Gene Mapping and Candidate Gene Analysis of Rice Early Senescence Mutant LS-es1
Chun Zhou1, Ran Jiao1, Ping Hu2, Han Lin1, Juan Hu1, Na Xu1, Xianmei Wu2, Yuchun Rao,1,*, Yuexing Wang,2,*1. College of Chemistry and Life Science, Zhejiang Normal University, Jinhua 321004, China 2. State Key Laboratory of Rice Biology, China National Rice Research Institute, Hangzhou 310006, China
Abstract Senescence is an autonomous and irreversible adaptive response at the end of plant development. The molecular mechanism related to premature senescence of leaves is important for rice genetic improvement and breeding of anti-aging varieties. LS-es1 is a stable hereditary premature early senescence mutant obtained by EMS mutagenesis of indica variety TP309. Phenotypic observation, physiological and biochemical analysis of LS-es1 and its wild type TP309 found that LS-es1 accumulated a large amount of reactive oxygen species and more cell death, while the yield-related agronomic traits of LS-es1 were significantly decreased compared to wild type TP309, which also verified the early senescence characteristics of LS-es1. Exogenous hormone treatment of LS-es1 and TP309 seedlings showed that LS-es1 was more sensitive to salicylic acid (SA), abscisic acid (ABA) and methyl jasmonate (MeJA). The LS-es1 gene was mapped to the 46.2 kb region of the long arm of rice chromosome 7 by map-based cloning, which included 8 open reading frames (ORFs). Bioinformatics analysis of the genes in this interval revealed that two candidate functional genes, Os07g0275300 and Os07g0276000, were associated with the early senescence pathway, and the expression levels of these two genes were significantly different between wild type and mutant. The results laid the foundation for further cloning of the LS-es1 gene and in-depth study of its biological function. Keywords:rice;early senescence;physiological characteristics;fine mapping;candidate genes
PDF (3748KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 周纯, 焦然, 胡萍, 林晗, 胡娟, 徐娜, 吴先美, 饶玉春, 王跃星. 水稻早衰突变体LS-es1的基因定位及候选基因分析. 植物学报, 2019, 54(5): 606-619 doi:10.11983/CBB19053 Zhou Chun, Jiao Ran, Hu Ping, Lin Han, Hu Juan, Xu Na, Wu Xianmei, Rao Yuchun, Wang Yuexing. Gene Mapping and Candidate Gene Analysis of Rice Early Senescence Mutant LS-es1. Chinese Bulletin of Botany, 2019, 54(5): 606-619 doi:10.11983/CBB19053
衰老是植物发育末期自主发生且不可逆的适应性反应机制, 也是植物发育过程中的必经阶段。 这一阶段植物细胞发生程序性死亡(programmed cell death, PCD), 器官逐渐退化, 导致植株整体的生物学功能和生命活动衰退。植物衰老的发生是受多种因素共同调控的复杂过程(Yoshida, 2003; Gan and Hörtensteiner, 2013; 徐娜等, 2017), 主要包括内部(遗传、基因调控和激素水平)和外部因素(温度、光照、水分和病虫害等)。水稻(Oryza sativa)叶片是植株进行光合作用的主要器官和重要场所, 为植株提供生长发育需要的大部分能量和有机物质, 而叶片衰老会使植株的光合作用减弱, 从而使供给植株的能量和有机物不足, 进而导致结实率下降、千粒重降低, 最终影响水稻产量和品质(Navabpour et al., 2003)。衰老通常涉及叶绿体的发育和降解、激素水平的变化和自由基代谢等途径(Rao et al., 2015), 影响叶片衰老的诸多因素组成复杂的调控网络起作用。水稻早衰相关基因的克隆是研究水稻早衰机制的基础, 也是构建水稻早衰调控网络的依据, 对水稻遗传育种具有重要意义。
(A) 苗期表型; (B) 分蘖期表型; (C) 成熟期表型。Bars=6 cm Figure 1Phenotypes of rice wild type (WT) TP309 and mu- tant LS-es1
(A) Phenotypes at seedling stage; (B) Phenotypes at tillering stage; (C) Phenotypes at maturity stage. Bars=6 cm
Table 2 表2 表2水稻野生型TP309和突变体LS-es1的农艺性状比较 Table 2The comparison of agronomic traits between rice wild type TP309 and mutant LS-es1
Agronomic traits
TP309
LS-es1
Effective number of panicle
13.40±5.41
8.20±0.84
Flag leaf length (cm)
39.46±8.16
35.68±4.08
Secondary branch number
29.80±6.87
18.00±4.47*
Tiller number
16.40±5.46
11.40±2.70
Filled grain number per panicle
169.60±8.08
125.60±24.83**
Seed-setting rate (%)
81.01±6.98
64.76±17.23
Data are means±SD (n=10). * and ** indicate significant differences between TP309 and LS-es1 at 0.05 and 0.01 level, respectively. 数据为平均值±标准差(n=10)。*和**分别表示TP309与LS-es1在0.05和0.01水平上差异显著。
*和**分别表示TP309与LS-es1在0.05和0.01水平上差异显著。 Figure 4Catalase (CAT) (A), peroxisome (POD) (B), and superoxide dismutase (SOD) (C) activities and malondialdehyde (MDA) (D) and H2O2 (E) contents of LS-es1 and wild type (WT) at heading stage of rice
* and ** indicate significant differences between TP309 and LS-es1 at 0.05 and 0.01 level, respectively.
(A) 水杨酸(SA)处理(上), 脱落酸(ABA)处理(中), 茉莉酸甲酯(MeJA)处理(下) (Bars=2 cm); (B) 激素处理后芽长的比较; (C) 激素处理后根长的比较。** 表示TP309与LS-es1在0.01水平上差异显著。 Figure 7Inhibition of exogenous hormone treatment on the growth of seedlings in rice wild type (WT) TP309 and mutant LS-es1
(A) Salicylic acid (SA) (top), abscisic acid (ABA) (middle), methyl jasmonate (MeJA) (bottom) (Bars=2 cm); (B) Comparison of shoot length of TP309 and LS-es1 seedlings treated with hormones; (C) Comparison of root length of TP309 and LS-es1 seedlings treated with hormones. ** indicate significant differences at 0.01 level between TP309 and LS-es1.
ChenY, XuYY, LuoW, LiWX, ChenN, ZhangDJ, ChongK (2013b). The F-box protein OsFBK12 targets OsSAMS1 for degradation and affects pleiotropic phenotypes including leaf senescence in rice Plant Physiol 163, 1673-1685. [本文引用: 1]
FanataWID, LeeKH, SonBH, YooJY, HarmokoR, KoKS, RamasamyNK, KimKH, OhDB, JungHS, KimJY, LeeSY, LeeKO (2013). N-glycan maturation is crucial for cytokinin-mediated development and cellulose synthesis in Oryza sativa Plant J 73, 966-979. [本文引用: 1]
GanS, AmasinoRM (1997). Making sense of senescence: molecular genetic regulation and manipulation of leaf senescence Plant Physiol 113, 313-319. [本文引用: 1]
GanSS, H?rtensteinerS (2013). Frontiers in plant senescence research: from bench to bank Plant Mol Biol 82, 503-504. [本文引用: 1]
HidegE, KálaiT, KósPB, AsadaK, HidegK (2006). Singlet oxygen in plants—its significance and possible detection with double (fluorescent and spin) indicator reagents Photochem Photobiol 82, 1211-1218. [本文引用: 1]
HuangLM, SunQW, QinFJ, LiC, ZhaoY, ZhouDX (2007). Down-regulation of a SILENT INFORMATION REGULATOR 2-related histone deacetylase gene, OsS- RT1, induces DNA fragmentation and cell death in rice Plant Physiol 144, 1508-1519. [本文引用: 1]
HuangQN, ShiYF, ZhangXB, SongLX, FengBH, WangHM, XuX, LiXH, GuoD, WuJL (2016). Single base substitution in OsCDC48 is responsible for premature senescence and death phenotype in rice J Integr Plant Biol 58, 12-28. [本文引用: 1]
JiangHW, LiMR, LiangNT, YanHB, WeiYB, XuXL, LiuJ, XuZF, ChenF, WuGJ (2007). Molecular cloning and function analysis of the stay green gene in rice Plant J 52, 197-209. [本文引用: 1]
JiaoBB, WangJJ, ZhuXD, ZengLJ, LiQ, HeZH (2012). A novel protein RLS1 with NB-ARM domains is involved in chloroplast degradation during leaf senescence in rice Mol Plant 5, 205-217. [本文引用: 1]
KariolaT, BraderG, LiJ, PalvaET (2005). Chlorophyllase 1, a damage control enzyme, affects the balance between defense pathways in plants Plant Cell 17, 282-294. [本文引用: 1]
KongZS, LiMN, YangWY, XuWY, XueYB (2006). A novel nuclear-localized CCCH-type zinc finger protein, OsDOS, is involved in delaying leaf senescence in rice Plant Physiol 141, 1376-1388. [本文引用: 1]
KusabaM, ItoH, MoritaR, IidaS, SatoY, FujimotoM, KawasakiS, TanakaR, HirochikaH, NishimuraM, TanakaA (2007). Rice NON-YELLOW COLORING1 is involved in light-harvesting complex II and grana degradation during leaf senescence Plant Cell 19, 1362-1375. [本文引用: 1]
LeeRH, LinMC, ChenSC (2004). A novel alkaline α-galactosidase gene is involved in rice leaf senescence Plant Mol Biol 55, 281-295. [本文引用: 1]
LeeRH, WangCH, HuangLT, ChenSCG (2001). Leaf senescence in rice plants: cloning and characterization of senescence up-regulated genes J Exp Bot 52, 1117-1121. [本文引用: 1]
LengYJ, YangYL, RenDY, HuangLC, DaiLP, WangYQ, ChenL, TuZJ, GaoYH, LiXY, ZhuL, HuJ, ZhangGH, GaoZY, GuoLB, KongZS, LinYJ, QianQ, ZengDL (2017). A rice PECTATE LYASE-LIKE gene is required for plant growth and leaf senescence Plant Physiol 174, 1151-1166. [本文引用: 1]
LiangCZ, WangYQ, ZhuYN, TangJY, HuB, LiuLC, OuSJ, WuHK, SunXH, ChuJF, ChuCC (2014). OsNAP connects abscisic acid and leaf senescence by fine-tuning abscisic acid biosynthesis and directly targeting senescence-associated genes in rice Proc Natl Acad Sci USA 111, 10013-10018. [本文引用: 1]
LinAH, WangYQ, TangJY, XueP, LiCL, LiuLC, HuB, YangFQ, LoakeGJ, ChuCC (2012). Nitric oxide and protein S-nitrosylation are integral to hydrogen peroxide-induced leaf cell death in rice Plant Physiol 158, 451-464. [本文引用: 1]
LinYH, TanLB, ZhaoL, SunXY, SunCQ (2016). RLS3, a protein with AAA+ domain localized in chloroplast, sustains leaf longevity in rice J Integr Plant Biol 58, 971-982. [本文引用: 1]
LivakKJ, SchmittgenTD (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2 -ΔΔCt method Methods 25, 402-408. [本文引用: 1]
LuanWJ, ShenA, JinZP, SongSS, LiZL, ShaAH (2013). Knockdown of OsHox33, a member of the class III homeodomain-leucine zipper gene family, accelerates leaf senescence in rice Sci China Life Sci 56, 1113-1123. [本文引用: 1]
MahalingamR, JambunathanN, GunjanSK, FaustinE, WengH, AyoubiP (2006). Analysis of oxidative signaling induced by ozone in Arabidopsis thaliana Plant Cell Environ 29, 1357-1371. [本文引用: 1]
McCabeMS, GarrattLC, SchepersF, JordiWJRM, StoopenGM, DavelaarE, van RhijnJHA, PowerJB, DaveyMR (2001). Effects of PSAG12- IPT gene expression on development and senescence in transgenic lettuce Plant Physiol 127, 505-516. [本文引用: 1]
MoritaR, SatoY, MasudaY, NishimuraM, KusabaM (2010). Defect in non-yellow coloring 3, an α/β hydrolase-fold family protein, causes a stay-green phenotype during leaf senescence in rice Plant J 59, 940-952. [本文引用: 1]
NavabpourS, MorrisK, AllenR, HarrisonE, A-H- MackernessS, Buchanan-WollastonV (2003). Expression of senescence-enhanced genes in response to oxidative stress J Exp Bot 54, 2285-2292. [本文引用: 1]
QiaoYL, JiangWZ, LeeJ, ParkBS, ChoiMS, PiaoRH, WooMO, RohJH, HanLZ, PaekNC, SeoHS, KohHJ (2010). SPL28 encodes a clathrin-associated adaptor protein complex 1, medium subunit μ1 (AP1M1) and is responsible for spotted leaf and early senescence in rice (Oryza sativa) New Phytol 185, 258-274. [本文引用: 1]
RaoYC, YangYL, XuJ, LiXJ, LengYJ, DaiLP, HuangLC, ShaoGS, RenDY, HuJ, GuoLB, PanJW, ZengDL (2015). EARLY SENESCENCE1 encodes a SCAR-LIKE PROTEIN2 that affects water loss in rice Plant Physiol 169, 1225-1239. [本文引用: 1]
SchippersJH, SchmidtR, WagstaffC, JingHC (2015). Living to die and dying to live: the survival strategy behind leaf senescence Plant Physiol 169, 914-930. [本文引用: 1]
SinghS, GiriMK, SinghPK, SiddiquiA, NandiAK (2013). Down-regulation of OsSAG12-1 results in enhanced senescence and pathogen-induced cell death in transgenic rice plants J Biosci 38, 583-592. [本文引用: 1]
SunLT, WangYH, LiuLL, WangCM, GanT, ZhangZY, WangYL, WangD, NiuM, LongWH, LiXH, ZhengM, JiangL, WanJM (2017). Isolation and characterization of a spotted leaf 32 mutant with early leaf senescence and enhanced defense response in rice Sci Rep 7, 41846. [本文引用: 1]
TamiruM, TakagiH, AbeA, YokotaT, KanzakiH, OkamotoH, SaitohH, TakahashiH, FujisakiK, OikawaK, UemuraA, NatsumeS, JikumaruY, MatsuuraH, UmemuraK, TerryMJ, TerauchiR (2016). A chloroplast-localized protein LESION AND LAMINA BENDING affects defence and growth responses in rice New Phytol 210, 1282-1297. [本文引用: 1]
TangYY, LiMR, ChenYP, WuPZ, WuGJ, JiangHW (2011). Knockdown of Os PAO and OsRCCR1 cause different plant death phenotypes in rice J Plant Physiol 168, 1952-1959. [本文引用: 1]
UndanJR, TamiruM, AbeA, YoshidaK, KosugiS, TakagiH, YoshidaK, KanzakiH, SaitohH, FekihR, SharmaS, UndanJ, YanoM, TerauchiR (2012). Mutation in OsLMS, a gene encoding a protein with two double-stranded RNA binding motifs, causes lesion mimic phenotype and early senescence in rice(Oryza sativa L.) Genes Genet Syst 87, 169-179. [本文引用: 1]
WangS, LeiCL, WangJL, MaJ, TangS, WangCL, ZhaoKJ, TianP, ZhangH, QiCY, ChengZJ, ZhangX, GuoXP, LiuLL, WuCY, WanJM (2017). SPL33, encoding an eEF1A-like protein, negatively regulates cell death and defense responses in rice J Exp Bot 68, 899-913. [本文引用: 1]
WuHB, WangB, ChenYL, LiuYG, ChenLT (2013). Characterization and fine mapping of the rice premature senescence mutant ospse1 Theor Appl Genet 126, 1897-1907. [本文引用: 1]
ZhouY, HuangWF, LiuL, ChenTY, ZhouF, LinYJ (2013). Identification and functional characterization of a rice NAC gene involved in the regulation of leaf senescence BMC Plant Biol 13, 132. [本文引用: 1]
ZhouY, LiuL, HuangWF, YuanM, ZhouF, LiXH, LinYJ (2014). Overexpression of OsSWEET5 in rice causes growth retardation and precocious senescence PLoS One 9, e94210. [本文引用: 1]
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
水稻叶片早衰成因及分子机理研究进展 1 2017
... 衰老是植物发育末期自主发生且不可逆的适应性反应机制, 也是植物发育过程中的必经阶段. 这一阶段植物细胞发生程序性死亡(programmed cell death, PCD), 器官逐渐退化, 导致植株整体的生物学功能和生命活动衰退.植物衰老的发生是受多种因素共同调控的复杂过程(Yoshida, 2003; Gan and Hörtensteiner, 2013; 徐娜等, 2017), 主要包括内部(遗传、基因调控和激素水平)和外部因素(温度、光照、水分和病虫害等).水稻(Oryza sativa)叶片是植株进行光合作用的主要器官和重要场所, 为植株提供生长发育需要的大部分能量和有机物质, 而叶片衰老会使植株的光合作用减弱, 从而使供给植株的能量和有机物不足, 进而导致结实率下降、千粒重降低, 最终影响水稻产量和品质(Navabpour et al., 2003).衰老通常涉及叶绿体的发育和降解、激素水平的变化和自由基代谢等途径(Rao et al., 2015), 影响叶片衰老的诸多因素组成复杂的调控网络起作用.水稻早衰相关基因的克隆是研究水稻早衰机制的基础, 也是构建水稻早衰调控网络的依据, 对水稻遗传育种具有重要意义. ...
水稻叶片衰老相关基因的分离 1 2000
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
A novel senescence-associated gene encoding γ-aminobutyric acid (GABA): pyruvate transaminase is upregulated during rice leaf senescence 1 2005
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
The Fd-GOGAT1 mutant gene lc7 confers resistance to Xanthomonas oryzae pv. oryzae in rice 1 2016
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
An S-domain receptor-like kinase, OsSIK2, confers abiotic stress tolerance and delays dark- induced leaf senescence in rice 1 2013a
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
The F-box protein OsFBK12 targets OsSAMS1 for degradation and affects pleiotropic phenotypes including leaf senescence in rice 1 2013
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
N-glycan maturation is crucial for cytokinin-mediated development and cellulose synthesis in Oryza sativa 1 2013
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
Making sense of senescence: molecular genetic regulation and manipulation of leaf senescence 1 1997
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
Frontiers in plant senescence research: from bench to bank 1 2013
... 衰老是植物发育末期自主发生且不可逆的适应性反应机制, 也是植物发育过程中的必经阶段. 这一阶段植物细胞发生程序性死亡(programmed cell death, PCD), 器官逐渐退化, 导致植株整体的生物学功能和生命活动衰退.植物衰老的发生是受多种因素共同调控的复杂过程(Yoshida, 2003; Gan and Hörtensteiner, 2013; 徐娜等, 2017), 主要包括内部(遗传、基因调控和激素水平)和外部因素(温度、光照、水分和病虫害等).水稻(Oryza sativa)叶片是植株进行光合作用的主要器官和重要场所, 为植株提供生长发育需要的大部分能量和有机物质, 而叶片衰老会使植株的光合作用减弱, 从而使供给植株的能量和有机物不足, 进而导致结实率下降、千粒重降低, 最终影响水稻产量和品质(Navabpour et al., 2003).衰老通常涉及叶绿体的发育和降解、激素水平的变化和自由基代谢等途径(Rao et al., 2015), 影响叶片衰老的诸多因素组成复杂的调控网络起作用.水稻早衰相关基因的克隆是研究水稻早衰机制的基础, 也是构建水稻早衰调控网络的依据, 对水稻遗传育种具有重要意义. ...
Singlet oxygen in plants—its significance and possible detection with double (fluorescent and spin) indicator reagents 1 2006
Down-regulation of a SILENT INFORMATION REGULATOR 2-related histone deacetylase gene, OsS- RT1, induces DNA fragmentation and cell death in rice 1 2007
Single base substitution in OsCDC48 is responsible for premature senescence and death phenotype in rice 1 2016
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
Molecular cloning and function analysis of the stay green gene in rice 1 2007
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
A novel protein RLS1 with NB-ARM domains is involved in chloroplast degradation during leaf senescence in rice 1 2012
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
Chlorophyllase 1, a damage control enzyme, affects the balance between defense pathways in plants 1 2005
A novel nuclear-localized CCCH-type zinc finger protein, OsDOS, is involved in delaying leaf senescence in rice 1 2006
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
Rice NON-YELLOW COLORING1 is involved in light-harvesting complex II and grana degradation during leaf senescence 1 2007
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
A novel alkaline α-galactosidase gene is involved in rice leaf senescence 1 2004
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
Leaf senescence in rice plants: cloning and characterization of senescence up-regulated genes 1 2001
... 衰老相关基因在水稻衰老过程中会不同程度地被诱导表达.Osh36编码一个与植株衰老相关的氨基酸转移酶; Osl85编码异柠檬酸裂解酶, 是水稻叶片衰老的标志基因(Lee et al., 2001); OsNAP是联系ABA和叶片衰老的重要纽带, 参与调控水稻衰老(Liang et al., 2014), 同时OsNAP的表达与衰老途径紧密相关, 通过茉莉酸通路来实现对叶片衰老的调控(Zhou et al., 2013); SGR编码叶绿体转运肽蛋白, 参与调控叶绿体合成及降解过程.在始穗期分别取野生型TP309和突变体LS-es1新鲜叶片, 提取RNA后用qRT-PCR对衰老相关基因在野生型和突变体中的表达量进行分析.结果表明, LS-es1中衰老相关基因Osh36、OsI85和OsNAP极显著上调, 其中水稻叶片衰老标志基因Osh36和OsI85的表达量分别上调2.1和20.4倍(图9); 叶片衰老的正调控基因OsNAP表达量上调了2.6倍; 同时叶绿素降解相关基因SGR也显著上调了9.7倍(图9), 表明LS-es1突变体发生严重早衰并伴随叶绿体的降解. ...
A rice PECTATE LYASE-LIKE gene is required for plant growth and leaf senescence 1 2017
Nitric oxide and protein S-nitrosylation are integral to hydrogen peroxide-induced leaf cell death in rice 1 2012
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
RLS3, a protein with AAA+ domain localized in chloroplast, sustains leaf longevity in rice 1 2016
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
Analysis of relative gene expression data using real-time quantitative PCR and the 2 -ΔΔCt method 1 2001
Knockdown of OsHox33, a member of the class III homeodomain-leucine zipper gene family, accelerates leaf senescence in rice 1 2013
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
Analysis of oxidative signaling induced by ozone in Arabidopsis thaliana 1 2006
... 使用氯化硝基四氮唑蓝液(nitrotetrazolium blue chloride, NBT)对水稻叶片进行染色可以反映细胞内的氧自由基含量.NBT染色主要参考Mahalingam等(2006)的方法, 实验前取0.05 g NBT染料, 用100 mL磷酸缓冲液进行溶解, 配制成0.5 mg∙mL-1 NBT染液.在抽穗期取LS-es1和TP309倒二叶叶片完全浸没在配制好的NBT染液中.抽真空, 并在真空状态维持5分钟后缓慢释放, 重复此操作3次后黑暗处理过夜(10小时), 用蒸馏水将样品冲洗干净后置于95%乙醇中, 加热脱色至叶绿素脱净后拍照. ...
Effects of PSAG12- IPT gene expression on development and senescence in transgenic lettuce 1 2001
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
Defect in non-yellow coloring 3, an α/β hydrolase-fold family protein, causes a stay-green phenotype during leaf senescence in rice 1 2010
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
Expression of senescence-enhanced genes in response to oxidative stress 1 2003
... 衰老是植物发育末期自主发生且不可逆的适应性反应机制, 也是植物发育过程中的必经阶段. 这一阶段植物细胞发生程序性死亡(programmed cell death, PCD), 器官逐渐退化, 导致植株整体的生物学功能和生命活动衰退.植物衰老的发生是受多种因素共同调控的复杂过程(Yoshida, 2003; Gan and Hörtensteiner, 2013; 徐娜等, 2017), 主要包括内部(遗传、基因调控和激素水平)和外部因素(温度、光照、水分和病虫害等).水稻(Oryza sativa)叶片是植株进行光合作用的主要器官和重要场所, 为植株提供生长发育需要的大部分能量和有机物质, 而叶片衰老会使植株的光合作用减弱, 从而使供给植株的能量和有机物不足, 进而导致结实率下降、千粒重降低, 最终影响水稻产量和品质(Navabpour et al., 2003).衰老通常涉及叶绿体的发育和降解、激素水平的变化和自由基代谢等途径(Rao et al., 2015), 影响叶片衰老的诸多因素组成复杂的调控网络起作用.水稻早衰相关基因的克隆是研究水稻早衰机制的基础, 也是构建水稻早衰调控网络的依据, 对水稻遗传育种具有重要意义. ...
SPL28 encodes a clathrin-associated adaptor protein complex 1, medium subunit μ1 (AP1M1) and is responsible for spotted leaf and early senescence in rice (Oryza sativa) 1 2010
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
EARLY SENESCENCE1 encodes a SCAR-LIKE PROTEIN2 that affects water loss in rice 1 2015
... 衰老是植物发育末期自主发生且不可逆的适应性反应机制, 也是植物发育过程中的必经阶段. 这一阶段植物细胞发生程序性死亡(programmed cell death, PCD), 器官逐渐退化, 导致植株整体的生物学功能和生命活动衰退.植物衰老的发生是受多种因素共同调控的复杂过程(Yoshida, 2003; Gan and Hörtensteiner, 2013; 徐娜等, 2017), 主要包括内部(遗传、基因调控和激素水平)和外部因素(温度、光照、水分和病虫害等).水稻(Oryza sativa)叶片是植株进行光合作用的主要器官和重要场所, 为植株提供生长发育需要的大部分能量和有机物质, 而叶片衰老会使植株的光合作用减弱, 从而使供给植株的能量和有机物不足, 进而导致结实率下降、千粒重降低, 最终影响水稻产量和品质(Navabpour et al., 2003).衰老通常涉及叶绿体的发育和降解、激素水平的变化和自由基代谢等途径(Rao et al., 2015), 影响叶片衰老的诸多因素组成复杂的调控网络起作用.水稻早衰相关基因的克隆是研究水稻早衰机制的基础, 也是构建水稻早衰调控网络的依据, 对水稻遗传育种具有重要意义. ...
Living to die and dying to live: the survival strategy behind leaf senescence 1 2015
Down-regulation of OsSAG12-1 results in enhanced senescence and pathogen-induced cell death in transgenic rice plants 1 2013
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
Isolation and characterization of a spotted leaf 32 mutant with early leaf senescence and enhanced defense response in rice 1 2017
A chloroplast-localized protein LESION AND LAMINA BENDING affects defence and growth responses in rice 1 2016
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
Knockdown of Os PAO and OsRCCR1 cause different plant death phenotypes in rice 1 2011
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
Mutation in OsLMS, a gene encoding a protein with two double-stranded RNA binding motifs, causes lesion mimic phenotype and early senescence in rice(Oryza sativa L.) 1 2012
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
SPL33, encoding an eEF1A-like protein, negatively regulates cell death and defense responses in rice 1 2017
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
Characterization and fine mapping of the rice premature senescence mutant ospse1 1 2013
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis 1 2007
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
Nyc4, the rice ortholog of Arabidopsis THF1, is involved in the degradation of chlorophyll-protein complexes during leaf senescence 1 2013
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
Molecular regulation of leaf senescence 1 2003
... 衰老是植物发育末期自主发生且不可逆的适应性反应机制, 也是植物发育过程中的必经阶段. 这一阶段植物细胞发生程序性死亡(programmed cell death, PCD), 器官逐渐退化, 导致植株整体的生物学功能和生命活动衰退.植物衰老的发生是受多种因素共同调控的复杂过程(Yoshida, 2003; Gan and Hörtensteiner, 2013; 徐娜等, 2017), 主要包括内部(遗传、基因调控和激素水平)和外部因素(温度、光照、水分和病虫害等).水稻(Oryza sativa)叶片是植株进行光合作用的主要器官和重要场所, 为植株提供生长发育需要的大部分能量和有机物质, 而叶片衰老会使植株的光合作用减弱, 从而使供给植株的能量和有机物不足, 进而导致结实率下降、千粒重降低, 最终影响水稻产量和品质(Navabpour et al., 2003).衰老通常涉及叶绿体的发育和降解、激素水平的变化和自由基代谢等途径(Rao et al., 2015), 影响叶片衰老的诸多因素组成复杂的调控网络起作用.水稻早衰相关基因的克隆是研究水稻早衰机制的基础, 也是构建水稻早衰调控网络的依据, 对水稻遗传育种具有重要意义. ...
Identification and functional characterization of a rice NAC gene involved in the regulation of leaf senescence 1 2013
... 衰老相关基因在水稻衰老过程中会不同程度地被诱导表达.Osh36编码一个与植株衰老相关的氨基酸转移酶; Osl85编码异柠檬酸裂解酶, 是水稻叶片衰老的标志基因(Lee et al., 2001); OsNAP是联系ABA和叶片衰老的重要纽带, 参与调控水稻衰老(Liang et al., 2014), 同时OsNAP的表达与衰老途径紧密相关, 通过茉莉酸通路来实现对叶片衰老的调控(Zhou et al., 2013); SGR编码叶绿体转运肽蛋白, 参与调控叶绿体合成及降解过程.在始穗期分别取野生型TP309和突变体LS-es1新鲜叶片, 提取RNA后用qRT-PCR对衰老相关基因在野生型和突变体中的表达量进行分析.结果表明, LS-es1中衰老相关基因Osh36、OsI85和OsNAP极显著上调, 其中水稻叶片衰老标志基因Osh36和OsI85的表达量分别上调2.1和20.4倍(图9); 叶片衰老的正调控基因OsNAP表达量上调了2.6倍; 同时叶绿素降解相关基因SGR也显著上调了9.7倍(图9), 表明LS-es1突变体发生严重早衰并伴随叶绿体的降解. ...
Overexpression of OsSWEET5 in rice causes growth retardation and precocious senescence 1 2014
... 随着水稻基因组测序的完成和分子生物技术的完善和发展, 前人对水稻衰老的调控机制进行了深入研究, 通过辐射诱变、化学诱变、转座子插入以及T-DNA插入等技术对水稻种子进行处理, 得到大量水稻叶片衰老相关突变体, 并进一步利用这些突变体定位或克隆了叶片衰老相关基因(senescence-associated genes, SAGs).综合前人的研究, 衰老相关基因主要包括3种: 第1种是下调基因, 目前发现的大多数SAGs都属于下调基因, 此类基因的mRNA水平在衰老的叶片中显著降低, 且其表达受到抑制; 第2种为I型SAG, 这类基因具有衰老特异性, 仅在衰老时期表达; 第3种基因为II型SAG, 这类基因的转录水平在衰老启动前后会大幅度升高(Gan and Amasino, 1997).根据功能、代谢途径和信号通路, 目前叶片衰老相关基因又可分为叶绿体发育和叶绿素降解相关基因(如SGR、OsPAO、Ygl1、OsLMS、Ospse1、PSL2、SUR1、NYC1、NYC3、NYC4、RLS1、Osh69和OSRCCR1) (张丽霞, 2000; Lee et al., 2004; Jiang et al., 2007; Kusaba et al., 2007; Morita et al., 2010; Tang et al., 2011; Jiao et al., 2012; Undan et al., 2012; 孙玉莹, 2013; Wu et al., 2013; Yamatani et al., 2013); 蛋白质合成、降解及转运途径相关基因(如GnT1、OsSAG12-1和OsSWEET5) (Fanata et al., 2013; Singh et al., 2013; Zhou et al., 2014); 激素途径相关基因(如OsDos、OsSAMS1和OsHox33) (Kong et al., 2006; Chen et al., 2013b; Luan et al., 2013); PCD途径相关基因(如SPL28、SPL33和OsCATC) (Qiao et al., 2010; Lin et al., 2012; Wang et al., 2017); 以及其它途径相关基因(如psd128、RLS3、Osl2和YGL1) (Ansari et al., 2005; Wu et al., 2007; Lin et al., 2016; Huang et al., 2016).水稻第7号染色体上已定位和克隆的早衰相关基因有LC7、LLB和OsSIK2.其中, LC7编码一个依赖于铁氧还蛋白的谷氨酸合酶(Fd-GOGAT1), 其突变体叶片叶绿体体积减小、数目减少, 且叶绿素合成途径有缺陷, 导致植株的光合作用效率低下.在表型上, 该突变体在四叶期叶片变黄并具有褐色条纹, 随后扩展到整个叶片表面(Chen et al., 2016).LLB编码一个亮氨酸羧基甲基转移酶, 其突变体相比野生型出现叶色变淡、类病斑、叶倾角增大及叶片叶绿素含量降低等表型.该基因主要在叶绿体中发挥功能, 可以直接或间接抑制茉莉酸和油菜素内酯介导的反应, 进而影响水稻的防卫反应和生长应答(Tamiru et al., 2016); OsSIK2是一个依赖Mn2+的蛋白激酶, OsSIK2可增强水稻非生物胁迫抗性并延缓黑暗诱导的叶片衰老, 通过整合发育过程中的胁迫信号使植物在不利环境条件下进行适应性生长.OsSIK2的表达受盐、干旱、冷和ABA诱导, 且在各组织中都有表达, 但在叶片和叶鞘中表达较强(Chen et al., 2013a).通过对衰老相关基因进行研究, 人们对植物衰老的调控途径和分子机理已有初步了解.Mccabe等(2001)将CK合成的限速酶异戊烯基转移酶(isopentenyl transferase, IPT)转入水稻, 其后代植株的衰老得到延缓, 结实率和千粒重也有改善, 但株高显著降低.此外, 衰老过程中还存在一系列生理生化变化, 叶片的衰老往往不是由单个基因调控, 而是由多个基因共同作用, 并且很多衰老相关基因也会受细胞内转录因子以及环境因素的调节. ...
 ,1,*, 王跃星
,1,*, 王跃星
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT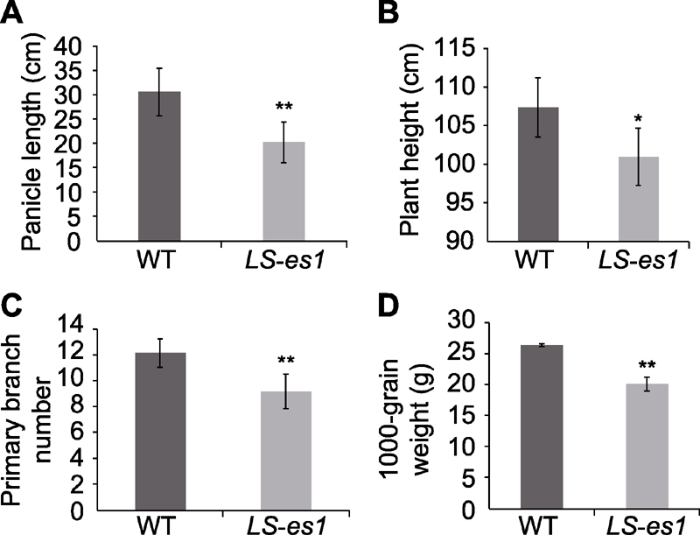 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT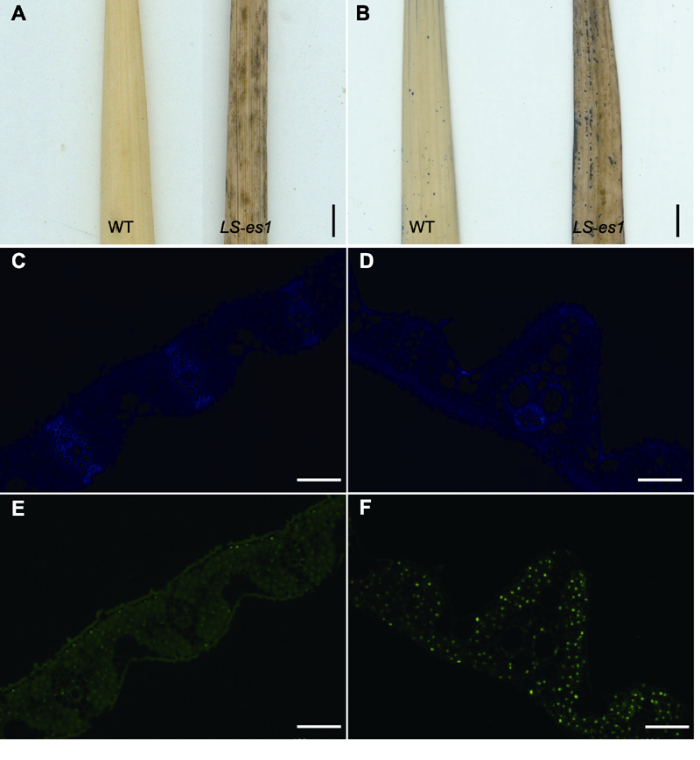 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT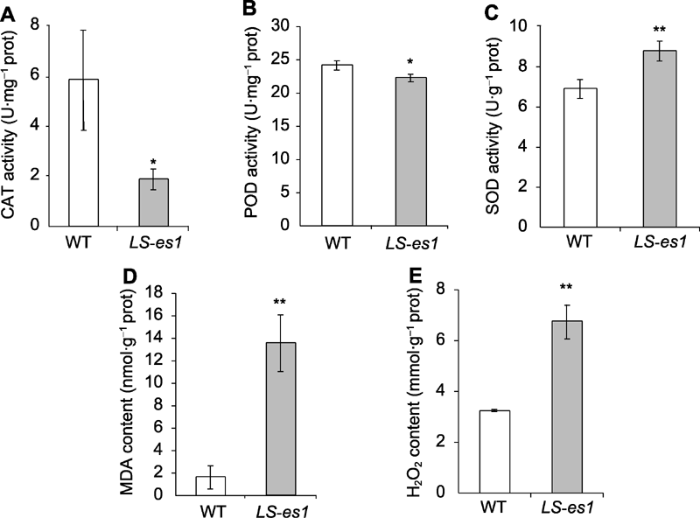 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT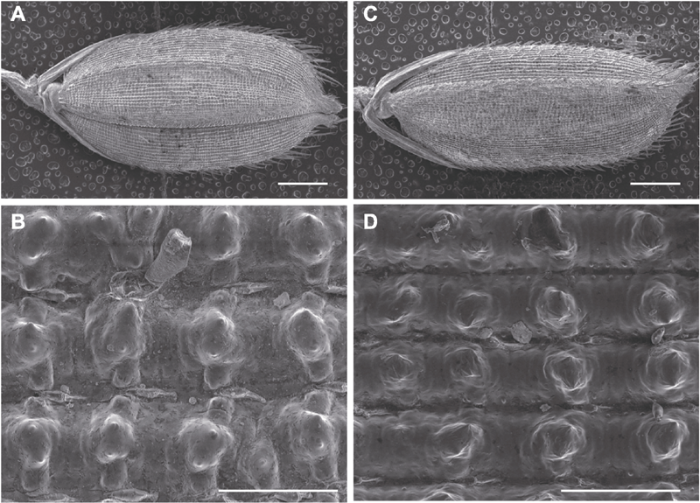 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT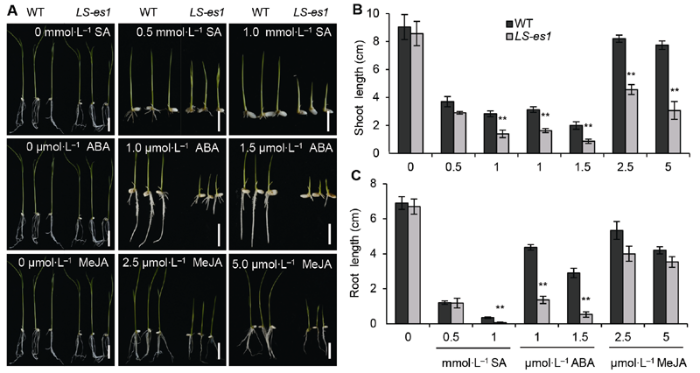 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT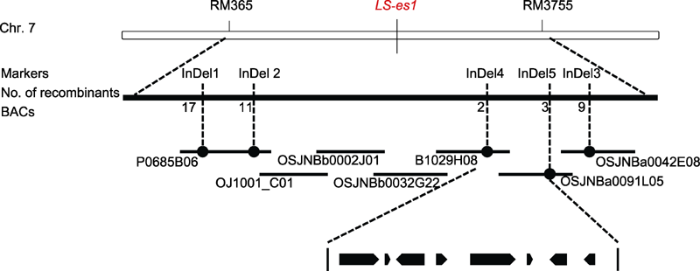 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT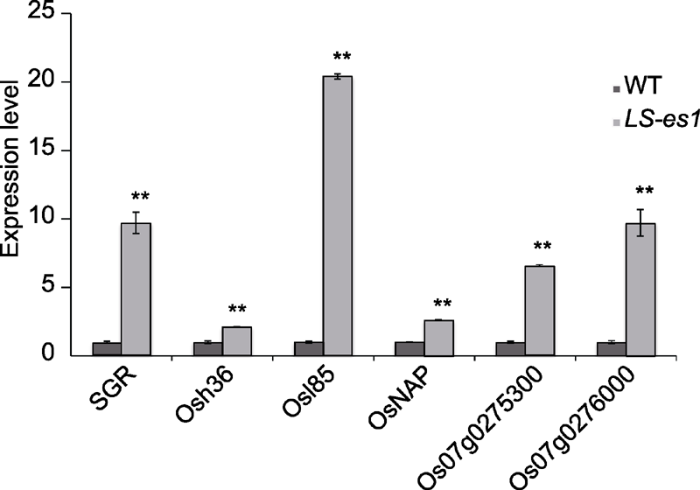 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}