Research Advances in Plant Science in China in 2018
Qian Qian1, Xiaoquan Qi2, Rongcheng Lin2, Shuhua Yang3, Aiwu Dong4, Jianru Zuo5, Fan Chen5, Langtao Xiao6, Hongya Gu7, Zhiduan Chen2, Yongfei Bai2, Xiaojing Wang8, Lei Wang2, Liwen Jiang9, Kang Chong,2,*, Tai Wang,2,*1 China National Rice Research Institute, Hangzhou 310006, China 2 Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 3 College of Biological Sciences, China Agricultural University, Beijing 100094, China 4 School of Life Sciences, Fudan University, Shanghai 200433, China 5 Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing 100101, China 6 College of Bioscience and Biotechnology, Hunan Agricultural University, Changsha 410128, China 7 College of Life Sciences, Peking University, Beijing 100871, China 8 College of Life Sciences, South China Normal University, Guangzhou 510631, China 9 The Chinese University of Hong Kong, Shatian, China
Abstract Plant science in China continues on the track of rapid development in 2018, with many remarkable achievements and a marked increase in number of original papers published in international top journals. The achivement “regulating the plant growth-metabolism balance to achieve sustainable agricultural development” was selected in the top ten progress of Chinese science in 2018, and “the history of angiosperm flora evolution in China” in the top ten progress of Chinese life science in 2018. Studies in rice and fruit and vegetable fields has been internationally leading. In this review, we summarize the significant progress in Chinese plant science in 2018, review the latest findings and hot events, and share the great achievements made by Chinese scientists. Keywords:China;plant science;research advances;2018
PDF (2396KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 钱前, 漆小泉, 林荣呈, 杨淑华, 董爱武, 左建儒, 陈凡, 萧浪涛, 顾红雅, 陈之端, 白永飞, 王小菁, 王雷, 姜里文, 种康, 王台. 2018年中国植物科学若干领域重要研究进展. 植物学报, 2019, 54(4): 405-440 doi:10.11983/CBB19165 Qian Qian, Qi Xiaoquan, Lin Rongcheng, Yang Shuhua, Dong Aiwu, Zuo Jianru, Chen Fan, Xiao Langtao, Gu Hongya, Chen Zhiduan, Bai Yongfei, Wang Xiaojing, Wang Lei, Jiang Liwen, Chong Kang, Wang Tai. Research Advances in Plant Science in China in 2018. Chinese Bulletin of Botany, 2019, 54(4): 405-440 doi:10.11983/CBB19165
新窗口打开|下载原图ZIP|生成PPT 图12016-2018年中国科学家在植物科学主流期刊Molecular Plant、The Plant Cell、Plant Physiology和The Plant Journal上发表论文的研究主题(数据来源: Web of Science, 绘制时间: 2019年7月2日)
使用VOSviewer软件对2016-2018年中国科学家在这4个学术期刊发表的研究论文进行关键词共现聚类。(A) 2016年的研究主题: 拟南芥蛋白激酶及盐胁迫下质膜依赖性内吞作用(蓝色); 水稻基因差异表达和编码基因预测(玉米和大麦)及基因组共线性和DNA甲基化与脱甲基化(红色); 丁香假单胞菌诱导的细胞死亡、系统获得性抗性等抗病性及植物免疫防御反应(橘色); 物质代谢关键酶(如交替氧化酶和合酶复合物)、NO对氧化胁迫的调节、PSII循环电子流状态转换及三角状五肽重复蛋白研究(绿色); 转录因子的功能及生物钟和根发育研究(黄绿色); 花发育和花器官特征及信号通路(淡粉色)。(B) 2017年的研究主题: 植物先天免疫、免疫激活、抗病性和防御机制(蓝色); PSI和PSII的叶绿素荧光, 线粒体、叶绿体基因家族及相关突变体(红色); 水稻基因组、相关代谢和自然变异, RNA序列的基因表达、进化发育与DNA甲基化(绿色); 拟南芥盐胁迫(黄绿色); 植物生长发育与水分利用效率(淡粉色)。(C) 2018年的研究主题: 丁香假单胞菌诱导的细胞死亡、抗病性鉴定及基因家族对逆境的响应(蓝色); 多倍体物种的进化和驯化、水稻开花时间和自然变异及玉米和大麦等的基因组, 小麦(节节麦)的遗传多样性, 序列的共线性(红色); 光合作用的类囊体膜(PSI和PSII)蛋白、体内磷酸化作用、同源重组及三酰甘油积累(绿色); 植物生长发育及其负调节子、叶绿素的生物合成与激素(生长素和脱落酸)蛋白(紫色); 拟南芥质膜蛋白及细胞组织动态(黄绿色)。 Figure 1Research themes of Chinese scientists published in major journals of plant sciences (Molecular Plant, The Plant Cell, Plant Physiology and The Plant Journal) from 2016 to 2018 (data sources: Web of Science; data collected by July 2, 2019)
Keyword co-occurence cluster of papers published by Chinese scientists in four plant science journals from 2016 to 2018 was constructed by VOSviewer. (A) The themes of 2016: Arabidopsis protein kinases and plasma membrane-dependent endocytosis under salt stress (blue); differentially expressed genes in rice and coding genes prediction (maize and barley), genome colinea- rity and DNA methylation and demethylation (red); Pseudomonas syringae induced cell death and systemic acquired resistance and immunity (orange); key enzymes of metabolism (such as alternative oxidase and synthase complexes), roles of NO in oxidative stress, cyclic electron flow of PSII and pentatricopeptide repeat (PPR) (green); transcription factors, circadian clock and root development (yellow-green); flower development and floral organ identity and signaling pathways (light pink). (B) The themes of 2017: plant innate immunity, immunity activation, disease resistance and defense mechanism (blue); chlorophyll fluorescence of PSI and PSII, mitochondria and chloroplast gene family and related mutants (red); rice genome, metabolism and natural variation, RNA-seq, evolution and DNA methylation (green); salt stress of Arabidopsis (yellow-green); plant growth and development and water use efficiency (light pink). (C) The themes of 2018: Pseudomonas syringae induced cell death, disease resistance recognition and gene family response on stress (blue); evolution and domestication of polyploid species, flowering time and natural variation of rice, genome (maize, barley, etc.), genetic diversity of wheat (Aegilops tauschii), and collinear analysis (red); thylakoid membrane protein of PSI and PSII; phosphorylation in vivo, homologous recombination and triacylglycerol accumulation (green); plant growth and development and negative regulators; biosynthesis of chlorophyll and hormone (auxin and abscisic acid) proteins (purple); Arabidopsis plasma membrane proteins and cell tissue dynamics (yellow green).
Table 1 表1 表12016–2018年中国与3个欧美国家在植物科学主流期刊(MP、PC、PP和PJ)的发文量比较(数据来源: Web of Science核心合集) Table 1The number of plant science publications originating from four countries (China, America, Germany and France) from 2016 to 2018, based on four plant science major journals (MP, PC, PP and PJ) (data sources: Web of Science)
2016年
2017年
2018年
文章数量 所占比例(%)
文章数量 所占比例(%)
文章数量 所占比例(%)
美国
369
36
373
35
366
35
中国
286
28
329
31
369
36
德国
198
19
208
19
180
17
法国
134
13
113
11
89
9
总载文量
1031
1072
1032
MP: Molecular Plant; PC: The Plant Cell; PP: Plant Physiology; PJ: The Plant Journal
2018年, 植物科学的亮点成果主要体现在如下6个方面。一是水稻生物学研究取得新突破。傅向东研究组发现水稻生长调节因子GRF4与生长抑制因子DELLA相互间的反向平衡赋予了植物生长与碳-氮代谢间的稳态共调节, 揭示了提高“绿色革命”品种的氮肥利用效率并增加谷物产量的遗传机制(Li et al., 2018g)。这一重大发现预示着一场新的绿色革命即将到来, 该成果入选2018年度中国科学十大进展。万建民研究组与相关单位合作, 运用自私基因模型揭示了水稻杂种不育现象, 探讨了“毒性-解毒”分子机制在水稻杂种不育中的普遍性, 揭示了自私基因在维持植物基因组的稳定性和促进新物种形成中的重要作用; 研究结果为创制“广亲和”材料, 克服杂种不育障碍, 充分利用杂种优势和野生种质资源, 提高水稻单产奠定了基础(Yu et al., 2018)。
二是植物系统进化研究取得了系列突破性成果。陈之端研究组重建了中国被子植物生命之树, 揭示了中国被子植物系统发育多样性形成的时空格局, 并明确了属、种水平分别应该重点保护的热点地区, 对我国保护区建设意义重大(Lu et al., 2018a)。该成果入选2018年度中国生命科学十大进展。凌宏清、梁承志、王道文和张爱民研究组完成了乌拉尔图小麦材料G1812的基因组测序和精细组装, 绘制出小麦A基因组7条染色体的分子图谱(Ling et al., 2018)。研究成果对进一步推动栽培小麦的遗传改良具有重要的理论意义和实用价值。叶凯研究组破译了罂粟基因组DNA密码, 揭示了其合成重要药用化合物的关键步骤, 这一发现为提高罂粟药用成分的产量及抗病性奠定了基础(Guo et al., 2018b)。黎志康研究组与相关单位合作, 深入探讨了水稻起源、分类以及驯化规律, 揭示了亚洲栽培稻的起源和群体基因组变异结构, 剖析了水稻核心种质资源的基因组遗传多样性(Wang et al., 2018o), 为水稻规模化基因发掘和复杂性状分子改良奠定了基础。
三是光生物学研究取得重要进展。李梅和章新政研究组解析了玉米PSI-LHCI-LHCII超级复合物3.3Å分辨率冷冻电镜结构(Pan et al., 2018)。该成果对人工光合作用体系的设计优化等应用研究均具促进作用。“中央研究院” (中国台湾)分子生物研究所李秀敏研究组揭示了叶绿体蛋白输入的新机制。他们以没有叶绿体内囊体的白色体为切入点, 发现了蛋白质穿越叶绿体外围双膜的连接桥梁——TIC236 (Chen et al., 2018j)。
四是植物抗病研究获得重要突破。陈学伟研究组与相关单位合作, 发现了水稻理想株型关键基因IPA1在水稻稻瘟病抗病过程中的作用, 揭示了IPA1既能提高水稻产量又能提高对稻瘟病抗性的分子机制(Wang et al., 2018f)。该研究打破了单个基因不可能同时实现增产和抗病的传统观点, 为高产高抗育种提供了实际应用新途径。
五是植物代谢组学研究取得重要进展。黄三文研究组与罗杰研究组合作, 利用多重组学大数据, 揭示了驯化和育种过程中番茄果实营养及风味物质的变化规律, 发现了调控这些物质的重要遗传位点(Zhu et al., 2018b)。这一成果不仅为番茄果实风味和营养物质遗传调控与全基因组设计育种提供了路线图, 而且为其它植物次生代谢资源的开发利用提供了重要思路。
六是农业绿色发展领域取得新成果。张福锁研究组证明绿色增产增效技术可大面积实现作物增产和环境减排双赢, 回答了持续增产是否必需依赖水肥资源的大量投入, 以及作物高产、养分资源高效和环境保护能否协同等国内外学术界一直争论的重大科学命题, 为中国农业走出一条产出高效、产品安全、资源节约和环境友好的现代化农业发展道路绘制了蓝图, 也为全球可持续集约化现代农业发展提供了范例(Cui et al., 2018b)。
目前, 中国科学家利用种间、亚种间和品种间杂交已先后培育了以D型等为代表的60余种水稻细胞质雄性不育系。D1型是一种新型孢子体细胞质雄性不育类型, 无花粉型败育, 且为中国所特有。颜龙安和蔡耀辉研究组成功克隆了D1型水稻细胞质雄性不育基因并揭示了其败育机理(Xie et al., 2018)。漆小泉研究组则发现了一种新型水稻雄性不育系, 并且阐释了OsOSC12/OsPTS1在水稻花粉包被形成过程中的关键作用, 对禾本科作物湿敏雄性不育材料的培育具有重要指导意义(Xue et al., 2018d)。
随着科学技术的发展, 基因组编辑技术开始应用于作物精准育种。然而由于缺乏高效去除含有Cas9表达盒及扩展性强、编辑效率高的单碱基编辑技术, 限制了其在分子育种中的应用。谢卡斌研究组利用生物体内的内含子剪切系统和tRNA加工系统开发出水稻中高效、多位点编辑且适用于CRISPR/Cpf1系统的新方法, 有望应用于动植物中(Ding et al., 2018a)。林宏辉和周焕斌研究组及朱健康研究组分别基于APOBEC1 酶的碱基编辑器及腺嘌呤的碱基编辑器(ABE7-10), 实现了水稻基因组4种不同碱基(A-G、T-C、C-T和G-A)的高效替换与水稻基因组特定位点的A·T碱基对高效转化为G·C碱基对, 不仅扩展了单碱基编辑技术在植物中的应用而且丰富了可用的单碱基编辑工具(Ren et al., 2018a; Hua et al., 2018)。赵云德研究组则利用自杀基因与CRISPR载体融合, 开发出高效的转基因自清除基因编辑系统(He et al., 2018d)。这些成果对深入研究水稻功能基因组具有重要推动作用。
1.2 水稻农艺及品质性状的遗传调控
水稻株型与产量密切相关。叶片作为水稻理想株型的组成要素之一, 其倾角对产量影响较大。薛红卫研究组新克隆了叶倾角调控基因LC3, 该基因编码1个含有SPOC结构域的转录抑制子, 其可与HIT锌指结构域蛋白LIP1互作, 通过调控生长素平衡和信号转导来调控植株叶倾角(Chen et al., 2018h)。李建雄研究组与田志宏研究组合作揭示了OsOFP19、OSH1与DLT蛋白可形成复合体, 调控BL信号转导及影响叶倾角发育(Yang et al., 2018b)。易可可研究组则解析了磷素有效性调控水稻叶倾角的分子机制。他们发现磷饥饿诱导的SPX1 (Syg1/Pho81/XPR1)与SPX2蛋白负调控叶倾角。SPX1的互作蛋白RLI1 (Regulator of leaf inclination 1)正调控叶倾角。RLI1可直接激活下游BU1 (Brassinosteroid upregulated 1)和BC1 (BU1-like1 complex 1)基因的表达以控制叶枕细胞的伸长, 从而使叶倾角增大(Ruan et al., 2018)。种康和徐云远研究组证实miR396d通过分别抑制其靶基因OsGRF4和OsGRF6的转录, 促进BL及抑制GA信号途径, 进而调控水稻叶夹角和株高(Tang et al., 2018)。邢永忠研究组则阐释了来自世界各地的水稻核心种质剑叶夹角的遗传基础, 发现bHLH转录因子家族中OsbHLH153和OsbHLH17能快速响应BL和IAA信号转导, 从而影响植株形态(Dong et al., 2018c)。王永红研究组揭示了水稻分蘖角度调控的分子机制。他们通过研究水稻茎重力反应过程中的动态转录组变化, 探索了一条水稻分蘖角度调控的核心途径(Zhang et al., 2018h)。该研究为解析水稻分蘖角度的调控网络进而挖掘有利用价值的基因提供了重要信息。谭禄宾研究组发现亚洲栽培稻PROG1基因相邻位置约有110 kb的染色体缺失。该缺失片段包含7个与PROG1基因相似的锌指基因, 其缺失推动了水稻由匍匐向直立转变, 最终实现产量大幅提高(Wu et al., 2018c)。
钱前研究组则报道了一种新的种质资源小薇。它具有生长周期短和株型矮小等表型, 可以像拟南芥一样在实验室内大规模种植。小薇的超矮秆表型是由于GA生物合成基因D18突变引起(Hu et al., 2018a)。该研究创制了“小薇日本晴”和“小薇93”, 为进行水稻功能基因组相关研究提供了重要资源。李来庚研究组与杨远柱研究组合作发现了一个新的特异调控水稻茎秆基部节长度的基因SBI, 该基因编码GA2氧化酶, 在培育水稻半矮秆性状, 提高抗倒伏能力, 增加水稻产量方面具重要的应用价值(Liu et al., 2018b)。周奕华研究组揭示了IIP4蛋白通过与NAC29/NAC31互作, 负向调控次生壁合成, 进而影响茎秆强度(Zhang et al., 2018b)。曹立勇和程式华研究组阐明了水稻叶片衰老的新机制。他们合作克隆了1个引起水稻叶片早衰的基因OsMTS1, 该基因编码褪黑素生物合成途径中的甲基转移酶, OsMTS1突变后可导致水稻叶片中褪黑素含量降低, 从而引起叶片早衰(Hong et al., 2018)。
丁勇研究组发现COMPASS-like复合体调控水稻抽穗期和花序结构, OsWDR5a和OsTrx1功能缺失可导致水稻花序二级枝梗减少及抽穗延迟(Jiang et al., 2018a)。此外, 他们还揭示了水稻SIP1招募OsTrx1到Ehd1基因调控开花时间的分子机制(Jiang et al., 2018b)。万建民和程治军研究组则解析了苹果酸抑制水稻穗顶部退化和OsALMT7转运苹果酸在水稻幼穗发育过程中起作用的分子机制, 为水稻高产品种的选育奠定了理论基础(Heng et al., 2018)。张大兵研究组与相关单位合作, 发现LOFSEP亚族基因OsMADS1、OsMADS34和OsMADS5共同调控花分生组织, 并正向调控其它MADS-box花的同源基因来确定小穗器官的特征(Wu et al., 2018a)。孙传清和朱作峰研究组对水稻驯化过程中二级枝梗数目转变的分子机制进行了研究, 发现栽培稻中FZP启动子的4 bp缺失影响了OsARFs的转录调控, 降低FZP的表达, 可提高水稻枝梗数和产量(Huang et al., 2018d)。林鸿宣和单军祥研究组则证实GSN1作为负调控因子调控水稻穗型发育的OsMKKK10-OsMKK4-OsMPK6级联信号通路。GSN1-MAPK通过整合下游植物激素信号, 精准调控水稻穗粒数和籽粒大小的协同发育(Guo et al., 2018c)。该研究为作物产量的遗传改良提供了新的分子模块与策略。
刘建祥和罗小金研究组、宋献军研究组、何予卿研究组同时克隆到与粒型和粒重相关的同一主效负调控QTL/基因qTGW3/TGW3/GL3.3, 并解析了相关分子机制(Hu et al., 2018c; Xia et al., 2018; Ying et al., 2018)。李云海研究组则与相关单位合作揭示了OsMKKK10-OsMKK4-OsMAPK6信号途径正向调控水稻粒型和粒重(Xu et al., 2018e); 并发现大粒基因LARGE8可与OsMAPK6直接互作使其失活, 进而抑制颖壳细胞增殖, 负向调控水稻粒型(Xu et al., 2018f)。余四斌研究组揭示了OsGBPs作为转录因子能够识别GAGA基序, 进而调控水稻粒长和植株高度(Gong et al., 2018)。梁国华研究组则证明OsSPMS1基因负向调控水稻种子萌发、粒型和单株产量, 是水稻增产育种的重要靶标基因(Tao et al., 2018)。刘巧泉研究组发现了水稻粒型与外观品质调控基因GS9, 其编码蛋白可与OsOFP14和OsOFP8蛋白互作, 通过改变细胞分裂调控水稻粒型(Zhao et al., 2018c)。傅向东研究组克隆了1个控制水稻产量和提升稻米品质的重要基因LGY3, 该基因编码转录因子OsMADS1。可变剪接蛋白OsMADS1lgy3可提高稻米粒长, 降低垩白率, 进而影响稻米产量和外观品质(Liu et al., 2018e)。
储成才研究组克隆了穗发芽调控基因PHS8, 该基因突变可导致胚乳中小分子糖积累, 从而抑制ABA信号通路中OsABI3和OsABI5的表达, 导致穗发芽表型(Du et al., 2018b)。刘春明研究组与相关单位合作创建了一种半粒种子筛选体系, 并利用该体系筛检获得了1个糊粉层增厚的水稻品系ta2。基因克隆发现DNA去甲基化酶基因OsROS1与糊粉层的发育相关, 有望开拓水稻营养品质育种新路径(Liu et al., 2018d)。
2 激素生物学
2.1 生长素
生长素在植物生长发育过程中起重要作用。赵忠研究组发现拟南芥茎顶端分生组织中的生长素具有促进干细胞分化的功能, 并维持干细胞的稳态, 揭示了生长素维持干细胞数目稳定的新机制(Luo et al., 2018b)。焦雨铃研究组与张磊研究组合作, 通过学科交叉研究发现了植物侧生器官可反馈调控茎尖干细胞, 并表明生长素长距离运输在反馈中发挥了重要作用(Shi et al., 2018a)。该研究有机整合了生物学与数学, 发挥了学科交叉研究解决动态生物学过程等前沿问题的优势。胡玉欣研究组对生长素启动愈伤组织形成的分子机制进行了探索, 揭示了控制愈伤组织形成中体细胞重编程的关键因子, 并建立了植物再生体系中生长素信号与细胞全能性获得的分子联系(Xu et al., 2018a)。该研究为探索植物再生的分子调控机制和发育程序提供了新线索。
2.2 芸薹素与脱落酸
芸薹素(BL)是植物特有的甾醇类激素。白明义研究组发现过氧化氢能氧化BL信号途径中的关键调控因子BZR1和BES1, 增强BZR1的转录活性, 从而促进植物的细胞伸长(Tian et al., 2018b)。该研究揭示了H2O2与BL协同调控植物生长发育的新机制, 为探索H2O2在植物中的功能提供了新的技术途径。李云海研究组与相关单位合作, 发现BL共受体BRI1和BAK1的互作与磷酸化受糖浓度调控。相关生化分析表明, BRI1和BAK1不仅可与G蛋白亚基互作, 还可磷酸化G蛋白亚基, 为BL途径与糖信号途径协同调控植物的生长发育建立了联系(Peng et al., 2018b)。许卫锋研究组与张建华研究组合作, 发现BL受体BRI1能与质子泵AHA2蛋白互作, 进而促进根系的向水性(Miao et al., 2018b)。
在苹果育种中, 矮化砧的选育具有重要意义。孔瑾研究组发现WRKY9在苹果矮化砧中高表达, 并直接抑制BL合成酶DWF4的转录从而减少BL合成, 介导植株矮化(Zheng et al., 2018c)。该研究揭示了在木本植物中WRKY9通过调控BL水平进而促进矮化的可能机制。
脱落酸(ABA)是控制植物生长和胁迫反应的关键激素。朱健康研究组发现水稻中ABA受体基因家族的特异受体基因突变导致胁迫反应减弱, 但在自然条件下表现出比野生型更好的生长状态(Miao et al., 2018a)。该研究为培育高产优质水稻品种提供了新思路。林宏辉和张大伟研究组发现, 拟南芥转录因子基因HAT1及其同源基因负调控ABA的生物合成, 从而参与调控对干旱胁迫的响应(Tan et al., 2018b)。该研究揭示了HAT1一种新的负调控机制, 对理解植物耐旱响应具有重要意义。
2.3 乙烯和茉莉素
乙烯(ETH)是重要的植物激素, 其对植物生长发育及果实成熟具重要调控作用。陈受宜和张劲松研究组对水稻乙烯不敏感系列突变体mhz2和mhz3进行了深入分析, 发现MHZ3可通过与乙烯信号通路关键调控因子OsEIN2的Nramp-like结构域结合稳定OsEIN2, 抑制其泛素化, 从而正调控乙烯信号转导(Ma et al., 2018a)。而MHZ2/SOR1编码一个E3泛素连接酶, 该酶可直接调控生长素信号通路中的转录抑制子OsIAA26降解, 介导乙烯对根生长的抑制效应(Chen et al., 2018b)。孙其信研究组则发现小麦TaWRKY51能结合到ETH合成基因TaACS的启动子区并抑制其表达, 进而促进侧根形成(Hu et al., 2018d)。
EBF和EIL均是乙烯信号转导的关键蛋白, 刘明春研究组发现番茄中SlEBF3基因受SlEIL调控, 过表达SlEBF3可使SlEIL含量降低, 并表现出ETH响应相关表型(Deng et al., 2018)。该研究揭示了一种新的番茄EBF基因在跃变型果实成熟中的作用, 为肉质果实成熟调控提供了基因资源。郝玉金研究组则发现苹果中MdEIL1能结合MdMYB1的启动子并激活其表达, 而MdMYB1能结合MdERF3的启动子调控乙烯的合成, 促进花青素积累与果实着色(An et al., 2018b)。
茉莉素(JA)是一类来源于环脂肪酸的植物激素, 可调节植物的防御反应和生长发育。谢道昕研究组发现健康植株中JJW (JAV1-JAZ8-WRKY51)复合体能够抑制JA合成基因的表达, 虫咬触发Ca2+/CAM依赖的JAV1磷酸化并促使JJW复合体解离而解除抑制, 进而促进JA合成并产生抗性(Yan et al., 2018a)。此外, 该研究组还发现COI1可识别JA的动态变化。COI1先识别JA形成COI1-JA复合体, 之后结合JAZs形成三元复合体, 进而促进JA的信号转导(Yan et al., 2018b)。
2.4 其它植物激素
一氧化氮(NO)是生物体内一种重要的信号分子, 参与调控众多的生物学过程。左建儒研究组发现, NO介导的亚硝基化修饰拟南芥GSNOR1, 导致其通过自噬途径被降解, 直接调控种子萌发过程中的低氧胁迫反应(Zhan et al., 2018)。该研究揭示了拟南芥中GSNOR1蛋白在低氧胁迫时发生选择性自噬降解的新机制。
赤霉素(GA)在植物发育的许多方面起关键作用。侯兴亮研究组发现GA信号抑制子DELLA可与胚胎发育晚期的关键调控因子LEC1 (LEAFY COTYLEDON 1)直接相互作用, 而GA介导的DELLA降解解除了其对LEC1的抑制, 从而促进胚胎发育。该研究揭示了GA在拟南芥胚胎发育晚期的作用机制(Hu et al., 2018b)。
2.5 植物激素互作与调控网络
植物激素之间存在复杂的调控网络。杨淑华研究组发现ABA信号关键蛋白激酶SnRK2s通过磷酸化细胞分裂素负调节子A型ARR5正调控其蛋白稳定, 揭示了植物激素信号调控网络在植物逆境响应胁迫中的精密调控机制(Huang et al., 2018c)。刘西岗研究组发现细胞分裂素的内稳态对于花分生组织(floral meristem, FM)的维持及分化非常重要。生长素可以通过促进ARF3 (Auxin Response Factor 3)的表达抑制细胞分裂素的活性进而促进FM的分化(Zhang et al., 2018e)。该研究揭示了转录因子AG通过植物激素信号途径协同调控花分生组织干细胞稳态的机理。王小菁研究组发现非洲菊锌指蛋白GhWIP2通过抑制GA和IAA信号及激活ABA信号, 进而抑制细胞生长并最终抑制花瓣伸展(Ren et al., 2018b)。该研究揭示了GA、ABA和生长素3种激素交叉互作网络共同调控非洲菊细胞伸展的作用机理。向成斌研究组发现乙烯响应基因HB52是乙烯和IAA信号互作的重要节点, 通过上调PIN2、WAG1和WAG2的表达抑制主根伸长(Miao et al., 2018c)。该研究揭示了乙烯与IAA交互作用的新机制。丁兆军研究组研究表明, BL合成途径突变体det2-9中乙烯合成大幅增加并积累了大量的超氧阴离子。该研究揭示了BL、乙烯与活性氧协同调控根生长的新机制(Lv et al., 2018)。ABA和BL信号通常相互拮抗。王学路研究组发现ABI1和ABI2通过与BIN2互作使其去磷酸化, 进而调控其对BES1的磷酸化(Wang et al., 2018c)。同时, 该研究组还发现BL信号途径中的GSK3为控制水稻中胚轴伸长的一个关键因子, 其通过磷酸化调控一类细胞周期蛋白CYC U2的稳定性促进细胞分裂。BL以及独脚金内酯(strigolactone)信号通路协同调控GSK3介导的细胞周期进程, 进而促进中胚轴伸长(Sun et al., 2018c)。
可变剪接是生物体内普遍存在的现象, 但是关于可变剪接是否也参与调控植物响应养分胁迫一直不很清楚。郑录庆研究组对不同矿质元素(Fe、Zn、Cu、Mn和P)缺乏条件下的水稻RNA-Seq数据进行系统分析, 结果表明可变剪接调控是植物响应营养缺乏的重要调控机制之一。丝氨酸精氨酸丰富(SR)蛋白是参与pre-mRNAs剪接的重要RNA结合蛋白。研究发现3个SR蛋白可能参与调控水稻对磷的吸收, 以及磷在叶片和地上部的再分配(Dong et al., 2018a)。该研究首次揭示了可变剪接在植物响应养分缺乏中的重要作用, 为该领域提供了新的研究思路。
3.4.3 重金属胁迫
镉是有毒重金属元素和致癌物, 土壤中的镉能够被农作物吸收进入农产品中, 从而对食品安全造成威胁。水稻作为重要粮食作物, 降低其籽粒中镉的积累对于保障食品安全至关重要。龚继明研究组与钱前研究组合作通过QTL定位找到了特异控制水稻叶片镉积累的一个重要基因CAL1 (Cadmium accumulation in leaf 1)。该基因主要在水稻根外皮层和木质部薄壁细胞中表达, CAL1在细胞质中螯合镉并将镉外排到细胞外, 从而降低细胞中镉的浓度, 驱动镉通过木质部导管长途转运。CAL1不影响籽粒中镉的积累且对其它必需元素也无显著影响(Luo et al., 2018a)。该研究为培育具有修复型和镉低积累的水稻新品种提供了理论依据。
4 发育、代谢与生殖生物学
4.1 植物发育生物学
茸毛作为植物体最外层结构, 具有很好的抗虫防病毒作用。叶志彪和杨长宪研究组分离了控制番茄腺体毛形成的关键基因H (Hair), 该基因编码C2H2锌指蛋白, 可能是茄科植物多细胞腺体毛形成的关键保守基因(Chang et al., 2018)。唐克轩研究组发现R2R3 MYB第9亚家族的转录因子AaMIXTA1参与调控青蒿分泌型腺毛发育, 为研究其它植物多细胞腺毛奠定了基础(Shi et al., 2018c)。此外, 他们还发现HD-ZIP IV转录因子AaHD8与AaMIXTA1相互作用形成复合体, 协同调控AaHD1和多个蜡质角质合成酶基因的表达, 从而促进蜡质合成和腺毛的起始发育(Yan et al., 2018d)。陈晓亚研究组与相关单位合作对可纺纤维形成的分子机制进行研究, 发现编码bHLH转录因子的棉花PRE1是可纺纤维进化的目标基因, 其正调控纤维伸长。PRE1基因启动子的变异可促进棉花可纺纤维的形成(Zhao et al., 2018a)。李学宝研究组鉴定了富含半胱氨酸的GhXLIM6蛋白, 其可与GhKNL1启动子的PAL-box结合而抑制该基因表达, 促进纤维素合成, 进而调控棉纤维伸长(Li et al., 2018i)。俞嘉宁研究组则揭示了棉花线粒体Ghatp1基因C1292和C1415位点发生RNA编辑可影响ATPase产生ATP, 从而促进表皮毛及纤维伸长(He et al., 2018c)。根毛是一种顶端生长的植物细胞, 微丝骨架对根毛的生长发育有极其重要的调控作用。贾红磊研究组揭示了H2S通过硫巯基化修饰扰乱正常的微丝骨架聚合及生长模式, 从而影响植物根毛的生长, 为进一步探索H2S调控植物生长发育的作用机制提供了理论依据(Li et al., 2018d)。
植物分枝是影响植物株型的关键因素。秦根基研究组与相关单位合作对植物分枝形成的分子调控机制进行研究, 发现了1个分枝正调控因子TIE1, 它与分枝重要调控因子BRC1 (BRANCHED1)发生互作, 抑制其转录活性。通过调节HB21、HB40和HB53等BRC1下游基因的转录, 发挥促进腋芽分枝的作用(Yang et al., 2018h)。开花是植物由营养生长转向生殖生长的标志。王前和张小兰研究组对热诱导开花的分子机理进行探索, 证明LsSOC1是热诱导花芽形成的激活子, 热激转录因子HsfA1e和HsfA4c通过结合在LsSOC1基因的启动子上, 影响其表达, 为开花相关机制研究奠定了理论基础(Chen et al., 2018k)。果实成熟过程对于果实品质形成和种子传播非常关键。陈建业和邝健飞研究组发现转录因子MaMYB3通过抑制淀粉降解相关基因以及MabHLH6转录因子的转录水平来调控淀粉降解, 丰富了香蕉果实成熟的转录调控网络(Fan et al., 2018)。朱本忠和罗云波研究组则发现, 在番茄rin突变体中, 基因组DNA片段部分缺失导致形成RIN-MC片段的嵌合, 该嵌合能够调控许多成熟相关基因的表达, 从而调节果实成熟(Li et al., 2018h)。此外, 吴鸿研究组与相关单位合作阐明了拟南芥角果开裂的调控机制。他们通过对纤维素酶CEL6和半纤维素酶MAN7以及果胶酶ADPG2基因的一系列突变体果荚离层进行细胞超微结构观察, 并结合基因表达的检测结果, 证明改变这些细胞壁酶的表达量和酶活性可以调控果荚开裂区细胞分化和离层区细胞降解(He et al., 2018a)。该研究为后期培育不同程度的果实抗开裂油料作物提供了技术支持。
大豆属于短日照植物, 对光周期极为敏感。李文滨研究组发现, 大豆中SKIP同源蛋白GmGBP1参与光周期介导的开花途径, 其启动子区域的自然变异调控大豆在不同环境中更早开花和成熟(Zhao et al., 2018d)。此外, 韩天富研究组发现, 大豆FT基因家族成员GmFT1a能够延迟大豆开花和成熟, 与开花促进基因GmFT2a/GmFT5a相互拮抗。基于本研究及前人的研究成果, 他们提出了FT基因家族精细调控大豆开花时间的跷跷板模型(Liu et al., 2018f)。生物钟是生物体适应环境昼夜周期变化而进化出的协调细胞内基因表达、代谢网络调控的分子系统。吴昌银研究组前期发现, OsELF3作为一个生物钟基因, 其昼夜表达相位是影响长日照条件下水稻抽穗期的关键因素。近期他们发现泛素化连接酶HAF1与OsELF3.1互作, 从而调控其蛋白昼夜节律性积累, 且单个氨基酸(L558S)变异影响二者的互作, 从而影响水稻的区域适应性(Zhu et al., 2018a)。王雷研究组则发现生物钟核心组分Evening Complex各组分的突变体叶片提前衰老。进一步研究发现, Evening Complex直接结合MYC2基因启动子并抑制其表达, 从而在时间维度精细调控茉莉酸诱导的植物叶片衰老进程(Zhang et al., 2018l)。
玉米胚乳是营养物质的主要储存组织。宋任涛研究组克隆了玉米突变体o11 (opaque11)的胚乳特异性bHLH转录因子O11。研究表明其不仅可直接调控胚乳发育的转录因子、cyPPDKs以及多个碳水化合物代谢酶, 还是ZmYoda的激活因子, 在胚乳发育和营养代谢调节网络中发挥中心调控作用(Feng et al., 2018); 另外, 该研究组还鉴定了1个与玉米储藏蛋白27 kDa γ-zein启动子相结合的bZIP型转录因子ZmbZIP22。该转录因子与PBF1、OHP1以及OHP2一起, 对27 kDa γ-zein进行转录调节, 影响胚乳发育(Li et al., 2018a)。
丁勇研究组则对拟南芥种子下胚轴发育进行研究, 发现参与拟南芥组蛋白修饰的MLK1和MLK2、GA信号途径负调控因子RGA, 以及与昼夜节律相关的CCA1形成蛋白复合体, 该复合体直接调控下游基因DWF4的表达, 进而影响下胚轴伸长(Zheng et al., 2018a)。李云海研究组以拟南芥为材料对控制植物器官大小的分子机制进行探索。他们发现E3连接酶F-盒组分SAP蛋白的2个底物KIX8和KIX9与PPD形成复合物行使功能, 其通过调控拟分生组织细胞增殖来控制器官大小(Li et al., 2018f)。玉米籽粒性状直接影响其产量。汤继华研究组与李文学研究组合作发现了1个控制玉米籽粒发育性状的基因ZmUrb2, 该基因主要通过影响核糖体的生物合成和核糖体rRNA前体加工来影响籽粒发育和整个营养生长过程(Wang et al., 2018d)。
4.2 植物代谢生物学
种子含油量是油菜的重要性状之一。郭亮研究组与相关单位合作对油菜籽中油脂合成的调控机制进行研究, 发现种子中脂质代谢相关基因在种子的不同部位及不同种子之间表达量存在显著差异。该研究对高油分油菜育种具有重要指导意义(Lu et al., 2018b)。王二涛研究组则对脂肪酸在植物和菌根真菌之间的转运调控机制进行探索, 发现在与丛枝菌根真菌共生过程中, 苜蓿转录因子WRI5a是脂肪酸碳源和磷营养交换的分子开关(Jiang et al., 2018e)。淀粉体中合成与储存的淀粉是谷物胚乳中的主要储能分子。万建民研究组与章文华研究组合作解析了淀粉体发育和淀粉合成的分子机制, 揭示了FSE1为磷脂酶样蛋白质, 其可调控水稻胚乳中半乳糖脂的合成, 为水稻胚乳发育过程中脂质代谢与淀粉合成之间的关系提供了新观点(Long et al., 2018)。
尼克酰胺腺嘌呤二核苷酸(NAD)从尼克酸(NA) 合成。目前已发现多种NA修饰(如糖基化)。王国栋研究组新发现一种NA修饰——甲酯化(MeNA)。MeNA是一种NAD前体, 可在植物不同组织间进行更长距离运输。该研究提出了植物中NAD生物合成与NA衍生物之间关系的新见解(Wu et al., 2018b)。萜类代谢物在植物生长发育等方面发挥重要作用。漆小泉研究组与相关单位合作发现一个新的、多产物OSC (2,3-氧化鲨烯环化酶) ——籼稻醇合酶(OsOS)。该酶合成一种新的五环三萜籼稻醇(orysatinol)的主产物及12种不同的三萜类化合物(Xue et al., 2018c)。王强研究组与相关单位合作发现玉米倍半萜合酶能直接催化(E, E)-法呢基二磷酸, 形成二羟基化的二醇产物(主要是eudesmane-2,11-diol) (Liang et al., 2018a)。该研究揭示了TPS的新型催化功能及形成二羟基化倍半萜的催化机制。
4-羟基-2,5-二甲基-3(2H)-呋喃酮(HDMF)是草莓果实的特征芳香组分, 其催化合成的关键酶是草莓醌氧化还原酶(FaQR)。陈昆松研究组发现乙烯应答因子(FaERF9)为FaQR启动子的正调节因子, 揭示了ERF-MYB复合物通过激活FaQR表达调节草莓中HDMF生物合成的分子机制(Zhang et al., 2018m)。苹果酸是苹果果实中的主要有机酸。韩振海和张新忠研究组获得了4个与苹果酸相关的主效QTLs; 并证明候选基因MdSAUR37等存在上位效应, 共同调控苹果酸的积累(Jia et al., 2018a)。草酸是最简单的二元酸, 在调控金属胁迫等方面起积极作用。巫永睿研究组克隆了玉米草酸降解途径中的草酰辅酶A基因, 阐明了玉米草酸代谢的前两步反应, 揭示了草酸代谢参与籽粒储藏物质积累和营养品质形成的分子机理, 为遗传改良草酸含量较高的蔬菜提供了候选基因(Yang et al., 2018e)。
类胡萝卜素是自然界广泛分布的一类次生代谢物。王涛涛研究组发现了番茄类胡萝卜素合成调控的关键转录因子SlBBX20, 其正调控番茄中类胡萝卜素的积累且稳定性受到泛素化调控(Xiong et al., 2019)。邓秀新研究组则获得一个调控类胡萝卜素合成途径关键基因LCYb的转录因子CsMADS6, 并证明其可通过直接调控类胡萝卜素代谢基因LCYb1等的表达, 协同正调控类胡萝卜素代谢(Lu et al., 2018c)。刘耀光研究组将双基因(sZmPSY1和sPaCrtI)、三基因(sZmPSY1、sPaCrtI和sCrBKT)和四基因(sZmPSY1、sPaCrtI、sCrBKT和sHpBHY)聚合转化水稻, 分别获得了筛选标记删除的富含黄色β-胡萝卜素、橙红色的角黄素和虾青素大米新种质(Zhu et al., 2018c)。此外, 陈学森研究组鉴定了PA1型 MYB转录因子MdMYBPA1。其在正常条件下能促进苹果原花青素的合成, 但在低温胁迫下促进花青苷的合成(Wang et al., 2018i)。
叶绿体是植物进行光合作用的细胞器, 其分裂由众多因子形成的复合物协助完成。张敏研究组发现MCD1在此过程中起重要作用。MCD1通过与ARC6作用, 识别FtsZ环, 并引导Min复合物整合到分裂环上, 从而调控叶绿体分裂(Chen et al., 2018e)。欧阳敏研究组则发现ECD1蛋白通过参与叶绿体基因rps14的RNA编辑调控叶绿体早期发育(Jiang et al., 2018d)。此外, 郁飞研究组和黄继荣研究组分别发现细胞质和叶绿体的翻译过程协同调控叶绿体发育, 及thf1突变体斑叶形成与质体翻译功能高度相关(Wang et al., 2018l; Huang et al., 2018b)。类囊体是叶绿体基质中由单位膜围成的扁平囊, 是光合作用光反应的场所。Kang Byung-Ho研究组揭示了类囊体蛋白复合体的组装以及CURT1和FZL蛋白在类囊体生物发生过程中的关键作用(Liang et al., 2018d)。隋森芳研究组与相关单位合作解析了红藻光系统I核心与捕光天线复合物(PSI-LHCR)的3.63?分辨率的三维结构, 发现红藻PSI核心既有蓝藻又有高等植物的特征(Pi et al., 2018)。该研究对阐明PSI的进化和功能具有重要意义。
5.2 光形态建成和信号转导
5.2.1 光受体调控机制
光对植物不仅是一种能源, 也是一种重要的环境信号。植物通过向光素(phot)、隐花色素(cry)和光敏色素(phy)等光受体来感应光信号。林金星研究组研究了蓝光对细胞质膜上向光素phot1的定位变化和动态性质, 在单分子水平上揭示了蓝光信号传递的起始事件及调控机制(Xue et al., 2018b)。刘宏涛研究组发现了隐花色素互作转录因子CIB1通过直接结合cry2和CO来介导cry2-CIB1-CO复合体的形成, 揭示了蓝光信号调控植物开花时间的新机制(Liu et al., 2018g)。杨洪全研究组发现, G蛋白β亚基AGB1通过与HY5互作抑制其DNA结合能力, 从而抑制光信号; 而蓝光信号通过CRY1与AGB1互作促进AGB1与HY5解离来抑制AGB1的功能(Lian et al., 2018); 并发现CRY1与去磷酸化形式的BL信号关键转录因子BES1发生依赖于蓝光的直接互作, 揭示了植物能根据光信号和内在BL信号有无或强弱的动态变化来优化其光形态建成(Wang et al., 2018n)。此外, 该研究组还发现光受体cry1、phyB竞争性与生长素信号通路抑制子Aux/IAA结合, 并磷酸化后者而负调控生长素信号转导, 进而精确调控植物光信号与生长素信号通路的平衡, 优化其生长(Xu et al., 2018c)。
李继刚和康定明研究组则筛选获得了远红光信号转导新组分TZP。TZP可与phyA和FHY1互作, 且在远红光下调控phyA、FHY1和HY5的蛋白水平, 并参与调控phyA的蛋白磷酸化(Zhang et al., 2018k)。李继刚研究组还与邓兴旺研究组合作证明了拟南芥phyA铰链区的3个位点(S590、T593和S602)对phyA行使功能非常重要(Zhou et al., 2018c)。吴素幸研究组发现, phyA和cry感知远红光和蓝光信号, 使负调控因子COP1失活, 从而激活生长素途径在TOR依赖的RPS6磷酸化中的作用(Chen et al., 2018a)。此外, 李琳研究组发现在遮阴, 尤其是严重遮阴条件下, phyA蛋白量增加, phyA与Aux/IAA互作影响了生长素受体TIR与Aux/IAA的互作, 从而阻止TIR介导的Aux/IAA降解, 导致生长素信号减弱, 通过调控光信号与生长素信号的平衡, 进而调节植物的生长发育(Yang et al., 2018c)。周艳虹研究组鉴定了番茄在光抑制和光保护过程中的光信号依赖性调控通路, 分析了phyA依赖的HY5-ABI5-RBOH1信号通路在缓解冷诱导的光抑制以及光保护中的作用(Wang et al., 2018a)。
5.2.2 光形态建成
邓兴旺研究组与许冬清研究组合作鉴定了光形态建成的正调节因子CSU4, 它能整合生物钟节律和光信号(Zhao et al., 2018g)。他们还发现BBX21通过第2个B-box结构域与HY5启动子中的T/G-box结合, 调控HY5及其靶基因的表达以促进植物光形态建成(Xu et al., 2018b)。而BBX28通过与HY5蛋白的C末端互作, 抑制HY5与下游靶基因启动子结合, 从而抑制HY5活性及其下游靶基因表达, 最终抑制植物光形态建成(Lin et al., 2018a)。此外, 邓兴旺和钟上威研究组揭示了PIFs、HY5和EIN3及EIL1对出土幼苗形态建成的调控作用, 为阐明幼苗从暗形态建成转向光形态建成的分子机制提供了新证据(Shi et al., 2018b)。吕应堂研究组则鉴定了光形态建成的正调控因子SRS5, 它直接结合HY5、BBX21和BBX22的启动子并激活其表达, 从而促进光形态建成(Yuan et al., 2018)。刘宏涛研究组发现UV-B照射后, UVR8形成单体并在细胞核中富集, UVR8通过抑制BES1/BIM1结合DNA的能力从而抑制下游生长相关基因的表达, 并抑制细胞伸长及光形态建成(Liang et al., 2018b)。同时, 发现UV-B激活的UVR8形成单体, 进入细胞核直接结合WRKY36而抑制其结合HY5启动子, 进而抑制下胚轴伸长(Yang et al., 2018g)。
光照和温度是影响植物生长发育非常重要的两个环境因素。林荣呈研究组发现了光信号与温度信号整合的新因子SEU, 揭示了SEU通过与PIF4互作形成转录调控复合物, 对生长素合成及与信号相关的靶基因进行调节, 从而实现对生长发育的调控(Huai et al., 2018)。方玉达研究组发现, 拟南芥miRNA加工复合体成员DCL1/HYL1与PIF4存在直接互作。PIF4在黑暗/红光转换过程中调控DCL1的稳定性及部分miRNA基因的转录, 进而调控光形态建成(Sun et al., 2018e)。黎家研究组则发现TCP17蛋白通过调控PIF4/PIF5依赖以及非依赖的生长素合成, 从而调控遮阴诱导下植物下胚轴的伸长生长(Zhou et al., 2018d)。
DNA甲基化主要指在DNA甲基转移酶的作用下, 将供体S-腺苷甲硫氨酸上的甲基转移至DNA受体碱基上的过程。基因组全局性DNA甲基化水平需要通过DNA甲基化和去甲基化的动态平衡调节。钱伟强研究组揭示了拟南芥中2种碱基切除修复蛋白APE2 (APURINIC/APYRIMIDINIC ENDONUCLEASE 2) 和ZDP (ZINC FINGER DNA 3'-PHOSPHOESTERASE)在DNA损伤修复以及环境胁迫中的生物学功能。全基因组胞嘧啶甲基化(5mC)分析表明, 缺失APE2和ZDP基因的双突变体中存在与单突变体不同的2 000余个DNA高甲基化位点; 且双突变体表现出生长发育缺陷及对DNA烷基化试剂超敏感。该研究证明了APE2和ZDP在维持植物表观基因组和基因组稳定性中的重要作用(Li et al., 2018e)。
表观遗传变异可促进植物对环境变化的适应, 但目前尚缺乏天然的表观遗传等位基因贡献于适应性进化的证据。朱健康和张庆祝研究组鉴定了一个名为NMR19 (天然存在的DNA甲基化变异区域19)的逆转录转座子。研究表明, NMR19-4的甲基化可抑制PPH基因的表达, 进而控制植物叶片衰老。NMR19-4的甲基化修饰水平与旱季的温度呈负相关, 暗示NMR19- 4可能通过调节PPH的表达帮助拟南芥适应环境的改变(He et al., 2018b)。
6mA是原核生物基因组DNA最常见的修饰, 近年来6mA修饰在真核生物中的作用也被陆续报道。谷晓峰研究组与相关单位合作发现6mA修饰在拟南芥基因组中广泛存在, 且水平与活跃基因的表达呈现出相关性(Liang et al., 2018c)。而周道绣和曾志雄研究组对水稻基因组中的6mA修饰进行了分析, 发现位于启动子区域的6mA与基因表达抑制相关, 而位于转录区的6mA与基因激活相关。转座子上的6mA与5mC呈现互补性, 表明DNA腺嘌呤甲基化与胞嘧啶甲基化在水稻中可以协同作用, 共同调控基因的表达(Zhou et al., 2018a)。
6.4 非编码RNA
非编码RNA是指生物体内一类不具有编码功能的RNA, 包括microRNA以及LncRNA等。它们对基因表达有调控作用, 主要通过调节mRNA的稳定性或者参与DNA甲基化等过程影响个体的生长发育。MicroRNA是一类长度为19-22 nt的非编码小RNA, 可参与非生物胁迫响应。朱健康研究组发现, 敲低microRNA166导致水稻形态发生变化, 气孔导度降低, 蒸腾速率下降, 抗旱性增强, 表明miRNA可以介导水稻的抗旱性(Zhang et al., 2018d)。此外, 郑丙莲研究组揭示了植物精子携带的miRNA参与早期胚胎发育。证明了具有高丰度miR159的精细胞与中央细胞融合后, 能快速抑制中央细胞传递的miR159的靶基因MYB33/65, 进而起始胚乳的第1次核分裂(Zhao et al., 2018j)。麻锦彪研究组与相关单位合作解析了SDN1的N端结构域以及催化结构域与RNA底物复合物的晶体结构, 阐明了SDN1识别剪切多种RNA底物的工作机制, 并提出了SDN1在生物体内修剪小非编码RNA的分子模型, 为设计和优化用于实验室研究的更加稳定的小非编码RNA奠定基础(Chen et al., 2018c)。
LncRNA是生物体中一类长度大于200 bp的非编码RNA, 其通过多种方式参与基因表达调控等重要生命活动。戚益军和李景睿研究组系统鉴定并分析了拟南芥中大量lncRNA, 发现可以调节开花时间的NAT-lncRNA, 并阐明了其正向调控正义链基因转录的作用机制(Zhao et al., 2018h)。杨金水和苏伟研究组则在水稻中鉴定到一例与产量性状相关且起转录激活作用的lncRNA, 过表达lncRNA会改变LRK1基因组位点的组蛋白修饰状态, 使水稻增产(Wang et al., 2018p)。
7 细胞骨架与囊泡运输
7.1 细胞骨架系统及其调控
细胞骨架(cytoskeleton)系统主要包括微管(MTs)和微丝(F-actin), 其在调控植物生长发育和细胞形态建成等生理过程中具重要作用。然而, 人们对微管和微丝的调控机理及其影响植物形态发生发育机制的认识还十分有限。高等植物已进化出多种(不同大小、形状和功能)细胞类型, 如锥形花瓣表皮细胞, 然而调控该种细胞形态建成的分子机制目前尚不清楚。林德书研究组证实AN (ANGUSTIFOLIA)-ROS途径与KTN1 (katanin)共同作用, 通过参与调节微管骨架的排列方式, 进而调控花瓣锥形细胞发育(Dang et al., 2018)。黄善金研究组则发现, 拟南芥成蛋白AtFH3 (Formin 3)和AtFH5参与调节花粉管顶端肌动蛋白的聚合和阵列构建, 对极性花粉管快速生长起关键作用(Lan et al., 2018)。此外, 他们还发现拟南芥成蛋白AtFH2特异定位于胞间连丝, 水稻的部分Formin家族成员也定位于胞间连丝, 该研究揭示了拟南芥胞间连丝处微丝的含量及稳定性对调控胞间连丝通透性具重要作用(Diao et al., 2018)。毛同林研究组从植物细胞骨架与环境因子互作方面进行了探索, 揭示了植物细胞受到高盐胁迫后, 乙烯通过上调微管稳定蛋白WDL5的表达调控微管重组(Dou et al., 2018), 及微管去稳定蛋白MDP60参与光和乙烯调控的下胚轴伸长(Ma et al., 2018c)。孔照胜研究组则利用标记了微丝骨架(actin cytoskeleton)的稳定转基因蒺藜苜蓿, 首次在活细胞水平解析了根瘤发育不同阶段, 微丝骨架调控共生界面形成过程中的排布结构和动态模式, 为探索根瘤发育和生物固氮提供了细胞生物学依据(Zhang et al., 2019)。
7.2 囊泡运输
细胞内的囊泡运输是生命活动中极其复杂的动态生物学过程, 参与多种植物的生长发育及对环境的响应。ESCRT为内体蛋白分选转运复合体, 关于调节ESCRT和控制多囊泡体/液泡前体细胞器形成及成熟并维持膜蛋白稳态的因子一直未知。姜里文和沈锦波研究组发现了1个含有Bro1结构域的新植物特异负调控因子BRAF (BRo1-domain protein As FREE1 suppressor)蛋白, 并证明其在多囊泡体/液泡前体的腔内小泡形成和膜蛋白降解分选过程中起负调控作用(Shen et al., 2018)。此外, 姜里文研究组还发现拟南芥可溶性NSF附着蛋白受体Qc-SNARE蛋白BET12在病程相关蛋白1 (Pathogenesis-Related 1, PR1)运输中具重要调节作用, 为植物抗病研究奠定了理论基础(Chung et al., 2018)。
8 植物系统进化
8.1 分子进化、比较基因组学和进化发育生物学
早期陆生植物形态结构相对简单, 但环境适应能力强。特有基因对物种适应特殊环境发挥了重要作用。黄锦岭研究组对小立碗藓的hemerythrin基因进行了研究, 发现其起源于真菌向早期陆生植物的水平基因转移事件。该基因在小立碗藓中通过改变移码突变, 演化出1个双编码基因YAN/AltYAN, 该基因介导了小立碗藓的脱水耐受性(Guan et al., 2018)。该研究对揭示陆生植物如何适应特殊环境提供了新观点及实验证据。基因组学研究为揭示一些物种特异现象提供了很好的切入点。例如, 卷柏(Selaginella tamariscina)是古老的复苏植物, 干旱时能存活数十年, 遇水而荣, 与已测序的其同属植物江南卷柏(S. moellendorffii) (干旱敏感型)截然不同。陈士林和宋经元研究组破译了卷柏基因组, 绘制了高杂合度卷柏的基因组图谱, 并与已发表的江南卷柏基因组进行了比较, 揭示了卷柏耐旱和复苏机制(Xu et al., 2018g)。维管系统(木质部等)是植物在水生到陆生的进化过程中产生的。拟南芥和小立碗藓中特定的NAC转录因子调控木质部发育以及水分运输。景海春研究组与才宏伟研究组合作从甜高粱中鉴定到1个在驯化过程中受到选择、控制茎秆持汁性的Dry基因(编码一个NAC转录因子), 并发现其功能缺失是甜高粱茎秆富含汁液的重要原因(Zhang et al., 2018f)。该研究不仅对促进作物茎秆持汁表型的遗传改良具重要作用, 而且为作物育种获得籽粒及秸秆“双丰收”提供了可能。
种子休眠减弱是一个典型的“驯化综合征”相关农艺性状。由于其表型难以鉴定, 关于作物驯化中受到选择的控制种子休眠的基因报道极少。田志喜和储成才研究组与相关单位合作鉴定了1个控制大豆种皮绿色的基因。该基因在大豆驯化过程中受到选择, 且与大豆种子休眠减弱相关。进一步研究发现, 其同源基因在水稻驯化中同样与种子休眠减弱相关。此外, 拟南芥野生群体中不同单倍型在休眠特性上也存在明显差异(Wang et al., 2018g)。该研究对新物种驯化具有重要的指导意义。
比较基因组学和全基因组关联分析(GWAS)不仅能揭示栽培植物的起源和演化历史, 还可以为分子辅助育种提供重要参考。梅花(Prunus mume)是蔷薇科植物, 已有3000多年的栽培历史。张启翔研究组与相关单位合作完成了梅花全基因组重测序, 对几百株梅花品种的重要观赏性状进行了GWAS分析, 为揭示梅花的演化历史, 开展其分子标记辅助育种打下坚实的基础(Zhang et al., 2018j)。此外, 杜雄明研究组与相关单位合作对419份陆地棉核心种质的基因组进行了重测序, 并结合多种农艺性状进行了GWAS分析, 为人工驯化选择导致棉花遗传多样性降低提供了直接的分子证据(Ma et al., 2018e)。张献龙研究组与相关单位合作对四倍体棉花(陆地棉和海岛棉)进行了精细测序, 将陆地棉和海岛棉基因组的连续性分别提高了55倍和90倍(Wang et al., 2019); 且绘制了棉花二倍体和四倍体的三维基因组图谱, 鉴定了棉花基因组中大量的拓扑结构域(TAD), 揭示了三维基因组的进化与转录调控之间的关系(Wang et al., 2018h)。该研究不仅促进了棉花功能基因组研究, 而且增加了人们对植物三维基因组结构的认知。
玉米是一个具有高度基因组多态性和杂种优势的重要作物。由B73自交系和Mo17自交系杂交产生的后代曾在全球范围内广泛种植。赖锦盛研究组对Mo17基因组进行了精细测序分析, 并与B73进行比较, 显示两个基因组约有10%的基因在染色体上的排序存在非共线性现象, 至少20%的基因存在可能导致蛋白编码功能变化的序列改变(Sun et al., 2018d)。该研究对玉米自交系间能够形成特别显著的杂种优势原因提供了一个新解释。
栽培稻和普通野生稻(Oryza rufipogon)丰富的遗传多样性是水稻育种的主要来源。黄学辉研究组对66个来自不同水稻类群的栽培稻品种和野生稻株系的精细基因组进行了比较分析, 鉴定出水稻基因组中各类复杂的遗传变异, 并发现很多功能基因存在多种等位基因类型(Zhao et al., 2018e)。发达的根系是旱粳稻和热带粳稻适应干旱环境最典型的根系形态特征。李自超研究组与黎志康研究组合作, 将795份栽培稻种质资源的根系性状和330多万个高质量的SNP数据进行关联分析, 获得几十个候选基因; 对这些候选基因的序列进行分析, 发现几乎所有位点在野生稻中都以低频存在。因此, 与旱粳稻长根和粗根等性状相关的基因是在高山干旱环境中通过正向选择及聚合而固定下来的(Zhao et al., 2018i)。此外, 陈明生研究组对约1 500万年前分化的短花药野生稻(Oryza brachyantha)和栽培稻着丝粒周围序列进行了比较, 发现该区域倾向有更多的共线基因丢失(大多转移到其它区域)。进一步分析表明, 这种基因丢失可能受着丝粒染色质动态变化(如扩展或微易位)驱动(Liao et al., 2018)。
QiXP, KuoLY, GuoCC, LiH, LiZY, QiJ, WangLB, HuY, XiangJY, ZhangCF, GuoJ, HuangCH, MaH ( 2018). A well-resolved fern nuclear phylogeny reveals the evolution history of numerous transcription factor families Mol Phylogenet Evol 127, 961-977. [本文引用: 1]
QinJ, ZhouX, SunL, WangK, YangF, LiaoH, RongW, YinJ, ChenH, ChenX, ZhangJ ( 2018). The Xanthomonas effector XopK harbours E3 ubiquitin-ligase activity that is required for virulence New Phytol 220, 219-231. [本文引用: 1]
RenB, YanF, KuangY, LiN, ZhangD, ZhouX, LinH, ZhouH ( 2018 a). Improved base editor for efficiently inducing genetic variations in rice with CRISPR/Cas9-guided hyperactive hAID mutant Mol Plant 11, 623-626. [本文引用: 1]
RenGP, LiLF, HuangYH, WangYQ, ZhangWB, ZhengRY, ZhongCM, WangXJ ( 2018 b). GhWIP2, a WIP zinc finger protein, suppresses cell expansion in Gerbera hybrida by mediating crosstalk between gibberellin, abscisic acid, and auxin New Phytol 219, 728-742. [本文引用: 1]
RenR, WangH, GuoC, ZhangN, ZengL, ChenY, MaH, QiJ ( 2018 c). Widespread whole genome duplications contribute to genome complexity and species diversity in angiosperms Mol Plant 11, 414-428. [本文引用: 1]
RuanW, GuoM, XuL, WangX, ZhaoH, WangJ, YiK ( 2018). An SPX-RLI1 module regulates leaf inclination in response to phosphate availability in rice Plant Cell 30, 853-870. [本文引用: 1]
ShenJB, ZhaoQ, WangXF, GaoCJ, ZhuY, ZengYL, JiangLW ( 2018). A plant Bro1 domain protein BRAF regulates multivesicular body biogenesis and membrane protein homeostasis Nat Commun 9, 3784. [本文引用: 1]
ShiB, GuoX, WangY, XiongY, WangJ, HayashiKI, LeiJ, ZhangL, JiaoY ( 2018 a). Feedback from lateral organs controls shoot apical meristem growth by modulating auxin transport Dev Cell 44, 204-216. [本文引用: 1]
ShiH, LyuM, LuoYW, LiuSC, LiY, HeH, WeiN, DengXW, ZhongSW ( 2018 b). Genome-wide regulation of light-controlled seedling morphogenesis by three families of transcription factors Proc Natl Acad Sci USA 115, 6482-6487. [本文引用: 1]
ShiP, FuX, ShenQ, LiuM, PanQ, TangY, JiangW, LvZ, YanT, MaY, ChenM, HaoX, LiuP, LiL, SunX, TangK ( 2018 c). The roles of AaMIXTA1 in regulating the initiation of glandular trichomes and cuticle biosynthesis in Artemisia annua New Phytol 217, 261-276. [本文引用: 1]
SunMH, MaQJ, HuDG, ZhuXP, YouCX, ShuHR, HaoYJ ( 2018 b). The glucose sensor MdHXK1 phosphorylates a tonoplast Na+/H+ exchanger to improve salt tolerance Plant Physiol 176, 2977-2990. [本文引用: 1]
SunS, WangT, WangL, LiX, JiaY, LiuC, HuangX, XieW, WangX ( 2018 c). Natural selection of a GSK3 determines rice mesocotyl domestication by coordinating strigolactone and brassinosteroid signaling Nat Commun 9, 2523. [本文引用: 1]
SunSL, ZhouYS, ChenJ, ShiJP, ZhaoHM, ZhaoHN, SongWB, ZhangM, CuiY, DongXM, LiuH, MaXX, JiaoYP, WangB, WeiXH, SteinJC, GlaubitzJC, LuF, YuGL, LiangCZ, FenglerK, LiBL, RafalskiA, SchnablePS, WareDH, BucklerES, LaiJS ( 2018 d). Extensive intraspecific gene order and gene structural variations between Mo17 and other maize genomes Nat Genet 50, 1289-1295. [本文引用: 1]
SunZF, LiM, ZhouY, GuoTT, LiuY, ZhangH, FangYD ( 2018 e). Coordinated regulation of Arabidopsis microRNA biogenesis and red light signaling through Dicer-like 1 and phytochrome-interacting factor 4 PLoS Genet 14, e1007-247. [本文引用: 1]
TanF, LuY, JiangW, WuT, ZhangRY, ZhaoY, ZhouDX ( 2018 a). DDM1 represses noncoding RNA expression and RNA-directed DNA methylation in heterochromatin Plant Physiol 177, 1187-1197. [本文引用: 1]
TanW, ZhangD, ZhouH, ZhengT, YinY, LinH ( 2018 b). Transcription factor HAT1 is a substrate of SnRK2.3 kinase and negatively regulates ABA synthesis and signaling in Arabidopsis responding to drought. PLoS Genet 14, e1007336. [本文引用: 1]
TangY, LiuH, GuoS, WangB, LiZ, ChongK, XuY ( 2018). OsmiR396d affects gibberellin and brassinosteroid signaling to regulate plant architecture in rice Plant Physiol 176, 946-959. [本文引用: 1]
TianS, KouYX, ZhangZR, YuanL, LiDR, Lopez-PujolJ, FanDM, ZhangZY ( 2018 a). Phylogeography of Eomecon chionantha in subtropical China: the dual roles of the Nanling Mountains as a glacial refugium and a dispersal corridor BMC Evol Biol 18, 20. [本文引用: 1]
WanT, LiuZM, LiLF, LeitchAR, LeitchIJ, LohausR, LiuZJ, XinHP, GongYB, LiuY, WangWC, ChenLY, YangY, KellyLJ, YangJ, HuangJL, LiZ, LiuP, ZhangL, LiuHM, WangH, DengSH, LiuM, LiJ, MaL, LiuY, LeiY, XuW, WuLQ, LiuF, MaQ, YuXR, JiangZ, ZhangGQ, LiSH, LiRQ, ZhangSZ, WangQF, Van de PeerY, ZhangJB, WangXM ( 2018). A genome for gnetophytes and early evolution of seed plants Nat Plants 4, 82-89. [本文引用: 1]
AnJP, LiR, QuFJ, YouCX, WangXF, HaoYJ ( 2018 a). R2R3-MYB transcription factor MdMYB23 is involved in the cold tolerance and proanthocyanidin accumulation in apple Plant J 96, 562-577. [本文引用: 1]
AnJP, WangXF, LiYY, SongLQ, ZhaoLL, YouCX, HaoYJ ( 2018 b). EIN3-LIKE1, MYB1, and ETHYLENE RESPONSE FACTOR 3 act in a regulatory loop that synergistically modulates ethylene biosynthesis and anthocyanin accumulation Plant Physiol 178, 808-823. [本文引用: 1]
WangF, WuN, ZhangLY, AhammedGJ, ChenXX, XiangX, ZhouJ, XiaX, ShiK, YuJQ, FoyerCH, ZhouYH ( 2018 a). Light signaling-dependent regulation of photoinhibition and photoprotection in tomato Plant Physiol 176, 1311-1326. [本文引用: 1]
WangFP, ShiGX, NicholasO, YaoBQ, JiMF, WangWY, MaZ, ZhouHK, ZhaoXQ ( 2018 b). Ecosystem nitrogen retention is regulated by plant community trait interactions with nutrient status in an alpine meadow J Ecol 106, 1570-1581. [本文引用: 1]
BiGZ, ZhouZY, WangWB, LiL, RaoSF, WuY, ZhangXJ, MenkeFLH, ChenS, ZhouJM ( 2018). Receptor-like cytoplasmic kinases directly link diverse pattern recognition receptors to the activation of mitogen-activated protein kinase cascades inArabidopsis Plant Cell 30, 1543-1561. [本文引用: 1]
CaoL, WangS, VenglatP, ZhaoLH, ChengY, YeeSJ, QinY, DatlaR, ZhouYM, WangH ( 2018). Arabidopsis ICK/KRP cyclin-dependent kinase inhibitors function to ensure the formation of one megaspore mother cell and one functional megaspore per ovule PLoS Genet 14, e1007230. [本文引用: 1]
WangHJ, TangJ, LiuJ, HuJ, LiuJJ, ChenYX, CaiZY, WangXL ( 2018 c). Abscisic acid signaling inhibits brassinosteroid signaling through dampening the dephosphorylation of BIN2 by ABI1 and ABI2 Mol Plant 11, 315-325. [本文引用: 1]
WangHQ, WangK, DuQG, WangYF, FuZY, GuoZY, KangDM, LiWX, TangJH ( 2018 d). Maize Urb2 protein is required for kernel development and vegetative growth by affecting pre-ribosomal RNA processing New Phytol 218, 1233-1246. [本文引用: 1]
ChenJ, WangP, de GraafBHJ, ZhangH, JiaoH, TangC, ZhangS, WuJ ( 2018 d). Phosphatidic acid counteracts S-RNase signaling in pollen by stabilizing the actin cytoskeleton Plant Cell 30, 1023-1039. [本文引用: 1]
ChenL, SunB, GaoW, ZhangQY, YuanH, ZhangM ( 2018 e). MCD1 associates with FtsZ filaments via the membrane-tethering protein ARC6 to guide chloroplast division Plant Cell 30, 1807-1823. [本文引用: 1]
WangMJ, WangPC, LinM, YeZX, LiGL, TuLL, ShenC, LiJY, YangQY, ZhangXL ( 2018 h). Evolutionary dynamics of 3D genome architecture following polyploidization in cotton Nat Plants 4, 90-97. [本文引用: 1]
WangN, QuC, JiangS, ChenZ, XuH, FangH, SuM, ZhangJ, WangY, LiuW, ZhangZ, LuN, ChenX ( 2018 i). The proanthocyanidin-specific transcription factor MdMYBPA1 initiates anthocyanin synthesis under low-temperature conditions in red-fleshed apples Plant J 96, 39-55. [本文引用: 1]
ChenL, ZhaoY, XuS, ZhangZ, XuY, ZhangJ, ChongK ( 2018 f). OsMADS57 together with OsTB1 coordinates transcription of its target OsWRKY94 and D14 to switch its organogenesis to defense for cold adaptation in rice New Phytol 218, 219-231. [本文引用: 1]
WangQL, SunAZ, ChenST, ChenLS, GuoFQ ( 2018 k). SPL6 represses signaling outputs of ER stress in control of panicle cell death in rice Nat Plants 4, 280-288. [本文引用: 1]
ChenSH, ZhouLJ, XuP, XueHW ( 2018 h). SPOC domain-containing protein leaf inclination3 interacts with LIP1 to regulate rice leaf inclination through auxin signaling PLoS Genet 14, e1007829. [本文引用: 1]
ChungKP, ZengYL, LiYM, JiCY, XiaYJ, JiangLW ( 2018). Signal motif-dependent ER export of the Qc- SNARE BET12 interacts with MEMB12 and affects PR1 trafficking in Arabidopsis J Cell Sci 131, UNSP jcs 202838. [本文引用: 1]
CuiY, HuC, ZhuY, ChengK, LiX, WeiZ, XueL, LinF, ShiH, YiJ, HouS, HeK, LiJ, GouX ( 2018 a). CIK receptor kinases determine cell fate specification during early anther development in Arabidopsis Plant Cell 30, 2383-2401. [本文引用: 1]
WangY, LuoXJ, SunF, HuJH, ZhaXJ, SuW, YangJS ( 2018 p). Overexpressing lncRNA LAIR increases grain yield and regulates neighbouring gene cluster expression in rice Nat Commun 9, 3516. [本文引用: 1]
WangZ, WangFX, HongYC, YaoJJ, RenZZ, ShiHZ, ZhuJK ( 2018 r). The flowering repressor SVP confers drought resistance in Arabidopsis by regulating abscisic acid catabolism Mol Plant 11, 1184-1197. [本文引用: 1]
WeiDH, LiuMJ, ChenH, ZhengY, LiuYX, WangX, YangSH, ZhouMQ, LinJ ( 2018). INDUCER OF CBF EXPRESSION 1 is a male fertility regulator impacting anther dehydration in Arabidopsis PLoS Genet 14, e1007695. [本文引用: 1]
WuD, LiangW, ZhuW, ChenM, FerrandizC, BurtonRA, DreniL, ZhangD ( 2018 a). Loss of LOFSEP Transcription factor function converts spikelet to leaf-like structures in rice Plant Physiol 176, 1646-1664. [本文引用: 1]
WuRR, ZhangFX, LiuLY, LiW, PicherskyE, WangGD ( 2018 b). MeNA, controlled by reversible methylation of nicotinate, is an NAD precursor that undergoes long-dist-ance transport in Arabidopsis Mol Plant 11, 1264-1277. [本文引用: 1]
DengH, PirrelloJ, ChenY, LiN, ZhuSH, ChirinosX, BouzayenM, LiuYS, LiuMC ( 2018). A novel tomato F-box protein, SlEBF3, is involved in tuning ethylene signaling during plant development and climacteric fruit ripening Plant J 95, 648-658. [本文引用: 1]
DiaoM, RenSL, WangQN, QianLC, ShenJF, LiuYL, HuangSJ ( 2018). Arabidopsis formin 2 regulates cell- to-cell trafficking by capping and stabilizing actin filaments at plasmodesmata eLife 7, e36316. [本文引用: 1]
WuY, ZhaoS, LiX, ZhangB, JiangL, TangY, ZhaoJ, MaX, CaiH, SunC, TanL ( 2018 c). Deletions linked to PROG1 gene participate in plant architecture domestication in Asian and African rice Nat Commun 9, 4157. [本文引用: 1]
WuZY, LiuJ, ProvanJ, WangH, ChenCJ, CadotteMW, LuoYH, AmorimBS, LiDZ, MilneRI ( 2018 d). Testing Darwin’s transoceanic dispersal hypothesis for the inland nettle family (Urticaceae) Ecol Lett 21, 1515-1529. [本文引用: 1]
DingD, ChenK, ChenY, LiH, XieK ( 2018 a). Engineering introns to express RNA guides for Cas9- and Cpf1-media- ted multiplex genome editing .Mol Plant 11, 542-552. [本文引用: 2]
DongC, HeF, BerkowitzO, LiuJ, CaoP, TangM, ShiH, WangW, LiQ, ShenZ, WhelanJ, ZhengL ( 2018 a). Alternative splicing plays a critical role in maintaining mineral nutrient homeostasis in rice (Oryza sativa) Plant Cell 30, 2267-2285. [本文引用: 1]
DongH, BaiL, ZhangY, ZhangGZ, MaoYQ, MinLL, XiangFY, QianDD, ZhuXH, SongCP ( 2018 b). Modulation of guard cell turgor and drought tolerance by a pero-xisomal acetate-malate shunt .Mol Plant 11, 1278-1291. [本文引用: 1]
XiaD, ZhouH, LiuRJ, DanWH, LiPB, WuB, ChenJX, WangLQ, GaoGJ, ZhangQL, HeYQ ( 2018). GL3.3, a novel QTL encoding a GSK3/SHAGGY-like kinase, epistatically interacts with GS3 to produce extra-long grains in rice Mol Plant 11, 754-756. [本文引用: 1]
XieC, XieDF, ZhongY, GuoXL, LiuQ, ZhouSD, HeXJ ( 2019). The effect of Hengduan Mountains Region (HMR) uplift to environmental changes in the HMR and its eastern adjacent area: tracing the evolutionary history of Allium section Sikkimensia( Amaryllidaceae) Mol Phylogenet Evol 130, 380-396. [本文引用: 1]
DongHJ, ZhaoH, LiSL, HanZM, HuG, LiuC, YangGY, WangGW, XieWB, XingYZ ( 2018 c). Genome-wide association studies reveal that members of bHLH subfamily 16 share a conserved function in regulating flag leaf angle in rice (Oryza sativa) PLoS Genet 14, e1007323. [本文引用: 1]
DongWP, XuC, WuP, ChengT, YuJ, ZhouSL, HongDY ( 2018 d). Resolving the systematic positions of enigmatic taxa: manipulating the chloroplast genome data of Saxifragales Mol Phylogenet Evol 126, 321-330. [本文引用: 1]
XuF, HeSB, ZhangJY, MaoZL, WangWX, LiT, HuaJ, DuSS, XuPB, LiL, LianHL, YangHQ ( 2018 c). Photoactivated CRY1 and phyB interact directly with AUX/IAA proteins to inhibit auxin signaling in Arabidopsis Mol Plant 11, 523-541. [本文引用: 1]
XuN, ChuY, ChenH, LiX, WuQ, JinL, WangG, HuangJ ( 2018 d). Rice transcription factor OsMADS25 modulates root growth and confers salinity tolerance via the ABA- mediated regulatory pathway and ROS scavenging PLoS Genet 14, e1007662. [本文引用: 2]
XuR, DuanPG, YuHY, ZhouZK, ZhangBL, WangRC, LiJ, ZhangGZ, ZhuangSS, LyuJ, LiN, ChaiTY, TianZX, YaoSG, LiYH ( 2018 e). Control of grain size and weight by the OsMKKK10-OsMKK4-OsMAPK6 signaling pathway in rice Mol Plant 11, 860-873. [本文引用: 1]
DuL, XuF, FangJ, GaoSP, TangJY, FangS, WangHR, TongHN, ZhangFX, ChuJF, WangGD, ChuCC ( 2018 b). Endosperm sugar accumulation caused by mutation of PHS8/ISA1 leads to pre-harvest sprouting in rice Plant J 95, 545-556. [本文引用: 1]
FanZQ, BaLJ, ShanW, XiaoYY, LuWJ, KuangJF, ChenJY ( 2018). A banana R2R3-MYB transcription factor MaMYB3 is involved in fruit ripening through modulation of starch degradation by repressing starch degradation-rela- ted genes andMabHLH6 Plant J 96, 1191-1205. [本文引用: 1]
XuR, YuHY, WangJM, DuanPG, ZhangBL, LiJ, LiY, XuJS, LyuJ, LiN, ChaiTY, LiYH ( 2018 f). A mitogen-activated protein kinase phosphatase influences grain size and weight in rice Plant J 95, 937-946. [本文引用: 1]
XuZC, XinTY, BartelsD, LiY, GuW, YaoH, LiuS, YuHY, PuXD, ZhouJG, XuJ, XiCC, LeiHT, SongJY, ChenSL ( 2018 g). Genome analysis of the ancient tracheophyte Selaginella tamariscina reveals evolutionary features relevant to the acquisition of desiccation tolerance Mol Plant 11, 983-994. [本文引用: 1]
FengF, QiWW, LvYD, YanSM, XuLM, YangAY, YuanY, ChenYH, ZhaoH, SongRT ( 2018). OPAQUE11 is a central hub of the regulatory network for maize endosperm development and nutrient metabolism Plant Cell 30, 375-396. [本文引用: 1]
GongR, CaoHS, ZhangJN, XieK, WangDW, YuSB ( 2018). Divergent functions of the GAGA-binding transcription factor family in rice Plant J 94, 32-47. [本文引用: 1]
GuoT, ChenK, DongNQ, ShiCL, YeWW, GaoJP, ShanJX, LinHX ( 2018 c). GRAIN SIZE AND NUMBER1 negatively regulates the OsMKKK10-OsMKK4-OsMPK6 cascade to coordinate the trade-off between grain number per panicle and grain size in rice Plant Cell 30, 871-888. [本文引用: 1]
GuoZ, YangW, ChangY, MaX, TuH, XiongF, JiangN, FengH, HuangC, YangP, ZhaoH, ChenG, LiuH, LuoL, HuH, LiuQ, XiongL ( 2018 d). Genome-wide association studies of image traits reveal genetic architecture of drought resistance in rice Mol Plant 11, 789-805.
YanJ, YaoR, ChenL, LiS, GuM, NanF, XieD ( 2018 b). Dynamic perception of jasmonates by the F-box protein COI1 Mol Plant 11, 1237-1247. [本文引用: 1]
YanMH, FritschPW, MooreMJ, FengT, MengAP, YangJ, DengT, ZhaoCX, YaoXH, SunH, WangHC ( 2018 c). Plastid phylogenomics resolves infrafamilial relationships of the Styracaceae and sheds light on the backbone relationships of the Ericales Mol Phylogenet Evol 121, 198-211. [本文引用: 1]
YanT, LiL, XieL, ChenM, ShenQ, PanQ, FuX, ShiP, TangY, HuangH, HuangY, HuangY, TangK ( 2018 d). A novel HD-ZIP IV/MIXTA complex promotes glandular trichome initiation and cuticle development in Artemisia annua New Phytol 218, 567-578. [本文引用: 1]
YangB, SongZ, LiC, JiangJ, ZhouY, WangR, WangQ, NiC, LiangQ, ChenH, FanLM ( 2018 a). RSM1, an Arabidopsis MYB protein, interacts with HY5/HYH to modulate seed germination and seedling development in response to abscisic acid and salinity PLoS Genet 14, e1007839. [本文引用: 1]
YangC, MaY, HeY, TianZ, LiJ ( 2018 b). OsOFP19 modulates plant architecture by integrating the cell division pattern and brassinosteroid signaling Plant J 93, 489-501. [本文引用: 1]
YangCW, XieFM, JiangYP, LiZ, HuangX, LiL ( 2018 c). Phytochrome A negatively regulates the shade avoidance response by increasing Auxin/Indole acidic acid protein stability Dev Cell 44, 29-41. [本文引用: 1]
HanJJ, SongZT, SunJL, YangZT, XianMJ, WangS, SunL, LiuJX ( 2018). Chromatin remodeling factor CHR18 interacts with replication protein RPA1A to regulate the DNA replication stress response in Arabidopsis New Phytol 220, 476-487. [本文引用: 1]
HeL, WuWW, ZintaG, YangL, WangD, LiuRY, ZhangHM, ZhengZM, HuangH, ZhangQZ, ZhuJK ( 2018 b). A naturally occurring epiallele associates with leaf senescence and local climate adaptation in Arabidopsis accessions Nat Commun 9, 460. [本文引用: 1]
HeP, XiaoG, LiuH, ZhangL, ZhaoL, TangM, HuangS, AnY, YuJ ( 2018 c). Two pivotal RNA editing sites in the mitochondrial atp1mRNA are required for ATP synthase to produce sufficient ATP for cotton fiber cell elongation New Phytol 218, 167-182. [本文引用: 1]
HeY, ZhuM, WangL, WuJ, WangQ, WangR, ZhaoY ( 2018 d). Programmed self-elimination of the CRISPR/ Cas9 construct greatly accelerates the isolation of edited and transgene-free rice plants Mol Plant 11, 1210-1213. [本文引用: 1]
HengY, WuC, LongY, LuoS, MaJ, ChenJ, LiuJ, ZhangH, RenY, WangM, TanJ, ZhuS, WangJ, LeiC, ZhangX, GuoX, WangH, ChengZ, WanJ ( 2018). OsALMT7 maintains panicle size and grain yield in rice by mediating malate transport Plant Cell 30, 889-906. [本文引用: 1]
HongY, ZhangY, SinumpornS, YuN, ZhanX, ShenX, ChenD, YuP, WuW, LiuQ, CaoZ, ZhaoC, ChengS, CaoL ( 2018). Premature leaf senescence 3, encoding a methyltransferase, is required for melatonin biosynthesis in rice Plant J 95, 877-891. [本文引用: 1]
HuS, HuX, HuJ, ShangL, DongG, ZengD, GuoL, QianQ ( 2018 a). Xiaowei, a new rice germplasm for large-scale indoor research Mol Plant 11, 1418-1420. [本文引用: 1]
HuY, ZhouL, HuangM, HeX, YangY, LiuX, LiY, HouX ( 2018 b). Gibberellins play an essential role in late embryogenesis of Arabidopsis Nat Plants 4, 289-298. [本文引用: 1]
HuZ, LuSJ, WangMJ, HeH, SunL, WangH, LiuXH, JiangL, SunJL, XinX, KongW, ChuC, XueHW, YangJ, LuoX, LiuJX ( 2018 c). A novel QTL qTGW3 encodes the GSK3/SHAGGY-like kinase OsGSK5/OsSK41 that interacts with OsARF4 to negatively regulate grain size and weight in rice Mol Plant 11, 736-749. [本文引用: 1]
HuZR, WangR, ZhengM, LiuXB, MengF, WuHL, YaoYY, XinMM, PengHR, NiZF, SunQX ( 2018 d). TaWRKY51 promotes lateral root formation through negative regulation of ethylene biosynthesis in wheat ( Triticum aestivum L.) Plant J 96, 372-388. [本文引用: 1]
HuaK, TaoX, YuanF, WangD, ZhuJK ( 2018). Precise A .T to G .C base editing in the rice genome Mol Plant 11, 627-630. [本文引用: 1]
HuaiJ, ZhangX, LiJ, MaT, ZhaP, JingY, LinR ( 2018). SEUSS and PIF4 coordinately regulate light and temperature signaling pathways to control plant growth Mol Plant 11, 928-942. [本文引用: 1]
HuangJ, ChenL, LuXY, PengQ, ZhangY, YangJ, ZhangBY, YangB, WaletichJR, YinWX, ZhengXB, WangYC, DongSM ( 2019). Natural allelic variations provide insights into host adaptation of Phytophthora avirulence effector PsAvr3c New Phytol 221, 1010-1022. [本文引用: 1]
HuangW, ZhuY, WuW, LiX, ZhangD, YinP, HuangJ ( 2018 b). The pentatricopeptide repeat protein SOT5/ EMB2279 is required for plastid rpl2 and trnK intron splicing Plant Physiol 177, 684-697. [本文引用: 1]
HuangX, HouL, MengJ, YouH, LiZ, GongZ, YangS, ShiY ( 2018 c). The antagonistic action of abscisic acid and cytokinin signaling mediates drought stress response in Arabidopsis Mol Plant 11, 970-982. [本文引用: 1]
HuangY, ZhaoS, FuY, SunH, MaX, TanL, LiuF, SunX, SunH, GuP, XieD, SunC, ZhuZ ( 2018 d). Variation in the regulatory region of FZP causes increases in secondary inflorescence branching and grain yield in rice domestication Plant J 96, 716-733. [本文引用: 1]
JiaDJ, ShenF, WangY, WuT, XuXF, ZhangXZ, HanZH ( 2018 a). Apple fruit acidity is genetically diversified by natural variations in three hierarchical epistatic genes: MdSAUR37, MdPP2CH and MdALMTII Plant J 95, 427-443. [本文引用: 1]
JiaPF, XueY, LiHJ, YangWC ( 2018 b). Golgi-localized LOT regulates trans-golgi network biogenesis and pollen tube growth Proc Natl Acad Sci USA 115, 12307-12312. [本文引用: 1]
JiangP, WangS, JiangH, ChengB, WuK, DingY ( 2018 a). The COMPASS-like complex promotes flowering and panicle branching in rice Plant Physiol 176, 2761-2771. [本文引用: 1]
JiangP, WangS, ZhengH, LiH, ZhangF, SuY, XuZ, LinH, QianQ, DingY ( 2018 b). SIP1 participates in regulation of flowering time in rice by recruiting OsTrx1 to Ehd1 New Phytol 219, 422-435. [本文引用: 1]
JiangSJ, LiuYJ, LuoJJ, QinMS, JohnsonNC, OpikM, VasarM, ChaiYX, ZhouXL, MaoL, DuGZ, AnLZ, FengHY ( 2018 c). Dynamics of arbuscular mycorrhizal fungal community structure and functioning along a nitrogen enrichment gradient in an alpine meadow ecosystem New Phytol 220, 1222-1235. [本文引用: 1]
JiangT, ZhangJ, RongLW, FengYJ, WangQ, SongQL, ZhangLX, OuyangM ( 2018 d). ECD1 functions as an RNA-editing transfactor of rps14-149 in plastids and is required for early chloroplast development in seedlings J Exp Bot 69, 3037-3051. [本文引用: 1]
JiangYN, XieQJ, WangWX, YangJ, ZhangXW, YuN, ZhouY, WangET ( 2018 e). Medicago AP2-domain transcription factor WRI5a is a master regulator of lipid biosynthesis and transfer during mycorrhizal symbiosis Mol Plant 11, 1344-1359. [本文引用: 1]
KouS, ChenL, TuW, ScossaF, WangY, LiuJ, FernieAR, SongB, XieC ( 2018). The arginine decarboxylase gene ADC1, associated to the putrescine pathway, plays an important role in potato cold-acclimated freezing tolerance as revealed by transcriptome and metabolome analyses Plant J 96, 1283-1298. [本文引用: 1]
LanYX, LiuXN, FuY, HuangSJ ( 2018). Arabidopsis class I formins control membrane-originated actin polymerization at pollen tube tips PLoS Genet 14, e1007789. [本文引用: 1]
LiC, YueY, ChenH, QiW, SongR ( 2018 a). The ZmbZIP22 transcription factor regulates 27-kD gamma-zein gene transcription during maize endosperm development Plant Cell 30, 2402-2424. [本文引用: 1]
LiH, LuoN, WangW, LiuZ, ChenJ, ZhaoL, TanL, WangC, QinY, LiC, XuT, YangZ ( 2018 b). The REN4 rheostat dynamically coordinates the apical and lateral domains of Arabidopsis pollen tubes Nat Commun 9, 2573. [本文引用: 1]
LiH, WangH, JingM, ZhuJ, GuoB, WangY, LinY, ChenH, KongL, MaZ, WangY, YeW, DongS, TylerB, WangY ( 2018 c). A phytophthora effector recruits a host cytoplasmic transacetylase into nuclear speckles to enhance plant susceptibility eLife 7, e40039. [本文引用: 1]
LiJC, LiangWJ, LiY, QianWQ ( 2018 e). APURINIC/ APYRIMIDINIC ENDONUCLEASE2 and ZINC FINGER DNA 3'-PHOSPHOESTERASE play overlapping roles in the maintenance of epigenome and genome stability Plant Cell 30, 1954-1970. [本文引用: 1]
YangJ, FuM, JiC, HuangY, WuY ( 2018 e). Maize Oxalyl-CoA Decarboxylase1 degrades oxalate and affects the seed metabolome and nutritional quality Plant Cell 30, 2447-2462. [本文引用: 1]
LiN, LiuZ, WangZ, RuL, GonzalezN, BaekelandtA, PauwelsL, GoossensA, XuR, ZhuZ, InzeD, LiY ( 2018 f). STERILE APETALA modulates the stability of a repressor protein complex to control organ size in Arabidopsis thaliana PLoS Genet 14, e1007218. [本文引用: 1]
LiS, XuH, JuZ, CaoD, ZhuH, FuD, GriersonD, QinG, LuoY, ZhuB ( 2018 h). The RIN-MC fusion of MADS-box transcription factors has transcriptional activity and modulates expression of many ripening genes Plant Physiol 176, 891-909. [本文引用: 1]
LiY, WangNN, WangY, LiuD, GaoY, LiL, LiXB ( 2018 i). The cotton XLIM protein (GhXLIM6) is required for fiber development via maintaining dynamic F-actin cytoskeleton and modulating cellulose biosynthesis Plant J 96, 1269-1282. [本文引用: 1]
LiYF, QinBX, ShenY, ZhangFF, LiuCZ, YouHL, DuGJ, TangD, ChengZK ( 2018 j). HEIP1 regulates crossover formation during meiosis in rice Proc Natl Acad Sci USA 115, 10810-10815. [本文引用: 1]
LiZ, JiangD, HeY ( 2018 k). FRIGIDA establishes a local chromosomal environment for FLOWERING LOCUS C mRNA production Nat Plants 4, 836-846. [本文引用: 1]
LiZC, FuX, WangYZ, LiuRY, HeYH ( 2018 l). Polycomb-mediated gene silencing by the BAH-EMF1 complex in plants Nat Genet 50, 1254-1261. [本文引用: 1]
LianHL, XuPB, HeSB, WuJ, PanJ, WangWX, XuF, WangS, PanJS, HuangJR, YangHQ ( 2018). Photoexcited CRYPTOCHROME 1 interacts directly with G-protein beta subunit AGB1 to regulate the DNA-binding activity of HY5 and photomorphogenesis in Arabidopsis Mol Plant 11, 1248-1263. [本文引用: 1]
LiangJ, LiuJ, BrownR, JiaM, ZhouK, PetersRJ, WangQ ( 2018 a). Direct production of dihydroxylated sesquiterpenoids by a maize terpene synthase Plant J 94, 847-856. [本文引用: 1]
LiangT, MeiSL, ShiC, YangY, PengY, MaLB, WangF, LiX, HuangX, YinYH, LiuHT ( 2018 b). UVR8 interacts with BES1 and BIM1 to regulate transcription and photomorphogenesis in Arabidopsis Dev Cell 44, 512-523. [本文引用: 1]
LiangZ, ShenLS, CuiXA, BaoSJ, GengYK, YuGL, LiangF, XieS, LuTG, GuXF, YuH ( 2018 c). DNA N-6-adenine methylation in Arabidopsis thaliana Dev Cell 45, 406-416. [本文引用: 1]
LiangZ, ZhuN, MaiKK, LiuZ, TzengD, OsteryoungKW, ZhongS, StaehelinLA, KangBH ( 2018 d). Thylakoid-bound polysomes and a dynamin-related protein, FZL, mediate critical stages of the linear chloroplast biogenesis program in greening Arabidopsis cotyledons Plant Cell 30, 1476-1495. [本文引用: 1]
LiaoY, ZhangXM, LiB, LiuTY, ChenJF, BaiZT, WangMJ, ShiJF, WallingJG, WingRA, JiangJM, ChenMS ( 2018). Comparison of Oryza sativa and Oryza brachyantha genomes reveals selection-driven gene escape from the centromeric regions Plant Cell 30, 1729-1744. [本文引用: 1]
LinF, JiangY, LiJ, YanT, FanL, LiangJ, ChenZJ, XuD, DengXW ( 2018 a). B-BOX DOMAIN PROTEIN28 negatively regulates photomorphogenesis by repressing the activity of transcription factor HY5 and undergoes COP1- mediated degradation Plant Cell 30, 2006-2019. [本文引用: 1]
LinXW, GuDD, ZhaoHT, PengY, ZhangGF, YuanTT, LiMG, WangZJ, WangXT, CuiSJ ( 2018 b). LFR is functionally associated with AS2 to mediate leaf development in Arabidopsis Plant J 95, 598-612. [本文引用: 1]
LiuC, ZhengS, GuiJ, FuC, YuH, SongD, ShenJ, QinP, LiuX, HanB, YangY, LiL ( 2018 b). Shortened basal internodes encodes a gibberellin 2-oxidase and contributes to lodging resistance in rice Mol Plant 11, 288-299. [本文引用: 1]
LiuHY, MiZR, LinL, WangYH, ZhangZH, ZhangFW, WangH, LiuLL, ZhuB, CaoGM, ZhaoXQ, SandersNJ, ClassenAT, ReichPB, HeJS ( 2018 c). Shifting plant species composition in response to climate change stabilizes grassland primary production Proc Natl Acad Sci USA 115, 4051-4056. [本文引用: 1]
LiuJX, WuXB, YaoXF, YuR, LarkinPJ, LiuCM ( 2018 d). Mutations in the DNA demethylase OsROS1 result in a thickened aleurone and improved nutritional value in rice grains Proc Natl Acad Sci USA 115, 11327-11332. [本文引用: 1]
LiuW, JiangB, MaL, ZhangS, ZhaiH, XuX, HouW, XiaZ, WuC, SunS, WuT, ChenL, HanT ( 2018 f). Functional diversification of Flowering Locus T homologs in soybean: GmFT1a and GmFT2a/5a have opposite roles in controlling flowering and maturation New Phytol 217, 1335-1345. [本文引用: 1]
LiuYW, LiX, MaDB, ChenZR, WangJW, LiuHT ( 2018 g). CIB1 and CO interact to mediate CRY2-dependent regulation of flowering EMBO Rep 19, e45762. [本文引用: 1]
LiuYY, CaoYL, ZhangQL, LiXH, WangSP ( 2018 h). A cytosolic triosephosphate isomerase is a key component in XA3/XA26-mediated resistance Plant Physiol 178, 923-935. [本文引用: 1]
YangM, ZhangY, XieX, YueN, LiJ, WangXB, HanC, YuJ, LiuY, LiD ( 2018 f). Barley stripe mosaic virus gammab protein subverts autophagy to promote viral infection by disrupting the ATG7-ATG8 interaction Plant Cell 30, 1582-1595. [本文引用: 1]
YangY, LiangT, ZhangLB, ShaoK, GuXX, ShangRX, ShiN, LiX, ZhangP, LiuHT ( 2018 g). UVR8 interacts with WRKY36 to regulate HY5 transcription and hypocotyl elongation in Arabidopsis Nat Plants 4, 98-107. [本文引用: 1]
LuoJS, HuangJ, ZengDL, PengJS, ZhangGB, MaHL, GuanY, YiHY, FuYL, HanB, LinHX, QianQ, GongJM ( 2018 a). A defensin-like protein drives cadmium efflux and allocation in rice Nat Commun 9, 645. [本文引用: 1]
LuoLJ, ZengJ, WuHJ, TianZX, ZhaoZ ( 2018 b). A molecular framework for auxin-controlled homeostasis of shoot stem cells in Arabidopsis. Mol Plant 11, 899-913. [本文引用: 1]
LuoWT, ZuoXA, MaW, XuC, LiA, YuQ, KnappAK, TognettiR, DijkstraFA, LiMH, HanGD, WangZW, HanXG ( 2018 c). Differential responses of canopy nutrients to experimental drought along a natural aridity gradient Ecology 99, 2230-2239. [本文引用: 1]
LvBS, TianHY, ZhangF, LiuJJ, LuSC, BaiMY, LiCY, DingZJ ( 2018). Brassinosteroids regulate root growth by controlling reactive oxygen species homeostasis and dual effect on ethylene synthesis in Arabidopsis PLoS Genet 14, e1007144. [本文引用: 1]
MaB, ZhouY, ChenH, HeSJ, HuangYH, ZhaoH, LuX, ZhangWK, PangJH, ChenSY, ZhangJS ( 2018 a). Membrane protein MHZ3 stabilizes OsEIN2 in rice by interacting with its Nramp-dlike domain Proc Natl Acad Sci USA 115, 2520-2525. [本文引用: 1]
MaH, LiuC, LiZ, RanQ, XieG, WangB, FangS, ChuJ, ZhangJ ( 2018 b). ZmbZIP4 contributes to stress resistance in maize by regulating ABA synthesis and root development Plant Physiol 178, 753-770. [本文引用: 1]
MaQQ, WangXH, SunJB, MaoTL ( 2018 c). Coordinated regulation of hypocotyl cell elongation by light and ethylene through a microtubule destabilizing protein Plant Physiol 176, 678-690. [本文引用: 1]
MaYZ, MinL, WangMJ, WangCZ, ZhaoYL, LiYY, FangQD, WuYL, XieS, DingYH, SuXJ, HuQ, ZhangQH, LiXY, ZhangXL ( 2018 d). Disrupted genome methylation in response to high temperature has distinct affects on microspore abortion and anther indehiscence Plant Cell 30, 1387-1403. [本文引用: 1]
YangZL, QianSM, ScheidRN, LuL, ChenXS, LiuR, DuXA, LvXC, BoersmaMD, ScalfM, SmithLM, DenuJM, DuJM, ZhongXH ( 2018 i). EBS is a bivalent histone reader that regulates floral phase transition in Arabidopsis Nat Genet 50, 1247-1253. [本文引用: 1]
MiaoR, WangM, YuanW, RenY, LiY, ZhangN, ZhangJH, KronzuckerHJ, XuWF ( 2018 b). Comparative analysis of Arabidopsis ecotypes reveals a role for brassinosteroids in root hydrotropism Plant Physiol 176, 2720-2736. [本文引用: 1]
MiaoZQ, ZhaoPX, MaoJL, YuLH, YuanY, TangH, LiuZB, XiangCB ( 2018 c). HOMEOBOX PROTEIN52 mediates the crosstalk between ethylene and auxin signaling during primary root elongation by modulating auxin transport-related gene expression Plant Cell 30, 2761-2778. [本文引用: 1]
YangZL, QiuQ, ChenW, JiaB, ChenXM, HuHM, HeKX, DengX, LiSS, TaoWA, CaoXF, DuJM ( 2018 j). Structure of the Arabidopsis JMJ14-H3K4me3 complex provides insight into the substrate specificity of KDM5 subfamily histone demethylases Plant Cell 30, 167-177. [本文引用: 1]
YaoXZ, TianL, YangJ, ZhaoYN, ZhuYX, DaiXH, ZhaoYD, YangZN ( 2018). Auxin production in diploid microsporocytes is necessary and sufficient for early stages of pollen development PLoS Genet 14, e1007397. [本文引用: 1]
NiuSL, ClassenAT, LuoYQ ( 2018). Functional traits along a transect Funct Ecol 32, 4-9. [本文引用: 1]
PanX, MaJ, SuX, CaoP, ChangW, LiuZ, ZhangX, LiM ( 2018). Structure of the maize photosystem I supercomplex with light-harvesting complexes I and II Science 360, 1109-1113. [本文引用: 1]
YeCL, ChenDM, HallSJ, PanS, YanXB, BaiTS, GuoH, ZhangY, BaiYF, HuSJ ( 2018 a). Reconciling multiple impacts of nitrogen enrichment on soil carbon: plant, microbial and geochemical controls Ecol Lett 21, 1162-1173. [本文引用: 1]
YeQ, WangH, SuT, WuWH, ChenYF ( 2018 b). The ubiquitin E3 Ligase PRU1 regulates WRKY6 degradation to modulate phosphate homeostasis in response to low-Pi stress in Arabidopsis Plant Cell 30, 1062-1076. [本文引用: 1]
YinLM, DijkstraFA, WangP, ZhuB, ChengWX ( 2018 a). Rhizosphere priming effects on soil carbon and nitrogen dynamics among tree species with and without intraspecific competition New Phytol 218, 1036-1048. [本文引用: 1]
YinXH, SterckF, HaoGY ( 2018 b). Divergent hydraulic strategies to cope with freezing in co-occurring temperate tree species with special reference to root and stem pressure generation New Phytol 219, 530-541. [本文引用: 1]
YingJZ, MaM, BaiC, HuangXH, LiuJL, FanYY, SongXJ ( 2018). TGW3, a major QTL that negatively modulates grain length and weight in rice Mol Plant 11, 750-753. [本文引用: 1]
ZengYF, ZhangJG, AbuduhamitiB, WangWT, JiaZQ ( 2018). Phylogeographic patterns of the desert poplar in Northwest China shaped by both geology and climatic oscillations BMC Evol Biol 18, 75. [本文引用: 1]
ZhangC, ShenY, TangD, ShiWQ, ZhangDM, DuGJ, ZhouYH, LiangGH, LiYF, ChengZK ( 2018 a). The zinc finger protein DCM1 is required for male meiotic cytokinesis by preserving callose in rice PLoS Genet 14, e1007769. [本文引用: 1]
PengML, LiZP, ZhouN, MaMM, JiangYP, DongAW, ShenWH, LiL ( 2018 a). Linking PHYTOCHROME-INTE-RACTING FACTOR to histone modification in plant shade avoidance Plant Physiol 176, 1341-1351. [本文引用: 1]
PengY, ChenL, LiS, ZhangY, XuR, LiuZ, LiuW, KongJ, HuangX, WangY, ChengB, ZhengL, LiY ( 2018 b). BRI1 and BAK1 interact with G proteins and regulate sugar-responsive growth and development in Arabidopsis Nat Commun 9, 1522. [本文引用: 1]
ZhangD, XuZ, CaoS, ChenK, LiS, LiuX, GaoC, ZhangB, ZhouY ( 2018 b). An uncanonical CCCH-tandem zinc- finger protein represses secondary wall synthesis and controls mechanical strength in rice Mol Plant 11, 163-174. [本文引用: 1]
ZhangH, YuP, ZhaoJ, JiangH, WangH, ZhuY, BotellaMA, SamajJ, LiC, LinJ ( 2018 c). Expression of tomato prosystemin gene in Arabidopsis reveals systemic translocation of its mRNA and confers necrotrophic fungal resistance New Phytol 217, 799-812. [本文引用: 1]
ZhangJS, ZhangH, SrivastavaAK, PanYJ, BaiJJ, FangJJ, ShiHZ, ZhuJK ( 2018 d). Knockdown of rice microRNA166 confers drought resistance by causing leaf rolling and altering stem xylem development Plant Physiol 176, 2082-2094. [本文引用: 1]
ZhangM, CaoY, WangZ, WangZQ, ShiJ, LiangX, SongW, ChenQ, LaiJ, JiangC ( 2018 g). A retrotransposon in an HKT1 family sodium transporter causes variation of leaf Na( +) exclusion and salt tolerance in maize New Phytol 217, 1161-1176. [本文引用: 1]
ZhangN, YuH, YuH, CaiY, HuangL, XuC, XiongG, MengX, WangJ, ChenH, LiuG, JingY, YuanY, LiangY, LiS, SmithSM, LiJ, WangY ( 2018 h). A core regulatory pathway controlling rice tiller angle mediated by the LAZY1-dependent asymmetric distribution of auxin Plant Cell 30, 1461-1475.
ZhangQ, MaC, ZhangY, GuZ, LiW, DuanX, WangS, HaoL, WangY, WangS, LiT ( 2018 i). A single-nucleotide polymorphism in the promoter of a hairpin RNA contributes to Alternaria alternata leaf spot resistance in apple( Malus x domestica) Plant Cell 30, 1924-1942. [本文引用: 2]
ZhaoTT, RenLJ, ChenXJ, YuHX, LiuCJ, ShenY, ShiWQ, TangD, DuGJ, LiYF, MaBJ, ChengZK ( 2018 f). The OsRR24/LEPTO1 type-B response regulator is essential for the organization of leptotene chromosomes in rice meiosis Plant Cell 30, 3024-3037. [本文引用: 1]
ZhaoXH, JiangY, LiJ, HuqE, ChenZJ, XuDQ, DengXW ( 2018 g). COP1 SUPPRESSOR 4 promotes seedling photomorphogenesis by repressing CCA1 and PIF4 expression in Arabidopsis Proc Natl Acad Sci USA 115, 11631-11636. [本文引用: 1]
ZhaoXY, LiJR, LianB, GuHQ, LiY, QiYJ ( 2018 h). Global identification of Arabidopsis lncRNAs reveals the regulation of MAF4 by a natural antisense RNA Nat Commun 9, 5056. [本文引用: 1]
ZhaoY, ZhangHL, XuJL, JiangCH, YinZG, XiongHY, XieJY, WangXQ, ZhuXY, LiY, ZhaoWP, RashidMAR, LiJJ, WangWS, FuBY, YeGY, GuoY, HuZQ, LiZK, LiZC ( 2018 i). Loci and natural alleles underlying robust roots and adaptive domestication of upland ecotype rice in aerobic conditions PLoS Genet 14, e1007521. [本文引用: 1]
ZhaoYS, WangSY, WuWY, LiL, JiangT, ZhengBL ( 2018 j). Clearance of maternal barriers by paternal miR159 to initiate endosperm nuclear division in Arabidopsis Nat Commun 9, 5011. [本文引用: 1]
ZhengH, ZhangF, WangS, SuY, JiX, JiangP, ChenR, HouS, DingY ( 2018 a). MLK1 and MLK2 coordinate RGA and CCA1 activity to regulate hypocotyl elongation in Arabidopsis thaliana Plant Cell 30, 67-82. [本文引用: 1]
ZhengX, KangS, JingY, RenZ, LiL, ZhouJM, BerkowitzG, ShiJ, FuA, LanW, ZhaoF, LuanS ( 2018 b). Danger-associated peptides close stomata by OST1-indepen-dent activation of anion channels in guard cells Plant Cell 30, 1132-1146. [本文引用: 1]
ZhengX, ZhaoY, ShanD, ShiK, WangL, LiQ, WangN, ZhouJ, YaoJ, XueY, FangS, ChuJ, GuoY, KongJ ( 2018 c). MdWRKY9 overexpression confers intensive dwarfing in the M26 rootstock of apple by directly inhibiting brassinosteroid synthetase MdDWF4 expression New Phytol 217, 1086-1098. [本文引用: 1]
ZhengYL, BurnsJH, LiaoZY, LiYP, YangJ, ChenYJ, ZhangJL, ZhengYG ( 2018 d). Species composition, functional and phylogenetic distances correlate with success of invasive Chromolaena odorata in an experimental test Ecol Lett 21, 1211-1220. [本文引用: 1]
ZhouC, WangCS, LiuHB, ZhouQW, LiuQ, GuoY, PengT, SongJM, ZhangJW, ChenLL, ZhaoY, ZengZX, ZhouDX ( 2018 a). Identification and analysis of adenine N-6-methylation sites in the rice genome Nat Plants 4, 554-563. [本文引用: 1]
ZhouX, LiaoH, ChernM, YinJ, ChenY, WangJ, ZhuX, ChenZ, YuanC, ZhaoW, WangJ, LiW, HeM, MaB, WangJ, QinP, ChenW, WangY, LiuJ, QianY, WangW, WuX, LiP, ZhuL, LiS, RonaldPC, ChenX ( 2018 b). Loss of function of a rice TPR-domain RNA-binding protein confers broad-spectrum disease resistance Proc Natl Acad Sci USA 115, 3174-3179.
ZhouY, YangL, DuanJ, ChengJ, ShenY, WangX, HanR, LiH, LiZ, WangL, TerzaghiW, ZhuD, ChenH, DengXW, LiJ ( 2018 c). Hinge region of Arabidopsis phyA plays an important role in regulating phyA function Proc Natl Acad Sci USA 115, E11864-E11873. [本文引用: 2]
ZhouY, ZhangD, AnJ, YinH, FangS, ChuJ, ZhaoY, LiJ ( 2018 d). TCP transcription factors regulate shade avoidance via directly mediating the expression of both PHYTOCHROME INTERACTING FACTORs and auxin biosynthetic genes Plant Physiol 176, 1850-1861. [本文引用: 1]
ZhouYB, LiuC, TangDY, YanL, WangD, YangYZ, GuiJS, ZhaoXY, LiLG, TangXD, YuF, LiJL, LiuLL, ZhuYH, LinJZ, LiuXM ( 2018 e). The receptor-like cytoplasmic kinase STRK1 phosphorylates and activates CatC, thereby regulating H2O2 homeostasis and improving salt tolerance in rice Plant Cell 30, 1100-1118.
ZhuC, PengQ, FuD, ZhuangD, YuY, DuanM, XieW, CaiY, OuyangY, LianX, WuC ( 2018 a). The E3 ubiquitin ligase HAF1 modulates circadian accumulation of EARLY FLOWERING3 to control heading date in rice under long- day conditions Plant Cell 30, 2352-2367. [本文引用: 1]
ZhuG, WangS, HuangZ, ZhangS, LiaoQ, ZhangC, LinT, QinM, PengM, YangC, CaoX, HanX, WangX, van der KnaapE, ZhangZ, CuiX, KleeH, FernieAR, LuoJ, HuangS ( 2018 b). Rewiring of the fruit metabolome in tomato breeding Cell 172, 249-261. [本文引用: 1]
ZhuQ, ZengD, YuS, CuiC, LiJ, LiH, ChenJ, ZhangR, ZhaoX, ChenL, LiuYG ( 2018 c). From golden rice to aSTARice: bioengineering astaxanthin biosynthesis in rice endosperm Mol Plant 11, 1440-1448. [本文引用: 2]
ZouS, WangH, LiY, KongZ, TangD ( 2018 a). The NB-LRR gene Pm60 confers powdery mildew resistance in wheat New Phytol 218, 298-309.
ZouYM, WangSF, ZhouYY, BaiJJ, HuangGZ, LiuXT, ZhangYY, TangDZ, LuDP ( 2018 b). Transcriptional regulation of the immune receptor FLS2 controls the ontogeny of plant innate immunity Plant Cell 30, 2779-2794. [本文引用: 1]
A well-resolved fern nuclear phylogeny reveals the evolution history of numerous transcription factor families 1 2018
Improved base editor for efficiently inducing genetic variations in rice with CRISPR/Cas9-guided hyperactive hAID mutant 1 2018
... 随着科学技术的发展, 基因组编辑技术开始应用于作物精准育种.然而由于缺乏高效去除含有Cas9表达盒及扩展性强、编辑效率高的单碱基编辑技术, 限制了其在分子育种中的应用.谢卡斌研究组利用生物体内的内含子剪切系统和tRNA加工系统开发出水稻中高效、多位点编辑且适用于CRISPR/Cpf1系统的新方法, 有望应用于动植物中(Ding et al., 2018a).林宏辉和周焕斌研究组及朱健康研究组分别基于APOBEC1 酶的碱基编辑器及腺嘌呤的碱基编辑器(ABE7-10), 实现了水稻基因组4种不同碱基(A-G、T-C、C-T和G-A)的高效替换与水稻基因组特定位点的A·T碱基对高效转化为G·C碱基对, 不仅扩展了单碱基编辑技术在植物中的应用而且丰富了可用的单碱基编辑工具(Ren et al., 2018a; Hua et al., 2018).赵云德研究组则利用自杀基因与CRISPR载体融合, 开发出高效的转基因自清除基因编辑系统(He et al., 2018d).这些成果对深入研究水稻功能基因组具有重要推动作用. ...
GhWIP2, a WIP zinc finger protein, suppresses cell expansion in Gerbera hybrida by mediating crosstalk between gibberellin, abscisic acid, and auxin 1 2018
... 植物激素之间存在复杂的调控网络.杨淑华研究组发现ABA信号关键蛋白激酶SnRK2s通过磷酸化细胞分裂素负调节子A型ARR5正调控其蛋白稳定, 揭示了植物激素信号调控网络在植物逆境响应胁迫中的精密调控机制(Huang et al., 2018c).刘西岗研究组发现细胞分裂素的内稳态对于花分生组织(floral meristem, FM)的维持及分化非常重要.生长素可以通过促进ARF3 (Auxin Response Factor 3)的表达抑制细胞分裂素的活性进而促进FM的分化(Zhang et al., 2018e).该研究揭示了转录因子AG通过植物激素信号途径协同调控花分生组织干细胞稳态的机理.王小菁研究组发现非洲菊锌指蛋白GhWIP2通过抑制GA和IAA信号及激活ABA信号, 进而抑制细胞生长并最终抑制花瓣伸展(Ren et al., 2018b).该研究揭示了GA、ABA和生长素3种激素交叉互作网络共同调控非洲菊细胞伸展的作用机理.向成斌研究组发现乙烯响应基因HB52是乙烯和IAA信号互作的重要节点, 通过上调PIN2、WAG1和WAG2的表达抑制主根伸长(Miao et al., 2018c).该研究揭示了乙烯与IAA交互作用的新机制.丁兆军研究组研究表明, BL合成途径突变体det2-9中乙烯合成大幅增加并积累了大量的超氧阴离子.该研究揭示了BL、乙烯与活性氧协同调控根生长的新机制(Lv et al., 2018).ABA和BL信号通常相互拮抗.王学路研究组发现ABI1和ABI2通过与BIN2互作使其去磷酸化, 进而调控其对BES1的磷酸化(Wang et al., 2018c).同时, 该研究组还发现BL信号途径中的GSK3为控制水稻中胚轴伸长的一个关键因子, 其通过磷酸化调控一类细胞周期蛋白CYC U2的稳定性促进细胞分裂.BL以及独脚金内酯(strigolactone)信号通路协同调控GSK3介导的细胞周期进程, 进而促进中胚轴伸长(Sun et al., 2018c). ...
Widespread whole genome duplications contribute to genome complexity and species diversity in angiosperms 1 2018
... 全基因组复制(WGD)长期以来被认为是物种多样性的主要推动力.戚继和马红研究组利用转录组数据对被子植物代表性植物进行了比较基因组学和系统基因组学分析, 验证了已报道的大部分WGD, 并检测到一些在被子植物演化过程中广泛传播的新WGD, 为证明WGD对被子植物辐射演化及增强适应环境变化能力具重要作用提供了充分证据(Ren et al., 2018c). ...
An SPX-RLI1 module regulates leaf inclination in response to phosphate availability in rice 1 2018
... 水稻株型与产量密切相关.叶片作为水稻理想株型的组成要素之一, 其倾角对产量影响较大.薛红卫研究组新克隆了叶倾角调控基因LC3, 该基因编码1个含有SPOC结构域的转录抑制子, 其可与HIT锌指结构域蛋白LIP1互作, 通过调控生长素平衡和信号转导来调控植株叶倾角(Chen et al., 2018h).李建雄研究组与田志宏研究组合作揭示了OsOFP19、OSH1与DLT蛋白可形成复合体, 调控BL信号转导及影响叶倾角发育(Yang et al., 2018b).易可可研究组则解析了磷素有效性调控水稻叶倾角的分子机制.他们发现磷饥饿诱导的SPX1 (Syg1/Pho81/XPR1)与SPX2蛋白负调控叶倾角.SPX1的互作蛋白RLI1 (Regulator of leaf inclination 1)正调控叶倾角.RLI1可直接激活下游BU1 (Brassinosteroid upregulated 1)和BC1 (BU1-like1 complex 1)基因的表达以控制叶枕细胞的伸长, 从而使叶倾角增大(Ruan et al., 2018).种康和徐云远研究组证实miR396d通过分别抑制其靶基因OsGRF4和OsGRF6的转录, 促进BL及抑制GA信号途径, 进而调控水稻叶夹角和株高(Tang et al., 2018).邢永忠研究组则阐释了来自世界各地的水稻核心种质剑叶夹角的遗传基础, 发现bHLH转录因子家族中OsbHLH153和OsbHLH17能快速响应BL和IAA信号转导, 从而影响植株形态(Dong et al., 2018c).王永红研究组揭示了水稻分蘖角度调控的分子机制.他们通过研究水稻茎重力反应过程中的动态转录组变化, 探索了一条水稻分蘖角度调控的核心途径(Zhang et al., 2018h).该研究为解析水稻分蘖角度的调控网络进而挖掘有利用价值的基因提供了重要信息.谭禄宾研究组发现亚洲栽培稻PROG1基因相邻位置约有110 kb的染色体缺失.该缺失片段包含7个与PROG1基因相似的锌指基因, 其缺失推动了水稻由匍匐向直立转变, 最终实现产量大幅提高(Wu et al., 2018c). ...
A plant Bro1 domain protein BRAF regulates multivesicular body biogenesis and membrane protein homeostasis 1 2018
... 细胞内的囊泡运输是生命活动中极其复杂的动态生物学过程, 参与多种植物的生长发育及对环境的响应.ESCRT为内体蛋白分选转运复合体, 关于调节ESCRT和控制多囊泡体/液泡前体细胞器形成及成熟并维持膜蛋白稳态的因子一直未知.姜里文和沈锦波研究组发现了1个含有Bro1结构域的新植物特异负调控因子BRAF (BRo1-domain protein As FREE1 suppressor)蛋白, 并证明其在多囊泡体/液泡前体的腔内小泡形成和膜蛋白降解分选过程中起负调控作用(Shen et al., 2018).此外, 姜里文研究组还发现拟南芥可溶性NSF附着蛋白受体Qc-SNARE蛋白BET12在病程相关蛋白1 (Pathogenesis-Related 1, PR1)运输中具重要调节作用, 为植物抗病研究奠定了理论基础(Chung et al., 2018). ...
Feedback from lateral organs controls shoot apical meristem growth by modulating auxin transport 1 2018
... 生长素在植物生长发育过程中起重要作用.赵忠研究组发现拟南芥茎顶端分生组织中的生长素具有促进干细胞分化的功能, 并维持干细胞的稳态, 揭示了生长素维持干细胞数目稳定的新机制(Luo et al., 2018b).焦雨铃研究组与张磊研究组合作, 通过学科交叉研究发现了植物侧生器官可反馈调控茎尖干细胞, 并表明生长素长距离运输在反馈中发挥了重要作用(Shi et al., 2018a).该研究有机整合了生物学与数学, 发挥了学科交叉研究解决动态生物学过程等前沿问题的优势.胡玉欣研究组对生长素启动愈伤组织形成的分子机制进行了探索, 揭示了控制愈伤组织形成中体细胞重编程的关键因子, 并建立了植物再生体系中生长素信号与细胞全能性获得的分子联系(Xu et al., 2018a).该研究为探索植物再生的分子调控机制和发育程序提供了新线索. ...
Genome-wide regulation of light-controlled seedling morphogenesis by three families of transcription factors 1 2018
... 邓兴旺研究组与许冬清研究组合作鉴定了光形态建成的正调节因子CSU4, 它能整合生物钟节律和光信号(Zhao et al., 2018g).他们还发现BBX21通过第2个B-box结构域与HY5启动子中的T/G-box结合, 调控HY5及其靶基因的表达以促进植物光形态建成(Xu et al., 2018b).而BBX28通过与HY5蛋白的C末端互作, 抑制HY5与下游靶基因启动子结合, 从而抑制HY5活性及其下游靶基因表达, 最终抑制植物光形态建成(Lin et al., 2018a).此外, 邓兴旺和钟上威研究组揭示了PIFs、HY5和EIN3及EIL1对出土幼苗形态建成的调控作用, 为阐明幼苗从暗形态建成转向光形态建成的分子机制提供了新证据(Shi et al., 2018b).吕应堂研究组则鉴定了光形态建成的正调控因子SRS5, 它直接结合HY5、BBX21和BBX22的启动子并激活其表达, 从而促进光形态建成(Yuan et al., 2018).刘宏涛研究组发现UV-B照射后, UVR8形成单体并在细胞核中富集, UVR8通过抑制BES1/BIM1结合DNA的能力从而抑制下游生长相关基因的表达, 并抑制细胞伸长及光形态建成(Liang et al., 2018b).同时, 发现UV-B激活的UVR8形成单体, 进入细胞核直接结合WRKY36而抑制其结合HY5启动子, 进而抑制下胚轴伸长(Yang et al., 2018g). ...
The roles of AaMIXTA1 in regulating the initiation of glandular trichomes and cuticle biosynthesis in Artemisia annua 1 2018
... 茸毛作为植物体最外层结构, 具有很好的抗虫防病毒作用.叶志彪和杨长宪研究组分离了控制番茄腺体毛形成的关键基因H (Hair), 该基因编码C2H2锌指蛋白, 可能是茄科植物多细胞腺体毛形成的关键保守基因(Chang et al., 2018).唐克轩研究组发现R2R3 MYB第9亚家族的转录因子AaMIXTA1参与调控青蒿分泌型腺毛发育, 为研究其它植物多细胞腺毛奠定了基础(Shi et al., 2018c).此外, 他们还发现HD-ZIP IV转录因子AaHD8与AaMIXTA1相互作用形成复合体, 协同调控AaHD1和多个蜡质角质合成酶基因的表达, 从而促进蜡质合成和腺毛的起始发育(Yan et al., 2018d).陈晓亚研究组与相关单位合作对可纺纤维形成的分子机制进行研究, 发现编码bHLH转录因子的棉花PRE1是可纺纤维进化的目标基因, 其正调控纤维伸长.PRE1基因启动子的变异可促进棉花可纺纤维的形成(Zhao et al., 2018a).李学宝研究组鉴定了富含半胱氨酸的GhXLIM6蛋白, 其可与GhKNL1启动子的PAL-box结合而抑制该基因表达, 促进纤维素合成, 进而调控棉纤维伸长(Li et al., 2018i).俞嘉宁研究组则揭示了棉花线粒体Ghatp1基因C1292和C1415位点发生RNA编辑可影响ATPase产生ATP, 从而促进表皮毛及纤维伸长(He et al., 2018c).根毛是一种顶端生长的植物细胞, 微丝骨架对根毛的生长发育有极其重要的调控作用.贾红磊研究组揭示了H2S通过硫巯基化修饰扰乱正常的微丝骨架聚合及生长模式, 从而影响植物根毛的生长, 为进一步探索H2S调控植物生长发育的作用机制提供了理论依据(Li et al., 2018d). ...
Large-scale gene losses underlie the genome evolution of parasitic plant Cuscuta australis 1 2018
The glucose sensor MdHXK1 phosphorylates a tonoplast Na+/H+ exchanger to improve salt tolerance 1 2018
... 土壤盐碱化极大地限制了粮食作物的稳产增收.如何将盐碱地变为高产粮田, 成为当下农业科学家研究的热点方向, 而解析植物应答盐碱胁迫的分子机制是提高植物抗盐碱的有效手段.朱健康研究组发现, 细胞壁LRX (Leucine-rich repeat extensins)蛋白家族多突变体植株表现出对盐胁迫非常敏感的表型.进一步研究发现, LRX蛋白与RALF多肽以及细胞膜受体类激酶FERONIA形成一个元件调控植物生长和耐盐性(Zhao et al., 2018b).该研究为培育抗逆高产作物指明了方向.此外, 朱健康和王镇研究组还探明了ABA对植物抗旱性的影响, 揭示了开花抑制子SVP (Short vegetative phase)通过调控ABA代谢提高拟南芥的抗旱性机制(Wang et al., 2018r).植物类受体激酶能感受外界环境胁迫信号, 并通过胞内激酶域激活下游效应子活性和调控细胞相关生理生化过程, 从而降低或消除逆境胁迫的影响.刘选明和林建中研究组破译了1个能降低土地盐碱化对水稻产量影响的类受体胞质激酶1 (salt tolerance receptor-like cytoplasmic kinase 1, STRK1), 其可通过磷酸化和激活CatC调节H2O2体内平衡来改善盐以及氧化胁迫的耐受性(Zhou et al., 2018e).该研究为进一步解析植物耐盐的分子机制奠定了基础.范六民研究组报道了MYB亚家族转录因子RSM1 (Radialis-like sant/myb 1)蛋白与HY5/HYH蛋白互作, 并作用于ABI5启动子区, 调控ABA和非生物逆境(盐)胁迫相关基因的应答(Yang et al., 2018a).黄俊丽研究组则对水稻MADS-box家族转录因子OsMADS25蛋白进行了研究, 发现其可与谷胱甘肽S-转移酶基因OsGST4的启动子直接结合, 通过ABA介导的调控通路和ROS清除共同调控水稻的根系生长和耐盐性(Xu et al., 2018d).蒋才富研究组利用QTL克隆了调控叶片Na+含量的主效基因ZmNC1 (Na+ content 1).该基因编码1个Na+选择性离子转运蛋白ZmHKT1, 其可通过抑制Na+由根向地上部运输提高玉米的耐盐能力(Zhang et al., 2018g).郝玉金研究组则发现苹果葡萄糖感知因子己糖激酶MdHXK1可磷酸化Na+/H+交换蛋白MdNHX1, 以提高其蛋白稳定性, 进而增强苹果的耐盐性(Sun et al., 2018b). ...
Natural selection of a GSK3 determines rice mesocotyl domestication by coordinating strigolactone and brassinosteroid signaling 1 2018
... 植物激素之间存在复杂的调控网络.杨淑华研究组发现ABA信号关键蛋白激酶SnRK2s通过磷酸化细胞分裂素负调节子A型ARR5正调控其蛋白稳定, 揭示了植物激素信号调控网络在植物逆境响应胁迫中的精密调控机制(Huang et al., 2018c).刘西岗研究组发现细胞分裂素的内稳态对于花分生组织(floral meristem, FM)的维持及分化非常重要.生长素可以通过促进ARF3 (Auxin Response Factor 3)的表达抑制细胞分裂素的活性进而促进FM的分化(Zhang et al., 2018e).该研究揭示了转录因子AG通过植物激素信号途径协同调控花分生组织干细胞稳态的机理.王小菁研究组发现非洲菊锌指蛋白GhWIP2通过抑制GA和IAA信号及激活ABA信号, 进而抑制细胞生长并最终抑制花瓣伸展(Ren et al., 2018b).该研究揭示了GA、ABA和生长素3种激素交叉互作网络共同调控非洲菊细胞伸展的作用机理.向成斌研究组发现乙烯响应基因HB52是乙烯和IAA信号互作的重要节点, 通过上调PIN2、WAG1和WAG2的表达抑制主根伸长(Miao et al., 2018c).该研究揭示了乙烯与IAA交互作用的新机制.丁兆军研究组研究表明, BL合成途径突变体det2-9中乙烯合成大幅增加并积累了大量的超氧阴离子.该研究揭示了BL、乙烯与活性氧协同调控根生长的新机制(Lv et al., 2018).ABA和BL信号通常相互拮抗.王学路研究组发现ABI1和ABI2通过与BIN2互作使其去磷酸化, 进而调控其对BES1的磷酸化(Wang et al., 2018c).同时, 该研究组还发现BL信号途径中的GSK3为控制水稻中胚轴伸长的一个关键因子, 其通过磷酸化调控一类细胞周期蛋白CYC U2的稳定性促进细胞分裂.BL以及独脚金内酯(strigolactone)信号通路协同调控GSK3介导的细胞周期进程, 进而促进中胚轴伸长(Sun et al., 2018c). ...
Extensive intraspecific gene order and gene structural variations between Mo17 and other maize genomes 1 2018
... 玉米是一个具有高度基因组多态性和杂种优势的重要作物.由B73自交系和Mo17自交系杂交产生的后代曾在全球范围内广泛种植.赖锦盛研究组对Mo17基因组进行了精细测序分析, 并与B73进行比较, 显示两个基因组约有10%的基因在染色体上的排序存在非共线性现象, 至少20%的基因存在可能导致蛋白编码功能变化的序列改变(Sun et al., 2018d).该研究对玉米自交系间能够形成特别显著的杂种优势原因提供了一个新解释. ...
Coordinated regulation of Arabidopsis microRNA biogenesis and red light signaling through Dicer-like 1 and phytochrome-interacting factor 4 1 2018
... 光照和温度是影响植物生长发育非常重要的两个环境因素.林荣呈研究组发现了光信号与温度信号整合的新因子SEU, 揭示了SEU通过与PIF4互作形成转录调控复合物, 对生长素合成及与信号相关的靶基因进行调节, 从而实现对生长发育的调控(Huai et al., 2018).方玉达研究组发现, 拟南芥miRNA加工复合体成员DCL1/HYL1与PIF4存在直接互作.PIF4在黑暗/红光转换过程中调控DCL1的稳定性及部分miRNA基因的转录, 进而调控光形态建成(Sun et al., 2018e).黎家研究组则发现TCP17蛋白通过调控PIF4/PIF5依赖以及非依赖的生长素合成, 从而调控遮阴诱导下植物下胚轴的伸长生长(Zhou et al., 2018d). ...
DDM1 represses noncoding RNA expression and RNA-directed DNA methylation in heterochromatin 1 2018
... 染色质重塑参与调控DNA复制、转录以及修复等多个生物学过程.染色质重塑因子可导致核小体位置和结构的改变, 引起染色质空间变化以及相关组蛋白修饰和DNA甲基化修饰变化, 但其是否参与植物DNA复制胁迫尚不清楚.刘建祥研究组针对该问题对拟南芥染色质重塑因子CHR18 (chromatin remodeling factor 18)进行了研究, 发现其功能缺失可导致拟南芥对DNA复制胁迫高度敏感.进一步分析发现, CHR18与DNA复制蛋白RPA1A在细胞核存在互作, 说明DNA复制相关蛋白RPA1A可招募染色质重塑因子CHR18到复制叉, 参与减轻DNA复制过程中遇到的逆境胁迫(Han et al., 2018).此外, 周道绣研究组对水稻染色质重塑因子DDM1 (DECREASE IN DNA METHYLATION 1)进行了研究, 揭示了OsDDM1在抑制非编码RNA (non-coding RNA)表达以及在RNA介导的DNA甲基化(RdDM)途径中的作用机制(Tan et al., 2018a). ...
Transcription factor HAT1 is a substrate of SnRK2.3 kinase and negatively regulates ABA synthesis and signaling in 1 2018
... 脱落酸(ABA)是控制植物生长和胁迫反应的关键激素.朱健康研究组发现水稻中ABA受体基因家族的特异受体基因突变导致胁迫反应减弱, 但在自然条件下表现出比野生型更好的生长状态(Miao et al., 2018a).该研究为培育高产优质水稻品种提供了新思路.林宏辉和张大伟研究组发现, 拟南芥转录因子基因HAT1及其同源基因负调控ABA的生物合成, 从而参与调控对干旱胁迫的响应(Tan et al., 2018b).该研究揭示了HAT1一种新的负调控机制, 对理解植物耐旱响应具有重要意义. ...
OsmiR396d affects gibberellin and brassinosteroid signaling to regulate plant architecture in rice 1 2018
... 水稻株型与产量密切相关.叶片作为水稻理想株型的组成要素之一, 其倾角对产量影响较大.薛红卫研究组新克隆了叶倾角调控基因LC3, 该基因编码1个含有SPOC结构域的转录抑制子, 其可与HIT锌指结构域蛋白LIP1互作, 通过调控生长素平衡和信号转导来调控植株叶倾角(Chen et al., 2018h).李建雄研究组与田志宏研究组合作揭示了OsOFP19、OSH1与DLT蛋白可形成复合体, 调控BL信号转导及影响叶倾角发育(Yang et al., 2018b).易可可研究组则解析了磷素有效性调控水稻叶倾角的分子机制.他们发现磷饥饿诱导的SPX1 (Syg1/Pho81/XPR1)与SPX2蛋白负调控叶倾角.SPX1的互作蛋白RLI1 (Regulator of leaf inclination 1)正调控叶倾角.RLI1可直接激活下游BU1 (Brassinosteroid upregulated 1)和BC1 (BU1-like1 complex 1)基因的表达以控制叶枕细胞的伸长, 从而使叶倾角增大(Ruan et al., 2018).种康和徐云远研究组证实miR396d通过分别抑制其靶基因OsGRF4和OsGRF6的转录, 促进BL及抑制GA信号途径, 进而调控水稻叶夹角和株高(Tang et al., 2018).邢永忠研究组则阐释了来自世界各地的水稻核心种质剑叶夹角的遗传基础, 发现bHLH转录因子家族中OsbHLH153和OsbHLH17能快速响应BL和IAA信号转导, 从而影响植株形态(Dong et al., 2018c).王永红研究组揭示了水稻分蘖角度调控的分子机制.他们通过研究水稻茎重力反应过程中的动态转录组变化, 探索了一条水稻分蘖角度调控的核心途径(Zhang et al., 2018h).该研究为解析水稻分蘖角度的调控网络进而挖掘有利用价值的基因提供了重要信息.谭禄宾研究组发现亚洲栽培稻PROG1基因相邻位置约有110 kb的染色体缺失.该缺失片段包含7个与PROG1基因相似的锌指基因, 其缺失推动了水稻由匍匐向直立转变, 最终实现产量大幅提高(Wu et al., 2018c). ...
The spermine synthase OsSPMS1 regulates seed germination, grain size, and yield 1 2018
... 刘建祥和罗小金研究组、宋献军研究组、何予卿研究组同时克隆到与粒型和粒重相关的同一主效负调控QTL/基因qTGW3/TGW3/GL3.3, 并解析了相关分子机制(Hu et al., 2018c; Xia et al., 2018; Ying et al., 2018).李云海研究组则与相关单位合作揭示了OsMKKK10-OsMKK4-OsMAPK6信号途径正向调控水稻粒型和粒重(Xu et al., 2018e); 并发现大粒基因LARGE8可与OsMAPK6直接互作使其失活, 进而抑制颖壳细胞增殖, 负向调控水稻粒型(Xu et al., 2018f).余四斌研究组揭示了OsGBPs作为转录因子能够识别GAGA基序, 进而调控水稻粒长和植株高度(Gong et al., 2018).梁国华研究组则证明OsSPMS1基因负向调控水稻种子萌发、粒型和单株产量, 是水稻增产育种的重要靶标基因(Tao et al., 2018).刘巧泉研究组发现了水稻粒型与外观品质调控基因GS9, 其编码蛋白可与OsOFP14和OsOFP8蛋白互作, 通过改变细胞分裂调控水稻粒型(Zhao et al., 2018c).傅向东研究组克隆了1个控制水稻产量和提升稻米品质的重要基因LGY3, 该基因编码转录因子OsMADS1.可变剪接蛋白OsMADS1lgy3可提高稻米粒长, 降低垩白率, 进而影响稻米产量和外观品质(Liu et al., 2018e). ...
Phylogeography of Eomecon chionantha in subtropical China: the dual roles of the Nanling Mountains as a glacial refugium and a dispersal corridor 1 2018
... 张志勇和范邓妹研究组则对分布于中国亚热带血水草(Eomecon chionantha)的谱系地理结构、居群历史动态和物种分布区进行了分析和模拟, 结果表明南岭在第四纪晚期不仅作为东西向的扩散通道, 而且是中国亚热带的冰期避难所(Tian et al., 2018a).该研究为充分认识山地在生物地理分布形成中扮演的角色提供了例证. ...
Hydrogen peroxide positively regulates brassinosteroid signaling through oxidation of the BRASSINAZOLE-RESISTANT1 transcription factor 1 2018
... 芸薹素(BL)是植物特有的甾醇类激素.白明义研究组发现过氧化氢能氧化BL信号途径中的关键调控因子BZR1和BES1, 增强BZR1的转录活性, 从而促进植物的细胞伸长(Tian et al., 2018b).该研究揭示了H2O2与BL协同调控植物生长发育的新机制, 为探索H2O2在植物中的功能提供了新的技术途径.李云海研究组与相关单位合作, 发现BL共受体BRI1和BAK1的互作与磷酸化受糖浓度调控.相关生化分析表明, BRI1和BAK1不仅可与G蛋白亚基互作, 还可磷酸化G蛋白亚基, 为BL途径与糖信号途径协同调控植物的生长发育建立了联系(Peng et al., 2018b).许卫锋研究组与张建华研究组合作, 发现BL受体BRI1能与质子泵AHA2蛋白互作, 进而促进根系的向水性(Miao et al., 2018b). ...
A genome for gnetophytes and early evolution of seed plants 1 2018
R2R3-MYB transcription factor MdMYB23 is involved in the cold tolerance and proanthocyanidin accumulation in apple 1 2018
... 随着全球气候的变化, 极度低温天气频发, 阶段性和局地性冷害加重, 严重影响了作物产量.因此, 开展作物耐低温研究意义重大.储成才研究组和王喜萍研究组合作发现了1个在进化过程中受到强烈选择的粳稻耐低温基因bZIP73.该基因编码区第511位单核苷酸多态性(single-nucleotide polymorphism, SNP)决定了籼、粳分化及低温的耐受性差异(Liu et al., 2018a).种康研究组则发现水稻MADS-box转录因子家族OsMADS57协同其互作蛋白OsTB1调控水稻的低温耐受性; 并发现二者对低温防御反应的调控依赖其共同靶基因OsWRKY94.常温下OsMADS57与OsTB1蛋白互作, 抑制独脚金内酯受体基因D14的转录, 促进水稻侧芽分化及分蘖形成; 低温下, OsMADS57与OsTB1通过直接激活OsWRKY94的转录, 启动防御反应.因此, OsMADS57具有平衡器官发生和防御反应的分子开关特性(Chen et al., 2018f).郝玉金和王小非研究组鉴定了苹果的R2R3-MYB转录因子MdMYB23, 发现过表达MdMYB23的转基因苹果愈伤组织和拟南芥耐寒性增强.MdMYB23可与MdCBF1和MdCBF2的启动子结合并激活其表达, 也能与原花青素生物合成的关键调节基因MdANR的启动子互作, 促进原花青素的积累和活性氧清除(An et al., 2018a).该研究揭示了MYB转录因子的功能及其在植物耐冷性调节中的潜在机制.宋波涛和谢从华研究组揭示了马铃薯精氨酸脱羧酶调控的腐胺合成是响应低温的重要途径, 首次证明转录因子CBF信号途径参与ADC1基因调控的马铃薯驯化抗寒(Kou et al., 2018).杨淑华研究组则发现了新生多肽链偶联蛋白复合体β亚基BTF3L (Basic transcription factor 3 like)正调控植物的低温应答.低温条件下, BTF3L蛋白被蛋白激酶OST1磷酸化, 导致BTF3L与CBFs蛋白互作增强, 提高了CBFs蛋白的稳定性, 进而增强植物的抗冻能力(Ding et al., 2018b).该研究揭示了OST1-BTF3L蛋白复合体调控植物响应低温胁迫的分子机制, 加深了人们对植物低温应答调控机理的理解.何跃辉研究组揭示了植物特异性支架蛋白FRI (FRIGIDA)介导拟南芥越冬习性形成的分子机制, 阐明了FRI通过超级复合体改变FLC位点的局部染色质环境, 从而激活FLC的表达, 促进转录延伸和共转录加工, 防止植物过冬前开花, 需经历冬季低温锻炼后才具备开花的潜力(Li et al., 2018k). ...
EIN3-LIKE1, MYB1, and ETHYLENE RESPONSE FACTOR 3 act in a regulatory loop that synergistically modulates ethylene biosynthesis and anthocyanin accumulation 1 2018
... EBF和EIL均是乙烯信号转导的关键蛋白, 刘明春研究组发现番茄中SlEBF3基因受SlEIL调控, 过表达SlEBF3可使SlEIL含量降低, 并表现出ETH响应相关表型(Deng et al., 2018).该研究揭示了一种新的番茄EBF基因在跃变型果实成熟中的作用, 为肉质果实成熟调控提供了基因资源.郝玉金研究组则发现苹果中MdEIL1能结合MdMYB1的启动子并激活其表达, 而MdMYB1能结合MdERF3的启动子调控乙烯的合成, 促进花青素积累与果实着色(An et al., 2018b). ...
Light signaling-dependent regulation of photoinhibition and photoprotection in tomato 1 2018
... 李继刚和康定明研究组则筛选获得了远红光信号转导新组分TZP.TZP可与phyA和FHY1互作, 且在远红光下调控phyA、FHY1和HY5的蛋白水平, 并参与调控phyA的蛋白磷酸化(Zhang et al., 2018k).李继刚研究组还与邓兴旺研究组合作证明了拟南芥phyA铰链区的3个位点(S590、T593和S602)对phyA行使功能非常重要(Zhou et al., 2018c).吴素幸研究组发现, phyA和cry感知远红光和蓝光信号, 使负调控因子COP1失活, 从而激活生长素途径在TOR依赖的RPS6磷酸化中的作用(Chen et al., 2018a).此外, 李琳研究组发现在遮阴, 尤其是严重遮阴条件下, phyA蛋白量增加, phyA与Aux/IAA互作影响了生长素受体TIR与Aux/IAA的互作, 从而阻止TIR介导的Aux/IAA降解, 导致生长素信号减弱, 通过调控光信号与生长素信号的平衡, 进而调节植物的生长发育(Yang et al., 2018c).周艳虹研究组鉴定了番茄在光抑制和光保护过程中的光信号依赖性调控通路, 分析了phyA依赖的HY5-ABI5-RBOH1信号通路在缓解冷诱导的光抑制以及光保护中的作用(Wang et al., 2018a). ...
Ecosystem nitrogen retention is regulated by plant community trait interactions with nutrient status in an alpine meadow 1 2018
... 在多种生态系统中, 氮素有效性正在急剧增加, 然而氮素水平的提高对自然生态系统中丛枝菌根(AM)真菌功能的影响目前仍不清楚.冯虎元和刘永俊研究组发现, N诱导的AM真菌群落组成变化与土壤氮素有效性和/或植物物种丰富度密切相关, 而菌根功能的变化与特定AM真菌谱系的群落相关, 非磷限制的氮富集生态系统可诱导菌根的寄生功能, 从而影响植物群落结构与生态系统的可持续性(Jiang et al., 2018c).周华坤和姚步青研究组则对植物群落特征影响生态系统N存留的方式进行了探讨, 发现植物群落特征对生态系统N存留的控制受到养分输入的作用, 并由群落水平功能性状介导(Wang et al., 2018b).该研究揭示了青藏高原高寒草甸生态系统氮素存留及其植被变化的机制, 对深刻理解高寒草地结构和功能稳定性调控及高寒草地退化演替机制具有重要意义. ...
Receptor-like cytoplasmic kinases directly link diverse pattern recognition receptors to the activation of mitogen-activated protein kinase cascades inArabidopsis 1 2018
Arabidopsis ICK/KRP cyclin-dependent kinase inhibitors function to ensure the formation of one megaspore mother cell and one functional megaspore per ovule 1 2018
MCD1 associates with FtsZ filaments via the membrane-tethering protein ARC6 to guide chloroplast division 1 2018
... 叶绿体是植物进行光合作用的细胞器, 其分裂由众多因子形成的复合物协助完成.张敏研究组发现MCD1在此过程中起重要作用.MCD1通过与ARC6作用, 识别FtsZ环, 并引导Min复合物整合到分裂环上, 从而调控叶绿体分裂(Chen et al., 2018e).欧阳敏研究组则发现ECD1蛋白通过参与叶绿体基因rps14的RNA编辑调控叶绿体早期发育(Jiang et al., 2018d).此外, 郁飞研究组和黄继荣研究组分别发现细胞质和叶绿体的翻译过程协同调控叶绿体发育, 及thf1突变体斑叶形成与质体翻译功能高度相关(Wang et al., 2018l; Huang et al., 2018b).类囊体是叶绿体基质中由单位膜围成的扁平囊, 是光合作用光反应的场所.Kang Byung-Ho研究组揭示了类囊体蛋白复合体的组装以及CURT1和FZL蛋白在类囊体生物发生过程中的关键作用(Liang et al., 2018d).隋森芳研究组与相关单位合作解析了红藻光系统I核心与捕光天线复合物(PSI-LHCR)的3.63?分辨率的三维结构, 发现红藻PSI核心既有蓝藻又有高等植物的特征(Pi et al., 2018).该研究对阐明PSI的进化和功能具有重要意义. ...
Evolutionary dynamics of 3D genome architecture following polyploidization in cotton 1 2018
... 比较基因组学和全基因组关联分析(GWAS)不仅能揭示栽培植物的起源和演化历史, 还可以为分子辅助育种提供重要参考.梅花(Prunus mume)是蔷薇科植物, 已有3000多年的栽培历史.张启翔研究组与相关单位合作完成了梅花全基因组重测序, 对几百株梅花品种的重要观赏性状进行了GWAS分析, 为揭示梅花的演化历史, 开展其分子标记辅助育种打下坚实的基础(Zhang et al., 2018j).此外, 杜雄明研究组与相关单位合作对419份陆地棉核心种质的基因组进行了重测序, 并结合多种农艺性状进行了GWAS分析, 为人工驯化选择导致棉花遗传多样性降低提供了直接的分子证据(Ma et al., 2018e).张献龙研究组与相关单位合作对四倍体棉花(陆地棉和海岛棉)进行了精细测序, 将陆地棉和海岛棉基因组的连续性分别提高了55倍和90倍(Wang et al., 2019); 且绘制了棉花二倍体和四倍体的三维基因组图谱, 鉴定了棉花基因组中大量的拓扑结构域(TAD), 揭示了三维基因组的进化与转录调控之间的关系(Wang et al., 2018h).该研究不仅促进了棉花功能基因组研究, 而且增加了人们对植物三维基因组结构的认知. ...
The proanthocyanidin-specific transcription factor MdMYBPA1 initiates anthocyanin synthesis under low-temperature conditions in red-fleshed apples 1 2018
... 类胡萝卜素是自然界广泛分布的一类次生代谢物.王涛涛研究组发现了番茄类胡萝卜素合成调控的关键转录因子SlBBX20, 其正调控番茄中类胡萝卜素的积累且稳定性受到泛素化调控(Xiong et al., 2019).邓秀新研究组则获得一个调控类胡萝卜素合成途径关键基因LCYb的转录因子CsMADS6, 并证明其可通过直接调控类胡萝卜素代谢基因LCYb1等的表达, 协同正调控类胡萝卜素代谢(Lu et al., 2018c).刘耀光研究组将双基因(sZmPSY1和sPaCrtI)、三基因(sZmPSY1、sPaCrtI和sCrBKT)和四基因(sZmPSY1、sPaCrtI、sCrBKT和sHpBHY)聚合转化水稻, 分别获得了筛选标记删除的富含黄色β-胡萝卜素、橙红色的角黄素和虾青素大米新种质(Zhu et al., 2018c).此外, 陈学森研究组鉴定了PA1型 MYB转录因子MdMYBPA1.其在正常条件下能促进苹果原花青素的合成, 但在低温胁迫下促进花青苷的合成(Wang et al., 2018i). ...
OsMADS57 together with OsTB1 coordinates transcription of its target OsWRKY94 and D14 to switch its organogenesis to defense for cold adaptation in rice 1 2018
... 随着全球气候的变化, 极度低温天气频发, 阶段性和局地性冷害加重, 严重影响了作物产量.因此, 开展作物耐低温研究意义重大.储成才研究组和王喜萍研究组合作发现了1个在进化过程中受到强烈选择的粳稻耐低温基因bZIP73.该基因编码区第511位单核苷酸多态性(single-nucleotide polymorphism, SNP)决定了籼、粳分化及低温的耐受性差异(Liu et al., 2018a).种康研究组则发现水稻MADS-box转录因子家族OsMADS57协同其互作蛋白OsTB1调控水稻的低温耐受性; 并发现二者对低温防御反应的调控依赖其共同靶基因OsWRKY94.常温下OsMADS57与OsTB1蛋白互作, 抑制独脚金内酯受体基因D14的转录, 促进水稻侧芽分化及分蘖形成; 低温下, OsMADS57与OsTB1通过直接激活OsWRKY94的转录, 启动防御反应.因此, OsMADS57具有平衡器官发生和防御反应的分子开关特性(Chen et al., 2018f).郝玉金和王小非研究组鉴定了苹果的R2R3-MYB转录因子MdMYB23, 发现过表达MdMYB23的转基因苹果愈伤组织和拟南芥耐寒性增强.MdMYB23可与MdCBF1和MdCBF2的启动子结合并激活其表达, 也能与原花青素生物合成的关键调节基因MdANR的启动子互作, 促进原花青素的积累和活性氧清除(An et al., 2018a).该研究揭示了MYB转录因子的功能及其在植物耐冷性调节中的潜在机制.宋波涛和谢从华研究组揭示了马铃薯精氨酸脱羧酶调控的腐胺合成是响应低温的重要途径, 首次证明转录因子CBF信号途径参与ADC1基因调控的马铃薯驯化抗寒(Kou et al., 2018).杨淑华研究组则发现了新生多肽链偶联蛋白复合体β亚基BTF3L (Basic transcription factor 3 like)正调控植物的低温应答.低温条件下, BTF3L蛋白被蛋白激酶OST1磷酸化, 导致BTF3L与CBFs蛋白互作增强, 提高了CBFs蛋白的稳定性, 进而增强植物的抗冻能力(Ding et al., 2018b).该研究揭示了OST1-BTF3L蛋白复合体调控植物响应低温胁迫的分子机制, 加深了人们对植物低温应答调控机理的理解.何跃辉研究组揭示了植物特异性支架蛋白FRI (FRIGIDA)介导拟南芥越冬习性形成的分子机制, 阐明了FRI通过超级复合体改变FLC位点的局部染色质环境, 从而激活FLC的表达, 促进转录延伸和共转录加工, 防止植物过冬前开花, 需经历冬季低温锻炼后才具备开花的潜力(Li et al., 2018k). ...
Plant diversity enhances productivity and soil carbon storage 1 2018
Alternative splicing plays a critical role in maintaining mineral nutrient homeostasis in rice (Oryza sativa) 1 2018
... 可变剪接是生物体内普遍存在的现象, 但是关于可变剪接是否也参与调控植物响应养分胁迫一直不很清楚.郑录庆研究组对不同矿质元素(Fe、Zn、Cu、Mn和P)缺乏条件下的水稻RNA-Seq数据进行系统分析, 结果表明可变剪接调控是植物响应营养缺乏的重要调控机制之一.丝氨酸精氨酸丰富(SR)蛋白是参与pre-mRNAs剪接的重要RNA结合蛋白.研究发现3个SR蛋白可能参与调控水稻对磷的吸收, 以及磷在叶片和地上部的再分配(Dong et al., 2018a).该研究首次揭示了可变剪接在植物响应养分缺乏中的重要作用, 为该领域提供了新的研究思路. ...
Modulation of guard cell turgor and drought tolerance by a pero-xisomal acetate-malate shunt 1 2018
... 植物的生长离不开合适的水分条件.干旱由于其发生频率高、周期长和影响范围广等特点, 成为我国农业生产的主要气象灾害.碱性亮氨酸拉链bZIP转录因子参与植物发育与胁迫响应.然而, 其在玉米中的功能研究尚少.张举仁研究组发现, 玉米bZIP转录因子ZmbZIP4不仅正调控逆境激素ABA的合成, 还通过调控根发育相关基因的表达, 正调控侧根的数量及主根伸长, 从而增强玉米抵抗干旱和盐胁迫的能力(Ma et al., 2018b).李自超研究组报道了ERF家族转录因子OsLG3正向调控水稻的干旱抗性, 揭示了OsLG3优质等位基因是水稻抗干旱品种育种研究中重要的遗传资源(Xiong et al., 2018).宋纯鹏研究组分离出一种与野生型植株相比气孔变小且抗旱性增强的拟南芥突变体bzu1.BZU1编码1种已知的乙酰辅酶A合成酶ACN1, 其在过氧化物酶体中乙酸转化为苹果酸代谢途径的第1步起作用.研究表明, BZU1/ACN1介导的乙酸-苹果酸旁路通过控制拟南芥保卫细胞的膨压进而调节其抗旱性(Dong et al., 2018b).熊立仲和杨万能研究组通过GWAS (genome-wide association study)方法分析了529个代表性的水稻种质资源, 为遗传解析和发现抗旱基因提供了一种新方法(Guo et al., 2018d).同时, 熊立仲研究组还探究了植物逃旱的分子机制, 发现轻度水分胁迫可诱发水稻发育早期提早开花和减少分蘖数, 阐明了水稻逃旱性由依赖以及不依赖ABA的多条途径协调控制(Du et al., 2018a). ...
GL3.3, a novel QTL encoding a GSK3/SHAGGY-like kinase, epistatically interacts with GS3 to produce extra-long grains in rice 1 2018
... 刘建祥和罗小金研究组、宋献军研究组、何予卿研究组同时克隆到与粒型和粒重相关的同一主效负调控QTL/基因qTGW3/TGW3/GL3.3, 并解析了相关分子机制(Hu et al., 2018c; Xia et al., 2018; Ying et al., 2018).李云海研究组则与相关单位合作揭示了OsMKKK10-OsMKK4-OsMAPK6信号途径正向调控水稻粒型和粒重(Xu et al., 2018e); 并发现大粒基因LARGE8可与OsMAPK6直接互作使其失活, 进而抑制颖壳细胞增殖, 负向调控水稻粒型(Xu et al., 2018f).余四斌研究组揭示了OsGBPs作为转录因子能够识别GAGA基序, 进而调控水稻粒长和植株高度(Gong et al., 2018).梁国华研究组则证明OsSPMS1基因负向调控水稻种子萌发、粒型和单株产量, 是水稻增产育种的重要靶标基因(Tao et al., 2018).刘巧泉研究组发现了水稻粒型与外观品质调控基因GS9, 其编码蛋白可与OsOFP14和OsOFP8蛋白互作, 通过改变细胞分裂调控水稻粒型(Zhao et al., 2018c).傅向东研究组克隆了1个控制水稻产量和提升稻米品质的重要基因LGY3, 该基因编码转录因子OsMADS1.可变剪接蛋白OsMADS1lgy3可提高稻米粒长, 降低垩白率, 进而影响稻米产量和外观品质(Liu et al., 2018e). ...
The effect of Hengduan Mountains Region (HMR) uplift to environmental changes in the HMR and its eastern adjacent area: tracing the evolutionary history of Allium section Sikkimensia( Amaryllidaceae) 1 2019
Genome-wide association studies reveal that members of bHLH subfamily 16 share a conserved function in regulating flag leaf angle in rice (Oryza sativa) 1 2018
... 水稻株型与产量密切相关.叶片作为水稻理想株型的组成要素之一, 其倾角对产量影响较大.薛红卫研究组新克隆了叶倾角调控基因LC3, 该基因编码1个含有SPOC结构域的转录抑制子, 其可与HIT锌指结构域蛋白LIP1互作, 通过调控生长素平衡和信号转导来调控植株叶倾角(Chen et al., 2018h).李建雄研究组与田志宏研究组合作揭示了OsOFP19、OSH1与DLT蛋白可形成复合体, 调控BL信号转导及影响叶倾角发育(Yang et al., 2018b).易可可研究组则解析了磷素有效性调控水稻叶倾角的分子机制.他们发现磷饥饿诱导的SPX1 (Syg1/Pho81/XPR1)与SPX2蛋白负调控叶倾角.SPX1的互作蛋白RLI1 (Regulator of leaf inclination 1)正调控叶倾角.RLI1可直接激活下游BU1 (Brassinosteroid upregulated 1)和BC1 (BU1-like1 complex 1)基因的表达以控制叶枕细胞的伸长, 从而使叶倾角增大(Ruan et al., 2018).种康和徐云远研究组证实miR396d通过分别抑制其靶基因OsGRF4和OsGRF6的转录, 促进BL及抑制GA信号途径, 进而调控水稻叶夹角和株高(Tang et al., 2018).邢永忠研究组则阐释了来自世界各地的水稻核心种质剑叶夹角的遗传基础, 发现bHLH转录因子家族中OsbHLH153和OsbHLH17能快速响应BL和IAA信号转导, 从而影响植株形态(Dong et al., 2018c).王永红研究组揭示了水稻分蘖角度调控的分子机制.他们通过研究水稻茎重力反应过程中的动态转录组变化, 探索了一条水稻分蘖角度调控的核心途径(Zhang et al., 2018h).该研究为解析水稻分蘖角度的调控网络进而挖掘有利用价值的基因提供了重要信息.谭禄宾研究组发现亚洲栽培稻PROG1基因相邻位置约有110 kb的染色体缺失.该缺失片段包含7个与PROG1基因相似的锌指基因, 其缺失推动了水稻由匍匐向直立转变, 最终实现产量大幅提高(Wu et al., 2018c). ...
Resolving the systematic positions of enigmatic taxa: manipulating the chloroplast genome data of Saxifragales 1 2018
The chimeric mitochondrial gene orf182 causes non-pollen-type abortion in Dongxiang cytoplasmic male-sterile rice 1 2018
... 目前, 中国科学家利用种间、亚种间和品种间杂交已先后培育了以D型等为代表的60余种水稻细胞质雄性不育系.D1型是一种新型孢子体细胞质雄性不育类型, 无花粉型败育, 且为中国所特有.颜龙安和蔡耀辉研究组成功克隆了D1型水稻细胞质雄性不育基因并揭示了其败育机理(Xie et al., 2018).漆小泉研究组则发现了一种新型水稻雄性不育系, 并且阐释了OsOSC12/OsPTS1在水稻花粉包被形成过程中的关键作用, 对禾本科作物湿敏雄性不育材料的培育具有重要指导意义(Xue et al., 2018d). ...
A tomato B-box protein SlBBX20 modulates carotenoid biosynthesis by directly activating PHYTOENE SYNTHASE 1, and is targeted for 26S proteasome-mediated degradation 1 2019
... 类胡萝卜素是自然界广泛分布的一类次生代谢物.王涛涛研究组发现了番茄类胡萝卜素合成调控的关键转录因子SlBBX20, 其正调控番茄中类胡萝卜素的积累且稳定性受到泛素化调控(Xiong et al., 2019).邓秀新研究组则获得一个调控类胡萝卜素合成途径关键基因LCYb的转录因子CsMADS6, 并证明其可通过直接调控类胡萝卜素代谢基因LCYb1等的表达, 协同正调控类胡萝卜素代谢(Lu et al., 2018c).刘耀光研究组将双基因(sZmPSY1和sPaCrtI)、三基因(sZmPSY1、sPaCrtI和sCrBKT)和四基因(sZmPSY1、sPaCrtI、sCrBKT和sHpBHY)聚合转化水稻, 分别获得了筛选标记删除的富含黄色β-胡萝卜素、橙红色的角黄素和虾青素大米新种质(Zhu et al., 2018c).此外, 陈学森研究组鉴定了PA1型 MYB转录因子MdMYBPA1.其在正常条件下能促进苹果原花青素的合成, 但在低温胁迫下促进花青苷的合成(Wang et al., 2018i). ...
Natural variation in OsLG3 increases drought tolerance in rice by inducing ROS scavenging 1 2018
... 植物的生长离不开合适的水分条件.干旱由于其发生频率高、周期长和影响范围广等特点, 成为我国农业生产的主要气象灾害.碱性亮氨酸拉链bZIP转录因子参与植物发育与胁迫响应.然而, 其在玉米中的功能研究尚少.张举仁研究组发现, 玉米bZIP转录因子ZmbZIP4不仅正调控逆境激素ABA的合成, 还通过调控根发育相关基因的表达, 正调控侧根的数量及主根伸长, 从而增强玉米抵抗干旱和盐胁迫的能力(Ma et al., 2018b).李自超研究组报道了ERF家族转录因子OsLG3正向调控水稻的干旱抗性, 揭示了OsLG3优质等位基因是水稻抗干旱品种育种研究中重要的遗传资源(Xiong et al., 2018).宋纯鹏研究组分离出一种与野生型植株相比气孔变小且抗旱性增强的拟南芥突变体bzu1.BZU1编码1种已知的乙酰辅酶A合成酶ACN1, 其在过氧化物酶体中乙酸转化为苹果酸代谢途径的第1步起作用.研究表明, BZU1/ACN1介导的乙酸-苹果酸旁路通过控制拟南芥保卫细胞的膨压进而调节其抗旱性(Dong et al., 2018b).熊立仲和杨万能研究组通过GWAS (genome-wide association study)方法分析了529个代表性的水稻种质资源, 为遗传解析和发现抗旱基因提供了一种新方法(Guo et al., 2018d).同时, 熊立仲研究组还探究了植物逃旱的分子机制, 发现轻度水分胁迫可诱发水稻发育早期提早开花和减少分蘖数, 阐明了水稻逃旱性由依赖以及不依赖ABA的多条途径协调控制(Du et al., 2018a). ...
Control of auxin-induced callus formation by bZIP59-LBD complex in Arabidopsis regeneration 1 2018
... 生长素在植物生长发育过程中起重要作用.赵忠研究组发现拟南芥茎顶端分生组织中的生长素具有促进干细胞分化的功能, 并维持干细胞的稳态, 揭示了生长素维持干细胞数目稳定的新机制(Luo et al., 2018b).焦雨铃研究组与张磊研究组合作, 通过学科交叉研究发现了植物侧生器官可反馈调控茎尖干细胞, 并表明生长素长距离运输在反馈中发挥了重要作用(Shi et al., 2018a).该研究有机整合了生物学与数学, 发挥了学科交叉研究解决动态生物学过程等前沿问题的优势.胡玉欣研究组对生长素启动愈伤组织形成的分子机制进行了探索, 揭示了控制愈伤组织形成中体细胞重编程的关键因子, 并建立了植物再生体系中生长素信号与细胞全能性获得的分子联系(Xu et al., 2018a).该研究为探索植物再生的分子调控机制和发育程序提供了新线索. ...
Ethylene signaling modulates cortical microtubule reassembly in response to salt stress 1 2018
... 细胞骨架(cytoskeleton)系统主要包括微管(MTs)和微丝(F-actin), 其在调控植物生长发育和细胞形态建成等生理过程中具重要作用.然而, 人们对微管和微丝的调控机理及其影响植物形态发生发育机制的认识还十分有限.高等植物已进化出多种(不同大小、形状和功能)细胞类型, 如锥形花瓣表皮细胞, 然而调控该种细胞形态建成的分子机制目前尚不清楚.林德书研究组证实AN (ANGUSTIFOLIA)-ROS途径与KTN1 (katanin)共同作用, 通过参与调节微管骨架的排列方式, 进而调控花瓣锥形细胞发育(Dang et al., 2018).黄善金研究组则发现, 拟南芥成蛋白AtFH3 (Formin 3)和AtFH5参与调节花粉管顶端肌动蛋白的聚合和阵列构建, 对极性花粉管快速生长起关键作用(Lan et al., 2018).此外, 他们还发现拟南芥成蛋白AtFH2特异定位于胞间连丝, 水稻的部分Formin家族成员也定位于胞间连丝, 该研究揭示了拟南芥胞间连丝处微丝的含量及稳定性对调控胞间连丝通透性具重要作用(Diao et al., 2018).毛同林研究组从植物细胞骨架与环境因子互作方面进行了探索, 揭示了植物细胞受到高盐胁迫后, 乙烯通过上调微管稳定蛋白WDL5的表达调控微管重组(Dou et al., 2018), 及微管去稳定蛋白MDP60参与光和乙烯调控的下胚轴伸长(Ma et al., 2018c).孔照胜研究组则利用标记了微丝骨架(actin cytoskeleton)的稳定转基因蒺藜苜蓿, 首次在活细胞水平解析了根瘤发育不同阶段, 微丝骨架调控共生界面形成过程中的排布结构和动态模式, 为探索根瘤发育和生物固氮提供了细胞生物学依据(Zhang et al., 2019). ...
Integrative regulation of drought escape through ABA- dependent and -independent pathways in rice 1 2018
... 植物的生长离不开合适的水分条件.干旱由于其发生频率高、周期长和影响范围广等特点, 成为我国农业生产的主要气象灾害.碱性亮氨酸拉链bZIP转录因子参与植物发育与胁迫响应.然而, 其在玉米中的功能研究尚少.张举仁研究组发现, 玉米bZIP转录因子ZmbZIP4不仅正调控逆境激素ABA的合成, 还通过调控根发育相关基因的表达, 正调控侧根的数量及主根伸长, 从而增强玉米抵抗干旱和盐胁迫的能力(Ma et al., 2018b).李自超研究组报道了ERF家族转录因子OsLG3正向调控水稻的干旱抗性, 揭示了OsLG3优质等位基因是水稻抗干旱品种育种研究中重要的遗传资源(Xiong et al., 2018).宋纯鹏研究组分离出一种与野生型植株相比气孔变小且抗旱性增强的拟南芥突变体bzu1.BZU1编码1种已知的乙酰辅酶A合成酶ACN1, 其在过氧化物酶体中乙酸转化为苹果酸代谢途径的第1步起作用.研究表明, BZU1/ACN1介导的乙酸-苹果酸旁路通过控制拟南芥保卫细胞的膨压进而调节其抗旱性(Dong et al., 2018b).熊立仲和杨万能研究组通过GWAS (genome-wide association study)方法分析了529个代表性的水稻种质资源, 为遗传解析和发现抗旱基因提供了一种新方法(Guo et al., 2018d).同时, 熊立仲研究组还探究了植物逃旱的分子机制, 发现轻度水分胁迫可诱发水稻发育早期提早开花和减少分蘖数, 阐明了水稻逃旱性由依赖以及不依赖ABA的多条途径协调控制(Du et al., 2018a). ...
The B-box domain protein BBX21 promotes photomorphogenesis 1 2018
... 邓兴旺研究组与许冬清研究组合作鉴定了光形态建成的正调节因子CSU4, 它能整合生物钟节律和光信号(Zhao et al., 2018g).他们还发现BBX21通过第2个B-box结构域与HY5启动子中的T/G-box结合, 调控HY5及其靶基因的表达以促进植物光形态建成(Xu et al., 2018b).而BBX28通过与HY5蛋白的C末端互作, 抑制HY5与下游靶基因启动子结合, 从而抑制HY5活性及其下游靶基因表达, 最终抑制植物光形态建成(Lin et al., 2018a).此外, 邓兴旺和钟上威研究组揭示了PIFs、HY5和EIN3及EIL1对出土幼苗形态建成的调控作用, 为阐明幼苗从暗形态建成转向光形态建成的分子机制提供了新证据(Shi et al., 2018b).吕应堂研究组则鉴定了光形态建成的正调控因子SRS5, 它直接结合HY5、BBX21和BBX22的启动子并激活其表达, 从而促进光形态建成(Yuan et al., 2018).刘宏涛研究组发现UV-B照射后, UVR8形成单体并在细胞核中富集, UVR8通过抑制BES1/BIM1结合DNA的能力从而抑制下游生长相关基因的表达, 并抑制细胞伸长及光形态建成(Liang et al., 2018b).同时, 发现UV-B激活的UVR8形成单体, 进入细胞核直接结合WRKY36而抑制其结合HY5启动子, 进而抑制下胚轴伸长(Yang et al., 2018g). ...
Photoactivated CRY1 and phyB interact directly with AUX/IAA proteins to inhibit auxin signaling in Arabidopsis 1 2018
... 光对植物不仅是一种能源, 也是一种重要的环境信号.植物通过向光素(phot)、隐花色素(cry)和光敏色素(phy)等光受体来感应光信号.林金星研究组研究了蓝光对细胞质膜上向光素phot1的定位变化和动态性质, 在单分子水平上揭示了蓝光信号传递的起始事件及调控机制(Xue et al., 2018b).刘宏涛研究组发现了隐花色素互作转录因子CIB1通过直接结合cry2和CO来介导cry2-CIB1-CO复合体的形成, 揭示了蓝光信号调控植物开花时间的新机制(Liu et al., 2018g).杨洪全研究组发现, G蛋白β亚基AGB1通过与HY5互作抑制其DNA结合能力, 从而抑制光信号; 而蓝光信号通过CRY1与AGB1互作促进AGB1与HY5解离来抑制AGB1的功能(Lian et al., 2018); 并发现CRY1与去磷酸化形式的BL信号关键转录因子BES1发生依赖于蓝光的直接互作, 揭示了植物能根据光信号和内在BL信号有无或强弱的动态变化来优化其光形态建成(Wang et al., 2018n).此外, 该研究组还发现光受体cry1、phyB竞争性与生长素信号通路抑制子Aux/IAA结合, 并磷酸化后者而负调控生长素信号转导, 进而精确调控植物光信号与生长素信号通路的平衡, 优化其生长(Xu et al., 2018c). ...
Rice transcription factor OsMADS25 modulates root growth and confers salinity tolerance via the ABA- mediated regulatory pathway and ROS scavenging 2 2018
... 土壤盐碱化极大地限制了粮食作物的稳产增收.如何将盐碱地变为高产粮田, 成为当下农业科学家研究的热点方向, 而解析植物应答盐碱胁迫的分子机制是提高植物抗盐碱的有效手段.朱健康研究组发现, 细胞壁LRX (Leucine-rich repeat extensins)蛋白家族多突变体植株表现出对盐胁迫非常敏感的表型.进一步研究发现, LRX蛋白与RALF多肽以及细胞膜受体类激酶FERONIA形成一个元件调控植物生长和耐盐性(Zhao et al., 2018b).该研究为培育抗逆高产作物指明了方向.此外, 朱健康和王镇研究组还探明了ABA对植物抗旱性的影响, 揭示了开花抑制子SVP (Short vegetative phase)通过调控ABA代谢提高拟南芥的抗旱性机制(Wang et al., 2018r).植物类受体激酶能感受外界环境胁迫信号, 并通过胞内激酶域激活下游效应子活性和调控细胞相关生理生化过程, 从而降低或消除逆境胁迫的影响.刘选明和林建中研究组破译了1个能降低土地盐碱化对水稻产量影响的类受体胞质激酶1 (salt tolerance receptor-like cytoplasmic kinase 1, STRK1), 其可通过磷酸化和激活CatC调节H2O2体内平衡来改善盐以及氧化胁迫的耐受性(Zhou et al., 2018e).该研究为进一步解析植物耐盐的分子机制奠定了基础.范六民研究组报道了MYB亚家族转录因子RSM1 (Radialis-like sant/myb 1)蛋白与HY5/HYH蛋白互作, 并作用于ABI5启动子区, 调控ABA和非生物逆境(盐)胁迫相关基因的应答(Yang et al., 2018a).黄俊丽研究组则对水稻MADS-box家族转录因子OsMADS25蛋白进行了研究, 发现其可与谷胱甘肽S-转移酶基因OsGST4的启动子直接结合, 通过ABA介导的调控通路和ROS清除共同调控水稻的根系生长和耐盐性(Xu et al., 2018d).蒋才富研究组利用QTL克隆了调控叶片Na+含量的主效基因ZmNC1 (Na+ content 1).该基因编码1个Na+选择性离子转运蛋白ZmHKT1, 其可通过抑制Na+由根向地上部运输提高玉米的耐盐能力(Zhang et al., 2018g).郝玉金研究组则发现苹果葡萄糖感知因子己糖激酶MdHXK1可磷酸化Na+/H+交换蛋白MdNHX1, 以提高其蛋白稳定性, 进而增强苹果的耐盐性(Sun et al., 2018b). ...
... 植物的生长离不开合适的水分条件.干旱由于其发生频率高、周期长和影响范围广等特点, 成为我国农业生产的主要气象灾害.碱性亮氨酸拉链bZIP转录因子参与植物发育与胁迫响应.然而, 其在玉米中的功能研究尚少.张举仁研究组发现, 玉米bZIP转录因子ZmbZIP4不仅正调控逆境激素ABA的合成, 还通过调控根发育相关基因的表达, 正调控侧根的数量及主根伸长, 从而增强玉米抵抗干旱和盐胁迫的能力(Ma et al., 2018b).李自超研究组报道了ERF家族转录因子OsLG3正向调控水稻的干旱抗性, 揭示了OsLG3优质等位基因是水稻抗干旱品种育种研究中重要的遗传资源(Xiong et al., 2018).宋纯鹏研究组分离出一种与野生型植株相比气孔变小且抗旱性增强的拟南芥突变体bzu1.BZU1编码1种已知的乙酰辅酶A合成酶ACN1, 其在过氧化物酶体中乙酸转化为苹果酸代谢途径的第1步起作用.研究表明, BZU1/ACN1介导的乙酸-苹果酸旁路通过控制拟南芥保卫细胞的膨压进而调节其抗旱性(Dong et al., 2018b).熊立仲和杨万能研究组通过GWAS (genome-wide association study)方法分析了529个代表性的水稻种质资源, 为遗传解析和发现抗旱基因提供了一种新方法(Guo et al., 2018d).同时, 熊立仲研究组还探究了植物逃旱的分子机制, 发现轻度水分胁迫可诱发水稻发育早期提早开花和减少分蘖数, 阐明了水稻逃旱性由依赖以及不依赖ABA的多条途径协调控制(Du et al., 2018a). ...
Control of grain size and weight by the OsMKKK10-OsMKK4-OsMAPK6 signaling pathway in rice 1 2018
... 刘建祥和罗小金研究组、宋献军研究组、何予卿研究组同时克隆到与粒型和粒重相关的同一主效负调控QTL/基因qTGW3/TGW3/GL3.3, 并解析了相关分子机制(Hu et al., 2018c; Xia et al., 2018; Ying et al., 2018).李云海研究组则与相关单位合作揭示了OsMKKK10-OsMKK4-OsMAPK6信号途径正向调控水稻粒型和粒重(Xu et al., 2018e); 并发现大粒基因LARGE8可与OsMAPK6直接互作使其失活, 进而抑制颖壳细胞增殖, 负向调控水稻粒型(Xu et al., 2018f).余四斌研究组揭示了OsGBPs作为转录因子能够识别GAGA基序, 进而调控水稻粒长和植株高度(Gong et al., 2018).梁国华研究组则证明OsSPMS1基因负向调控水稻种子萌发、粒型和单株产量, 是水稻增产育种的重要靶标基因(Tao et al., 2018).刘巧泉研究组发现了水稻粒型与外观品质调控基因GS9, 其编码蛋白可与OsOFP14和OsOFP8蛋白互作, 通过改变细胞分裂调控水稻粒型(Zhao et al., 2018c).傅向东研究组克隆了1个控制水稻产量和提升稻米品质的重要基因LGY3, 该基因编码转录因子OsMADS1.可变剪接蛋白OsMADS1lgy3可提高稻米粒长, 降低垩白率, 进而影响稻米产量和外观品质(Liu et al., 2018e). ...
Endosperm sugar accumulation caused by mutation of PHS8/ISA1 leads to pre-harvest sprouting in rice 1 2018
... 储成才研究组克隆了穗发芽调控基因PHS8, 该基因突变可导致胚乳中小分子糖积累, 从而抑制ABA信号通路中OsABI3和OsABI5的表达, 导致穗发芽表型(Du et al., 2018b).刘春明研究组与相关单位合作创建了一种半粒种子筛选体系, 并利用该体系筛检获得了1个糊粉层增厚的水稻品系ta2.基因克隆发现DNA去甲基化酶基因OsROS1与糊粉层的发育相关, 有望开拓水稻营养品质育种新路径(Liu et al., 2018d). ...
A banana R2R3-MYB transcription factor MaMYB3 is involved in fruit ripening through modulation of starch degradation by repressing starch degradation-rela- ted genes andMabHLH6 1 2018
... 植物分枝是影响植物株型的关键因素.秦根基研究组与相关单位合作对植物分枝形成的分子调控机制进行研究, 发现了1个分枝正调控因子TIE1, 它与分枝重要调控因子BRC1 (BRANCHED1)发生互作, 抑制其转录活性.通过调节HB21、HB40和HB53等BRC1下游基因的转录, 发挥促进腋芽分枝的作用(Yang et al., 2018h).开花是植物由营养生长转向生殖生长的标志.王前和张小兰研究组对热诱导开花的分子机理进行探索, 证明LsSOC1是热诱导花芽形成的激活子, 热激转录因子HsfA1e和HsfA4c通过结合在LsSOC1基因的启动子上, 影响其表达, 为开花相关机制研究奠定了理论基础(Chen et al., 2018k).果实成熟过程对于果实品质形成和种子传播非常关键.陈建业和邝健飞研究组发现转录因子MaMYB3通过抑制淀粉降解相关基因以及MabHLH6转录因子的转录水平来调控淀粉降解, 丰富了香蕉果实成熟的转录调控网络(Fan et al., 2018).朱本忠和罗云波研究组则发现, 在番茄rin突变体中, 基因组DNA片段部分缺失导致形成RIN-MC片段的嵌合, 该嵌合能够调控许多成熟相关基因的表达, 从而调节果实成熟(Li et al., 2018h).此外, 吴鸿研究组与相关单位合作阐明了拟南芥角果开裂的调控机制.他们通过对纤维素酶CEL6和半纤维素酶MAN7以及果胶酶ADPG2基因的一系列突变体果荚离层进行细胞超微结构观察, 并结合基因表达的检测结果, 证明改变这些细胞壁酶的表达量和酶活性可以调控果荚开裂区细胞分化和离层区细胞降解(He et al., 2018a).该研究为后期培育不同程度的果实抗开裂油料作物提供了技术支持. ...
A mitogen-activated protein kinase phosphatase influences grain size and weight in rice 1 2018
... 刘建祥和罗小金研究组、宋献军研究组、何予卿研究组同时克隆到与粒型和粒重相关的同一主效负调控QTL/基因qTGW3/TGW3/GL3.3, 并解析了相关分子机制(Hu et al., 2018c; Xia et al., 2018; Ying et al., 2018).李云海研究组则与相关单位合作揭示了OsMKKK10-OsMKK4-OsMAPK6信号途径正向调控水稻粒型和粒重(Xu et al., 2018e); 并发现大粒基因LARGE8可与OsMAPK6直接互作使其失活, 进而抑制颖壳细胞增殖, 负向调控水稻粒型(Xu et al., 2018f).余四斌研究组揭示了OsGBPs作为转录因子能够识别GAGA基序, 进而调控水稻粒长和植株高度(Gong et al., 2018).梁国华研究组则证明OsSPMS1基因负向调控水稻种子萌发、粒型和单株产量, 是水稻增产育种的重要靶标基因(Tao et al., 2018).刘巧泉研究组发现了水稻粒型与外观品质调控基因GS9, 其编码蛋白可与OsOFP14和OsOFP8蛋白互作, 通过改变细胞分裂调控水稻粒型(Zhao et al., 2018c).傅向东研究组克隆了1个控制水稻产量和提升稻米品质的重要基因LGY3, 该基因编码转录因子OsMADS1.可变剪接蛋白OsMADS1lgy3可提高稻米粒长, 降低垩白率, 进而影响稻米产量和外观品质(Liu et al., 2018e). ...
Genome analysis of the ancient tracheophyte Selaginella tamariscina reveals evolutionary features relevant to the acquisition of desiccation tolerance 1 2018
... 早期陆生植物形态结构相对简单, 但环境适应能力强.特有基因对物种适应特殊环境发挥了重要作用.黄锦岭研究组对小立碗藓的hemerythrin基因进行了研究, 发现其起源于真菌向早期陆生植物的水平基因转移事件.该基因在小立碗藓中通过改变移码突变, 演化出1个双编码基因YAN/AltYAN, 该基因介导了小立碗藓的脱水耐受性(Guan et al., 2018).该研究对揭示陆生植物如何适应特殊环境提供了新观点及实验证据.基因组学研究为揭示一些物种特异现象提供了很好的切入点.例如, 卷柏(Selaginella tamariscina)是古老的复苏植物, 干旱时能存活数十年, 遇水而荣, 与已测序的其同属植物江南卷柏(S. moellendorffii) (干旱敏感型)截然不同.陈士林和宋经元研究组破译了卷柏基因组, 绘制了高杂合度卷柏的基因组图谱, 并与已发表的江南卷柏基因组进行了比较, 揭示了卷柏耐旱和复苏机制(Xu et al., 2018g).维管系统(木质部等)是植物在水生到陆生的进化过程中产生的.拟南芥和小立碗藓中特定的NAC转录因子调控木质部发育以及水分运输.景海春研究组与才宏伟研究组合作从甜高粱中鉴定到1个在驯化过程中受到选择、控制茎秆持汁性的Dry基因(编码一个NAC转录因子), 并发现其功能缺失是甜高粱茎秆富含汁液的重要原因(Zhang et al., 2018f).该研究不仅对促进作物茎秆持汁表型的遗传改良具重要作用, 而且为作物育种获得籽粒及秸秆“双丰收”提供了可能. ...
OPAQUE11 is a central hub of the regulatory network for maize endosperm development and nutrient metabolism 1 2018
... 玉米胚乳是营养物质的主要储存组织.宋任涛研究组克隆了玉米突变体o11 (opaque11)的胚乳特异性bHLH转录因子O11.研究表明其不仅可直接调控胚乳发育的转录因子、cyPPDKs以及多个碳水化合物代谢酶, 还是ZmYoda的激活因子, 在胚乳发育和营养代谢调节网络中发挥中心调控作用(Feng et al., 2018); 另外, 该研究组还鉴定了1个与玉米储藏蛋白27 kDa γ-zein启动子相结合的bZIP型转录因子ZmbZIP22.该转录因子与PBF1、OHP1以及OHP2一起, 对27 kDa γ-zein进行转录调节, 影响胚乳发育(Li et al., 2018a). ...
Divergent functions of the GAGA-binding transcription factor family in rice 1 2018
... 刘建祥和罗小金研究组、宋献军研究组、何予卿研究组同时克隆到与粒型和粒重相关的同一主效负调控QTL/基因qTGW3/TGW3/GL3.3, 并解析了相关分子机制(Hu et al., 2018c; Xia et al., 2018; Ying et al., 2018).李云海研究组则与相关单位合作揭示了OsMKKK10-OsMKK4-OsMAPK6信号途径正向调控水稻粒型和粒重(Xu et al., 2018e); 并发现大粒基因LARGE8可与OsMAPK6直接互作使其失活, 进而抑制颖壳细胞增殖, 负向调控水稻粒型(Xu et al., 2018f).余四斌研究组揭示了OsGBPs作为转录因子能够识别GAGA基序, 进而调控水稻粒长和植株高度(Gong et al., 2018).梁国华研究组则证明OsSPMS1基因负向调控水稻种子萌发、粒型和单株产量, 是水稻增产育种的重要靶标基因(Tao et al., 2018).刘巧泉研究组发现了水稻粒型与外观品质调控基因GS9, 其编码蛋白可与OsOFP14和OsOFP8蛋白互作, 通过改变细胞分裂调控水稻粒型(Zhao et al., 2018c).傅向东研究组克隆了1个控制水稻产量和提升稻米品质的重要基因LGY3, 该基因编码转录因子OsMADS1.可变剪接蛋白OsMADS1lgy3可提高稻米粒长, 降低垩白率, 进而影响稻米产量和外观品质(Liu et al., 2018e). ...
Plant functional diversity modulates global environmental change effects on grassland productivity 1 2018
... 青藏高原近年来经历了快速(2倍于全球平均值)升温过程, 且降水格局也发生了显著变化, 然而剧烈的气候变化将如何影响高寒草地生态系统的结构和功能, 一直以来备受国内外研究者的关注.贺金生研究组发现尽管过去32年来海北站气候呈现暖干化趋势, 但草地生产力无显著变化, 功能群组成改变是生产力稳定性维持的主要原因(Liu et al., 2018c).该研究表明物种多样性在气候变化下生产力维持过程中起着关键作用.姜勇研究组与相关单位合作探讨了环境变化影响我国温带典型草原生产力的直接和间接过程.他们发现增加氮、水不仅直接提高生产力, 而且还通过增加植物功能多样性的2个组分间接驱动生产力(Xu et al., 2018h).该研究表明群落中优势物种的性状及物种间的性状分布在调节生态系统功能中均具重要作用. ...
The number of meiotic double-strand breaks influences crossover distribution in Arabidopsis 1 2018
... 减数分裂是真核生物有性生殖所必需的环节.程祝宽和李亚非研究组对水稻减数分裂时胞质分裂的遗传调控机制进行了探索.鉴定出1个调控花粉母细胞胞质分裂的蛋白DCM1 (Defective Callose in Meiosis 1).该蛋白的C端含有5个串联的CCCH锌指结构域, 并与核多聚腺苷酸结合蛋白OsPABNs存在互作.dcm1突变体的花粉母细胞在胞质分裂时, 由于胼胝质板提前降解, 胞质分裂无法进行, 使得减数第二次分裂的纺锤体取向异常, 并最终表现为雄性不育(Zhang et al., 2018a).此外, 程祝宽研究组在水稻中成功分离了1个新的减数分裂交叉结形成相关蛋白HEIP1 (HEI10 Interaction Protein 1).实验证明该蛋白与HEI10、MSH5和ZIP4等交叉结形成促进因子相互作用并共定位, 调控减数分裂一类交叉的形成(Li et al., 2018j).该研究为深入揭示减数分裂同源重组形成的分子机制奠定了基础.该研究组还与马伯军研究组合作通过筛选水稻减数分裂缺陷的不育突变体, 发现LEPTO1 (LEPTOTENE1)参与水稻减数分裂细线期染色体的形态建成.lepto1突变体的花粉母细胞染色体停滞在前细线期状态, 不能进一步组装进入典型的细线期染色体状态.性母细胞中没有DSB形成, 也没有减数分裂特异蛋白的组装, 观察不到胼胝质的积累.LEPTO1编码OsRR24, 属于B类响应调节因子, N端具有保守的DDK和MYB结构域, C端具有转录激活活性, 表明LEPTO1可能通过调控相关基因的表达, 进而调节减数分裂细线期染色体的形态建成(Zhao et al., 2018f).王应祥研究组与贺岩研究组合作探究了减数分裂重组过程中DNA双链断裂(double- strand breaks, DSB)的数目对重组稳态和最终分布的影响.他们通过转基因技术在DSB缺失突变体(spo11-/-)背景下操控DSB发生的数目, 揭示了减数分裂中DSB形成的数目对重组的稳态及分布有重要调控作用(Xue et al., 2018a). ...
Transgenerationally precipitated meiotic chromosome instability fuels rapid karyotypic evolution and phenotypic diversity in an artificially constructed allotetraploid wheat (AADD) 1 2018
... 多倍体在生物界广泛存在, 很多作物都是多倍体.刘宝研究组利用六倍体小麦基因组中2个基因组——乌拉尔图小麦(Triticum urartu, AA)和节节麦(Aegilops tauschii, DD)构建了人工异源多倍体(AADD), 并对其连续12代个体的核型和表型变异进行了检测, 揭示了异源多倍化催生了大量可遗传的核型异质性和表型多样性, 为“全基因组重复能加大生物的演化潜力”观点提供了鲜活的实验证据(Gou et al., 2018).辛明明研究组则对山羊草属(Aegilops)中一些二倍体, 以及小麦属(Triticum)中一些四倍体和六倍体胚乳中亲本特异性基因的表达模式进行了分析, 结果表明小麦多倍体化过程中的基因组印记(一种表观遗传现象, 导致基因根据其亲本来源出现差异表达)在杂交多倍化演化过程中是保守的(Yang et al., 2018d). ...
Gene refashioning through innovative shifting of reading frames in mosses 1 2018
... 早期陆生植物形态结构相对简单, 但环境适应能力强.特有基因对物种适应特殊环境发挥了重要作用.黄锦岭研究组对小立碗藓的hemerythrin基因进行了研究, 发现其起源于真菌向早期陆生植物的水平基因转移事件.该基因在小立碗藓中通过改变移码突变, 演化出1个双编码基因YAN/AltYAN, 该基因介导了小立碗藓的脱水耐受性(Guan et al., 2018).该研究对揭示陆生植物如何适应特殊环境提供了新观点及实验证据.基因组学研究为揭示一些物种特异现象提供了很好的切入点.例如, 卷柏(Selaginella tamariscina)是古老的复苏植物, 干旱时能存活数十年, 遇水而荣, 与已测序的其同属植物江南卷柏(S. moellendorffii) (干旱敏感型)截然不同.陈士林和宋经元研究组破译了卷柏基因组, 绘制了高杂合度卷柏的基因组图谱, 并与已发表的江南卷柏基因组进行了比较, 揭示了卷柏耐旱和复苏机制(Xu et al., 2018g).维管系统(木质部等)是植物在水生到陆生的进化过程中产生的.拟南芥和小立碗藓中特定的NAC转录因子调控木质部发育以及水分运输.景海春研究组与才宏伟研究组合作从甜高粱中鉴定到1个在驯化过程中受到选择、控制茎秆持汁性的Dry基因(编码一个NAC转录因子), 并发现其功能缺失是甜高粱茎秆富含汁液的重要原因(Zhang et al., 2018f).该研究不仅对促进作物茎秆持汁表型的遗传改良具重要作用, 而且为作物育种获得籽粒及秸秆“双丰收”提供了可能. ...
Arabidopsis blue light receptor Phototropin 1 undergoes blue light-induced activation in membrane microdomains 1 2018
... 光对植物不仅是一种能源, 也是一种重要的环境信号.植物通过向光素(phot)、隐花色素(cry)和光敏色素(phy)等光受体来感应光信号.林金星研究组研究了蓝光对细胞质膜上向光素phot1的定位变化和动态性质, 在单分子水平上揭示了蓝光信号传递的起始事件及调控机制(Xue et al., 2018b).刘宏涛研究组发现了隐花色素互作转录因子CIB1通过直接结合cry2和CO来介导cry2-CIB1-CO复合体的形成, 揭示了蓝光信号调控植物开花时间的新机制(Liu et al., 2018g).杨洪全研究组发现, G蛋白β亚基AGB1通过与HY5互作抑制其DNA结合能力, 从而抑制光信号; 而蓝光信号通过CRY1与AGB1互作促进AGB1与HY5解离来抑制AGB1的功能(Lian et al., 2018); 并发现CRY1与去磷酸化形式的BL信号关键转录因子BES1发生依赖于蓝光的直接互作, 揭示了植物能根据光信号和内在BL信号有无或强弱的动态变化来优化其光形态建成(Wang et al., 2018n).此外, 该研究组还发现光受体cry1、phyB竞争性与生长素信号通路抑制子Aux/IAA结合, 并磷酸化后者而负调控生长素信号转导, 进而精确调控植物光信号与生长素信号通路的平衡, 优化其生长(Xu et al., 2018c). ...
Identification of key amino acid residues determining product specificity of 2,3-oxidosqualene cyclase in oryza species 1 2018
... 尼克酰胺腺嘌呤二核苷酸(NAD)从尼克酸(NA) 合成.目前已发现多种NA修饰(如糖基化).王国栋研究组新发现一种NA修饰——甲酯化(MeNA).MeNA是一种NAD前体, 可在植物不同组织间进行更长距离运输.该研究提出了植物中NAD生物合成与NA衍生物之间关系的新见解(Wu et al., 2018b).萜类代谢物在植物生长发育等方面发挥重要作用.漆小泉研究组与相关单位合作发现一个新的、多产物OSC (2,3-氧化鲨烯环化酶) ——籼稻醇合酶(OsOS).该酶合成一种新的五环三萜籼稻醇(orysatinol)的主产物及12种不同的三萜类化合物(Xue et al., 2018c).王强研究组与相关单位合作发现玉米倍半萜合酶能直接催化(E, E)-法呢基二磷酸, 形成二羟基化的二醇产物(主要是eudesmane-2,11-diol) (Liang et al., 2018a).该研究揭示了TPS的新型催化功能及形成二羟基化倍半萜的催化机制. ...
A chromatin loop represses WUSCHEL expression in Arabidopsis 0 2018
The opium poppy genome and morphinan production 2 2018
... 二是植物系统进化研究取得了系列突破性成果.陈之端研究组重建了中国被子植物生命之树, 揭示了中国被子植物系统发育多样性形成的时空格局, 并明确了属、种水平分别应该重点保护的热点地区, 对我国保护区建设意义重大(Lu et al., 2018a).该成果入选2018年度中国生命科学十大进展.凌宏清、梁承志、王道文和张爱民研究组完成了乌拉尔图小麦材料G1812的基因组测序和精细组装, 绘制出小麦A基因组7条染色体的分子图谱(Ling et al., 2018).研究成果对进一步推动栽培小麦的遗传改良具有重要的理论意义和实用价值.叶凯研究组破译了罂粟基因组DNA密码, 揭示了其合成重要药用化合物的关键步骤, 这一发现为提高罂粟药用成分的产量及抗病性奠定了基础(Guo et al., 2018b).黎志康研究组与相关单位合作, 深入探讨了水稻起源、分类以及驯化规律, 揭示了亚洲栽培稻的起源和群体基因组变异结构, 剖析了水稻核心种质资源的基因组遗传多样性(Wang et al., 2018o), 为水稻规模化基因发掘和复杂性状分子改良奠定了基础. ...
Deficiency of a triterpene pathway results in humidity-sensitive genic male sterility in rice 1 2018
... 目前, 中国科学家利用种间、亚种间和品种间杂交已先后培育了以D型等为代表的60余种水稻细胞质雄性不育系.D1型是一种新型孢子体细胞质雄性不育类型, 无花粉型败育, 且为中国所特有.颜龙安和蔡耀辉研究组成功克隆了D1型水稻细胞质雄性不育基因并揭示了其败育机理(Xie et al., 2018).漆小泉研究组则发现了一种新型水稻雄性不育系, 并且阐释了OsOSC12/OsPTS1在水稻花粉包被形成过程中的关键作用, 对禾本科作物湿敏雄性不育材料的培育具有重要指导意义(Xue et al., 2018d). ...
Injury activates Ca( 2+)/calmodulin-dependent phosphorylation of JAV1-JAZ8-WRKY51 complex for jasmonate biosynthesis 1 2018
... 茉莉素(JA)是一类来源于环脂肪酸的植物激素, 可调节植物的防御反应和生长发育.谢道昕研究组发现健康植株中JJW (JAV1-JAZ8-WRKY51)复合体能够抑制JA合成基因的表达, 虫咬触发Ca2+/CAM依赖的JAV1磷酸化并促使JJW复合体解离而解除抑制, 进而促进JA合成并产生抗性(Yan et al., 2018a).此外, 该研究组还发现COI1可识别JA的动态变化.COI1先识别JA形成COI1-JA复合体, 之后结合JAZs形成三元复合体, 进而促进JA的信号转导(Yan et al., 2018b). ...
GRAIN SIZE AND NUMBER1 negatively regulates the OsMKKK10-OsMKK4-OsMPK6 cascade to coordinate the trade-off between grain number per panicle and grain size in rice 1 2018
... 丁勇研究组发现COMPASS-like复合体调控水稻抽穗期和花序结构, OsWDR5a和OsTrx1功能缺失可导致水稻花序二级枝梗减少及抽穗延迟(Jiang et al., 2018a).此外, 他们还揭示了水稻SIP1招募OsTrx1到Ehd1基因调控开花时间的分子机制(Jiang et al., 2018b).万建民和程治军研究组则解析了苹果酸抑制水稻穗顶部退化和OsALMT7转运苹果酸在水稻幼穗发育过程中起作用的分子机制, 为水稻高产品种的选育奠定了理论基础(Heng et al., 2018).张大兵研究组与相关单位合作, 发现LOFSEP亚族基因OsMADS1、OsMADS34和OsMADS5共同调控花分生组织, 并正向调控其它MADS-box花的同源基因来确定小穗器官的特征(Wu et al., 2018a).孙传清和朱作峰研究组对水稻驯化过程中二级枝梗数目转变的分子机制进行了研究, 发现栽培稻中FZP启动子的4 bp缺失影响了OsARFs的转录调控, 降低FZP的表达, 可提高水稻枝梗数和产量(Huang et al., 2018d).林鸿宣和单军祥研究组则证实GSN1作为负调控因子调控水稻穗型发育的OsMKKK10-OsMKK4-OsMPK6级联信号通路.GSN1-MAPK通过整合下游植物激素信号, 精准调控水稻穗粒数和籽粒大小的协同发育(Guo et al., 2018c).该研究为作物产量的遗传改良提供了新的分子模块与策略. ...
Genome-wide association studies of image traits reveal genetic architecture of drought resistance in rice 0 2018
Dynamic perception of jasmonates by the F-box protein COI1 1 2018
... 茉莉素(JA)是一类来源于环脂肪酸的植物激素, 可调节植物的防御反应和生长发育.谢道昕研究组发现健康植株中JJW (JAV1-JAZ8-WRKY51)复合体能够抑制JA合成基因的表达, 虫咬触发Ca2+/CAM依赖的JAV1磷酸化并促使JJW复合体解离而解除抑制, 进而促进JA合成并产生抗性(Yan et al., 2018a).此外, 该研究组还发现COI1可识别JA的动态变化.COI1先识别JA形成COI1-JA复合体, 之后结合JAZs形成三元复合体, 进而促进JA的信号转导(Yan et al., 2018b). ...
Plastid phylogenomics resolves infrafamilial relationships of the Styracaceae and sheds light on the backbone relationships of the Ericales 1 2018
A novel HD-ZIP IV/MIXTA complex promotes glandular trichome initiation and cuticle development in Artemisia annua 1 2018
... 茸毛作为植物体最外层结构, 具有很好的抗虫防病毒作用.叶志彪和杨长宪研究组分离了控制番茄腺体毛形成的关键基因H (Hair), 该基因编码C2H2锌指蛋白, 可能是茄科植物多细胞腺体毛形成的关键保守基因(Chang et al., 2018).唐克轩研究组发现R2R3 MYB第9亚家族的转录因子AaMIXTA1参与调控青蒿分泌型腺毛发育, 为研究其它植物多细胞腺毛奠定了基础(Shi et al., 2018c).此外, 他们还发现HD-ZIP IV转录因子AaHD8与AaMIXTA1相互作用形成复合体, 协同调控AaHD1和多个蜡质角质合成酶基因的表达, 从而促进蜡质合成和腺毛的起始发育(Yan et al., 2018d).陈晓亚研究组与相关单位合作对可纺纤维形成的分子机制进行研究, 发现编码bHLH转录因子的棉花PRE1是可纺纤维进化的目标基因, 其正调控纤维伸长.PRE1基因启动子的变异可促进棉花可纺纤维的形成(Zhao et al., 2018a).李学宝研究组鉴定了富含半胱氨酸的GhXLIM6蛋白, 其可与GhKNL1启动子的PAL-box结合而抑制该基因表达, 促进纤维素合成, 进而调控棉纤维伸长(Li et al., 2018i).俞嘉宁研究组则揭示了棉花线粒体Ghatp1基因C1292和C1415位点发生RNA编辑可影响ATPase产生ATP, 从而促进表皮毛及纤维伸长(He et al., 2018c).根毛是一种顶端生长的植物细胞, 微丝骨架对根毛的生长发育有极其重要的调控作用.贾红磊研究组揭示了H2S通过硫巯基化修饰扰乱正常的微丝骨架聚合及生长模式, 从而影响植物根毛的生长, 为进一步探索H2S调控植物生长发育的作用机制提供了理论依据(Li et al., 2018d). ...
RSM1, an Arabidopsis MYB protein, interacts with HY5/HYH to modulate seed germination and seedling development in response to abscisic acid and salinity 1 2018
... 土壤盐碱化极大地限制了粮食作物的稳产增收.如何将盐碱地变为高产粮田, 成为当下农业科学家研究的热点方向, 而解析植物应答盐碱胁迫的分子机制是提高植物抗盐碱的有效手段.朱健康研究组发现, 细胞壁LRX (Leucine-rich repeat extensins)蛋白家族多突变体植株表现出对盐胁迫非常敏感的表型.进一步研究发现, LRX蛋白与RALF多肽以及细胞膜受体类激酶FERONIA形成一个元件调控植物生长和耐盐性(Zhao et al., 2018b).该研究为培育抗逆高产作物指明了方向.此外, 朱健康和王镇研究组还探明了ABA对植物抗旱性的影响, 揭示了开花抑制子SVP (Short vegetative phase)通过调控ABA代谢提高拟南芥的抗旱性机制(Wang et al., 2018r).植物类受体激酶能感受外界环境胁迫信号, 并通过胞内激酶域激活下游效应子活性和调控细胞相关生理生化过程, 从而降低或消除逆境胁迫的影响.刘选明和林建中研究组破译了1个能降低土地盐碱化对水稻产量影响的类受体胞质激酶1 (salt tolerance receptor-like cytoplasmic kinase 1, STRK1), 其可通过磷酸化和激活CatC调节H2O2体内平衡来改善盐以及氧化胁迫的耐受性(Zhou et al., 2018e).该研究为进一步解析植物耐盐的分子机制奠定了基础.范六民研究组报道了MYB亚家族转录因子RSM1 (Radialis-like sant/myb 1)蛋白与HY5/HYH蛋白互作, 并作用于ABI5启动子区, 调控ABA和非生物逆境(盐)胁迫相关基因的应答(Yang et al., 2018a).黄俊丽研究组则对水稻MADS-box家族转录因子OsMADS25蛋白进行了研究, 发现其可与谷胱甘肽S-转移酶基因OsGST4的启动子直接结合, 通过ABA介导的调控通路和ROS清除共同调控水稻的根系生长和耐盐性(Xu et al., 2018d).蒋才富研究组利用QTL克隆了调控叶片Na+含量的主效基因ZmNC1 (Na+ content 1).该基因编码1个Na+选择性离子转运蛋白ZmHKT1, 其可通过抑制Na+由根向地上部运输提高玉米的耐盐能力(Zhang et al., 2018g).郝玉金研究组则发现苹果葡萄糖感知因子己糖激酶MdHXK1可磷酸化Na+/H+交换蛋白MdNHX1, 以提高其蛋白稳定性, 进而增强苹果的耐盐性(Sun et al., 2018b). ...
OsOFP19 modulates plant architecture by integrating the cell division pattern and brassinosteroid signaling 1 2018
... 水稻株型与产量密切相关.叶片作为水稻理想株型的组成要素之一, 其倾角对产量影响较大.薛红卫研究组新克隆了叶倾角调控基因LC3, 该基因编码1个含有SPOC结构域的转录抑制子, 其可与HIT锌指结构域蛋白LIP1互作, 通过调控生长素平衡和信号转导来调控植株叶倾角(Chen et al., 2018h).李建雄研究组与田志宏研究组合作揭示了OsOFP19、OSH1与DLT蛋白可形成复合体, 调控BL信号转导及影响叶倾角发育(Yang et al., 2018b).易可可研究组则解析了磷素有效性调控水稻叶倾角的分子机制.他们发现磷饥饿诱导的SPX1 (Syg1/Pho81/XPR1)与SPX2蛋白负调控叶倾角.SPX1的互作蛋白RLI1 (Regulator of leaf inclination 1)正调控叶倾角.RLI1可直接激活下游BU1 (Brassinosteroid upregulated 1)和BC1 (BU1-like1 complex 1)基因的表达以控制叶枕细胞的伸长, 从而使叶倾角增大(Ruan et al., 2018).种康和徐云远研究组证实miR396d通过分别抑制其靶基因OsGRF4和OsGRF6的转录, 促进BL及抑制GA信号途径, 进而调控水稻叶夹角和株高(Tang et al., 2018).邢永忠研究组则阐释了来自世界各地的水稻核心种质剑叶夹角的遗传基础, 发现bHLH转录因子家族中OsbHLH153和OsbHLH17能快速响应BL和IAA信号转导, 从而影响植株形态(Dong et al., 2018c).王永红研究组揭示了水稻分蘖角度调控的分子机制.他们通过研究水稻茎重力反应过程中的动态转录组变化, 探索了一条水稻分蘖角度调控的核心途径(Zhang et al., 2018h).该研究为解析水稻分蘖角度的调控网络进而挖掘有利用价值的基因提供了重要信息.谭禄宾研究组发现亚洲栽培稻PROG1基因相邻位置约有110 kb的染色体缺失.该缺失片段包含7个与PROG1基因相似的锌指基因, 其缺失推动了水稻由匍匐向直立转变, 最终实现产量大幅提高(Wu et al., 2018c). ...
Phytochrome A negatively regulates the shade avoidance response by increasing Auxin/Indole acidic acid protein stability 1 2018
... 李继刚和康定明研究组则筛选获得了远红光信号转导新组分TZP.TZP可与phyA和FHY1互作, 且在远红光下调控phyA、FHY1和HY5的蛋白水平, 并参与调控phyA的蛋白磷酸化(Zhang et al., 2018k).李继刚研究组还与邓兴旺研究组合作证明了拟南芥phyA铰链区的3个位点(S590、T593和S602)对phyA行使功能非常重要(Zhou et al., 2018c).吴素幸研究组发现, phyA和cry感知远红光和蓝光信号, 使负调控因子COP1失活, 从而激活生长素途径在TOR依赖的RPS6磷酸化中的作用(Chen et al., 2018a).此外, 李琳研究组发现在遮阴, 尤其是严重遮阴条件下, phyA蛋白量增加, phyA与Aux/IAA互作影响了生长素受体TIR与Aux/IAA的互作, 从而阻止TIR介导的Aux/IAA降解, 导致生长素信号减弱, 通过调控光信号与生长素信号的平衡, 进而调节植物的生长发育(Yang et al., 2018c).周艳虹研究组鉴定了番茄在光抑制和光保护过程中的光信号依赖性调控通路, 分析了phyA依赖的HY5-ABI5-RBOH1信号通路在缓解冷诱导的光抑制以及光保护中的作用(Wang et al., 2018a). ...
Chromatin remodeling factor CHR18 interacts with replication protein RPA1A to regulate the DNA replication stress response in Arabidopsis 1 2018
... 染色质重塑参与调控DNA复制、转录以及修复等多个生物学过程.染色质重塑因子可导致核小体位置和结构的改变, 引起染色质空间变化以及相关组蛋白修饰和DNA甲基化修饰变化, 但其是否参与植物DNA复制胁迫尚不清楚.刘建祥研究组针对该问题对拟南芥染色质重塑因子CHR18 (chromatin remodeling factor 18)进行了研究, 发现其功能缺失可导致拟南芥对DNA复制胁迫高度敏感.进一步分析发现, CHR18与DNA复制蛋白RPA1A在细胞核存在互作, 说明DNA复制相关蛋白RPA1A可招募染色质重塑因子CHR18到复制叉, 参与减轻DNA复制过程中遇到的逆境胁迫(Han et al., 2018).此外, 周道绣研究组对水稻染色质重塑因子DDM1 (DECREASE IN DNA METHYLATION 1)进行了研究, 揭示了OsDDM1在抑制非编码RNA (non-coding RNA)表达以及在RNA介导的DNA甲基化(RdDM)途径中的作用机制(Tan et al., 2018a). ...
CELLULASE6 and MANNANASE7 affect cell differentiation and silique dehiscence 1 2018
... 植物分枝是影响植物株型的关键因素.秦根基研究组与相关单位合作对植物分枝形成的分子调控机制进行研究, 发现了1个分枝正调控因子TIE1, 它与分枝重要调控因子BRC1 (BRANCHED1)发生互作, 抑制其转录活性.通过调节HB21、HB40和HB53等BRC1下游基因的转录, 发挥促进腋芽分枝的作用(Yang et al., 2018h).开花是植物由营养生长转向生殖生长的标志.王前和张小兰研究组对热诱导开花的分子机理进行探索, 证明LsSOC1是热诱导花芽形成的激活子, 热激转录因子HsfA1e和HsfA4c通过结合在LsSOC1基因的启动子上, 影响其表达, 为开花相关机制研究奠定了理论基础(Chen et al., 2018k).果实成熟过程对于果实品质形成和种子传播非常关键.陈建业和邝健飞研究组发现转录因子MaMYB3通过抑制淀粉降解相关基因以及MabHLH6转录因子的转录水平来调控淀粉降解, 丰富了香蕉果实成熟的转录调控网络(Fan et al., 2018).朱本忠和罗云波研究组则发现, 在番茄rin突变体中, 基因组DNA片段部分缺失导致形成RIN-MC片段的嵌合, 该嵌合能够调控许多成熟相关基因的表达, 从而调节果实成熟(Li et al., 2018h).此外, 吴鸿研究组与相关单位合作阐明了拟南芥角果开裂的调控机制.他们通过对纤维素酶CEL6和半纤维素酶MAN7以及果胶酶ADPG2基因的一系列突变体果荚离层进行细胞超微结构观察, 并结合基因表达的检测结果, 证明改变这些细胞壁酶的表达量和酶活性可以调控果荚开裂区细胞分化和离层区细胞降解(He et al., 2018a).该研究为后期培育不同程度的果实抗开裂油料作物提供了技术支持. ...
A naturally occurring epiallele associates with leaf senescence and local climate adaptation in Arabidopsis accessions 1 2018
Two pivotal RNA editing sites in the mitochondrial atp1mRNA are required for ATP synthase to produce sufficient ATP for cotton fiber cell elongation 1 2018
... 茸毛作为植物体最外层结构, 具有很好的抗虫防病毒作用.叶志彪和杨长宪研究组分离了控制番茄腺体毛形成的关键基因H (Hair), 该基因编码C2H2锌指蛋白, 可能是茄科植物多细胞腺体毛形成的关键保守基因(Chang et al., 2018).唐克轩研究组发现R2R3 MYB第9亚家族的转录因子AaMIXTA1参与调控青蒿分泌型腺毛发育, 为研究其它植物多细胞腺毛奠定了基础(Shi et al., 2018c).此外, 他们还发现HD-ZIP IV转录因子AaHD8与AaMIXTA1相互作用形成复合体, 协同调控AaHD1和多个蜡质角质合成酶基因的表达, 从而促进蜡质合成和腺毛的起始发育(Yan et al., 2018d).陈晓亚研究组与相关单位合作对可纺纤维形成的分子机制进行研究, 发现编码bHLH转录因子的棉花PRE1是可纺纤维进化的目标基因, 其正调控纤维伸长.PRE1基因启动子的变异可促进棉花可纺纤维的形成(Zhao et al., 2018a).李学宝研究组鉴定了富含半胱氨酸的GhXLIM6蛋白, 其可与GhKNL1启动子的PAL-box结合而抑制该基因表达, 促进纤维素合成, 进而调控棉纤维伸长(Li et al., 2018i).俞嘉宁研究组则揭示了棉花线粒体Ghatp1基因C1292和C1415位点发生RNA编辑可影响ATPase产生ATP, 从而促进表皮毛及纤维伸长(He et al., 2018c).根毛是一种顶端生长的植物细胞, 微丝骨架对根毛的生长发育有极其重要的调控作用.贾红磊研究组揭示了H2S通过硫巯基化修饰扰乱正常的微丝骨架聚合及生长模式, 从而影响植物根毛的生长, 为进一步探索H2S调控植物生长发育的作用机制提供了理论依据(Li et al., 2018d). ...
Programmed self-elimination of the CRISPR/ Cas9 construct greatly accelerates the isolation of edited and transgene-free rice plants 1 2018
... 随着科学技术的发展, 基因组编辑技术开始应用于作物精准育种.然而由于缺乏高效去除含有Cas9表达盒及扩展性强、编辑效率高的单碱基编辑技术, 限制了其在分子育种中的应用.谢卡斌研究组利用生物体内的内含子剪切系统和tRNA加工系统开发出水稻中高效、多位点编辑且适用于CRISPR/Cpf1系统的新方法, 有望应用于动植物中(Ding et al., 2018a).林宏辉和周焕斌研究组及朱健康研究组分别基于APOBEC1 酶的碱基编辑器及腺嘌呤的碱基编辑器(ABE7-10), 实现了水稻基因组4种不同碱基(A-G、T-C、C-T和G-A)的高效替换与水稻基因组特定位点的A·T碱基对高效转化为G·C碱基对, 不仅扩展了单碱基编辑技术在植物中的应用而且丰富了可用的单碱基编辑工具(Ren et al., 2018a; Hua et al., 2018).赵云德研究组则利用自杀基因与CRISPR载体融合, 开发出高效的转基因自清除基因编辑系统(He et al., 2018d).这些成果对深入研究水稻功能基因组具有重要推动作用. ...
OsALMT7 maintains panicle size and grain yield in rice by mediating malate transport 1 2018
... 丁勇研究组发现COMPASS-like复合体调控水稻抽穗期和花序结构, OsWDR5a和OsTrx1功能缺失可导致水稻花序二级枝梗减少及抽穗延迟(Jiang et al., 2018a).此外, 他们还揭示了水稻SIP1招募OsTrx1到Ehd1基因调控开花时间的分子机制(Jiang et al., 2018b).万建民和程治军研究组则解析了苹果酸抑制水稻穗顶部退化和OsALMT7转运苹果酸在水稻幼穗发育过程中起作用的分子机制, 为水稻高产品种的选育奠定了理论基础(Heng et al., 2018).张大兵研究组与相关单位合作, 发现LOFSEP亚族基因OsMADS1、OsMADS34和OsMADS5共同调控花分生组织, 并正向调控其它MADS-box花的同源基因来确定小穗器官的特征(Wu et al., 2018a).孙传清和朱作峰研究组对水稻驯化过程中二级枝梗数目转变的分子机制进行了研究, 发现栽培稻中FZP启动子的4 bp缺失影响了OsARFs的转录调控, 降低FZP的表达, 可提高水稻枝梗数和产量(Huang et al., 2018d).林鸿宣和单军祥研究组则证实GSN1作为负调控因子调控水稻穗型发育的OsMKKK10-OsMKK4-OsMPK6级联信号通路.GSN1-MAPK通过整合下游植物激素信号, 精准调控水稻穗粒数和籽粒大小的协同发育(Guo et al., 2018c).该研究为作物产量的遗传改良提供了新的分子模块与策略. ...
Premature leaf senescence 3, encoding a methyltransferase, is required for melatonin biosynthesis in rice 1 2018
... 钱前研究组则报道了一种新的种质资源小薇.它具有生长周期短和株型矮小等表型, 可以像拟南芥一样在实验室内大规模种植.小薇的超矮秆表型是由于GA生物合成基因D18突变引起(Hu et al., 2018a).该研究创制了“小薇日本晴”和“小薇93”, 为进行水稻功能基因组相关研究提供了重要资源.李来庚研究组与杨远柱研究组合作发现了一个新的特异调控水稻茎秆基部节长度的基因SBI, 该基因编码GA2氧化酶, 在培育水稻半矮秆性状, 提高抗倒伏能力, 增加水稻产量方面具重要的应用价值(Liu et al., 2018b).周奕华研究组揭示了IIP4蛋白通过与NAC29/NAC31互作, 负向调控次生壁合成, 进而影响茎秆强度(Zhang et al., 2018b).曹立勇和程式华研究组阐明了水稻叶片衰老的新机制.他们合作克隆了1个引起水稻叶片早衰的基因OsMTS1, 该基因编码褪黑素生物合成途径中的甲基转移酶, OsMTS1突变后可导致水稻叶片中褪黑素含量降低, 从而引起叶片早衰(Hong et al., 2018). ...
Xiaowei, a new rice germplasm for large-scale indoor research 1 2018
... 钱前研究组则报道了一种新的种质资源小薇.它具有生长周期短和株型矮小等表型, 可以像拟南芥一样在实验室内大规模种植.小薇的超矮秆表型是由于GA生物合成基因D18突变引起(Hu et al., 2018a).该研究创制了“小薇日本晴”和“小薇93”, 为进行水稻功能基因组相关研究提供了重要资源.李来庚研究组与杨远柱研究组合作发现了一个新的特异调控水稻茎秆基部节长度的基因SBI, 该基因编码GA2氧化酶, 在培育水稻半矮秆性状, 提高抗倒伏能力, 增加水稻产量方面具重要的应用价值(Liu et al., 2018b).周奕华研究组揭示了IIP4蛋白通过与NAC29/NAC31互作, 负向调控次生壁合成, 进而影响茎秆强度(Zhang et al., 2018b).曹立勇和程式华研究组阐明了水稻叶片衰老的新机制.他们合作克隆了1个引起水稻叶片早衰的基因OsMTS1, 该基因编码褪黑素生物合成途径中的甲基转移酶, OsMTS1突变后可导致水稻叶片中褪黑素含量降低, 从而引起叶片早衰(Hong et al., 2018). ...
Gibberellins play an essential role in late embryogenesis of Arabidopsis 1 2018
A novel QTL qTGW3 encodes the GSK3/SHAGGY-like kinase OsGSK5/OsSK41 that interacts with OsARF4 to negatively regulate grain size and weight in rice 1 2018
... 刘建祥和罗小金研究组、宋献军研究组、何予卿研究组同时克隆到与粒型和粒重相关的同一主效负调控QTL/基因qTGW3/TGW3/GL3.3, 并解析了相关分子机制(Hu et al., 2018c; Xia et al., 2018; Ying et al., 2018).李云海研究组则与相关单位合作揭示了OsMKKK10-OsMKK4-OsMAPK6信号途径正向调控水稻粒型和粒重(Xu et al., 2018e); 并发现大粒基因LARGE8可与OsMAPK6直接互作使其失活, 进而抑制颖壳细胞增殖, 负向调控水稻粒型(Xu et al., 2018f).余四斌研究组揭示了OsGBPs作为转录因子能够识别GAGA基序, 进而调控水稻粒长和植株高度(Gong et al., 2018).梁国华研究组则证明OsSPMS1基因负向调控水稻种子萌发、粒型和单株产量, 是水稻增产育种的重要靶标基因(Tao et al., 2018).刘巧泉研究组发现了水稻粒型与外观品质调控基因GS9, 其编码蛋白可与OsOFP14和OsOFP8蛋白互作, 通过改变细胞分裂调控水稻粒型(Zhao et al., 2018c).傅向东研究组克隆了1个控制水稻产量和提升稻米品质的重要基因LGY3, 该基因编码转录因子OsMADS1.可变剪接蛋白OsMADS1lgy3可提高稻米粒长, 降低垩白率, 进而影响稻米产量和外观品质(Liu et al., 2018e). ...
TaWRKY51 promotes lateral root formation through negative regulation of ethylene biosynthesis in wheat ( Triticum aestivum L.) 1 2018
... 乙烯(ETH)是重要的植物激素, 其对植物生长发育及果实成熟具重要调控作用.陈受宜和张劲松研究组对水稻乙烯不敏感系列突变体mhz2和mhz3进行了深入分析, 发现MHZ3可通过与乙烯信号通路关键调控因子OsEIN2的Nramp-like结构域结合稳定OsEIN2, 抑制其泛素化, 从而正调控乙烯信号转导(Ma et al., 2018a).而MHZ2/SOR1编码一个E3泛素连接酶, 该酶可直接调控生长素信号通路中的转录抑制子OsIAA26降解, 介导乙烯对根生长的抑制效应(Chen et al., 2018b).孙其信研究组则发现小麦TaWRKY51能结合到ETH合成基因TaACS的启动子区并抑制其表达, 进而促进侧根形成(Hu et al., 2018d). ...
Precise A .T to G .C base editing in the rice genome 1 2018
... 随着科学技术的发展, 基因组编辑技术开始应用于作物精准育种.然而由于缺乏高效去除含有Cas9表达盒及扩展性强、编辑效率高的单碱基编辑技术, 限制了其在分子育种中的应用.谢卡斌研究组利用生物体内的内含子剪切系统和tRNA加工系统开发出水稻中高效、多位点编辑且适用于CRISPR/Cpf1系统的新方法, 有望应用于动植物中(Ding et al., 2018a).林宏辉和周焕斌研究组及朱健康研究组分别基于APOBEC1 酶的碱基编辑器及腺嘌呤的碱基编辑器(ABE7-10), 实现了水稻基因组4种不同碱基(A-G、T-C、C-T和G-A)的高效替换与水稻基因组特定位点的A·T碱基对高效转化为G·C碱基对, 不仅扩展了单碱基编辑技术在植物中的应用而且丰富了可用的单碱基编辑工具(Ren et al., 2018a; Hua et al., 2018).赵云德研究组则利用自杀基因与CRISPR载体融合, 开发出高效的转基因自清除基因编辑系统(He et al., 2018d).这些成果对深入研究水稻功能基因组具有重要推动作用. ...
SEUSS and PIF4 coordinately regulate light and temperature signaling pathways to control plant growth 1 2018
... 光照和温度是影响植物生长发育非常重要的两个环境因素.林荣呈研究组发现了光信号与温度信号整合的新因子SEU, 揭示了SEU通过与PIF4互作形成转录调控复合物, 对生长素合成及与信号相关的靶基因进行调节, 从而实现对生长发育的调控(Huai et al., 2018).方玉达研究组发现, 拟南芥miRNA加工复合体成员DCL1/HYL1与PIF4存在直接互作.PIF4在黑暗/红光转换过程中调控DCL1的稳定性及部分miRNA基因的转录, 进而调控光形态建成(Sun et al., 2018e).黎家研究组则发现TCP17蛋白通过调控PIF4/PIF5依赖以及非依赖的生长素合成, 从而调控遮阴诱导下植物下胚轴的伸长生长(Zhou et al., 2018d). ...
Rice actin binding protein RMD controls crown root angle in response to external phosphate 1 2018
Natural allelic variations provide insights into host adaptation of Phytophthora avirulence effector PsAvr3c 1 2019
... 作物疫病是农业生产上的毁灭性病害.董莎萌研究组对大豆疫霉菌逃避寄主抗病基因识别的分子机制进行了探索, 发现疫霉中保守的效应子PsAvr3c中第174位丝氨酸突变为甘氨酸是疫霉菌逃避抗病基因Rps3c识别的重要遗传位点.进一步研究发现, 该位点突变导致PsAvr3c与GmSKRPs的互作亲和度显著降低, 进而逃避了寄主抗病基因Rps3c的识别(Huang et al., 2019).该研究揭示了一种病菌通过遗传变异逃避植物免疫系统的机制, 为植物抗病育种提供了重要的新分子靶标. ...
The pentatricopeptide repeat protein SOT5/ EMB2279 is required for plastid rpl2 and trnK intron splicing 1 2018
... 叶绿体是植物进行光合作用的细胞器, 其分裂由众多因子形成的复合物协助完成.张敏研究组发现MCD1在此过程中起重要作用.MCD1通过与ARC6作用, 识别FtsZ环, 并引导Min复合物整合到分裂环上, 从而调控叶绿体分裂(Chen et al., 2018e).欧阳敏研究组则发现ECD1蛋白通过参与叶绿体基因rps14的RNA编辑调控叶绿体早期发育(Jiang et al., 2018d).此外, 郁飞研究组和黄继荣研究组分别发现细胞质和叶绿体的翻译过程协同调控叶绿体发育, 及thf1突变体斑叶形成与质体翻译功能高度相关(Wang et al., 2018l; Huang et al., 2018b).类囊体是叶绿体基质中由单位膜围成的扁平囊, 是光合作用光反应的场所.Kang Byung-Ho研究组揭示了类囊体蛋白复合体的组装以及CURT1和FZL蛋白在类囊体生物发生过程中的关键作用(Liang et al., 2018d).隋森芳研究组与相关单位合作解析了红藻光系统I核心与捕光天线复合物(PSI-LHCR)的3.63?分辨率的三维结构, 发现红藻PSI核心既有蓝藻又有高等植物的特征(Pi et al., 2018).该研究对阐明PSI的进化和功能具有重要意义. ...
The antagonistic action of abscisic acid and cytokinin signaling mediates drought stress response in Arabidopsis 1 2018
... 植物激素之间存在复杂的调控网络.杨淑华研究组发现ABA信号关键蛋白激酶SnRK2s通过磷酸化细胞分裂素负调节子A型ARR5正调控其蛋白稳定, 揭示了植物激素信号调控网络在植物逆境响应胁迫中的精密调控机制(Huang et al., 2018c).刘西岗研究组发现细胞分裂素的内稳态对于花分生组织(floral meristem, FM)的维持及分化非常重要.生长素可以通过促进ARF3 (Auxin Response Factor 3)的表达抑制细胞分裂素的活性进而促进FM的分化(Zhang et al., 2018e).该研究揭示了转录因子AG通过植物激素信号途径协同调控花分生组织干细胞稳态的机理.王小菁研究组发现非洲菊锌指蛋白GhWIP2通过抑制GA和IAA信号及激活ABA信号, 进而抑制细胞生长并最终抑制花瓣伸展(Ren et al., 2018b).该研究揭示了GA、ABA和生长素3种激素交叉互作网络共同调控非洲菊细胞伸展的作用机理.向成斌研究组发现乙烯响应基因HB52是乙烯和IAA信号互作的重要节点, 通过上调PIN2、WAG1和WAG2的表达抑制主根伸长(Miao et al., 2018c).该研究揭示了乙烯与IAA交互作用的新机制.丁兆军研究组研究表明, BL合成途径突变体det2-9中乙烯合成大幅增加并积累了大量的超氧阴离子.该研究揭示了BL、乙烯与活性氧协同调控根生长的新机制(Lv et al., 2018).ABA和BL信号通常相互拮抗.王学路研究组发现ABI1和ABI2通过与BIN2互作使其去磷酸化, 进而调控其对BES1的磷酸化(Wang et al., 2018c).同时, 该研究组还发现BL信号途径中的GSK3为控制水稻中胚轴伸长的一个关键因子, 其通过磷酸化调控一类细胞周期蛋白CYC U2的稳定性促进细胞分裂.BL以及独脚金内酯(strigolactone)信号通路协同调控GSK3介导的细胞周期进程, 进而促进中胚轴伸长(Sun et al., 2018c). ...
Variation in the regulatory region of FZP causes increases in secondary inflorescence branching and grain yield in rice domestication 1 2018
... 丁勇研究组发现COMPASS-like复合体调控水稻抽穗期和花序结构, OsWDR5a和OsTrx1功能缺失可导致水稻花序二级枝梗减少及抽穗延迟(Jiang et al., 2018a).此外, 他们还揭示了水稻SIP1招募OsTrx1到Ehd1基因调控开花时间的分子机制(Jiang et al., 2018b).万建民和程治军研究组则解析了苹果酸抑制水稻穗顶部退化和OsALMT7转运苹果酸在水稻幼穗发育过程中起作用的分子机制, 为水稻高产品种的选育奠定了理论基础(Heng et al., 2018).张大兵研究组与相关单位合作, 发现LOFSEP亚族基因OsMADS1、OsMADS34和OsMADS5共同调控花分生组织, 并正向调控其它MADS-box花的同源基因来确定小穗器官的特征(Wu et al., 2018a).孙传清和朱作峰研究组对水稻驯化过程中二级枝梗数目转变的分子机制进行了研究, 发现栽培稻中FZP启动子的4 bp缺失影响了OsARFs的转录调控, 降低FZP的表达, 可提高水稻枝梗数和产量(Huang et al., 2018d).林鸿宣和单军祥研究组则证实GSN1作为负调控因子调控水稻穗型发育的OsMKKK10-OsMKK4-OsMPK6级联信号通路.GSN1-MAPK通过整合下游植物激素信号, 精准调控水稻穗粒数和籽粒大小的协同发育(Guo et al., 2018c).该研究为作物产量的遗传改良提供了新的分子模块与策略. ...
Apple fruit acidity is genetically diversified by natural variations in three hierarchical epistatic genes: MdSAUR37, MdPP2CH and MdALMTII 1 2018
... 4-羟基-2,5-二甲基-3(2H)-呋喃酮(HDMF)是草莓果实的特征芳香组分, 其催化合成的关键酶是草莓醌氧化还原酶(FaQR).陈昆松研究组发现乙烯应答因子(FaERF9)为FaQR启动子的正调节因子, 揭示了ERF-MYB复合物通过激活FaQR表达调节草莓中HDMF生物合成的分子机制(Zhang et al., 2018m).苹果酸是苹果果实中的主要有机酸.韩振海和张新忠研究组获得了4个与苹果酸相关的主效QTLs; 并证明候选基因MdSAUR37等存在上位效应, 共同调控苹果酸的积累(Jia et al., 2018a).草酸是最简单的二元酸, 在调控金属胁迫等方面起积极作用.巫永睿研究组克隆了玉米草酸降解途径中的草酰辅酶A基因, 阐明了玉米草酸代谢的前两步反应, 揭示了草酸代谢参与籽粒储藏物质积累和营养品质形成的分子机理, 为遗传改良草酸含量较高的蔬菜提供了候选基因(Yang et al., 2018e). ...
Golgi-localized LOT regulates trans-golgi network biogenesis and pollen tube growth 1 2018
... 花粉管生长是典型的细胞极性延伸现象.关跃峰研究组近期发现1个胞质定位的糖酵解关键酶——磷酸甘油酸激酶(PGKc).其突变体的花粉萌发速度大幅提升, 但花粉管极性形态异常.进一步研究发现, PGKc对花粉管极性的调控并非通过对细胞总体能量代谢的影响, 而是通过糖酵解途径(Chen et al., 2018i).杨贞标研究组则以拟南芥花粉管为模式系统, 阐明了植物小G蛋白ROP1、细胞骨架与胞吞作用共同形成调节花粉管极性生长的细胞与分子反馈调控网络, 揭示了网格蛋白介导的内吞作用调控花粉管顶端膜与侧面膜分子的极性分布及细胞顶端极性生长的分子机理(Li et al., 2018b).杨维才和李红菊研究组鉴定了1个影响高尔基体反面网络结构(trans-Golgi network, TGN)形成的蛋白LOT (Loss of TGN).在lot突变体中, 花粉管在柱头上的生长被抑制, 植物表现出严重的不育表型.进一步研究发现, 突变体中高尔基体膜囊过度堆积, 且源于高尔基体的分泌小泡以及TGN的数目显著减少.此外, 突变体花粉管中细胞壁组分——果胶质(pectin)等的形成均受到抑制, 细胞内吞作用也被显著阻碍, 从而影响花粉管的生长(Jia et al., 2018b). ...
The COMPASS-like complex promotes flowering and panicle branching in rice 1 2018
... 丁勇研究组发现COMPASS-like复合体调控水稻抽穗期和花序结构, OsWDR5a和OsTrx1功能缺失可导致水稻花序二级枝梗减少及抽穗延迟(Jiang et al., 2018a).此外, 他们还揭示了水稻SIP1招募OsTrx1到Ehd1基因调控开花时间的分子机制(Jiang et al., 2018b).万建民和程治军研究组则解析了苹果酸抑制水稻穗顶部退化和OsALMT7转运苹果酸在水稻幼穗发育过程中起作用的分子机制, 为水稻高产品种的选育奠定了理论基础(Heng et al., 2018).张大兵研究组与相关单位合作, 发现LOFSEP亚族基因OsMADS1、OsMADS34和OsMADS5共同调控花分生组织, 并正向调控其它MADS-box花的同源基因来确定小穗器官的特征(Wu et al., 2018a).孙传清和朱作峰研究组对水稻驯化过程中二级枝梗数目转变的分子机制进行了研究, 发现栽培稻中FZP启动子的4 bp缺失影响了OsARFs的转录调控, 降低FZP的表达, 可提高水稻枝梗数和产量(Huang et al., 2018d).林鸿宣和单军祥研究组则证实GSN1作为负调控因子调控水稻穗型发育的OsMKKK10-OsMKK4-OsMPK6级联信号通路.GSN1-MAPK通过整合下游植物激素信号, 精准调控水稻穗粒数和籽粒大小的协同发育(Guo et al., 2018c).该研究为作物产量的遗传改良提供了新的分子模块与策略. ...
SIP1 participates in regulation of flowering time in rice by recruiting OsTrx1 to Ehd1 1 2018
... 丁勇研究组发现COMPASS-like复合体调控水稻抽穗期和花序结构, OsWDR5a和OsTrx1功能缺失可导致水稻花序二级枝梗减少及抽穗延迟(Jiang et al., 2018a).此外, 他们还揭示了水稻SIP1招募OsTrx1到Ehd1基因调控开花时间的分子机制(Jiang et al., 2018b).万建民和程治军研究组则解析了苹果酸抑制水稻穗顶部退化和OsALMT7转运苹果酸在水稻幼穗发育过程中起作用的分子机制, 为水稻高产品种的选育奠定了理论基础(Heng et al., 2018).张大兵研究组与相关单位合作, 发现LOFSEP亚族基因OsMADS1、OsMADS34和OsMADS5共同调控花分生组织, 并正向调控其它MADS-box花的同源基因来确定小穗器官的特征(Wu et al., 2018a).孙传清和朱作峰研究组对水稻驯化过程中二级枝梗数目转变的分子机制进行了研究, 发现栽培稻中FZP启动子的4 bp缺失影响了OsARFs的转录调控, 降低FZP的表达, 可提高水稻枝梗数和产量(Huang et al., 2018d).林鸿宣和单军祥研究组则证实GSN1作为负调控因子调控水稻穗型发育的OsMKKK10-OsMKK4-OsMPK6级联信号通路.GSN1-MAPK通过整合下游植物激素信号, 精准调控水稻穗粒数和籽粒大小的协同发育(Guo et al., 2018c).该研究为作物产量的遗传改良提供了新的分子模块与策略. ...
Dynamics of arbuscular mycorrhizal fungal community structure and functioning along a nitrogen enrichment gradient in an alpine meadow ecosystem 1 2018
... 在多种生态系统中, 氮素有效性正在急剧增加, 然而氮素水平的提高对自然生态系统中丛枝菌根(AM)真菌功能的影响目前仍不清楚.冯虎元和刘永俊研究组发现, N诱导的AM真菌群落组成变化与土壤氮素有效性和/或植物物种丰富度密切相关, 而菌根功能的变化与特定AM真菌谱系的群落相关, 非磷限制的氮富集生态系统可诱导菌根的寄生功能, 从而影响植物群落结构与生态系统的可持续性(Jiang et al., 2018c).周华坤和姚步青研究组则对植物群落特征影响生态系统N存留的方式进行了探讨, 发现植物群落特征对生态系统N存留的控制受到养分输入的作用, 并由群落水平功能性状介导(Wang et al., 2018b).该研究揭示了青藏高原高寒草甸生态系统氮素存留及其植被变化的机制, 对深刻理解高寒草地结构和功能稳定性调控及高寒草地退化演替机制具有重要意义. ...
ECD1 functions as an RNA-editing transfactor of rps14-149 in plastids and is required for early chloroplast development in seedlings 1 2018
... 叶绿体是植物进行光合作用的细胞器, 其分裂由众多因子形成的复合物协助完成.张敏研究组发现MCD1在此过程中起重要作用.MCD1通过与ARC6作用, 识别FtsZ环, 并引导Min复合物整合到分裂环上, 从而调控叶绿体分裂(Chen et al., 2018e).欧阳敏研究组则发现ECD1蛋白通过参与叶绿体基因rps14的RNA编辑调控叶绿体早期发育(Jiang et al., 2018d).此外, 郁飞研究组和黄继荣研究组分别发现细胞质和叶绿体的翻译过程协同调控叶绿体发育, 及thf1突变体斑叶形成与质体翻译功能高度相关(Wang et al., 2018l; Huang et al., 2018b).类囊体是叶绿体基质中由单位膜围成的扁平囊, 是光合作用光反应的场所.Kang Byung-Ho研究组揭示了类囊体蛋白复合体的组装以及CURT1和FZL蛋白在类囊体生物发生过程中的关键作用(Liang et al., 2018d).隋森芳研究组与相关单位合作解析了红藻光系统I核心与捕光天线复合物(PSI-LHCR)的3.63?分辨率的三维结构, 发现红藻PSI核心既有蓝藻又有高等植物的特征(Pi et al., 2018).该研究对阐明PSI的进化和功能具有重要意义. ...
Medicago AP2-domain transcription factor WRI5a is a master regulator of lipid biosynthesis and transfer during mycorrhizal symbiosis 1 2018
... 种子含油量是油菜的重要性状之一.郭亮研究组与相关单位合作对油菜籽中油脂合成的调控机制进行研究, 发现种子中脂质代谢相关基因在种子的不同部位及不同种子之间表达量存在显著差异.该研究对高油分油菜育种具有重要指导意义(Lu et al., 2018b).王二涛研究组则对脂肪酸在植物和菌根真菌之间的转运调控机制进行探索, 发现在与丛枝菌根真菌共生过程中, 苜蓿转录因子WRI5a是脂肪酸碳源和磷营养交换的分子开关(Jiang et al., 2018e).淀粉体中合成与储存的淀粉是谷物胚乳中的主要储能分子.万建民研究组与章文华研究组合作解析了淀粉体发育和淀粉合成的分子机制, 揭示了FSE1为磷脂酶样蛋白质, 其可调控水稻胚乳中半乳糖脂的合成, 为水稻胚乳发育过程中脂质代谢与淀粉合成之间的关系提供了新观点(Long et al., 2018). ...
The arginine decarboxylase gene ADC1, associated to the putrescine pathway, plays an important role in potato cold-acclimated freezing tolerance as revealed by transcriptome and metabolome analyses 1 2018
... 随着全球气候的变化, 极度低温天气频发, 阶段性和局地性冷害加重, 严重影响了作物产量.因此, 开展作物耐低温研究意义重大.储成才研究组和王喜萍研究组合作发现了1个在进化过程中受到强烈选择的粳稻耐低温基因bZIP73.该基因编码区第511位单核苷酸多态性(single-nucleotide polymorphism, SNP)决定了籼、粳分化及低温的耐受性差异(Liu et al., 2018a).种康研究组则发现水稻MADS-box转录因子家族OsMADS57协同其互作蛋白OsTB1调控水稻的低温耐受性; 并发现二者对低温防御反应的调控依赖其共同靶基因OsWRKY94.常温下OsMADS57与OsTB1蛋白互作, 抑制独脚金内酯受体基因D14的转录, 促进水稻侧芽分化及分蘖形成; 低温下, OsMADS57与OsTB1通过直接激活OsWRKY94的转录, 启动防御反应.因此, OsMADS57具有平衡器官发生和防御反应的分子开关特性(Chen et al., 2018f).郝玉金和王小非研究组鉴定了苹果的R2R3-MYB转录因子MdMYB23, 发现过表达MdMYB23的转基因苹果愈伤组织和拟南芥耐寒性增强.MdMYB23可与MdCBF1和MdCBF2的启动子结合并激活其表达, 也能与原花青素生物合成的关键调节基因MdANR的启动子互作, 促进原花青素的积累和活性氧清除(An et al., 2018a).该研究揭示了MYB转录因子的功能及其在植物耐冷性调节中的潜在机制.宋波涛和谢从华研究组揭示了马铃薯精氨酸脱羧酶调控的腐胺合成是响应低温的重要途径, 首次证明转录因子CBF信号途径参与ADC1基因调控的马铃薯驯化抗寒(Kou et al., 2018).杨淑华研究组则发现了新生多肽链偶联蛋白复合体β亚基BTF3L (Basic transcription factor 3 like)正调控植物的低温应答.低温条件下, BTF3L蛋白被蛋白激酶OST1磷酸化, 导致BTF3L与CBFs蛋白互作增强, 提高了CBFs蛋白的稳定性, 进而增强植物的抗冻能力(Ding et al., 2018b).该研究揭示了OST1-BTF3L蛋白复合体调控植物响应低温胁迫的分子机制, 加深了人们对植物低温应答调控机理的理解.何跃辉研究组揭示了植物特异性支架蛋白FRI (FRIGIDA)介导拟南芥越冬习性形成的分子机制, 阐明了FRI通过超级复合体改变FLC位点的局部染色质环境, 从而激活FLC的表达, 促进转录延伸和共转录加工, 防止植物过冬前开花, 需经历冬季低温锻炼后才具备开花的潜力(Li et al., 2018k). ...
Arabidopsis class I formins control membrane-originated actin polymerization at pollen tube tips 1 2018
... 细胞骨架(cytoskeleton)系统主要包括微管(MTs)和微丝(F-actin), 其在调控植物生长发育和细胞形态建成等生理过程中具重要作用.然而, 人们对微管和微丝的调控机理及其影响植物形态发生发育机制的认识还十分有限.高等植物已进化出多种(不同大小、形状和功能)细胞类型, 如锥形花瓣表皮细胞, 然而调控该种细胞形态建成的分子机制目前尚不清楚.林德书研究组证实AN (ANGUSTIFOLIA)-ROS途径与KTN1 (katanin)共同作用, 通过参与调节微管骨架的排列方式, 进而调控花瓣锥形细胞发育(Dang et al., 2018).黄善金研究组则发现, 拟南芥成蛋白AtFH3 (Formin 3)和AtFH5参与调节花粉管顶端肌动蛋白的聚合和阵列构建, 对极性花粉管快速生长起关键作用(Lan et al., 2018).此外, 他们还发现拟南芥成蛋白AtFH2特异定位于胞间连丝, 水稻的部分Formin家族成员也定位于胞间连丝, 该研究揭示了拟南芥胞间连丝处微丝的含量及稳定性对调控胞间连丝通透性具重要作用(Diao et al., 2018).毛同林研究组从植物细胞骨架与环境因子互作方面进行了探索, 揭示了植物细胞受到高盐胁迫后, 乙烯通过上调微管稳定蛋白WDL5的表达调控微管重组(Dou et al., 2018), 及微管去稳定蛋白MDP60参与光和乙烯调控的下胚轴伸长(Ma et al., 2018c).孔照胜研究组则利用标记了微丝骨架(actin cytoskeleton)的稳定转基因蒺藜苜蓿, 首次在活细胞水平解析了根瘤发育不同阶段, 微丝骨架调控共生界面形成过程中的排布结构和动态模式, 为探索根瘤发育和生物固氮提供了细胞生物学依据(Zhang et al., 2019). ...
The ZmbZIP22 transcription factor regulates 27-kD gamma-zein gene transcription during maize endosperm development 1 2018
... 玉米胚乳是营养物质的主要储存组织.宋任涛研究组克隆了玉米突变体o11 (opaque11)的胚乳特异性bHLH转录因子O11.研究表明其不仅可直接调控胚乳发育的转录因子、cyPPDKs以及多个碳水化合物代谢酶, 还是ZmYoda的激活因子, 在胚乳发育和营养代谢调节网络中发挥中心调控作用(Feng et al., 2018); 另外, 该研究组还鉴定了1个与玉米储藏蛋白27 kDa γ-zein启动子相结合的bZIP型转录因子ZmbZIP22.该转录因子与PBF1、OHP1以及OHP2一起, 对27 kDa γ-zein进行转录调节, 影响胚乳发育(Li et al., 2018a). ...
The REN4 rheostat dynamically coordinates the apical and lateral domains of Arabidopsis pollen tubes 1 2018
... 花粉管生长是典型的细胞极性延伸现象.关跃峰研究组近期发现1个胞质定位的糖酵解关键酶——磷酸甘油酸激酶(PGKc).其突变体的花粉萌发速度大幅提升, 但花粉管极性形态异常.进一步研究发现, PGKc对花粉管极性的调控并非通过对细胞总体能量代谢的影响, 而是通过糖酵解途径(Chen et al., 2018i).杨贞标研究组则以拟南芥花粉管为模式系统, 阐明了植物小G蛋白ROP1、细胞骨架与胞吞作用共同形成调节花粉管极性生长的细胞与分子反馈调控网络, 揭示了网格蛋白介导的内吞作用调控花粉管顶端膜与侧面膜分子的极性分布及细胞顶端极性生长的分子机理(Li et al., 2018b).杨维才和李红菊研究组鉴定了1个影响高尔基体反面网络结构(trans-Golgi network, TGN)形成的蛋白LOT (Loss of TGN).在lot突变体中, 花粉管在柱头上的生长被抑制, 植物表现出严重的不育表型.进一步研究发现, 突变体中高尔基体膜囊过度堆积, 且源于高尔基体的分泌小泡以及TGN的数目显著减少.此外, 突变体花粉管中细胞壁组分——果胶质(pectin)等的形成均受到抑制, 细胞内吞作用也被显著阻碍, 从而影响花粉管的生长(Jia et al., 2018b). ...
A phytophthora effector recruits a host cytoplasmic transacetylase into nuclear speckles to enhance plant susceptibility 1 2018
... 王源超研究组则基于病毒诱导基因沉默技术的植物LRR类受体功能的高通量分析体系, 成功鉴定到植物中识别疫病菌模式分子XEG1的受体蛋白RXEG1.该蛋白是植物识别XEG1后产生细胞坏死及防卫反应的关键因子.激活RXEG1能显著提高植物对疫病菌的抗病性(Wang et al., 2018q).此外, 该研究组还发现大豆疫霉在侵染早期分泌的效应分子Avh52能够“挟持”大豆的组蛋白乙酰转移酶蛋白GmTAP1, 使其由细胞质进入细胞核, 通过乙酰化植物感病基因启动子区组蛋白H2A及H3上的激活位点, 激活寄主感病基因的表达, 促进病原菌成功侵染(Li et al., 2018c). ...
Hydrogen sulfide disturbs actin polymerization via S-sulfhydration resulting in stunted root hair growth 1 2018
... 茸毛作为植物体最外层结构, 具有很好的抗虫防病毒作用.叶志彪和杨长宪研究组分离了控制番茄腺体毛形成的关键基因H (Hair), 该基因编码C2H2锌指蛋白, 可能是茄科植物多细胞腺体毛形成的关键保守基因(Chang et al., 2018).唐克轩研究组发现R2R3 MYB第9亚家族的转录因子AaMIXTA1参与调控青蒿分泌型腺毛发育, 为研究其它植物多细胞腺毛奠定了基础(Shi et al., 2018c).此外, 他们还发现HD-ZIP IV转录因子AaHD8与AaMIXTA1相互作用形成复合体, 协同调控AaHD1和多个蜡质角质合成酶基因的表达, 从而促进蜡质合成和腺毛的起始发育(Yan et al., 2018d).陈晓亚研究组与相关单位合作对可纺纤维形成的分子机制进行研究, 发现编码bHLH转录因子的棉花PRE1是可纺纤维进化的目标基因, 其正调控纤维伸长.PRE1基因启动子的变异可促进棉花可纺纤维的形成(Zhao et al., 2018a).李学宝研究组鉴定了富含半胱氨酸的GhXLIM6蛋白, 其可与GhKNL1启动子的PAL-box结合而抑制该基因表达, 促进纤维素合成, 进而调控棉纤维伸长(Li et al., 2018i).俞嘉宁研究组则揭示了棉花线粒体Ghatp1基因C1292和C1415位点发生RNA编辑可影响ATPase产生ATP, 从而促进表皮毛及纤维伸长(He et al., 2018c).根毛是一种顶端生长的植物细胞, 微丝骨架对根毛的生长发育有极其重要的调控作用.贾红磊研究组揭示了H2S通过硫巯基化修饰扰乱正常的微丝骨架聚合及生长模式, 从而影响植物根毛的生长, 为进一步探索H2S调控植物生长发育的作用机制提供了理论依据(Li et al., 2018d). ...
APURINIC/ APYRIMIDINIC ENDONUCLEASE2 and ZINC FINGER DNA 3'-PHOSPHOESTERASE play overlapping roles in the maintenance of epigenome and genome stability 1 2018
... DNA甲基化主要指在DNA甲基转移酶的作用下, 将供体S-腺苷甲硫氨酸上的甲基转移至DNA受体碱基上的过程.基因组全局性DNA甲基化水平需要通过DNA甲基化和去甲基化的动态平衡调节.钱伟强研究组揭示了拟南芥中2种碱基切除修复蛋白APE2 (APURINIC/APYRIMIDINIC ENDONUCLEASE 2) 和ZDP (ZINC FINGER DNA 3'-PHOSPHOESTERASE)在DNA损伤修复以及环境胁迫中的生物学功能.全基因组胞嘧啶甲基化(5mC)分析表明, 缺失APE2和ZDP基因的双突变体中存在与单突变体不同的2 000余个DNA高甲基化位点; 且双突变体表现出生长发育缺陷及对DNA烷基化试剂超敏感.该研究证明了APE2和ZDP在维持植物表观基因组和基因组稳定性中的重要作用(Li et al., 2018e). ...
Genomic imprinting was evolutionarily conserved during wheat polyploidization 1 2018
... 多倍体在生物界广泛存在, 很多作物都是多倍体.刘宝研究组利用六倍体小麦基因组中2个基因组——乌拉尔图小麦(Triticum urartu, AA)和节节麦(Aegilops tauschii, DD)构建了人工异源多倍体(AADD), 并对其连续12代个体的核型和表型变异进行了检测, 揭示了异源多倍化催生了大量可遗传的核型异质性和表型多样性, 为“全基因组重复能加大生物的演化潜力”观点提供了鲜活的实验证据(Gou et al., 2018).辛明明研究组则对山羊草属(Aegilops)中一些二倍体, 以及小麦属(Triticum)中一些四倍体和六倍体胚乳中亲本特异性基因的表达模式进行了分析, 结果表明小麦多倍体化过程中的基因组印记(一种表观遗传现象, 导致基因根据其亲本来源出现差异表达)在杂交多倍化演化过程中是保守的(Yang et al., 2018d). ...
Maize Oxalyl-CoA Decarboxylase1 degrades oxalate and affects the seed metabolome and nutritional quality 1 2018
... 4-羟基-2,5-二甲基-3(2H)-呋喃酮(HDMF)是草莓果实的特征芳香组分, 其催化合成的关键酶是草莓醌氧化还原酶(FaQR).陈昆松研究组发现乙烯应答因子(FaERF9)为FaQR启动子的正调节因子, 揭示了ERF-MYB复合物通过激活FaQR表达调节草莓中HDMF生物合成的分子机制(Zhang et al., 2018m).苹果酸是苹果果实中的主要有机酸.韩振海和张新忠研究组获得了4个与苹果酸相关的主效QTLs; 并证明候选基因MdSAUR37等存在上位效应, 共同调控苹果酸的积累(Jia et al., 2018a).草酸是最简单的二元酸, 在调控金属胁迫等方面起积极作用.巫永睿研究组克隆了玉米草酸降解途径中的草酰辅酶A基因, 阐明了玉米草酸代谢的前两步反应, 揭示了草酸代谢参与籽粒储藏物质积累和营养品质形成的分子机理, 为遗传改良草酸含量较高的蔬菜提供了候选基因(Yang et al., 2018e). ...
STERILE APETALA modulates the stability of a repressor protein complex to control organ size in Arabidopsis thaliana 1 2018
... 丁勇研究组则对拟南芥种子下胚轴发育进行研究, 发现参与拟南芥组蛋白修饰的MLK1和MLK2、GA信号途径负调控因子RGA, 以及与昼夜节律相关的CCA1形成蛋白复合体, 该复合体直接调控下游基因DWF4的表达, 进而影响下胚轴伸长(Zheng et al., 2018a).李云海研究组以拟南芥为材料对控制植物器官大小的分子机制进行探索.他们发现E3连接酶F-盒组分SAP蛋白的2个底物KIX8和KIX9与PPD形成复合物行使功能, 其通过调控拟分生组织细胞增殖来控制器官大小(Li et al., 2018f).玉米籽粒性状直接影响其产量.汤继华研究组与李文学研究组合作发现了1个控制玉米籽粒发育性状的基因ZmUrb2, 该基因主要通过影响核糖体的生物合成和核糖体rRNA前体加工来影响籽粒发育和整个营养生长过程(Wang et al., 2018d). ...
Modulating plant growth-metabolism coordination for sustainable agriculture 1 2018
... 2018年, 植物科学的亮点成果主要体现在如下6个方面.一是水稻生物学研究取得新突破.傅向东研究组发现水稻生长调节因子GRF4与生长抑制因子DELLA相互间的反向平衡赋予了植物生长与碳-氮代谢间的稳态共调节, 揭示了提高“绿色革命”品种的氮肥利用效率并增加谷物产量的遗传机制(Li et al., 2018g).这一重大发现预示着一场新的绿色革命即将到来, 该成果入选2018年度中国科学十大进展.万建民研究组与相关单位合作, 运用自私基因模型揭示了水稻杂种不育现象, 探讨了“毒性-解毒”分子机制在水稻杂种不育中的普遍性, 揭示了自私基因在维持植物基因组的稳定性和促进新物种形成中的重要作用; 研究结果为创制“广亲和”材料, 克服杂种不育障碍, 充分利用杂种优势和野生种质资源, 提高水稻单产奠定了基础(Yu et al., 2018). ...
The RIN-MC fusion of MADS-box transcription factors has transcriptional activity and modulates expression of many ripening genes 1 2018
... 植物分枝是影响植物株型的关键因素.秦根基研究组与相关单位合作对植物分枝形成的分子调控机制进行研究, 发现了1个分枝正调控因子TIE1, 它与分枝重要调控因子BRC1 (BRANCHED1)发生互作, 抑制其转录活性.通过调节HB21、HB40和HB53等BRC1下游基因的转录, 发挥促进腋芽分枝的作用(Yang et al., 2018h).开花是植物由营养生长转向生殖生长的标志.王前和张小兰研究组对热诱导开花的分子机理进行探索, 证明LsSOC1是热诱导花芽形成的激活子, 热激转录因子HsfA1e和HsfA4c通过结合在LsSOC1基因的启动子上, 影响其表达, 为开花相关机制研究奠定了理论基础(Chen et al., 2018k).果实成熟过程对于果实品质形成和种子传播非常关键.陈建业和邝健飞研究组发现转录因子MaMYB3通过抑制淀粉降解相关基因以及MabHLH6转录因子的转录水平来调控淀粉降解, 丰富了香蕉果实成熟的转录调控网络(Fan et al., 2018).朱本忠和罗云波研究组则发现, 在番茄rin突变体中, 基因组DNA片段部分缺失导致形成RIN-MC片段的嵌合, 该嵌合能够调控许多成熟相关基因的表达, 从而调节果实成熟(Li et al., 2018h).此外, 吴鸿研究组与相关单位合作阐明了拟南芥角果开裂的调控机制.他们通过对纤维素酶CEL6和半纤维素酶MAN7以及果胶酶ADPG2基因的一系列突变体果荚离层进行细胞超微结构观察, 并结合基因表达的检测结果, 证明改变这些细胞壁酶的表达量和酶活性可以调控果荚开裂区细胞分化和离层区细胞降解(He et al., 2018a).该研究为后期培育不同程度的果实抗开裂油料作物提供了技术支持. ...
The cotton XLIM protein (GhXLIM6) is required for fiber development via maintaining dynamic F-actin cytoskeleton and modulating cellulose biosynthesis 1 2018
... 茸毛作为植物体最外层结构, 具有很好的抗虫防病毒作用.叶志彪和杨长宪研究组分离了控制番茄腺体毛形成的关键基因H (Hair), 该基因编码C2H2锌指蛋白, 可能是茄科植物多细胞腺体毛形成的关键保守基因(Chang et al., 2018).唐克轩研究组发现R2R3 MYB第9亚家族的转录因子AaMIXTA1参与调控青蒿分泌型腺毛发育, 为研究其它植物多细胞腺毛奠定了基础(Shi et al., 2018c).此外, 他们还发现HD-ZIP IV转录因子AaHD8与AaMIXTA1相互作用形成复合体, 协同调控AaHD1和多个蜡质角质合成酶基因的表达, 从而促进蜡质合成和腺毛的起始发育(Yan et al., 2018d).陈晓亚研究组与相关单位合作对可纺纤维形成的分子机制进行研究, 发现编码bHLH转录因子的棉花PRE1是可纺纤维进化的目标基因, 其正调控纤维伸长.PRE1基因启动子的变异可促进棉花可纺纤维的形成(Zhao et al., 2018a).李学宝研究组鉴定了富含半胱氨酸的GhXLIM6蛋白, 其可与GhKNL1启动子的PAL-box结合而抑制该基因表达, 促进纤维素合成, 进而调控棉纤维伸长(Li et al., 2018i).俞嘉宁研究组则揭示了棉花线粒体Ghatp1基因C1292和C1415位点发生RNA编辑可影响ATPase产生ATP, 从而促进表皮毛及纤维伸长(He et al., 2018c).根毛是一种顶端生长的植物细胞, 微丝骨架对根毛的生长发育有极其重要的调控作用.贾红磊研究组揭示了H2S通过硫巯基化修饰扰乱正常的微丝骨架聚合及生长模式, 从而影响植物根毛的生长, 为进一步探索H2S调控植物生长发育的作用机制提供了理论依据(Li et al., 2018d). ...
HEIP1 regulates crossover formation during meiosis in rice 1 2018
... 减数分裂是真核生物有性生殖所必需的环节.程祝宽和李亚非研究组对水稻减数分裂时胞质分裂的遗传调控机制进行了探索.鉴定出1个调控花粉母细胞胞质分裂的蛋白DCM1 (Defective Callose in Meiosis 1).该蛋白的C端含有5个串联的CCCH锌指结构域, 并与核多聚腺苷酸结合蛋白OsPABNs存在互作.dcm1突变体的花粉母细胞在胞质分裂时, 由于胼胝质板提前降解, 胞质分裂无法进行, 使得减数第二次分裂的纺锤体取向异常, 并最终表现为雄性不育(Zhang et al., 2018a).此外, 程祝宽研究组在水稻中成功分离了1个新的减数分裂交叉结形成相关蛋白HEIP1 (HEI10 Interaction Protein 1).实验证明该蛋白与HEI10、MSH5和ZIP4等交叉结形成促进因子相互作用并共定位, 调控减数分裂一类交叉的形成(Li et al., 2018j).该研究为深入揭示减数分裂同源重组形成的分子机制奠定了基础.该研究组还与马伯军研究组合作通过筛选水稻减数分裂缺陷的不育突变体, 发现LEPTO1 (LEPTOTENE1)参与水稻减数分裂细线期染色体的形态建成.lepto1突变体的花粉母细胞染色体停滞在前细线期状态, 不能进一步组装进入典型的细线期染色体状态.性母细胞中没有DSB形成, 也没有减数分裂特异蛋白的组装, 观察不到胼胝质的积累.LEPTO1编码OsRR24, 属于B类响应调节因子, N端具有保守的DDK和MYB结构域, C端具有转录激活活性, 表明LEPTO1可能通过调控相关基因的表达, 进而调节减数分裂细线期染色体的形态建成(Zhao et al., 2018f).王应祥研究组与贺岩研究组合作探究了减数分裂重组过程中DNA双链断裂(double- strand breaks, DSB)的数目对重组稳态和最终分布的影响.他们通过转基因技术在DSB缺失突变体(spo11-/-)背景下操控DSB发生的数目, 揭示了减数分裂中DSB形成的数目对重组的稳态及分布有重要调控作用(Xue et al., 2018a). ...
FRIGIDA establishes a local chromosomal environment for FLOWERING LOCUS C mRNA production 1 2018
... 随着全球气候的变化, 极度低温天气频发, 阶段性和局地性冷害加重, 严重影响了作物产量.因此, 开展作物耐低温研究意义重大.储成才研究组和王喜萍研究组合作发现了1个在进化过程中受到强烈选择的粳稻耐低温基因bZIP73.该基因编码区第511位单核苷酸多态性(single-nucleotide polymorphism, SNP)决定了籼、粳分化及低温的耐受性差异(Liu et al., 2018a).种康研究组则发现水稻MADS-box转录因子家族OsMADS57协同其互作蛋白OsTB1调控水稻的低温耐受性; 并发现二者对低温防御反应的调控依赖其共同靶基因OsWRKY94.常温下OsMADS57与OsTB1蛋白互作, 抑制独脚金内酯受体基因D14的转录, 促进水稻侧芽分化及分蘖形成; 低温下, OsMADS57与OsTB1通过直接激活OsWRKY94的转录, 启动防御反应.因此, OsMADS57具有平衡器官发生和防御反应的分子开关特性(Chen et al., 2018f).郝玉金和王小非研究组鉴定了苹果的R2R3-MYB转录因子MdMYB23, 发现过表达MdMYB23的转基因苹果愈伤组织和拟南芥耐寒性增强.MdMYB23可与MdCBF1和MdCBF2的启动子结合并激活其表达, 也能与原花青素生物合成的关键调节基因MdANR的启动子互作, 促进原花青素的积累和活性氧清除(An et al., 2018a).该研究揭示了MYB转录因子的功能及其在植物耐冷性调节中的潜在机制.宋波涛和谢从华研究组揭示了马铃薯精氨酸脱羧酶调控的腐胺合成是响应低温的重要途径, 首次证明转录因子CBF信号途径参与ADC1基因调控的马铃薯驯化抗寒(Kou et al., 2018).杨淑华研究组则发现了新生多肽链偶联蛋白复合体β亚基BTF3L (Basic transcription factor 3 like)正调控植物的低温应答.低温条件下, BTF3L蛋白被蛋白激酶OST1磷酸化, 导致BTF3L与CBFs蛋白互作增强, 提高了CBFs蛋白的稳定性, 进而增强植物的抗冻能力(Ding et al., 2018b).该研究揭示了OST1-BTF3L蛋白复合体调控植物响应低温胁迫的分子机制, 加深了人们对植物低温应答调控机理的理解.何跃辉研究组揭示了植物特异性支架蛋白FRI (FRIGIDA)介导拟南芥越冬习性形成的分子机制, 阐明了FRI通过超级复合体改变FLC位点的局部染色质环境, 从而激活FLC的表达, 促进转录延伸和共转录加工, 防止植物过冬前开花, 需经历冬季低温锻炼后才具备开花的潜力(Li et al., 2018k). ...
l). Polycomb-mediated gene silencing by the BAH-EMF1 complex in plants 1 2018
... 染色质的基本单位为核小体, 核小体核心组蛋白的N端存在多种共价修饰, 这些修饰在植物生长发育过程中不可或缺.Polycomb家族蛋白所建立的H3K27me3修饰在基因沉默过程中行使重要功能.何跃辉研究组利用酵母双杂交技术鉴定到拟南芥EMF1 (EMBRYONIC FLOWER 1)的互作蛋白SHL (SHORT LIFE)和EBS (EARLY BOLTING IN SHORT DAYS). SHL以及EBS的BAM结构域能够特异性识别H3K27me3, 影响全基因组范围H3K27me3的建立, 进而抑制基因的表达(Li et al., 2018l).同时, 杜嘉木研究组与相关单位合作证明了EBS的BAH和PHD结构域分别识别H3K27me3及H3K4me3修饰.EBS通过一种特殊的自动抑制调控方式来平衡与两种拮抗型修饰的结合, 破坏BAH-H3K27me3或PHD-H3K4me3的结合均会导致拟南芥出现早花表型(Yang et al., 2018i).该研究揭示了识别蛋白通过识别两种不同类型甲基化修饰调控植物开花的新机制.此外, 该研究组还与曹晓风研究组合作解析了拟南芥KDM5亚家族组蛋白去甲基化酶JMJ14 (JUMONJI14)以及H3K4me3复合物的晶体结构.JMJ14蛋白与底物复合物结构的解析, 为了解JMJ14在开花等生理过程中发挥作用的分子基础提供了理论依据(Yang et al., 2018j). ...
Photoexcited CRYPTOCHROME 1 interacts directly with G-protein beta subunit AGB1 to regulate the DNA-binding activity of HY5 and photomorphogenesis in Arabidopsis 1 2018
... 光对植物不仅是一种能源, 也是一种重要的环境信号.植物通过向光素(phot)、隐花色素(cry)和光敏色素(phy)等光受体来感应光信号.林金星研究组研究了蓝光对细胞质膜上向光素phot1的定位变化和动态性质, 在单分子水平上揭示了蓝光信号传递的起始事件及调控机制(Xue et al., 2018b).刘宏涛研究组发现了隐花色素互作转录因子CIB1通过直接结合cry2和CO来介导cry2-CIB1-CO复合体的形成, 揭示了蓝光信号调控植物开花时间的新机制(Liu et al., 2018g).杨洪全研究组发现, G蛋白β亚基AGB1通过与HY5互作抑制其DNA结合能力, 从而抑制光信号; 而蓝光信号通过CRY1与AGB1互作促进AGB1与HY5解离来抑制AGB1的功能(Lian et al., 2018); 并发现CRY1与去磷酸化形式的BL信号关键转录因子BES1发生依赖于蓝光的直接互作, 揭示了植物能根据光信号和内在BL信号有无或强弱的动态变化来优化其光形态建成(Wang et al., 2018n).此外, 该研究组还发现光受体cry1、phyB竞争性与生长素信号通路抑制子Aux/IAA结合, 并磷酸化后者而负调控生长素信号转导, 进而精确调控植物光信号与生长素信号通路的平衡, 优化其生长(Xu et al., 2018c). ...
Direct production of dihydroxylated sesquiterpenoids by a maize terpene synthase 1 2018
... 尼克酰胺腺嘌呤二核苷酸(NAD)从尼克酸(NA) 合成.目前已发现多种NA修饰(如糖基化).王国栋研究组新发现一种NA修饰——甲酯化(MeNA).MeNA是一种NAD前体, 可在植物不同组织间进行更长距离运输.该研究提出了植物中NAD生物合成与NA衍生物之间关系的新见解(Wu et al., 2018b).萜类代谢物在植物生长发育等方面发挥重要作用.漆小泉研究组与相关单位合作发现一个新的、多产物OSC (2,3-氧化鲨烯环化酶) ——籼稻醇合酶(OsOS).该酶合成一种新的五环三萜籼稻醇(orysatinol)的主产物及12种不同的三萜类化合物(Xue et al., 2018c).王强研究组与相关单位合作发现玉米倍半萜合酶能直接催化(E, E)-法呢基二磷酸, 形成二羟基化的二醇产物(主要是eudesmane-2,11-diol) (Liang et al., 2018a).该研究揭示了TPS的新型催化功能及形成二羟基化倍半萜的催化机制. ...
UVR8 interacts with BES1 and BIM1 to regulate transcription and photomorphogenesis in Arabidopsis 1 2018
... 邓兴旺研究组与许冬清研究组合作鉴定了光形态建成的正调节因子CSU4, 它能整合生物钟节律和光信号(Zhao et al., 2018g).他们还发现BBX21通过第2个B-box结构域与HY5启动子中的T/G-box结合, 调控HY5及其靶基因的表达以促进植物光形态建成(Xu et al., 2018b).而BBX28通过与HY5蛋白的C末端互作, 抑制HY5与下游靶基因启动子结合, 从而抑制HY5活性及其下游靶基因表达, 最终抑制植物光形态建成(Lin et al., 2018a).此外, 邓兴旺和钟上威研究组揭示了PIFs、HY5和EIN3及EIL1对出土幼苗形态建成的调控作用, 为阐明幼苗从暗形态建成转向光形态建成的分子机制提供了新证据(Shi et al., 2018b).吕应堂研究组则鉴定了光形态建成的正调控因子SRS5, 它直接结合HY5、BBX21和BBX22的启动子并激活其表达, 从而促进光形态建成(Yuan et al., 2018).刘宏涛研究组发现UV-B照射后, UVR8形成单体并在细胞核中富集, UVR8通过抑制BES1/BIM1结合DNA的能力从而抑制下游生长相关基因的表达, 并抑制细胞伸长及光形态建成(Liang et al., 2018b).同时, 发现UV-B激活的UVR8形成单体, 进入细胞核直接结合WRKY36而抑制其结合HY5启动子, 进而抑制下胚轴伸长(Yang et al., 2018g). ...
DNA N-6-adenine methylation in Arabidopsis thaliana 1 2018
... 6mA是原核生物基因组DNA最常见的修饰, 近年来6mA修饰在真核生物中的作用也被陆续报道.谷晓峰研究组与相关单位合作发现6mA修饰在拟南芥基因组中广泛存在, 且水平与活跃基因的表达呈现出相关性(Liang et al., 2018c).而周道绣和曾志雄研究组对水稻基因组中的6mA修饰进行了分析, 发现位于启动子区域的6mA与基因表达抑制相关, 而位于转录区的6mA与基因激活相关.转座子上的6mA与5mC呈现互补性, 表明DNA腺嘌呤甲基化与胞嘧啶甲基化在水稻中可以协同作用, 共同调控基因的表达(Zhou et al., 2018a). ...
Thylakoid-bound polysomes and a dynamin-related protein, FZL, mediate critical stages of the linear chloroplast biogenesis program in greening Arabidopsis cotyledons 1 2018
... 叶绿体是植物进行光合作用的细胞器, 其分裂由众多因子形成的复合物协助完成.张敏研究组发现MCD1在此过程中起重要作用.MCD1通过与ARC6作用, 识别FtsZ环, 并引导Min复合物整合到分裂环上, 从而调控叶绿体分裂(Chen et al., 2018e).欧阳敏研究组则发现ECD1蛋白通过参与叶绿体基因rps14的RNA编辑调控叶绿体早期发育(Jiang et al., 2018d).此外, 郁飞研究组和黄继荣研究组分别发现细胞质和叶绿体的翻译过程协同调控叶绿体发育, 及thf1突变体斑叶形成与质体翻译功能高度相关(Wang et al., 2018l; Huang et al., 2018b).类囊体是叶绿体基质中由单位膜围成的扁平囊, 是光合作用光反应的场所.Kang Byung-Ho研究组揭示了类囊体蛋白复合体的组装以及CURT1和FZL蛋白在类囊体生物发生过程中的关键作用(Liang et al., 2018d).隋森芳研究组与相关单位合作解析了红藻光系统I核心与捕光天线复合物(PSI-LHCR)的3.63?分辨率的三维结构, 发现红藻PSI核心既有蓝藻又有高等植物的特征(Pi et al., 2018).该研究对阐明PSI的进化和功能具有重要意义. ...
Comparison of Oryza sativa and Oryza brachyantha genomes reveals selection-driven gene escape from the centromeric regions 1 2018
... 栽培稻和普通野生稻(Oryza rufipogon)丰富的遗传多样性是水稻育种的主要来源.黄学辉研究组对66个来自不同水稻类群的栽培稻品种和野生稻株系的精细基因组进行了比较分析, 鉴定出水稻基因组中各类复杂的遗传变异, 并发现很多功能基因存在多种等位基因类型(Zhao et al., 2018e).发达的根系是旱粳稻和热带粳稻适应干旱环境最典型的根系形态特征.李自超研究组与黎志康研究组合作, 将795份栽培稻种质资源的根系性状和330多万个高质量的SNP数据进行关联分析, 获得几十个候选基因; 对这些候选基因的序列进行分析, 发现几乎所有位点在野生稻中都以低频存在.因此, 与旱粳稻长根和粗根等性状相关的基因是在高山干旱环境中通过正向选择及聚合而固定下来的(Zhao et al., 2018i).此外, 陈明生研究组对约1 500万年前分化的短花药野生稻(Oryza brachyantha)和栽培稻着丝粒周围序列进行了比较, 发现该区域倾向有更多的共线基因丢失(大多转移到其它区域).进一步分析表明, 这种基因丢失可能受着丝粒染色质动态变化(如扩展或微易位)驱动(Liao et al., 2018). ...
B-BOX DOMAIN PROTEIN28 negatively regulates photomorphogenesis by repressing the activity of transcription factor HY5 and undergoes COP1- mediated degradation 1 2018
... 邓兴旺研究组与许冬清研究组合作鉴定了光形态建成的正调节因子CSU4, 它能整合生物钟节律和光信号(Zhao et al., 2018g).他们还发现BBX21通过第2个B-box结构域与HY5启动子中的T/G-box结合, 调控HY5及其靶基因的表达以促进植物光形态建成(Xu et al., 2018b).而BBX28通过与HY5蛋白的C末端互作, 抑制HY5与下游靶基因启动子结合, 从而抑制HY5活性及其下游靶基因表达, 最终抑制植物光形态建成(Lin et al., 2018a).此外, 邓兴旺和钟上威研究组揭示了PIFs、HY5和EIN3及EIL1对出土幼苗形态建成的调控作用, 为阐明幼苗从暗形态建成转向光形态建成的分子机制提供了新证据(Shi et al., 2018b).吕应堂研究组则鉴定了光形态建成的正调控因子SRS5, 它直接结合HY5、BBX21和BBX22的启动子并激活其表达, 从而促进光形态建成(Yuan et al., 2018).刘宏涛研究组发现UV-B照射后, UVR8形成单体并在细胞核中富集, UVR8通过抑制BES1/BIM1结合DNA的能力从而抑制下游生长相关基因的表达, 并抑制细胞伸长及光形态建成(Liang et al., 2018b).同时, 发现UV-B激活的UVR8形成单体, 进入细胞核直接结合WRKY36而抑制其结合HY5启动子, 进而抑制下胚轴伸长(Yang et al., 2018g). ...
LFR is functionally associated with AS2 to mediate leaf development in Arabidopsis 1 2018
Genome sequence of the progenitor of wheat A subgenome Triticum urartu 1 2018
... 二是植物系统进化研究取得了系列突破性成果.陈之端研究组重建了中国被子植物生命之树, 揭示了中国被子植物系统发育多样性形成的时空格局, 并明确了属、种水平分别应该重点保护的热点地区, 对我国保护区建设意义重大(Lu et al., 2018a).该成果入选2018年度中国生命科学十大进展.凌宏清、梁承志、王道文和张爱民研究组完成了乌拉尔图小麦材料G1812的基因组测序和精细组装, 绘制出小麦A基因组7条染色体的分子图谱(Ling et al., 2018).研究成果对进一步推动栽培小麦的遗传改良具有重要的理论意义和实用价值.叶凯研究组破译了罂粟基因组DNA密码, 揭示了其合成重要药用化合物的关键步骤, 这一发现为提高罂粟药用成分的产量及抗病性奠定了基础(Guo et al., 2018b).黎志康研究组与相关单位合作, 深入探讨了水稻起源、分类以及驯化规律, 揭示了亚洲栽培稻的起源和群体基因组变异结构, 剖析了水稻核心种质资源的基因组遗传多样性(Wang et al., 2018o), 为水稻规模化基因发掘和复杂性状分子改良奠定了基础. ...
Early selection of bZIP73 facilitated adaptation ofjaponica rice to cold climates 1 2018
... 随着全球气候的变化, 极度低温天气频发, 阶段性和局地性冷害加重, 严重影响了作物产量.因此, 开展作物耐低温研究意义重大.储成才研究组和王喜萍研究组合作发现了1个在进化过程中受到强烈选择的粳稻耐低温基因bZIP73.该基因编码区第511位单核苷酸多态性(single-nucleotide polymorphism, SNP)决定了籼、粳分化及低温的耐受性差异(Liu et al., 2018a).种康研究组则发现水稻MADS-box转录因子家族OsMADS57协同其互作蛋白OsTB1调控水稻的低温耐受性; 并发现二者对低温防御反应的调控依赖其共同靶基因OsWRKY94.常温下OsMADS57与OsTB1蛋白互作, 抑制独脚金内酯受体基因D14的转录, 促进水稻侧芽分化及分蘖形成; 低温下, OsMADS57与OsTB1通过直接激活OsWRKY94的转录, 启动防御反应.因此, OsMADS57具有平衡器官发生和防御反应的分子开关特性(Chen et al., 2018f).郝玉金和王小非研究组鉴定了苹果的R2R3-MYB转录因子MdMYB23, 发现过表达MdMYB23的转基因苹果愈伤组织和拟南芥耐寒性增强.MdMYB23可与MdCBF1和MdCBF2的启动子结合并激活其表达, 也能与原花青素生物合成的关键调节基因MdANR的启动子互作, 促进原花青素的积累和活性氧清除(An et al., 2018a).该研究揭示了MYB转录因子的功能及其在植物耐冷性调节中的潜在机制.宋波涛和谢从华研究组揭示了马铃薯精氨酸脱羧酶调控的腐胺合成是响应低温的重要途径, 首次证明转录因子CBF信号途径参与ADC1基因调控的马铃薯驯化抗寒(Kou et al., 2018).杨淑华研究组则发现了新生多肽链偶联蛋白复合体β亚基BTF3L (Basic transcription factor 3 like)正调控植物的低温应答.低温条件下, BTF3L蛋白被蛋白激酶OST1磷酸化, 导致BTF3L与CBFs蛋白互作增强, 提高了CBFs蛋白的稳定性, 进而增强植物的抗冻能力(Ding et al., 2018b).该研究揭示了OST1-BTF3L蛋白复合体调控植物响应低温胁迫的分子机制, 加深了人们对植物低温应答调控机理的理解.何跃辉研究组揭示了植物特异性支架蛋白FRI (FRIGIDA)介导拟南芥越冬习性形成的分子机制, 阐明了FRI通过超级复合体改变FLC位点的局部染色质环境, 从而激活FLC的表达, 促进转录延伸和共转录加工, 防止植物过冬前开花, 需经历冬季低温锻炼后才具备开花的潜力(Li et al., 2018k). ...
Shortened basal internodes encodes a gibberellin 2-oxidase and contributes to lodging resistance in rice 1 2018
... 钱前研究组则报道了一种新的种质资源小薇.它具有生长周期短和株型矮小等表型, 可以像拟南芥一样在实验室内大规模种植.小薇的超矮秆表型是由于GA生物合成基因D18突变引起(Hu et al., 2018a).该研究创制了“小薇日本晴”和“小薇93”, 为进行水稻功能基因组相关研究提供了重要资源.李来庚研究组与杨远柱研究组合作发现了一个新的特异调控水稻茎秆基部节长度的基因SBI, 该基因编码GA2氧化酶, 在培育水稻半矮秆性状, 提高抗倒伏能力, 增加水稻产量方面具重要的应用价值(Liu et al., 2018b).周奕华研究组揭示了IIP4蛋白通过与NAC29/NAC31互作, 负向调控次生壁合成, 进而影响茎秆强度(Zhang et al., 2018b).曹立勇和程式华研究组阐明了水稻叶片衰老的新机制.他们合作克隆了1个引起水稻叶片早衰的基因OsMTS1, 该基因编码褪黑素生物合成途径中的甲基转移酶, OsMTS1突变后可导致水稻叶片中褪黑素含量降低, 从而引起叶片早衰(Hong et al., 2018). ...
Shifting plant species composition in response to climate change stabilizes grassland primary production 1 2018
... 青藏高原近年来经历了快速(2倍于全球平均值)升温过程, 且降水格局也发生了显著变化, 然而剧烈的气候变化将如何影响高寒草地生态系统的结构和功能, 一直以来备受国内外研究者的关注.贺金生研究组发现尽管过去32年来海北站气候呈现暖干化趋势, 但草地生产力无显著变化, 功能群组成改变是生产力稳定性维持的主要原因(Liu et al., 2018c).该研究表明物种多样性在气候变化下生产力维持过程中起着关键作用.姜勇研究组与相关单位合作探讨了环境变化影响我国温带典型草原生产力的直接和间接过程.他们发现增加氮、水不仅直接提高生产力, 而且还通过增加植物功能多样性的2个组分间接驱动生产力(Xu et al., 2018h).该研究表明群落中优势物种的性状及物种间的性状分布在调节生态系统功能中均具重要作用. ...
Mutations in the DNA demethylase OsROS1 result in a thickened aleurone and improved nutritional value in rice grains 1 2018
... 储成才研究组克隆了穗发芽调控基因PHS8, 该基因突变可导致胚乳中小分子糖积累, 从而抑制ABA信号通路中OsABI3和OsABI5的表达, 导致穗发芽表型(Du et al., 2018b).刘春明研究组与相关单位合作创建了一种半粒种子筛选体系, 并利用该体系筛检获得了1个糊粉层增厚的水稻品系ta2.基因克隆发现DNA去甲基化酶基因OsROS1与糊粉层的发育相关, 有望开拓水稻营养品质育种新路径(Liu et al., 2018d). ...
G-protein betagamma subunits determine grain size through interaction with MADS-domain transcription factors in rice 1 2018
... 刘建祥和罗小金研究组、宋献军研究组、何予卿研究组同时克隆到与粒型和粒重相关的同一主效负调控QTL/基因qTGW3/TGW3/GL3.3, 并解析了相关分子机制(Hu et al., 2018c; Xia et al., 2018; Ying et al., 2018).李云海研究组则与相关单位合作揭示了OsMKKK10-OsMKK4-OsMAPK6信号途径正向调控水稻粒型和粒重(Xu et al., 2018e); 并发现大粒基因LARGE8可与OsMAPK6直接互作使其失活, 进而抑制颖壳细胞增殖, 负向调控水稻粒型(Xu et al., 2018f).余四斌研究组揭示了OsGBPs作为转录因子能够识别GAGA基序, 进而调控水稻粒长和植株高度(Gong et al., 2018).梁国华研究组则证明OsSPMS1基因负向调控水稻种子萌发、粒型和单株产量, 是水稻增产育种的重要靶标基因(Tao et al., 2018).刘巧泉研究组发现了水稻粒型与外观品质调控基因GS9, 其编码蛋白可与OsOFP14和OsOFP8蛋白互作, 通过改变细胞分裂调控水稻粒型(Zhao et al., 2018c).傅向东研究组克隆了1个控制水稻产量和提升稻米品质的重要基因LGY3, 该基因编码转录因子OsMADS1.可变剪接蛋白OsMADS1lgy3可提高稻米粒长, 降低垩白率, 进而影响稻米产量和外观品质(Liu et al., 2018e). ...
Functional diversification of Flowering Locus T homologs in soybean: GmFT1a and GmFT2a/5a have opposite roles in controlling flowering and maturation 1 2018
... 大豆属于短日照植物, 对光周期极为敏感.李文滨研究组发现, 大豆中SKIP同源蛋白GmGBP1参与光周期介导的开花途径, 其启动子区域的自然变异调控大豆在不同环境中更早开花和成熟(Zhao et al., 2018d).此外, 韩天富研究组发现, 大豆FT基因家族成员GmFT1a能够延迟大豆开花和成熟, 与开花促进基因GmFT2a/GmFT5a相互拮抗.基于本研究及前人的研究成果, 他们提出了FT基因家族精细调控大豆开花时间的跷跷板模型(Liu et al., 2018f).生物钟是生物体适应环境昼夜周期变化而进化出的协调细胞内基因表达、代谢网络调控的分子系统.吴昌银研究组前期发现, OsELF3作为一个生物钟基因, 其昼夜表达相位是影响长日照条件下水稻抽穗期的关键因素.近期他们发现泛素化连接酶HAF1与OsELF3.1互作, 从而调控其蛋白昼夜节律性积累, 且单个氨基酸(L558S)变异影响二者的互作, 从而影响水稻的区域适应性(Zhu et al., 2018a).王雷研究组则发现生物钟核心组分Evening Complex各组分的突变体叶片提前衰老.进一步研究发现, Evening Complex直接结合MYC2基因启动子并抑制其表达, 从而在时间维度精细调控茉莉酸诱导的植物叶片衰老进程(Zhang et al., 2018l). ...
CIB1 and CO interact to mediate CRY2-dependent regulation of flowering 1 2018
... 光对植物不仅是一种能源, 也是一种重要的环境信号.植物通过向光素(phot)、隐花色素(cry)和光敏色素(phy)等光受体来感应光信号.林金星研究组研究了蓝光对细胞质膜上向光素phot1的定位变化和动态性质, 在单分子水平上揭示了蓝光信号传递的起始事件及调控机制(Xue et al., 2018b).刘宏涛研究组发现了隐花色素互作转录因子CIB1通过直接结合cry2和CO来介导cry2-CIB1-CO复合体的形成, 揭示了蓝光信号调控植物开花时间的新机制(Liu et al., 2018g).杨洪全研究组发现, G蛋白β亚基AGB1通过与HY5互作抑制其DNA结合能力, 从而抑制光信号; 而蓝光信号通过CRY1与AGB1互作促进AGB1与HY5解离来抑制AGB1的功能(Lian et al., 2018); 并发现CRY1与去磷酸化形式的BL信号关键转录因子BES1发生依赖于蓝光的直接互作, 揭示了植物能根据光信号和内在BL信号有无或强弱的动态变化来优化其光形态建成(Wang et al., 2018n).此外, 该研究组还发现光受体cry1、phyB竞争性与生长素信号通路抑制子Aux/IAA结合, 并磷酸化后者而负调控生长素信号转导, 进而精确调控植物光信号与生长素信号通路的平衡, 优化其生长(Xu et al., 2018c). ...
A cytosolic triosephosphate isomerase is a key component in XA3/XA26-mediated resistance 1 2018
FLOURY SHRUNKEN ENDOSPERM1 connects phospholipid metabolism and amyloplast development in rice 1 2018
... 种子含油量是油菜的重要性状之一.郭亮研究组与相关单位合作对油菜籽中油脂合成的调控机制进行研究, 发现种子中脂质代谢相关基因在种子的不同部位及不同种子之间表达量存在显著差异.该研究对高油分油菜育种具有重要指导意义(Lu et al., 2018b).王二涛研究组则对脂肪酸在植物和菌根真菌之间的转运调控机制进行探索, 发现在与丛枝菌根真菌共生过程中, 苜蓿转录因子WRI5a是脂肪酸碳源和磷营养交换的分子开关(Jiang et al., 2018e).淀粉体中合成与储存的淀粉是谷物胚乳中的主要储能分子.万建民研究组与章文华研究组合作解析了淀粉体发育和淀粉合成的分子机制, 揭示了FSE1为磷脂酶样蛋白质, 其可调控水稻胚乳中半乳糖脂的合成, 为水稻胚乳发育过程中脂质代谢与淀粉合成之间的关系提供了新观点(Long et al., 2018). ...
Evolutionary history of the angiosperm flora of China 1 2018
... 二是植物系统进化研究取得了系列突破性成果.陈之端研究组重建了中国被子植物生命之树, 揭示了中国被子植物系统发育多样性形成的时空格局, 并明确了属、种水平分别应该重点保护的热点地区, 对我国保护区建设意义重大(Lu et al., 2018a).该成果入选2018年度中国生命科学十大进展.凌宏清、梁承志、王道文和张爱民研究组完成了乌拉尔图小麦材料G1812的基因组测序和精细组装, 绘制出小麦A基因组7条染色体的分子图谱(Ling et al., 2018).研究成果对进一步推动栽培小麦的遗传改良具有重要的理论意义和实用价值.叶凯研究组破译了罂粟基因组DNA密码, 揭示了其合成重要药用化合物的关键步骤, 这一发现为提高罂粟药用成分的产量及抗病性奠定了基础(Guo et al., 2018b).黎志康研究组与相关单位合作, 深入探讨了水稻起源、分类以及驯化规律, 揭示了亚洲栽培稻的起源和群体基因组变异结构, 剖析了水稻核心种质资源的基因组遗传多样性(Wang et al., 2018o), 为水稻规模化基因发掘和复杂性状分子改良奠定了基础. ...
Spatial analysis of lipid metabolites and expressed genes reveals tissue-specific heterogeneity of lipid metabolism in high- and low-oil Brassica napus L. seeds 1 2018
... 种子含油量是油菜的重要性状之一.郭亮研究组与相关单位合作对油菜籽中油脂合成的调控机制进行研究, 发现种子中脂质代谢相关基因在种子的不同部位及不同种子之间表达量存在显著差异.该研究对高油分油菜育种具有重要指导意义(Lu et al., 2018b).王二涛研究组则对脂肪酸在植物和菌根真菌之间的转运调控机制进行探索, 发现在与丛枝菌根真菌共生过程中, 苜蓿转录因子WRI5a是脂肪酸碳源和磷营养交换的分子开关(Jiang et al., 2018e).淀粉体中合成与储存的淀粉是谷物胚乳中的主要储能分子.万建民研究组与章文华研究组合作解析了淀粉体发育和淀粉合成的分子机制, 揭示了FSE1为磷脂酶样蛋白质, 其可调控水稻胚乳中半乳糖脂的合成, 为水稻胚乳发育过程中脂质代谢与淀粉合成之间的关系提供了新观点(Long et al., 2018). ...
The citrus transcription factor CsMADS6 modulates carotenoid metabolism by directly regulating carotenogenic genes 1 2018
... 类胡萝卜素是自然界广泛分布的一类次生代谢物.王涛涛研究组发现了番茄类胡萝卜素合成调控的关键转录因子SlBBX20, 其正调控番茄中类胡萝卜素的积累且稳定性受到泛素化调控(Xiong et al., 2019).邓秀新研究组则获得一个调控类胡萝卜素合成途径关键基因LCYb的转录因子CsMADS6, 并证明其可通过直接调控类胡萝卜素代谢基因LCYb1等的表达, 协同正调控类胡萝卜素代谢(Lu et al., 2018c).刘耀光研究组将双基因(sZmPSY1和sPaCrtI)、三基因(sZmPSY1、sPaCrtI和sCrBKT)和四基因(sZmPSY1、sPaCrtI、sCrBKT和sHpBHY)聚合转化水稻, 分别获得了筛选标记删除的富含黄色β-胡萝卜素、橙红色的角黄素和虾青素大米新种质(Zhu et al., 2018c).此外, 陈学森研究组鉴定了PA1型 MYB转录因子MdMYBPA1.其在正常条件下能促进苹果原花青素的合成, 但在低温胁迫下促进花青苷的合成(Wang et al., 2018i). ...
Barley stripe mosaic virus gammab protein subverts autophagy to promote viral infection by disrupting the ATG7-ATG8 interaction 1 2018
Resequencing a core collection of upland cotton identifies genomic variation and loci influencing fiber quality and yield 1 2018
... 比较基因组学和全基因组关联分析(GWAS)不仅能揭示栽培植物的起源和演化历史, 还可以为分子辅助育种提供重要参考.梅花(Prunus mume)是蔷薇科植物, 已有3000多年的栽培历史.张启翔研究组与相关单位合作完成了梅花全基因组重测序, 对几百株梅花品种的重要观赏性状进行了GWAS分析, 为揭示梅花的演化历史, 开展其分子标记辅助育种打下坚实的基础(Zhang et al., 2018j).此外, 杜雄明研究组与相关单位合作对419份陆地棉核心种质的基因组进行了重测序, 并结合多种农艺性状进行了GWAS分析, 为人工驯化选择导致棉花遗传多样性降低提供了直接的分子证据(Ma et al., 2018e).张献龙研究组与相关单位合作对四倍体棉花(陆地棉和海岛棉)进行了精细测序, 将陆地棉和海岛棉基因组的连续性分别提高了55倍和90倍(Wang et al., 2019); 且绘制了棉花二倍体和四倍体的三维基因组图谱, 鉴定了棉花基因组中大量的拓扑结构域(TAD), 揭示了三维基因组的进化与转录调控之间的关系(Wang et al., 2018h).该研究不仅促进了棉花功能基因组研究, 而且增加了人们对植物三维基因组结构的认知. ...
Mutations in a subfamily of abscisic acid receptor genes promote rice growth and productivity 1 2018
... 脱落酸(ABA)是控制植物生长和胁迫反应的关键激素.朱健康研究组发现水稻中ABA受体基因家族的特异受体基因突变导致胁迫反应减弱, 但在自然条件下表现出比野生型更好的生长状态(Miao et al., 2018a).该研究为培育高产优质水稻品种提供了新思路.林宏辉和张大伟研究组发现, 拟南芥转录因子基因HAT1及其同源基因负调控ABA的生物合成, 从而参与调控对干旱胁迫的响应(Tan et al., 2018b).该研究揭示了HAT1一种新的负调控机制, 对理解植物耐旱响应具有重要意义. ...
The TIE1 transcriptional repressor controls shoot branching by directly repressing BRANCHED1 in Arabidopsis 1 2018
... 植物分枝是影响植物株型的关键因素.秦根基研究组与相关单位合作对植物分枝形成的分子调控机制进行研究, 发现了1个分枝正调控因子TIE1, 它与分枝重要调控因子BRC1 (BRANCHED1)发生互作, 抑制其转录活性.通过调节HB21、HB40和HB53等BRC1下游基因的转录, 发挥促进腋芽分枝的作用(Yang et al., 2018h).开花是植物由营养生长转向生殖生长的标志.王前和张小兰研究组对热诱导开花的分子机理进行探索, 证明LsSOC1是热诱导花芽形成的激活子, 热激转录因子HsfA1e和HsfA4c通过结合在LsSOC1基因的启动子上, 影响其表达, 为开花相关机制研究奠定了理论基础(Chen et al., 2018k).果实成熟过程对于果实品质形成和种子传播非常关键.陈建业和邝健飞研究组发现转录因子MaMYB3通过抑制淀粉降解相关基因以及MabHLH6转录因子的转录水平来调控淀粉降解, 丰富了香蕉果实成熟的转录调控网络(Fan et al., 2018).朱本忠和罗云波研究组则发现, 在番茄rin突变体中, 基因组DNA片段部分缺失导致形成RIN-MC片段的嵌合, 该嵌合能够调控许多成熟相关基因的表达, 从而调节果实成熟(Li et al., 2018h).此外, 吴鸿研究组与相关单位合作阐明了拟南芥角果开裂的调控机制.他们通过对纤维素酶CEL6和半纤维素酶MAN7以及果胶酶ADPG2基因的一系列突变体果荚离层进行细胞超微结构观察, 并结合基因表达的检测结果, 证明改变这些细胞壁酶的表达量和酶活性可以调控果荚开裂区细胞分化和离层区细胞降解(He et al., 2018a).该研究为后期培育不同程度的果实抗开裂油料作物提供了技术支持. ...
EBS is a bivalent histone reader that regulates floral phase transition in Arabidopsis 1 2018
... 染色质的基本单位为核小体, 核小体核心组蛋白的N端存在多种共价修饰, 这些修饰在植物生长发育过程中不可或缺.Polycomb家族蛋白所建立的H3K27me3修饰在基因沉默过程中行使重要功能.何跃辉研究组利用酵母双杂交技术鉴定到拟南芥EMF1 (EMBRYONIC FLOWER 1)的互作蛋白SHL (SHORT LIFE)和EBS (EARLY BOLTING IN SHORT DAYS). SHL以及EBS的BAM结构域能够特异性识别H3K27me3, 影响全基因组范围H3K27me3的建立, 进而抑制基因的表达(Li et al., 2018l).同时, 杜嘉木研究组与相关单位合作证明了EBS的BAH和PHD结构域分别识别H3K27me3及H3K4me3修饰.EBS通过一种特殊的自动抑制调控方式来平衡与两种拮抗型修饰的结合, 破坏BAH-H3K27me3或PHD-H3K4me3的结合均会导致拟南芥出现早花表型(Yang et al., 2018i).该研究揭示了识别蛋白通过识别两种不同类型甲基化修饰调控植物开花的新机制.此外, 该研究组还与曹晓风研究组合作解析了拟南芥KDM5亚家族组蛋白去甲基化酶JMJ14 (JUMONJI14)以及H3K4me3复合物的晶体结构.JMJ14蛋白与底物复合物结构的解析, 为了解JMJ14在开花等生理过程中发挥作用的分子基础提供了理论依据(Yang et al., 2018j). ...
Comparative analysis of Arabidopsis ecotypes reveals a role for brassinosteroids in root hydrotropism 1 2018
... 芸薹素(BL)是植物特有的甾醇类激素.白明义研究组发现过氧化氢能氧化BL信号途径中的关键调控因子BZR1和BES1, 增强BZR1的转录活性, 从而促进植物的细胞伸长(Tian et al., 2018b).该研究揭示了H2O2与BL协同调控植物生长发育的新机制, 为探索H2O2在植物中的功能提供了新的技术途径.李云海研究组与相关单位合作, 发现BL共受体BRI1和BAK1的互作与磷酸化受糖浓度调控.相关生化分析表明, BRI1和BAK1不仅可与G蛋白亚基互作, 还可磷酸化G蛋白亚基, 为BL途径与糖信号途径协同调控植物的生长发育建立了联系(Peng et al., 2018b).许卫锋研究组与张建华研究组合作, 发现BL受体BRI1能与质子泵AHA2蛋白互作, 进而促进根系的向水性(Miao et al., 2018b). ...
HOMEOBOX PROTEIN52 mediates the crosstalk between ethylene and auxin signaling during primary root elongation by modulating auxin transport-related gene expression 1 2018
... 植物激素之间存在复杂的调控网络.杨淑华研究组发现ABA信号关键蛋白激酶SnRK2s通过磷酸化细胞分裂素负调节子A型ARR5正调控其蛋白稳定, 揭示了植物激素信号调控网络在植物逆境响应胁迫中的精密调控机制(Huang et al., 2018c).刘西岗研究组发现细胞分裂素的内稳态对于花分生组织(floral meristem, FM)的维持及分化非常重要.生长素可以通过促进ARF3 (Auxin Response Factor 3)的表达抑制细胞分裂素的活性进而促进FM的分化(Zhang et al., 2018e).该研究揭示了转录因子AG通过植物激素信号途径协同调控花分生组织干细胞稳态的机理.王小菁研究组发现非洲菊锌指蛋白GhWIP2通过抑制GA和IAA信号及激活ABA信号, 进而抑制细胞生长并最终抑制花瓣伸展(Ren et al., 2018b).该研究揭示了GA、ABA和生长素3种激素交叉互作网络共同调控非洲菊细胞伸展的作用机理.向成斌研究组发现乙烯响应基因HB52是乙烯和IAA信号互作的重要节点, 通过上调PIN2、WAG1和WAG2的表达抑制主根伸长(Miao et al., 2018c).该研究揭示了乙烯与IAA交互作用的新机制.丁兆军研究组研究表明, BL合成途径突变体det2-9中乙烯合成大幅增加并积累了大量的超氧阴离子.该研究揭示了BL、乙烯与活性氧协同调控根生长的新机制(Lv et al., 2018).ABA和BL信号通常相互拮抗.王学路研究组发现ABI1和ABI2通过与BIN2互作使其去磷酸化, 进而调控其对BES1的磷酸化(Wang et al., 2018c).同时, 该研究组还发现BL信号途径中的GSK3为控制水稻中胚轴伸长的一个关键因子, 其通过磷酸化调控一类细胞周期蛋白CYC U2的稳定性促进细胞分裂.BL以及独脚金内酯(strigolactone)信号通路协同调控GSK3介导的细胞周期进程, 进而促进中胚轴伸长(Sun et al., 2018c). ...
Structure of the Arabidopsis JMJ14-H3K4me3 complex provides insight into the substrate specificity of KDM5 subfamily histone demethylases 1 2018
... 染色质的基本单位为核小体, 核小体核心组蛋白的N端存在多种共价修饰, 这些修饰在植物生长发育过程中不可或缺.Polycomb家族蛋白所建立的H3K27me3修饰在基因沉默过程中行使重要功能.何跃辉研究组利用酵母双杂交技术鉴定到拟南芥EMF1 (EMBRYONIC FLOWER 1)的互作蛋白SHL (SHORT LIFE)和EBS (EARLY BOLTING IN SHORT DAYS). SHL以及EBS的BAM结构域能够特异性识别H3K27me3, 影响全基因组范围H3K27me3的建立, 进而抑制基因的表达(Li et al., 2018l).同时, 杜嘉木研究组与相关单位合作证明了EBS的BAH和PHD结构域分别识别H3K27me3及H3K4me3修饰.EBS通过一种特殊的自动抑制调控方式来平衡与两种拮抗型修饰的结合, 破坏BAH-H3K27me3或PHD-H3K4me3的结合均会导致拟南芥出现早花表型(Yang et al., 2018i).该研究揭示了识别蛋白通过识别两种不同类型甲基化修饰调控植物开花的新机制.此外, 该研究组还与曹晓风研究组合作解析了拟南芥KDM5亚家族组蛋白去甲基化酶JMJ14 (JUMONJI14)以及H3K4me3复合物的晶体结构.JMJ14蛋白与底物复合物结构的解析, 为了解JMJ14在开花等生理过程中发挥作用的分子基础提供了理论依据(Yang et al., 2018j). ...
Auxin production in diploid microsporocytes is necessary and sufficient for early stages of pollen development 1 2018
Divergent hydraulic strategies to cope with freezing in co-occurring temperate tree species with special reference to root and stem pressure generation 1 2018
... 冻融造成的水力传输失败是我国北方地区树木受害的重要原因之一, 其与森林生产力降低和衰退死亡等制约林业发展的重大问题密切相关.郝广友研究组以长白山阔叶红松林中的3个功能类群的典型树种为对象, 进行了树木水力结构对比研究, 发现不同功能类群树种在木质部导水率、对冻融造成的木质部功能损伤的抵抗力及对干旱诱导栓塞的抵抗力方面存在显著差异, 而不同功能类群间木质部组织水平和纹孔水平的结构特征显著分化是导致上述差异的主要原因(Yin et al., 2018b).该研究为探讨温带森林树种的环境适应性提供了新视角. ...
TGW3, a major QTL that negatively modulates grain length and weight in rice 1 2018
... 刘建祥和罗小金研究组、宋献军研究组、何予卿研究组同时克隆到与粒型和粒重相关的同一主效负调控QTL/基因qTGW3/TGW3/GL3.3, 并解析了相关分子机制(Hu et al., 2018c; Xia et al., 2018; Ying et al., 2018).李云海研究组则与相关单位合作揭示了OsMKKK10-OsMKK4-OsMAPK6信号途径正向调控水稻粒型和粒重(Xu et al., 2018e); 并发现大粒基因LARGE8可与OsMAPK6直接互作使其失活, 进而抑制颖壳细胞增殖, 负向调控水稻粒型(Xu et al., 2018f).余四斌研究组揭示了OsGBPs作为转录因子能够识别GAGA基序, 进而调控水稻粒长和植株高度(Gong et al., 2018).梁国华研究组则证明OsSPMS1基因负向调控水稻种子萌发、粒型和单株产量, 是水稻增产育种的重要靶标基因(Tao et al., 2018).刘巧泉研究组发现了水稻粒型与外观品质调控基因GS9, 其编码蛋白可与OsOFP14和OsOFP8蛋白互作, 通过改变细胞分裂调控水稻粒型(Zhao et al., 2018c).傅向东研究组克隆了1个控制水稻产量和提升稻米品质的重要基因LGY3, 该基因编码转录因子OsMADS1.可变剪接蛋白OsMADS1lgy3可提高稻米粒长, 降低垩白率, 进而影响稻米产量和外观品质(Liu et al., 2018e). ...
A selfish genetic element confers non-Mendelian inheritance in rice 1 2018
... 2018年, 植物科学的亮点成果主要体现在如下6个方面.一是水稻生物学研究取得新突破.傅向东研究组发现水稻生长调节因子GRF4与生长抑制因子DELLA相互间的反向平衡赋予了植物生长与碳-氮代谢间的稳态共调节, 揭示了提高“绿色革命”品种的氮肥利用效率并增加谷物产量的遗传机制(Li et al., 2018g).这一重大发现预示着一场新的绿色革命即将到来, 该成果入选2018年度中国科学十大进展.万建民研究组与相关单位合作, 运用自私基因模型揭示了水稻杂种不育现象, 探讨了“毒性-解毒”分子机制在水稻杂种不育中的普遍性, 揭示了自私基因在维持植物基因组的稳定性和促进新物种形成中的重要作用; 研究结果为创制“广亲和”材料, 克服杂种不育障碍, 充分利用杂种优势和野生种质资源, 提高水稻单产奠定了基础(Yu et al., 2018). ...
Knockdown of rice microRNA166 confers drought resistance by causing leaf rolling and altering stem xylem development 1 2018
... 非编码RNA是指生物体内一类不具有编码功能的RNA, 包括microRNA以及LncRNA等.它们对基因表达有调控作用, 主要通过调节mRNA的稳定性或者参与DNA甲基化等过程影响个体的生长发育.MicroRNA是一类长度为19-22 nt的非编码小RNA, 可参与非生物胁迫响应.朱健康研究组发现, 敲低microRNA166导致水稻形态发生变化, 气孔导度降低, 蒸腾速率下降, 抗旱性增强, 表明miRNA可以介导水稻的抗旱性(Zhang et al., 2018d).此外, 郑丙莲研究组揭示了植物精子携带的miRNA参与早期胚胎发育.证明了具有高丰度miR159的精细胞与中央细胞融合后, 能快速抑制中央细胞传递的miR159的靶基因MYB33/65, 进而起始胚乳的第1次核分裂(Zhao et al., 2018j).麻锦彪研究组与相关单位合作解析了SDN1的N端结构域以及催化结构域与RNA底物复合物的晶体结构, 阐明了SDN1识别剪切多种RNA底物的工作机制, 并提出了SDN1在生物体内修剪小非编码RNA的分子模型, 为设计和优化用于实验室研究的更加稳定的小非编码RNA奠定基础(Chen et al., 2018c). ...
AUXIN RESPONSE FACTOR3 regulates floral meristem determinacy by repressing cytokinin biosynthesis and signaling 1 2018
... 植物激素之间存在复杂的调控网络.杨淑华研究组发现ABA信号关键蛋白激酶SnRK2s通过磷酸化细胞分裂素负调节子A型ARR5正调控其蛋白稳定, 揭示了植物激素信号调控网络在植物逆境响应胁迫中的精密调控机制(Huang et al., 2018c).刘西岗研究组发现细胞分裂素的内稳态对于花分生组织(floral meristem, FM)的维持及分化非常重要.生长素可以通过促进ARF3 (Auxin Response Factor 3)的表达抑制细胞分裂素的活性进而促进FM的分化(Zhang et al., 2018e).该研究揭示了转录因子AG通过植物激素信号途径协同调控花分生组织干细胞稳态的机理.王小菁研究组发现非洲菊锌指蛋白GhWIP2通过抑制GA和IAA信号及激活ABA信号, 进而抑制细胞生长并最终抑制花瓣伸展(Ren et al., 2018b).该研究揭示了GA、ABA和生长素3种激素交叉互作网络共同调控非洲菊细胞伸展的作用机理.向成斌研究组发现乙烯响应基因HB52是乙烯和IAA信号互作的重要节点, 通过上调PIN2、WAG1和WAG2的表达抑制主根伸长(Miao et al., 2018c).该研究揭示了乙烯与IAA交互作用的新机制.丁兆军研究组研究表明, BL合成途径突变体det2-9中乙烯合成大幅增加并积累了大量的超氧阴离子.该研究揭示了BL、乙烯与活性氧协同调控根生长的新机制(Lv et al., 2018).ABA和BL信号通常相互拮抗.王学路研究组发现ABI1和ABI2通过与BIN2互作使其去磷酸化, 进而调控其对BES1的磷酸化(Wang et al., 2018c).同时, 该研究组还发现BL信号途径中的GSK3为控制水稻中胚轴伸长的一个关键因子, 其通过磷酸化调控一类细胞周期蛋白CYC U2的稳定性促进细胞分裂.BL以及独脚金内酯(strigolactone)信号通路协同调控GSK3介导的细胞周期进程, 进而促进中胚轴伸长(Sun et al., 2018c). ...
Unique organization of photosystem I-light-harvesting supercomplex revealed by cryo-EM from a red alga 1 2018
... 叶绿体是植物进行光合作用的细胞器, 其分裂由众多因子形成的复合物协助完成.张敏研究组发现MCD1在此过程中起重要作用.MCD1通过与ARC6作用, 识别FtsZ环, 并引导Min复合物整合到分裂环上, 从而调控叶绿体分裂(Chen et al., 2018e).欧阳敏研究组则发现ECD1蛋白通过参与叶绿体基因rps14的RNA编辑调控叶绿体早期发育(Jiang et al., 2018d).此外, 郁飞研究组和黄继荣研究组分别发现细胞质和叶绿体的翻译过程协同调控叶绿体发育, 及thf1突变体斑叶形成与质体翻译功能高度相关(Wang et al., 2018l; Huang et al., 2018b).类囊体是叶绿体基质中由单位膜围成的扁平囊, 是光合作用光反应的场所.Kang Byung-Ho研究组揭示了类囊体蛋白复合体的组装以及CURT1和FZL蛋白在类囊体生物发生过程中的关键作用(Liang et al., 2018d).隋森芳研究组与相关单位合作解析了红藻光系统I核心与捕光天线复合物(PSI-LHCR)的3.63?分辨率的三维结构, 发现红藻PSI核心既有蓝藻又有高等植物的特征(Pi et al., 2018).该研究对阐明PSI的进化和功能具有重要意义. ...
Sweet sorghum originated through selection of dry, a plant-specific NAC transcription factor gene 1 2018
... 早期陆生植物形态结构相对简单, 但环境适应能力强.特有基因对物种适应特殊环境发挥了重要作用.黄锦岭研究组对小立碗藓的hemerythrin基因进行了研究, 发现其起源于真菌向早期陆生植物的水平基因转移事件.该基因在小立碗藓中通过改变移码突变, 演化出1个双编码基因YAN/AltYAN, 该基因介导了小立碗藓的脱水耐受性(Guan et al., 2018).该研究对揭示陆生植物如何适应特殊环境提供了新观点及实验证据.基因组学研究为揭示一些物种特异现象提供了很好的切入点.例如, 卷柏(Selaginella tamariscina)是古老的复苏植物, 干旱时能存活数十年, 遇水而荣, 与已测序的其同属植物江南卷柏(S. moellendorffii) (干旱敏感型)截然不同.陈士林和宋经元研究组破译了卷柏基因组, 绘制了高杂合度卷柏的基因组图谱, 并与已发表的江南卷柏基因组进行了比较, 揭示了卷柏耐旱和复苏机制(Xu et al., 2018g).维管系统(木质部等)是植物在水生到陆生的进化过程中产生的.拟南芥和小立碗藓中特定的NAC转录因子调控木质部发育以及水分运输.景海春研究组与才宏伟研究组合作从甜高粱中鉴定到1个在驯化过程中受到选择、控制茎秆持汁性的Dry基因(编码一个NAC转录因子), 并发现其功能缺失是甜高粱茎秆富含汁液的重要原因(Zhang et al., 2018f).该研究不仅对促进作物茎秆持汁表型的遗传改良具重要作用, 而且为作物育种获得籽粒及秸秆“双丰收”提供了可能. ...
A retrotransposon in an HKT1 family sodium transporter causes variation of leaf Na( +) exclusion and salt tolerance in maize 1 2018
... 土壤盐碱化极大地限制了粮食作物的稳产增收.如何将盐碱地变为高产粮田, 成为当下农业科学家研究的热点方向, 而解析植物应答盐碱胁迫的分子机制是提高植物抗盐碱的有效手段.朱健康研究组发现, 细胞壁LRX (Leucine-rich repeat extensins)蛋白家族多突变体植株表现出对盐胁迫非常敏感的表型.进一步研究发现, LRX蛋白与RALF多肽以及细胞膜受体类激酶FERONIA形成一个元件调控植物生长和耐盐性(Zhao et al., 2018b).该研究为培育抗逆高产作物指明了方向.此外, 朱健康和王镇研究组还探明了ABA对植物抗旱性的影响, 揭示了开花抑制子SVP (Short vegetative phase)通过调控ABA代谢提高拟南芥的抗旱性机制(Wang et al., 2018r).植物类受体激酶能感受外界环境胁迫信号, 并通过胞内激酶域激活下游效应子活性和调控细胞相关生理生化过程, 从而降低或消除逆境胁迫的影响.刘选明和林建中研究组破译了1个能降低土地盐碱化对水稻产量影响的类受体胞质激酶1 (salt tolerance receptor-like cytoplasmic kinase 1, STRK1), 其可通过磷酸化和激活CatC调节H2O2体内平衡来改善盐以及氧化胁迫的耐受性(Zhou et al., 2018e).该研究为进一步解析植物耐盐的分子机制奠定了基础.范六民研究组报道了MYB亚家族转录因子RSM1 (Radialis-like sant/myb 1)蛋白与HY5/HYH蛋白互作, 并作用于ABI5启动子区, 调控ABA和非生物逆境(盐)胁迫相关基因的应答(Yang et al., 2018a).黄俊丽研究组则对水稻MADS-box家族转录因子OsMADS25蛋白进行了研究, 发现其可与谷胱甘肽S-转移酶基因OsGST4的启动子直接结合, 通过ABA介导的调控通路和ROS清除共同调控水稻的根系生长和耐盐性(Xu et al., 2018d).蒋才富研究组利用QTL克隆了调控叶片Na+含量的主效基因ZmNC1 (Na+ content 1).该基因编码1个Na+选择性离子转运蛋白ZmHKT1, 其可通过抑制Na+由根向地上部运输提高玉米的耐盐能力(Zhang et al., 2018g).郝玉金研究组则发现苹果葡萄糖感知因子己糖激酶MdHXK1可磷酸化Na+/H+交换蛋白MdNHX1, 以提高其蛋白稳定性, 进而增强苹果的耐盐性(Sun et al., 2018b). ...
A core regulatory pathway controlling rice tiller angle mediated by the LAZY1-dependent asymmetric distribution of auxin 0 2018
A single-nucleotide polymorphism in the promoter of a hairpin RNA contributes to Alternaria alternata leaf spot resistance in apple( Malus x domestica) 2 2018
... 水稻株型与产量密切相关.叶片作为水稻理想株型的组成要素之一, 其倾角对产量影响较大.薛红卫研究组新克隆了叶倾角调控基因LC3, 该基因编码1个含有SPOC结构域的转录抑制子, 其可与HIT锌指结构域蛋白LIP1互作, 通过调控生长素平衡和信号转导来调控植株叶倾角(Chen et al., 2018h).李建雄研究组与田志宏研究组合作揭示了OsOFP19、OSH1与DLT蛋白可形成复合体, 调控BL信号转导及影响叶倾角发育(Yang et al., 2018b).易可可研究组则解析了磷素有效性调控水稻叶倾角的分子机制.他们发现磷饥饿诱导的SPX1 (Syg1/Pho81/XPR1)与SPX2蛋白负调控叶倾角.SPX1的互作蛋白RLI1 (Regulator of leaf inclination 1)正调控叶倾角.RLI1可直接激活下游BU1 (Brassinosteroid upregulated 1)和BC1 (BU1-like1 complex 1)基因的表达以控制叶枕细胞的伸长, 从而使叶倾角增大(Ruan et al., 2018).种康和徐云远研究组证实miR396d通过分别抑制其靶基因OsGRF4和OsGRF6的转录, 促进BL及抑制GA信号途径, 进而调控水稻叶夹角和株高(Tang et al., 2018).邢永忠研究组则阐释了来自世界各地的水稻核心种质剑叶夹角的遗传基础, 发现bHLH转录因子家族中OsbHLH153和OsbHLH17能快速响应BL和IAA信号转导, 从而影响植株形态(Dong et al., 2018c).王永红研究组揭示了水稻分蘖角度调控的分子机制.他们通过研究水稻茎重力反应过程中的动态转录组变化, 探索了一条水稻分蘖角度调控的核心途径(Zhang et al., 2018h).该研究为解析水稻分蘖角度的调控网络进而挖掘有利用价值的基因提供了重要信息.谭禄宾研究组发现亚洲栽培稻PROG1基因相邻位置约有110 kb的染色体缺失.该缺失片段包含7个与PROG1基因相似的锌指基因, 其缺失推动了水稻由匍匐向直立转变, 最终实现产量大幅提高(Wu et al., 2018c). ...
Core cis-element variation confers subgenome-biased expression of a transcription factor that functions in cotton fiber elongation 1 2018
... 茸毛作为植物体最外层结构, 具有很好的抗虫防病毒作用.叶志彪和杨长宪研究组分离了控制番茄腺体毛形成的关键基因H (Hair), 该基因编码C2H2锌指蛋白, 可能是茄科植物多细胞腺体毛形成的关键保守基因(Chang et al., 2018).唐克轩研究组发现R2R3 MYB第9亚家族的转录因子AaMIXTA1参与调控青蒿分泌型腺毛发育, 为研究其它植物多细胞腺毛奠定了基础(Shi et al., 2018c).此外, 他们还发现HD-ZIP IV转录因子AaHD8与AaMIXTA1相互作用形成复合体, 协同调控AaHD1和多个蜡质角质合成酶基因的表达, 从而促进蜡质合成和腺毛的起始发育(Yan et al., 2018d).陈晓亚研究组与相关单位合作对可纺纤维形成的分子机制进行研究, 发现编码bHLH转录因子的棉花PRE1是可纺纤维进化的目标基因, 其正调控纤维伸长.PRE1基因启动子的变异可促进棉花可纺纤维的形成(Zhao et al., 2018a).李学宝研究组鉴定了富含半胱氨酸的GhXLIM6蛋白, 其可与GhKNL1启动子的PAL-box结合而抑制该基因表达, 促进纤维素合成, 进而调控棉纤维伸长(Li et al., 2018i).俞嘉宁研究组则揭示了棉花线粒体Ghatp1基因C1292和C1415位点发生RNA编辑可影响ATPase产生ATP, 从而促进表皮毛及纤维伸长(He et al., 2018c).根毛是一种顶端生长的植物细胞, 微丝骨架对根毛的生长发育有极其重要的调控作用.贾红磊研究组揭示了H2S通过硫巯基化修饰扰乱正常的微丝骨架聚合及生长模式, 从而影响植物根毛的生长, 为进一步探索H2S调控植物生长发育的作用机制提供了理论依据(Li et al., 2018d). ...
Leucine-rich repeat extensin proteins regulate plant salt tolerance in Arabidopsis 1 2018
... 土壤盐碱化极大地限制了粮食作物的稳产增收.如何将盐碱地变为高产粮田, 成为当下农业科学家研究的热点方向, 而解析植物应答盐碱胁迫的分子机制是提高植物抗盐碱的有效手段.朱健康研究组发现, 细胞壁LRX (Leucine-rich repeat extensins)蛋白家族多突变体植株表现出对盐胁迫非常敏感的表型.进一步研究发现, LRX蛋白与RALF多肽以及细胞膜受体类激酶FERONIA形成一个元件调控植物生长和耐盐性(Zhao et al., 2018b).该研究为培育抗逆高产作物指明了方向.此外, 朱健康和王镇研究组还探明了ABA对植物抗旱性的影响, 揭示了开花抑制子SVP (Short vegetative phase)通过调控ABA代谢提高拟南芥的抗旱性机制(Wang et al., 2018r).植物类受体激酶能感受外界环境胁迫信号, 并通过胞内激酶域激活下游效应子活性和调控细胞相关生理生化过程, 从而降低或消除逆境胁迫的影响.刘选明和林建中研究组破译了1个能降低土地盐碱化对水稻产量影响的类受体胞质激酶1 (salt tolerance receptor-like cytoplasmic kinase 1, STRK1), 其可通过磷酸化和激活CatC调节H2O2体内平衡来改善盐以及氧化胁迫的耐受性(Zhou et al., 2018e).该研究为进一步解析植物耐盐的分子机制奠定了基础.范六民研究组报道了MYB亚家族转录因子RSM1 (Radialis-like sant/myb 1)蛋白与HY5/HYH蛋白互作, 并作用于ABI5启动子区, 调控ABA和非生物逆境(盐)胁迫相关基因的应答(Yang et al., 2018a).黄俊丽研究组则对水稻MADS-box家族转录因子OsMADS25蛋白进行了研究, 发现其可与谷胱甘肽S-转移酶基因OsGST4的启动子直接结合, 通过ABA介导的调控通路和ROS清除共同调控水稻的根系生长和耐盐性(Xu et al., 2018d).蒋才富研究组利用QTL克隆了调控叶片Na+含量的主效基因ZmNC1 (Na+ content 1).该基因编码1个Na+选择性离子转运蛋白ZmHKT1, 其可通过抑制Na+由根向地上部运输提高玉米的耐盐能力(Zhang et al., 2018g).郝玉金研究组则发现苹果葡萄糖感知因子己糖激酶MdHXK1可磷酸化Na+/H+交换蛋白MdNHX1, 以提高其蛋白稳定性, 进而增强苹果的耐盐性(Sun et al., 2018b). ...
GS9 acts as a transcriptional activator to regulate rice grain shape and appearance quality 1 2018
... 刘建祥和罗小金研究组、宋献军研究组、何予卿研究组同时克隆到与粒型和粒重相关的同一主效负调控QTL/基因qTGW3/TGW3/GL3.3, 并解析了相关分子机制(Hu et al., 2018c; Xia et al., 2018; Ying et al., 2018).李云海研究组则与相关单位合作揭示了OsMKKK10-OsMKK4-OsMAPK6信号途径正向调控水稻粒型和粒重(Xu et al., 2018e); 并发现大粒基因LARGE8可与OsMAPK6直接互作使其失活, 进而抑制颖壳细胞增殖, 负向调控水稻粒型(Xu et al., 2018f).余四斌研究组揭示了OsGBPs作为转录因子能够识别GAGA基序, 进而调控水稻粒长和植株高度(Gong et al., 2018).梁国华研究组则证明OsSPMS1基因负向调控水稻种子萌发、粒型和单株产量, 是水稻增产育种的重要靶标基因(Tao et al., 2018).刘巧泉研究组发现了水稻粒型与外观品质调控基因GS9, 其编码蛋白可与OsOFP14和OsOFP8蛋白互作, 通过改变细胞分裂调控水稻粒型(Zhao et al., 2018c).傅向东研究组克隆了1个控制水稻产量和提升稻米品质的重要基因LGY3, 该基因编码转录因子OsMADS1.可变剪接蛋白OsMADS1lgy3可提高稻米粒长, 降低垩白率, 进而影响稻米产量和外观品质(Liu et al., 2018e). ...
Natural variation in GmGBP1 promoter affects photoperiod control of flowering time and maturity in soybean 1 2018
... 大豆属于短日照植物, 对光周期极为敏感.李文滨研究组发现, 大豆中SKIP同源蛋白GmGBP1参与光周期介导的开花途径, 其启动子区域的自然变异调控大豆在不同环境中更早开花和成熟(Zhao et al., 2018d).此外, 韩天富研究组发现, 大豆FT基因家族成员GmFT1a能够延迟大豆开花和成熟, 与开花促进基因GmFT2a/GmFT5a相互拮抗.基于本研究及前人的研究成果, 他们提出了FT基因家族精细调控大豆开花时间的跷跷板模型(Liu et al., 2018f).生物钟是生物体适应环境昼夜周期变化而进化出的协调细胞内基因表达、代谢网络调控的分子系统.吴昌银研究组前期发现, OsELF3作为一个生物钟基因, 其昼夜表达相位是影响长日照条件下水稻抽穗期的关键因素.近期他们发现泛素化连接酶HAF1与OsELF3.1互作, 从而调控其蛋白昼夜节律性积累, 且单个氨基酸(L558S)变异影响二者的互作, 从而影响水稻的区域适应性(Zhu et al., 2018a).王雷研究组则发现生物钟核心组分Evening Complex各组分的突变体叶片提前衰老.进一步研究发现, Evening Complex直接结合MYC2基因启动子并抑制其表达, 从而在时间维度精细调控茉莉酸诱导的植物叶片衰老进程(Zhang et al., 2018l). ...
Pan-genome analysis highlights the extent of genomic variation in cultivated and wild rice 1 2018
... 栽培稻和普通野生稻(Oryza rufipogon)丰富的遗传多样性是水稻育种的主要来源.黄学辉研究组对66个来自不同水稻类群的栽培稻品种和野生稻株系的精细基因组进行了比较分析, 鉴定出水稻基因组中各类复杂的遗传变异, 并发现很多功能基因存在多种等位基因类型(Zhao et al., 2018e).发达的根系是旱粳稻和热带粳稻适应干旱环境最典型的根系形态特征.李自超研究组与黎志康研究组合作, 将795份栽培稻种质资源的根系性状和330多万个高质量的SNP数据进行关联分析, 获得几十个候选基因; 对这些候选基因的序列进行分析, 发现几乎所有位点在野生稻中都以低频存在.因此, 与旱粳稻长根和粗根等性状相关的基因是在高山干旱环境中通过正向选择及聚合而固定下来的(Zhao et al., 2018i).此外, 陈明生研究组对约1 500万年前分化的短花药野生稻(Oryza brachyantha)和栽培稻着丝粒周围序列进行了比较, 发现该区域倾向有更多的共线基因丢失(大多转移到其它区域).进一步分析表明, 这种基因丢失可能受着丝粒染色质动态变化(如扩展或微易位)驱动(Liao et al., 2018). ...
The OsRR24/LEPTO1 type-B response regulator is essential for the organization of leptotene chromosomes in rice meiosis 1 2018
... 减数分裂是真核生物有性生殖所必需的环节.程祝宽和李亚非研究组对水稻减数分裂时胞质分裂的遗传调控机制进行了探索.鉴定出1个调控花粉母细胞胞质分裂的蛋白DCM1 (Defective Callose in Meiosis 1).该蛋白的C端含有5个串联的CCCH锌指结构域, 并与核多聚腺苷酸结合蛋白OsPABNs存在互作.dcm1突变体的花粉母细胞在胞质分裂时, 由于胼胝质板提前降解, 胞质分裂无法进行, 使得减数第二次分裂的纺锤体取向异常, 并最终表现为雄性不育(Zhang et al., 2018a).此外, 程祝宽研究组在水稻中成功分离了1个新的减数分裂交叉结形成相关蛋白HEIP1 (HEI10 Interaction Protein 1).实验证明该蛋白与HEI10、MSH5和ZIP4等交叉结形成促进因子相互作用并共定位, 调控减数分裂一类交叉的形成(Li et al., 2018j).该研究为深入揭示减数分裂同源重组形成的分子机制奠定了基础.该研究组还与马伯军研究组合作通过筛选水稻减数分裂缺陷的不育突变体, 发现LEPTO1 (LEPTOTENE1)参与水稻减数分裂细线期染色体的形态建成.lepto1突变体的花粉母细胞染色体停滞在前细线期状态, 不能进一步组装进入典型的细线期染色体状态.性母细胞中没有DSB形成, 也没有减数分裂特异蛋白的组装, 观察不到胼胝质的积累.LEPTO1编码OsRR24, 属于B类响应调节因子, N端具有保守的DDK和MYB结构域, C端具有转录激活活性, 表明LEPTO1可能通过调控相关基因的表达, 进而调节减数分裂细线期染色体的形态建成(Zhao et al., 2018f).王应祥研究组与贺岩研究组合作探究了减数分裂重组过程中DNA双链断裂(double- strand breaks, DSB)的数目对重组稳态和最终分布的影响.他们通过转基因技术在DSB缺失突变体(spo11-/-)背景下操控DSB发生的数目, 揭示了减数分裂中DSB形成的数目对重组的稳态及分布有重要调控作用(Xue et al., 2018a). ...
COP1 SUPPRESSOR 4 promotes seedling photomorphogenesis by repressing CCA1 and PIF4 expression in Arabidopsis 1 2018
... 邓兴旺研究组与许冬清研究组合作鉴定了光形态建成的正调节因子CSU4, 它能整合生物钟节律和光信号(Zhao et al., 2018g).他们还发现BBX21通过第2个B-box结构域与HY5启动子中的T/G-box结合, 调控HY5及其靶基因的表达以促进植物光形态建成(Xu et al., 2018b).而BBX28通过与HY5蛋白的C末端互作, 抑制HY5与下游靶基因启动子结合, 从而抑制HY5活性及其下游靶基因表达, 最终抑制植物光形态建成(Lin et al., 2018a).此外, 邓兴旺和钟上威研究组揭示了PIFs、HY5和EIN3及EIL1对出土幼苗形态建成的调控作用, 为阐明幼苗从暗形态建成转向光形态建成的分子机制提供了新证据(Shi et al., 2018b).吕应堂研究组则鉴定了光形态建成的正调控因子SRS5, 它直接结合HY5、BBX21和BBX22的启动子并激活其表达, 从而促进光形态建成(Yuan et al., 2018).刘宏涛研究组发现UV-B照射后, UVR8形成单体并在细胞核中富集, UVR8通过抑制BES1/BIM1结合DNA的能力从而抑制下游生长相关基因的表达, 并抑制细胞伸长及光形态建成(Liang et al., 2018b).同时, 发现UV-B激活的UVR8形成单体, 进入细胞核直接结合WRKY36而抑制其结合HY5启动子, 进而抑制下胚轴伸长(Yang et al., 2018g). ...
Global identification of Arabidopsis lncRNAs reveals the regulation of MAF4 by a natural antisense RNA 1 2018
... LncRNA是生物体中一类长度大于200 bp的非编码RNA, 其通过多种方式参与基因表达调控等重要生命活动.戚益军和李景睿研究组系统鉴定并分析了拟南芥中大量lncRNA, 发现可以调节开花时间的NAT-lncRNA, 并阐明了其正向调控正义链基因转录的作用机制(Zhao et al., 2018h).杨金水和苏伟研究组则在水稻中鉴定到一例与产量性状相关且起转录激活作用的lncRNA, 过表达lncRNA会改变LRK1基因组位点的组蛋白修饰状态, 使水稻增产(Wang et al., 2018p). ...
Loci and natural alleles underlying robust roots and adaptive domestication of upland ecotype rice in aerobic conditions 1 2018
... 栽培稻和普通野生稻(Oryza rufipogon)丰富的遗传多样性是水稻育种的主要来源.黄学辉研究组对66个来自不同水稻类群的栽培稻品种和野生稻株系的精细基因组进行了比较分析, 鉴定出水稻基因组中各类复杂的遗传变异, 并发现很多功能基因存在多种等位基因类型(Zhao et al., 2018e).发达的根系是旱粳稻和热带粳稻适应干旱环境最典型的根系形态特征.李自超研究组与黎志康研究组合作, 将795份栽培稻种质资源的根系性状和330多万个高质量的SNP数据进行关联分析, 获得几十个候选基因; 对这些候选基因的序列进行分析, 发现几乎所有位点在野生稻中都以低频存在.因此, 与旱粳稻长根和粗根等性状相关的基因是在高山干旱环境中通过正向选择及聚合而固定下来的(Zhao et al., 2018i).此外, 陈明生研究组对约1 500万年前分化的短花药野生稻(Oryza brachyantha)和栽培稻着丝粒周围序列进行了比较, 发现该区域倾向有更多的共线基因丢失(大多转移到其它区域).进一步分析表明, 这种基因丢失可能受着丝粒染色质动态变化(如扩展或微易位)驱动(Liao et al., 2018). ...
Clearance of maternal barriers by paternal miR159 to initiate endosperm nuclear division in Arabidopsis 1 2018
... 非编码RNA是指生物体内一类不具有编码功能的RNA, 包括microRNA以及LncRNA等.它们对基因表达有调控作用, 主要通过调节mRNA的稳定性或者参与DNA甲基化等过程影响个体的生长发育.MicroRNA是一类长度为19-22 nt的非编码小RNA, 可参与非生物胁迫响应.朱健康研究组发现, 敲低microRNA166导致水稻形态发生变化, 气孔导度降低, 蒸腾速率下降, 抗旱性增强, 表明miRNA可以介导水稻的抗旱性(Zhang et al., 2018d).此外, 郑丙莲研究组揭示了植物精子携带的miRNA参与早期胚胎发育.证明了具有高丰度miR159的精细胞与中央细胞融合后, 能快速抑制中央细胞传递的miR159的靶基因MYB33/65, 进而起始胚乳的第1次核分裂(Zhao et al., 2018j).麻锦彪研究组与相关单位合作解析了SDN1的N端结构域以及催化结构域与RNA底物复合物的晶体结构, 阐明了SDN1识别剪切多种RNA底物的工作机制, 并提出了SDN1在生物体内修剪小非编码RNA的分子模型, 为设计和优化用于实验室研究的更加稳定的小非编码RNA奠定基础(Chen et al., 2018c). ...
MLK1 and MLK2 coordinate RGA and CCA1 activity to regulate hypocotyl elongation in Arabidopsis thaliana 1 2018
... 丁勇研究组则对拟南芥种子下胚轴发育进行研究, 发现参与拟南芥组蛋白修饰的MLK1和MLK2、GA信号途径负调控因子RGA, 以及与昼夜节律相关的CCA1形成蛋白复合体, 该复合体直接调控下游基因DWF4的表达, 进而影响下胚轴伸长(Zheng et al., 2018a).李云海研究组以拟南芥为材料对控制植物器官大小的分子机制进行探索.他们发现E3连接酶F-盒组分SAP蛋白的2个底物KIX8和KIX9与PPD形成复合物行使功能, 其通过调控拟分生组织细胞增殖来控制器官大小(Li et al., 2018f).玉米籽粒性状直接影响其产量.汤继华研究组与李文学研究组合作发现了1个控制玉米籽粒发育性状的基因ZmUrb2, 该基因主要通过影响核糖体的生物合成和核糖体rRNA前体加工来影响籽粒发育和整个营养生长过程(Wang et al., 2018d). ...
Danger-associated peptides close stomata by OST1-indepen-dent activation of anion channels in guard cells 1 2018
MdWRKY9 overexpression confers intensive dwarfing in the M26 rootstock of apple by directly inhibiting brassinosteroid synthetase MdDWF4 expression 1 2018
... 在苹果育种中, 矮化砧的选育具有重要意义.孔瑾研究组发现WRKY9在苹果矮化砧中高表达, 并直接抑制BL合成酶DWF4的转录从而减少BL合成, 介导植株矮化(Zheng et al., 2018c).该研究揭示了在木本植物中WRKY9通过调控BL水平进而促进矮化的可能机制. ...
Species composition, functional and phylogenetic distances correlate with success of invasive Chromolaena odorata in an experimental test 1 2018
... 李继刚和康定明研究组则筛选获得了远红光信号转导新组分TZP.TZP可与phyA和FHY1互作, 且在远红光下调控phyA、FHY1和HY5的蛋白水平, 并参与调控phyA的蛋白磷酸化(Zhang et al., 2018k).李继刚研究组还与邓兴旺研究组合作证明了拟南芥phyA铰链区的3个位点(S590、T593和S602)对phyA行使功能非常重要(Zhou et al., 2018c).吴素幸研究组发现, phyA和cry感知远红光和蓝光信号, 使负调控因子COP1失活, 从而激活生长素途径在TOR依赖的RPS6磷酸化中的作用(Chen et al., 2018a).此外, 李琳研究组发现在遮阴, 尤其是严重遮阴条件下, phyA蛋白量增加, phyA与Aux/IAA互作影响了生长素受体TIR与Aux/IAA的互作, 从而阻止TIR介导的Aux/IAA降解, 导致生长素信号减弱, 通过调控光信号与生长素信号的平衡, 进而调节植物的生长发育(Yang et al., 2018c).周艳虹研究组鉴定了番茄在光抑制和光保护过程中的光信号依赖性调控通路, 分析了phyA依赖的HY5-ABI5-RBOH1信号通路在缓解冷诱导的光抑制以及光保护中的作用(Wang et al., 2018a). ...
TCP transcription factors regulate shade avoidance via directly mediating the expression of both PHYTOCHROME INTERACTING FACTORs and auxin biosynthetic genes 1 2018
... 光照和温度是影响植物生长发育非常重要的两个环境因素.林荣呈研究组发现了光信号与温度信号整合的新因子SEU, 揭示了SEU通过与PIF4互作形成转录调控复合物, 对生长素合成及与信号相关的靶基因进行调节, 从而实现对生长发育的调控(Huai et al., 2018).方玉达研究组发现, 拟南芥miRNA加工复合体成员DCL1/HYL1与PIF4存在直接互作.PIF4在黑暗/红光转换过程中调控DCL1的稳定性及部分miRNA基因的转录, 进而调控光形态建成(Sun et al., 2018e).黎家研究组则发现TCP17蛋白通过调控PIF4/PIF5依赖以及非依赖的生长素合成, 从而调控遮阴诱导下植物下胚轴的伸长生长(Zhou et al., 2018d). ...
The receptor-like cytoplasmic kinase STRK1 phosphorylates and activates CatC, thereby regulating H2O2 homeostasis and improving salt tolerance in rice 0 2018
The E3 ubiquitin ligase HAF1 modulates circadian accumulation of EARLY FLOWERING3 to control heading date in rice under long- day conditions 1 2018
... 土壤盐碱化极大地限制了粮食作物的稳产增收.如何将盐碱地变为高产粮田, 成为当下农业科学家研究的热点方向, 而解析植物应答盐碱胁迫的分子机制是提高植物抗盐碱的有效手段.朱健康研究组发现, 细胞壁LRX (Leucine-rich repeat extensins)蛋白家族多突变体植株表现出对盐胁迫非常敏感的表型.进一步研究发现, LRX蛋白与RALF多肽以及细胞膜受体类激酶FERONIA形成一个元件调控植物生长和耐盐性(Zhao et al., 2018b).该研究为培育抗逆高产作物指明了方向.此外, 朱健康和王镇研究组还探明了ABA对植物抗旱性的影响, 揭示了开花抑制子SVP (Short vegetative phase)通过调控ABA代谢提高拟南芥的抗旱性机制(Wang et al., 2018r).植物类受体激酶能感受外界环境胁迫信号, 并通过胞内激酶域激活下游效应子活性和调控细胞相关生理生化过程, 从而降低或消除逆境胁迫的影响.刘选明和林建中研究组破译了1个能降低土地盐碱化对水稻产量影响的类受体胞质激酶1 (salt tolerance receptor-like cytoplasmic kinase 1, STRK1), 其可通过磷酸化和激活CatC调节H2O2体内平衡来改善盐以及氧化胁迫的耐受性(Zhou et al., 2018e).该研究为进一步解析植物耐盐的分子机制奠定了基础.范六民研究组报道了MYB亚家族转录因子RSM1 (Radialis-like sant/myb 1)蛋白与HY5/HYH蛋白互作, 并作用于ABI5启动子区, 调控ABA和非生物逆境(盐)胁迫相关基因的应答(Yang et al., 2018a).黄俊丽研究组则对水稻MADS-box家族转录因子OsMADS25蛋白进行了研究, 发现其可与谷胱甘肽S-转移酶基因OsGST4的启动子直接结合, 通过ABA介导的调控通路和ROS清除共同调控水稻的根系生长和耐盐性(Xu et al., 2018d).蒋才富研究组利用QTL克隆了调控叶片Na+含量的主效基因ZmNC1 (Na+ content 1).该基因编码1个Na+选择性离子转运蛋白ZmHKT1, 其可通过抑制Na+由根向地上部运输提高玉米的耐盐能力(Zhang et al., 2018g).郝玉金研究组则发现苹果葡萄糖感知因子己糖激酶MdHXK1可磷酸化Na+/H+交换蛋白MdNHX1, 以提高其蛋白稳定性, 进而增强苹果的耐盐性(Sun et al., 2018b). ...
Rewiring of the fruit metabolome in tomato breeding 1 2018
... 大豆属于短日照植物, 对光周期极为敏感.李文滨研究组发现, 大豆中SKIP同源蛋白GmGBP1参与光周期介导的开花途径, 其启动子区域的自然变异调控大豆在不同环境中更早开花和成熟(Zhao et al., 2018d).此外, 韩天富研究组发现, 大豆FT基因家族成员GmFT1a能够延迟大豆开花和成熟, 与开花促进基因GmFT2a/GmFT5a相互拮抗.基于本研究及前人的研究成果, 他们提出了FT基因家族精细调控大豆开花时间的跷跷板模型(Liu et al., 2018f).生物钟是生物体适应环境昼夜周期变化而进化出的协调细胞内基因表达、代谢网络调控的分子系统.吴昌银研究组前期发现, OsELF3作为一个生物钟基因, 其昼夜表达相位是影响长日照条件下水稻抽穗期的关键因素.近期他们发现泛素化连接酶HAF1与OsELF3.1互作, 从而调控其蛋白昼夜节律性积累, 且单个氨基酸(L558S)变异影响二者的互作, 从而影响水稻的区域适应性(Zhu et al., 2018a).王雷研究组则发现生物钟核心组分Evening Complex各组分的突变体叶片提前衰老.进一步研究发现, Evening Complex直接结合MYC2基因启动子并抑制其表达, 从而在时间维度精细调控茉莉酸诱导的植物叶片衰老进程(Zhang et al., 2018l). ...
From golden rice to aSTARice: bioengineering astaxanthin biosynthesis in rice endosperm 2 2018
... 五是植物代谢组学研究取得重要进展.黄三文研究组与罗杰研究组合作, 利用多重组学大数据, 揭示了驯化和育种过程中番茄果实营养及风味物质的变化规律, 发现了调控这些物质的重要遗传位点(Zhu et al., 2018b).这一成果不仅为番茄果实风味和营养物质遗传调控与全基因组设计育种提供了路线图, 而且为其它植物次生代谢资源的开发利用提供了重要思路. ...
... 类胡萝卜素是自然界广泛分布的一类次生代谢物.王涛涛研究组发现了番茄类胡萝卜素合成调控的关键转录因子SlBBX20, 其正调控番茄中类胡萝卜素的积累且稳定性受到泛素化调控(Xiong et al., 2019).邓秀新研究组则获得一个调控类胡萝卜素合成途径关键基因LCYb的转录因子CsMADS6, 并证明其可通过直接调控类胡萝卜素代谢基因LCYb1等的表达, 协同正调控类胡萝卜素代谢(Lu et al., 2018c).刘耀光研究组将双基因(sZmPSY1和sPaCrtI)、三基因(sZmPSY1、sPaCrtI和sCrBKT)和四基因(sZmPSY1、sPaCrtI、sCrBKT和sHpBHY)聚合转化水稻, 分别获得了筛选标记删除的富含黄色β-胡萝卜素、橙红色的角黄素和虾青素大米新种质(Zhu et al., 2018c).此外, 陈学森研究组鉴定了PA1型 MYB转录因子MdMYBPA1.其在正常条件下能促进苹果原花青素的合成, 但在低温胁迫下促进花青苷的合成(Wang et al., 2018i). ...
The NB-LRR gene Pm60 confers powdery mildew resistance in wheat 0 2018
Transcriptional regulation of the immune receptor FLS2 controls the ontogeny of plant innate immunity 1 2018
 ,2,*, 王台
,2,*, 王台
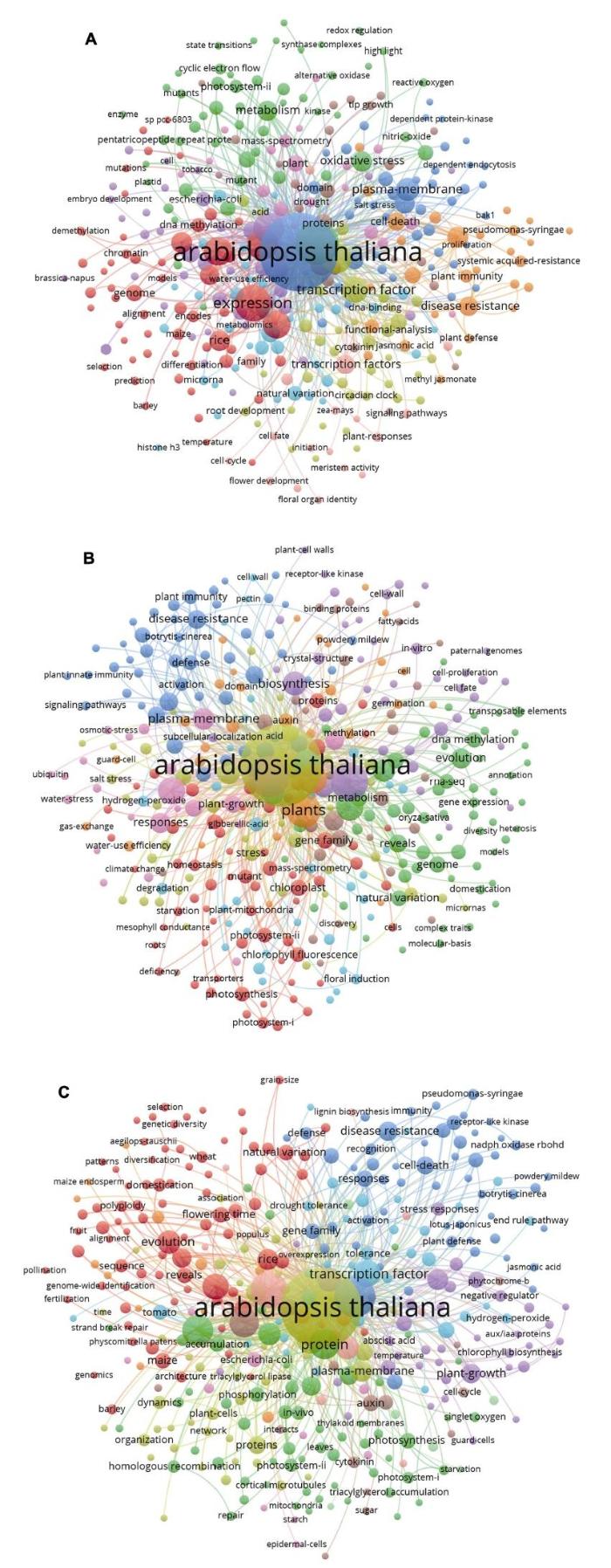 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}