Abstract Under field conditions, biotic and abiotic stresses usually occur simultaneously, and threaten global food security. Uncovering the mechanisms underlying plant response to combinations of two or more stress conditions holds the potential to breed new crop varieties with enhanced stress tolerance. Recent studies have revealed that the response of plants to stress combinations is unique and cannot be directly extrapolated from the response of plants to each of the different stresses. The responses of plants to different combined stresses might integrate with different signaling pathways at multiple levels, including defence responses, transcription factors, hormone signaling and osmolyte biosynthesis. Here, we review the molecular and physiological responses and adaptations of plants to different stress combinations, and provide an update on multi-omics approaches to study combined stresses. Keywords:stress combinations;environmental stress;plant growth and development;yield;omics
PDF (1159KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 郭倩倩, 周文彬. 植物响应联合胁迫机制的研究进展. 植物学报, 2019, 54(5): 662-673 doi:10.11983/CBB19100 Guo Qianqian, Zhou Wenbin. Advances in the Mechanism Underlying Plant Response to Stress Combination. Chinese Bulletin of Botany, 2019, 54(5): 662-673 doi:10.11983/CBB19100
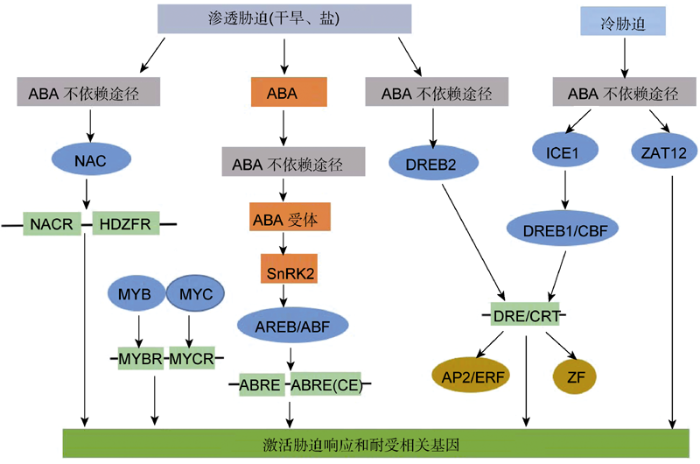
Figure 1Cross-talk network in ABA-dependent and ABA-independent pathways during abiotic stress (drought, salinity and low temperature) (modified from Roychoudhury et al., 2013)
Table 1 表1 表1联合胁迫之间的相互作用 Table 1Interactions of combined stresses
AchuoaEA, PrinsenbE, H?fteM (2006). Influence of drought, salt stress and abscisic acid on the resistance of tomato to Botrytis cinerea and Oidium neolycopersici Plant Pathol 55, 178-186. [本文引用: 1]
AhmedIM, DaiHX, ZhengWT, CaoFB, ZhangGP, SunDF, WuFB (2013). Genotypic differences in physiological characteristics in the tolerance to drought and salinity combined stress between Tibetan wild and cultivated barley Plant Physiol Biochem 63, 49-60. [本文引用: 1]
AinsworthEA, RogersA, LeakeyADB (2008). Targets for crop biotechnology in a future high-CO2 and high-O3 world Plant Physiol 147, 13-19. [本文引用: 1]
AllakhverdievSI, HayashiH, NishiyamaY, IvanovAG, AlievJA, KlimovVV, MurataN, CarpentierR (2003). Glycinebetaine protects the D1/D2/Cytb559 complex of photosystem II against photo-induced and heat-induced inactivation J Plant Physiol 160, 41-49. [本文引用: 1]
AtkinsonNJ, LilleyCJ, UrwinPE (2013). Identification of genes involved in the response of Arabidopsis to simultaneous biotic and abiotic stresses Plant Physiol 162, 2028-2041. [本文引用: 1]
AveryanovAA, LapikovaVP, NikolaevON, StepanovAI (2000). Active oxygen-associated control of rice blast disease by riboflavin and roseoflavin Biochemistry 11, 1292-1298. [本文引用: 1]
BaiYL, KissoudisC, YanZ, VisserRGF, van der LindenG (2018). Plant behaviour under combined stress: tomato responses to combined salinity and pathogen stress Plant J 93, 781-793. [本文引用: 1]
BandurskaH, NiedzielaJ, ChadzinikolauT (2013). Separate and combined responses to water deficit and UV-B radiation Plant Sci 213, 98-105. [本文引用: 1]
BerensML, WolinskaKW, SpaepenS, ZieglerJ, NoboriT, NairA, KrülerV, WinkelmüllerTM, WangYM, MineA, BeckerD, Garrido-OterR, Schulze-LefertP, TsudaK (2019). Balancing trade-offs between biotic and abiotic stress responses through leaf age-dependent variation in stress hormone cross-talk Proc Natl Acad Sci USA 116, 2364-2373. [本文引用: 1]
BookerFL, FiscusEL (2005). The role of ozone flux and antioxidants in the suppression of ozone injury by elevated CO2 in soybean J Exp Bot 56, 2139-2151. [本文引用: 1]
BrouderSM, VolenecJJ (2008). Impact of climate change on crop nutrient and water use efficiencies Physiol Plant 133, 705-724. [本文引用: 1]
Carmo-SilvaAE, GoreMA, Andrade-SanchezP, FrenchAN, HunsakerDJ, SalvucciME (2012). Decreased CO2 availability and inactivation of Rubisco limit photosynthesis in cotton plants under heat and drought stress in the field Environ Exp Bot 83, 1-11. [本文引用: 1]
ChavesMM, MarocoJP, PereiraJS (2003). Understanding plant responses to drought—from genes to the whole plant Funct Plant Biol 30, 239-264. [本文引用: 1]
ChenTH, MurataN (2008). Glycinebetaine: an effective protectant against abiotic stress in plants Trends Plant Sci 13, 499-505. [本文引用: 1]
CoolenS, ProiettiS, HickmanR, Davila OlivasNH, HuangPP, Van VerkMC, Van PeltJA, WittenbergAHJ, De VosM, PrinsM, Van LoonJJA, AartsMGM, DickeM, PieterseCMJ, Van WeesSCM (2016). Transcriptome dynamics of Arabidopsis during sequential biotic and abiotic stresses Plant J 86, 249-267. [本文引用: 1]
CraufurdPQ, PeacockJM (1993). Effect of heat and drought stress on sorghum (Sorghum bicolor). II. Grain yield Exp Agric 29, 77-86. [本文引用: 1]
de SilvaNDG, CholewaE, RyserP (2012). Effects of combined drought and heavy metal stresses on xylem structure and hydraulic conductivity in red maple ( Acer rubrum L.) J Exp Bot 63, 5957-5966. [本文引用: 1]
DuanBL, RanF, ZhangXL, ZhangYB, KorpelainenH, LiCY (2011). Long-term acclimation of mesophyll conductance, carbon isotope discrimination and growth in two contrasting Picea asperata populations exposed to drought and enhanced UV-B radiation for three years Agric Forest Meteor 151, 116-126. [本文引用: 1]
FanJ, HillL, CrooksC, DoernerP, LambC (2009). Abscisic acid has a key role in modulating diverse plant- pathogen interactions Plant Physiol 150, 1750-1761. [本文引用: 1]
FigueroaP, BrowseJ (2012). The Arabidopsis JAZ2 promoter contains a G-box and thymidine-rich module that are necessary and sufficient for jasmonate-dependent activation by MYC transcription factors and repression by JAZ proteins Plant Cell Physiol 53, 330-343. [本文引用: 1]
GaoQM, VenugopalS, NavarreD, KachrooA (2011). Low oleic acid-derived repression of jasmonic acid-inducible defense responses requires the WRKY50 and WRKY51 proteins Plant Physiol 155, 464-476. [本文引用: 1]
GargAK, KimJK, OwensTG, RanwalaAP, Do ChoiY, KochianLV, WuRJ (2002). Trehalose accumulation in rice plants confers high tolerance levels to different abiotic stresses Proc Natl Acad Sci USA 99, 15898-15903. [本文引用: 1]
GhandiA, AdiM, LiliaF, LinoyA, RenaG (2016). Tomato yellow leaf curl virus infection mitigates the heat stress response of plants grown at high temperatures Sci Rep 6, 19715. [本文引用: 1]
GiraudE, HoLHM, CliftonR, CarrollA, EstavilloG, TanYF, HowellKA, IvanovaA, PogsonBJ, MillarAH, WhelanJ (2008). The absence of ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147, 595-610. [本文引用: 2]
GuptaA, HisanoH, HojoY, MatsuuraT, IkedaY, MoriIC, Senthil-KumarM (2017). Global profiling of phytohormone dynamics during combined drought and pathogen stress in Arabidopsis thaliana reveals ABA and JA as major regulators Sci Rep 7, 4017. [本文引用: 1]
HaghjouMM, ShariatiM, SmirnoffN (2009). The effect of acute high light and low temperature stresses on the ascorbate-glutathione cycle and superoxide dismutase activity in two Dunaliella salina strains Physiol Plant 135, 272-280. [本文引用: 1]
HanSH, ParkYJ, ParkCM (2019). Light primes the thermally induced detoxification of reactive oxygen species during thermotolerance development in Arabidopsis Plant Cell Physiol 60, 230-241. [本文引用: 1]
HartikainenK, NergAM, Kivim?enp??M, Kontunen-SoppelaS, M?enp??M, OksanenE, RousiM, HolopainenT (2009). Emissions of volatile organic compounds and leaf structural characteristics of European aspen (Populus tremula) grown under elevated ozone and temperature Tree Physiol 29, 1163-1173. [本文引用: 1]
HeweziT, LégerM, GentzbittelL (2008). A comprehensive analysis of the combined effects of high light and high temperature stresses on gene expression in sunflower Ann Bot 102, 127-140. [本文引用: 1]
HofmannRW, CampbellBD, FountainDF (2003). Sensitivity of white clover to UV-B radiation depends on water availability, plant productivity and duration of stress Glob Chang Biol 9, 473-477. [本文引用: 1]
HubbardM, GermidaJJ, VujanovicV (2014). Fungal endophytes enhance wheat heat and drought tolerance in terms of grain yield and second-generation seed viability J Appl Microbiol 116, 109-122. [本文引用: 1]
IPCC ( 2014). Summary for policymakers. In: Climate change 2014: Impacts, Adaptation, and Vulnerability. Summaries, Frequently Asked Questions, and Cross-Chapter Boxes. A Contribution of Working Group II to the Fifth Assessment Report of the IPCC. Cambridge and New York: Cam- bridge University Press. pp. 190. [本文引用: 2]
IyerNJ, TangYH, MahalingamR (2013). Physiological, biochemical and molecular responses to a combination of drought and ozone in Medicago truncatula Plant Cell Environ 36, 706-720. [本文引用: 2]
JiangYW, HuangBR (2001). Drought and heat stress injury to two cool-season turfgrasses in relation to antioxidant metabolism and lipid peroxidation Crop Sci 41, 436-442. [本文引用: 1]
KasurinenA, BiasiC, HolopainenT, RousiM, M?enp??M, OksanenE (2012). Interactive effects of elevated ozone and temperature on carbon allocation of silver birch (Betula pendula) genotypes in an open-air field exposure Tree Physiol 32, 737-751. [本文引用: 3]
KazanK, MannersJM (2012). JAZ repressors and the orchestration of phytohormone crosstalk Trends Plant Sci 17, 22-31.
Kele?Y, ?ncelI (2002). Response of antioxidative defence system to temperature and water stress combinations in wheat seedlings Plant Sci 163, 783-790. [本文引用: 5]
KoussevitzkyS, SuzukiN, HuntingtonS, ArmijoL, ShaW, CortesD, ShulaevV, MittlerR (2008). Ascorbate peroxidase 1 plays a key role in the response of Arabidopsis thaliana to stress combination J Biol Chem 283, 34197-34203. [本文引用: 1]
KumarD, DattaR, HazraS, SultanaA, MukhopadhyayR, ChattopadhyayS (2015). Transcriptomic profiling of Arabidopsis thaliana mutant pad2.1 in response to combined cold and osmotic stress PLoS One 10, e0122690. [本文引用: 1]
LunnJE (2007). Gene families and evolution of trehalose metabolism in plants Funct Plant Biol 34, 550-563. [本文引用: 1]
MadgwickJW, WestJS, WhiteRP, SemenovMA, TownsendJA, TurnerJA, FittBDL (2011). Impacts of climate change on wheat anthesis and fusarium ear blight in the UK Eur J Plant Pathol 130, 117-131. [本文引用: 1]
MahrookashaniA, SiebertS, HügingH, EwertF (2017). Independent and combined effects of high temperature and drought stress around anthesis on wheat J Agron Crop Sci 203, 453-463. [本文引用: 1]
Martínez-BallestaMDC, BastíasE, CarvajalM (2008). Combined effect of boron and salinity on water transport: the role of aquaporins Plant Signal Behav 3, 844-845. [本文引用: 1]
MittlerR, BlumwaldE (2010). Genetic engineering for modern agriculture: challenges and perspectives Annu Rev Plant Biol 61, 443-462. [本文引用: 2]
ObataT, WittS, LisecJ, Palacios-RojasN, Florez-SarasaI, YousfiS, ArausJL, CairnsJE, FernieAR (2015). Metabolite profiles of maize leaves in drought, heat, and combined stress field trials reveal the relationship between metabolism and grain yield Plant Physiol 169, 2665-2683.
OttmanMJ, KimballBA, PinterPJ, WallGW, VanderlipRL, LeavittSW, LaMorteRL, MatthiasAD, BrooksTJ (2001). Elevated CO2 increases sorghum biomass under drought conditions New Phytol 150, 261-273. [本文引用: 1]
P??kk?nenE, VahalaJ, PohjolaM, HolopainenT, K?renlampiL (1998). Physiological, stomatal and ultrastructural ozone responses in birch ( Betula pendula Roth.) are modified by water stress Plant Cell Environ 21, 671-684. [本文引用: 2]
PandeyP, RamegowdaV, Senthil-KumarM (2015). Shared and unique responses of plants to multiple individual stresses and stress combinations: physiological and molecular mechanisms Front Plant Sci 6, 723. [本文引用: 1]
ParentB, BonneauJ, MaphosaL, KovalchukA, LangridgeP, FleuryD (2017). Quantifying wheat sensitivities to environmental constraints to dissect genotype × environ- ment interactions in the field Plant Physiol 174, 1669-1682.
Pérez-LópezU, Miranda-ApodacaJ, Mu?oz-RuedaA, Mena-PetiteA (2013). Lettuce production and antioxidant capacity are differentially modified by salt stress and light intensity under ambient and elevated CO2 J Plant Physiol 170, 1517-1525. [本文引用: 2]
PoulsonME, BoegerMRT, DonahueRA (2006). Response of photosynthesis to high light and drought for Arabidopsis thaliana grown under a UV-B enhanced light regime Photosynth Res 90, 79-90. [本文引用: 2]
PrasadPVV, PisipatiSR, Mom?ilovi?I, RisticZ (2011). Independent and combined effects of high temperature and drought stress during grain filling on plant yield and chloroplast EF-Tu expression in spring wheat J Agron Crop Sci 197, 430-441. [本文引用: 4]
PraschCM, SonnewaldU (2013). Simultaneous application of heat, drought, and virus to Arabidopsis plants reveals significant shifts in signaling networks Plant Physiol 162, 1849-1866. [本文引用: 2]
RamegowdaV, Senthil-KumarM (2015). The interactive effects of simultaneous biotic and abiotic stresses on plants: mechanistic understanding from drought and pathogen combination J Plant Physiol 176, 47-54. [本文引用: 1]
RasmussenS, BarahP, Suarez-RodriguezMC, BressendorffS, FriisP, CostantinoP, BonesAM, NielsenHB, MundyJ (2013). Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161, 1783-1794. [本文引用: 2]
RiveroRM, MestreTC, MittlerR, RubioF, Garcia- SanchezF, MartinezV (2014). The combined effect of salinity and heat reveals a specific physiological, biochemical and molecular response in tomato plants Plant Cell Environ 37, 1059-1073. [本文引用: 1]
RizhskyL, DavletovaS, LiangHJ, MittlerR (2004). The zinc finger protein Zat12 is required for cytosolic ascorbate peroxidase 1 expression during oxidative stress in Arabidopsis J Biol Chem 279, 11736-11743. [本文引用: 2]
RizhskyL, LiangHJ, MittlerR (2002). The combined effect of drought stress and heat shock on gene expression in tobacco Plant Physiol 130, 1143-1151. [本文引用: 3]
RollinsJA, HabteE, TemplerSE, ColbyT, SchmidtJ, Von KorffM (2013). Leaf proteome alterations in the context of physiological and morphological responses to drought and heat stress in barley ( Hordeum vulgare L.) J Exp Bot 64, 3201-3212. [本文引用: 1]
RoychoudhuryA, PaulS, BasuS (2013). Cross-talk between abscisic acid-dependent and abscisic acid-independent pathways during abiotic stress Plant Cell Rep 32, 985-1006. [本文引用: 2]
SalesCRG, RibeiroRV, SilveiraJAG, MachadoEC, MartinsMO, Lag?aAMMA (2013). Superoxide dismutase and ascorbate peroxidase improve the recovery of photosynthesis in sugarcane plants subjected to water deficit and low substrate temperature Plant Physiol Biochem 73, 326-336. [本文引用: 2]
SalvucciME, Crafts-BrandnerSJ (2004). Relationship bet- ween the heat tolerance of photosynthesis and the thermal stability of Rubisco activase in plants from contrasting thermal environments Plant Physiol 134, 1460-1470.
SangtarashMH, QaderiMM, ChinnappaCC, ReidDM (2009). Differential sensitivity of canola ( Brassica napus) seedlings to ultraviolet-B radiation, water stress and abscisic acid Environ Exp Bot 66, 212-219. [本文引用: 2]
SchenkeD, B?ettcherC, ScheelD (2011). Crosstalk between abiotic ultraviolet-B stress and biotic (flg22) stress signaling in Arabidopsis prevents flavonol accumulation in favor of pathogen defence compound production Plant Cell Environ 34, 1849-1864.
SrivastavaG, KumarS, DubeyG, MishraV, PrasadSM (2012). Nickel and ultraviolet-B stresses induce differential growth and photosynthetic responses in Pisum sativum L. seedlings Biol Trace Elem Res 149, 86-96. [本文引用: 2]
SuzukiN, RiveroRM, ShulaevV, BlumwaldE, MittlerR (2014). Abiotic and biotic stress combinations New Phytol 203, 32-43.
TurtolaS, RousiM, PuseniusJ, YamajiK, HeiskaS, TirkkonenV, MeierB, Julkunen-TiittoR (2006). Genotypic variation in drought response of willows grown under ambient and enhanced UV-B radiation Environ Exp Bot 56, 80-86. [本文引用: 2]
Van IttersumMK, CassmanKG, GrassiniP, WolfJ, TittonellP, HochmanZ (2013). Yield gap analysis with local to global relevance—a review Field Crops Res 143, 4-17. [本文引用: 1]
VileD, PerventM, BelluauM, VasseurF, BressonJ, MullerB, GranierC, SimonneauT (2012). Arabidopsis growth under prolonged high temperature and water deficit: independent or interactive effects? Plant Cell Environ 35, 702-718. [本文引用: 2]
WallerF (2005). The endophytic fungus Pirifomospora indica reprograms barley to salt-stress tolerance, disease resistance, and higher yield Proc Natl Acad Sci USA 38, 13386-13391.
WalterMH (1989). The induction of phenylpropanoid biosynthetic enzymes by ultraviolet light or fungal elicitor in cultured parsley cells is overriden by a heat-shock treatment Planta 177, 1-8. [本文引用: 1]
WayDA, OrenR (2010). Differential responses to changes in growth temperature between trees from different functional groups and biomes: a review and synthesis of data Tree Physiol 30, 669-688.
WelfareK, YeoAR, FlowersTJ (2002). Effects of salinity and ozone, individually and in combination, on the growth and ion contents of two chickpea ( Cicer arietinum L.) varieties Environ Pollut 120, 397-403. [本文引用: 2]
WenXG, QiuNW, LuQT, LuCM (2005). Enhanced thermotolerance of photosystem II in salt-adapted plants of the halophyte Artemisia anethifolia Planta 220, 486-497. [本文引用: 1]
ZandalinasSI, BalfagónD, ArbonaV, Gómez-CadenasA, InupakutikaMA, MittlerR (2016a). ABA is required for the accumulation of APX1 and MBF1c during a combination of water deficit and heat stress J Exp Bot 67, 5381-5390. [本文引用: 2]
ZandalinasSI, RiveroRM, MartínezV, Gómez-CadenasA, ArbonaV (2016b). Tolerance of citrus plants to the combination of high temperatures and drought is associated to the increase in transpiration modulated by a reduction in abscisic acid levels BMC Plant Biol 16, 105. [本文引用: 1]
ZandalinasSI, SalesC, BeltránJ, Gómez-CadenasA, ArbonaV (2017). Activation of secondary metabolism in citrus plants is associated to sensitivity to combined drought and high temperatures Front Plant Sci 7, 1954. [本文引用: 2]
ZhangHN, SonnewaldU (2017). Differences and commonalities of plant responses to single and combined stresses Plant J 90, 839-855.
ZintaG, AbdElgawadH, DomagalskaMA, VergauwenL, KnapenD, NijsI, JanssensIA, BeemsterGTS, AsardH (2014). Physiological, biochemical, and genome-wide transcriptional analysis reveals that elevated CO2 mitigates the impact of combined heat wave and drought stress in Arabidopsis thaliana at multiple organizational levels Glob Chang Biol 20, 3670-3685. [本文引用: 1]
Influence of drought, salt stress and abscisic acid on the resistance of tomato to Botrytis cinerea and Oidium neolycopersici 1 2006
... 联合胁迫的正向还是负向相互作用需要具体情况具体分析, 不能一概而论.有研究表明, 非生物胁迫因子可提高或削弱植物对病原菌的抗性.热胁迫导致活性氧积累, 增强水稻(Oryza sativa)对稻瘟病的抗性(Averyanov et al., 2000); 干旱和盐胁迫处理也能提高植物对病原菌的抗性(Achuoa et al., 2006).而生物胁迫使得植物对非生物胁迫的耐受性降低.例如, 有研究表明, 病毒处理的植物在联合干旱和热胁迫下, 其防卫基因表达下调(Prasch and Sonnewald, 2013), 且高温促进病原菌扩散(Luck et al., 2011; Madgwick et al., 2011); Ghandi等(2016)研究发现, 受黄曲叶病毒侵染的番茄对热胁迫敏感, 且热胁迫敏感的增强与植物热激蛋白或因子下调密切相关, 表明非生物胁迫可能弱化植物对生物胁迫的抗性.此外, 也有生物胁迫促进非生物胁迫的报道.例如, 侵染内生真菌的小麦对干旱以及热胁迫的耐受性增强(Hubbard et al., 2014); 大麦感染内生真菌Piriformis indica后提高了其盐胁迫的耐受性和抗病性(Waller et al., 2005).高温能够促使拟南芥提前开花, 并抑制其对丁香假单胞菌(Pseudomonas syringae) PstDC3000 (avrRpt2)的抗性(Liu et al., 2019).可见, 非生物胁迫弱化还是增强生物胁迫的抗性依赖于单个胁迫的本质、时间、程度以及它们之间的相互作用. ...
Genotypic differences in physiological characteristics in the tolerance to drought and salinity combined stress between Tibetan wild and cultivated barley 1 2013
Pääkkönen et al., 1998; Löw et al., 2006; Iyer et al., 2013
干旱+高CO2
高粱
Ottman et al., 2001; Brouder and Volenec, 2008
盐+高温
番茄(Solanum lycopersicon)
Rivero et al., 2014
盐+高CO2
莴苣(Lactuca sativa)
Pérez-López et al., 2013
盐+硼
玉米(Zea mays)
Martínez-Ballesta et al., 2008
臭氧+高CO2
大豆(Glycine max)
Booker and Fiscus, 2005; Ainsworth et al., 2008
高CO2+高光
莴苣
Pérez-López et al., 2013
1 联合胁迫的负向相互作用 自然界中, 植物同时响应多种胁迫时, 胁迫之间的响应是拮抗的, 造成对植物更严重的不利影响, 称为联合胁迫的负向相互作用(negative interactions).干旱和高温胁迫作为联合胁迫负向作用的代表, 已在多种植物中进行了研究.结果表明, 植物对干旱和高温胁迫的响应相互拮抗.高温胁迫下, 植物能增加气孔导度从而通过蒸腾作用降低叶片温度.干旱胁迫通常导致气孔关闭.当干旱和高温同时发生时, 植物不能打开气孔, 叶片温度会相对较高, 从而对植株生长发育造成更严重的伤害(Vile et al., 2012).相对于单一的干旱或高温胁迫, 联合胁迫对小麦穗数、叶绿素含量、产量和收获指数造成了更严重的负面影响(Prasad et al., 2011; Mahrookashani et al., 2017).植物处于高温和盐联合胁迫时, 蒸腾速率的提高会导致盐离子吸收效率更高, 从而造成更严重的盐胁迫伤害(Keles and Öncel, 2002; Wen et al., 2005).微量元素营养胁迫与其它非生物胁迫联合发生时, 由于微量元素为抗活性氧酶类合成所必需, 从而加重了对植物生长发育的不利影响.上述研究结果表明, 联合胁迫的负向相互作用对植物生长发育造成了更加严重的影响(Martínez-Ballesta et al., 2008). ...
Decreased CO2 availability and inactivation of Rubisco limit photosynthesis in cotton plants under heat and drought stress in the field 1 2012
... 植物光合作用对联合胁迫的响应通常也表现出负向作用.以干旱和高温联合胁迫为例.高温胁迫下, 棉花(Gossypium spp.)的光合速率和气孔导度均下降, 叶片温度升高.干旱和高温复合胁迫下, 其影响程度更加严重(Carmo-Silva et al., 2012), 原因是干旱胁迫下, 叶片的气孔通常处于关闭状态, 导致CO2可利用浓度降低, 进而限制了光合作用(Chaves et al., 2003).高温胁迫主要通过改变电子传递能力和RuBisCO酶活性抑制光合作用(Salvucci and Crafts- Brandner, 2004; Way and Oren, 2010). ...
Understanding plant responses to drought—from genes to the whole plant 1 2003
... 植物光合作用对联合胁迫的响应通常也表现出负向作用.以干旱和高温联合胁迫为例.高温胁迫下, 棉花(Gossypium spp.)的光合速率和气孔导度均下降, 叶片温度升高.干旱和高温复合胁迫下, 其影响程度更加严重(Carmo-Silva et al., 2012), 原因是干旱胁迫下, 叶片的气孔通常处于关闭状态, 导致CO2可利用浓度降低, 进而限制了光合作用(Chaves et al., 2003).高温胁迫主要通过改变电子传递能力和RuBisCO酶活性抑制光合作用(Salvucci and Crafts- Brandner, 2004; Way and Oren, 2010). ...
Glycinebetaine: an effective protectant against abiotic stress in plants 1 2008
... 联合胁迫之间的正向或负向相互作用因植物种类不同而异.盐和高温联合胁迫对小麦造成了负向作用(Keles and Öncel, 2002).然而, 在番茄(Solanum lycopersicon)中, 盐和高温联合胁迫却能缓解单一盐胁迫对番茄的影响.盐胁迫下, 气孔关闭导致光合效率下降; 而叶片中积累的果糖和葡萄糖等反馈抑制光合作用.高温胁迫主要影响光合作用中心电子传递以及二氧化碳同化等, 如RuBisCO酶活、类囊体膜和光系统(PS II)反应中心均受到不同程度的影响.在盐和高温联合胁迫下, 番茄积累大量的甜菜碱和海藻糖来应对联合胁迫, 从而具有较高的光合效率和叶片水势.甜菜碱等物质的积累可以维持高K+浓度, 从而保证了较低的Na+/K+比值, 使植株免受盐胁迫的损伤(Rivero et al., 2014).此外, 甜菜碱还可保护PSII免受高温诱导的失活(Allakhverdiev et al., 2003), 以及抑制盐诱导的K+流出(Cuin and Shabala, 2005).海藻糖被证明在保持植物光合能力和PSII效率上具有重要作用(Lunn, 2007); 并有助于细胞抵抗氧化胁迫(Garg et al., 2002; Chen and Murata, 2008).同时, 在盐和高温联合胁迫下, H2O2的积累与蛋白氧化损伤受到抑制, 进而保护植物免受氧化胁迫(Rivero et al., 2014). ...
Transcriptome dynamics of Arabidopsis during sequential biotic and abiotic stresses 1 2016
... 研究表明, 生物和非生物联合胁迫发生时, 严重的非生物胁迫通常主导植物对联合胁迫的响应.例如, 严重的干旱胁迫和线虫侵染时, 在联合胁迫中96%表达上调的基因同时也受到干旱诱导, 而只有2%的基因受线虫感染上调(Atkinson et al., 2013).此外, 也有关于生物胁迫主导非生物胁迫响应的报道.拟南芥在干旱、灰霉病 (Botrytis cinerea)真菌侵染以及菜粉蝶(Pieris rapae)取食连续胁迫时, 转录组显示基因表达主要受菜粉蝶取食调控(Coolen et al., 2016).显然, 这些联合胁迫就不能够简单地区分为正向或负向相互作用. ...
Effect of heat and drought stress on sorghum (Sorghum bicolor). II. Grain yield 1 1993
Pääkkönen et al., 1998; Löw et al., 2006; Iyer et al., 2013
干旱+高CO2
高粱
Ottman et al., 2001; Brouder and Volenec, 2008
盐+高温
番茄(Solanum lycopersicon)
Rivero et al., 2014
盐+高CO2
莴苣(Lactuca sativa)
Pérez-López et al., 2013
盐+硼
玉米(Zea mays)
Martínez-Ballesta et al., 2008
臭氧+高CO2
大豆(Glycine max)
Booker and Fiscus, 2005; Ainsworth et al., 2008
高CO2+高光
莴苣
Pérez-López et al., 2013
1 联合胁迫的负向相互作用 自然界中, 植物同时响应多种胁迫时, 胁迫之间的响应是拮抗的, 造成对植物更严重的不利影响, 称为联合胁迫的负向相互作用(negative interactions).干旱和高温胁迫作为联合胁迫负向作用的代表, 已在多种植物中进行了研究.结果表明, 植物对干旱和高温胁迫的响应相互拮抗.高温胁迫下, 植物能增加气孔导度从而通过蒸腾作用降低叶片温度.干旱胁迫通常导致气孔关闭.当干旱和高温同时发生时, 植物不能打开气孔, 叶片温度会相对较高, 从而对植株生长发育造成更严重的伤害(Vile et al., 2012).相对于单一的干旱或高温胁迫, 联合胁迫对小麦穗数、叶绿素含量、产量和收获指数造成了更严重的负面影响(Prasad et al., 2011; Mahrookashani et al., 2017).植物处于高温和盐联合胁迫时, 蒸腾速率的提高会导致盐离子吸收效率更高, 从而造成更严重的盐胁迫伤害(Keles and Öncel, 2002; Wen et al., 2005).微量元素营养胁迫与其它非生物胁迫联合发生时, 由于微量元素为抗活性氧酶类合成所必需, 从而加重了对植物生长发育的不利影响.上述研究结果表明, 联合胁迫的负向相互作用对植物生长发育造成了更加严重的影响(Martínez-Ballesta et al., 2008). ...
Long-term acclimation of mesophyll conductance, carbon isotope discrimination and growth in two contrasting Picea asperata populations exposed to drought and enhanced UV-B radiation for three years 1 2011
The Arabidopsis JAZ2 promoter contains a G-box and thymidine-rich module that are necessary and sufficient for jasmonate-dependent activation by MYC transcription factors and repression by JAZ proteins 1 2012
... Iyer等(2013)对苜蓿进行了研究, 发现其在基因转录水平特异性地响应单一的干旱或O3胁迫以及二者的联合胁迫.干旱条件下, 有关脱落酸(ABA)信号和脯氨酸合成, 以及响应高光、高温和氧化胁迫的基因转录上调; 而O3特异性地诱导葡萄糖、蔗糖和葡聚糖代谢相关基因.在联合胁迫下, 茉莉酸(JA)信号通路和固有免疫相关基因转录上调, 一些转录因子(WRKY和MYC3)特异性上调.转录因子WRKY主要响应JA、生物和非生物胁迫的一类转录因子(Gao et al., 2011).JAZ1和JAZ2的抑制子MYC3也参与JA信号通路(Figueroa and Browse, 2012; Kazan and Manners, 2012). ...
Low oleic acid-derived repression of jasmonic acid-inducible defense responses requires the WRKY50 and WRKY51 proteins 1 2011
... Iyer等(2013)对苜蓿进行了研究, 发现其在基因转录水平特异性地响应单一的干旱或O3胁迫以及二者的联合胁迫.干旱条件下, 有关脱落酸(ABA)信号和脯氨酸合成, 以及响应高光、高温和氧化胁迫的基因转录上调; 而O3特异性地诱导葡萄糖、蔗糖和葡聚糖代谢相关基因.在联合胁迫下, 茉莉酸(JA)信号通路和固有免疫相关基因转录上调, 一些转录因子(WRKY和MYC3)特异性上调.转录因子WRKY主要响应JA、生物和非生物胁迫的一类转录因子(Gao et al., 2011).JAZ1和JAZ2的抑制子MYC3也参与JA信号通路(Figueroa and Browse, 2012; Kazan and Manners, 2012). ...
Trehalose accumulation in rice plants confers high tolerance levels to different abiotic stresses 1 2002
... 联合胁迫之间的正向或负向相互作用因植物种类不同而异.盐和高温联合胁迫对小麦造成了负向作用(Keles and Öncel, 2002).然而, 在番茄(Solanum lycopersicon)中, 盐和高温联合胁迫却能缓解单一盐胁迫对番茄的影响.盐胁迫下, 气孔关闭导致光合效率下降; 而叶片中积累的果糖和葡萄糖等反馈抑制光合作用.高温胁迫主要影响光合作用中心电子传递以及二氧化碳同化等, 如RuBisCO酶活、类囊体膜和光系统(PS II)反应中心均受到不同程度的影响.在盐和高温联合胁迫下, 番茄积累大量的甜菜碱和海藻糖来应对联合胁迫, 从而具有较高的光合效率和叶片水势.甜菜碱等物质的积累可以维持高K+浓度, 从而保证了较低的Na+/K+比值, 使植株免受盐胁迫的损伤(Rivero et al., 2014).此外, 甜菜碱还可保护PSII免受高温诱导的失活(Allakhverdiev et al., 2003), 以及抑制盐诱导的K+流出(Cuin and Shabala, 2005).海藻糖被证明在保持植物光合能力和PSII效率上具有重要作用(Lunn, 2007); 并有助于细胞抵抗氧化胁迫(Garg et al., 2002; Chen and Murata, 2008).同时, 在盐和高温联合胁迫下, H2O2的积累与蛋白氧化损伤受到抑制, 进而保护植物免受氧化胁迫(Rivero et al., 2014). ...
Tomato yellow leaf curl virus infection mitigates the heat stress response of plants grown at high temperatures 1 2016
... 联合胁迫的正向还是负向相互作用需要具体情况具体分析, 不能一概而论.有研究表明, 非生物胁迫因子可提高或削弱植物对病原菌的抗性.热胁迫导致活性氧积累, 增强水稻(Oryza sativa)对稻瘟病的抗性(Averyanov et al., 2000); 干旱和盐胁迫处理也能提高植物对病原菌的抗性(Achuoa et al., 2006).而生物胁迫使得植物对非生物胁迫的耐受性降低.例如, 有研究表明, 病毒处理的植物在联合干旱和热胁迫下, 其防卫基因表达下调(Prasch and Sonnewald, 2013), 且高温促进病原菌扩散(Luck et al., 2011; Madgwick et al., 2011); Ghandi等(2016)研究发现, 受黄曲叶病毒侵染的番茄对热胁迫敏感, 且热胁迫敏感的增强与植物热激蛋白或因子下调密切相关, 表明非生物胁迫可能弱化植物对生物胁迫的抗性.此外, 也有生物胁迫促进非生物胁迫的报道.例如, 侵染内生真菌的小麦对干旱以及热胁迫的耐受性增强(Hubbard et al., 2014); 大麦感染内生真菌Piriformis indica后提高了其盐胁迫的耐受性和抗病性(Waller et al., 2005).高温能够促使拟南芥提前开花, 并抑制其对丁香假单胞菌(Pseudomonas syringae) PstDC3000 (avrRpt2)的抗性(Liu et al., 2019).可见, 非生物胁迫弱化还是增强生物胁迫的抗性依赖于单个胁迫的本质、时间、程度以及它们之间的相互作用. ...
The absence of ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress 2 2008
Global profiling of phytohormone dynamics during combined drought and pathogen stress in Arabidopsis thaliana reveals ABA and JA as major regulators 1 2017
The effect of acute high light and low temperature stresses on the ascorbate-glutathione cycle and superoxide dismutase activity in two Dunaliella salina strains 1 2009
Emissions of volatile organic compounds and leaf structural characteristics of European aspen (Populus tremula) grown under elevated ozone and temperature 1 2009
Pääkkönen et al., 1998; Löw et al., 2006; Iyer et al., 2013
干旱+高CO2
高粱
Ottman et al., 2001; Brouder and Volenec, 2008
盐+高温
番茄(Solanum lycopersicon)
Rivero et al., 2014
盐+高CO2
莴苣(Lactuca sativa)
Pérez-López et al., 2013
盐+硼
玉米(Zea mays)
Martínez-Ballesta et al., 2008
臭氧+高CO2
大豆(Glycine max)
Booker and Fiscus, 2005; Ainsworth et al., 2008
高CO2+高光
莴苣
Pérez-López et al., 2013
1 联合胁迫的负向相互作用 自然界中, 植物同时响应多种胁迫时, 胁迫之间的响应是拮抗的, 造成对植物更严重的不利影响, 称为联合胁迫的负向相互作用(negative interactions).干旱和高温胁迫作为联合胁迫负向作用的代表, 已在多种植物中进行了研究.结果表明, 植物对干旱和高温胁迫的响应相互拮抗.高温胁迫下, 植物能增加气孔导度从而通过蒸腾作用降低叶片温度.干旱胁迫通常导致气孔关闭.当干旱和高温同时发生时, 植物不能打开气孔, 叶片温度会相对较高, 从而对植株生长发育造成更严重的伤害(Vile et al., 2012).相对于单一的干旱或高温胁迫, 联合胁迫对小麦穗数、叶绿素含量、产量和收获指数造成了更严重的负面影响(Prasad et al., 2011; Mahrookashani et al., 2017).植物处于高温和盐联合胁迫时, 蒸腾速率的提高会导致盐离子吸收效率更高, 从而造成更严重的盐胁迫伤害(Keles and Öncel, 2002; Wen et al., 2005).微量元素营养胁迫与其它非生物胁迫联合发生时, 由于微量元素为抗活性氧酶类合成所必需, 从而加重了对植物生长发育的不利影响.上述研究结果表明, 联合胁迫的负向相互作用对植物生长发育造成了更加严重的影响(Martínez-Ballesta et al., 2008). ...
Interactive effects of elevated ozone and temperature on carbon allocation of silver birch (Betula pendula) genotypes in an open-air field exposure 3 2012
Metabolite profiles of maize leaves in drought, heat, and combined stress field trials reveal the relationship between metabolism and grain yield 0 2015
Elevated CO2 increases sorghum biomass under drought conditions 1 2001
Pääkkönen et al., 1998; Löw et al., 2006; Iyer et al., 2013
干旱+高CO2
高粱
Ottman et al., 2001; Brouder and Volenec, 2008
盐+高温
番茄(Solanum lycopersicon)
Rivero et al., 2014
盐+高CO2
莴苣(Lactuca sativa)
Pérez-López et al., 2013
盐+硼
玉米(Zea mays)
Martínez-Ballesta et al., 2008
臭氧+高CO2
大豆(Glycine max)
Booker and Fiscus, 2005; Ainsworth et al., 2008
高CO2+高光
莴苣
Pérez-López et al., 2013
1 联合胁迫的负向相互作用 自然界中, 植物同时响应多种胁迫时, 胁迫之间的响应是拮抗的, 造成对植物更严重的不利影响, 称为联合胁迫的负向相互作用(negative interactions).干旱和高温胁迫作为联合胁迫负向作用的代表, 已在多种植物中进行了研究.结果表明, 植物对干旱和高温胁迫的响应相互拮抗.高温胁迫下, 植物能增加气孔导度从而通过蒸腾作用降低叶片温度.干旱胁迫通常导致气孔关闭.当干旱和高温同时发生时, 植物不能打开气孔, 叶片温度会相对较高, 从而对植株生长发育造成更严重的伤害(Vile et al., 2012).相对于单一的干旱或高温胁迫, 联合胁迫对小麦穗数、叶绿素含量、产量和收获指数造成了更严重的负面影响(Prasad et al., 2011; Mahrookashani et al., 2017).植物处于高温和盐联合胁迫时, 蒸腾速率的提高会导致盐离子吸收效率更高, 从而造成更严重的盐胁迫伤害(Keles and Öncel, 2002; Wen et al., 2005).微量元素营养胁迫与其它非生物胁迫联合发生时, 由于微量元素为抗活性氧酶类合成所必需, 从而加重了对植物生长发育的不利影响.上述研究结果表明, 联合胁迫的负向相互作用对植物生长发育造成了更加严重的影响(Martínez-Ballesta et al., 2008). ...
Physiological, stomatal and ultrastructural ozone responses in birch ( Betula pendula Roth.) are modified by water stress 2 1998
... 从胁迫时间上, 联合胁迫分为连续胁迫(consecutive or sequential stress)和平行胁迫(simulta-neous or parallel stress)两种(Zhang and Sonnewald, 2017).连续胁迫指多种胁迫在发生时间上有一定的先后顺序, 而平行胁迫指在同一时间内发生多种胁迫.从胁迫类型上, 其可分为非生物胁迫之间、生物胁迫之间以及非生物与生物胁迫之间的联合胁迫.植物应对一些胁迫的响应信号机制之间存在一定的交互作用.例如, 含有DRE元件的rd29A基因能够整合干旱、盐以及冷胁迫诱导的信号通路(图1) (Roychoudhury et al., 2013).这种胁迫响应通路之间的内在联系, 使得植物应对两种或多种胁迫的响应通常是特异的, 不能够简单地从单一胁迫响应叠加来推断(Ramegowda and Senthil-Kumar, 2015; Zhang and Sonnewald, 2017).联合胁迫发生时, 植物各种生理、代谢和信号途径相互作用, 对植物可能是正向、负向和中性的影响, 但也有可能是新的未知的变化(Pandey et al., 2015; Prasch and Sonnewald, 2013), 使得植物响应联合胁迫异常复杂.联合胁迫之间的正向或负向互作受多种因素影响, 包括不同作物、基因型或胁迫类型、处理时间及强度等(Suzuki et al., 2014) (表1). ...
Pääkkönen et al., 1998; Löw et al., 2006; Iyer et al., 2013
干旱+高CO2
高粱
Ottman et al., 2001; Brouder and Volenec, 2008
盐+高温
番茄(Solanum lycopersicon)
Rivero et al., 2014
盐+高CO2
莴苣(Lactuca sativa)
Pérez-López et al., 2013
盐+硼
玉米(Zea mays)
Martínez-Ballesta et al., 2008
臭氧+高CO2
大豆(Glycine max)
Booker and Fiscus, 2005; Ainsworth et al., 2008
高CO2+高光
莴苣
Pérez-López et al., 2013
1 联合胁迫的负向相互作用 自然界中, 植物同时响应多种胁迫时, 胁迫之间的响应是拮抗的, 造成对植物更严重的不利影响, 称为联合胁迫的负向相互作用(negative interactions).干旱和高温胁迫作为联合胁迫负向作用的代表, 已在多种植物中进行了研究.结果表明, 植物对干旱和高温胁迫的响应相互拮抗.高温胁迫下, 植物能增加气孔导度从而通过蒸腾作用降低叶片温度.干旱胁迫通常导致气孔关闭.当干旱和高温同时发生时, 植物不能打开气孔, 叶片温度会相对较高, 从而对植株生长发育造成更严重的伤害(Vile et al., 2012).相对于单一的干旱或高温胁迫, 联合胁迫对小麦穗数、叶绿素含量、产量和收获指数造成了更严重的负面影响(Prasad et al., 2011; Mahrookashani et al., 2017).植物处于高温和盐联合胁迫时, 蒸腾速率的提高会导致盐离子吸收效率更高, 从而造成更严重的盐胁迫伤害(Keles and Öncel, 2002; Wen et al., 2005).微量元素营养胁迫与其它非生物胁迫联合发生时, 由于微量元素为抗活性氧酶类合成所必需, 从而加重了对植物生长发育的不利影响.上述研究结果表明, 联合胁迫的负向相互作用对植物生长发育造成了更加严重的影响(Martínez-Ballesta et al., 2008). ...
... 自然界中, 植物同时响应多种胁迫时, 胁迫之间的响应是拮抗的, 造成对植物更严重的不利影响, 称为联合胁迫的负向相互作用(negative interactions).干旱和高温胁迫作为联合胁迫负向作用的代表, 已在多种植物中进行了研究.结果表明, 植物对干旱和高温胁迫的响应相互拮抗.高温胁迫下, 植物能增加气孔导度从而通过蒸腾作用降低叶片温度.干旱胁迫通常导致气孔关闭.当干旱和高温同时发生时, 植物不能打开气孔, 叶片温度会相对较高, 从而对植株生长发育造成更严重的伤害(Vile et al., 2012).相对于单一的干旱或高温胁迫, 联合胁迫对小麦穗数、叶绿素含量、产量和收获指数造成了更严重的负面影响(Prasad et al., 2011; Mahrookashani et al., 2017).植物处于高温和盐联合胁迫时, 蒸腾速率的提高会导致盐离子吸收效率更高, 从而造成更严重的盐胁迫伤害(Keles and Öncel, 2002; Wen et al., 2005).微量元素营养胁迫与其它非生物胁迫联合发生时, 由于微量元素为抗活性氧酶类合成所必需, 从而加重了对植物生长发育的不利影响.上述研究结果表明, 联合胁迫的负向相互作用对植物生长发育造成了更加严重的影响(Martínez-Ballesta et al., 2008). ...
Independent and combined effects of high temperature and drought stress during grain filling on plant yield and chloroplast EF-Tu expression in spring wheat 4 2011
... 从胁迫时间上, 联合胁迫分为连续胁迫(consecutive or sequential stress)和平行胁迫(simulta-neous or parallel stress)两种(Zhang and Sonnewald, 2017).连续胁迫指多种胁迫在发生时间上有一定的先后顺序, 而平行胁迫指在同一时间内发生多种胁迫.从胁迫类型上, 其可分为非生物胁迫之间、生物胁迫之间以及非生物与生物胁迫之间的联合胁迫.植物应对一些胁迫的响应信号机制之间存在一定的交互作用.例如, 含有DRE元件的rd29A基因能够整合干旱、盐以及冷胁迫诱导的信号通路(图1) (Roychoudhury et al., 2013).这种胁迫响应通路之间的内在联系, 使得植物应对两种或多种胁迫的响应通常是特异的, 不能够简单地从单一胁迫响应叠加来推断(Ramegowda and Senthil-Kumar, 2015; Zhang and Sonnewald, 2017).联合胁迫发生时, 植物各种生理、代谢和信号途径相互作用, 对植物可能是正向、负向和中性的影响, 但也有可能是新的未知的变化(Pandey et al., 2015; Prasch and Sonnewald, 2013), 使得植物响应联合胁迫异常复杂.联合胁迫之间的正向或负向互作受多种因素影响, 包括不同作物、基因型或胁迫类型、处理时间及强度等(Suzuki et al., 2014) (表1). ...
Pääkkönen et al., 1998; Löw et al., 2006; Iyer et al., 2013
干旱+高CO2
高粱
Ottman et al., 2001; Brouder and Volenec, 2008
盐+高温
番茄(Solanum lycopersicon)
Rivero et al., 2014
盐+高CO2
莴苣(Lactuca sativa)
Pérez-López et al., 2013
盐+硼
玉米(Zea mays)
Martínez-Ballesta et al., 2008
臭氧+高CO2
大豆(Glycine max)
Booker and Fiscus, 2005; Ainsworth et al., 2008
高CO2+高光
莴苣
Pérez-López et al., 2013
1 联合胁迫的负向相互作用 自然界中, 植物同时响应多种胁迫时, 胁迫之间的响应是拮抗的, 造成对植物更严重的不利影响, 称为联合胁迫的负向相互作用(negative interactions).干旱和高温胁迫作为联合胁迫负向作用的代表, 已在多种植物中进行了研究.结果表明, 植物对干旱和高温胁迫的响应相互拮抗.高温胁迫下, 植物能增加气孔导度从而通过蒸腾作用降低叶片温度.干旱胁迫通常导致气孔关闭.当干旱和高温同时发生时, 植物不能打开气孔, 叶片温度会相对较高, 从而对植株生长发育造成更严重的伤害(Vile et al., 2012).相对于单一的干旱或高温胁迫, 联合胁迫对小麦穗数、叶绿素含量、产量和收获指数造成了更严重的负面影响(Prasad et al., 2011; Mahrookashani et al., 2017).植物处于高温和盐联合胁迫时, 蒸腾速率的提高会导致盐离子吸收效率更高, 从而造成更严重的盐胁迫伤害(Keles and Öncel, 2002; Wen et al., 2005).微量元素营养胁迫与其它非生物胁迫联合发生时, 由于微量元素为抗活性氧酶类合成所必需, 从而加重了对植物生长发育的不利影响.上述研究结果表明, 联合胁迫的负向相互作用对植物生长发育造成了更加严重的影响(Martínez-Ballesta et al., 2008). ...
... 联合胁迫的正向还是负向相互作用需要具体情况具体分析, 不能一概而论.有研究表明, 非生物胁迫因子可提高或削弱植物对病原菌的抗性.热胁迫导致活性氧积累, 增强水稻(Oryza sativa)对稻瘟病的抗性(Averyanov et al., 2000); 干旱和盐胁迫处理也能提高植物对病原菌的抗性(Achuoa et al., 2006).而生物胁迫使得植物对非生物胁迫的耐受性降低.例如, 有研究表明, 病毒处理的植物在联合干旱和热胁迫下, 其防卫基因表达下调(Prasch and Sonnewald, 2013), 且高温促进病原菌扩散(Luck et al., 2011; Madgwick et al., 2011); Ghandi等(2016)研究发现, 受黄曲叶病毒侵染的番茄对热胁迫敏感, 且热胁迫敏感的增强与植物热激蛋白或因子下调密切相关, 表明非生物胁迫可能弱化植物对生物胁迫的抗性.此外, 也有生物胁迫促进非生物胁迫的报道.例如, 侵染内生真菌的小麦对干旱以及热胁迫的耐受性增强(Hubbard et al., 2014); 大麦感染内生真菌Piriformis indica后提高了其盐胁迫的耐受性和抗病性(Waller et al., 2005).高温能够促使拟南芥提前开花, 并抑制其对丁香假单胞菌(Pseudomonas syringae) PstDC3000 (avrRpt2)的抗性(Liu et al., 2019).可见, 非生物胁迫弱化还是增强生物胁迫的抗性依赖于单个胁迫的本质、时间、程度以及它们之间的相互作用. ...
... 后基因组时代, 利用基因组信息, 结合表型组学、转录组学、蛋白组学及代谢组学, 筛选、鉴定和分离植物重要性状调控基因或元件, 选择最佳调控通路, 实现对植物定向分子设计, 是当前研究的前沿与热点.在联合胁迫下, 植物通常整合不同的信号(如激素、钙离子、活性氧)及糖代谢等信号通路来响应联合胁迫, 是一个系统性响应(Schenke et al., 2011; Prasch and Sonnewald, 2013).因此, 应用多组学方法解析植物响应联合胁迫的关键信号通路及应对联合胁迫的调控网络, 对阐明植物应对联合胁迫的生理及分子机理显得非常重要. ...
Simultaneous application of heat, drought, and virus to Arabidopsis plants reveals significant shifts in signaling networks 2 2013
... 从胁迫时间上, 联合胁迫分为连续胁迫(consecutive or sequential stress)和平行胁迫(simulta-neous or parallel stress)两种(Zhang and Sonnewald, 2017).连续胁迫指多种胁迫在发生时间上有一定的先后顺序, 而平行胁迫指在同一时间内发生多种胁迫.从胁迫类型上, 其可分为非生物胁迫之间、生物胁迫之间以及非生物与生物胁迫之间的联合胁迫.植物应对一些胁迫的响应信号机制之间存在一定的交互作用.例如, 含有DRE元件的rd29A基因能够整合干旱、盐以及冷胁迫诱导的信号通路(图1) (Roychoudhury et al., 2013).这种胁迫响应通路之间的内在联系, 使得植物应对两种或多种胁迫的响应通常是特异的, 不能够简单地从单一胁迫响应叠加来推断(Ramegowda and Senthil-Kumar, 2015; Zhang and Sonnewald, 2017).联合胁迫发生时, 植物各种生理、代谢和信号途径相互作用, 对植物可能是正向、负向和中性的影响, 但也有可能是新的未知的变化(Pandey et al., 2015; Prasch and Sonnewald, 2013), 使得植物响应联合胁迫异常复杂.联合胁迫之间的正向或负向互作受多种因素影响, 包括不同作物、基因型或胁迫类型、处理时间及强度等(Suzuki et al., 2014) (表1). ...
Leaf proteome alterations in the context of physiological and morphological responses to drought and heat stress in barley ( Hordeum vulgare L.) 1 2013
... 从胁迫时间上, 联合胁迫分为连续胁迫(consecutive or sequential stress)和平行胁迫(simulta-neous or parallel stress)两种(Zhang and Sonnewald, 2017).连续胁迫指多种胁迫在发生时间上有一定的先后顺序, 而平行胁迫指在同一时间内发生多种胁迫.从胁迫类型上, 其可分为非生物胁迫之间、生物胁迫之间以及非生物与生物胁迫之间的联合胁迫.植物应对一些胁迫的响应信号机制之间存在一定的交互作用.例如, 含有DRE元件的rd29A基因能够整合干旱、盐以及冷胁迫诱导的信号通路(图1) (Roychoudhury et al., 2013).这种胁迫响应通路之间的内在联系, 使得植物应对两种或多种胁迫的响应通常是特异的, 不能够简单地从单一胁迫响应叠加来推断(Ramegowda and Senthil-Kumar, 2015; Zhang and Sonnewald, 2017).联合胁迫发生时, 植物各种生理、代谢和信号途径相互作用, 对植物可能是正向、负向和中性的影响, 但也有可能是新的未知的变化(Pandey et al., 2015; Prasch and Sonnewald, 2013), 使得植物响应联合胁迫异常复杂.联合胁迫之间的正向或负向互作受多种因素影响, 包括不同作物、基因型或胁迫类型、处理时间及强度等(Suzuki et al., 2014) (表1). ...
Cross-talk between abscisic acid-dependent and abscisic acid-independent pathways during abiotic stress 2 2013
... 从胁迫时间上, 联合胁迫分为连续胁迫(consecutive or sequential stress)和平行胁迫(simulta-neous or parallel stress)两种(Zhang and Sonnewald, 2017).连续胁迫指多种胁迫在发生时间上有一定的先后顺序, 而平行胁迫指在同一时间内发生多种胁迫.从胁迫类型上, 其可分为非生物胁迫之间、生物胁迫之间以及非生物与生物胁迫之间的联合胁迫.植物应对一些胁迫的响应信号机制之间存在一定的交互作用.例如, 含有DRE元件的rd29A基因能够整合干旱、盐以及冷胁迫诱导的信号通路(图1) (Roychoudhury et al., 2013).这种胁迫响应通路之间的内在联系, 使得植物应对两种或多种胁迫的响应通常是特异的, 不能够简单地从单一胁迫响应叠加来推断(Ramegowda and Senthil-Kumar, 2015; Zhang and Sonnewald, 2017).联合胁迫发生时, 植物各种生理、代谢和信号途径相互作用, 对植物可能是正向、负向和中性的影响, 但也有可能是新的未知的变化(Pandey et al., 2015; Prasch and Sonnewald, 2013), 使得植物响应联合胁迫异常复杂.联合胁迫之间的正向或负向互作受多种因素影响, 包括不同作物、基因型或胁迫类型、处理时间及强度等(Suzuki et al., 2014) (表1). 10.11983/CBB19100.F001
植物响应非生物胁迫(干旱、盐和冷害)的ABA依赖与ABA不依赖途径的交互作用网络(改自<xref ref-type="bibr" rid="b66">Roychoudhury et al., 2013</xref>)
Cross-talk network in ABA-dependent and ABA-independent pathways during abiotic stress (drought, salinity and low temperature) (<xref ref-type="bibr" rid="b66">modified from Roychoudhury et al., 2013</xref>)Figure 1 10.11983/CBB19100.t0001
联合胁迫之间的相互作用 ...
... Cross-talk network in ABA-dependent and ABA-independent pathways during abiotic stress (drought, salinity and low temperature) (modified from Roychoudhury et al., 2013) Figure 1 10.11983/CBB19100.t0001
联合胁迫之间的相互作用 ...
Superoxide dismutase and ascorbate peroxidase improve the recovery of photosynthesis in sugarcane plants subjected to water deficit and low substrate temperature 2 2013
Pääkkönen et al., 1998; Löw et al., 2006; Iyer et al., 2013
干旱+高CO2
高粱
Ottman et al., 2001; Brouder and Volenec, 2008
盐+高温
番茄(Solanum lycopersicon)
Rivero et al., 2014
盐+高CO2
莴苣(Lactuca sativa)
Pérez-López et al., 2013
盐+硼
玉米(Zea mays)
Martínez-Ballesta et al., 2008
臭氧+高CO2
大豆(Glycine max)
Booker and Fiscus, 2005; Ainsworth et al., 2008
高CO2+高光
莴苣
Pérez-López et al., 2013
1 联合胁迫的负向相互作用 自然界中, 植物同时响应多种胁迫时, 胁迫之间的响应是拮抗的, 造成对植物更严重的不利影响, 称为联合胁迫的负向相互作用(negative interactions).干旱和高温胁迫作为联合胁迫负向作用的代表, 已在多种植物中进行了研究.结果表明, 植物对干旱和高温胁迫的响应相互拮抗.高温胁迫下, 植物能增加气孔导度从而通过蒸腾作用降低叶片温度.干旱胁迫通常导致气孔关闭.当干旱和高温同时发生时, 植物不能打开气孔, 叶片温度会相对较高, 从而对植株生长发育造成更严重的伤害(Vile et al., 2012).相对于单一的干旱或高温胁迫, 联合胁迫对小麦穗数、叶绿素含量、产量和收获指数造成了更严重的负面影响(Prasad et al., 2011; Mahrookashani et al., 2017).植物处于高温和盐联合胁迫时, 蒸腾速率的提高会导致盐离子吸收效率更高, 从而造成更严重的盐胁迫伤害(Keles and Öncel, 2002; Wen et al., 2005).微量元素营养胁迫与其它非生物胁迫联合发生时, 由于微量元素为抗活性氧酶类合成所必需, 从而加重了对植物生长发育的不利影响.上述研究结果表明, 联合胁迫的负向相互作用对植物生长发育造成了更加严重的影响(Martínez-Ballesta et al., 2008). ...
... 植物光合作用对联合胁迫的响应通常也表现出负向作用.以干旱和高温联合胁迫为例.高温胁迫下, 棉花(Gossypium spp.)的光合速率和气孔导度均下降, 叶片温度升高.干旱和高温复合胁迫下, 其影响程度更加严重(Carmo-Silva et al., 2012), 原因是干旱胁迫下, 叶片的气孔通常处于关闭状态, 导致CO2可利用浓度降低, 进而限制了光合作用(Chaves et al., 2003).高温胁迫主要通过改变电子传递能力和RuBisCO酶活性抑制光合作用(Salvucci and Crafts- Brandner, 2004; Way and Oren, 2010). ...
Relationship bet- ween the heat tolerance of photosynthesis and the thermal stability of Rubisco activase in plants from contrasting thermal environments 0 2004
Differential sensitivity of canola ( Brassica napus) seedlings to ultraviolet-B radiation, water stress and abscisic acid 2 2009
Pääkkönen et al., 1998; Löw et al., 2006; Iyer et al., 2013
干旱+高CO2
高粱
Ottman et al., 2001; Brouder and Volenec, 2008
盐+高温
番茄(Solanum lycopersicon)
Rivero et al., 2014
盐+高CO2
莴苣(Lactuca sativa)
Pérez-López et al., 2013
盐+硼
玉米(Zea mays)
Martínez-Ballesta et al., 2008
臭氧+高CO2
大豆(Glycine max)
Booker and Fiscus, 2005; Ainsworth et al., 2008
高CO2+高光
莴苣
Pérez-López et al., 2013
1 联合胁迫的负向相互作用 自然界中, 植物同时响应多种胁迫时, 胁迫之间的响应是拮抗的, 造成对植物更严重的不利影响, 称为联合胁迫的负向相互作用(negative interactions).干旱和高温胁迫作为联合胁迫负向作用的代表, 已在多种植物中进行了研究.结果表明, 植物对干旱和高温胁迫的响应相互拮抗.高温胁迫下, 植物能增加气孔导度从而通过蒸腾作用降低叶片温度.干旱胁迫通常导致气孔关闭.当干旱和高温同时发生时, 植物不能打开气孔, 叶片温度会相对较高, 从而对植株生长发育造成更严重的伤害(Vile et al., 2012).相对于单一的干旱或高温胁迫, 联合胁迫对小麦穗数、叶绿素含量、产量和收获指数造成了更严重的负面影响(Prasad et al., 2011; Mahrookashani et al., 2017).植物处于高温和盐联合胁迫时, 蒸腾速率的提高会导致盐离子吸收效率更高, 从而造成更严重的盐胁迫伤害(Keles and Öncel, 2002; Wen et al., 2005).微量元素营养胁迫与其它非生物胁迫联合发生时, 由于微量元素为抗活性氧酶类合成所必需, 从而加重了对植物生长发育的不利影响.上述研究结果表明, 联合胁迫的负向相互作用对植物生长发育造成了更加严重的影响(Martínez-Ballesta et al., 2008). ...
... 后基因组时代, 利用基因组信息, 结合表型组学、转录组学、蛋白组学及代谢组学, 筛选、鉴定和分离植物重要性状调控基因或元件, 选择最佳调控通路, 实现对植物定向分子设计, 是当前研究的前沿与热点.在联合胁迫下, 植物通常整合不同的信号(如激素、钙离子、活性氧)及糖代谢等信号通路来响应联合胁迫, 是一个系统性响应(Schenke et al., 2011; Prasch and Sonnewald, 2013).因此, 应用多组学方法解析植物响应联合胁迫的关键信号通路及应对联合胁迫的调控网络, 对阐明植物应对联合胁迫的生理及分子机理显得非常重要. ...
Crosstalk between abiotic ultraviolet-B stress and biotic (flg22) stress signaling in Arabidopsis prevents flavonol accumulation in favor of pathogen defence compound production 0 2011
Nickel and ultraviolet-B stresses induce differential growth and photosynthetic responses in Pisum sativum L. seedlings 2 2012
... 从胁迫时间上, 联合胁迫分为连续胁迫(consecutive or sequential stress)和平行胁迫(simulta-neous or parallel stress)两种(Zhang and Sonnewald, 2017).连续胁迫指多种胁迫在发生时间上有一定的先后顺序, 而平行胁迫指在同一时间内发生多种胁迫.从胁迫类型上, 其可分为非生物胁迫之间、生物胁迫之间以及非生物与生物胁迫之间的联合胁迫.植物应对一些胁迫的响应信号机制之间存在一定的交互作用.例如, 含有DRE元件的rd29A基因能够整合干旱、盐以及冷胁迫诱导的信号通路(图1) (Roychoudhury et al., 2013).这种胁迫响应通路之间的内在联系, 使得植物应对两种或多种胁迫的响应通常是特异的, 不能够简单地从单一胁迫响应叠加来推断(Ramegowda and Senthil-Kumar, 2015; Zhang and Sonnewald, 2017).联合胁迫发生时, 植物各种生理、代谢和信号途径相互作用, 对植物可能是正向、负向和中性的影响, 但也有可能是新的未知的变化(Pandey et al., 2015; Prasch and Sonnewald, 2013), 使得植物响应联合胁迫异常复杂.联合胁迫之间的正向或负向互作受多种因素影响, 包括不同作物、基因型或胁迫类型、处理时间及强度等(Suzuki et al., 2014) (表1). ...
Pääkkönen et al., 1998; Löw et al., 2006; Iyer et al., 2013
干旱+高CO2
高粱
Ottman et al., 2001; Brouder and Volenec, 2008
盐+高温
番茄(Solanum lycopersicon)
Rivero et al., 2014
盐+高CO2
莴苣(Lactuca sativa)
Pérez-López et al., 2013
盐+硼
玉米(Zea mays)
Martínez-Ballesta et al., 2008
臭氧+高CO2
大豆(Glycine max)
Booker and Fiscus, 2005; Ainsworth et al., 2008
高CO2+高光
莴苣
Pérez-López et al., 2013
1 联合胁迫的负向相互作用 自然界中, 植物同时响应多种胁迫时, 胁迫之间的响应是拮抗的, 造成对植物更严重的不利影响, 称为联合胁迫的负向相互作用(negative interactions).干旱和高温胁迫作为联合胁迫负向作用的代表, 已在多种植物中进行了研究.结果表明, 植物对干旱和高温胁迫的响应相互拮抗.高温胁迫下, 植物能增加气孔导度从而通过蒸腾作用降低叶片温度.干旱胁迫通常导致气孔关闭.当干旱和高温同时发生时, 植物不能打开气孔, 叶片温度会相对较高, 从而对植株生长发育造成更严重的伤害(Vile et al., 2012).相对于单一的干旱或高温胁迫, 联合胁迫对小麦穗数、叶绿素含量、产量和收获指数造成了更严重的负面影响(Prasad et al., 2011; Mahrookashani et al., 2017).植物处于高温和盐联合胁迫时, 蒸腾速率的提高会导致盐离子吸收效率更高, 从而造成更严重的盐胁迫伤害(Keles and Öncel, 2002; Wen et al., 2005).微量元素营养胁迫与其它非生物胁迫联合发生时, 由于微量元素为抗活性氧酶类合成所必需, 从而加重了对植物生长发育的不利影响.上述研究结果表明, 联合胁迫的负向相互作用对植物生长发育造成了更加严重的影响(Martínez-Ballesta et al., 2008). ...
... 联合胁迫的正向还是负向相互作用需要具体情况具体分析, 不能一概而论.有研究表明, 非生物胁迫因子可提高或削弱植物对病原菌的抗性.热胁迫导致活性氧积累, 增强水稻(Oryza sativa)对稻瘟病的抗性(Averyanov et al., 2000); 干旱和盐胁迫处理也能提高植物对病原菌的抗性(Achuoa et al., 2006).而生物胁迫使得植物对非生物胁迫的耐受性降低.例如, 有研究表明, 病毒处理的植物在联合干旱和热胁迫下, 其防卫基因表达下调(Prasch and Sonnewald, 2013), 且高温促进病原菌扩散(Luck et al., 2011; Madgwick et al., 2011); Ghandi等(2016)研究发现, 受黄曲叶病毒侵染的番茄对热胁迫敏感, 且热胁迫敏感的增强与植物热激蛋白或因子下调密切相关, 表明非生物胁迫可能弱化植物对生物胁迫的抗性.此外, 也有生物胁迫促进非生物胁迫的报道.例如, 侵染内生真菌的小麦对干旱以及热胁迫的耐受性增强(Hubbard et al., 2014); 大麦感染内生真菌Piriformis indica后提高了其盐胁迫的耐受性和抗病性(Waller et al., 2005).高温能够促使拟南芥提前开花, 并抑制其对丁香假单胞菌(Pseudomonas syringae) PstDC3000 (avrRpt2)的抗性(Liu et al., 2019).可见, 非生物胁迫弱化还是增强生物胁迫的抗性依赖于单个胁迫的本质、时间、程度以及它们之间的相互作用. ...
The endophytic fungus Pirifomospora indica reprograms barley to salt-stress tolerance, disease resistance, and higher yield 0 2005
The induction of phenylpropanoid biosynthetic enzymes by ultraviolet light or fungal elicitor in cultured parsley cells is overriden by a heat-shock treatment 1 1989
... 植物光合作用对联合胁迫的响应通常也表现出负向作用.以干旱和高温联合胁迫为例.高温胁迫下, 棉花(Gossypium spp.)的光合速率和气孔导度均下降, 叶片温度升高.干旱和高温复合胁迫下, 其影响程度更加严重(Carmo-Silva et al., 2012), 原因是干旱胁迫下, 叶片的气孔通常处于关闭状态, 导致CO2可利用浓度降低, 进而限制了光合作用(Chaves et al., 2003).高温胁迫主要通过改变电子传递能力和RuBisCO酶活性抑制光合作用(Salvucci and Crafts- Brandner, 2004; Way and Oren, 2010). ...
Differential responses to changes in growth temperature between trees from different functional groups and biomes: a review and synthesis of data 0 2010
Effects of salinity and ozone, individually and in combination, on the growth and ion contents of two chickpea ( Cicer arietinum L.) varieties 2 2002
Tolerance of citrus plants to the combination of high temperatures and drought is associated to the increase in transpiration modulated by a reduction in abscisic acid levels 1 2016
... 需要指出的是, 连续胁迫中, 植物对变换两种胁迫的先后顺序的响应通常相似.例如, 在拟南芥(Arabidopsis thaliana) (Zandalinas et al., 2016a)、大麦(Hordeum vulgare) (Rollins et al., 2013)、烟草(Nicotiana tabacum) (Rizhsky et al., 2002)和柑橘(Citrus reticulata) (Zandalinas et al., 2016b)中, 无论先进行干旱胁迫再进行高温胁迫; 还是先进行高温胁迫再进行干旱胁迫, 它们对这两种胁迫的先后处理表现出的生理响应相似, 均在联合胁迫下受到比单一胁迫更严重的损伤, 表明不同植物应对干旱和高温联合胁迫的响应机制相对保守. ...
Activation of secondary metabolism in citrus plants is associated to sensitivity to combined drought and high temperatures 2 2017
... 从胁迫时间上, 联合胁迫分为连续胁迫(consecutive or sequential stress)和平行胁迫(simulta-neous or parallel stress)两种(Zhang and Sonnewald, 2017).连续胁迫指多种胁迫在发生时间上有一定的先后顺序, 而平行胁迫指在同一时间内发生多种胁迫.从胁迫类型上, 其可分为非生物胁迫之间、生物胁迫之间以及非生物与生物胁迫之间的联合胁迫.植物应对一些胁迫的响应信号机制之间存在一定的交互作用.例如, 含有DRE元件的rd29A基因能够整合干旱、盐以及冷胁迫诱导的信号通路(图1) (Roychoudhury et al., 2013).这种胁迫响应通路之间的内在联系, 使得植物应对两种或多种胁迫的响应通常是特异的, 不能够简单地从单一胁迫响应叠加来推断(Ramegowda and Senthil-Kumar, 2015; Zhang and Sonnewald, 2017).联合胁迫发生时, 植物各种生理、代谢和信号途径相互作用, 对植物可能是正向、负向和中性的影响, 但也有可能是新的未知的变化(Pandey et al., 2015; Prasch and Sonnewald, 2013), 使得植物响应联合胁迫异常复杂.联合胁迫之间的正向或负向互作受多种因素影响, 包括不同作物、基因型或胁迫类型、处理时间及强度等(Suzuki et al., 2014) (表1). ...
... ; Zhang and Sonnewald, 2017).联合胁迫发生时, 植物各种生理、代谢和信号途径相互作用, 对植物可能是正向、负向和中性的影响, 但也有可能是新的未知的变化(Pandey et al., 2015; Prasch and Sonnewald, 2013), 使得植物响应联合胁迫异常复杂.联合胁迫之间的正向或负向互作受多种因素影响, 包括不同作物、基因型或胁迫类型、处理时间及强度等(Suzuki et al., 2014) (表1). ...
Differences and commonalities of plant responses to single and combined stresses 0 2017
Physiological, biochemical, and genome-wide transcriptional analysis reveals that elevated CO2 mitigates the impact of combined heat wave and drought stress in Arabidopsis thaliana at multiple organizational levels 1 2014
 ,*中国农业科学院作物科学研究所, 北京 100081
,*中国农业科学院作物科学研究所, 北京 100081
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}