Abstract Plants have evolved to maintain the dormancy of freshly harvested seeds, which ensures that seeds do not germinate until environmental conditions are optimal. Therefore, dormancy helps seeds spread over long distances to ensure the survival of species. The transition from dormancy to germination is crucial to plant survival and for promoting yield and quality in agricultural production. Seed dormancy and germination are precisely regulated by diverse endogenous hormones and light signals. Light cues regulate seed dormancy and germination by affecting abscisic acid/gibberellic acid biosynthesis and signals. In this review, we summarize the key roles of the hormone pathway and light signal transduction pathways in regulating seed dormancy and germination. We also discuss the interactions (crosstalk) between phytohormone signals and light signals in seed dormancy and germination, in order to apply reference for regulating seed dormancy and germination by using light and hormones in agricultural production. Keywords:seed dormancy;seed germination;light signal;hormones;crosstalk
PDF (1575KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 杨立文, 刘双荣, 林荣呈. 光信号与激素调控种子休眠和萌发研究进展. 植物学报, 2019, 54(5): 569-581 doi:10.11983/CBB19038 Yang Liwen, Liu Shuangrong, Lin Rongcheng. Advances in Light and Hormones in Regulating Seed Dormancy and Germination. Chinese Bulletin of Botany, 2019, 54(5): 569-581 doi:10.11983/CBB19038
种子休眠是指有活力的种子在适宜的条件下暂时不萌发的现象(Bewley, 1997; Finch-Savage and Leubner-Metzger, 2006)。种子的休眠包括休眠的诱导、维持和释放3个阶段(Shu et al., 2016)。成熟的种子释放休眠后, 在适宜的条件下即可萌发。DOG1 (DELAY OF GERMINATION 1)在种子休眠诱导和维持中发挥重要作用。它既能通过抑制脱落酸(abscisic acid, ABA)通路的负调控因子——AHG1(ABA HYPERSENSITIVE GERMINATION 1)的磷酸酶活性促进种子休眠(Nishimura et al., 2018); 也能通过调控响应赤霉素(gibberellin, GA)信号的细胞壁重塑相关基因的表达促进种子休眠(Graeber et al., 2014)。在种子吸胀初期, 胚乳中PHYB (PHYTOCHROME B)吸收远红光后以生理失活型存在, 此时PIF1 (PIF3-LIKE 5, PIL5)蛋白逐渐积累, 使胚乳中ABA水平逐渐升高。胚乳中的ABA信号能够释放到胚中并抑制GA的生物合成, 使种子保持休眠状态。随着种子吸胀时间的延长, 胚乳中ABA水平逐渐下调。此时胚中的PHYA介导远红光促进GA的生物合成, 进而促进种子萌发(Lee et al., 2012)。因此, 种子的休眠和萌发是两个相互独立的生物学过程。
研究表明, 种子的休眠及萌发受到内源激素和外源光信号的共同调控(Bassel, 2016; Shu et al., 2016)。本文主要从外源光信号和内源激素对种子休眠和萌发的调控以及两类信号通路的互作3方面对模式植物拟南芥(Arabidopsis thaliana)种子休眠和萌发的调控机制进行综述, 以期为农业生产中利用光和激素调控种子休眠与萌发提供参考。
2 激素调控种子休眠和萌发
2.1 ABA-GA动态平衡是调控种子休眠和萌发的关键
激素在种子休眠和萌发过程中发挥重要调控作用。其中, ABA和GA在调控种子休眠和萌发上行使主要功能。ABA促进种子休眠, 而GA促进种子萌发(Shu et al., 2016; Née et al., 2017a)。在种子成熟过程中, ABA在种子内逐渐积累, 使种子的休眠水平逐渐升高。相比之下, 当种子受到吸胀或层积处理时, ABA含量逐渐降低, GA含量逐渐升高, 使种子萌发。与野生型种子相比, ABA合成缺陷突变体种子萌发更快(Frey et al., 2012); 而过表达ABA合成基因的种子以及ABA代谢突变体种子保持较高的休眠水平(Matakiadis et al., 2009; Martinez-Andújar et al., 2011; Nonogaki et al., 2014), 表明ABA能够通过其生物合成通路调控种子的休眠和萌发。ABA信号通过PYR/PYL/RCAR (PYRABACTIN RESISTANCE 1/PYR-LIKE PROTEINS/REGULATORY COMPONENTS OF ABA RECEPTORS)-PP2C (PROTEIN PHOSPHATASE 2C)-SnRKs (SNF1-RELATED PROTEIN KINASES)级联反应向下传递(Cutler et al., 2010; Hubbard et al., 2010)。目前鉴定到的PP2C类蛋白包括ABI1 (ABSCISIC ACID INSENSITIVE 1)、ABI2、HON (HONSU)和RDO5 (REDUCED DORMANCY 5)。其中, ABI1和ABI2通过与ABA信号受体蛋白PYR/PYL/RCAR互作进而抑制ABA信号转导(Ma et al., 2009; Park et al., 2009)。HON蛋白能够通过抑制ABA信号转导及促进GA信号转导抑制种子休眠(Kim et al., 2013), 说明HON能够整合ABA和GA信号以调控种子休眠。rdo5突变体的休眠水平降低, 但其ABA含量和ABA敏感性并未发生显著变化(Xiang et al., 2014), 说明RDO5通过不依赖ABA的途径调控种子休眠。ABI3、ABI4和ABI5是ABA信号通路下游的关键组分, 能够抑制种子萌发、促进种子休眠(Bentsink and Koornneef, 2008; Kanai et al., 2010; Shu et al., 2013)。响应ABA信号的R2R3型MYB转录因子MYB96通过调控ABI4以及ABA合成相关基因NCED2 (9-CIS-EPOXYCAROTENOID DIOXYGENASE 2)和NCED6的转录进而促进种子休眠、抑制种子萌发(Lee et al., 2015a, 2015b)。上述研究表明, ABA不仅通过其生物合成通路调控种子的休眠和萌发, 还能以ABA信号的形式发挥作用。
GA能够通过抑制ABA诱导的种子休眠促进种子萌发(Gubler et al., 2005; Graeber et al., 2012)。在植物体内, 具有生物活性的GA主要包括GA1和GA4, 它们通过软化种皮、促进胚乳层细胞的水解和胚的生长打破种子休眠, 进而促进种子萌发(Holdsworth et al., 2008)。GA缺失突变体ga1 (gibberellic acid- requiring 1)和ga2表现出强烈的休眠表型, 需要外源施加GA方可萌发(Lee et al., 2002; Shu et al., 2013)。GA20ox (GA 20-OXIDASE)和GA3ox基因编码参与GA生物合成的氧化酶; GA2ox编码的GA2-氧化酶主要参与GA的代谢过程。在PHYB-off条件下, ga2ox突变体萌发水平升高(Oh et al., 2006; Yamauchi et al., 2007)。上述研究结果表明, GA能通过其生物合成通路调控种子的休眠和萌发。在GA信号转导过程中, DELLA蛋白扮演着重要角色, 它能响应GA信号快速降解, 对GA信号转导起限速作用(Silverstone et al., 1998; Itoh et al., 2002; Zentella et al., 2007; Nemoto et al., 2017)。GA受体蛋白GID1 (GA-INSENSITIVE DWARF 1)能够负调控DELLA蛋白的稳定性, 促进GA信号转导(Davière and Achard, 2013)。拟南芥SLY1 (SLEEPY1)和水稻GID2 (GA-INSENSITIVE DWARF2)是SCF聚合体中的F-box亚基, 依赖其C端的GGF和LSL基序与DELLA蛋白C端VHIID和LHR2基序结合, 促进DELLA多聚泛素化(Hirano et al., 2010; Ariizumi et al., 2011)。研究表明, DELLA蛋白GAI (GA-INSENSITIVE)能够通过抑制TCP14 (TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR 14)和TCP15的表达减缓细胞周期的进程, 最终使种子保持休眠状态(Resentini et al., 2015)。作为转录调节子, DELLA如何实现对下游基因的转录调控? 有研究指出, DELLA能够通过与染色质重塑因子互作调控下游基因的转录。DELLA蛋白RGL2 (RGA-LIKE 2)和RGL3与染色质重塑因子SWI3C (SWITCH3C)发生互作, 进而影响GID1a以及GA3ox基因的转录(Sarnowska et al., 2013)。此外, DELLA还能通过与种子萌发相关转录因子互作进而实现其对萌发的调控。RGL2通过与转录因子NF-YC互作, 诱导下游ABI5的转录, 进而抑制种子萌发(Liu et al., 2016)。此外, RGL2还能通过与转录因子DOF6 (BINDING1 ZINC FINGER 6)互作诱导GATA12的表达, 进而促进种子休眠(Ravindran et al., 2017)。DELLA通过与ICE1 (INDUCER OF CBF EXPRESSION 1)互作解除其对ABA响应基因EM1 (LATE EMBRYOGENESIS ABUNDANT 1)和EM6的转录抑制作用, 最终抑制种子萌发(Hu et al., 2019)。
植物依赖光受体蛋白识别外源环境中的光信号。根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011)。近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展。其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录。此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路。PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018)。此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014)。在UVR8介导的信号通路中, COP1作为正调控因子发挥作用。UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017)。最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解。RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019)。此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018)。UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b)。
(A) Light signal regulates seed germination via ABA and GA pathway. PHYB regulates seed germination through promoting the degradation of PIF1 protein. The accumulation of PIF1 in nucleus activates the transcription of DAG1 and SOM, which indirectly regulates the expression of GA biosynthesis gene or directly induces RGA and GAI (DELLA protein encoding genes) transcription and leads to repressing seed germination. Similarly, PIF1 stimulates ABA biosynthesis and ABA signaling pathway to suppress seed germination. PIF1 induces ABA biosynthesis via SOM-dependent pathway to repress seed germination; or it induces the transcription of ABI3 and ABI5 in order to stimulate ABA signaling. Expect for PIF1, PHYB also promotes seed germination via inhibiting RVE1 transcription which indirectly promote GA biosynthesis. SPT and CSN complex could regulate seed germination in an ABI5-dependent manner. SPT suppresses ABI5 transcript to destroy ABA pathway. CSN1 stimulates RGL2 degradation to inhibit ABI5 activity, while CSN5a directly decreases the accumulation of ABI5 in order to provoke seed germination. JAZ3 promotes seed germination by repressing the transcriptional activity of ABI5 which activates ABA-responsing gene EM1 expression. (B) Light signal controls seed dormancy via ABA and GA pathway. PHYB mediates red light to repress the transcription level of GA3ox2, inhibiting seed dormancy. SPT plays different roles in regulating seed dormancy under Col and Ler background of Arabidopsis. SPT promotes seed dormancy through activating the expression of RGL3 and ABI5 under Col background (green line), however, SPT suppresses seed dormancy via inhibiting RGA and ABI4 transcript under Ler background (red line). In addition, PIF6 is also involved in controlling seed dormancy.
Alonso-BlancoC, BentsinkL, HanhartCJ, Blankestijn-de VriesH, KoornneefM (2003). Analysis of natural allelic variation at seed dormancy loci of Arabidopsis thaliana Genetics 164, 711-729. [本文引用: 2]
ArgyrisJ, TrucoMJ, OchoaO, KnappSJ, StillDW, KnappSJ, StillDW, LenssenGM, SchutJW, MichelmoreRW, BradfordKJ (2005). Quantitative trait loci associated with seed and seedling traits in Lactuca Theor Appl Genet 111, 1365-1376. [本文引用: 1]
AriizumiT, LawrencePK, SteberCM (2011). The role of two F-box proteins, SLEEPY1 and SNEEZY, in Arabidopsis gibberellin signaling Plant Physiol 155, 765-775. [本文引用: 1]
BasselGW (2016). To grow or not to grow? Trends Plant Sci 21, 498-505. [本文引用: 1]
BentsinkL, JowettJ, HanhartCJ, KoornneefM (2006). Cloning of DOG1, a quantitative trait locus controlling seed dormancy in Arabidopsis Proc Natl Acad Sci USA 103, 17042-17047. [本文引用: 1]
BentsinkL, KoornneefM (2008). Seed dormancy and germination Arabidopsis Book 6, e0119. [本文引用: 1]
BriggsWR, HualaE (1999). Blue-light photoreceptors in higher plants Annu Rev Cell Dev Biol 15, 33-62. [本文引用: 1]
CashmoreAR (2003). Cryptochromes: enabling plants and animals to determine circadian time Cell 114, 537-543. [本文引用: 1]
ChenF, LiBS, LiG, CharronJB, DaiMQ, ShiXR, DengXW (2014). Arabidopsis phytochrome A directly targets numerous promoters for individualized modulation of genes in a wide range of pathways Plant Cell 26, 1949-1966. [本文引用: 1]
ChengWH, ChiangMH, HwangSG, LinPC (2009). Antagonism between abscisic acid and ethylene in Arabidopsis acts in parallel with the reciprocal regulation of their metabolism and signaling pathways Plant Mol Biol 71, 61-80. [本文引用: 1]
ClackT, MathewsS, SharrockRA (1994). The phytochrome apoprotein family in Arabidopsis is encoded by five genes: the sequences and expression of PHYD and PHYE Plant Mol Biol 25, 413-427. [本文引用: 1]
CorbineauF, XiaQ, BaillyC, El-Maarouf-BouteauH (2014). Ethylene, a key factor in the regulation of seed dormancy Front Plant Sci 5, 539. [本文引用: 1]
CutlerSR, RodriguezPL, FinkelsteinRR, AbramsSR (2010). Abscisic acid: emergence of a core signaling network Annu Rev Plant Biol 61, 651-679. [本文引用: 1]
DavièreJM, AchardP (2013). Gibberellin signaling in plants Development 140, 1147-1151. [本文引用: 1]
de WitM, Galv?oVC, FankhauserC (2016). Light-mediated hormonal regulation of plant growth and development Annu Rev Plant Biol 67, 513-537. [本文引用: 2]
DuqueP, ChuaNH (2003). IMB1, a bromodomain protein induced during seed imbibition, regulates ABA- and phyA- mediated responses of germination in Arabidopsis Plant J 35, 787-799. [本文引用: 3]
Finch-SavageWE, Leubner-MetzgerG (2006). Seed dormancy and the control of germination New Phytol 171, 501-523. [本文引用: 2]
FreyA, EffroyD, LefebvreV, SeoM, PerreauF, BergerA, SechetJ, ToA, NorthHM, Marion-PollA (2012). Epoxycarotenoid cleavage by NCED5 fine-tunes ABA accumulation and affects seed dormancy and drought tolerance with other NCED family members Plant J 70, 501-512. [本文引用: 1]
GabrieleS, RizzaA, MartoneJ, CircelliP, CostantinoP, VittoriosoP (2010). The Dof protein DAG1 mediates PIL5 activity on seed germination by negatively regulating GA biosynthetic gene AtGA3ox1 Plant J 61, 312-323. [本文引用: 2]
GraeberK, LinkiesA, SteinbrecherT, MummenhoffK, TarkowskáD, Ture?kováV, IgnatzM, SperberK, VoegeleA, de JongH, UrbanováT, StrnadM, Leubner-MetzgerG (2014). DELAY OF GERMINATION 1 mediates a conserved coat-dormancy mechanism for the temperature- and gibberellin-dependent control of seed germination Proc Natl Acad Sci USA 111, E3571-E3580. [本文引用: 2]
GraeberK, NakabayashiK, MiattonE, Leubner-MetzgerG, SoppeWJJ (2012). Molecular mechanisms of seed dormancy Plant Cell Environ 35, 1769-1786. [本文引用: 1]
GuXY, KianianSF, FoleyME (2006). Dormancy genes from weedy rice respond divergently to seed development environments Genetics 172, 1199-1211. [本文引用: 1]
GuanCM, WangXC, FengJ, HongSL, LiangY, RenB, ZuoJ (2014). Cytokinin antagonizes abscisic acidmediated inhibition of cotyledon greening by promoting the degradation of abscisic acid insensitive5 protein in Arabidopsis Plant Physiol 164, 1515-1526. [本文引用: 1]
GublerF, MillarAA, JacobsenJV (2005). Dormancy release, ABA and pre-harvest sprouting Curr Opin Plant Biol 8, 183-187. [本文引用: 1]
HardtkeCS, GohdaK, OsterlundMT, OyamaT, OkadaK, DengXW (2000). HY5 stability and activity in Arabidopsis is regulated by phosphorylation in its COP1 binding domain EMBO J 19, 4997-5006. [本文引用: 1]
HennigL, StoddartWM, DieterleM, WhitelamGC, Sch?ferE (2002) Phytochrome E controls light-induced germination of Arabidopsis Plant Physiol 128, 194-200. [本文引用: 3]
HiranoK, AsanoK, TsujiH, KawamuraM, MoriH, KitanoH, Ueguchi-TanakaM, MatsuokaM (2010). Characterization of the molecular mechanism underlying gibberellin perception complex formation in rice Plant Cell 22, 2680-2696. [本文引用: 1]
HoldsworthMJ, BentsinkL, SoppeWJJ (2008). Molecular networks regulating Arabidopsis seed maturation, afterripening, dormancy and germination New Phytol 179, 33-54. [本文引用: 1]
HuYR, HanX, YangML, ZhangMH, PanJJ, YuDQ (2019). The transcription factor INDUCER OF CBF EXPRESSION 1 interacts with ABSCISIC ACID INSENSITIVE 5 and DELLA proteins to fine-tune abscisic acid signaling during seed germination in Arabidopsis Plant Cell 31, 1520-1538. [本文引用: 1]
HuYR, YuDQ (2014). BRASSINOSTEROID INSENSITIVE 2 interacts with ABSCISIC ACID INSENSITIVE 5 to mediate the antagonism of brassinosteroids to abscisic acid during seed germination in Arabidopsis Plant Cell 26, 4394-4408. [本文引用: 1]
HuangX, OuyangXH, YangPY, LauOS, ChenLB, WeiN, DengXW (2013). Conversion from CUL4-based COP1- SPA E3 apparatus to UVR8-COP1-SPA complexes underlies a distinct biochemical function of COP1 under UV-B Proc Natl Acad Sci USA 110, 16669-16674. [本文引用: 1]
HubbardKE, NishimuraN, HitomiK, GetzoffED, SchroederJI (2010). Early abscisic acid signal transduction mechanisms: newly discovered components and newly emerging questions Genes Dev 24, 1695-1708. [本文引用: 1]
ItohH, Ueguchi-TanakaM, SatoY, AshikariM, MatsuokaM (2002). The gibberellin signaling pathway is regulated by the appearance and disappearance of SLENDER RICE1 in nuclei Plant Cell 14, 57-70. [本文引用: 1]
JacobsenJV, BarreroJM, HughesT, JulkowskaM, TaylorJM, XuQ, GublerF (2013). Roles for blue light, jasmonate and nitric oxide in the regulation of dormancy and germination in wheat grain ( Triticum aestivum L.) Planta 238, 121-138. [本文引用: 1]
JangIC, YangJY, SeoHS, ChuaNH (2005). HFR1 is targeted by COP1 E3 ligase for post-translational proteolysis during phytochrome A signaling Genes Dev 19, 593-602. [本文引用: 1]
JangK, LeeHG, JungSJ, PaekNC, SeoPJ (2015). The E3 ubiquitin ligase COP1 regulates thermosensory flowering by triggering GI degradation in Arabidopsis Sci Rep 5, 12071. [本文引用: 1]
JiangZM, XuG, JingYJ, TangWJ, LinRC (2016). Phytochrome B and REVEILLE1/2-mediated signaling controls seed dormancy and germination in Arabidopsis Nat Commun 7, 12377. [本文引用: 4]
JinD, WuM, LiBS, BückerB, KeilP, ZhangSM, LiJG, KangDM, LiuJ, DongJ, DengXW, IrishV, WeiN (2018). The COP9 signalosome regulates seed germination by facilitating protein degradation of RGL2 and ABI5 PLoS Genet 14, e1007237. [本文引用: 2]
JuL, JingYX, ShiPT, LiuJ, ChenJS, YanJJ, ChuJF, ChenKM, SunJQ (2019). JAZ proteins modulate seed germination through interaction with ABI5 in bread wheat and Arabidopsis New Phytol 223, 246-260. [本文引用: 1]
KanaiM, NishimuraM, HayashiM (2010). A peroxisomal ABC transporter promotes seed germination by inducing pectin degradation under the control of ABI5 Plant J 62, 936-947. [本文引用: 1]
KimDH, YamaguchiS, LimS, OhE, ParkJ, HanadaA, KamiyaY, ChoiG (2008). SOMNUS, a CCCH-type zinc finger protein in Arabidopsis, negatively regulates light- dependent seed germination downstream of PIL5 Plant Cell 20, 1260-1277. [本文引用: 2]
KimW, LeeY, ParkJ, LeeN, ChoiG (2013). HONSU, a protein phosphatase 2C, regulates seed dormancy by inhibiting ABA signaling in Arabidopsis Plant Cell Physiol 54, 555-572. [本文引用: 1]
LeeHG, LeeK, SeoPJ (2015a). The Arabidopsis MYB96 transcription factor plays a role in seed dormancy Plant Mol Biol 87, 371-381. [本文引用: 1]
LeeK, LeeHG, YoonS, KimHU, SeoPJ (2015b). The Arabidopsis MYB96 transcription factor is a positive regulator of ABSCISIC ACID-INSENSITIVE 4 in the control of seed germination Plant Physiol 168, 677-689. [本文引用: 1]
LeeKP, PiskurewiczU, Ture?kováV, CaratS, ChappuisR, StrnadM, FankhauserC, Lopez-MolinaL (2012). Spatially and genetically distinct control of seed germination by phytochromes A and B Genes Dev 26, 1984-1996. [本文引用: 1]
LeeS, ChengH, KingKE, WangW, HeYW, HussainA, LoJ, HarberdNP, PengJR (2002). Gibberellin regulates Arabidopsis seed germination via RGL2, a GAIIRGA-like gene whose expression is up-regulated following imbibition Genes Dev 16, 646-658. [本文引用: 1]
LeeS, KimSG, ParkCM (2010). Salicylic acid promotes seed germination under high salinity by modulating antioxidant activity in Arabidopsis New Phytol 188, 626-637. [本文引用: 1]
LiXY, ChenTT, LiY, WangZ, CaoH, ChenFY, LiY, SoppeWJJ, LiWL, LiuYX (2019). ETR1/RDO3 regulates seed dormancy by relieving the inhibitory effect of the ERF12-TPL complex on DELAY OF GERMINATION 1 expression Plant Cell 31, 832-847. [本文引用: 1]
LiangT, MeiSL, ShiC, YangY, PengY, MaLB, WangF, LiX, HuangX, YinYH, LiuHT (2018). UVR8 interacts with BES1 and BIM1 to regulate transcription and photomorphogenesis in Arabidopsis Dev Cell 44, 512-523. [本文引用: 1]
LinCT, ShalitinD (2003). Cryptochrome structure and signal transduction Annu Rev Plant Biol 54, 469-496. [本文引用: 1]
LinkiesA, MüllerK, MorrisK, Ture?kováV, WenkM, CadmanCSC, CorbineauF, StrnadM, LynnJR, Finch-SavageWE, Leubner-MetzgerG (2009). Ethylene interacts with abscisic acid to regulate endosperm rupture during germination: a comparative approach using Lepidium sativum and Arabidopsis thaliana Plant Cell 21, 3803-3822. [本文引用: 1]
LiuHT, YuXH, LiKW, KlejnotJ, YangHY, LisieroD, LinCT (2008a). Photoexcited CRY2 interacts with CIB1 to regulate transcription and floral initiation in Arabidopsis Science 322, 1535-1539. [本文引用: 1]
LiuLJ, ZhangYC, LiQH, SangY, MaoJ, LianHL, WangL, YangHQ (2008b). COP1-mediated ubiquitination of CONSTANS is implicated in cryptochrome regulation of flowering in Arabidopsis Plant Cell 20, 292-306. [本文引用: 1]
LiuX, HuP, HuangM, TangY, LiY, LiL, HouX (2016). The NF-YC-RGL2 module integrates GA and ABA signaling to regulate seed germination in Arabidopsis Nat Commun 7, 12768. [本文引用: 1]
LiuX, WangJ, YuY, KongLN, LiuYM, LiuZQ, LiHY, WeiPW, LiuML, ZhouH, BuQY, FangJ (2019). Identification and characterization of the rice pre-harvest sprouting mutants involved in molybdenum cofactor biosynthesis New Phytol 222, 275-285. [本文引用: 1]
LiuXD, ZhangH, ZhaoY, FengZY, LiQ, YangHQ, LuanS, LiJM, HeZH (2013a). Auxin controls seed dormancy through stimulation of abscisic acid signaling by inducing ARF-mediated ABI3 activation in Arabidopsis Proc Natl Acad Sci USA 110, 15485-15490. [本文引用: 2]
LiuYW, LiX, LiKW, LiuHT, LinCT (2013b). Multiple bHLH proteins form heterodimers to mediate CRY2-dependent regulation of flowering-time in Arabidopsis PLoS Genet 9, e1003861. [本文引用: 1]
LuoQ, LianHL, HeSB, LiL, JiaKP, YangHQ (2014). COP1 and phyB physically interact with PIL1 to regulate its stability and photomorphogenic development in Arabidopsis Plant Cell 26, 2441-2456. [本文引用: 1]
MaDB, LiX, GuoYX, ChuJF, FangS, YanCY, NoelJP, LiuHT (2016). Cryptochrome 1 interacts with PIF4 to regulate high temperature-mediated hypocotyl elongation in response to blue light Proc Natl Acad Sci USA 113, 224-229. [本文引用: 1]
MaY, SzostkiewiczI, KorteA, MoesD, YangY, ChristmannA, GrillE (2009). Regulators of PP2C phosphatase activity function as abscisic acid sensors Science 324, 1064-1068. [本文引用: 1]
Martinez-AndújarC, OrdizMI, HuangZL, NonogakiM, BeachyRN, NonogakiH (2011). Induction of 9- cis-epoxycarotenoid dioxygenase in Arabidopsis thaliana seeds enhances seed dormancy Proc Natl Acad Sci USA 108, 17225-17229. [本文引用: 1]
MatakiadisT, AlboresiA, JikumaruY, TatematsuK, PichonO, RenouJP, KamiyaY, NambaraE, TruongHN (2009). The Arabidopsis abscisic acid catabolic gene CYP707A2 plays a key role in nitrate control of seed dormancy Plant Physiol 149, 949-960. [本文引用: 1]
NambaraE, OkamotoM, TatematsuK, YanoR, SeoM, KamiyaY (2010). Abscisic acid and the control of seed dormancy and germination Seed Sci Res 20, 55-67. [本文引用: 1]
NéeG, KramerK, NakabayashiK, YuanBJ, XiangY, MiattonE, FinkemeierI, SoppeWJJ (2017b). DELAY OF GERMINATION 1 requires PP2C phosphatases of the ABA signaling pathway to control seed dormancy Nat Commun 8, 72. [本文引用: 2]
NéeG, XiangY, SoppeWJJ (2017a). The release of dormancy, a wake-up call for seeds to germinate Curr Opin Plant Biol 35, 8-14. [本文引用: 1]
NemotoK, RamadanA, ArimuraGI, ImaiK, TomiiK, ShinozakiK, SawasakiT (2017). Tyrosine phosphorylation of the GARU E3 ubiquitin ligase promotes gibberellin signaling by preventing GID1 degradation Nat Commun 8, 1004. [本文引用: 1]
NishimuraN, TsuchiyaW, MorescoJJ, HayashiY, SatohK, KaiwaN, IrisaT, KinoshitaT, SchroederJI, Yates IIIJR, HirayamaT, YamazakiT (2018). Control of seed dormancy and germination by DOG1-AHG1 PP2C phosphatase complex via binding to heme Nat Commun 9, 2132. [本文引用: 2]
NonogakiM, SallK, NambaraE, NonogakiH (2014). Amplification of ABA biosynthesis and signaling through a positive feedback mechanism in seeds Plant J 78, 527-539. [本文引用: 1]
OhE, KimJ, ParkE, KimJI, KangC, ChoiG (2004). PIL5, a phytochrome-interacting basic helix-loop-helix protein, is a key negative regulator of seed germination in Arabidopsis thaliana Plant Cell 16, 3045-3058. [本文引用: 1]
OhE, YamaguchiS, HuJH, YusukeJ, JungB, PaikI, LeeHS, SunTP, KamiyaY, ChoiG (2007). PIL5, a phytochrome-interacting bHLH protein, regulates gibberellin responsiveness by binding directly to the GAI and RGA promoters in Arabidopsis seeds Plant Cell 19, 1192-1208. [本文引用: 2]
OhE, YamaguchiS, KamiyaY, BaeG, ChungWI, ChoiG (2006). Light activates the degradation of PIL5 protein to promote seed germination through gibberellin in Arabidopsis Plant J 47, 124-139. [本文引用: 5]
ParkJ, LeeN, KimW, LimS, ChoiG (2011). ABI3 and PIL5 collaboratively activate the expression of SOMNUS by directly binding to its promoter in imbibed Arabidopsis seeds Plant Cell 23, 1404-1415. [本文引用: 1]
ParkSY, FungP, NishimuraN, JensenDR, FujiiH, ZhaoY, LumbaS, SantiagoJ, RodriguesA, ChowTFF, AlfredSE, BonettaD, FinkelsteinR, ProvartNJ, DesveauxD, RodriguezPL, McCourtP, ZhuJK, SchroederJI, VolkmanBF, CutlerSR (2009). Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins Science 324, 1068-1071. [本文引用: 1]
PedmaleUV, HuangSSC, ZanderM, ColeBJ, HetzelJ, LjungK, ReisPAB, SrideviP, NitoK, NeryJR, EckerJR, ChoryJ (2016). Cryptochromes interact directly with PIFs to control plant growth in limiting blue light Cell 164, 233-245. [本文引用: 1]
PenfieldS, HallA (2009). A role for multiple circadian clock genes in the response to signals that break seed dormancy in Arabidopsis Plant Cell 21, 1722-1732. [本文引用: 1]
PenfieldS, JosseEM, HallidayKJ (2010). A role for an alternative splice variant of PIF6 in the control of Arabidopsis primary seed dormancy Plant Mol Biol 73, 89-95. [本文引用: 1]
QuailPH, BoylanMT, ParksBM, ShortTW, XuY, WagnerD (1995). Phytochromes: photosensory perception and signal transduction Science 268, 675-680. [本文引用: 1]
RavindranP, VermaV, StammP, KumarPP (2017). A novel RGL2-DOF6 complex contributes to primary seed dormancy in Arabidopsis thaliana by regulating a GATA transcription factor Mol Plant 10, 1307-1320. [本文引用: 1]
RenH, HanJP, YangPY, MaoWW, LiuX, QiuLL, QianCZ, LiuY, ChenZR, OuyangXH, ChenX, DengXW, HuangX (2019). Two E3 ligases antagonistically regulate the UV-B response in Arabidopsis Proc Natl Acad Sci USA 116, 4722-4731. [本文引用: 1]
ResentiniF, Felipo-BenaventA, ColomboL, BlázquezMA, AlabadíD, MasieroS (2015). TCP14 and TCP15 mediate the promotion of seed germination by gibberellins in Arabidopsis thaliana Mol Plant 8, 482-485. [本文引用: 1]
RizziniL, FavoryJJ, CloixC, FaggionatoD, O’HaraA, KaiserliE, BaumeisterR, Sch?ferE, NagyF, JenkinsGI, UlmR (2011). Perception of UV-B by the Arabidopsis UVR8 protein Science 332, 103-106. [本文引用: 1]
SeoHS, YangJY, IshikawaM, BolleC, BallesterosML, ChuaNH (2003). LAF1 ubiquitination by COP1 controls photomorphogenesis and is stimulated by SPA1 Nature 423, 995-999. [本文引用: 1]
SeoM, NambaraE, ChoiG, YamaguchiS (2009). Interaction of light and hormone signals in germinating seeds Plant Mol Biol 69, 463-472. [本文引用: 1]
SharrockRA, QuailPH (1989). Novel phytochrome sequences in Arabidopsis thaliana: structure, evolution, and differential expression of a plant regulatory photoreceptor family Gene Dev 3, 1745-1757. [本文引用: 1]
ShiH, WangX, MoXR, TangC, ZhongSW, DengXW (2015). Arabidopsis DET1 degrades HFR1 but stabilizes PIF1 to precisely regulate seed germination Proc Natl Acad Sci USA 112, 3817-3822. [本文引用: 1]
ShiH, ZhongSW, MoXR, LiuN, NezamesCD, DengXW (2013). HFR1 sequesters PIF1 to govern the transcriptional network underlying light-initiated seed germination in Arabidopsis Plant Cell 25, 3770-3784. [本文引用: 1]
ShinomuraT, NagataniA, ChoryJ, FuruyaM (1994). The induction of seed germination in Arabidopsis thaliana is regulated principally by Phytochrome B and secondarily by Phytochrome A Plant Physiol 104, 363-371. [本文引用: 1]
ShinomuraT, NagataniA, HanzawaH, KubotaM, WatanabeM, FuruyaM (1996). Action spectra for phytochrome A- and B-specific photoinduction of seed germination in Arabidopsis thaliana Proc Natl Acad Sci USA 93, 8129-8133. [本文引用: 1]
ShuK, LiuXD, XieQ, HeZH (2016). Two faces of one seed: hormonal regulation of dormancy and germination Mol Plant 9, 34-45. [本文引用: 4]
ShuK, MengYJ, ShuaiHW, LiuWG, DuJB, LiuJ, YangWY (2015). Dormancy and germination: how does the crop seed decide? Plant Biol 17, 1104-1112. [本文引用: 1]
ShuK, ZhangHW, WangSF, ChenML, WuYR, TangSY, LiuCY, FengYQ, CaoXF, XieQ (2013). ABI4 regulates primary seed dormancy by regulating the biogenesis of abscisic acid and gibberellins in Arabidopsis PLoS Genet 9, e1003577. [本文引用: 2]
SilverstoneAL, CiampaglioCN, SunTP (1998). The Arabidopsis RGA gene encodes a transcriptional regulator repressing the gibberellin signal transduction pathway Plant Cell 10, 155-169. [本文引用: 1]
SimsekS, OhmJB, LuHY, RuggM, BerzonskyW, AlamriMS, MergoumM (2014). Effect of pre-harvest sprouting on physicochemical changes of proteins in wheat J Sci Food Agric 94, 205-212. [本文引用: 1]
StangaJP, SmithSM, BriggsWR, NelsonDC (2013). SUPPRESSOR OF MORE AXILLARY GROWTH2 1 controls seed germination and seedling development in Arabidopsis Plant Physiol 163, 318-330. [本文引用: 1]
TohS, KamiyaY, KawakamiN, NambaraE, McCourtP, TsuchiyaY (2012). Thermoinhibition uncovers a role for strigolactones in Arabidopsis seed germination Plant Cell Physiol 53, 107-117. [本文引用: 1]
VaistijFE, Barros-Galv?oT, ColeAF, GildayAD, HeZS, LiY, HarveyD, LarsonTR, GrahamlA (2018). MOTHER-OF-FT-AND-TFL1 represses seed germination under far-red light by modulating phytohormone responses in Arabidopsis thaliana Proc Natl Acad Sci USA 115, 8442-8447. [本文引用: 2]
VaistijFE, GanYB, PenfieldS, GildayAD, DaveA, HeZS, JosseEM, ChoiG, HallidayKJ, GrahamIA (2013). Differential control of seed primary dormancy in Arabidopsis ecotypes by the transcription factor SPATULA Proc Natl Acad Sci USA 110, 10866-10871. [本文引用: 2]
WangYP, LiL, YeTT, ZhaoSJ, LiuZ, FengYQ, WuY (2011). Cytokinin antagonizes ABA suppression to seed germination of Arabidopsis by downregulating ABI5 expression Plant J 68, 249-261. [本文引用: 1]
WeiN, DengXW (2003). The COP9 signalosome Annu Rev Cell Dev Biol 19, 261-286. [本文引用: 1]
WilsonRL, KimH, BakshiA, BinderBM (2014). The ethylene receptors ETHYLENE RESPONSE 1 and ETHYLENE RESPONSE 2 have contrasting roles in seed germination of Arabidopsis during salt stress Plant Physiol 165, 1353-1366. [本文引用: 1]
XiWY, LiuC, HouXL, YuH (2010). MOTHER OF FT AND TFL1 regulates seed germination through a negative feedback loop modulating ABA signaling in Arabidopsis Plant Cell 22, 1733-1748. [本文引用: 1]
XiWY, YuH (2010). Mother of FT and TFL1 regulates seed germination and fertility relevant to the brassinosteroid signaling pathway Plant Signal Behav 5, 1315-1317. [本文引用: 1]
XiangY, NakabayashiK, DingJ, HeF, BentsinkL, SoppeWJJ (2014). REDUCED DORMANCY 5 encodes a protein phosphatase 2C that is required for seed dormancy in Arabidopsis Plant Cell 26, 4362-4375. [本文引用: 1]
XieZ, ZhangZL, HanzlikS, CookE, ShenQJ (2007). Salicylic acid inhibits gibberellin-induced alpha-amylase expression and seed germination via a pathway involving an abscisic-acid-inducible WRKY gene Plant Mol Biol 64, 293-303. [本文引用: 1]
XuF, HeSB, ZhangJY, MaoZL, WangWX, LiT, HuaJ, DuSS, XuPB, LiL, LianHL, YangHQ (2018). Photoactivated CRY1 and phyB interact directly with AUX/IAA proteins to inhibit auxin signaling in Arabidopsis Mol Plant 11, 523-541. [本文引用: 1]
YamauchiY, Takeda-KamiyaN, HanadaA, OgawaM, KuwaharaA, SeoM, KamiyaY, YamaguchiS (2007). Contribution of gibberellin deactivation by AtGA2ox2 to the suppression of germination of dark-imbibed Arabidopsis thaliana seeds Plant Cell Physiol 48, 555-561. [本文引用: 1]
YangCW, XieFM, JiangYP, LiZ, HuangX, LiL (2018a). Phytochrome a negatively regulates the shade avoidance response by increasing auxin/indole acidic acid protein stability Dev Cell 44, 29-41. [本文引用: 1]
YangY, LiangT, ZhangLB, ShaoK, GuXX, ShangRX, ShiN, LiX, ZhangP, LiuHT (2018b). UVR8 interacts with WRKY36 to regulate HY5 transcription and hypocotyl elongation in Arabidopsis Nat Plants 4, 98-107. [本文引用: 1]
ZentellaR, ZhangZL, ParkM, ThomasSG, EndoA, MuraseK, FleetCM, JikumaruY, NambaraE, KamiyaY, SunTP (2007). Global analysis of DELLA direct targets in early gibberellin signaling in Arabidopsis Plant Cell 19, 3037-3057. [本文引用: 1]
我国植物光信号转导研究进展概述 1 2017
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
光受体介导信号转导调控植物开花研究进展 1 2019
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
Analysis of natural allelic variation at seed dormancy loci of Arabidopsis thaliana 2 2003
... 与野生种相比, 经过驯化的栽培种种子休眠水平下调, 如水稻(Oryza sativa)、小麦(Triticum aestivum)、大麦(Hordeum vulgare)和玉米(Zea mays)等禾本科作物(Bewley and Black, 1994).近年来, 育种学家们利用遗传学或组学的方法鉴定到一些与种子休眠相关的位点、转录本或蛋白(Alonso-Blanco et al., 2003; Argyris et al., 2005; Gu et al., 2006; Née et al., 2017b).例如, 研究者利用QTL方法鉴定到调控种子休眠的关键基因——DOG1 (Alonso-Blanco et al., 2003; Bentsink et al., 2006).后续研究发现, DOG1能够参与调控ABA信号通路以及GA的生物合成, 进而调控种子休眠(Graeber et al., 2014; Nishimura et al., 2018).那么, DOG1能否参与调控GA信号通路进而促进种子休眠? DOG1是否受到外源光信号和内源激素的调控? 针对上述问题的研究不仅有助于人们更加清晰地理解种子如何监测环境因子的变化, 从而决定是否完成从休眠到萌发的发育转变; 还为利用光信号或激素人工调控种子的休眠和萌发, 解决农业生产中“胎萌”以及穗发芽等问题提供了理论支撑. ...
... (Alonso-Blanco et al., 2003; Bentsink et al., 2006).后续研究发现, DOG1能够参与调控ABA信号通路以及GA的生物合成, 进而调控种子休眠(Graeber et al., 2014; Nishimura et al., 2018).那么, DOG1能否参与调控GA信号通路进而促进种子休眠? DOG1是否受到外源光信号和内源激素的调控? 针对上述问题的研究不仅有助于人们更加清晰地理解种子如何监测环境因子的变化, 从而决定是否完成从休眠到萌发的发育转变; 还为利用光信号或激素人工调控种子的休眠和萌发, 解决农业生产中“胎萌”以及穗发芽等问题提供了理论支撑. ...
Quantitative trait loci associated with seed and seedling traits in Lactuca 1 2005
... 与野生种相比, 经过驯化的栽培种种子休眠水平下调, 如水稻(Oryza sativa)、小麦(Triticum aestivum)、大麦(Hordeum vulgare)和玉米(Zea mays)等禾本科作物(Bewley and Black, 1994).近年来, 育种学家们利用遗传学或组学的方法鉴定到一些与种子休眠相关的位点、转录本或蛋白(Alonso-Blanco et al., 2003; Argyris et al., 2005; Gu et al., 2006; Née et al., 2017b).例如, 研究者利用QTL方法鉴定到调控种子休眠的关键基因——DOG1 (Alonso-Blanco et al., 2003; Bentsink et al., 2006).后续研究发现, DOG1能够参与调控ABA信号通路以及GA的生物合成, 进而调控种子休眠(Graeber et al., 2014; Nishimura et al., 2018).那么, DOG1能否参与调控GA信号通路进而促进种子休眠? DOG1是否受到外源光信号和内源激素的调控? 针对上述问题的研究不仅有助于人们更加清晰地理解种子如何监测环境因子的变化, 从而决定是否完成从休眠到萌发的发育转变; 还为利用光信号或激素人工调控种子的休眠和萌发, 解决农业生产中“胎萌”以及穗发芽等问题提供了理论支撑. ...
The role of two F-box proteins, SLEEPY1 and SNEEZY, in Arabidopsis gibberellin signaling 1 2011
... GA能够通过抑制ABA诱导的种子休眠促进种子萌发(Gubler et al., 2005; Graeber et al., 2012).在植物体内, 具有生物活性的GA主要包括GA1和GA4, 它们通过软化种皮、促进胚乳层细胞的水解和胚的生长打破种子休眠, 进而促进种子萌发(Holdsworth et al., 2008).GA缺失突变体ga1 (gibberellic acid- requiring 1)和ga2表现出强烈的休眠表型, 需要外源施加GA方可萌发(Lee et al., 2002; Shu et al., 2013).GA20ox (GA 20-OXIDASE)和GA3ox基因编码参与GA生物合成的氧化酶; GA2ox编码的GA2-氧化酶主要参与GA的代谢过程.在PHYB-off条件下, ga2ox突变体萌发水平升高(Oh et al., 2006; Yamauchi et al., 2007).上述研究结果表明, GA能通过其生物合成通路调控种子的休眠和萌发.在GA信号转导过程中, DELLA蛋白扮演着重要角色, 它能响应GA信号快速降解, 对GA信号转导起限速作用(Silverstone et al., 1998; Itoh et al., 2002; Zentella et al., 2007; Nemoto et al., 2017).GA受体蛋白GID1 (GA-INSENSITIVE DWARF 1)能够负调控DELLA蛋白的稳定性, 促进GA信号转导(Davière and Achard, 2013).拟南芥SLY1 (SLEEPY1)和水稻GID2 (GA-INSENSITIVE DWARF2)是SCF聚合体中的F-box亚基, 依赖其C端的GGF和LSL基序与DELLA蛋白C端VHIID和LHR2基序结合, 促进DELLA多聚泛素化(Hirano et al., 2010; Ariizumi et al., 2011).研究表明, DELLA蛋白GAI (GA-INSENSITIVE)能够通过抑制TCP14 (TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR 14)和TCP15的表达减缓细胞周期的进程, 最终使种子保持休眠状态(Resentini et al., 2015).作为转录调节子, DELLA如何实现对下游基因的转录调控? 有研究指出, DELLA能够通过与染色质重塑因子互作调控下游基因的转录.DELLA蛋白RGL2 (RGA-LIKE 2)和RGL3与染色质重塑因子SWI3C (SWITCH3C)发生互作, 进而影响GID1a以及GA3ox基因的转录(Sarnowska et al., 2013).此外, DELLA还能通过与种子萌发相关转录因子互作进而实现其对萌发的调控.RGL2通过与转录因子NF-YC互作, 诱导下游ABI5的转录, 进而抑制种子萌发(Liu et al., 2016).此外, RGL2还能通过与转录因子DOF6 (BINDING1 ZINC FINGER 6)互作诱导GATA12的表达, 进而促进种子休眠(Ravindran et al., 2017).DELLA通过与ICE1 (INDUCER OF CBF EXPRESSION 1)互作解除其对ABA响应基因EM1 (LATE EMBRYOGENESIS ABUNDANT 1)和EM6的转录抑制作用, 最终抑制种子萌发(Hu et al., 2019). ...
To grow or not to grow? 1 2016
... 研究表明, 种子的休眠及萌发受到内源激素和外源光信号的共同调控(Bassel, 2016; Shu et al., 2016).本文主要从外源光信号和内源激素对种子休眠和萌发的调控以及两类信号通路的互作3方面对模式植物拟南芥(Arabidopsis thaliana)种子休眠和萌发的调控机制进行综述, 以期为农业生产中利用光和激素调控种子休眠与萌发提供参考. ...
Cloning of DOG1, a quantitative trait locus controlling seed dormancy in Arabidopsis 1 2006
... 与野生种相比, 经过驯化的栽培种种子休眠水平下调, 如水稻(Oryza sativa)、小麦(Triticum aestivum)、大麦(Hordeum vulgare)和玉米(Zea mays)等禾本科作物(Bewley and Black, 1994).近年来, 育种学家们利用遗传学或组学的方法鉴定到一些与种子休眠相关的位点、转录本或蛋白(Alonso-Blanco et al., 2003; Argyris et al., 2005; Gu et al., 2006; Née et al., 2017b).例如, 研究者利用QTL方法鉴定到调控种子休眠的关键基因——DOG1 (Alonso-Blanco et al., 2003; Bentsink et al., 2006).后续研究发现, DOG1能够参与调控ABA信号通路以及GA的生物合成, 进而调控种子休眠(Graeber et al., 2014; Nishimura et al., 2018).那么, DOG1能否参与调控GA信号通路进而促进种子休眠? DOG1是否受到外源光信号和内源激素的调控? 针对上述问题的研究不仅有助于人们更加清晰地理解种子如何监测环境因子的变化, 从而决定是否完成从休眠到萌发的发育转变; 还为利用光信号或激素人工调控种子的休眠和萌发, 解决农业生产中“胎萌”以及穗发芽等问题提供了理论支撑. ...
Seed dormancy and germination 1 2008
... 激素在种子休眠和萌发过程中发挥重要调控作用.其中, ABA和GA在调控种子休眠和萌发上行使主要功能.ABA促进种子休眠, 而GA促进种子萌发(Shu et al., 2016; Née et al., 2017a).在种子成熟过程中, ABA在种子内逐渐积累, 使种子的休眠水平逐渐升高.相比之下, 当种子受到吸胀或层积处理时, ABA含量逐渐降低, GA含量逐渐升高, 使种子萌发.与野生型种子相比, ABA合成缺陷突变体种子萌发更快(Frey et al., 2012); 而过表达ABA合成基因的种子以及ABA代谢突变体种子保持较高的休眠水平(Matakiadis et al., 2009; Martinez-Andújar et al., 2011; Nonogaki et al., 2014), 表明ABA能够通过其生物合成通路调控种子的休眠和萌发.ABA信号通过PYR/PYL/RCAR (PYRABACTIN RESISTANCE 1/PYR-LIKE PROTEINS/REGULATORY COMPONENTS OF ABA RECEPTORS)-PP2C (PROTEIN PHOSPHATASE 2C)-SnRKs (SNF1-RELATED PROTEIN KINASES)级联反应向下传递(Cutler et al., 2010; Hubbard et al., 2010).目前鉴定到的PP2C类蛋白包括ABI1 (ABSCISIC ACID INSENSITIVE 1)、ABI2、HON (HONSU)和RDO5 (REDUCED DORMANCY 5).其中, ABI1和ABI2通过与ABA信号受体蛋白PYR/PYL/RCAR互作进而抑制ABA信号转导(Ma et al., 2009; Park et al., 2009).HON蛋白能够通过抑制ABA信号转导及促进GA信号转导抑制种子休眠(Kim et al., 2013), 说明HON能够整合ABA和GA信号以调控种子休眠.rdo5突变体的休眠水平降低, 但其ABA含量和ABA敏感性并未发生显著变化(Xiang et al., 2014), 说明RDO5通过不依赖ABA的途径调控种子休眠.ABI3、ABI4和ABI5是ABA信号通路下游的关键组分, 能够抑制种子萌发、促进种子休眠(Bentsink and Koornneef, 2008; Kanai et al., 2010; Shu et al., 2013).响应ABA信号的R2R3型MYB转录因子MYB96通过调控ABI4以及ABA合成相关基因NCED2 (9-CIS-EPOXYCAROTENOID DIOXYGENASE 2)和NCED6的转录进而促进种子休眠、抑制种子萌发(Lee et al., 2015a, 2015b).上述研究表明, ABA不仅通过其生物合成通路调控种子的休眠和萌发, 还能以ABA信号的形式发挥作用. ...
Seed germination and dormancy 1 1997
... 种子休眠是指有活力的种子在适宜的条件下暂时不萌发的现象(Bewley, 1997; Finch-Savage and Leubner-Metzger, 2006).种子的休眠包括休眠的诱导、维持和释放3个阶段(Shu et al., 2016).成熟的种子释放休眠后, 在适宜的条件下即可萌发.DOG1 (DELAY OF GERMINATION 1)在种子休眠诱导和维持中发挥重要作用.它既能通过抑制脱落酸(abscisic acid, ABA)通路的负调控因子——AHG1(ABA HYPERSENSITIVE GERMINATION 1)的磷酸酶活性促进种子休眠(Nishimura et al., 2018); 也能通过调控响应赤霉素(gibberellin, GA)信号的细胞壁重塑相关基因的表达促进种子休眠(Graeber et al., 2014).在种子吸胀初期, 胚乳中PHYB (PHYTOCHROME B)吸收远红光后以生理失活型存在, 此时PIF1 (PIF3-LIKE 5, PIL5)蛋白逐渐积累, 使胚乳中ABA水平逐渐升高.胚乳中的ABA信号能够释放到胚中并抑制GA的生物合成, 使种子保持休眠状态.随着种子吸胀时间的延长, 胚乳中ABA水平逐渐下调.此时胚中的PHYA介导远红光促进GA的生物合成, 进而促进种子萌发(Lee et al., 2012).因此, 种子的休眠和萌发是两个相互独立的生物学过程. ...
1 1994
... 与野生种相比, 经过驯化的栽培种种子休眠水平下调, 如水稻(Oryza sativa)、小麦(Triticum aestivum)、大麦(Hordeum vulgare)和玉米(Zea mays)等禾本科作物(Bewley and Black, 1994).近年来, 育种学家们利用遗传学或组学的方法鉴定到一些与种子休眠相关的位点、转录本或蛋白(Alonso-Blanco et al., 2003; Argyris et al., 2005; Gu et al., 2006; Née et al., 2017b).例如, 研究者利用QTL方法鉴定到调控种子休眠的关键基因——DOG1 (Alonso-Blanco et al., 2003; Bentsink et al., 2006).后续研究发现, DOG1能够参与调控ABA信号通路以及GA的生物合成, 进而调控种子休眠(Graeber et al., 2014; Nishimura et al., 2018).那么, DOG1能否参与调控GA信号通路进而促进种子休眠? DOG1是否受到外源光信号和内源激素的调控? 针对上述问题的研究不仅有助于人们更加清晰地理解种子如何监测环境因子的变化, 从而决定是否完成从休眠到萌发的发育转变; 还为利用光信号或激素人工调控种子的休眠和萌发, 解决农业生产中“胎萌”以及穗发芽等问题提供了理论支撑. ...
A reversible photoreaction controlling seed germination 1 1952
... PHY在黑暗条件下以生理失活的红光吸收型(Pr)存在, 吸收红光之后转变成其生理激活型(Pfr).两种光吸收型的PHY在Pr和Pfr两种状态间相互转变.早在20世纪50年代, 研究人员以莴苣(Lactuca sativa)种子为材料, 研究红光和远红光对其萌发的影响.结果表明红光促进种子萌发, 而远红光能够逆转红光的作用(Borthwick et al., 1952).红光和远红光对种子萌发的可逆调控暗示着PHY参与调控种子的萌发过程(Shinomura et al., 1994; Hennig et al., 2002).拟南芥PHY基因家族包含5个成员——PHYA-PHYE (Sharrock and Quail, 1989; Clack et al., 1994).种子萌发受到PHYA和PHYB的调控.其中, PHYB发挥主要功能.研究表明, PHYA蛋白在干种子中表达量很低, 随着种子在黑暗条件下吸胀时间的延长其蛋白表达量逐渐增加, 说明PHYA参与调控种子的萌发过程(Shinomura et al., 1996).PHYA主要在种子吸胀后期通过介导红光和远红光条件下的极低辐照度反应(VLFR)和远红光下的高辐照度反应(FR-HIR)调控种子萌发.相比之下, PHYB在干种子和吸胀种子中表达量均很高, 能够在种子吸胀初期(几个小时以内)介导红光和远红光下的低辐照度反应(LFR)调控种子萌发(Seo et al., 2009; Li et al., 2011).除了PHYA和PHYB外, PHYE也参与光调控的种子萌发过程, 而且三者在调控种子萌发方面功能冗余(Hennig et al., 2002).最近有研究表明, PHYB除了调控种子萌发, 还参与调控种子休眠(Jiang et al., 2016). ...
Blue-light photoreceptors in higher plants 1 1999
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
Cryptochromes: enabling plants and animals to determine circadian time 1 2003
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
Arabidopsis phytochrome A directly targets numerous promoters for individualized modulation of genes in a wide range of pathways 1 2014
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
Antagonism between abscisic acid and ethylene in Arabidopsis acts in parallel with the reciprocal regulation of their metabolism and signaling pathways 1 2009
... 除ABA和GA之外, 种子的休眠和萌发还受到7类激素的调控, 包括生长素(auxin)、乙烯(ethylene, ET)、油菜素内酯(brassinosteroids, BRs)、水杨酸(salicylic acid, SA)、茉莉酸(jasmonic acid, JA)、细胞分裂素(cytokinins, CTKs)和独脚金内酯(strigolactones, SLs).这些激素均通过影响ABA或GA通路间接调控种子的休眠和萌发(Shu et al., 2016; Née et al., 2017a).生长素能够通过调控ABI3的转录水平影响种子的休眠和萌发.当内源生长素水平升高时, 生长素信号通路中的负调控因子AXR2/3 (AUXIN- RESISTANT 2/3)发生降解.此时AXR2/3对响应生长素信号的ARF10 (AUXIN RESPONSE FACTOR 10)和ARF16基因的转录抑制作用被解除, 而ARF10和ARF16能够激活ABI3的表达, 从而使种子保持休眠状态(Liu et al., 2013a).然而, ARF10和ARF16不能直接结合在ABI3的启动子上(Liu et al., 2013a), 因此有待深入研究以揭示完整的生长素信号传递链.ET能够通过ABA生物合成和信号转导通路调控种子萌发(Cheng et al., 2009; Linkies et al., 2009; Corbineau et al., 2014), 但具体机制还不清楚.在盐胁迫条件下, ET受体蛋白ETR1 (ETHYLENE RESPONSE 1)和ETR2可能通过不依赖ET信号的通路调控ABA生物合成, 进而影响种子萌发(Wilson et al., 2014).最近的研究发现, ETR1通过解除ERF12 (ETHYLENE RESPONSE FACTOR 12)/TPL (TOPLESS)模块对DOG1基因的转录抑制作用而促进DOG1的转录, 进而促进种子休眠(Li et al., 2019).BR通过依赖MFT (MOTHER OFFT AND TFL 1)的途径拮抗ABA信号转导通路, 促进种子萌发(Xi and Yu, 2010; Xi et al., 2010).进一步的研究发现, BR信号通路中的负调控因子——BIN2 (BRASSINOSTEROID INSENSITIVE 2), 能够磷酸化并稳定ABI5蛋白, 进而促进ABA信号转导.BR信号通过抑制BIN2-ABI5互作解除ABA对种子萌发的抑制作用(Hu and Yu, 2014).SA在调控种子萌发上发挥双重作用.在正常生长条件下, SA通过抑制GA诱导的α淀粉酶编码基因的表达抑制种子萌发(Xie et al., 2007); 而在高盐胁迫下, SA通过其它通路促进种子萌发(Lee et al., 2010).CTKs通过抑制ABI5的转录或促进ABI5蛋白的降解拮抗ABA效应, 从而促进种子萌发(Wang et al., 2011; Guan et al., 2014).外源施加JA能够延迟种子萌发(Nambara et al., 2010), 表明JA能够抑制种子萌发.有研究表明, JA通过抑制ABA合成相关基因的表达、促进ABA代谢相关基因的表达拮抗ABA, 从而促进种子萌发(Jacobsen et al., 2013).JAZ3 (JASMONATE-ZIM DOMAIN PROTEIN 3)通过与ABI5互作抑制ABI5的转录激活活性, 进而促进种子萌发(Ju et al., 2019).目前关于JA调控种子萌发功能上存在矛盾的原因还有待深入探究.SLs通过降低ABA/GA值促进种子萌发(Toh et al., 2012).此外, SLs信号通路组分也能参与调控种子萌发, 如SMAX1 (SUPPRESSOR OF MORE AXILLARY GROWTH2 1) (Stanga et al., 2013).但目前SLs调控种子萌发的具体机制还不清楚.综上, 生长素、ET、BR、CTKs、JA和SLs通过调控ABA生物合成或信号通路调控种子的休眠和萌发.然而生长素、ET、BR、CTKs、JA与GA之间的交互作用还有待深入研究. ...
The phytochrome apoprotein family in Arabidopsis is encoded by five genes: the sequences and expression of PHYD and PHYE 1 1994
... PHY在黑暗条件下以生理失活的红光吸收型(Pr)存在, 吸收红光之后转变成其生理激活型(Pfr).两种光吸收型的PHY在Pr和Pfr两种状态间相互转变.早在20世纪50年代, 研究人员以莴苣(Lactuca sativa)种子为材料, 研究红光和远红光对其萌发的影响.结果表明红光促进种子萌发, 而远红光能够逆转红光的作用(Borthwick et al., 1952).红光和远红光对种子萌发的可逆调控暗示着PHY参与调控种子的萌发过程(Shinomura et al., 1994; Hennig et al., 2002).拟南芥PHY基因家族包含5个成员——PHYA-PHYE (Sharrock and Quail, 1989; Clack et al., 1994).种子萌发受到PHYA和PHYB的调控.其中, PHYB发挥主要功能.研究表明, PHYA蛋白在干种子中表达量很低, 随着种子在黑暗条件下吸胀时间的延长其蛋白表达量逐渐增加, 说明PHYA参与调控种子的萌发过程(Shinomura et al., 1996).PHYA主要在种子吸胀后期通过介导红光和远红光条件下的极低辐照度反应(VLFR)和远红光下的高辐照度反应(FR-HIR)调控种子萌发.相比之下, PHYB在干种子和吸胀种子中表达量均很高, 能够在种子吸胀初期(几个小时以内)介导红光和远红光下的低辐照度反应(LFR)调控种子萌发(Seo et al., 2009; Li et al., 2011).除了PHYA和PHYB外, PHYE也参与光调控的种子萌发过程, 而且三者在调控种子萌发方面功能冗余(Hennig et al., 2002).最近有研究表明, PHYB除了调控种子萌发, 还参与调控种子休眠(Jiang et al., 2016). ...
Ethylene, a key factor in the regulation of seed dormancy 1 2014
... 除ABA和GA之外, 种子的休眠和萌发还受到7类激素的调控, 包括生长素(auxin)、乙烯(ethylene, ET)、油菜素内酯(brassinosteroids, BRs)、水杨酸(salicylic acid, SA)、茉莉酸(jasmonic acid, JA)、细胞分裂素(cytokinins, CTKs)和独脚金内酯(strigolactones, SLs).这些激素均通过影响ABA或GA通路间接调控种子的休眠和萌发(Shu et al., 2016; Née et al., 2017a).生长素能够通过调控ABI3的转录水平影响种子的休眠和萌发.当内源生长素水平升高时, 生长素信号通路中的负调控因子AXR2/3 (AUXIN- RESISTANT 2/3)发生降解.此时AXR2/3对响应生长素信号的ARF10 (AUXIN RESPONSE FACTOR 10)和ARF16基因的转录抑制作用被解除, 而ARF10和ARF16能够激活ABI3的表达, 从而使种子保持休眠状态(Liu et al., 2013a).然而, ARF10和ARF16不能直接结合在ABI3的启动子上(Liu et al., 2013a), 因此有待深入研究以揭示完整的生长素信号传递链.ET能够通过ABA生物合成和信号转导通路调控种子萌发(Cheng et al., 2009; Linkies et al., 2009; Corbineau et al., 2014), 但具体机制还不清楚.在盐胁迫条件下, ET受体蛋白ETR1 (ETHYLENE RESPONSE 1)和ETR2可能通过不依赖ET信号的通路调控ABA生物合成, 进而影响种子萌发(Wilson et al., 2014).最近的研究发现, ETR1通过解除ERF12 (ETHYLENE RESPONSE FACTOR 12)/TPL (TOPLESS)模块对DOG1基因的转录抑制作用而促进DOG1的转录, 进而促进种子休眠(Li et al., 2019).BR通过依赖MFT (MOTHER OFFT AND TFL 1)的途径拮抗ABA信号转导通路, 促进种子萌发(Xi and Yu, 2010; Xi et al., 2010).进一步的研究发现, BR信号通路中的负调控因子——BIN2 (BRASSINOSTEROID INSENSITIVE 2), 能够磷酸化并稳定ABI5蛋白, 进而促进ABA信号转导.BR信号通过抑制BIN2-ABI5互作解除ABA对种子萌发的抑制作用(Hu and Yu, 2014).SA在调控种子萌发上发挥双重作用.在正常生长条件下, SA通过抑制GA诱导的α淀粉酶编码基因的表达抑制种子萌发(Xie et al., 2007); 而在高盐胁迫下, SA通过其它通路促进种子萌发(Lee et al., 2010).CTKs通过抑制ABI5的转录或促进ABI5蛋白的降解拮抗ABA效应, 从而促进种子萌发(Wang et al., 2011; Guan et al., 2014).外源施加JA能够延迟种子萌发(Nambara et al., 2010), 表明JA能够抑制种子萌发.有研究表明, JA通过抑制ABA合成相关基因的表达、促进ABA代谢相关基因的表达拮抗ABA, 从而促进种子萌发(Jacobsen et al., 2013).JAZ3 (JASMONATE-ZIM DOMAIN PROTEIN 3)通过与ABI5互作抑制ABI5的转录激活活性, 进而促进种子萌发(Ju et al., 2019).目前关于JA调控种子萌发功能上存在矛盾的原因还有待深入探究.SLs通过降低ABA/GA值促进种子萌发(Toh et al., 2012).此外, SLs信号通路组分也能参与调控种子萌发, 如SMAX1 (SUPPRESSOR OF MORE AXILLARY GROWTH2 1) (Stanga et al., 2013).但目前SLs调控种子萌发的具体机制还不清楚.综上, 生长素、ET、BR、CTKs、JA和SLs通过调控ABA生物合成或信号通路调控种子的休眠和萌发.然而生长素、ET、BR、CTKs、JA与GA之间的交互作用还有待深入研究. ...
Abscisic acid: emergence of a core signaling network 1 2010
... 激素在种子休眠和萌发过程中发挥重要调控作用.其中, ABA和GA在调控种子休眠和萌发上行使主要功能.ABA促进种子休眠, 而GA促进种子萌发(Shu et al., 2016; Née et al., 2017a).在种子成熟过程中, ABA在种子内逐渐积累, 使种子的休眠水平逐渐升高.相比之下, 当种子受到吸胀或层积处理时, ABA含量逐渐降低, GA含量逐渐升高, 使种子萌发.与野生型种子相比, ABA合成缺陷突变体种子萌发更快(Frey et al., 2012); 而过表达ABA合成基因的种子以及ABA代谢突变体种子保持较高的休眠水平(Matakiadis et al., 2009; Martinez-Andújar et al., 2011; Nonogaki et al., 2014), 表明ABA能够通过其生物合成通路调控种子的休眠和萌发.ABA信号通过PYR/PYL/RCAR (PYRABACTIN RESISTANCE 1/PYR-LIKE PROTEINS/REGULATORY COMPONENTS OF ABA RECEPTORS)-PP2C (PROTEIN PHOSPHATASE 2C)-SnRKs (SNF1-RELATED PROTEIN KINASES)级联反应向下传递(Cutler et al., 2010; Hubbard et al., 2010).目前鉴定到的PP2C类蛋白包括ABI1 (ABSCISIC ACID INSENSITIVE 1)、ABI2、HON (HONSU)和RDO5 (REDUCED DORMANCY 5).其中, ABI1和ABI2通过与ABA信号受体蛋白PYR/PYL/RCAR互作进而抑制ABA信号转导(Ma et al., 2009; Park et al., 2009).HON蛋白能够通过抑制ABA信号转导及促进GA信号转导抑制种子休眠(Kim et al., 2013), 说明HON能够整合ABA和GA信号以调控种子休眠.rdo5突变体的休眠水平降低, 但其ABA含量和ABA敏感性并未发生显著变化(Xiang et al., 2014), 说明RDO5通过不依赖ABA的途径调控种子休眠.ABI3、ABI4和ABI5是ABA信号通路下游的关键组分, 能够抑制种子萌发、促进种子休眠(Bentsink and Koornneef, 2008; Kanai et al., 2010; Shu et al., 2013).响应ABA信号的R2R3型MYB转录因子MYB96通过调控ABI4以及ABA合成相关基因NCED2 (9-CIS-EPOXYCAROTENOID DIOXYGENASE 2)和NCED6的转录进而促进种子休眠、抑制种子萌发(Lee et al., 2015a, 2015b).上述研究表明, ABA不仅通过其生物合成通路调控种子的休眠和萌发, 还能以ABA信号的形式发挥作用. ...
Gibberellin signaling in plants 1 2013
... GA能够通过抑制ABA诱导的种子休眠促进种子萌发(Gubler et al., 2005; Graeber et al., 2012).在植物体内, 具有生物活性的GA主要包括GA1和GA4, 它们通过软化种皮、促进胚乳层细胞的水解和胚的生长打破种子休眠, 进而促进种子萌发(Holdsworth et al., 2008).GA缺失突变体ga1 (gibberellic acid- requiring 1)和ga2表现出强烈的休眠表型, 需要外源施加GA方可萌发(Lee et al., 2002; Shu et al., 2013).GA20ox (GA 20-OXIDASE)和GA3ox基因编码参与GA生物合成的氧化酶; GA2ox编码的GA2-氧化酶主要参与GA的代谢过程.在PHYB-off条件下, ga2ox突变体萌发水平升高(Oh et al., 2006; Yamauchi et al., 2007).上述研究结果表明, GA能通过其生物合成通路调控种子的休眠和萌发.在GA信号转导过程中, DELLA蛋白扮演着重要角色, 它能响应GA信号快速降解, 对GA信号转导起限速作用(Silverstone et al., 1998; Itoh et al., 2002; Zentella et al., 2007; Nemoto et al., 2017).GA受体蛋白GID1 (GA-INSENSITIVE DWARF 1)能够负调控DELLA蛋白的稳定性, 促进GA信号转导(Davière and Achard, 2013).拟南芥SLY1 (SLEEPY1)和水稻GID2 (GA-INSENSITIVE DWARF2)是SCF聚合体中的F-box亚基, 依赖其C端的GGF和LSL基序与DELLA蛋白C端VHIID和LHR2基序结合, 促进DELLA多聚泛素化(Hirano et al., 2010; Ariizumi et al., 2011).研究表明, DELLA蛋白GAI (GA-INSENSITIVE)能够通过抑制TCP14 (TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR 14)和TCP15的表达减缓细胞周期的进程, 最终使种子保持休眠状态(Resentini et al., 2015).作为转录调节子, DELLA如何实现对下游基因的转录调控? 有研究指出, DELLA能够通过与染色质重塑因子互作调控下游基因的转录.DELLA蛋白RGL2 (RGA-LIKE 2)和RGL3与染色质重塑因子SWI3C (SWITCH3C)发生互作, 进而影响GID1a以及GA3ox基因的转录(Sarnowska et al., 2013).此外, DELLA还能通过与种子萌发相关转录因子互作进而实现其对萌发的调控.RGL2通过与转录因子NF-YC互作, 诱导下游ABI5的转录, 进而抑制种子萌发(Liu et al., 2016).此外, RGL2还能通过与转录因子DOF6 (BINDING1 ZINC FINGER 6)互作诱导GATA12的表达, 进而促进种子休眠(Ravindran et al., 2017).DELLA通过与ICE1 (INDUCER OF CBF EXPRESSION 1)互作解除其对ABA响应基因EM1 (LATE EMBRYOGENESIS ABUNDANT 1)和EM6的转录抑制作用, 最终抑制种子萌发(Hu et al., 2019). ...
Light-mediated hormonal regulation of plant growth and development 2 2016
Epoxycarotenoid cleavage by NCED5 fine-tunes ABA accumulation and affects seed dormancy and drought tolerance with other NCED family members 1 2012
... 激素在种子休眠和萌发过程中发挥重要调控作用.其中, ABA和GA在调控种子休眠和萌发上行使主要功能.ABA促进种子休眠, 而GA促进种子萌发(Shu et al., 2016; Née et al., 2017a).在种子成熟过程中, ABA在种子内逐渐积累, 使种子的休眠水平逐渐升高.相比之下, 当种子受到吸胀或层积处理时, ABA含量逐渐降低, GA含量逐渐升高, 使种子萌发.与野生型种子相比, ABA合成缺陷突变体种子萌发更快(Frey et al., 2012); 而过表达ABA合成基因的种子以及ABA代谢突变体种子保持较高的休眠水平(Matakiadis et al., 2009; Martinez-Andújar et al., 2011; Nonogaki et al., 2014), 表明ABA能够通过其生物合成通路调控种子的休眠和萌发.ABA信号通过PYR/PYL/RCAR (PYRABACTIN RESISTANCE 1/PYR-LIKE PROTEINS/REGULATORY COMPONENTS OF ABA RECEPTORS)-PP2C (PROTEIN PHOSPHATASE 2C)-SnRKs (SNF1-RELATED PROTEIN KINASES)级联反应向下传递(Cutler et al., 2010; Hubbard et al., 2010).目前鉴定到的PP2C类蛋白包括ABI1 (ABSCISIC ACID INSENSITIVE 1)、ABI2、HON (HONSU)和RDO5 (REDUCED DORMANCY 5).其中, ABI1和ABI2通过与ABA信号受体蛋白PYR/PYL/RCAR互作进而抑制ABA信号转导(Ma et al., 2009; Park et al., 2009).HON蛋白能够通过抑制ABA信号转导及促进GA信号转导抑制种子休眠(Kim et al., 2013), 说明HON能够整合ABA和GA信号以调控种子休眠.rdo5突变体的休眠水平降低, 但其ABA含量和ABA敏感性并未发生显著变化(Xiang et al., 2014), 说明RDO5通过不依赖ABA的途径调控种子休眠.ABI3、ABI4和ABI5是ABA信号通路下游的关键组分, 能够抑制种子萌发、促进种子休眠(Bentsink and Koornneef, 2008; Kanai et al., 2010; Shu et al., 2013).响应ABA信号的R2R3型MYB转录因子MYB96通过调控ABI4以及ABA合成相关基因NCED2 (9-CIS-EPOXYCAROTENOID DIOXYGENASE 2)和NCED6的转录进而促进种子休眠、抑制种子萌发(Lee et al., 2015a, 2015b).上述研究表明, ABA不仅通过其生物合成通路调控种子的休眠和萌发, 还能以ABA信号的形式发挥作用. ...
The Dof protein DAG1 mediates PIL5 activity on seed germination by negatively regulating GA biosynthetic gene AtGA3ox1 2 2010
... PIF1能够介导外源光信号, 通过调控ABA和GA的生物合成调控种子萌发(图1A).统计Col和pif1突变体的种子在PHYB-on和PHYB-off条件下的萌发率, 发现Col只能在PHYB-on条件下萌发, pif1在PHYB-on和PHYB-off条件下均能萌发; 使用GA合成抑制剂PAC (paclobutrazol)能够抑制Col和pif1萌发.上述研究结果表明, PIF1在PHYB介导的种子萌发过程中发挥重要作用, 它可能通过调控GA的生物合成抑制种子萌发.进一步研究证实, PIF1能够抑制GA合成相关基因GA3ox1和GA3ox2的表达, 促进GA代谢相关基因GA2ox2的表达, 进而下调内源GA4的水平, 抑制种子萌发(Oh et al., 2006).与ga1单突变体相比, pif1/ga1双突变体中ABA合成相关基因的表达下调; 而ABA代谢相关基因的表达水平上调, 致使pif1/ga1双突变体中ABA的水平降低, 表现出持续萌发的表型(Oh et al., 2007).那么, PIF1如何调控ABA和GA生物合成相关基因的转录? 后续的研究发现, PIF1能够通过诱导SOM (SOMNUS)和DAG1 (DOF AFFECTING GERMINATION 1)的转录间接抑制GA合成、促进ABA合成, 最终抑制种子萌发(Kim et al., 2008; Gabriele et al., 2010) (图1A).其中, CCCH型锌指蛋白SOM通过调控组蛋白精氨酸去甲基化酶编码基因JMJ20 (JUMONJI 20)和JMJ22的表达, 进而影响GA合成基因GA3ox1和GA3ox2的甲基化水平, 最终抑制种子的萌发过程.此外, SOM还能激活GA代谢基因GA2ox2以及ABA合成基因ABA1 (ABA- DEFICIENT 1)、NCED6和NCED9的表达, 抑制ABA代谢基因CYP707A2表达, 从而抑制GA生物合成、促进ABA合成, 最终抑制种子萌发(Kim et al., 2008; de Wit et al., 2016).包含Dof结构域的DAG1蛋白直接结合在GA3ox1启动子上抑制其表达, 进而抑制种子萌发(Gabriele et al., 2010). ...
... 启动子上抑制其表达, 进而抑制种子萌发(Gabriele et al., 2010). ...
DELAY OF GERMINATION 1 mediates a conserved coat-dormancy mechanism for the temperature- and gibberellin-dependent control of seed germination 2 2014
... 种子休眠是指有活力的种子在适宜的条件下暂时不萌发的现象(Bewley, 1997; Finch-Savage and Leubner-Metzger, 2006).种子的休眠包括休眠的诱导、维持和释放3个阶段(Shu et al., 2016).成熟的种子释放休眠后, 在适宜的条件下即可萌发.DOG1 (DELAY OF GERMINATION 1)在种子休眠诱导和维持中发挥重要作用.它既能通过抑制脱落酸(abscisic acid, ABA)通路的负调控因子——AHG1(ABA HYPERSENSITIVE GERMINATION 1)的磷酸酶活性促进种子休眠(Nishimura et al., 2018); 也能通过调控响应赤霉素(gibberellin, GA)信号的细胞壁重塑相关基因的表达促进种子休眠(Graeber et al., 2014).在种子吸胀初期, 胚乳中PHYB (PHYTOCHROME B)吸收远红光后以生理失活型存在, 此时PIF1 (PIF3-LIKE 5, PIL5)蛋白逐渐积累, 使胚乳中ABA水平逐渐升高.胚乳中的ABA信号能够释放到胚中并抑制GA的生物合成, 使种子保持休眠状态.随着种子吸胀时间的延长, 胚乳中ABA水平逐渐下调.此时胚中的PHYA介导远红光促进GA的生物合成, 进而促进种子萌发(Lee et al., 2012).因此, 种子的休眠和萌发是两个相互独立的生物学过程. ...
... 与野生种相比, 经过驯化的栽培种种子休眠水平下调, 如水稻(Oryza sativa)、小麦(Triticum aestivum)、大麦(Hordeum vulgare)和玉米(Zea mays)等禾本科作物(Bewley and Black, 1994).近年来, 育种学家们利用遗传学或组学的方法鉴定到一些与种子休眠相关的位点、转录本或蛋白(Alonso-Blanco et al., 2003; Argyris et al., 2005; Gu et al., 2006; Née et al., 2017b).例如, 研究者利用QTL方法鉴定到调控种子休眠的关键基因——DOG1 (Alonso-Blanco et al., 2003; Bentsink et al., 2006).后续研究发现, DOG1能够参与调控ABA信号通路以及GA的生物合成, 进而调控种子休眠(Graeber et al., 2014; Nishimura et al., 2018).那么, DOG1能否参与调控GA信号通路进而促进种子休眠? DOG1是否受到外源光信号和内源激素的调控? 针对上述问题的研究不仅有助于人们更加清晰地理解种子如何监测环境因子的变化, 从而决定是否完成从休眠到萌发的发育转变; 还为利用光信号或激素人工调控种子的休眠和萌发, 解决农业生产中“胎萌”以及穗发芽等问题提供了理论支撑. ...
Molecular mechanisms of seed dormancy 1 2012
... GA能够通过抑制ABA诱导的种子休眠促进种子萌发(Gubler et al., 2005; Graeber et al., 2012).在植物体内, 具有生物活性的GA主要包括GA1和GA4, 它们通过软化种皮、促进胚乳层细胞的水解和胚的生长打破种子休眠, 进而促进种子萌发(Holdsworth et al., 2008).GA缺失突变体ga1 (gibberellic acid- requiring 1)和ga2表现出强烈的休眠表型, 需要外源施加GA方可萌发(Lee et al., 2002; Shu et al., 2013).GA20ox (GA 20-OXIDASE)和GA3ox基因编码参与GA生物合成的氧化酶; GA2ox编码的GA2-氧化酶主要参与GA的代谢过程.在PHYB-off条件下, ga2ox突变体萌发水平升高(Oh et al., 2006; Yamauchi et al., 2007).上述研究结果表明, GA能通过其生物合成通路调控种子的休眠和萌发.在GA信号转导过程中, DELLA蛋白扮演着重要角色, 它能响应GA信号快速降解, 对GA信号转导起限速作用(Silverstone et al., 1998; Itoh et al., 2002; Zentella et al., 2007; Nemoto et al., 2017).GA受体蛋白GID1 (GA-INSENSITIVE DWARF 1)能够负调控DELLA蛋白的稳定性, 促进GA信号转导(Davière and Achard, 2013).拟南芥SLY1 (SLEEPY1)和水稻GID2 (GA-INSENSITIVE DWARF2)是SCF聚合体中的F-box亚基, 依赖其C端的GGF和LSL基序与DELLA蛋白C端VHIID和LHR2基序结合, 促进DELLA多聚泛素化(Hirano et al., 2010; Ariizumi et al., 2011).研究表明, DELLA蛋白GAI (GA-INSENSITIVE)能够通过抑制TCP14 (TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR 14)和TCP15的表达减缓细胞周期的进程, 最终使种子保持休眠状态(Resentini et al., 2015).作为转录调节子, DELLA如何实现对下游基因的转录调控? 有研究指出, DELLA能够通过与染色质重塑因子互作调控下游基因的转录.DELLA蛋白RGL2 (RGA-LIKE 2)和RGL3与染色质重塑因子SWI3C (SWITCH3C)发生互作, 进而影响GID1a以及GA3ox基因的转录(Sarnowska et al., 2013).此外, DELLA还能通过与种子萌发相关转录因子互作进而实现其对萌发的调控.RGL2通过与转录因子NF-YC互作, 诱导下游ABI5的转录, 进而抑制种子萌发(Liu et al., 2016).此外, RGL2还能通过与转录因子DOF6 (BINDING1 ZINC FINGER 6)互作诱导GATA12的表达, 进而促进种子休眠(Ravindran et al., 2017).DELLA通过与ICE1 (INDUCER OF CBF EXPRESSION 1)互作解除其对ABA响应基因EM1 (LATE EMBRYOGENESIS ABUNDANT 1)和EM6的转录抑制作用, 最终抑制种子萌发(Hu et al., 2019). ...
Dormancy genes from weedy rice respond divergently to seed development environments 1 2006
... 与野生种相比, 经过驯化的栽培种种子休眠水平下调, 如水稻(Oryza sativa)、小麦(Triticum aestivum)、大麦(Hordeum vulgare)和玉米(Zea mays)等禾本科作物(Bewley and Black, 1994).近年来, 育种学家们利用遗传学或组学的方法鉴定到一些与种子休眠相关的位点、转录本或蛋白(Alonso-Blanco et al., 2003; Argyris et al., 2005; Gu et al., 2006; Née et al., 2017b).例如, 研究者利用QTL方法鉴定到调控种子休眠的关键基因——DOG1 (Alonso-Blanco et al., 2003; Bentsink et al., 2006).后续研究发现, DOG1能够参与调控ABA信号通路以及GA的生物合成, 进而调控种子休眠(Graeber et al., 2014; Nishimura et al., 2018).那么, DOG1能否参与调控GA信号通路进而促进种子休眠? DOG1是否受到外源光信号和内源激素的调控? 针对上述问题的研究不仅有助于人们更加清晰地理解种子如何监测环境因子的变化, 从而决定是否完成从休眠到萌发的发育转变; 还为利用光信号或激素人工调控种子的休眠和萌发, 解决农业生产中“胎萌”以及穗发芽等问题提供了理论支撑. ...
Cytokinin antagonizes abscisic acidmediated inhibition of cotyledon greening by promoting the degradation of abscisic acid insensitive5 protein in Arabidopsis 1 2014
... 除ABA和GA之外, 种子的休眠和萌发还受到7类激素的调控, 包括生长素(auxin)、乙烯(ethylene, ET)、油菜素内酯(brassinosteroids, BRs)、水杨酸(salicylic acid, SA)、茉莉酸(jasmonic acid, JA)、细胞分裂素(cytokinins, CTKs)和独脚金内酯(strigolactones, SLs).这些激素均通过影响ABA或GA通路间接调控种子的休眠和萌发(Shu et al., 2016; Née et al., 2017a).生长素能够通过调控ABI3的转录水平影响种子的休眠和萌发.当内源生长素水平升高时, 生长素信号通路中的负调控因子AXR2/3 (AUXIN- RESISTANT 2/3)发生降解.此时AXR2/3对响应生长素信号的ARF10 (AUXIN RESPONSE FACTOR 10)和ARF16基因的转录抑制作用被解除, 而ARF10和ARF16能够激活ABI3的表达, 从而使种子保持休眠状态(Liu et al., 2013a).然而, ARF10和ARF16不能直接结合在ABI3的启动子上(Liu et al., 2013a), 因此有待深入研究以揭示完整的生长素信号传递链.ET能够通过ABA生物合成和信号转导通路调控种子萌发(Cheng et al., 2009; Linkies et al., 2009; Corbineau et al., 2014), 但具体机制还不清楚.在盐胁迫条件下, ET受体蛋白ETR1 (ETHYLENE RESPONSE 1)和ETR2可能通过不依赖ET信号的通路调控ABA生物合成, 进而影响种子萌发(Wilson et al., 2014).最近的研究发现, ETR1通过解除ERF12 (ETHYLENE RESPONSE FACTOR 12)/TPL (TOPLESS)模块对DOG1基因的转录抑制作用而促进DOG1的转录, 进而促进种子休眠(Li et al., 2019).BR通过依赖MFT (MOTHER OFFT AND TFL 1)的途径拮抗ABA信号转导通路, 促进种子萌发(Xi and Yu, 2010; Xi et al., 2010).进一步的研究发现, BR信号通路中的负调控因子——BIN2 (BRASSINOSTEROID INSENSITIVE 2), 能够磷酸化并稳定ABI5蛋白, 进而促进ABA信号转导.BR信号通过抑制BIN2-ABI5互作解除ABA对种子萌发的抑制作用(Hu and Yu, 2014).SA在调控种子萌发上发挥双重作用.在正常生长条件下, SA通过抑制GA诱导的α淀粉酶编码基因的表达抑制种子萌发(Xie et al., 2007); 而在高盐胁迫下, SA通过其它通路促进种子萌发(Lee et al., 2010).CTKs通过抑制ABI5的转录或促进ABI5蛋白的降解拮抗ABA效应, 从而促进种子萌发(Wang et al., 2011; Guan et al., 2014).外源施加JA能够延迟种子萌发(Nambara et al., 2010), 表明JA能够抑制种子萌发.有研究表明, JA通过抑制ABA合成相关基因的表达、促进ABA代谢相关基因的表达拮抗ABA, 从而促进种子萌发(Jacobsen et al., 2013).JAZ3 (JASMONATE-ZIM DOMAIN PROTEIN 3)通过与ABI5互作抑制ABI5的转录激活活性, 进而促进种子萌发(Ju et al., 2019).目前关于JA调控种子萌发功能上存在矛盾的原因还有待深入探究.SLs通过降低ABA/GA值促进种子萌发(Toh et al., 2012).此外, SLs信号通路组分也能参与调控种子萌发, 如SMAX1 (SUPPRESSOR OF MORE AXILLARY GROWTH2 1) (Stanga et al., 2013).但目前SLs调控种子萌发的具体机制还不清楚.综上, 生长素、ET、BR、CTKs、JA和SLs通过调控ABA生物合成或信号通路调控种子的休眠和萌发.然而生长素、ET、BR、CTKs、JA与GA之间的交互作用还有待深入研究. ...
Dormancy release, ABA and pre-harvest sprouting 1 2005
... GA能够通过抑制ABA诱导的种子休眠促进种子萌发(Gubler et al., 2005; Graeber et al., 2012).在植物体内, 具有生物活性的GA主要包括GA1和GA4, 它们通过软化种皮、促进胚乳层细胞的水解和胚的生长打破种子休眠, 进而促进种子萌发(Holdsworth et al., 2008).GA缺失突变体ga1 (gibberellic acid- requiring 1)和ga2表现出强烈的休眠表型, 需要外源施加GA方可萌发(Lee et al., 2002; Shu et al., 2013).GA20ox (GA 20-OXIDASE)和GA3ox基因编码参与GA生物合成的氧化酶; GA2ox编码的GA2-氧化酶主要参与GA的代谢过程.在PHYB-off条件下, ga2ox突变体萌发水平升高(Oh et al., 2006; Yamauchi et al., 2007).上述研究结果表明, GA能通过其生物合成通路调控种子的休眠和萌发.在GA信号转导过程中, DELLA蛋白扮演着重要角色, 它能响应GA信号快速降解, 对GA信号转导起限速作用(Silverstone et al., 1998; Itoh et al., 2002; Zentella et al., 2007; Nemoto et al., 2017).GA受体蛋白GID1 (GA-INSENSITIVE DWARF 1)能够负调控DELLA蛋白的稳定性, 促进GA信号转导(Davière and Achard, 2013).拟南芥SLY1 (SLEEPY1)和水稻GID2 (GA-INSENSITIVE DWARF2)是SCF聚合体中的F-box亚基, 依赖其C端的GGF和LSL基序与DELLA蛋白C端VHIID和LHR2基序结合, 促进DELLA多聚泛素化(Hirano et al., 2010; Ariizumi et al., 2011).研究表明, DELLA蛋白GAI (GA-INSENSITIVE)能够通过抑制TCP14 (TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR 14)和TCP15的表达减缓细胞周期的进程, 最终使种子保持休眠状态(Resentini et al., 2015).作为转录调节子, DELLA如何实现对下游基因的转录调控? 有研究指出, DELLA能够通过与染色质重塑因子互作调控下游基因的转录.DELLA蛋白RGL2 (RGA-LIKE 2)和RGL3与染色质重塑因子SWI3C (SWITCH3C)发生互作, 进而影响GID1a以及GA3ox基因的转录(Sarnowska et al., 2013).此外, DELLA还能通过与种子萌发相关转录因子互作进而实现其对萌发的调控.RGL2通过与转录因子NF-YC互作, 诱导下游ABI5的转录, 进而抑制种子萌发(Liu et al., 2016).此外, RGL2还能通过与转录因子DOF6 (BINDING1 ZINC FINGER 6)互作诱导GATA12的表达, 进而促进种子休眠(Ravindran et al., 2017).DELLA通过与ICE1 (INDUCER OF CBF EXPRESSION 1)互作解除其对ABA响应基因EM1 (LATE EMBRYOGENESIS ABUNDANT 1)和EM6的转录抑制作用, 最终抑制种子萌发(Hu et al., 2019). ...
HY5 stability and activity in Arabidopsis is regulated by phosphorylation in its COP1 binding domain 1 2000
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
Phytochrome E controls light-induced germination of Arabidopsis 3 2002
... PHY在黑暗条件下以生理失活的红光吸收型(Pr)存在, 吸收红光之后转变成其生理激活型(Pfr).两种光吸收型的PHY在Pr和Pfr两种状态间相互转变.早在20世纪50年代, 研究人员以莴苣(Lactuca sativa)种子为材料, 研究红光和远红光对其萌发的影响.结果表明红光促进种子萌发, 而远红光能够逆转红光的作用(Borthwick et al., 1952).红光和远红光对种子萌发的可逆调控暗示着PHY参与调控种子的萌发过程(Shinomura et al., 1994; Hennig et al., 2002).拟南芥PHY基因家族包含5个成员——PHYA-PHYE (Sharrock and Quail, 1989; Clack et al., 1994).种子萌发受到PHYA和PHYB的调控.其中, PHYB发挥主要功能.研究表明, PHYA蛋白在干种子中表达量很低, 随着种子在黑暗条件下吸胀时间的延长其蛋白表达量逐渐增加, 说明PHYA参与调控种子的萌发过程(Shinomura et al., 1996).PHYA主要在种子吸胀后期通过介导红光和远红光条件下的极低辐照度反应(VLFR)和远红光下的高辐照度反应(FR-HIR)调控种子萌发.相比之下, PHYB在干种子和吸胀种子中表达量均很高, 能够在种子吸胀初期(几个小时以内)介导红光和远红光下的低辐照度反应(LFR)调控种子萌发(Seo et al., 2009; Li et al., 2011).除了PHYA和PHYB外, PHYE也参与光调控的种子萌发过程, 而且三者在调控种子萌发方面功能冗余(Hennig et al., 2002).最近有研究表明, PHYB除了调控种子萌发, 还参与调控种子休眠(Jiang et al., 2016). ...
... ).除了PHYA和PHYB外, PHYE也参与光调控的种子萌发过程, 而且三者在调控种子萌发方面功能冗余(Hennig et al., 2002).最近有研究表明, PHYB除了调控种子萌发, 还参与调控种子休眠(Jiang et al., 2016). ...
... GA能够通过抑制ABA诱导的种子休眠促进种子萌发(Gubler et al., 2005; Graeber et al., 2012).在植物体内, 具有生物活性的GA主要包括GA1和GA4, 它们通过软化种皮、促进胚乳层细胞的水解和胚的生长打破种子休眠, 进而促进种子萌发(Holdsworth et al., 2008).GA缺失突变体ga1 (gibberellic acid- requiring 1)和ga2表现出强烈的休眠表型, 需要外源施加GA方可萌发(Lee et al., 2002; Shu et al., 2013).GA20ox (GA 20-OXIDASE)和GA3ox基因编码参与GA生物合成的氧化酶; GA2ox编码的GA2-氧化酶主要参与GA的代谢过程.在PHYB-off条件下, ga2ox突变体萌发水平升高(Oh et al., 2006; Yamauchi et al., 2007).上述研究结果表明, GA能通过其生物合成通路调控种子的休眠和萌发.在GA信号转导过程中, DELLA蛋白扮演着重要角色, 它能响应GA信号快速降解, 对GA信号转导起限速作用(Silverstone et al., 1998; Itoh et al., 2002; Zentella et al., 2007; Nemoto et al., 2017).GA受体蛋白GID1 (GA-INSENSITIVE DWARF 1)能够负调控DELLA蛋白的稳定性, 促进GA信号转导(Davière and Achard, 2013).拟南芥SLY1 (SLEEPY1)和水稻GID2 (GA-INSENSITIVE DWARF2)是SCF聚合体中的F-box亚基, 依赖其C端的GGF和LSL基序与DELLA蛋白C端VHIID和LHR2基序结合, 促进DELLA多聚泛素化(Hirano et al., 2010; Ariizumi et al., 2011).研究表明, DELLA蛋白GAI (GA-INSENSITIVE)能够通过抑制TCP14 (TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR 14)和TCP15的表达减缓细胞周期的进程, 最终使种子保持休眠状态(Resentini et al., 2015).作为转录调节子, DELLA如何实现对下游基因的转录调控? 有研究指出, DELLA能够通过与染色质重塑因子互作调控下游基因的转录.DELLA蛋白RGL2 (RGA-LIKE 2)和RGL3与染色质重塑因子SWI3C (SWITCH3C)发生互作, 进而影响GID1a以及GA3ox基因的转录(Sarnowska et al., 2013).此外, DELLA还能通过与种子萌发相关转录因子互作进而实现其对萌发的调控.RGL2通过与转录因子NF-YC互作, 诱导下游ABI5的转录, 进而抑制种子萌发(Liu et al., 2016).此外, RGL2还能通过与转录因子DOF6 (BINDING1 ZINC FINGER 6)互作诱导GATA12的表达, 进而促进种子休眠(Ravindran et al., 2017).DELLA通过与ICE1 (INDUCER OF CBF EXPRESSION 1)互作解除其对ABA响应基因EM1 (LATE EMBRYOGENESIS ABUNDANT 1)和EM6的转录抑制作用, 最终抑制种子萌发(Hu et al., 2019). ...
The transcription factor INDUCER OF CBF EXPRESSION 1 interacts with ABSCISIC ACID INSENSITIVE 5 and DELLA proteins to fine-tune abscisic acid signaling during seed germination in Arabidopsis 1 2019
... GA能够通过抑制ABA诱导的种子休眠促进种子萌发(Gubler et al., 2005; Graeber et al., 2012).在植物体内, 具有生物活性的GA主要包括GA1和GA4, 它们通过软化种皮、促进胚乳层细胞的水解和胚的生长打破种子休眠, 进而促进种子萌发(Holdsworth et al., 2008).GA缺失突变体ga1 (gibberellic acid- requiring 1)和ga2表现出强烈的休眠表型, 需要外源施加GA方可萌发(Lee et al., 2002; Shu et al., 2013).GA20ox (GA 20-OXIDASE)和GA3ox基因编码参与GA生物合成的氧化酶; GA2ox编码的GA2-氧化酶主要参与GA的代谢过程.在PHYB-off条件下, ga2ox突变体萌发水平升高(Oh et al., 2006; Yamauchi et al., 2007).上述研究结果表明, GA能通过其生物合成通路调控种子的休眠和萌发.在GA信号转导过程中, DELLA蛋白扮演着重要角色, 它能响应GA信号快速降解, 对GA信号转导起限速作用(Silverstone et al., 1998; Itoh et al., 2002; Zentella et al., 2007; Nemoto et al., 2017).GA受体蛋白GID1 (GA-INSENSITIVE DWARF 1)能够负调控DELLA蛋白的稳定性, 促进GA信号转导(Davière and Achard, 2013).拟南芥SLY1 (SLEEPY1)和水稻GID2 (GA-INSENSITIVE DWARF2)是SCF聚合体中的F-box亚基, 依赖其C端的GGF和LSL基序与DELLA蛋白C端VHIID和LHR2基序结合, 促进DELLA多聚泛素化(Hirano et al., 2010; Ariizumi et al., 2011).研究表明, DELLA蛋白GAI (GA-INSENSITIVE)能够通过抑制TCP14 (TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR 14)和TCP15的表达减缓细胞周期的进程, 最终使种子保持休眠状态(Resentini et al., 2015).作为转录调节子, DELLA如何实现对下游基因的转录调控? 有研究指出, DELLA能够通过与染色质重塑因子互作调控下游基因的转录.DELLA蛋白RGL2 (RGA-LIKE 2)和RGL3与染色质重塑因子SWI3C (SWITCH3C)发生互作, 进而影响GID1a以及GA3ox基因的转录(Sarnowska et al., 2013).此外, DELLA还能通过与种子萌发相关转录因子互作进而实现其对萌发的调控.RGL2通过与转录因子NF-YC互作, 诱导下游ABI5的转录, 进而抑制种子萌发(Liu et al., 2016).此外, RGL2还能通过与转录因子DOF6 (BINDING1 ZINC FINGER 6)互作诱导GATA12的表达, 进而促进种子休眠(Ravindran et al., 2017).DELLA通过与ICE1 (INDUCER OF CBF EXPRESSION 1)互作解除其对ABA响应基因EM1 (LATE EMBRYOGENESIS ABUNDANT 1)和EM6的转录抑制作用, 最终抑制种子萌发(Hu et al., 2019). ...
BRASSINOSTEROID INSENSITIVE 2 interacts with ABSCISIC ACID INSENSITIVE 5 to mediate the antagonism of brassinosteroids to abscisic acid during seed germination in Arabidopsis 1 2014
... 除ABA和GA之外, 种子的休眠和萌发还受到7类激素的调控, 包括生长素(auxin)、乙烯(ethylene, ET)、油菜素内酯(brassinosteroids, BRs)、水杨酸(salicylic acid, SA)、茉莉酸(jasmonic acid, JA)、细胞分裂素(cytokinins, CTKs)和独脚金内酯(strigolactones, SLs).这些激素均通过影响ABA或GA通路间接调控种子的休眠和萌发(Shu et al., 2016; Née et al., 2017a).生长素能够通过调控ABI3的转录水平影响种子的休眠和萌发.当内源生长素水平升高时, 生长素信号通路中的负调控因子AXR2/3 (AUXIN- RESISTANT 2/3)发生降解.此时AXR2/3对响应生长素信号的ARF10 (AUXIN RESPONSE FACTOR 10)和ARF16基因的转录抑制作用被解除, 而ARF10和ARF16能够激活ABI3的表达, 从而使种子保持休眠状态(Liu et al., 2013a).然而, ARF10和ARF16不能直接结合在ABI3的启动子上(Liu et al., 2013a), 因此有待深入研究以揭示完整的生长素信号传递链.ET能够通过ABA生物合成和信号转导通路调控种子萌发(Cheng et al., 2009; Linkies et al., 2009; Corbineau et al., 2014), 但具体机制还不清楚.在盐胁迫条件下, ET受体蛋白ETR1 (ETHYLENE RESPONSE 1)和ETR2可能通过不依赖ET信号的通路调控ABA生物合成, 进而影响种子萌发(Wilson et al., 2014).最近的研究发现, ETR1通过解除ERF12 (ETHYLENE RESPONSE FACTOR 12)/TPL (TOPLESS)模块对DOG1基因的转录抑制作用而促进DOG1的转录, 进而促进种子休眠(Li et al., 2019).BR通过依赖MFT (MOTHER OFFT AND TFL 1)的途径拮抗ABA信号转导通路, 促进种子萌发(Xi and Yu, 2010; Xi et al., 2010).进一步的研究发现, BR信号通路中的负调控因子——BIN2 (BRASSINOSTEROID INSENSITIVE 2), 能够磷酸化并稳定ABI5蛋白, 进而促进ABA信号转导.BR信号通过抑制BIN2-ABI5互作解除ABA对种子萌发的抑制作用(Hu and Yu, 2014).SA在调控种子萌发上发挥双重作用.在正常生长条件下, SA通过抑制GA诱导的α淀粉酶编码基因的表达抑制种子萌发(Xie et al., 2007); 而在高盐胁迫下, SA通过其它通路促进种子萌发(Lee et al., 2010).CTKs通过抑制ABI5的转录或促进ABI5蛋白的降解拮抗ABA效应, 从而促进种子萌发(Wang et al., 2011; Guan et al., 2014).外源施加JA能够延迟种子萌发(Nambara et al., 2010), 表明JA能够抑制种子萌发.有研究表明, JA通过抑制ABA合成相关基因的表达、促进ABA代谢相关基因的表达拮抗ABA, 从而促进种子萌发(Jacobsen et al., 2013).JAZ3 (JASMONATE-ZIM DOMAIN PROTEIN 3)通过与ABI5互作抑制ABI5的转录激活活性, 进而促进种子萌发(Ju et al., 2019).目前关于JA调控种子萌发功能上存在矛盾的原因还有待深入探究.SLs通过降低ABA/GA值促进种子萌发(Toh et al., 2012).此外, SLs信号通路组分也能参与调控种子萌发, 如SMAX1 (SUPPRESSOR OF MORE AXILLARY GROWTH2 1) (Stanga et al., 2013).但目前SLs调控种子萌发的具体机制还不清楚.综上, 生长素、ET、BR、CTKs、JA和SLs通过调控ABA生物合成或信号通路调控种子的休眠和萌发.然而生长素、ET、BR、CTKs、JA与GA之间的交互作用还有待深入研究. ...
Conversion from CUL4-based COP1- SPA E3 apparatus to UVR8-COP1-SPA complexes underlies a distinct biochemical function of COP1 under UV-B 1 2013
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
Early abscisic acid signal transduction mechanisms: newly discovered components and newly emerging questions 1 2010
... 激素在种子休眠和萌发过程中发挥重要调控作用.其中, ABA和GA在调控种子休眠和萌发上行使主要功能.ABA促进种子休眠, 而GA促进种子萌发(Shu et al., 2016; Née et al., 2017a).在种子成熟过程中, ABA在种子内逐渐积累, 使种子的休眠水平逐渐升高.相比之下, 当种子受到吸胀或层积处理时, ABA含量逐渐降低, GA含量逐渐升高, 使种子萌发.与野生型种子相比, ABA合成缺陷突变体种子萌发更快(Frey et al., 2012); 而过表达ABA合成基因的种子以及ABA代谢突变体种子保持较高的休眠水平(Matakiadis et al., 2009; Martinez-Andújar et al., 2011; Nonogaki et al., 2014), 表明ABA能够通过其生物合成通路调控种子的休眠和萌发.ABA信号通过PYR/PYL/RCAR (PYRABACTIN RESISTANCE 1/PYR-LIKE PROTEINS/REGULATORY COMPONENTS OF ABA RECEPTORS)-PP2C (PROTEIN PHOSPHATASE 2C)-SnRKs (SNF1-RELATED PROTEIN KINASES)级联反应向下传递(Cutler et al., 2010; Hubbard et al., 2010).目前鉴定到的PP2C类蛋白包括ABI1 (ABSCISIC ACID INSENSITIVE 1)、ABI2、HON (HONSU)和RDO5 (REDUCED DORMANCY 5).其中, ABI1和ABI2通过与ABA信号受体蛋白PYR/PYL/RCAR互作进而抑制ABA信号转导(Ma et al., 2009; Park et al., 2009).HON蛋白能够通过抑制ABA信号转导及促进GA信号转导抑制种子休眠(Kim et al., 2013), 说明HON能够整合ABA和GA信号以调控种子休眠.rdo5突变体的休眠水平降低, 但其ABA含量和ABA敏感性并未发生显著变化(Xiang et al., 2014), 说明RDO5通过不依赖ABA的途径调控种子休眠.ABI3、ABI4和ABI5是ABA信号通路下游的关键组分, 能够抑制种子萌发、促进种子休眠(Bentsink and Koornneef, 2008; Kanai et al., 2010; Shu et al., 2013).响应ABA信号的R2R3型MYB转录因子MYB96通过调控ABI4以及ABA合成相关基因NCED2 (9-CIS-EPOXYCAROTENOID DIOXYGENASE 2)和NCED6的转录进而促进种子休眠、抑制种子萌发(Lee et al., 2015a, 2015b).上述研究表明, ABA不仅通过其生物合成通路调控种子的休眠和萌发, 还能以ABA信号的形式发挥作用. ...
The gibberellin signaling pathway is regulated by the appearance and disappearance of SLENDER RICE1 in nuclei 1 2002
... GA能够通过抑制ABA诱导的种子休眠促进种子萌发(Gubler et al., 2005; Graeber et al., 2012).在植物体内, 具有生物活性的GA主要包括GA1和GA4, 它们通过软化种皮、促进胚乳层细胞的水解和胚的生长打破种子休眠, 进而促进种子萌发(Holdsworth et al., 2008).GA缺失突变体ga1 (gibberellic acid- requiring 1)和ga2表现出强烈的休眠表型, 需要外源施加GA方可萌发(Lee et al., 2002; Shu et al., 2013).GA20ox (GA 20-OXIDASE)和GA3ox基因编码参与GA生物合成的氧化酶; GA2ox编码的GA2-氧化酶主要参与GA的代谢过程.在PHYB-off条件下, ga2ox突变体萌发水平升高(Oh et al., 2006; Yamauchi et al., 2007).上述研究结果表明, GA能通过其生物合成通路调控种子的休眠和萌发.在GA信号转导过程中, DELLA蛋白扮演着重要角色, 它能响应GA信号快速降解, 对GA信号转导起限速作用(Silverstone et al., 1998; Itoh et al., 2002; Zentella et al., 2007; Nemoto et al., 2017).GA受体蛋白GID1 (GA-INSENSITIVE DWARF 1)能够负调控DELLA蛋白的稳定性, 促进GA信号转导(Davière and Achard, 2013).拟南芥SLY1 (SLEEPY1)和水稻GID2 (GA-INSENSITIVE DWARF2)是SCF聚合体中的F-box亚基, 依赖其C端的GGF和LSL基序与DELLA蛋白C端VHIID和LHR2基序结合, 促进DELLA多聚泛素化(Hirano et al., 2010; Ariizumi et al., 2011).研究表明, DELLA蛋白GAI (GA-INSENSITIVE)能够通过抑制TCP14 (TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR 14)和TCP15的表达减缓细胞周期的进程, 最终使种子保持休眠状态(Resentini et al., 2015).作为转录调节子, DELLA如何实现对下游基因的转录调控? 有研究指出, DELLA能够通过与染色质重塑因子互作调控下游基因的转录.DELLA蛋白RGL2 (RGA-LIKE 2)和RGL3与染色质重塑因子SWI3C (SWITCH3C)发生互作, 进而影响GID1a以及GA3ox基因的转录(Sarnowska et al., 2013).此外, DELLA还能通过与种子萌发相关转录因子互作进而实现其对萌发的调控.RGL2通过与转录因子NF-YC互作, 诱导下游ABI5的转录, 进而抑制种子萌发(Liu et al., 2016).此外, RGL2还能通过与转录因子DOF6 (BINDING1 ZINC FINGER 6)互作诱导GATA12的表达, 进而促进种子休眠(Ravindran et al., 2017).DELLA通过与ICE1 (INDUCER OF CBF EXPRESSION 1)互作解除其对ABA响应基因EM1 (LATE EMBRYOGENESIS ABUNDANT 1)和EM6的转录抑制作用, 最终抑制种子萌发(Hu et al., 2019). ...
Roles for blue light, jasmonate and nitric oxide in the regulation of dormancy and germination in wheat grain ( Triticum aestivum L.) 1 2013
... 除ABA和GA之外, 种子的休眠和萌发还受到7类激素的调控, 包括生长素(auxin)、乙烯(ethylene, ET)、油菜素内酯(brassinosteroids, BRs)、水杨酸(salicylic acid, SA)、茉莉酸(jasmonic acid, JA)、细胞分裂素(cytokinins, CTKs)和独脚金内酯(strigolactones, SLs).这些激素均通过影响ABA或GA通路间接调控种子的休眠和萌发(Shu et al., 2016; Née et al., 2017a).生长素能够通过调控ABI3的转录水平影响种子的休眠和萌发.当内源生长素水平升高时, 生长素信号通路中的负调控因子AXR2/3 (AUXIN- RESISTANT 2/3)发生降解.此时AXR2/3对响应生长素信号的ARF10 (AUXIN RESPONSE FACTOR 10)和ARF16基因的转录抑制作用被解除, 而ARF10和ARF16能够激活ABI3的表达, 从而使种子保持休眠状态(Liu et al., 2013a).然而, ARF10和ARF16不能直接结合在ABI3的启动子上(Liu et al., 2013a), 因此有待深入研究以揭示完整的生长素信号传递链.ET能够通过ABA生物合成和信号转导通路调控种子萌发(Cheng et al., 2009; Linkies et al., 2009; Corbineau et al., 2014), 但具体机制还不清楚.在盐胁迫条件下, ET受体蛋白ETR1 (ETHYLENE RESPONSE 1)和ETR2可能通过不依赖ET信号的通路调控ABA生物合成, 进而影响种子萌发(Wilson et al., 2014).最近的研究发现, ETR1通过解除ERF12 (ETHYLENE RESPONSE FACTOR 12)/TPL (TOPLESS)模块对DOG1基因的转录抑制作用而促进DOG1的转录, 进而促进种子休眠(Li et al., 2019).BR通过依赖MFT (MOTHER OFFT AND TFL 1)的途径拮抗ABA信号转导通路, 促进种子萌发(Xi and Yu, 2010; Xi et al., 2010).进一步的研究发现, BR信号通路中的负调控因子——BIN2 (BRASSINOSTEROID INSENSITIVE 2), 能够磷酸化并稳定ABI5蛋白, 进而促进ABA信号转导.BR信号通过抑制BIN2-ABI5互作解除ABA对种子萌发的抑制作用(Hu and Yu, 2014).SA在调控种子萌发上发挥双重作用.在正常生长条件下, SA通过抑制GA诱导的α淀粉酶编码基因的表达抑制种子萌发(Xie et al., 2007); 而在高盐胁迫下, SA通过其它通路促进种子萌发(Lee et al., 2010).CTKs通过抑制ABI5的转录或促进ABI5蛋白的降解拮抗ABA效应, 从而促进种子萌发(Wang et al., 2011; Guan et al., 2014).外源施加JA能够延迟种子萌发(Nambara et al., 2010), 表明JA能够抑制种子萌发.有研究表明, JA通过抑制ABA合成相关基因的表达、促进ABA代谢相关基因的表达拮抗ABA, 从而促进种子萌发(Jacobsen et al., 2013).JAZ3 (JASMONATE-ZIM DOMAIN PROTEIN 3)通过与ABI5互作抑制ABI5的转录激活活性, 进而促进种子萌发(Ju et al., 2019).目前关于JA调控种子萌发功能上存在矛盾的原因还有待深入探究.SLs通过降低ABA/GA值促进种子萌发(Toh et al., 2012).此外, SLs信号通路组分也能参与调控种子萌发, 如SMAX1 (SUPPRESSOR OF MORE AXILLARY GROWTH2 1) (Stanga et al., 2013).但目前SLs调控种子萌发的具体机制还不清楚.综上, 生长素、ET、BR、CTKs、JA和SLs通过调控ABA生物合成或信号通路调控种子的休眠和萌发.然而生长素、ET、BR、CTKs、JA与GA之间的交互作用还有待深入研究. ...
HFR1 is targeted by COP1 E3 ligase for post-translational proteolysis during phytochrome A signaling 1 2005
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
The E3 ubiquitin ligase COP1 regulates thermosensory flowering by triggering GI degradation in Arabidopsis 1 2015
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
Phytochrome B and REVEILLE1/2-mediated signaling controls seed dormancy and germination in Arabidopsis 4 2016
... PHY在黑暗条件下以生理失活的红光吸收型(Pr)存在, 吸收红光之后转变成其生理激活型(Pfr).两种光吸收型的PHY在Pr和Pfr两种状态间相互转变.早在20世纪50年代, 研究人员以莴苣(Lactuca sativa)种子为材料, 研究红光和远红光对其萌发的影响.结果表明红光促进种子萌发, 而远红光能够逆转红光的作用(Borthwick et al., 1952).红光和远红光对种子萌发的可逆调控暗示着PHY参与调控种子的萌发过程(Shinomura et al., 1994; Hennig et al., 2002).拟南芥PHY基因家族包含5个成员——PHYA-PHYE (Sharrock and Quail, 1989; Clack et al., 1994).种子萌发受到PHYA和PHYB的调控.其中, PHYB发挥主要功能.研究表明, PHYA蛋白在干种子中表达量很低, 随着种子在黑暗条件下吸胀时间的延长其蛋白表达量逐渐增加, 说明PHYA参与调控种子的萌发过程(Shinomura et al., 1996).PHYA主要在种子吸胀后期通过介导红光和远红光条件下的极低辐照度反应(VLFR)和远红光下的高辐照度反应(FR-HIR)调控种子萌发.相比之下, PHYB在干种子和吸胀种子中表达量均很高, 能够在种子吸胀初期(几个小时以内)介导红光和远红光下的低辐照度反应(LFR)调控种子萌发(Seo et al., 2009; Li et al., 2011).除了PHYA和PHYB外, PHYE也参与光调控的种子萌发过程, 而且三者在调控种子萌发方面功能冗余(Hennig et al., 2002).最近有研究表明, PHYB除了调控种子萌发, 还参与调控种子休眠(Jiang et al., 2016). ...
... 拟南芥基因组编码8个PIFs (PHYTOCHROME- INTERACTING FACTORs, PIF1-PIF8)蛋白.其中, PIF1和PIF6分别调控种子萌发和休眠(Oh et al., 2006; Penfield et al., 2010).PIF1作为种子萌发的负调控因子, 能够将内源激素和外源光信号连接起来, 在光介导的种子萌发过程中发挥关键作用.研究表明, 外源光信号通过调控PIF1蛋白稳定性或转录活性影响其对下游基因的转录调控.光照条件下, PIF1蛋白能够通过与Pfr形式的PHY互作, 进而发生泛素化降解(Oh et al., 2004; 2006); HFR1 (LONG HYPOCOTYL IN FAR-RED 1)通过PIF1的C端与之发生互作, 进而干扰PIF1的转录活性, 最终促进种子萌发(Shi et al., 2013).在黑暗条件下, DET1 (DE-ETIOLATED 1)和COP10以一种未知的机制稳定PIF1蛋白的活性(Shi et al., 2015).bHLH转录因子SPT (SPATULA)作为种子萌发的正调控因子, 在种子萌发中也发挥重要作用(Vaistij et al., 2018).此外, SPT还能调控种子休眠.然而, 在不同的生态型背景下SPT调控种子休眠的功能不同.在Ler背景下, SPT抑制种子休眠; 而在Col背景下, SPT促进种子休眠(Vaistij et al., 2013).外源光信号能否影响SPT对下游基因的转录调控? 目前还需阐释PHY调控SPT转录活性的作用机制, 以解答上述问题.最近的研究表明, 昼夜节律钟的关键组分CCA1 (CIRCADIAN CLOCK ASSOCIATED 1)、LHY (LATE ELONGATED HYPOCOTYL)以及RVE1 (REVEILLE1)均能介导外源光信号调控的种子休眠(Penfield and Hall, 2009; Jiang et al., 2016).此外, RVE1还能调控种子萌发(Jiang et al., 2016).IMB1 (IMBIBITION-INDUCIBLE1)是染色质域蛋白家族成员, 在PHYA介导的种子萌发中发挥作用.IMB1在干种子中的表达量很低, 随着种子吸胀呈现上调表达, 暗示IMB1能够促进种子萌发(Duque and Chua, 2003).目前, 关于IMB1调控PHYA介导的种子萌发的分子机制还有待深入探究.CSN (COP9 SIGNALOSOME)蛋白复合体是一类保守的蛋白复合体, 能够调控RING型E3泛素化连接酶的活性.CSN包含8个亚基(CSN1-CSN8) (Wei and Deng, 2003).其中, CSN1和CSN5参与调控种子萌发.csn1-10和csn5a-1突变体呈现出种子延迟萌发的表型(Jin et al., 2018). ...
... ).此外, RVE1还能调控种子萌发(Jiang et al., 2016).IMB1 (IMBIBITION-INDUCIBLE1)是染色质域蛋白家族成员, 在PHYA介导的种子萌发中发挥作用.IMB1在干种子中的表达量很低, 随着种子吸胀呈现上调表达, 暗示IMB1能够促进种子萌发(Duque and Chua, 2003).目前, 关于IMB1调控PHYA介导的种子萌发的分子机制还有待深入探究.CSN (COP9 SIGNALOSOME)蛋白复合体是一类保守的蛋白复合体, 能够调控RING型E3泛素化连接酶的活性.CSN包含8个亚基(CSN1-CSN8) (Wei and Deng, 2003).其中, CSN1和CSN5参与调控种子萌发.csn1-10和csn5a-1突变体呈现出种子延迟萌发的表型(Jin et al., 2018). ...
The COP9 signalosome regulates seed germination by facilitating protein degradation of RGL2 and ABI5 2 2018
... 拟南芥基因组编码8个PIFs (PHYTOCHROME- INTERACTING FACTORs, PIF1-PIF8)蛋白.其中, PIF1和PIF6分别调控种子萌发和休眠(Oh et al., 2006; Penfield et al., 2010).PIF1作为种子萌发的负调控因子, 能够将内源激素和外源光信号连接起来, 在光介导的种子萌发过程中发挥关键作用.研究表明, 外源光信号通过调控PIF1蛋白稳定性或转录活性影响其对下游基因的转录调控.光照条件下, PIF1蛋白能够通过与Pfr形式的PHY互作, 进而发生泛素化降解(Oh et al., 2004; 2006); HFR1 (LONG HYPOCOTYL IN FAR-RED 1)通过PIF1的C端与之发生互作, 进而干扰PIF1的转录活性, 最终促进种子萌发(Shi et al., 2013).在黑暗条件下, DET1 (DE-ETIOLATED 1)和COP10以一种未知的机制稳定PIF1蛋白的活性(Shi et al., 2015).bHLH转录因子SPT (SPATULA)作为种子萌发的正调控因子, 在种子萌发中也发挥重要作用(Vaistij et al., 2018).此外, SPT还能调控种子休眠.然而, 在不同的生态型背景下SPT调控种子休眠的功能不同.在Ler背景下, SPT抑制种子休眠; 而在Col背景下, SPT促进种子休眠(Vaistij et al., 2013).外源光信号能否影响SPT对下游基因的转录调控? 目前还需阐释PHY调控SPT转录活性的作用机制, 以解答上述问题.最近的研究表明, 昼夜节律钟的关键组分CCA1 (CIRCADIAN CLOCK ASSOCIATED 1)、LHY (LATE ELONGATED HYPOCOTYL)以及RVE1 (REVEILLE1)均能介导外源光信号调控的种子休眠(Penfield and Hall, 2009; Jiang et al., 2016).此外, RVE1还能调控种子萌发(Jiang et al., 2016).IMB1 (IMBIBITION-INDUCIBLE1)是染色质域蛋白家族成员, 在PHYA介导的种子萌发中发挥作用.IMB1在干种子中的表达量很低, 随着种子吸胀呈现上调表达, 暗示IMB1能够促进种子萌发(Duque and Chua, 2003).目前, 关于IMB1调控PHYA介导的种子萌发的分子机制还有待深入探究.CSN (COP9 SIGNALOSOME)蛋白复合体是一类保守的蛋白复合体, 能够调控RING型E3泛素化连接酶的活性.CSN包含8个亚基(CSN1-CSN8) (Wei and Deng, 2003).其中, CSN1和CSN5参与调控种子萌发.csn1-10和csn5a-1突变体呈现出种子延迟萌发的表型(Jin et al., 2018). ...
... 为了进一步探究外源光信号能否通过调控GA的信号转导影响种子萌发, 研究者检测了ga1突变体和pif1/ga1双突变体种子萌发对GA3的敏感性.结果发现, 与ga1突变体相比, pif1/ga1双突变体对GA3的敏感性升高.说明PIF1可能参与调控GA信号转导.进一步检测GA信号转导通路中关键组分的转录水平, 发现PIF1能够促进DELLA蛋白编码基因GAI和RGA (REPRESSOR OF GA1-3)的表达.该研究进一步证实PIF1能够直接结合在GAI和RGA的启动子上.上述结果表明, 黑暗条件下, PIF1蛋白能够直接诱导GAI和RGA的表达, 从而抑制GA信号转导, 最终抑制种子萌发(Oh et al., 2007).此外, PIF1还能通过诱导ABI3和ABI5的转录促进ABA信号转导, 从而抑制种子萌发(Park et al., 2011).SPT在不同生态型拟南芥背景下调控种子休眠的功能和作用机制不同.在Col背景下, SPT诱导ABI5和RGL3的表达, 从而促进ABA、抑制GA的信号转导, 最终促进种子休眠.在Ler背景下, SPT抑制ABI4和RGA的表达, 从而抑制ABA、促进GA的信号转导, 最终抑制种子休眠(Vaistij et al., 2013).此外, SPT还能通过抑制ABI5表达促进种子萌发(Vaistij et al., 2018).imb1功能缺失突变体的萌发率降低, 对ABA超敏感.进一步研究发现, ABI5转录本的积累可能是造成imb1突变体对ABA超敏感的原因之一(Duque and Chua, 2003).上述结果暗示, IMB1可能通过抑制ABA信号转导通路促进种子萌发(Duque and Chua, 2003).CSN蛋白复合体亚基CSN1和CSN5a的功能缺失突变体表现出延迟萌发的表型.去除种皮后csn1-10和csn5a-1能够正常萌发, 说明csn1-10和csn5a-1延迟萌发的表型依赖于种皮.种皮抑制的种子萌发与胚乳中RGL2相关, RGL2通过促进ABA的合成以及稳定ABI5蛋白进而抑制种子萌发.说明CSN1和CSN5a可能通过调控GA和ABA的信号转导促进种子萌发.进一步研究发现, CSN1通过促进RGL2的泛素化降解促进GA信号转导, 从而促进种子萌发; CSN5a通过抑制ABI5蛋白的稳定性进而抑制ABA信号转导, 最终促进种子萌发(Jin et al., 2018).上述研究结果表明, 外源光信号还能通过调控GA和ABA信号转导影响种子的休眠与萌发(图1A, B). ...
JAZ proteins modulate seed germination through interaction with ABI5 in bread wheat and Arabidopsis 1 2019
... 除ABA和GA之外, 种子的休眠和萌发还受到7类激素的调控, 包括生长素(auxin)、乙烯(ethylene, ET)、油菜素内酯(brassinosteroids, BRs)、水杨酸(salicylic acid, SA)、茉莉酸(jasmonic acid, JA)、细胞分裂素(cytokinins, CTKs)和独脚金内酯(strigolactones, SLs).这些激素均通过影响ABA或GA通路间接调控种子的休眠和萌发(Shu et al., 2016; Née et al., 2017a).生长素能够通过调控ABI3的转录水平影响种子的休眠和萌发.当内源生长素水平升高时, 生长素信号通路中的负调控因子AXR2/3 (AUXIN- RESISTANT 2/3)发生降解.此时AXR2/3对响应生长素信号的ARF10 (AUXIN RESPONSE FACTOR 10)和ARF16基因的转录抑制作用被解除, 而ARF10和ARF16能够激活ABI3的表达, 从而使种子保持休眠状态(Liu et al., 2013a).然而, ARF10和ARF16不能直接结合在ABI3的启动子上(Liu et al., 2013a), 因此有待深入研究以揭示完整的生长素信号传递链.ET能够通过ABA生物合成和信号转导通路调控种子萌发(Cheng et al., 2009; Linkies et al., 2009; Corbineau et al., 2014), 但具体机制还不清楚.在盐胁迫条件下, ET受体蛋白ETR1 (ETHYLENE RESPONSE 1)和ETR2可能通过不依赖ET信号的通路调控ABA生物合成, 进而影响种子萌发(Wilson et al., 2014).最近的研究发现, ETR1通过解除ERF12 (ETHYLENE RESPONSE FACTOR 12)/TPL (TOPLESS)模块对DOG1基因的转录抑制作用而促进DOG1的转录, 进而促进种子休眠(Li et al., 2019).BR通过依赖MFT (MOTHER OFFT AND TFL 1)的途径拮抗ABA信号转导通路, 促进种子萌发(Xi and Yu, 2010; Xi et al., 2010).进一步的研究发现, BR信号通路中的负调控因子——BIN2 (BRASSINOSTEROID INSENSITIVE 2), 能够磷酸化并稳定ABI5蛋白, 进而促进ABA信号转导.BR信号通过抑制BIN2-ABI5互作解除ABA对种子萌发的抑制作用(Hu and Yu, 2014).SA在调控种子萌发上发挥双重作用.在正常生长条件下, SA通过抑制GA诱导的α淀粉酶编码基因的表达抑制种子萌发(Xie et al., 2007); 而在高盐胁迫下, SA通过其它通路促进种子萌发(Lee et al., 2010).CTKs通过抑制ABI5的转录或促进ABI5蛋白的降解拮抗ABA效应, 从而促进种子萌发(Wang et al., 2011; Guan et al., 2014).外源施加JA能够延迟种子萌发(Nambara et al., 2010), 表明JA能够抑制种子萌发.有研究表明, JA通过抑制ABA合成相关基因的表达、促进ABA代谢相关基因的表达拮抗ABA, 从而促进种子萌发(Jacobsen et al., 2013).JAZ3 (JASMONATE-ZIM DOMAIN PROTEIN 3)通过与ABI5互作抑制ABI5的转录激活活性, 进而促进种子萌发(Ju et al., 2019).目前关于JA调控种子萌发功能上存在矛盾的原因还有待深入探究.SLs通过降低ABA/GA值促进种子萌发(Toh et al., 2012).此外, SLs信号通路组分也能参与调控种子萌发, 如SMAX1 (SUPPRESSOR OF MORE AXILLARY GROWTH2 1) (Stanga et al., 2013).但目前SLs调控种子萌发的具体机制还不清楚.综上, 生长素、ET、BR、CTKs、JA和SLs通过调控ABA生物合成或信号通路调控种子的休眠和萌发.然而生长素、ET、BR、CTKs、JA与GA之间的交互作用还有待深入研究. ...
A peroxisomal ABC transporter promotes seed germination by inducing pectin degradation under the control of ABI5 1 2010
... 激素在种子休眠和萌发过程中发挥重要调控作用.其中, ABA和GA在调控种子休眠和萌发上行使主要功能.ABA促进种子休眠, 而GA促进种子萌发(Shu et al., 2016; Née et al., 2017a).在种子成熟过程中, ABA在种子内逐渐积累, 使种子的休眠水平逐渐升高.相比之下, 当种子受到吸胀或层积处理时, ABA含量逐渐降低, GA含量逐渐升高, 使种子萌发.与野生型种子相比, ABA合成缺陷突变体种子萌发更快(Frey et al., 2012); 而过表达ABA合成基因的种子以及ABA代谢突变体种子保持较高的休眠水平(Matakiadis et al., 2009; Martinez-Andújar et al., 2011; Nonogaki et al., 2014), 表明ABA能够通过其生物合成通路调控种子的休眠和萌发.ABA信号通过PYR/PYL/RCAR (PYRABACTIN RESISTANCE 1/PYR-LIKE PROTEINS/REGULATORY COMPONENTS OF ABA RECEPTORS)-PP2C (PROTEIN PHOSPHATASE 2C)-SnRKs (SNF1-RELATED PROTEIN KINASES)级联反应向下传递(Cutler et al., 2010; Hubbard et al., 2010).目前鉴定到的PP2C类蛋白包括ABI1 (ABSCISIC ACID INSENSITIVE 1)、ABI2、HON (HONSU)和RDO5 (REDUCED DORMANCY 5).其中, ABI1和ABI2通过与ABA信号受体蛋白PYR/PYL/RCAR互作进而抑制ABA信号转导(Ma et al., 2009; Park et al., 2009).HON蛋白能够通过抑制ABA信号转导及促进GA信号转导抑制种子休眠(Kim et al., 2013), 说明HON能够整合ABA和GA信号以调控种子休眠.rdo5突变体的休眠水平降低, 但其ABA含量和ABA敏感性并未发生显著变化(Xiang et al., 2014), 说明RDO5通过不依赖ABA的途径调控种子休眠.ABI3、ABI4和ABI5是ABA信号通路下游的关键组分, 能够抑制种子萌发、促进种子休眠(Bentsink and Koornneef, 2008; Kanai et al., 2010; Shu et al., 2013).响应ABA信号的R2R3型MYB转录因子MYB96通过调控ABI4以及ABA合成相关基因NCED2 (9-CIS-EPOXYCAROTENOID DIOXYGENASE 2)和NCED6的转录进而促进种子休眠、抑制种子萌发(Lee et al., 2015a, 2015b).上述研究表明, ABA不仅通过其生物合成通路调控种子的休眠和萌发, 还能以ABA信号的形式发挥作用. ...
SOMNUS, a CCCH-type zinc finger protein in Arabidopsis, negatively regulates light- dependent seed germination downstream of PIL5 2 2008
... PIF1能够介导外源光信号, 通过调控ABA和GA的生物合成调控种子萌发(图1A).统计Col和pif1突变体的种子在PHYB-on和PHYB-off条件下的萌发率, 发现Col只能在PHYB-on条件下萌发, pif1在PHYB-on和PHYB-off条件下均能萌发; 使用GA合成抑制剂PAC (paclobutrazol)能够抑制Col和pif1萌发.上述研究结果表明, PIF1在PHYB介导的种子萌发过程中发挥重要作用, 它可能通过调控GA的生物合成抑制种子萌发.进一步研究证实, PIF1能够抑制GA合成相关基因GA3ox1和GA3ox2的表达, 促进GA代谢相关基因GA2ox2的表达, 进而下调内源GA4的水平, 抑制种子萌发(Oh et al., 2006).与ga1单突变体相比, pif1/ga1双突变体中ABA合成相关基因的表达下调; 而ABA代谢相关基因的表达水平上调, 致使pif1/ga1双突变体中ABA的水平降低, 表现出持续萌发的表型(Oh et al., 2007).那么, PIF1如何调控ABA和GA生物合成相关基因的转录? 后续的研究发现, PIF1能够通过诱导SOM (SOMNUS)和DAG1 (DOF AFFECTING GERMINATION 1)的转录间接抑制GA合成、促进ABA合成, 最终抑制种子萌发(Kim et al., 2008; Gabriele et al., 2010) (图1A).其中, CCCH型锌指蛋白SOM通过调控组蛋白精氨酸去甲基化酶编码基因JMJ20 (JUMONJI 20)和JMJ22的表达, 进而影响GA合成基因GA3ox1和GA3ox2的甲基化水平, 最终抑制种子的萌发过程.此外, SOM还能激活GA代谢基因GA2ox2以及ABA合成基因ABA1 (ABA- DEFICIENT 1)、NCED6和NCED9的表达, 抑制ABA代谢基因CYP707A2表达, 从而抑制GA生物合成、促进ABA合成, 最终抑制种子萌发(Kim et al., 2008; de Wit et al., 2016).包含Dof结构域的DAG1蛋白直接结合在GA3ox1启动子上抑制其表达, 进而抑制种子萌发(Gabriele et al., 2010). ...
... 表达, 从而抑制GA生物合成、促进ABA合成, 最终抑制种子萌发(Kim et al., 2008; de Wit et al., 2016).包含Dof结构域的DAG1蛋白直接结合在GA3ox1启动子上抑制其表达, 进而抑制种子萌发(Gabriele et al., 2010). ...
HONSU, a protein phosphatase 2C, regulates seed dormancy by inhibiting ABA signaling in Arabidopsis 1 2013
... 激素在种子休眠和萌发过程中发挥重要调控作用.其中, ABA和GA在调控种子休眠和萌发上行使主要功能.ABA促进种子休眠, 而GA促进种子萌发(Shu et al., 2016; Née et al., 2017a).在种子成熟过程中, ABA在种子内逐渐积累, 使种子的休眠水平逐渐升高.相比之下, 当种子受到吸胀或层积处理时, ABA含量逐渐降低, GA含量逐渐升高, 使种子萌发.与野生型种子相比, ABA合成缺陷突变体种子萌发更快(Frey et al., 2012); 而过表达ABA合成基因的种子以及ABA代谢突变体种子保持较高的休眠水平(Matakiadis et al., 2009; Martinez-Andújar et al., 2011; Nonogaki et al., 2014), 表明ABA能够通过其生物合成通路调控种子的休眠和萌发.ABA信号通过PYR/PYL/RCAR (PYRABACTIN RESISTANCE 1/PYR-LIKE PROTEINS/REGULATORY COMPONENTS OF ABA RECEPTORS)-PP2C (PROTEIN PHOSPHATASE 2C)-SnRKs (SNF1-RELATED PROTEIN KINASES)级联反应向下传递(Cutler et al., 2010; Hubbard et al., 2010).目前鉴定到的PP2C类蛋白包括ABI1 (ABSCISIC ACID INSENSITIVE 1)、ABI2、HON (HONSU)和RDO5 (REDUCED DORMANCY 5).其中, ABI1和ABI2通过与ABA信号受体蛋白PYR/PYL/RCAR互作进而抑制ABA信号转导(Ma et al., 2009; Park et al., 2009).HON蛋白能够通过抑制ABA信号转导及促进GA信号转导抑制种子休眠(Kim et al., 2013), 说明HON能够整合ABA和GA信号以调控种子休眠.rdo5突变体的休眠水平降低, 但其ABA含量和ABA敏感性并未发生显著变化(Xiang et al., 2014), 说明RDO5通过不依赖ABA的途径调控种子休眠.ABI3、ABI4和ABI5是ABA信号通路下游的关键组分, 能够抑制种子萌发、促进种子休眠(Bentsink and Koornneef, 2008; Kanai et al., 2010; Shu et al., 2013).响应ABA信号的R2R3型MYB转录因子MYB96通过调控ABI4以及ABA合成相关基因NCED2 (9-CIS-EPOXYCAROTENOID DIOXYGENASE 2)和NCED6的转录进而促进种子休眠、抑制种子萌发(Lee et al., 2015a, 2015b).上述研究表明, ABA不仅通过其生物合成通路调控种子的休眠和萌发, 还能以ABA信号的形式发挥作用. ...
The Arabidopsis MYB96 transcription factor plays a role in seed dormancy 1 2015a
... 激素在种子休眠和萌发过程中发挥重要调控作用.其中, ABA和GA在调控种子休眠和萌发上行使主要功能.ABA促进种子休眠, 而GA促进种子萌发(Shu et al., 2016; Née et al., 2017a).在种子成熟过程中, ABA在种子内逐渐积累, 使种子的休眠水平逐渐升高.相比之下, 当种子受到吸胀或层积处理时, ABA含量逐渐降低, GA含量逐渐升高, 使种子萌发.与野生型种子相比, ABA合成缺陷突变体种子萌发更快(Frey et al., 2012); 而过表达ABA合成基因的种子以及ABA代谢突变体种子保持较高的休眠水平(Matakiadis et al., 2009; Martinez-Andújar et al., 2011; Nonogaki et al., 2014), 表明ABA能够通过其生物合成通路调控种子的休眠和萌发.ABA信号通过PYR/PYL/RCAR (PYRABACTIN RESISTANCE 1/PYR-LIKE PROTEINS/REGULATORY COMPONENTS OF ABA RECEPTORS)-PP2C (PROTEIN PHOSPHATASE 2C)-SnRKs (SNF1-RELATED PROTEIN KINASES)级联反应向下传递(Cutler et al., 2010; Hubbard et al., 2010).目前鉴定到的PP2C类蛋白包括ABI1 (ABSCISIC ACID INSENSITIVE 1)、ABI2、HON (HONSU)和RDO5 (REDUCED DORMANCY 5).其中, ABI1和ABI2通过与ABA信号受体蛋白PYR/PYL/RCAR互作进而抑制ABA信号转导(Ma et al., 2009; Park et al., 2009).HON蛋白能够通过抑制ABA信号转导及促进GA信号转导抑制种子休眠(Kim et al., 2013), 说明HON能够整合ABA和GA信号以调控种子休眠.rdo5突变体的休眠水平降低, 但其ABA含量和ABA敏感性并未发生显著变化(Xiang et al., 2014), 说明RDO5通过不依赖ABA的途径调控种子休眠.ABI3、ABI4和ABI5是ABA信号通路下游的关键组分, 能够抑制种子萌发、促进种子休眠(Bentsink and Koornneef, 2008; Kanai et al., 2010; Shu et al., 2013).响应ABA信号的R2R3型MYB转录因子MYB96通过调控ABI4以及ABA合成相关基因NCED2 (9-CIS-EPOXYCAROTENOID DIOXYGENASE 2)和NCED6的转录进而促进种子休眠、抑制种子萌发(Lee et al., 2015a, 2015b).上述研究表明, ABA不仅通过其生物合成通路调控种子的休眠和萌发, 还能以ABA信号的形式发挥作用. ...
The Arabidopsis MYB96 transcription factor is a positive regulator of ABSCISIC ACID-INSENSITIVE 4 in the control of seed germination 1 2015b
... 激素在种子休眠和萌发过程中发挥重要调控作用.其中, ABA和GA在调控种子休眠和萌发上行使主要功能.ABA促进种子休眠, 而GA促进种子萌发(Shu et al., 2016; Née et al., 2017a).在种子成熟过程中, ABA在种子内逐渐积累, 使种子的休眠水平逐渐升高.相比之下, 当种子受到吸胀或层积处理时, ABA含量逐渐降低, GA含量逐渐升高, 使种子萌发.与野生型种子相比, ABA合成缺陷突变体种子萌发更快(Frey et al., 2012); 而过表达ABA合成基因的种子以及ABA代谢突变体种子保持较高的休眠水平(Matakiadis et al., 2009; Martinez-Andújar et al., 2011; Nonogaki et al., 2014), 表明ABA能够通过其生物合成通路调控种子的休眠和萌发.ABA信号通过PYR/PYL/RCAR (PYRABACTIN RESISTANCE 1/PYR-LIKE PROTEINS/REGULATORY COMPONENTS OF ABA RECEPTORS)-PP2C (PROTEIN PHOSPHATASE 2C)-SnRKs (SNF1-RELATED PROTEIN KINASES)级联反应向下传递(Cutler et al., 2010; Hubbard et al., 2010).目前鉴定到的PP2C类蛋白包括ABI1 (ABSCISIC ACID INSENSITIVE 1)、ABI2、HON (HONSU)和RDO5 (REDUCED DORMANCY 5).其中, ABI1和ABI2通过与ABA信号受体蛋白PYR/PYL/RCAR互作进而抑制ABA信号转导(Ma et al., 2009; Park et al., 2009).HON蛋白能够通过抑制ABA信号转导及促进GA信号转导抑制种子休眠(Kim et al., 2013), 说明HON能够整合ABA和GA信号以调控种子休眠.rdo5突变体的休眠水平降低, 但其ABA含量和ABA敏感性并未发生显著变化(Xiang et al., 2014), 说明RDO5通过不依赖ABA的途径调控种子休眠.ABI3、ABI4和ABI5是ABA信号通路下游的关键组分, 能够抑制种子萌发、促进种子休眠(Bentsink and Koornneef, 2008; Kanai et al., 2010; Shu et al., 2013).响应ABA信号的R2R3型MYB转录因子MYB96通过调控ABI4以及ABA合成相关基因NCED2 (9-CIS-EPOXYCAROTENOID DIOXYGENASE 2)和NCED6的转录进而促进种子休眠、抑制种子萌发(Lee et al., 2015a, 2015b).上述研究表明, ABA不仅通过其生物合成通路调控种子的休眠和萌发, 还能以ABA信号的形式发挥作用. ...
Spatially and genetically distinct control of seed germination by phytochromes A and B 1 2012
... 种子休眠是指有活力的种子在适宜的条件下暂时不萌发的现象(Bewley, 1997; Finch-Savage and Leubner-Metzger, 2006).种子的休眠包括休眠的诱导、维持和释放3个阶段(Shu et al., 2016).成熟的种子释放休眠后, 在适宜的条件下即可萌发.DOG1 (DELAY OF GERMINATION 1)在种子休眠诱导和维持中发挥重要作用.它既能通过抑制脱落酸(abscisic acid, ABA)通路的负调控因子——AHG1(ABA HYPERSENSITIVE GERMINATION 1)的磷酸酶活性促进种子休眠(Nishimura et al., 2018); 也能通过调控响应赤霉素(gibberellin, GA)信号的细胞壁重塑相关基因的表达促进种子休眠(Graeber et al., 2014).在种子吸胀初期, 胚乳中PHYB (PHYTOCHROME B)吸收远红光后以生理失活型存在, 此时PIF1 (PIF3-LIKE 5, PIL5)蛋白逐渐积累, 使胚乳中ABA水平逐渐升高.胚乳中的ABA信号能够释放到胚中并抑制GA的生物合成, 使种子保持休眠状态.随着种子吸胀时间的延长, 胚乳中ABA水平逐渐下调.此时胚中的PHYA介导远红光促进GA的生物合成, 进而促进种子萌发(Lee et al., 2012).因此, 种子的休眠和萌发是两个相互独立的生物学过程. ...
Gibberellin regulates Arabidopsis seed germination via RGL2, a GAIIRGA-like gene whose expression is up-regulated following imbibition 1 2002
... GA能够通过抑制ABA诱导的种子休眠促进种子萌发(Gubler et al., 2005; Graeber et al., 2012).在植物体内, 具有生物活性的GA主要包括GA1和GA4, 它们通过软化种皮、促进胚乳层细胞的水解和胚的生长打破种子休眠, 进而促进种子萌发(Holdsworth et al., 2008).GA缺失突变体ga1 (gibberellic acid- requiring 1)和ga2表现出强烈的休眠表型, 需要外源施加GA方可萌发(Lee et al., 2002; Shu et al., 2013).GA20ox (GA 20-OXIDASE)和GA3ox基因编码参与GA生物合成的氧化酶; GA2ox编码的GA2-氧化酶主要参与GA的代谢过程.在PHYB-off条件下, ga2ox突变体萌发水平升高(Oh et al., 2006; Yamauchi et al., 2007).上述研究结果表明, GA能通过其生物合成通路调控种子的休眠和萌发.在GA信号转导过程中, DELLA蛋白扮演着重要角色, 它能响应GA信号快速降解, 对GA信号转导起限速作用(Silverstone et al., 1998; Itoh et al., 2002; Zentella et al., 2007; Nemoto et al., 2017).GA受体蛋白GID1 (GA-INSENSITIVE DWARF 1)能够负调控DELLA蛋白的稳定性, 促进GA信号转导(Davière and Achard, 2013).拟南芥SLY1 (SLEEPY1)和水稻GID2 (GA-INSENSITIVE DWARF2)是SCF聚合体中的F-box亚基, 依赖其C端的GGF和LSL基序与DELLA蛋白C端VHIID和LHR2基序结合, 促进DELLA多聚泛素化(Hirano et al., 2010; Ariizumi et al., 2011).研究表明, DELLA蛋白GAI (GA-INSENSITIVE)能够通过抑制TCP14 (TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR 14)和TCP15的表达减缓细胞周期的进程, 最终使种子保持休眠状态(Resentini et al., 2015).作为转录调节子, DELLA如何实现对下游基因的转录调控? 有研究指出, DELLA能够通过与染色质重塑因子互作调控下游基因的转录.DELLA蛋白RGL2 (RGA-LIKE 2)和RGL3与染色质重塑因子SWI3C (SWITCH3C)发生互作, 进而影响GID1a以及GA3ox基因的转录(Sarnowska et al., 2013).此外, DELLA还能通过与种子萌发相关转录因子互作进而实现其对萌发的调控.RGL2通过与转录因子NF-YC互作, 诱导下游ABI5的转录, 进而抑制种子萌发(Liu et al., 2016).此外, RGL2还能通过与转录因子DOF6 (BINDING1 ZINC FINGER 6)互作诱导GATA12的表达, 进而促进种子休眠(Ravindran et al., 2017).DELLA通过与ICE1 (INDUCER OF CBF EXPRESSION 1)互作解除其对ABA响应基因EM1 (LATE EMBRYOGENESIS ABUNDANT 1)和EM6的转录抑制作用, 最终抑制种子萌发(Hu et al., 2019). ...
Salicylic acid promotes seed germination under high salinity by modulating antioxidant activity in Arabidopsis 1 2010
... 除ABA和GA之外, 种子的休眠和萌发还受到7类激素的调控, 包括生长素(auxin)、乙烯(ethylene, ET)、油菜素内酯(brassinosteroids, BRs)、水杨酸(salicylic acid, SA)、茉莉酸(jasmonic acid, JA)、细胞分裂素(cytokinins, CTKs)和独脚金内酯(strigolactones, SLs).这些激素均通过影响ABA或GA通路间接调控种子的休眠和萌发(Shu et al., 2016; Née et al., 2017a).生长素能够通过调控ABI3的转录水平影响种子的休眠和萌发.当内源生长素水平升高时, 生长素信号通路中的负调控因子AXR2/3 (AUXIN- RESISTANT 2/3)发生降解.此时AXR2/3对响应生长素信号的ARF10 (AUXIN RESPONSE FACTOR 10)和ARF16基因的转录抑制作用被解除, 而ARF10和ARF16能够激活ABI3的表达, 从而使种子保持休眠状态(Liu et al., 2013a).然而, ARF10和ARF16不能直接结合在ABI3的启动子上(Liu et al., 2013a), 因此有待深入研究以揭示完整的生长素信号传递链.ET能够通过ABA生物合成和信号转导通路调控种子萌发(Cheng et al., 2009; Linkies et al., 2009; Corbineau et al., 2014), 但具体机制还不清楚.在盐胁迫条件下, ET受体蛋白ETR1 (ETHYLENE RESPONSE 1)和ETR2可能通过不依赖ET信号的通路调控ABA生物合成, 进而影响种子萌发(Wilson et al., 2014).最近的研究发现, ETR1通过解除ERF12 (ETHYLENE RESPONSE FACTOR 12)/TPL (TOPLESS)模块对DOG1基因的转录抑制作用而促进DOG1的转录, 进而促进种子休眠(Li et al., 2019).BR通过依赖MFT (MOTHER OFFT AND TFL 1)的途径拮抗ABA信号转导通路, 促进种子萌发(Xi and Yu, 2010; Xi et al., 2010).进一步的研究发现, BR信号通路中的负调控因子——BIN2 (BRASSINOSTEROID INSENSITIVE 2), 能够磷酸化并稳定ABI5蛋白, 进而促进ABA信号转导.BR信号通过抑制BIN2-ABI5互作解除ABA对种子萌发的抑制作用(Hu and Yu, 2014).SA在调控种子萌发上发挥双重作用.在正常生长条件下, SA通过抑制GA诱导的α淀粉酶编码基因的表达抑制种子萌发(Xie et al., 2007); 而在高盐胁迫下, SA通过其它通路促进种子萌发(Lee et al., 2010).CTKs通过抑制ABI5的转录或促进ABI5蛋白的降解拮抗ABA效应, 从而促进种子萌发(Wang et al., 2011; Guan et al., 2014).外源施加JA能够延迟种子萌发(Nambara et al., 2010), 表明JA能够抑制种子萌发.有研究表明, JA通过抑制ABA合成相关基因的表达、促进ABA代谢相关基因的表达拮抗ABA, 从而促进种子萌发(Jacobsen et al., 2013).JAZ3 (JASMONATE-ZIM DOMAIN PROTEIN 3)通过与ABI5互作抑制ABI5的转录激活活性, 进而促进种子萌发(Ju et al., 2019).目前关于JA调控种子萌发功能上存在矛盾的原因还有待深入探究.SLs通过降低ABA/GA值促进种子萌发(Toh et al., 2012).此外, SLs信号通路组分也能参与调控种子萌发, 如SMAX1 (SUPPRESSOR OF MORE AXILLARY GROWTH2 1) (Stanga et al., 2013).但目前SLs调控种子萌发的具体机制还不清楚.综上, 生长素、ET、BR、CTKs、JA和SLs通过调控ABA生物合成或信号通路调控种子的休眠和萌发.然而生长素、ET、BR、CTKs、JA与GA之间的交互作用还有待深入研究. ...
Molecular mechanisms involved in convergent crop domestication 1 2013
... 休眠是种子植物在长期进化过程中产生的适应性性状, 植物通过抑制种子萌发保证其在不适宜的环境中生存, 因此休眠对于种子能够“适时萌发”具有重要意义.在实际生产中, 为了保证种子的高萌发率, 大部分作物的种子往往呈现出较低的休眠水平(Lenser and Theißen, 2013).然而过低的休眠水平导致成熟的种子在母体上直接萌发(“胎萌”)或出现穗发芽现象, 致使农业生产中种子的产量和质量下降(Simsek et al., 2014; Shu et al., 2015; Liu et al., 2019).因此, 研究种子休眠和萌发的调控机制对农业生产具有重要指导意义. ...
Phytochrome signaling mechanisms 1 2011
... PHY在黑暗条件下以生理失活的红光吸收型(Pr)存在, 吸收红光之后转变成其生理激活型(Pfr).两种光吸收型的PHY在Pr和Pfr两种状态间相互转变.早在20世纪50年代, 研究人员以莴苣(Lactuca sativa)种子为材料, 研究红光和远红光对其萌发的影响.结果表明红光促进种子萌发, 而远红光能够逆转红光的作用(Borthwick et al., 1952).红光和远红光对种子萌发的可逆调控暗示着PHY参与调控种子的萌发过程(Shinomura et al., 1994; Hennig et al., 2002).拟南芥PHY基因家族包含5个成员——PHYA-PHYE (Sharrock and Quail, 1989; Clack et al., 1994).种子萌发受到PHYA和PHYB的调控.其中, PHYB发挥主要功能.研究表明, PHYA蛋白在干种子中表达量很低, 随着种子在黑暗条件下吸胀时间的延长其蛋白表达量逐渐增加, 说明PHYA参与调控种子的萌发过程(Shinomura et al., 1996).PHYA主要在种子吸胀后期通过介导红光和远红光条件下的极低辐照度反应(VLFR)和远红光下的高辐照度反应(FR-HIR)调控种子萌发.相比之下, PHYB在干种子和吸胀种子中表达量均很高, 能够在种子吸胀初期(几个小时以内)介导红光和远红光下的低辐照度反应(LFR)调控种子萌发(Seo et al., 2009; Li et al., 2011).除了PHYA和PHYB外, PHYE也参与光调控的种子萌发过程, 而且三者在调控种子萌发方面功能冗余(Hennig et al., 2002).最近有研究表明, PHYB除了调控种子萌发, 还参与调控种子休眠(Jiang et al., 2016). ...
ETR1/RDO3 regulates seed dormancy by relieving the inhibitory effect of the ERF12-TPL complex on DELAY OF GERMINATION 1 expression 1 2019
... 除ABA和GA之外, 种子的休眠和萌发还受到7类激素的调控, 包括生长素(auxin)、乙烯(ethylene, ET)、油菜素内酯(brassinosteroids, BRs)、水杨酸(salicylic acid, SA)、茉莉酸(jasmonic acid, JA)、细胞分裂素(cytokinins, CTKs)和独脚金内酯(strigolactones, SLs).这些激素均通过影响ABA或GA通路间接调控种子的休眠和萌发(Shu et al., 2016; Née et al., 2017a).生长素能够通过调控ABI3的转录水平影响种子的休眠和萌发.当内源生长素水平升高时, 生长素信号通路中的负调控因子AXR2/3 (AUXIN- RESISTANT 2/3)发生降解.此时AXR2/3对响应生长素信号的ARF10 (AUXIN RESPONSE FACTOR 10)和ARF16基因的转录抑制作用被解除, 而ARF10和ARF16能够激活ABI3的表达, 从而使种子保持休眠状态(Liu et al., 2013a).然而, ARF10和ARF16不能直接结合在ABI3的启动子上(Liu et al., 2013a), 因此有待深入研究以揭示完整的生长素信号传递链.ET能够通过ABA生物合成和信号转导通路调控种子萌发(Cheng et al., 2009; Linkies et al., 2009; Corbineau et al., 2014), 但具体机制还不清楚.在盐胁迫条件下, ET受体蛋白ETR1 (ETHYLENE RESPONSE 1)和ETR2可能通过不依赖ET信号的通路调控ABA生物合成, 进而影响种子萌发(Wilson et al., 2014).最近的研究发现, ETR1通过解除ERF12 (ETHYLENE RESPONSE FACTOR 12)/TPL (TOPLESS)模块对DOG1基因的转录抑制作用而促进DOG1的转录, 进而促进种子休眠(Li et al., 2019).BR通过依赖MFT (MOTHER OFFT AND TFL 1)的途径拮抗ABA信号转导通路, 促进种子萌发(Xi and Yu, 2010; Xi et al., 2010).进一步的研究发现, BR信号通路中的负调控因子——BIN2 (BRASSINOSTEROID INSENSITIVE 2), 能够磷酸化并稳定ABI5蛋白, 进而促进ABA信号转导.BR信号通过抑制BIN2-ABI5互作解除ABA对种子萌发的抑制作用(Hu and Yu, 2014).SA在调控种子萌发上发挥双重作用.在正常生长条件下, SA通过抑制GA诱导的α淀粉酶编码基因的表达抑制种子萌发(Xie et al., 2007); 而在高盐胁迫下, SA通过其它通路促进种子萌发(Lee et al., 2010).CTKs通过抑制ABI5的转录或促进ABI5蛋白的降解拮抗ABA效应, 从而促进种子萌发(Wang et al., 2011; Guan et al., 2014).外源施加JA能够延迟种子萌发(Nambara et al., 2010), 表明JA能够抑制种子萌发.有研究表明, JA通过抑制ABA合成相关基因的表达、促进ABA代谢相关基因的表达拮抗ABA, 从而促进种子萌发(Jacobsen et al., 2013).JAZ3 (JASMONATE-ZIM DOMAIN PROTEIN 3)通过与ABI5互作抑制ABI5的转录激活活性, 进而促进种子萌发(Ju et al., 2019).目前关于JA调控种子萌发功能上存在矛盾的原因还有待深入探究.SLs通过降低ABA/GA值促进种子萌发(Toh et al., 2012).此外, SLs信号通路组分也能参与调控种子萌发, 如SMAX1 (SUPPRESSOR OF MORE AXILLARY GROWTH2 1) (Stanga et al., 2013).但目前SLs调控种子萌发的具体机制还不清楚.综上, 生长素、ET、BR、CTKs、JA和SLs通过调控ABA生物合成或信号通路调控种子的休眠和萌发.然而生长素、ET、BR、CTKs、JA与GA之间的交互作用还有待深入研究. ...
UVR8 interacts with BES1 and BIM1 to regulate transcription and photomorphogenesis in Arabidopsis 1 2018
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
Cryptochrome structure and signal transduction 1 2003
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
Ethylene interacts with abscisic acid to regulate endosperm rupture during germination: a comparative approach using Lepidium sativum and Arabidopsis thaliana 1 2009
... 除ABA和GA之外, 种子的休眠和萌发还受到7类激素的调控, 包括生长素(auxin)、乙烯(ethylene, ET)、油菜素内酯(brassinosteroids, BRs)、水杨酸(salicylic acid, SA)、茉莉酸(jasmonic acid, JA)、细胞分裂素(cytokinins, CTKs)和独脚金内酯(strigolactones, SLs).这些激素均通过影响ABA或GA通路间接调控种子的休眠和萌发(Shu et al., 2016; Née et al., 2017a).生长素能够通过调控ABI3的转录水平影响种子的休眠和萌发.当内源生长素水平升高时, 生长素信号通路中的负调控因子AXR2/3 (AUXIN- RESISTANT 2/3)发生降解.此时AXR2/3对响应生长素信号的ARF10 (AUXIN RESPONSE FACTOR 10)和ARF16基因的转录抑制作用被解除, 而ARF10和ARF16能够激活ABI3的表达, 从而使种子保持休眠状态(Liu et al., 2013a).然而, ARF10和ARF16不能直接结合在ABI3的启动子上(Liu et al., 2013a), 因此有待深入研究以揭示完整的生长素信号传递链.ET能够通过ABA生物合成和信号转导通路调控种子萌发(Cheng et al., 2009; Linkies et al., 2009; Corbineau et al., 2014), 但具体机制还不清楚.在盐胁迫条件下, ET受体蛋白ETR1 (ETHYLENE RESPONSE 1)和ETR2可能通过不依赖ET信号的通路调控ABA生物合成, 进而影响种子萌发(Wilson et al., 2014).最近的研究发现, ETR1通过解除ERF12 (ETHYLENE RESPONSE FACTOR 12)/TPL (TOPLESS)模块对DOG1基因的转录抑制作用而促进DOG1的转录, 进而促进种子休眠(Li et al., 2019).BR通过依赖MFT (MOTHER OFFT AND TFL 1)的途径拮抗ABA信号转导通路, 促进种子萌发(Xi and Yu, 2010; Xi et al., 2010).进一步的研究发现, BR信号通路中的负调控因子——BIN2 (BRASSINOSTEROID INSENSITIVE 2), 能够磷酸化并稳定ABI5蛋白, 进而促进ABA信号转导.BR信号通过抑制BIN2-ABI5互作解除ABA对种子萌发的抑制作用(Hu and Yu, 2014).SA在调控种子萌发上发挥双重作用.在正常生长条件下, SA通过抑制GA诱导的α淀粉酶编码基因的表达抑制种子萌发(Xie et al., 2007); 而在高盐胁迫下, SA通过其它通路促进种子萌发(Lee et al., 2010).CTKs通过抑制ABI5的转录或促进ABI5蛋白的降解拮抗ABA效应, 从而促进种子萌发(Wang et al., 2011; Guan et al., 2014).外源施加JA能够延迟种子萌发(Nambara et al., 2010), 表明JA能够抑制种子萌发.有研究表明, JA通过抑制ABA合成相关基因的表达、促进ABA代谢相关基因的表达拮抗ABA, 从而促进种子萌发(Jacobsen et al., 2013).JAZ3 (JASMONATE-ZIM DOMAIN PROTEIN 3)通过与ABI5互作抑制ABI5的转录激活活性, 进而促进种子萌发(Ju et al., 2019).目前关于JA调控种子萌发功能上存在矛盾的原因还有待深入探究.SLs通过降低ABA/GA值促进种子萌发(Toh et al., 2012).此外, SLs信号通路组分也能参与调控种子萌发, 如SMAX1 (SUPPRESSOR OF MORE AXILLARY GROWTH2 1) (Stanga et al., 2013).但目前SLs调控种子萌发的具体机制还不清楚.综上, 生长素、ET、BR、CTKs、JA和SLs通过调控ABA生物合成或信号通路调控种子的休眠和萌发.然而生长素、ET、BR、CTKs、JA与GA之间的交互作用还有待深入研究. ...
Photoexcited CRY2 interacts with CIB1 to regulate transcription and floral initiation in Arabidopsis 1 2008a
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
COP1-mediated ubiquitination of CONSTANS is implicated in cryptochrome regulation of flowering in Arabidopsis 1 2008b
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
The NF-YC-RGL2 module integrates GA and ABA signaling to regulate seed germination in Arabidopsis 1 2016
... GA能够通过抑制ABA诱导的种子休眠促进种子萌发(Gubler et al., 2005; Graeber et al., 2012).在植物体内, 具有生物活性的GA主要包括GA1和GA4, 它们通过软化种皮、促进胚乳层细胞的水解和胚的生长打破种子休眠, 进而促进种子萌发(Holdsworth et al., 2008).GA缺失突变体ga1 (gibberellic acid- requiring 1)和ga2表现出强烈的休眠表型, 需要外源施加GA方可萌发(Lee et al., 2002; Shu et al., 2013).GA20ox (GA 20-OXIDASE)和GA3ox基因编码参与GA生物合成的氧化酶; GA2ox编码的GA2-氧化酶主要参与GA的代谢过程.在PHYB-off条件下, ga2ox突变体萌发水平升高(Oh et al., 2006; Yamauchi et al., 2007).上述研究结果表明, GA能通过其生物合成通路调控种子的休眠和萌发.在GA信号转导过程中, DELLA蛋白扮演着重要角色, 它能响应GA信号快速降解, 对GA信号转导起限速作用(Silverstone et al., 1998; Itoh et al., 2002; Zentella et al., 2007; Nemoto et al., 2017).GA受体蛋白GID1 (GA-INSENSITIVE DWARF 1)能够负调控DELLA蛋白的稳定性, 促进GA信号转导(Davière and Achard, 2013).拟南芥SLY1 (SLEEPY1)和水稻GID2 (GA-INSENSITIVE DWARF2)是SCF聚合体中的F-box亚基, 依赖其C端的GGF和LSL基序与DELLA蛋白C端VHIID和LHR2基序结合, 促进DELLA多聚泛素化(Hirano et al., 2010; Ariizumi et al., 2011).研究表明, DELLA蛋白GAI (GA-INSENSITIVE)能够通过抑制TCP14 (TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR 14)和TCP15的表达减缓细胞周期的进程, 最终使种子保持休眠状态(Resentini et al., 2015).作为转录调节子, DELLA如何实现对下游基因的转录调控? 有研究指出, DELLA能够通过与染色质重塑因子互作调控下游基因的转录.DELLA蛋白RGL2 (RGA-LIKE 2)和RGL3与染色质重塑因子SWI3C (SWITCH3C)发生互作, 进而影响GID1a以及GA3ox基因的转录(Sarnowska et al., 2013).此外, DELLA还能通过与种子萌发相关转录因子互作进而实现其对萌发的调控.RGL2通过与转录因子NF-YC互作, 诱导下游ABI5的转录, 进而抑制种子萌发(Liu et al., 2016).此外, RGL2还能通过与转录因子DOF6 (BINDING1 ZINC FINGER 6)互作诱导GATA12的表达, 进而促进种子休眠(Ravindran et al., 2017).DELLA通过与ICE1 (INDUCER OF CBF EXPRESSION 1)互作解除其对ABA响应基因EM1 (LATE EMBRYOGENESIS ABUNDANT 1)和EM6的转录抑制作用, 最终抑制种子萌发(Hu et al., 2019). ...
Identification and characterization of the rice pre-harvest sprouting mutants involved in molybdenum cofactor biosynthesis 1 2019
... 休眠是种子植物在长期进化过程中产生的适应性性状, 植物通过抑制种子萌发保证其在不适宜的环境中生存, 因此休眠对于种子能够“适时萌发”具有重要意义.在实际生产中, 为了保证种子的高萌发率, 大部分作物的种子往往呈现出较低的休眠水平(Lenser and Theißen, 2013).然而过低的休眠水平导致成熟的种子在母体上直接萌发(“胎萌”)或出现穗发芽现象, 致使农业生产中种子的产量和质量下降(Simsek et al., 2014; Shu et al., 2015; Liu et al., 2019).因此, 研究种子休眠和萌发的调控机制对农业生产具有重要指导意义. ...
Auxin controls seed dormancy through stimulation of abscisic acid signaling by inducing ARF-mediated ABI3 activation in Arabidopsis 2 2013a
... 除ABA和GA之外, 种子的休眠和萌发还受到7类激素的调控, 包括生长素(auxin)、乙烯(ethylene, ET)、油菜素内酯(brassinosteroids, BRs)、水杨酸(salicylic acid, SA)、茉莉酸(jasmonic acid, JA)、细胞分裂素(cytokinins, CTKs)和独脚金内酯(strigolactones, SLs).这些激素均通过影响ABA或GA通路间接调控种子的休眠和萌发(Shu et al., 2016; Née et al., 2017a).生长素能够通过调控ABI3的转录水平影响种子的休眠和萌发.当内源生长素水平升高时, 生长素信号通路中的负调控因子AXR2/3 (AUXIN- RESISTANT 2/3)发生降解.此时AXR2/3对响应生长素信号的ARF10 (AUXIN RESPONSE FACTOR 10)和ARF16基因的转录抑制作用被解除, 而ARF10和ARF16能够激活ABI3的表达, 从而使种子保持休眠状态(Liu et al., 2013a).然而, ARF10和ARF16不能直接结合在ABI3的启动子上(Liu et al., 2013a), 因此有待深入研究以揭示完整的生长素信号传递链.ET能够通过ABA生物合成和信号转导通路调控种子萌发(Cheng et al., 2009; Linkies et al., 2009; Corbineau et al., 2014), 但具体机制还不清楚.在盐胁迫条件下, ET受体蛋白ETR1 (ETHYLENE RESPONSE 1)和ETR2可能通过不依赖ET信号的通路调控ABA生物合成, 进而影响种子萌发(Wilson et al., 2014).最近的研究发现, ETR1通过解除ERF12 (ETHYLENE RESPONSE FACTOR 12)/TPL (TOPLESS)模块对DOG1基因的转录抑制作用而促进DOG1的转录, 进而促进种子休眠(Li et al., 2019).BR通过依赖MFT (MOTHER OFFT AND TFL 1)的途径拮抗ABA信号转导通路, 促进种子萌发(Xi and Yu, 2010; Xi et al., 2010).进一步的研究发现, BR信号通路中的负调控因子——BIN2 (BRASSINOSTEROID INSENSITIVE 2), 能够磷酸化并稳定ABI5蛋白, 进而促进ABA信号转导.BR信号通过抑制BIN2-ABI5互作解除ABA对种子萌发的抑制作用(Hu and Yu, 2014).SA在调控种子萌发上发挥双重作用.在正常生长条件下, SA通过抑制GA诱导的α淀粉酶编码基因的表达抑制种子萌发(Xie et al., 2007); 而在高盐胁迫下, SA通过其它通路促进种子萌发(Lee et al., 2010).CTKs通过抑制ABI5的转录或促进ABI5蛋白的降解拮抗ABA效应, 从而促进种子萌发(Wang et al., 2011; Guan et al., 2014).外源施加JA能够延迟种子萌发(Nambara et al., 2010), 表明JA能够抑制种子萌发.有研究表明, JA通过抑制ABA合成相关基因的表达、促进ABA代谢相关基因的表达拮抗ABA, 从而促进种子萌发(Jacobsen et al., 2013).JAZ3 (JASMONATE-ZIM DOMAIN PROTEIN 3)通过与ABI5互作抑制ABI5的转录激活活性, 进而促进种子萌发(Ju et al., 2019).目前关于JA调控种子萌发功能上存在矛盾的原因还有待深入探究.SLs通过降低ABA/GA值促进种子萌发(Toh et al., 2012).此外, SLs信号通路组分也能参与调控种子萌发, 如SMAX1 (SUPPRESSOR OF MORE AXILLARY GROWTH2 1) (Stanga et al., 2013).但目前SLs调控种子萌发的具体机制还不清楚.综上, 生长素、ET、BR、CTKs、JA和SLs通过调控ABA生物合成或信号通路调控种子的休眠和萌发.然而生长素、ET、BR、CTKs、JA与GA之间的交互作用还有待深入研究. ...
... 的启动子上(Liu et al., 2013a), 因此有待深入研究以揭示完整的生长素信号传递链.ET能够通过ABA生物合成和信号转导通路调控种子萌发(Cheng et al., 2009; Linkies et al., 2009; Corbineau et al., 2014), 但具体机制还不清楚.在盐胁迫条件下, ET受体蛋白ETR1 (ETHYLENE RESPONSE 1)和ETR2可能通过不依赖ET信号的通路调控ABA生物合成, 进而影响种子萌发(Wilson et al., 2014).最近的研究发现, ETR1通过解除ERF12 (ETHYLENE RESPONSE FACTOR 12)/TPL (TOPLESS)模块对DOG1基因的转录抑制作用而促进DOG1的转录, 进而促进种子休眠(Li et al., 2019).BR通过依赖MFT (MOTHER OFFT AND TFL 1)的途径拮抗ABA信号转导通路, 促进种子萌发(Xi and Yu, 2010; Xi et al., 2010).进一步的研究发现, BR信号通路中的负调控因子——BIN2 (BRASSINOSTEROID INSENSITIVE 2), 能够磷酸化并稳定ABI5蛋白, 进而促进ABA信号转导.BR信号通过抑制BIN2-ABI5互作解除ABA对种子萌发的抑制作用(Hu and Yu, 2014).SA在调控种子萌发上发挥双重作用.在正常生长条件下, SA通过抑制GA诱导的α淀粉酶编码基因的表达抑制种子萌发(Xie et al., 2007); 而在高盐胁迫下, SA通过其它通路促进种子萌发(Lee et al., 2010).CTKs通过抑制ABI5的转录或促进ABI5蛋白的降解拮抗ABA效应, 从而促进种子萌发(Wang et al., 2011; Guan et al., 2014).外源施加JA能够延迟种子萌发(Nambara et al., 2010), 表明JA能够抑制种子萌发.有研究表明, JA通过抑制ABA合成相关基因的表达、促进ABA代谢相关基因的表达拮抗ABA, 从而促进种子萌发(Jacobsen et al., 2013).JAZ3 (JASMONATE-ZIM DOMAIN PROTEIN 3)通过与ABI5互作抑制ABI5的转录激活活性, 进而促进种子萌发(Ju et al., 2019).目前关于JA调控种子萌发功能上存在矛盾的原因还有待深入探究.SLs通过降低ABA/GA值促进种子萌发(Toh et al., 2012).此外, SLs信号通路组分也能参与调控种子萌发, 如SMAX1 (SUPPRESSOR OF MORE AXILLARY GROWTH2 1) (Stanga et al., 2013).但目前SLs调控种子萌发的具体机制还不清楚.综上, 生长素、ET、BR、CTKs、JA和SLs通过调控ABA生物合成或信号通路调控种子的休眠和萌发.然而生长素、ET、BR、CTKs、JA与GA之间的交互作用还有待深入研究. ...
Multiple bHLH proteins form heterodimers to mediate CRY2-dependent regulation of flowering-time in Arabidopsis 1 2013b
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
COP1 and phyB physically interact with PIL1 to regulate its stability and photomorphogenic development in Arabidopsis 1 2014
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
Cryptochrome 1 interacts with PIF4 to regulate high temperature-mediated hypocotyl elongation in response to blue light 1 2016
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
Regulators of PP2C phosphatase activity function as abscisic acid sensors 1 2009
... 激素在种子休眠和萌发过程中发挥重要调控作用.其中, ABA和GA在调控种子休眠和萌发上行使主要功能.ABA促进种子休眠, 而GA促进种子萌发(Shu et al., 2016; Née et al., 2017a).在种子成熟过程中, ABA在种子内逐渐积累, 使种子的休眠水平逐渐升高.相比之下, 当种子受到吸胀或层积处理时, ABA含量逐渐降低, GA含量逐渐升高, 使种子萌发.与野生型种子相比, ABA合成缺陷突变体种子萌发更快(Frey et al., 2012); 而过表达ABA合成基因的种子以及ABA代谢突变体种子保持较高的休眠水平(Matakiadis et al., 2009; Martinez-Andújar et al., 2011; Nonogaki et al., 2014), 表明ABA能够通过其生物合成通路调控种子的休眠和萌发.ABA信号通过PYR/PYL/RCAR (PYRABACTIN RESISTANCE 1/PYR-LIKE PROTEINS/REGULATORY COMPONENTS OF ABA RECEPTORS)-PP2C (PROTEIN PHOSPHATASE 2C)-SnRKs (SNF1-RELATED PROTEIN KINASES)级联反应向下传递(Cutler et al., 2010; Hubbard et al., 2010).目前鉴定到的PP2C类蛋白包括ABI1 (ABSCISIC ACID INSENSITIVE 1)、ABI2、HON (HONSU)和RDO5 (REDUCED DORMANCY 5).其中, ABI1和ABI2通过与ABA信号受体蛋白PYR/PYL/RCAR互作进而抑制ABA信号转导(Ma et al., 2009; Park et al., 2009).HON蛋白能够通过抑制ABA信号转导及促进GA信号转导抑制种子休眠(Kim et al., 2013), 说明HON能够整合ABA和GA信号以调控种子休眠.rdo5突变体的休眠水平降低, 但其ABA含量和ABA敏感性并未发生显著变化(Xiang et al., 2014), 说明RDO5通过不依赖ABA的途径调控种子休眠.ABI3、ABI4和ABI5是ABA信号通路下游的关键组分, 能够抑制种子萌发、促进种子休眠(Bentsink and Koornneef, 2008; Kanai et al., 2010; Shu et al., 2013).响应ABA信号的R2R3型MYB转录因子MYB96通过调控ABI4以及ABA合成相关基因NCED2 (9-CIS-EPOXYCAROTENOID DIOXYGENASE 2)和NCED6的转录进而促进种子休眠、抑制种子萌发(Lee et al., 2015a, 2015b).上述研究表明, ABA不仅通过其生物合成通路调控种子的休眠和萌发, 还能以ABA信号的形式发挥作用. ...
Induction of 9- cis-epoxycarotenoid dioxygenase in Arabidopsis thaliana seeds enhances seed dormancy 1 2011
... 激素在种子休眠和萌发过程中发挥重要调控作用.其中, ABA和GA在调控种子休眠和萌发上行使主要功能.ABA促进种子休眠, 而GA促进种子萌发(Shu et al., 2016; Née et al., 2017a).在种子成熟过程中, ABA在种子内逐渐积累, 使种子的休眠水平逐渐升高.相比之下, 当种子受到吸胀或层积处理时, ABA含量逐渐降低, GA含量逐渐升高, 使种子萌发.与野生型种子相比, ABA合成缺陷突变体种子萌发更快(Frey et al., 2012); 而过表达ABA合成基因的种子以及ABA代谢突变体种子保持较高的休眠水平(Matakiadis et al., 2009; Martinez-Andújar et al., 2011; Nonogaki et al., 2014), 表明ABA能够通过其生物合成通路调控种子的休眠和萌发.ABA信号通过PYR/PYL/RCAR (PYRABACTIN RESISTANCE 1/PYR-LIKE PROTEINS/REGULATORY COMPONENTS OF ABA RECEPTORS)-PP2C (PROTEIN PHOSPHATASE 2C)-SnRKs (SNF1-RELATED PROTEIN KINASES)级联反应向下传递(Cutler et al., 2010; Hubbard et al., 2010).目前鉴定到的PP2C类蛋白包括ABI1 (ABSCISIC ACID INSENSITIVE 1)、ABI2、HON (HONSU)和RDO5 (REDUCED DORMANCY 5).其中, ABI1和ABI2通过与ABA信号受体蛋白PYR/PYL/RCAR互作进而抑制ABA信号转导(Ma et al., 2009; Park et al., 2009).HON蛋白能够通过抑制ABA信号转导及促进GA信号转导抑制种子休眠(Kim et al., 2013), 说明HON能够整合ABA和GA信号以调控种子休眠.rdo5突变体的休眠水平降低, 但其ABA含量和ABA敏感性并未发生显著变化(Xiang et al., 2014), 说明RDO5通过不依赖ABA的途径调控种子休眠.ABI3、ABI4和ABI5是ABA信号通路下游的关键组分, 能够抑制种子萌发、促进种子休眠(Bentsink and Koornneef, 2008; Kanai et al., 2010; Shu et al., 2013).响应ABA信号的R2R3型MYB转录因子MYB96通过调控ABI4以及ABA合成相关基因NCED2 (9-CIS-EPOXYCAROTENOID DIOXYGENASE 2)和NCED6的转录进而促进种子休眠、抑制种子萌发(Lee et al., 2015a, 2015b).上述研究表明, ABA不仅通过其生物合成通路调控种子的休眠和萌发, 还能以ABA信号的形式发挥作用. ...
The Arabidopsis abscisic acid catabolic gene CYP707A2 plays a key role in nitrate control of seed dormancy 1 2009
... 激素在种子休眠和萌发过程中发挥重要调控作用.其中, ABA和GA在调控种子休眠和萌发上行使主要功能.ABA促进种子休眠, 而GA促进种子萌发(Shu et al., 2016; Née et al., 2017a).在种子成熟过程中, ABA在种子内逐渐积累, 使种子的休眠水平逐渐升高.相比之下, 当种子受到吸胀或层积处理时, ABA含量逐渐降低, GA含量逐渐升高, 使种子萌发.与野生型种子相比, ABA合成缺陷突变体种子萌发更快(Frey et al., 2012); 而过表达ABA合成基因的种子以及ABA代谢突变体种子保持较高的休眠水平(Matakiadis et al., 2009; Martinez-Andújar et al., 2011; Nonogaki et al., 2014), 表明ABA能够通过其生物合成通路调控种子的休眠和萌发.ABA信号通过PYR/PYL/RCAR (PYRABACTIN RESISTANCE 1/PYR-LIKE PROTEINS/REGULATORY COMPONENTS OF ABA RECEPTORS)-PP2C (PROTEIN PHOSPHATASE 2C)-SnRKs (SNF1-RELATED PROTEIN KINASES)级联反应向下传递(Cutler et al., 2010; Hubbard et al., 2010).目前鉴定到的PP2C类蛋白包括ABI1 (ABSCISIC ACID INSENSITIVE 1)、ABI2、HON (HONSU)和RDO5 (REDUCED DORMANCY 5).其中, ABI1和ABI2通过与ABA信号受体蛋白PYR/PYL/RCAR互作进而抑制ABA信号转导(Ma et al., 2009; Park et al., 2009).HON蛋白能够通过抑制ABA信号转导及促进GA信号转导抑制种子休眠(Kim et al., 2013), 说明HON能够整合ABA和GA信号以调控种子休眠.rdo5突变体的休眠水平降低, 但其ABA含量和ABA敏感性并未发生显著变化(Xiang et al., 2014), 说明RDO5通过不依赖ABA的途径调控种子休眠.ABI3、ABI4和ABI5是ABA信号通路下游的关键组分, 能够抑制种子萌发、促进种子休眠(Bentsink and Koornneef, 2008; Kanai et al., 2010; Shu et al., 2013).响应ABA信号的R2R3型MYB转录因子MYB96通过调控ABI4以及ABA合成相关基因NCED2 (9-CIS-EPOXYCAROTENOID DIOXYGENASE 2)和NCED6的转录进而促进种子休眠、抑制种子萌发(Lee et al., 2015a, 2015b).上述研究表明, ABA不仅通过其生物合成通路调控种子的休眠和萌发, 还能以ABA信号的形式发挥作用. ...
Abscisic acid and the control of seed dormancy and germination 1 2010
... 除ABA和GA之外, 种子的休眠和萌发还受到7类激素的调控, 包括生长素(auxin)、乙烯(ethylene, ET)、油菜素内酯(brassinosteroids, BRs)、水杨酸(salicylic acid, SA)、茉莉酸(jasmonic acid, JA)、细胞分裂素(cytokinins, CTKs)和独脚金内酯(strigolactones, SLs).这些激素均通过影响ABA或GA通路间接调控种子的休眠和萌发(Shu et al., 2016; Née et al., 2017a).生长素能够通过调控ABI3的转录水平影响种子的休眠和萌发.当内源生长素水平升高时, 生长素信号通路中的负调控因子AXR2/3 (AUXIN- RESISTANT 2/3)发生降解.此时AXR2/3对响应生长素信号的ARF10 (AUXIN RESPONSE FACTOR 10)和ARF16基因的转录抑制作用被解除, 而ARF10和ARF16能够激活ABI3的表达, 从而使种子保持休眠状态(Liu et al., 2013a).然而, ARF10和ARF16不能直接结合在ABI3的启动子上(Liu et al., 2013a), 因此有待深入研究以揭示完整的生长素信号传递链.ET能够通过ABA生物合成和信号转导通路调控种子萌发(Cheng et al., 2009; Linkies et al., 2009; Corbineau et al., 2014), 但具体机制还不清楚.在盐胁迫条件下, ET受体蛋白ETR1 (ETHYLENE RESPONSE 1)和ETR2可能通过不依赖ET信号的通路调控ABA生物合成, 进而影响种子萌发(Wilson et al., 2014).最近的研究发现, ETR1通过解除ERF12 (ETHYLENE RESPONSE FACTOR 12)/TPL (TOPLESS)模块对DOG1基因的转录抑制作用而促进DOG1的转录, 进而促进种子休眠(Li et al., 2019).BR通过依赖MFT (MOTHER OFFT AND TFL 1)的途径拮抗ABA信号转导通路, 促进种子萌发(Xi and Yu, 2010; Xi et al., 2010).进一步的研究发现, BR信号通路中的负调控因子——BIN2 (BRASSINOSTEROID INSENSITIVE 2), 能够磷酸化并稳定ABI5蛋白, 进而促进ABA信号转导.BR信号通过抑制BIN2-ABI5互作解除ABA对种子萌发的抑制作用(Hu and Yu, 2014).SA在调控种子萌发上发挥双重作用.在正常生长条件下, SA通过抑制GA诱导的α淀粉酶编码基因的表达抑制种子萌发(Xie et al., 2007); 而在高盐胁迫下, SA通过其它通路促进种子萌发(Lee et al., 2010).CTKs通过抑制ABI5的转录或促进ABI5蛋白的降解拮抗ABA效应, 从而促进种子萌发(Wang et al., 2011; Guan et al., 2014).外源施加JA能够延迟种子萌发(Nambara et al., 2010), 表明JA能够抑制种子萌发.有研究表明, JA通过抑制ABA合成相关基因的表达、促进ABA代谢相关基因的表达拮抗ABA, 从而促进种子萌发(Jacobsen et al., 2013).JAZ3 (JASMONATE-ZIM DOMAIN PROTEIN 3)通过与ABI5互作抑制ABI5的转录激活活性, 进而促进种子萌发(Ju et al., 2019).目前关于JA调控种子萌发功能上存在矛盾的原因还有待深入探究.SLs通过降低ABA/GA值促进种子萌发(Toh et al., 2012).此外, SLs信号通路组分也能参与调控种子萌发, 如SMAX1 (SUPPRESSOR OF MORE AXILLARY GROWTH2 1) (Stanga et al., 2013).但目前SLs调控种子萌发的具体机制还不清楚.综上, 生长素、ET、BR、CTKs、JA和SLs通过调控ABA生物合成或信号通路调控种子的休眠和萌发.然而生长素、ET、BR、CTKs、JA与GA之间的交互作用还有待深入研究. ...
DELAY OF GERMINATION 1 requires PP2C phosphatases of the ABA signaling pathway to control seed dormancy 2 2017b
... 激素在种子休眠和萌发过程中发挥重要调控作用.其中, ABA和GA在调控种子休眠和萌发上行使主要功能.ABA促进种子休眠, 而GA促进种子萌发(Shu et al., 2016; Née et al., 2017a).在种子成熟过程中, ABA在种子内逐渐积累, 使种子的休眠水平逐渐升高.相比之下, 当种子受到吸胀或层积处理时, ABA含量逐渐降低, GA含量逐渐升高, 使种子萌发.与野生型种子相比, ABA合成缺陷突变体种子萌发更快(Frey et al., 2012); 而过表达ABA合成基因的种子以及ABA代谢突变体种子保持较高的休眠水平(Matakiadis et al., 2009; Martinez-Andújar et al., 2011; Nonogaki et al., 2014), 表明ABA能够通过其生物合成通路调控种子的休眠和萌发.ABA信号通过PYR/PYL/RCAR (PYRABACTIN RESISTANCE 1/PYR-LIKE PROTEINS/REGULATORY COMPONENTS OF ABA RECEPTORS)-PP2C (PROTEIN PHOSPHATASE 2C)-SnRKs (SNF1-RELATED PROTEIN KINASES)级联反应向下传递(Cutler et al., 2010; Hubbard et al., 2010).目前鉴定到的PP2C类蛋白包括ABI1 (ABSCISIC ACID INSENSITIVE 1)、ABI2、HON (HONSU)和RDO5 (REDUCED DORMANCY 5).其中, ABI1和ABI2通过与ABA信号受体蛋白PYR/PYL/RCAR互作进而抑制ABA信号转导(Ma et al., 2009; Park et al., 2009).HON蛋白能够通过抑制ABA信号转导及促进GA信号转导抑制种子休眠(Kim et al., 2013), 说明HON能够整合ABA和GA信号以调控种子休眠.rdo5突变体的休眠水平降低, 但其ABA含量和ABA敏感性并未发生显著变化(Xiang et al., 2014), 说明RDO5通过不依赖ABA的途径调控种子休眠.ABI3、ABI4和ABI5是ABA信号通路下游的关键组分, 能够抑制种子萌发、促进种子休眠(Bentsink and Koornneef, 2008; Kanai et al., 2010; Shu et al., 2013).响应ABA信号的R2R3型MYB转录因子MYB96通过调控ABI4以及ABA合成相关基因NCED2 (9-CIS-EPOXYCAROTENOID DIOXYGENASE 2)和NCED6的转录进而促进种子休眠、抑制种子萌发(Lee et al., 2015a, 2015b).上述研究表明, ABA不仅通过其生物合成通路调控种子的休眠和萌发, 还能以ABA信号的形式发挥作用. ...
... 除ABA和GA之外, 种子的休眠和萌发还受到7类激素的调控, 包括生长素(auxin)、乙烯(ethylene, ET)、油菜素内酯(brassinosteroids, BRs)、水杨酸(salicylic acid, SA)、茉莉酸(jasmonic acid, JA)、细胞分裂素(cytokinins, CTKs)和独脚金内酯(strigolactones, SLs).这些激素均通过影响ABA或GA通路间接调控种子的休眠和萌发(Shu et al., 2016; Née et al., 2017a).生长素能够通过调控ABI3的转录水平影响种子的休眠和萌发.当内源生长素水平升高时, 生长素信号通路中的负调控因子AXR2/3 (AUXIN- RESISTANT 2/3)发生降解.此时AXR2/3对响应生长素信号的ARF10 (AUXIN RESPONSE FACTOR 10)和ARF16基因的转录抑制作用被解除, 而ARF10和ARF16能够激活ABI3的表达, 从而使种子保持休眠状态(Liu et al., 2013a).然而, ARF10和ARF16不能直接结合在ABI3的启动子上(Liu et al., 2013a), 因此有待深入研究以揭示完整的生长素信号传递链.ET能够通过ABA生物合成和信号转导通路调控种子萌发(Cheng et al., 2009; Linkies et al., 2009; Corbineau et al., 2014), 但具体机制还不清楚.在盐胁迫条件下, ET受体蛋白ETR1 (ETHYLENE RESPONSE 1)和ETR2可能通过不依赖ET信号的通路调控ABA生物合成, 进而影响种子萌发(Wilson et al., 2014).最近的研究发现, ETR1通过解除ERF12 (ETHYLENE RESPONSE FACTOR 12)/TPL (TOPLESS)模块对DOG1基因的转录抑制作用而促进DOG1的转录, 进而促进种子休眠(Li et al., 2019).BR通过依赖MFT (MOTHER OFFT AND TFL 1)的途径拮抗ABA信号转导通路, 促进种子萌发(Xi and Yu, 2010; Xi et al., 2010).进一步的研究发现, BR信号通路中的负调控因子——BIN2 (BRASSINOSTEROID INSENSITIVE 2), 能够磷酸化并稳定ABI5蛋白, 进而促进ABA信号转导.BR信号通过抑制BIN2-ABI5互作解除ABA对种子萌发的抑制作用(Hu and Yu, 2014).SA在调控种子萌发上发挥双重作用.在正常生长条件下, SA通过抑制GA诱导的α淀粉酶编码基因的表达抑制种子萌发(Xie et al., 2007); 而在高盐胁迫下, SA通过其它通路促进种子萌发(Lee et al., 2010).CTKs通过抑制ABI5的转录或促进ABI5蛋白的降解拮抗ABA效应, 从而促进种子萌发(Wang et al., 2011; Guan et al., 2014).外源施加JA能够延迟种子萌发(Nambara et al., 2010), 表明JA能够抑制种子萌发.有研究表明, JA通过抑制ABA合成相关基因的表达、促进ABA代谢相关基因的表达拮抗ABA, 从而促进种子萌发(Jacobsen et al., 2013).JAZ3 (JASMONATE-ZIM DOMAIN PROTEIN 3)通过与ABI5互作抑制ABI5的转录激活活性, 进而促进种子萌发(Ju et al., 2019).目前关于JA调控种子萌发功能上存在矛盾的原因还有待深入探究.SLs通过降低ABA/GA值促进种子萌发(Toh et al., 2012).此外, SLs信号通路组分也能参与调控种子萌发, 如SMAX1 (SUPPRESSOR OF MORE AXILLARY GROWTH2 1) (Stanga et al., 2013).但目前SLs调控种子萌发的具体机制还不清楚.综上, 生长素、ET、BR、CTKs、JA和SLs通过调控ABA生物合成或信号通路调控种子的休眠和萌发.然而生长素、ET、BR、CTKs、JA与GA之间的交互作用还有待深入研究. ...
The release of dormancy, a wake-up call for seeds to germinate 1 2017a
... 与野生种相比, 经过驯化的栽培种种子休眠水平下调, 如水稻(Oryza sativa)、小麦(Triticum aestivum)、大麦(Hordeum vulgare)和玉米(Zea mays)等禾本科作物(Bewley and Black, 1994).近年来, 育种学家们利用遗传学或组学的方法鉴定到一些与种子休眠相关的位点、转录本或蛋白(Alonso-Blanco et al., 2003; Argyris et al., 2005; Gu et al., 2006; Née et al., 2017b).例如, 研究者利用QTL方法鉴定到调控种子休眠的关键基因——DOG1 (Alonso-Blanco et al., 2003; Bentsink et al., 2006).后续研究发现, DOG1能够参与调控ABA信号通路以及GA的生物合成, 进而调控种子休眠(Graeber et al., 2014; Nishimura et al., 2018).那么, DOG1能否参与调控GA信号通路进而促进种子休眠? DOG1是否受到外源光信号和内源激素的调控? 针对上述问题的研究不仅有助于人们更加清晰地理解种子如何监测环境因子的变化, 从而决定是否完成从休眠到萌发的发育转变; 还为利用光信号或激素人工调控种子的休眠和萌发, 解决农业生产中“胎萌”以及穗发芽等问题提供了理论支撑. ...
Tyrosine phosphorylation of the GARU E3 ubiquitin ligase promotes gibberellin signaling by preventing GID1 degradation 1 2017
... GA能够通过抑制ABA诱导的种子休眠促进种子萌发(Gubler et al., 2005; Graeber et al., 2012).在植物体内, 具有生物活性的GA主要包括GA1和GA4, 它们通过软化种皮、促进胚乳层细胞的水解和胚的生长打破种子休眠, 进而促进种子萌发(Holdsworth et al., 2008).GA缺失突变体ga1 (gibberellic acid- requiring 1)和ga2表现出强烈的休眠表型, 需要外源施加GA方可萌发(Lee et al., 2002; Shu et al., 2013).GA20ox (GA 20-OXIDASE)和GA3ox基因编码参与GA生物合成的氧化酶; GA2ox编码的GA2-氧化酶主要参与GA的代谢过程.在PHYB-off条件下, ga2ox突变体萌发水平升高(Oh et al., 2006; Yamauchi et al., 2007).上述研究结果表明, GA能通过其生物合成通路调控种子的休眠和萌发.在GA信号转导过程中, DELLA蛋白扮演着重要角色, 它能响应GA信号快速降解, 对GA信号转导起限速作用(Silverstone et al., 1998; Itoh et al., 2002; Zentella et al., 2007; Nemoto et al., 2017).GA受体蛋白GID1 (GA-INSENSITIVE DWARF 1)能够负调控DELLA蛋白的稳定性, 促进GA信号转导(Davière and Achard, 2013).拟南芥SLY1 (SLEEPY1)和水稻GID2 (GA-INSENSITIVE DWARF2)是SCF聚合体中的F-box亚基, 依赖其C端的GGF和LSL基序与DELLA蛋白C端VHIID和LHR2基序结合, 促进DELLA多聚泛素化(Hirano et al., 2010; Ariizumi et al., 2011).研究表明, DELLA蛋白GAI (GA-INSENSITIVE)能够通过抑制TCP14 (TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR 14)和TCP15的表达减缓细胞周期的进程, 最终使种子保持休眠状态(Resentini et al., 2015).作为转录调节子, DELLA如何实现对下游基因的转录调控? 有研究指出, DELLA能够通过与染色质重塑因子互作调控下游基因的转录.DELLA蛋白RGL2 (RGA-LIKE 2)和RGL3与染色质重塑因子SWI3C (SWITCH3C)发生互作, 进而影响GID1a以及GA3ox基因的转录(Sarnowska et al., 2013).此外, DELLA还能通过与种子萌发相关转录因子互作进而实现其对萌发的调控.RGL2通过与转录因子NF-YC互作, 诱导下游ABI5的转录, 进而抑制种子萌发(Liu et al., 2016).此外, RGL2还能通过与转录因子DOF6 (BINDING1 ZINC FINGER 6)互作诱导GATA12的表达, 进而促进种子休眠(Ravindran et al., 2017).DELLA通过与ICE1 (INDUCER OF CBF EXPRESSION 1)互作解除其对ABA响应基因EM1 (LATE EMBRYOGENESIS ABUNDANT 1)和EM6的转录抑制作用, 最终抑制种子萌发(Hu et al., 2019). ...
Control of seed dormancy and germination by DOG1-AHG1 PP2C phosphatase complex via binding to heme 2 2018
... 种子休眠是指有活力的种子在适宜的条件下暂时不萌发的现象(Bewley, 1997; Finch-Savage and Leubner-Metzger, 2006).种子的休眠包括休眠的诱导、维持和释放3个阶段(Shu et al., 2016).成熟的种子释放休眠后, 在适宜的条件下即可萌发.DOG1 (DELAY OF GERMINATION 1)在种子休眠诱导和维持中发挥重要作用.它既能通过抑制脱落酸(abscisic acid, ABA)通路的负调控因子——AHG1(ABA HYPERSENSITIVE GERMINATION 1)的磷酸酶活性促进种子休眠(Nishimura et al., 2018); 也能通过调控响应赤霉素(gibberellin, GA)信号的细胞壁重塑相关基因的表达促进种子休眠(Graeber et al., 2014).在种子吸胀初期, 胚乳中PHYB (PHYTOCHROME B)吸收远红光后以生理失活型存在, 此时PIF1 (PIF3-LIKE 5, PIL5)蛋白逐渐积累, 使胚乳中ABA水平逐渐升高.胚乳中的ABA信号能够释放到胚中并抑制GA的生物合成, 使种子保持休眠状态.随着种子吸胀时间的延长, 胚乳中ABA水平逐渐下调.此时胚中的PHYA介导远红光促进GA的生物合成, 进而促进种子萌发(Lee et al., 2012).因此, 种子的休眠和萌发是两个相互独立的生物学过程. ...
... 与野生种相比, 经过驯化的栽培种种子休眠水平下调, 如水稻(Oryza sativa)、小麦(Triticum aestivum)、大麦(Hordeum vulgare)和玉米(Zea mays)等禾本科作物(Bewley and Black, 1994).近年来, 育种学家们利用遗传学或组学的方法鉴定到一些与种子休眠相关的位点、转录本或蛋白(Alonso-Blanco et al., 2003; Argyris et al., 2005; Gu et al., 2006; Née et al., 2017b).例如, 研究者利用QTL方法鉴定到调控种子休眠的关键基因——DOG1 (Alonso-Blanco et al., 2003; Bentsink et al., 2006).后续研究发现, DOG1能够参与调控ABA信号通路以及GA的生物合成, 进而调控种子休眠(Graeber et al., 2014; Nishimura et al., 2018).那么, DOG1能否参与调控GA信号通路进而促进种子休眠? DOG1是否受到外源光信号和内源激素的调控? 针对上述问题的研究不仅有助于人们更加清晰地理解种子如何监测环境因子的变化, 从而决定是否完成从休眠到萌发的发育转变; 还为利用光信号或激素人工调控种子的休眠和萌发, 解决农业生产中“胎萌”以及穗发芽等问题提供了理论支撑. ...
Amplification of ABA biosynthesis and signaling through a positive feedback mechanism in seeds 1 2014
... 激素在种子休眠和萌发过程中发挥重要调控作用.其中, ABA和GA在调控种子休眠和萌发上行使主要功能.ABA促进种子休眠, 而GA促进种子萌发(Shu et al., 2016; Née et al., 2017a).在种子成熟过程中, ABA在种子内逐渐积累, 使种子的休眠水平逐渐升高.相比之下, 当种子受到吸胀或层积处理时, ABA含量逐渐降低, GA含量逐渐升高, 使种子萌发.与野生型种子相比, ABA合成缺陷突变体种子萌发更快(Frey et al., 2012); 而过表达ABA合成基因的种子以及ABA代谢突变体种子保持较高的休眠水平(Matakiadis et al., 2009; Martinez-Andújar et al., 2011; Nonogaki et al., 2014), 表明ABA能够通过其生物合成通路调控种子的休眠和萌发.ABA信号通过PYR/PYL/RCAR (PYRABACTIN RESISTANCE 1/PYR-LIKE PROTEINS/REGULATORY COMPONENTS OF ABA RECEPTORS)-PP2C (PROTEIN PHOSPHATASE 2C)-SnRKs (SNF1-RELATED PROTEIN KINASES)级联反应向下传递(Cutler et al., 2010; Hubbard et al., 2010).目前鉴定到的PP2C类蛋白包括ABI1 (ABSCISIC ACID INSENSITIVE 1)、ABI2、HON (HONSU)和RDO5 (REDUCED DORMANCY 5).其中, ABI1和ABI2通过与ABA信号受体蛋白PYR/PYL/RCAR互作进而抑制ABA信号转导(Ma et al., 2009; Park et al., 2009).HON蛋白能够通过抑制ABA信号转导及促进GA信号转导抑制种子休眠(Kim et al., 2013), 说明HON能够整合ABA和GA信号以调控种子休眠.rdo5突变体的休眠水平降低, 但其ABA含量和ABA敏感性并未发生显著变化(Xiang et al., 2014), 说明RDO5通过不依赖ABA的途径调控种子休眠.ABI3、ABI4和ABI5是ABA信号通路下游的关键组分, 能够抑制种子萌发、促进种子休眠(Bentsink and Koornneef, 2008; Kanai et al., 2010; Shu et al., 2013).响应ABA信号的R2R3型MYB转录因子MYB96通过调控ABI4以及ABA合成相关基因NCED2 (9-CIS-EPOXYCAROTENOID DIOXYGENASE 2)和NCED6的转录进而促进种子休眠、抑制种子萌发(Lee et al., 2015a, 2015b).上述研究表明, ABA不仅通过其生物合成通路调控种子的休眠和萌发, 还能以ABA信号的形式发挥作用. ...
PIL5, a phytochrome-interacting basic helix-loop-helix protein, is a key negative regulator of seed germination in Arabidopsis thaliana 1 2004
... 拟南芥基因组编码8个PIFs (PHYTOCHROME- INTERACTING FACTORs, PIF1-PIF8)蛋白.其中, PIF1和PIF6分别调控种子萌发和休眠(Oh et al., 2006; Penfield et al., 2010).PIF1作为种子萌发的负调控因子, 能够将内源激素和外源光信号连接起来, 在光介导的种子萌发过程中发挥关键作用.研究表明, 外源光信号通过调控PIF1蛋白稳定性或转录活性影响其对下游基因的转录调控.光照条件下, PIF1蛋白能够通过与Pfr形式的PHY互作, 进而发生泛素化降解(Oh et al., 2004; 2006); HFR1 (LONG HYPOCOTYL IN FAR-RED 1)通过PIF1的C端与之发生互作, 进而干扰PIF1的转录活性, 最终促进种子萌发(Shi et al., 2013).在黑暗条件下, DET1 (DE-ETIOLATED 1)和COP10以一种未知的机制稳定PIF1蛋白的活性(Shi et al., 2015).bHLH转录因子SPT (SPATULA)作为种子萌发的正调控因子, 在种子萌发中也发挥重要作用(Vaistij et al., 2018).此外, SPT还能调控种子休眠.然而, 在不同的生态型背景下SPT调控种子休眠的功能不同.在Ler背景下, SPT抑制种子休眠; 而在Col背景下, SPT促进种子休眠(Vaistij et al., 2013).外源光信号能否影响SPT对下游基因的转录调控? 目前还需阐释PHY调控SPT转录活性的作用机制, 以解答上述问题.最近的研究表明, 昼夜节律钟的关键组分CCA1 (CIRCADIAN CLOCK ASSOCIATED 1)、LHY (LATE ELONGATED HYPOCOTYL)以及RVE1 (REVEILLE1)均能介导外源光信号调控的种子休眠(Penfield and Hall, 2009; Jiang et al., 2016).此外, RVE1还能调控种子萌发(Jiang et al., 2016).IMB1 (IMBIBITION-INDUCIBLE1)是染色质域蛋白家族成员, 在PHYA介导的种子萌发中发挥作用.IMB1在干种子中的表达量很低, 随着种子吸胀呈现上调表达, 暗示IMB1能够促进种子萌发(Duque and Chua, 2003).目前, 关于IMB1调控PHYA介导的种子萌发的分子机制还有待深入探究.CSN (COP9 SIGNALOSOME)蛋白复合体是一类保守的蛋白复合体, 能够调控RING型E3泛素化连接酶的活性.CSN包含8个亚基(CSN1-CSN8) (Wei and Deng, 2003).其中, CSN1和CSN5参与调控种子萌发.csn1-10和csn5a-1突变体呈现出种子延迟萌发的表型(Jin et al., 2018). ...
PIL5, a phytochrome-interacting bHLH protein, regulates gibberellin responsiveness by binding directly to the GAI and RGA promoters in Arabidopsis seeds 2 2007
... PIF1能够介导外源光信号, 通过调控ABA和GA的生物合成调控种子萌发(图1A).统计Col和pif1突变体的种子在PHYB-on和PHYB-off条件下的萌发率, 发现Col只能在PHYB-on条件下萌发, pif1在PHYB-on和PHYB-off条件下均能萌发; 使用GA合成抑制剂PAC (paclobutrazol)能够抑制Col和pif1萌发.上述研究结果表明, PIF1在PHYB介导的种子萌发过程中发挥重要作用, 它可能通过调控GA的生物合成抑制种子萌发.进一步研究证实, PIF1能够抑制GA合成相关基因GA3ox1和GA3ox2的表达, 促进GA代谢相关基因GA2ox2的表达, 进而下调内源GA4的水平, 抑制种子萌发(Oh et al., 2006).与ga1单突变体相比, pif1/ga1双突变体中ABA合成相关基因的表达下调; 而ABA代谢相关基因的表达水平上调, 致使pif1/ga1双突变体中ABA的水平降低, 表现出持续萌发的表型(Oh et al., 2007).那么, PIF1如何调控ABA和GA生物合成相关基因的转录? 后续的研究发现, PIF1能够通过诱导SOM (SOMNUS)和DAG1 (DOF AFFECTING GERMINATION 1)的转录间接抑制GA合成、促进ABA合成, 最终抑制种子萌发(Kim et al., 2008; Gabriele et al., 2010) (图1A).其中, CCCH型锌指蛋白SOM通过调控组蛋白精氨酸去甲基化酶编码基因JMJ20 (JUMONJI 20)和JMJ22的表达, 进而影响GA合成基因GA3ox1和GA3ox2的甲基化水平, 最终抑制种子的萌发过程.此外, SOM还能激活GA代谢基因GA2ox2以及ABA合成基因ABA1 (ABA- DEFICIENT 1)、NCED6和NCED9的表达, 抑制ABA代谢基因CYP707A2表达, 从而抑制GA生物合成、促进ABA合成, 最终抑制种子萌发(Kim et al., 2008; de Wit et al., 2016).包含Dof结构域的DAG1蛋白直接结合在GA3ox1启动子上抑制其表达, 进而抑制种子萌发(Gabriele et al., 2010). ...
... 为了进一步探究外源光信号能否通过调控GA的信号转导影响种子萌发, 研究者检测了ga1突变体和pif1/ga1双突变体种子萌发对GA3的敏感性.结果发现, 与ga1突变体相比, pif1/ga1双突变体对GA3的敏感性升高.说明PIF1可能参与调控GA信号转导.进一步检测GA信号转导通路中关键组分的转录水平, 发现PIF1能够促进DELLA蛋白编码基因GAI和RGA (REPRESSOR OF GA1-3)的表达.该研究进一步证实PIF1能够直接结合在GAI和RGA的启动子上.上述结果表明, 黑暗条件下, PIF1蛋白能够直接诱导GAI和RGA的表达, 从而抑制GA信号转导, 最终抑制种子萌发(Oh et al., 2007).此外, PIF1还能通过诱导ABI3和ABI5的转录促进ABA信号转导, 从而抑制种子萌发(Park et al., 2011).SPT在不同生态型拟南芥背景下调控种子休眠的功能和作用机制不同.在Col背景下, SPT诱导ABI5和RGL3的表达, 从而促进ABA、抑制GA的信号转导, 最终促进种子休眠.在Ler背景下, SPT抑制ABI4和RGA的表达, 从而抑制ABA、促进GA的信号转导, 最终抑制种子休眠(Vaistij et al., 2013).此外, SPT还能通过抑制ABI5表达促进种子萌发(Vaistij et al., 2018).imb1功能缺失突变体的萌发率降低, 对ABA超敏感.进一步研究发现, ABI5转录本的积累可能是造成imb1突变体对ABA超敏感的原因之一(Duque and Chua, 2003).上述结果暗示, IMB1可能通过抑制ABA信号转导通路促进种子萌发(Duque and Chua, 2003).CSN蛋白复合体亚基CSN1和CSN5a的功能缺失突变体表现出延迟萌发的表型.去除种皮后csn1-10和csn5a-1能够正常萌发, 说明csn1-10和csn5a-1延迟萌发的表型依赖于种皮.种皮抑制的种子萌发与胚乳中RGL2相关, RGL2通过促进ABA的合成以及稳定ABI5蛋白进而抑制种子萌发.说明CSN1和CSN5a可能通过调控GA和ABA的信号转导促进种子萌发.进一步研究发现, CSN1通过促进RGL2的泛素化降解促进GA信号转导, 从而促进种子萌发; CSN5a通过抑制ABI5蛋白的稳定性进而抑制ABA信号转导, 最终促进种子萌发(Jin et al., 2018).上述研究结果表明, 外源光信号还能通过调控GA和ABA信号转导影响种子的休眠与萌发(图1A, B). ...
Light activates the degradation of PIL5 protein to promote seed germination through gibberellin in Arabidopsis 5 2006
... GA能够通过抑制ABA诱导的种子休眠促进种子萌发(Gubler et al., 2005; Graeber et al., 2012).在植物体内, 具有生物活性的GA主要包括GA1和GA4, 它们通过软化种皮、促进胚乳层细胞的水解和胚的生长打破种子休眠, 进而促进种子萌发(Holdsworth et al., 2008).GA缺失突变体ga1 (gibberellic acid- requiring 1)和ga2表现出强烈的休眠表型, 需要外源施加GA方可萌发(Lee et al., 2002; Shu et al., 2013).GA20ox (GA 20-OXIDASE)和GA3ox基因编码参与GA生物合成的氧化酶; GA2ox编码的GA2-氧化酶主要参与GA的代谢过程.在PHYB-off条件下, ga2ox突变体萌发水平升高(Oh et al., 2006; Yamauchi et al., 2007).上述研究结果表明, GA能通过其生物合成通路调控种子的休眠和萌发.在GA信号转导过程中, DELLA蛋白扮演着重要角色, 它能响应GA信号快速降解, 对GA信号转导起限速作用(Silverstone et al., 1998; Itoh et al., 2002; Zentella et al., 2007; Nemoto et al., 2017).GA受体蛋白GID1 (GA-INSENSITIVE DWARF 1)能够负调控DELLA蛋白的稳定性, 促进GA信号转导(Davière and Achard, 2013).拟南芥SLY1 (SLEEPY1)和水稻GID2 (GA-INSENSITIVE DWARF2)是SCF聚合体中的F-box亚基, 依赖其C端的GGF和LSL基序与DELLA蛋白C端VHIID和LHR2基序结合, 促进DELLA多聚泛素化(Hirano et al., 2010; Ariizumi et al., 2011).研究表明, DELLA蛋白GAI (GA-INSENSITIVE)能够通过抑制TCP14 (TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR 14)和TCP15的表达减缓细胞周期的进程, 最终使种子保持休眠状态(Resentini et al., 2015).作为转录调节子, DELLA如何实现对下游基因的转录调控? 有研究指出, DELLA能够通过与染色质重塑因子互作调控下游基因的转录.DELLA蛋白RGL2 (RGA-LIKE 2)和RGL3与染色质重塑因子SWI3C (SWITCH3C)发生互作, 进而影响GID1a以及GA3ox基因的转录(Sarnowska et al., 2013).此外, DELLA还能通过与种子萌发相关转录因子互作进而实现其对萌发的调控.RGL2通过与转录因子NF-YC互作, 诱导下游ABI5的转录, 进而抑制种子萌发(Liu et al., 2016).此外, RGL2还能通过与转录因子DOF6 (BINDING1 ZINC FINGER 6)互作诱导GATA12的表达, 进而促进种子休眠(Ravindran et al., 2017).DELLA通过与ICE1 (INDUCER OF CBF EXPRESSION 1)互作解除其对ABA响应基因EM1 (LATE EMBRYOGENESIS ABUNDANT 1)和EM6的转录抑制作用, 最终抑制种子萌发(Hu et al., 2019). ...
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
... 拟南芥基因组编码8个PIFs (PHYTOCHROME- INTERACTING FACTORs, PIF1-PIF8)蛋白.其中, PIF1和PIF6分别调控种子萌发和休眠(Oh et al., 2006; Penfield et al., 2010).PIF1作为种子萌发的负调控因子, 能够将内源激素和外源光信号连接起来, 在光介导的种子萌发过程中发挥关键作用.研究表明, 外源光信号通过调控PIF1蛋白稳定性或转录活性影响其对下游基因的转录调控.光照条件下, PIF1蛋白能够通过与Pfr形式的PHY互作, 进而发生泛素化降解(Oh et al., 2004; 2006); HFR1 (LONG HYPOCOTYL IN FAR-RED 1)通过PIF1的C端与之发生互作, 进而干扰PIF1的转录活性, 最终促进种子萌发(Shi et al., 2013).在黑暗条件下, DET1 (DE-ETIOLATED 1)和COP10以一种未知的机制稳定PIF1蛋白的活性(Shi et al., 2015).bHLH转录因子SPT (SPATULA)作为种子萌发的正调控因子, 在种子萌发中也发挥重要作用(Vaistij et al., 2018).此外, SPT还能调控种子休眠.然而, 在不同的生态型背景下SPT调控种子休眠的功能不同.在Ler背景下, SPT抑制种子休眠; 而在Col背景下, SPT促进种子休眠(Vaistij et al., 2013).外源光信号能否影响SPT对下游基因的转录调控? 目前还需阐释PHY调控SPT转录活性的作用机制, 以解答上述问题.最近的研究表明, 昼夜节律钟的关键组分CCA1 (CIRCADIAN CLOCK ASSOCIATED 1)、LHY (LATE ELONGATED HYPOCOTYL)以及RVE1 (REVEILLE1)均能介导外源光信号调控的种子休眠(Penfield and Hall, 2009; Jiang et al., 2016).此外, RVE1还能调控种子萌发(Jiang et al., 2016).IMB1 (IMBIBITION-INDUCIBLE1)是染色质域蛋白家族成员, 在PHYA介导的种子萌发中发挥作用.IMB1在干种子中的表达量很低, 随着种子吸胀呈现上调表达, 暗示IMB1能够促进种子萌发(Duque and Chua, 2003).目前, 关于IMB1调控PHYA介导的种子萌发的分子机制还有待深入探究.CSN (COP9 SIGNALOSOME)蛋白复合体是一类保守的蛋白复合体, 能够调控RING型E3泛素化连接酶的活性.CSN包含8个亚基(CSN1-CSN8) (Wei and Deng, 2003).其中, CSN1和CSN5参与调控种子萌发.csn1-10和csn5a-1突变体呈现出种子延迟萌发的表型(Jin et al., 2018). ...
... ; 2006); HFR1 (LONG HYPOCOTYL IN FAR-RED 1)通过PIF1的C端与之发生互作, 进而干扰PIF1的转录活性, 最终促进种子萌发(Shi et al., 2013).在黑暗条件下, DET1 (DE-ETIOLATED 1)和COP10以一种未知的机制稳定PIF1蛋白的活性(Shi et al., 2015).bHLH转录因子SPT (SPATULA)作为种子萌发的正调控因子, 在种子萌发中也发挥重要作用(Vaistij et al., 2018).此外, SPT还能调控种子休眠.然而, 在不同的生态型背景下SPT调控种子休眠的功能不同.在Ler背景下, SPT抑制种子休眠; 而在Col背景下, SPT促进种子休眠(Vaistij et al., 2013).外源光信号能否影响SPT对下游基因的转录调控? 目前还需阐释PHY调控SPT转录活性的作用机制, 以解答上述问题.最近的研究表明, 昼夜节律钟的关键组分CCA1 (CIRCADIAN CLOCK ASSOCIATED 1)、LHY (LATE ELONGATED HYPOCOTYL)以及RVE1 (REVEILLE1)均能介导外源光信号调控的种子休眠(Penfield and Hall, 2009; Jiang et al., 2016).此外, RVE1还能调控种子萌发(Jiang et al., 2016).IMB1 (IMBIBITION-INDUCIBLE1)是染色质域蛋白家族成员, 在PHYA介导的种子萌发中发挥作用.IMB1在干种子中的表达量很低, 随着种子吸胀呈现上调表达, 暗示IMB1能够促进种子萌发(Duque and Chua, 2003).目前, 关于IMB1调控PHYA介导的种子萌发的分子机制还有待深入探究.CSN (COP9 SIGNALOSOME)蛋白复合体是一类保守的蛋白复合体, 能够调控RING型E3泛素化连接酶的活性.CSN包含8个亚基(CSN1-CSN8) (Wei and Deng, 2003).其中, CSN1和CSN5参与调控种子萌发.csn1-10和csn5a-1突变体呈现出种子延迟萌发的表型(Jin et al., 2018). ...
... PIF1能够介导外源光信号, 通过调控ABA和GA的生物合成调控种子萌发(图1A).统计Col和pif1突变体的种子在PHYB-on和PHYB-off条件下的萌发率, 发现Col只能在PHYB-on条件下萌发, pif1在PHYB-on和PHYB-off条件下均能萌发; 使用GA合成抑制剂PAC (paclobutrazol)能够抑制Col和pif1萌发.上述研究结果表明, PIF1在PHYB介导的种子萌发过程中发挥重要作用, 它可能通过调控GA的生物合成抑制种子萌发.进一步研究证实, PIF1能够抑制GA合成相关基因GA3ox1和GA3ox2的表达, 促进GA代谢相关基因GA2ox2的表达, 进而下调内源GA4的水平, 抑制种子萌发(Oh et al., 2006).与ga1单突变体相比, pif1/ga1双突变体中ABA合成相关基因的表达下调; 而ABA代谢相关基因的表达水平上调, 致使pif1/ga1双突变体中ABA的水平降低, 表现出持续萌发的表型(Oh et al., 2007).那么, PIF1如何调控ABA和GA生物合成相关基因的转录? 后续的研究发现, PIF1能够通过诱导SOM (SOMNUS)和DAG1 (DOF AFFECTING GERMINATION 1)的转录间接抑制GA合成、促进ABA合成, 最终抑制种子萌发(Kim et al., 2008; Gabriele et al., 2010) (图1A).其中, CCCH型锌指蛋白SOM通过调控组蛋白精氨酸去甲基化酶编码基因JMJ20 (JUMONJI 20)和JMJ22的表达, 进而影响GA合成基因GA3ox1和GA3ox2的甲基化水平, 最终抑制种子的萌发过程.此外, SOM还能激活GA代谢基因GA2ox2以及ABA合成基因ABA1 (ABA- DEFICIENT 1)、NCED6和NCED9的表达, 抑制ABA代谢基因CYP707A2表达, 从而抑制GA生物合成、促进ABA合成, 最终抑制种子萌发(Kim et al., 2008; de Wit et al., 2016).包含Dof结构域的DAG1蛋白直接结合在GA3ox1启动子上抑制其表达, 进而抑制种子萌发(Gabriele et al., 2010). ...
ABI3 and PIL5 collaboratively activate the expression of SOMNUS by directly binding to its promoter in imbibed Arabidopsis seeds 1 2011
... 为了进一步探究外源光信号能否通过调控GA的信号转导影响种子萌发, 研究者检测了ga1突变体和pif1/ga1双突变体种子萌发对GA3的敏感性.结果发现, 与ga1突变体相比, pif1/ga1双突变体对GA3的敏感性升高.说明PIF1可能参与调控GA信号转导.进一步检测GA信号转导通路中关键组分的转录水平, 发现PIF1能够促进DELLA蛋白编码基因GAI和RGA (REPRESSOR OF GA1-3)的表达.该研究进一步证实PIF1能够直接结合在GAI和RGA的启动子上.上述结果表明, 黑暗条件下, PIF1蛋白能够直接诱导GAI和RGA的表达, 从而抑制GA信号转导, 最终抑制种子萌发(Oh et al., 2007).此外, PIF1还能通过诱导ABI3和ABI5的转录促进ABA信号转导, 从而抑制种子萌发(Park et al., 2011).SPT在不同生态型拟南芥背景下调控种子休眠的功能和作用机制不同.在Col背景下, SPT诱导ABI5和RGL3的表达, 从而促进ABA、抑制GA的信号转导, 最终促进种子休眠.在Ler背景下, SPT抑制ABI4和RGA的表达, 从而抑制ABA、促进GA的信号转导, 最终抑制种子休眠(Vaistij et al., 2013).此外, SPT还能通过抑制ABI5表达促进种子萌发(Vaistij et al., 2018).imb1功能缺失突变体的萌发率降低, 对ABA超敏感.进一步研究发现, ABI5转录本的积累可能是造成imb1突变体对ABA超敏感的原因之一(Duque and Chua, 2003).上述结果暗示, IMB1可能通过抑制ABA信号转导通路促进种子萌发(Duque and Chua, 2003).CSN蛋白复合体亚基CSN1和CSN5a的功能缺失突变体表现出延迟萌发的表型.去除种皮后csn1-10和csn5a-1能够正常萌发, 说明csn1-10和csn5a-1延迟萌发的表型依赖于种皮.种皮抑制的种子萌发与胚乳中RGL2相关, RGL2通过促进ABA的合成以及稳定ABI5蛋白进而抑制种子萌发.说明CSN1和CSN5a可能通过调控GA和ABA的信号转导促进种子萌发.进一步研究发现, CSN1通过促进RGL2的泛素化降解促进GA信号转导, 从而促进种子萌发; CSN5a通过抑制ABI5蛋白的稳定性进而抑制ABA信号转导, 最终促进种子萌发(Jin et al., 2018).上述研究结果表明, 外源光信号还能通过调控GA和ABA信号转导影响种子的休眠与萌发(图1A, B). ...
Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins 1 2009
... 激素在种子休眠和萌发过程中发挥重要调控作用.其中, ABA和GA在调控种子休眠和萌发上行使主要功能.ABA促进种子休眠, 而GA促进种子萌发(Shu et al., 2016; Née et al., 2017a).在种子成熟过程中, ABA在种子内逐渐积累, 使种子的休眠水平逐渐升高.相比之下, 当种子受到吸胀或层积处理时, ABA含量逐渐降低, GA含量逐渐升高, 使种子萌发.与野生型种子相比, ABA合成缺陷突变体种子萌发更快(Frey et al., 2012); 而过表达ABA合成基因的种子以及ABA代谢突变体种子保持较高的休眠水平(Matakiadis et al., 2009; Martinez-Andújar et al., 2011; Nonogaki et al., 2014), 表明ABA能够通过其生物合成通路调控种子的休眠和萌发.ABA信号通过PYR/PYL/RCAR (PYRABACTIN RESISTANCE 1/PYR-LIKE PROTEINS/REGULATORY COMPONENTS OF ABA RECEPTORS)-PP2C (PROTEIN PHOSPHATASE 2C)-SnRKs (SNF1-RELATED PROTEIN KINASES)级联反应向下传递(Cutler et al., 2010; Hubbard et al., 2010).目前鉴定到的PP2C类蛋白包括ABI1 (ABSCISIC ACID INSENSITIVE 1)、ABI2、HON (HONSU)和RDO5 (REDUCED DORMANCY 5).其中, ABI1和ABI2通过与ABA信号受体蛋白PYR/PYL/RCAR互作进而抑制ABA信号转导(Ma et al., 2009; Park et al., 2009).HON蛋白能够通过抑制ABA信号转导及促进GA信号转导抑制种子休眠(Kim et al., 2013), 说明HON能够整合ABA和GA信号以调控种子休眠.rdo5突变体的休眠水平降低, 但其ABA含量和ABA敏感性并未发生显著变化(Xiang et al., 2014), 说明RDO5通过不依赖ABA的途径调控种子休眠.ABI3、ABI4和ABI5是ABA信号通路下游的关键组分, 能够抑制种子萌发、促进种子休眠(Bentsink and Koornneef, 2008; Kanai et al., 2010; Shu et al., 2013).响应ABA信号的R2R3型MYB转录因子MYB96通过调控ABI4以及ABA合成相关基因NCED2 (9-CIS-EPOXYCAROTENOID DIOXYGENASE 2)和NCED6的转录进而促进种子休眠、抑制种子萌发(Lee et al., 2015a, 2015b).上述研究表明, ABA不仅通过其生物合成通路调控种子的休眠和萌发, 还能以ABA信号的形式发挥作用. ...
Cryptochromes interact directly with PIFs to control plant growth in limiting blue light 1 2016
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
A role for multiple circadian clock genes in the response to signals that break seed dormancy in Arabidopsis 1 2009
... 拟南芥基因组编码8个PIFs (PHYTOCHROME- INTERACTING FACTORs, PIF1-PIF8)蛋白.其中, PIF1和PIF6分别调控种子萌发和休眠(Oh et al., 2006; Penfield et al., 2010).PIF1作为种子萌发的负调控因子, 能够将内源激素和外源光信号连接起来, 在光介导的种子萌发过程中发挥关键作用.研究表明, 外源光信号通过调控PIF1蛋白稳定性或转录活性影响其对下游基因的转录调控.光照条件下, PIF1蛋白能够通过与Pfr形式的PHY互作, 进而发生泛素化降解(Oh et al., 2004; 2006); HFR1 (LONG HYPOCOTYL IN FAR-RED 1)通过PIF1的C端与之发生互作, 进而干扰PIF1的转录活性, 最终促进种子萌发(Shi et al., 2013).在黑暗条件下, DET1 (DE-ETIOLATED 1)和COP10以一种未知的机制稳定PIF1蛋白的活性(Shi et al., 2015).bHLH转录因子SPT (SPATULA)作为种子萌发的正调控因子, 在种子萌发中也发挥重要作用(Vaistij et al., 2018).此外, SPT还能调控种子休眠.然而, 在不同的生态型背景下SPT调控种子休眠的功能不同.在Ler背景下, SPT抑制种子休眠; 而在Col背景下, SPT促进种子休眠(Vaistij et al., 2013).外源光信号能否影响SPT对下游基因的转录调控? 目前还需阐释PHY调控SPT转录活性的作用机制, 以解答上述问题.最近的研究表明, 昼夜节律钟的关键组分CCA1 (CIRCADIAN CLOCK ASSOCIATED 1)、LHY (LATE ELONGATED HYPOCOTYL)以及RVE1 (REVEILLE1)均能介导外源光信号调控的种子休眠(Penfield and Hall, 2009; Jiang et al., 2016).此外, RVE1还能调控种子萌发(Jiang et al., 2016).IMB1 (IMBIBITION-INDUCIBLE1)是染色质域蛋白家族成员, 在PHYA介导的种子萌发中发挥作用.IMB1在干种子中的表达量很低, 随着种子吸胀呈现上调表达, 暗示IMB1能够促进种子萌发(Duque and Chua, 2003).目前, 关于IMB1调控PHYA介导的种子萌发的分子机制还有待深入探究.CSN (COP9 SIGNALOSOME)蛋白复合体是一类保守的蛋白复合体, 能够调控RING型E3泛素化连接酶的活性.CSN包含8个亚基(CSN1-CSN8) (Wei and Deng, 2003).其中, CSN1和CSN5参与调控种子萌发.csn1-10和csn5a-1突变体呈现出种子延迟萌发的表型(Jin et al., 2018). ...
A role for an alternative splice variant of PIF6 in the control of Arabidopsis primary seed dormancy 1 2010
... 拟南芥基因组编码8个PIFs (PHYTOCHROME- INTERACTING FACTORs, PIF1-PIF8)蛋白.其中, PIF1和PIF6分别调控种子萌发和休眠(Oh et al., 2006; Penfield et al., 2010).PIF1作为种子萌发的负调控因子, 能够将内源激素和外源光信号连接起来, 在光介导的种子萌发过程中发挥关键作用.研究表明, 外源光信号通过调控PIF1蛋白稳定性或转录活性影响其对下游基因的转录调控.光照条件下, PIF1蛋白能够通过与Pfr形式的PHY互作, 进而发生泛素化降解(Oh et al., 2004; 2006); HFR1 (LONG HYPOCOTYL IN FAR-RED 1)通过PIF1的C端与之发生互作, 进而干扰PIF1的转录活性, 最终促进种子萌发(Shi et al., 2013).在黑暗条件下, DET1 (DE-ETIOLATED 1)和COP10以一种未知的机制稳定PIF1蛋白的活性(Shi et al., 2015).bHLH转录因子SPT (SPATULA)作为种子萌发的正调控因子, 在种子萌发中也发挥重要作用(Vaistij et al., 2018).此外, SPT还能调控种子休眠.然而, 在不同的生态型背景下SPT调控种子休眠的功能不同.在Ler背景下, SPT抑制种子休眠; 而在Col背景下, SPT促进种子休眠(Vaistij et al., 2013).外源光信号能否影响SPT对下游基因的转录调控? 目前还需阐释PHY调控SPT转录活性的作用机制, 以解答上述问题.最近的研究表明, 昼夜节律钟的关键组分CCA1 (CIRCADIAN CLOCK ASSOCIATED 1)、LHY (LATE ELONGATED HYPOCOTYL)以及RVE1 (REVEILLE1)均能介导外源光信号调控的种子休眠(Penfield and Hall, 2009; Jiang et al., 2016).此外, RVE1还能调控种子萌发(Jiang et al., 2016).IMB1 (IMBIBITION-INDUCIBLE1)是染色质域蛋白家族成员, 在PHYA介导的种子萌发中发挥作用.IMB1在干种子中的表达量很低, 随着种子吸胀呈现上调表达, 暗示IMB1能够促进种子萌发(Duque and Chua, 2003).目前, 关于IMB1调控PHYA介导的种子萌发的分子机制还有待深入探究.CSN (COP9 SIGNALOSOME)蛋白复合体是一类保守的蛋白复合体, 能够调控RING型E3泛素化连接酶的活性.CSN包含8个亚基(CSN1-CSN8) (Wei and Deng, 2003).其中, CSN1和CSN5参与调控种子萌发.csn1-10和csn5a-1突变体呈现出种子延迟萌发的表型(Jin et al., 2018). ...
Phytochromes: photosensory perception and signal transduction 1 1995
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
A novel RGL2-DOF6 complex contributes to primary seed dormancy in Arabidopsis thaliana by regulating a GATA transcription factor 1 2017
... GA能够通过抑制ABA诱导的种子休眠促进种子萌发(Gubler et al., 2005; Graeber et al., 2012).在植物体内, 具有生物活性的GA主要包括GA1和GA4, 它们通过软化种皮、促进胚乳层细胞的水解和胚的生长打破种子休眠, 进而促进种子萌发(Holdsworth et al., 2008).GA缺失突变体ga1 (gibberellic acid- requiring 1)和ga2表现出强烈的休眠表型, 需要外源施加GA方可萌发(Lee et al., 2002; Shu et al., 2013).GA20ox (GA 20-OXIDASE)和GA3ox基因编码参与GA生物合成的氧化酶; GA2ox编码的GA2-氧化酶主要参与GA的代谢过程.在PHYB-off条件下, ga2ox突变体萌发水平升高(Oh et al., 2006; Yamauchi et al., 2007).上述研究结果表明, GA能通过其生物合成通路调控种子的休眠和萌发.在GA信号转导过程中, DELLA蛋白扮演着重要角色, 它能响应GA信号快速降解, 对GA信号转导起限速作用(Silverstone et al., 1998; Itoh et al., 2002; Zentella et al., 2007; Nemoto et al., 2017).GA受体蛋白GID1 (GA-INSENSITIVE DWARF 1)能够负调控DELLA蛋白的稳定性, 促进GA信号转导(Davière and Achard, 2013).拟南芥SLY1 (SLEEPY1)和水稻GID2 (GA-INSENSITIVE DWARF2)是SCF聚合体中的F-box亚基, 依赖其C端的GGF和LSL基序与DELLA蛋白C端VHIID和LHR2基序结合, 促进DELLA多聚泛素化(Hirano et al., 2010; Ariizumi et al., 2011).研究表明, DELLA蛋白GAI (GA-INSENSITIVE)能够通过抑制TCP14 (TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR 14)和TCP15的表达减缓细胞周期的进程, 最终使种子保持休眠状态(Resentini et al., 2015).作为转录调节子, DELLA如何实现对下游基因的转录调控? 有研究指出, DELLA能够通过与染色质重塑因子互作调控下游基因的转录.DELLA蛋白RGL2 (RGA-LIKE 2)和RGL3与染色质重塑因子SWI3C (SWITCH3C)发生互作, 进而影响GID1a以及GA3ox基因的转录(Sarnowska et al., 2013).此外, DELLA还能通过与种子萌发相关转录因子互作进而实现其对萌发的调控.RGL2通过与转录因子NF-YC互作, 诱导下游ABI5的转录, 进而抑制种子萌发(Liu et al., 2016).此外, RGL2还能通过与转录因子DOF6 (BINDING1 ZINC FINGER 6)互作诱导GATA12的表达, 进而促进种子休眠(Ravindran et al., 2017).DELLA通过与ICE1 (INDUCER OF CBF EXPRESSION 1)互作解除其对ABA响应基因EM1 (LATE EMBRYOGENESIS ABUNDANT 1)和EM6的转录抑制作用, 最终抑制种子萌发(Hu et al., 2019). ...
Two E3 ligases antagonistically regulate the UV-B response in Arabidopsis 1 2019
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
TCP14 and TCP15 mediate the promotion of seed germination by gibberellins in Arabidopsis thaliana 1 2015
... GA能够通过抑制ABA诱导的种子休眠促进种子萌发(Gubler et al., 2005; Graeber et al., 2012).在植物体内, 具有生物活性的GA主要包括GA1和GA4, 它们通过软化种皮、促进胚乳层细胞的水解和胚的生长打破种子休眠, 进而促进种子萌发(Holdsworth et al., 2008).GA缺失突变体ga1 (gibberellic acid- requiring 1)和ga2表现出强烈的休眠表型, 需要外源施加GA方可萌发(Lee et al., 2002; Shu et al., 2013).GA20ox (GA 20-OXIDASE)和GA3ox基因编码参与GA生物合成的氧化酶; GA2ox编码的GA2-氧化酶主要参与GA的代谢过程.在PHYB-off条件下, ga2ox突变体萌发水平升高(Oh et al., 2006; Yamauchi et al., 2007).上述研究结果表明, GA能通过其生物合成通路调控种子的休眠和萌发.在GA信号转导过程中, DELLA蛋白扮演着重要角色, 它能响应GA信号快速降解, 对GA信号转导起限速作用(Silverstone et al., 1998; Itoh et al., 2002; Zentella et al., 2007; Nemoto et al., 2017).GA受体蛋白GID1 (GA-INSENSITIVE DWARF 1)能够负调控DELLA蛋白的稳定性, 促进GA信号转导(Davière and Achard, 2013).拟南芥SLY1 (SLEEPY1)和水稻GID2 (GA-INSENSITIVE DWARF2)是SCF聚合体中的F-box亚基, 依赖其C端的GGF和LSL基序与DELLA蛋白C端VHIID和LHR2基序结合, 促进DELLA多聚泛素化(Hirano et al., 2010; Ariizumi et al., 2011).研究表明, DELLA蛋白GAI (GA-INSENSITIVE)能够通过抑制TCP14 (TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR 14)和TCP15的表达减缓细胞周期的进程, 最终使种子保持休眠状态(Resentini et al., 2015).作为转录调节子, DELLA如何实现对下游基因的转录调控? 有研究指出, DELLA能够通过与染色质重塑因子互作调控下游基因的转录.DELLA蛋白RGL2 (RGA-LIKE 2)和RGL3与染色质重塑因子SWI3C (SWITCH3C)发生互作, 进而影响GID1a以及GA3ox基因的转录(Sarnowska et al., 2013).此外, DELLA还能通过与种子萌发相关转录因子互作进而实现其对萌发的调控.RGL2通过与转录因子NF-YC互作, 诱导下游ABI5的转录, 进而抑制种子萌发(Liu et al., 2016).此外, RGL2还能通过与转录因子DOF6 (BINDING1 ZINC FINGER 6)互作诱导GATA12的表达, 进而促进种子休眠(Ravindran et al., 2017).DELLA通过与ICE1 (INDUCER OF CBF EXPRESSION 1)互作解除其对ABA响应基因EM1 (LATE EMBRYOGENESIS ABUNDANT 1)和EM6的转录抑制作用, 最终抑制种子萌发(Hu et al., 2019). ...
Perception of UV-B by the Arabidopsis UVR8 protein 1 2011
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
DELLA-interacting SWI3C core subunit of switch/sucrose nonfermenting chromatin remodeling complex modulates gibberellin responses and hormonal cross talk inArabidopsis 1 2013
... GA能够通过抑制ABA诱导的种子休眠促进种子萌发(Gubler et al., 2005; Graeber et al., 2012).在植物体内, 具有生物活性的GA主要包括GA1和GA4, 它们通过软化种皮、促进胚乳层细胞的水解和胚的生长打破种子休眠, 进而促进种子萌发(Holdsworth et al., 2008).GA缺失突变体ga1 (gibberellic acid- requiring 1)和ga2表现出强烈的休眠表型, 需要外源施加GA方可萌发(Lee et al., 2002; Shu et al., 2013).GA20ox (GA 20-OXIDASE)和GA3ox基因编码参与GA生物合成的氧化酶; GA2ox编码的GA2-氧化酶主要参与GA的代谢过程.在PHYB-off条件下, ga2ox突变体萌发水平升高(Oh et al., 2006; Yamauchi et al., 2007).上述研究结果表明, GA能通过其生物合成通路调控种子的休眠和萌发.在GA信号转导过程中, DELLA蛋白扮演着重要角色, 它能响应GA信号快速降解, 对GA信号转导起限速作用(Silverstone et al., 1998; Itoh et al., 2002; Zentella et al., 2007; Nemoto et al., 2017).GA受体蛋白GID1 (GA-INSENSITIVE DWARF 1)能够负调控DELLA蛋白的稳定性, 促进GA信号转导(Davière and Achard, 2013).拟南芥SLY1 (SLEEPY1)和水稻GID2 (GA-INSENSITIVE DWARF2)是SCF聚合体中的F-box亚基, 依赖其C端的GGF和LSL基序与DELLA蛋白C端VHIID和LHR2基序结合, 促进DELLA多聚泛素化(Hirano et al., 2010; Ariizumi et al., 2011).研究表明, DELLA蛋白GAI (GA-INSENSITIVE)能够通过抑制TCP14 (TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR 14)和TCP15的表达减缓细胞周期的进程, 最终使种子保持休眠状态(Resentini et al., 2015).作为转录调节子, DELLA如何实现对下游基因的转录调控? 有研究指出, DELLA能够通过与染色质重塑因子互作调控下游基因的转录.DELLA蛋白RGL2 (RGA-LIKE 2)和RGL3与染色质重塑因子SWI3C (SWITCH3C)发生互作, 进而影响GID1a以及GA3ox基因的转录(Sarnowska et al., 2013).此外, DELLA还能通过与种子萌发相关转录因子互作进而实现其对萌发的调控.RGL2通过与转录因子NF-YC互作, 诱导下游ABI5的转录, 进而抑制种子萌发(Liu et al., 2016).此外, RGL2还能通过与转录因子DOF6 (BINDING1 ZINC FINGER 6)互作诱导GATA12的表达, 进而促进种子休眠(Ravindran et al., 2017).DELLA通过与ICE1 (INDUCER OF CBF EXPRESSION 1)互作解除其对ABA响应基因EM1 (LATE EMBRYOGENESIS ABUNDANT 1)和EM6的转录抑制作用, 最终抑制种子萌发(Hu et al., 2019). ...
LAF1 ubiquitination by COP1 controls photomorphogenesis and is stimulated by SPA1 1 2003
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
Interaction of light and hormone signals in germinating seeds 1 2009
... PHY在黑暗条件下以生理失活的红光吸收型(Pr)存在, 吸收红光之后转变成其生理激活型(Pfr).两种光吸收型的PHY在Pr和Pfr两种状态间相互转变.早在20世纪50年代, 研究人员以莴苣(Lactuca sativa)种子为材料, 研究红光和远红光对其萌发的影响.结果表明红光促进种子萌发, 而远红光能够逆转红光的作用(Borthwick et al., 1952).红光和远红光对种子萌发的可逆调控暗示着PHY参与调控种子的萌发过程(Shinomura et al., 1994; Hennig et al., 2002).拟南芥PHY基因家族包含5个成员——PHYA-PHYE (Sharrock and Quail, 1989; Clack et al., 1994).种子萌发受到PHYA和PHYB的调控.其中, PHYB发挥主要功能.研究表明, PHYA蛋白在干种子中表达量很低, 随着种子在黑暗条件下吸胀时间的延长其蛋白表达量逐渐增加, 说明PHYA参与调控种子的萌发过程(Shinomura et al., 1996).PHYA主要在种子吸胀后期通过介导红光和远红光条件下的极低辐照度反应(VLFR)和远红光下的高辐照度反应(FR-HIR)调控种子萌发.相比之下, PHYB在干种子和吸胀种子中表达量均很高, 能够在种子吸胀初期(几个小时以内)介导红光和远红光下的低辐照度反应(LFR)调控种子萌发(Seo et al., 2009; Li et al., 2011).除了PHYA和PHYB外, PHYE也参与光调控的种子萌发过程, 而且三者在调控种子萌发方面功能冗余(Hennig et al., 2002).最近有研究表明, PHYB除了调控种子萌发, 还参与调控种子休眠(Jiang et al., 2016). ...
Novel phytochrome sequences in Arabidopsis thaliana: structure, evolution, and differential expression of a plant regulatory photoreceptor family 1 1989
... PHY在黑暗条件下以生理失活的红光吸收型(Pr)存在, 吸收红光之后转变成其生理激活型(Pfr).两种光吸收型的PHY在Pr和Pfr两种状态间相互转变.早在20世纪50年代, 研究人员以莴苣(Lactuca sativa)种子为材料, 研究红光和远红光对其萌发的影响.结果表明红光促进种子萌发, 而远红光能够逆转红光的作用(Borthwick et al., 1952).红光和远红光对种子萌发的可逆调控暗示着PHY参与调控种子的萌发过程(Shinomura et al., 1994; Hennig et al., 2002).拟南芥PHY基因家族包含5个成员——PHYA-PHYE (Sharrock and Quail, 1989; Clack et al., 1994).种子萌发受到PHYA和PHYB的调控.其中, PHYB发挥主要功能.研究表明, PHYA蛋白在干种子中表达量很低, 随着种子在黑暗条件下吸胀时间的延长其蛋白表达量逐渐增加, 说明PHYA参与调控种子的萌发过程(Shinomura et al., 1996).PHYA主要在种子吸胀后期通过介导红光和远红光条件下的极低辐照度反应(VLFR)和远红光下的高辐照度反应(FR-HIR)调控种子萌发.相比之下, PHYB在干种子和吸胀种子中表达量均很高, 能够在种子吸胀初期(几个小时以内)介导红光和远红光下的低辐照度反应(LFR)调控种子萌发(Seo et al., 2009; Li et al., 2011).除了PHYA和PHYB外, PHYE也参与光调控的种子萌发过程, 而且三者在调控种子萌发方面功能冗余(Hennig et al., 2002).最近有研究表明, PHYB除了调控种子萌发, 还参与调控种子休眠(Jiang et al., 2016). ...
Arabidopsis DET1 degrades HFR1 but stabilizes PIF1 to precisely regulate seed germination 1 2015
... 拟南芥基因组编码8个PIFs (PHYTOCHROME- INTERACTING FACTORs, PIF1-PIF8)蛋白.其中, PIF1和PIF6分别调控种子萌发和休眠(Oh et al., 2006; Penfield et al., 2010).PIF1作为种子萌发的负调控因子, 能够将内源激素和外源光信号连接起来, 在光介导的种子萌发过程中发挥关键作用.研究表明, 外源光信号通过调控PIF1蛋白稳定性或转录活性影响其对下游基因的转录调控.光照条件下, PIF1蛋白能够通过与Pfr形式的PHY互作, 进而发生泛素化降解(Oh et al., 2004; 2006); HFR1 (LONG HYPOCOTYL IN FAR-RED 1)通过PIF1的C端与之发生互作, 进而干扰PIF1的转录活性, 最终促进种子萌发(Shi et al., 2013).在黑暗条件下, DET1 (DE-ETIOLATED 1)和COP10以一种未知的机制稳定PIF1蛋白的活性(Shi et al., 2015).bHLH转录因子SPT (SPATULA)作为种子萌发的正调控因子, 在种子萌发中也发挥重要作用(Vaistij et al., 2018).此外, SPT还能调控种子休眠.然而, 在不同的生态型背景下SPT调控种子休眠的功能不同.在Ler背景下, SPT抑制种子休眠; 而在Col背景下, SPT促进种子休眠(Vaistij et al., 2013).外源光信号能否影响SPT对下游基因的转录调控? 目前还需阐释PHY调控SPT转录活性的作用机制, 以解答上述问题.最近的研究表明, 昼夜节律钟的关键组分CCA1 (CIRCADIAN CLOCK ASSOCIATED 1)、LHY (LATE ELONGATED HYPOCOTYL)以及RVE1 (REVEILLE1)均能介导外源光信号调控的种子休眠(Penfield and Hall, 2009; Jiang et al., 2016).此外, RVE1还能调控种子萌发(Jiang et al., 2016).IMB1 (IMBIBITION-INDUCIBLE1)是染色质域蛋白家族成员, 在PHYA介导的种子萌发中发挥作用.IMB1在干种子中的表达量很低, 随着种子吸胀呈现上调表达, 暗示IMB1能够促进种子萌发(Duque and Chua, 2003).目前, 关于IMB1调控PHYA介导的种子萌发的分子机制还有待深入探究.CSN (COP9 SIGNALOSOME)蛋白复合体是一类保守的蛋白复合体, 能够调控RING型E3泛素化连接酶的活性.CSN包含8个亚基(CSN1-CSN8) (Wei and Deng, 2003).其中, CSN1和CSN5参与调控种子萌发.csn1-10和csn5a-1突变体呈现出种子延迟萌发的表型(Jin et al., 2018). ...
HFR1 sequesters PIF1 to govern the transcriptional network underlying light-initiated seed germination in Arabidopsis 1 2013
... 拟南芥基因组编码8个PIFs (PHYTOCHROME- INTERACTING FACTORs, PIF1-PIF8)蛋白.其中, PIF1和PIF6分别调控种子萌发和休眠(Oh et al., 2006; Penfield et al., 2010).PIF1作为种子萌发的负调控因子, 能够将内源激素和外源光信号连接起来, 在光介导的种子萌发过程中发挥关键作用.研究表明, 外源光信号通过调控PIF1蛋白稳定性或转录活性影响其对下游基因的转录调控.光照条件下, PIF1蛋白能够通过与Pfr形式的PHY互作, 进而发生泛素化降解(Oh et al., 2004; 2006); HFR1 (LONG HYPOCOTYL IN FAR-RED 1)通过PIF1的C端与之发生互作, 进而干扰PIF1的转录活性, 最终促进种子萌发(Shi et al., 2013).在黑暗条件下, DET1 (DE-ETIOLATED 1)和COP10以一种未知的机制稳定PIF1蛋白的活性(Shi et al., 2015).bHLH转录因子SPT (SPATULA)作为种子萌发的正调控因子, 在种子萌发中也发挥重要作用(Vaistij et al., 2018).此外, SPT还能调控种子休眠.然而, 在不同的生态型背景下SPT调控种子休眠的功能不同.在Ler背景下, SPT抑制种子休眠; 而在Col背景下, SPT促进种子休眠(Vaistij et al., 2013).外源光信号能否影响SPT对下游基因的转录调控? 目前还需阐释PHY调控SPT转录活性的作用机制, 以解答上述问题.最近的研究表明, 昼夜节律钟的关键组分CCA1 (CIRCADIAN CLOCK ASSOCIATED 1)、LHY (LATE ELONGATED HYPOCOTYL)以及RVE1 (REVEILLE1)均能介导外源光信号调控的种子休眠(Penfield and Hall, 2009; Jiang et al., 2016).此外, RVE1还能调控种子萌发(Jiang et al., 2016).IMB1 (IMBIBITION-INDUCIBLE1)是染色质域蛋白家族成员, 在PHYA介导的种子萌发中发挥作用.IMB1在干种子中的表达量很低, 随着种子吸胀呈现上调表达, 暗示IMB1能够促进种子萌发(Duque and Chua, 2003).目前, 关于IMB1调控PHYA介导的种子萌发的分子机制还有待深入探究.CSN (COP9 SIGNALOSOME)蛋白复合体是一类保守的蛋白复合体, 能够调控RING型E3泛素化连接酶的活性.CSN包含8个亚基(CSN1-CSN8) (Wei and Deng, 2003).其中, CSN1和CSN5参与调控种子萌发.csn1-10和csn5a-1突变体呈现出种子延迟萌发的表型(Jin et al., 2018). ...
The induction of seed germination in Arabidopsis thaliana is regulated principally by Phytochrome B and secondarily by Phytochrome A 1 1994
... PHY在黑暗条件下以生理失活的红光吸收型(Pr)存在, 吸收红光之后转变成其生理激活型(Pfr).两种光吸收型的PHY在Pr和Pfr两种状态间相互转变.早在20世纪50年代, 研究人员以莴苣(Lactuca sativa)种子为材料, 研究红光和远红光对其萌发的影响.结果表明红光促进种子萌发, 而远红光能够逆转红光的作用(Borthwick et al., 1952).红光和远红光对种子萌发的可逆调控暗示着PHY参与调控种子的萌发过程(Shinomura et al., 1994; Hennig et al., 2002).拟南芥PHY基因家族包含5个成员——PHYA-PHYE (Sharrock and Quail, 1989; Clack et al., 1994).种子萌发受到PHYA和PHYB的调控.其中, PHYB发挥主要功能.研究表明, PHYA蛋白在干种子中表达量很低, 随着种子在黑暗条件下吸胀时间的延长其蛋白表达量逐渐增加, 说明PHYA参与调控种子的萌发过程(Shinomura et al., 1996).PHYA主要在种子吸胀后期通过介导红光和远红光条件下的极低辐照度反应(VLFR)和远红光下的高辐照度反应(FR-HIR)调控种子萌发.相比之下, PHYB在干种子和吸胀种子中表达量均很高, 能够在种子吸胀初期(几个小时以内)介导红光和远红光下的低辐照度反应(LFR)调控种子萌发(Seo et al., 2009; Li et al., 2011).除了PHYA和PHYB外, PHYE也参与光调控的种子萌发过程, 而且三者在调控种子萌发方面功能冗余(Hennig et al., 2002).最近有研究表明, PHYB除了调控种子萌发, 还参与调控种子休眠(Jiang et al., 2016). ...
Action spectra for phytochrome A- and B-specific photoinduction of seed germination in Arabidopsis thaliana 1 1996
... PHY在黑暗条件下以生理失活的红光吸收型(Pr)存在, 吸收红光之后转变成其生理激活型(Pfr).两种光吸收型的PHY在Pr和Pfr两种状态间相互转变.早在20世纪50年代, 研究人员以莴苣(Lactuca sativa)种子为材料, 研究红光和远红光对其萌发的影响.结果表明红光促进种子萌发, 而远红光能够逆转红光的作用(Borthwick et al., 1952).红光和远红光对种子萌发的可逆调控暗示着PHY参与调控种子的萌发过程(Shinomura et al., 1994; Hennig et al., 2002).拟南芥PHY基因家族包含5个成员——PHYA-PHYE (Sharrock and Quail, 1989; Clack et al., 1994).种子萌发受到PHYA和PHYB的调控.其中, PHYB发挥主要功能.研究表明, PHYA蛋白在干种子中表达量很低, 随着种子在黑暗条件下吸胀时间的延长其蛋白表达量逐渐增加, 说明PHYA参与调控种子的萌发过程(Shinomura et al., 1996).PHYA主要在种子吸胀后期通过介导红光和远红光条件下的极低辐照度反应(VLFR)和远红光下的高辐照度反应(FR-HIR)调控种子萌发.相比之下, PHYB在干种子和吸胀种子中表达量均很高, 能够在种子吸胀初期(几个小时以内)介导红光和远红光下的低辐照度反应(LFR)调控种子萌发(Seo et al., 2009; Li et al., 2011).除了PHYA和PHYB外, PHYE也参与光调控的种子萌发过程, 而且三者在调控种子萌发方面功能冗余(Hennig et al., 2002).最近有研究表明, PHYB除了调控种子萌发, 还参与调控种子休眠(Jiang et al., 2016). ...
Two faces of one seed: hormonal regulation of dormancy and germination 4 2016
... 种子休眠是指有活力的种子在适宜的条件下暂时不萌发的现象(Bewley, 1997; Finch-Savage and Leubner-Metzger, 2006).种子的休眠包括休眠的诱导、维持和释放3个阶段(Shu et al., 2016).成熟的种子释放休眠后, 在适宜的条件下即可萌发.DOG1 (DELAY OF GERMINATION 1)在种子休眠诱导和维持中发挥重要作用.它既能通过抑制脱落酸(abscisic acid, ABA)通路的负调控因子——AHG1(ABA HYPERSENSITIVE GERMINATION 1)的磷酸酶活性促进种子休眠(Nishimura et al., 2018); 也能通过调控响应赤霉素(gibberellin, GA)信号的细胞壁重塑相关基因的表达促进种子休眠(Graeber et al., 2014).在种子吸胀初期, 胚乳中PHYB (PHYTOCHROME B)吸收远红光后以生理失活型存在, 此时PIF1 (PIF3-LIKE 5, PIL5)蛋白逐渐积累, 使胚乳中ABA水平逐渐升高.胚乳中的ABA信号能够释放到胚中并抑制GA的生物合成, 使种子保持休眠状态.随着种子吸胀时间的延长, 胚乳中ABA水平逐渐下调.此时胚中的PHYA介导远红光促进GA的生物合成, 进而促进种子萌发(Lee et al., 2012).因此, 种子的休眠和萌发是两个相互独立的生物学过程. ...
... 研究表明, 种子的休眠及萌发受到内源激素和外源光信号的共同调控(Bassel, 2016; Shu et al., 2016).本文主要从外源光信号和内源激素对种子休眠和萌发的调控以及两类信号通路的互作3方面对模式植物拟南芥(Arabidopsis thaliana)种子休眠和萌发的调控机制进行综述, 以期为农业生产中利用光和激素调控种子休眠与萌发提供参考. ...
... 激素在种子休眠和萌发过程中发挥重要调控作用.其中, ABA和GA在调控种子休眠和萌发上行使主要功能.ABA促进种子休眠, 而GA促进种子萌发(Shu et al., 2016; Née et al., 2017a).在种子成熟过程中, ABA在种子内逐渐积累, 使种子的休眠水平逐渐升高.相比之下, 当种子受到吸胀或层积处理时, ABA含量逐渐降低, GA含量逐渐升高, 使种子萌发.与野生型种子相比, ABA合成缺陷突变体种子萌发更快(Frey et al., 2012); 而过表达ABA合成基因的种子以及ABA代谢突变体种子保持较高的休眠水平(Matakiadis et al., 2009; Martinez-Andújar et al., 2011; Nonogaki et al., 2014), 表明ABA能够通过其生物合成通路调控种子的休眠和萌发.ABA信号通过PYR/PYL/RCAR (PYRABACTIN RESISTANCE 1/PYR-LIKE PROTEINS/REGULATORY COMPONENTS OF ABA RECEPTORS)-PP2C (PROTEIN PHOSPHATASE 2C)-SnRKs (SNF1-RELATED PROTEIN KINASES)级联反应向下传递(Cutler et al., 2010; Hubbard et al., 2010).目前鉴定到的PP2C类蛋白包括ABI1 (ABSCISIC ACID INSENSITIVE 1)、ABI2、HON (HONSU)和RDO5 (REDUCED DORMANCY 5).其中, ABI1和ABI2通过与ABA信号受体蛋白PYR/PYL/RCAR互作进而抑制ABA信号转导(Ma et al., 2009; Park et al., 2009).HON蛋白能够通过抑制ABA信号转导及促进GA信号转导抑制种子休眠(Kim et al., 2013), 说明HON能够整合ABA和GA信号以调控种子休眠.rdo5突变体的休眠水平降低, 但其ABA含量和ABA敏感性并未发生显著变化(Xiang et al., 2014), 说明RDO5通过不依赖ABA的途径调控种子休眠.ABI3、ABI4和ABI5是ABA信号通路下游的关键组分, 能够抑制种子萌发、促进种子休眠(Bentsink and Koornneef, 2008; Kanai et al., 2010; Shu et al., 2013).响应ABA信号的R2R3型MYB转录因子MYB96通过调控ABI4以及ABA合成相关基因NCED2 (9-CIS-EPOXYCAROTENOID DIOXYGENASE 2)和NCED6的转录进而促进种子休眠、抑制种子萌发(Lee et al., 2015a, 2015b).上述研究表明, ABA不仅通过其生物合成通路调控种子的休眠和萌发, 还能以ABA信号的形式发挥作用. ...
... 除ABA和GA之外, 种子的休眠和萌发还受到7类激素的调控, 包括生长素(auxin)、乙烯(ethylene, ET)、油菜素内酯(brassinosteroids, BRs)、水杨酸(salicylic acid, SA)、茉莉酸(jasmonic acid, JA)、细胞分裂素(cytokinins, CTKs)和独脚金内酯(strigolactones, SLs).这些激素均通过影响ABA或GA通路间接调控种子的休眠和萌发(Shu et al., 2016; Née et al., 2017a).生长素能够通过调控ABI3的转录水平影响种子的休眠和萌发.当内源生长素水平升高时, 生长素信号通路中的负调控因子AXR2/3 (AUXIN- RESISTANT 2/3)发生降解.此时AXR2/3对响应生长素信号的ARF10 (AUXIN RESPONSE FACTOR 10)和ARF16基因的转录抑制作用被解除, 而ARF10和ARF16能够激活ABI3的表达, 从而使种子保持休眠状态(Liu et al., 2013a).然而, ARF10和ARF16不能直接结合在ABI3的启动子上(Liu et al., 2013a), 因此有待深入研究以揭示完整的生长素信号传递链.ET能够通过ABA生物合成和信号转导通路调控种子萌发(Cheng et al., 2009; Linkies et al., 2009; Corbineau et al., 2014), 但具体机制还不清楚.在盐胁迫条件下, ET受体蛋白ETR1 (ETHYLENE RESPONSE 1)和ETR2可能通过不依赖ET信号的通路调控ABA生物合成, 进而影响种子萌发(Wilson et al., 2014).最近的研究发现, ETR1通过解除ERF12 (ETHYLENE RESPONSE FACTOR 12)/TPL (TOPLESS)模块对DOG1基因的转录抑制作用而促进DOG1的转录, 进而促进种子休眠(Li et al., 2019).BR通过依赖MFT (MOTHER OFFT AND TFL 1)的途径拮抗ABA信号转导通路, 促进种子萌发(Xi and Yu, 2010; Xi et al., 2010).进一步的研究发现, BR信号通路中的负调控因子——BIN2 (BRASSINOSTEROID INSENSITIVE 2), 能够磷酸化并稳定ABI5蛋白, 进而促进ABA信号转导.BR信号通过抑制BIN2-ABI5互作解除ABA对种子萌发的抑制作用(Hu and Yu, 2014).SA在调控种子萌发上发挥双重作用.在正常生长条件下, SA通过抑制GA诱导的α淀粉酶编码基因的表达抑制种子萌发(Xie et al., 2007); 而在高盐胁迫下, SA通过其它通路促进种子萌发(Lee et al., 2010).CTKs通过抑制ABI5的转录或促进ABI5蛋白的降解拮抗ABA效应, 从而促进种子萌发(Wang et al., 2011; Guan et al., 2014).外源施加JA能够延迟种子萌发(Nambara et al., 2010), 表明JA能够抑制种子萌发.有研究表明, JA通过抑制ABA合成相关基因的表达、促进ABA代谢相关基因的表达拮抗ABA, 从而促进种子萌发(Jacobsen et al., 2013).JAZ3 (JASMONATE-ZIM DOMAIN PROTEIN 3)通过与ABI5互作抑制ABI5的转录激活活性, 进而促进种子萌发(Ju et al., 2019).目前关于JA调控种子萌发功能上存在矛盾的原因还有待深入探究.SLs通过降低ABA/GA值促进种子萌发(Toh et al., 2012).此外, SLs信号通路组分也能参与调控种子萌发, 如SMAX1 (SUPPRESSOR OF MORE AXILLARY GROWTH2 1) (Stanga et al., 2013).但目前SLs调控种子萌发的具体机制还不清楚.综上, 生长素、ET、BR、CTKs、JA和SLs通过调控ABA生物合成或信号通路调控种子的休眠和萌发.然而生长素、ET、BR、CTKs、JA与GA之间的交互作用还有待深入研究. ...
Dormancy and germination: how does the crop seed decide? 1 2015
... 休眠是种子植物在长期进化过程中产生的适应性性状, 植物通过抑制种子萌发保证其在不适宜的环境中生存, 因此休眠对于种子能够“适时萌发”具有重要意义.在实际生产中, 为了保证种子的高萌发率, 大部分作物的种子往往呈现出较低的休眠水平(Lenser and Theißen, 2013).然而过低的休眠水平导致成熟的种子在母体上直接萌发(“胎萌”)或出现穗发芽现象, 致使农业生产中种子的产量和质量下降(Simsek et al., 2014; Shu et al., 2015; Liu et al., 2019).因此, 研究种子休眠和萌发的调控机制对农业生产具有重要指导意义. ...
ABI4 regulates primary seed dormancy by regulating the biogenesis of abscisic acid and gibberellins in Arabidopsis 2 2013
... 激素在种子休眠和萌发过程中发挥重要调控作用.其中, ABA和GA在调控种子休眠和萌发上行使主要功能.ABA促进种子休眠, 而GA促进种子萌发(Shu et al., 2016; Née et al., 2017a).在种子成熟过程中, ABA在种子内逐渐积累, 使种子的休眠水平逐渐升高.相比之下, 当种子受到吸胀或层积处理时, ABA含量逐渐降低, GA含量逐渐升高, 使种子萌发.与野生型种子相比, ABA合成缺陷突变体种子萌发更快(Frey et al., 2012); 而过表达ABA合成基因的种子以及ABA代谢突变体种子保持较高的休眠水平(Matakiadis et al., 2009; Martinez-Andújar et al., 2011; Nonogaki et al., 2014), 表明ABA能够通过其生物合成通路调控种子的休眠和萌发.ABA信号通过PYR/PYL/RCAR (PYRABACTIN RESISTANCE 1/PYR-LIKE PROTEINS/REGULATORY COMPONENTS OF ABA RECEPTORS)-PP2C (PROTEIN PHOSPHATASE 2C)-SnRKs (SNF1-RELATED PROTEIN KINASES)级联反应向下传递(Cutler et al., 2010; Hubbard et al., 2010).目前鉴定到的PP2C类蛋白包括ABI1 (ABSCISIC ACID INSENSITIVE 1)、ABI2、HON (HONSU)和RDO5 (REDUCED DORMANCY 5).其中, ABI1和ABI2通过与ABA信号受体蛋白PYR/PYL/RCAR互作进而抑制ABA信号转导(Ma et al., 2009; Park et al., 2009).HON蛋白能够通过抑制ABA信号转导及促进GA信号转导抑制种子休眠(Kim et al., 2013), 说明HON能够整合ABA和GA信号以调控种子休眠.rdo5突变体的休眠水平降低, 但其ABA含量和ABA敏感性并未发生显著变化(Xiang et al., 2014), 说明RDO5通过不依赖ABA的途径调控种子休眠.ABI3、ABI4和ABI5是ABA信号通路下游的关键组分, 能够抑制种子萌发、促进种子休眠(Bentsink and Koornneef, 2008; Kanai et al., 2010; Shu et al., 2013).响应ABA信号的R2R3型MYB转录因子MYB96通过调控ABI4以及ABA合成相关基因NCED2 (9-CIS-EPOXYCAROTENOID DIOXYGENASE 2)和NCED6的转录进而促进种子休眠、抑制种子萌发(Lee et al., 2015a, 2015b).上述研究表明, ABA不仅通过其生物合成通路调控种子的休眠和萌发, 还能以ABA信号的形式发挥作用. ...
... GA能够通过抑制ABA诱导的种子休眠促进种子萌发(Gubler et al., 2005; Graeber et al., 2012).在植物体内, 具有生物活性的GA主要包括GA1和GA4, 它们通过软化种皮、促进胚乳层细胞的水解和胚的生长打破种子休眠, 进而促进种子萌发(Holdsworth et al., 2008).GA缺失突变体ga1 (gibberellic acid- requiring 1)和ga2表现出强烈的休眠表型, 需要外源施加GA方可萌发(Lee et al., 2002; Shu et al., 2013).GA20ox (GA 20-OXIDASE)和GA3ox基因编码参与GA生物合成的氧化酶; GA2ox编码的GA2-氧化酶主要参与GA的代谢过程.在PHYB-off条件下, ga2ox突变体萌发水平升高(Oh et al., 2006; Yamauchi et al., 2007).上述研究结果表明, GA能通过其生物合成通路调控种子的休眠和萌发.在GA信号转导过程中, DELLA蛋白扮演着重要角色, 它能响应GA信号快速降解, 对GA信号转导起限速作用(Silverstone et al., 1998; Itoh et al., 2002; Zentella et al., 2007; Nemoto et al., 2017).GA受体蛋白GID1 (GA-INSENSITIVE DWARF 1)能够负调控DELLA蛋白的稳定性, 促进GA信号转导(Davière and Achard, 2013).拟南芥SLY1 (SLEEPY1)和水稻GID2 (GA-INSENSITIVE DWARF2)是SCF聚合体中的F-box亚基, 依赖其C端的GGF和LSL基序与DELLA蛋白C端VHIID和LHR2基序结合, 促进DELLA多聚泛素化(Hirano et al., 2010; Ariizumi et al., 2011).研究表明, DELLA蛋白GAI (GA-INSENSITIVE)能够通过抑制TCP14 (TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR 14)和TCP15的表达减缓细胞周期的进程, 最终使种子保持休眠状态(Resentini et al., 2015).作为转录调节子, DELLA如何实现对下游基因的转录调控? 有研究指出, DELLA能够通过与染色质重塑因子互作调控下游基因的转录.DELLA蛋白RGL2 (RGA-LIKE 2)和RGL3与染色质重塑因子SWI3C (SWITCH3C)发生互作, 进而影响GID1a以及GA3ox基因的转录(Sarnowska et al., 2013).此外, DELLA还能通过与种子萌发相关转录因子互作进而实现其对萌发的调控.RGL2通过与转录因子NF-YC互作, 诱导下游ABI5的转录, 进而抑制种子萌发(Liu et al., 2016).此外, RGL2还能通过与转录因子DOF6 (BINDING1 ZINC FINGER 6)互作诱导GATA12的表达, 进而促进种子休眠(Ravindran et al., 2017).DELLA通过与ICE1 (INDUCER OF CBF EXPRESSION 1)互作解除其对ABA响应基因EM1 (LATE EMBRYOGENESIS ABUNDANT 1)和EM6的转录抑制作用, 最终抑制种子萌发(Hu et al., 2019). ...
The Arabidopsis RGA gene encodes a transcriptional regulator repressing the gibberellin signal transduction pathway 1 1998
... GA能够通过抑制ABA诱导的种子休眠促进种子萌发(Gubler et al., 2005; Graeber et al., 2012).在植物体内, 具有生物活性的GA主要包括GA1和GA4, 它们通过软化种皮、促进胚乳层细胞的水解和胚的生长打破种子休眠, 进而促进种子萌发(Holdsworth et al., 2008).GA缺失突变体ga1 (gibberellic acid- requiring 1)和ga2表现出强烈的休眠表型, 需要外源施加GA方可萌发(Lee et al., 2002; Shu et al., 2013).GA20ox (GA 20-OXIDASE)和GA3ox基因编码参与GA生物合成的氧化酶; GA2ox编码的GA2-氧化酶主要参与GA的代谢过程.在PHYB-off条件下, ga2ox突变体萌发水平升高(Oh et al., 2006; Yamauchi et al., 2007).上述研究结果表明, GA能通过其生物合成通路调控种子的休眠和萌发.在GA信号转导过程中, DELLA蛋白扮演着重要角色, 它能响应GA信号快速降解, 对GA信号转导起限速作用(Silverstone et al., 1998; Itoh et al., 2002; Zentella et al., 2007; Nemoto et al., 2017).GA受体蛋白GID1 (GA-INSENSITIVE DWARF 1)能够负调控DELLA蛋白的稳定性, 促进GA信号转导(Davière and Achard, 2013).拟南芥SLY1 (SLEEPY1)和水稻GID2 (GA-INSENSITIVE DWARF2)是SCF聚合体中的F-box亚基, 依赖其C端的GGF和LSL基序与DELLA蛋白C端VHIID和LHR2基序结合, 促进DELLA多聚泛素化(Hirano et al., 2010; Ariizumi et al., 2011).研究表明, DELLA蛋白GAI (GA-INSENSITIVE)能够通过抑制TCP14 (TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR 14)和TCP15的表达减缓细胞周期的进程, 最终使种子保持休眠状态(Resentini et al., 2015).作为转录调节子, DELLA如何实现对下游基因的转录调控? 有研究指出, DELLA能够通过与染色质重塑因子互作调控下游基因的转录.DELLA蛋白RGL2 (RGA-LIKE 2)和RGL3与染色质重塑因子SWI3C (SWITCH3C)发生互作, 进而影响GID1a以及GA3ox基因的转录(Sarnowska et al., 2013).此外, DELLA还能通过与种子萌发相关转录因子互作进而实现其对萌发的调控.RGL2通过与转录因子NF-YC互作, 诱导下游ABI5的转录, 进而抑制种子萌发(Liu et al., 2016).此外, RGL2还能通过与转录因子DOF6 (BINDING1 ZINC FINGER 6)互作诱导GATA12的表达, 进而促进种子休眠(Ravindran et al., 2017).DELLA通过与ICE1 (INDUCER OF CBF EXPRESSION 1)互作解除其对ABA响应基因EM1 (LATE EMBRYOGENESIS ABUNDANT 1)和EM6的转录抑制作用, 最终抑制种子萌发(Hu et al., 2019). ...
Effect of pre-harvest sprouting on physicochemical changes of proteins in wheat 1 2014
... 休眠是种子植物在长期进化过程中产生的适应性性状, 植物通过抑制种子萌发保证其在不适宜的环境中生存, 因此休眠对于种子能够“适时萌发”具有重要意义.在实际生产中, 为了保证种子的高萌发率, 大部分作物的种子往往呈现出较低的休眠水平(Lenser and Theißen, 2013).然而过低的休眠水平导致成熟的种子在母体上直接萌发(“胎萌”)或出现穗发芽现象, 致使农业生产中种子的产量和质量下降(Simsek et al., 2014; Shu et al., 2015; Liu et al., 2019).因此, 研究种子休眠和萌发的调控机制对农业生产具有重要指导意义. ...
SUPPRESSOR OF MORE AXILLARY GROWTH2 1 controls seed germination and seedling development in Arabidopsis 1 2013
... 除ABA和GA之外, 种子的休眠和萌发还受到7类激素的调控, 包括生长素(auxin)、乙烯(ethylene, ET)、油菜素内酯(brassinosteroids, BRs)、水杨酸(salicylic acid, SA)、茉莉酸(jasmonic acid, JA)、细胞分裂素(cytokinins, CTKs)和独脚金内酯(strigolactones, SLs).这些激素均通过影响ABA或GA通路间接调控种子的休眠和萌发(Shu et al., 2016; Née et al., 2017a).生长素能够通过调控ABI3的转录水平影响种子的休眠和萌发.当内源生长素水平升高时, 生长素信号通路中的负调控因子AXR2/3 (AUXIN- RESISTANT 2/3)发生降解.此时AXR2/3对响应生长素信号的ARF10 (AUXIN RESPONSE FACTOR 10)和ARF16基因的转录抑制作用被解除, 而ARF10和ARF16能够激活ABI3的表达, 从而使种子保持休眠状态(Liu et al., 2013a).然而, ARF10和ARF16不能直接结合在ABI3的启动子上(Liu et al., 2013a), 因此有待深入研究以揭示完整的生长素信号传递链.ET能够通过ABA生物合成和信号转导通路调控种子萌发(Cheng et al., 2009; Linkies et al., 2009; Corbineau et al., 2014), 但具体机制还不清楚.在盐胁迫条件下, ET受体蛋白ETR1 (ETHYLENE RESPONSE 1)和ETR2可能通过不依赖ET信号的通路调控ABA生物合成, 进而影响种子萌发(Wilson et al., 2014).最近的研究发现, ETR1通过解除ERF12 (ETHYLENE RESPONSE FACTOR 12)/TPL (TOPLESS)模块对DOG1基因的转录抑制作用而促进DOG1的转录, 进而促进种子休眠(Li et al., 2019).BR通过依赖MFT (MOTHER OFFT AND TFL 1)的途径拮抗ABA信号转导通路, 促进种子萌发(Xi and Yu, 2010; Xi et al., 2010).进一步的研究发现, BR信号通路中的负调控因子——BIN2 (BRASSINOSTEROID INSENSITIVE 2), 能够磷酸化并稳定ABI5蛋白, 进而促进ABA信号转导.BR信号通过抑制BIN2-ABI5互作解除ABA对种子萌发的抑制作用(Hu and Yu, 2014).SA在调控种子萌发上发挥双重作用.在正常生长条件下, SA通过抑制GA诱导的α淀粉酶编码基因的表达抑制种子萌发(Xie et al., 2007); 而在高盐胁迫下, SA通过其它通路促进种子萌发(Lee et al., 2010).CTKs通过抑制ABI5的转录或促进ABI5蛋白的降解拮抗ABA效应, 从而促进种子萌发(Wang et al., 2011; Guan et al., 2014).外源施加JA能够延迟种子萌发(Nambara et al., 2010), 表明JA能够抑制种子萌发.有研究表明, JA通过抑制ABA合成相关基因的表达、促进ABA代谢相关基因的表达拮抗ABA, 从而促进种子萌发(Jacobsen et al., 2013).JAZ3 (JASMONATE-ZIM DOMAIN PROTEIN 3)通过与ABI5互作抑制ABI5的转录激活活性, 进而促进种子萌发(Ju et al., 2019).目前关于JA调控种子萌发功能上存在矛盾的原因还有待深入探究.SLs通过降低ABA/GA值促进种子萌发(Toh et al., 2012).此外, SLs信号通路组分也能参与调控种子萌发, 如SMAX1 (SUPPRESSOR OF MORE AXILLARY GROWTH2 1) (Stanga et al., 2013).但目前SLs调控种子萌发的具体机制还不清楚.综上, 生长素、ET、BR、CTKs、JA和SLs通过调控ABA生物合成或信号通路调控种子的休眠和萌发.然而生长素、ET、BR、CTKs、JA与GA之间的交互作用还有待深入研究. ...
Thermoinhibition uncovers a role for strigolactones in Arabidopsis seed germination 1 2012
... 除ABA和GA之外, 种子的休眠和萌发还受到7类激素的调控, 包括生长素(auxin)、乙烯(ethylene, ET)、油菜素内酯(brassinosteroids, BRs)、水杨酸(salicylic acid, SA)、茉莉酸(jasmonic acid, JA)、细胞分裂素(cytokinins, CTKs)和独脚金内酯(strigolactones, SLs).这些激素均通过影响ABA或GA通路间接调控种子的休眠和萌发(Shu et al., 2016; Née et al., 2017a).生长素能够通过调控ABI3的转录水平影响种子的休眠和萌发.当内源生长素水平升高时, 生长素信号通路中的负调控因子AXR2/3 (AUXIN- RESISTANT 2/3)发生降解.此时AXR2/3对响应生长素信号的ARF10 (AUXIN RESPONSE FACTOR 10)和ARF16基因的转录抑制作用被解除, 而ARF10和ARF16能够激活ABI3的表达, 从而使种子保持休眠状态(Liu et al., 2013a).然而, ARF10和ARF16不能直接结合在ABI3的启动子上(Liu et al., 2013a), 因此有待深入研究以揭示完整的生长素信号传递链.ET能够通过ABA生物合成和信号转导通路调控种子萌发(Cheng et al., 2009; Linkies et al., 2009; Corbineau et al., 2014), 但具体机制还不清楚.在盐胁迫条件下, ET受体蛋白ETR1 (ETHYLENE RESPONSE 1)和ETR2可能通过不依赖ET信号的通路调控ABA生物合成, 进而影响种子萌发(Wilson et al., 2014).最近的研究发现, ETR1通过解除ERF12 (ETHYLENE RESPONSE FACTOR 12)/TPL (TOPLESS)模块对DOG1基因的转录抑制作用而促进DOG1的转录, 进而促进种子休眠(Li et al., 2019).BR通过依赖MFT (MOTHER OFFT AND TFL 1)的途径拮抗ABA信号转导通路, 促进种子萌发(Xi and Yu, 2010; Xi et al., 2010).进一步的研究发现, BR信号通路中的负调控因子——BIN2 (BRASSINOSTEROID INSENSITIVE 2), 能够磷酸化并稳定ABI5蛋白, 进而促进ABA信号转导.BR信号通过抑制BIN2-ABI5互作解除ABA对种子萌发的抑制作用(Hu and Yu, 2014).SA在调控种子萌发上发挥双重作用.在正常生长条件下, SA通过抑制GA诱导的α淀粉酶编码基因的表达抑制种子萌发(Xie et al., 2007); 而在高盐胁迫下, SA通过其它通路促进种子萌发(Lee et al., 2010).CTKs通过抑制ABI5的转录或促进ABI5蛋白的降解拮抗ABA效应, 从而促进种子萌发(Wang et al., 2011; Guan et al., 2014).外源施加JA能够延迟种子萌发(Nambara et al., 2010), 表明JA能够抑制种子萌发.有研究表明, JA通过抑制ABA合成相关基因的表达、促进ABA代谢相关基因的表达拮抗ABA, 从而促进种子萌发(Jacobsen et al., 2013).JAZ3 (JASMONATE-ZIM DOMAIN PROTEIN 3)通过与ABI5互作抑制ABI5的转录激活活性, 进而促进种子萌发(Ju et al., 2019).目前关于JA调控种子萌发功能上存在矛盾的原因还有待深入探究.SLs通过降低ABA/GA值促进种子萌发(Toh et al., 2012).此外, SLs信号通路组分也能参与调控种子萌发, 如SMAX1 (SUPPRESSOR OF MORE AXILLARY GROWTH2 1) (Stanga et al., 2013).但目前SLs调控种子萌发的具体机制还不清楚.综上, 生长素、ET、BR、CTKs、JA和SLs通过调控ABA生物合成或信号通路调控种子的休眠和萌发.然而生长素、ET、BR、CTKs、JA与GA之间的交互作用还有待深入研究. ...
MOTHER-OF-FT-AND-TFL1 represses seed germination under far-red light by modulating phytohormone responses in Arabidopsis thaliana 2 2018
... 拟南芥基因组编码8个PIFs (PHYTOCHROME- INTERACTING FACTORs, PIF1-PIF8)蛋白.其中, PIF1和PIF6分别调控种子萌发和休眠(Oh et al., 2006; Penfield et al., 2010).PIF1作为种子萌发的负调控因子, 能够将内源激素和外源光信号连接起来, 在光介导的种子萌发过程中发挥关键作用.研究表明, 外源光信号通过调控PIF1蛋白稳定性或转录活性影响其对下游基因的转录调控.光照条件下, PIF1蛋白能够通过与Pfr形式的PHY互作, 进而发生泛素化降解(Oh et al., 2004; 2006); HFR1 (LONG HYPOCOTYL IN FAR-RED 1)通过PIF1的C端与之发生互作, 进而干扰PIF1的转录活性, 最终促进种子萌发(Shi et al., 2013).在黑暗条件下, DET1 (DE-ETIOLATED 1)和COP10以一种未知的机制稳定PIF1蛋白的活性(Shi et al., 2015).bHLH转录因子SPT (SPATULA)作为种子萌发的正调控因子, 在种子萌发中也发挥重要作用(Vaistij et al., 2018).此外, SPT还能调控种子休眠.然而, 在不同的生态型背景下SPT调控种子休眠的功能不同.在Ler背景下, SPT抑制种子休眠; 而在Col背景下, SPT促进种子休眠(Vaistij et al., 2013).外源光信号能否影响SPT对下游基因的转录调控? 目前还需阐释PHY调控SPT转录活性的作用机制, 以解答上述问题.最近的研究表明, 昼夜节律钟的关键组分CCA1 (CIRCADIAN CLOCK ASSOCIATED 1)、LHY (LATE ELONGATED HYPOCOTYL)以及RVE1 (REVEILLE1)均能介导外源光信号调控的种子休眠(Penfield and Hall, 2009; Jiang et al., 2016).此外, RVE1还能调控种子萌发(Jiang et al., 2016).IMB1 (IMBIBITION-INDUCIBLE1)是染色质域蛋白家族成员, 在PHYA介导的种子萌发中发挥作用.IMB1在干种子中的表达量很低, 随着种子吸胀呈现上调表达, 暗示IMB1能够促进种子萌发(Duque and Chua, 2003).目前, 关于IMB1调控PHYA介导的种子萌发的分子机制还有待深入探究.CSN (COP9 SIGNALOSOME)蛋白复合体是一类保守的蛋白复合体, 能够调控RING型E3泛素化连接酶的活性.CSN包含8个亚基(CSN1-CSN8) (Wei and Deng, 2003).其中, CSN1和CSN5参与调控种子萌发.csn1-10和csn5a-1突变体呈现出种子延迟萌发的表型(Jin et al., 2018). ...
... 为了进一步探究外源光信号能否通过调控GA的信号转导影响种子萌发, 研究者检测了ga1突变体和pif1/ga1双突变体种子萌发对GA3的敏感性.结果发现, 与ga1突变体相比, pif1/ga1双突变体对GA3的敏感性升高.说明PIF1可能参与调控GA信号转导.进一步检测GA信号转导通路中关键组分的转录水平, 发现PIF1能够促进DELLA蛋白编码基因GAI和RGA (REPRESSOR OF GA1-3)的表达.该研究进一步证实PIF1能够直接结合在GAI和RGA的启动子上.上述结果表明, 黑暗条件下, PIF1蛋白能够直接诱导GAI和RGA的表达, 从而抑制GA信号转导, 最终抑制种子萌发(Oh et al., 2007).此外, PIF1还能通过诱导ABI3和ABI5的转录促进ABA信号转导, 从而抑制种子萌发(Park et al., 2011).SPT在不同生态型拟南芥背景下调控种子休眠的功能和作用机制不同.在Col背景下, SPT诱导ABI5和RGL3的表达, 从而促进ABA、抑制GA的信号转导, 最终促进种子休眠.在Ler背景下, SPT抑制ABI4和RGA的表达, 从而抑制ABA、促进GA的信号转导, 最终抑制种子休眠(Vaistij et al., 2013).此外, SPT还能通过抑制ABI5表达促进种子萌发(Vaistij et al., 2018).imb1功能缺失突变体的萌发率降低, 对ABA超敏感.进一步研究发现, ABI5转录本的积累可能是造成imb1突变体对ABA超敏感的原因之一(Duque and Chua, 2003).上述结果暗示, IMB1可能通过抑制ABA信号转导通路促进种子萌发(Duque and Chua, 2003).CSN蛋白复合体亚基CSN1和CSN5a的功能缺失突变体表现出延迟萌发的表型.去除种皮后csn1-10和csn5a-1能够正常萌发, 说明csn1-10和csn5a-1延迟萌发的表型依赖于种皮.种皮抑制的种子萌发与胚乳中RGL2相关, RGL2通过促进ABA的合成以及稳定ABI5蛋白进而抑制种子萌发.说明CSN1和CSN5a可能通过调控GA和ABA的信号转导促进种子萌发.进一步研究发现, CSN1通过促进RGL2的泛素化降解促进GA信号转导, 从而促进种子萌发; CSN5a通过抑制ABI5蛋白的稳定性进而抑制ABA信号转导, 最终促进种子萌发(Jin et al., 2018).上述研究结果表明, 外源光信号还能通过调控GA和ABA信号转导影响种子的休眠与萌发(图1A, B). ...
Differential control of seed primary dormancy in Arabidopsis ecotypes by the transcription factor SPATULA 2 2013
... 拟南芥基因组编码8个PIFs (PHYTOCHROME- INTERACTING FACTORs, PIF1-PIF8)蛋白.其中, PIF1和PIF6分别调控种子萌发和休眠(Oh et al., 2006; Penfield et al., 2010).PIF1作为种子萌发的负调控因子, 能够将内源激素和外源光信号连接起来, 在光介导的种子萌发过程中发挥关键作用.研究表明, 外源光信号通过调控PIF1蛋白稳定性或转录活性影响其对下游基因的转录调控.光照条件下, PIF1蛋白能够通过与Pfr形式的PHY互作, 进而发生泛素化降解(Oh et al., 2004; 2006); HFR1 (LONG HYPOCOTYL IN FAR-RED 1)通过PIF1的C端与之发生互作, 进而干扰PIF1的转录活性, 最终促进种子萌发(Shi et al., 2013).在黑暗条件下, DET1 (DE-ETIOLATED 1)和COP10以一种未知的机制稳定PIF1蛋白的活性(Shi et al., 2015).bHLH转录因子SPT (SPATULA)作为种子萌发的正调控因子, 在种子萌发中也发挥重要作用(Vaistij et al., 2018).此外, SPT还能调控种子休眠.然而, 在不同的生态型背景下SPT调控种子休眠的功能不同.在Ler背景下, SPT抑制种子休眠; 而在Col背景下, SPT促进种子休眠(Vaistij et al., 2013).外源光信号能否影响SPT对下游基因的转录调控? 目前还需阐释PHY调控SPT转录活性的作用机制, 以解答上述问题.最近的研究表明, 昼夜节律钟的关键组分CCA1 (CIRCADIAN CLOCK ASSOCIATED 1)、LHY (LATE ELONGATED HYPOCOTYL)以及RVE1 (REVEILLE1)均能介导外源光信号调控的种子休眠(Penfield and Hall, 2009; Jiang et al., 2016).此外, RVE1还能调控种子萌发(Jiang et al., 2016).IMB1 (IMBIBITION-INDUCIBLE1)是染色质域蛋白家族成员, 在PHYA介导的种子萌发中发挥作用.IMB1在干种子中的表达量很低, 随着种子吸胀呈现上调表达, 暗示IMB1能够促进种子萌发(Duque and Chua, 2003).目前, 关于IMB1调控PHYA介导的种子萌发的分子机制还有待深入探究.CSN (COP9 SIGNALOSOME)蛋白复合体是一类保守的蛋白复合体, 能够调控RING型E3泛素化连接酶的活性.CSN包含8个亚基(CSN1-CSN8) (Wei and Deng, 2003).其中, CSN1和CSN5参与调控种子萌发.csn1-10和csn5a-1突变体呈现出种子延迟萌发的表型(Jin et al., 2018). ...
... 为了进一步探究外源光信号能否通过调控GA的信号转导影响种子萌发, 研究者检测了ga1突变体和pif1/ga1双突变体种子萌发对GA3的敏感性.结果发现, 与ga1突变体相比, pif1/ga1双突变体对GA3的敏感性升高.说明PIF1可能参与调控GA信号转导.进一步检测GA信号转导通路中关键组分的转录水平, 发现PIF1能够促进DELLA蛋白编码基因GAI和RGA (REPRESSOR OF GA1-3)的表达.该研究进一步证实PIF1能够直接结合在GAI和RGA的启动子上.上述结果表明, 黑暗条件下, PIF1蛋白能够直接诱导GAI和RGA的表达, 从而抑制GA信号转导, 最终抑制种子萌发(Oh et al., 2007).此外, PIF1还能通过诱导ABI3和ABI5的转录促进ABA信号转导, 从而抑制种子萌发(Park et al., 2011).SPT在不同生态型拟南芥背景下调控种子休眠的功能和作用机制不同.在Col背景下, SPT诱导ABI5和RGL3的表达, 从而促进ABA、抑制GA的信号转导, 最终促进种子休眠.在Ler背景下, SPT抑制ABI4和RGA的表达, 从而抑制ABA、促进GA的信号转导, 最终抑制种子休眠(Vaistij et al., 2013).此外, SPT还能通过抑制ABI5表达促进种子萌发(Vaistij et al., 2018).imb1功能缺失突变体的萌发率降低, 对ABA超敏感.进一步研究发现, ABI5转录本的积累可能是造成imb1突变体对ABA超敏感的原因之一(Duque and Chua, 2003).上述结果暗示, IMB1可能通过抑制ABA信号转导通路促进种子萌发(Duque and Chua, 2003).CSN蛋白复合体亚基CSN1和CSN5a的功能缺失突变体表现出延迟萌发的表型.去除种皮后csn1-10和csn5a-1能够正常萌发, 说明csn1-10和csn5a-1延迟萌发的表型依赖于种皮.种皮抑制的种子萌发与胚乳中RGL2相关, RGL2通过促进ABA的合成以及稳定ABI5蛋白进而抑制种子萌发.说明CSN1和CSN5a可能通过调控GA和ABA的信号转导促进种子萌发.进一步研究发现, CSN1通过促进RGL2的泛素化降解促进GA信号转导, 从而促进种子萌发; CSN5a通过抑制ABI5蛋白的稳定性进而抑制ABA信号转导, 最终促进种子萌发(Jin et al., 2018).上述研究结果表明, 外源光信号还能通过调控GA和ABA信号转导影响种子的休眠与萌发(图1A, B). ...
Cytokinin antagonizes ABA suppression to seed germination of Arabidopsis by downregulating ABI5 expression 1 2011
... 除ABA和GA之外, 种子的休眠和萌发还受到7类激素的调控, 包括生长素(auxin)、乙烯(ethylene, ET)、油菜素内酯(brassinosteroids, BRs)、水杨酸(salicylic acid, SA)、茉莉酸(jasmonic acid, JA)、细胞分裂素(cytokinins, CTKs)和独脚金内酯(strigolactones, SLs).这些激素均通过影响ABA或GA通路间接调控种子的休眠和萌发(Shu et al., 2016; Née et al., 2017a).生长素能够通过调控ABI3的转录水平影响种子的休眠和萌发.当内源生长素水平升高时, 生长素信号通路中的负调控因子AXR2/3 (AUXIN- RESISTANT 2/3)发生降解.此时AXR2/3对响应生长素信号的ARF10 (AUXIN RESPONSE FACTOR 10)和ARF16基因的转录抑制作用被解除, 而ARF10和ARF16能够激活ABI3的表达, 从而使种子保持休眠状态(Liu et al., 2013a).然而, ARF10和ARF16不能直接结合在ABI3的启动子上(Liu et al., 2013a), 因此有待深入研究以揭示完整的生长素信号传递链.ET能够通过ABA生物合成和信号转导通路调控种子萌发(Cheng et al., 2009; Linkies et al., 2009; Corbineau et al., 2014), 但具体机制还不清楚.在盐胁迫条件下, ET受体蛋白ETR1 (ETHYLENE RESPONSE 1)和ETR2可能通过不依赖ET信号的通路调控ABA生物合成, 进而影响种子萌发(Wilson et al., 2014).最近的研究发现, ETR1通过解除ERF12 (ETHYLENE RESPONSE FACTOR 12)/TPL (TOPLESS)模块对DOG1基因的转录抑制作用而促进DOG1的转录, 进而促进种子休眠(Li et al., 2019).BR通过依赖MFT (MOTHER OFFT AND TFL 1)的途径拮抗ABA信号转导通路, 促进种子萌发(Xi and Yu, 2010; Xi et al., 2010).进一步的研究发现, BR信号通路中的负调控因子——BIN2 (BRASSINOSTEROID INSENSITIVE 2), 能够磷酸化并稳定ABI5蛋白, 进而促进ABA信号转导.BR信号通过抑制BIN2-ABI5互作解除ABA对种子萌发的抑制作用(Hu and Yu, 2014).SA在调控种子萌发上发挥双重作用.在正常生长条件下, SA通过抑制GA诱导的α淀粉酶编码基因的表达抑制种子萌发(Xie et al., 2007); 而在高盐胁迫下, SA通过其它通路促进种子萌发(Lee et al., 2010).CTKs通过抑制ABI5的转录或促进ABI5蛋白的降解拮抗ABA效应, 从而促进种子萌发(Wang et al., 2011; Guan et al., 2014).外源施加JA能够延迟种子萌发(Nambara et al., 2010), 表明JA能够抑制种子萌发.有研究表明, JA通过抑制ABA合成相关基因的表达、促进ABA代谢相关基因的表达拮抗ABA, 从而促进种子萌发(Jacobsen et al., 2013).JAZ3 (JASMONATE-ZIM DOMAIN PROTEIN 3)通过与ABI5互作抑制ABI5的转录激活活性, 进而促进种子萌发(Ju et al., 2019).目前关于JA调控种子萌发功能上存在矛盾的原因还有待深入探究.SLs通过降低ABA/GA值促进种子萌发(Toh et al., 2012).此外, SLs信号通路组分也能参与调控种子萌发, 如SMAX1 (SUPPRESSOR OF MORE AXILLARY GROWTH2 1) (Stanga et al., 2013).但目前SLs调控种子萌发的具体机制还不清楚.综上, 生长素、ET、BR、CTKs、JA和SLs通过调控ABA生物合成或信号通路调控种子的休眠和萌发.然而生长素、ET、BR、CTKs、JA与GA之间的交互作用还有待深入研究. ...
The COP9 signalosome 1 2003
... 拟南芥基因组编码8个PIFs (PHYTOCHROME- INTERACTING FACTORs, PIF1-PIF8)蛋白.其中, PIF1和PIF6分别调控种子萌发和休眠(Oh et al., 2006; Penfield et al., 2010).PIF1作为种子萌发的负调控因子, 能够将内源激素和外源光信号连接起来, 在光介导的种子萌发过程中发挥关键作用.研究表明, 外源光信号通过调控PIF1蛋白稳定性或转录活性影响其对下游基因的转录调控.光照条件下, PIF1蛋白能够通过与Pfr形式的PHY互作, 进而发生泛素化降解(Oh et al., 2004; 2006); HFR1 (LONG HYPOCOTYL IN FAR-RED 1)通过PIF1的C端与之发生互作, 进而干扰PIF1的转录活性, 最终促进种子萌发(Shi et al., 2013).在黑暗条件下, DET1 (DE-ETIOLATED 1)和COP10以一种未知的机制稳定PIF1蛋白的活性(Shi et al., 2015).bHLH转录因子SPT (SPATULA)作为种子萌发的正调控因子, 在种子萌发中也发挥重要作用(Vaistij et al., 2018).此外, SPT还能调控种子休眠.然而, 在不同的生态型背景下SPT调控种子休眠的功能不同.在Ler背景下, SPT抑制种子休眠; 而在Col背景下, SPT促进种子休眠(Vaistij et al., 2013).外源光信号能否影响SPT对下游基因的转录调控? 目前还需阐释PHY调控SPT转录活性的作用机制, 以解答上述问题.最近的研究表明, 昼夜节律钟的关键组分CCA1 (CIRCADIAN CLOCK ASSOCIATED 1)、LHY (LATE ELONGATED HYPOCOTYL)以及RVE1 (REVEILLE1)均能介导外源光信号调控的种子休眠(Penfield and Hall, 2009; Jiang et al., 2016).此外, RVE1还能调控种子萌发(Jiang et al., 2016).IMB1 (IMBIBITION-INDUCIBLE1)是染色质域蛋白家族成员, 在PHYA介导的种子萌发中发挥作用.IMB1在干种子中的表达量很低, 随着种子吸胀呈现上调表达, 暗示IMB1能够促进种子萌发(Duque and Chua, 2003).目前, 关于IMB1调控PHYA介导的种子萌发的分子机制还有待深入探究.CSN (COP9 SIGNALOSOME)蛋白复合体是一类保守的蛋白复合体, 能够调控RING型E3泛素化连接酶的活性.CSN包含8个亚基(CSN1-CSN8) (Wei and Deng, 2003).其中, CSN1和CSN5参与调控种子萌发.csn1-10和csn5a-1突变体呈现出种子延迟萌发的表型(Jin et al., 2018). ...
The ethylene receptors ETHYLENE RESPONSE 1 and ETHYLENE RESPONSE 2 have contrasting roles in seed germination of Arabidopsis during salt stress 1 2014
... 除ABA和GA之外, 种子的休眠和萌发还受到7类激素的调控, 包括生长素(auxin)、乙烯(ethylene, ET)、油菜素内酯(brassinosteroids, BRs)、水杨酸(salicylic acid, SA)、茉莉酸(jasmonic acid, JA)、细胞分裂素(cytokinins, CTKs)和独脚金内酯(strigolactones, SLs).这些激素均通过影响ABA或GA通路间接调控种子的休眠和萌发(Shu et al., 2016; Née et al., 2017a).生长素能够通过调控ABI3的转录水平影响种子的休眠和萌发.当内源生长素水平升高时, 生长素信号通路中的负调控因子AXR2/3 (AUXIN- RESISTANT 2/3)发生降解.此时AXR2/3对响应生长素信号的ARF10 (AUXIN RESPONSE FACTOR 10)和ARF16基因的转录抑制作用被解除, 而ARF10和ARF16能够激活ABI3的表达, 从而使种子保持休眠状态(Liu et al., 2013a).然而, ARF10和ARF16不能直接结合在ABI3的启动子上(Liu et al., 2013a), 因此有待深入研究以揭示完整的生长素信号传递链.ET能够通过ABA生物合成和信号转导通路调控种子萌发(Cheng et al., 2009; Linkies et al., 2009; Corbineau et al., 2014), 但具体机制还不清楚.在盐胁迫条件下, ET受体蛋白ETR1 (ETHYLENE RESPONSE 1)和ETR2可能通过不依赖ET信号的通路调控ABA生物合成, 进而影响种子萌发(Wilson et al., 2014).最近的研究发现, ETR1通过解除ERF12 (ETHYLENE RESPONSE FACTOR 12)/TPL (TOPLESS)模块对DOG1基因的转录抑制作用而促进DOG1的转录, 进而促进种子休眠(Li et al., 2019).BR通过依赖MFT (MOTHER OFFT AND TFL 1)的途径拮抗ABA信号转导通路, 促进种子萌发(Xi and Yu, 2010; Xi et al., 2010).进一步的研究发现, BR信号通路中的负调控因子——BIN2 (BRASSINOSTEROID INSENSITIVE 2), 能够磷酸化并稳定ABI5蛋白, 进而促进ABA信号转导.BR信号通过抑制BIN2-ABI5互作解除ABA对种子萌发的抑制作用(Hu and Yu, 2014).SA在调控种子萌发上发挥双重作用.在正常生长条件下, SA通过抑制GA诱导的α淀粉酶编码基因的表达抑制种子萌发(Xie et al., 2007); 而在高盐胁迫下, SA通过其它通路促进种子萌发(Lee et al., 2010).CTKs通过抑制ABI5的转录或促进ABI5蛋白的降解拮抗ABA效应, 从而促进种子萌发(Wang et al., 2011; Guan et al., 2014).外源施加JA能够延迟种子萌发(Nambara et al., 2010), 表明JA能够抑制种子萌发.有研究表明, JA通过抑制ABA合成相关基因的表达、促进ABA代谢相关基因的表达拮抗ABA, 从而促进种子萌发(Jacobsen et al., 2013).JAZ3 (JASMONATE-ZIM DOMAIN PROTEIN 3)通过与ABI5互作抑制ABI5的转录激活活性, 进而促进种子萌发(Ju et al., 2019).目前关于JA调控种子萌发功能上存在矛盾的原因还有待深入探究.SLs通过降低ABA/GA值促进种子萌发(Toh et al., 2012).此外, SLs信号通路组分也能参与调控种子萌发, 如SMAX1 (SUPPRESSOR OF MORE AXILLARY GROWTH2 1) (Stanga et al., 2013).但目前SLs调控种子萌发的具体机制还不清楚.综上, 生长素、ET、BR、CTKs、JA和SLs通过调控ABA生物合成或信号通路调控种子的休眠和萌发.然而生长素、ET、BR、CTKs、JA与GA之间的交互作用还有待深入研究. ...
MOTHER OF FT AND TFL1 regulates seed germination through a negative feedback loop modulating ABA signaling in Arabidopsis 1 2010
... 除ABA和GA之外, 种子的休眠和萌发还受到7类激素的调控, 包括生长素(auxin)、乙烯(ethylene, ET)、油菜素内酯(brassinosteroids, BRs)、水杨酸(salicylic acid, SA)、茉莉酸(jasmonic acid, JA)、细胞分裂素(cytokinins, CTKs)和独脚金内酯(strigolactones, SLs).这些激素均通过影响ABA或GA通路间接调控种子的休眠和萌发(Shu et al., 2016; Née et al., 2017a).生长素能够通过调控ABI3的转录水平影响种子的休眠和萌发.当内源生长素水平升高时, 生长素信号通路中的负调控因子AXR2/3 (AUXIN- RESISTANT 2/3)发生降解.此时AXR2/3对响应生长素信号的ARF10 (AUXIN RESPONSE FACTOR 10)和ARF16基因的转录抑制作用被解除, 而ARF10和ARF16能够激活ABI3的表达, 从而使种子保持休眠状态(Liu et al., 2013a).然而, ARF10和ARF16不能直接结合在ABI3的启动子上(Liu et al., 2013a), 因此有待深入研究以揭示完整的生长素信号传递链.ET能够通过ABA生物合成和信号转导通路调控种子萌发(Cheng et al., 2009; Linkies et al., 2009; Corbineau et al., 2014), 但具体机制还不清楚.在盐胁迫条件下, ET受体蛋白ETR1 (ETHYLENE RESPONSE 1)和ETR2可能通过不依赖ET信号的通路调控ABA生物合成, 进而影响种子萌发(Wilson et al., 2014).最近的研究发现, ETR1通过解除ERF12 (ETHYLENE RESPONSE FACTOR 12)/TPL (TOPLESS)模块对DOG1基因的转录抑制作用而促进DOG1的转录, 进而促进种子休眠(Li et al., 2019).BR通过依赖MFT (MOTHER OFFT AND TFL 1)的途径拮抗ABA信号转导通路, 促进种子萌发(Xi and Yu, 2010; Xi et al., 2010).进一步的研究发现, BR信号通路中的负调控因子——BIN2 (BRASSINOSTEROID INSENSITIVE 2), 能够磷酸化并稳定ABI5蛋白, 进而促进ABA信号转导.BR信号通过抑制BIN2-ABI5互作解除ABA对种子萌发的抑制作用(Hu and Yu, 2014).SA在调控种子萌发上发挥双重作用.在正常生长条件下, SA通过抑制GA诱导的α淀粉酶编码基因的表达抑制种子萌发(Xie et al., 2007); 而在高盐胁迫下, SA通过其它通路促进种子萌发(Lee et al., 2010).CTKs通过抑制ABI5的转录或促进ABI5蛋白的降解拮抗ABA效应, 从而促进种子萌发(Wang et al., 2011; Guan et al., 2014).外源施加JA能够延迟种子萌发(Nambara et al., 2010), 表明JA能够抑制种子萌发.有研究表明, JA通过抑制ABA合成相关基因的表达、促进ABA代谢相关基因的表达拮抗ABA, 从而促进种子萌发(Jacobsen et al., 2013).JAZ3 (JASMONATE-ZIM DOMAIN PROTEIN 3)通过与ABI5互作抑制ABI5的转录激活活性, 进而促进种子萌发(Ju et al., 2019).目前关于JA调控种子萌发功能上存在矛盾的原因还有待深入探究.SLs通过降低ABA/GA值促进种子萌发(Toh et al., 2012).此外, SLs信号通路组分也能参与调控种子萌发, 如SMAX1 (SUPPRESSOR OF MORE AXILLARY GROWTH2 1) (Stanga et al., 2013).但目前SLs调控种子萌发的具体机制还不清楚.综上, 生长素、ET、BR、CTKs、JA和SLs通过调控ABA生物合成或信号通路调控种子的休眠和萌发.然而生长素、ET、BR、CTKs、JA与GA之间的交互作用还有待深入研究. ...
Mother of FT and TFL1 regulates seed germination and fertility relevant to the brassinosteroid signaling pathway 1 2010
... 除ABA和GA之外, 种子的休眠和萌发还受到7类激素的调控, 包括生长素(auxin)、乙烯(ethylene, ET)、油菜素内酯(brassinosteroids, BRs)、水杨酸(salicylic acid, SA)、茉莉酸(jasmonic acid, JA)、细胞分裂素(cytokinins, CTKs)和独脚金内酯(strigolactones, SLs).这些激素均通过影响ABA或GA通路间接调控种子的休眠和萌发(Shu et al., 2016; Née et al., 2017a).生长素能够通过调控ABI3的转录水平影响种子的休眠和萌发.当内源生长素水平升高时, 生长素信号通路中的负调控因子AXR2/3 (AUXIN- RESISTANT 2/3)发生降解.此时AXR2/3对响应生长素信号的ARF10 (AUXIN RESPONSE FACTOR 10)和ARF16基因的转录抑制作用被解除, 而ARF10和ARF16能够激活ABI3的表达, 从而使种子保持休眠状态(Liu et al., 2013a).然而, ARF10和ARF16不能直接结合在ABI3的启动子上(Liu et al., 2013a), 因此有待深入研究以揭示完整的生长素信号传递链.ET能够通过ABA生物合成和信号转导通路调控种子萌发(Cheng et al., 2009; Linkies et al., 2009; Corbineau et al., 2014), 但具体机制还不清楚.在盐胁迫条件下, ET受体蛋白ETR1 (ETHYLENE RESPONSE 1)和ETR2可能通过不依赖ET信号的通路调控ABA生物合成, 进而影响种子萌发(Wilson et al., 2014).最近的研究发现, ETR1通过解除ERF12 (ETHYLENE RESPONSE FACTOR 12)/TPL (TOPLESS)模块对DOG1基因的转录抑制作用而促进DOG1的转录, 进而促进种子休眠(Li et al., 2019).BR通过依赖MFT (MOTHER OFFT AND TFL 1)的途径拮抗ABA信号转导通路, 促进种子萌发(Xi and Yu, 2010; Xi et al., 2010).进一步的研究发现, BR信号通路中的负调控因子——BIN2 (BRASSINOSTEROID INSENSITIVE 2), 能够磷酸化并稳定ABI5蛋白, 进而促进ABA信号转导.BR信号通过抑制BIN2-ABI5互作解除ABA对种子萌发的抑制作用(Hu and Yu, 2014).SA在调控种子萌发上发挥双重作用.在正常生长条件下, SA通过抑制GA诱导的α淀粉酶编码基因的表达抑制种子萌发(Xie et al., 2007); 而在高盐胁迫下, SA通过其它通路促进种子萌发(Lee et al., 2010).CTKs通过抑制ABI5的转录或促进ABI5蛋白的降解拮抗ABA效应, 从而促进种子萌发(Wang et al., 2011; Guan et al., 2014).外源施加JA能够延迟种子萌发(Nambara et al., 2010), 表明JA能够抑制种子萌发.有研究表明, JA通过抑制ABA合成相关基因的表达、促进ABA代谢相关基因的表达拮抗ABA, 从而促进种子萌发(Jacobsen et al., 2013).JAZ3 (JASMONATE-ZIM DOMAIN PROTEIN 3)通过与ABI5互作抑制ABI5的转录激活活性, 进而促进种子萌发(Ju et al., 2019).目前关于JA调控种子萌发功能上存在矛盾的原因还有待深入探究.SLs通过降低ABA/GA值促进种子萌发(Toh et al., 2012).此外, SLs信号通路组分也能参与调控种子萌发, 如SMAX1 (SUPPRESSOR OF MORE AXILLARY GROWTH2 1) (Stanga et al., 2013).但目前SLs调控种子萌发的具体机制还不清楚.综上, 生长素、ET、BR、CTKs、JA和SLs通过调控ABA生物合成或信号通路调控种子的休眠和萌发.然而生长素、ET、BR、CTKs、JA与GA之间的交互作用还有待深入研究. ...
REDUCED DORMANCY 5 encodes a protein phosphatase 2C that is required for seed dormancy in Arabidopsis 1 2014
... 激素在种子休眠和萌发过程中发挥重要调控作用.其中, ABA和GA在调控种子休眠和萌发上行使主要功能.ABA促进种子休眠, 而GA促进种子萌发(Shu et al., 2016; Née et al., 2017a).在种子成熟过程中, ABA在种子内逐渐积累, 使种子的休眠水平逐渐升高.相比之下, 当种子受到吸胀或层积处理时, ABA含量逐渐降低, GA含量逐渐升高, 使种子萌发.与野生型种子相比, ABA合成缺陷突变体种子萌发更快(Frey et al., 2012); 而过表达ABA合成基因的种子以及ABA代谢突变体种子保持较高的休眠水平(Matakiadis et al., 2009; Martinez-Andújar et al., 2011; Nonogaki et al., 2014), 表明ABA能够通过其生物合成通路调控种子的休眠和萌发.ABA信号通过PYR/PYL/RCAR (PYRABACTIN RESISTANCE 1/PYR-LIKE PROTEINS/REGULATORY COMPONENTS OF ABA RECEPTORS)-PP2C (PROTEIN PHOSPHATASE 2C)-SnRKs (SNF1-RELATED PROTEIN KINASES)级联反应向下传递(Cutler et al., 2010; Hubbard et al., 2010).目前鉴定到的PP2C类蛋白包括ABI1 (ABSCISIC ACID INSENSITIVE 1)、ABI2、HON (HONSU)和RDO5 (REDUCED DORMANCY 5).其中, ABI1和ABI2通过与ABA信号受体蛋白PYR/PYL/RCAR互作进而抑制ABA信号转导(Ma et al., 2009; Park et al., 2009).HON蛋白能够通过抑制ABA信号转导及促进GA信号转导抑制种子休眠(Kim et al., 2013), 说明HON能够整合ABA和GA信号以调控种子休眠.rdo5突变体的休眠水平降低, 但其ABA含量和ABA敏感性并未发生显著变化(Xiang et al., 2014), 说明RDO5通过不依赖ABA的途径调控种子休眠.ABI3、ABI4和ABI5是ABA信号通路下游的关键组分, 能够抑制种子萌发、促进种子休眠(Bentsink and Koornneef, 2008; Kanai et al., 2010; Shu et al., 2013).响应ABA信号的R2R3型MYB转录因子MYB96通过调控ABI4以及ABA合成相关基因NCED2 (9-CIS-EPOXYCAROTENOID DIOXYGENASE 2)和NCED6的转录进而促进种子休眠、抑制种子萌发(Lee et al., 2015a, 2015b).上述研究表明, ABA不仅通过其生物合成通路调控种子的休眠和萌发, 还能以ABA信号的形式发挥作用. ...
Salicylic acid inhibits gibberellin-induced alpha-amylase expression and seed germination via a pathway involving an abscisic-acid-inducible WRKY gene 1 2007
... 除ABA和GA之外, 种子的休眠和萌发还受到7类激素的调控, 包括生长素(auxin)、乙烯(ethylene, ET)、油菜素内酯(brassinosteroids, BRs)、水杨酸(salicylic acid, SA)、茉莉酸(jasmonic acid, JA)、细胞分裂素(cytokinins, CTKs)和独脚金内酯(strigolactones, SLs).这些激素均通过影响ABA或GA通路间接调控种子的休眠和萌发(Shu et al., 2016; Née et al., 2017a).生长素能够通过调控ABI3的转录水平影响种子的休眠和萌发.当内源生长素水平升高时, 生长素信号通路中的负调控因子AXR2/3 (AUXIN- RESISTANT 2/3)发生降解.此时AXR2/3对响应生长素信号的ARF10 (AUXIN RESPONSE FACTOR 10)和ARF16基因的转录抑制作用被解除, 而ARF10和ARF16能够激活ABI3的表达, 从而使种子保持休眠状态(Liu et al., 2013a).然而, ARF10和ARF16不能直接结合在ABI3的启动子上(Liu et al., 2013a), 因此有待深入研究以揭示完整的生长素信号传递链.ET能够通过ABA生物合成和信号转导通路调控种子萌发(Cheng et al., 2009; Linkies et al., 2009; Corbineau et al., 2014), 但具体机制还不清楚.在盐胁迫条件下, ET受体蛋白ETR1 (ETHYLENE RESPONSE 1)和ETR2可能通过不依赖ET信号的通路调控ABA生物合成, 进而影响种子萌发(Wilson et al., 2014).最近的研究发现, ETR1通过解除ERF12 (ETHYLENE RESPONSE FACTOR 12)/TPL (TOPLESS)模块对DOG1基因的转录抑制作用而促进DOG1的转录, 进而促进种子休眠(Li et al., 2019).BR通过依赖MFT (MOTHER OFFT AND TFL 1)的途径拮抗ABA信号转导通路, 促进种子萌发(Xi and Yu, 2010; Xi et al., 2010).进一步的研究发现, BR信号通路中的负调控因子——BIN2 (BRASSINOSTEROID INSENSITIVE 2), 能够磷酸化并稳定ABI5蛋白, 进而促进ABA信号转导.BR信号通过抑制BIN2-ABI5互作解除ABA对种子萌发的抑制作用(Hu and Yu, 2014).SA在调控种子萌发上发挥双重作用.在正常生长条件下, SA通过抑制GA诱导的α淀粉酶编码基因的表达抑制种子萌发(Xie et al., 2007); 而在高盐胁迫下, SA通过其它通路促进种子萌发(Lee et al., 2010).CTKs通过抑制ABI5的转录或促进ABI5蛋白的降解拮抗ABA效应, 从而促进种子萌发(Wang et al., 2011; Guan et al., 2014).外源施加JA能够延迟种子萌发(Nambara et al., 2010), 表明JA能够抑制种子萌发.有研究表明, JA通过抑制ABA合成相关基因的表达、促进ABA代谢相关基因的表达拮抗ABA, 从而促进种子萌发(Jacobsen et al., 2013).JAZ3 (JASMONATE-ZIM DOMAIN PROTEIN 3)通过与ABI5互作抑制ABI5的转录激活活性, 进而促进种子萌发(Ju et al., 2019).目前关于JA调控种子萌发功能上存在矛盾的原因还有待深入探究.SLs通过降低ABA/GA值促进种子萌发(Toh et al., 2012).此外, SLs信号通路组分也能参与调控种子萌发, 如SMAX1 (SUPPRESSOR OF MORE AXILLARY GROWTH2 1) (Stanga et al., 2013).但目前SLs调控种子萌发的具体机制还不清楚.综上, 生长素、ET、BR、CTKs、JA和SLs通过调控ABA生物合成或信号通路调控种子的休眠和萌发.然而生长素、ET、BR、CTKs、JA与GA之间的交互作用还有待深入研究. ...
Photoactivated CRY1 and phyB interact directly with AUX/IAA proteins to inhibit auxin signaling in Arabidopsis 1 2018
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
Contribution of gibberellin deactivation by AtGA2ox2 to the suppression of germination of dark-imbibed Arabidopsis thaliana seeds 1 2007
... GA能够通过抑制ABA诱导的种子休眠促进种子萌发(Gubler et al., 2005; Graeber et al., 2012).在植物体内, 具有生物活性的GA主要包括GA1和GA4, 它们通过软化种皮、促进胚乳层细胞的水解和胚的生长打破种子休眠, 进而促进种子萌发(Holdsworth et al., 2008).GA缺失突变体ga1 (gibberellic acid- requiring 1)和ga2表现出强烈的休眠表型, 需要外源施加GA方可萌发(Lee et al., 2002; Shu et al., 2013).GA20ox (GA 20-OXIDASE)和GA3ox基因编码参与GA生物合成的氧化酶; GA2ox编码的GA2-氧化酶主要参与GA的代谢过程.在PHYB-off条件下, ga2ox突变体萌发水平升高(Oh et al., 2006; Yamauchi et al., 2007).上述研究结果表明, GA能通过其生物合成通路调控种子的休眠和萌发.在GA信号转导过程中, DELLA蛋白扮演着重要角色, 它能响应GA信号快速降解, 对GA信号转导起限速作用(Silverstone et al., 1998; Itoh et al., 2002; Zentella et al., 2007; Nemoto et al., 2017).GA受体蛋白GID1 (GA-INSENSITIVE DWARF 1)能够负调控DELLA蛋白的稳定性, 促进GA信号转导(Davière and Achard, 2013).拟南芥SLY1 (SLEEPY1)和水稻GID2 (GA-INSENSITIVE DWARF2)是SCF聚合体中的F-box亚基, 依赖其C端的GGF和LSL基序与DELLA蛋白C端VHIID和LHR2基序结合, 促进DELLA多聚泛素化(Hirano et al., 2010; Ariizumi et al., 2011).研究表明, DELLA蛋白GAI (GA-INSENSITIVE)能够通过抑制TCP14 (TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR 14)和TCP15的表达减缓细胞周期的进程, 最终使种子保持休眠状态(Resentini et al., 2015).作为转录调节子, DELLA如何实现对下游基因的转录调控? 有研究指出, DELLA能够通过与染色质重塑因子互作调控下游基因的转录.DELLA蛋白RGL2 (RGA-LIKE 2)和RGL3与染色质重塑因子SWI3C (SWITCH3C)发生互作, 进而影响GID1a以及GA3ox基因的转录(Sarnowska et al., 2013).此外, DELLA还能通过与种子萌发相关转录因子互作进而实现其对萌发的调控.RGL2通过与转录因子NF-YC互作, 诱导下游ABI5的转录, 进而抑制种子萌发(Liu et al., 2016).此外, RGL2还能通过与转录因子DOF6 (BINDING1 ZINC FINGER 6)互作诱导GATA12的表达, 进而促进种子休眠(Ravindran et al., 2017).DELLA通过与ICE1 (INDUCER OF CBF EXPRESSION 1)互作解除其对ABA响应基因EM1 (LATE EMBRYOGENESIS ABUNDANT 1)和EM6的转录抑制作用, 最终抑制种子萌发(Hu et al., 2019). ...
Phytochrome a negatively regulates the shade avoidance response by increasing auxin/indole acidic acid protein stability 1 2018a
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
UVR8 interacts with WRKY36 to regulate HY5 transcription and hypocotyl elongation in Arabidopsis 1 2018b
... 植物依赖光受体蛋白识别外源环境中的光信号.根据吸收光谱成分的不同可以将植物光受体分为3类: 吸收红光/远红光(600-750 nm)的光敏色素(PHY) (Quail et al., 1995)、吸收蓝光/UV-A (320-500 nm)的向光素(PHOTOTROPIN, PHOT)、隐花色素(CRYPTOCHROME, CRY)和ZTL (ZEITLUPE)/FKF1 (FLAVIN BINDING KELCH REPEAT F-BOX 1)/ LKP2 (LOV KELCH PROTEIN 2)基因家族(Briggs and Huala, 1999; Cashmore, 2003; Lin and Shalitin, 2003)以及吸收UV-B (280-320 nm)的UVR8 (UV RESISTANCE LOCUS 8) (Rizzini et al., 2011).近年来, 人们在PHY、CRY以及UVR8介导的光信号转导通路研究中取得了重要进展.其中, PHY和CRY均能通过与转录因子互作进而直接调控下游基因的转录.此类信号通路主要包括: PHYB-PIFs通路、CRY-PIF4/5通路、CRY2-CIBs (CRYPTOCHROME- INTERACTING BASIC-HELIX-LOOP-HELIX)信号通路、PHYA-AUX/IAA (AUXIN/INDOLE-3-ACETIC ACID)信号通路以及PHYB/CRY1-AUX/IAA信号通路.PHYB通过与PIFs互作促使PIFs发生泛素化降解, 进而调控PIFs下游基因的转录(Oh et al., 2006); PHYA/B和CRY1/2通过与PIFs互作进而影响PIFs对靶基因的转录调控(Chen et al., 2014; Pedmale et al., 2016; Ma et al., 2016); CRY2通过与CIBs蛋白互作提高CIBs对FT基因的激活水平(Liu et al., 2008a, 2013b; 马朝峰和戴思兰, 2019); PHYA通过与生长素信号转导通路的负调控因子AUX/IAA互作稳定其蛋白活性, 进而调控植物的避荫反应(Yang et al., 2018a); PHYB和CRY1分别介导红光和蓝光, 通过抑制AUX/IAA的泛素化降解抑制生长素信号转导(Xu et al., 2018).此外, PHY和CRY亦可通过与COP1 (CONSTITUTIVE PHOTOMORPHO-GENESIS 1)互作抑制其E3泛素连接酶活性, 促进COP1靶蛋白的积累, 从而间接调控基因转录(Hardtke et al., 2000; Seo et al., 2003; Jang et al., 2005, 2015; Liu et al., 2008b; Luo et al., 2014).在UVR8介导的信号通路中, COP1作为正调控因子发挥作用.UVR8通过与COP1互作促进下游HY5 (LONG HYPOCOTYL 5)蛋白的积累, 进而诱导光响应基因的转录(Huang et al., 2013; 景艳军和林荣呈, 2017).最近的研究发现, RUP1 (REPRESSOR OF UV-B PHOTOMORPHO-GENESIS 1)和RUP2作为UVR8介导的信号转导通路中的负调控因子, 能够促进HY5蛋白降解; 而COP1能够与RUP1/RUP2互作介导其泛素化降解.RUP1/RUP2-HY5以及COP1-RUP1/RUP2构成植物响应UVB信号的分子开关(Ren et al., 2019).此外, UVR8通过与BES1 (BRI1-EMS-SUPPRESSOR 1)/ BIM1 (BES1-INTERACTING MYC-LIKE 1)蛋白互作抑制BES1/BIM1对下游BR响应基因的转录激活活性(Liang et al., 2018).UVR8通过与WRKY36 (WRKY DNA-BINDING PROTEIN 36)互作进而解除WRKY36对HY5的转录抑制作用, 最终促进HY5转录和植物光形态建成(Yang et al., 2018b). ...
Global analysis of DELLA direct targets in early gibberellin signaling in Arabidopsis 1 2007
... GA能够通过抑制ABA诱导的种子休眠促进种子萌发(Gubler et al., 2005; Graeber et al., 2012).在植物体内, 具有生物活性的GA主要包括GA1和GA4, 它们通过软化种皮、促进胚乳层细胞的水解和胚的生长打破种子休眠, 进而促进种子萌发(Holdsworth et al., 2008).GA缺失突变体ga1 (gibberellic acid- requiring 1)和ga2表现出强烈的休眠表型, 需要外源施加GA方可萌发(Lee et al., 2002; Shu et al., 2013).GA20ox (GA 20-OXIDASE)和GA3ox基因编码参与GA生物合成的氧化酶; GA2ox编码的GA2-氧化酶主要参与GA的代谢过程.在PHYB-off条件下, ga2ox突变体萌发水平升高(Oh et al., 2006; Yamauchi et al., 2007).上述研究结果表明, GA能通过其生物合成通路调控种子的休眠和萌发.在GA信号转导过程中, DELLA蛋白扮演着重要角色, 它能响应GA信号快速降解, 对GA信号转导起限速作用(Silverstone et al., 1998; Itoh et al., 2002; Zentella et al., 2007; Nemoto et al., 2017).GA受体蛋白GID1 (GA-INSENSITIVE DWARF 1)能够负调控DELLA蛋白的稳定性, 促进GA信号转导(Davière and Achard, 2013).拟南芥SLY1 (SLEEPY1)和水稻GID2 (GA-INSENSITIVE DWARF2)是SCF聚合体中的F-box亚基, 依赖其C端的GGF和LSL基序与DELLA蛋白C端VHIID和LHR2基序结合, 促进DELLA多聚泛素化(Hirano et al., 2010; Ariizumi et al., 2011).研究表明, DELLA蛋白GAI (GA-INSENSITIVE)能够通过抑制TCP14 (TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR 14)和TCP15的表达减缓细胞周期的进程, 最终使种子保持休眠状态(Resentini et al., 2015).作为转录调节子, DELLA如何实现对下游基因的转录调控? 有研究指出, DELLA能够通过与染色质重塑因子互作调控下游基因的转录.DELLA蛋白RGL2 (RGA-LIKE 2)和RGL3与染色质重塑因子SWI3C (SWITCH3C)发生互作, 进而影响GID1a以及GA3ox基因的转录(Sarnowska et al., 2013).此外, DELLA还能通过与种子萌发相关转录因子互作进而实现其对萌发的调控.RGL2通过与转录因子NF-YC互作, 诱导下游ABI5的转录, 进而抑制种子萌发(Liu et al., 2016).此外, RGL2还能通过与转录因子DOF6 (BINDING1 ZINC FINGER 6)互作诱导GATA12的表达, 进而促进种子休眠(Ravindran et al., 2017).DELLA通过与ICE1 (INDUCER OF CBF EXPRESSION 1)互作解除其对ABA响应基因EM1 (LATE EMBRYOGENESIS ABUNDANT 1)和EM6的转录抑制作用, 最终抑制种子萌发(Hu et al., 2019). ...
 ,*中国科学院植物研究所光生物学重点实验室, 北京 100093
,*中国科学院植物研究所光生物学重点实验室, 北京 100093
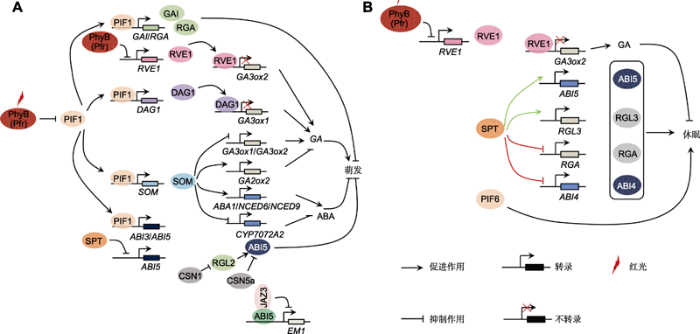 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}