这些研究仅推断出木犀科部分属(如Fraxinus、Syringa、Ligustrum和Olea) (Jeandroz et al., 1997; Li et al., 2002; Wallander, 2008; Besnard et al., 2009)或种间的系统发育关系(Besnard et al., 2002; Baali cherif and Besnard, 2005).随着二代测序技术的不断发展, 有关木犀科叶绿体基因组方面的研究也逐渐兴起.Lee等(2007)主要分析了Jasminum和Menodora结构重排及其形成原因, 内容上更侧重于叶绿体基因组的结构变异.时至今日, 数据库中木犀科植物叶绿体基因组数据在不断更新, 但尚未见对该科叶绿体基因组结构及进化关系的系统研究.本研究在数据库共享信息不断增加的基础上, 拟通过下载整理数据库中木犀科19个物种的序列, 从叶绿体基因组的角度分析这19个物种的种间关系, 并通过比较这19个物种(11个属)的叶绿体基因组数据, 揭示该科植物叶绿体基因组的结构特征和变异情况. ...
Vista: computational tools for comparative genomics 1 2004
... 叶绿体基因组序列共线性分析显示, 木犀科Jasminum属3个种的叶绿体基因组部分片段有重排现象.研究表明Jasminum属部分基因位置变化具有物种特异性, 且可能与基因组结构有关(Lee et al., 2007; Greiner et al., 2008).Lee等(2007)认为J. nudiflorum中的部分基因序列重排是由2段序列的先后倒位(inversion)所致, 该研究还标注了基因重排的具体位置并估算了倒位发生的大概时间.也有研究表明重复序列可能是倒位的原因之一(Palmer, 1985; Knox et al., 1993).长且复杂的重复序列可能在基因组的重排或重组中起重要作用(Ogihara et al., 1988; Weng et al., 2014), 甚至重复序列之间的重组也可能诱导重排(Gray et al., 2009).之前牻牛儿苗科发生了基因重排的天竺葵属和牻牛儿苗属中都含有大量的大片段重复序列(Guisinger et al., 2011).本研究中出现基因重排的Jasminum属3种类型的重复序列无论数量还是长度均与其余物种有较大差异(图1A; 附表2, 附表3), 出现大量大于100 bp的重复序列, 数量也明显多, 与前人结论相符.在木犀科Jasminum属的3个物种中检测到的SSR数量与其余物种无明显差异, 但重复单元较长的3-6核苷酸重复大多分布在这3个物种中(附表4).同其它植物类似(李巧丽等, 2018; Zhang et al., 2018), 本研究检测到的大量SSR也可作为潜在的分子标记用于该类群植物的后续研究.此外, 在多个物种中存在accD基因缺失也可能与基因重排有关(Hiratsuka et al., 1989; Maier et al.,1995; Katayama and Ogihara, 1996). ...
Plastid genome sequencing, comparative genomics, and phylogenomics: current status and prospects 1 2010
... 叶绿体基因组序列共线性分析显示, 木犀科Jasminum属3个种的叶绿体基因组部分片段有重排现象.研究表明Jasminum属部分基因位置变化具有物种特异性, 且可能与基因组结构有关(Lee et al., 2007; Greiner et al., 2008).Lee等(2007)认为J. nudiflorum中的部分基因序列重排是由2段序列的先后倒位(inversion)所致, 该研究还标注了基因重排的具体位置并估算了倒位发生的大概时间.也有研究表明重复序列可能是倒位的原因之一(Palmer, 1985; Knox et al., 1993).长且复杂的重复序列可能在基因组的重排或重组中起重要作用(Ogihara et al., 1988; Weng et al., 2014), 甚至重复序列之间的重组也可能诱导重排(Gray et al., 2009).之前牻牛儿苗科发生了基因重排的天竺葵属和牻牛儿苗属中都含有大量的大片段重复序列(Guisinger et al., 2011).本研究中出现基因重排的Jasminum属3种类型的重复序列无论数量还是长度均与其余物种有较大差异(图1A; 附表2, 附表3), 出现大量大于100 bp的重复序列, 数量也明显多, 与前人结论相符.在木犀科Jasminum属的3个物种中检测到的SSR数量与其余物种无明显差异, 但重复单元较长的3-6核苷酸重复大多分布在这3个物种中(附表4).同其它植物类似(李巧丽等, 2018; Zhang et al., 2018), 本研究检测到的大量SSR也可作为潜在的分子标记用于该类群植物的后续研究.此外, 在多个物种中存在accD基因缺失也可能与基因重排有关(Hiratsuka et al., 1989; Maier et al.,1995; Katayama and Ogihara, 1996). ...
Extensive homologous recombination between introduced and native regulatory plastid DNA elements in transplastomic plants 3 2009
... 叶绿体是绿色植物进行光合作用的细胞器, 具有合成蛋白质、淀粉及色素等功能, 其基因组可自主遗传, 在植物漫长的进化历史中起重要作用.在被子植物中, 多数叶绿体基因组(cpDNA)为母系遗传, 少数为双亲遗传或父系遗传(Hu et al., 2008).叶绿体基因组结构和序列都比较保守, 大部分基因组结构为双链环状结构, 包括2个反向重复区(inverted repeats region, IRa/IRb)、1个大单拷贝区(large single copy region, LSC)和1个小单拷贝区(small single copy region, SSC) 4部分, 大小一般在120-180 kb之间(Zhang et al., 2012).虽然与核基因组和线粒体基因组相比, cpDNA在基因组成和结构上相对保守, 但近年的研究也表明许多cpDNA发生基因变异, 如基因或内含子片段的丢失(Gao et al., 2010; 唐萍等, 2011)、反向重复区的长度变异或部分片段的插入/缺失(Chumley et al., 2006)、整个反向重复区的扩张或缺失(Chumley et al., 2006)及基因重排(Guisinger et al., 2011; Blazier et al., 2016).由于基因组较小且结构相对保守, cpDNA已经成为进化和比较基因组研究的理想模型(Dong et al., 2013), 为揭示植物类群的系统位置和遗传发育关系提供更多有利依据. ...
... 被子植物的叶绿体基因组大小为120-180 kb, IR区长度为20-30 kb (Zhang et al., 2012).19个(11属)木犀科物种的叶绿体基因组大小为154-165 kb, IR区长25-29 kb, 与被子植物叶绿体基因组长度相吻合.本研究表明, 19个基因组大小差异约为10 kb, LSC、SSC与IR各区域的差异为LSC (7 kb)>SSC (4.7 kb)>IR (4 kb), 可见木犀科不同种间基因组长度的差异主要是LSC的差异.研究表明, 反向互补区可增强叶绿体基因组的稳定性和保守性(Khakhlova and Bock, 2006).本研究中木犀科11属19种植物, 除Jasminum属3个种外, 其余16个种的叶绿体基因组IR区差异均很小, 未发现IR区域过长、过短或丢失(Chumley et al., 2006; Hirao et al., 2008; Guisinger et al., 2011), 整体较保守且一致. ...
... 叶绿体基因组序列共线性分析显示, 木犀科Jasminum属3个种的叶绿体基因组部分片段有重排现象.研究表明Jasminum属部分基因位置变化具有物种特异性, 且可能与基因组结构有关(Lee et al., 2007; Greiner et al., 2008).Lee等(2007)认为J. nudiflorum中的部分基因序列重排是由2段序列的先后倒位(inversion)所致, 该研究还标注了基因重排的具体位置并估算了倒位发生的大概时间.也有研究表明重复序列可能是倒位的原因之一(Palmer, 1985; Knox et al., 1993).长且复杂的重复序列可能在基因组的重排或重组中起重要作用(Ogihara et al., 1988; Weng et al., 2014), 甚至重复序列之间的重组也可能诱导重排(Gray et al., 2009).之前牻牛儿苗科发生了基因重排的天竺葵属和牻牛儿苗属中都含有大量的大片段重复序列(Guisinger et al., 2011).本研究中出现基因重排的Jasminum属3种类型的重复序列无论数量还是长度均与其余物种有较大差异(图1A; 附表2, 附表3), 出现大量大于100 bp的重复序列, 数量也明显多, 与前人结论相符.在木犀科Jasminum属的3个物种中检测到的SSR数量与其余物种无明显差异, 但重复单元较长的3-6核苷酸重复大多分布在这3个物种中(附表4).同其它植物类似(李巧丽等, 2018; Zhang et al., 2018), 本研究检测到的大量SSR也可作为潜在的分子标记用于该类群植物的后续研究.此外, 在多个物种中存在accD基因缺失也可能与基因重排有关(Hiratsuka et al., 1989; Maier et al.,1995; Katayama and Ogihara, 1996). ...
The complete nucleotide sequences of the five genetically distinct plastid genomes of Oenothera, subsection Oenothera. I. Sequence evaluation and plastome evolution 1 2008
Extreme reconfiguration of plastid genomes in the angiosperm family Geraniaceae: rearrangements, repeats, and codon usage 0 2011
1 2007
... 被子植物的叶绿体基因组大小为120-180 kb, IR区长度为20-30 kb (Zhang et al., 2012).19个(11属)木犀科物种的叶绿体基因组大小为154-165 kb, IR区长25-29 kb, 与被子植物叶绿体基因组长度相吻合.本研究表明, 19个基因组大小差异约为10 kb, LSC、SSC与IR各区域的差异为LSC (7 kb)>SSC (4.7 kb)>IR (4 kb), 可见木犀科不同种间基因组长度的差异主要是LSC的差异.研究表明, 反向互补区可增强叶绿体基因组的稳定性和保守性(Khakhlova and Bock, 2006).本研究中木犀科11属19种植物, 除Jasminum属3个种外, 其余16个种的叶绿体基因组IR区差异均很小, 未发现IR区域过长、过短或丢失(Chumley et al., 2006; Hirao et al., 2008; Guisinger et al., 2011), 整体较保守且一致. ...
Characterization of the complete chloroplast genome of chinese fringetree (Chionanthus retusus) 1 2017
... 叶绿体基因组序列共线性分析显示, 木犀科Jasminum属3个种的叶绿体基因组部分片段有重排现象.研究表明Jasminum属部分基因位置变化具有物种特异性, 且可能与基因组结构有关(Lee et al., 2007; Greiner et al., 2008).Lee等(2007)认为J. nudiflorum中的部分基因序列重排是由2段序列的先后倒位(inversion)所致, 该研究还标注了基因重排的具体位置并估算了倒位发生的大概时间.也有研究表明重复序列可能是倒位的原因之一(Palmer, 1985; Knox et al., 1993).长且复杂的重复序列可能在基因组的重排或重组中起重要作用(Ogihara et al., 1988; Weng et al., 2014), 甚至重复序列之间的重组也可能诱导重排(Gray et al., 2009).之前牻牛儿苗科发生了基因重排的天竺葵属和牻牛儿苗属中都含有大量的大片段重复序列(Guisinger et al., 2011).本研究中出现基因重排的Jasminum属3种类型的重复序列无论数量还是长度均与其余物种有较大差异(图1A; 附表2, 附表3), 出现大量大于100 bp的重复序列, 数量也明显多, 与前人结论相符.在木犀科Jasminum属的3个物种中检测到的SSR数量与其余物种无明显差异, 但重复单元较长的3-6核苷酸重复大多分布在这3个物种中(附表4).同其它植物类似(李巧丽等, 2018; Zhang et al., 2018), 本研究检测到的大量SSR也可作为潜在的分子标记用于该类群植物的后续研究.此外, 在多个物种中存在accD基因缺失也可能与基因重排有关(Hiratsuka et al., 1989; Maier et al.,1995; Katayama and Ogihara, 1996). ...
Complete nucleotide sequence of the Cryptomeria japonica D. Don. chloroplast genome and comparative chloroplast genomics: diversified genomic structure of coniferous species 1 2008
... 叶绿体是绿色植物进行光合作用的细胞器, 具有合成蛋白质、淀粉及色素等功能, 其基因组可自主遗传, 在植物漫长的进化历史中起重要作用.在被子植物中, 多数叶绿体基因组(cpDNA)为母系遗传, 少数为双亲遗传或父系遗传(Hu et al., 2008).叶绿体基因组结构和序列都比较保守, 大部分基因组结构为双链环状结构, 包括2个反向重复区(inverted repeats region, IRa/IRb)、1个大单拷贝区(large single copy region, LSC)和1个小单拷贝区(small single copy region, SSC) 4部分, 大小一般在120-180 kb之间(Zhang et al., 2012).虽然与核基因组和线粒体基因组相比, cpDNA在基因组成和结构上相对保守, 但近年的研究也表明许多cpDNA发生基因变异, 如基因或内含子片段的丢失(Gao et al., 2010; 唐萍等, 2011)、反向重复区的长度变异或部分片段的插入/缺失(Chumley et al., 2006)、整个反向重复区的扩张或缺失(Chumley et al., 2006)及基因重排(Guisinger et al., 2011; Blazier et al., 2016).由于基因组较小且结构相对保守, cpDNA已经成为进化和比较基因组研究的理想模型(Dong et al., 2013), 为揭示植物类群的系统位置和遗传发育关系提供更多有利依据. ...
The complete sequence of the rice (Oryza sativa) chloroplast genome: intermolecular recombination between distinct tRNA genes accounts for a major plastid DNA inversion during the evolution of the cereals 1 1989
Occurrence of plastids in the sperm cells of Caprifoliaceae: biparental plastid inheritance in angiosperms is unilaterally derived from maternal inheritance 1 2008
... 木犀科植物多为乔木, 直立或藤状灌木, 属于被子植物中合瓣花亚纲的木犀亚目, 包含28属400余种, 广泛分布于温带和热带, 我国有10属160余种(Flora of China Editorial Committee, 1996).该科的茉莉(Jasminum sambac)、流苏(Chionanthus retusus)和迎春花(J. nudiflorum)等均是重要的园林植物; 连翘(Forsythia suspensa)则是重要的药用植物, 其果具清热解毒、消肿散结等功效.近年来, 随着分子系统学的发展, 有关木犀科的系统发育研究也有所增加(Wallander and Albert, 2000), 但多数研究仅基于核基因ITS或叶绿体基因片段, 且这些研究仅推断出木犀科部分属(如Fraxinus、Syringa、Ligustrum和Olea) (Jeandroz et al., 1997; Li et al., 2002; Wallander, 2008; Besnard et al., 2009)或种间的系统发育关系(Besnard et al., 2002; Baali cherif and Besnard, 2005).随着二代测序技术的不断发展, 有关木犀科叶绿体基因组方面的研究也逐渐兴起.Lee等(2007)主要分析了Jasminum和Menodora结构重排及其形成原因, 内容上更侧重于叶绿体基因组的结构变异.时至今日, 数据库中木犀科植物叶绿体基因组数据在不断更新, 但尚未见对该科叶绿体基因组结构及进化关系的系统研究.本研究在数据库共享信息不断增加的基础上, 拟通过下载整理数据库中木犀科19个物种的序列, 从叶绿体基因组的角度分析这19个物种的种间关系, 并通过比较这19个物种(11个属)的叶绿体基因组数据, 揭示该科植物叶绿体基因组的结构特征和变异情况. ...
Thirteen Camellia chloroplast genome sequences determined by high-throughput sequencing: genome structure and phylogenetic relationships 1 2014
... 叶绿体基因组序列共线性分析显示, 木犀科Jasminum属3个种的叶绿体基因组部分片段有重排现象.研究表明Jasminum属部分基因位置变化具有物种特异性, 且可能与基因组结构有关(Lee et al., 2007; Greiner et al., 2008).Lee等(2007)认为J. nudiflorum中的部分基因序列重排是由2段序列的先后倒位(inversion)所致, 该研究还标注了基因重排的具体位置并估算了倒位发生的大概时间.也有研究表明重复序列可能是倒位的原因之一(Palmer, 1985; Knox et al., 1993).长且复杂的重复序列可能在基因组的重排或重组中起重要作用(Ogihara et al., 1988; Weng et al., 2014), 甚至重复序列之间的重组也可能诱导重排(Gray et al., 2009).之前牻牛儿苗科发生了基因重排的天竺葵属和牻牛儿苗属中都含有大量的大片段重复序列(Guisinger et al., 2011).本研究中出现基因重排的Jasminum属3种类型的重复序列无论数量还是长度均与其余物种有较大差异(图1A; 附表2, 附表3), 出现大量大于100 bp的重复序列, 数量也明显多, 与前人结论相符.在木犀科Jasminum属的3个物种中检测到的SSR数量与其余物种无明显差异, 但重复单元较长的3-6核苷酸重复大多分布在这3个物种中(附表4).同其它植物类似(李巧丽等, 2018; Zhang et al., 2018), 本研究检测到的大量SSR也可作为潜在的分子标记用于该类群植物的后续研究.此外, 在多个物种中存在accD基因缺失也可能与基因重排有关(Hiratsuka et al., 1989; Maier et al.,1995; Katayama and Ogihara, 1996). ...
Phylogeny and phylogeography of the circumpolar genus Fraxinus( Oleaceae) based on internal transcribed spacer sequences of nuclear ribosomal DNA 1 1997
... 被子植物的叶绿体基因组大小为120-180 kb, IR区长度为20-30 kb (Zhang et al., 2012).19个(11属)木犀科物种的叶绿体基因组大小为154-165 kb, IR区长25-29 kb, 与被子植物叶绿体基因组长度相吻合.本研究表明, 19个基因组大小差异约为10 kb, LSC、SSC与IR各区域的差异为LSC (7 kb)>SSC (4.7 kb)>IR (4 kb), 可见木犀科不同种间基因组长度的差异主要是LSC的差异.研究表明, 反向互补区可增强叶绿体基因组的稳定性和保守性(Khakhlova and Bock, 2006).本研究中木犀科11属19种植物, 除Jasminum属3个种外, 其余16个种的叶绿体基因组IR区差异均很小, 未发现IR区域过长、过短或丢失(Chumley et al., 2006; Hirao et al., 2008; Guisinger et al., 2011), 整体较保守且一致. ...
Phylogenetic affinities of the grasses to other monocots as revealed by molecular analysis of chloroplast DNA 0 1996
Elimination of deleterious mutations in plastid genomes by gene conversion 0 2006
Characterization of the complete chloroplast genome of Fraxinus chiisanensis( Oleaceae), an endemic to Korea 1 2018
... 叶绿体基因组序列共线性分析显示, 木犀科Jasminum属3个种的叶绿体基因组部分片段有重排现象.研究表明Jasminum属部分基因位置变化具有物种特异性, 且可能与基因组结构有关(Lee et al., 2007; Greiner et al., 2008).Lee等(2007)认为J. nudiflorum中的部分基因序列重排是由2段序列的先后倒位(inversion)所致, 该研究还标注了基因重排的具体位置并估算了倒位发生的大概时间.也有研究表明重复序列可能是倒位的原因之一(Palmer, 1985; Knox et al., 1993).长且复杂的重复序列可能在基因组的重排或重组中起重要作用(Ogihara et al., 1988; Weng et al., 2014), 甚至重复序列之间的重组也可能诱导重排(Gray et al., 2009).之前牻牛儿苗科发生了基因重排的天竺葵属和牻牛儿苗属中都含有大量的大片段重复序列(Guisinger et al., 2011).本研究中出现基因重排的Jasminum属3种类型的重复序列无论数量还是长度均与其余物种有较大差异(图1A; 附表2, 附表3), 出现大量大于100 bp的重复序列, 数量也明显多, 与前人结论相符.在木犀科Jasminum属的3个物种中检测到的SSR数量与其余物种无明显差异, 但重复单元较长的3-6核苷酸重复大多分布在这3个物种中(附表4).同其它植物类似(李巧丽等, 2018; Zhang et al., 2018), 本研究检测到的大量SSR也可作为潜在的分子标记用于该类群植物的后续研究.此外, 在多个物种中存在accD基因缺失也可能与基因重排有关(Hiratsuka et al., 1989; Maier et al.,1995; Katayama and Ogihara, 1996). ...
Complete plastid genome sequences of Abeliophyllum distichum Nakai (Oleaceae), a Korea endemic genus 1 2016
... 叶绿体基因组序列共线性分析显示, 木犀科Jasminum属3个种的叶绿体基因组部分片段有重排现象.研究表明Jasminum属部分基因位置变化具有物种特异性, 且可能与基因组结构有关(Lee et al., 2007; Greiner et al., 2008).Lee等(2007)认为J. nudiflorum中的部分基因序列重排是由2段序列的先后倒位(inversion)所致, 该研究还标注了基因重排的具体位置并估算了倒位发生的大概时间.也有研究表明重复序列可能是倒位的原因之一(Palmer, 1985; Knox et al., 1993).长且复杂的重复序列可能在基因组的重排或重组中起重要作用(Ogihara et al., 1988; Weng et al., 2014), 甚至重复序列之间的重组也可能诱导重排(Gray et al., 2009).之前牻牛儿苗科发生了基因重排的天竺葵属和牻牛儿苗属中都含有大量的大片段重复序列(Guisinger et al., 2011).本研究中出现基因重排的Jasminum属3种类型的重复序列无论数量还是长度均与其余物种有较大差异(图1A; 附表2, 附表3), 出现大量大于100 bp的重复序列, 数量也明显多, 与前人结论相符.在木犀科Jasminum属的3个物种中检测到的SSR数量与其余物种无明显差异, 但重复单元较长的3-6核苷酸重复大多分布在这3个物种中(附表4).同其它植物类似(李巧丽等, 2018; Zhang et al., 2018), 本研究检测到的大量SSR也可作为潜在的分子标记用于该类群植物的后续研究.此外, 在多个物种中存在accD基因缺失也可能与基因重排有关(Hiratsuka et al., 1989; Maier et al.,1995; Katayama and Ogihara, 1996). ...
... ; Maier et al.,1995; Katayama and Ogihara, 1996). ...
Paraphyletic Syringa( Oleaceae): evidence from sequences of nuclear ribosomal DNA ITS and ETS regions 1 2002
... 叶绿体基因组序列共线性分析显示, 木犀科Jasminum属3个种的叶绿体基因组部分片段有重排现象.研究表明Jasminum属部分基因位置变化具有物种特异性, 且可能与基因组结构有关(Lee et al., 2007; Greiner et al., 2008).Lee等(2007)认为J. nudiflorum中的部分基因序列重排是由2段序列的先后倒位(inversion)所致, 该研究还标注了基因重排的具体位置并估算了倒位发生的大概时间.也有研究表明重复序列可能是倒位的原因之一(Palmer, 1985; Knox et al., 1993).长且复杂的重复序列可能在基因组的重排或重组中起重要作用(Ogihara et al., 1988; Weng et al., 2014), 甚至重复序列之间的重组也可能诱导重排(Gray et al., 2009).之前牻牛儿苗科发生了基因重排的天竺葵属和牻牛儿苗属中都含有大量的大片段重复序列(Guisinger et al., 2011).本研究中出现基因重排的Jasminum属3种类型的重复序列无论数量还是长度均与其余物种有较大差异(图1A; 附表2, 附表3), 出现大量大于100 bp的重复序列, 数量也明显多, 与前人结论相符.在木犀科Jasminum属的3个物种中检测到的SSR数量与其余物种无明显差异, 但重复单元较长的3-6核苷酸重复大多分布在这3个物种中(附表4).同其它植物类似(李巧丽等, 2018; Zhang et al., 2018), 本研究检测到的大量SSR也可作为潜在的分子标记用于该类群植物的后续研究.此外, 在多个物种中存在accD基因缺失也可能与基因重排有关(Hiratsuka et al., 1989; Maier et al.,1995; Katayama and Ogihara, 1996). ...
Complete sequence of the maize chloroplast genome: gene content, hotspots of divergence and fine tuning of genetic information by transcript editing 1 1995
... 叶绿体基因组序列共线性分析显示, 木犀科Jasminum属3个种的叶绿体基因组部分片段有重排现象.研究表明Jasminum属部分基因位置变化具有物种特异性, 且可能与基因组结构有关(Lee et al., 2007; Greiner et al., 2008).Lee等(2007)认为J. nudiflorum中的部分基因序列重排是由2段序列的先后倒位(inversion)所致, 该研究还标注了基因重排的具体位置并估算了倒位发生的大概时间.也有研究表明重复序列可能是倒位的原因之一(Palmer, 1985; Knox et al., 1993).长且复杂的重复序列可能在基因组的重排或重组中起重要作用(Ogihara et al., 1988; Weng et al., 2014), 甚至重复序列之间的重组也可能诱导重排(Gray et al., 2009).之前牻牛儿苗科发生了基因重排的天竺葵属和牻牛儿苗属中都含有大量的大片段重复序列(Guisinger et al., 2011).本研究中出现基因重排的Jasminum属3种类型的重复序列无论数量还是长度均与其余物种有较大差异(图1A; 附表2, 附表3), 出现大量大于100 bp的重复序列, 数量也明显多, 与前人结论相符.在木犀科Jasminum属的3个物种中检测到的SSR数量与其余物种无明显差异, 但重复单元较长的3-6核苷酸重复大多分布在这3个物种中(附表4).同其它植物类似(李巧丽等, 2018; Zhang et al., 2018), 本研究检测到的大量SSR也可作为潜在的分子标记用于该类群植物的后续研究.此外, 在多个物种中存在accD基因缺失也可能与基因重排有关(Hiratsuka et al., 1989; Maier et al.,1995; Katayama and Ogihara, 1996). ...
Intramolecular recombination of chloroplast genome mediated by short direct-repeat sequences in wheat species 1 1988
A rapid bootstrap algorithm for the raxml web servers 0 2008
Exploiting EST databases for the development and characterization of gene-derived SSR-markers in barley (Hordeum vulgare L.) 1 2003
... 木犀科植物多为乔木, 直立或藤状灌木, 属于被子植物中合瓣花亚纲的木犀亚目, 包含28属400余种, 广泛分布于温带和热带, 我国有10属160余种(Flora of China Editorial Committee, 1996).该科的茉莉(Jasminum sambac)、流苏(Chionanthus retusus)和迎春花(J. nudiflorum)等均是重要的园林植物; 连翘(Forsythia suspensa)则是重要的药用植物, 其果具清热解毒、消肿散结等功效.近年来, 随着分子系统学的发展, 有关木犀科的系统发育研究也有所增加(Wallander and Albert, 2000), 但多数研究仅基于核基因ITS或叶绿体基因片段, 且这些研究仅推断出木犀科部分属(如Fraxinus、Syringa、Ligustrum和Olea) (Jeandroz et al., 1997; Li et al., 2002; Wallander, 2008; Besnard et al., 2009)或种间的系统发育关系(Besnard et al., 2002; Baali cherif and Besnard, 2005).随着二代测序技术的不断发展, 有关木犀科叶绿体基因组方面的研究也逐渐兴起.Lee等(2007)主要分析了Jasminum和Menodora结构重排及其形成原因, 内容上更侧重于叶绿体基因组的结构变异.时至今日, 数据库中木犀科植物叶绿体基因组数据在不断更新, 但尚未见对该科叶绿体基因组结构及进化关系的系统研究.本研究在数据库共享信息不断增加的基础上, 拟通过下载整理数据库中木犀科19个物种的序列, 从叶绿体基因组的角度分析这19个物种的种间关系, 并通过比较这19个物种(11个属)的叶绿体基因组数据, 揭示该科植物叶绿体基因组的结构特征和变异情况. ...
Prospects on the evolutionary mitogenomics of plants: a case study on the olive family (Oleaceae) 2 2018
... 木犀科植物多为乔木, 直立或藤状灌木, 属于被子植物中合瓣花亚纲的木犀亚目, 包含28属400余种, 广泛分布于温带和热带, 我国有10属160余种(Flora of China Editorial Committee, 1996).该科的茉莉(Jasminum sambac)、流苏(Chionanthus retusus)和迎春花(J. nudiflorum)等均是重要的园林植物; 连翘(Forsythia suspensa)则是重要的药用植物, 其果具清热解毒、消肿散结等功效.近年来, 随着分子系统学的发展, 有关木犀科的系统发育研究也有所增加(Wallander and Albert, 2000), 但多数研究仅基于核基因ITS或叶绿体基因片段, 且这些研究仅推断出木犀科部分属(如Fraxinus、Syringa、Ligustrum和Olea) (Jeandroz et al., 1997; Li et al., 2002; Wallander, 2008; Besnard et al., 2009)或种间的系统发育关系(Besnard et al., 2002; Baali cherif and Besnard, 2005).随着二代测序技术的不断发展, 有关木犀科叶绿体基因组方面的研究也逐渐兴起.Lee等(2007)主要分析了Jasminum和Menodora结构重排及其形成原因, 内容上更侧重于叶绿体基因组的结构变异.时至今日, 数据库中木犀科植物叶绿体基因组数据在不断更新, 但尚未见对该科叶绿体基因组结构及进化关系的系统研究.本研究在数据库共享信息不断增加的基础上, 拟通过下载整理数据库中木犀科19个物种的序列, 从叶绿体基因组的角度分析这19个物种的种间关系, 并通过比较这19个物种(11个属)的叶绿体基因组数据, 揭示该科植物叶绿体基因组的结构特征和变异情况. ...
... 本研究基于木犀科19个物种的叶绿体基因组共有的蛋白编码基因(CDS)数据构建系统发育树, 该科各属的关系均得到较好的处理, 11个属各自聚为一支且各节点均获得了较高的支持率.A. distichum和F. suspensa位于进化树的基部, 属木犀科中较为原始的类群.Jasminum中出现基因重排, 但基于保守的CDS序列构建的系统发育树表明该属并不是木犀科中的原始类群.ndhF和rbcL联合序列的系统发育分析也表明, Abeliophyllum、Forsythia、Fontanesia、Myxopyrum和Nyctanthes属为原始类群, 其分化起始于白垩纪早期(本研究中无后3个属的基因组数据), 支持Jasminum是单系类群且其分化起始于白垩纪晚期(78.3 MYA), 同时第1次DNA序列倒位也发生于该时期, 该属其它物种分化于始新世中期(42.1 MYA) (Lee et al., 2007).Wallander和Albert (2000)曾用rps16和trnL-F序列对木犀科76个物种(25属)进行系统发育分析, 结果表明Abeliophyllum和Forsythia聚在一起并位于进化树基部.本研究结果与其相符, 进一步在叶绿体基因组水平证明了这2个属在木犀科中的原始地位.但如上所述, 该文中Fontanesia和Nactanthes位于系统进化树最基部, 因数据缺乏, 我们无法对这2个研究中的系统发育关系进行更具体的比较.本研究中, Olea、Nestegis、Chionanthus、Forestiera、Schrebera、Fraxinus和Syringa属均为较晚出的分支, 但其具体的系统位置在不同序列构建的系统发育树中稍有不同(Wallander and Albert, 2000; Lee et al., 2007), 可能是所采用的数据集信息不同所致.此外, 本研究中样品数量不足也可能影响分析结果. ...
Systematics of Fraxinus( Oleaceae) and evolution of dioecy 0 2008
Phylogeny and classification of Oleaceae based on rps16 and trnL-F sequence data 2 2000
... 叶绿体基因组序列共线性分析显示, 木犀科Jasminum属3个种的叶绿体基因组部分片段有重排现象.研究表明Jasminum属部分基因位置变化具有物种特异性, 且可能与基因组结构有关(Lee et al., 2007; Greiner et al., 2008).Lee等(2007)认为J. nudiflorum中的部分基因序列重排是由2段序列的先后倒位(inversion)所致, 该研究还标注了基因重排的具体位置并估算了倒位发生的大概时间.也有研究表明重复序列可能是倒位的原因之一(Palmer, 1985; Knox et al., 1993).长且复杂的重复序列可能在基因组的重排或重组中起重要作用(Ogihara et al., 1988; Weng et al., 2014), 甚至重复序列之间的重组也可能诱导重排(Gray et al., 2009).之前牻牛儿苗科发生了基因重排的天竺葵属和牻牛儿苗属中都含有大量的大片段重复序列(Guisinger et al., 2011).本研究中出现基因重排的Jasminum属3种类型的重复序列无论数量还是长度均与其余物种有较大差异(图1A; 附表2, 附表3), 出现大量大于100 bp的重复序列, 数量也明显多, 与前人结论相符.在木犀科Jasminum属的3个物种中检测到的SSR数量与其余物种无明显差异, 但重复单元较长的3-6核苷酸重复大多分布在这3个物种中(附表4).同其它植物类似(李巧丽等, 2018; Zhang et al., 2018), 本研究检测到的大量SSR也可作为潜在的分子标记用于该类群植物的后续研究.此外, 在多个物种中存在accD基因缺失也可能与基因重排有关(Hiratsuka et al., 1989; Maier et al.,1995; Katayama and Ogihara, 1996). ...
... 本研究基于木犀科19个物种的叶绿体基因组共有的蛋白编码基因(CDS)数据构建系统发育树, 该科各属的关系均得到较好的处理, 11个属各自聚为一支且各节点均获得了较高的支持率.A. distichum和F. suspensa位于进化树的基部, 属木犀科中较为原始的类群.Jasminum中出现基因重排, 但基于保守的CDS序列构建的系统发育树表明该属并不是木犀科中的原始类群.ndhF和rbcL联合序列的系统发育分析也表明, Abeliophyllum、Forsythia、Fontanesia、Myxopyrum和Nyctanthes属为原始类群, 其分化起始于白垩纪早期(本研究中无后3个属的基因组数据), 支持Jasminum是单系类群且其分化起始于白垩纪晚期(78.3 MYA), 同时第1次DNA序列倒位也发生于该时期, 该属其它物种分化于始新世中期(42.1 MYA) (Lee et al., 2007).Wallander和Albert (2000)曾用rps16和trnL-F序列对木犀科76个物种(25属)进行系统发育分析, 结果表明Abeliophyllum和Forsythia聚在一起并位于进化树基部.本研究结果与其相符, 进一步在叶绿体基因组水平证明了这2个属在木犀科中的原始地位.但如上所述, 该文中Fontanesia和Nactanthes位于系统进化树最基部, 因数据缺乏, 我们无法对这2个研究中的系统发育关系进行更具体的比较.本研究中, Olea、Nestegis、Chionanthus、Forestiera、Schrebera、Fraxinus和Syringa属均为较晚出的分支, 但其具体的系统位置在不同序列构建的系统发育树中稍有不同(Wallander and Albert, 2000; Lee et al., 2007), 可能是所采用的数据集信息不同所致.此外, 本研究中样品数量不足也可能影响分析结果. ...
The complete chloroplast genome sequences of the medicinal plant Forsythia suspensa( Oleaceae) 2 2017
... 叶绿体是绿色植物进行光合作用的细胞器, 具有合成蛋白质、淀粉及色素等功能, 其基因组可自主遗传, 在植物漫长的进化历史中起重要作用.在被子植物中, 多数叶绿体基因组(cpDNA)为母系遗传, 少数为双亲遗传或父系遗传(Hu et al., 2008).叶绿体基因组结构和序列都比较保守, 大部分基因组结构为双链环状结构, 包括2个反向重复区(inverted repeats region, IRa/IRb)、1个大单拷贝区(large single copy region, LSC)和1个小单拷贝区(small single copy region, SSC) 4部分, 大小一般在120-180 kb之间(Zhang et al., 2012).虽然与核基因组和线粒体基因组相比, cpDNA在基因组成和结构上相对保守, 但近年的研究也表明许多cpDNA发生基因变异, 如基因或内含子片段的丢失(Gao et al., 2010; 唐萍等, 2011)、反向重复区的长度变异或部分片段的插入/缺失(Chumley et al., 2006)、整个反向重复区的扩张或缺失(Chumley et al., 2006)及基因重排(Guisinger et al., 2011; Blazier et al., 2016).由于基因组较小且结构相对保守, cpDNA已经成为进化和比较基因组研究的理想模型(Dong et al., 2013), 为揭示植物类群的系统位置和遗传发育关系提供更多有利依据. ...
... 被子植物的叶绿体基因组大小为120-180 kb, IR区长度为20-30 kb (Zhang et al., 2012).19个(11属)木犀科物种的叶绿体基因组大小为154-165 kb, IR区长25-29 kb, 与被子植物叶绿体基因组长度相吻合.本研究表明, 19个基因组大小差异约为10 kb, LSC、SSC与IR各区域的差异为LSC (7 kb)>SSC (4.7 kb)>IR (4 kb), 可见木犀科不同种间基因组长度的差异主要是LSC的差异.研究表明, 反向互补区可增强叶绿体基因组的稳定性和保守性(Khakhlova and Bock, 2006).本研究中木犀科11属19种植物, 除Jasminum属3个种外, 其余16个种的叶绿体基因组IR区差异均很小, 未发现IR区域过长、过短或丢失(Chumley et al., 2006; Hirao et al., 2008; Guisinger et al., 2011), 整体较保守且一致. ...
Reconstruction of the ancestral plastid genome in Geraniaceae reveals a correlation between genome rearrangements, repeats, and nucleotide substitution rates 1 2014
... 叶绿体基因组序列共线性分析显示, 木犀科Jasminum属3个种的叶绿体基因组部分片段有重排现象.研究表明Jasminum属部分基因位置变化具有物种特异性, 且可能与基因组结构有关(Lee et al., 2007; Greiner et al., 2008).Lee等(2007)认为J. nudiflorum中的部分基因序列重排是由2段序列的先后倒位(inversion)所致, 该研究还标注了基因重排的具体位置并估算了倒位发生的大概时间.也有研究表明重复序列可能是倒位的原因之一(Palmer, 1985; Knox et al., 1993).长且复杂的重复序列可能在基因组的重排或重组中起重要作用(Ogihara et al., 1988; Weng et al., 2014), 甚至重复序列之间的重组也可能诱导重排(Gray et al., 2009).之前牻牛儿苗科发生了基因重排的天竺葵属和牻牛儿苗属中都含有大量的大片段重复序列(Guisinger et al., 2011).本研究中出现基因重排的Jasminum属3种类型的重复序列无论数量还是长度均与其余物种有较大差异(图1A; 附表2, 附表3), 出现大量大于100 bp的重复序列, 数量也明显多, 与前人结论相符.在木犀科Jasminum属的3个物种中检测到的SSR数量与其余物种无明显差异, 但重复单元较长的3-6核苷酸重复大多分布在这3个物种中(附表4).同其它植物类似(李巧丽等, 2018; Zhang et al., 2018), 本研究检测到的大量SSR也可作为潜在的分子标记用于该类群植物的后续研究.此外, 在多个物种中存在accD基因缺失也可能与基因重排有关(Hiratsuka et al., 1989; Maier et al.,1995; Katayama and Ogihara, 1996). ...
The complete chloroplast and mitochondrial genome sequences of Boea hygrometrica: insights into the evolution of plant organellar genomes 0 2012
Comparative analyses of chloroplast genomes of Cucurbitaceae species: lights into selective pressures and phylogenetic relationships 0 2018
 ,3,*
,3,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT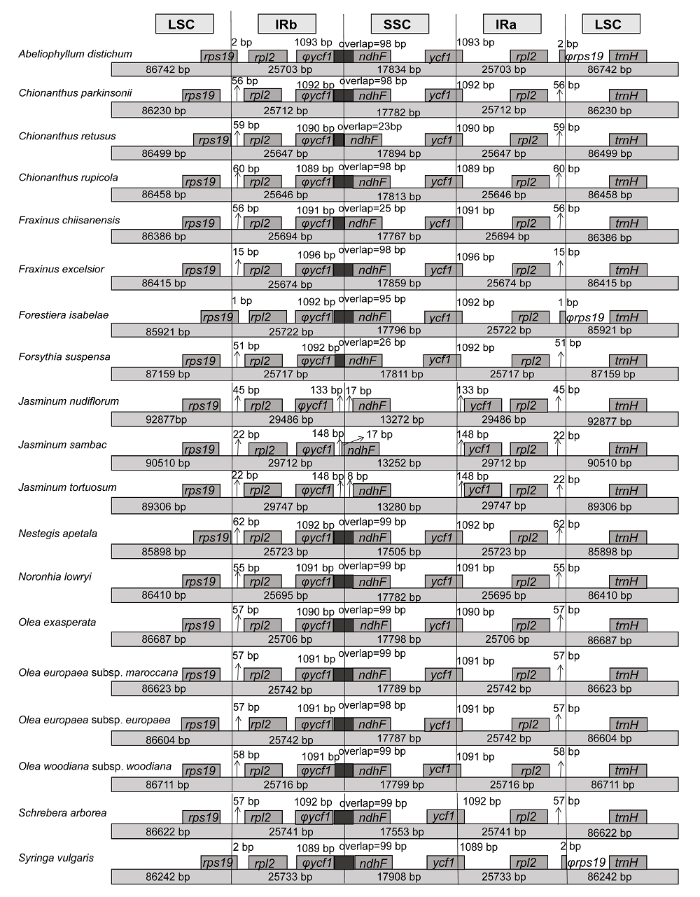 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}