Advances in Biofunctions of the ABCB Subfamily in Plants
Zhenmei He1, Dongming Li2, Yanhua Qi,1,2,*1 State Key Laboratory of Plant Physiology and Biochemistry, Institute of Plant Biology, College of Life Sciences, Zhejiang University, Hangzhou 310058, China 2 Key Laboratory of Herbage & Endemic Crop Biotechnology, Ministry of Education, School of Life Sciences, Inner Mongolia University, Huhhot 010000, China;
Abstract The superfamily of ABC (ATP-binding cassette) transporters, which contains eight subfamilies from ABCA to ABCH, has diverse structures and complex functions. ABCB transporters, are mostly located in the plasma membrane, while others are located in the mitochondrial membrane or chloroplast membrane. ABCB transporters, together with AUX1/LAX (AUXIN1/LIKE AUXIN) and PIN (PIN-FORMED), coordinate and participate in the polar transport of auxin, and play an important role in regulating plant growth and development. ABCB transporters also function in plant tropism and resistance to heavy metals. In recent years, with the completion of whole-genome sequencing in different plants, research on ABCB genes is no longer confined to the model plant Arabidopsis thaliana, rather, preliminary studies have been carried out to explore the functions of ABCB genes in cereal including rice, maize, and sorghum. However, the functions for most of the plant ABCB transporters remain elusive. Here we reviewed the research progress and future development of ABCB subfamily transporters in Arabidopsis and cereal, in the hope of providing clues for fully revealing biofunctions of the ABCB subfamily. Keywords:ABCB transporter;cereal crops; Arabidopsis thaliana;auxin;biofunctions
PDF (2669KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 贺祯媚, 李东明, 齐艳华. 植物ABCB亚家族生物学功能研究进展. 植物学报, 2019, 54(6): 688-698 doi:10.11983/CBB19140 He Zhenmei, Li Dongming, Qi Yanhua. Advances in Biofunctions of the ABCB Subfamily in Plants. Chinese Bulletin of Botany, 2019, 54(6): 688-698 doi:10.11983/CBB19140
ABC (ATP-binding cassette)蛋白超家族是现阶段已知最大的蛋白质家族之一, 包括ABCA、ABCB和ABCC等8个亚家族。早期ABC转运蛋白在动物的耐药性研究中取得较大进展(Theodoulou and Kerr, 2015)。因此, 植物中ABC转运蛋白的初步研究也主要集中于外源物质及次生代谢物的转运在解毒和病原体防御中的作用方面(Hwang et al., 2016)。随着研究的不断深入, 研究者发现植物ABC转运蛋白在生长素和重金属转运以及植物抗病性等方面同样发挥重要作用(Theodoulou and Kerr, 2015), 其亚家族ABCB转运蛋白在植物生长发育特别是生长素转运中的作用更显著。
自1992年从拟南芥(Arabidopsis thaliana)中克隆到第1个ABCB亚家族的基因——AtPGP1 (AtABCB1)以来, 植物中的ABCB亚家族研究备受关注(Dudler and Hertig, 1992)。近30年的研究发现, ABCB作为转运蛋白与AUX1/LAX (AUXIN1/LIKE AUXIN)和PIN (PIN-FORMED)协同调控植物生长素的极性运输。例如, AtABCB1、AtABCB19、AtABCB4、AtABCB14和AtABCB21 (Sidler et al.,1998; Noh et al., 2001; Geisler et al., 2005; Santelia et al., 2005; Kaneda et al., 2011; Kamimoto et al., 2012)等相继被报道与生长素的运输相关。此外, 也有研究表明, ABCB转运蛋白参与植物对重金属和铝毒的抗性等过程。例如, Kim等(2016)报道了ABCB半分子转运蛋白ATM3 (AtABCB24)可提高拟南芥对镉和铅的抗性。而ALS1/TAP2 (AtABCB27)被报道参与拟南芥的铝转运, 进而提高其铝毒抗性(Larsen et al., 2007)。单子叶植物玉米(Zea mays)、高粱(Sorghum bicolor)和水稻(Oryza sativa)中的ABCB蛋白也具有生长素转运功能。玉米ZmABCB1 (Brachytic2, BR2)和高粱SbABCB1 (Dwarf1, DW3)参与生长素的向基运输(Multani et al., 2003); 水稻OsABCB14则参与生长素的向顶运输(Xu et al., 2014)。单子叶植物的转录组数据分析显示, ABCB亚家族蛋白可能与植物的非生物胁迫响应密切相关(Shen et al., 2010; Pang et al., 2013; Chai and Subudhi, 2016)。可见, ABCB转运蛋白的功能在植物中具有一定的保守性。植物ABCB转运蛋白的数量较多, 然而目前的研究主要集中在双子叶模式植物拟南芥中, 单子叶植物中的研究尚少, 大部分ABCB转运蛋白的功能有待进一步探索。本文主要综述了ABCB亚家族蛋白在双子叶植物拟南芥以及单子叶植物玉米、高粱、小麦(Triticum aestivum)和水稻中的研究进展, 旨在为ABCB基因功能的揭示提供新线索。
1 ABCB转运蛋白的结构及命名
ABC蛋白由于结构中包含有核酸结合域(nucleotide binding domain, NBD), 故又被称作ATP结合盒式蛋白。NBD含有(ABC特征基序, Walker A和B序列, H环和Q环)几个高度保守的基序(Sánchez-Fernández et al., 2001; Higgins and Linton, 2004)。除了NBD外, ABC转运蛋白的结构中还包含有跨膜结构域(trans membrane domain, TMD), 每个跨膜结构域均由几个疏水性α-螺旋组成。当ABCB转运蛋白转运底物时, NBD负责结合并水解ATP, 为底物的运输提供动力; TMD则负责底物的识别及参与底物的跨膜转运。根据蛋白的结构, ABC转运蛋白可以分为3类, 即全分子转运蛋白、半分子转运蛋白以及可溶性转运蛋白(Sánchez-Fernández et al., 2001; Verrier et al., 2008)。其中, ABCB亚家族蛋白只存在前两种结构(全分子和半分子转运蛋白)。全分子转运蛋白包含2个NBD和2个TMD; 半分子转运蛋白则只包括1个NBD和TMD (图1)。
(A) TWD1在ABCB从内质网膜到质膜的运输过程中发挥分子伴侣作用; (B) AGC激酶通过磷酸化ABCB蛋白, 使其转运活性下降; (C) PIN与ABCB蛋白互作。 Figure 2Regulation of ABCB transporter activity in Arabidopsis thaliana
(A) TWD1 acts as a molecular chaperone during the transport of ABCB from the endoplasmic reticulum membrane to the plasma membrane; (B) AGC kinase phosphorylates ABCB protein to reduce its transport activity; (C) PIN interacts with ABCB.
AGC蛋白激酶可调节ABCB转运蛋白的活性。在HeLa细胞表达系统中, 蛋白激酶AGC4/Phot1能与AtABCB19蛋白的NBD区结合使其磷酸化, 进而降低其活性(Christie et al., 2011) (图2)。Co-IP和BRET实验证明, AGC3/PID与TWD1存在互作, PID通过与TWD1互作直接磷酸化AtABCB1, 从而使其生长素转运活性受抑制(Henrichs et al., 2012)。然而, PID磷酸化ABCB是否必须TWD1介导目前尚不清楚。现有的研究表明AGC激酶可磷酸化ABCB, 降低其与TWD1间的互作, 从而改变ABCB蛋白跨膜转运过程中TWD1的分子伴侣作用, 影响其活性。
PIN与ABCB蛋白存在互作。植物生长素极性运输主要依赖ABCB、PIN以及AUX1/LAX转运蛋白家族。目前, 对PIN蛋白家族的研究最为深入。PIN的一个重要特征是亚细胞极性定位。Blakeslee等(2007)研究表明, PIN在极性胚胎发育和器官形成等需要定向生长素极性运输的过程中起决定作用, ABCB则在顶端组织生长素转运及长距离运输中起重要作用, 二者在调控生长素的转运上具有一定的独立性。另有研究发现, ABCB1和ABCB19在拟南芥的茎尖中与PIN1共定位, 在根中与PIN1和PIN2共定位(Bandyopadhyay et al., 2007)。酵母双杂交和免疫共沉淀实验均表明, ABCB1、ABCB19与PIN1、PIN2间存在物理互作(Blakeslee et al., 2007; Kim et al., 2010; Geisler et al., 2017); 且共表达PIN1-ABCB1和PIN1-ABCB19的植株均表现出生长素输出率及底物特异性提高(Blakeslee et al., 2007) (图2)。尽管Geisler等(2017)提出PIN与ABCB互作的3种假设: (1) PIN发挥运输作用, ABCB发挥调节作用; (2) ABCB发挥运输作用, PIN发挥调节作用; (3) PIN和ABCB不单独发挥转运作用, 而是形成复合体行使转运功能, 但具体机制仍不清楚, 尚需进一步探索。
CecchettiV, BrunettiP, NapoliN, FattoriniL, AltamuraMM, CostantinoP, CardarelliM ( 2015). ABCB1 and ABCB19 auxin transporters have synergistic effects on early and late Arabidopsis anther development J Integr Plant Biol 57, 1089-1098. [本文引用: 1]
ChaiCL, SubudhiPK ( 2016). Comprehensive analysis and expression profiling of the OsLAX and OsABCB auxin transporter gene families in rice(Oryza sativa) under phytohormone stimuli and abiotic stresses Front Plant Sci 7, 593. [本文引用: 1]
ChenSX, Sánchez-FernándezR, LyverER, DancisA, ReaPA ( 2007). Functional characterization of AtATM1, AtATM2, and AtATM3, a subfamily of Arabidopsis half- molecule ATP-binding cassette transporters implicated in iron homeostasis J Biol Chem 282, 21561-21571. [本文引用: 4]
ChristieJM, YangHB, RichterGL, SullivanS, ThomsonCE, LinJS, TitapiwatanakunB, EnnisM, KaiserliE, LeeOR, AdamecJ, PeerWA, MurphyAS ( 2011). phot1 inhibition of ABCB19 primes lateral auxin fluxes in the shoot apex required for phototropism PLoS Biol 9, e100 1076. [本文引用: 4]
DeanM, RzhetskyA, AllikmetsR ( 2001). The human ATP-binding cassette (ABC) transporter superfamily Genome Res 11, 1156-1166. [本文引用: 1]
DudlerR, HertigC ( 1992). Structure of an mdr-like gene from Arabidopsis thaliana. Evolutionary implications J Biol Chem 267, 5882-5888. [本文引用: 2]
FerroM, BrugièreS, SalviD, Seigneurin-BernyD, CourtM, MoyetL, RamusC, MirasS, MellalM, Le GallS, Kieffer-JaquinodS, BruleyC, GarinJ, JoyardJ, MasselonC, RollandN ( 2010). AT_CHLORO, a comprehensive chloroplast proteome database with subplastidial localization and curated information on envelope proteins Mol Cell Proteomics 9, 1063-1084. [本文引用: 2]
GeislerM, AryalB, di DonatoM, HaoPC ( 2017). A critical view on ABC transporters and their interacting partners in auxin transport Plant Cell Physiol 58, 1601-1614. [本文引用: 2]
GeislerM, BaillyA, IvanchenkoM ( 2016). Master and servant: regulation of auxin transporters by FKBPs and cyclophilins Plant Sci 245, 1-10. [本文引用: 1]
GranzinJ, EckhoffA, WeiergraberOH ( 2006). Crystal structure of a multi-domain immunophilin from Arabidopsis thaliana: a paradigm for regulation of plant ABC transporters J Mol Biol 364, 799-809. [本文引用: 1]
JennessMK, CarraroN, PritchardCA, MurphyAS ( 2019). The Arabidopsis ATP-binding cassette transporter ABCB21 regulates auxin levels in cotyledons, the root pericycle, and leaves Front Plant Sci 10, 806. [本文引用: 2]
JensenPJ, HangarterRP, EstelleM ( 1998). Auxin trans-port is required for hypocotyl elongation in light-grown but not dark-grownArabidopsis Plant Physiol 116, 455-462. [本文引用: 1]
KanedaM, SchuetzM, LinBS, ChanisC, HambergerB, WesternTL, EhltingJ, SamuelsAL ( 2011). ABC transporters coordinately expressed during lignification of Arabidopsis stems include a set of ABCBs associated with auxin transport J Exp Bot 62, 2063-2077. [本文引用: 5]
KimDY, BovetL, KushnirS, NohEW, MartinoiaE, LeeY ( 2006). AtATM3 is involved in heavy metal resistance in Arabidopsis Plant Physiol 140, 922-932. [本文引用: 3]
KimJY, HenrichsS, BaillyA, VincenzettiV, SoveroV, MancusoS, PollmannS, KimD, GeislerM, NamHG ( 2010). Identification of an ABCB/P-glycoprotein-specific inhibitor of auxin transport by chemical genomics J Biol Chem 285, 23309-23317. [本文引用: 2]
LarsenPB, CancelJ, RoundsM, OchoaV ( 2007). Arabidopsis ALS1 encodes a root tip and stele localized half type ABC transporter required for root growth in an aluminum toxic environment Planta 225, 1447-1458. [本文引用: 3]
LeeM, ChoiY, BurlaB, KimYY, JeonB, MaeshimaM, YooJY, MartinoiaE, LeeY ( 2008). The ABC transporter AtABCB14 is a malate importer and modulates stomatal response to CO2 Nat Cell Biol 10, 1217-1223. [本文引用: 2]
LewisDR, MillerND, SplittBL, WuGS, SpaldingEP ( 2007). Separating the roles of acropetal and basipetal auxin transport on gravitropism with mutations in two Arabidopsis multidrug resistance-like ABC transporter genes Plant Cell 19, 1838-1850. [本文引用: 3]
LinRC, WangHY ( 2005). Two homologous ATP-binding cassette transporter proteins, AtMDR1 and AtPGP1, regulate Arabidopsis photomorphogenesis and root development by mediating polar auxin transport Plant Physiol 138, 949-964. [本文引用: 1]
MultaniDS, BriggsSP, ChamberlinMA, BlakesleeJJ, MurphyAS, JohalGS ( 2003). Loss of an MDR transporter in compact stalks of maize br2 and sorghum dw3 mutants Science 302, 81-84. [本文引用: 4]
NagashimaA, SuzukiG, UeharaY, SajiK, FurukawaT, KoshibaT, SekimotoM, FujiokaS, KurohaT, KojimaM, SakakibaraH, FujisawaN, OkadaK, SakaiT ( 2008). Phytochromes and cryptochromes regulate the differential growth of Arabidopsis hypocotyls in both a PGP19-dependent and a PGP19-independent manner Plant J 53, 516-529. [本文引用: 1]
NguyenVNT, MoonS, JungKH ( 2014). Genome-wide expression analysis of rice ABC transporter family across spatio-temporal samples and in response to abiotic stresses J Plant Physiol 171, 1276-1288. [本文引用: 1]
NohB, MurphyAS, SpaldingEP ( 2001). Multidrug resistance-like genes of Arabidopsis required for auxin transport and auxin-mediated development Plant Cell 13, 2441-2454. [本文引用: 3]
OkamotoK, UedaH, ShimadaT, TamuraK, KoumotoY, TasakaM, MoritaMT, Hara-NishimuraI ( 2016). An ABC transporter B family protein, ABCB19, is required for cytoplasmic streaming and gravitropism of the inflorescence stems Plant Signal Behav 11, e1010947. [本文引用: 1]
PangKY, LiYJ, LiuMH, MengZD, YuYL ( 2013). Inventory and general analysis of the ATP-binding cassette (ABC) gene superfamily in maize (Zea mays L.) Gene 526, 411-428. [本文引用: 4]
ParksBM, SpaldingEP ( 1999). Sequential and coordinated action of phytochromes A and B during Arabidopsis stem growth revealed by kinetic analysis Proc Natl Acad Sci USA 96, 14142-14146. [本文引用: 1]
SahaJ, SenguptaA, GuptaK, GuptaB ( 2015). Molecular phylogenetic study and expression analysis of ATP- binding cassette transporter gene family in Oryza sativa in response to salt stress Comput Biol Chem 54, 18-32. [本文引用: 1]
SalisburyFJ, HallA, GriersonCS, HallidayKJ ( 2007). Phytochrome coordinates Arabidopsis shoot and root development Plant J 50, 429-438. [本文引用: 1]
Sánchez-FernándezR, DaviesTGE, ColemanJOD, ReaPA ( 2001). The Arabidopsis thaliana ABC protein superfamily, a complete inventory J Biol Chem 276, 30231-30244. [本文引用: 3]
SanteliaD, VincenzettiV, AzzarelloE, BovetL, FukaoY, DüchtigP, MancusoS, MartinoiaE, GeislerM ( 2005). MDR-like ABC transporter AtPGP4 is involved in auxin- mediated lateral root and root hair development FEBS Lett 579, 5399-5406. [本文引用: 3]
SasakiT, EzakiB, MatsumotoH ( 2002). A gene encoding multidrug resistance (MDR)-like protein is induced by aluminum and inhibitors of calcium flux in wheat Plant Cell Physiol 43, 177-185. [本文引用: 2]
ShenCJ, BaiYH, WangSK, ZhangSN, WuYR, ChenM, JiangDA, QiYH ( 2010). Expression profile of PIN, AUX/LAX and PGP auxin transporter gene families in Sorghum bicolor under phytohormone and abiotic stress FEBS J 277, 2954-2969. [本文引用: 2]
SidlerM, HassaP, HasanS, RingliC, DudlerR ( 1998). Involvement of an ABC transporter in a developmental pathway regulating hypocotyl cell elongation in the light Plant Cell 10, 1623-1636. [本文引用: 3]
SukumarP, MaloneyGS, MudayGK ( 2013). Localized induction of the ATP-binding cassette B19 auxin transporter enhances adventitious root formation in Arabidopsis Plant Physiol 162, 1392-1405. [本文引用: 2]
TerasakaK, BlakesleeJJ, TitapiwatanakunB, PeerWA, BandyopadhyayA, MakamSN, LeeOR, RichardsEL, MurphyAS, SatoF, YazakiK ( 2005). PGP4, an ATP binding cassette P-glycoprotein, catalyzes auxin transport in Arabidopsis thaliana roots Plant Cell 17, 2922-2939. [本文引用: 3]
TheodoulouFL, KerrID ( 2015). ABC transporter research: going strong 40 years on Biochem Soc Trans 43, 1033-1040. [本文引用: 2]
WangSK, ShenCJ, ZhangSN, XuYX, JiangDA, QiYH ( 2011). Analysis of subcellular localization of auxin carriers PIN, AUX/LAX and PGP in Sorghum bicolor Plant Signal Behav 6, 2023-2025. [本文引用: 1]
WuGS, CameronJN, LjungK, SpaldingEP ( 2010). A role for ABCB19-mediated polar auxin transport in seedling photomorphogenesis mediated by cryptochrome 1 and phytochrome B Plant J 62, 179-191. [本文引用: 4]
XuYX, ZhangSN, GuoHP, WangSK, XuLG, LiCY, QianQ, ChenF, GeislerM, QiYH, JiangDA ( 2014). OsABCB14 functions in auxin transport and iron homeostasis in rice ( Oryza sativa L.) Plant J 79, 106-117. [本文引用: 4]
ZhangYQ, NasserV, PisantyO, OmaryM, WulffN, Di DonatoM, TalI, HauserF, HaoPC, RothO, FrommH, SchroederJI, GeislerM, Nour-EldinHH, ShaniE ( 2018). A transportome-scale amiRNA-based screen identifies redundant roles of Arabidopsis ABCB6 and ABCB20 in auxin transport Nat Commun 9, 4204. [本文引用: 4]
ZhaoHT, LiuL, MoHX, QianLT, CaoY, CuiSJ, LiX, MaLG ( 2013). The ATP-binding cassette transporter ABCB19 regulates postembryonic organ separation in Arabidopsis PLoS One 8, e60809. [本文引用: 2]
ZhuJS, BaillyA, ZwiewkaM, SoveroV, Di DonatoM, GeP, OehriJ, AryalB, HaoPC, LinnertM, BurgardtNI, LückeC, WeiwadM, MichelM, Weiergr?berOH, PollmannS, AzzarelloE, MancusoS, FerroN, FukaoY, HoffmannC, Wedlich-S?ldnerR, FrimlJ, ThomasC, GeislerM ( 2016). TWISTED DWARF1 mediates the action of auxin transport inhibitors on actin cytoskeleton dynamics Plant Cell 28, 930-948. [本文引用: 3]
Structural model of ABCB transporter (modified from <xref ref-type="bibr" rid="b2">Wang et al., 2007</xref>, in Chinese) (A) Structural model of ABCB full-molecular transporter; (B) Structural model of ABCB half-molecular transporter ...
拟南芥ABC转运蛋白研究进展 4 2017
... 拟南芥和水稻ABCB转运蛋白的分类及命名(Verrier et al., 2008; 王晓珠等, 2017) ...
... Classification and nomenclature of ABCB transporters in Arabidopsis thaliana and Oryza sativa (Verrier et al., 2008; Wang et al., 2017, in Chinese) ...
... 拟南芥ABCB基因的生物学功能(王晓珠等, 2017) ...
... Biofunctions of ABCB genes in Arabidopsis thaliana (Wang et al., 2017, in Chinese) ...
水稻ABCB转运蛋白基因的分子进化和表达分析 1 2012
... 水稻ABCB基因的研究也一直处于缓慢发展状态.水稻OsABCB14与拟南芥AtABCB19同源性较高, 且OsABCB14在水稻根尖、叶鞘、中柱、叶片、节、节间、根茎结合部和花等器官中均有表达, 其中在维管组织中的表达量最高(Xu et al., 2014).osabcb14突变体生长素的向顶运输速率降低, 且离子含量分析显示其铁离子浓度明显高于野生型, 表明OsABCB14不仅参与生长素的转运, 还与水稻铁离子的平衡密切相关(Xu et al., 2014).水稻全基因组表达分析显示, OsABCB10在花药和花粉中表达水平较高, 暗示其在花药和花粉的发育过程中发挥重要作用(Nguyen et al., 2014).非生物胁迫下的表达分析表明大部分ABCB基因的表达量发生变化.例如, OsABCB11、OsABCB8、OsABCB13、OsABCB23和OsABCB24受干旱胁迫诱导, 而OsABCB6、OsABCB9以及OsABCB8受盐胁迫诱导, 但ABCB亚家族基因在水稻非生物胁迫响应中的作用还有待进一步验证(徐杏等, 2012; Saha et al., 2015).此外, 定位于液泡膜上的TAP型转运蛋白OsABCB27 (OsALS1)参与水稻的铝胁迫响应(Huang et al., 2009), 而定位于线粒体的ATM型转运蛋白OsABCB23 (ATM3)在铁硫聚簇装配中起重要作用, 表明线粒体ATM3转运蛋白对水稻铁离子的稳态至关重要(Zuo et al., 2017). ...
Cryptochrome photoreceptors cry1 and cry2 antagonistically regulate primary root elongation in Arabidopsis thaliana 1 2006
... 20世纪20年代提出的Cholodny-Went理论认为, 植物向光性运动的产生是由于单侧光照射下生长素逐渐向背光一侧移动, 进而使生长素在植物茎中分布不均引起.后来该假说也被用于解释植物的向性生长.光敏色素(Phy)和隐花色素(Cry)是植物体内的光受体, 影响植物的向光性甚至光形态建成.许多研究表明, 光敏色素和隐花色素调节下胚轴顶端到根部的生长素向基运输(Jensen et al., 1998; Canamero et al., 2006; Salisbury et al., 2007).ABCB依赖光受体(PhyA、Phot和PhyB)调控光下生长素的极性运输, 进而调节下胚轴生长(Wu et al., 2010; Christie et al., 2011; 刘广超和丁兆军, 2018).Lin和Wang (2005)研究表明, atabcb1和atabcb19突变体对远红光、红光及蓝光较为敏感, 表现出下胚轴生长受抑制的表型, 说明AtABCB1和AtABCB19通过调控生长素的极性运输来影响拟南芥的光形态建成.此外, Wu等(2010)研究发现, 过表达AtABCB19可显著增加下胚轴中生长素的含量, 且红光和蓝光均可减弱其对下胚轴伸长的抑制作用.AtABCB19突变则增强了光敏色素和隐花色素诱导的拟南芥下胚轴弯曲, 说明光敏色素和隐花色素可能通过抑制AtABCB19的表达, 进而抑制下胚轴中生长素的极性运输(Nagashima et al., 2008).研究者认为上述调控模型中首先是红光通过PhyA抑制下胚轴伸长, 而后PhyB作用于AtABCB19影响幼苗中生长素的极性转运(Parks and Spalding, 1999; Wu et al., 2010). ...
ABCB1 and ABCB19 auxin transporters have synergistic effects on early and late Arabidopsis anther development 1 2015
... ABCB亚家族成员在植物生长发育的各个方面发挥不同作用(表2).植物根切除导致吲哚-3-乙酸(IAA)转运快速增加, 并在切除点以上的下胚轴中积累, 进而诱导不定根的形成.Sukumar等(2013)研究发现, 相对于野生型株系atabcb19突变体的不定根数量显著减少, 过表达株系则出现较多的不定根, 且AtABCB19的转录及蛋白表达水平与局部IAA积累的时空变化模式密切相关, 因此认为AtABCB19的局部诱导可促进拟南芥不定根的形成.此外, 有研究表明, AtABCB19还参与调控拟南芥胚后器官的分离(Zhao et al., 2013)及花序轴的细胞质流(Okamoto et al., 2016).AtABCB1在花药发育过程中起主要作用, AtABCB19则起协同作用(Cecchetti et al., 2015).Kaneda等(2011)发现, atabcb14花序轴中维管发育不正常, 说明AtABCB14可能参与拟南芥花序轴的木质化过程.而之前有报道显示, AtABCB14可将苹果酸盐从质外体转运至保卫细胞, 调节渗透压, 从而调控细胞对二氧化碳的响应, 影响气孔开闭(Lee et al., 2008).总之, ABCB蛋白功能复杂多样, 仍有待进一步研究. ...
Comprehensive analysis and expression profiling of the OsLAX and OsABCB auxin transporter gene families in rice(Oryza sativa) under phytohormone stimuli and abiotic stresses 1 2016
... 自1992年从拟南芥(Arabidopsis thaliana)中克隆到第1个ABCB亚家族的基因——AtPGP1 (AtABCB1)以来, 植物中的ABCB亚家族研究备受关注(Dudler and Hertig, 1992).近30年的研究发现, ABCB作为转运蛋白与AUX1/LAX (AUXIN1/LIKE AUXIN)和PIN (PIN-FORMED)协同调控植物生长素的极性运输.例如, AtABCB1、AtABCB19、AtABCB4、AtABCB14和AtABCB21 (Sidler et al.,1998; Noh et al., 2001; Geisler et al., 2005; Santelia et al., 2005; Kaneda et al., 2011; Kamimoto et al., 2012)等相继被报道与生长素的运输相关.此外, 也有研究表明, ABCB转运蛋白参与植物对重金属和铝毒的抗性等过程.例如, Kim等(2016)报道了ABCB半分子转运蛋白ATM3 (AtABCB24)可提高拟南芥对镉和铅的抗性.而ALS1/TAP2 (AtABCB27)被报道参与拟南芥的铝转运, 进而提高其铝毒抗性(Larsen et al., 2007).单子叶植物玉米(Zea mays)、高粱(Sorghum bicolor)和水稻(Oryza sativa)中的ABCB蛋白也具有生长素转运功能.玉米ZmABCB1 (Brachytic2, BR2)和高粱SbABCB1 (Dwarf1, DW3)参与生长素的向基运输(Multani et al., 2003); 水稻OsABCB14则参与生长素的向顶运输(Xu et al., 2014).单子叶植物的转录组数据分析显示, ABCB亚家族蛋白可能与植物的非生物胁迫响应密切相关(Shen et al., 2010; Pang et al., 2013; Chai and Subudhi, 2016).可见, ABCB转运蛋白的功能在植物中具有一定的保守性.植物ABCB转运蛋白的数量较多, 然而目前的研究主要集中在双子叶模式植物拟南芥中, 单子叶植物中的研究尚少, 大部分ABCB转运蛋白的功能有待进一步探索.本文主要综述了ABCB亚家族蛋白在双子叶植物拟南芥以及单子叶植物玉米、高粱、小麦(Triticum aestivum)和水稻中的研究进展, 旨在为ABCB基因功能的揭示提供新线索. ...
Functional characterization of AtATM1, AtATM2, and AtATM3, a subfamily of Arabidopsis half- molecule ATP-binding cassette transporters implicated in iron homeostasis 4 2007
... Biofunctions of ABCB genes in Arabidopsis thaliana (Wang et al., 2017, in Chinese) Table 2
类型
基因名称
功能
参考文献
MDR (multidrug resistance)
AtABCB1/PGP1
参与地上部(花序轴、下胚轴)生长素输出, 促进下胚轴伸长
Sidler et al.,1998; Geisler et al., 2005
AtABCB4/PGP4
在拟南芥根中表现为生长素兼性转运载体, 介导侧根及根毛发育
Santelia et al., 2005; Terasaka et al., 2005; Cho et al., 2007; Lewis et al., 2007; Kubeš et al., 2012
AtABCB6
与AtABCB20在功能上冗余, 参与花序轴中生长素的向基转运
Zhang et al., 2018
AtABCB14/MDR12
参与花序轴中生长素运输, 与花序轴维管发育和气孔开闭有关
Lee et al., 2008; Kaneda et al., 2011
AtABCB15
参与花序轴中生长素运输
Kaneda et al., 2011
AtABCB19/PGP19
参与地上部生长素向基运输以及根的向顶运输, 调控不定根发育及胚后器官分离
Noh et al., 2001; Lewis et al., 2007; Sukumar et al., 2013; Zhao et al., 2013
AtABCB20
与AtABCB6在功能上冗余, 参与花序轴中生长素向基转运
Zhang et al., 2018
AtABCB21
地上部生长素兼性运输载体, 参与生长素向顶转运, 调控生长素水平
Kamimoto et al., 2012; Jenness et al., 2019
ATM (ABC transporter of mitochondria)
AtABCB23/ATM1
参与铁硫簇输出
Chen et al., 2007
AtABCB24/ATM2
未知
Chen et al., 2007
AtABCB25/ATM3
参与铁硫簇输出, 重金属抗性, 并参与调控钼辅因子合成
Kim et al., 2006; Chen et al., 2007; Bernard et al., 2009; Teschner et al., 2010
TAP (transporter associated with antigen processing)
AtABCB26/TAP1 AtABCB27/TAP2/ALS1
未知 调控铝毒环境中根的发育
Ferro et al., 2010 Larsen et al., 2007; Zhu et al., 2013
半分子转运蛋白AtABCB23 (AtATM1)、AtAB-CB24 (AtATM2)以及AtABCB25 (AtATM3)与酵母线粒体膜蛋白ATM1 (ScATM1)同源.其中, AtATM3以及AtATM1参与线粒体合成时铁硫簇的输出(Chen et al., 2007); 且AtATM3还与拟南芥的重金属抗性有关, 过表达AtATM3可提高拟南芥对镉和铅的抗性(Kim et al., 2006).TAP型转运蛋白成员AtABCB27 (ALS1/TAP2)定位于液泡膜, 在铝毒环境下根系的发 ...
... Chen et al., 2007
AtABCB25/ATM3
参与铁硫簇输出, 重金属抗性, 并参与调控钼辅因子合成
Kim et al., 2006; Chen et al., 2007; Bernard et al., 2009; Teschner et al., 2010
TAP (transporter associated with antigen processing)
AtABCB26/TAP1 AtABCB27/TAP2/ALS1
未知 调控铝毒环境中根的发育
Ferro et al., 2010 Larsen et al., 2007; Zhu et al., 2013
半分子转运蛋白AtABCB23 (AtATM1)、AtAB-CB24 (AtATM2)以及AtABCB25 (AtATM3)与酵母线粒体膜蛋白ATM1 (ScATM1)同源.其中, AtATM3以及AtATM1参与线粒体合成时铁硫簇的输出(Chen et al., 2007); 且AtATM3还与拟南芥的重金属抗性有关, 过表达AtATM3可提高拟南芥对镉和铅的抗性(Kim et al., 2006).TAP型转运蛋白成员AtABCB27 (ALS1/TAP2)定位于液泡膜, 在铝毒环境下根系的发 ...
... ; Chen et al., 2007; Bernard et al., 2009; Teschner et al., 2010
TAP (transporter associated with antigen processing)
AtABCB26/TAP1 AtABCB27/TAP2/ALS1
未知 调控铝毒环境中根的发育
Ferro et al., 2010 Larsen et al., 2007; Zhu et al., 2013
半分子转运蛋白AtABCB23 (AtATM1)、AtAB-CB24 (AtATM2)以及AtABCB25 (AtATM3)与酵母线粒体膜蛋白ATM1 (ScATM1)同源.其中, AtATM3以及AtATM1参与线粒体合成时铁硫簇的输出(Chen et al., 2007); 且AtATM3还与拟南芥的重金属抗性有关, 过表达AtATM3可提高拟南芥对镉和铅的抗性(Kim et al., 2006).TAP型转运蛋白成员AtABCB27 (ALS1/TAP2)定位于液泡膜, 在铝毒环境下根系的发 ...
... 半分子转运蛋白AtABCB23 (AtATM1)、AtAB-CB24 (AtATM2)以及AtABCB25 (AtATM3)与酵母线粒体膜蛋白ATM1 (ScATM1)同源.其中, AtATM3以及AtATM1参与线粒体合成时铁硫簇的输出(Chen et al., 2007); 且AtATM3还与拟南芥的重金属抗性有关, 过表达AtATM3可提高拟南芥对镉和铅的抗性(Kim et al., 2006).TAP型转运蛋白成员AtABCB27 (ALS1/TAP2)定位于液泡膜, 在铝毒环境下根系的发 ...
P-glycoprotein4 displays auxin efflux transporter-like action in Arabidopsis root hair cells and tobacco cells 3 2007
... Lewis等(2007)发现mdr1 (abcb19)根中生长素向顶运输减少80%, 但是向基运输不受影响; mdr4 (abcb4)向基运输减少50%, 而向顶运输不受影响, 说明不同ABCB蛋白在生长素转运中可能起着截然不同的作用.此外, AtPGP4最早被报道参与生长素介导的侧根及根毛发育, 酵母中异源表达AtPGP4可导致其对NPA的敏感性升高; 且atpgp4对生长素及NPA的敏感性下降, 该突变体中游离生长素水平升高而生长素运输水平下降, 表明AtPGP4直接参与调控生长素的转运, 进而影响侧根及根毛的发育(Santelia et al., 2005).同年, Terasaka等(2005)证实, AtPGP4主要参与调控根表皮层细胞生长素的输入过程(Terasaka et al., 2005); 而Cho等(2007)研究发现, AtPGP4在拟南芥根毛细胞以及烟草(Nicotiana taba- cum)细胞中都表现出生长素输出的作用(Cho et al., 2007).Kubeš等(2012)对AtPGP4蛋白进行了研究, 发现其是一种双向生长素转运蛋白, 当细胞中生长素水平较低时促进生长素输入, 较高时则促进生长素输出.AtABCB4的同源蛋白AtABCB21与AtPGP4类似, 也是双向生长素转运蛋白(Kamimoto et al., 2012).最新研究显示, AtABCB21能够调控拟南芥子叶、根的韧皮部以及叶片中的生长素水平, 在拟南芥地上部生长素向顶运输以及植株的向光性运动中发挥作用(Jenness et al., 2019). ...
... )细胞中都表现出生长素输出的作用(Cho et al., 2007).Kubeš等(2012)对AtPGP4蛋白进行了研究, 发现其是一种双向生长素转运蛋白, 当细胞中生长素水平较低时促进生长素输入, 较高时则促进生长素输出.AtABCB4的同源蛋白AtABCB21与AtPGP4类似, 也是双向生长素转运蛋白(Kamimoto et al., 2012).最新研究显示, AtABCB21能够调控拟南芥子叶、根的韧皮部以及叶片中的生长素水平, 在拟南芥地上部生长素向顶运输以及植株的向光性运动中发挥作用(Jenness et al., 2019). ...
... Biofunctions of ABCB genes in Arabidopsis thaliana (Wang et al., 2017, in Chinese) Table 2
类型
基因名称
功能
参考文献
MDR (multidrug resistance)
AtABCB1/PGP1
参与地上部(花序轴、下胚轴)生长素输出, 促进下胚轴伸长
Sidler et al.,1998; Geisler et al., 2005
AtABCB4/PGP4
在拟南芥根中表现为生长素兼性转运载体, 介导侧根及根毛发育
Santelia et al., 2005; Terasaka et al., 2005; Cho et al., 2007; Lewis et al., 2007; Kubeš et al., 2012
AtABCB6
与AtABCB20在功能上冗余, 参与花序轴中生长素的向基转运
Zhang et al., 2018
AtABCB14/MDR12
参与花序轴中生长素运输, 与花序轴维管发育和气孔开闭有关
Lee et al., 2008; Kaneda et al., 2011
AtABCB15
参与花序轴中生长素运输
Kaneda et al., 2011
AtABCB19/PGP19
参与地上部生长素向基运输以及根的向顶运输, 调控不定根发育及胚后器官分离
Noh et al., 2001; Lewis et al., 2007; Sukumar et al., 2013; Zhao et al., 2013
AtABCB20
与AtABCB6在功能上冗余, 参与花序轴中生长素向基转运
Zhang et al., 2018
AtABCB21
地上部生长素兼性运输载体, 参与生长素向顶转运, 调控生长素水平
Kamimoto et al., 2012; Jenness et al., 2019
ATM (ABC transporter of mitochondria)
AtABCB23/ATM1
参与铁硫簇输出
Chen et al., 2007
AtABCB24/ATM2
未知
Chen et al., 2007
AtABCB25/ATM3
参与铁硫簇输出, 重金属抗性, 并参与调控钼辅因子合成
Kim et al., 2006; Chen et al., 2007; Bernard et al., 2009; Teschner et al., 2010
TAP (transporter associated with antigen processing)
AtABCB26/TAP1 AtABCB27/TAP2/ALS1
未知 调控铝毒环境中根的发育
Ferro et al., 2010 Larsen et al., 2007; Zhu et al., 2013
半分子转运蛋白AtABCB23 (AtATM1)、AtAB-CB24 (AtATM2)以及AtABCB25 (AtATM3)与酵母线粒体膜蛋白ATM1 (ScATM1)同源.其中, AtATM3以及AtATM1参与线粒体合成时铁硫簇的输出(Chen et al., 2007); 且AtATM3还与拟南芥的重金属抗性有关, 过表达AtATM3可提高拟南芥对镉和铅的抗性(Kim et al., 2006).TAP型转运蛋白成员AtABCB27 (ALS1/TAP2)定位于液泡膜, 在铝毒环境下根系的发 ...
phot1 inhibition of ABCB19 primes lateral auxin fluxes in the shoot apex required for phototropism 4 2011
... 20世纪20年代提出的Cholodny-Went理论认为, 植物向光性运动的产生是由于单侧光照射下生长素逐渐向背光一侧移动, 进而使生长素在植物茎中分布不均引起.后来该假说也被用于解释植物的向性生长.光敏色素(Phy)和隐花色素(Cry)是植物体内的光受体, 影响植物的向光性甚至光形态建成.许多研究表明, 光敏色素和隐花色素调节下胚轴顶端到根部的生长素向基运输(Jensen et al., 1998; Canamero et al., 2006; Salisbury et al., 2007).ABCB依赖光受体(PhyA、Phot和PhyB)调控光下生长素的极性运输, 进而调节下胚轴生长(Wu et al., 2010; Christie et al., 2011; 刘广超和丁兆军, 2018).Lin和Wang (2005)研究表明, atabcb1和atabcb19突变体对远红光、红光及蓝光较为敏感, 表现出下胚轴生长受抑制的表型, 说明AtABCB1和AtABCB19通过调控生长素的极性运输来影响拟南芥的光形态建成.此外, Wu等(2010)研究发现, 过表达AtABCB19可显著增加下胚轴中生长素的含量, 且红光和蓝光均可减弱其对下胚轴伸长的抑制作用.AtABCB19突变则增强了光敏色素和隐花色素诱导的拟南芥下胚轴弯曲, 说明光敏色素和隐花色素可能通过抑制AtABCB19的表达, 进而抑制下胚轴中生长素的极性运输(Nagashima et al., 2008).研究者认为上述调控模型中首先是红光通过PhyA抑制下胚轴伸长, 而后PhyB作用于AtABCB19影响幼苗中生长素的极性转运(Parks and Spalding, 1999; Wu et al., 2010). ...
... 另外, 光受体激酶AGC4/Phot1可直接磷酸化AtABCB19, 使AtABCB19的生长素输出活性受抑制, 下胚轴顶部生长素水平升高.PIN3 (PIN-FORMED3)将生长素转运至伸长区并参与横向运输, 影响植物的向光性(Christie et al., 2011).早期已有PIN与ABCB之间存在互作的报道, 但是关于生长素横向浓度梯度产生的机制至今仍不清楚(Bandyopadhyay et al., 2007).Blakeslee等(2007)发现AtABCB19可与PIN1蛋白互作, 影响PIN在质膜微区的稳定性, 进而提高转运蛋白的底物特异性.转运蛋白通常并不单独发挥作用, 而是与其它蛋白协同调控植物的生长, 多个不同的生长素转运蛋白协作可能是导致生长素横向分布不均的一个重要因素(Christie et al., 2011). ...
... 发现AtABCB19可与PIN1蛋白互作, 影响PIN在质膜微区的稳定性, 进而提高转运蛋白的底物特异性.转运蛋白通常并不单独发挥作用, 而是与其它蛋白协同调控植物的生长, 多个不同的生长素转运蛋白协作可能是导致生长素横向分布不均的一个重要因素(Christie et al., 2011). ...
... AGC蛋白激酶可调节ABCB转运蛋白的活性.在HeLa细胞表达系统中, 蛋白激酶AGC4/Phot1能与AtABCB19蛋白的NBD区结合使其磷酸化, 进而降低其活性(Christie et al., 2011) (图2).Co-IP和BRET实验证明, AGC3/PID与TWD1存在互作, PID通过与TWD1互作直接磷酸化AtABCB1, 从而使其生长素转运活性受抑制(Henrichs et al., 2012).然而, PID磷酸化ABCB是否必须TWD1介导目前尚不清楚.现有的研究表明AGC激酶可磷酸化ABCB, 降低其与TWD1间的互作, 从而改变ABCB蛋白跨膜转运过程中TWD1的分子伴侣作用, 影响其活性. ...
The human ATP-binding cassette (ABC) transporter superfamily 1 2001
... 早期, HUGO命名系统中, ABC转运蛋白被分为ABCA-ABCH八个亚家族(Dean et al., 2001).而拟南芥的ABC转运蛋白, 根据分子大小、方向、结构域组成以及同源性不同分成13个亚家族, 分别为ATH (ABC2 homolog)、ATM (ABC transporter of the mitochondria)、MRP (multidrug resistance associa- ted protein)、PDR (pleiotropic drug resistance)、TAP (transporter associated with antigen processing)、AOH (ABC1 homolog)、NAP (non-intrinsic ABC protein)、PMP (peroxisomal membrane protein)、WBC (white-brown complex homolog)、RLI (RNase L inhibitor)、GCN (general control non-repressible)、MDR (multidrug resistance)和SMC (structural maintenance of chromosomes) (Sánchez-Fernández et al., 2001).随着科学技术的不断进步, 越来越多的蛋白基因被克隆.而不同的命名方式造成了基因名称的混乱, Verrier等(2008)整理并统一了植物中ABC转运蛋白的命名方式.其中, ABCB亚家族分为全分子和半分子转运蛋白两类.全分子转运蛋白仅包括MDR型转运蛋白, 而半分子转运蛋白包括HMT/ATM、TAP和LLP (prokaryotic lipid A-like exporters, putative)型3种(表1). ...
Structure of an mdr-like gene from Arabidopsis thaliana. Evolutionary implications 2 1992
... 自1992年从拟南芥(Arabidopsis thaliana)中克隆到第1个ABCB亚家族的基因——AtPGP1 (AtABCB1)以来, 植物中的ABCB亚家族研究备受关注(Dudler and Hertig, 1992).近30年的研究发现, ABCB作为转运蛋白与AUX1/LAX (AUXIN1/LIKE AUXIN)和PIN (PIN-FORMED)协同调控植物生长素的极性运输.例如, AtABCB1、AtABCB19、AtABCB4、AtABCB14和AtABCB21 (Sidler et al.,1998; Noh et al., 2001; Geisler et al., 2005; Santelia et al., 2005; Kaneda et al., 2011; Kamimoto et al., 2012)等相继被报道与生长素的运输相关.此外, 也有研究表明, ABCB转运蛋白参与植物对重金属和铝毒的抗性等过程.例如, Kim等(2016)报道了ABCB半分子转运蛋白ATM3 (AtABCB24)可提高拟南芥对镉和铅的抗性.而ALS1/TAP2 (AtABCB27)被报道参与拟南芥的铝转运, 进而提高其铝毒抗性(Larsen et al., 2007).单子叶植物玉米(Zea mays)、高粱(Sorghum bicolor)和水稻(Oryza sativa)中的ABCB蛋白也具有生长素转运功能.玉米ZmABCB1 (Brachytic2, BR2)和高粱SbABCB1 (Dwarf1, DW3)参与生长素的向基运输(Multani et al., 2003); 水稻OsABCB14则参与生长素的向顶运输(Xu et al., 2014).单子叶植物的转录组数据分析显示, ABCB亚家族蛋白可能与植物的非生物胁迫响应密切相关(Shen et al., 2010; Pang et al., 2013; Chai and Subudhi, 2016).可见, ABCB转运蛋白的功能在植物中具有一定的保守性.植物ABCB转运蛋白的数量较多, 然而目前的研究主要集中在双子叶模式植物拟南芥中, 单子叶植物中的研究尚少, 大部分ABCB转运蛋白的功能有待进一步探索.本文主要综述了ABCB亚家族蛋白在双子叶植物拟南芥以及单子叶植物玉米、高粱、小麦(Triticum aestivum)和水稻中的研究进展, 旨在为ABCB基因功能的揭示提供新线索. ...
Master and servant: regulation of auxin transporters by FKBPs and cyclophilins 1 2016
... 免疫亲和素可与ABCB蛋白互作, 间接影响生长素的运输(Geisler et al., 2004, 2016; Granzin et al., 2006).而分子伴侣TWD1最初是在酵母双杂交中被发现的与ABCB1互作的蛋白(Geisler et al., 2003).后来, 通过免疫共沉淀(Co-immunoprecipitation, Co- IP)以及生物发光能量共振转移(BRET)技术进一步证明TWD1和ABCB1在酵母及植物表达系统中存在互作(Wang et al., 2013).TWD1作为分子伴侣可通过促进AtABCB1重折叠改变其构型, 增强其ATP及底物结合能力(Geisler et al., 2004).此外, twd1突变体中被错误定位到内质网膜上的AtABCB1、AtABCB4和AtABCB19最终会被降解, 表明TWD1主要在ABCB蛋白从内质网膜到质膜的转运过程中发挥分子伴侣作用(Wu et al., 2010; Wang et al., 2013) (图2).近年, 有研究表明NPA和BUM等生长素非竞争性抑制剂可通过结合TWD1, 导致其与ABCB的结合受抑制, 从而使生长素运输减少(Kim et al., 2010; Zhu et al., 2016).Zhu等(2016)研究发现, 肌动蛋白ACTIN7与TWD1之间存在物理互作, 二者可能在不同层次控制蛋白的转运(Zhu et al., 2016). ...
Cellular efflux of auxin catalyzed by the Arabidopsis MDR/PGP transporter AtPGP1 3 2005
... 自1992年从拟南芥(Arabidopsis thaliana)中克隆到第1个ABCB亚家族的基因——AtPGP1 (AtABCB1)以来, 植物中的ABCB亚家族研究备受关注(Dudler and Hertig, 1992).近30年的研究发现, ABCB作为转运蛋白与AUX1/LAX (AUXIN1/LIKE AUXIN)和PIN (PIN-FORMED)协同调控植物生长素的极性运输.例如, AtABCB1、AtABCB19、AtABCB4、AtABCB14和AtABCB21 (Sidler et al.,1998; Noh et al., 2001; Geisler et al., 2005; Santelia et al., 2005; Kaneda et al., 2011; Kamimoto et al., 2012)等相继被报道与生长素的运输相关.此外, 也有研究表明, ABCB转运蛋白参与植物对重金属和铝毒的抗性等过程.例如, Kim等(2016)报道了ABCB半分子转运蛋白ATM3 (AtABCB24)可提高拟南芥对镉和铅的抗性.而ALS1/TAP2 (AtABCB27)被报道参与拟南芥的铝转运, 进而提高其铝毒抗性(Larsen et al., 2007).单子叶植物玉米(Zea mays)、高粱(Sorghum bicolor)和水稻(Oryza sativa)中的ABCB蛋白也具有生长素转运功能.玉米ZmABCB1 (Brachytic2, BR2)和高粱SbABCB1 (Dwarf1, DW3)参与生长素的向基运输(Multani et al., 2003); 水稻OsABCB14则参与生长素的向顶运输(Xu et al., 2014).单子叶植物的转录组数据分析显示, ABCB亚家族蛋白可能与植物的非生物胁迫响应密切相关(Shen et al., 2010; Pang et al., 2013; Chai and Subudhi, 2016).可见, ABCB转运蛋白的功能在植物中具有一定的保守性.植物ABCB转运蛋白的数量较多, 然而目前的研究主要集中在双子叶模式植物拟南芥中, 单子叶植物中的研究尚少, 大部分ABCB转运蛋白的功能有待进一步探索.本文主要综述了ABCB亚家族蛋白在双子叶植物拟南芥以及单子叶植物玉米、高粱、小麦(Triticum aestivum)和水稻中的研究进展, 旨在为ABCB基因功能的揭示提供新线索. ...
... 免疫亲和素可与ABCB蛋白互作, 间接影响生长素的运输(Geisler et al., 2004, 2016; Granzin et al., 2006).而分子伴侣TWD1最初是在酵母双杂交中被发现的与ABCB1互作的蛋白(Geisler et al., 2003).后来, 通过免疫共沉淀(Co-immunoprecipitation, Co- IP)以及生物发光能量共振转移(BRET)技术进一步证明TWD1和ABCB1在酵母及植物表达系统中存在互作(Wang et al., 2013).TWD1作为分子伴侣可通过促进AtABCB1重折叠改变其构型, 增强其ATP及底物结合能力(Geisler et al., 2004).此外, twd1突变体中被错误定位到内质网膜上的AtABCB1、AtABCB4和AtABCB19最终会被降解, 表明TWD1主要在ABCB蛋白从内质网膜到质膜的转运过程中发挥分子伴侣作用(Wu et al., 2010; Wang et al., 2013) (图2).近年, 有研究表明NPA和BUM等生长素非竞争性抑制剂可通过结合TWD1, 导致其与ABCB的结合受抑制, 从而使生长素运输减少(Kim et al., 2010; Zhu et al., 2016).Zhu等(2016)研究发现, 肌动蛋白ACTIN7与TWD1之间存在物理互作, 二者可能在不同层次控制蛋白的转运(Zhu et al., 2016). ...
... ).TWD1作为分子伴侣可通过促进AtABCB1重折叠改变其构型, 增强其ATP及底物结合能力(Geisler et al., 2004).此外, twd1突变体中被错误定位到内质网膜上的AtABCB1、AtABCB4和AtABCB19最终会被降解, 表明TWD1主要在ABCB蛋白从内质网膜到质膜的转运过程中发挥分子伴侣作用(Wu et al., 2010; Wang et al., 2013) (图2).近年, 有研究表明NPA和BUM等生长素非竞争性抑制剂可通过结合TWD1, 导致其与ABCB的结合受抑制, 从而使生长素运输减少(Kim et al., 2010; Zhu et al., 2016).Zhu等(2016)研究发现, 肌动蛋白ACTIN7与TWD1之间存在物理互作, 二者可能在不同层次控制蛋白的转运(Zhu et al., 2016). ...
TWISTED DWARF1, a unique plasma membrane-anchored immunophilin-like protein, interacts with Arabidopsis multidrug resistance-like transporters AtPGP1 and AtPGP19 1 2003
... 免疫亲和素可与ABCB蛋白互作, 间接影响生长素的运输(Geisler et al., 2004, 2016; Granzin et al., 2006).而分子伴侣TWD1最初是在酵母双杂交中被发现的与ABCB1互作的蛋白(Geisler et al., 2003).后来, 通过免疫共沉淀(Co-immunoprecipitation, Co- IP)以及生物发光能量共振转移(BRET)技术进一步证明TWD1和ABCB1在酵母及植物表达系统中存在互作(Wang et al., 2013).TWD1作为分子伴侣可通过促进AtABCB1重折叠改变其构型, 增强其ATP及底物结合能力(Geisler et al., 2004).此外, twd1突变体中被错误定位到内质网膜上的AtABCB1、AtABCB4和AtABCB19最终会被降解, 表明TWD1主要在ABCB蛋白从内质网膜到质膜的转运过程中发挥分子伴侣作用(Wu et al., 2010; Wang et al., 2013) (图2).近年, 有研究表明NPA和BUM等生长素非竞争性抑制剂可通过结合TWD1, 导致其与ABCB的结合受抑制, 从而使生长素运输减少(Kim et al., 2010; Zhu et al., 2016).Zhu等(2016)研究发现, 肌动蛋白ACTIN7与TWD1之间存在物理互作, 二者可能在不同层次控制蛋白的转运(Zhu et al., 2016). ...
Crystal structure of a multi-domain immunophilin from Arabidopsis thaliana: a paradigm for regulation of plant ABC transporters 1 2006
... 免疫亲和素可与ABCB蛋白互作, 间接影响生长素的运输(Geisler et al., 2004, 2016; Granzin et al., 2006).而分子伴侣TWD1最初是在酵母双杂交中被发现的与ABCB1互作的蛋白(Geisler et al., 2003).后来, 通过免疫共沉淀(Co-immunoprecipitation, Co- IP)以及生物发光能量共振转移(BRET)技术进一步证明TWD1和ABCB1在酵母及植物表达系统中存在互作(Wang et al., 2013).TWD1作为分子伴侣可通过促进AtABCB1重折叠改变其构型, 增强其ATP及底物结合能力(Geisler et al., 2004).此外, twd1突变体中被错误定位到内质网膜上的AtABCB1、AtABCB4和AtABCB19最终会被降解, 表明TWD1主要在ABCB蛋白从内质网膜到质膜的转运过程中发挥分子伴侣作用(Wu et al., 2010; Wang et al., 2013) (图2).近年, 有研究表明NPA和BUM等生长素非竞争性抑制剂可通过结合TWD1, 导致其与ABCB的结合受抑制, 从而使生长素运输减少(Kim et al., 2010; Zhu et al., 2016).Zhu等(2016)研究发现, 肌动蛋白ACTIN7与TWD1之间存在物理互作, 二者可能在不同层次控制蛋白的转运(Zhu et al., 2016). ...
Regulation of ABCB1/PGP1-catalysed auxin transport by linker phosphorylation 1 2012
... AGC蛋白激酶可调节ABCB转运蛋白的活性.在HeLa细胞表达系统中, 蛋白激酶AGC4/Phot1能与AtABCB19蛋白的NBD区结合使其磷酸化, 进而降低其活性(Christie et al., 2011) (图2).Co-IP和BRET实验证明, AGC3/PID与TWD1存在互作, PID通过与TWD1互作直接磷酸化AtABCB1, 从而使其生长素转运活性受抑制(Henrichs et al., 2012).然而, PID磷酸化ABCB是否必须TWD1介导目前尚不清楚.现有的研究表明AGC激酶可磷酸化ABCB, 降低其与TWD1间的互作, 从而改变ABCB蛋白跨膜转运过程中TWD1的分子伴侣作用, 影响其活性. ...
The ATP switch model for ABC transporters 1 2004
... ABC蛋白由于结构中包含有核酸结合域(nucleotide binding domain, NBD), 故又被称作ATP结合盒式蛋白.NBD含有(ABC特征基序, Walker A和B序列, H环和Q环)几个高度保守的基序(Sánchez-Fernández et al., 2001; Higgins and Linton, 2004).除了NBD外, ABC转运蛋白的结构中还包含有跨膜结构域(trans membrane domain, TMD), 每个跨膜结构域均由几个疏水性α-螺旋组成.当ABCB转运蛋白转运底物时, NBD负责结合并水解ATP, 为底物的运输提供动力; TMD则负责底物的识别及参与底物的跨膜转运.根据蛋白的结构, ABC转运蛋白可以分为3类, 即全分子转运蛋白、半分子转运蛋白以及可溶性转运蛋白(Sánchez-Fernández et al., 2001; Verrier et al., 2008).其中, ABCB亚家族蛋白只存在前两种结构(全分子和半分子转运蛋白).全分子转运蛋白包含2个NBD和2个TMD; 半分子转运蛋白则只包括1个NBD和TMD (图1). ...
A bacterial-type ABC transporter is involved in aluminum tolerance in rice 2 2009
... Biofunctions of ABCB genes in cereal crops Table 3
物种
基因名称
功能
参考文献
玉米
ZmABCB1
参与地上部生长素运输
Multani et al., 2003
高粱
SbABCB1
参与地上部生长素运输
Multani et al., 2003
小麦
TaMDR1
抗铝毒害
Sasaki et al., 2002
水稻
OsABCB14
参与根中生长素向顶运输, 铁离子平衡
Xu et al., 2014
OsABCB23
铁硫聚簇装配
Zuo et al., 2017
OsABCB27
铝胁迫响应
Huang et al., 2009
3.1 高粱和玉米中ABCB亚家族基因的研究进展 玉米ABCB亚家族蛋白共有31个成员, 包括18个全分子转运蛋白、12个半分子转运蛋白及1个仅编码不完全NBD的成员(ZmABCB19) (Pang et al., 2013).玉米ZmABCB1 (Brachytic2, BR2)和高粱SbABCB1 (Dwarf3, DW3)与拟南芥AtABCB1基因同源, 其突变可导致生长素向基运输减少, 植株矮化, 该表型与拟南芥突变体atabcb1类似, 表明在单子叶植物中ABCB蛋白可能同样发挥调控生长素运输的作用(Multani et al., 2003).此外, 玉米基因组分析显示, 与AtMDR11同源的基因ZmABCB10及与其分布位置重叠的基因ZmABCB16和ZmABCB26在生长旺盛的组织中表达, 说明这些基因也可能与生长素的运输相关(Pang et al., 2013).此外, ZmABCB7和ZmABCB8受干旱胁迫的显著诱导, ZmABCB18则受到盐胁迫的显著抑制(Pang et al., 2013). ...
... 水稻ABCB基因的研究也一直处于缓慢发展状态.水稻OsABCB14与拟南芥AtABCB19同源性较高, 且OsABCB14在水稻根尖、叶鞘、中柱、叶片、节、节间、根茎结合部和花等器官中均有表达, 其中在维管组织中的表达量最高(Xu et al., 2014).osabcb14突变体生长素的向顶运输速率降低, 且离子含量分析显示其铁离子浓度明显高于野生型, 表明OsABCB14不仅参与生长素的转运, 还与水稻铁离子的平衡密切相关(Xu et al., 2014).水稻全基因组表达分析显示, OsABCB10在花药和花粉中表达水平较高, 暗示其在花药和花粉的发育过程中发挥重要作用(Nguyen et al., 2014).非生物胁迫下的表达分析表明大部分ABCB基因的表达量发生变化.例如, OsABCB11、OsABCB8、OsABCB13、OsABCB23和OsABCB24受干旱胁迫诱导, 而OsABCB6、OsABCB9以及OsABCB8受盐胁迫诱导, 但ABCB亚家族基因在水稻非生物胁迫响应中的作用还有待进一步验证(徐杏等, 2012; Saha et al., 2015).此外, 定位于液泡膜上的TAP型转运蛋白OsABCB27 (OsALS1)参与水稻的铝胁迫响应(Huang et al., 2009), 而定位于线粒体的ATM型转运蛋白OsABCB23 (ATM3)在铁硫聚簇装配中起重要作用, 表明线粒体ATM3转运蛋白对水稻铁离子的稳态至关重要(Zuo et al., 2017). ...
Plant ABC transporters enable many unique aspects of a terrestrial plant’s lifestyle 1 2016
... ABC (ATP-binding cassette)蛋白超家族是现阶段已知最大的蛋白质家族之一, 包括ABCA、ABCB和ABCC等8个亚家族.早期ABC转运蛋白在动物的耐药性研究中取得较大进展(Theodoulou and Kerr, 2015).因此, 植物中ABC转运蛋白的初步研究也主要集中于外源物质及次生代谢物的转运在解毒和病原体防御中的作用方面(Hwang et al., 2016).随着研究的不断深入, 研究者发现植物ABC转运蛋白在生长素和重金属转运以及植物抗病性等方面同样发挥重要作用(Theodoulou and Kerr, 2015), 其亚家族ABCB转运蛋白在植物生长发育特别是生长素转运中的作用更显著. ...
The Arabidopsis ATP-binding cassette transporter ABCB21 regulates auxin levels in cotyledons, the root pericycle, and leaves 2 2019
... Lewis等(2007)发现mdr1 (abcb19)根中生长素向顶运输减少80%, 但是向基运输不受影响; mdr4 (abcb4)向基运输减少50%, 而向顶运输不受影响, 说明不同ABCB蛋白在生长素转运中可能起着截然不同的作用.此外, AtPGP4最早被报道参与生长素介导的侧根及根毛发育, 酵母中异源表达AtPGP4可导致其对NPA的敏感性升高; 且atpgp4对生长素及NPA的敏感性下降, 该突变体中游离生长素水平升高而生长素运输水平下降, 表明AtPGP4直接参与调控生长素的转运, 进而影响侧根及根毛的发育(Santelia et al., 2005).同年, Terasaka等(2005)证实, AtPGP4主要参与调控根表皮层细胞生长素的输入过程(Terasaka et al., 2005); 而Cho等(2007)研究发现, AtPGP4在拟南芥根毛细胞以及烟草(Nicotiana taba- cum)细胞中都表现出生长素输出的作用(Cho et al., 2007).Kubeš等(2012)对AtPGP4蛋白进行了研究, 发现其是一种双向生长素转运蛋白, 当细胞中生长素水平较低时促进生长素输入, 较高时则促进生长素输出.AtABCB4的同源蛋白AtABCB21与AtPGP4类似, 也是双向生长素转运蛋白(Kamimoto et al., 2012).最新研究显示, AtABCB21能够调控拟南芥子叶、根的韧皮部以及叶片中的生长素水平, 在拟南芥地上部生长素向顶运输以及植株的向光性运动中发挥作用(Jenness et al., 2019). ...
... Biofunctions of ABCB genes in Arabidopsis thaliana (Wang et al., 2017, in Chinese) Table 2
类型
基因名称
功能
参考文献
MDR (multidrug resistance)
AtABCB1/PGP1
参与地上部(花序轴、下胚轴)生长素输出, 促进下胚轴伸长
Sidler et al.,1998; Geisler et al., 2005
AtABCB4/PGP4
在拟南芥根中表现为生长素兼性转运载体, 介导侧根及根毛发育
Santelia et al., 2005; Terasaka et al., 2005; Cho et al., 2007; Lewis et al., 2007; Kubeš et al., 2012
AtABCB6
与AtABCB20在功能上冗余, 参与花序轴中生长素的向基转运
Zhang et al., 2018
AtABCB14/MDR12
参与花序轴中生长素运输, 与花序轴维管发育和气孔开闭有关
Lee et al., 2008; Kaneda et al., 2011
AtABCB15
参与花序轴中生长素运输
Kaneda et al., 2011
AtABCB19/PGP19
参与地上部生长素向基运输以及根的向顶运输, 调控不定根发育及胚后器官分离
Noh et al., 2001; Lewis et al., 2007; Sukumar et al., 2013; Zhao et al., 2013
AtABCB20
与AtABCB6在功能上冗余, 参与花序轴中生长素向基转运
Zhang et al., 2018
AtABCB21
地上部生长素兼性运输载体, 参与生长素向顶转运, 调控生长素水平
Kamimoto et al., 2012; Jenness et al., 2019
ATM (ABC transporter of mitochondria)
AtABCB23/ATM1
参与铁硫簇输出
Chen et al., 2007
AtABCB24/ATM2
未知
Chen et al., 2007
AtABCB25/ATM3
参与铁硫簇输出, 重金属抗性, 并参与调控钼辅因子合成
Kim et al., 2006; Chen et al., 2007; Bernard et al., 2009; Teschner et al., 2010
TAP (transporter associated with antigen processing)
AtABCB26/TAP1 AtABCB27/TAP2/ALS1
未知 调控铝毒环境中根的发育
Ferro et al., 2010 Larsen et al., 2007; Zhu et al., 2013
半分子转运蛋白AtABCB23 (AtATM1)、AtAB-CB24 (AtATM2)以及AtABCB25 (AtATM3)与酵母线粒体膜蛋白ATM1 (ScATM1)同源.其中, AtATM3以及AtATM1参与线粒体合成时铁硫簇的输出(Chen et al., 2007); 且AtATM3还与拟南芥的重金属抗性有关, 过表达AtATM3可提高拟南芥对镉和铅的抗性(Kim et al., 2006).TAP型转运蛋白成员AtABCB27 (ALS1/TAP2)定位于液泡膜, 在铝毒环境下根系的发 ...
Auxin trans-port is required for hypocotyl elongation in light-grown but not dark-grownArabidopsis 1 1998
... 20世纪20年代提出的Cholodny-Went理论认为, 植物向光性运动的产生是由于单侧光照射下生长素逐渐向背光一侧移动, 进而使生长素在植物茎中分布不均引起.后来该假说也被用于解释植物的向性生长.光敏色素(Phy)和隐花色素(Cry)是植物体内的光受体, 影响植物的向光性甚至光形态建成.许多研究表明, 光敏色素和隐花色素调节下胚轴顶端到根部的生长素向基运输(Jensen et al., 1998; Canamero et al., 2006; Salisbury et al., 2007).ABCB依赖光受体(PhyA、Phot和PhyB)调控光下生长素的极性运输, 进而调节下胚轴生长(Wu et al., 2010; Christie et al., 2011; 刘广超和丁兆军, 2018).Lin和Wang (2005)研究表明, atabcb1和atabcb19突变体对远红光、红光及蓝光较为敏感, 表现出下胚轴生长受抑制的表型, 说明AtABCB1和AtABCB19通过调控生长素的极性运输来影响拟南芥的光形态建成.此外, Wu等(2010)研究发现, 过表达AtABCB19可显著增加下胚轴中生长素的含量, 且红光和蓝光均可减弱其对下胚轴伸长的抑制作用.AtABCB19突变则增强了光敏色素和隐花色素诱导的拟南芥下胚轴弯曲, 说明光敏色素和隐花色素可能通过抑制AtABCB19的表达, 进而抑制下胚轴中生长素的极性运输(Nagashima et al., 2008).研究者认为上述调控模型中首先是红光通过PhyA抑制下胚轴伸长, 而后PhyB作用于AtABCB19影响幼苗中生长素的极性转运(Parks and Spalding, 1999; Wu et al., 2010). ...
Ara-bidopsis ABCB21 is a facultative auxin importer/exporter regulated by cytoplasmic auxin concentration 3 2012
... 自1992年从拟南芥(Arabidopsis thaliana)中克隆到第1个ABCB亚家族的基因——AtPGP1 (AtABCB1)以来, 植物中的ABCB亚家族研究备受关注(Dudler and Hertig, 1992).近30年的研究发现, ABCB作为转运蛋白与AUX1/LAX (AUXIN1/LIKE AUXIN)和PIN (PIN-FORMED)协同调控植物生长素的极性运输.例如, AtABCB1、AtABCB19、AtABCB4、AtABCB14和AtABCB21 (Sidler et al.,1998; Noh et al., 2001; Geisler et al., 2005; Santelia et al., 2005; Kaneda et al., 2011; Kamimoto et al., 2012)等相继被报道与生长素的运输相关.此外, 也有研究表明, ABCB转运蛋白参与植物对重金属和铝毒的抗性等过程.例如, Kim等(2016)报道了ABCB半分子转运蛋白ATM3 (AtABCB24)可提高拟南芥对镉和铅的抗性.而ALS1/TAP2 (AtABCB27)被报道参与拟南芥的铝转运, 进而提高其铝毒抗性(Larsen et al., 2007).单子叶植物玉米(Zea mays)、高粱(Sorghum bicolor)和水稻(Oryza sativa)中的ABCB蛋白也具有生长素转运功能.玉米ZmABCB1 (Brachytic2, BR2)和高粱SbABCB1 (Dwarf1, DW3)参与生长素的向基运输(Multani et al., 2003); 水稻OsABCB14则参与生长素的向顶运输(Xu et al., 2014).单子叶植物的转录组数据分析显示, ABCB亚家族蛋白可能与植物的非生物胁迫响应密切相关(Shen et al., 2010; Pang et al., 2013; Chai and Subudhi, 2016).可见, ABCB转运蛋白的功能在植物中具有一定的保守性.植物ABCB转运蛋白的数量较多, 然而目前的研究主要集中在双子叶模式植物拟南芥中, 单子叶植物中的研究尚少, 大部分ABCB转运蛋白的功能有待进一步探索.本文主要综述了ABCB亚家族蛋白在双子叶植物拟南芥以及单子叶植物玉米、高粱、小麦(Triticum aestivum)和水稻中的研究进展, 旨在为ABCB基因功能的揭示提供新线索. ...
... Lewis等(2007)发现mdr1 (abcb19)根中生长素向顶运输减少80%, 但是向基运输不受影响; mdr4 (abcb4)向基运输减少50%, 而向顶运输不受影响, 说明不同ABCB蛋白在生长素转运中可能起着截然不同的作用.此外, AtPGP4最早被报道参与生长素介导的侧根及根毛发育, 酵母中异源表达AtPGP4可导致其对NPA的敏感性升高; 且atpgp4对生长素及NPA的敏感性下降, 该突变体中游离生长素水平升高而生长素运输水平下降, 表明AtPGP4直接参与调控生长素的转运, 进而影响侧根及根毛的发育(Santelia et al., 2005).同年, Terasaka等(2005)证实, AtPGP4主要参与调控根表皮层细胞生长素的输入过程(Terasaka et al., 2005); 而Cho等(2007)研究发现, AtPGP4在拟南芥根毛细胞以及烟草(Nicotiana taba- cum)细胞中都表现出生长素输出的作用(Cho et al., 2007).Kubeš等(2012)对AtPGP4蛋白进行了研究, 发现其是一种双向生长素转运蛋白, 当细胞中生长素水平较低时促进生长素输入, 较高时则促进生长素输出.AtABCB4的同源蛋白AtABCB21与AtPGP4类似, 也是双向生长素转运蛋白(Kamimoto et al., 2012).最新研究显示, AtABCB21能够调控拟南芥子叶、根的韧皮部以及叶片中的生长素水平, 在拟南芥地上部生长素向顶运输以及植株的向光性运动中发挥作用(Jenness et al., 2019). ...
... Biofunctions of ABCB genes in Arabidopsis thaliana (Wang et al., 2017, in Chinese) Table 2
类型
基因名称
功能
参考文献
MDR (multidrug resistance)
AtABCB1/PGP1
参与地上部(花序轴、下胚轴)生长素输出, 促进下胚轴伸长
Sidler et al.,1998; Geisler et al., 2005
AtABCB4/PGP4
在拟南芥根中表现为生长素兼性转运载体, 介导侧根及根毛发育
Santelia et al., 2005; Terasaka et al., 2005; Cho et al., 2007; Lewis et al., 2007; Kubeš et al., 2012
AtABCB6
与AtABCB20在功能上冗余, 参与花序轴中生长素的向基转运
Zhang et al., 2018
AtABCB14/MDR12
参与花序轴中生长素运输, 与花序轴维管发育和气孔开闭有关
Lee et al., 2008; Kaneda et al., 2011
AtABCB15
参与花序轴中生长素运输
Kaneda et al., 2011
AtABCB19/PGP19
参与地上部生长素向基运输以及根的向顶运输, 调控不定根发育及胚后器官分离
Noh et al., 2001; Lewis et al., 2007; Sukumar et al., 2013; Zhao et al., 2013
AtABCB20
与AtABCB6在功能上冗余, 参与花序轴中生长素向基转运
Zhang et al., 2018
AtABCB21
地上部生长素兼性运输载体, 参与生长素向顶转运, 调控生长素水平
Kamimoto et al., 2012; Jenness et al., 2019
ATM (ABC transporter of mitochondria)
AtABCB23/ATM1
参与铁硫簇输出
Chen et al., 2007
AtABCB24/ATM2
未知
Chen et al., 2007
AtABCB25/ATM3
参与铁硫簇输出, 重金属抗性, 并参与调控钼辅因子合成
Kim et al., 2006; Chen et al., 2007; Bernard et al., 2009; Teschner et al., 2010
TAP (transporter associated with antigen processing)
AtABCB26/TAP1 AtABCB27/TAP2/ALS1
未知 调控铝毒环境中根的发育
Ferro et al., 2010 Larsen et al., 2007; Zhu et al., 2013
半分子转运蛋白AtABCB23 (AtATM1)、AtAB-CB24 (AtATM2)以及AtABCB25 (AtATM3)与酵母线粒体膜蛋白ATM1 (ScATM1)同源.其中, AtATM3以及AtATM1参与线粒体合成时铁硫簇的输出(Chen et al., 2007); 且AtATM3还与拟南芥的重金属抗性有关, 过表达AtATM3可提高拟南芥对镉和铅的抗性(Kim et al., 2006).TAP型转运蛋白成员AtABCB27 (ALS1/TAP2)定位于液泡膜, 在铝毒环境下根系的发 ...
ABC transporters coordinately expressed during lignification of Arabidopsis stems include a set of ABCBs associated with auxin transport 5 2011
... 自1992年从拟南芥(Arabidopsis thaliana)中克隆到第1个ABCB亚家族的基因——AtPGP1 (AtABCB1)以来, 植物中的ABCB亚家族研究备受关注(Dudler and Hertig, 1992).近30年的研究发现, ABCB作为转运蛋白与AUX1/LAX (AUXIN1/LIKE AUXIN)和PIN (PIN-FORMED)协同调控植物生长素的极性运输.例如, AtABCB1、AtABCB19、AtABCB4、AtABCB14和AtABCB21 (Sidler et al.,1998; Noh et al., 2001; Geisler et al., 2005; Santelia et al., 2005; Kaneda et al., 2011; Kamimoto et al., 2012)等相继被报道与生长素的运输相关.此外, 也有研究表明, ABCB转运蛋白参与植物对重金属和铝毒的抗性等过程.例如, Kim等(2016)报道了ABCB半分子转运蛋白ATM3 (AtABCB24)可提高拟南芥对镉和铅的抗性.而ALS1/TAP2 (AtABCB27)被报道参与拟南芥的铝转运, 进而提高其铝毒抗性(Larsen et al., 2007).单子叶植物玉米(Zea mays)、高粱(Sorghum bicolor)和水稻(Oryza sativa)中的ABCB蛋白也具有生长素转运功能.玉米ZmABCB1 (Brachytic2, BR2)和高粱SbABCB1 (Dwarf1, DW3)参与生长素的向基运输(Multani et al., 2003); 水稻OsABCB14则参与生长素的向顶运输(Xu et al., 2014).单子叶植物的转录组数据分析显示, ABCB亚家族蛋白可能与植物的非生物胁迫响应密切相关(Shen et al., 2010; Pang et al., 2013; Chai and Subudhi, 2016).可见, ABCB转运蛋白的功能在植物中具有一定的保守性.植物ABCB转运蛋白的数量较多, 然而目前的研究主要集中在双子叶模式植物拟南芥中, 单子叶植物中的研究尚少, 大部分ABCB转运蛋白的功能有待进一步探索.本文主要综述了ABCB亚家族蛋白在双子叶植物拟南芥以及单子叶植物玉米、高粱、小麦(Triticum aestivum)和水稻中的研究进展, 旨在为ABCB基因功能的揭示提供新线索. ...
... ABCB亚家族成员在植物生长发育的各个方面发挥不同作用(表2).植物根切除导致吲哚-3-乙酸(IAA)转运快速增加, 并在切除点以上的下胚轴中积累, 进而诱导不定根的形成.Sukumar等(2013)研究发现, 相对于野生型株系atabcb19突变体的不定根数量显著减少, 过表达株系则出现较多的不定根, 且AtABCB19的转录及蛋白表达水平与局部IAA积累的时空变化模式密切相关, 因此认为AtABCB19的局部诱导可促进拟南芥不定根的形成.此外, 有研究表明, AtABCB19还参与调控拟南芥胚后器官的分离(Zhao et al., 2013)及花序轴的细胞质流(Okamoto et al., 2016).AtABCB1在花药发育过程中起主要作用, AtABCB19则起协同作用(Cecchetti et al., 2015).Kaneda等(2011)发现, atabcb14花序轴中维管发育不正常, 说明AtABCB14可能参与拟南芥花序轴的木质化过程.而之前有报道显示, AtABCB14可将苹果酸盐从质外体转运至保卫细胞, 调节渗透压, 从而调控细胞对二氧化碳的响应, 影响气孔开闭(Lee et al., 2008).总之, ABCB蛋白功能复杂多样, 仍有待进一步研究. ...
... Biofunctions of ABCB genes in Arabidopsis thaliana (Wang et al., 2017, in Chinese) Table 2
类型
基因名称
功能
参考文献
MDR (multidrug resistance)
AtABCB1/PGP1
参与地上部(花序轴、下胚轴)生长素输出, 促进下胚轴伸长
Sidler et al.,1998; Geisler et al., 2005
AtABCB4/PGP4
在拟南芥根中表现为生长素兼性转运载体, 介导侧根及根毛发育
Santelia et al., 2005; Terasaka et al., 2005; Cho et al., 2007; Lewis et al., 2007; Kubeš et al., 2012
AtABCB6
与AtABCB20在功能上冗余, 参与花序轴中生长素的向基转运
Zhang et al., 2018
AtABCB14/MDR12
参与花序轴中生长素运输, 与花序轴维管发育和气孔开闭有关
Lee et al., 2008; Kaneda et al., 2011
AtABCB15
参与花序轴中生长素运输
Kaneda et al., 2011
AtABCB19/PGP19
参与地上部生长素向基运输以及根的向顶运输, 调控不定根发育及胚后器官分离
Noh et al., 2001; Lewis et al., 2007; Sukumar et al., 2013; Zhao et al., 2013
AtABCB20
与AtABCB6在功能上冗余, 参与花序轴中生长素向基转运
Zhang et al., 2018
AtABCB21
地上部生长素兼性运输载体, 参与生长素向顶转运, 调控生长素水平
Kamimoto et al., 2012; Jenness et al., 2019
ATM (ABC transporter of mitochondria)
AtABCB23/ATM1
参与铁硫簇输出
Chen et al., 2007
AtABCB24/ATM2
未知
Chen et al., 2007
AtABCB25/ATM3
参与铁硫簇输出, 重金属抗性, 并参与调控钼辅因子合成
Kim et al., 2006; Chen et al., 2007; Bernard et al., 2009; Teschner et al., 2010
TAP (transporter associated with antigen processing)
AtABCB26/TAP1 AtABCB27/TAP2/ALS1
未知 调控铝毒环境中根的发育
Ferro et al., 2010 Larsen et al., 2007; Zhu et al., 2013
半分子转运蛋白AtABCB23 (AtATM1)、AtAB-CB24 (AtATM2)以及AtABCB25 (AtATM3)与酵母线粒体膜蛋白ATM1 (ScATM1)同源.其中, AtATM3以及AtATM1参与线粒体合成时铁硫簇的输出(Chen et al., 2007); 且AtATM3还与拟南芥的重金属抗性有关, 过表达AtATM3可提高拟南芥对镉和铅的抗性(Kim et al., 2006).TAP型转运蛋白成员AtABCB27 (ALS1/TAP2)定位于液泡膜, 在铝毒环境下根系的发 ...
... Kaneda et al., 2011
AtABCB19/PGP19
参与地上部生长素向基运输以及根的向顶运输, 调控不定根发育及胚后器官分离
Noh et al., 2001; Lewis et al., 2007; Sukumar et al., 2013; Zhao et al., 2013
AtABCB20
与AtABCB6在功能上冗余, 参与花序轴中生长素向基转运
Zhang et al., 2018
AtABCB21
地上部生长素兼性运输载体, 参与生长素向顶转运, 调控生长素水平
Kamimoto et al., 2012; Jenness et al., 2019
ATM (ABC transporter of mitochondria)
AtABCB23/ATM1
参与铁硫簇输出
Chen et al., 2007
AtABCB24/ATM2
未知
Chen et al., 2007
AtABCB25/ATM3
参与铁硫簇输出, 重金属抗性, 并参与调控钼辅因子合成
Kim et al., 2006; Chen et al., 2007; Bernard et al., 2009; Teschner et al., 2010
TAP (transporter associated with antigen processing)
AtABCB26/TAP1 AtABCB27/TAP2/ALS1
未知 调控铝毒环境中根的发育
Ferro et al., 2010 Larsen et al., 2007; Zhu et al., 2013
半分子转运蛋白AtABCB23 (AtATM1)、AtAB-CB24 (AtATM2)以及AtABCB25 (AtATM3)与酵母线粒体膜蛋白ATM1 (ScATM1)同源.其中, AtATM3以及AtATM1参与线粒体合成时铁硫簇的输出(Chen et al., 2007); 且AtATM3还与拟南芥的重金属抗性有关, 过表达AtATM3可提高拟南芥对镉和铅的抗性(Kim et al., 2006).TAP型转运蛋白成员AtABCB27 (ALS1/TAP2)定位于液泡膜, 在铝毒环境下根系的发 ...
AtATM3 is involved in heavy metal resistance in Arabidopsis 3 2006
... 自1992年从拟南芥(Arabidopsis thaliana)中克隆到第1个ABCB亚家族的基因——AtPGP1 (AtABCB1)以来, 植物中的ABCB亚家族研究备受关注(Dudler and Hertig, 1992).近30年的研究发现, ABCB作为转运蛋白与AUX1/LAX (AUXIN1/LIKE AUXIN)和PIN (PIN-FORMED)协同调控植物生长素的极性运输.例如, AtABCB1、AtABCB19、AtABCB4、AtABCB14和AtABCB21 (Sidler et al.,1998; Noh et al., 2001; Geisler et al., 2005; Santelia et al., 2005; Kaneda et al., 2011; Kamimoto et al., 2012)等相继被报道与生长素的运输相关.此外, 也有研究表明, ABCB转运蛋白参与植物对重金属和铝毒的抗性等过程.例如, Kim等(2016)报道了ABCB半分子转运蛋白ATM3 (AtABCB24)可提高拟南芥对镉和铅的抗性.而ALS1/TAP2 (AtABCB27)被报道参与拟南芥的铝转运, 进而提高其铝毒抗性(Larsen et al., 2007).单子叶植物玉米(Zea mays)、高粱(Sorghum bicolor)和水稻(Oryza sativa)中的ABCB蛋白也具有生长素转运功能.玉米ZmABCB1 (Brachytic2, BR2)和高粱SbABCB1 (Dwarf1, DW3)参与生长素的向基运输(Multani et al., 2003); 水稻OsABCB14则参与生长素的向顶运输(Xu et al., 2014).单子叶植物的转录组数据分析显示, ABCB亚家族蛋白可能与植物的非生物胁迫响应密切相关(Shen et al., 2010; Pang et al., 2013; Chai and Subudhi, 2016).可见, ABCB转运蛋白的功能在植物中具有一定的保守性.植物ABCB转运蛋白的数量较多, 然而目前的研究主要集中在双子叶模式植物拟南芥中, 单子叶植物中的研究尚少, 大部分ABCB转运蛋白的功能有待进一步探索.本文主要综述了ABCB亚家族蛋白在双子叶植物拟南芥以及单子叶植物玉米、高粱、小麦(Triticum aestivum)和水稻中的研究进展, 旨在为ABCB基因功能的揭示提供新线索. ...
... Biofunctions of ABCB genes in Arabidopsis thaliana (Wang et al., 2017, in Chinese) Table 2
类型
基因名称
功能
参考文献
MDR (multidrug resistance)
AtABCB1/PGP1
参与地上部(花序轴、下胚轴)生长素输出, 促进下胚轴伸长
Sidler et al.,1998; Geisler et al., 2005
AtABCB4/PGP4
在拟南芥根中表现为生长素兼性转运载体, 介导侧根及根毛发育
Santelia et al., 2005; Terasaka et al., 2005; Cho et al., 2007; Lewis et al., 2007; Kubeš et al., 2012
AtABCB6
与AtABCB20在功能上冗余, 参与花序轴中生长素的向基转运
Zhang et al., 2018
AtABCB14/MDR12
参与花序轴中生长素运输, 与花序轴维管发育和气孔开闭有关
Lee et al., 2008; Kaneda et al., 2011
AtABCB15
参与花序轴中生长素运输
Kaneda et al., 2011
AtABCB19/PGP19
参与地上部生长素向基运输以及根的向顶运输, 调控不定根发育及胚后器官分离
Noh et al., 2001; Lewis et al., 2007; Sukumar et al., 2013; Zhao et al., 2013
AtABCB20
与AtABCB6在功能上冗余, 参与花序轴中生长素向基转运
Zhang et al., 2018
AtABCB21
地上部生长素兼性运输载体, 参与生长素向顶转运, 调控生长素水平
Kamimoto et al., 2012; Jenness et al., 2019
ATM (ABC transporter of mitochondria)
AtABCB23/ATM1
参与铁硫簇输出
Chen et al., 2007
AtABCB24/ATM2
未知
Chen et al., 2007
AtABCB25/ATM3
参与铁硫簇输出, 重金属抗性, 并参与调控钼辅因子合成
Kim et al., 2006; Chen et al., 2007; Bernard et al., 2009; Teschner et al., 2010
TAP (transporter associated with antigen processing)
AtABCB26/TAP1 AtABCB27/TAP2/ALS1
未知 调控铝毒环境中根的发育
Ferro et al., 2010 Larsen et al., 2007; Zhu et al., 2013
半分子转运蛋白AtABCB23 (AtATM1)、AtAB-CB24 (AtATM2)以及AtABCB25 (AtATM3)与酵母线粒体膜蛋白ATM1 (ScATM1)同源.其中, AtATM3以及AtATM1参与线粒体合成时铁硫簇的输出(Chen et al., 2007); 且AtATM3还与拟南芥的重金属抗性有关, 过表达AtATM3可提高拟南芥对镉和铅的抗性(Kim et al., 2006).TAP型转运蛋白成员AtABCB27 (ALS1/TAP2)定位于液泡膜, 在铝毒环境下根系的发 ...
... 半分子转运蛋白AtABCB23 (AtATM1)、AtAB-CB24 (AtATM2)以及AtABCB25 (AtATM3)与酵母线粒体膜蛋白ATM1 (ScATM1)同源.其中, AtATM3以及AtATM1参与线粒体合成时铁硫簇的输出(Chen et al., 2007); 且AtATM3还与拟南芥的重金属抗性有关, 过表达AtATM3可提高拟南芥对镉和铅的抗性(Kim et al., 2006).TAP型转运蛋白成员AtABCB27 (ALS1/TAP2)定位于液泡膜, 在铝毒环境下根系的发 ...
Identification of an ABCB/P-glycoprotein-specific inhibitor of auxin transport by chemical genomics 2 2010
... 免疫亲和素可与ABCB蛋白互作, 间接影响生长素的运输(Geisler et al., 2004, 2016; Granzin et al., 2006).而分子伴侣TWD1最初是在酵母双杂交中被发现的与ABCB1互作的蛋白(Geisler et al., 2003).后来, 通过免疫共沉淀(Co-immunoprecipitation, Co- IP)以及生物发光能量共振转移(BRET)技术进一步证明TWD1和ABCB1在酵母及植物表达系统中存在互作(Wang et al., 2013).TWD1作为分子伴侣可通过促进AtABCB1重折叠改变其构型, 增强其ATP及底物结合能力(Geisler et al., 2004).此外, twd1突变体中被错误定位到内质网膜上的AtABCB1、AtABCB4和AtABCB19最终会被降解, 表明TWD1主要在ABCB蛋白从内质网膜到质膜的转运过程中发挥分子伴侣作用(Wu et al., 2010; Wang et al., 2013) (图2).近年, 有研究表明NPA和BUM等生长素非竞争性抑制剂可通过结合TWD1, 导致其与ABCB的结合受抑制, 从而使生长素运输减少(Kim et al., 2010; Zhu et al., 2016).Zhu等(2016)研究发现, 肌动蛋白ACTIN7与TWD1之间存在物理互作, 二者可能在不同层次控制蛋白的转运(Zhu et al., 2016). ...
... PIN与ABCB蛋白存在互作.植物生长素极性运输主要依赖ABCB、PIN以及AUX1/LAX转运蛋白家族.目前, 对PIN蛋白家族的研究最为深入.PIN的一个重要特征是亚细胞极性定位.Blakeslee等(2007)研究表明, PIN在极性胚胎发育和器官形成等需要定向生长素极性运输的过程中起决定作用, ABCB则在顶端组织生长素转运及长距离运输中起重要作用, 二者在调控生长素的转运上具有一定的独立性.另有研究发现, ABCB1和ABCB19在拟南芥的茎尖中与PIN1共定位, 在根中与PIN1和PIN2共定位(Bandyopadhyay et al., 2007).酵母双杂交和免疫共沉淀实验均表明, ABCB1、ABCB19与PIN1、PIN2间存在物理互作(Blakeslee et al., 2007; Kim et al., 2010; Geisler et al., 2017); 且共表达PIN1-ABCB1和PIN1-ABCB19的植株均表现出生长素输出率及底物特异性提高(Blakeslee et al., 2007) (图2).尽管Geisler等(2017)提出PIN与ABCB互作的3种假设: (1) PIN发挥运输作用, ABCB发挥调节作用; (2) ABCB发挥运输作用, PIN发挥调节作用; (3) PIN和ABCB不单独发挥转运作用, 而是形成复合体行使转运功能, 但具体机制仍不清楚, 尚需进一步探索. ...
The Arabidopsis concentration-dependent influx/efflux transporter ABCB4 regulates cellular auxin levels in the root epidermis 2 2012
... Lewis等(2007)发现mdr1 (abcb19)根中生长素向顶运输减少80%, 但是向基运输不受影响; mdr4 (abcb4)向基运输减少50%, 而向顶运输不受影响, 说明不同ABCB蛋白在生长素转运中可能起着截然不同的作用.此外, AtPGP4最早被报道参与生长素介导的侧根及根毛发育, 酵母中异源表达AtPGP4可导致其对NPA的敏感性升高; 且atpgp4对生长素及NPA的敏感性下降, 该突变体中游离生长素水平升高而生长素运输水平下降, 表明AtPGP4直接参与调控生长素的转运, 进而影响侧根及根毛的发育(Santelia et al., 2005).同年, Terasaka等(2005)证实, AtPGP4主要参与调控根表皮层细胞生长素的输入过程(Terasaka et al., 2005); 而Cho等(2007)研究发现, AtPGP4在拟南芥根毛细胞以及烟草(Nicotiana taba- cum)细胞中都表现出生长素输出的作用(Cho et al., 2007).Kubeš等(2012)对AtPGP4蛋白进行了研究, 发现其是一种双向生长素转运蛋白, 当细胞中生长素水平较低时促进生长素输入, 较高时则促进生长素输出.AtABCB4的同源蛋白AtABCB21与AtPGP4类似, 也是双向生长素转运蛋白(Kamimoto et al., 2012).最新研究显示, AtABCB21能够调控拟南芥子叶、根的韧皮部以及叶片中的生长素水平, 在拟南芥地上部生长素向顶运输以及植株的向光性运动中发挥作用(Jenness et al., 2019). ...
... Biofunctions of ABCB genes in Arabidopsis thaliana (Wang et al., 2017, in Chinese) Table 2
类型
基因名称
功能
参考文献
MDR (multidrug resistance)
AtABCB1/PGP1
参与地上部(花序轴、下胚轴)生长素输出, 促进下胚轴伸长
Sidler et al.,1998; Geisler et al., 2005
AtABCB4/PGP4
在拟南芥根中表现为生长素兼性转运载体, 介导侧根及根毛发育
Santelia et al., 2005; Terasaka et al., 2005; Cho et al., 2007; Lewis et al., 2007; Kubeš et al., 2012
AtABCB6
与AtABCB20在功能上冗余, 参与花序轴中生长素的向基转运
Zhang et al., 2018
AtABCB14/MDR12
参与花序轴中生长素运输, 与花序轴维管发育和气孔开闭有关
Lee et al., 2008; Kaneda et al., 2011
AtABCB15
参与花序轴中生长素运输
Kaneda et al., 2011
AtABCB19/PGP19
参与地上部生长素向基运输以及根的向顶运输, 调控不定根发育及胚后器官分离
Noh et al., 2001; Lewis et al., 2007; Sukumar et al., 2013; Zhao et al., 2013
AtABCB20
与AtABCB6在功能上冗余, 参与花序轴中生长素向基转运
Zhang et al., 2018
AtABCB21
地上部生长素兼性运输载体, 参与生长素向顶转运, 调控生长素水平
Kamimoto et al., 2012; Jenness et al., 2019
ATM (ABC transporter of mitochondria)
AtABCB23/ATM1
参与铁硫簇输出
Chen et al., 2007
AtABCB24/ATM2
未知
Chen et al., 2007
AtABCB25/ATM3
参与铁硫簇输出, 重金属抗性, 并参与调控钼辅因子合成
Kim et al., 2006; Chen et al., 2007; Bernard et al., 2009; Teschner et al., 2010
TAP (transporter associated with antigen processing)
AtABCB26/TAP1 AtABCB27/TAP2/ALS1
未知 调控铝毒环境中根的发育
Ferro et al., 2010 Larsen et al., 2007; Zhu et al., 2013
半分子转运蛋白AtABCB23 (AtATM1)、AtAB-CB24 (AtATM2)以及AtABCB25 (AtATM3)与酵母线粒体膜蛋白ATM1 (ScATM1)同源.其中, AtATM3以及AtATM1参与线粒体合成时铁硫簇的输出(Chen et al., 2007); 且AtATM3还与拟南芥的重金属抗性有关, 过表达AtATM3可提高拟南芥对镉和铅的抗性(Kim et al., 2006).TAP型转运蛋白成员AtABCB27 (ALS1/TAP2)定位于液泡膜, 在铝毒环境下根系的发 ...
Arabidopsis ALS1 encodes a root tip and stele localized half type ABC transporter required for root growth in an aluminum toxic environment 3 2007
... 自1992年从拟南芥(Arabidopsis thaliana)中克隆到第1个ABCB亚家族的基因——AtPGP1 (AtABCB1)以来, 植物中的ABCB亚家族研究备受关注(Dudler and Hertig, 1992).近30年的研究发现, ABCB作为转运蛋白与AUX1/LAX (AUXIN1/LIKE AUXIN)和PIN (PIN-FORMED)协同调控植物生长素的极性运输.例如, AtABCB1、AtABCB19、AtABCB4、AtABCB14和AtABCB21 (Sidler et al.,1998; Noh et al., 2001; Geisler et al., 2005; Santelia et al., 2005; Kaneda et al., 2011; Kamimoto et al., 2012)等相继被报道与生长素的运输相关.此外, 也有研究表明, ABCB转运蛋白参与植物对重金属和铝毒的抗性等过程.例如, Kim等(2016)报道了ABCB半分子转运蛋白ATM3 (AtABCB24)可提高拟南芥对镉和铅的抗性.而ALS1/TAP2 (AtABCB27)被报道参与拟南芥的铝转运, 进而提高其铝毒抗性(Larsen et al., 2007).单子叶植物玉米(Zea mays)、高粱(Sorghum bicolor)和水稻(Oryza sativa)中的ABCB蛋白也具有生长素转运功能.玉米ZmABCB1 (Brachytic2, BR2)和高粱SbABCB1 (Dwarf1, DW3)参与生长素的向基运输(Multani et al., 2003); 水稻OsABCB14则参与生长素的向顶运输(Xu et al., 2014).单子叶植物的转录组数据分析显示, ABCB亚家族蛋白可能与植物的非生物胁迫响应密切相关(Shen et al., 2010; Pang et al., 2013; Chai and Subudhi, 2016).可见, ABCB转运蛋白的功能在植物中具有一定的保守性.植物ABCB转运蛋白的数量较多, 然而目前的研究主要集中在双子叶模式植物拟南芥中, 单子叶植物中的研究尚少, 大部分ABCB转运蛋白的功能有待进一步探索.本文主要综述了ABCB亚家族蛋白在双子叶植物拟南芥以及单子叶植物玉米、高粱、小麦(Triticum aestivum)和水稻中的研究进展, 旨在为ABCB基因功能的揭示提供新线索. ...
... Biofunctions of ABCB genes in Arabidopsis thaliana (Wang et al., 2017, in Chinese) Table 2
类型
基因名称
功能
参考文献
MDR (multidrug resistance)
AtABCB1/PGP1
参与地上部(花序轴、下胚轴)生长素输出, 促进下胚轴伸长
Sidler et al.,1998; Geisler et al., 2005
AtABCB4/PGP4
在拟南芥根中表现为生长素兼性转运载体, 介导侧根及根毛发育
Santelia et al., 2005; Terasaka et al., 2005; Cho et al., 2007; Lewis et al., 2007; Kubeš et al., 2012
AtABCB6
与AtABCB20在功能上冗余, 参与花序轴中生长素的向基转运
Zhang et al., 2018
AtABCB14/MDR12
参与花序轴中生长素运输, 与花序轴维管发育和气孔开闭有关
Lee et al., 2008; Kaneda et al., 2011
AtABCB15
参与花序轴中生长素运输
Kaneda et al., 2011
AtABCB19/PGP19
参与地上部生长素向基运输以及根的向顶运输, 调控不定根发育及胚后器官分离
Noh et al., 2001; Lewis et al., 2007; Sukumar et al., 2013; Zhao et al., 2013
AtABCB20
与AtABCB6在功能上冗余, 参与花序轴中生长素向基转运
Zhang et al., 2018
AtABCB21
地上部生长素兼性运输载体, 参与生长素向顶转运, 调控生长素水平
Kamimoto et al., 2012; Jenness et al., 2019
ATM (ABC transporter of mitochondria)
AtABCB23/ATM1
参与铁硫簇输出
Chen et al., 2007
AtABCB24/ATM2
未知
Chen et al., 2007
AtABCB25/ATM3
参与铁硫簇输出, 重金属抗性, 并参与调控钼辅因子合成
Kim et al., 2006; Chen et al., 2007; Bernard et al., 2009; Teschner et al., 2010
TAP (transporter associated with antigen processing)
AtABCB26/TAP1 AtABCB27/TAP2/ALS1
未知 调控铝毒环境中根的发育
Ferro et al., 2010 Larsen et al., 2007; Zhu et al., 2013
半分子转运蛋白AtABCB23 (AtATM1)、AtAB-CB24 (AtATM2)以及AtABCB25 (AtATM3)与酵母线粒体膜蛋白ATM1 (ScATM1)同源.其中, AtATM3以及AtATM1参与线粒体合成时铁硫簇的输出(Chen et al., 2007); 且AtATM3还与拟南芥的重金属抗性有关, 过表达AtATM3可提高拟南芥对镉和铅的抗性(Kim et al., 2006).TAP型转运蛋白成员AtABCB27 (ALS1/TAP2)定位于液泡膜, 在铝毒环境下根系的发 ...
... 育方面发挥重要作用(Larsen et al., 2007; Zhu et al., 2013); AtABCB26 (TAP1)则定位于叶绿体膜, 具体功能未知(Ferro et al., 2010). ...
The ABC transporter AtABCB14 is a malate importer and modulates stomatal response to CO2 2 2008
... ABCB亚家族成员在植物生长发育的各个方面发挥不同作用(表2).植物根切除导致吲哚-3-乙酸(IAA)转运快速增加, 并在切除点以上的下胚轴中积累, 进而诱导不定根的形成.Sukumar等(2013)研究发现, 相对于野生型株系atabcb19突变体的不定根数量显著减少, 过表达株系则出现较多的不定根, 且AtABCB19的转录及蛋白表达水平与局部IAA积累的时空变化模式密切相关, 因此认为AtABCB19的局部诱导可促进拟南芥不定根的形成.此外, 有研究表明, AtABCB19还参与调控拟南芥胚后器官的分离(Zhao et al., 2013)及花序轴的细胞质流(Okamoto et al., 2016).AtABCB1在花药发育过程中起主要作用, AtABCB19则起协同作用(Cecchetti et al., 2015).Kaneda等(2011)发现, atabcb14花序轴中维管发育不正常, 说明AtABCB14可能参与拟南芥花序轴的木质化过程.而之前有报道显示, AtABCB14可将苹果酸盐从质外体转运至保卫细胞, 调节渗透压, 从而调控细胞对二氧化碳的响应, 影响气孔开闭(Lee et al., 2008).总之, ABCB蛋白功能复杂多样, 仍有待进一步研究. ...
... Biofunctions of ABCB genes in Arabidopsis thaliana (Wang et al., 2017, in Chinese) Table 2
类型
基因名称
功能
参考文献
MDR (multidrug resistance)
AtABCB1/PGP1
参与地上部(花序轴、下胚轴)生长素输出, 促进下胚轴伸长
Sidler et al.,1998; Geisler et al., 2005
AtABCB4/PGP4
在拟南芥根中表现为生长素兼性转运载体, 介导侧根及根毛发育
Santelia et al., 2005; Terasaka et al., 2005; Cho et al., 2007; Lewis et al., 2007; Kubeš et al., 2012
AtABCB6
与AtABCB20在功能上冗余, 参与花序轴中生长素的向基转运
Zhang et al., 2018
AtABCB14/MDR12
参与花序轴中生长素运输, 与花序轴维管发育和气孔开闭有关
Lee et al., 2008; Kaneda et al., 2011
AtABCB15
参与花序轴中生长素运输
Kaneda et al., 2011
AtABCB19/PGP19
参与地上部生长素向基运输以及根的向顶运输, 调控不定根发育及胚后器官分离
Noh et al., 2001; Lewis et al., 2007; Sukumar et al., 2013; Zhao et al., 2013
AtABCB20
与AtABCB6在功能上冗余, 参与花序轴中生长素向基转运
Zhang et al., 2018
AtABCB21
地上部生长素兼性运输载体, 参与生长素向顶转运, 调控生长素水平
Kamimoto et al., 2012; Jenness et al., 2019
ATM (ABC transporter of mitochondria)
AtABCB23/ATM1
参与铁硫簇输出
Chen et al., 2007
AtABCB24/ATM2
未知
Chen et al., 2007
AtABCB25/ATM3
参与铁硫簇输出, 重金属抗性, 并参与调控钼辅因子合成
Kim et al., 2006; Chen et al., 2007; Bernard et al., 2009; Teschner et al., 2010
TAP (transporter associated with antigen processing)
AtABCB26/TAP1 AtABCB27/TAP2/ALS1
未知 调控铝毒环境中根的发育
Ferro et al., 2010 Larsen et al., 2007; Zhu et al., 2013
半分子转运蛋白AtABCB23 (AtATM1)、AtAB-CB24 (AtATM2)以及AtABCB25 (AtATM3)与酵母线粒体膜蛋白ATM1 (ScATM1)同源.其中, AtATM3以及AtATM1参与线粒体合成时铁硫簇的输出(Chen et al., 2007); 且AtATM3还与拟南芥的重金属抗性有关, 过表达AtATM3可提高拟南芥对镉和铅的抗性(Kim et al., 2006).TAP型转运蛋白成员AtABCB27 (ALS1/TAP2)定位于液泡膜, 在铝毒环境下根系的发 ...
Separating the roles of acropetal and basipetal auxin transport on gravitropism with mutations in two Arabidopsis multidrug resistance-like ABC transporter genes 3 2007
... Lewis等(2007)发现mdr1 (abcb19)根中生长素向顶运输减少80%, 但是向基运输不受影响; mdr4 (abcb4)向基运输减少50%, 而向顶运输不受影响, 说明不同ABCB蛋白在生长素转运中可能起着截然不同的作用.此外, AtPGP4最早被报道参与生长素介导的侧根及根毛发育, 酵母中异源表达AtPGP4可导致其对NPA的敏感性升高; 且atpgp4对生长素及NPA的敏感性下降, 该突变体中游离生长素水平升高而生长素运输水平下降, 表明AtPGP4直接参与调控生长素的转运, 进而影响侧根及根毛的发育(Santelia et al., 2005).同年, Terasaka等(2005)证实, AtPGP4主要参与调控根表皮层细胞生长素的输入过程(Terasaka et al., 2005); 而Cho等(2007)研究发现, AtPGP4在拟南芥根毛细胞以及烟草(Nicotiana taba- cum)细胞中都表现出生长素输出的作用(Cho et al., 2007).Kubeš等(2012)对AtPGP4蛋白进行了研究, 发现其是一种双向生长素转运蛋白, 当细胞中生长素水平较低时促进生长素输入, 较高时则促进生长素输出.AtABCB4的同源蛋白AtABCB21与AtPGP4类似, 也是双向生长素转运蛋白(Kamimoto et al., 2012).最新研究显示, AtABCB21能够调控拟南芥子叶、根的韧皮部以及叶片中的生长素水平, 在拟南芥地上部生长素向顶运输以及植株的向光性运动中发挥作用(Jenness et al., 2019). ...
... Biofunctions of ABCB genes in Arabidopsis thaliana (Wang et al., 2017, in Chinese) Table 2
类型
基因名称
功能
参考文献
MDR (multidrug resistance)
AtABCB1/PGP1
参与地上部(花序轴、下胚轴)生长素输出, 促进下胚轴伸长
Sidler et al.,1998; Geisler et al., 2005
AtABCB4/PGP4
在拟南芥根中表现为生长素兼性转运载体, 介导侧根及根毛发育
Santelia et al., 2005; Terasaka et al., 2005; Cho et al., 2007; Lewis et al., 2007; Kubeš et al., 2012
AtABCB6
与AtABCB20在功能上冗余, 参与花序轴中生长素的向基转运
Zhang et al., 2018
AtABCB14/MDR12
参与花序轴中生长素运输, 与花序轴维管发育和气孔开闭有关
Lee et al., 2008; Kaneda et al., 2011
AtABCB15
参与花序轴中生长素运输
Kaneda et al., 2011
AtABCB19/PGP19
参与地上部生长素向基运输以及根的向顶运输, 调控不定根发育及胚后器官分离
Noh et al., 2001; Lewis et al., 2007; Sukumar et al., 2013; Zhao et al., 2013
AtABCB20
与AtABCB6在功能上冗余, 参与花序轴中生长素向基转运
Zhang et al., 2018
AtABCB21
地上部生长素兼性运输载体, 参与生长素向顶转运, 调控生长素水平
Kamimoto et al., 2012; Jenness et al., 2019
ATM (ABC transporter of mitochondria)
AtABCB23/ATM1
参与铁硫簇输出
Chen et al., 2007
AtABCB24/ATM2
未知
Chen et al., 2007
AtABCB25/ATM3
参与铁硫簇输出, 重金属抗性, 并参与调控钼辅因子合成
Kim et al., 2006; Chen et al., 2007; Bernard et al., 2009; Teschner et al., 2010
TAP (transporter associated with antigen processing)
AtABCB26/TAP1 AtABCB27/TAP2/ALS1
未知 调控铝毒环境中根的发育
Ferro et al., 2010 Larsen et al., 2007; Zhu et al., 2013
半分子转运蛋白AtABCB23 (AtATM1)、AtAB-CB24 (AtATM2)以及AtABCB25 (AtATM3)与酵母线粒体膜蛋白ATM1 (ScATM1)同源.其中, AtATM3以及AtATM1参与线粒体合成时铁硫簇的输出(Chen et al., 2007); 且AtATM3还与拟南芥的重金属抗性有关, 过表达AtATM3可提高拟南芥对镉和铅的抗性(Kim et al., 2006).TAP型转运蛋白成员AtABCB27 (ALS1/TAP2)定位于液泡膜, 在铝毒环境下根系的发 ...
... ; Lewis et al., 2007; Sukumar et al., 2013; Zhao et al., 2013
AtABCB20
与AtABCB6在功能上冗余, 参与花序轴中生长素向基转运
Zhang et al., 2018
AtABCB21
地上部生长素兼性运输载体, 参与生长素向顶转运, 调控生长素水平
Kamimoto et al., 2012; Jenness et al., 2019
ATM (ABC transporter of mitochondria)
AtABCB23/ATM1
参与铁硫簇输出
Chen et al., 2007
AtABCB24/ATM2
未知
Chen et al., 2007
AtABCB25/ATM3
参与铁硫簇输出, 重金属抗性, 并参与调控钼辅因子合成
Kim et al., 2006; Chen et al., 2007; Bernard et al., 2009; Teschner et al., 2010
TAP (transporter associated with antigen processing)
AtABCB26/TAP1 AtABCB27/TAP2/ALS1
未知 调控铝毒环境中根的发育
Ferro et al., 2010 Larsen et al., 2007; Zhu et al., 2013
半分子转运蛋白AtABCB23 (AtATM1)、AtAB-CB24 (AtATM2)以及AtABCB25 (AtATM3)与酵母线粒体膜蛋白ATM1 (ScATM1)同源.其中, AtATM3以及AtATM1参与线粒体合成时铁硫簇的输出(Chen et al., 2007); 且AtATM3还与拟南芥的重金属抗性有关, 过表达AtATM3可提高拟南芥对镉和铅的抗性(Kim et al., 2006).TAP型转运蛋白成员AtABCB27 (ALS1/TAP2)定位于液泡膜, 在铝毒环境下根系的发 ...
Two homologous ATP-binding cassette transporter proteins, AtMDR1 and AtPGP1, regulate Arabidopsis photomorphogenesis and root development by mediating polar auxin transport 1 2005
... 20世纪20年代提出的Cholodny-Went理论认为, 植物向光性运动的产生是由于单侧光照射下生长素逐渐向背光一侧移动, 进而使生长素在植物茎中分布不均引起.后来该假说也被用于解释植物的向性生长.光敏色素(Phy)和隐花色素(Cry)是植物体内的光受体, 影响植物的向光性甚至光形态建成.许多研究表明, 光敏色素和隐花色素调节下胚轴顶端到根部的生长素向基运输(Jensen et al., 1998; Canamero et al., 2006; Salisbury et al., 2007).ABCB依赖光受体(PhyA、Phot和PhyB)调控光下生长素的极性运输, 进而调节下胚轴生长(Wu et al., 2010; Christie et al., 2011; 刘广超和丁兆军, 2018).Lin和Wang (2005)研究表明, atabcb1和atabcb19突变体对远红光、红光及蓝光较为敏感, 表现出下胚轴生长受抑制的表型, 说明AtABCB1和AtABCB19通过调控生长素的极性运输来影响拟南芥的光形态建成.此外, Wu等(2010)研究发现, 过表达AtABCB19可显著增加下胚轴中生长素的含量, 且红光和蓝光均可减弱其对下胚轴伸长的抑制作用.AtABCB19突变则增强了光敏色素和隐花色素诱导的拟南芥下胚轴弯曲, 说明光敏色素和隐花色素可能通过抑制AtABCB19的表达, 进而抑制下胚轴中生长素的极性运输(Nagashima et al., 2008).研究者认为上述调控模型中首先是红光通过PhyA抑制下胚轴伸长, 而后PhyB作用于AtABCB19影响幼苗中生长素的极性转运(Parks and Spalding, 1999; Wu et al., 2010). ...
Loss of an MDR transporter in compact stalks of maize br2 and sorghum dw3 mutants 4 2003
... 自1992年从拟南芥(Arabidopsis thaliana)中克隆到第1个ABCB亚家族的基因——AtPGP1 (AtABCB1)以来, 植物中的ABCB亚家族研究备受关注(Dudler and Hertig, 1992).近30年的研究发现, ABCB作为转运蛋白与AUX1/LAX (AUXIN1/LIKE AUXIN)和PIN (PIN-FORMED)协同调控植物生长素的极性运输.例如, AtABCB1、AtABCB19、AtABCB4、AtABCB14和AtABCB21 (Sidler et al.,1998; Noh et al., 2001; Geisler et al., 2005; Santelia et al., 2005; Kaneda et al., 2011; Kamimoto et al., 2012)等相继被报道与生长素的运输相关.此外, 也有研究表明, ABCB转运蛋白参与植物对重金属和铝毒的抗性等过程.例如, Kim等(2016)报道了ABCB半分子转运蛋白ATM3 (AtABCB24)可提高拟南芥对镉和铅的抗性.而ALS1/TAP2 (AtABCB27)被报道参与拟南芥的铝转运, 进而提高其铝毒抗性(Larsen et al., 2007).单子叶植物玉米(Zea mays)、高粱(Sorghum bicolor)和水稻(Oryza sativa)中的ABCB蛋白也具有生长素转运功能.玉米ZmABCB1 (Brachytic2, BR2)和高粱SbABCB1 (Dwarf1, DW3)参与生长素的向基运输(Multani et al., 2003); 水稻OsABCB14则参与生长素的向顶运输(Xu et al., 2014).单子叶植物的转录组数据分析显示, ABCB亚家族蛋白可能与植物的非生物胁迫响应密切相关(Shen et al., 2010; Pang et al., 2013; Chai and Subudhi, 2016).可见, ABCB转运蛋白的功能在植物中具有一定的保守性.植物ABCB转运蛋白的数量较多, 然而目前的研究主要集中在双子叶模式植物拟南芥中, 单子叶植物中的研究尚少, 大部分ABCB转运蛋白的功能有待进一步探索.本文主要综述了ABCB亚家族蛋白在双子叶植物拟南芥以及单子叶植物玉米、高粱、小麦(Triticum aestivum)和水稻中的研究进展, 旨在为ABCB基因功能的揭示提供新线索. ...
... Biofunctions of ABCB genes in cereal crops Table 3
物种
基因名称
功能
参考文献
玉米
ZmABCB1
参与地上部生长素运输
Multani et al., 2003
高粱
SbABCB1
参与地上部生长素运输
Multani et al., 2003
小麦
TaMDR1
抗铝毒害
Sasaki et al., 2002
水稻
OsABCB14
参与根中生长素向顶运输, 铁离子平衡
Xu et al., 2014
OsABCB23
铁硫聚簇装配
Zuo et al., 2017
OsABCB27
铝胁迫响应
Huang et al., 2009
3.1 高粱和玉米中ABCB亚家族基因的研究进展 玉米ABCB亚家族蛋白共有31个成员, 包括18个全分子转运蛋白、12个半分子转运蛋白及1个仅编码不完全NBD的成员(ZmABCB19) (Pang et al., 2013).玉米ZmABCB1 (Brachytic2, BR2)和高粱SbABCB1 (Dwarf3, DW3)与拟南芥AtABCB1基因同源, 其突变可导致生长素向基运输减少, 植株矮化, 该表型与拟南芥突变体atabcb1类似, 表明在单子叶植物中ABCB蛋白可能同样发挥调控生长素运输的作用(Multani et al., 2003).此外, 玉米基因组分析显示, 与AtMDR11同源的基因ZmABCB10及与其分布位置重叠的基因ZmABCB16和ZmABCB26在生长旺盛的组织中表达, 说明这些基因也可能与生长素的运输相关(Pang et al., 2013).此外, ZmABCB7和ZmABCB8受干旱胁迫的显著诱导, ZmABCB18则受到盐胁迫的显著抑制(Pang et al., 2013). ...
... Multani et al., 2003
小麦
TaMDR1
抗铝毒害
Sasaki et al., 2002
水稻
OsABCB14
参与根中生长素向顶运输, 铁离子平衡
Xu et al., 2014
OsABCB23
铁硫聚簇装配
Zuo et al., 2017
OsABCB27
铝胁迫响应
Huang et al., 2009
3.1 高粱和玉米中ABCB亚家族基因的研究进展 玉米ABCB亚家族蛋白共有31个成员, 包括18个全分子转运蛋白、12个半分子转运蛋白及1个仅编码不完全NBD的成员(ZmABCB19) (Pang et al., 2013).玉米ZmABCB1 (Brachytic2, BR2)和高粱SbABCB1 (Dwarf3, DW3)与拟南芥AtABCB1基因同源, 其突变可导致生长素向基运输减少, 植株矮化, 该表型与拟南芥突变体atabcb1类似, 表明在单子叶植物中ABCB蛋白可能同样发挥调控生长素运输的作用(Multani et al., 2003).此外, 玉米基因组分析显示, 与AtMDR11同源的基因ZmABCB10及与其分布位置重叠的基因ZmABCB16和ZmABCB26在生长旺盛的组织中表达, 说明这些基因也可能与生长素的运输相关(Pang et al., 2013).此外, ZmABCB7和ZmABCB8受干旱胁迫的显著诱导, ZmABCB18则受到盐胁迫的显著抑制(Pang et al., 2013). ...
... 玉米ABCB亚家族蛋白共有31个成员, 包括18个全分子转运蛋白、12个半分子转运蛋白及1个仅编码不完全NBD的成员(ZmABCB19) (Pang et al., 2013).玉米ZmABCB1 (Brachytic2, BR2)和高粱SbABCB1 (Dwarf3, DW3)与拟南芥AtABCB1基因同源, 其突变可导致生长素向基运输减少, 植株矮化, 该表型与拟南芥突变体atabcb1类似, 表明在单子叶植物中ABCB蛋白可能同样发挥调控生长素运输的作用(Multani et al., 2003).此外, 玉米基因组分析显示, 与AtMDR11同源的基因ZmABCB10及与其分布位置重叠的基因ZmABCB16和ZmABCB26在生长旺盛的组织中表达, 说明这些基因也可能与生长素的运输相关(Pang et al., 2013).此外, ZmABCB7和ZmABCB8受干旱胁迫的显著诱导, ZmABCB18则受到盐胁迫的显著抑制(Pang et al., 2013). ...
Phytochromes and cryptochromes regulate the differential growth of Arabidopsis hypocotyls in both a PGP19-dependent and a PGP19-independent manner 1 2008
... 20世纪20年代提出的Cholodny-Went理论认为, 植物向光性运动的产生是由于单侧光照射下生长素逐渐向背光一侧移动, 进而使生长素在植物茎中分布不均引起.后来该假说也被用于解释植物的向性生长.光敏色素(Phy)和隐花色素(Cry)是植物体内的光受体, 影响植物的向光性甚至光形态建成.许多研究表明, 光敏色素和隐花色素调节下胚轴顶端到根部的生长素向基运输(Jensen et al., 1998; Canamero et al., 2006; Salisbury et al., 2007).ABCB依赖光受体(PhyA、Phot和PhyB)调控光下生长素的极性运输, 进而调节下胚轴生长(Wu et al., 2010; Christie et al., 2011; 刘广超和丁兆军, 2018).Lin和Wang (2005)研究表明, atabcb1和atabcb19突变体对远红光、红光及蓝光较为敏感, 表现出下胚轴生长受抑制的表型, 说明AtABCB1和AtABCB19通过调控生长素的极性运输来影响拟南芥的光形态建成.此外, Wu等(2010)研究发现, 过表达AtABCB19可显著增加下胚轴中生长素的含量, 且红光和蓝光均可减弱其对下胚轴伸长的抑制作用.AtABCB19突变则增强了光敏色素和隐花色素诱导的拟南芥下胚轴弯曲, 说明光敏色素和隐花色素可能通过抑制AtABCB19的表达, 进而抑制下胚轴中生长素的极性运输(Nagashima et al., 2008).研究者认为上述调控模型中首先是红光通过PhyA抑制下胚轴伸长, 而后PhyB作用于AtABCB19影响幼苗中生长素的极性转运(Parks and Spalding, 1999; Wu et al., 2010). ...
Genome-wide expression analysis of rice ABC transporter family across spatio-temporal samples and in response to abiotic stresses 1 2014
... 水稻ABCB基因的研究也一直处于缓慢发展状态.水稻OsABCB14与拟南芥AtABCB19同源性较高, 且OsABCB14在水稻根尖、叶鞘、中柱、叶片、节、节间、根茎结合部和花等器官中均有表达, 其中在维管组织中的表达量最高(Xu et al., 2014).osabcb14突变体生长素的向顶运输速率降低, 且离子含量分析显示其铁离子浓度明显高于野生型, 表明OsABCB14不仅参与生长素的转运, 还与水稻铁离子的平衡密切相关(Xu et al., 2014).水稻全基因组表达分析显示, OsABCB10在花药和花粉中表达水平较高, 暗示其在花药和花粉的发育过程中发挥重要作用(Nguyen et al., 2014).非生物胁迫下的表达分析表明大部分ABCB基因的表达量发生变化.例如, OsABCB11、OsABCB8、OsABCB13、OsABCB23和OsABCB24受干旱胁迫诱导, 而OsABCB6、OsABCB9以及OsABCB8受盐胁迫诱导, 但ABCB亚家族基因在水稻非生物胁迫响应中的作用还有待进一步验证(徐杏等, 2012; Saha et al., 2015).此外, 定位于液泡膜上的TAP型转运蛋白OsABCB27 (OsALS1)参与水稻的铝胁迫响应(Huang et al., 2009), 而定位于线粒体的ATM型转运蛋白OsABCB23 (ATM3)在铁硫聚簇装配中起重要作用, 表明线粒体ATM3转运蛋白对水稻铁离子的稳态至关重要(Zuo et al., 2017). ...
Multidrug resistance-like genes of Arabidopsis required for auxin transport and auxin-mediated development 3 2001
... 自1992年从拟南芥(Arabidopsis thaliana)中克隆到第1个ABCB亚家族的基因——AtPGP1 (AtABCB1)以来, 植物中的ABCB亚家族研究备受关注(Dudler and Hertig, 1992).近30年的研究发现, ABCB作为转运蛋白与AUX1/LAX (AUXIN1/LIKE AUXIN)和PIN (PIN-FORMED)协同调控植物生长素的极性运输.例如, AtABCB1、AtABCB19、AtABCB4、AtABCB14和AtABCB21 (Sidler et al.,1998; Noh et al., 2001; Geisler et al., 2005; Santelia et al., 2005; Kaneda et al., 2011; Kamimoto et al., 2012)等相继被报道与生长素的运输相关.此外, 也有研究表明, ABCB转运蛋白参与植物对重金属和铝毒的抗性等过程.例如, Kim等(2016)报道了ABCB半分子转运蛋白ATM3 (AtABCB24)可提高拟南芥对镉和铅的抗性.而ALS1/TAP2 (AtABCB27)被报道参与拟南芥的铝转运, 进而提高其铝毒抗性(Larsen et al., 2007).单子叶植物玉米(Zea mays)、高粱(Sorghum bicolor)和水稻(Oryza sativa)中的ABCB蛋白也具有生长素转运功能.玉米ZmABCB1 (Brachytic2, BR2)和高粱SbABCB1 (Dwarf1, DW3)参与生长素的向基运输(Multani et al., 2003); 水稻OsABCB14则参与生长素的向顶运输(Xu et al., 2014).单子叶植物的转录组数据分析显示, ABCB亚家族蛋白可能与植物的非生物胁迫响应密切相关(Shen et al., 2010; Pang et al., 2013; Chai and Subudhi, 2016).可见, ABCB转运蛋白的功能在植物中具有一定的保守性.植物ABCB转运蛋白的数量较多, 然而目前的研究主要集中在双子叶模式植物拟南芥中, 单子叶植物中的研究尚少, 大部分ABCB转运蛋白的功能有待进一步探索.本文主要综述了ABCB亚家族蛋白在双子叶植物拟南芥以及单子叶植物玉米、高粱、小麦(Triticum aestivum)和水稻中的研究进展, 旨在为ABCB基因功能的揭示提供新线索. ...
... Biofunctions of ABCB genes in Arabidopsis thaliana (Wang et al., 2017, in Chinese) Table 2
类型
基因名称
功能
参考文献
MDR (multidrug resistance)
AtABCB1/PGP1
参与地上部(花序轴、下胚轴)生长素输出, 促进下胚轴伸长
Sidler et al.,1998; Geisler et al., 2005
AtABCB4/PGP4
在拟南芥根中表现为生长素兼性转运载体, 介导侧根及根毛发育
Santelia et al., 2005; Terasaka et al., 2005; Cho et al., 2007; Lewis et al., 2007; Kubeš et al., 2012
AtABCB6
与AtABCB20在功能上冗余, 参与花序轴中生长素的向基转运
Zhang et al., 2018
AtABCB14/MDR12
参与花序轴中生长素运输, 与花序轴维管发育和气孔开闭有关
Lee et al., 2008; Kaneda et al., 2011
AtABCB15
参与花序轴中生长素运输
Kaneda et al., 2011
AtABCB19/PGP19
参与地上部生长素向基运输以及根的向顶运输, 调控不定根发育及胚后器官分离
Noh et al., 2001; Lewis et al., 2007; Sukumar et al., 2013; Zhao et al., 2013
AtABCB20
与AtABCB6在功能上冗余, 参与花序轴中生长素向基转运
Zhang et al., 2018
AtABCB21
地上部生长素兼性运输载体, 参与生长素向顶转运, 调控生长素水平
Kamimoto et al., 2012; Jenness et al., 2019
ATM (ABC transporter of mitochondria)
AtABCB23/ATM1
参与铁硫簇输出
Chen et al., 2007
AtABCB24/ATM2
未知
Chen et al., 2007
AtABCB25/ATM3
参与铁硫簇输出, 重金属抗性, 并参与调控钼辅因子合成
Kim et al., 2006; Chen et al., 2007; Bernard et al., 2009; Teschner et al., 2010
TAP (transporter associated with antigen processing)
AtABCB26/TAP1 AtABCB27/TAP2/ALS1
未知 调控铝毒环境中根的发育
Ferro et al., 2010 Larsen et al., 2007; Zhu et al., 2013
半分子转运蛋白AtABCB23 (AtATM1)、AtAB-CB24 (AtATM2)以及AtABCB25 (AtATM3)与酵母线粒体膜蛋白ATM1 (ScATM1)同源.其中, AtATM3以及AtATM1参与线粒体合成时铁硫簇的输出(Chen et al., 2007); 且AtATM3还与拟南芥的重金属抗性有关, 过表达AtATM3可提高拟南芥对镉和铅的抗性(Kim et al., 2006).TAP型转运蛋白成员AtABCB27 (ALS1/TAP2)定位于液泡膜, 在铝毒环境下根系的发 ...
An ABC transporter B family protein, ABCB19, is required for cytoplasmic streaming and gravitropism of the inflorescence stems 1 2016
... ABCB亚家族成员在植物生长发育的各个方面发挥不同作用(表2).植物根切除导致吲哚-3-乙酸(IAA)转运快速增加, 并在切除点以上的下胚轴中积累, 进而诱导不定根的形成.Sukumar等(2013)研究发现, 相对于野生型株系atabcb19突变体的不定根数量显著减少, 过表达株系则出现较多的不定根, 且AtABCB19的转录及蛋白表达水平与局部IAA积累的时空变化模式密切相关, 因此认为AtABCB19的局部诱导可促进拟南芥不定根的形成.此外, 有研究表明, AtABCB19还参与调控拟南芥胚后器官的分离(Zhao et al., 2013)及花序轴的细胞质流(Okamoto et al., 2016).AtABCB1在花药发育过程中起主要作用, AtABCB19则起协同作用(Cecchetti et al., 2015).Kaneda等(2011)发现, atabcb14花序轴中维管发育不正常, 说明AtABCB14可能参与拟南芥花序轴的木质化过程.而之前有报道显示, AtABCB14可将苹果酸盐从质外体转运至保卫细胞, 调节渗透压, 从而调控细胞对二氧化碳的响应, 影响气孔开闭(Lee et al., 2008).总之, ABCB蛋白功能复杂多样, 仍有待进一步研究. ...
Inventory and general analysis of the ATP-binding cassette (ABC) gene superfamily in maize (Zea mays L.) 4 2013
... 自1992年从拟南芥(Arabidopsis thaliana)中克隆到第1个ABCB亚家族的基因——AtPGP1 (AtABCB1)以来, 植物中的ABCB亚家族研究备受关注(Dudler and Hertig, 1992).近30年的研究发现, ABCB作为转运蛋白与AUX1/LAX (AUXIN1/LIKE AUXIN)和PIN (PIN-FORMED)协同调控植物生长素的极性运输.例如, AtABCB1、AtABCB19、AtABCB4、AtABCB14和AtABCB21 (Sidler et al.,1998; Noh et al., 2001; Geisler et al., 2005; Santelia et al., 2005; Kaneda et al., 2011; Kamimoto et al., 2012)等相继被报道与生长素的运输相关.此外, 也有研究表明, ABCB转运蛋白参与植物对重金属和铝毒的抗性等过程.例如, Kim等(2016)报道了ABCB半分子转运蛋白ATM3 (AtABCB24)可提高拟南芥对镉和铅的抗性.而ALS1/TAP2 (AtABCB27)被报道参与拟南芥的铝转运, 进而提高其铝毒抗性(Larsen et al., 2007).单子叶植物玉米(Zea mays)、高粱(Sorghum bicolor)和水稻(Oryza sativa)中的ABCB蛋白也具有生长素转运功能.玉米ZmABCB1 (Brachytic2, BR2)和高粱SbABCB1 (Dwarf1, DW3)参与生长素的向基运输(Multani et al., 2003); 水稻OsABCB14则参与生长素的向顶运输(Xu et al., 2014).单子叶植物的转录组数据分析显示, ABCB亚家族蛋白可能与植物的非生物胁迫响应密切相关(Shen et al., 2010; Pang et al., 2013; Chai and Subudhi, 2016).可见, ABCB转运蛋白的功能在植物中具有一定的保守性.植物ABCB转运蛋白的数量较多, 然而目前的研究主要集中在双子叶模式植物拟南芥中, 单子叶植物中的研究尚少, 大部分ABCB转运蛋白的功能有待进一步探索.本文主要综述了ABCB亚家族蛋白在双子叶植物拟南芥以及单子叶植物玉米、高粱、小麦(Triticum aestivum)和水稻中的研究进展, 旨在为ABCB基因功能的揭示提供新线索. ...
... 玉米ABCB亚家族蛋白共有31个成员, 包括18个全分子转运蛋白、12个半分子转运蛋白及1个仅编码不完全NBD的成员(ZmABCB19) (Pang et al., 2013).玉米ZmABCB1 (Brachytic2, BR2)和高粱SbABCB1 (Dwarf3, DW3)与拟南芥AtABCB1基因同源, 其突变可导致生长素向基运输减少, 植株矮化, 该表型与拟南芥突变体atabcb1类似, 表明在单子叶植物中ABCB蛋白可能同样发挥调控生长素运输的作用(Multani et al., 2003).此外, 玉米基因组分析显示, 与AtMDR11同源的基因ZmABCB10及与其分布位置重叠的基因ZmABCB16和ZmABCB26在生长旺盛的组织中表达, 说明这些基因也可能与生长素的运输相关(Pang et al., 2013).此外, ZmABCB7和ZmABCB8受干旱胁迫的显著诱导, ZmABCB18则受到盐胁迫的显著抑制(Pang et al., 2013). ...
... 在生长旺盛的组织中表达, 说明这些基因也可能与生长素的运输相关(Pang et al., 2013).此外, ZmABCB7和ZmABCB8受干旱胁迫的显著诱导, ZmABCB18则受到盐胁迫的显著抑制(Pang et al., 2013). ...
... 则受到盐胁迫的显著抑制(Pang et al., 2013). ...
Sequential and coordinated action of phytochromes A and B during Arabidopsis stem growth revealed by kinetic analysis 1 1999
... 20世纪20年代提出的Cholodny-Went理论认为, 植物向光性运动的产生是由于单侧光照射下生长素逐渐向背光一侧移动, 进而使生长素在植物茎中分布不均引起.后来该假说也被用于解释植物的向性生长.光敏色素(Phy)和隐花色素(Cry)是植物体内的光受体, 影响植物的向光性甚至光形态建成.许多研究表明, 光敏色素和隐花色素调节下胚轴顶端到根部的生长素向基运输(Jensen et al., 1998; Canamero et al., 2006; Salisbury et al., 2007).ABCB依赖光受体(PhyA、Phot和PhyB)调控光下生长素的极性运输, 进而调节下胚轴生长(Wu et al., 2010; Christie et al., 2011; 刘广超和丁兆军, 2018).Lin和Wang (2005)研究表明, atabcb1和atabcb19突变体对远红光、红光及蓝光较为敏感, 表现出下胚轴生长受抑制的表型, 说明AtABCB1和AtABCB19通过调控生长素的极性运输来影响拟南芥的光形态建成.此外, Wu等(2010)研究发现, 过表达AtABCB19可显著增加下胚轴中生长素的含量, 且红光和蓝光均可减弱其对下胚轴伸长的抑制作用.AtABCB19突变则增强了光敏色素和隐花色素诱导的拟南芥下胚轴弯曲, 说明光敏色素和隐花色素可能通过抑制AtABCB19的表达, 进而抑制下胚轴中生长素的极性运输(Nagashima et al., 2008).研究者认为上述调控模型中首先是红光通过PhyA抑制下胚轴伸长, 而后PhyB作用于AtABCB19影响幼苗中生长素的极性转运(Parks and Spalding, 1999; Wu et al., 2010). ...
Molecular phylogenetic study and expression analysis of ATP- binding cassette transporter gene family in Oryza sativa in response to salt stress 1 2015
... 水稻ABCB基因的研究也一直处于缓慢发展状态.水稻OsABCB14与拟南芥AtABCB19同源性较高, 且OsABCB14在水稻根尖、叶鞘、中柱、叶片、节、节间、根茎结合部和花等器官中均有表达, 其中在维管组织中的表达量最高(Xu et al., 2014).osabcb14突变体生长素的向顶运输速率降低, 且离子含量分析显示其铁离子浓度明显高于野生型, 表明OsABCB14不仅参与生长素的转运, 还与水稻铁离子的平衡密切相关(Xu et al., 2014).水稻全基因组表达分析显示, OsABCB10在花药和花粉中表达水平较高, 暗示其在花药和花粉的发育过程中发挥重要作用(Nguyen et al., 2014).非生物胁迫下的表达分析表明大部分ABCB基因的表达量发生变化.例如, OsABCB11、OsABCB8、OsABCB13、OsABCB23和OsABCB24受干旱胁迫诱导, 而OsABCB6、OsABCB9以及OsABCB8受盐胁迫诱导, 但ABCB亚家族基因在水稻非生物胁迫响应中的作用还有待进一步验证(徐杏等, 2012; Saha et al., 2015).此外, 定位于液泡膜上的TAP型转运蛋白OsABCB27 (OsALS1)参与水稻的铝胁迫响应(Huang et al., 2009), 而定位于线粒体的ATM型转运蛋白OsABCB23 (ATM3)在铁硫聚簇装配中起重要作用, 表明线粒体ATM3转运蛋白对水稻铁离子的稳态至关重要(Zuo et al., 2017). ...
Phytochrome coordinates Arabidopsis shoot and root development 1 2007
... 20世纪20年代提出的Cholodny-Went理论认为, 植物向光性运动的产生是由于单侧光照射下生长素逐渐向背光一侧移动, 进而使生长素在植物茎中分布不均引起.后来该假说也被用于解释植物的向性生长.光敏色素(Phy)和隐花色素(Cry)是植物体内的光受体, 影响植物的向光性甚至光形态建成.许多研究表明, 光敏色素和隐花色素调节下胚轴顶端到根部的生长素向基运输(Jensen et al., 1998; Canamero et al., 2006; Salisbury et al., 2007).ABCB依赖光受体(PhyA、Phot和PhyB)调控光下生长素的极性运输, 进而调节下胚轴生长(Wu et al., 2010; Christie et al., 2011; 刘广超和丁兆军, 2018).Lin和Wang (2005)研究表明, atabcb1和atabcb19突变体对远红光、红光及蓝光较为敏感, 表现出下胚轴生长受抑制的表型, 说明AtABCB1和AtABCB19通过调控生长素的极性运输来影响拟南芥的光形态建成.此外, Wu等(2010)研究发现, 过表达AtABCB19可显著增加下胚轴中生长素的含量, 且红光和蓝光均可减弱其对下胚轴伸长的抑制作用.AtABCB19突变则增强了光敏色素和隐花色素诱导的拟南芥下胚轴弯曲, 说明光敏色素和隐花色素可能通过抑制AtABCB19的表达, 进而抑制下胚轴中生长素的极性运输(Nagashima et al., 2008).研究者认为上述调控模型中首先是红光通过PhyA抑制下胚轴伸长, 而后PhyB作用于AtABCB19影响幼苗中生长素的极性转运(Parks and Spalding, 1999; Wu et al., 2010). ...
The Arabidopsis thaliana ABC protein superfamily, a complete inventory 3 2001
... ABC蛋白由于结构中包含有核酸结合域(nucleotide binding domain, NBD), 故又被称作ATP结合盒式蛋白.NBD含有(ABC特征基序, Walker A和B序列, H环和Q环)几个高度保守的基序(Sánchez-Fernández et al., 2001; Higgins and Linton, 2004).除了NBD外, ABC转运蛋白的结构中还包含有跨膜结构域(trans membrane domain, TMD), 每个跨膜结构域均由几个疏水性α-螺旋组成.当ABCB转运蛋白转运底物时, NBD负责结合并水解ATP, 为底物的运输提供动力; TMD则负责底物的识别及参与底物的跨膜转运.根据蛋白的结构, ABC转运蛋白可以分为3类, 即全分子转运蛋白、半分子转运蛋白以及可溶性转运蛋白(Sánchez-Fernández et al., 2001; Verrier et al., 2008).其中, ABCB亚家族蛋白只存在前两种结构(全分子和半分子转运蛋白).全分子转运蛋白包含2个NBD和2个TMD; 半分子转运蛋白则只包括1个NBD和TMD (图1). ...
... Biofunctions of ABCB genes in Arabidopsis thaliana (Wang et al., 2017, in Chinese) Table 2
类型
基因名称
功能
参考文献
MDR (multidrug resistance)
AtABCB1/PGP1
参与地上部(花序轴、下胚轴)生长素输出, 促进下胚轴伸长
Sidler et al.,1998; Geisler et al., 2005
AtABCB4/PGP4
在拟南芥根中表现为生长素兼性转运载体, 介导侧根及根毛发育
Santelia et al., 2005; Terasaka et al., 2005; Cho et al., 2007; Lewis et al., 2007; Kubeš et al., 2012
AtABCB6
与AtABCB20在功能上冗余, 参与花序轴中生长素的向基转运
Zhang et al., 2018
AtABCB14/MDR12
参与花序轴中生长素运输, 与花序轴维管发育和气孔开闭有关
Lee et al., 2008; Kaneda et al., 2011
AtABCB15
参与花序轴中生长素运输
Kaneda et al., 2011
AtABCB19/PGP19
参与地上部生长素向基运输以及根的向顶运输, 调控不定根发育及胚后器官分离
Noh et al., 2001; Lewis et al., 2007; Sukumar et al., 2013; Zhao et al., 2013
AtABCB20
与AtABCB6在功能上冗余, 参与花序轴中生长素向基转运
Zhang et al., 2018
AtABCB21
地上部生长素兼性运输载体, 参与生长素向顶转运, 调控生长素水平
Kamimoto et al., 2012; Jenness et al., 2019
ATM (ABC transporter of mitochondria)
AtABCB23/ATM1
参与铁硫簇输出
Chen et al., 2007
AtABCB24/ATM2
未知
Chen et al., 2007
AtABCB25/ATM3
参与铁硫簇输出, 重金属抗性, 并参与调控钼辅因子合成
Kim et al., 2006; Chen et al., 2007; Bernard et al., 2009; Teschner et al., 2010
TAP (transporter associated with antigen processing)
AtABCB26/TAP1 AtABCB27/TAP2/ALS1
未知 调控铝毒环境中根的发育
Ferro et al., 2010 Larsen et al., 2007; Zhu et al., 2013
半分子转运蛋白AtABCB23 (AtATM1)、AtAB-CB24 (AtATM2)以及AtABCB25 (AtATM3)与酵母线粒体膜蛋白ATM1 (ScATM1)同源.其中, AtATM3以及AtATM1参与线粒体合成时铁硫簇的输出(Chen et al., 2007); 且AtATM3还与拟南芥的重金属抗性有关, 过表达AtATM3可提高拟南芥对镉和铅的抗性(Kim et al., 2006).TAP型转运蛋白成员AtABCB27 (ALS1/TAP2)定位于液泡膜, 在铝毒环境下根系的发 ...
Localized induction of the ATP-binding cassette B19 auxin transporter enhances adventitious root formation in Arabidopsis 2 2013
... ABCB亚家族成员在植物生长发育的各个方面发挥不同作用(表2).植物根切除导致吲哚-3-乙酸(IAA)转运快速增加, 并在切除点以上的下胚轴中积累, 进而诱导不定根的形成.Sukumar等(2013)研究发现, 相对于野生型株系atabcb19突变体的不定根数量显著减少, 过表达株系则出现较多的不定根, 且AtABCB19的转录及蛋白表达水平与局部IAA积累的时空变化模式密切相关, 因此认为AtABCB19的局部诱导可促进拟南芥不定根的形成.此外, 有研究表明, AtABCB19还参与调控拟南芥胚后器官的分离(Zhao et al., 2013)及花序轴的细胞质流(Okamoto et al., 2016).AtABCB1在花药发育过程中起主要作用, AtABCB19则起协同作用(Cecchetti et al., 2015).Kaneda等(2011)发现, atabcb14花序轴中维管发育不正常, 说明AtABCB14可能参与拟南芥花序轴的木质化过程.而之前有报道显示, AtABCB14可将苹果酸盐从质外体转运至保卫细胞, 调节渗透压, 从而调控细胞对二氧化碳的响应, 影响气孔开闭(Lee et al., 2008).总之, ABCB蛋白功能复杂多样, 仍有待进一步研究. ...
... Biofunctions of ABCB genes in Arabidopsis thaliana (Wang et al., 2017, in Chinese) Table 2
类型
基因名称
功能
参考文献
MDR (multidrug resistance)
AtABCB1/PGP1
参与地上部(花序轴、下胚轴)生长素输出, 促进下胚轴伸长
Sidler et al.,1998; Geisler et al., 2005
AtABCB4/PGP4
在拟南芥根中表现为生长素兼性转运载体, 介导侧根及根毛发育
Santelia et al., 2005; Terasaka et al., 2005; Cho et al., 2007; Lewis et al., 2007; Kubeš et al., 2012
AtABCB6
与AtABCB20在功能上冗余, 参与花序轴中生长素的向基转运
Zhang et al., 2018
AtABCB14/MDR12
参与花序轴中生长素运输, 与花序轴维管发育和气孔开闭有关
Lee et al., 2008; Kaneda et al., 2011
AtABCB15
参与花序轴中生长素运输
Kaneda et al., 2011
AtABCB19/PGP19
参与地上部生长素向基运输以及根的向顶运输, 调控不定根发育及胚后器官分离
Noh et al., 2001; Lewis et al., 2007; Sukumar et al., 2013; Zhao et al., 2013
AtABCB20
与AtABCB6在功能上冗余, 参与花序轴中生长素向基转运
Zhang et al., 2018
AtABCB21
地上部生长素兼性运输载体, 参与生长素向顶转运, 调控生长素水平
Kamimoto et al., 2012; Jenness et al., 2019
ATM (ABC transporter of mitochondria)
AtABCB23/ATM1
参与铁硫簇输出
Chen et al., 2007
AtABCB24/ATM2
未知
Chen et al., 2007
AtABCB25/ATM3
参与铁硫簇输出, 重金属抗性, 并参与调控钼辅因子合成
Kim et al., 2006; Chen et al., 2007; Bernard et al., 2009; Teschner et al., 2010
TAP (transporter associated with antigen processing)
AtABCB26/TAP1 AtABCB27/TAP2/ALS1
未知 调控铝毒环境中根的发育
Ferro et al., 2010 Larsen et al., 2007; Zhu et al., 2013
半分子转运蛋白AtABCB23 (AtATM1)、AtAB-CB24 (AtATM2)以及AtABCB25 (AtATM3)与酵母线粒体膜蛋白ATM1 (ScATM1)同源.其中, AtATM3以及AtATM1参与线粒体合成时铁硫簇的输出(Chen et al., 2007); 且AtATM3还与拟南芥的重金属抗性有关, 过表达AtATM3可提高拟南芥对镉和铅的抗性(Kim et al., 2006).TAP型转运蛋白成员AtABCB27 (ALS1/TAP2)定位于液泡膜, 在铝毒环境下根系的发 ...
PGP4, an ATP binding cassette P-glycoprotein, catalyzes auxin transport in Arabidopsis thaliana roots 3 2005
... Lewis等(2007)发现mdr1 (abcb19)根中生长素向顶运输减少80%, 但是向基运输不受影响; mdr4 (abcb4)向基运输减少50%, 而向顶运输不受影响, 说明不同ABCB蛋白在生长素转运中可能起着截然不同的作用.此外, AtPGP4最早被报道参与生长素介导的侧根及根毛发育, 酵母中异源表达AtPGP4可导致其对NPA的敏感性升高; 且atpgp4对生长素及NPA的敏感性下降, 该突变体中游离生长素水平升高而生长素运输水平下降, 表明AtPGP4直接参与调控生长素的转运, 进而影响侧根及根毛的发育(Santelia et al., 2005).同年, Terasaka等(2005)证实, AtPGP4主要参与调控根表皮层细胞生长素的输入过程(Terasaka et al., 2005); 而Cho等(2007)研究发现, AtPGP4在拟南芥根毛细胞以及烟草(Nicotiana taba- cum)细胞中都表现出生长素输出的作用(Cho et al., 2007).Kubeš等(2012)对AtPGP4蛋白进行了研究, 发现其是一种双向生长素转运蛋白, 当细胞中生长素水平较低时促进生长素输入, 较高时则促进生长素输出.AtABCB4的同源蛋白AtABCB21与AtPGP4类似, 也是双向生长素转运蛋白(Kamimoto et al., 2012).最新研究显示, AtABCB21能够调控拟南芥子叶、根的韧皮部以及叶片中的生长素水平, 在拟南芥地上部生长素向顶运输以及植株的向光性运动中发挥作用(Jenness et al., 2019). ...
... 证实, AtPGP4主要参与调控根表皮层细胞生长素的输入过程(Terasaka et al., 2005); 而Cho等(2007)研究发现, AtPGP4在拟南芥根毛细胞以及烟草(Nicotiana taba- cum)细胞中都表现出生长素输出的作用(Cho et al., 2007).Kubeš等(2012)对AtPGP4蛋白进行了研究, 发现其是一种双向生长素转运蛋白, 当细胞中生长素水平较低时促进生长素输入, 较高时则促进生长素输出.AtABCB4的同源蛋白AtABCB21与AtPGP4类似, 也是双向生长素转运蛋白(Kamimoto et al., 2012).最新研究显示, AtABCB21能够调控拟南芥子叶、根的韧皮部以及叶片中的生长素水平, 在拟南芥地上部生长素向顶运输以及植株的向光性运动中发挥作用(Jenness et al., 2019). ...
... Biofunctions of ABCB genes in Arabidopsis thaliana (Wang et al., 2017, in Chinese) Table 2
类型
基因名称
功能
参考文献
MDR (multidrug resistance)
AtABCB1/PGP1
参与地上部(花序轴、下胚轴)生长素输出, 促进下胚轴伸长
Sidler et al.,1998; Geisler et al., 2005
AtABCB4/PGP4
在拟南芥根中表现为生长素兼性转运载体, 介导侧根及根毛发育
Santelia et al., 2005; Terasaka et al., 2005; Cho et al., 2007; Lewis et al., 2007; Kubeš et al., 2012
AtABCB6
与AtABCB20在功能上冗余, 参与花序轴中生长素的向基转运
Zhang et al., 2018
AtABCB14/MDR12
参与花序轴中生长素运输, 与花序轴维管发育和气孔开闭有关
Lee et al., 2008; Kaneda et al., 2011
AtABCB15
参与花序轴中生长素运输
Kaneda et al., 2011
AtABCB19/PGP19
参与地上部生长素向基运输以及根的向顶运输, 调控不定根发育及胚后器官分离
Noh et al., 2001; Lewis et al., 2007; Sukumar et al., 2013; Zhao et al., 2013
AtABCB20
与AtABCB6在功能上冗余, 参与花序轴中生长素向基转运
Zhang et al., 2018
AtABCB21
地上部生长素兼性运输载体, 参与生长素向顶转运, 调控生长素水平
Kamimoto et al., 2012; Jenness et al., 2019
ATM (ABC transporter of mitochondria)
AtABCB23/ATM1
参与铁硫簇输出
Chen et al., 2007
AtABCB24/ATM2
未知
Chen et al., 2007
AtABCB25/ATM3
参与铁硫簇输出, 重金属抗性, 并参与调控钼辅因子合成
Kim et al., 2006; Chen et al., 2007; Bernard et al., 2009; Teschner et al., 2010
TAP (transporter associated with antigen processing)
AtABCB26/TAP1 AtABCB27/TAP2/ALS1
未知 调控铝毒环境中根的发育
Ferro et al., 2010 Larsen et al., 2007; Zhu et al., 2013
半分子转运蛋白AtABCB23 (AtATM1)、AtAB-CB24 (AtATM2)以及AtABCB25 (AtATM3)与酵母线粒体膜蛋白ATM1 (ScATM1)同源.其中, AtATM3以及AtATM1参与线粒体合成时铁硫簇的输出(Chen et al., 2007); 且AtATM3还与拟南芥的重金属抗性有关, 过表达AtATM3可提高拟南芥对镉和铅的抗性(Kim et al., 2006).TAP型转运蛋白成员AtABCB27 (ALS1/TAP2)定位于液泡膜, 在铝毒环境下根系的发 ...
ABC transporter research: going strong 40 years on 2 2015
... ABC (ATP-binding cassette)蛋白超家族是现阶段已知最大的蛋白质家族之一, 包括ABCA、ABCB和ABCC等8个亚家族.早期ABC转运蛋白在动物的耐药性研究中取得较大进展(Theodoulou and Kerr, 2015).因此, 植物中ABC转运蛋白的初步研究也主要集中于外源物质及次生代谢物的转运在解毒和病原体防御中的作用方面(Hwang et al., 2016).随着研究的不断深入, 研究者发现植物ABC转运蛋白在生长素和重金属转运以及植物抗病性等方面同样发挥重要作用(Theodoulou and Kerr, 2015), 其亚家族ABCB转运蛋白在植物生长发育特别是生长素转运中的作用更显著. ...
... ).随着研究的不断深入, 研究者发现植物ABC转运蛋白在生长素和重金属转运以及植物抗病性等方面同样发挥重要作用(Theodoulou and Kerr, 2015), 其亚家族ABCB转运蛋白在植物生长发育特别是生长素转运中的作用更显著. ...
Plant ABC proteins-a unified nomenclature and updated inventory 5 2008
... ABC蛋白由于结构中包含有核酸结合域(nucleotide binding domain, NBD), 故又被称作ATP结合盒式蛋白.NBD含有(ABC特征基序, Walker A和B序列, H环和Q环)几个高度保守的基序(Sánchez-Fernández et al., 2001; Higgins and Linton, 2004).除了NBD外, ABC转运蛋白的结构中还包含有跨膜结构域(trans membrane domain, TMD), 每个跨膜结构域均由几个疏水性α-螺旋组成.当ABCB转运蛋白转运底物时, NBD负责结合并水解ATP, 为底物的运输提供动力; TMD则负责底物的识别及参与底物的跨膜转运.根据蛋白的结构, ABC转运蛋白可以分为3类, 即全分子转运蛋白、半分子转运蛋白以及可溶性转运蛋白(Sánchez-Fernández et al., 2001; Verrier et al., 2008).其中, ABCB亚家族蛋白只存在前两种结构(全分子和半分子转运蛋白).全分子转运蛋白包含2个NBD和2个TMD; 半分子转运蛋白则只包括1个NBD和TMD (图1). ...
... 早期, HUGO命名系统中, ABC转运蛋白被分为ABCA-ABCH八个亚家族(Dean et al., 2001).而拟南芥的ABC转运蛋白, 根据分子大小、方向、结构域组成以及同源性不同分成13个亚家族, 分别为ATH (ABC2 homolog)、ATM (ABC transporter of the mitochondria)、MRP (multidrug resistance associa- ted protein)、PDR (pleiotropic drug resistance)、TAP (transporter associated with antigen processing)、AOH (ABC1 homolog)、NAP (non-intrinsic ABC protein)、PMP (peroxisomal membrane protein)、WBC (white-brown complex homolog)、RLI (RNase L inhibitor)、GCN (general control non-repressible)、MDR (multidrug resistance)和SMC (structural maintenance of chromosomes) (Sánchez-Fernández et al., 2001).随着科学技术的不断进步, 越来越多的蛋白基因被克隆.而不同的命名方式造成了基因名称的混乱, Verrier等(2008)整理并统一了植物中ABC转运蛋白的命名方式.其中, ABCB亚家族分为全分子和半分子转运蛋白两类.全分子转运蛋白仅包括MDR型转运蛋白, 而半分子转运蛋白包括HMT/ATM、TAP和LLP (prokaryotic lipid A-like exporters, putative)型3种(表1). ...
... 拟南芥和水稻ABCB转运蛋白的分类及命名(Verrier et al., 2008; 王晓珠等, 2017) ...
... Classification and nomenclature of ABCB transporters in Arabidopsis thaliana and Oryza sativa (Verrier et al., 2008; Wang et al., 2017, in Chinese) ...
... 拟南芥共有29个ABCB成员, 其中22个药物抗性相关蛋白(multidrug resistance, MDR)为全分子转运蛋白, 其余则为半分子转运蛋白, 即线粒体ABC转运蛋白(ABC transporter of the mitochondria, ATM) 3个、抗原加工相关转运蛋白(transporter associated with antigen processing, TAP) 3个和脂质A输出蛋白(prokaryotic lipid A-like exporters, LLP) 1个(Verrier et al., 2008).目前, 近一半成员得到鉴定, 部分成员的功能得到了初步研究. 但由于ABCB转运蛋白功能复杂, 故仍需进一步探索.现有的研究表明, ABCB转运蛋白在拟南芥生长素的极性运输、向光性、向地性、器官发育和重金属转运等生理过程中均发挥作用. ...
Arabidopsis TWISTED DWARF1 functionally interacts with auxin exporter ABCB1 on the root plasma membrane 2 2013
... 免疫亲和素可与ABCB蛋白互作, 间接影响生长素的运输(Geisler et al., 2004, 2016; Granzin et al., 2006).而分子伴侣TWD1最初是在酵母双杂交中被发现的与ABCB1互作的蛋白(Geisler et al., 2003).后来, 通过免疫共沉淀(Co-immunoprecipitation, Co- IP)以及生物发光能量共振转移(BRET)技术进一步证明TWD1和ABCB1在酵母及植物表达系统中存在互作(Wang et al., 2013).TWD1作为分子伴侣可通过促进AtABCB1重折叠改变其构型, 增强其ATP及底物结合能力(Geisler et al., 2004).此外, twd1突变体中被错误定位到内质网膜上的AtABCB1、AtABCB4和AtABCB19最终会被降解, 表明TWD1主要在ABCB蛋白从内质网膜到质膜的转运过程中发挥分子伴侣作用(Wu et al., 2010; Wang et al., 2013) (图2).近年, 有研究表明NPA和BUM等生长素非竞争性抑制剂可通过结合TWD1, 导致其与ABCB的结合受抑制, 从而使生长素运输减少(Kim et al., 2010; Zhu et al., 2016).Zhu等(2016)研究发现, 肌动蛋白ACTIN7与TWD1之间存在物理互作, 二者可能在不同层次控制蛋白的转运(Zhu et al., 2016). ...
... ; Wang et al., 2013) (图2).近年, 有研究表明NPA和BUM等生长素非竞争性抑制剂可通过结合TWD1, 导致其与ABCB的结合受抑制, 从而使生长素运输减少(Kim et al., 2010; Zhu et al., 2016).Zhu等(2016)研究发现, 肌动蛋白ACTIN7与TWD1之间存在物理互作, 二者可能在不同层次控制蛋白的转运(Zhu et al., 2016). ...
Analysis of subcellular localization of auxin carriers PIN, AUX/LAX and PGP in Sorghum bicolor 1 2011
... 高粱ABC蛋白有24个ABCB亚家族成员, 亚细胞定位预测分析显示, 除了SbPGP5、SbPGP11和SbPGP13外的ABCB蛋白可能定位于细胞质膜上(Wang et al., 2011).此外, 有研究表明, IAA处理可上调SbPGP1、SbPGP13、SbPGP15、SbPGP18和SbPGP23在根中表达, 下调SbPGP5、SbPGP11、SbPGP12、SbPGP17、SbPGP19和SbPGP24表达; 而非生物胁迫条件下, 多个高粱ABCB基因表达出现了显著变化, 说明ABCB家族可能在生长素响应与非生物胁迫之间的信号交互中发挥作用(Shen et al., 2010). ...
A role for ABCB19-mediated polar auxin transport in seedling photomorphogenesis mediated by cryptochrome 1 and phytochrome B 4 2010
... 20世纪20年代提出的Cholodny-Went理论认为, 植物向光性运动的产生是由于单侧光照射下生长素逐渐向背光一侧移动, 进而使生长素在植物茎中分布不均引起.后来该假说也被用于解释植物的向性生长.光敏色素(Phy)和隐花色素(Cry)是植物体内的光受体, 影响植物的向光性甚至光形态建成.许多研究表明, 光敏色素和隐花色素调节下胚轴顶端到根部的生长素向基运输(Jensen et al., 1998; Canamero et al., 2006; Salisbury et al., 2007).ABCB依赖光受体(PhyA、Phot和PhyB)调控光下生长素的极性运输, 进而调节下胚轴生长(Wu et al., 2010; Christie et al., 2011; 刘广超和丁兆军, 2018).Lin和Wang (2005)研究表明, atabcb1和atabcb19突变体对远红光、红光及蓝光较为敏感, 表现出下胚轴生长受抑制的表型, 说明AtABCB1和AtABCB19通过调控生长素的极性运输来影响拟南芥的光形态建成.此外, Wu等(2010)研究发现, 过表达AtABCB19可显著增加下胚轴中生长素的含量, 且红光和蓝光均可减弱其对下胚轴伸长的抑制作用.AtABCB19突变则增强了光敏色素和隐花色素诱导的拟南芥下胚轴弯曲, 说明光敏色素和隐花色素可能通过抑制AtABCB19的表达, 进而抑制下胚轴中生长素的极性运输(Nagashima et al., 2008).研究者认为上述调控模型中首先是红光通过PhyA抑制下胚轴伸长, 而后PhyB作用于AtABCB19影响幼苗中生长素的极性转运(Parks and Spalding, 1999; Wu et al., 2010). ...
... 突变体对远红光、红光及蓝光较为敏感, 表现出下胚轴生长受抑制的表型, 说明AtABCB1和AtABCB19通过调控生长素的极性运输来影响拟南芥的光形态建成.此外, Wu等(2010)研究发现, 过表达AtABCB19可显著增加下胚轴中生长素的含量, 且红光和蓝光均可减弱其对下胚轴伸长的抑制作用.AtABCB19突变则增强了光敏色素和隐花色素诱导的拟南芥下胚轴弯曲, 说明光敏色素和隐花色素可能通过抑制AtABCB19的表达, 进而抑制下胚轴中生长素的极性运输(Nagashima et al., 2008).研究者认为上述调控模型中首先是红光通过PhyA抑制下胚轴伸长, 而后PhyB作用于AtABCB19影响幼苗中生长素的极性转运(Parks and Spalding, 1999; Wu et al., 2010). ...
... ; Wu et al., 2010). ...
... 免疫亲和素可与ABCB蛋白互作, 间接影响生长素的运输(Geisler et al., 2004, 2016; Granzin et al., 2006).而分子伴侣TWD1最初是在酵母双杂交中被发现的与ABCB1互作的蛋白(Geisler et al., 2003).后来, 通过免疫共沉淀(Co-immunoprecipitation, Co- IP)以及生物发光能量共振转移(BRET)技术进一步证明TWD1和ABCB1在酵母及植物表达系统中存在互作(Wang et al., 2013).TWD1作为分子伴侣可通过促进AtABCB1重折叠改变其构型, 增强其ATP及底物结合能力(Geisler et al., 2004).此外, twd1突变体中被错误定位到内质网膜上的AtABCB1、AtABCB4和AtABCB19最终会被降解, 表明TWD1主要在ABCB蛋白从内质网膜到质膜的转运过程中发挥分子伴侣作用(Wu et al., 2010; Wang et al., 2013) (图2).近年, 有研究表明NPA和BUM等生长素非竞争性抑制剂可通过结合TWD1, 导致其与ABCB的结合受抑制, 从而使生长素运输减少(Kim et al., 2010; Zhu et al., 2016).Zhu等(2016)研究发现, 肌动蛋白ACTIN7与TWD1之间存在物理互作, 二者可能在不同层次控制蛋白的转运(Zhu et al., 2016). ...
OsABCB14 functions in auxin transport and iron homeostasis in rice ( Oryza sativa L.) 4 2014
... 自1992年从拟南芥(Arabidopsis thaliana)中克隆到第1个ABCB亚家族的基因——AtPGP1 (AtABCB1)以来, 植物中的ABCB亚家族研究备受关注(Dudler and Hertig, 1992).近30年的研究发现, ABCB作为转运蛋白与AUX1/LAX (AUXIN1/LIKE AUXIN)和PIN (PIN-FORMED)协同调控植物生长素的极性运输.例如, AtABCB1、AtABCB19、AtABCB4、AtABCB14和AtABCB21 (Sidler et al.,1998; Noh et al., 2001; Geisler et al., 2005; Santelia et al., 2005; Kaneda et al., 2011; Kamimoto et al., 2012)等相继被报道与生长素的运输相关.此外, 也有研究表明, ABCB转运蛋白参与植物对重金属和铝毒的抗性等过程.例如, Kim等(2016)报道了ABCB半分子转运蛋白ATM3 (AtABCB24)可提高拟南芥对镉和铅的抗性.而ALS1/TAP2 (AtABCB27)被报道参与拟南芥的铝转运, 进而提高其铝毒抗性(Larsen et al., 2007).单子叶植物玉米(Zea mays)、高粱(Sorghum bicolor)和水稻(Oryza sativa)中的ABCB蛋白也具有生长素转运功能.玉米ZmABCB1 (Brachytic2, BR2)和高粱SbABCB1 (Dwarf1, DW3)参与生长素的向基运输(Multani et al., 2003); 水稻OsABCB14则参与生长素的向顶运输(Xu et al., 2014).单子叶植物的转录组数据分析显示, ABCB亚家族蛋白可能与植物的非生物胁迫响应密切相关(Shen et al., 2010; Pang et al., 2013; Chai and Subudhi, 2016).可见, ABCB转运蛋白的功能在植物中具有一定的保守性.植物ABCB转运蛋白的数量较多, 然而目前的研究主要集中在双子叶模式植物拟南芥中, 单子叶植物中的研究尚少, 大部分ABCB转运蛋白的功能有待进一步探索.本文主要综述了ABCB亚家族蛋白在双子叶植物拟南芥以及单子叶植物玉米、高粱、小麦(Triticum aestivum)和水稻中的研究进展, 旨在为ABCB基因功能的揭示提供新线索. ...
... Biofunctions of ABCB genes in cereal crops Table 3
物种
基因名称
功能
参考文献
玉米
ZmABCB1
参与地上部生长素运输
Multani et al., 2003
高粱
SbABCB1
参与地上部生长素运输
Multani et al., 2003
小麦
TaMDR1
抗铝毒害
Sasaki et al., 2002
水稻
OsABCB14
参与根中生长素向顶运输, 铁离子平衡
Xu et al., 2014
OsABCB23
铁硫聚簇装配
Zuo et al., 2017
OsABCB27
铝胁迫响应
Huang et al., 2009
3.1 高粱和玉米中ABCB亚家族基因的研究进展 玉米ABCB亚家族蛋白共有31个成员, 包括18个全分子转运蛋白、12个半分子转运蛋白及1个仅编码不完全NBD的成员(ZmABCB19) (Pang et al., 2013).玉米ZmABCB1 (Brachytic2, BR2)和高粱SbABCB1 (Dwarf3, DW3)与拟南芥AtABCB1基因同源, 其突变可导致生长素向基运输减少, 植株矮化, 该表型与拟南芥突变体atabcb1类似, 表明在单子叶植物中ABCB蛋白可能同样发挥调控生长素运输的作用(Multani et al., 2003).此外, 玉米基因组分析显示, 与AtMDR11同源的基因ZmABCB10及与其分布位置重叠的基因ZmABCB16和ZmABCB26在生长旺盛的组织中表达, 说明这些基因也可能与生长素的运输相关(Pang et al., 2013).此外, ZmABCB7和ZmABCB8受干旱胁迫的显著诱导, ZmABCB18则受到盐胁迫的显著抑制(Pang et al., 2013). ...
... 水稻ABCB基因的研究也一直处于缓慢发展状态.水稻OsABCB14与拟南芥AtABCB19同源性较高, 且OsABCB14在水稻根尖、叶鞘、中柱、叶片、节、节间、根茎结合部和花等器官中均有表达, 其中在维管组织中的表达量最高(Xu et al., 2014).osabcb14突变体生长素的向顶运输速率降低, 且离子含量分析显示其铁离子浓度明显高于野生型, 表明OsABCB14不仅参与生长素的转运, 还与水稻铁离子的平衡密切相关(Xu et al., 2014).水稻全基因组表达分析显示, OsABCB10在花药和花粉中表达水平较高, 暗示其在花药和花粉的发育过程中发挥重要作用(Nguyen et al., 2014).非生物胁迫下的表达分析表明大部分ABCB基因的表达量发生变化.例如, OsABCB11、OsABCB8、OsABCB13、OsABCB23和OsABCB24受干旱胁迫诱导, 而OsABCB6、OsABCB9以及OsABCB8受盐胁迫诱导, 但ABCB亚家族基因在水稻非生物胁迫响应中的作用还有待进一步验证(徐杏等, 2012; Saha et al., 2015).此外, 定位于液泡膜上的TAP型转运蛋白OsABCB27 (OsALS1)参与水稻的铝胁迫响应(Huang et al., 2009), 而定位于线粒体的ATM型转运蛋白OsABCB23 (ATM3)在铁硫聚簇装配中起重要作用, 表明线粒体ATM3转运蛋白对水稻铁离子的稳态至关重要(Zuo et al., 2017). ...
... 突变体生长素的向顶运输速率降低, 且离子含量分析显示其铁离子浓度明显高于野生型, 表明OsABCB14不仅参与生长素的转运, 还与水稻铁离子的平衡密切相关(Xu et al., 2014).水稻全基因组表达分析显示, OsABCB10在花药和花粉中表达水平较高, 暗示其在花药和花粉的发育过程中发挥重要作用(Nguyen et al., 2014).非生物胁迫下的表达分析表明大部分ABCB基因的表达量发生变化.例如, OsABCB11、OsABCB8、OsABCB13、OsABCB23和OsABCB24受干旱胁迫诱导, 而OsABCB6、OsABCB9以及OsABCB8受盐胁迫诱导, 但ABCB亚家族基因在水稻非生物胁迫响应中的作用还有待进一步验证(徐杏等, 2012; Saha et al., 2015).此外, 定位于液泡膜上的TAP型转运蛋白OsABCB27 (OsALS1)参与水稻的铝胁迫响应(Huang et al., 2009), 而定位于线粒体的ATM型转运蛋白OsABCB23 (ATM3)在铁硫聚簇装配中起重要作用, 表明线粒体ATM3转运蛋白对水稻铁离子的稳态至关重要(Zuo et al., 2017). ...
A transportome-scale amiRNA-based screen identifies redundant roles of Arabidopsis ABCB6 and ABCB20 in auxin transport 4 2018
The ATP-binding cassette transporter ABCB19 regulates postembryonic organ separation in Arabidopsis 2 2013
... ABCB亚家族成员在植物生长发育的各个方面发挥不同作用(表2).植物根切除导致吲哚-3-乙酸(IAA)转运快速增加, 并在切除点以上的下胚轴中积累, 进而诱导不定根的形成.Sukumar等(2013)研究发现, 相对于野生型株系atabcb19突变体的不定根数量显著减少, 过表达株系则出现较多的不定根, 且AtABCB19的转录及蛋白表达水平与局部IAA积累的时空变化模式密切相关, 因此认为AtABCB19的局部诱导可促进拟南芥不定根的形成.此外, 有研究表明, AtABCB19还参与调控拟南芥胚后器官的分离(Zhao et al., 2013)及花序轴的细胞质流(Okamoto et al., 2016).AtABCB1在花药发育过程中起主要作用, AtABCB19则起协同作用(Cecchetti et al., 2015).Kaneda等(2011)发现, atabcb14花序轴中维管发育不正常, 说明AtABCB14可能参与拟南芥花序轴的木质化过程.而之前有报道显示, AtABCB14可将苹果酸盐从质外体转运至保卫细胞, 调节渗透压, 从而调控细胞对二氧化碳的响应, 影响气孔开闭(Lee et al., 2008).总之, ABCB蛋白功能复杂多样, 仍有待进一步研究. ...
... Biofunctions of ABCB genes in Arabidopsis thaliana (Wang et al., 2017, in Chinese) Table 2
类型
基因名称
功能
参考文献
MDR (multidrug resistance)
AtABCB1/PGP1
参与地上部(花序轴、下胚轴)生长素输出, 促进下胚轴伸长
Sidler et al.,1998; Geisler et al., 2005
AtABCB4/PGP4
在拟南芥根中表现为生长素兼性转运载体, 介导侧根及根毛发育
Santelia et al., 2005; Terasaka et al., 2005; Cho et al., 2007; Lewis et al., 2007; Kubeš et al., 2012
AtABCB6
与AtABCB20在功能上冗余, 参与花序轴中生长素的向基转运
Zhang et al., 2018
AtABCB14/MDR12
参与花序轴中生长素运输, 与花序轴维管发育和气孔开闭有关
Lee et al., 2008; Kaneda et al., 2011
AtABCB15
参与花序轴中生长素运输
Kaneda et al., 2011
AtABCB19/PGP19
参与地上部生长素向基运输以及根的向顶运输, 调控不定根发育及胚后器官分离
Noh et al., 2001; Lewis et al., 2007; Sukumar et al., 2013; Zhao et al., 2013
AtABCB20
与AtABCB6在功能上冗余, 参与花序轴中生长素向基转运
Zhang et al., 2018
AtABCB21
地上部生长素兼性运输载体, 参与生长素向顶转运, 调控生长素水平
Kamimoto et al., 2012; Jenness et al., 2019
ATM (ABC transporter of mitochondria)
AtABCB23/ATM1
参与铁硫簇输出
Chen et al., 2007
AtABCB24/ATM2
未知
Chen et al., 2007
AtABCB25/ATM3
参与铁硫簇输出, 重金属抗性, 并参与调控钼辅因子合成
Kim et al., 2006; Chen et al., 2007; Bernard et al., 2009; Teschner et al., 2010
TAP (transporter associated with antigen processing)
AtABCB26/TAP1 AtABCB27/TAP2/ALS1
未知 调控铝毒环境中根的发育
Ferro et al., 2010 Larsen et al., 2007; Zhu et al., 2013
半分子转运蛋白AtABCB23 (AtATM1)、AtAB-CB24 (AtATM2)以及AtABCB25 (AtATM3)与酵母线粒体膜蛋白ATM1 (ScATM1)同源.其中, AtATM3以及AtATM1参与线粒体合成时铁硫簇的输出(Chen et al., 2007); 且AtATM3还与拟南芥的重金属抗性有关, 过表达AtATM3可提高拟南芥对镉和铅的抗性(Kim et al., 2006).TAP型转运蛋白成员AtABCB27 (ALS1/TAP2)定位于液泡膜, 在铝毒环境下根系的发 ...
TWISTED DWARF1 mediates the action of auxin transport inhibitors on actin cytoskeleton dynamics 3 2016
... 免疫亲和素可与ABCB蛋白互作, 间接影响生长素的运输(Geisler et al., 2004, 2016; Granzin et al., 2006).而分子伴侣TWD1最初是在酵母双杂交中被发现的与ABCB1互作的蛋白(Geisler et al., 2003).后来, 通过免疫共沉淀(Co-immunoprecipitation, Co- IP)以及生物发光能量共振转移(BRET)技术进一步证明TWD1和ABCB1在酵母及植物表达系统中存在互作(Wang et al., 2013).TWD1作为分子伴侣可通过促进AtABCB1重折叠改变其构型, 增强其ATP及底物结合能力(Geisler et al., 2004).此外, twd1突变体中被错误定位到内质网膜上的AtABCB1、AtABCB4和AtABCB19最终会被降解, 表明TWD1主要在ABCB蛋白从内质网膜到质膜的转运过程中发挥分子伴侣作用(Wu et al., 2010; Wang et al., 2013) (图2).近年, 有研究表明NPA和BUM等生长素非竞争性抑制剂可通过结合TWD1, 导致其与ABCB的结合受抑制, 从而使生长素运输减少(Kim et al., 2010; Zhu et al., 2016).Zhu等(2016)研究发现, 肌动蛋白ACTIN7与TWD1之间存在物理互作, 二者可能在不同层次控制蛋白的转运(Zhu et al., 2016). ...
... ).Zhu等(2016)研究发现, 肌动蛋白ACTIN7与TWD1之间存在物理互作, 二者可能在不同层次控制蛋白的转运(Zhu et al., 2016). ...
... 研究发现, 肌动蛋白ACTIN7与TWD1之间存在物理互作, 二者可能在不同层次控制蛋白的转运(Zhu et al., 2016). ...
Coordination between apoplastic and symplastic detoxification confers plant aluminum resistance 2 2013
... Biofunctions of ABCB genes in Arabidopsis thaliana (Wang et al., 2017, in Chinese) Table 2
类型
基因名称
功能
参考文献
MDR (multidrug resistance)
AtABCB1/PGP1
参与地上部(花序轴、下胚轴)生长素输出, 促进下胚轴伸长
Sidler et al.,1998; Geisler et al., 2005
AtABCB4/PGP4
在拟南芥根中表现为生长素兼性转运载体, 介导侧根及根毛发育
Santelia et al., 2005; Terasaka et al., 2005; Cho et al., 2007; Lewis et al., 2007; Kubeš et al., 2012
AtABCB6
与AtABCB20在功能上冗余, 参与花序轴中生长素的向基转运
Zhang et al., 2018
AtABCB14/MDR12
参与花序轴中生长素运输, 与花序轴维管发育和气孔开闭有关
Lee et al., 2008; Kaneda et al., 2011
AtABCB15
参与花序轴中生长素运输
Kaneda et al., 2011
AtABCB19/PGP19
参与地上部生长素向基运输以及根的向顶运输, 调控不定根发育及胚后器官分离
Noh et al., 2001; Lewis et al., 2007; Sukumar et al., 2013; Zhao et al., 2013
AtABCB20
与AtABCB6在功能上冗余, 参与花序轴中生长素向基转运
Zhang et al., 2018
AtABCB21
地上部生长素兼性运输载体, 参与生长素向顶转运, 调控生长素水平
Kamimoto et al., 2012; Jenness et al., 2019
ATM (ABC transporter of mitochondria)
AtABCB23/ATM1
参与铁硫簇输出
Chen et al., 2007
AtABCB24/ATM2
未知
Chen et al., 2007
AtABCB25/ATM3
参与铁硫簇输出, 重金属抗性, 并参与调控钼辅因子合成
Kim et al., 2006; Chen et al., 2007; Bernard et al., 2009; Teschner et al., 2010
TAP (transporter associated with antigen processing)
AtABCB26/TAP1 AtABCB27/TAP2/ALS1
未知 调控铝毒环境中根的发育
Ferro et al., 2010 Larsen et al., 2007; Zhu et al., 2013
半分子转运蛋白AtABCB23 (AtATM1)、AtAB-CB24 (AtATM2)以及AtABCB25 (AtATM3)与酵母线粒体膜蛋白ATM1 (ScATM1)同源.其中, AtATM3以及AtATM1参与线粒体合成时铁硫簇的输出(Chen et al., 2007); 且AtATM3还与拟南芥的重金属抗性有关, 过表达AtATM3可提高拟南芥对镉和铅的抗性(Kim et al., 2006).TAP型转运蛋白成员AtABCB27 (ALS1/TAP2)定位于液泡膜, 在铝毒环境下根系的发 ...
... 育方面发挥重要作用(Larsen et al., 2007; Zhu et al., 2013); AtABCB26 (TAP1)则定位于叶绿体膜, 具体功能未知(Ferro et al., 2010). ...
Mitochondrial ABC transporter ATM3 is essential for cytosolic iron-sulfur cluster assembly 2 2017
... Biofunctions of ABCB genes in cereal crops Table 3
物种
基因名称
功能
参考文献
玉米
ZmABCB1
参与地上部生长素运输
Multani et al., 2003
高粱
SbABCB1
参与地上部生长素运输
Multani et al., 2003
小麦
TaMDR1
抗铝毒害
Sasaki et al., 2002
水稻
OsABCB14
参与根中生长素向顶运输, 铁离子平衡
Xu et al., 2014
OsABCB23
铁硫聚簇装配
Zuo et al., 2017
OsABCB27
铝胁迫响应
Huang et al., 2009
3.1 高粱和玉米中ABCB亚家族基因的研究进展 玉米ABCB亚家族蛋白共有31个成员, 包括18个全分子转运蛋白、12个半分子转运蛋白及1个仅编码不完全NBD的成员(ZmABCB19) (Pang et al., 2013).玉米ZmABCB1 (Brachytic2, BR2)和高粱SbABCB1 (Dwarf3, DW3)与拟南芥AtABCB1基因同源, 其突变可导致生长素向基运输减少, 植株矮化, 该表型与拟南芥突变体atabcb1类似, 表明在单子叶植物中ABCB蛋白可能同样发挥调控生长素运输的作用(Multani et al., 2003).此外, 玉米基因组分析显示, 与AtMDR11同源的基因ZmABCB10及与其分布位置重叠的基因ZmABCB16和ZmABCB26在生长旺盛的组织中表达, 说明这些基因也可能与生长素的运输相关(Pang et al., 2013).此外, ZmABCB7和ZmABCB8受干旱胁迫的显著诱导, ZmABCB18则受到盐胁迫的显著抑制(Pang et al., 2013). ...
... 水稻ABCB基因的研究也一直处于缓慢发展状态.水稻OsABCB14与拟南芥AtABCB19同源性较高, 且OsABCB14在水稻根尖、叶鞘、中柱、叶片、节、节间、根茎结合部和花等器官中均有表达, 其中在维管组织中的表达量最高(Xu et al., 2014).osabcb14突变体生长素的向顶运输速率降低, 且离子含量分析显示其铁离子浓度明显高于野生型, 表明OsABCB14不仅参与生长素的转运, 还与水稻铁离子的平衡密切相关(Xu et al., 2014).水稻全基因组表达分析显示, OsABCB10在花药和花粉中表达水平较高, 暗示其在花药和花粉的发育过程中发挥重要作用(Nguyen et al., 2014).非生物胁迫下的表达分析表明大部分ABCB基因的表达量发生变化.例如, OsABCB11、OsABCB8、OsABCB13、OsABCB23和OsABCB24受干旱胁迫诱导, 而OsABCB6、OsABCB9以及OsABCB8受盐胁迫诱导, 但ABCB亚家族基因在水稻非生物胁迫响应中的作用还有待进一步验证(徐杏等, 2012; Saha et al., 2015).此外, 定位于液泡膜上的TAP型转运蛋白OsABCB27 (OsALS1)参与水稻的铝胁迫响应(Huang et al., 2009), 而定位于线粒体的ATM型转运蛋白OsABCB23 (ATM3)在铁硫聚簇装配中起重要作用, 表明线粒体ATM3转运蛋白对水稻铁离子的稳态至关重要(Zuo et al., 2017). ...
 ,1,2,*
,1,2,*
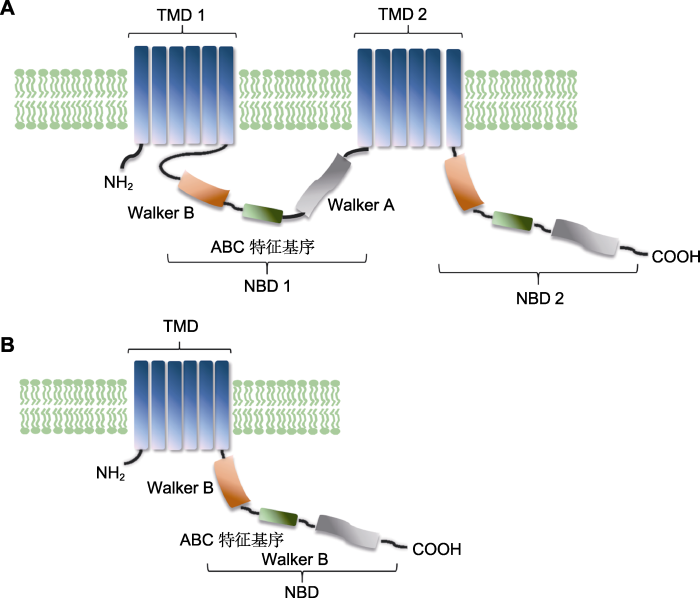 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT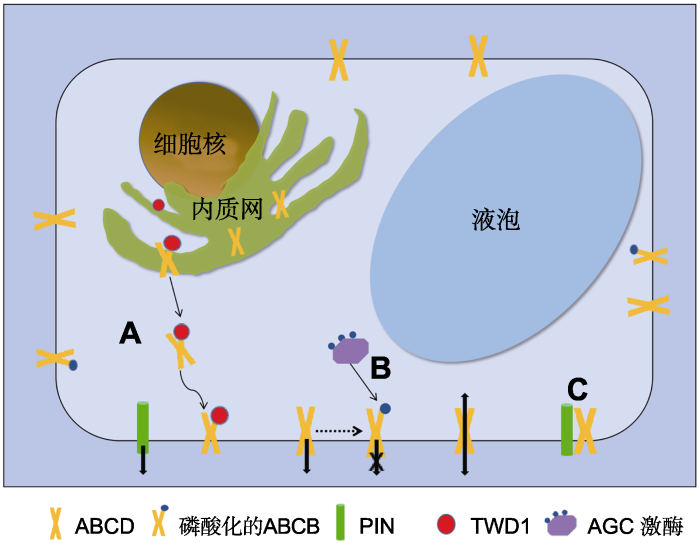 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}