 ,1,*, 吴树敬,1,*
,1,*, 吴树敬,1,*Identifying Genes Responsive to Jasmonates in Apple Based on Transcriptome Analysis
Na Zhang1,2, Xiuxia Liu1, Xuesen Chen,1,*, Shujing Wu,1,*通讯作者:
责任编辑: 朱亚娜
收稿日期:2018-11-4接受日期:2019-02-11网络出版日期:2019-11-01
| 基金资助: |
Corresponding authors:
Received:2018-11-4Accepted:2019-02-11Online:2019-11-01

摘要
关键词:
Abstract
Keywords:
PDF (2830KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
张娜, 刘秀霞, 陈学森, 吴树敬. 基于转录组分析鉴定苹果茉莉素响应基因. 植物学报, 2019, 54(6): 733-743 doi:10.11983/CBB18235
Zhang Na, Liu Xiuxia, Chen Xuesen, Wu Shujing.
苹果(Malus domestica)是世界上栽培面积和产量最大的果树种类之一, 其产量和品质易遭受病原菌等生物胁迫的影响, 给苹果产业带来巨大的经济损失(国立耘等, 2009; 陈学森等, 2015)。使用化学药剂是防治苹果病虫害的有效手段, 但容易造成药物残留及生态破坏。抗性诱导, 即采用植物生长调节剂诱导增强苹果对病原菌的抗性是极其重要且可行的农艺措施(王英珍等, 2016)。有研究表明, 外源水杨酸(salicylic acid, SA)能提高苹果叶片对炭疽病的抗性(Zhang et al., 2016); 在富士苹果中过表达SA信号途径的关键调控因子NPR1能提高苹果对白粉病的抗性(Chen et al., 2012); 此外, SA还能强烈诱导WRKY40b的表达以提高富士苹果对白粉病的抗性(罗昌国等, 2013)。
茉莉素(jasmonates, JA)可诱导植物体内产生凝集素和植保素等一系列具有防御功能的次生代谢物, 或者诱导抗病基因表达以提高植物的抗病性(李梦莎和阎秀峰, 2014)。茉莉酸甲酯(MeJA)对蓝莓(Vaccinium vitis-idaea) (许晴晴等, 2014)、杧果(Mangifera indica) (弓德强等, 2013)、葡萄(Vitis vinifera × V. labrusca) (汪开拓等, 2012)、香蕉(Musa spp. cv. ‘Brazilian’) (麻宝成和朱世江, 2006)和梨(Pyrus pyrifolia) (Zhang et al., 2009)等多种果树均有抗病诱导功能, 但外源JA/MeJA诱导苹果抗病性方面的研究较少。李灿婴等(2015)发现, 富士苹果采后用MeJA处理可增强果实对青霉病的抑制效果。刘志等(2009)研究发现, MeJA处理可以诱导苹果悬浮细胞产生NO, 进而激发防卫反应, 通过产生植保素提高抗性。然而, 外源MeJA诱导苹果抗病性的分子机制仍有待阐明。
RNA-seq高通量转录组测序技术可高效研究植物细胞的转录调控网络, 有助于挖掘关键代谢途径及调控基因, 对解析MeJA诱导的苹果抗病性分子机制具有重要意义。本研究以MeJA处理的苹果叶片为试材, 通过RNA-seq获得苹果中茉莉素响应基因信息, 为深入研究MeJA诱导的苹果抗病分子机制奠定基础。
1 材料与方法
1.1 材料与处理
实验材料为Gala苹果(Malus domestica (Suckow) Borkh. cv. ‘Gala’)组培苗, 培养温度为24°C, 光周期为12小时光照/12小时黑暗。培养30天, 取长势一致的叶片, 将离体叶片浸入100 μmol∙L-1 MeJA溶液中, 24°C处理12小时后取样, 用滤纸吸干表面液体, 在液氮中速冻后保存于-80°C冰箱以备提取RNA。以无菌水处理的叶片为对照(CTRL1), 实验重复3次。1.2 RNA提取及测序
利用RNA提取试剂盒(天根, Cat No.DP424)提取样本总RNA。参考Sun等(2018)的方法富集mRNA、合成cDNA并建立文库。采用Agilent 2100 Bioanalyzer和ABI StepOnePlusRealTime PCR System检测文库产量和质量。测序所得数据经碱基识别分析后得到原始测序结果, 去除含接头以及含N比例大于10%等低质量的序列后得到适合分析的数据。使用BWA (Li and Durbin, 2009)和Bowtie (Langmead et al., 2009)软件将测序数据与金冠苹果基因组进行对比。1.3 差异表达基因分析
利用RSEM工具进行基因表达定量, 结果以FPKM (fragments per kilobase of transcript per million fragments mapped)为单位, FDR (false discovery rate)≤0.001且FC (fold change)≥2的基因为差异表达基因, 将差异表达基因与Nr数据库比对后进行注释。1.4 差异表达基因功能分析
将所有差异表达基因提交到Gene Ontology数据库(http://www.geneontology.org/), 用WEGO软件(Ye et al., 2006)对其进行GO功能分类。数据同时提交KEGG (Kyoto Encyclopedia of Genes and Genomes)数据库(http://www.kegg.jp/), 以KEGG途径为单位, Q-value≤0.05的途径定义为在差异表达基因中显著富集的途径, 分析差异表达基因参与的最主要代谢途径。1.5 差异表达基因的荧光定量分析
采用RevertAid First Strand cDNA Synthesis Kit试剂盒(Thermo, Cat No.K1622)对1.2节中的RNA进行反转录, 以SYBRGreen染料进行荧光定量PCR (qRT- PCR)分析。以苹果ACTIN (序列号为GQ339778)为内参基因, 通过2-ΔΔCt法计算基因的相对表达量。实验设3次重复。差异表达基因的特异性引物由生工生物工程有限公司合成。2 结果与讨论
2.1 RNA-Seq质量评估及差异表达基因筛选
对照(CTRL1)和处理(MeJA)分别得到751 477 132 bp和751 612 960 bp的测序数据, 其中分别有71.28%和72.06%定位于苹果基因组, 44.23%和45.12%的序列与金冠苹果基因组(Velasco et al., 2010)中的基因完全匹配(表1)。Q20介于99.29%-99.32%之间, 所有测序结果均达到质控要求(图1A), 可用于后续分析。我们共筛选到937个差异表达基因, 其中上调表达基因480个, 下调表达基因457个(图1B)。Table 1
表1
表1供试样品(Gala)与苹果(金冠)基因组序列的比对结果
Table 1
| Sample | Total reads | Total base pairs | Total mapped reads | Perfect match | Mismatch | Unique match | Muti-position match | Total unmapped reads |
|---|---|---|---|---|---|---|---|---|
| CTRL1 | 15336268 (100.00%) | 751477132 (100.00%) | 10932142 (71.28%) | 6783455 (44.23%) | 4148687 (27.05%) | 9149425 (59.66%) | 1782717 (11.62%) | 4404126 (28.72%) |
| MeJA | 15339040 (100.00%) | 751612960 (100.00%) | 11052987 (72.06%) | 6921067 (45.12%) | 4131920 (26.94%) | 9177064 (59.83%) | 1875923 (12.23%) | 4286053 (27.94%) |
新窗口打开|下载CSV
图1
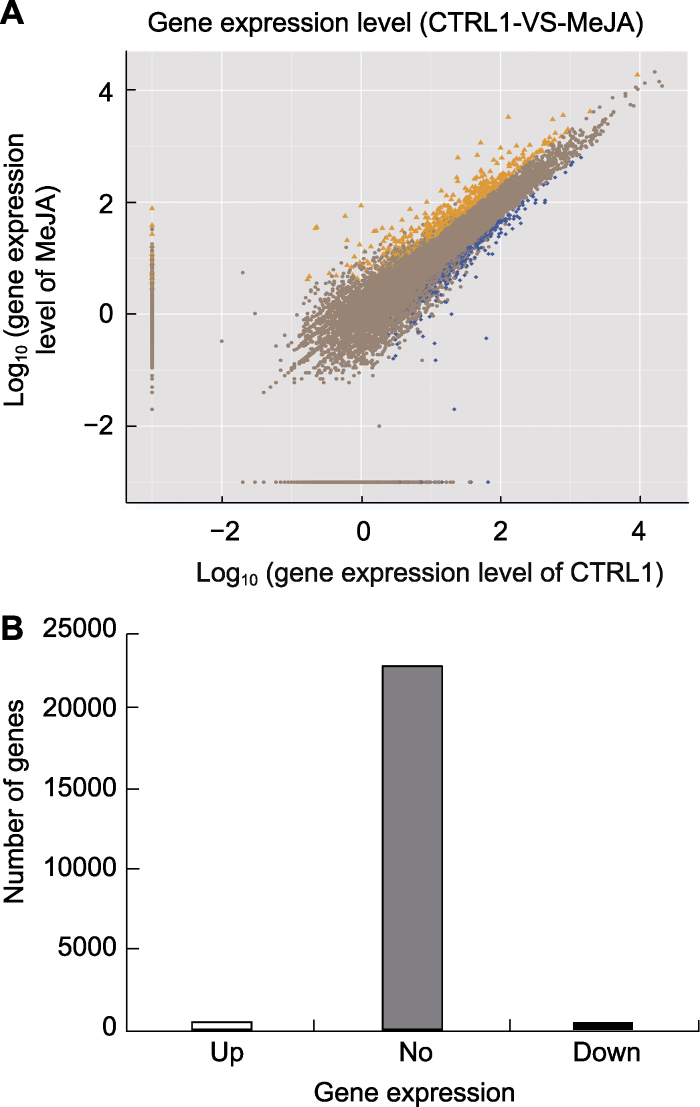 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1茉莉酸甲酯处理下苹果叶片差异表达基因筛选
(A) 基因表达水平。横坐标表示CTRL1基因表达量的对数值, 纵坐标表示MeJA处理基因表达量的对数值(橙色三角代表上调基因, 蓝色方块代表下调基因, 褐色圆点代表非显著差异基因); (B) 基因差异表达情况。Up: 上调基因; No: 非显著差异表达基因; Down: 下调基因
Figure 1The screening of differentially expressed genes in leaves of apple treated by MeJA
(A) Gene expression level. X-axis represents log10 (gene expression level of CTRL1), Y-axis represents log10 (gene expression level of MeJA) (The orange triangle represents up-regulated genes, the blue block represents down-regu- lated genes, and the brown dot represents no significant difference genes); (B) Differentially expressed genes. Up: Up-regulated genes; No: No significant difference genes; Down: Down-regulated genes
2.2 差异表达基因的GO和KEGG分析
GO分析结果表明, 差异表达基因涉及生物学过程(biological process, 5 971)、分子功能(molecular function, 7 605)和细胞成分(cellular component, 3 652) 3类(图2A)。生物学过程涉及15种, 其中调控代谢过程(metabolic process)的基因所占比例最高(4 521, 75.7%), 其次为细胞过程(cellular process, 3 296, 55.2%), 再次为单物种过程(single-organism process, 1 971, 33.0%)。分子功能包含9种, 其中绑定(binding, 4 314, 56.7%)及催化活性(catalytic activity, 5 360, 70.5%)差异基因所占比例最高, 其次为转运活性(transporter activity, 489, 6.4%)。细胞成分包含8种, 其中细胞(cell, 2 669, 73.1%)、细胞组分(cell part, 2 669, 73.1%)所占比例最高, 其次为膜(membrane, 1 226, 33.6%)以及细胞器(organelle, 1 415, 38.7%)。图2
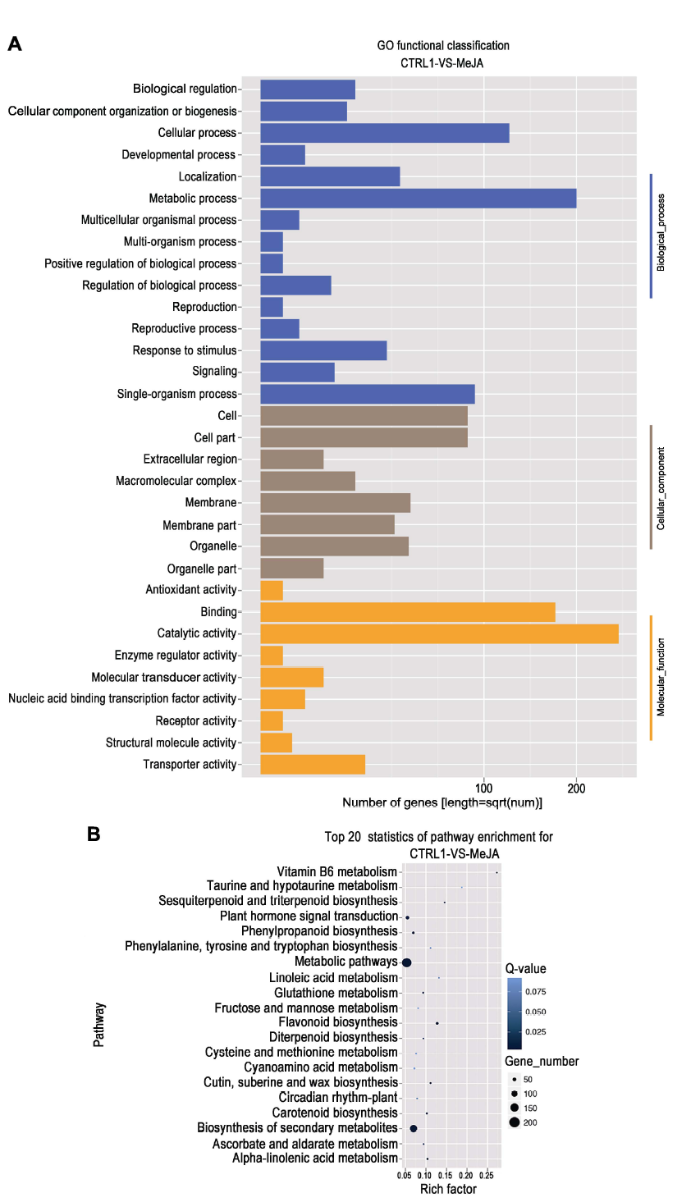 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2茉莉酸甲酯处理苹果叶片差异表达基因的GO功能分类(A)和KEGG富集分析(B)
Figure 2GO functional classification (A) and KEGG enrichment classification (B) of differentially expressed genes in apple leaves under MeJA treatment
采用KEGG富集分析可以确定差异表达基因参与的最主要代谢途径。根据P≤0.01, 找到前20个与整个苹果基因组背景相比显著富集的途径(图2B)。KEGG分析结果发现, 差异表达基因显著富集途径主要包括3个, 其中代谢途径(metabolic pathways)差异表达基因202个, 占35.94%; 次生代谢物合成途径(biosynthesis of secondary metabolites)差异表达基因150个, Q-value最小为1.96×10-15, 占26.69%; 植物激素信号转导途径(plant hormone signal transduction)差异表达基因57个, Q-value为6.5×10-3, 占10.14%。KEGG富集分析结果表明, 代谢过程是最主要的富集途径, 这与GO分析结果相同。因此, 在MeJA诱导下, 代谢过程中的次生代谢物合成途径是最主要的富集途径。
2.3 植物次生代谢物合成途径KEGG转录分析
次生代谢物生物合成途径在植物抵抗胁迫中起重要调控作用。本研究发现, 茉莉酸甲酯处理下, 在苹果叶片次生代谢合成途径中, 113个基因上调表达, 37个基因下调表达。在18个表达比率(Log2 (MeJA/CTRL))绝对值≥3的基因中(表2), 有15个基因上调表达, 3个基因下调表达, 其中有5个基因(MDP0000702120、MDP000205617、MDP0000919962、MDP0000225361和MDP0000248152)参与调控倍半萜类和三萜的生物合成(sesquiterpenoid and triterpenoid biosynthesis); 4个基因(MDP0000692178、MDP0000503940、MDP0000225939和MDP0000523477)参与类黄酮的生物合成(flavonoid biosynthesis); 2个基因(MDP000317152和MDP0000153822)参与角质、木栓质和蜡质的生物合成(cutin, suberine and wax biosynthesis); 2个基因(MDP0000218691和MDP0000280265)参与氨基糖和核苷酸糖代谢(amino sugar and nucleotide sugar metabolism); 2个基因(MDP0000813805和MDP0000228070)参与类胡萝卜素的生物合成(carotenoid biosynthesis); 2个基因(MDP0000687619和MDP0000539956)参与苯丙烷的生物合成途径; 1个基因(MDP0000313394)参与异黄酮的生物合成(isoflavonoid biosynthesis)。上述结果表明, MeJA主要调控苹果叶片倍半萜类、三萜及类黄酮的生物合成途径。Table 2
Table 2The regulated genes involved in biosynthesis of secondary metabolites pathway of apple leaves under MeJA treatment (Log2 (MeJA/CTRL)≥3)
| Gene ID | Log2 ratio (MeJA/CTRL1) | KEGG orthology | Blast nr | Pathway |
|---|---|---|---|---|
| MDP0000702120 | 12.32 | K14181 | (-)-alpha-pinene synthase-like | Sesquiterpenoid and triterpenoid biosynthesis |
| MDP0000205617 | 7.28 | K14181 | Putative pinene synthase | Sesquiterpenoid and triterpenoid biosynthesis |
| MDP0000919962 | 6.57 | K14181 | (-)-alpha-pinene synthase-like | Sesquiterpenoid and triterpenoid biosynthesis |
| MDP0000225361 | 3.72 | K14181 | (-)-germacrene D synthase-like | Sesquiterpenoid and triterpenoid biosynthesis |
| MDP0000248152 | 3.24 | K14181 | (-)-germacrene D synthase-like | Sesquiterpenoid and triterpenoid biosynthesis |
| MDP0000692178 | 12.12 | K00517 | Cytochrome P450 82G1-like | Flavonoid biosynthesis |
| MDP0000503940 | 4.67 | K05277 | Protein SRG1-like | Flavonoid biosynthesis |
| MDP0000225939 | 3.55 | K05277 | Flavonol synthase/flavanone 3-hydroxylase-like | Flavonoid biosynthesis |
| MDP0000523477 | -3.17 | K01859 | Chalcone-flavonone isomerase-like isoform X5 | Flavonoid biosynthesis |
| MDP0000317152 | 13.53 | K13407 | Cytochrome P450 94C1-like | Cutin, suberine and wax biosynthesis |
| MDP0000153822 | 3.25 | K13407 | Cytochrome P450 94B3-like | Cutin, suberine and wax biosynthesis |
| MDP0000218691 | 3.89 | K01183 | Acidic endochitinase-like | Amino sugar and nucleotide sugar metabolism |
| MDP0000280265 | 3.22 | K01183 | Acidic endochitinase-like | Amino sugar and nucleotide sugar metabolism |
| MDP0000687619 | 12.96 | K09755 | Cytochrome P450 CYP736A12-like | Phenylpropanoid biosynthesis |
| MDP0000539956 | 3.9 | K09755 | Cytochrome P450 CYP736A12-like | Phenylpropanoid biosynthesis |
| MDP0000313394 | 6.44 | K13267 | Cytochrome P450 CYP736A12-like | Isoflavonoid biosynthesis |
| MDP0000813805 | -6.29 | K09840 | 9-cis-epoxycarotenoid dioxygenase NCED3, chloroplastic-like | Carotenoid biosynthesis |
| MDP0000228070 | -5.24 | K09840 | 9-cis-epoxycarotenoid dioxygenase NCED1, chloroplastic-like | Carotenoid biosynthesis |
新窗口打开|下载CSV
2.4 植物激素信号转导途径KEGG分析
在MeJA处理下, 苹果叶片中植物激素信号转导途径发生了显著变化(图3)。共有56个基因显著差异表达, 其中涉及芸薹素信号转导途径的基因最多(17个), 其次为茉莉素信号转导途径(10个) (图3; 表3)。图3
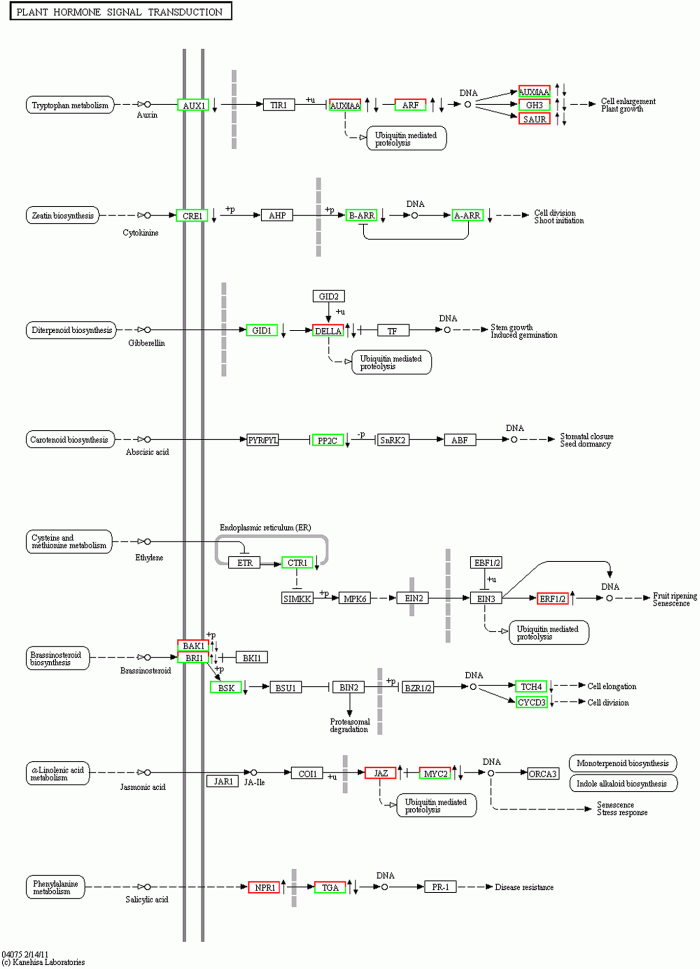 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3茉莉酸甲酯处理对苹果叶片植物激素信号转导途径的影响
红色边框右侧: 上调; 绿色边框右侧
 : 同时包含上调和下调基因。
: 同时包含上调和下调基因。Figure 3Effect of MeJA on plant hormone signaling pathways in leaves of apple
on the right side of red frame: Up-regulated;
 on the right side of red-green frame: Include both up-regulated and down-regulated genes.
on the right side of red-green frame: Include both up-regulated and down-regulated genes.Table 3
表3
表3茉莉酸甲酯处理苹果叶片植物激素信号转导途径中涉及的显著差异表达基因
Table 3
| Plant hormone signaling pathway | Gene ID | Log2 ratio (MeJA/CTRL1) |
|---|---|---|
| Auxin | MDP0000300452 | -1.4 |
| MDP0000255223 MDP0000213864 | 1.1 -1.1 | |
| MDP0000143749 MDP0000221322 | 1.3 -1.6 | |
| MDP0000873893 MDP0000214081 | 1.5 -1.1 | |
| MDP0000786165 | 1.1 | |
| Cytokinine | MDP0000146863 | -1.1 |
| MDP0000285242 MDP0000225179 | 1.6 -1.0 | |
| MDP0000250737 | -1.5 | |
| Gibberellin | MDP0000227827 MDP0000868088 | -1.3 -1.0 |
| MDP0000284679 MDP0000227056 MDP0000397638 | 1.4 1.3 -2.2 | |
| Abscisic acid | MDP0000437033 MDP0000203818 MDP0000283274 | -1.6 -1.3 -1.2 |
| Ethylene | MDP0000275915 | -2.4 |
| MDP0000855671 MDP0000127134 MDP0000235313 MDP0000805422 MDP0000787281 | 3.1 2.5 1.8 1.2 1.0 | |
| Brassinosteroid | MDP0000123379 MDP0000319460 MDP0000319708 MDP0000904826 MDP0000196862 | 12.5 1.9 1.0 1.0 -1.8 |
| MDP0000217213 MDP0000897962 MDP0000290950 MDP0000620422 MDP0000303744 MDP0000189315 MDP0000741253 MDP0000223726 MDP0000189841 | 1.9 1.3 -2.3 -1.8 -1.6 -1.1 -1.0 -1.0 -1.0 | |
| MDP0000130143 | -12.8 | |
| MDP0000320017 | -1.4 | |
| MDP0000135392 | -1.0 | |
| Jasmonic acid | MDP0000565690 MDP0000187921 MDP0000889413 MDP0000301927 | 2.8 2.6 1.7 1.2 |
| MDP0000603546 MDP0000242554 MDP0000029168 MDP0000900024 MDP0000226497 MDP0000290263 | 3.6 2.2 1.6 1.6 1.0 -1.0 | |
| Salicylic acid | MDP0000292425 | 1.8 |
| MDP0000529682 MDP0000375992 | 1.1 -4.3 |
新窗口打开|下载CSV
在5个表达比率绝对值≥3的基因中(表4), MDP0000123379上调幅度最大(12.46), MDP0000130143下调幅度最大(-12.82), 二者均编码丝氨酸/苏氨酸蛋白激酶(serine/threonine-protein kinase), 从而调控芸薹素信号转导。MDP0000375992显著下调(-4.27), 编码U-box结构域蛋白(U-box domain-containing protein), 调控TGA活性进而调节水杨酸信号转导途径。MDP0000603546显著上调(3.56), 编码MYC2进而影响茉莉素信号转导途径。MDP0000855671显著上调(3.1), 编码乙烯响应因子(ethylene- responsive transcription factor 1B-like)而影响ERF1/ 2活性, 从而调控乙烯信号转导途径。
Table 4
表4
表4茉莉酸甲酯处理苹果叶片次生代谢途径中的调控基因(Log2 (MeJA/CTRL)≥3)
Table 4
| Gene ID | Log2 ratio (MeJA/CTRL) | Annotation |
|---|---|---|
| MDP0000123379 | 12.46 | Putative serine/threonine- protein kinase |
| MDP0000603546 | 3.56 | Transcription factor MYC2- like |
| MDP0000855671 | 3.1 | Ethylene-responsive transcription factor 1B-like |
| MDP0000130143 | -12.82 | Probable serine/threonine- protein kinase At5g41260 |
| MDP0000375992 | -4.27 | U-box domain-containing protein 19-like |
新窗口打开|下载CSV
2.5 植物-病原菌互作途径转录分析
植物对病害的抗性包括病原菌入侵之前就具备的基础抗性以及受病原菌激发获得的主动抗性。植物与病原菌互作途径在植物早期抗病反应中发挥重要作用。KEGG分析表明, 在植物-病原菌互作途径中, 共有63个基因显著差异表达, 共有17个作用位点受到诱导调控, 参与基因最多的调控位点为一类富含亮氨酸拉链的丝氨酸/苏氨酸类受体激酶(LRR receptor-like serine/threonine-protein kinase), 有17个基因参与调控; 其次为另一种丝氨酸/苏氨酸蛋白激酶PBS1 (serine/threonine-protein kinase PBS1), 有7个基因参与调控。表达量差异较大的基因有MDP0000123379 (12.5)、MDP0000263349 (-12.2)和MDP0000263768 (5.0) (图4)。MDP0000263349 (-12.2)和MDP0000263768 (5.0)共同调控WRKY25转录因子, 负调控防御相关基因(defense- related gene)的诱导表达。MDP0000123379编码的蛋白激酶则在WRKY25的上游发挥早期抗病诱导作用。图4
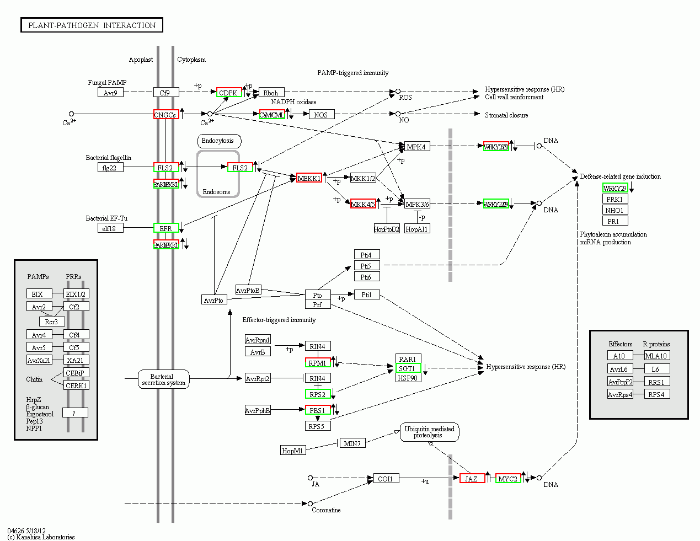 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4茉莉酸甲酯处理对苹果叶片植物-病原菌互作途径的影响
红色边框右侧: 上调; 绿色边框右侧
 : 同时包含上调和下调基因。
: 同时包含上调和下调基因。Figure 4Effect of MeJA on plant-pathogen interaction pathway in leaves of apple
on the right side of red frame: Up-regulated;
 on the right side of red-green frame: Include both up-regulated and down-regulated genes.
on the right side of red-green frame: Include both up-regulated and down-regulated genes.2.6 差异表达基因荧光定量分析
随机选定12个差异表达基因(7个上调基因, 5个下调基因)进行qRT-PCR分析。结果(图5)表明, 7个上调基因均受MeJA诱导显著上调, 5个下调基因也受MeJA诱导显著下调, 即qRT-PCR结果与转录组分析结果趋势一致, 只是在差异倍数上与转录组数据有所差别。图5
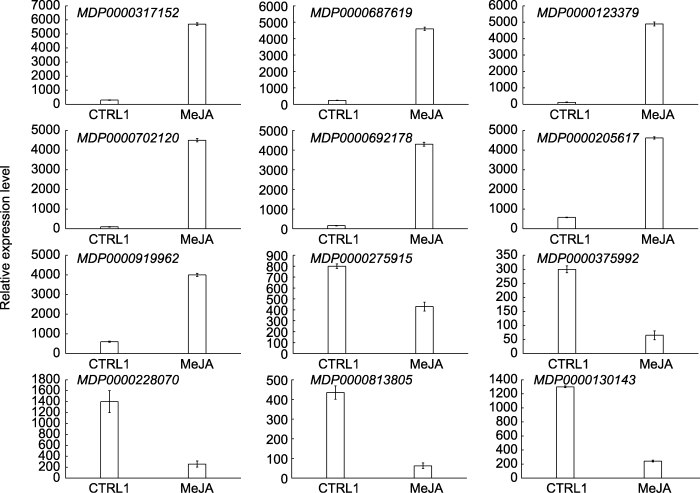 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5茉莉酸甲酯处理苹果叶片中差异表达基因的qRT-PCR分析
Figure 5qRT-PCR analysis of differentially expressed genes in leaves of apple under MeJA treatment
2.7 讨论
2.7.1 MeJA诱导的苹果叶片中主要代谢途径及重要功能基因外源施加MeJA有利于提高植物的抗病性(宾金华和潘瑞炽, 1995), MeJA对苹果抗病性的诱导作用已在少数研究中得到证实(刘志等, 2009; 李灿婴等, 2015), 但具体分子调控机制尚不明确, 涉及的关键基因也未探明。本研究利用外源MeJA处理苹果叶片并进行转录组测序, 探讨MeJA处理对苹果叶片主要代谢通路的影响, 为揭示MeJA调控抗病反应的分子机制奠定基础。
本研究发现, 经MeJA处理后, 苹果叶片中的绝大多数基因表达并未发生显著变化。KEGG富集分析结果表明, 经MeJA诱导的差异表达基因显著富集途径主要包括次生代谢物合成和植物激素信号转导途径, 并不涉及植物-病原菌互作途径(图2B), 这表明MeJA诱导的苹果叶片抗病性可能并不直接影响植物与病原菌互作中的基因表达, 而是主要通过影响特定次生代谢产物合成及调控植物激素信号转导发挥抗病诱导功能, 其具体分子机制尚待深入研究。
在次生代谢途径中, MeJA主要调控苹果叶片倍半萜类和三萜以及类黄酮的生物合成。涉及倍半萜类和三萜生物合成途径的基因主要调控蒎烯合成酶(pinene synthase)合成, 最主要的调控基因为MDP0000702120 (12.32, K14181)。K14181编码6-磷酸海藻糖合成酶1 (trehalose-6-phosphate synthase 1, Tps1), 而Tps1可通过调节海藻糖代谢产生抗真菌抑制剂已经在水稻(Oryza sativa)中得到证实(Xue et al., 2014)。在类黄酮生物合成途径中MeJA主要调控细胞色素P450, 最主要的调控基因为MDP0000692178 (12.12, K13083)。
MeJA处理后苹果叶片中8种植物激素信号转导途径均存在不同程度的变化(图3), 其中以芸薹素信号转导途径和以茉莉素的活性形式(茉莉酸异亮氨酸配合物(jasmonoyl-L-isoleucine, JA-Ile))介导的茉莉素信号转导途径基因最多, 表明MeJA很可能通过调控芸薹素信号转导途径和茉莉素信号转导途径间接诱导叶片的抗病性。进一步分析表明, 在芸薹素信号转导途径中的5个调节比率绝对值≥3的基因中(表4), 上调幅度最大的基因是MDP0000123379, 其通过编码丝氨酸/苏氨酸蛋白激酶来调控细胞信号转导; 下调幅度最大的基因是MDP0000130143, 主要通过调控编码丝氨酸/苏氨酸蛋白激酶活性来调节芸薹素信号转导途径。此外, 芸薹素信号转导途径中, 活性发生显著性改变的基因数多于茉莉素信号转导途径, 因此可以推测, MeJA对芸薹素信号转导途径的调控较茉莉素信号转导途径更为重要, 且主要调控蛋白激酶等。
2.7.2 MeJA诱导的苹果叶片植物-病原菌互作途径
植物-病原菌互作途径虽然不是受MeJA调控的主要途径, 但也发生了显著变化。其中表达量差异较大的基因有MDP0000123379 (K13416, 12.46), 该基因编码丝氨酸/苏氨酸蛋白激酶; 而MDP0000263349
(K13424, -12.2)则编码WRKY33转录因子。本课题组在前期研究中发现, WRKY33转录因子的表达与轮纹病病程相关基因MdPRs的表达高度协同(张芮, 2015), 因此MeJA可能对病程相关基因的表达也存在正向调控作用, 从而诱导叶片产生抗病性。MDP0000123379 (K13416, 12.46)既参与芸薹素信号转导途径, 又参与植物-病原菌互作途径, 是联系植物激素信号转导途径和植物-病原菌互作途径的关键调控基因。
3 结论
外源MeJA主要影响苹果叶片倍半萜类、三萜及类黄酮的生物合成, 也通过信号转导途径间接诱导抗病性。其中, 倍半萜类、三萜和类黄酮生物合成途径中的关键基因为MDP0000702120和MDP0000692178; MDP0000123379是联系芸薹素信号转导途径和植物-病原菌互作途径的关键调控基因。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
茉莉酸甲酯的生理生化及在植物抗病中的作用
1
1995
... 外源施加MeJA有利于提高植物的抗病性(
主要果树果实品质遗传改良与提升实践
1
2015
... 苹果(Malus domestica)是世界上栽培面积和产量最大的果树种类之一, 其产量和品质易遭受病原菌等生物胁迫的影响, 给苹果产业带来巨大的经济损失(
杧果采前喷施茉莉酸甲酯对其抗病性和采后品质的影响
1
2013
... 茉莉素(jasmonates, JA)可诱导植物体内产生凝集素和植保素等一系列具有防御功能的次生代谢物, 或者诱导抗病基因表达以提高植物的抗病性(
中国苹果枝干轮纹病发生和防治情况
1
2009
... 苹果(Malus domestica)是世界上栽培面积和产量最大的果树种类之一, 其产量和品质易遭受病原菌等生物胁迫的影响, 给苹果产业带来巨大的经济损失(
采后茉莉酸甲酯处理对富士苹果青霉病和贮藏品质的影响
2
2015
... 茉莉素(jasmonates, JA)可诱导植物体内产生凝集素和植保素等一系列具有防御功能的次生代谢物, 或者诱导抗病基因表达以提高植物的抗病性(
... 外源施加MeJA有利于提高植物的抗病性(
植物的环境信号分子茉莉酸及其生物学功能
1
2014
... 茉莉素(jasmonates, JA)可诱导植物体内产生凝集素和植保素等一系列具有防御功能的次生代谢物, 或者诱导抗病基因表达以提高植物的抗病性(
茉莉酸甲酯诱导的苹果悬浮细胞的防卫响应
2
2009
... 茉莉素(jasmonates, JA)可诱导植物体内产生凝集素和植保素等一系列具有防御功能的次生代谢物, 或者诱导抗病基因表达以提高植物的抗病性(
... 外源施加MeJA有利于提高植物的抗病性(
富士苹果MdWRKY40b基因克隆及其对白粉病的抗性分析
1
2013
... 苹果(Malus domestica)是世界上栽培面积和产量最大的果树种类之一, 其产量和品质易遭受病原菌等生物胁迫的影响, 给苹果产业带来巨大的经济损失(
苯丙噻重氮和茉莉酸甲酯对采后香蕉果实抗病性及相关酶活性的影响
1
2006
... 茉莉素(jasmonates, JA)可诱导植物体内产生凝集素和植保素等一系列具有防御功能的次生代谢物, 或者诱导抗病基因表达以提高植物的抗病性(
茉莉酸甲酯处理对葡萄果实NO和H2O2水平及植保素合成的影响
1
2012
... 茉莉素(jasmonates, JA)可诱导植物体内产生凝集素和植保素等一系列具有防御功能的次生代谢物, 或者诱导抗病基因表达以提高植物的抗病性(
采前茉莉酸甲酯(MeJA)处理对梨果实抗病性的影响
1
2016
... 苹果(Malus domestica)是世界上栽培面积和产量最大的果树种类之一, 其产量和品质易遭受病原菌等生物胁迫的影响, 给苹果产业带来巨大的经济损失(
茉莉酸甲酯对蓝莓贮藏品质及抗病相关酶活性的影响
1
2014
... 茉莉素(jasmonates, JA)可诱导植物体内产生凝集素和植保素等一系列具有防御功能的次生代谢物, 或者诱导抗病基因表达以提高植物的抗病性(
苹果MdWRKY33基因在轮纹病抗性形成中的作用机制研究
1
2015
... (K13424, -12.2)则编码WRKY33转录因子.本课题组在前期研究中发现, WRKY33转录因子的表达与轮纹病病程相关基因MdPRs的表达高度协同(
Overexpressing MhNPR1 in transgenic Fuji apples enhances resistance to apple powdery mildew
1
2012
... 苹果(Malus domestica)是世界上栽培面积和产量最大的果树种类之一, 其产量和品质易遭受病原菌等生物胁迫的影响, 给苹果产业带来巨大的经济损失(
Ultrafast and memory-efficient alignment of short DNA sequences to the human genome
1
2009
... 利用RNA提取试剂盒(天根, Cat No.DP424)提取样本总RNA.参考
Fast and accurate short read alignment with Burrows-Wheeler transform
1
2009
... 利用RNA提取试剂盒(天根, Cat No.DP424)提取样本总RNA.参考
The GARP/MYB-related grape transcription factor AQUILO improves cold tolerance and promotes the accumulation of raffinose family oligosaccharides
1
2018
... 利用RNA提取试剂盒(天根, Cat No.DP424)提取样本总RNA.参考
The genome of the domesticated apple ( Malus × domestica Borkh.)
1
2010
... 对照(CTRL1)和处理(MeJA)分别得到751 477 132 bp和751 612 960 bp的测序数据, 其中分别有71.28%和72.06%定位于苹果基因组, 44.23%和45.12%的序列与金冠苹果基因组(
TPS1 drug design for rice blast disease in Magnaporthe oryzae
1
2014
... 在次生代谢途径中, MeJA主要调控苹果叶片倍半萜类和三萜以及类黄酮的生物合成.涉及倍半萜类和三萜生物合成途径的基因主要调控蒎烯合成酶(pinene synthase)合成, 最主要的调控基因为MDP0000702120 (12.32, K14181).K14181编码6-磷酸海藻糖合成酶1 (trehalose-6-phosphate synthase 1, Tps1), 而Tps1可通过调节海藻糖代谢产生抗真菌抑制剂已经在水稻(Oryza sativa)中得到证实(
WEGO: a web tool for plotting GO annotations
1
2006
... 将所有差异表达基因提交到Gene Ontology数据库(http://www.geneontology.org/), 用WEGO软件(
Methyl jasmonate enhances biocontrol efficacy of Rhodotorula glutinis to postharvest blue mold decay of pears
1
2009
... 茉莉素(jasmonates, JA)可诱导植物体内产生凝集素和植保素等一系列具有防御功能的次生代谢物, 或者诱导抗病基因表达以提高植物的抗病性(
Salicylic acid confers enhanced resistance to Glomerella leaf spot in apple
1
2016
... 苹果(Malus domestica)是世界上栽培面积和产量最大的果树种类之一, 其产量和品质易遭受病原菌等生物胁迫的影响, 给苹果产业带来巨大的经济损失(
备案号: 京ICP备16067583号-21
版权所有 © 2021 《植物学报》编辑部
地址:北京香山南辛村20号 邮编:100093
电话:010-62836135 010-62836131 E-mail:cbb@ibcas.ac.cn
本系统由北京玛格泰克科技发展有限公司设计开发

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}