Abstract Increasing plant density is an important approach to boost crop yield, and leaf angle is one of the key factors affecting plant density. Recently, Feng Tian’s lab from China Agricultural University cloned and characterized two major QTLs (UPA1 and UPA2) regulating leaf angle in maize. The underlying genes are brd1 and ZmRAVL1, respectively, and both of them are involved in the brassinosteroid (BR) pathway to regulate leaf angle. UPA2 is located 9.5 kb upstream of ZmRAVL1 and is bound by DRL1. LG1, another leaf angle protein, directly activates the expression of ZmRAVL1. DRL1 and LG1 physically interact and the resulting complex in turn represses the LG1-activated expression of ZmRAVL1. The teosinte allele of UPA2 has a higher binding affinity with DRL1, resulting in the reduced ZmRAVL1 expression, which consequently down-regulates the brd1 expression and leads to the decreased brassinosteroid level, thereby reducing the leaf angle. The introgression of UPA2 teosinte allele into maize and the manipulation of ZmRAVL1 significantly increase maize yield with increased plant density. These findings have paved a new avenue for molecular breeding of high-yield maize varieties. Keywords:maize;plant density;yield;UPA1; UPA2
PDF (638KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 刘杰, 严建兵. 大刍草稀有等位基因促进玉米密植高产. 植物学报, 2019, 54(5): 554-557 doi:10.11983/CBB19119 Liu Jie, Yan Jianbing. A Teosinte Rare Allele Increases Maize Plant Density and Yield. Chinese Bulletin of Botany, 2019, 54(5): 554-557 doi:10.11983/CBB19119
玉米(Zea mays)种植密度的增加是玉米增产的重要途径之一。美国玉米的种植密度由20世纪30年代的35 000株∙hm-2提高到了21世纪初的80 000株∙hm-2 (Lee and Tollenaar, 2007); 中国玉米的种植密度在最近10年也有显著增加, 以黄淮海夏玉米主产区为例, 由2005年的58 200株∙hm-2增加到2016年的62 100株∙hm-2 (明博等, 2017)。叶夹角是影响玉米种植密度的重要因素, 已有多项研究克隆了控制玉米叶夹角的基因, 包括lg1 (liguleless1) (Moreno et al., 1997)、lg2 (Walsh et al., 1998)、ZmTAC1 (Ku et al., 2011)、ZmCLA4 (Zhang et al., 2014)、na2 (nana plant2) (Best et al., 2016)、drl1 (drooping leaf 1)和drl2 (Strable et al., 2017)。lg1编码含SBP结构域的蛋白, 该基因突变体的叶舌和叶耳缺失、叶片直立; lg2编码bZIP转录因子; ZmTAC1是水稻(Oryza sativa)中控制叶夹角基因OsTAC1 (Yu et al., 2007)的同源基因; ZmCLA4是水稻中OsLAZY1 (Li et al., 2007)的同源基因, 通过影响向重力性调控叶夹角; na2编码油菜素内酯(brassinosteroid, BR)生物合成途径的关键酶, 是拟南芥(Arabidopsis thaliana)中DWARF1 (Choe et al., 1999)的同源基因; drl1和drl2均编码YABBY家族转录因子, 且drl2可以增强drl1的作用。
lg1和lg2双突变体的遗传分析表明, 这2个基因位于同一条叶发育途径, 但是发挥不同功能, 且lg2发挥作用早于lg1 (Harper and Freeling, 1996)。na2调控BR合成, 而BR则可能调控drl1和drl2的活性(Strable et al., 2017), 表明这3个基因可能通过BR相关途径来调控叶夹角。上述研究结果表明, BR信号途径在调控玉米叶夹角中有非常重要的作用, 但这3个基因参与BR途径的具体方式以及它们之间是如何互作的仍然不清楚。
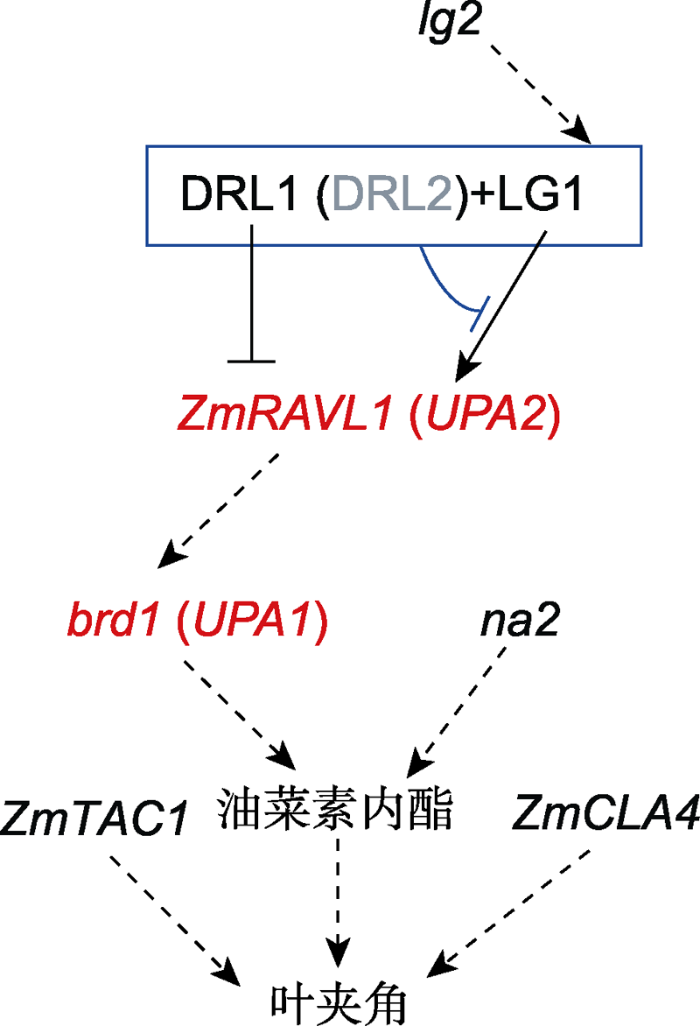
lg2调控lg1, 但具体机制未知。LG1激活ZmRAVL1的表达, DRL1抑制ZmRAVL1的表达, DRL1与LG1蛋白互作抑制LG1对ZmRAVL1的激活作用(DRL2可能具有与DRL1类似的功能)。ZmRAVL1调控下游brd1的表达, brd1和nana plant2 (na2)都参与油菜素内酯(brassinosteroid, BR)的生物合成。这些基因均通过BR途径调控玉米叶夹角形成。实线表示机制已被阐明, 虚线表示机制未知。 Figure 1A proposed pathway regulating the leaf angle in maize
lg2 regulates lg1 with an unknown mechanism. LG1 and DRL1 activates and represses the expression of ZmRAVL1, respectively. The DRL1-LG1 complex represses the LG1- activated ZmRAVL1 expression (DRL2 may have a similar function as DRL1). ZmRAVL1 regulates the expression of brd1, which, together with nana plant2 (na2), are involved in the biosynthesis of brassinosteroid (BR) and eventually regulate leaf angle. Solid and dash lines indicate the clear and unclear regulatory mechanism, respectively.
BestNB, HartwigT, BudkaJ, FujiokaS, JohalG, SchulzB, DilkesBP (2016). nana plant2 encodes a maize ortholog of the Arabidopsis brassinosteroid biosynthesis gene DWARF1, identifying developmental interactions between brassinosteroids and gibberellins Plant Physiol 171, 2633-2647. [本文引用: 1]
ChoeS, DilkesBP, GregoryBD, RossAS, YuanH, NoguchiT, FujiokaS, TakatsutoS, TanakaA, YoshidaS, TaxFE, FeldmannKA (1999). The Arabidopsis dwarf1 mutant is defective in the conversion of 24-methylenecholesterol to campesterol in brassinosteroid biosynthesis Plant Physiol 119, 897-907. [本文引用: 1]
HarperL, FreelingM (1996). Interactions of liguleless1 and liguleless2 function during ligule induction in maize Genetics 144, 1871-1882. [本文引用: 1]
JeBI, PiaoHL, ParkSJ, ParkSH, KimCM, XuanYH, ParkSH, HuangJ, Do ChoiY, AnG, WongHL, FujiokaS, KimMC, ShimamotoK, HanCD (2010). RAV-Like1 maintains brassinosteroid homeostasis via the coordinated activation of BRI1 and biosynthetic genes in rice Plant Cell 22, 1777-1791. [本文引用: 1]
KuL, WeiX, ZhangS, ZhangJ, GuoS, ChenY (2011). Cloning and characterization of a putative TAC1 ortholog associated with leaf angle in maize(Zea mays L.) PLoS One 6, e20621. [本文引用: 1]
LeeEA, TollenaarM (2007). Physiological basis of successful breeding strategies for maize grain yield Crop Sci 47, S202-S215. [本文引用: 1]
LiP, WangY, QianQ, FuZ, WangM, ZengD, LiB, WangX, LiJ (2007). LAZY1 controls rice shoot gravitropism through regulating polar auxin transport Cell Res 17, 402-410. [本文引用: 1]
MorenoMA, HarperLC, KruegerRW, DellaportaSL, FreelingM (1997). liguleless1 encodes a nuclear-localized protein required for induction of ligules and auricles during maize leaf organogenesis Genes Dev 11, 616-628. [本文引用: 1]
WalshJ, WatersCA, FreelingM (1998). The maize gene liguleless2 encodes a basic leucine zipper protein involved in the establishment of the leaf blade-sheath boundary Genes Dev 12, 208-218. [本文引用: 1]
YuB, LinZ, LiH, LiX, LiJ, WangY, ZhangX, ZhuZ, ZhaiW, WangX, XieD, SunC (2007). TAC1, a major quantitative trait locus controlling tiller angle in rice Plant J 52, 891-898. [本文引用: 1]
ZhangJ, KuLX, HanZP, GuoSL, LiuHJ, ZhangZZ, CaoLR, CuiXJ, ChenYH (2014). The ZmCLA4 gene in the qLA4-1 QTL controls leaf angle in maize( Zea mays L.) J Exp Bot 65, 5063-5076. [本文引用: 1]
2005-2016年中国玉米种植密度变化分析 1 2017
... 玉米(Zea mays)种植密度的增加是玉米增产的重要途径之一.美国玉米的种植密度由20世纪30年代的35 000株∙hm-2提高到了21世纪初的80 000株∙hm-2 (Lee and Tollenaar, 2007); 中国玉米的种植密度在最近10年也有显著增加, 以黄淮海夏玉米主产区为例, 由2005年的58 200株∙hm-2增加到2016年的62 100株∙hm-2 (明博等, 2017).叶夹角是影响玉米种植密度的重要因素, 已有多项研究克隆了控制玉米叶夹角的基因, 包括lg1 (liguleless1) (Moreno et al., 1997)、lg2 (Walsh et al., 1998)、ZmTAC1 (Ku et al., 2011)、ZmCLA4 (Zhang et al., 2014)、na2 (nana plant2) (Best et al., 2016)、drl1 (drooping leaf 1)和drl2 (Strable et al., 2017).lg1编码含SBP结构域的蛋白, 该基因突变体的叶舌和叶耳缺失、叶片直立; lg2编码bZIP转录因子; ZmTAC1是水稻(Oryza sativa)中控制叶夹角基因OsTAC1 (Yu et al., 2007)的同源基因; ZmCLA4是水稻中OsLAZY1 (Li et al., 2007)的同源基因, 通过影响向重力性调控叶夹角; na2编码油菜素内酯(brassinosteroid, BR)生物合成途径的关键酶, 是拟南芥(Arabidopsis thaliana)中DWARF1 (Choe et al., 1999)的同源基因; drl1和drl2均编码YABBY家族转录因子, 且drl2可以增强drl1的作用. ...
nana plant2 encodes a maize ortholog of the Arabidopsis brassinosteroid biosynthesis gene DWARF1, identifying developmental interactions between brassinosteroids and gibberellins 1 2016
... 玉米(Zea mays)种植密度的增加是玉米增产的重要途径之一.美国玉米的种植密度由20世纪30年代的35 000株∙hm-2提高到了21世纪初的80 000株∙hm-2 (Lee and Tollenaar, 2007); 中国玉米的种植密度在最近10年也有显著增加, 以黄淮海夏玉米主产区为例, 由2005年的58 200株∙hm-2增加到2016年的62 100株∙hm-2 (明博等, 2017).叶夹角是影响玉米种植密度的重要因素, 已有多项研究克隆了控制玉米叶夹角的基因, 包括lg1 (liguleless1) (Moreno et al., 1997)、lg2 (Walsh et al., 1998)、ZmTAC1 (Ku et al., 2011)、ZmCLA4 (Zhang et al., 2014)、na2 (nana plant2) (Best et al., 2016)、drl1 (drooping leaf 1)和drl2 (Strable et al., 2017).lg1编码含SBP结构域的蛋白, 该基因突变体的叶舌和叶耳缺失、叶片直立; lg2编码bZIP转录因子; ZmTAC1是水稻(Oryza sativa)中控制叶夹角基因OsTAC1 (Yu et al., 2007)的同源基因; ZmCLA4是水稻中OsLAZY1 (Li et al., 2007)的同源基因, 通过影响向重力性调控叶夹角; na2编码油菜素内酯(brassinosteroid, BR)生物合成途径的关键酶, 是拟南芥(Arabidopsis thaliana)中DWARF1 (Choe et al., 1999)的同源基因; drl1和drl2均编码YABBY家族转录因子, 且drl2可以增强drl1的作用. ...
The Arabidopsis dwarf1 mutant is defective in the conversion of 24-methylenecholesterol to campesterol in brassinosteroid biosynthesis 1 1999
... 玉米(Zea mays)种植密度的增加是玉米增产的重要途径之一.美国玉米的种植密度由20世纪30年代的35 000株∙hm-2提高到了21世纪初的80 000株∙hm-2 (Lee and Tollenaar, 2007); 中国玉米的种植密度在最近10年也有显著增加, 以黄淮海夏玉米主产区为例, 由2005年的58 200株∙hm-2增加到2016年的62 100株∙hm-2 (明博等, 2017).叶夹角是影响玉米种植密度的重要因素, 已有多项研究克隆了控制玉米叶夹角的基因, 包括lg1 (liguleless1) (Moreno et al., 1997)、lg2 (Walsh et al., 1998)、ZmTAC1 (Ku et al., 2011)、ZmCLA4 (Zhang et al., 2014)、na2 (nana plant2) (Best et al., 2016)、drl1 (drooping leaf 1)和drl2 (Strable et al., 2017).lg1编码含SBP结构域的蛋白, 该基因突变体的叶舌和叶耳缺失、叶片直立; lg2编码bZIP转录因子; ZmTAC1是水稻(Oryza sativa)中控制叶夹角基因OsTAC1 (Yu et al., 2007)的同源基因; ZmCLA4是水稻中OsLAZY1 (Li et al., 2007)的同源基因, 通过影响向重力性调控叶夹角; na2编码油菜素内酯(brassinosteroid, BR)生物合成途径的关键酶, 是拟南芥(Arabidopsis thaliana)中DWARF1 (Choe et al., 1999)的同源基因; drl1和drl2均编码YABBY家族转录因子, 且drl2可以增强drl1的作用. ...
Interactions of liguleless1 and liguleless2 function during ligule induction in maize 1 1996
... lg1和lg2双突变体的遗传分析表明, 这2个基因位于同一条叶发育途径, 但是发挥不同功能, 且lg2发挥作用早于lg1 (Harper and Freeling, 1996).na2调控BR合成, 而BR则可能调控drl1和drl2的活性(Strable et al., 2017), 表明这3个基因可能通过BR相关途径来调控叶夹角.上述研究结果表明, BR信号途径在调控玉米叶夹角中有非常重要的作用, 但这3个基因参与BR途径的具体方式以及它们之间是如何互作的仍然不清楚. ...
RAV-Like1 maintains brassinosteroid homeostasis via the coordinated activation of BRI1 and biosynthetic genes in rice 1 2010
 ,*华中农业大学, 作物遗传改良国家重点实验室, 武汉 430070
,*华中农业大学, 作物遗传改良国家重点实验室, 武汉 430070
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}