Research Advances on Lectin Receptor-like Kinases in Plants
Menglong Wang1, Xiaoqun Peng1, Zhufeng Chen2, Xiaoyan Tang,1,*1Guangdong Provincial Key Laboratory of Biotechnology for Plant Development, School of Life Sciences, South China Normal University, Guangzhou 510631, China 2Shenzhen Institute of Molecular Crop Design, Shenzhen 518107, China
Abstract Plant growth and development are affected by various environmental factors. In response to various environmental changes, plants have evolved a series of signal recognition and transduction proteins, such as the plasma membrane-localized receptor-like kinases (RLKs), to cope with the environmental conditions. The lectin receptor-like kinases (LecRLKs) are a subfamily of RLKs that contain three structural domains: the extracellular lectin domain, transmembrane domain, and the intracellular kinase domain. Based on the structural difference of the extracellular lectin domain, LecRLKs are classified into three subclasses: L-, G-, and C-type. Recent studies have shown that LecRLKs play a vital role in plant development and biotic/abiotic stress responses. In this review, we discribe the research history, structural features and classification, and biological functions of LecRLKs, and emphasize on the functions of LecRLKs in plants in response to biotic/abiotic stresses and in regulating development. This review provides a view for future functional study on LecRLKs and crop improvement by elaborating different types and functions of LecRLKs. Keywords:receptor-like kinase;lectin receptor-like kinase;biotic/abiotic stress;plant growth and development
PDF (948KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 王梦龙, 彭小群, 陈竹锋, 唐晓艳. 植物凝集素类受体蛋白激酶研究进展. 植物学报, 2020, 55(1): 96-105 doi:10.11983/CBB19130 Wang Menglong, Peng Xiaoqun, Chen Zhufeng, Tang Xiaoyan. Research Advances on Lectin Receptor-like Kinases in Plants. Chinese Bulletin of Botany, 2020, 55(1): 96-105 doi:10.11983/CBB19130
受体蛋白激酶(receptor protein kinases, RPKs)是一类能够与配基结合并具有酶活性的受体(Hunter, 1991)。RPKs蛋白在动物中研究较早, 主要分为酪氨酸激酶(receptor tyrosine kinases, RTKs)和丝/苏氨酸激酶(serine/threonine kinase receptors, STRKs) 2种。与动物不同, 植物中受体蛋白激酶主要属于丝/苏氨酸激酶。目前, 已在植物中陆续鉴定到多个与动物受体蛋白激酶结构相似的蛋白, 这类蛋白绝大部分功能未知, 被称为类受体蛋白激酶(receptor-like kinases, RLKs) (Walker, 1994; Shiu and Bleecker, 2001)。RLKs是一种普遍存在于植物体内的蛋白, 能够感知和传递各种信号, 在拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)中分别有超过600和1 100个该类蛋白(Shiu and Bleecker, 2001)。大部分RLKs属于细胞质膜蛋白, 主要功能是通过它们的细胞外配体识别结构域和细胞内激酶结构域传递各种信号, 最终产生一系列生物效应(Vaid et al., 2013)。
BecraftPW, StinardPS, McCartyDR (1996). CRINKLY4: a TNFR-like receptor kinase involved in maize epidermal differentiation Science 273, 1406-1409. [本文引用: 1]
BellandeK, BonoJJ, SavelliB, JametE, CanutH (2017). Plant lectins and lectin receptor-like kinases: how do they sense the outside? Int J Mol Sci 18, 1164. [本文引用: 2]
BonaventureG (2011). The Nicotiana attenuata lectin receptor kinase 1 is involved in the perception of insect feeding Plant Signal Behav 6, 2060-2063. [本文引用: 3]
ChoiJ, TanakaK, CaoYR, QiY, QiuJ, LiangY, LeeSY, StaceyG (2014). Identi?cation of a plant receptor for extracellular ATP Science 343, 290-294. [本文引用: 1]
DengKQ, WangQM, ZengJX, GuoXH, ZhaoXY, TangDY, LiuXM (2009). A lectin receptor kinase positively regulates ABA response during seed germination and is involved in salt and osmotic stress response J Plant Biol 52, 493-500. [本文引用: 1]
EdelmanGM, WangJL (1978). Binding and functional properties of concanavalin A and its derivatives. III. Interactions with indoleacetic acid and other hydrophobic ligands J Biol Chem 253, 3016-3022. [本文引用: 1]
EpsteinJ, EichbaumQ, SheriffS, EzekowitzRAB (1996). The collectins in innate immunity Curr Opin Immunol 8, 29-35. [本文引用: 2]
Gouhier-DarimontC, SchmiesingA, BonnetC, LassueurS, ReymondP (2013). Signaling of Arabidopsis thaliana response to Pieris brassicae eggs shares similarities with PAMP-triggered immunity J Exp Bot 64, 665-674. [本文引用: 2]
HawgoodS, AkiyamaJ, BrownC, AllenL, LiG, PoulainFR (2001). GM-CSF mediates alveolar macrophage proliferation and type II cell hypertrophy in SP-D gene-targeted mice Am J Physiol Lung Cell Mol Physiol 280, L1148-L1156. [本文引用: 2]
HeXJ, ZhangZG, YanDQ, ZhangJS, ChenSY (2004). A salt-responsive receptor-like kinase gene regulated by the ethylene signaling pathway encodes a plasma membrane serine/threonine kinase Theor Appl Genet 109, 377-383. [本文引用: 2]
HervéC, dabosP, GalaudJP, RougéP, LescureB (1996). Characterization of an Arabidopsis thaliana gene that defines a new class of putative plant receptor kinases with an extracellular lectin-like domain J Mol Biol 258, 778-788. [本文引用: 1]
HervéC, SerresJ, DabosP, CanutH, BarreA, RougéP, LescureB (1999). Characterization of the Arabidopsis lecRK-a genes: members of a superfamily encoding putative receptors with an extracellular domain homologous to legume lectins Plant Mol Biol 39, 671-682. [本文引用: 3]
HuX, ReddyASN (1997). Cloning and expression of a PR5-like protein from Arabidopsis: inhibition of fungal growth by bacterially expressed protein Plant Mol Biol 34, 949-959. [本文引用: 1]
JoshiA, DangHD, VaidN, TutejaN (2010). Pea lectin receptor-like kinase promotes high salinity stress tolerance in bacteria and expresses in response to stress in planta Glycoconj J 27, 133-150. [本文引用: 1]
KanzakiH, SaitohH, TakahashiY, BerberichT, ItoA, KamounS, TerauchiR (2008). NbLRK1, a lectin-like receptor kinase protein of Nicotiana benthamiana, interacts with Phytophthora infestans INF1 elicitin and mediates INF1-induced cell death Planta 228, 977-987. [本文引用: 1]
KobeB, KajavaAV (2001). The leucine-rich repeat as a protein recognition motif Curr Opin Struct Biol 11, 725-732. [本文引用: 1]
KusabaM, DwyerK, HendershotJ, VrebalovJ, NasrallahJB, NasrallahME (2001). Self-incompatibility in the genus Arabidopsis: characterization of the S locus in the outcrossing A. lyrata and its autogamous relative A. thaliana Plant Cell 13, 627-643. [本文引用: 1]
LiCH, WangG, ZhaoJL, ZhangLQ, AiLF, HanYF, SunDY, ZhangSW, SunY (2014). The receptor-like kinase SIT1 mediates salt sensitivity by activating MAPK3/6 and regulating ethylene homeostasis in rice Plant Cell 26, 2538-2553. [本文引用: 3]
LiHY, GrayJE (1997). Pollination-enhanced expression of a receptor-like protein kinase related gene in tobacco styles Plant Mol Biol 33, 653-665. [本文引用: 1]
LorisR (2002). Principles of structures of animal and plant lectins Biochim Biophys Acta 1572, 198-208. [本文引用: 1]
MaN, LiuCX, LiH, WangJY, ZhangBL, LinJ, ChangYH (2018). Genome-wide identification of lectin receptor kinases in pear: functional characterization of the L-type LecRLK gene PbLRK138 Gene 661, 11-21. [本文引用: 2]
MorilloSA, TaxFE (2006). Functional analysis of receptor-like kinases in monocots and dicots Curr Opin Plant Biol 9, 460-469. [本文引用: 1]
NaithaniS, ChookajornT, RipollDR, NasrallahJB (2007). Structural modules for receptor dimerization in the S-locus receptor kinase extracellular domain Proc Natl Acad Sci USA 104, 12211-12216. [本文引用: 1]
Navarro-GochicoaMT, CamutS, TimmersACJ, NiebelA, HervéC, BoutetE, BonoJJ, ImbertyA, CullimoreJV (2003). Characterization of four lectin-like receptor kinases expressed in roots of Medicago truncatula. Structure, location, regulation of expression, and potential role in the symbiosis with Sinorhizobium meliloti Plant Physiol 133, 1893-1910. [本文引用: 3]
NewmanT, de BruijnFJ, GreenP, KeegstraK, KendeH, McIntoshL, OhlroggeJ, RaikhelN, SomervilleS, ThomashowM, RetzelE, SomervilleC (1994). Genes galore: a summary of methods for accessing results from large-scale partial sequencing of anonymous Arabidopsis cDNA clones Plant Physiol 106, 1241-1255. [本文引用: 1]
NishiguchiM, YoshidaK, SumizonoT, TazakiK (2002). A receptor-like protein kinase with a lectin-like domain from Lombardy poplar: gene expression in response to wounding and characterization of phosphorylation activity Mol Genet Genomics 267, 506-514. [本文引用: 2]
PeumansWJ, van DammeEJM (1995). The role of lectins in plant defence Histochem J 27, 253-271. [本文引用: 2]
RiouC, HervéC, PacquitV, DabosP, LescureB (2002). Expression of an Arabidopsis lectin kinase receptor gene, lecRK-a1, is induced during senescence, wounding and in response to oligogalacturonic acids Plant Physiol Biochem 40, 431-438. [本文引用: 3]
SanabriaNM, van HeerdenH, DuberyIA (2012). Molecular characterisation and regulation of a Nicotiana tabacum S- domain receptor-like kinase gene induced during an early rapid response to lipopolysaccharides Gene 501, 39-48. [本文引用: 1]
Sherman-BroylesS, BoggsN, FarkasA, LiuP, VrebalovJ, NasrallahME, NasrallahJB (2007). S locus genes and the evolution of self-fertility in Arabidopsis thaliana Plant Cell 19, 94-106. [本文引用: 1]
ShiuSH, BleeckerAB (2001). Receptor-like kinases from Arabidopsis form a monophyletic gene family related to animal receptor kinases Proc Natl Acad Sci USA 98, 10763-10768. [本文引用: 6]
SinghP, KuoYC, MishraS, TsaiCH, ChienCC, ChenCW, Desclos-TheveniauM, ChuPW, SchulzeB, ChinchillaD, BollerT, ZimmerliL (2012). The lectin receptor kinase-VI.2 is required for priming and positively regulates Arabidopsis pattern-triggered immunity Plant Cell 24, 1256-1270. [本文引用: 3]
SunXL, YuQY, TangLL, JiW, BaiX, CaiH, LiuXF, DingXD, ZhuYM (2013). GsSRK, a G-type lectin S-receptor- like serine/threonine protein kinase, is a positive regulator of plant tolerance to salt stress J Plant Physiol 170, 505-515. [本文引用: 1]
TanksleySD, Loaiza-FigueroaF (1985). Gametophytic self-incompatibility is controlled by a single major locus on chromosome 1 in Lycopersicon peruvianum Proc Natl Acad Sci USA 82, 5093-5096. [本文引用: 1]
TordaiH, BányaiL, PatthyL (1999). The PAN module: the N-terminal domains of plasminogen and hepatocyte growth factor are homologous with the apple domains of the prekallikrein family and with a novel domain found in numerous nematode proteins FEBS Lett 461, 63-67. [本文引用: 1]
VaidN, MacoveiA, TutejaN (2013). Knights in action: lectin receptor-like kinases in plant development and stress responses Mol Plant 6, 1405-1418. [本文引用: 2]
VaidN, PandeyP, SrivastavaVK, TutejaN (2015). Pea lectin receptor-like kinase functions in salinity adaptation without yield penalty, by alleviating osmotic and ionic stresses and upregulating stress-responsive genes Plant Mol Biol 88, 193-206. [本文引用: 1]
VaidN, PandeyPK, TutejaN (2012). Genome-wide analysis of lectin receptor-like kinase family from Arabidopsis and rice Plant Mol Biol 80, 365-388. [本文引用: 7]
WalkerJC (1994). Structure and function of the receptor-like protein kinases of higher plants Plant Mol Biol 26, 1599-1609. [本文引用: 3]
WalkerJC, ZhangR (1990). Relationship of a putative receptor protein kinase from maize to the S-locus glycoproteins of Brassica Nature 345, 743-746. [本文引用: 1]
WanJR, PatelA, MathieuM, KimSY, XuD, StaceyG (2008). A lectin receptor-like kinase is required for pollen development in Arabidopsis Plant Mol Biol 67, 469-482. [本文引用: 2]
WangGL, RuanDL, SongWY, SiderisS, ChenLL, PiLY, ZhangSP, ZhangZ, FauquetC, GautBS, WhalenMC, RonaldPC (1998). Xa21D encodes a receptor-like molecule with a leucine-rich repeat domain that determines race-specific recognition and is subject to adaptive evolution Plant Cell 10, 765-779. [本文引用: 1]
WangY, CordewenerJHG, AmericaAHP, ShanWX, BouwmeesterK, GoversF (2015). Arabidopsis lectin receptor kinases LecRK-IX.1 and LecRK-IX.2 are functional analogs in regulating Phytophthora resistance and plant cell death Mol Plant Microbe Interact 28, 1032-1048. [本文引用: 2]
XinZY, WangA, YangGH, GaoP, ZhengZL (2009). The Arabidopsis A4 subfamily of lectin receptor kinases negatively regulates abscisic acid response in seed germination Plant Physiol 149, 434-444. [本文引用: 2]
ZuoKJ, ZhaoJY, WangJ, SunXF, TanKX (2004). Molecular cloning and characterization of GhLecRK, a novel kinase gene with lectin-like domain from Gossypium hirsutum DNA Seq 15, 58-65. [本文引用: 2]
... 凝集素类受体蛋白激酶主要由胞外区(包含凝集素结构域)、跨膜区以及胞内区(包含近膜区和激酶结构域)组成(图1).凝集素结构域可与糖类分子特异可逆结合, 该结构域在不同受体中差异较大, 可分为不同类型, 主要参与信号识别和结合(Peumans and van Damme, 1995; Vaid et al., 2012).跨膜结构域一般由18-25个氨基酸组成, 不具有明显的保守性, 但该部位对于凝集素类受体蛋白激酶的功能极其重要(Morillo and Tax, 2006).例如, 水稻Pi-d2蛋白跨膜结构域中的1个氨基酸发生改变, 可导致该蛋白完全丧失功能(Chen et al., 2006).近膜区主要是将跨膜结构域和激酶结构域分开, 该区由50个左右的氨基酸组成, 保守性较高, 其C端一般包含PxPFxYxELxxATxG (x表示任一氨基酸)的未知功能亲水区(Walker, 1994; Barre et al., 2002).激酶结构域是高度保守的结构域, 具有磷酸化结合位点, 位于细胞质区域, 一般由250- 300个氨基酸组成, 主要负责传递外界信号(Li and Gray, 1997).激酶结构域的N端包含GxGxxG结构域, 能够影响核苷酸的结合; 而激酶结构域的C端由43- 66个氨基酸组成, 为激酶的催化活性所必需(Barre et al., 2002).与酪氨酸类受体蛋白激酶含有DLAARN和FPIKWMAPE基序不同, 凝集素类受体蛋白激酶的激酶结构域中含有DIKPAN和GT(FIL)GYIAPE基序, 属于丝/苏氨酸激酶(Hervé et al., 1999). ...
... ).激酶结构域的N端包含GxGxxG结构域, 能够影响核苷酸的结合; 而激酶结构域的C端由43- 66个氨基酸组成, 为激酶的催化活性所必需(Barre et al., 2002).与酪氨酸类受体蛋白激酶含有DLAARN和FPIKWMAPE基序不同, 凝集素类受体蛋白激酶的激酶结构域中含有DIKPAN和GT(FIL)GYIAPE基序, 属于丝/苏氨酸激酶(Hervé et al., 1999). ...
... L型凝集素类受体蛋白激酶的细胞外结构含legume-lectin结构域, 位于N端, 大约由250个氨基酸组成, 该结构域呈现出典型的β-sandwich折叠, 具有糖类结合位点, 并保留了疏水腔结构, 可能涉及与疏水配体之间的互作(Edelman and Wang, 1978; Hervé et al., 1999; Barre et al., 2002).此外, 在该结构域中也包含Ca2+和Mn2+, 可稳定糖类结合位点(Bellande et al., 2017).大部分L型凝集素类受体蛋白激酶的凝集素结构域的糖类结合位点和与Ca2+、Mn2+结合的氨基酸都不保守(Hervé et al., 1999; Barre et al., 2002).L型凝集素类受体蛋白激酶通常以LecRLK形式命名. ...
... ; Barre et al., 2002).L型凝集素类受体蛋白激酶通常以LecRLK形式命名. ...
CRINKLY4: a TNFR-like receptor kinase involved in maize epidermal differentiation 1 1996
... 根据RLKs细胞外结构域的不同特点, 同时参照动物RLKs, 人们将植物RLKs分为6种类型: 富含亮氨酸重复序列型(leucine-rich repeats, LRRs)、S-结构域型(S-domain)、类凝集素型(lectin-like)、类表皮生长因子型(epidermal growth factor-like, EGF)、类肿瘤坏死因子型(tumor-necrosis factor receptor-like, TNFR)和类PR5型(pathogenesis related protein-5 like receptor kinases, PR5K) (Walker, 1994; Becraft et al., 1996; Hu and Reddy, 1997; Wang et al., 1998; Kobe and Kajava, 2001; Shiu and Bleecker, 2001).凝集素类受体蛋白激酶(lectin receptor-like kinases, LecRLKs)是RLKs家族中的一个亚族, 是指含有1个或多个可与单糖或寡聚糖特异可逆结合的非催化结构域的植物蛋白(Peumans and van Damme, 1995).已知在拟南芥和水稻中分别存在77和173个LecRLKs (Bouwmeester and Govers, 2009; Vaid et al., 2012).到目前为止, 在酵母(Saccharomyces cerevisiae)和人体基因组中还未找到LecRLKs的同源基因(Navarro-Gochicoa et al., 2003).已有研究表明, LecRLKs蛋白可参与多种生物/非生物胁迫途径以及植物发育调控(Vaid et al., 2012).本文简要阐述植物凝集素类受体蛋白激酶的研究历史、结构特点与分类及其生物学功能, 以期增进对凝集素类受体蛋白激酶的认识. ...
Plant lectins and lectin receptor-like kinases: how do they sense the outside? 2 2017
... L型凝集素类受体蛋白激酶的细胞外结构含legume-lectin结构域, 位于N端, 大约由250个氨基酸组成, 该结构域呈现出典型的β-sandwich折叠, 具有糖类结合位点, 并保留了疏水腔结构, 可能涉及与疏水配体之间的互作(Edelman and Wang, 1978; Hervé et al., 1999; Barre et al., 2002).此外, 在该结构域中也包含Ca2+和Mn2+, 可稳定糖类结合位点(Bellande et al., 2017).大部分L型凝集素类受体蛋白激酶的凝集素结构域的糖类结合位点和与Ca2+、Mn2+结合的氨基酸都不保守(Hervé et al., 1999; Barre et al., 2002).L型凝集素类受体蛋白激酶通常以LecRLK形式命名. ...
... 凝集素蛋白在植物整个生命过程中的作用非常重要, 并且在细胞与细胞的交流、植物的发育和防御反应中扮演重要角色(Bellande et al., 2017).已知凝集素蛋白可参与多种生理学过程, 包括蛋白分选、识别、胚胎发生和发育; 凝集素蛋白可与外源糖基结合, 从而保护植物免受食草动物或植食性无脊椎动物的侵害(Brewin and Kardailsky, 1997; Brill et al., 2001; Rudiger and Gabius, 2001).由于凝集素类受体蛋白激酶的凝集素识别结构域与凝集素蛋白相似, 推测其可能具有识别和转导糖信号的功能(Navarro-Gochicoa et al., 2003).近期的研究表明, 凝集素类受体蛋白激酶在响应胁迫信号、抗病虫害和调控植物发育方面都具有非常重要的功能(表1; 图2). ...
The Nicotiana attenuata lectin receptor kinase 1 is involved in the perception of insect feeding 3 2011
... ZmPK1是第1个被鉴定到的类受体蛋白激酶, 它属于G型凝集素类受体蛋白激酶.该激酶的细胞外结构域与芸薹属自交不亲和性位点编码的糖蛋白类似, 参与调控花粉与柱头之间的识别(Walker and Zhang, 1990).Hervé等(1996)在拟南芥中鉴定到第1个L型凝集素类受体蛋白激酶Ath.LecRK1, 该蛋白激酶具有豆类类凝集素结构, 可能在植物低聚糖和激素信号转导过程中发挥作用(Newman et al., 1994).根据LecRLKs细胞外凝集素结构域的特点, LecRLKs被归类为RLKs的一个亚家族(Shiu and Bleecker, 2001; Goff et al., 2002; Yu et al., 2002).虽然LecRLKs基因发现较早, 但对这些基因家族的系统分类却相对较晚, 而早期对于LecRLKs基因的研究也主要集中在基因表达等较为初步的层面(Nishiguchi et al., 2002; Riou et al., 2002; Navarro-Gochicoa et al., 2003; He et al., 2004; Zuo et al., 2004).近年来, 越来越多的研究表明, LecRLKs蛋白的功能极其广泛, 可参与调控各种生物/非生物胁迫和植物发育(Chen et al., 2006; Kanzaki et al., 2008; Joshi et al., 2010; Bonaventure, 2011; Desclos-Theveniau et al., 2012; Gouhier-Darimont et al., 2013; Li et al., 2014; Ranf et al., 2015; Luo et al., 2017; Ma et al., 2018; Labbé et al., 2019) (表1). ...
... 凝集素类受体蛋白激酶对于植物的抗虫性也极其重要.例如, 在野生烟草(N. attenuata)中, LecRK1基因在烟草天蛾触发所引起的植物防御反应中发挥重要作用(Gilardoni et al., 2011).RNA干涉LecRK1基因表达表明, LecRK1基因对于全面防御抵抗烟草天蛾是必需的.例如, 生长在LecRK1基因干涉植株中的烟草天蛾幼虫比生长在野生型上的幼虫体积大40%-100%; 相对于野生型植株, 昆虫诱导积累的尼古丁、二萜糖甙、胰蛋白酶抑制剂和苏氨酸脱氨酶在干涉植株中的含量也明显减少(Bonaventure, 2011; Gilardoni et al., 2011).在拟南芥中, 昆虫卵衍生的诱导物能激活植物的防御反应, 该反应类似于植物识别PAMPs (pathogen-associated molecular pat- terns)所产生的PTI反应.例如, LecRK-I.8基因在昆虫卵衍生的诱导物识别中发挥重要功能, 该基因突变后会抑制昆虫卵所诱导的抗病基因PR-1的表达(Gouhier- Darimont et al., 2013). ...
... 凝集素类受体蛋白激酶参与激素信号响应.(1)参与脱落酸(ABA)信号响应.在拟南芥中, 过表达凝集素类受体蛋白激酶基因LecRK-V.5可抑制植物对ABA的响应能力, 从而影响气孔开闭(Desclos-Theveniau et al., 2012).Xin等(2009)发现, L型凝集素类受体蛋白激酶VI亚家族成员基因LecRK-VI.2、LecRK-VI.3和LecRK-VI.4突变后都会微弱增加ABA对种子萌发的抑制作用.相对于LecRK- VI.2单基因突变, LecRK- VI.2和LecRK-VI.3双基因突变时ABA对种子萌发的抑制作用更显著.基因表达谱和网络调控分析表明, LecRK-VI.2和LecRK-VI.3基因还可调控ABA相关基因的表达(Xin et al., 2009).Zhang等(2019)通过对拟南芥凝集素类受体蛋白激酶基因LecRK-VI.4进行深入研究后, 发现该基因在ABA介导的气孔开闭中行使重要功能.(2) 参与茉莉酸(JA)信号响应.野生烟草中LecRK1基因突变会抑制由JA引起的防御途径(Bonaventure, 2011; Gilardoni et al., 2011).(3) 参与水杨酸(SA)信号响应.在拟南芥中过表达LecRK-IX.2基因导致SA积累, 从而引起细胞死亡(Luo et al., 2017). ...
... 根据RLKs细胞外结构域的不同特点, 同时参照动物RLKs, 人们将植物RLKs分为6种类型: 富含亮氨酸重复序列型(leucine-rich repeats, LRRs)、S-结构域型(S-domain)、类凝集素型(lectin-like)、类表皮生长因子型(epidermal growth factor-like, EGF)、类肿瘤坏死因子型(tumor-necrosis factor receptor-like, TNFR)和类PR5型(pathogenesis related protein-5 like receptor kinases, PR5K) (Walker, 1994; Becraft et al., 1996; Hu and Reddy, 1997; Wang et al., 1998; Kobe and Kajava, 2001; Shiu and Bleecker, 2001).凝集素类受体蛋白激酶(lectin receptor-like kinases, LecRLKs)是RLKs家族中的一个亚族, 是指含有1个或多个可与单糖或寡聚糖特异可逆结合的非催化结构域的植物蛋白(Peumans and van Damme, 1995).已知在拟南芥和水稻中分别存在77和173个LecRLKs (Bouwmeester and Govers, 2009; Vaid et al., 2012).到目前为止, 在酵母(Saccharomyces cerevisiae)和人体基因组中还未找到LecRLKs的同源基因(Navarro-Gochicoa et al., 2003).已有研究表明, LecRLKs蛋白可参与多种生物/非生物胁迫途径以及植物发育调控(Vaid et al., 2012).本文简要阐述植物凝集素类受体蛋白激酶的研究历史、结构特点与分类及其生物学功能, 以期增进对凝集素类受体蛋白激酶的认识. ...
... 根据细胞外凝集素结构域的多样性可将凝集素类受体蛋白激酶分为L、G和C型(图1), 其中L和G型为植物特有, 而C型主要存在于哺乳动物中(Wan et al., 2008; Vaid et al., 2013).研究显示, 拟南芥中包含77个凝集素类受体蛋白激酶.而Vaid等(2012)明确指出拟南芥中L、G和C型凝集素类受体蛋白激酶分别为42、32和1个, 其中L型凝集素类受体蛋白激酶不包含LecRK S.2和LecRK S.3两个蛋白激酶(Bouwmeester and Govers, 2009).LecRK S.2和LecRK S.3蛋白含激酶结构域, 无胞外结构域和跨膜结构域.目前暂无明确证据证明这2个类受体蛋白激酶不属于凝集素类受体蛋白激酶亚家族(Bouwmeester and Govers, 2009).在水稻中, L、G和C型凝集素类受体蛋白激酶分别为72、100和1个(Vaid et al., 2012). ...
... ).LecRK S.2和LecRK S.3蛋白含激酶结构域, 无胞外结构域和跨膜结构域.目前暂无明确证据证明这2个类受体蛋白激酶不属于凝集素类受体蛋白激酶亚家族(Bouwmeester and Govers, 2009).在水稻中, L、G和C型凝集素类受体蛋白激酶分别为72、100和1个(Vaid et al., 2012). ...
... C型凝集素类受体蛋白激酶为钙依赖的凝集素类受体蛋白激酶, 是哺乳动物中的一个大家族蛋白, 主要参与先天性免疫反应以及病原体的识别(Epstein et al., 1996; Hawgood et al., 2001; Cambi et al., 2005).该类型凝集素类受体蛋白激酶的凝集素结构域大量存在于哺乳动物中, 通常通过自我和非自我识别参与免疫反应(Epstein et al., 1996; Hawgood et al., 2001).在植物中, C型凝集素类受体蛋白激酶十分稀少, 属于凝集素类受体蛋白激酶家族中最小的类群.到目前为止, 在水稻和拟南芥中分别仅发现1个C型凝集素类受体蛋白激酶(Bouwmeester and Govers, 2009; Vaid et al., 2012). ...
... 由于凝集素类受体蛋白激酶的细胞外结构域与凝集素蛋白相似, 而凝集素蛋白可以与真菌和细菌细胞壁的成分结合, 因此推测凝集素类受体蛋白激酶可以参与对生物胁迫的响应(Bouwmeester and Govers, 2009). ...
Legume lectins and nodulation by Rhizobium 1 1997
... 凝集素蛋白在植物整个生命过程中的作用非常重要, 并且在细胞与细胞的交流、植物的发育和防御反应中扮演重要角色(Bellande et al., 2017).已知凝集素蛋白可参与多种生理学过程, 包括蛋白分选、识别、胚胎发生和发育; 凝集素蛋白可与外源糖基结合, 从而保护植物免受食草动物或植食性无脊椎动物的侵害(Brewin and Kardailsky, 1997; Brill et al., 2001; Rudiger and Gabius, 2001).由于凝集素类受体蛋白激酶的凝集素识别结构域与凝集素蛋白相似, 推测其可能具有识别和转导糖信号的功能(Navarro-Gochicoa et al., 2003).近期的研究表明, 凝集素类受体蛋白激酶在响应胁迫信号、抗病虫害和调控植物发育方面都具有非常重要的功能(表1; 图2). ...
Expression of MsLEC1- and MsLEC2-antisense genes in alfalfa plant lines causes severe embryogenic, developmental and reproductive abnormalities 1 2001
... 凝集素蛋白在植物整个生命过程中的作用非常重要, 并且在细胞与细胞的交流、植物的发育和防御反应中扮演重要角色(Bellande et al., 2017).已知凝集素蛋白可参与多种生理学过程, 包括蛋白分选、识别、胚胎发生和发育; 凝集素蛋白可与外源糖基结合, 从而保护植物免受食草动物或植食性无脊椎动物的侵害(Brewin and Kardailsky, 1997; Brill et al., 2001; Rudiger and Gabius, 2001).由于凝集素类受体蛋白激酶的凝集素识别结构域与凝集素蛋白相似, 推测其可能具有识别和转导糖信号的功能(Navarro-Gochicoa et al., 2003).近期的研究表明, 凝集素类受体蛋白激酶在响应胁迫信号、抗病虫害和调控植物发育方面都具有非常重要的功能(表1; 图2). ...
How C-type lectins detect pathogens 1 2005
... C型凝集素类受体蛋白激酶为钙依赖的凝集素类受体蛋白激酶, 是哺乳动物中的一个大家族蛋白, 主要参与先天性免疫反应以及病原体的识别(Epstein et al., 1996; Hawgood et al., 2001; Cambi et al., 2005).该类型凝集素类受体蛋白激酶的凝集素结构域大量存在于哺乳动物中, 通常通过自我和非自我识别参与免疫反应(Epstein et al., 1996; Hawgood et al., 2001).在植物中, C型凝集素类受体蛋白激酶十分稀少, 属于凝集素类受体蛋白激酶家族中最小的类群.到目前为止, 在水稻和拟南芥中分别仅发现1个C型凝集素类受体蛋白激酶(Bouwmeester and Govers, 2009; Vaid et al., 2012). ...
... ZmPK1是第1个被鉴定到的类受体蛋白激酶, 它属于G型凝集素类受体蛋白激酶.该激酶的细胞外结构域与芸薹属自交不亲和性位点编码的糖蛋白类似, 参与调控花粉与柱头之间的识别(Walker and Zhang, 1990).Hervé等(1996)在拟南芥中鉴定到第1个L型凝集素类受体蛋白激酶Ath.LecRK1, 该蛋白激酶具有豆类类凝集素结构, 可能在植物低聚糖和激素信号转导过程中发挥作用(Newman et al., 1994).根据LecRLKs细胞外凝集素结构域的特点, LecRLKs被归类为RLKs的一个亚家族(Shiu and Bleecker, 2001; Goff et al., 2002; Yu et al., 2002).虽然LecRLKs基因发现较早, 但对这些基因家族的系统分类却相对较晚, 而早期对于LecRLKs基因的研究也主要集中在基因表达等较为初步的层面(Nishiguchi et al., 2002; Riou et al., 2002; Navarro-Gochicoa et al., 2003; He et al., 2004; Zuo et al., 2004).近年来, 越来越多的研究表明, LecRLKs蛋白的功能极其广泛, 可参与调控各种生物/非生物胁迫和植物发育(Chen et al., 2006; Kanzaki et al., 2008; Joshi et al., 2010; Bonaventure, 2011; Desclos-Theveniau et al., 2012; Gouhier-Darimont et al., 2013; Li et al., 2014; Ranf et al., 2015; Luo et al., 2017; Ma et al., 2018; Labbé et al., 2019) (表1). ...
... 凝集素类受体蛋白激酶主要由胞外区(包含凝集素结构域)、跨膜区以及胞内区(包含近膜区和激酶结构域)组成(图1).凝集素结构域可与糖类分子特异可逆结合, 该结构域在不同受体中差异较大, 可分为不同类型, 主要参与信号识别和结合(Peumans and van Damme, 1995; Vaid et al., 2012).跨膜结构域一般由18-25个氨基酸组成, 不具有明显的保守性, 但该部位对于凝集素类受体蛋白激酶的功能极其重要(Morillo and Tax, 2006).例如, 水稻Pi-d2蛋白跨膜结构域中的1个氨基酸发生改变, 可导致该蛋白完全丧失功能(Chen et al., 2006).近膜区主要是将跨膜结构域和激酶结构域分开, 该区由50个左右的氨基酸组成, 保守性较高, 其C端一般包含PxPFxYxELxxATxG (x表示任一氨基酸)的未知功能亲水区(Walker, 1994; Barre et al., 2002).激酶结构域是高度保守的结构域, 具有磷酸化结合位点, 位于细胞质区域, 一般由250- 300个氨基酸组成, 主要负责传递外界信号(Li and Gray, 1997).激酶结构域的N端包含GxGxxG结构域, 能够影响核苷酸的结合; 而激酶结构域的C端由43- 66个氨基酸组成, 为激酶的催化活性所必需(Barre et al., 2002).与酪氨酸类受体蛋白激酶含有DLAARN和FPIKWMAPE基序不同, 凝集素类受体蛋白激酶的激酶结构域中含有DIKPAN和GT(FIL)GYIAPE基序, 属于丝/苏氨酸激酶(Hervé et al., 1999). ...
... 凝集素类受体蛋白激酶参与植物对病原菌的响应.例如, 水稻Pi-d2蛋白属于G型凝集素类受体蛋白激酶, 其转基因植株对中国稻瘟病菌小种ZB15具有较高的抗性(Chen et al., 2006).Fan等(2018)通过EMS诱变筛选获得E3泛素连接酶基因SPL11突变所诱导细胞死亡的抑制子SDS2, 该基因编码1个S-domain类受体蛋白激酶, 属于G型凝集素类受体蛋白激酶.研究表明, SDS2基因突变可降低植物的免疫反应, 易感染稻瘟病菌; 相反, 过表达SDS2基因导致细胞程序性死亡、增强免疫反应和对稻瘟病菌的抗性.此外, SDS2蛋白可与细胞质类受体蛋白激酶OsRLCK118/176互作, 通过磷酸化调控NADPH氧化酶OsRbohB, 刺激ROS产生, 从而正向调控免疫反应(Fan et al., 2018).在烟草(Nicotiana tabacum)中, G型凝集素类受体蛋白激酶基因Nt-Sd-RLK受脂多糖诱导表达, 在免疫信号的识别和防御过程中发挥重要作用(Sanabria et al., 2012).Ranf等(2015)通过筛选脂多糖不敏感型缺陷突变体, 获得了1个S-domain类受体蛋白激酶基因LORE.LORE基因突变导致植物易感染丁香假单胞杆菌, 它在介导脂多糖识别引起的免疫反应中发挥功能.在拟南芥中, 缺失L型凝集素类受体蛋白激酶基因LecRK-V.5会增强植物对表面接种Pst DC3000而产生的抗性, 但对通过渗透方法接种所产生的抗性无影响, 提示LecRK-V.5基因可能在植物早期免疫阶段发挥功能; 相反, 过表达LecRK- V.5基因导致植物更易感染Pst DC3000 (Desclos- Theveniau et al., 2012).研究表明, LecRK-V.5基因突变引起的抗病性增强表型与气孔的组成型关闭相关, 而LecRK-V.5基因过表达引起的易感病表型与早期气孔的重新打开有关.LecRK-V.5基因突变和过表达表型结果表明, LecRK-V.5基因可以逆转由于细菌感染诱导的气孔关闭, 从而减轻植株因缺乏CO2而造成的伤害(Desclos-Theveniau et al., 2012).另一个凝集素类受体蛋白激酶基因LecRK-VI.2被报道在抵抗丁香假单胞杆菌和胡萝卜软腐果胶杆菌方面具有重要功能(Singh et al., 2012).LecRK-VI.2基因突变导致PTI (pattern-triggered immunity)反应相关抗病基因表达量降低、胼胝质沉淀形成受阻和气孔关闭出现缺陷等.LecRK-VI.2基因过表达和全基因组分析实验表明, LecRK-VI.2基因正向调控PTI反应.Singh等(2012)研究表明, LecRK-VI.2蛋白位于MAPK信号的上游, 但独立于ROS信号和BIK1的磷酸化调控信号途径.此外, LecRK-VI.2蛋白对于β-氨基丁酸诱导的抗性反应和PTI反应是必需的(Singh et al., 2012).在拟南芥中, LecRK-I.9基因突变导致植物对ATP不敏感, 从而降低ATP引起的防御反应.研究发现, LecRK-I.9蛋白与ATP结合且高度亲和, 它们的结合对于ATP诱导的钙离子响应、MAPK的激活和基因表达是必需的(Choi et al., 2014).Wang等(2015)研究发现, 拟南芥LecRK-IX.1和LecRK-IX.2基因突变可降低植物对疫霉菌的抵抗能力, 且双突变会加剧表型; 相反, 过表达LecRK-IX.1和LecRK-IX.2基因能增强植物的抗病性, 但同时导致自发性的细胞死亡.此外, LecRK-IX.1和LecRK-IX.2蛋白的凝集素结构域和激酶结构域对于抗疫霉菌是必需的; 而凝集素结构域对于诱导细胞死亡为非必需(Wang et al., 2015). ...
Identi?cation of a plant receptor for extracellular ATP 1 2014
... 凝集素类受体蛋白激酶参与植物对病原菌的响应.例如, 水稻Pi-d2蛋白属于G型凝集素类受体蛋白激酶, 其转基因植株对中国稻瘟病菌小种ZB15具有较高的抗性(Chen et al., 2006).Fan等(2018)通过EMS诱变筛选获得E3泛素连接酶基因SPL11突变所诱导细胞死亡的抑制子SDS2, 该基因编码1个S-domain类受体蛋白激酶, 属于G型凝集素类受体蛋白激酶.研究表明, SDS2基因突变可降低植物的免疫反应, 易感染稻瘟病菌; 相反, 过表达SDS2基因导致细胞程序性死亡、增强免疫反应和对稻瘟病菌的抗性.此外, SDS2蛋白可与细胞质类受体蛋白激酶OsRLCK118/176互作, 通过磷酸化调控NADPH氧化酶OsRbohB, 刺激ROS产生, 从而正向调控免疫反应(Fan et al., 2018).在烟草(Nicotiana tabacum)中, G型凝集素类受体蛋白激酶基因Nt-Sd-RLK受脂多糖诱导表达, 在免疫信号的识别和防御过程中发挥重要作用(Sanabria et al., 2012).Ranf等(2015)通过筛选脂多糖不敏感型缺陷突变体, 获得了1个S-domain类受体蛋白激酶基因LORE.LORE基因突变导致植物易感染丁香假单胞杆菌, 它在介导脂多糖识别引起的免疫反应中发挥功能.在拟南芥中, 缺失L型凝集素类受体蛋白激酶基因LecRK-V.5会增强植物对表面接种Pst DC3000而产生的抗性, 但对通过渗透方法接种所产生的抗性无影响, 提示LecRK-V.5基因可能在植物早期免疫阶段发挥功能; 相反, 过表达LecRK- V.5基因导致植物更易感染Pst DC3000 (Desclos- Theveniau et al., 2012).研究表明, LecRK-V.5基因突变引起的抗病性增强表型与气孔的组成型关闭相关, 而LecRK-V.5基因过表达引起的易感病表型与早期气孔的重新打开有关.LecRK-V.5基因突变和过表达表型结果表明, LecRK-V.5基因可以逆转由于细菌感染诱导的气孔关闭, 从而减轻植株因缺乏CO2而造成的伤害(Desclos-Theveniau et al., 2012).另一个凝集素类受体蛋白激酶基因LecRK-VI.2被报道在抵抗丁香假单胞杆菌和胡萝卜软腐果胶杆菌方面具有重要功能(Singh et al., 2012).LecRK-VI.2基因突变导致PTI (pattern-triggered immunity)反应相关抗病基因表达量降低、胼胝质沉淀形成受阻和气孔关闭出现缺陷等.LecRK-VI.2基因过表达和全基因组分析实验表明, LecRK-VI.2基因正向调控PTI反应.Singh等(2012)研究表明, LecRK-VI.2蛋白位于MAPK信号的上游, 但独立于ROS信号和BIK1的磷酸化调控信号途径.此外, LecRK-VI.2蛋白对于β-氨基丁酸诱导的抗性反应和PTI反应是必需的(Singh et al., 2012).在拟南芥中, LecRK-I.9基因突变导致植物对ATP不敏感, 从而降低ATP引起的防御反应.研究发现, LecRK-I.9蛋白与ATP结合且高度亲和, 它们的结合对于ATP诱导的钙离子响应、MAPK的激活和基因表达是必需的(Choi et al., 2014).Wang等(2015)研究发现, 拟南芥LecRK-IX.1和LecRK-IX.2基因突变可降低植物对疫霉菌的抵抗能力, 且双突变会加剧表型; 相反, 过表达LecRK-IX.1和LecRK-IX.2基因能增强植物的抗病性, 但同时导致自发性的细胞死亡.此外, LecRK-IX.1和LecRK-IX.2蛋白的凝集素结构域和激酶结构域对于抗疫霉菌是必需的; 而凝集素结构域对于诱导细胞死亡为非必需(Wang et al., 2015). ...
A lectin receptor kinase positively regulates ABA response during seed germination and is involved in salt and osmotic stress response 1 2009
... 凝集素类受体蛋白激酶参与盐胁迫响应.拟南芥LecRK-V.2基因缺失会降低植物早期发育阶段对盐的敏感性(Deng et al., 2009).He等(2004)发现, 拟南芥LecRK2基因在盐胁迫时其表达量有不同程度的上调, 并进一步证明它受到乙烯(ET)信号通路的调控.豌豆(Pisum sativum) PsLecRLK基因在盐胁迫下被诱导表达, 而PsLecRLK基因过表达可缓解离子平衡和渗透压, 从而增强植物的耐盐性(Vaid et al., 2015).Sun等(2013)发现野大豆(Glycine soja) GsSRK基因的表达受盐胁迫诱导, 而过表达该基因可增强植物的耐盐性, 并使植物在耐盐环境中表现出更高的产量, 同时还伴随叶绿素含量增加、离子泄漏降低、株高增加和果实增多等表型.在水稻中, 盐胁迫能够增加SIT1蛋白激酶活性, 从而磷酸化其下游效应蛋白MAPK3和MAPK6; 此外, SIT1蛋白还可介导乙烯的生成和盐胁迫诱导的乙烯信号途径(Li et al., 2014).而在拟南芥中, SIT1蛋白激酶在盐胁迫下还可促进活性氧的积累, 最终导致植物生长受抑制甚至死亡, 该过程依赖于MAPK3/6和乙烯信号途径(Li et al., 2014).Ma等(2018)发现在本氏烟草(N. benthamiana)中过表达豆梨(Pyrus calleryana) L型凝集素类受体蛋白激酶基因PbLRK138导致细胞死亡, 从而增强植物的耐盐性. ...
The Arabidopsis lectin receptor kinase LecRK-V.5 represses stomatal immunity induced by Pseudomonas syringae pv. tomato DC3000 4 2012
... ZmPK1是第1个被鉴定到的类受体蛋白激酶, 它属于G型凝集素类受体蛋白激酶.该激酶的细胞外结构域与芸薹属自交不亲和性位点编码的糖蛋白类似, 参与调控花粉与柱头之间的识别(Walker and Zhang, 1990).Hervé等(1996)在拟南芥中鉴定到第1个L型凝集素类受体蛋白激酶Ath.LecRK1, 该蛋白激酶具有豆类类凝集素结构, 可能在植物低聚糖和激素信号转导过程中发挥作用(Newman et al., 1994).根据LecRLKs细胞外凝集素结构域的特点, LecRLKs被归类为RLKs的一个亚家族(Shiu and Bleecker, 2001; Goff et al., 2002; Yu et al., 2002).虽然LecRLKs基因发现较早, 但对这些基因家族的系统分类却相对较晚, 而早期对于LecRLKs基因的研究也主要集中在基因表达等较为初步的层面(Nishiguchi et al., 2002; Riou et al., 2002; Navarro-Gochicoa et al., 2003; He et al., 2004; Zuo et al., 2004).近年来, 越来越多的研究表明, LecRLKs蛋白的功能极其广泛, 可参与调控各种生物/非生物胁迫和植物发育(Chen et al., 2006; Kanzaki et al., 2008; Joshi et al., 2010; Bonaventure, 2011; Desclos-Theveniau et al., 2012; Gouhier-Darimont et al., 2013; Li et al., 2014; Ranf et al., 2015; Luo et al., 2017; Ma et al., 2018; Labbé et al., 2019) (表1). ...
... 凝集素类受体蛋白激酶参与植物对病原菌的响应.例如, 水稻Pi-d2蛋白属于G型凝集素类受体蛋白激酶, 其转基因植株对中国稻瘟病菌小种ZB15具有较高的抗性(Chen et al., 2006).Fan等(2018)通过EMS诱变筛选获得E3泛素连接酶基因SPL11突变所诱导细胞死亡的抑制子SDS2, 该基因编码1个S-domain类受体蛋白激酶, 属于G型凝集素类受体蛋白激酶.研究表明, SDS2基因突变可降低植物的免疫反应, 易感染稻瘟病菌; 相反, 过表达SDS2基因导致细胞程序性死亡、增强免疫反应和对稻瘟病菌的抗性.此外, SDS2蛋白可与细胞质类受体蛋白激酶OsRLCK118/176互作, 通过磷酸化调控NADPH氧化酶OsRbohB, 刺激ROS产生, 从而正向调控免疫反应(Fan et al., 2018).在烟草(Nicotiana tabacum)中, G型凝集素类受体蛋白激酶基因Nt-Sd-RLK受脂多糖诱导表达, 在免疫信号的识别和防御过程中发挥重要作用(Sanabria et al., 2012).Ranf等(2015)通过筛选脂多糖不敏感型缺陷突变体, 获得了1个S-domain类受体蛋白激酶基因LORE.LORE基因突变导致植物易感染丁香假单胞杆菌, 它在介导脂多糖识别引起的免疫反应中发挥功能.在拟南芥中, 缺失L型凝集素类受体蛋白激酶基因LecRK-V.5会增强植物对表面接种Pst DC3000而产生的抗性, 但对通过渗透方法接种所产生的抗性无影响, 提示LecRK-V.5基因可能在植物早期免疫阶段发挥功能; 相反, 过表达LecRK- V.5基因导致植物更易感染Pst DC3000 (Desclos- Theveniau et al., 2012).研究表明, LecRK-V.5基因突变引起的抗病性增强表型与气孔的组成型关闭相关, 而LecRK-V.5基因过表达引起的易感病表型与早期气孔的重新打开有关.LecRK-V.5基因突变和过表达表型结果表明, LecRK-V.5基因可以逆转由于细菌感染诱导的气孔关闭, 从而减轻植株因缺乏CO2而造成的伤害(Desclos-Theveniau et al., 2012).另一个凝集素类受体蛋白激酶基因LecRK-VI.2被报道在抵抗丁香假单胞杆菌和胡萝卜软腐果胶杆菌方面具有重要功能(Singh et al., 2012).LecRK-VI.2基因突变导致PTI (pattern-triggered immunity)反应相关抗病基因表达量降低、胼胝质沉淀形成受阻和气孔关闭出现缺陷等.LecRK-VI.2基因过表达和全基因组分析实验表明, LecRK-VI.2基因正向调控PTI反应.Singh等(2012)研究表明, LecRK-VI.2蛋白位于MAPK信号的上游, 但独立于ROS信号和BIK1的磷酸化调控信号途径.此外, LecRK-VI.2蛋白对于β-氨基丁酸诱导的抗性反应和PTI反应是必需的(Singh et al., 2012).在拟南芥中, LecRK-I.9基因突变导致植物对ATP不敏感, 从而降低ATP引起的防御反应.研究发现, LecRK-I.9蛋白与ATP结合且高度亲和, 它们的结合对于ATP诱导的钙离子响应、MAPK的激活和基因表达是必需的(Choi et al., 2014).Wang等(2015)研究发现, 拟南芥LecRK-IX.1和LecRK-IX.2基因突变可降低植物对疫霉菌的抵抗能力, 且双突变会加剧表型; 相反, 过表达LecRK-IX.1和LecRK-IX.2基因能增强植物的抗病性, 但同时导致自发性的细胞死亡.此外, LecRK-IX.1和LecRK-IX.2蛋白的凝集素结构域和激酶结构域对于抗疫霉菌是必需的; 而凝集素结构域对于诱导细胞死亡为非必需(Wang et al., 2015). ...
... 而造成的伤害(Desclos-Theveniau et al., 2012).另一个凝集素类受体蛋白激酶基因LecRK-VI.2被报道在抵抗丁香假单胞杆菌和胡萝卜软腐果胶杆菌方面具有重要功能(Singh et al., 2012).LecRK-VI.2基因突变导致PTI (pattern-triggered immunity)反应相关抗病基因表达量降低、胼胝质沉淀形成受阻和气孔关闭出现缺陷等.LecRK-VI.2基因过表达和全基因组分析实验表明, LecRK-VI.2基因正向调控PTI反应.Singh等(2012)研究表明, LecRK-VI.2蛋白位于MAPK信号的上游, 但独立于ROS信号和BIK1的磷酸化调控信号途径.此外, LecRK-VI.2蛋白对于β-氨基丁酸诱导的抗性反应和PTI反应是必需的(Singh et al., 2012).在拟南芥中, LecRK-I.9基因突变导致植物对ATP不敏感, 从而降低ATP引起的防御反应.研究发现, LecRK-I.9蛋白与ATP结合且高度亲和, 它们的结合对于ATP诱导的钙离子响应、MAPK的激活和基因表达是必需的(Choi et al., 2014).Wang等(2015)研究发现, 拟南芥LecRK-IX.1和LecRK-IX.2基因突变可降低植物对疫霉菌的抵抗能力, 且双突变会加剧表型; 相反, 过表达LecRK-IX.1和LecRK-IX.2基因能增强植物的抗病性, 但同时导致自发性的细胞死亡.此外, LecRK-IX.1和LecRK-IX.2蛋白的凝集素结构域和激酶结构域对于抗疫霉菌是必需的; 而凝集素结构域对于诱导细胞死亡为非必需(Wang et al., 2015). ...
... 凝集素类受体蛋白激酶参与激素信号响应.(1)参与脱落酸(ABA)信号响应.在拟南芥中, 过表达凝集素类受体蛋白激酶基因LecRK-V.5可抑制植物对ABA的响应能力, 从而影响气孔开闭(Desclos-Theveniau et al., 2012).Xin等(2009)发现, L型凝集素类受体蛋白激酶VI亚家族成员基因LecRK-VI.2、LecRK-VI.3和LecRK-VI.4突变后都会微弱增加ABA对种子萌发的抑制作用.相对于LecRK- VI.2单基因突变, LecRK- VI.2和LecRK-VI.3双基因突变时ABA对种子萌发的抑制作用更显著.基因表达谱和网络调控分析表明, LecRK-VI.2和LecRK-VI.3基因还可调控ABA相关基因的表达(Xin et al., 2009).Zhang等(2019)通过对拟南芥凝集素类受体蛋白激酶基因LecRK-VI.4进行深入研究后, 发现该基因在ABA介导的气孔开闭中行使重要功能.(2) 参与茉莉酸(JA)信号响应.野生烟草中LecRK1基因突变会抑制由JA引起的防御途径(Bonaventure, 2011; Gilardoni et al., 2011).(3) 参与水杨酸(SA)信号响应.在拟南芥中过表达LecRK-IX.2基因导致SA积累, 从而引起细胞死亡(Luo et al., 2017). ...
Binding and functional properties of concanavalin A and its derivatives. III. Interactions with indoleacetic acid and other hydrophobic ligands 1 1978
... L型凝集素类受体蛋白激酶的细胞外结构含legume-lectin结构域, 位于N端, 大约由250个氨基酸组成, 该结构域呈现出典型的β-sandwich折叠, 具有糖类结合位点, 并保留了疏水腔结构, 可能涉及与疏水配体之间的互作(Edelman and Wang, 1978; Hervé et al., 1999; Barre et al., 2002).此外, 在该结构域中也包含Ca2+和Mn2+, 可稳定糖类结合位点(Bellande et al., 2017).大部分L型凝集素类受体蛋白激酶的凝集素结构域的糖类结合位点和与Ca2+、Mn2+结合的氨基酸都不保守(Hervé et al., 1999; Barre et al., 2002).L型凝集素类受体蛋白激酶通常以LecRLK形式命名. ...
The collectins in innate immunity 2 1996
... C型凝集素类受体蛋白激酶为钙依赖的凝集素类受体蛋白激酶, 是哺乳动物中的一个大家族蛋白, 主要参与先天性免疫反应以及病原体的识别(Epstein et al., 1996; Hawgood et al., 2001; Cambi et al., 2005).该类型凝集素类受体蛋白激酶的凝集素结构域大量存在于哺乳动物中, 通常通过自我和非自我识别参与免疫反应(Epstein et al., 1996; Hawgood et al., 2001).在植物中, C型凝集素类受体蛋白激酶十分稀少, 属于凝集素类受体蛋白激酶家族中最小的类群.到目前为止, 在水稻和拟南芥中分别仅发现1个C型凝集素类受体蛋白激酶(Bouwmeester and Govers, 2009; Vaid et al., 2012). ...
... ).该类型凝集素类受体蛋白激酶的凝集素结构域大量存在于哺乳动物中, 通常通过自我和非自我识别参与免疫反应(Epstein et al., 1996; Hawgood et al., 2001).在植物中, C型凝集素类受体蛋白激酶十分稀少, 属于凝集素类受体蛋白激酶家族中最小的类群.到目前为止, 在水稻和拟南芥中分别仅发现1个C型凝集素类受体蛋白激酶(Bouwmeester and Govers, 2009; Vaid et al., 2012). ...
The monocot-specific receptor-like kinase SDS2 controls cell death and immunity in rice 2 2018
... 凝集素类受体蛋白激酶参与植物对病原菌的响应.例如, 水稻Pi-d2蛋白属于G型凝集素类受体蛋白激酶, 其转基因植株对中国稻瘟病菌小种ZB15具有较高的抗性(Chen et al., 2006).Fan等(2018)通过EMS诱变筛选获得E3泛素连接酶基因SPL11突变所诱导细胞死亡的抑制子SDS2, 该基因编码1个S-domain类受体蛋白激酶, 属于G型凝集素类受体蛋白激酶.研究表明, SDS2基因突变可降低植物的免疫反应, 易感染稻瘟病菌; 相反, 过表达SDS2基因导致细胞程序性死亡、增强免疫反应和对稻瘟病菌的抗性.此外, SDS2蛋白可与细胞质类受体蛋白激酶OsRLCK118/176互作, 通过磷酸化调控NADPH氧化酶OsRbohB, 刺激ROS产生, 从而正向调控免疫反应(Fan et al., 2018).在烟草(Nicotiana tabacum)中, G型凝集素类受体蛋白激酶基因Nt-Sd-RLK受脂多糖诱导表达, 在免疫信号的识别和防御过程中发挥重要作用(Sanabria et al., 2012).Ranf等(2015)通过筛选脂多糖不敏感型缺陷突变体, 获得了1个S-domain类受体蛋白激酶基因LORE.LORE基因突变导致植物易感染丁香假单胞杆菌, 它在介导脂多糖识别引起的免疫反应中发挥功能.在拟南芥中, 缺失L型凝集素类受体蛋白激酶基因LecRK-V.5会增强植物对表面接种Pst DC3000而产生的抗性, 但对通过渗透方法接种所产生的抗性无影响, 提示LecRK-V.5基因可能在植物早期免疫阶段发挥功能; 相反, 过表达LecRK- V.5基因导致植物更易感染Pst DC3000 (Desclos- Theveniau et al., 2012).研究表明, LecRK-V.5基因突变引起的抗病性增强表型与气孔的组成型关闭相关, 而LecRK-V.5基因过表达引起的易感病表型与早期气孔的重新打开有关.LecRK-V.5基因突变和过表达表型结果表明, LecRK-V.5基因可以逆转由于细菌感染诱导的气孔关闭, 从而减轻植株因缺乏CO2而造成的伤害(Desclos-Theveniau et al., 2012).另一个凝集素类受体蛋白激酶基因LecRK-VI.2被报道在抵抗丁香假单胞杆菌和胡萝卜软腐果胶杆菌方面具有重要功能(Singh et al., 2012).LecRK-VI.2基因突变导致PTI (pattern-triggered immunity)反应相关抗病基因表达量降低、胼胝质沉淀形成受阻和气孔关闭出现缺陷等.LecRK-VI.2基因过表达和全基因组分析实验表明, LecRK-VI.2基因正向调控PTI反应.Singh等(2012)研究表明, LecRK-VI.2蛋白位于MAPK信号的上游, 但独立于ROS信号和BIK1的磷酸化调控信号途径.此外, LecRK-VI.2蛋白对于β-氨基丁酸诱导的抗性反应和PTI反应是必需的(Singh et al., 2012).在拟南芥中, LecRK-I.9基因突变导致植物对ATP不敏感, 从而降低ATP引起的防御反应.研究发现, LecRK-I.9蛋白与ATP结合且高度亲和, 它们的结合对于ATP诱导的钙离子响应、MAPK的激活和基因表达是必需的(Choi et al., 2014).Wang等(2015)研究发现, 拟南芥LecRK-IX.1和LecRK-IX.2基因突变可降低植物对疫霉菌的抵抗能力, 且双突变会加剧表型; 相反, 过表达LecRK-IX.1和LecRK-IX.2基因能增强植物的抗病性, 但同时导致自发性的细胞死亡.此外, LecRK-IX.1和LecRK-IX.2蛋白的凝集素结构域和激酶结构域对于抗疫霉菌是必需的; 而凝集素结构域对于诱导细胞死亡为非必需(Wang et al., 2015). ...
... 基因导致细胞程序性死亡、增强免疫反应和对稻瘟病菌的抗性.此外, SDS2蛋白可与细胞质类受体蛋白激酶OsRLCK118/176互作, 通过磷酸化调控NADPH氧化酶OsRbohB, 刺激ROS产生, 从而正向调控免疫反应(Fan et al., 2018).在烟草(Nicotiana tabacum)中, G型凝集素类受体蛋白激酶基因Nt-Sd-RLK受脂多糖诱导表达, 在免疫信号的识别和防御过程中发挥重要作用(Sanabria et al., 2012).Ranf等(2015)通过筛选脂多糖不敏感型缺陷突变体, 获得了1个S-domain类受体蛋白激酶基因LORE.LORE基因突变导致植物易感染丁香假单胞杆菌, 它在介导脂多糖识别引起的免疫反应中发挥功能.在拟南芥中, 缺失L型凝集素类受体蛋白激酶基因LecRK-V.5会增强植物对表面接种Pst DC3000而产生的抗性, 但对通过渗透方法接种所产生的抗性无影响, 提示LecRK-V.5基因可能在植物早期免疫阶段发挥功能; 相反, 过表达LecRK- V.5基因导致植物更易感染Pst DC3000 (Desclos- Theveniau et al., 2012).研究表明, LecRK-V.5基因突变引起的抗病性增强表型与气孔的组成型关闭相关, 而LecRK-V.5基因过表达引起的易感病表型与早期气孔的重新打开有关.LecRK-V.5基因突变和过表达表型结果表明, LecRK-V.5基因可以逆转由于细菌感染诱导的气孔关闭, 从而减轻植株因缺乏CO2而造成的伤害(Desclos-Theveniau et al., 2012).另一个凝集素类受体蛋白激酶基因LecRK-VI.2被报道在抵抗丁香假单胞杆菌和胡萝卜软腐果胶杆菌方面具有重要功能(Singh et al., 2012).LecRK-VI.2基因突变导致PTI (pattern-triggered immunity)反应相关抗病基因表达量降低、胼胝质沉淀形成受阻和气孔关闭出现缺陷等.LecRK-VI.2基因过表达和全基因组分析实验表明, LecRK-VI.2基因正向调控PTI反应.Singh等(2012)研究表明, LecRK-VI.2蛋白位于MAPK信号的上游, 但独立于ROS信号和BIK1的磷酸化调控信号途径.此外, LecRK-VI.2蛋白对于β-氨基丁酸诱导的抗性反应和PTI反应是必需的(Singh et al., 2012).在拟南芥中, LecRK-I.9基因突变导致植物对ATP不敏感, 从而降低ATP引起的防御反应.研究发现, LecRK-I.9蛋白与ATP结合且高度亲和, 它们的结合对于ATP诱导的钙离子响应、MAPK的激活和基因表达是必需的(Choi et al., 2014).Wang等(2015)研究发现, 拟南芥LecRK-IX.1和LecRK-IX.2基因突变可降低植物对疫霉菌的抵抗能力, 且双突变会加剧表型; 相反, 过表达LecRK-IX.1和LecRK-IX.2基因能增强植物的抗病性, 但同时导致自发性的细胞死亡.此外, LecRK-IX.1和LecRK-IX.2蛋白的凝集素结构域和激酶结构域对于抗疫霉菌是必需的; 而凝集素结构域对于诱导细胞死亡为非必需(Wang et al., 2015). ...
Nicotiana attenuata LECTIN RECEPTOR KINASE 1 suppresses the insect-mediated inhibition of induced defense responses during Manduca sexta herbivory 3 2011
... 凝集素类受体蛋白激酶对于植物的抗虫性也极其重要.例如, 在野生烟草(N. attenuata)中, LecRK1基因在烟草天蛾触发所引起的植物防御反应中发挥重要作用(Gilardoni et al., 2011).RNA干涉LecRK1基因表达表明, LecRK1基因对于全面防御抵抗烟草天蛾是必需的.例如, 生长在LecRK1基因干涉植株中的烟草天蛾幼虫比生长在野生型上的幼虫体积大40%-100%; 相对于野生型植株, 昆虫诱导积累的尼古丁、二萜糖甙、胰蛋白酶抑制剂和苏氨酸脱氨酶在干涉植株中的含量也明显减少(Bonaventure, 2011; Gilardoni et al., 2011).在拟南芥中, 昆虫卵衍生的诱导物能激活植物的防御反应, 该反应类似于植物识别PAMPs (pathogen-associated molecular pat- terns)所产生的PTI反应.例如, LecRK-I.8基因在昆虫卵衍生的诱导物识别中发挥重要功能, 该基因突变后会抑制昆虫卵所诱导的抗病基因PR-1的表达(Gouhier- Darimont et al., 2013). ...
... ; Gilardoni et al., 2011).在拟南芥中, 昆虫卵衍生的诱导物能激活植物的防御反应, 该反应类似于植物识别PAMPs (pathogen-associated molecular pat- terns)所产生的PTI反应.例如, LecRK-I.8基因在昆虫卵衍生的诱导物识别中发挥重要功能, 该基因突变后会抑制昆虫卵所诱导的抗病基因PR-1的表达(Gouhier- Darimont et al., 2013). ...
... 凝集素类受体蛋白激酶参与激素信号响应.(1)参与脱落酸(ABA)信号响应.在拟南芥中, 过表达凝集素类受体蛋白激酶基因LecRK-V.5可抑制植物对ABA的响应能力, 从而影响气孔开闭(Desclos-Theveniau et al., 2012).Xin等(2009)发现, L型凝集素类受体蛋白激酶VI亚家族成员基因LecRK-VI.2、LecRK-VI.3和LecRK-VI.4突变后都会微弱增加ABA对种子萌发的抑制作用.相对于LecRK- VI.2单基因突变, LecRK- VI.2和LecRK-VI.3双基因突变时ABA对种子萌发的抑制作用更显著.基因表达谱和网络调控分析表明, LecRK-VI.2和LecRK-VI.3基因还可调控ABA相关基因的表达(Xin et al., 2009).Zhang等(2019)通过对拟南芥凝集素类受体蛋白激酶基因LecRK-VI.4进行深入研究后, 发现该基因在ABA介导的气孔开闭中行使重要功能.(2) 参与茉莉酸(JA)信号响应.野生烟草中LecRK1基因突变会抑制由JA引起的防御途径(Bonaventure, 2011; Gilardoni et al., 2011).(3) 参与水杨酸(SA)信号响应.在拟南芥中过表达LecRK-IX.2基因导致SA积累, 从而引起细胞死亡(Luo et al., 2017). ...
A draft sequence of the rice genome (Oryza sativa L. ssp. japonica) 1 2002
... ZmPK1是第1个被鉴定到的类受体蛋白激酶, 它属于G型凝集素类受体蛋白激酶.该激酶的细胞外结构域与芸薹属自交不亲和性位点编码的糖蛋白类似, 参与调控花粉与柱头之间的识别(Walker and Zhang, 1990).Hervé等(1996)在拟南芥中鉴定到第1个L型凝集素类受体蛋白激酶Ath.LecRK1, 该蛋白激酶具有豆类类凝集素结构, 可能在植物低聚糖和激素信号转导过程中发挥作用(Newman et al., 1994).根据LecRLKs细胞外凝集素结构域的特点, LecRLKs被归类为RLKs的一个亚家族(Shiu and Bleecker, 2001; Goff et al., 2002; Yu et al., 2002).虽然LecRLKs基因发现较早, 但对这些基因家族的系统分类却相对较晚, 而早期对于LecRLKs基因的研究也主要集中在基因表达等较为初步的层面(Nishiguchi et al., 2002; Riou et al., 2002; Navarro-Gochicoa et al., 2003; He et al., 2004; Zuo et al., 2004).近年来, 越来越多的研究表明, LecRLKs蛋白的功能极其广泛, 可参与调控各种生物/非生物胁迫和植物发育(Chen et al., 2006; Kanzaki et al., 2008; Joshi et al., 2010; Bonaventure, 2011; Desclos-Theveniau et al., 2012; Gouhier-Darimont et al., 2013; Li et al., 2014; Ranf et al., 2015; Luo et al., 2017; Ma et al., 2018; Labbé et al., 2019) (表1). ...
Signaling of Arabidopsis thaliana response to Pieris brassicae eggs shares similarities with PAMP-triggered immunity 2 2013
... ZmPK1是第1个被鉴定到的类受体蛋白激酶, 它属于G型凝集素类受体蛋白激酶.该激酶的细胞外结构域与芸薹属自交不亲和性位点编码的糖蛋白类似, 参与调控花粉与柱头之间的识别(Walker and Zhang, 1990).Hervé等(1996)在拟南芥中鉴定到第1个L型凝集素类受体蛋白激酶Ath.LecRK1, 该蛋白激酶具有豆类类凝集素结构, 可能在植物低聚糖和激素信号转导过程中发挥作用(Newman et al., 1994).根据LecRLKs细胞外凝集素结构域的特点, LecRLKs被归类为RLKs的一个亚家族(Shiu and Bleecker, 2001; Goff et al., 2002; Yu et al., 2002).虽然LecRLKs基因发现较早, 但对这些基因家族的系统分类却相对较晚, 而早期对于LecRLKs基因的研究也主要集中在基因表达等较为初步的层面(Nishiguchi et al., 2002; Riou et al., 2002; Navarro-Gochicoa et al., 2003; He et al., 2004; Zuo et al., 2004).近年来, 越来越多的研究表明, LecRLKs蛋白的功能极其广泛, 可参与调控各种生物/非生物胁迫和植物发育(Chen et al., 2006; Kanzaki et al., 2008; Joshi et al., 2010; Bonaventure, 2011; Desclos-Theveniau et al., 2012; Gouhier-Darimont et al., 2013; Li et al., 2014; Ranf et al., 2015; Luo et al., 2017; Ma et al., 2018; Labbé et al., 2019) (表1). ...
... 凝集素类受体蛋白激酶对于植物的抗虫性也极其重要.例如, 在野生烟草(N. attenuata)中, LecRK1基因在烟草天蛾触发所引起的植物防御反应中发挥重要作用(Gilardoni et al., 2011).RNA干涉LecRK1基因表达表明, LecRK1基因对于全面防御抵抗烟草天蛾是必需的.例如, 生长在LecRK1基因干涉植株中的烟草天蛾幼虫比生长在野生型上的幼虫体积大40%-100%; 相对于野生型植株, 昆虫诱导积累的尼古丁、二萜糖甙、胰蛋白酶抑制剂和苏氨酸脱氨酶在干涉植株中的含量也明显减少(Bonaventure, 2011; Gilardoni et al., 2011).在拟南芥中, 昆虫卵衍生的诱导物能激活植物的防御反应, 该反应类似于植物识别PAMPs (pathogen-associated molecular pat- terns)所产生的PTI反应.例如, LecRK-I.8基因在昆虫卵衍生的诱导物识别中发挥重要功能, 该基因突变后会抑制昆虫卵所诱导的抗病基因PR-1的表达(Gouhier- Darimont et al., 2013). ...
GM-CSF mediates alveolar macrophage proliferation and type II cell hypertrophy in SP-D gene-targeted mice 2 2001
... C型凝集素类受体蛋白激酶为钙依赖的凝集素类受体蛋白激酶, 是哺乳动物中的一个大家族蛋白, 主要参与先天性免疫反应以及病原体的识别(Epstein et al., 1996; Hawgood et al., 2001; Cambi et al., 2005).该类型凝集素类受体蛋白激酶的凝集素结构域大量存在于哺乳动物中, 通常通过自我和非自我识别参与免疫反应(Epstein et al., 1996; Hawgood et al., 2001).在植物中, C型凝集素类受体蛋白激酶十分稀少, 属于凝集素类受体蛋白激酶家族中最小的类群.到目前为止, 在水稻和拟南芥中分别仅发现1个C型凝集素类受体蛋白激酶(Bouwmeester and Govers, 2009; Vaid et al., 2012). ...
... ; Hawgood et al., 2001).在植物中, C型凝集素类受体蛋白激酶十分稀少, 属于凝集素类受体蛋白激酶家族中最小的类群.到目前为止, 在水稻和拟南芥中分别仅发现1个C型凝集素类受体蛋白激酶(Bouwmeester and Govers, 2009; Vaid et al., 2012). ...
A salt-responsive receptor-like kinase gene regulated by the ethylene signaling pathway encodes a plasma membrane serine/threonine kinase 2 2004
... ZmPK1是第1个被鉴定到的类受体蛋白激酶, 它属于G型凝集素类受体蛋白激酶.该激酶的细胞外结构域与芸薹属自交不亲和性位点编码的糖蛋白类似, 参与调控花粉与柱头之间的识别(Walker and Zhang, 1990).Hervé等(1996)在拟南芥中鉴定到第1个L型凝集素类受体蛋白激酶Ath.LecRK1, 该蛋白激酶具有豆类类凝集素结构, 可能在植物低聚糖和激素信号转导过程中发挥作用(Newman et al., 1994).根据LecRLKs细胞外凝集素结构域的特点, LecRLKs被归类为RLKs的一个亚家族(Shiu and Bleecker, 2001; Goff et al., 2002; Yu et al., 2002).虽然LecRLKs基因发现较早, 但对这些基因家族的系统分类却相对较晚, 而早期对于LecRLKs基因的研究也主要集中在基因表达等较为初步的层面(Nishiguchi et al., 2002; Riou et al., 2002; Navarro-Gochicoa et al., 2003; He et al., 2004; Zuo et al., 2004).近年来, 越来越多的研究表明, LecRLKs蛋白的功能极其广泛, 可参与调控各种生物/非生物胁迫和植物发育(Chen et al., 2006; Kanzaki et al., 2008; Joshi et al., 2010; Bonaventure, 2011; Desclos-Theveniau et al., 2012; Gouhier-Darimont et al., 2013; Li et al., 2014; Ranf et al., 2015; Luo et al., 2017; Ma et al., 2018; Labbé et al., 2019) (表1). ...
... 凝集素类受体蛋白激酶参与盐胁迫响应.拟南芥LecRK-V.2基因缺失会降低植物早期发育阶段对盐的敏感性(Deng et al., 2009).He等(2004)发现, 拟南芥LecRK2基因在盐胁迫时其表达量有不同程度的上调, 并进一步证明它受到乙烯(ET)信号通路的调控.豌豆(Pisum sativum) PsLecRLK基因在盐胁迫下被诱导表达, 而PsLecRLK基因过表达可缓解离子平衡和渗透压, 从而增强植物的耐盐性(Vaid et al., 2015).Sun等(2013)发现野大豆(Glycine soja) GsSRK基因的表达受盐胁迫诱导, 而过表达该基因可增强植物的耐盐性, 并使植物在耐盐环境中表现出更高的产量, 同时还伴随叶绿素含量增加、离子泄漏降低、株高增加和果实增多等表型.在水稻中, 盐胁迫能够增加SIT1蛋白激酶活性, 从而磷酸化其下游效应蛋白MAPK3和MAPK6; 此外, SIT1蛋白还可介导乙烯的生成和盐胁迫诱导的乙烯信号途径(Li et al., 2014).而在拟南芥中, SIT1蛋白激酶在盐胁迫下还可促进活性氧的积累, 最终导致植物生长受抑制甚至死亡, 该过程依赖于MAPK3/6和乙烯信号途径(Li et al., 2014).Ma等(2018)发现在本氏烟草(N. benthamiana)中过表达豆梨(Pyrus calleryana) L型凝集素类受体蛋白激酶基因PbLRK138导致细胞死亡, 从而增强植物的耐盐性. ...
Characterization of an Arabidopsis thaliana gene that defines a new class of putative plant receptor kinases with an extracellular lectin-like domain 1 1996
... ZmPK1是第1个被鉴定到的类受体蛋白激酶, 它属于G型凝集素类受体蛋白激酶.该激酶的细胞外结构域与芸薹属自交不亲和性位点编码的糖蛋白类似, 参与调控花粉与柱头之间的识别(Walker and Zhang, 1990).Hervé等(1996)在拟南芥中鉴定到第1个L型凝集素类受体蛋白激酶Ath.LecRK1, 该蛋白激酶具有豆类类凝集素结构, 可能在植物低聚糖和激素信号转导过程中发挥作用(Newman et al., 1994).根据LecRLKs细胞外凝集素结构域的特点, LecRLKs被归类为RLKs的一个亚家族(Shiu and Bleecker, 2001; Goff et al., 2002; Yu et al., 2002).虽然LecRLKs基因发现较早, 但对这些基因家族的系统分类却相对较晚, 而早期对于LecRLKs基因的研究也主要集中在基因表达等较为初步的层面(Nishiguchi et al., 2002; Riou et al., 2002; Navarro-Gochicoa et al., 2003; He et al., 2004; Zuo et al., 2004).近年来, 越来越多的研究表明, LecRLKs蛋白的功能极其广泛, 可参与调控各种生物/非生物胁迫和植物发育(Chen et al., 2006; Kanzaki et al., 2008; Joshi et al., 2010; Bonaventure, 2011; Desclos-Theveniau et al., 2012; Gouhier-Darimont et al., 2013; Li et al., 2014; Ranf et al., 2015; Luo et al., 2017; Ma et al., 2018; Labbé et al., 2019) (表1). ...
Characterization of the Arabidopsis lecRK-a genes: members of a superfamily encoding putative receptors with an extracellular domain homologous to legume lectins 3 1999
... 凝集素类受体蛋白激酶主要由胞外区(包含凝集素结构域)、跨膜区以及胞内区(包含近膜区和激酶结构域)组成(图1).凝集素结构域可与糖类分子特异可逆结合, 该结构域在不同受体中差异较大, 可分为不同类型, 主要参与信号识别和结合(Peumans and van Damme, 1995; Vaid et al., 2012).跨膜结构域一般由18-25个氨基酸组成, 不具有明显的保守性, 但该部位对于凝集素类受体蛋白激酶的功能极其重要(Morillo and Tax, 2006).例如, 水稻Pi-d2蛋白跨膜结构域中的1个氨基酸发生改变, 可导致该蛋白完全丧失功能(Chen et al., 2006).近膜区主要是将跨膜结构域和激酶结构域分开, 该区由50个左右的氨基酸组成, 保守性较高, 其C端一般包含PxPFxYxELxxATxG (x表示任一氨基酸)的未知功能亲水区(Walker, 1994; Barre et al., 2002).激酶结构域是高度保守的结构域, 具有磷酸化结合位点, 位于细胞质区域, 一般由250- 300个氨基酸组成, 主要负责传递外界信号(Li and Gray, 1997).激酶结构域的N端包含GxGxxG结构域, 能够影响核苷酸的结合; 而激酶结构域的C端由43- 66个氨基酸组成, 为激酶的催化活性所必需(Barre et al., 2002).与酪氨酸类受体蛋白激酶含有DLAARN和FPIKWMAPE基序不同, 凝集素类受体蛋白激酶的激酶结构域中含有DIKPAN和GT(FIL)GYIAPE基序, 属于丝/苏氨酸激酶(Hervé et al., 1999). ...
... L型凝集素类受体蛋白激酶的细胞外结构含legume-lectin结构域, 位于N端, 大约由250个氨基酸组成, 该结构域呈现出典型的β-sandwich折叠, 具有糖类结合位点, 并保留了疏水腔结构, 可能涉及与疏水配体之间的互作(Edelman and Wang, 1978; Hervé et al., 1999; Barre et al., 2002).此外, 在该结构域中也包含Ca2+和Mn2+, 可稳定糖类结合位点(Bellande et al., 2017).大部分L型凝集素类受体蛋白激酶的凝集素结构域的糖类结合位点和与Ca2+、Mn2+结合的氨基酸都不保守(Hervé et al., 1999; Barre et al., 2002).L型凝集素类受体蛋白激酶通常以LecRLK形式命名. ...
... 结合的氨基酸都不保守(Hervé et al., 1999; Barre et al., 2002).L型凝集素类受体蛋白激酶通常以LecRLK形式命名. ...
Cloning and expression of a PR5-like protein from Arabidopsis: inhibition of fungal growth by bacterially expressed protein 1 1997
... 根据RLKs细胞外结构域的不同特点, 同时参照动物RLKs, 人们将植物RLKs分为6种类型: 富含亮氨酸重复序列型(leucine-rich repeats, LRRs)、S-结构域型(S-domain)、类凝集素型(lectin-like)、类表皮生长因子型(epidermal growth factor-like, EGF)、类肿瘤坏死因子型(tumor-necrosis factor receptor-like, TNFR)和类PR5型(pathogenesis related protein-5 like receptor kinases, PR5K) (Walker, 1994; Becraft et al., 1996; Hu and Reddy, 1997; Wang et al., 1998; Kobe and Kajava, 2001; Shiu and Bleecker, 2001).凝集素类受体蛋白激酶(lectin receptor-like kinases, LecRLKs)是RLKs家族中的一个亚族, 是指含有1个或多个可与单糖或寡聚糖特异可逆结合的非催化结构域的植物蛋白(Peumans and van Damme, 1995).已知在拟南芥和水稻中分别存在77和173个LecRLKs (Bouwmeester and Govers, 2009; Vaid et al., 2012).到目前为止, 在酵母(Saccharomyces cerevisiae)和人体基因组中还未找到LecRLKs的同源基因(Navarro-Gochicoa et al., 2003).已有研究表明, LecRLKs蛋白可参与多种生物/非生物胁迫途径以及植物发育调控(Vaid et al., 2012).本文简要阐述植物凝集素类受体蛋白激酶的研究历史、结构特点与分类及其生物学功能, 以期增进对凝集素类受体蛋白激酶的认识. ...
Protein Kinase Classication 1 1991
... 受体蛋白激酶(receptor protein kinases, RPKs)是一类能够与配基结合并具有酶活性的受体(Hunter, 1991).RPKs蛋白在动物中研究较早, 主要分为酪氨酸激酶(receptor tyrosine kinases, RTKs)和丝/苏氨酸激酶(serine/threonine kinase receptors, STRKs) 2种.与动物不同, 植物中受体蛋白激酶主要属于丝/苏氨酸激酶.目前, 已在植物中陆续鉴定到多个与动物受体蛋白激酶结构相似的蛋白, 这类蛋白绝大部分功能未知, 被称为类受体蛋白激酶(receptor-like kinases, RLKs) (Walker, 1994; Shiu and Bleecker, 2001).RLKs是一种普遍存在于植物体内的蛋白, 能够感知和传递各种信号, 在拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)中分别有超过600和1 100个该类蛋白(Shiu and Bleecker, 2001).大部分RLKs属于细胞质膜蛋白, 主要功能是通过它们的细胞外配体识别结构域和细胞内激酶结构域传递各种信号, 最终产生一系列生物效应(Vaid et al., 2013). ...
Pea lectin receptor-like kinase promotes high salinity stress tolerance in bacteria and expresses in response to stress in planta 1 2010
... ZmPK1是第1个被鉴定到的类受体蛋白激酶, 它属于G型凝集素类受体蛋白激酶.该激酶的细胞外结构域与芸薹属自交不亲和性位点编码的糖蛋白类似, 参与调控花粉与柱头之间的识别(Walker and Zhang, 1990).Hervé等(1996)在拟南芥中鉴定到第1个L型凝集素类受体蛋白激酶Ath.LecRK1, 该蛋白激酶具有豆类类凝集素结构, 可能在植物低聚糖和激素信号转导过程中发挥作用(Newman et al., 1994).根据LecRLKs细胞外凝集素结构域的特点, LecRLKs被归类为RLKs的一个亚家族(Shiu and Bleecker, 2001; Goff et al., 2002; Yu et al., 2002).虽然LecRLKs基因发现较早, 但对这些基因家族的系统分类却相对较晚, 而早期对于LecRLKs基因的研究也主要集中在基因表达等较为初步的层面(Nishiguchi et al., 2002; Riou et al., 2002; Navarro-Gochicoa et al., 2003; He et al., 2004; Zuo et al., 2004).近年来, 越来越多的研究表明, LecRLKs蛋白的功能极其广泛, 可参与调控各种生物/非生物胁迫和植物发育(Chen et al., 2006; Kanzaki et al., 2008; Joshi et al., 2010; Bonaventure, 2011; Desclos-Theveniau et al., 2012; Gouhier-Darimont et al., 2013; Li et al., 2014; Ranf et al., 2015; Luo et al., 2017; Ma et al., 2018; Labbé et al., 2019) (表1). ...
NbLRK1, a lectin-like receptor kinase protein of Nicotiana benthamiana, interacts with Phytophthora infestans INF1 elicitin and mediates INF1-induced cell death 1 2008
... ZmPK1是第1个被鉴定到的类受体蛋白激酶, 它属于G型凝集素类受体蛋白激酶.该激酶的细胞外结构域与芸薹属自交不亲和性位点编码的糖蛋白类似, 参与调控花粉与柱头之间的识别(Walker and Zhang, 1990).Hervé等(1996)在拟南芥中鉴定到第1个L型凝集素类受体蛋白激酶Ath.LecRK1, 该蛋白激酶具有豆类类凝集素结构, 可能在植物低聚糖和激素信号转导过程中发挥作用(Newman et al., 1994).根据LecRLKs细胞外凝集素结构域的特点, LecRLKs被归类为RLKs的一个亚家族(Shiu and Bleecker, 2001; Goff et al., 2002; Yu et al., 2002).虽然LecRLKs基因发现较早, 但对这些基因家族的系统分类却相对较晚, 而早期对于LecRLKs基因的研究也主要集中在基因表达等较为初步的层面(Nishiguchi et al., 2002; Riou et al., 2002; Navarro-Gochicoa et al., 2003; He et al., 2004; Zuo et al., 2004).近年来, 越来越多的研究表明, LecRLKs蛋白的功能极其广泛, 可参与调控各种生物/非生物胁迫和植物发育(Chen et al., 2006; Kanzaki et al., 2008; Joshi et al., 2010; Bonaventure, 2011; Desclos-Theveniau et al., 2012; Gouhier-Darimont et al., 2013; Li et al., 2014; Ranf et al., 2015; Luo et al., 2017; Ma et al., 2018; Labbé et al., 2019) (表1). ...
The leucine-rich repeat as a protein recognition motif 1 2001
... 根据RLKs细胞外结构域的不同特点, 同时参照动物RLKs, 人们将植物RLKs分为6种类型: 富含亮氨酸重复序列型(leucine-rich repeats, LRRs)、S-结构域型(S-domain)、类凝集素型(lectin-like)、类表皮生长因子型(epidermal growth factor-like, EGF)、类肿瘤坏死因子型(tumor-necrosis factor receptor-like, TNFR)和类PR5型(pathogenesis related protein-5 like receptor kinases, PR5K) (Walker, 1994; Becraft et al., 1996; Hu and Reddy, 1997; Wang et al., 1998; Kobe and Kajava, 2001; Shiu and Bleecker, 2001).凝集素类受体蛋白激酶(lectin receptor-like kinases, LecRLKs)是RLKs家族中的一个亚族, 是指含有1个或多个可与单糖或寡聚糖特异可逆结合的非催化结构域的植物蛋白(Peumans and van Damme, 1995).已知在拟南芥和水稻中分别存在77和173个LecRLKs (Bouwmeester and Govers, 2009; Vaid et al., 2012).到目前为止, 在酵母(Saccharomyces cerevisiae)和人体基因组中还未找到LecRLKs的同源基因(Navarro-Gochicoa et al., 2003).已有研究表明, LecRLKs蛋白可参与多种生物/非生物胁迫途径以及植物发育调控(Vaid et al., 2012).本文简要阐述植物凝集素类受体蛋白激酶的研究历史、结构特点与分类及其生物学功能, 以期增进对凝集素类受体蛋白激酶的认识. ...
Self-incompatibility in the genus Arabidopsis: characterization of the S locus in the outcrossing A. lyrata and its autogamous relative A. thaliana 1 2001
... G型凝集素类受体蛋白激酶曾被称为B-type LecRLK, 其细胞外凝集素结构域具有β-barre结构, 属于α-D甘露醇特异的植物凝集素类受体蛋白激酶(Shiu and Bleecker, 2001).除凝集素结构域外, G型凝集素类受体蛋白激酶的细胞外结构还包含EGF和/或PAN (plasminogen-apple-nematode)结构域(Vaid et al., 2012) (图1).其中, EGF结构域也被称作S-locus结构域, 在十字花科家族的自交不亲和中起作用(Tanksley and Loaiza-Figueroa, 1985; Kusaba et al., 2001).S-domain RLK因被发现参与有花植物自交不亲和(SI)而又被称为S-locus receptor kinase SRK, 属于G型凝集素类受体蛋白激酶(Sherman- Broyles et al., 2007).而PAN为富含半胱氨酸的结构域, 参与形成二硫键, 且PAN结构域的复杂功能是通过蛋白与蛋白以及蛋白与碳水化合物之间的互作来介导(Tordai et al., 1999; Shiu and Bleecker, 2001; Loris, 2002; Naithani et al., 2007). ...
Mediation of plant-mycorrhizal interaction by a lectin receptor-like kinase 2 2019
... ZmPK1是第1个被鉴定到的类受体蛋白激酶, 它属于G型凝集素类受体蛋白激酶.该激酶的细胞外结构域与芸薹属自交不亲和性位点编码的糖蛋白类似, 参与调控花粉与柱头之间的识别(Walker and Zhang, 1990).Hervé等(1996)在拟南芥中鉴定到第1个L型凝集素类受体蛋白激酶Ath.LecRK1, 该蛋白激酶具有豆类类凝集素结构, 可能在植物低聚糖和激素信号转导过程中发挥作用(Newman et al., 1994).根据LecRLKs细胞外凝集素结构域的特点, LecRLKs被归类为RLKs的一个亚家族(Shiu and Bleecker, 2001; Goff et al., 2002; Yu et al., 2002).虽然LecRLKs基因发现较早, 但对这些基因家族的系统分类却相对较晚, 而早期对于LecRLKs基因的研究也主要集中在基因表达等较为初步的层面(Nishiguchi et al., 2002; Riou et al., 2002; Navarro-Gochicoa et al., 2003; He et al., 2004; Zuo et al., 2004).近年来, 越来越多的研究表明, LecRLKs蛋白的功能极其广泛, 可参与调控各种生物/非生物胁迫和植物发育(Chen et al., 2006; Kanzaki et al., 2008; Joshi et al., 2010; Bonaventure, 2011; Desclos-Theveniau et al., 2012; Gouhier-Darimont et al., 2013; Li et al., 2014; Ranf et al., 2015; Luo et al., 2017; Ma et al., 2018; Labbé et al., 2019) (表1). ...
The receptor-like kinase SIT1 mediates salt sensitivity by activating MAPK3/6 and regulating ethylene homeostasis in rice 3 2014
... ZmPK1是第1个被鉴定到的类受体蛋白激酶, 它属于G型凝集素类受体蛋白激酶.该激酶的细胞外结构域与芸薹属自交不亲和性位点编码的糖蛋白类似, 参与调控花粉与柱头之间的识别(Walker and Zhang, 1990).Hervé等(1996)在拟南芥中鉴定到第1个L型凝集素类受体蛋白激酶Ath.LecRK1, 该蛋白激酶具有豆类类凝集素结构, 可能在植物低聚糖和激素信号转导过程中发挥作用(Newman et al., 1994).根据LecRLKs细胞外凝集素结构域的特点, LecRLKs被归类为RLKs的一个亚家族(Shiu and Bleecker, 2001; Goff et al., 2002; Yu et al., 2002).虽然LecRLKs基因发现较早, 但对这些基因家族的系统分类却相对较晚, 而早期对于LecRLKs基因的研究也主要集中在基因表达等较为初步的层面(Nishiguchi et al., 2002; Riou et al., 2002; Navarro-Gochicoa et al., 2003; He et al., 2004; Zuo et al., 2004).近年来, 越来越多的研究表明, LecRLKs蛋白的功能极其广泛, 可参与调控各种生物/非生物胁迫和植物发育(Chen et al., 2006; Kanzaki et al., 2008; Joshi et al., 2010; Bonaventure, 2011; Desclos-Theveniau et al., 2012; Gouhier-Darimont et al., 2013; Li et al., 2014; Ranf et al., 2015; Luo et al., 2017; Ma et al., 2018; Labbé et al., 2019) (表1). ...
... 凝集素类受体蛋白激酶参与盐胁迫响应.拟南芥LecRK-V.2基因缺失会降低植物早期发育阶段对盐的敏感性(Deng et al., 2009).He等(2004)发现, 拟南芥LecRK2基因在盐胁迫时其表达量有不同程度的上调, 并进一步证明它受到乙烯(ET)信号通路的调控.豌豆(Pisum sativum) PsLecRLK基因在盐胁迫下被诱导表达, 而PsLecRLK基因过表达可缓解离子平衡和渗透压, 从而增强植物的耐盐性(Vaid et al., 2015).Sun等(2013)发现野大豆(Glycine soja) GsSRK基因的表达受盐胁迫诱导, 而过表达该基因可增强植物的耐盐性, 并使植物在耐盐环境中表现出更高的产量, 同时还伴随叶绿素含量增加、离子泄漏降低、株高增加和果实增多等表型.在水稻中, 盐胁迫能够增加SIT1蛋白激酶活性, 从而磷酸化其下游效应蛋白MAPK3和MAPK6; 此外, SIT1蛋白还可介导乙烯的生成和盐胁迫诱导的乙烯信号途径(Li et al., 2014).而在拟南芥中, SIT1蛋白激酶在盐胁迫下还可促进活性氧的积累, 最终导致植物生长受抑制甚至死亡, 该过程依赖于MAPK3/6和乙烯信号途径(Li et al., 2014).Ma等(2018)发现在本氏烟草(N. benthamiana)中过表达豆梨(Pyrus calleryana) L型凝集素类受体蛋白激酶基因PbLRK138导致细胞死亡, 从而增强植物的耐盐性. ...
... ZmPK1是第1个被鉴定到的类受体蛋白激酶, 它属于G型凝集素类受体蛋白激酶.该激酶的细胞外结构域与芸薹属自交不亲和性位点编码的糖蛋白类似, 参与调控花粉与柱头之间的识别(Walker and Zhang, 1990).Hervé等(1996)在拟南芥中鉴定到第1个L型凝集素类受体蛋白激酶Ath.LecRK1, 该蛋白激酶具有豆类类凝集素结构, 可能在植物低聚糖和激素信号转导过程中发挥作用(Newman et al., 1994).根据LecRLKs细胞外凝集素结构域的特点, LecRLKs被归类为RLKs的一个亚家族(Shiu and Bleecker, 2001; Goff et al., 2002; Yu et al., 2002).虽然LecRLKs基因发现较早, 但对这些基因家族的系统分类却相对较晚, 而早期对于LecRLKs基因的研究也主要集中在基因表达等较为初步的层面(Nishiguchi et al., 2002; Riou et al., 2002; Navarro-Gochicoa et al., 2003; He et al., 2004; Zuo et al., 2004).近年来, 越来越多的研究表明, LecRLKs蛋白的功能极其广泛, 可参与调控各种生物/非生物胁迫和植物发育(Chen et al., 2006; Kanzaki et al., 2008; Joshi et al., 2010; Bonaventure, 2011; Desclos-Theveniau et al., 2012; Gouhier-Darimont et al., 2013; Li et al., 2014; Ranf et al., 2015; Luo et al., 2017; Ma et al., 2018; Labbé et al., 2019) (表1). ...
... 凝集素类受体蛋白激酶参与激素信号响应.(1)参与脱落酸(ABA)信号响应.在拟南芥中, 过表达凝集素类受体蛋白激酶基因LecRK-V.5可抑制植物对ABA的响应能力, 从而影响气孔开闭(Desclos-Theveniau et al., 2012).Xin等(2009)发现, L型凝集素类受体蛋白激酶VI亚家族成员基因LecRK-VI.2、LecRK-VI.3和LecRK-VI.4突变后都会微弱增加ABA对种子萌发的抑制作用.相对于LecRK- VI.2单基因突变, LecRK- VI.2和LecRK-VI.3双基因突变时ABA对种子萌发的抑制作用更显著.基因表达谱和网络调控分析表明, LecRK-VI.2和LecRK-VI.3基因还可调控ABA相关基因的表达(Xin et al., 2009).Zhang等(2019)通过对拟南芥凝集素类受体蛋白激酶基因LecRK-VI.4进行深入研究后, 发现该基因在ABA介导的气孔开闭中行使重要功能.(2) 参与茉莉酸(JA)信号响应.野生烟草中LecRK1基因突变会抑制由JA引起的防御途径(Bonaventure, 2011; Gilardoni et al., 2011).(3) 参与水杨酸(SA)信号响应.在拟南芥中过表达LecRK-IX.2基因导致SA积累, 从而引起细胞死亡(Luo et al., 2017). ...
Genome-wide identification of lectin receptor kinases in pear: functional characterization of the L-type LecRLK gene PbLRK138 2 2018
... ZmPK1是第1个被鉴定到的类受体蛋白激酶, 它属于G型凝集素类受体蛋白激酶.该激酶的细胞外结构域与芸薹属自交不亲和性位点编码的糖蛋白类似, 参与调控花粉与柱头之间的识别(Walker and Zhang, 1990).Hervé等(1996)在拟南芥中鉴定到第1个L型凝集素类受体蛋白激酶Ath.LecRK1, 该蛋白激酶具有豆类类凝集素结构, 可能在植物低聚糖和激素信号转导过程中发挥作用(Newman et al., 1994).根据LecRLKs细胞外凝集素结构域的特点, LecRLKs被归类为RLKs的一个亚家族(Shiu and Bleecker, 2001; Goff et al., 2002; Yu et al., 2002).虽然LecRLKs基因发现较早, 但对这些基因家族的系统分类却相对较晚, 而早期对于LecRLKs基因的研究也主要集中在基因表达等较为初步的层面(Nishiguchi et al., 2002; Riou et al., 2002; Navarro-Gochicoa et al., 2003; He et al., 2004; Zuo et al., 2004).近年来, 越来越多的研究表明, LecRLKs蛋白的功能极其广泛, 可参与调控各种生物/非生物胁迫和植物发育(Chen et al., 2006; Kanzaki et al., 2008; Joshi et al., 2010; Bonaventure, 2011; Desclos-Theveniau et al., 2012; Gouhier-Darimont et al., 2013; Li et al., 2014; Ranf et al., 2015; Luo et al., 2017; Ma et al., 2018; Labbé et al., 2019) (表1). ...
... 凝集素类受体蛋白激酶参与盐胁迫响应.拟南芥LecRK-V.2基因缺失会降低植物早期发育阶段对盐的敏感性(Deng et al., 2009).He等(2004)发现, 拟南芥LecRK2基因在盐胁迫时其表达量有不同程度的上调, 并进一步证明它受到乙烯(ET)信号通路的调控.豌豆(Pisum sativum) PsLecRLK基因在盐胁迫下被诱导表达, 而PsLecRLK基因过表达可缓解离子平衡和渗透压, 从而增强植物的耐盐性(Vaid et al., 2015).Sun等(2013)发现野大豆(Glycine soja) GsSRK基因的表达受盐胁迫诱导, 而过表达该基因可增强植物的耐盐性, 并使植物在耐盐环境中表现出更高的产量, 同时还伴随叶绿素含量增加、离子泄漏降低、株高增加和果实增多等表型.在水稻中, 盐胁迫能够增加SIT1蛋白激酶活性, 从而磷酸化其下游效应蛋白MAPK3和MAPK6; 此外, SIT1蛋白还可介导乙烯的生成和盐胁迫诱导的乙烯信号途径(Li et al., 2014).而在拟南芥中, SIT1蛋白激酶在盐胁迫下还可促进活性氧的积累, 最终导致植物生长受抑制甚至死亡, 该过程依赖于MAPK3/6和乙烯信号途径(Li et al., 2014).Ma等(2018)发现在本氏烟草(N. benthamiana)中过表达豆梨(Pyrus calleryana) L型凝集素类受体蛋白激酶基因PbLRK138导致细胞死亡, 从而增强植物的耐盐性. ...
Functional analysis of receptor-like kinases in monocots and dicots 1 2006
... 凝集素类受体蛋白激酶主要由胞外区(包含凝集素结构域)、跨膜区以及胞内区(包含近膜区和激酶结构域)组成(图1).凝集素结构域可与糖类分子特异可逆结合, 该结构域在不同受体中差异较大, 可分为不同类型, 主要参与信号识别和结合(Peumans and van Damme, 1995; Vaid et al., 2012).跨膜结构域一般由18-25个氨基酸组成, 不具有明显的保守性, 但该部位对于凝集素类受体蛋白激酶的功能极其重要(Morillo and Tax, 2006).例如, 水稻Pi-d2蛋白跨膜结构域中的1个氨基酸发生改变, 可导致该蛋白完全丧失功能(Chen et al., 2006).近膜区主要是将跨膜结构域和激酶结构域分开, 该区由50个左右的氨基酸组成, 保守性较高, 其C端一般包含PxPFxYxELxxATxG (x表示任一氨基酸)的未知功能亲水区(Walker, 1994; Barre et al., 2002).激酶结构域是高度保守的结构域, 具有磷酸化结合位点, 位于细胞质区域, 一般由250- 300个氨基酸组成, 主要负责传递外界信号(Li and Gray, 1997).激酶结构域的N端包含GxGxxG结构域, 能够影响核苷酸的结合; 而激酶结构域的C端由43- 66个氨基酸组成, 为激酶的催化活性所必需(Barre et al., 2002).与酪氨酸类受体蛋白激酶含有DLAARN和FPIKWMAPE基序不同, 凝集素类受体蛋白激酶的激酶结构域中含有DIKPAN和GT(FIL)GYIAPE基序, 属于丝/苏氨酸激酶(Hervé et al., 1999). ...
Structural modules for receptor dimerization in the S-locus receptor kinase extracellular domain 1 2007
... G型凝集素类受体蛋白激酶曾被称为B-type LecRLK, 其细胞外凝集素结构域具有β-barre结构, 属于α-D甘露醇特异的植物凝集素类受体蛋白激酶(Shiu and Bleecker, 2001).除凝集素结构域外, G型凝集素类受体蛋白激酶的细胞外结构还包含EGF和/或PAN (plasminogen-apple-nematode)结构域(Vaid et al., 2012) (图1).其中, EGF结构域也被称作S-locus结构域, 在十字花科家族的自交不亲和中起作用(Tanksley and Loaiza-Figueroa, 1985; Kusaba et al., 2001).S-domain RLK因被发现参与有花植物自交不亲和(SI)而又被称为S-locus receptor kinase SRK, 属于G型凝集素类受体蛋白激酶(Sherman- Broyles et al., 2007).而PAN为富含半胱氨酸的结构域, 参与形成二硫键, 且PAN结构域的复杂功能是通过蛋白与蛋白以及蛋白与碳水化合物之间的互作来介导(Tordai et al., 1999; Shiu and Bleecker, 2001; Loris, 2002; Naithani et al., 2007). ...
Characterization of four lectin-like receptor kinases expressed in roots of Medicago truncatula. Structure, location, regulation of expression, and potential role in the symbiosis with Sinorhizobium meliloti 3 2003
... 根据RLKs细胞外结构域的不同特点, 同时参照动物RLKs, 人们将植物RLKs分为6种类型: 富含亮氨酸重复序列型(leucine-rich repeats, LRRs)、S-结构域型(S-domain)、类凝集素型(lectin-like)、类表皮生长因子型(epidermal growth factor-like, EGF)、类肿瘤坏死因子型(tumor-necrosis factor receptor-like, TNFR)和类PR5型(pathogenesis related protein-5 like receptor kinases, PR5K) (Walker, 1994; Becraft et al., 1996; Hu and Reddy, 1997; Wang et al., 1998; Kobe and Kajava, 2001; Shiu and Bleecker, 2001).凝集素类受体蛋白激酶(lectin receptor-like kinases, LecRLKs)是RLKs家族中的一个亚族, 是指含有1个或多个可与单糖或寡聚糖特异可逆结合的非催化结构域的植物蛋白(Peumans and van Damme, 1995).已知在拟南芥和水稻中分别存在77和173个LecRLKs (Bouwmeester and Govers, 2009; Vaid et al., 2012).到目前为止, 在酵母(Saccharomyces cerevisiae)和人体基因组中还未找到LecRLKs的同源基因(Navarro-Gochicoa et al., 2003).已有研究表明, LecRLKs蛋白可参与多种生物/非生物胁迫途径以及植物发育调控(Vaid et al., 2012).本文简要阐述植物凝集素类受体蛋白激酶的研究历史、结构特点与分类及其生物学功能, 以期增进对凝集素类受体蛋白激酶的认识. ...
... ZmPK1是第1个被鉴定到的类受体蛋白激酶, 它属于G型凝集素类受体蛋白激酶.该激酶的细胞外结构域与芸薹属自交不亲和性位点编码的糖蛋白类似, 参与调控花粉与柱头之间的识别(Walker and Zhang, 1990).Hervé等(1996)在拟南芥中鉴定到第1个L型凝集素类受体蛋白激酶Ath.LecRK1, 该蛋白激酶具有豆类类凝集素结构, 可能在植物低聚糖和激素信号转导过程中发挥作用(Newman et al., 1994).根据LecRLKs细胞外凝集素结构域的特点, LecRLKs被归类为RLKs的一个亚家族(Shiu and Bleecker, 2001; Goff et al., 2002; Yu et al., 2002).虽然LecRLKs基因发现较早, 但对这些基因家族的系统分类却相对较晚, 而早期对于LecRLKs基因的研究也主要集中在基因表达等较为初步的层面(Nishiguchi et al., 2002; Riou et al., 2002; Navarro-Gochicoa et al., 2003; He et al., 2004; Zuo et al., 2004).近年来, 越来越多的研究表明, LecRLKs蛋白的功能极其广泛, 可参与调控各种生物/非生物胁迫和植物发育(Chen et al., 2006; Kanzaki et al., 2008; Joshi et al., 2010; Bonaventure, 2011; Desclos-Theveniau et al., 2012; Gouhier-Darimont et al., 2013; Li et al., 2014; Ranf et al., 2015; Luo et al., 2017; Ma et al., 2018; Labbé et al., 2019) (表1). ...
... 凝集素蛋白在植物整个生命过程中的作用非常重要, 并且在细胞与细胞的交流、植物的发育和防御反应中扮演重要角色(Bellande et al., 2017).已知凝集素蛋白可参与多种生理学过程, 包括蛋白分选、识别、胚胎发生和发育; 凝集素蛋白可与外源糖基结合, 从而保护植物免受食草动物或植食性无脊椎动物的侵害(Brewin and Kardailsky, 1997; Brill et al., 2001; Rudiger and Gabius, 2001).由于凝集素类受体蛋白激酶的凝集素识别结构域与凝集素蛋白相似, 推测其可能具有识别和转导糖信号的功能(Navarro-Gochicoa et al., 2003).近期的研究表明, 凝集素类受体蛋白激酶在响应胁迫信号、抗病虫害和调控植物发育方面都具有非常重要的功能(表1; 图2). ...
Genes galore: a summary of methods for accessing results from large-scale partial sequencing of anonymous Arabidopsis cDNA clones 1 1994
... ZmPK1是第1个被鉴定到的类受体蛋白激酶, 它属于G型凝集素类受体蛋白激酶.该激酶的细胞外结构域与芸薹属自交不亲和性位点编码的糖蛋白类似, 参与调控花粉与柱头之间的识别(Walker and Zhang, 1990).Hervé等(1996)在拟南芥中鉴定到第1个L型凝集素类受体蛋白激酶Ath.LecRK1, 该蛋白激酶具有豆类类凝集素结构, 可能在植物低聚糖和激素信号转导过程中发挥作用(Newman et al., 1994).根据LecRLKs细胞外凝集素结构域的特点, LecRLKs被归类为RLKs的一个亚家族(Shiu and Bleecker, 2001; Goff et al., 2002; Yu et al., 2002).虽然LecRLKs基因发现较早, 但对这些基因家族的系统分类却相对较晚, 而早期对于LecRLKs基因的研究也主要集中在基因表达等较为初步的层面(Nishiguchi et al., 2002; Riou et al., 2002; Navarro-Gochicoa et al., 2003; He et al., 2004; Zuo et al., 2004).近年来, 越来越多的研究表明, LecRLKs蛋白的功能极其广泛, 可参与调控各种生物/非生物胁迫和植物发育(Chen et al., 2006; Kanzaki et al., 2008; Joshi et al., 2010; Bonaventure, 2011; Desclos-Theveniau et al., 2012; Gouhier-Darimont et al., 2013; Li et al., 2014; Ranf et al., 2015; Luo et al., 2017; Ma et al., 2018; Labbé et al., 2019) (表1). ...
A receptor-like protein kinase with a lectin-like domain from Lombardy poplar: gene expression in response to wounding and characterization of phosphorylation activity 2 2002
... ZmPK1是第1个被鉴定到的类受体蛋白激酶, 它属于G型凝集素类受体蛋白激酶.该激酶的细胞外结构域与芸薹属自交不亲和性位点编码的糖蛋白类似, 参与调控花粉与柱头之间的识别(Walker and Zhang, 1990).Hervé等(1996)在拟南芥中鉴定到第1个L型凝集素类受体蛋白激酶Ath.LecRK1, 该蛋白激酶具有豆类类凝集素结构, 可能在植物低聚糖和激素信号转导过程中发挥作用(Newman et al., 1994).根据LecRLKs细胞外凝集素结构域的特点, LecRLKs被归类为RLKs的一个亚家族(Shiu and Bleecker, 2001; Goff et al., 2002; Yu et al., 2002).虽然LecRLKs基因发现较早, 但对这些基因家族的系统分类却相对较晚, 而早期对于LecRLKs基因的研究也主要集中在基因表达等较为初步的层面(Nishiguchi et al., 2002; Riou et al., 2002; Navarro-Gochicoa et al., 2003; He et al., 2004; Zuo et al., 2004).近年来, 越来越多的研究表明, LecRLKs蛋白的功能极其广泛, 可参与调控各种生物/非生物胁迫和植物发育(Chen et al., 2006; Kanzaki et al., 2008; Joshi et al., 2010; Bonaventure, 2011; Desclos-Theveniau et al., 2012; Gouhier-Darimont et al., 2013; Li et al., 2014; Ranf et al., 2015; Luo et al., 2017; Ma et al., 2018; Labbé et al., 2019) (表1). ...
... 凝集素类受体蛋白激酶参与机械损伤引起的胁迫反应.在拟南芥中, LecRK-V.5基因的表达被局部激活以响应机械损伤信号(Riou et al., 2002).在黑杨(Populus nigra)受到机械损伤的幼叶中, PnLPK基因的mRNA丰度增加(Nishiguchi et al., 2002). ...
The role of lectins in plant defence 2 1995
... 根据RLKs细胞外结构域的不同特点, 同时参照动物RLKs, 人们将植物RLKs分为6种类型: 富含亮氨酸重复序列型(leucine-rich repeats, LRRs)、S-结构域型(S-domain)、类凝集素型(lectin-like)、类表皮生长因子型(epidermal growth factor-like, EGF)、类肿瘤坏死因子型(tumor-necrosis factor receptor-like, TNFR)和类PR5型(pathogenesis related protein-5 like receptor kinases, PR5K) (Walker, 1994; Becraft et al., 1996; Hu and Reddy, 1997; Wang et al., 1998; Kobe and Kajava, 2001; Shiu and Bleecker, 2001).凝集素类受体蛋白激酶(lectin receptor-like kinases, LecRLKs)是RLKs家族中的一个亚族, 是指含有1个或多个可与单糖或寡聚糖特异可逆结合的非催化结构域的植物蛋白(Peumans and van Damme, 1995).已知在拟南芥和水稻中分别存在77和173个LecRLKs (Bouwmeester and Govers, 2009; Vaid et al., 2012).到目前为止, 在酵母(Saccharomyces cerevisiae)和人体基因组中还未找到LecRLKs的同源基因(Navarro-Gochicoa et al., 2003).已有研究表明, LecRLKs蛋白可参与多种生物/非生物胁迫途径以及植物发育调控(Vaid et al., 2012).本文简要阐述植物凝集素类受体蛋白激酶的研究历史、结构特点与分类及其生物学功能, 以期增进对凝集素类受体蛋白激酶的认识. ...
... 凝集素类受体蛋白激酶主要由胞外区(包含凝集素结构域)、跨膜区以及胞内区(包含近膜区和激酶结构域)组成(图1).凝集素结构域可与糖类分子特异可逆结合, 该结构域在不同受体中差异较大, 可分为不同类型, 主要参与信号识别和结合(Peumans and van Damme, 1995; Vaid et al., 2012).跨膜结构域一般由18-25个氨基酸组成, 不具有明显的保守性, 但该部位对于凝集素类受体蛋白激酶的功能极其重要(Morillo and Tax, 2006).例如, 水稻Pi-d2蛋白跨膜结构域中的1个氨基酸发生改变, 可导致该蛋白完全丧失功能(Chen et al., 2006).近膜区主要是将跨膜结构域和激酶结构域分开, 该区由50个左右的氨基酸组成, 保守性较高, 其C端一般包含PxPFxYxELxxATxG (x表示任一氨基酸)的未知功能亲水区(Walker, 1994; Barre et al., 2002).激酶结构域是高度保守的结构域, 具有磷酸化结合位点, 位于细胞质区域, 一般由250- 300个氨基酸组成, 主要负责传递外界信号(Li and Gray, 1997).激酶结构域的N端包含GxGxxG结构域, 能够影响核苷酸的结合; 而激酶结构域的C端由43- 66个氨基酸组成, 为激酶的催化活性所必需(Barre et al., 2002).与酪氨酸类受体蛋白激酶含有DLAARN和FPIKWMAPE基序不同, 凝集素类受体蛋白激酶的激酶结构域中含有DIKPAN和GT(FIL)GYIAPE基序, 属于丝/苏氨酸激酶(Hervé et al., 1999). ...
A lectin S-domain receptor kinase mediates lipopolysaccharide sensing in Arabidopsis thaliana 2 2015
... ZmPK1是第1个被鉴定到的类受体蛋白激酶, 它属于G型凝集素类受体蛋白激酶.该激酶的细胞外结构域与芸薹属自交不亲和性位点编码的糖蛋白类似, 参与调控花粉与柱头之间的识别(Walker and Zhang, 1990).Hervé等(1996)在拟南芥中鉴定到第1个L型凝集素类受体蛋白激酶Ath.LecRK1, 该蛋白激酶具有豆类类凝集素结构, 可能在植物低聚糖和激素信号转导过程中发挥作用(Newman et al., 1994).根据LecRLKs细胞外凝集素结构域的特点, LecRLKs被归类为RLKs的一个亚家族(Shiu and Bleecker, 2001; Goff et al., 2002; Yu et al., 2002).虽然LecRLKs基因发现较早, 但对这些基因家族的系统分类却相对较晚, 而早期对于LecRLKs基因的研究也主要集中在基因表达等较为初步的层面(Nishiguchi et al., 2002; Riou et al., 2002; Navarro-Gochicoa et al., 2003; He et al., 2004; Zuo et al., 2004).近年来, 越来越多的研究表明, LecRLKs蛋白的功能极其广泛, 可参与调控各种生物/非生物胁迫和植物发育(Chen et al., 2006; Kanzaki et al., 2008; Joshi et al., 2010; Bonaventure, 2011; Desclos-Theveniau et al., 2012; Gouhier-Darimont et al., 2013; Li et al., 2014; Ranf et al., 2015; Luo et al., 2017; Ma et al., 2018; Labbé et al., 2019) (表1). ...
... 凝集素类受体蛋白激酶参与植物对病原菌的响应.例如, 水稻Pi-d2蛋白属于G型凝集素类受体蛋白激酶, 其转基因植株对中国稻瘟病菌小种ZB15具有较高的抗性(Chen et al., 2006).Fan等(2018)通过EMS诱变筛选获得E3泛素连接酶基因SPL11突变所诱导细胞死亡的抑制子SDS2, 该基因编码1个S-domain类受体蛋白激酶, 属于G型凝集素类受体蛋白激酶.研究表明, SDS2基因突变可降低植物的免疫反应, 易感染稻瘟病菌; 相反, 过表达SDS2基因导致细胞程序性死亡、增强免疫反应和对稻瘟病菌的抗性.此外, SDS2蛋白可与细胞质类受体蛋白激酶OsRLCK118/176互作, 通过磷酸化调控NADPH氧化酶OsRbohB, 刺激ROS产生, 从而正向调控免疫反应(Fan et al., 2018).在烟草(Nicotiana tabacum)中, G型凝集素类受体蛋白激酶基因Nt-Sd-RLK受脂多糖诱导表达, 在免疫信号的识别和防御过程中发挥重要作用(Sanabria et al., 2012).Ranf等(2015)通过筛选脂多糖不敏感型缺陷突变体, 获得了1个S-domain类受体蛋白激酶基因LORE.LORE基因突变导致植物易感染丁香假单胞杆菌, 它在介导脂多糖识别引起的免疫反应中发挥功能.在拟南芥中, 缺失L型凝集素类受体蛋白激酶基因LecRK-V.5会增强植物对表面接种Pst DC3000而产生的抗性, 但对通过渗透方法接种所产生的抗性无影响, 提示LecRK-V.5基因可能在植物早期免疫阶段发挥功能; 相反, 过表达LecRK- V.5基因导致植物更易感染Pst DC3000 (Desclos- Theveniau et al., 2012).研究表明, LecRK-V.5基因突变引起的抗病性增强表型与气孔的组成型关闭相关, 而LecRK-V.5基因过表达引起的易感病表型与早期气孔的重新打开有关.LecRK-V.5基因突变和过表达表型结果表明, LecRK-V.5基因可以逆转由于细菌感染诱导的气孔关闭, 从而减轻植株因缺乏CO2而造成的伤害(Desclos-Theveniau et al., 2012).另一个凝集素类受体蛋白激酶基因LecRK-VI.2被报道在抵抗丁香假单胞杆菌和胡萝卜软腐果胶杆菌方面具有重要功能(Singh et al., 2012).LecRK-VI.2基因突变导致PTI (pattern-triggered immunity)反应相关抗病基因表达量降低、胼胝质沉淀形成受阻和气孔关闭出现缺陷等.LecRK-VI.2基因过表达和全基因组分析实验表明, LecRK-VI.2基因正向调控PTI反应.Singh等(2012)研究表明, LecRK-VI.2蛋白位于MAPK信号的上游, 但独立于ROS信号和BIK1的磷酸化调控信号途径.此外, LecRK-VI.2蛋白对于β-氨基丁酸诱导的抗性反应和PTI反应是必需的(Singh et al., 2012).在拟南芥中, LecRK-I.9基因突变导致植物对ATP不敏感, 从而降低ATP引起的防御反应.研究发现, LecRK-I.9蛋白与ATP结合且高度亲和, 它们的结合对于ATP诱导的钙离子响应、MAPK的激活和基因表达是必需的(Choi et al., 2014).Wang等(2015)研究发现, 拟南芥LecRK-IX.1和LecRK-IX.2基因突变可降低植物对疫霉菌的抵抗能力, 且双突变会加剧表型; 相反, 过表达LecRK-IX.1和LecRK-IX.2基因能增强植物的抗病性, 但同时导致自发性的细胞死亡.此外, LecRK-IX.1和LecRK-IX.2蛋白的凝集素结构域和激酶结构域对于抗疫霉菌是必需的; 而凝集素结构域对于诱导细胞死亡为非必需(Wang et al., 2015). ...
Expression of an Arabidopsis lectin kinase receptor gene, lecRK-a1, is induced during senescence, wounding and in response to oligogalacturonic acids 3 2002
... ZmPK1是第1个被鉴定到的类受体蛋白激酶, 它属于G型凝集素类受体蛋白激酶.该激酶的细胞外结构域与芸薹属自交不亲和性位点编码的糖蛋白类似, 参与调控花粉与柱头之间的识别(Walker and Zhang, 1990).Hervé等(1996)在拟南芥中鉴定到第1个L型凝集素类受体蛋白激酶Ath.LecRK1, 该蛋白激酶具有豆类类凝集素结构, 可能在植物低聚糖和激素信号转导过程中发挥作用(Newman et al., 1994).根据LecRLKs细胞外凝集素结构域的特点, LecRLKs被归类为RLKs的一个亚家族(Shiu and Bleecker, 2001; Goff et al., 2002; Yu et al., 2002).虽然LecRLKs基因发现较早, 但对这些基因家族的系统分类却相对较晚, 而早期对于LecRLKs基因的研究也主要集中在基因表达等较为初步的层面(Nishiguchi et al., 2002; Riou et al., 2002; Navarro-Gochicoa et al., 2003; He et al., 2004; Zuo et al., 2004).近年来, 越来越多的研究表明, LecRLKs蛋白的功能极其广泛, 可参与调控各种生物/非生物胁迫和植物发育(Chen et al., 2006; Kanzaki et al., 2008; Joshi et al., 2010; Bonaventure, 2011; Desclos-Theveniau et al., 2012; Gouhier-Darimont et al., 2013; Li et al., 2014; Ranf et al., 2015; Luo et al., 2017; Ma et al., 2018; Labbé et al., 2019) (表1). ...
... 凝集素类受体蛋白激酶参与机械损伤引起的胁迫反应.在拟南芥中, LecRK-V.5基因的表达被局部激活以响应机械损伤信号(Riou et al., 2002).在黑杨(Populus nigra)受到机械损伤的幼叶中, PnLPK基因的mRNA丰度增加(Nishiguchi et al., 2002). ...
... 在拟南芥中, LecRK-V.5基因在衰老组织和受伤的细胞周围大量表达, 推测其可能通过调控细胞分裂来修复受损细胞, 表明LecRK-V.5基因可能在植物发育过程中发挥重要功能(Riou et al., 2002).棉花(Gossypium hirsutum) GhLecRK基因在嫩枝和发育的纤维中大量表达, 对于纤维的发育至关重要(Zuo et al., 2004). ...
Plant lectins: occurrence, biochemistry, functions and applications 1 2001
... 凝集素蛋白在植物整个生命过程中的作用非常重要, 并且在细胞与细胞的交流、植物的发育和防御反应中扮演重要角色(Bellande et al., 2017).已知凝集素蛋白可参与多种生理学过程, 包括蛋白分选、识别、胚胎发生和发育; 凝集素蛋白可与外源糖基结合, 从而保护植物免受食草动物或植食性无脊椎动物的侵害(Brewin and Kardailsky, 1997; Brill et al., 2001; Rudiger and Gabius, 2001).由于凝集素类受体蛋白激酶的凝集素识别结构域与凝集素蛋白相似, 推测其可能具有识别和转导糖信号的功能(Navarro-Gochicoa et al., 2003).近期的研究表明, 凝集素类受体蛋白激酶在响应胁迫信号、抗病虫害和调控植物发育方面都具有非常重要的功能(表1; 图2). ...
Molecular characterisation and regulation of a Nicotiana tabacum S- domain receptor-like kinase gene induced during an early rapid response to lipopolysaccharides 1 2012
... 凝集素类受体蛋白激酶参与植物对病原菌的响应.例如, 水稻Pi-d2蛋白属于G型凝集素类受体蛋白激酶, 其转基因植株对中国稻瘟病菌小种ZB15具有较高的抗性(Chen et al., 2006).Fan等(2018)通过EMS诱变筛选获得E3泛素连接酶基因SPL11突变所诱导细胞死亡的抑制子SDS2, 该基因编码1个S-domain类受体蛋白激酶, 属于G型凝集素类受体蛋白激酶.研究表明, SDS2基因突变可降低植物的免疫反应, 易感染稻瘟病菌; 相反, 过表达SDS2基因导致细胞程序性死亡、增强免疫反应和对稻瘟病菌的抗性.此外, SDS2蛋白可与细胞质类受体蛋白激酶OsRLCK118/176互作, 通过磷酸化调控NADPH氧化酶OsRbohB, 刺激ROS产生, 从而正向调控免疫反应(Fan et al., 2018).在烟草(Nicotiana tabacum)中, G型凝集素类受体蛋白激酶基因Nt-Sd-RLK受脂多糖诱导表达, 在免疫信号的识别和防御过程中发挥重要作用(Sanabria et al., 2012).Ranf等(2015)通过筛选脂多糖不敏感型缺陷突变体, 获得了1个S-domain类受体蛋白激酶基因LORE.LORE基因突变导致植物易感染丁香假单胞杆菌, 它在介导脂多糖识别引起的免疫反应中发挥功能.在拟南芥中, 缺失L型凝集素类受体蛋白激酶基因LecRK-V.5会增强植物对表面接种Pst DC3000而产生的抗性, 但对通过渗透方法接种所产生的抗性无影响, 提示LecRK-V.5基因可能在植物早期免疫阶段发挥功能; 相反, 过表达LecRK- V.5基因导致植物更易感染Pst DC3000 (Desclos- Theveniau et al., 2012).研究表明, LecRK-V.5基因突变引起的抗病性增强表型与气孔的组成型关闭相关, 而LecRK-V.5基因过表达引起的易感病表型与早期气孔的重新打开有关.LecRK-V.5基因突变和过表达表型结果表明, LecRK-V.5基因可以逆转由于细菌感染诱导的气孔关闭, 从而减轻植株因缺乏CO2而造成的伤害(Desclos-Theveniau et al., 2012).另一个凝集素类受体蛋白激酶基因LecRK-VI.2被报道在抵抗丁香假单胞杆菌和胡萝卜软腐果胶杆菌方面具有重要功能(Singh et al., 2012).LecRK-VI.2基因突变导致PTI (pattern-triggered immunity)反应相关抗病基因表达量降低、胼胝质沉淀形成受阻和气孔关闭出现缺陷等.LecRK-VI.2基因过表达和全基因组分析实验表明, LecRK-VI.2基因正向调控PTI反应.Singh等(2012)研究表明, LecRK-VI.2蛋白位于MAPK信号的上游, 但独立于ROS信号和BIK1的磷酸化调控信号途径.此外, LecRK-VI.2蛋白对于β-氨基丁酸诱导的抗性反应和PTI反应是必需的(Singh et al., 2012).在拟南芥中, LecRK-I.9基因突变导致植物对ATP不敏感, 从而降低ATP引起的防御反应.研究发现, LecRK-I.9蛋白与ATP结合且高度亲和, 它们的结合对于ATP诱导的钙离子响应、MAPK的激活和基因表达是必需的(Choi et al., 2014).Wang等(2015)研究发现, 拟南芥LecRK-IX.1和LecRK-IX.2基因突变可降低植物对疫霉菌的抵抗能力, 且双突变会加剧表型; 相反, 过表达LecRK-IX.1和LecRK-IX.2基因能增强植物的抗病性, 但同时导致自发性的细胞死亡.此外, LecRK-IX.1和LecRK-IX.2蛋白的凝集素结构域和激酶结构域对于抗疫霉菌是必需的; 而凝集素结构域对于诱导细胞死亡为非必需(Wang et al., 2015). ...
S locus genes and the evolution of self-fertility in Arabidopsis thaliana 1 2007
... G型凝集素类受体蛋白激酶曾被称为B-type LecRLK, 其细胞外凝集素结构域具有β-barre结构, 属于α-D甘露醇特异的植物凝集素类受体蛋白激酶(Shiu and Bleecker, 2001).除凝集素结构域外, G型凝集素类受体蛋白激酶的细胞外结构还包含EGF和/或PAN (plasminogen-apple-nematode)结构域(Vaid et al., 2012) (图1).其中, EGF结构域也被称作S-locus结构域, 在十字花科家族的自交不亲和中起作用(Tanksley and Loaiza-Figueroa, 1985; Kusaba et al., 2001).S-domain RLK因被发现参与有花植物自交不亲和(SI)而又被称为S-locus receptor kinase SRK, 属于G型凝集素类受体蛋白激酶(Sherman- Broyles et al., 2007).而PAN为富含半胱氨酸的结构域, 参与形成二硫键, 且PAN结构域的复杂功能是通过蛋白与蛋白以及蛋白与碳水化合物之间的互作来介导(Tordai et al., 1999; Shiu and Bleecker, 2001; Loris, 2002; Naithani et al., 2007). ...
Receptor-like kinases from Arabidopsis form a monophyletic gene family related to animal receptor kinases 6 2001
... 受体蛋白激酶(receptor protein kinases, RPKs)是一类能够与配基结合并具有酶活性的受体(Hunter, 1991).RPKs蛋白在动物中研究较早, 主要分为酪氨酸激酶(receptor tyrosine kinases, RTKs)和丝/苏氨酸激酶(serine/threonine kinase receptors, STRKs) 2种.与动物不同, 植物中受体蛋白激酶主要属于丝/苏氨酸激酶.目前, 已在植物中陆续鉴定到多个与动物受体蛋白激酶结构相似的蛋白, 这类蛋白绝大部分功能未知, 被称为类受体蛋白激酶(receptor-like kinases, RLKs) (Walker, 1994; Shiu and Bleecker, 2001).RLKs是一种普遍存在于植物体内的蛋白, 能够感知和传递各种信号, 在拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)中分别有超过600和1 100个该类蛋白(Shiu and Bleecker, 2001).大部分RLKs属于细胞质膜蛋白, 主要功能是通过它们的细胞外配体识别结构域和细胞内激酶结构域传递各种信号, 最终产生一系列生物效应(Vaid et al., 2013). ...
... )中分别有超过600和1 100个该类蛋白(Shiu and Bleecker, 2001).大部分RLKs属于细胞质膜蛋白, 主要功能是通过它们的细胞外配体识别结构域和细胞内激酶结构域传递各种信号, 最终产生一系列生物效应(Vaid et al., 2013). ...
... 根据RLKs细胞外结构域的不同特点, 同时参照动物RLKs, 人们将植物RLKs分为6种类型: 富含亮氨酸重复序列型(leucine-rich repeats, LRRs)、S-结构域型(S-domain)、类凝集素型(lectin-like)、类表皮生长因子型(epidermal growth factor-like, EGF)、类肿瘤坏死因子型(tumor-necrosis factor receptor-like, TNFR)和类PR5型(pathogenesis related protein-5 like receptor kinases, PR5K) (Walker, 1994; Becraft et al., 1996; Hu and Reddy, 1997; Wang et al., 1998; Kobe and Kajava, 2001; Shiu and Bleecker, 2001).凝集素类受体蛋白激酶(lectin receptor-like kinases, LecRLKs)是RLKs家族中的一个亚族, 是指含有1个或多个可与单糖或寡聚糖特异可逆结合的非催化结构域的植物蛋白(Peumans and van Damme, 1995).已知在拟南芥和水稻中分别存在77和173个LecRLKs (Bouwmeester and Govers, 2009; Vaid et al., 2012).到目前为止, 在酵母(Saccharomyces cerevisiae)和人体基因组中还未找到LecRLKs的同源基因(Navarro-Gochicoa et al., 2003).已有研究表明, LecRLKs蛋白可参与多种生物/非生物胁迫途径以及植物发育调控(Vaid et al., 2012).本文简要阐述植物凝集素类受体蛋白激酶的研究历史、结构特点与分类及其生物学功能, 以期增进对凝集素类受体蛋白激酶的认识. ...
... ZmPK1是第1个被鉴定到的类受体蛋白激酶, 它属于G型凝集素类受体蛋白激酶.该激酶的细胞外结构域与芸薹属自交不亲和性位点编码的糖蛋白类似, 参与调控花粉与柱头之间的识别(Walker and Zhang, 1990).Hervé等(1996)在拟南芥中鉴定到第1个L型凝集素类受体蛋白激酶Ath.LecRK1, 该蛋白激酶具有豆类类凝集素结构, 可能在植物低聚糖和激素信号转导过程中发挥作用(Newman et al., 1994).根据LecRLKs细胞外凝集素结构域的特点, LecRLKs被归类为RLKs的一个亚家族(Shiu and Bleecker, 2001; Goff et al., 2002; Yu et al., 2002).虽然LecRLKs基因发现较早, 但对这些基因家族的系统分类却相对较晚, 而早期对于LecRLKs基因的研究也主要集中在基因表达等较为初步的层面(Nishiguchi et al., 2002; Riou et al., 2002; Navarro-Gochicoa et al., 2003; He et al., 2004; Zuo et al., 2004).近年来, 越来越多的研究表明, LecRLKs蛋白的功能极其广泛, 可参与调控各种生物/非生物胁迫和植物发育(Chen et al., 2006; Kanzaki et al., 2008; Joshi et al., 2010; Bonaventure, 2011; Desclos-Theveniau et al., 2012; Gouhier-Darimont et al., 2013; Li et al., 2014; Ranf et al., 2015; Luo et al., 2017; Ma et al., 2018; Labbé et al., 2019) (表1). ...
... G型凝集素类受体蛋白激酶曾被称为B-type LecRLK, 其细胞外凝集素结构域具有β-barre结构, 属于α-D甘露醇特异的植物凝集素类受体蛋白激酶(Shiu and Bleecker, 2001).除凝集素结构域外, G型凝集素类受体蛋白激酶的细胞外结构还包含EGF和/或PAN (plasminogen-apple-nematode)结构域(Vaid et al., 2012) (图1).其中, EGF结构域也被称作S-locus结构域, 在十字花科家族的自交不亲和中起作用(Tanksley and Loaiza-Figueroa, 1985; Kusaba et al., 2001).S-domain RLK因被发现参与有花植物自交不亲和(SI)而又被称为S-locus receptor kinase SRK, 属于G型凝集素类受体蛋白激酶(Sherman- Broyles et al., 2007).而PAN为富含半胱氨酸的结构域, 参与形成二硫键, 且PAN结构域的复杂功能是通过蛋白与蛋白以及蛋白与碳水化合物之间的互作来介导(Tordai et al., 1999; Shiu and Bleecker, 2001; Loris, 2002; Naithani et al., 2007). ...
... ; Shiu and Bleecker, 2001; Loris, 2002; Naithani et al., 2007). ...
The lectin receptor kinase-VI.2 is required for priming and positively regulates Arabidopsis pattern-triggered immunity 3 2012
... 凝集素类受体蛋白激酶参与植物对病原菌的响应.例如, 水稻Pi-d2蛋白属于G型凝集素类受体蛋白激酶, 其转基因植株对中国稻瘟病菌小种ZB15具有较高的抗性(Chen et al., 2006).Fan等(2018)通过EMS诱变筛选获得E3泛素连接酶基因SPL11突变所诱导细胞死亡的抑制子SDS2, 该基因编码1个S-domain类受体蛋白激酶, 属于G型凝集素类受体蛋白激酶.研究表明, SDS2基因突变可降低植物的免疫反应, 易感染稻瘟病菌; 相反, 过表达SDS2基因导致细胞程序性死亡、增强免疫反应和对稻瘟病菌的抗性.此外, SDS2蛋白可与细胞质类受体蛋白激酶OsRLCK118/176互作, 通过磷酸化调控NADPH氧化酶OsRbohB, 刺激ROS产生, 从而正向调控免疫反应(Fan et al., 2018).在烟草(Nicotiana tabacum)中, G型凝集素类受体蛋白激酶基因Nt-Sd-RLK受脂多糖诱导表达, 在免疫信号的识别和防御过程中发挥重要作用(Sanabria et al., 2012).Ranf等(2015)通过筛选脂多糖不敏感型缺陷突变体, 获得了1个S-domain类受体蛋白激酶基因LORE.LORE基因突变导致植物易感染丁香假单胞杆菌, 它在介导脂多糖识别引起的免疫反应中发挥功能.在拟南芥中, 缺失L型凝集素类受体蛋白激酶基因LecRK-V.5会增强植物对表面接种Pst DC3000而产生的抗性, 但对通过渗透方法接种所产生的抗性无影响, 提示LecRK-V.5基因可能在植物早期免疫阶段发挥功能; 相反, 过表达LecRK- V.5基因导致植物更易感染Pst DC3000 (Desclos- Theveniau et al., 2012).研究表明, LecRK-V.5基因突变引起的抗病性增强表型与气孔的组成型关闭相关, 而LecRK-V.5基因过表达引起的易感病表型与早期气孔的重新打开有关.LecRK-V.5基因突变和过表达表型结果表明, LecRK-V.5基因可以逆转由于细菌感染诱导的气孔关闭, 从而减轻植株因缺乏CO2而造成的伤害(Desclos-Theveniau et al., 2012).另一个凝集素类受体蛋白激酶基因LecRK-VI.2被报道在抵抗丁香假单胞杆菌和胡萝卜软腐果胶杆菌方面具有重要功能(Singh et al., 2012).LecRK-VI.2基因突变导致PTI (pattern-triggered immunity)反应相关抗病基因表达量降低、胼胝质沉淀形成受阻和气孔关闭出现缺陷等.LecRK-VI.2基因过表达和全基因组分析实验表明, LecRK-VI.2基因正向调控PTI反应.Singh等(2012)研究表明, LecRK-VI.2蛋白位于MAPK信号的上游, 但独立于ROS信号和BIK1的磷酸化调控信号途径.此外, LecRK-VI.2蛋白对于β-氨基丁酸诱导的抗性反应和PTI反应是必需的(Singh et al., 2012).在拟南芥中, LecRK-I.9基因突变导致植物对ATP不敏感, 从而降低ATP引起的防御反应.研究发现, LecRK-I.9蛋白与ATP结合且高度亲和, 它们的结合对于ATP诱导的钙离子响应、MAPK的激活和基因表达是必需的(Choi et al., 2014).Wang等(2015)研究发现, 拟南芥LecRK-IX.1和LecRK-IX.2基因突变可降低植物对疫霉菌的抵抗能力, 且双突变会加剧表型; 相反, 过表达LecRK-IX.1和LecRK-IX.2基因能增强植物的抗病性, 但同时导致自发性的细胞死亡.此外, LecRK-IX.1和LecRK-IX.2蛋白的凝集素结构域和激酶结构域对于抗疫霉菌是必需的; 而凝集素结构域对于诱导细胞死亡为非必需(Wang et al., 2015). ...
... 2基因正向调控PTI反应.Singh等(2012)研究表明, LecRK-VI.2蛋白位于MAPK信号的上游, 但独立于ROS信号和BIK1的磷酸化调控信号途径.此外, LecRK-VI.2蛋白对于β-氨基丁酸诱导的抗性反应和PTI反应是必需的(Singh et al., 2012).在拟南芥中, LecRK-I.9基因突变导致植物对ATP不敏感, 从而降低ATP引起的防御反应.研究发现, LecRK-I.9蛋白与ATP结合且高度亲和, 它们的结合对于ATP诱导的钙离子响应、MAPK的激活和基因表达是必需的(Choi et al., 2014).Wang等(2015)研究发现, 拟南芥LecRK-IX.1和LecRK-IX.2基因突变可降低植物对疫霉菌的抵抗能力, 且双突变会加剧表型; 相反, 过表达LecRK-IX.1和LecRK-IX.2基因能增强植物的抗病性, 但同时导致自发性的细胞死亡.此外, LecRK-IX.1和LecRK-IX.2蛋白的凝集素结构域和激酶结构域对于抗疫霉菌是必需的; 而凝集素结构域对于诱导细胞死亡为非必需(Wang et al., 2015). ...
... 研究表明, LecRK-VI.2蛋白位于MAPK信号的上游, 但独立于ROS信号和BIK1的磷酸化调控信号途径.此外, LecRK-VI.2蛋白对于β-氨基丁酸诱导的抗性反应和PTI反应是必需的(Singh et al., 2012).在拟南芥中, LecRK-I.9基因突变导致植物对ATP不敏感, 从而降低ATP引起的防御反应.研究发现, LecRK-I.9蛋白与ATP结合且高度亲和, 它们的结合对于ATP诱导的钙离子响应、MAPK的激活和基因表达是必需的(Choi et al., 2014).Wang等(2015)研究发现, 拟南芥LecRK-IX.1和LecRK-IX.2基因突变可降低植物对疫霉菌的抵抗能力, 且双突变会加剧表型; 相反, 过表达LecRK-IX.1和LecRK-IX.2基因能增强植物的抗病性, 但同时导致自发性的细胞死亡.此外, LecRK-IX.1和LecRK-IX.2蛋白的凝集素结构域和激酶结构域对于抗疫霉菌是必需的; 而凝集素结构域对于诱导细胞死亡为非必需(Wang et al., 2015). ...
GsSRK, a G-type lectin S-receptor- like serine/threonine protein kinase, is a positive regulator of plant tolerance to salt stress 1 2013
... 凝集素类受体蛋白激酶参与盐胁迫响应.拟南芥LecRK-V.2基因缺失会降低植物早期发育阶段对盐的敏感性(Deng et al., 2009).He等(2004)发现, 拟南芥LecRK2基因在盐胁迫时其表达量有不同程度的上调, 并进一步证明它受到乙烯(ET)信号通路的调控.豌豆(Pisum sativum) PsLecRLK基因在盐胁迫下被诱导表达, 而PsLecRLK基因过表达可缓解离子平衡和渗透压, 从而增强植物的耐盐性(Vaid et al., 2015).Sun等(2013)发现野大豆(Glycine soja) GsSRK基因的表达受盐胁迫诱导, 而过表达该基因可增强植物的耐盐性, 并使植物在耐盐环境中表现出更高的产量, 同时还伴随叶绿素含量增加、离子泄漏降低、株高增加和果实增多等表型.在水稻中, 盐胁迫能够增加SIT1蛋白激酶活性, 从而磷酸化其下游效应蛋白MAPK3和MAPK6; 此外, SIT1蛋白还可介导乙烯的生成和盐胁迫诱导的乙烯信号途径(Li et al., 2014).而在拟南芥中, SIT1蛋白激酶在盐胁迫下还可促进活性氧的积累, 最终导致植物生长受抑制甚至死亡, 该过程依赖于MAPK3/6和乙烯信号途径(Li et al., 2014).Ma等(2018)发现在本氏烟草(N. benthamiana)中过表达豆梨(Pyrus calleryana) L型凝集素类受体蛋白激酶基因PbLRK138导致细胞死亡, 从而增强植物的耐盐性. ...
Gametophytic self-incompatibility is controlled by a single major locus on chromosome 1 in Lycopersicon peruvianum 1 1985
... G型凝集素类受体蛋白激酶曾被称为B-type LecRLK, 其细胞外凝集素结构域具有β-barre结构, 属于α-D甘露醇特异的植物凝集素类受体蛋白激酶(Shiu and Bleecker, 2001).除凝集素结构域外, G型凝集素类受体蛋白激酶的细胞外结构还包含EGF和/或PAN (plasminogen-apple-nematode)结构域(Vaid et al., 2012) (图1).其中, EGF结构域也被称作S-locus结构域, 在十字花科家族的自交不亲和中起作用(Tanksley and Loaiza-Figueroa, 1985; Kusaba et al., 2001).S-domain RLK因被发现参与有花植物自交不亲和(SI)而又被称为S-locus receptor kinase SRK, 属于G型凝集素类受体蛋白激酶(Sherman- Broyles et al., 2007).而PAN为富含半胱氨酸的结构域, 参与形成二硫键, 且PAN结构域的复杂功能是通过蛋白与蛋白以及蛋白与碳水化合物之间的互作来介导(Tordai et al., 1999; Shiu and Bleecker, 2001; Loris, 2002; Naithani et al., 2007). ...
The PAN module: the N-terminal domains of plasminogen and hepatocyte growth factor are homologous with the apple domains of the prekallikrein family and with a novel domain found in numerous nematode proteins 1 1999
... G型凝集素类受体蛋白激酶曾被称为B-type LecRLK, 其细胞外凝集素结构域具有β-barre结构, 属于α-D甘露醇特异的植物凝集素类受体蛋白激酶(Shiu and Bleecker, 2001).除凝集素结构域外, G型凝集素类受体蛋白激酶的细胞外结构还包含EGF和/或PAN (plasminogen-apple-nematode)结构域(Vaid et al., 2012) (图1).其中, EGF结构域也被称作S-locus结构域, 在十字花科家族的自交不亲和中起作用(Tanksley and Loaiza-Figueroa, 1985; Kusaba et al., 2001).S-domain RLK因被发现参与有花植物自交不亲和(SI)而又被称为S-locus receptor kinase SRK, 属于G型凝集素类受体蛋白激酶(Sherman- Broyles et al., 2007).而PAN为富含半胱氨酸的结构域, 参与形成二硫键, 且PAN结构域的复杂功能是通过蛋白与蛋白以及蛋白与碳水化合物之间的互作来介导(Tordai et al., 1999; Shiu and Bleecker, 2001; Loris, 2002; Naithani et al., 2007). ...
Knights in action: lectin receptor-like kinases in plant development and stress responses 2 2013
... 受体蛋白激酶(receptor protein kinases, RPKs)是一类能够与配基结合并具有酶活性的受体(Hunter, 1991).RPKs蛋白在动物中研究较早, 主要分为酪氨酸激酶(receptor tyrosine kinases, RTKs)和丝/苏氨酸激酶(serine/threonine kinase receptors, STRKs) 2种.与动物不同, 植物中受体蛋白激酶主要属于丝/苏氨酸激酶.目前, 已在植物中陆续鉴定到多个与动物受体蛋白激酶结构相似的蛋白, 这类蛋白绝大部分功能未知, 被称为类受体蛋白激酶(receptor-like kinases, RLKs) (Walker, 1994; Shiu and Bleecker, 2001).RLKs是一种普遍存在于植物体内的蛋白, 能够感知和传递各种信号, 在拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)中分别有超过600和1 100个该类蛋白(Shiu and Bleecker, 2001).大部分RLKs属于细胞质膜蛋白, 主要功能是通过它们的细胞外配体识别结构域和细胞内激酶结构域传递各种信号, 最终产生一系列生物效应(Vaid et al., 2013). ...
... 根据细胞外凝集素结构域的多样性可将凝集素类受体蛋白激酶分为L、G和C型(图1), 其中L和G型为植物特有, 而C型主要存在于哺乳动物中(Wan et al., 2008; Vaid et al., 2013).研究显示, 拟南芥中包含77个凝集素类受体蛋白激酶.而Vaid等(2012)明确指出拟南芥中L、G和C型凝集素类受体蛋白激酶分别为42、32和1个, 其中L型凝集素类受体蛋白激酶不包含LecRK S.2和LecRK S.3两个蛋白激酶(Bouwmeester and Govers, 2009).LecRK S.2和LecRK S.3蛋白含激酶结构域, 无胞外结构域和跨膜结构域.目前暂无明确证据证明这2个类受体蛋白激酶不属于凝集素类受体蛋白激酶亚家族(Bouwmeester and Govers, 2009).在水稻中, L、G和C型凝集素类受体蛋白激酶分别为72、100和1个(Vaid et al., 2012). ...
Pea lectin receptor-like kinase functions in salinity adaptation without yield penalty, by alleviating osmotic and ionic stresses and upregulating stress-responsive genes 1 2015
... 凝集素类受体蛋白激酶参与盐胁迫响应.拟南芥LecRK-V.2基因缺失会降低植物早期发育阶段对盐的敏感性(Deng et al., 2009).He等(2004)发现, 拟南芥LecRK2基因在盐胁迫时其表达量有不同程度的上调, 并进一步证明它受到乙烯(ET)信号通路的调控.豌豆(Pisum sativum) PsLecRLK基因在盐胁迫下被诱导表达, 而PsLecRLK基因过表达可缓解离子平衡和渗透压, 从而增强植物的耐盐性(Vaid et al., 2015).Sun等(2013)发现野大豆(Glycine soja) GsSRK基因的表达受盐胁迫诱导, 而过表达该基因可增强植物的耐盐性, 并使植物在耐盐环境中表现出更高的产量, 同时还伴随叶绿素含量增加、离子泄漏降低、株高增加和果实增多等表型.在水稻中, 盐胁迫能够增加SIT1蛋白激酶活性, 从而磷酸化其下游效应蛋白MAPK3和MAPK6; 此外, SIT1蛋白还可介导乙烯的生成和盐胁迫诱导的乙烯信号途径(Li et al., 2014).而在拟南芥中, SIT1蛋白激酶在盐胁迫下还可促进活性氧的积累, 最终导致植物生长受抑制甚至死亡, 该过程依赖于MAPK3/6和乙烯信号途径(Li et al., 2014).Ma等(2018)发现在本氏烟草(N. benthamiana)中过表达豆梨(Pyrus calleryana) L型凝集素类受体蛋白激酶基因PbLRK138导致细胞死亡, 从而增强植物的耐盐性. ...
Genome-wide analysis of lectin receptor-like kinase family from Arabidopsis and rice 7 2012
... 根据RLKs细胞外结构域的不同特点, 同时参照动物RLKs, 人们将植物RLKs分为6种类型: 富含亮氨酸重复序列型(leucine-rich repeats, LRRs)、S-结构域型(S-domain)、类凝集素型(lectin-like)、类表皮生长因子型(epidermal growth factor-like, EGF)、类肿瘤坏死因子型(tumor-necrosis factor receptor-like, TNFR)和类PR5型(pathogenesis related protein-5 like receptor kinases, PR5K) (Walker, 1994; Becraft et al., 1996; Hu and Reddy, 1997; Wang et al., 1998; Kobe and Kajava, 2001; Shiu and Bleecker, 2001).凝集素类受体蛋白激酶(lectin receptor-like kinases, LecRLKs)是RLKs家族中的一个亚族, 是指含有1个或多个可与单糖或寡聚糖特异可逆结合的非催化结构域的植物蛋白(Peumans and van Damme, 1995).已知在拟南芥和水稻中分别存在77和173个LecRLKs (Bouwmeester and Govers, 2009; Vaid et al., 2012).到目前为止, 在酵母(Saccharomyces cerevisiae)和人体基因组中还未找到LecRLKs的同源基因(Navarro-Gochicoa et al., 2003).已有研究表明, LecRLKs蛋白可参与多种生物/非生物胁迫途径以及植物发育调控(Vaid et al., 2012).本文简要阐述植物凝集素类受体蛋白激酶的研究历史、结构特点与分类及其生物学功能, 以期增进对凝集素类受体蛋白激酶的认识. ...
... ).已有研究表明, LecRLKs蛋白可参与多种生物/非生物胁迫途径以及植物发育调控(Vaid et al., 2012).本文简要阐述植物凝集素类受体蛋白激酶的研究历史、结构特点与分类及其生物学功能, 以期增进对凝集素类受体蛋白激酶的认识. ...
... 凝集素类受体蛋白激酶主要由胞外区(包含凝集素结构域)、跨膜区以及胞内区(包含近膜区和激酶结构域)组成(图1).凝集素结构域可与糖类分子特异可逆结合, 该结构域在不同受体中差异较大, 可分为不同类型, 主要参与信号识别和结合(Peumans and van Damme, 1995; Vaid et al., 2012).跨膜结构域一般由18-25个氨基酸组成, 不具有明显的保守性, 但该部位对于凝集素类受体蛋白激酶的功能极其重要(Morillo and Tax, 2006).例如, 水稻Pi-d2蛋白跨膜结构域中的1个氨基酸发生改变, 可导致该蛋白完全丧失功能(Chen et al., 2006).近膜区主要是将跨膜结构域和激酶结构域分开, 该区由50个左右的氨基酸组成, 保守性较高, 其C端一般包含PxPFxYxELxxATxG (x表示任一氨基酸)的未知功能亲水区(Walker, 1994; Barre et al., 2002).激酶结构域是高度保守的结构域, 具有磷酸化结合位点, 位于细胞质区域, 一般由250- 300个氨基酸组成, 主要负责传递外界信号(Li and Gray, 1997).激酶结构域的N端包含GxGxxG结构域, 能够影响核苷酸的结合; 而激酶结构域的C端由43- 66个氨基酸组成, 为激酶的催化活性所必需(Barre et al., 2002).与酪氨酸类受体蛋白激酶含有DLAARN和FPIKWMAPE基序不同, 凝集素类受体蛋白激酶的激酶结构域中含有DIKPAN和GT(FIL)GYIAPE基序, 属于丝/苏氨酸激酶(Hervé et al., 1999). ...
... 根据细胞外凝集素结构域的多样性可将凝集素类受体蛋白激酶分为L、G和C型(图1), 其中L和G型为植物特有, 而C型主要存在于哺乳动物中(Wan et al., 2008; Vaid et al., 2013).研究显示, 拟南芥中包含77个凝集素类受体蛋白激酶.而Vaid等(2012)明确指出拟南芥中L、G和C型凝集素类受体蛋白激酶分别为42、32和1个, 其中L型凝集素类受体蛋白激酶不包含LecRK S.2和LecRK S.3两个蛋白激酶(Bouwmeester and Govers, 2009).LecRK S.2和LecRK S.3蛋白含激酶结构域, 无胞外结构域和跨膜结构域.目前暂无明确证据证明这2个类受体蛋白激酶不属于凝集素类受体蛋白激酶亚家族(Bouwmeester and Govers, 2009).在水稻中, L、G和C型凝集素类受体蛋白激酶分别为72、100和1个(Vaid et al., 2012). ...
... G型凝集素类受体蛋白激酶曾被称为B-type LecRLK, 其细胞外凝集素结构域具有β-barre结构, 属于α-D甘露醇特异的植物凝集素类受体蛋白激酶(Shiu and Bleecker, 2001).除凝集素结构域外, G型凝集素类受体蛋白激酶的细胞外结构还包含EGF和/或PAN (plasminogen-apple-nematode)结构域(Vaid et al., 2012) (图1).其中, EGF结构域也被称作S-locus结构域, 在十字花科家族的自交不亲和中起作用(Tanksley and Loaiza-Figueroa, 1985; Kusaba et al., 2001).S-domain RLK因被发现参与有花植物自交不亲和(SI)而又被称为S-locus receptor kinase SRK, 属于G型凝集素类受体蛋白激酶(Sherman- Broyles et al., 2007).而PAN为富含半胱氨酸的结构域, 参与形成二硫键, 且PAN结构域的复杂功能是通过蛋白与蛋白以及蛋白与碳水化合物之间的互作来介导(Tordai et al., 1999; Shiu and Bleecker, 2001; Loris, 2002; Naithani et al., 2007). ...
... C型凝集素类受体蛋白激酶为钙依赖的凝集素类受体蛋白激酶, 是哺乳动物中的一个大家族蛋白, 主要参与先天性免疫反应以及病原体的识别(Epstein et al., 1996; Hawgood et al., 2001; Cambi et al., 2005).该类型凝集素类受体蛋白激酶的凝集素结构域大量存在于哺乳动物中, 通常通过自我和非自我识别参与免疫反应(Epstein et al., 1996; Hawgood et al., 2001).在植物中, C型凝集素类受体蛋白激酶十分稀少, 属于凝集素类受体蛋白激酶家族中最小的类群.到目前为止, 在水稻和拟南芥中分别仅发现1个C型凝集素类受体蛋白激酶(Bouwmeester and Govers, 2009; Vaid et al., 2012). ...
... 许多研究表明, 凝集素类受体蛋白激酶可参与多种非生物胁迫响应(Vaid et al., 2012). ...
Structure and function of the receptor-like protein kinases of higher plants 3 1994
... 受体蛋白激酶(receptor protein kinases, RPKs)是一类能够与配基结合并具有酶活性的受体(Hunter, 1991).RPKs蛋白在动物中研究较早, 主要分为酪氨酸激酶(receptor tyrosine kinases, RTKs)和丝/苏氨酸激酶(serine/threonine kinase receptors, STRKs) 2种.与动物不同, 植物中受体蛋白激酶主要属于丝/苏氨酸激酶.目前, 已在植物中陆续鉴定到多个与动物受体蛋白激酶结构相似的蛋白, 这类蛋白绝大部分功能未知, 被称为类受体蛋白激酶(receptor-like kinases, RLKs) (Walker, 1994; Shiu and Bleecker, 2001).RLKs是一种普遍存在于植物体内的蛋白, 能够感知和传递各种信号, 在拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)中分别有超过600和1 100个该类蛋白(Shiu and Bleecker, 2001).大部分RLKs属于细胞质膜蛋白, 主要功能是通过它们的细胞外配体识别结构域和细胞内激酶结构域传递各种信号, 最终产生一系列生物效应(Vaid et al., 2013). ...
... 根据RLKs细胞外结构域的不同特点, 同时参照动物RLKs, 人们将植物RLKs分为6种类型: 富含亮氨酸重复序列型(leucine-rich repeats, LRRs)、S-结构域型(S-domain)、类凝集素型(lectin-like)、类表皮生长因子型(epidermal growth factor-like, EGF)、类肿瘤坏死因子型(tumor-necrosis factor receptor-like, TNFR)和类PR5型(pathogenesis related protein-5 like receptor kinases, PR5K) (Walker, 1994; Becraft et al., 1996; Hu and Reddy, 1997; Wang et al., 1998; Kobe and Kajava, 2001; Shiu and Bleecker, 2001).凝集素类受体蛋白激酶(lectin receptor-like kinases, LecRLKs)是RLKs家族中的一个亚族, 是指含有1个或多个可与单糖或寡聚糖特异可逆结合的非催化结构域的植物蛋白(Peumans and van Damme, 1995).已知在拟南芥和水稻中分别存在77和173个LecRLKs (Bouwmeester and Govers, 2009; Vaid et al., 2012).到目前为止, 在酵母(Saccharomyces cerevisiae)和人体基因组中还未找到LecRLKs的同源基因(Navarro-Gochicoa et al., 2003).已有研究表明, LecRLKs蛋白可参与多种生物/非生物胁迫途径以及植物发育调控(Vaid et al., 2012).本文简要阐述植物凝集素类受体蛋白激酶的研究历史、结构特点与分类及其生物学功能, 以期增进对凝集素类受体蛋白激酶的认识. ...
... 凝集素类受体蛋白激酶主要由胞外区(包含凝集素结构域)、跨膜区以及胞内区(包含近膜区和激酶结构域)组成(图1).凝集素结构域可与糖类分子特异可逆结合, 该结构域在不同受体中差异较大, 可分为不同类型, 主要参与信号识别和结合(Peumans and van Damme, 1995; Vaid et al., 2012).跨膜结构域一般由18-25个氨基酸组成, 不具有明显的保守性, 但该部位对于凝集素类受体蛋白激酶的功能极其重要(Morillo and Tax, 2006).例如, 水稻Pi-d2蛋白跨膜结构域中的1个氨基酸发生改变, 可导致该蛋白完全丧失功能(Chen et al., 2006).近膜区主要是将跨膜结构域和激酶结构域分开, 该区由50个左右的氨基酸组成, 保守性较高, 其C端一般包含PxPFxYxELxxATxG (x表示任一氨基酸)的未知功能亲水区(Walker, 1994; Barre et al., 2002).激酶结构域是高度保守的结构域, 具有磷酸化结合位点, 位于细胞质区域, 一般由250- 300个氨基酸组成, 主要负责传递外界信号(Li and Gray, 1997).激酶结构域的N端包含GxGxxG结构域, 能够影响核苷酸的结合; 而激酶结构域的C端由43- 66个氨基酸组成, 为激酶的催化活性所必需(Barre et al., 2002).与酪氨酸类受体蛋白激酶含有DLAARN和FPIKWMAPE基序不同, 凝集素类受体蛋白激酶的激酶结构域中含有DIKPAN和GT(FIL)GYIAPE基序, 属于丝/苏氨酸激酶(Hervé et al., 1999). ...
Relationship of a putative receptor protein kinase from maize to the S-locus glycoproteins of Brassica 1 1990
... ZmPK1是第1个被鉴定到的类受体蛋白激酶, 它属于G型凝集素类受体蛋白激酶.该激酶的细胞外结构域与芸薹属自交不亲和性位点编码的糖蛋白类似, 参与调控花粉与柱头之间的识别(Walker and Zhang, 1990).Hervé等(1996)在拟南芥中鉴定到第1个L型凝集素类受体蛋白激酶Ath.LecRK1, 该蛋白激酶具有豆类类凝集素结构, 可能在植物低聚糖和激素信号转导过程中发挥作用(Newman et al., 1994).根据LecRLKs细胞外凝集素结构域的特点, LecRLKs被归类为RLKs的一个亚家族(Shiu and Bleecker, 2001; Goff et al., 2002; Yu et al., 2002).虽然LecRLKs基因发现较早, 但对这些基因家族的系统分类却相对较晚, 而早期对于LecRLKs基因的研究也主要集中在基因表达等较为初步的层面(Nishiguchi et al., 2002; Riou et al., 2002; Navarro-Gochicoa et al., 2003; He et al., 2004; Zuo et al., 2004).近年来, 越来越多的研究表明, LecRLKs蛋白的功能极其广泛, 可参与调控各种生物/非生物胁迫和植物发育(Chen et al., 2006; Kanzaki et al., 2008; Joshi et al., 2010; Bonaventure, 2011; Desclos-Theveniau et al., 2012; Gouhier-Darimont et al., 2013; Li et al., 2014; Ranf et al., 2015; Luo et al., 2017; Ma et al., 2018; Labbé et al., 2019) (表1). ...
A lectin receptor-like kinase is required for pollen development in Arabidopsis 2 2008
... 根据细胞外凝集素结构域的多样性可将凝集素类受体蛋白激酶分为L、G和C型(图1), 其中L和G型为植物特有, 而C型主要存在于哺乳动物中(Wan et al., 2008; Vaid et al., 2013).研究显示, 拟南芥中包含77个凝集素类受体蛋白激酶.而Vaid等(2012)明确指出拟南芥中L、G和C型凝集素类受体蛋白激酶分别为42、32和1个, 其中L型凝集素类受体蛋白激酶不包含LecRK S.2和LecRK S.3两个蛋白激酶(Bouwmeester and Govers, 2009).LecRK S.2和LecRK S.3蛋白含激酶结构域, 无胞外结构域和跨膜结构域.目前暂无明确证据证明这2个类受体蛋白激酶不属于凝集素类受体蛋白激酶亚家族(Bouwmeester and Govers, 2009).在水稻中, L、G和C型凝集素类受体蛋白激酶分别为72、100和1个(Vaid et al., 2012). ...
Xa21D encodes a receptor-like molecule with a leucine-rich repeat domain that determines race-specific recognition and is subject to adaptive evolution 1 1998
... 根据RLKs细胞外结构域的不同特点, 同时参照动物RLKs, 人们将植物RLKs分为6种类型: 富含亮氨酸重复序列型(leucine-rich repeats, LRRs)、S-结构域型(S-domain)、类凝集素型(lectin-like)、类表皮生长因子型(epidermal growth factor-like, EGF)、类肿瘤坏死因子型(tumor-necrosis factor receptor-like, TNFR)和类PR5型(pathogenesis related protein-5 like receptor kinases, PR5K) (Walker, 1994; Becraft et al., 1996; Hu and Reddy, 1997; Wang et al., 1998; Kobe and Kajava, 2001; Shiu and Bleecker, 2001).凝集素类受体蛋白激酶(lectin receptor-like kinases, LecRLKs)是RLKs家族中的一个亚族, 是指含有1个或多个可与单糖或寡聚糖特异可逆结合的非催化结构域的植物蛋白(Peumans and van Damme, 1995).已知在拟南芥和水稻中分别存在77和173个LecRLKs (Bouwmeester and Govers, 2009; Vaid et al., 2012).到目前为止, 在酵母(Saccharomyces cerevisiae)和人体基因组中还未找到LecRLKs的同源基因(Navarro-Gochicoa et al., 2003).已有研究表明, LecRLKs蛋白可参与多种生物/非生物胁迫途径以及植物发育调控(Vaid et al., 2012).本文简要阐述植物凝集素类受体蛋白激酶的研究历史、结构特点与分类及其生物学功能, 以期增进对凝集素类受体蛋白激酶的认识. ...
Arabidopsis lectin receptor kinases LecRK-IX.1 and LecRK-IX.2 are functional analogs in regulating Phytophthora resistance and plant cell death 2 2015
... 凝集素类受体蛋白激酶参与植物对病原菌的响应.例如, 水稻Pi-d2蛋白属于G型凝集素类受体蛋白激酶, 其转基因植株对中国稻瘟病菌小种ZB15具有较高的抗性(Chen et al., 2006).Fan等(2018)通过EMS诱变筛选获得E3泛素连接酶基因SPL11突变所诱导细胞死亡的抑制子SDS2, 该基因编码1个S-domain类受体蛋白激酶, 属于G型凝集素类受体蛋白激酶.研究表明, SDS2基因突变可降低植物的免疫反应, 易感染稻瘟病菌; 相反, 过表达SDS2基因导致细胞程序性死亡、增强免疫反应和对稻瘟病菌的抗性.此外, SDS2蛋白可与细胞质类受体蛋白激酶OsRLCK118/176互作, 通过磷酸化调控NADPH氧化酶OsRbohB, 刺激ROS产生, 从而正向调控免疫反应(Fan et al., 2018).在烟草(Nicotiana tabacum)中, G型凝集素类受体蛋白激酶基因Nt-Sd-RLK受脂多糖诱导表达, 在免疫信号的识别和防御过程中发挥重要作用(Sanabria et al., 2012).Ranf等(2015)通过筛选脂多糖不敏感型缺陷突变体, 获得了1个S-domain类受体蛋白激酶基因LORE.LORE基因突变导致植物易感染丁香假单胞杆菌, 它在介导脂多糖识别引起的免疫反应中发挥功能.在拟南芥中, 缺失L型凝集素类受体蛋白激酶基因LecRK-V.5会增强植物对表面接种Pst DC3000而产生的抗性, 但对通过渗透方法接种所产生的抗性无影响, 提示LecRK-V.5基因可能在植物早期免疫阶段发挥功能; 相反, 过表达LecRK- V.5基因导致植物更易感染Pst DC3000 (Desclos- Theveniau et al., 2012).研究表明, LecRK-V.5基因突变引起的抗病性增强表型与气孔的组成型关闭相关, 而LecRK-V.5基因过表达引起的易感病表型与早期气孔的重新打开有关.LecRK-V.5基因突变和过表达表型结果表明, LecRK-V.5基因可以逆转由于细菌感染诱导的气孔关闭, 从而减轻植株因缺乏CO2而造成的伤害(Desclos-Theveniau et al., 2012).另一个凝集素类受体蛋白激酶基因LecRK-VI.2被报道在抵抗丁香假单胞杆菌和胡萝卜软腐果胶杆菌方面具有重要功能(Singh et al., 2012).LecRK-VI.2基因突变导致PTI (pattern-triggered immunity)反应相关抗病基因表达量降低、胼胝质沉淀形成受阻和气孔关闭出现缺陷等.LecRK-VI.2基因过表达和全基因组分析实验表明, LecRK-VI.2基因正向调控PTI反应.Singh等(2012)研究表明, LecRK-VI.2蛋白位于MAPK信号的上游, 但独立于ROS信号和BIK1的磷酸化调控信号途径.此外, LecRK-VI.2蛋白对于β-氨基丁酸诱导的抗性反应和PTI反应是必需的(Singh et al., 2012).在拟南芥中, LecRK-I.9基因突变导致植物对ATP不敏感, 从而降低ATP引起的防御反应.研究发现, LecRK-I.9蛋白与ATP结合且高度亲和, 它们的结合对于ATP诱导的钙离子响应、MAPK的激活和基因表达是必需的(Choi et al., 2014).Wang等(2015)研究发现, 拟南芥LecRK-IX.1和LecRK-IX.2基因突变可降低植物对疫霉菌的抵抗能力, 且双突变会加剧表型; 相反, 过表达LecRK-IX.1和LecRK-IX.2基因能增强植物的抗病性, 但同时导致自发性的细胞死亡.此外, LecRK-IX.1和LecRK-IX.2蛋白的凝集素结构域和激酶结构域对于抗疫霉菌是必需的; 而凝集素结构域对于诱导细胞死亡为非必需(Wang et al., 2015). ...
... 基因能增强植物的抗病性, 但同时导致自发性的细胞死亡.此外, LecRK-IX.1和LecRK-IX.2蛋白的凝集素结构域和激酶结构域对于抗疫霉菌是必需的; 而凝集素结构域对于诱导细胞死亡为非必需(Wang et al., 2015). ...
The Arabidopsis A4 subfamily of lectin receptor kinases negatively regulates abscisic acid response in seed germination 2 2009
... 凝集素类受体蛋白激酶参与激素信号响应.(1)参与脱落酸(ABA)信号响应.在拟南芥中, 过表达凝集素类受体蛋白激酶基因LecRK-V.5可抑制植物对ABA的响应能力, 从而影响气孔开闭(Desclos-Theveniau et al., 2012).Xin等(2009)发现, L型凝集素类受体蛋白激酶VI亚家族成员基因LecRK-VI.2、LecRK-VI.3和LecRK-VI.4突变后都会微弱增加ABA对种子萌发的抑制作用.相对于LecRK- VI.2单基因突变, LecRK- VI.2和LecRK-VI.3双基因突变时ABA对种子萌发的抑制作用更显著.基因表达谱和网络调控分析表明, LecRK-VI.2和LecRK-VI.3基因还可调控ABA相关基因的表达(Xin et al., 2009).Zhang等(2019)通过对拟南芥凝集素类受体蛋白激酶基因LecRK-VI.4进行深入研究后, 发现该基因在ABA介导的气孔开闭中行使重要功能.(2) 参与茉莉酸(JA)信号响应.野生烟草中LecRK1基因突变会抑制由JA引起的防御途径(Bonaventure, 2011; Gilardoni et al., 2011).(3) 参与水杨酸(SA)信号响应.在拟南芥中过表达LecRK-IX.2基因导致SA积累, 从而引起细胞死亡(Luo et al., 2017). ...
... 基因还可调控ABA相关基因的表达(Xin et al., 2009).Zhang等(2019)通过对拟南芥凝集素类受体蛋白激酶基因LecRK-VI.4进行深入研究后, 发现该基因在ABA介导的气孔开闭中行使重要功能.(2) 参与茉莉酸(JA)信号响应.野生烟草中LecRK1基因突变会抑制由JA引起的防御途径(Bonaventure, 2011; Gilardoni et al., 2011).(3) 参与水杨酸(SA)信号响应.在拟南芥中过表达LecRK-IX.2基因导致SA积累, 从而引起细胞死亡(Luo et al., 2017). ...
A draft sequence of the rice genome (Oryza sativa L. ssp. indica) 1 2002
... ZmPK1是第1个被鉴定到的类受体蛋白激酶, 它属于G型凝集素类受体蛋白激酶.该激酶的细胞外结构域与芸薹属自交不亲和性位点编码的糖蛋白类似, 参与调控花粉与柱头之间的识别(Walker and Zhang, 1990).Hervé等(1996)在拟南芥中鉴定到第1个L型凝集素类受体蛋白激酶Ath.LecRK1, 该蛋白激酶具有豆类类凝集素结构, 可能在植物低聚糖和激素信号转导过程中发挥作用(Newman et al., 1994).根据LecRLKs细胞外凝集素结构域的特点, LecRLKs被归类为RLKs的一个亚家族(Shiu and Bleecker, 2001; Goff et al., 2002; Yu et al., 2002).虽然LecRLKs基因发现较早, 但对这些基因家族的系统分类却相对较晚, 而早期对于LecRLKs基因的研究也主要集中在基因表达等较为初步的层面(Nishiguchi et al., 2002; Riou et al., 2002; Navarro-Gochicoa et al., 2003; He et al., 2004; Zuo et al., 2004).近年来, 越来越多的研究表明, LecRLKs蛋白的功能极其广泛, 可参与调控各种生物/非生物胁迫和植物发育(Chen et al., 2006; Kanzaki et al., 2008; Joshi et al., 2010; Bonaventure, 2011; Desclos-Theveniau et al., 2012; Gouhier-Darimont et al., 2013; Li et al., 2014; Ranf et al., 2015; Luo et al., 2017; Ma et al., 2018; Labbé et al., 2019) (表1). ...
Quantitative phosphoproteomics of lectin receptor-like kinase VI.4 dependent abscisic acid response in Arabidopsis thaliana 1 2019
... 凝集素类受体蛋白激酶参与激素信号响应.(1)参与脱落酸(ABA)信号响应.在拟南芥中, 过表达凝集素类受体蛋白激酶基因LecRK-V.5可抑制植物对ABA的响应能力, 从而影响气孔开闭(Desclos-Theveniau et al., 2012).Xin等(2009)发现, L型凝集素类受体蛋白激酶VI亚家族成员基因LecRK-VI.2、LecRK-VI.3和LecRK-VI.4突变后都会微弱增加ABA对种子萌发的抑制作用.相对于LecRK- VI.2单基因突变, LecRK- VI.2和LecRK-VI.3双基因突变时ABA对种子萌发的抑制作用更显著.基因表达谱和网络调控分析表明, LecRK-VI.2和LecRK-VI.3基因还可调控ABA相关基因的表达(Xin et al., 2009).Zhang等(2019)通过对拟南芥凝集素类受体蛋白激酶基因LecRK-VI.4进行深入研究后, 发现该基因在ABA介导的气孔开闭中行使重要功能.(2) 参与茉莉酸(JA)信号响应.野生烟草中LecRK1基因突变会抑制由JA引起的防御途径(Bonaventure, 2011; Gilardoni et al., 2011).(3) 参与水杨酸(SA)信号响应.在拟南芥中过表达LecRK-IX.2基因导致SA积累, 从而引起细胞死亡(Luo et al., 2017). ...
Molecular cloning and characterization of GhLecRK, a novel kinase gene with lectin-like domain from Gossypium hirsutum 2 2004
... ZmPK1是第1个被鉴定到的类受体蛋白激酶, 它属于G型凝集素类受体蛋白激酶.该激酶的细胞外结构域与芸薹属自交不亲和性位点编码的糖蛋白类似, 参与调控花粉与柱头之间的识别(Walker and Zhang, 1990).Hervé等(1996)在拟南芥中鉴定到第1个L型凝集素类受体蛋白激酶Ath.LecRK1, 该蛋白激酶具有豆类类凝集素结构, 可能在植物低聚糖和激素信号转导过程中发挥作用(Newman et al., 1994).根据LecRLKs细胞外凝集素结构域的特点, LecRLKs被归类为RLKs的一个亚家族(Shiu and Bleecker, 2001; Goff et al., 2002; Yu et al., 2002).虽然LecRLKs基因发现较早, 但对这些基因家族的系统分类却相对较晚, 而早期对于LecRLKs基因的研究也主要集中在基因表达等较为初步的层面(Nishiguchi et al., 2002; Riou et al., 2002; Navarro-Gochicoa et al., 2003; He et al., 2004; Zuo et al., 2004).近年来, 越来越多的研究表明, LecRLKs蛋白的功能极其广泛, 可参与调控各种生物/非生物胁迫和植物发育(Chen et al., 2006; Kanzaki et al., 2008; Joshi et al., 2010; Bonaventure, 2011; Desclos-Theveniau et al., 2012; Gouhier-Darimont et al., 2013; Li et al., 2014; Ranf et al., 2015; Luo et al., 2017; Ma et al., 2018; Labbé et al., 2019) (表1). ...
... 在拟南芥中, LecRK-V.5基因在衰老组织和受伤的细胞周围大量表达, 推测其可能通过调控细胞分裂来修复受损细胞, 表明LecRK-V.5基因可能在植物发育过程中发挥重要功能(Riou et al., 2002).棉花(Gossypium hirsutum) GhLecRK基因在嫩枝和发育的纤维中大量表达, 对于纤维的发育至关重要(Zuo et al., 2004). ...
 ,1,*
,1,*
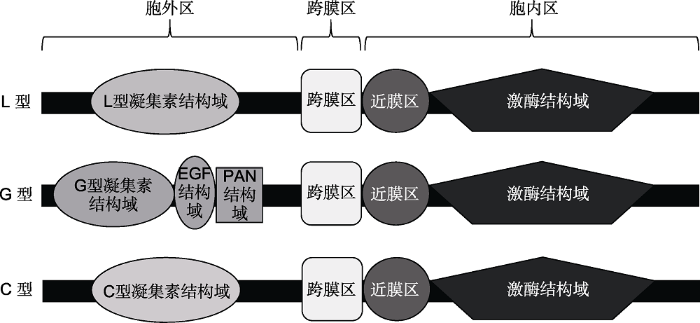 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT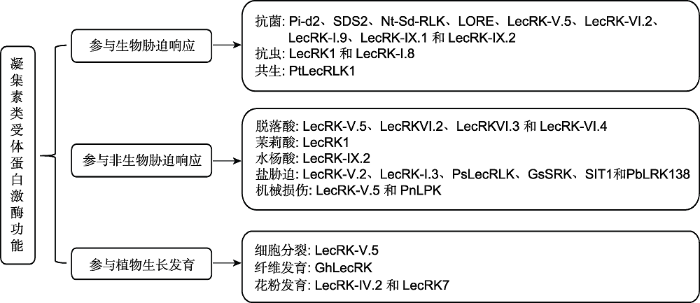 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}