The Dynamic Regulation Mechanism of the Endo-genous ABA in Plant
Wei Chen, Xiaoxian Zeng, Chuping Xie, Chang’en Tian, Yuping Zhou,*Guangzhou Key Laboratory for Functional Study on Plant Stress-resistant Genes, School of Life Sciences, Guangzhou University, Guangzhou 510006, China
Abstract Abscisic acid (ABA) plays important roles in regulating plant growth and development, and in responding rapidly to various environmental stimuli. The endogenous ABA level in plants is regulated sophisticatedly by the ABA biosynthesis, catabolism, and transportation pathways. This paper reviewed the most recent advancements in ABA de novo biosynthesis, ABA hydroxylation catabolism, reversible glycosylation metabolism, and ABA transportation pathway in plants, with emphasis on the expression regulation mechanism of the ABA biosynthetic and catabolic genes. Prospectives for research directions in the future were also suggested. Keywords:abscisic acid;ABA biosynthesis;ABA metabolism;ABA transportation
PDF (1176KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 陈唯, 曾晓贤, 谢楚萍, 田长恩, 周玉萍. 植物内源ABA水平的动态调控机制. 植物学报, 2019, 54(6): 677-687 doi:10.11983/CBB19092 Chen Wei, Zeng Xiaoxian, Xie Chuping, Tian Chang’en, Zhou Yuping. The Dynamic Regulation Mechanism of the Endo-genous ABA in Plant. Chinese Bulletin of Botany, 2019, 54(6): 677-687 doi:10.11983/CBB19092
植物激素脱落酸(abscisic acid, ABA)能调节种子成熟、休眠与萌发、气孔运动及叶片衰老等重要生理过程, 并介导植物对环境胁迫的快速反应。植物特定组织中内源ABA水平受ABA从头合成、羟基化代谢、糖基化失活和去糖基活化以及ABA长距离转运等的动态调控(魏开发等, 2012; 伍静辉等, 2018; Ma et al., 2018) (图1A)。近20年来, 植物ABA合成、代谢和信号转导研究取得了重要进展, 但ABA合成与代谢基因的启动和表达调控机制的研究却相对滞后。本文依据近年来植物内源ABA水平的动态调控机制的相关研究, 综述了植物ABA从头合成、羟基化与糖基化代谢以及ABA转运等领域的最新研究成果, 着重讨论ABA合成与代谢基因的表达调控机制(图1B), 并展望了今后的研究方向, 旨在为植物抗逆研究提供理论参考。
(A) ABA合成、代谢和转运途径; (B) ABA合成与代谢基因的表达调控。椭圆表示转录因子, 矩形表示调节蛋白。 Figure 1The ABA biosynthesis, catabolism and transportation pathway and the expression regulation of the ABA biosynthetic and catabolic genes in plant
(A) The ABA biosynthesis, catabolism and transportation pathway; (B) The expression regulation of the ABA biosynthetic and catabolic genes. The ellipses represent transcription factors, the rectangles represent regulatory proteins.
1 植物ABA从头合成的调控机制
1.1 ABA合成酶基因表达谱及突变体表型
高等植物ABA从头(de novo)合成的第1步是玉米黄质(zeaxanthin)被玉米黄质环氧酶(zeaxanthin epoxidase, ZEP)催化形成环氧玉米黄质(antheraxanthin), 继而环氧化为紫黄质(violaxanthin), 紫黄质异构化为9′-顺式-紫黄质(9′-cis-violaxanthin), 或被新黄素合成酶(neoxanthin synthase, ABA4)催化为反式-新黄质(trans-neoxanthin), 进而异构化为9′-顺式-新黄质(9′-cis-neoxanthin) (Nambara and Marion- Poll, 2005)。自1996年以来相继从烟草(Nicotiana plumbaginifolia) (NpABA2)、拟南芥(Arabidopsis thaliana) (AtABA1/LOS6)、水稻(Oryza sativa) (OsABA1)、番茄(Solanum lycopersicum) (SlHP3)和长豇豆(Vigna unguiculata) (VuABA1)中克隆了ZEP基因, 不同物种的ZEP功能缺失突变体都表现为内源ABA含量降低、种子不休眠、植株呈萎蔫状及干旱不能诱导ABA合成等特征(Endo et al., 2014)。拟南芥aba1突变体株型矮小, 叶片形状和内部结构畸形; 而ABA1过表达植株的抗盐和抗旱性增强, 在光照条件下气孔开度小于野生型, 说明ABA具有调节植物生长发育和抗逆性的重要功能(Barrero et al., 2005; Park et al., 2008)。植物ABA4蛋白高度保守, 定位于叶绿体内囊体膜。拟南芥aba4突变体仅营养组织与野生型有差异, 脱水不能诱导合成ABA, 推测ABA4参与脱水胁迫下的ABA合成(North et al., 2007)。
拟南芥AtCYP707As转录本广泛存在于各种器官组织, 但表达量有所不同。例如, 花和角果中AtCYP707A1表达量最高, AtCYP707A2和AtCYP707A3在叶、茎和根中表达较高, 但AtCYP707A4在所有组织中表达都较低, 种子吸胀增强AtCYP707A2的表达, 干旱抑制而复水促进所有AtCYP707As表达(Kushiro et al., 2004)。AtCYP707A1主要调节成熟中期种子ABA水平, 而AtCYP707A2调控成熟晚期至萌发阶段的ABA代谢(Okamoto et al., 2006)。AtCYP707A3是主要的干旱应答基因, 正常生长环境中的cyp707a3突变体内源ABA含量高于野生型、叶片蒸腾速率降低并对外源ABA敏感; 干旱促进突变体积累更多ABA, 植株耐旱性显著增强, 但过表达植株表型相反(Umezawa et al., 2006)。在正常生长环境中, cyp707a1和cyp707a3突变体叶片的气孔导度都较低; 高湿环境显著增强AtCYP707A1/3表达、促进ABA降解, cyp707a3突变体可以积累更高水平的ABA, 然而cyp707a1突变体的ABA水平与野生型相似。研究表明, 在高湿环境中AtCYP707A1使保卫细胞内的ABA失活, AtCYP707A3则主要降解维管组织中移动的ABA, 植物气孔运动受到来自维管组织的ABA和保卫细胞内的ABA代谢共同调控(Okamoto et al., 2009)。
2.2 ABA代谢酶基因的表达调控
已有研究表明若干转录因子参与ABA代谢基因的表达调控。拟南芥CHO1 (CHOTTO1)为AP2/ERF型转录因子, 促进AtABA1和AtNCED9、抑制AtCYP707A2/3转录, 调控吸胀期种子ABA抑制的赤霉素合成(Yano et al., 2009)。AP2/ERF型转录因子ABI4促进ABA积累和种子休眠, 通过结合AtCYP707A1/2启动子并抑制其转录来实现(Shu et al., 2013)。拟南芥NLP8 (NIN-like protein 8)以硝酸盐依赖的方式结合AtCYP707A2的启动子并激活其转录, 降低种子ABA水平, 促进种子萌发(Matakiadis et al., 2009; Yan et al., 2016)。bZIP家族转录因子VIP1 (VirE2-interacting protein 1)为渗透胁迫的调节因子, VIP1直接结合AtCYP707A1/3启动子并激活其转录(Tsugama et al., 2012)。拟南芥bHLH122是干旱、NaCl和渗透胁迫的正调节因子, bHLH122结合AtCYP707A3启动子G-/E-box元件并抑制其表达, 进而抑制ABA代谢(Liu et al., 2014)。拟南芥成花抑制因子SVP (short vegetative phase)直接结合AtCYP707A1/3和AtBG1启动子的CArG Motif元件, 抑制AtCYP707A1/3, 促进AtBG1转录, 表明SVP是干旱胁迫下ABA代谢的重要调控因子(Wang et al., 2018)。
3 植物ABA可逆糖基化代谢的表达调控
内源ABA水平还受到ABA特异的葡萄糖基转移酶(ABA-uridine diphosphate glucosyltransferase, ABAUGT)和β-葡萄糖苷酶(β-glucosidase, BG)介导的可逆糖基化代谢调节。研究人员从赤豆(V. angularis)中克隆了编码ABA-UGT酶的基因AOG, UGT可催化ABA与UDPG结合生成ABA葡萄糖苷酯(ABA-glucoside ester, ABA-GE); ABA-GE缺乏生物学活性, 在液泡或质外体中积累(Xu et al., 2002; Priest et al., 2006)。拟南芥UGT家族的E亚家族中UGT71B6/7/8参与调节内源ABA水平, 影响AtCYP707A1-4表达, 在脱水和高盐胁迫以及种子萌发和生长发育进程中发挥重要调节作用(Dong et al., 2014)。此外, AtUGT71C5也可催化ABA转化为ABA-GE, 与野生型相比, ugt71c5突变体的种子萌发延迟且叶片气孔变小, 抗旱性增强(Liu et al., 2015)。
干旱条件下, 液泡或质外体贮存的ABA-GE迅速被BG水解, 生成活性ABA以适应变化的环境。拟南芥AtBG1在内质网将ABA-GE水解生成ABA, bg1突变体内源ABA含量和抗旱性降低; 脱水迅速诱导BG1蛋白多聚化后显著增强BG活性, 从而加速ABA生成, ABA正反馈激活BG1蛋白多聚化(Lee et al., 2006)。AtBG2以复合物形式储存在中央液泡, 脱水诱导AtBG2表达, bg2突变体对脱水和盐胁迫敏感, AtBG2过表达可互补bg1表型(Xu et al., 2012)。在低湿环境下, 拟南芥aba3-1突变体中AtBG1转录水平极显著高于野生型, 说明细胞内ABA-GE水解机制可部分补偿ABA从头合成的缺陷(Bauer et al., 2013)。此外, AtBGLU10编码的BG酶分布在液泡中, bglu10突变体的BG酶活性和ABA含量均降低, 且叶面温度和胁迫反应基因表达量也降低, 而AtBGLU10过表达株系抗旱性增强(Wang et al., 2011a)。
4 植物ABA转运的表达调控
4.1 ABA合成位点和作用位点
ABA从头合成途径的最后步骤由SDR1和AAO酶催化, 因此, 检测SDR1和AAO蛋白定位就可确定ABA合成位点。GFP-和免疫荧光显示, 拟南芥AAO3在根尖、韧皮部伴胞细胞和木质部薄壁细胞中含量最高, 保卫细胞也有少量蛋白(Koiwai et al., 2004)。免疫组化也发现叶脉薄壁细胞含有大量的AAO3和ABA2蛋白, 失水可使薄壁细胞内NCED3蛋白显著增加, 证明干旱胁迫下NCED3酶是ABA合成的关键酶(Endo et al., 2008)。GUS染色证明NCED3基因在叶肉细胞、保卫细胞、维管束及根系中都有表达, 在根尖表达量最高, 证明NCED3酶促进根源ABA合成, 但保卫细胞ABA合成量低于其它组织(Tan et al., 2003)。进一步研究表明, 保卫细胞ABA合成受到维管细胞合成的ABA正反馈调节, 低湿环境下保卫细胞合成的ABA是诱导气孔关闭充分且必要因子(Bauer et al., 2013)。种子内源ABA分别来源于母体维管组织、胚和胚乳细胞(Lefebvre et al., 2006)。总之, 植物ABA合成位点包括根尖、维管束伴胞细胞和薄壁细胞、叶肉细胞、保卫细胞、种胚和胚乳细胞。
BarreroJM, PiquerasP, González-GuzmánM, SerranoR, RodríguezPL, PonceMR, MicolJL ( 2005). A mutational analysis of the ABA1 gene of Arabidopsis thaliana highlights the involvement of ABA in vegetative development J Exp Bot 56, 2071-2083. [本文引用: 1]
BoursiacY, LeranS, Corratgé-FaillieC, GojonA, KroukG, LacombeB ( 2013). ABA transport and transporters Trends Plant Sci 18, 325-333. [本文引用: 1]
CaoMJ, WangZ, ZhaoQ, MaoJL, SpeiserA, WirtzM, HellR, ZhuJK, XiangCB ( 2014). Sulfate availability affects ABA levels and germination response to ABA and salt stress in Arabidopsis thaliana Plant J 77, 604-615. [本文引用: 1]
ChenHY, HsiehEJ, ChengMC, ChenCY, HwangSY, LinTP ( 2016). ORA47 (octadecanoid-responsive AP2/ERF- domain transcription factor 47) regulates jasmonic acid and abscisic acid biosynthesis and signaling through binding to a novel cis-element New Phytol 211, 599-613. [本文引用: 1]
CuiP, ZhangS, DingF, AliS, XiongL ( 2014). Dynamic regulation of genome-wide pre-mRNA splicing and stress tolerance by the Sm-like protein LSm5 in Arabidopsis Genome Biol 15, R1. [本文引用: 1]
DongT, XuZY, ParkY, KimDH, LeeY, HwangI ( 2014). Abscisic acid uridine diphosphate glucosyltransferases play a crucial role in abscisic acid homeostasis in Arabidopsis Plant Physiol 165, 277-289. [本文引用: 1]
EndoA, OkamotoM, KoshibaT (2014). ABA biosynthetic and catabolic pathways In: Zhang DP, ed. Abscisic Acid: Metabolism, Transport and Signaling. Dordrecht: Springer Press. pp. 21-45. [本文引用: 4]
FreyA, EffroyD, LefebvreV, SeoM, PerreauF, BergerA, SechetJ, ToA, NorthHM, Marion-PollA ( 2012). Epoxycarotenoid cleavage by NCED5 fine-tunes ABA accumulation and affects seed dormancy and drought tolerance with other NCED family members Plant J 70, 501-512. [本文引用: 1]
HaoGP, ZhangXH, WangYQ, WuZY, HuangCL ( 2009). Nucleotide variation in the NCED3 region of Arabidopsis thaliana and its association study with abscisic acid content under drought stress J Integr Plant Biol 51, 175-183. [本文引用: 1]
HarrisonE, BurbidgeA, OkyereJP, ThompsonAJ, TaylorIB ( 2011). Identification of the tomato ABA-deficient mutant sitiens as a member of the ABA-aldehyde oxidase gene family using genetic and genomic analysis Plant Growth Regul 64, 301-309. [本文引用: 1]
JeJ, ChenH, SongC, LimCO ( 2014). Arabidopsis DREB2C modulates ABA biosynthesis during germination Biochem Biophys Res Commun 452, 91-98. [本文引用: 1]
KannoY, HanadaA, ChibaY, IchikawaT, NakazawaM, MatsuiM, KoshibaT, KamiyaY, SeoM ( 2012). Identification of an abscisic acid transporter by functional screening using the receptor complex as a sensor Proc Natl Acad Sci USA 109, 9653-9658. [本文引用: 1]
KoJH, YangSH, HanKH ( 2006). Upregulation of an Arabidopsis RING-H2 gene, XERICO, confers drought tolerance through increased abscisic acid biosynthesis Plant J 47, 343-355. [本文引用: 1]
KoiwaiH, NakaminamiK, SeoM, MitsuhashiW, ToyomasuT, KoshibaT ( 2004). Tissue-specific localization of an abscisic acid biosynthetic enzyme, AAO3, in Arabidopsis Plant Physiol 134, 1697-1707. [本文引用: 1]
KuromoriT, FujitaM, UranoK, TanabataT, SugimotoE, ShinozakiK ( 2016). Overexpression of AtABCG25 enhances the abscisic acid signal in guard cells and improves plant water use efficiency Plant Sci 251, 75-81. [本文引用: 1]
KuromoriT, MiyajiT, YabuuchiH, ShimizuH, SugimotoE, KamiyaA, MoriyamaY, ShinozakiK ( 2010). ABC transporter AtABCG25 is involved in abscisic acid transport and responses Proc Natl Acad Sci USA 107, 2361-2366. [本文引用: 1]
KuromoriT, SeoM, ShinozakiK ( 2018). ABA transport and plant water stress responses Trends Plant Sci 23, 513-522. [本文引用: 2]
KuromoriT, SugimotoE, ShinozakiK ( 2011). Arabidopsis mutants of AtABCG22, an ABC transporter gene, increase water transpiration and drought susceptibility Plant J 67, 885-894. [本文引用: 1]
KushiroT, OkamotoM, NakabayashiK, YamagishiK, KitamuraS, AsamiT, HiraiN, KoshibaT, KamiyaY, NambaraE ( 2004). The Arabidopsis cytochrome P450 CYP707A encodes ABA 8′-hydroxylases: key enzymes in ABA catabolism EMBO J 23, 1647-1656. [本文引用: 1]
Le HirR, CastelainM, ChakrabortiD, MoritzT, DinantS, BelliniC ( 2017). At bHLH68 transcription factor contributes to the regulation of ABA homeostasis and drought stress tolerance in Arabidopsis thaliana Physiol Plant 160, 312-327. [本文引用: 1]
LeeHG, LeeK, SeoPJ ( 2015). The Arabidopsis MYB96 transcription factor plays a role in seed dormancy Plant Mol Biol 87, 371-381. [本文引用: 1]
LeeKH, PiaoHL, KimHY, ChoiSM, JiangF, HartungW, HwangI, KwakJM, LeeIJ, HwangI ( 2006). Activation of glucosidase via stress-induced polymerization rapidly increases active pools of abscisic acid Cell 126, 1109-1120. [本文引用: 1]
LefebvreV, NorthH, FreyA, SottaB, SeoM, OkamotoM, NambaraE, Marion-PollA ( 2006). Functional analysis of Arabidopsis NCED6 and NCED9 genes indicates that ABA synthesized in the endosperm is involved in the induction of seed dormancy Plant J 45, 309-319. [本文引用: 2]
LiW, De OllasC, DoddIC ( 2018). Long-distance ABA transport can mediate distal tissue responses by affecting local ABA concentrations J Integr Plant Biol 60, 16-33. [本文引用: 1]
LinPC, HwangSG, EndoA, OkamotoM, KoshibaT, ChengWH ( 2007). Ectopic expression of ABSCISIC ACID 2/GLUCOSE INSENSITIVE 1 in Arabidopsis promotes seed dormancy and stress tolerance Plant Physiol 143, 745-758. [本文引用: 1]
LissoJ, Schr?derF, FisahnJ, MüssigC ( 2011). NFX1-LIKE2 (NFXL2) suppresses abscisic acid accumulation and stomatal closure in Arabidopsis thaliana PLoS One 6, e26982. [本文引用: 1]
LiuS, LiM, SuL, GeK, LiL, LiX, LiuX, LiL ( 2016). Negative feedback regulation of ABA biosynthesis in peanut (Arachis hypogaea): a transcription factor complex inhibits AhNCED1 expression during water stress Sci Rep 6, 37943. [本文引用: 1]
LiuW, TaiH, LiS, GaoW, ZhaoM, XieC, LiWX ( 2014). bHLH122 is important for drought and osmotic stress resistance in Arabidopsis and in the repression of ABA catabolism New Phytol 201, 1192-1204. [本文引用: 1]
LiuZ, YanJP, LiDK, LuoQ, YanQ, LiuZB, YeLM, WangJM, LiXF, YangY ( 2015). UDP-glucosyltransferase71C5, a major glucosyltransferase, mediates abscisic acid homeostasis in Arabidopsis Plant Physiol 167, 1659-1670. [本文引用: 1]
MaYL, CaoJ, HeJH, ChenQQ, LiXF, YangY ( 2018). Molecular mechanism for the regulation of ABA homeostasis during plant development and stress responses Int J Mol Sci 19, 3643. [本文引用: 1]
MalcheskaF, AhmadA, BatoolS, MüllerHM, Ludwig- MüllerJ, KreuzwieserJ, RandewigD, H?nschR, MendelRR, HellR, WirtzM, GeigerD, AcheP, HedrichR, HerschbachC, RennenbergH ( 2017). Drought-enhanced xylem sap sulfate closes stomata by affecting ALMT12 and guard cell ABA synthesis Plant Physiol 174, 798-814. [本文引用: 1]
MatakiadisT, AlboresiA, JikumaruY, TatematsuK, PichonO, RenouJP, KamiyaY, NambaraE, TruongHN ( 2009). The Arabidopsis abscisic acid catabolic gene CYP707A2 plays a key role in nitrate control of seed dormancy Plant Physiol 149, 949-960. [本文引用: 1]
NambaraE, Marion-PollA ( 2005). Abscisic acid biosynthesis and catabolism Annu Rev Plant Biol 56, 165-185. [本文引用: 1]
NorthHM, De AlmeidaA, BoutinJP, FreyA, ToA, BotranL, SottaB, Marion-PollA ( 2007). The Arabidopsis ABA- deficient mutant aba4 demonstrates that the major route for stress-induced ABA accumulation is via neoxanthin isomers Plant J 50, 810-824. [本文引用: 1]
OkamotoM, KuwaharaA, SeoM, KushiroT, AsamiT, HiraiN, KamiyaY, KoshibaT, NambaraE ( 2006). CYP707A1 and CYP707A2, which encode abscisic acid 8′-hydroxylases, are indispensable for proper control of seed dormancy and germination in Arabidopsis Plant Physiol 141, 97-107. [本文引用: 1]
OkamotoM, TanakaY, AbramsSR, KamiyaY, SekiM, NambaraE ( 2009). High humidity induces abscisic acid 8′-hydroxylase in stomata and vasculature to regulate local and systemic abscisic acid responses in Arabidopsis Plant Physiol 149, 825-834. [本文引用: 1]
ParkHY, SeokHY, ParkBK, KimSH, GohCH, LeeBH, LeeCH, MoonYH ( 2008). Overexpression of Arabidopsis ZEP enhances tolerance to osmotic stress Biochem Biophys Res Commun 375, 80-85. [本文引用: 1]
ParkY, XuZY, KimSY, LeeJ, ChoiB, LeeJ, KimH, SimHJ, HwangI ( 2016). Spatial regulation of ABCG25, an ABA exporter, is an important component of the mechanism controlling cellular ABA levels Plant Cell 28, 2528-2544. [本文引用: 1]
PriestDM, AmbroseSJ, VaistijFE, EliasL, HigginsGS, RossARS, AbramsS, BowlesDJ ( 2006). Use of the glucosyltransferase UGT71B6 to disturb abscisic acid homeostasis in Arabidopsis thaliana Plant J 46, 492-502. [本文引用: 1]
RenT, WangJ, ZhaoM, GongX, WangS, WangG, ZhouC ( 2018). Involvement of NAC transcription factor SiNAC1 in a positive feedback loop via ABA biosynthesis and leaf senescence in foxtail millet Planta 247, 53-68. [本文引用: 1]
SatoH, TakasakiH, TakahashiF, SuzukiT, IuchiS, MitsudaN, Ohme-TakagiM, IkedaM, SeoM, Yamaguchi-ShinozakiK, ShinozakiK ( 2018). Arabidopsis thaliana NGATHA1 transcription factor induces ABA biosynthesis by activating NCED3 gene during dehydration stress Proc Natl Acad Sci USA 115, E11178-E11187. [本文引用: 1]
ScholzSS, ReicheltM, VadasseryJ, Mith?ferA ( 2015). Calmodulin-like protein CML37 is a positive regulator of ABA during drought stress in Arabidopsis Plant Signal Behav 10, e1011951. [本文引用: 1]
SchwartzSH, TanBC, GageDA, ZeevaartJAD, McCartyDR ( 1997). Specific oxidative cleavage of carotenoids by VP14 of maize Science 276, 1872-1874. [本文引用: 1]
SeoM, AokiK, KoiwaiH, KamiyaY, NambaraE, KoshibaT ( 2004). Comparative studies on the Arabidopsis aldehyde oxidase (AAO) gene family revealed a major role of AAO3 in ABA biosynthesis in seeds Plant Cell Physiol 45, 1694-1703. [本文引用: 1]
ShuK, ZhangH, WangS, ChenM, WuY, TangS, LiuC, FengY, CaoX, XieQ ( 2013). ABI4 regulates primary seed dormancy by regulating the biogenesis of abscisic acid and gibberellins in Arabidopsis PLoS Genet 9, e1003577. [本文引用: 1]
SussmilchFC, BrodribbTJ, McadamSAM ( 2017). Up-regulation of NCED3 and ABA biosynthesis occur within minutes of a decrease in leaf turgor but AHK1 is not required J Exp Bot 68, 2913-2918. [本文引用: 1]
TakahashiF, SuzukiT, OsakabeY, BetsuyakuS, KondoY, DohmaeN, FukudaH, Yamaguchi-ShinozakiK, ShinozakiK ( 2018). A small peptide modulates stomatal control via abscisic acid in long-distance signaling Nature 556, 235-238. [本文引用: 1]
TanBC, JosephLM, DengWT, LiuLJ, LiQB, ClineK, McCartyDR ( 2003). Molecular characterization of the Arabidopsis 9-cis epoxycarotenoid dioxygenase gene family Plant J 35, 44-56. [本文引用: 2]
TanBC, SchwartzSH, ZeevaartJAD, MccartyDR ( 1997). Genetic control of abscisic acid biosynthesis in maize Proc Natl Acad Sci USA 94, 12235-12240. [本文引用: 1]
TanWR, ZhangDW, ZhouHP, ZhengT, YinYH, LinHH ( 2018). Transcription factor HAT1 is a substrate of SnRK2.3 kinase and negatively regulates ABA synthesis and signaling in Arabidopsis responding to drought PLoS Genet 14, e1007336. [本文引用: 1]
TsugamaD, LiuSK, TakanoT ( 2012). A bZIP protein, VIP1, is a regulator of osmosensory signaling in Arabidopsis Plant Physiol 159, 144-155. [本文引用: 1]
UmezawaT, OkamotoM, KushiroT, NambaraE, OonoY, SekiM, KobayashiM, KoshibaT, KamiyaY, ShinozakiK ( 2006). CYP707A3, a major ABA 80-hydroxylase involved in dehydration and rehydration response in Arabido- psis thaliana Plant J 46, 171-182. [本文引用: 1]
VallabhaneniR, WurtzelET ( 2010). From epoxycarotenoids to ABA: the role of ABA 8′-hydroxylases in drought- stressed maize roots Arch Biochem Biophys 504, 112-117. [本文引用: 1]
WangPT, LiuH, HuaHJ, WangL, SongCP ( 2011a). A vacuole localized β-glucosidase contributes to drought tolerance in Arabidopsis Chin Sci Bull 56, 3538-3546. [本文引用: 1]
WangZ, WangFX, HongYC, YaoJJ, RenZZ, ShiHZ, ZhuJK ( 2018). The flowering repressor SVP confers drought resistance in Arabidopsis by regulating abscisic acid catabolism Mol Plant 11, 1184-1197. [本文引用: 1]
WangZY, XiongL, LiW, ZhuJK, ZhuJ ( 2011b). The plant cuticle is required for osmotic stress regulation of abscisic acid biosynthesis and osmotic stress tolerance in Arabidopsis Plant Cell 23, 1971-1984. [本文引用: 1]
XiongL, GongZ, RockCD, SubramanianS, GuoY, XuW, GalbraithD, ZhuJK ( 2001). Modulation of abscisic acid signal transduction and biosynthesis by an smlike protein in Arabidopsis Cell 1, 771-781. [本文引用: 2]
XuZJ, NakajimaM, SuzukiY, YamaguchiI ( 2002). Cloning and characterization of the abscisic acid-specific glucosyltransferase gene from adzuki bean seedlings Plant Physiol 129, 1285-1295. [本文引用: 1]
XuZY, LeeKH, DongT, JeongJC, JinJB, KannoY, KimDH, KimSY, SeoM, BressanRA, YunDJ, HwangI ( 2012). A vacuolar β-glucosidase homolog that possesses glucose-conjugated abscisic acid hydrolyzing activity plays an important role in osmotic stress responses in Arabidopsis Plant Cell 24, 2184-2199. [本文引用: 1]
YanD, EaswaranV, ChauV, OkamotoM, IerulloM, KimuraM, EndoA, YanoR, PashaA, GongY, BiYM, ProvartN, GuttmanD, KrappA, RothsteinSJ, NambaraE ( 2016). NIN-like protein 8 is a master regulator of nitrate-promoted seed germination in Arabidopsis Nat Commun 7, 13179. [本文引用: 1]
YangJ, WorleyE, UdvardiM ( 2014). A NAP-AAO3 regulatory module promotes chlorophyll degradation via ABA biosynthesis in Arabidopsis leaves Plant Cell 26, 4862-4874. [本文引用: 1]
YanoR, KannoY, JikumaruY, NakabayashiK, KamiyaY, NambaraE ( 2009). CHOTTO1, a putative double APETALA2 repeat transcription factor, is involved in abscisic acid-mediated repression of gibberellin biosynthesis during seed germination in Arabidopsis Plant Physiol 151, 641-654. [本文引用: 1]
YueY, ZhangM, ZhangJ, TianX, DuanL, LiZ ( 2012). Overexpression of the AtLOS5 gene increased abscisic acid level and drought tolerance in transgenic cotton J Exp Bot 63, 3741-3748. [本文引用: 1]
Zdunek-ZastockaE, SobczakM ( 2013). Expression of Pisum sativum PsAO3 gene, which encodes an aldehyde oxidase utilizing abscisic aldehyde, is induced under progressively but not rapidly imposed drought stress Plant Physiol Biochem 71(2), 57-66. [本文引用: 1]
ZhangH, ZhuH, PanY, YuY, LuanS, LiL ( 2014). A DTX/MATE-type transporter facilitates abscisic acid efflux and modulates ABA sensitivity and drought tolerance in Arabidopsis Mol Plant 7, 1522-1532. [本文引用: 1]
ZhouYP, WuJH, XiaoWH, ChenW, ChenQH, FanT, XieCP, TianCE ( 2018). Arabidopsis IQM4, a novel calmodulin-binding protein, is involved with seed dormancy and germination in Arabidopsis Front Plant Sci 9, 721. [本文引用: 1]
ZhuG, YeN, ZhangJ ( 2009). Glucose-induced delay of seed germination in rice is mediated by the suppression of ABA catabolism rather than an enhancement of ABA biosynthesis Plant Cell Physiol 50, 644-651. [本文引用: 1]
ChenW, ZengXX, XieCP, TianCE, ZhouYP ( 2019). The dynamic regulation mechanism of the endogenous ABA in plant Chin Bull Bot 54, 677-687.
NCED3基因的持续诱导及ABA合成与代谢的协同调控在拟南芥ABA信号积累中的作用 1 2007
... Schwartz等(1997)证明重组的VP14蛋白可切割9′-顺式-紫黄质形成黄质醛, 推测VP14具有NCED功能.目前, 已证明植物NCED蛋白是一个多基因家族, 先后从水稻(OsNCED1-5)、拟南芥(AtNCED2/3/5/6/9)、四季豆(Phaseolus vulgaris) (PvNCED1)、鳄梨(Persea americana) (PaNCED1/3)以及菟丝子(Cuscuta reflexa) (CrNCED1/2)中克隆到NCED基因(Endo et al., 2014).水稻OsNCED1在叶片中表达量最高, OsNCED2主要在种子中表达, OsNCED5参与水稻籽粒灌浆期ABA的合成, OsNCED3转基因拟南芥ABA含量增加, 叶片形态发生改变(Zhu et al., 2009).拟南芥基因组有9个VP14同源序列(AtNCED1-9), 其中AtNCED2/3/5/6/9编码蛋白具有NCED酶活性, AtNCED2/3在植物根和叶中表达量较高.干旱迅速诱导AtNCED3基因表达并促进ABA合成, AtNCED3过表达株系的内源ABA水平较高、叶片蒸腾速率较低、抗旱性较强, 且ABA响应基因的转录本增加(任慧波等, 2007; Hao et al., 2009).种子成熟晚期AtNCED5/6/9表达显著增强, AtNCED6在胚乳中表达, AtNCED5/9在胚乳和胚中表达.nced6/nced9双突变体种子内源ABA含量和休眠程度显著降低, 说明AtNCED6/9主要调控种子休眠(Tan et al., 2003; Lefebvre et al., 2006).nced3/nced5双突变体内源ABA含量减少、营养器官变小且植株抗旱性降低, 说明AtNCED3/5参与植物生长发育和胁迫响应(Frey et al., 2012). ...
内源ABA信号水平动态调控的分子机制 1 2012
... 植物激素脱落酸(abscisic acid, ABA)能调节种子成熟、休眠与萌发、气孔运动及叶片衰老等重要生理过程, 并介导植物对环境胁迫的快速反应.植物特定组织中内源ABA水平受ABA从头合成、羟基化代谢、糖基化失活和去糖基活化以及ABA长距离转运等的动态调控(魏开发等, 2012; 伍静辉等, 2018; Ma et al., 2018) (图1A).近20年来, 植物ABA合成、代谢和信号转导研究取得了重要进展, 但ABA合成与代谢基因的启动和表达调控机制的研究却相对滞后.本文依据近年来植物内源ABA水平的动态调控机制的相关研究, 综述了植物ABA从头合成、羟基化与糖基化代谢以及ABA转运等领域的最新研究成果, 着重讨论ABA合成与代谢基因的表达调控机制(图1B), 并展望了今后的研究方向, 旨在为植物抗逆研究提供理论参考. ...
脱落酸调控种子休眠和萌发的分子机制 1 2018
... 植物激素脱落酸(abscisic acid, ABA)能调节种子成熟、休眠与萌发、气孔运动及叶片衰老等重要生理过程, 并介导植物对环境胁迫的快速反应.植物特定组织中内源ABA水平受ABA从头合成、羟基化代谢、糖基化失活和去糖基活化以及ABA长距离转运等的动态调控(魏开发等, 2012; 伍静辉等, 2018; Ma et al., 2018) (图1A).近20年来, 植物ABA合成、代谢和信号转导研究取得了重要进展, 但ABA合成与代谢基因的启动和表达调控机制的研究却相对滞后.本文依据近年来植物内源ABA水平的动态调控机制的相关研究, 综述了植物ABA从头合成、羟基化与糖基化代谢以及ABA转运等领域的最新研究成果, 着重讨论ABA合成与代谢基因的表达调控机制(图1B), 并展望了今后的研究方向, 旨在为植物抗逆研究提供理论参考. ...
A mutational analysis of the ABA1 gene of Arabidopsis thaliana highlights the involvement of ABA in vegetative development 1 2005
... 高等植物ABA从头(de novo)合成的第1步是玉米黄质(zeaxanthin)被玉米黄质环氧酶(zeaxanthin epoxidase, ZEP)催化形成环氧玉米黄质(antheraxanthin), 继而环氧化为紫黄质(violaxanthin), 紫黄质异构化为9′-顺式-紫黄质(9′-cis-violaxanthin), 或被新黄素合成酶(neoxanthin synthase, ABA4)催化为反式-新黄质(trans-neoxanthin), 进而异构化为9′-顺式-新黄质(9′-cis-neoxanthin) (Nambara and Marion- Poll, 2005).自1996年以来相继从烟草(Nicotiana plumbaginifolia) (NpABA2)、拟南芥(Arabidopsis thaliana) (AtABA1/LOS6)、水稻(Oryza sativa) (OsABA1)、番茄(Solanum lycopersicum) (SlHP3)和长豇豆(Vigna unguiculata) (VuABA1)中克隆了ZEP基因, 不同物种的ZEP功能缺失突变体都表现为内源ABA含量降低、种子不休眠、植株呈萎蔫状及干旱不能诱导ABA合成等特征(Endo et al., 2014).拟南芥aba1突变体株型矮小, 叶片形状和内部结构畸形; 而ABA1过表达植株的抗盐和抗旱性增强, 在光照条件下气孔开度小于野生型, 说明ABA具有调节植物生长发育和抗逆性的重要功能(Barrero et al., 2005; Park et al., 2008).植物ABA4蛋白高度保守, 定位于叶绿体内囊体膜.拟南芥aba4突变体仅营养组织与野生型有差异, 脱水不能诱导合成ABA, 推测ABA4参与脱水胁迫下的ABA合成(North et al., 2007). ...
The stomatal response to reduced relative humidity requires guard cell-autonomous ABA synthesis 2 2013
... 干旱条件下, 液泡或质外体贮存的ABA-GE迅速被BG水解, 生成活性ABA以适应变化的环境.拟南芥AtBG1在内质网将ABA-GE水解生成ABA, bg1突变体内源ABA含量和抗旱性降低; 脱水迅速诱导BG1蛋白多聚化后显著增强BG活性, 从而加速ABA生成, ABA正反馈激活BG1蛋白多聚化(Lee et al., 2006).AtBG2以复合物形式储存在中央液泡, 脱水诱导AtBG2表达, bg2突变体对脱水和盐胁迫敏感, AtBG2过表达可互补bg1表型(Xu et al., 2012).在低湿环境下, 拟南芥aba3-1突变体中AtBG1转录水平极显著高于野生型, 说明细胞内ABA-GE水解机制可部分补偿ABA从头合成的缺陷(Bauer et al., 2013).此外, AtBGLU10编码的BG酶分布在液泡中, bglu10突变体的BG酶活性和ABA含量均降低, 且叶面温度和胁迫反应基因表达量也降低, 而AtBGLU10过表达株系抗旱性增强(Wang et al., 2011a). ...
... ABA从头合成途径的最后步骤由SDR1和AAO酶催化, 因此, 检测SDR1和AAO蛋白定位就可确定ABA合成位点.GFP-和免疫荧光显示, 拟南芥AAO3在根尖、韧皮部伴胞细胞和木质部薄壁细胞中含量最高, 保卫细胞也有少量蛋白(Koiwai et al., 2004).免疫组化也发现叶脉薄壁细胞含有大量的AAO3和ABA2蛋白, 失水可使薄壁细胞内NCED3蛋白显著增加, 证明干旱胁迫下NCED3酶是ABA合成的关键酶(Endo et al., 2008).GUS染色证明NCED3基因在叶肉细胞、保卫细胞、维管束及根系中都有表达, 在根尖表达量最高, 证明NCED3酶促进根源ABA合成, 但保卫细胞ABA合成量低于其它组织(Tan et al., 2003).进一步研究表明, 保卫细胞ABA合成受到维管细胞合成的ABA正反馈调节, 低湿环境下保卫细胞合成的ABA是诱导气孔关闭充分且必要因子(Bauer et al., 2013).种子内源ABA分别来源于母体维管组织、胚和胚乳细胞(Lefebvre et al., 2006).总之, 植物ABA合成位点包括根尖、维管束伴胞细胞和薄壁细胞、叶肉细胞、保卫细胞、种胚和胚乳细胞. ...
Sulfate availability affects ABA levels and germination response to ABA and salt stress in Arabidopsis thaliana 1 2014
... 此外, 硫元素供应、角质层合成基因、钙调素信号蛋白也参与ABA生物合成的调控.叶绿体硫转运蛋白AtSULTR3;1功能缺失显著降低幼苗AAO酶活性和内源ABA水平, 硫元素供应可提高AtNCED3和AtABA3转录水平; ABA促进AtSULTR3;1和其它硫代谢基因转录, 表明硫在植物响应环境胁迫时发挥重要作用(Cao et al., 2014).干旱使木质部汁液硫酸盐含量增加, 进而促进保卫细胞AtNCED3表达和ABA合成, 引起气孔关闭(Malcheska et al., 2017).已知植物角质层作为物理屏障减少水分流失并介导ABA合成和信号转导, 进而调控植物渗透胁迫反应.拟南芥ced1 (9-cis-epoxycarotenoid dioxygenase defective 1)突变体对轻微渗透胁迫极为敏感, CED1/BDG1基因促进ABA合成基因的表达(Wang et al., 2011b).植物钙调素类似蛋白AtCML37 (calmodulin-like 37)和AtCML42拮抗调控干旱胁迫诱导的ABA积累, AtCML37为正调节因子, 而AtCML42为负调节因子(Scholz et al., 2015).我们也发现钙调素结合蛋白AtIQM4能促进种子ABA合成, iqm4突变体种子ABA含量降低, 角果AtNCED6/9表达量显著降低(Zhou et al., 2018). ...
ORA47 (octadecanoid-responsive AP2/ERF- domain transcription factor 47) regulates jasmonic acid and abscisic acid biosynthesis and signaling through binding to a novel cis-element 1 2016
... 已有研究表明, 植物多个转录因子家族参与ABA合成酶基因的转录调控, 其中MYB、AP2/ERF、WRKY、NAC、bHLH和NGA等家族的转录因子促进ABA合成基因转录, 而NFX和HD-ZIP等转录因子抑制ABA合成基因的表达.例如, 拟南芥AtMYB96通过结合AtNCED2/6启动子促进ABA合成和种子休眠(Lee et al., 2015).AP2/ERF (APETALA2/ethylene-responsive factor)家族的AtORA47 (octadecanoid-responsive AP2/ERF-domain transcription factor 47)结合AtNCED3/9启动子的O-box元件, 促进损伤时ABA合成与信号转导(Chen et al., 2016); 而AtDREB2C (dehydration-responsive element binding factor)通过结合AtNCED9启动子的DRE元件激活高温胁迫下种子的ABA合成(Je et al., 2014).拟南芥AtWRKY57结合AtNCED3启动子, 促进干旱诱导的ABA合成、提高植物的耐旱性(Jiang et al., 2012).NAC转录因子中的AtATAF1结合AtNCED3启动子, 促进ABA合成(Jensen et al., 2013).AtNAP (NAC-like, activated by AP3/PI)结合AtAAO3启动子, 促进衰老叶片ABA合成与叶绿素降解(Yang et al., 2014).与AtNAP同源的谷子 (Setaria italica) SiNAC1促进ABA合成, 是叶片衰老的正调节因子(Ren et al., 2018).AtbHLH68过表达株系的抗旱性增强、植株侧根减少, 推测AtbHLH68通过ABA依赖途径调节ABA代谢和/或信号(Le Hir et al., 2017).NGAs (NGA THAs)家族转录因子AtNGA1结合AtNCED3启动子的NBE (NGA-binding element)顺式元件, 通过激活AtNCED3转录促进干旱诱导的ABA合成(Sato et al., 2018).拟南芥AtNFXL2 (nuclear transcription factor X-box binding protein-like 2)抑制内源ABA合成和信号, atnfxl2缺失突变体气孔导度降低, 耐旱性增强; AtNFLX2同工型AtNFLX2-78直接抑制SHINE (SHINE1/SHN1, SHN2, SHN3)基因和BDG1 (BODYGUARD1)基因转录, SHINE与表皮蜡质合成相关, BDG1调控表皮角质层发育, 二者同时促进胁迫环境下植物ABA的合成(Lisso et al., 2011, 2012).除正反馈调节外, ABA合成还存在负反馈调节, 如花生(Arachis hypogaea) AhNAC2和AhAREB1蛋白复合体介导ABA合成的负反馈调节, 外源ABA促进AhNAC2和AhAREB1结合到AhNCED1启动子, 进而抑制AhNCED1转录, 其中AhAREB1是主要负调节因子(Liu et al., 2016).HD-ZIPII转录因子AtHAT1 (homeodomain-leucine zipper protein 1)结合AtABA3和AtNCED3的启动子, 抑制ABA合成和信号(Tan et al., 2018). ...
Dynamic regulation of genome-wide pre-mRNA splicing and stress tolerance by the Sm-like protein LSm5 in Arabidopsis 1 2014
... 植物LSM (like-Sm protein)和XERICO (greek for ‘drought tolerant’)蛋白分别在mRNA代谢和蛋白水平调控内源ABA水平.LSMs是真核生物中高度保守的蛋白家族, 参与RNA代谢(如剪切、脱帽和降解).拟南芥sad1 (supersensitive to ABA and drought)突变体对干旱和外源ABA敏感性增强, sad1植物缺失ABA合成的正反馈调节和干旱诱导的ABA合成, 图位克隆确定SAD1基因编码LSM5蛋白(Xiong et al., 2001).转录组分析表明, SAD1/LSM5蛋白可动态调控拟南芥转录组前体mRNA剪切效率和剪切位点的识别, 表明SAD1/LSM5可能通过调控胁迫基因的表达而介导植物胁迫反应(Cui et al., 2014).LSM1-7复合体通过促进mRNA脱帽和随后的5'-3'降解参与精确的mRNA流转(turnover)和P-bodies形成.当拟南芥暴露在冷和高盐环境时, LSM1-7复合体通过差异化调控AtNCED3和AtNCED5 mRNA流转而调节内源ABA水平(Perea-Resa et al., 2016).拟南芥XERICO蛋白C端含RING-H2锌指基序, XERICO表达上调能显著提高细胞ABA水平并增强植物的抗旱性, 已知很多RING-H2蛋白具有E3泛素连接酶功能, 而酵母双杂交实验显示, AtUBC8 (E2 ubiquitin-conjugating enzyme)和AtTLP9 (ASK1-interacting F-box protein)是XERICO潜在的互作蛋白, 推测XERICO在蛋白水平调节细胞ABA水平(Ko et al., 2006). ...
Abscisic acid uridine diphosphate glucosyltransferases play a crucial role in abscisic acid homeostasis in Arabidopsis 1 2014
... 内源ABA水平还受到ABA特异的葡萄糖基转移酶(ABA-uridine diphosphate glucosyltransferase, ABAUGT)和β-葡萄糖苷酶(β-glucosidase, BG)介导的可逆糖基化代谢调节.研究人员从赤豆(V. angularis)中克隆了编码ABA-UGT酶的基因AOG, UGT可催化ABA与UDPG结合生成ABA葡萄糖苷酯(ABA-glucoside ester, ABA-GE); ABA-GE缺乏生物学活性, 在液泡或质外体中积累(Xu et al., 2002; Priest et al., 2006).拟南芥UGT家族的E亚家族中UGT71B6/7/8参与调节内源ABA水平, 影响AtCYP707A1-4表达, 在脱水和高盐胁迫以及种子萌发和生长发育进程中发挥重要调节作用(Dong et al., 2014).此外, AtUGT71C5也可催化ABA转化为ABA-GE, 与野生型相比, ugt71c5突变体的种子萌发延迟且叶片气孔变小, 抗旱性增强(Liu et al., 2015). ...
ABA biosynthetic and catabolic pathways 4 2014
... 高等植物ABA从头(de novo)合成的第1步是玉米黄质(zeaxanthin)被玉米黄质环氧酶(zeaxanthin epoxidase, ZEP)催化形成环氧玉米黄质(antheraxanthin), 继而环氧化为紫黄质(violaxanthin), 紫黄质异构化为9′-顺式-紫黄质(9′-cis-violaxanthin), 或被新黄素合成酶(neoxanthin synthase, ABA4)催化为反式-新黄质(trans-neoxanthin), 进而异构化为9′-顺式-新黄质(9′-cis-neoxanthin) (Nambara and Marion- Poll, 2005).自1996年以来相继从烟草(Nicotiana plumbaginifolia) (NpABA2)、拟南芥(Arabidopsis thaliana) (AtABA1/LOS6)、水稻(Oryza sativa) (OsABA1)、番茄(Solanum lycopersicum) (SlHP3)和长豇豆(Vigna unguiculata) (VuABA1)中克隆了ZEP基因, 不同物种的ZEP功能缺失突变体都表现为内源ABA含量降低、种子不休眠、植株呈萎蔫状及干旱不能诱导ABA合成等特征(Endo et al., 2014).拟南芥aba1突变体株型矮小, 叶片形状和内部结构畸形; 而ABA1过表达植株的抗盐和抗旱性增强, 在光照条件下气孔开度小于野生型, 说明ABA具有调节植物生长发育和抗逆性的重要功能(Barrero et al., 2005; Park et al., 2008).植物ABA4蛋白高度保守, 定位于叶绿体内囊体膜.拟南芥aba4突变体仅营养组织与野生型有差异, 脱水不能诱导合成ABA, 推测ABA4参与脱水胁迫下的ABA合成(North et al., 2007). ...
... Schwartz等(1997)证明重组的VP14蛋白可切割9′-顺式-紫黄质形成黄质醛, 推测VP14具有NCED功能.目前, 已证明植物NCED蛋白是一个多基因家族, 先后从水稻(OsNCED1-5)、拟南芥(AtNCED2/3/5/6/9)、四季豆(Phaseolus vulgaris) (PvNCED1)、鳄梨(Persea americana) (PaNCED1/3)以及菟丝子(Cuscuta reflexa) (CrNCED1/2)中克隆到NCED基因(Endo et al., 2014).水稻OsNCED1在叶片中表达量最高, OsNCED2主要在种子中表达, OsNCED5参与水稻籽粒灌浆期ABA的合成, OsNCED3转基因拟南芥ABA含量增加, 叶片形态发生改变(Zhu et al., 2009).拟南芥基因组有9个VP14同源序列(AtNCED1-9), 其中AtNCED2/3/5/6/9编码蛋白具有NCED酶活性, AtNCED2/3在植物根和叶中表达量较高.干旱迅速诱导AtNCED3基因表达并促进ABA合成, AtNCED3过表达株系的内源ABA水平较高、叶片蒸腾速率较低、抗旱性较强, 且ABA响应基因的转录本增加(任慧波等, 2007; Hao et al., 2009).种子成熟晚期AtNCED5/6/9表达显著增强, AtNCED6在胚乳中表达, AtNCED5/9在胚乳和胚中表达.nced6/nced9双突变体种子内源ABA含量和休眠程度显著降低, 说明AtNCED6/9主要调控种子休眠(Tan et al., 2003; Lefebvre et al., 2006).nced3/nced5双突变体内源ABA含量减少、营养器官变小且植株抗旱性降低, 说明AtNCED3/5参与植物生长发育和胁迫响应(Frey et al., 2012). ...
... ABA合成最后2步是胞质溶胶中的黄质醛被短链脱氢酶/还原酶(short chain dehydrogenase/reductase-like, SDR1)还原为ABA醛(ABA aldehyde), 继而被ABA醛氧化酶(ABA-aldehyde oxidase, AAO)和钼辅因子硫化酶(molybdenum cofactor sulfurase, MoCo)转化为活性ABA.拟南芥仅有1个AtABA/SDR1 (abscisic acid 2)基因, 在所有器官中组成型表达, 且不受ABA、盐和甘露醇诱导.aba2突变体内源ABA含量减少、种子不休眠、植株呈萎蔫状且主根变短, 外源ABA可恢复其主根根长; 而AtABA2过表达株系的表型与之相反(Lin et al., 2007).拟南芥有4个AAO基因(AtAAO1-4), AtAAO1和AtAAO4分别在干种子和发育中的角果大量表达; AtAAO3是种子和失水叶片中ABA合成关键酶, aao3突变体植株呈萎蔫状、种子休眠程度略有降低(Seo et al., 2004).番茄sitiens突变体不能将ABA醛转化为活性ABA, 导致内源ABA缺失而呈萎蔫状, AtAAO3异位表达能恢复番茄sitiens表型(Harrison et al., 2011).豌豆(Pisum sativum)中有3个PsAO基因, 重组PsAO3蛋白显示AAO酶活性, 干旱使豌豆叶和根的PsAO3基因表达量增加(Zdunek-Zastocka and Sobczak, 2013).拟南芥AtLOS5/ABA3、番茄SiFLACCA和烟草NpABA1/ CKP1基因编码MoCo酶, 可将ABA醛转化为ABA (Endo et al., 2014).AtLOS5/ABA3在所有器官中均表达, 受干旱、盐和ABA诱导, los5突变体内源ABA含量减少、叶片蒸腾速率增加, 抗冷、抗盐及抗旱性减弱, 但AtLOS5过表达可提高转基因棉花(Gossy- pium hirsutum)的内源ABA水平和抗旱性(Xiong et al., 2001; Yue et al., 2012). ...
Drought induction of Arabidopsis 9-cis-epoxycarotenoid dioxygenase occurs in vascular parenchyma cells 1 2008
... ABA从头合成途径的最后步骤由SDR1和AAO酶催化, 因此, 检测SDR1和AAO蛋白定位就可确定ABA合成位点.GFP-和免疫荧光显示, 拟南芥AAO3在根尖、韧皮部伴胞细胞和木质部薄壁细胞中含量最高, 保卫细胞也有少量蛋白(Koiwai et al., 2004).免疫组化也发现叶脉薄壁细胞含有大量的AAO3和ABA2蛋白, 失水可使薄壁细胞内NCED3蛋白显著增加, 证明干旱胁迫下NCED3酶是ABA合成的关键酶(Endo et al., 2008).GUS染色证明NCED3基因在叶肉细胞、保卫细胞、维管束及根系中都有表达, 在根尖表达量最高, 证明NCED3酶促进根源ABA合成, 但保卫细胞ABA合成量低于其它组织(Tan et al., 2003).进一步研究表明, 保卫细胞ABA合成受到维管细胞合成的ABA正反馈调节, 低湿环境下保卫细胞合成的ABA是诱导气孔关闭充分且必要因子(Bauer et al., 2013).种子内源ABA分别来源于母体维管组织、胚和胚乳细胞(Lefebvre et al., 2006).总之, 植物ABA合成位点包括根尖、维管束伴胞细胞和薄壁细胞、叶肉细胞、保卫细胞、种胚和胚乳细胞. ...
Epoxycarotenoid cleavage by NCED5 fine-tunes ABA accumulation and affects seed dormancy and drought tolerance with other NCED family members 1 2012
... Schwartz等(1997)证明重组的VP14蛋白可切割9′-顺式-紫黄质形成黄质醛, 推测VP14具有NCED功能.目前, 已证明植物NCED蛋白是一个多基因家族, 先后从水稻(OsNCED1-5)、拟南芥(AtNCED2/3/5/6/9)、四季豆(Phaseolus vulgaris) (PvNCED1)、鳄梨(Persea americana) (PaNCED1/3)以及菟丝子(Cuscuta reflexa) (CrNCED1/2)中克隆到NCED基因(Endo et al., 2014).水稻OsNCED1在叶片中表达量最高, OsNCED2主要在种子中表达, OsNCED5参与水稻籽粒灌浆期ABA的合成, OsNCED3转基因拟南芥ABA含量增加, 叶片形态发生改变(Zhu et al., 2009).拟南芥基因组有9个VP14同源序列(AtNCED1-9), 其中AtNCED2/3/5/6/9编码蛋白具有NCED酶活性, AtNCED2/3在植物根和叶中表达量较高.干旱迅速诱导AtNCED3基因表达并促进ABA合成, AtNCED3过表达株系的内源ABA水平较高、叶片蒸腾速率较低、抗旱性较强, 且ABA响应基因的转录本增加(任慧波等, 2007; Hao et al., 2009).种子成熟晚期AtNCED5/6/9表达显著增强, AtNCED6在胚乳中表达, AtNCED5/9在胚乳和胚中表达.nced6/nced9双突变体种子内源ABA含量和休眠程度显著降低, 说明AtNCED6/9主要调控种子休眠(Tan et al., 2003; Lefebvre et al., 2006).nced3/nced5双突变体内源ABA含量减少、营养器官变小且植株抗旱性降低, 说明AtNCED3/5参与植物生长发育和胁迫响应(Frey et al., 2012). ...
Nucleotide variation in the NCED3 region of Arabidopsis thaliana and its association study with abscisic acid content under drought stress 1 2009
... Schwartz等(1997)证明重组的VP14蛋白可切割9′-顺式-紫黄质形成黄质醛, 推测VP14具有NCED功能.目前, 已证明植物NCED蛋白是一个多基因家族, 先后从水稻(OsNCED1-5)、拟南芥(AtNCED2/3/5/6/9)、四季豆(Phaseolus vulgaris) (PvNCED1)、鳄梨(Persea americana) (PaNCED1/3)以及菟丝子(Cuscuta reflexa) (CrNCED1/2)中克隆到NCED基因(Endo et al., 2014).水稻OsNCED1在叶片中表达量最高, OsNCED2主要在种子中表达, OsNCED5参与水稻籽粒灌浆期ABA的合成, OsNCED3转基因拟南芥ABA含量增加, 叶片形态发生改变(Zhu et al., 2009).拟南芥基因组有9个VP14同源序列(AtNCED1-9), 其中AtNCED2/3/5/6/9编码蛋白具有NCED酶活性, AtNCED2/3在植物根和叶中表达量较高.干旱迅速诱导AtNCED3基因表达并促进ABA合成, AtNCED3过表达株系的内源ABA水平较高、叶片蒸腾速率较低、抗旱性较强, 且ABA响应基因的转录本增加(任慧波等, 2007; Hao et al., 2009).种子成熟晚期AtNCED5/6/9表达显著增强, AtNCED6在胚乳中表达, AtNCED5/9在胚乳和胚中表达.nced6/nced9双突变体种子内源ABA含量和休眠程度显著降低, 说明AtNCED6/9主要调控种子休眠(Tan et al., 2003; Lefebvre et al., 2006).nced3/nced5双突变体内源ABA含量减少、营养器官变小且植株抗旱性降低, 说明AtNCED3/5参与植物生长发育和胁迫响应(Frey et al., 2012). ...
Identification of the tomato ABA-deficient mutant sitiens as a member of the ABA-aldehyde oxidase gene family using genetic and genomic analysis 1 2011
... ABA合成最后2步是胞质溶胶中的黄质醛被短链脱氢酶/还原酶(short chain dehydrogenase/reductase-like, SDR1)还原为ABA醛(ABA aldehyde), 继而被ABA醛氧化酶(ABA-aldehyde oxidase, AAO)和钼辅因子硫化酶(molybdenum cofactor sulfurase, MoCo)转化为活性ABA.拟南芥仅有1个AtABA/SDR1 (abscisic acid 2)基因, 在所有器官中组成型表达, 且不受ABA、盐和甘露醇诱导.aba2突变体内源ABA含量减少、种子不休眠、植株呈萎蔫状且主根变短, 外源ABA可恢复其主根根长; 而AtABA2过表达株系的表型与之相反(Lin et al., 2007).拟南芥有4个AAO基因(AtAAO1-4), AtAAO1和AtAAO4分别在干种子和发育中的角果大量表达; AtAAO3是种子和失水叶片中ABA合成关键酶, aao3突变体植株呈萎蔫状、种子休眠程度略有降低(Seo et al., 2004).番茄sitiens突变体不能将ABA醛转化为活性ABA, 导致内源ABA缺失而呈萎蔫状, AtAAO3异位表达能恢复番茄sitiens表型(Harrison et al., 2011).豌豆(Pisum sativum)中有3个PsAO基因, 重组PsAO3蛋白显示AAO酶活性, 干旱使豌豆叶和根的PsAO3基因表达量增加(Zdunek-Zastocka and Sobczak, 2013).拟南芥AtLOS5/ABA3、番茄SiFLACCA和烟草NpABA1/ CKP1基因编码MoCo酶, 可将ABA醛转化为ABA (Endo et al., 2014).AtLOS5/ABA3在所有器官中均表达, 受干旱、盐和ABA诱导, los5突变体内源ABA含量减少、叶片蒸腾速率增加, 抗冷、抗盐及抗旱性减弱, 但AtLOS5过表达可提高转基因棉花(Gossy- pium hirsutum)的内源ABA水平和抗旱性(Xiong et al., 2001; Yue et al., 2012). ...
Arabidopsis DREB2C modulates ABA biosynthesis during germination 1 2014
... 已有研究表明, 植物多个转录因子家族参与ABA合成酶基因的转录调控, 其中MYB、AP2/ERF、WRKY、NAC、bHLH和NGA等家族的转录因子促进ABA合成基因转录, 而NFX和HD-ZIP等转录因子抑制ABA合成基因的表达.例如, 拟南芥AtMYB96通过结合AtNCED2/6启动子促进ABA合成和种子休眠(Lee et al., 2015).AP2/ERF (APETALA2/ethylene-responsive factor)家族的AtORA47 (octadecanoid-responsive AP2/ERF-domain transcription factor 47)结合AtNCED3/9启动子的O-box元件, 促进损伤时ABA合成与信号转导(Chen et al., 2016); 而AtDREB2C (dehydration-responsive element binding factor)通过结合AtNCED9启动子的DRE元件激活高温胁迫下种子的ABA合成(Je et al., 2014).拟南芥AtWRKY57结合AtNCED3启动子, 促进干旱诱导的ABA合成、提高植物的耐旱性(Jiang et al., 2012).NAC转录因子中的AtATAF1结合AtNCED3启动子, 促进ABA合成(Jensen et al., 2013).AtNAP (NAC-like, activated by AP3/PI)结合AtAAO3启动子, 促进衰老叶片ABA合成与叶绿素降解(Yang et al., 2014).与AtNAP同源的谷子 (Setaria italica) SiNAC1促进ABA合成, 是叶片衰老的正调节因子(Ren et al., 2018).AtbHLH68过表达株系的抗旱性增强、植株侧根减少, 推测AtbHLH68通过ABA依赖途径调节ABA代谢和/或信号(Le Hir et al., 2017).NGAs (NGA THAs)家族转录因子AtNGA1结合AtNCED3启动子的NBE (NGA-binding element)顺式元件, 通过激活AtNCED3转录促进干旱诱导的ABA合成(Sato et al., 2018).拟南芥AtNFXL2 (nuclear transcription factor X-box binding protein-like 2)抑制内源ABA合成和信号, atnfxl2缺失突变体气孔导度降低, 耐旱性增强; AtNFLX2同工型AtNFLX2-78直接抑制SHINE (SHINE1/SHN1, SHN2, SHN3)基因和BDG1 (BODYGUARD1)基因转录, SHINE与表皮蜡质合成相关, BDG1调控表皮角质层发育, 二者同时促进胁迫环境下植物ABA的合成(Lisso et al., 2011, 2012).除正反馈调节外, ABA合成还存在负反馈调节, 如花生(Arachis hypogaea) AhNAC2和AhAREB1蛋白复合体介导ABA合成的负反馈调节, 外源ABA促进AhNAC2和AhAREB1结合到AhNCED1启动子, 进而抑制AhNCED1转录, 其中AhAREB1是主要负调节因子(Liu et al., 2016).HD-ZIPII转录因子AtHAT1 (homeodomain-leucine zipper protein 1)结合AtABA3和AtNCED3的启动子, 抑制ABA合成和信号(Tan et al., 2018). ...
... AtABCG40/PDR12 (pleiotropic drug resistance 12)定位在保卫细胞和其它细胞的质膜, abcg40突变体原生质体对ABA吸收能力显著下降(Kang et al., 2010).AtABCG25是ABA输出转运蛋白, AtABCG25在维管组织中表达, 过表达植株保卫细胞ABA信号增强、植株水分利用率提高(Kuromori et al., 2010, 2016).AtNRT1.2/AIT1介导质膜ABA输入转运, nrt1.2/ait1突变体对外源ABA敏感性降低(Kanno et al., 2012).AtABCG40和AtNRT1.2/AIT1介导细胞吸收ABA的空间表达谱不同, AtABCG40在保卫细胞中表达, 而AtNRT1.2/AIT1在维管细胞中表达; AtNRT1.2/AIT1与AtABCG25共同调节维管组织向保卫细胞转运ABA, 且AtNRT1.2/AIT1介导ABA进入维管组织可能与植物通过木质部和韧皮部进行ABA再分配相关(Kuromori et al., 2018).拟南芥MATE (DTX/multidrug and toxin compound extrusion)家族成员DTX50蛋白定位在维管细胞和保卫细胞质膜, 介导维管细胞和保卫细胞向外转运ABA; 外源ABA促进DTX50表达, dtx50突变体对ABA敏感、气孔导度较低、抗旱性增强(Zhang et al., 2014).AtABCG22定位于质膜, 具有ABA转运功能, 参与ABA转运、合成和信号转导(Kuromori et al., 2011).此外, 研究表明AtABCG25/31/30/40协同参与种子的ABA转运, 其中胚乳表达的AtABCG25/31为ABA输出转运蛋白, 协同将胚乳细胞合成的ABA转运出去, 而种胚表达的AtABCG30/40为输入转运蛋白, 共同将ABA转运入种胚细胞(Kang et al., 2015). ...
Arabidopsis mutants of AtABCG22, an ABC transporter gene, increase water transpiration and drought susceptibility 1 2011
... AtABCG40/PDR12 (pleiotropic drug resistance 12)定位在保卫细胞和其它细胞的质膜, abcg40突变体原生质体对ABA吸收能力显著下降(Kang et al., 2010).AtABCG25是ABA输出转运蛋白, AtABCG25在维管组织中表达, 过表达植株保卫细胞ABA信号增强、植株水分利用率提高(Kuromori et al., 2010, 2016).AtNRT1.2/AIT1介导质膜ABA输入转运, nrt1.2/ait1突变体对外源ABA敏感性降低(Kanno et al., 2012).AtABCG40和AtNRT1.2/AIT1介导细胞吸收ABA的空间表达谱不同, AtABCG40在保卫细胞中表达, 而AtNRT1.2/AIT1在维管细胞中表达; AtNRT1.2/AIT1与AtABCG25共同调节维管组织向保卫细胞转运ABA, 且AtNRT1.2/AIT1介导ABA进入维管组织可能与植物通过木质部和韧皮部进行ABA再分配相关(Kuromori et al., 2018).拟南芥MATE (DTX/multidrug and toxin compound extrusion)家族成员DTX50蛋白定位在维管细胞和保卫细胞质膜, 介导维管细胞和保卫细胞向外转运ABA; 外源ABA促进DTX50表达, dtx50突变体对ABA敏感、气孔导度较低、抗旱性增强(Zhang et al., 2014).AtABCG22定位于质膜, 具有ABA转运功能, 参与ABA转运、合成和信号转导(Kuromori et al., 2011).此外, 研究表明AtABCG25/31/30/40协同参与种子的ABA转运, 其中胚乳表达的AtABCG25/31为ABA输出转运蛋白, 协同将胚乳细胞合成的ABA转运出去, 而种胚表达的AtABCG30/40为输入转运蛋白, 共同将ABA转运入种胚细胞(Kang et al., 2015). ...
The Arabidopsis cytochrome P450 CYP707A encodes ABA 8′-hydroxylases: key enzymes in ABA catabolism 1 2004
... 拟南芥AtCYP707As转录本广泛存在于各种器官组织, 但表达量有所不同.例如, 花和角果中AtCYP707A1表达量最高, AtCYP707A2和AtCYP707A3在叶、茎和根中表达较高, 但AtCYP707A4在所有组织中表达都较低, 种子吸胀增强AtCYP707A2的表达, 干旱抑制而复水促进所有AtCYP707As表达(Kushiro et al., 2004).AtCYP707A1主要调节成熟中期种子ABA水平, 而AtCYP707A2调控成熟晚期至萌发阶段的ABA代谢(Okamoto et al., 2006).AtCYP707A3是主要的干旱应答基因, 正常生长环境中的cyp707a3突变体内源ABA含量高于野生型、叶片蒸腾速率降低并对外源ABA敏感; 干旱促进突变体积累更多ABA, 植株耐旱性显著增强, 但过表达植株表型相反(Umezawa et al., 2006).在正常生长环境中, cyp707a1和cyp707a3突变体叶片的气孔导度都较低; 高湿环境显著增强AtCYP707A1/3表达、促进ABA降解, cyp707a3突变体可以积累更高水平的ABA, 然而cyp707a1突变体的ABA水平与野生型相似.研究表明, 在高湿环境中AtCYP707A1使保卫细胞内的ABA失活, AtCYP707A3则主要降解维管组织中移动的ABA, 植物气孔运动受到来自维管组织的ABA和保卫细胞内的ABA代谢共同调控(Okamoto et al., 2009). ...
At bHLH68 transcription factor contributes to the regulation of ABA homeostasis and drought stress tolerance in Arabidopsis thaliana 1 2017
... 已有研究表明, 植物多个转录因子家族参与ABA合成酶基因的转录调控, 其中MYB、AP2/ERF、WRKY、NAC、bHLH和NGA等家族的转录因子促进ABA合成基因转录, 而NFX和HD-ZIP等转录因子抑制ABA合成基因的表达.例如, 拟南芥AtMYB96通过结合AtNCED2/6启动子促进ABA合成和种子休眠(Lee et al., 2015).AP2/ERF (APETALA2/ethylene-responsive factor)家族的AtORA47 (octadecanoid-responsive AP2/ERF-domain transcription factor 47)结合AtNCED3/9启动子的O-box元件, 促进损伤时ABA合成与信号转导(Chen et al., 2016); 而AtDREB2C (dehydration-responsive element binding factor)通过结合AtNCED9启动子的DRE元件激活高温胁迫下种子的ABA合成(Je et al., 2014).拟南芥AtWRKY57结合AtNCED3启动子, 促进干旱诱导的ABA合成、提高植物的耐旱性(Jiang et al., 2012).NAC转录因子中的AtATAF1结合AtNCED3启动子, 促进ABA合成(Jensen et al., 2013).AtNAP (NAC-like, activated by AP3/PI)结合AtAAO3启动子, 促进衰老叶片ABA合成与叶绿素降解(Yang et al., 2014).与AtNAP同源的谷子 (Setaria italica) SiNAC1促进ABA合成, 是叶片衰老的正调节因子(Ren et al., 2018).AtbHLH68过表达株系的抗旱性增强、植株侧根减少, 推测AtbHLH68通过ABA依赖途径调节ABA代谢和/或信号(Le Hir et al., 2017).NGAs (NGA THAs)家族转录因子AtNGA1结合AtNCED3启动子的NBE (NGA-binding element)顺式元件, 通过激活AtNCED3转录促进干旱诱导的ABA合成(Sato et al., 2018).拟南芥AtNFXL2 (nuclear transcription factor X-box binding protein-like 2)抑制内源ABA合成和信号, atnfxl2缺失突变体气孔导度降低, 耐旱性增强; AtNFLX2同工型AtNFLX2-78直接抑制SHINE (SHINE1/SHN1, SHN2, SHN3)基因和BDG1 (BODYGUARD1)基因转录, SHINE与表皮蜡质合成相关, BDG1调控表皮角质层发育, 二者同时促进胁迫环境下植物ABA的合成(Lisso et al., 2011, 2012).除正反馈调节外, ABA合成还存在负反馈调节, 如花生(Arachis hypogaea) AhNAC2和AhAREB1蛋白复合体介导ABA合成的负反馈调节, 外源ABA促进AhNAC2和AhAREB1结合到AhNCED1启动子, 进而抑制AhNCED1转录, 其中AhAREB1是主要负调节因子(Liu et al., 2016).HD-ZIPII转录因子AtHAT1 (homeodomain-leucine zipper protein 1)结合AtABA3和AtNCED3的启动子, 抑制ABA合成和信号(Tan et al., 2018). ...
The Arabidopsis MYB96 transcription factor plays a role in seed dormancy 1 2015
... 已有研究表明, 植物多个转录因子家族参与ABA合成酶基因的转录调控, 其中MYB、AP2/ERF、WRKY、NAC、bHLH和NGA等家族的转录因子促进ABA合成基因转录, 而NFX和HD-ZIP等转录因子抑制ABA合成基因的表达.例如, 拟南芥AtMYB96通过结合AtNCED2/6启动子促进ABA合成和种子休眠(Lee et al., 2015).AP2/ERF (APETALA2/ethylene-responsive factor)家族的AtORA47 (octadecanoid-responsive AP2/ERF-domain transcription factor 47)结合AtNCED3/9启动子的O-box元件, 促进损伤时ABA合成与信号转导(Chen et al., 2016); 而AtDREB2C (dehydration-responsive element binding factor)通过结合AtNCED9启动子的DRE元件激活高温胁迫下种子的ABA合成(Je et al., 2014).拟南芥AtWRKY57结合AtNCED3启动子, 促进干旱诱导的ABA合成、提高植物的耐旱性(Jiang et al., 2012).NAC转录因子中的AtATAF1结合AtNCED3启动子, 促进ABA合成(Jensen et al., 2013).AtNAP (NAC-like, activated by AP3/PI)结合AtAAO3启动子, 促进衰老叶片ABA合成与叶绿素降解(Yang et al., 2014).与AtNAP同源的谷子 (Setaria italica) SiNAC1促进ABA合成, 是叶片衰老的正调节因子(Ren et al., 2018).AtbHLH68过表达株系的抗旱性增强、植株侧根减少, 推测AtbHLH68通过ABA依赖途径调节ABA代谢和/或信号(Le Hir et al., 2017).NGAs (NGA THAs)家族转录因子AtNGA1结合AtNCED3启动子的NBE (NGA-binding element)顺式元件, 通过激活AtNCED3转录促进干旱诱导的ABA合成(Sato et al., 2018).拟南芥AtNFXL2 (nuclear transcription factor X-box binding protein-like 2)抑制内源ABA合成和信号, atnfxl2缺失突变体气孔导度降低, 耐旱性增强; AtNFLX2同工型AtNFLX2-78直接抑制SHINE (SHINE1/SHN1, SHN2, SHN3)基因和BDG1 (BODYGUARD1)基因转录, SHINE与表皮蜡质合成相关, BDG1调控表皮角质层发育, 二者同时促进胁迫环境下植物ABA的合成(Lisso et al., 2011, 2012).除正反馈调节外, ABA合成还存在负反馈调节, 如花生(Arachis hypogaea) AhNAC2和AhAREB1蛋白复合体介导ABA合成的负反馈调节, 外源ABA促进AhNAC2和AhAREB1结合到AhNCED1启动子, 进而抑制AhNCED1转录, 其中AhAREB1是主要负调节因子(Liu et al., 2016).HD-ZIPII转录因子AtHAT1 (homeodomain-leucine zipper protein 1)结合AtABA3和AtNCED3的启动子, 抑制ABA合成和信号(Tan et al., 2018). ...
Activation of glucosidase via stress-induced polymerization rapidly increases active pools of abscisic acid 1 2006
... 干旱条件下, 液泡或质外体贮存的ABA-GE迅速被BG水解, 生成活性ABA以适应变化的环境.拟南芥AtBG1在内质网将ABA-GE水解生成ABA, bg1突变体内源ABA含量和抗旱性降低; 脱水迅速诱导BG1蛋白多聚化后显著增强BG活性, 从而加速ABA生成, ABA正反馈激活BG1蛋白多聚化(Lee et al., 2006).AtBG2以复合物形式储存在中央液泡, 脱水诱导AtBG2表达, bg2突变体对脱水和盐胁迫敏感, AtBG2过表达可互补bg1表型(Xu et al., 2012).在低湿环境下, 拟南芥aba3-1突变体中AtBG1转录水平极显著高于野生型, 说明细胞内ABA-GE水解机制可部分补偿ABA从头合成的缺陷(Bauer et al., 2013).此外, AtBGLU10编码的BG酶分布在液泡中, bglu10突变体的BG酶活性和ABA含量均降低, 且叶面温度和胁迫反应基因表达量也降低, 而AtBGLU10过表达株系抗旱性增强(Wang et al., 2011a). ...
Functional analysis of Arabidopsis NCED6 and NCED9 genes indicates that ABA synthesized in the endosperm is involved in the induction of seed dormancy 2 2006
... Schwartz等(1997)证明重组的VP14蛋白可切割9′-顺式-紫黄质形成黄质醛, 推测VP14具有NCED功能.目前, 已证明植物NCED蛋白是一个多基因家族, 先后从水稻(OsNCED1-5)、拟南芥(AtNCED2/3/5/6/9)、四季豆(Phaseolus vulgaris) (PvNCED1)、鳄梨(Persea americana) (PaNCED1/3)以及菟丝子(Cuscuta reflexa) (CrNCED1/2)中克隆到NCED基因(Endo et al., 2014).水稻OsNCED1在叶片中表达量最高, OsNCED2主要在种子中表达, OsNCED5参与水稻籽粒灌浆期ABA的合成, OsNCED3转基因拟南芥ABA含量增加, 叶片形态发生改变(Zhu et al., 2009).拟南芥基因组有9个VP14同源序列(AtNCED1-9), 其中AtNCED2/3/5/6/9编码蛋白具有NCED酶活性, AtNCED2/3在植物根和叶中表达量较高.干旱迅速诱导AtNCED3基因表达并促进ABA合成, AtNCED3过表达株系的内源ABA水平较高、叶片蒸腾速率较低、抗旱性较强, 且ABA响应基因的转录本增加(任慧波等, 2007; Hao et al., 2009).种子成熟晚期AtNCED5/6/9表达显著增强, AtNCED6在胚乳中表达, AtNCED5/9在胚乳和胚中表达.nced6/nced9双突变体种子内源ABA含量和休眠程度显著降低, 说明AtNCED6/9主要调控种子休眠(Tan et al., 2003; Lefebvre et al., 2006).nced3/nced5双突变体内源ABA含量减少、营养器官变小且植株抗旱性降低, 说明AtNCED3/5参与植物生长发育和胁迫响应(Frey et al., 2012). ...
... ABA从头合成途径的最后步骤由SDR1和AAO酶催化, 因此, 检测SDR1和AAO蛋白定位就可确定ABA合成位点.GFP-和免疫荧光显示, 拟南芥AAO3在根尖、韧皮部伴胞细胞和木质部薄壁细胞中含量最高, 保卫细胞也有少量蛋白(Koiwai et al., 2004).免疫组化也发现叶脉薄壁细胞含有大量的AAO3和ABA2蛋白, 失水可使薄壁细胞内NCED3蛋白显著增加, 证明干旱胁迫下NCED3酶是ABA合成的关键酶(Endo et al., 2008).GUS染色证明NCED3基因在叶肉细胞、保卫细胞、维管束及根系中都有表达, 在根尖表达量最高, 证明NCED3酶促进根源ABA合成, 但保卫细胞ABA合成量低于其它组织(Tan et al., 2003).进一步研究表明, 保卫细胞ABA合成受到维管细胞合成的ABA正反馈调节, 低湿环境下保卫细胞合成的ABA是诱导气孔关闭充分且必要因子(Bauer et al., 2013).种子内源ABA分别来源于母体维管组织、胚和胚乳细胞(Lefebvre et al., 2006).总之, 植物ABA合成位点包括根尖、维管束伴胞细胞和薄壁细胞、叶肉细胞、保卫细胞、种胚和胚乳细胞. ...
Long-distance ABA transport can mediate distal tissue responses by affecting local ABA concentrations 1 2018
Ectopic expression of ABSCISIC ACID 2/GLUCOSE INSENSITIVE 1 in Arabidopsis promotes seed dormancy and stress tolerance 1 2007
... ABA合成最后2步是胞质溶胶中的黄质醛被短链脱氢酶/还原酶(short chain dehydrogenase/reductase-like, SDR1)还原为ABA醛(ABA aldehyde), 继而被ABA醛氧化酶(ABA-aldehyde oxidase, AAO)和钼辅因子硫化酶(molybdenum cofactor sulfurase, MoCo)转化为活性ABA.拟南芥仅有1个AtABA/SDR1 (abscisic acid 2)基因, 在所有器官中组成型表达, 且不受ABA、盐和甘露醇诱导.aba2突变体内源ABA含量减少、种子不休眠、植株呈萎蔫状且主根变短, 外源ABA可恢复其主根根长; 而AtABA2过表达株系的表型与之相反(Lin et al., 2007).拟南芥有4个AAO基因(AtAAO1-4), AtAAO1和AtAAO4分别在干种子和发育中的角果大量表达; AtAAO3是种子和失水叶片中ABA合成关键酶, aao3突变体植株呈萎蔫状、种子休眠程度略有降低(Seo et al., 2004).番茄sitiens突变体不能将ABA醛转化为活性ABA, 导致内源ABA缺失而呈萎蔫状, AtAAO3异位表达能恢复番茄sitiens表型(Harrison et al., 2011).豌豆(Pisum sativum)中有3个PsAO基因, 重组PsAO3蛋白显示AAO酶活性, 干旱使豌豆叶和根的PsAO3基因表达量增加(Zdunek-Zastocka and Sobczak, 2013).拟南芥AtLOS5/ABA3、番茄SiFLACCA和烟草NpABA1/ CKP1基因编码MoCo酶, 可将ABA醛转化为ABA (Endo et al., 2014).AtLOS5/ABA3在所有器官中均表达, 受干旱、盐和ABA诱导, los5突变体内源ABA含量减少、叶片蒸腾速率增加, 抗冷、抗盐及抗旱性减弱, 但AtLOS5过表达可提高转基因棉花(Gossy- pium hirsutum)的内源ABA水平和抗旱性(Xiong et al., 2001; Yue et al., 2012). ...
NFX1-LIKE2 (NFXL2) suppresses abscisic acid accumulation and stomatal closure in Arabidopsis thaliana 1 2011
... 已有研究表明, 植物多个转录因子家族参与ABA合成酶基因的转录调控, 其中MYB、AP2/ERF、WRKY、NAC、bHLH和NGA等家族的转录因子促进ABA合成基因转录, 而NFX和HD-ZIP等转录因子抑制ABA合成基因的表达.例如, 拟南芥AtMYB96通过结合AtNCED2/6启动子促进ABA合成和种子休眠(Lee et al., 2015).AP2/ERF (APETALA2/ethylene-responsive factor)家族的AtORA47 (octadecanoid-responsive AP2/ERF-domain transcription factor 47)结合AtNCED3/9启动子的O-box元件, 促进损伤时ABA合成与信号转导(Chen et al., 2016); 而AtDREB2C (dehydration-responsive element binding factor)通过结合AtNCED9启动子的DRE元件激活高温胁迫下种子的ABA合成(Je et al., 2014).拟南芥AtWRKY57结合AtNCED3启动子, 促进干旱诱导的ABA合成、提高植物的耐旱性(Jiang et al., 2012).NAC转录因子中的AtATAF1结合AtNCED3启动子, 促进ABA合成(Jensen et al., 2013).AtNAP (NAC-like, activated by AP3/PI)结合AtAAO3启动子, 促进衰老叶片ABA合成与叶绿素降解(Yang et al., 2014).与AtNAP同源的谷子 (Setaria italica) SiNAC1促进ABA合成, 是叶片衰老的正调节因子(Ren et al., 2018).AtbHLH68过表达株系的抗旱性增强、植株侧根减少, 推测AtbHLH68通过ABA依赖途径调节ABA代谢和/或信号(Le Hir et al., 2017).NGAs (NGA THAs)家族转录因子AtNGA1结合AtNCED3启动子的NBE (NGA-binding element)顺式元件, 通过激活AtNCED3转录促进干旱诱导的ABA合成(Sato et al., 2018).拟南芥AtNFXL2 (nuclear transcription factor X-box binding protein-like 2)抑制内源ABA合成和信号, atnfxl2缺失突变体气孔导度降低, 耐旱性增强; AtNFLX2同工型AtNFLX2-78直接抑制SHINE (SHINE1/SHN1, SHN2, SHN3)基因和BDG1 (BODYGUARD1)基因转录, SHINE与表皮蜡质合成相关, BDG1调控表皮角质层发育, 二者同时促进胁迫环境下植物ABA的合成(Lisso et al., 2011, 2012).除正反馈调节外, ABA合成还存在负反馈调节, 如花生(Arachis hypogaea) AhNAC2和AhAREB1蛋白复合体介导ABA合成的负反馈调节, 外源ABA促进AhNAC2和AhAREB1结合到AhNCED1启动子, 进而抑制AhNCED1转录, 其中AhAREB1是主要负调节因子(Liu et al., 2016).HD-ZIPII转录因子AtHAT1 (homeodomain-leucine zipper protein 1)结合AtABA3和AtNCED3的启动子, 抑制ABA合成和信号(Tan et al., 2018). ...
Nfxl2 modifies cuticle properties in Arabidopsis 1 2012
... 已有研究表明, 植物多个转录因子家族参与ABA合成酶基因的转录调控, 其中MYB、AP2/ERF、WRKY、NAC、bHLH和NGA等家族的转录因子促进ABA合成基因转录, 而NFX和HD-ZIP等转录因子抑制ABA合成基因的表达.例如, 拟南芥AtMYB96通过结合AtNCED2/6启动子促进ABA合成和种子休眠(Lee et al., 2015).AP2/ERF (APETALA2/ethylene-responsive factor)家族的AtORA47 (octadecanoid-responsive AP2/ERF-domain transcription factor 47)结合AtNCED3/9启动子的O-box元件, 促进损伤时ABA合成与信号转导(Chen et al., 2016); 而AtDREB2C (dehydration-responsive element binding factor)通过结合AtNCED9启动子的DRE元件激活高温胁迫下种子的ABA合成(Je et al., 2014).拟南芥AtWRKY57结合AtNCED3启动子, 促进干旱诱导的ABA合成、提高植物的耐旱性(Jiang et al., 2012).NAC转录因子中的AtATAF1结合AtNCED3启动子, 促进ABA合成(Jensen et al., 2013).AtNAP (NAC-like, activated by AP3/PI)结合AtAAO3启动子, 促进衰老叶片ABA合成与叶绿素降解(Yang et al., 2014).与AtNAP同源的谷子 (Setaria italica) SiNAC1促进ABA合成, 是叶片衰老的正调节因子(Ren et al., 2018).AtbHLH68过表达株系的抗旱性增强、植株侧根减少, 推测AtbHLH68通过ABA依赖途径调节ABA代谢和/或信号(Le Hir et al., 2017).NGAs (NGA THAs)家族转录因子AtNGA1结合AtNCED3启动子的NBE (NGA-binding element)顺式元件, 通过激活AtNCED3转录促进干旱诱导的ABA合成(Sato et al., 2018).拟南芥AtNFXL2 (nuclear transcription factor X-box binding protein-like 2)抑制内源ABA合成和信号, atnfxl2缺失突变体气孔导度降低, 耐旱性增强; AtNFLX2同工型AtNFLX2-78直接抑制SHINE (SHINE1/SHN1, SHN2, SHN3)基因和BDG1 (BODYGUARD1)基因转录, SHINE与表皮蜡质合成相关, BDG1调控表皮角质层发育, 二者同时促进胁迫环境下植物ABA的合成(Lisso et al., 2011, 2012).除正反馈调节外, ABA合成还存在负反馈调节, 如花生(Arachis hypogaea) AhNAC2和AhAREB1蛋白复合体介导ABA合成的负反馈调节, 外源ABA促进AhNAC2和AhAREB1结合到AhNCED1启动子, 进而抑制AhNCED1转录, 其中AhAREB1是主要负调节因子(Liu et al., 2016).HD-ZIPII转录因子AtHAT1 (homeodomain-leucine zipper protein 1)结合AtABA3和AtNCED3的启动子, 抑制ABA合成和信号(Tan et al., 2018). ...
Negative feedback regulation of ABA biosynthesis in peanut (Arachis hypogaea): a transcription factor complex inhibits AhNCED1 expression during water stress 1 2016
... 已有研究表明, 植物多个转录因子家族参与ABA合成酶基因的转录调控, 其中MYB、AP2/ERF、WRKY、NAC、bHLH和NGA等家族的转录因子促进ABA合成基因转录, 而NFX和HD-ZIP等转录因子抑制ABA合成基因的表达.例如, 拟南芥AtMYB96通过结合AtNCED2/6启动子促进ABA合成和种子休眠(Lee et al., 2015).AP2/ERF (APETALA2/ethylene-responsive factor)家族的AtORA47 (octadecanoid-responsive AP2/ERF-domain transcription factor 47)结合AtNCED3/9启动子的O-box元件, 促进损伤时ABA合成与信号转导(Chen et al., 2016); 而AtDREB2C (dehydration-responsive element binding factor)通过结合AtNCED9启动子的DRE元件激活高温胁迫下种子的ABA合成(Je et al., 2014).拟南芥AtWRKY57结合AtNCED3启动子, 促进干旱诱导的ABA合成、提高植物的耐旱性(Jiang et al., 2012).NAC转录因子中的AtATAF1结合AtNCED3启动子, 促进ABA合成(Jensen et al., 2013).AtNAP (NAC-like, activated by AP3/PI)结合AtAAO3启动子, 促进衰老叶片ABA合成与叶绿素降解(Yang et al., 2014).与AtNAP同源的谷子 (Setaria italica) SiNAC1促进ABA合成, 是叶片衰老的正调节因子(Ren et al., 2018).AtbHLH68过表达株系的抗旱性增强、植株侧根减少, 推测AtbHLH68通过ABA依赖途径调节ABA代谢和/或信号(Le Hir et al., 2017).NGAs (NGA THAs)家族转录因子AtNGA1结合AtNCED3启动子的NBE (NGA-binding element)顺式元件, 通过激活AtNCED3转录促进干旱诱导的ABA合成(Sato et al., 2018).拟南芥AtNFXL2 (nuclear transcription factor X-box binding protein-like 2)抑制内源ABA合成和信号, atnfxl2缺失突变体气孔导度降低, 耐旱性增强; AtNFLX2同工型AtNFLX2-78直接抑制SHINE (SHINE1/SHN1, SHN2, SHN3)基因和BDG1 (BODYGUARD1)基因转录, SHINE与表皮蜡质合成相关, BDG1调控表皮角质层发育, 二者同时促进胁迫环境下植物ABA的合成(Lisso et al., 2011, 2012).除正反馈调节外, ABA合成还存在负反馈调节, 如花生(Arachis hypogaea) AhNAC2和AhAREB1蛋白复合体介导ABA合成的负反馈调节, 外源ABA促进AhNAC2和AhAREB1结合到AhNCED1启动子, 进而抑制AhNCED1转录, 其中AhAREB1是主要负调节因子(Liu et al., 2016).HD-ZIPII转录因子AtHAT1 (homeodomain-leucine zipper protein 1)结合AtABA3和AtNCED3的启动子, 抑制ABA合成和信号(Tan et al., 2018). ...
bHLH122 is important for drought and osmotic stress resistance in Arabidopsis and in the repression of ABA catabolism 1 2014
... 已有研究表明若干转录因子参与ABA代谢基因的表达调控.拟南芥CHO1 (CHOTTO1)为AP2/ERF型转录因子, 促进AtABA1和AtNCED9、抑制AtCYP707A2/3转录, 调控吸胀期种子ABA抑制的赤霉素合成(Yano et al., 2009).AP2/ERF型转录因子ABI4促进ABA积累和种子休眠, 通过结合AtCYP707A1/2启动子并抑制其转录来实现(Shu et al., 2013).拟南芥NLP8 (NIN-like protein 8)以硝酸盐依赖的方式结合AtCYP707A2的启动子并激活其转录, 降低种子ABA水平, 促进种子萌发(Matakiadis et al., 2009; Yan et al., 2016).bZIP家族转录因子VIP1 (VirE2-interacting protein 1)为渗透胁迫的调节因子, VIP1直接结合AtCYP707A1/3启动子并激活其转录(Tsugama et al., 2012).拟南芥bHLH122是干旱、NaCl和渗透胁迫的正调节因子, bHLH122结合AtCYP707A3启动子G-/E-box元件并抑制其表达, 进而抑制ABA代谢(Liu et al., 2014).拟南芥成花抑制因子SVP (short vegetative phase)直接结合AtCYP707A1/3和AtBG1启动子的CArG Motif元件, 抑制AtCYP707A1/3, 促进AtBG1转录, 表明SVP是干旱胁迫下ABA代谢的重要调控因子(Wang et al., 2018). ...
UDP-glucosyltransferase71C5, a major glucosyltransferase, mediates abscisic acid homeostasis in Arabidopsis 1 2015
... 内源ABA水平还受到ABA特异的葡萄糖基转移酶(ABA-uridine diphosphate glucosyltransferase, ABAUGT)和β-葡萄糖苷酶(β-glucosidase, BG)介导的可逆糖基化代谢调节.研究人员从赤豆(V. angularis)中克隆了编码ABA-UGT酶的基因AOG, UGT可催化ABA与UDPG结合生成ABA葡萄糖苷酯(ABA-glucoside ester, ABA-GE); ABA-GE缺乏生物学活性, 在液泡或质外体中积累(Xu et al., 2002; Priest et al., 2006).拟南芥UGT家族的E亚家族中UGT71B6/7/8参与调节内源ABA水平, 影响AtCYP707A1-4表达, 在脱水和高盐胁迫以及种子萌发和生长发育进程中发挥重要调节作用(Dong et al., 2014).此外, AtUGT71C5也可催化ABA转化为ABA-GE, 与野生型相比, ugt71c5突变体的种子萌发延迟且叶片气孔变小, 抗旱性增强(Liu et al., 2015). ...
Molecular mechanism for the regulation of ABA homeostasis during plant development and stress responses 1 2018
... 植物激素脱落酸(abscisic acid, ABA)能调节种子成熟、休眠与萌发、气孔运动及叶片衰老等重要生理过程, 并介导植物对环境胁迫的快速反应.植物特定组织中内源ABA水平受ABA从头合成、羟基化代谢、糖基化失活和去糖基活化以及ABA长距离转运等的动态调控(魏开发等, 2012; 伍静辉等, 2018; Ma et al., 2018) (图1A).近20年来, 植物ABA合成、代谢和信号转导研究取得了重要进展, 但ABA合成与代谢基因的启动和表达调控机制的研究却相对滞后.本文依据近年来植物内源ABA水平的动态调控机制的相关研究, 综述了植物ABA从头合成、羟基化与糖基化代谢以及ABA转运等领域的最新研究成果, 着重讨论ABA合成与代谢基因的表达调控机制(图1B), 并展望了今后的研究方向, 旨在为植物抗逆研究提供理论参考. ...
Drought-enhanced xylem sap sulfate closes stomata by affecting ALMT12 and guard cell ABA synthesis 1 2017
... 此外, 硫元素供应、角质层合成基因、钙调素信号蛋白也参与ABA生物合成的调控.叶绿体硫转运蛋白AtSULTR3;1功能缺失显著降低幼苗AAO酶活性和内源ABA水平, 硫元素供应可提高AtNCED3和AtABA3转录水平; ABA促进AtSULTR3;1和其它硫代谢基因转录, 表明硫在植物响应环境胁迫时发挥重要作用(Cao et al., 2014).干旱使木质部汁液硫酸盐含量增加, 进而促进保卫细胞AtNCED3表达和ABA合成, 引起气孔关闭(Malcheska et al., 2017).已知植物角质层作为物理屏障减少水分流失并介导ABA合成和信号转导, 进而调控植物渗透胁迫反应.拟南芥ced1 (9-cis-epoxycarotenoid dioxygenase defective 1)突变体对轻微渗透胁迫极为敏感, CED1/BDG1基因促进ABA合成基因的表达(Wang et al., 2011b).植物钙调素类似蛋白AtCML37 (calmodulin-like 37)和AtCML42拮抗调控干旱胁迫诱导的ABA积累, AtCML37为正调节因子, 而AtCML42为负调节因子(Scholz et al., 2015).我们也发现钙调素结合蛋白AtIQM4能促进种子ABA合成, iqm4突变体种子ABA含量降低, 角果AtNCED6/9表达量显著降低(Zhou et al., 2018). ...
The Arabidopsis abscisic acid catabolic gene CYP707A2 plays a key role in nitrate control of seed dormancy 1 2009
... 已有研究表明若干转录因子参与ABA代谢基因的表达调控.拟南芥CHO1 (CHOTTO1)为AP2/ERF型转录因子, 促进AtABA1和AtNCED9、抑制AtCYP707A2/3转录, 调控吸胀期种子ABA抑制的赤霉素合成(Yano et al., 2009).AP2/ERF型转录因子ABI4促进ABA积累和种子休眠, 通过结合AtCYP707A1/2启动子并抑制其转录来实现(Shu et al., 2013).拟南芥NLP8 (NIN-like protein 8)以硝酸盐依赖的方式结合AtCYP707A2的启动子并激活其转录, 降低种子ABA水平, 促进种子萌发(Matakiadis et al., 2009; Yan et al., 2016).bZIP家族转录因子VIP1 (VirE2-interacting protein 1)为渗透胁迫的调节因子, VIP1直接结合AtCYP707A1/3启动子并激活其转录(Tsugama et al., 2012).拟南芥bHLH122是干旱、NaCl和渗透胁迫的正调节因子, bHLH122结合AtCYP707A3启动子G-/E-box元件并抑制其表达, 进而抑制ABA代谢(Liu et al., 2014).拟南芥成花抑制因子SVP (short vegetative phase)直接结合AtCYP707A1/3和AtBG1启动子的CArG Motif元件, 抑制AtCYP707A1/3, 促进AtBG1转录, 表明SVP是干旱胁迫下ABA代谢的重要调控因子(Wang et al., 2018). ...
Abscisic acid biosynthesis and catabolism 1 2005
... 高等植物ABA从头(de novo)合成的第1步是玉米黄质(zeaxanthin)被玉米黄质环氧酶(zeaxanthin epoxidase, ZEP)催化形成环氧玉米黄质(antheraxanthin), 继而环氧化为紫黄质(violaxanthin), 紫黄质异构化为9′-顺式-紫黄质(9′-cis-violaxanthin), 或被新黄素合成酶(neoxanthin synthase, ABA4)催化为反式-新黄质(trans-neoxanthin), 进而异构化为9′-顺式-新黄质(9′-cis-neoxanthin) (Nambara and Marion- Poll, 2005).自1996年以来相继从烟草(Nicotiana plumbaginifolia) (NpABA2)、拟南芥(Arabidopsis thaliana) (AtABA1/LOS6)、水稻(Oryza sativa) (OsABA1)、番茄(Solanum lycopersicum) (SlHP3)和长豇豆(Vigna unguiculata) (VuABA1)中克隆了ZEP基因, 不同物种的ZEP功能缺失突变体都表现为内源ABA含量降低、种子不休眠、植株呈萎蔫状及干旱不能诱导ABA合成等特征(Endo et al., 2014).拟南芥aba1突变体株型矮小, 叶片形状和内部结构畸形; 而ABA1过表达植株的抗盐和抗旱性增强, 在光照条件下气孔开度小于野生型, 说明ABA具有调节植物生长发育和抗逆性的重要功能(Barrero et al., 2005; Park et al., 2008).植物ABA4蛋白高度保守, 定位于叶绿体内囊体膜.拟南芥aba4突变体仅营养组织与野生型有差异, 脱水不能诱导合成ABA, 推测ABA4参与脱水胁迫下的ABA合成(North et al., 2007). ...
The Arabidopsis ABA- deficient mutant aba4 demonstrates that the major route for stress-induced ABA accumulation is via neoxanthin isomers 1 2007
... 高等植物ABA从头(de novo)合成的第1步是玉米黄质(zeaxanthin)被玉米黄质环氧酶(zeaxanthin epoxidase, ZEP)催化形成环氧玉米黄质(antheraxanthin), 继而环氧化为紫黄质(violaxanthin), 紫黄质异构化为9′-顺式-紫黄质(9′-cis-violaxanthin), 或被新黄素合成酶(neoxanthin synthase, ABA4)催化为反式-新黄质(trans-neoxanthin), 进而异构化为9′-顺式-新黄质(9′-cis-neoxanthin) (Nambara and Marion- Poll, 2005).自1996年以来相继从烟草(Nicotiana plumbaginifolia) (NpABA2)、拟南芥(Arabidopsis thaliana) (AtABA1/LOS6)、水稻(Oryza sativa) (OsABA1)、番茄(Solanum lycopersicum) (SlHP3)和长豇豆(Vigna unguiculata) (VuABA1)中克隆了ZEP基因, 不同物种的ZEP功能缺失突变体都表现为内源ABA含量降低、种子不休眠、植株呈萎蔫状及干旱不能诱导ABA合成等特征(Endo et al., 2014).拟南芥aba1突变体株型矮小, 叶片形状和内部结构畸形; 而ABA1过表达植株的抗盐和抗旱性增强, 在光照条件下气孔开度小于野生型, 说明ABA具有调节植物生长发育和抗逆性的重要功能(Barrero et al., 2005; Park et al., 2008).植物ABA4蛋白高度保守, 定位于叶绿体内囊体膜.拟南芥aba4突变体仅营养组织与野生型有差异, 脱水不能诱导合成ABA, 推测ABA4参与脱水胁迫下的ABA合成(North et al., 2007). ...
CYP707A1 and CYP707A2, which encode abscisic acid 8′-hydroxylases, are indispensable for proper control of seed dormancy and germination in Arabidopsis 1 2006
... 拟南芥AtCYP707As转录本广泛存在于各种器官组织, 但表达量有所不同.例如, 花和角果中AtCYP707A1表达量最高, AtCYP707A2和AtCYP707A3在叶、茎和根中表达较高, 但AtCYP707A4在所有组织中表达都较低, 种子吸胀增强AtCYP707A2的表达, 干旱抑制而复水促进所有AtCYP707As表达(Kushiro et al., 2004).AtCYP707A1主要调节成熟中期种子ABA水平, 而AtCYP707A2调控成熟晚期至萌发阶段的ABA代谢(Okamoto et al., 2006).AtCYP707A3是主要的干旱应答基因, 正常生长环境中的cyp707a3突变体内源ABA含量高于野生型、叶片蒸腾速率降低并对外源ABA敏感; 干旱促进突变体积累更多ABA, 植株耐旱性显著增强, 但过表达植株表型相反(Umezawa et al., 2006).在正常生长环境中, cyp707a1和cyp707a3突变体叶片的气孔导度都较低; 高湿环境显著增强AtCYP707A1/3表达、促进ABA降解, cyp707a3突变体可以积累更高水平的ABA, 然而cyp707a1突变体的ABA水平与野生型相似.研究表明, 在高湿环境中AtCYP707A1使保卫细胞内的ABA失活, AtCYP707A3则主要降解维管组织中移动的ABA, 植物气孔运动受到来自维管组织的ABA和保卫细胞内的ABA代谢共同调控(Okamoto et al., 2009). ...
High humidity induces abscisic acid 8′-hydroxylase in stomata and vasculature to regulate local and systemic abscisic acid responses in Arabidopsis 1 2009
... 拟南芥AtCYP707As转录本广泛存在于各种器官组织, 但表达量有所不同.例如, 花和角果中AtCYP707A1表达量最高, AtCYP707A2和AtCYP707A3在叶、茎和根中表达较高, 但AtCYP707A4在所有组织中表达都较低, 种子吸胀增强AtCYP707A2的表达, 干旱抑制而复水促进所有AtCYP707As表达(Kushiro et al., 2004).AtCYP707A1主要调节成熟中期种子ABA水平, 而AtCYP707A2调控成熟晚期至萌发阶段的ABA代谢(Okamoto et al., 2006).AtCYP707A3是主要的干旱应答基因, 正常生长环境中的cyp707a3突变体内源ABA含量高于野生型、叶片蒸腾速率降低并对外源ABA敏感; 干旱促进突变体积累更多ABA, 植株耐旱性显著增强, 但过表达植株表型相反(Umezawa et al., 2006).在正常生长环境中, cyp707a1和cyp707a3突变体叶片的气孔导度都较低; 高湿环境显著增强AtCYP707A1/3表达、促进ABA降解, cyp707a3突变体可以积累更高水平的ABA, 然而cyp707a1突变体的ABA水平与野生型相似.研究表明, 在高湿环境中AtCYP707A1使保卫细胞内的ABA失活, AtCYP707A3则主要降解维管组织中移动的ABA, 植物气孔运动受到来自维管组织的ABA和保卫细胞内的ABA代谢共同调控(Okamoto et al., 2009). ...
Overexpression of Arabidopsis ZEP enhances tolerance to osmotic stress 1 2008
... 高等植物ABA从头(de novo)合成的第1步是玉米黄质(zeaxanthin)被玉米黄质环氧酶(zeaxanthin epoxidase, ZEP)催化形成环氧玉米黄质(antheraxanthin), 继而环氧化为紫黄质(violaxanthin), 紫黄质异构化为9′-顺式-紫黄质(9′-cis-violaxanthin), 或被新黄素合成酶(neoxanthin synthase, ABA4)催化为反式-新黄质(trans-neoxanthin), 进而异构化为9′-顺式-新黄质(9′-cis-neoxanthin) (Nambara and Marion- Poll, 2005).自1996年以来相继从烟草(Nicotiana plumbaginifolia) (NpABA2)、拟南芥(Arabidopsis thaliana) (AtABA1/LOS6)、水稻(Oryza sativa) (OsABA1)、番茄(Solanum lycopersicum) (SlHP3)和长豇豆(Vigna unguiculata) (VuABA1)中克隆了ZEP基因, 不同物种的ZEP功能缺失突变体都表现为内源ABA含量降低、种子不休眠、植株呈萎蔫状及干旱不能诱导ABA合成等特征(Endo et al., 2014).拟南芥aba1突变体株型矮小, 叶片形状和内部结构畸形; 而ABA1过表达植株的抗盐和抗旱性增强, 在光照条件下气孔开度小于野生型, 说明ABA具有调节植物生长发育和抗逆性的重要功能(Barrero et al., 2005; Park et al., 2008).植物ABA4蛋白高度保守, 定位于叶绿体内囊体膜.拟南芥aba4突变体仅营养组织与野生型有差异, 脱水不能诱导合成ABA, 推测ABA4参与脱水胁迫下的ABA合成(North et al., 2007). ...
Spatial regulation of ABCG25, an ABA exporter, is an important component of the mechanism controlling cellular ABA levels 1 2016
The LSM1-7 complex differentially regulates Arabidopsis tolerance to abiotic stress conditions by promoting selective mRNA decapping 1 2016
... 植物LSM (like-Sm protein)和XERICO (greek for ‘drought tolerant’)蛋白分别在mRNA代谢和蛋白水平调控内源ABA水平.LSMs是真核生物中高度保守的蛋白家族, 参与RNA代谢(如剪切、脱帽和降解).拟南芥sad1 (supersensitive to ABA and drought)突变体对干旱和外源ABA敏感性增强, sad1植物缺失ABA合成的正反馈调节和干旱诱导的ABA合成, 图位克隆确定SAD1基因编码LSM5蛋白(Xiong et al., 2001).转录组分析表明, SAD1/LSM5蛋白可动态调控拟南芥转录组前体mRNA剪切效率和剪切位点的识别, 表明SAD1/LSM5可能通过调控胁迫基因的表达而介导植物胁迫反应(Cui et al., 2014).LSM1-7复合体通过促进mRNA脱帽和随后的5'-3'降解参与精确的mRNA流转(turnover)和P-bodies形成.当拟南芥暴露在冷和高盐环境时, LSM1-7复合体通过差异化调控AtNCED3和AtNCED5 mRNA流转而调节内源ABA水平(Perea-Resa et al., 2016).拟南芥XERICO蛋白C端含RING-H2锌指基序, XERICO表达上调能显著提高细胞ABA水平并增强植物的抗旱性, 已知很多RING-H2蛋白具有E3泛素连接酶功能, 而酵母双杂交实验显示, AtUBC8 (E2 ubiquitin-conjugating enzyme)和AtTLP9 (ASK1-interacting F-box protein)是XERICO潜在的互作蛋白, 推测XERICO在蛋白水平调节细胞ABA水平(Ko et al., 2006). ...
Use of the glucosyltransferase UGT71B6 to disturb abscisic acid homeostasis in Arabidopsis thaliana 1 2006
... 内源ABA水平还受到ABA特异的葡萄糖基转移酶(ABA-uridine diphosphate glucosyltransferase, ABAUGT)和β-葡萄糖苷酶(β-glucosidase, BG)介导的可逆糖基化代谢调节.研究人员从赤豆(V. angularis)中克隆了编码ABA-UGT酶的基因AOG, UGT可催化ABA与UDPG结合生成ABA葡萄糖苷酯(ABA-glucoside ester, ABA-GE); ABA-GE缺乏生物学活性, 在液泡或质外体中积累(Xu et al., 2002; Priest et al., 2006).拟南芥UGT家族的E亚家族中UGT71B6/7/8参与调节内源ABA水平, 影响AtCYP707A1-4表达, 在脱水和高盐胁迫以及种子萌发和生长发育进程中发挥重要调节作用(Dong et al., 2014).此外, AtUGT71C5也可催化ABA转化为ABA-GE, 与野生型相比, ugt71c5突变体的种子萌发延迟且叶片气孔变小, 抗旱性增强(Liu et al., 2015). ...
Involvement of NAC transcription factor SiNAC1 in a positive feedback loop via ABA biosynthesis and leaf senescence in foxtail millet 1 2018
... 已有研究表明, 植物多个转录因子家族参与ABA合成酶基因的转录调控, 其中MYB、AP2/ERF、WRKY、NAC、bHLH和NGA等家族的转录因子促进ABA合成基因转录, 而NFX和HD-ZIP等转录因子抑制ABA合成基因的表达.例如, 拟南芥AtMYB96通过结合AtNCED2/6启动子促进ABA合成和种子休眠(Lee et al., 2015).AP2/ERF (APETALA2/ethylene-responsive factor)家族的AtORA47 (octadecanoid-responsive AP2/ERF-domain transcription factor 47)结合AtNCED3/9启动子的O-box元件, 促进损伤时ABA合成与信号转导(Chen et al., 2016); 而AtDREB2C (dehydration-responsive element binding factor)通过结合AtNCED9启动子的DRE元件激活高温胁迫下种子的ABA合成(Je et al., 2014).拟南芥AtWRKY57结合AtNCED3启动子, 促进干旱诱导的ABA合成、提高植物的耐旱性(Jiang et al., 2012).NAC转录因子中的AtATAF1结合AtNCED3启动子, 促进ABA合成(Jensen et al., 2013).AtNAP (NAC-like, activated by AP3/PI)结合AtAAO3启动子, 促进衰老叶片ABA合成与叶绿素降解(Yang et al., 2014).与AtNAP同源的谷子 (Setaria italica) SiNAC1促进ABA合成, 是叶片衰老的正调节因子(Ren et al., 2018).AtbHLH68过表达株系的抗旱性增强、植株侧根减少, 推测AtbHLH68通过ABA依赖途径调节ABA代谢和/或信号(Le Hir et al., 2017).NGAs (NGA THAs)家族转录因子AtNGA1结合AtNCED3启动子的NBE (NGA-binding element)顺式元件, 通过激活AtNCED3转录促进干旱诱导的ABA合成(Sato et al., 2018).拟南芥AtNFXL2 (nuclear transcription factor X-box binding protein-like 2)抑制内源ABA合成和信号, atnfxl2缺失突变体气孔导度降低, 耐旱性增强; AtNFLX2同工型AtNFLX2-78直接抑制SHINE (SHINE1/SHN1, SHN2, SHN3)基因和BDG1 (BODYGUARD1)基因转录, SHINE与表皮蜡质合成相关, BDG1调控表皮角质层发育, 二者同时促进胁迫环境下植物ABA的合成(Lisso et al., 2011, 2012).除正反馈调节外, ABA合成还存在负反馈调节, 如花生(Arachis hypogaea) AhNAC2和AhAREB1蛋白复合体介导ABA合成的负反馈调节, 外源ABA促进AhNAC2和AhAREB1结合到AhNCED1启动子, 进而抑制AhNCED1转录, 其中AhAREB1是主要负调节因子(Liu et al., 2016).HD-ZIPII转录因子AtHAT1 (homeodomain-leucine zipper protein 1)结合AtABA3和AtNCED3的启动子, 抑制ABA合成和信号(Tan et al., 2018). ...
Arabidopsis thaliana NGATHA1 transcription factor induces ABA biosynthesis by activating NCED3 gene during dehydration stress 1 2018
... 已有研究表明, 植物多个转录因子家族参与ABA合成酶基因的转录调控, 其中MYB、AP2/ERF、WRKY、NAC、bHLH和NGA等家族的转录因子促进ABA合成基因转录, 而NFX和HD-ZIP等转录因子抑制ABA合成基因的表达.例如, 拟南芥AtMYB96通过结合AtNCED2/6启动子促进ABA合成和种子休眠(Lee et al., 2015).AP2/ERF (APETALA2/ethylene-responsive factor)家族的AtORA47 (octadecanoid-responsive AP2/ERF-domain transcription factor 47)结合AtNCED3/9启动子的O-box元件, 促进损伤时ABA合成与信号转导(Chen et al., 2016); 而AtDREB2C (dehydration-responsive element binding factor)通过结合AtNCED9启动子的DRE元件激活高温胁迫下种子的ABA合成(Je et al., 2014).拟南芥AtWRKY57结合AtNCED3启动子, 促进干旱诱导的ABA合成、提高植物的耐旱性(Jiang et al., 2012).NAC转录因子中的AtATAF1结合AtNCED3启动子, 促进ABA合成(Jensen et al., 2013).AtNAP (NAC-like, activated by AP3/PI)结合AtAAO3启动子, 促进衰老叶片ABA合成与叶绿素降解(Yang et al., 2014).与AtNAP同源的谷子 (Setaria italica) SiNAC1促进ABA合成, 是叶片衰老的正调节因子(Ren et al., 2018).AtbHLH68过表达株系的抗旱性增强、植株侧根减少, 推测AtbHLH68通过ABA依赖途径调节ABA代谢和/或信号(Le Hir et al., 2017).NGAs (NGA THAs)家族转录因子AtNGA1结合AtNCED3启动子的NBE (NGA-binding element)顺式元件, 通过激活AtNCED3转录促进干旱诱导的ABA合成(Sato et al., 2018).拟南芥AtNFXL2 (nuclear transcription factor X-box binding protein-like 2)抑制内源ABA合成和信号, atnfxl2缺失突变体气孔导度降低, 耐旱性增强; AtNFLX2同工型AtNFLX2-78直接抑制SHINE (SHINE1/SHN1, SHN2, SHN3)基因和BDG1 (BODYGUARD1)基因转录, SHINE与表皮蜡质合成相关, BDG1调控表皮角质层发育, 二者同时促进胁迫环境下植物ABA的合成(Lisso et al., 2011, 2012).除正反馈调节外, ABA合成还存在负反馈调节, 如花生(Arachis hypogaea) AhNAC2和AhAREB1蛋白复合体介导ABA合成的负反馈调节, 外源ABA促进AhNAC2和AhAREB1结合到AhNCED1启动子, 进而抑制AhNCED1转录, 其中AhAREB1是主要负调节因子(Liu et al., 2016).HD-ZIPII转录因子AtHAT1 (homeodomain-leucine zipper protein 1)结合AtABA3和AtNCED3的启动子, 抑制ABA合成和信号(Tan et al., 2018). ...
Calmodulin-like protein CML37 is a positive regulator of ABA during drought stress in Arabidopsis 1 2015
... 此外, 硫元素供应、角质层合成基因、钙调素信号蛋白也参与ABA生物合成的调控.叶绿体硫转运蛋白AtSULTR3;1功能缺失显著降低幼苗AAO酶活性和内源ABA水平, 硫元素供应可提高AtNCED3和AtABA3转录水平; ABA促进AtSULTR3;1和其它硫代谢基因转录, 表明硫在植物响应环境胁迫时发挥重要作用(Cao et al., 2014).干旱使木质部汁液硫酸盐含量增加, 进而促进保卫细胞AtNCED3表达和ABA合成, 引起气孔关闭(Malcheska et al., 2017).已知植物角质层作为物理屏障减少水分流失并介导ABA合成和信号转导, 进而调控植物渗透胁迫反应.拟南芥ced1 (9-cis-epoxycarotenoid dioxygenase defective 1)突变体对轻微渗透胁迫极为敏感, CED1/BDG1基因促进ABA合成基因的表达(Wang et al., 2011b).植物钙调素类似蛋白AtCML37 (calmodulin-like 37)和AtCML42拮抗调控干旱胁迫诱导的ABA积累, AtCML37为正调节因子, 而AtCML42为负调节因子(Scholz et al., 2015).我们也发现钙调素结合蛋白AtIQM4能促进种子ABA合成, iqm4突变体种子ABA含量降低, 角果AtNCED6/9表达量显著降低(Zhou et al., 2018). ...
Specific oxidative cleavage of carotenoids by VP14 of maize 1 1997
... Schwartz等(1997)证明重组的VP14蛋白可切割9′-顺式-紫黄质形成黄质醛, 推测VP14具有NCED功能.目前, 已证明植物NCED蛋白是一个多基因家族, 先后从水稻(OsNCED1-5)、拟南芥(AtNCED2/3/5/6/9)、四季豆(Phaseolus vulgaris) (PvNCED1)、鳄梨(Persea americana) (PaNCED1/3)以及菟丝子(Cuscuta reflexa) (CrNCED1/2)中克隆到NCED基因(Endo et al., 2014).水稻OsNCED1在叶片中表达量最高, OsNCED2主要在种子中表达, OsNCED5参与水稻籽粒灌浆期ABA的合成, OsNCED3转基因拟南芥ABA含量增加, 叶片形态发生改变(Zhu et al., 2009).拟南芥基因组有9个VP14同源序列(AtNCED1-9), 其中AtNCED2/3/5/6/9编码蛋白具有NCED酶活性, AtNCED2/3在植物根和叶中表达量较高.干旱迅速诱导AtNCED3基因表达并促进ABA合成, AtNCED3过表达株系的内源ABA水平较高、叶片蒸腾速率较低、抗旱性较强, 且ABA响应基因的转录本增加(任慧波等, 2007; Hao et al., 2009).种子成熟晚期AtNCED5/6/9表达显著增强, AtNCED6在胚乳中表达, AtNCED5/9在胚乳和胚中表达.nced6/nced9双突变体种子内源ABA含量和休眠程度显著降低, 说明AtNCED6/9主要调控种子休眠(Tan et al., 2003; Lefebvre et al., 2006).nced3/nced5双突变体内源ABA含量减少、营养器官变小且植株抗旱性降低, 说明AtNCED3/5参与植物生长发育和胁迫响应(Frey et al., 2012). ...
Comparative studies on the Arabidopsis aldehyde oxidase (AAO) gene family revealed a major role of AAO3 in ABA biosynthesis in seeds 1 2004
... ABA合成最后2步是胞质溶胶中的黄质醛被短链脱氢酶/还原酶(short chain dehydrogenase/reductase-like, SDR1)还原为ABA醛(ABA aldehyde), 继而被ABA醛氧化酶(ABA-aldehyde oxidase, AAO)和钼辅因子硫化酶(molybdenum cofactor sulfurase, MoCo)转化为活性ABA.拟南芥仅有1个AtABA/SDR1 (abscisic acid 2)基因, 在所有器官中组成型表达, 且不受ABA、盐和甘露醇诱导.aba2突变体内源ABA含量减少、种子不休眠、植株呈萎蔫状且主根变短, 外源ABA可恢复其主根根长; 而AtABA2过表达株系的表型与之相反(Lin et al., 2007).拟南芥有4个AAO基因(AtAAO1-4), AtAAO1和AtAAO4分别在干种子和发育中的角果大量表达; AtAAO3是种子和失水叶片中ABA合成关键酶, aao3突变体植株呈萎蔫状、种子休眠程度略有降低(Seo et al., 2004).番茄sitiens突变体不能将ABA醛转化为活性ABA, 导致内源ABA缺失而呈萎蔫状, AtAAO3异位表达能恢复番茄sitiens表型(Harrison et al., 2011).豌豆(Pisum sativum)中有3个PsAO基因, 重组PsAO3蛋白显示AAO酶活性, 干旱使豌豆叶和根的PsAO3基因表达量增加(Zdunek-Zastocka and Sobczak, 2013).拟南芥AtLOS5/ABA3、番茄SiFLACCA和烟草NpABA1/ CKP1基因编码MoCo酶, 可将ABA醛转化为ABA (Endo et al., 2014).AtLOS5/ABA3在所有器官中均表达, 受干旱、盐和ABA诱导, los5突变体内源ABA含量减少、叶片蒸腾速率增加, 抗冷、抗盐及抗旱性减弱, 但AtLOS5过表达可提高转基因棉花(Gossy- pium hirsutum)的内源ABA水平和抗旱性(Xiong et al., 2001; Yue et al., 2012). ...
ABI4 regulates primary seed dormancy by regulating the biogenesis of abscisic acid and gibberellins in Arabidopsis 1 2013
... 已有研究表明若干转录因子参与ABA代谢基因的表达调控.拟南芥CHO1 (CHOTTO1)为AP2/ERF型转录因子, 促进AtABA1和AtNCED9、抑制AtCYP707A2/3转录, 调控吸胀期种子ABA抑制的赤霉素合成(Yano et al., 2009).AP2/ERF型转录因子ABI4促进ABA积累和种子休眠, 通过结合AtCYP707A1/2启动子并抑制其转录来实现(Shu et al., 2013).拟南芥NLP8 (NIN-like protein 8)以硝酸盐依赖的方式结合AtCYP707A2的启动子并激活其转录, 降低种子ABA水平, 促进种子萌发(Matakiadis et al., 2009; Yan et al., 2016).bZIP家族转录因子VIP1 (VirE2-interacting protein 1)为渗透胁迫的调节因子, VIP1直接结合AtCYP707A1/3启动子并激活其转录(Tsugama et al., 2012).拟南芥bHLH122是干旱、NaCl和渗透胁迫的正调节因子, bHLH122结合AtCYP707A3启动子G-/E-box元件并抑制其表达, 进而抑制ABA代谢(Liu et al., 2014).拟南芥成花抑制因子SVP (short vegetative phase)直接结合AtCYP707A1/3和AtBG1启动子的CArG Motif元件, 抑制AtCYP707A1/3, 促进AtBG1转录, 表明SVP是干旱胁迫下ABA代谢的重要调控因子(Wang et al., 2018). ...
Up-regulation of NCED3 and ABA biosynthesis occur within minutes of a decrease in leaf turgor but AHK1 is not required 1 2017
Transcription factor HAT1 is a substrate of SnRK2.3 kinase and negatively regulates ABA synthesis and signaling in Arabidopsis responding to drought 1 2018
... 已有研究表明, 植物多个转录因子家族参与ABA合成酶基因的转录调控, 其中MYB、AP2/ERF、WRKY、NAC、bHLH和NGA等家族的转录因子促进ABA合成基因转录, 而NFX和HD-ZIP等转录因子抑制ABA合成基因的表达.例如, 拟南芥AtMYB96通过结合AtNCED2/6启动子促进ABA合成和种子休眠(Lee et al., 2015).AP2/ERF (APETALA2/ethylene-responsive factor)家族的AtORA47 (octadecanoid-responsive AP2/ERF-domain transcription factor 47)结合AtNCED3/9启动子的O-box元件, 促进损伤时ABA合成与信号转导(Chen et al., 2016); 而AtDREB2C (dehydration-responsive element binding factor)通过结合AtNCED9启动子的DRE元件激活高温胁迫下种子的ABA合成(Je et al., 2014).拟南芥AtWRKY57结合AtNCED3启动子, 促进干旱诱导的ABA合成、提高植物的耐旱性(Jiang et al., 2012).NAC转录因子中的AtATAF1结合AtNCED3启动子, 促进ABA合成(Jensen et al., 2013).AtNAP (NAC-like, activated by AP3/PI)结合AtAAO3启动子, 促进衰老叶片ABA合成与叶绿素降解(Yang et al., 2014).与AtNAP同源的谷子 (Setaria italica) SiNAC1促进ABA合成, 是叶片衰老的正调节因子(Ren et al., 2018).AtbHLH68过表达株系的抗旱性增强、植株侧根减少, 推测AtbHLH68通过ABA依赖途径调节ABA代谢和/或信号(Le Hir et al., 2017).NGAs (NGA THAs)家族转录因子AtNGA1结合AtNCED3启动子的NBE (NGA-binding element)顺式元件, 通过激活AtNCED3转录促进干旱诱导的ABA合成(Sato et al., 2018).拟南芥AtNFXL2 (nuclear transcription factor X-box binding protein-like 2)抑制内源ABA合成和信号, atnfxl2缺失突变体气孔导度降低, 耐旱性增强; AtNFLX2同工型AtNFLX2-78直接抑制SHINE (SHINE1/SHN1, SHN2, SHN3)基因和BDG1 (BODYGUARD1)基因转录, SHINE与表皮蜡质合成相关, BDG1调控表皮角质层发育, 二者同时促进胁迫环境下植物ABA的合成(Lisso et al., 2011, 2012).除正反馈调节外, ABA合成还存在负反馈调节, 如花生(Arachis hypogaea) AhNAC2和AhAREB1蛋白复合体介导ABA合成的负反馈调节, 外源ABA促进AhNAC2和AhAREB1结合到AhNCED1启动子, 进而抑制AhNCED1转录, 其中AhAREB1是主要负调节因子(Liu et al., 2016).HD-ZIPII转录因子AtHAT1 (homeodomain-leucine zipper protein 1)结合AtABA3和AtNCED3的启动子, 抑制ABA合成和信号(Tan et al., 2018). ...
A bZIP protein, VIP1, is a regulator of osmosensory signaling in Arabidopsis 1 2012
... 已有研究表明若干转录因子参与ABA代谢基因的表达调控.拟南芥CHO1 (CHOTTO1)为AP2/ERF型转录因子, 促进AtABA1和AtNCED9、抑制AtCYP707A2/3转录, 调控吸胀期种子ABA抑制的赤霉素合成(Yano et al., 2009).AP2/ERF型转录因子ABI4促进ABA积累和种子休眠, 通过结合AtCYP707A1/2启动子并抑制其转录来实现(Shu et al., 2013).拟南芥NLP8 (NIN-like protein 8)以硝酸盐依赖的方式结合AtCYP707A2的启动子并激活其转录, 降低种子ABA水平, 促进种子萌发(Matakiadis et al., 2009; Yan et al., 2016).bZIP家族转录因子VIP1 (VirE2-interacting protein 1)为渗透胁迫的调节因子, VIP1直接结合AtCYP707A1/3启动子并激活其转录(Tsugama et al., 2012).拟南芥bHLH122是干旱、NaCl和渗透胁迫的正调节因子, bHLH122结合AtCYP707A3启动子G-/E-box元件并抑制其表达, 进而抑制ABA代谢(Liu et al., 2014).拟南芥成花抑制因子SVP (short vegetative phase)直接结合AtCYP707A1/3和AtBG1启动子的CArG Motif元件, 抑制AtCYP707A1/3, 促进AtBG1转录, 表明SVP是干旱胁迫下ABA代谢的重要调控因子(Wang et al., 2018). ...
CYP707A3, a major ABA 80-hydroxylase involved in dehydration and rehydration response in Arabido- psis thaliana 1 2006
... 拟南芥AtCYP707As转录本广泛存在于各种器官组织, 但表达量有所不同.例如, 花和角果中AtCYP707A1表达量最高, AtCYP707A2和AtCYP707A3在叶、茎和根中表达较高, 但AtCYP707A4在所有组织中表达都较低, 种子吸胀增强AtCYP707A2的表达, 干旱抑制而复水促进所有AtCYP707As表达(Kushiro et al., 2004).AtCYP707A1主要调节成熟中期种子ABA水平, 而AtCYP707A2调控成熟晚期至萌发阶段的ABA代谢(Okamoto et al., 2006).AtCYP707A3是主要的干旱应答基因, 正常生长环境中的cyp707a3突变体内源ABA含量高于野生型、叶片蒸腾速率降低并对外源ABA敏感; 干旱促进突变体积累更多ABA, 植株耐旱性显著增强, 但过表达植株表型相反(Umezawa et al., 2006).在正常生长环境中, cyp707a1和cyp707a3突变体叶片的气孔导度都较低; 高湿环境显著增强AtCYP707A1/3表达、促进ABA降解, cyp707a3突变体可以积累更高水平的ABA, 然而cyp707a1突变体的ABA水平与野生型相似.研究表明, 在高湿环境中AtCYP707A1使保卫细胞内的ABA失活, AtCYP707A3则主要降解维管组织中移动的ABA, 植物气孔运动受到来自维管组织的ABA和保卫细胞内的ABA代谢共同调控(Okamoto et al., 2009). ...
From epoxycarotenoids to ABA: the role of ABA 8′-hydroxylases in drought- stressed maize roots 1 2010
A vacuole localized β-glucosidase contributes to drought tolerance in Arabidopsis 1 2011
... 干旱条件下, 液泡或质外体贮存的ABA-GE迅速被BG水解, 生成活性ABA以适应变化的环境.拟南芥AtBG1在内质网将ABA-GE水解生成ABA, bg1突变体内源ABA含量和抗旱性降低; 脱水迅速诱导BG1蛋白多聚化后显著增强BG活性, 从而加速ABA生成, ABA正反馈激活BG1蛋白多聚化(Lee et al., 2006).AtBG2以复合物形式储存在中央液泡, 脱水诱导AtBG2表达, bg2突变体对脱水和盐胁迫敏感, AtBG2过表达可互补bg1表型(Xu et al., 2012).在低湿环境下, 拟南芥aba3-1突变体中AtBG1转录水平极显著高于野生型, 说明细胞内ABA-GE水解机制可部分补偿ABA从头合成的缺陷(Bauer et al., 2013).此外, AtBGLU10编码的BG酶分布在液泡中, bglu10突变体的BG酶活性和ABA含量均降低, 且叶面温度和胁迫反应基因表达量也降低, 而AtBGLU10过表达株系抗旱性增强(Wang et al., 2011a). ...
The flowering repressor SVP confers drought resistance in Arabidopsis by regulating abscisic acid catabolism 1 2018
... 已有研究表明若干转录因子参与ABA代谢基因的表达调控.拟南芥CHO1 (CHOTTO1)为AP2/ERF型转录因子, 促进AtABA1和AtNCED9、抑制AtCYP707A2/3转录, 调控吸胀期种子ABA抑制的赤霉素合成(Yano et al., 2009).AP2/ERF型转录因子ABI4促进ABA积累和种子休眠, 通过结合AtCYP707A1/2启动子并抑制其转录来实现(Shu et al., 2013).拟南芥NLP8 (NIN-like protein 8)以硝酸盐依赖的方式结合AtCYP707A2的启动子并激活其转录, 降低种子ABA水平, 促进种子萌发(Matakiadis et al., 2009; Yan et al., 2016).bZIP家族转录因子VIP1 (VirE2-interacting protein 1)为渗透胁迫的调节因子, VIP1直接结合AtCYP707A1/3启动子并激活其转录(Tsugama et al., 2012).拟南芥bHLH122是干旱、NaCl和渗透胁迫的正调节因子, bHLH122结合AtCYP707A3启动子G-/E-box元件并抑制其表达, 进而抑制ABA代谢(Liu et al., 2014).拟南芥成花抑制因子SVP (short vegetative phase)直接结合AtCYP707A1/3和AtBG1启动子的CArG Motif元件, 抑制AtCYP707A1/3, 促进AtBG1转录, 表明SVP是干旱胁迫下ABA代谢的重要调控因子(Wang et al., 2018). ...
The plant cuticle is required for osmotic stress regulation of abscisic acid biosynthesis and osmotic stress tolerance in Arabidopsis 1 2011
... 此外, 硫元素供应、角质层合成基因、钙调素信号蛋白也参与ABA生物合成的调控.叶绿体硫转运蛋白AtSULTR3;1功能缺失显著降低幼苗AAO酶活性和内源ABA水平, 硫元素供应可提高AtNCED3和AtABA3转录水平; ABA促进AtSULTR3;1和其它硫代谢基因转录, 表明硫在植物响应环境胁迫时发挥重要作用(Cao et al., 2014).干旱使木质部汁液硫酸盐含量增加, 进而促进保卫细胞AtNCED3表达和ABA合成, 引起气孔关闭(Malcheska et al., 2017).已知植物角质层作为物理屏障减少水分流失并介导ABA合成和信号转导, 进而调控植物渗透胁迫反应.拟南芥ced1 (9-cis-epoxycarotenoid dioxygenase defective 1)突变体对轻微渗透胁迫极为敏感, CED1/BDG1基因促进ABA合成基因的表达(Wang et al., 2011b).植物钙调素类似蛋白AtCML37 (calmodulin-like 37)和AtCML42拮抗调控干旱胁迫诱导的ABA积累, AtCML37为正调节因子, 而AtCML42为负调节因子(Scholz et al., 2015).我们也发现钙调素结合蛋白AtIQM4能促进种子ABA合成, iqm4突变体种子ABA含量降低, 角果AtNCED6/9表达量显著降低(Zhou et al., 2018). ...
Modulation of abscisic acid signal transduction and biosynthesis by an smlike protein in Arabidopsis 2 2001
... ABA合成最后2步是胞质溶胶中的黄质醛被短链脱氢酶/还原酶(short chain dehydrogenase/reductase-like, SDR1)还原为ABA醛(ABA aldehyde), 继而被ABA醛氧化酶(ABA-aldehyde oxidase, AAO)和钼辅因子硫化酶(molybdenum cofactor sulfurase, MoCo)转化为活性ABA.拟南芥仅有1个AtABA/SDR1 (abscisic acid 2)基因, 在所有器官中组成型表达, 且不受ABA、盐和甘露醇诱导.aba2突变体内源ABA含量减少、种子不休眠、植株呈萎蔫状且主根变短, 外源ABA可恢复其主根根长; 而AtABA2过表达株系的表型与之相反(Lin et al., 2007).拟南芥有4个AAO基因(AtAAO1-4), AtAAO1和AtAAO4分别在干种子和发育中的角果大量表达; AtAAO3是种子和失水叶片中ABA合成关键酶, aao3突变体植株呈萎蔫状、种子休眠程度略有降低(Seo et al., 2004).番茄sitiens突变体不能将ABA醛转化为活性ABA, 导致内源ABA缺失而呈萎蔫状, AtAAO3异位表达能恢复番茄sitiens表型(Harrison et al., 2011).豌豆(Pisum sativum)中有3个PsAO基因, 重组PsAO3蛋白显示AAO酶活性, 干旱使豌豆叶和根的PsAO3基因表达量增加(Zdunek-Zastocka and Sobczak, 2013).拟南芥AtLOS5/ABA3、番茄SiFLACCA和烟草NpABA1/ CKP1基因编码MoCo酶, 可将ABA醛转化为ABA (Endo et al., 2014).AtLOS5/ABA3在所有器官中均表达, 受干旱、盐和ABA诱导, los5突变体内源ABA含量减少、叶片蒸腾速率增加, 抗冷、抗盐及抗旱性减弱, 但AtLOS5过表达可提高转基因棉花(Gossy- pium hirsutum)的内源ABA水平和抗旱性(Xiong et al., 2001; Yue et al., 2012). ...
... 植物LSM (like-Sm protein)和XERICO (greek for ‘drought tolerant’)蛋白分别在mRNA代谢和蛋白水平调控内源ABA水平.LSMs是真核生物中高度保守的蛋白家族, 参与RNA代谢(如剪切、脱帽和降解).拟南芥sad1 (supersensitive to ABA and drought)突变体对干旱和外源ABA敏感性增强, sad1植物缺失ABA合成的正反馈调节和干旱诱导的ABA合成, 图位克隆确定SAD1基因编码LSM5蛋白(Xiong et al., 2001).转录组分析表明, SAD1/LSM5蛋白可动态调控拟南芥转录组前体mRNA剪切效率和剪切位点的识别, 表明SAD1/LSM5可能通过调控胁迫基因的表达而介导植物胁迫反应(Cui et al., 2014).LSM1-7复合体通过促进mRNA脱帽和随后的5'-3'降解参与精确的mRNA流转(turnover)和P-bodies形成.当拟南芥暴露在冷和高盐环境时, LSM1-7复合体通过差异化调控AtNCED3和AtNCED5 mRNA流转而调节内源ABA水平(Perea-Resa et al., 2016).拟南芥XERICO蛋白C端含RING-H2锌指基序, XERICO表达上调能显著提高细胞ABA水平并增强植物的抗旱性, 已知很多RING-H2蛋白具有E3泛素连接酶功能, 而酵母双杂交实验显示, AtUBC8 (E2 ubiquitin-conjugating enzyme)和AtTLP9 (ASK1-interacting F-box protein)是XERICO潜在的互作蛋白, 推测XERICO在蛋白水平调节细胞ABA水平(Ko et al., 2006). ...
Cloning and characterization of the abscisic acid-specific glucosyltransferase gene from adzuki bean seedlings 1 2002
... 内源ABA水平还受到ABA特异的葡萄糖基转移酶(ABA-uridine diphosphate glucosyltransferase, ABAUGT)和β-葡萄糖苷酶(β-glucosidase, BG)介导的可逆糖基化代谢调节.研究人员从赤豆(V. angularis)中克隆了编码ABA-UGT酶的基因AOG, UGT可催化ABA与UDPG结合生成ABA葡萄糖苷酯(ABA-glucoside ester, ABA-GE); ABA-GE缺乏生物学活性, 在液泡或质外体中积累(Xu et al., 2002; Priest et al., 2006).拟南芥UGT家族的E亚家族中UGT71B6/7/8参与调节内源ABA水平, 影响AtCYP707A1-4表达, 在脱水和高盐胁迫以及种子萌发和生长发育进程中发挥重要调节作用(Dong et al., 2014).此外, AtUGT71C5也可催化ABA转化为ABA-GE, 与野生型相比, ugt71c5突变体的种子萌发延迟且叶片气孔变小, 抗旱性增强(Liu et al., 2015). ...
A vacuolar β-glucosidase homolog that possesses glucose-conjugated abscisic acid hydrolyzing activity plays an important role in osmotic stress responses in Arabidopsis 1 2012
... 干旱条件下, 液泡或质外体贮存的ABA-GE迅速被BG水解, 生成活性ABA以适应变化的环境.拟南芥AtBG1在内质网将ABA-GE水解生成ABA, bg1突变体内源ABA含量和抗旱性降低; 脱水迅速诱导BG1蛋白多聚化后显著增强BG活性, 从而加速ABA生成, ABA正反馈激活BG1蛋白多聚化(Lee et al., 2006).AtBG2以复合物形式储存在中央液泡, 脱水诱导AtBG2表达, bg2突变体对脱水和盐胁迫敏感, AtBG2过表达可互补bg1表型(Xu et al., 2012).在低湿环境下, 拟南芥aba3-1突变体中AtBG1转录水平极显著高于野生型, 说明细胞内ABA-GE水解机制可部分补偿ABA从头合成的缺陷(Bauer et al., 2013).此外, AtBGLU10编码的BG酶分布在液泡中, bglu10突变体的BG酶活性和ABA含量均降低, 且叶面温度和胁迫反应基因表达量也降低, 而AtBGLU10过表达株系抗旱性增强(Wang et al., 2011a). ...
NIN-like protein 8 is a master regulator of nitrate-promoted seed germination in Arabidopsis 1 2016
... 已有研究表明若干转录因子参与ABA代谢基因的表达调控.拟南芥CHO1 (CHOTTO1)为AP2/ERF型转录因子, 促进AtABA1和AtNCED9、抑制AtCYP707A2/3转录, 调控吸胀期种子ABA抑制的赤霉素合成(Yano et al., 2009).AP2/ERF型转录因子ABI4促进ABA积累和种子休眠, 通过结合AtCYP707A1/2启动子并抑制其转录来实现(Shu et al., 2013).拟南芥NLP8 (NIN-like protein 8)以硝酸盐依赖的方式结合AtCYP707A2的启动子并激活其转录, 降低种子ABA水平, 促进种子萌发(Matakiadis et al., 2009; Yan et al., 2016).bZIP家族转录因子VIP1 (VirE2-interacting protein 1)为渗透胁迫的调节因子, VIP1直接结合AtCYP707A1/3启动子并激活其转录(Tsugama et al., 2012).拟南芥bHLH122是干旱、NaCl和渗透胁迫的正调节因子, bHLH122结合AtCYP707A3启动子G-/E-box元件并抑制其表达, 进而抑制ABA代谢(Liu et al., 2014).拟南芥成花抑制因子SVP (short vegetative phase)直接结合AtCYP707A1/3和AtBG1启动子的CArG Motif元件, 抑制AtCYP707A1/3, 促进AtBG1转录, 表明SVP是干旱胁迫下ABA代谢的重要调控因子(Wang et al., 2018). ...
A NAP-AAO3 regulatory module promotes chlorophyll degradation via ABA biosynthesis in Arabidopsis leaves 1 2014
... 已有研究表明, 植物多个转录因子家族参与ABA合成酶基因的转录调控, 其中MYB、AP2/ERF、WRKY、NAC、bHLH和NGA等家族的转录因子促进ABA合成基因转录, 而NFX和HD-ZIP等转录因子抑制ABA合成基因的表达.例如, 拟南芥AtMYB96通过结合AtNCED2/6启动子促进ABA合成和种子休眠(Lee et al., 2015).AP2/ERF (APETALA2/ethylene-responsive factor)家族的AtORA47 (octadecanoid-responsive AP2/ERF-domain transcription factor 47)结合AtNCED3/9启动子的O-box元件, 促进损伤时ABA合成与信号转导(Chen et al., 2016); 而AtDREB2C (dehydration-responsive element binding factor)通过结合AtNCED9启动子的DRE元件激活高温胁迫下种子的ABA合成(Je et al., 2014).拟南芥AtWRKY57结合AtNCED3启动子, 促进干旱诱导的ABA合成、提高植物的耐旱性(Jiang et al., 2012).NAC转录因子中的AtATAF1结合AtNCED3启动子, 促进ABA合成(Jensen et al., 2013).AtNAP (NAC-like, activated by AP3/PI)结合AtAAO3启动子, 促进衰老叶片ABA合成与叶绿素降解(Yang et al., 2014).与AtNAP同源的谷子 (Setaria italica) SiNAC1促进ABA合成, 是叶片衰老的正调节因子(Ren et al., 2018).AtbHLH68过表达株系的抗旱性增强、植株侧根减少, 推测AtbHLH68通过ABA依赖途径调节ABA代谢和/或信号(Le Hir et al., 2017).NGAs (NGA THAs)家族转录因子AtNGA1结合AtNCED3启动子的NBE (NGA-binding element)顺式元件, 通过激活AtNCED3转录促进干旱诱导的ABA合成(Sato et al., 2018).拟南芥AtNFXL2 (nuclear transcription factor X-box binding protein-like 2)抑制内源ABA合成和信号, atnfxl2缺失突变体气孔导度降低, 耐旱性增强; AtNFLX2同工型AtNFLX2-78直接抑制SHINE (SHINE1/SHN1, SHN2, SHN3)基因和BDG1 (BODYGUARD1)基因转录, SHINE与表皮蜡质合成相关, BDG1调控表皮角质层发育, 二者同时促进胁迫环境下植物ABA的合成(Lisso et al., 2011, 2012).除正反馈调节外, ABA合成还存在负反馈调节, 如花生(Arachis hypogaea) AhNAC2和AhAREB1蛋白复合体介导ABA合成的负反馈调节, 外源ABA促进AhNAC2和AhAREB1结合到AhNCED1启动子, 进而抑制AhNCED1转录, 其中AhAREB1是主要负调节因子(Liu et al., 2016).HD-ZIPII转录因子AtHAT1 (homeodomain-leucine zipper protein 1)结合AtABA3和AtNCED3的启动子, 抑制ABA合成和信号(Tan et al., 2018). ...
CHOTTO1, a putative double APETALA2 repeat transcription factor, is involved in abscisic acid-mediated repression of gibberellin biosynthesis during seed germination in Arabidopsis 1 2009
... 已有研究表明若干转录因子参与ABA代谢基因的表达调控.拟南芥CHO1 (CHOTTO1)为AP2/ERF型转录因子, 促进AtABA1和AtNCED9、抑制AtCYP707A2/3转录, 调控吸胀期种子ABA抑制的赤霉素合成(Yano et al., 2009).AP2/ERF型转录因子ABI4促进ABA积累和种子休眠, 通过结合AtCYP707A1/2启动子并抑制其转录来实现(Shu et al., 2013).拟南芥NLP8 (NIN-like protein 8)以硝酸盐依赖的方式结合AtCYP707A2的启动子并激活其转录, 降低种子ABA水平, 促进种子萌发(Matakiadis et al., 2009; Yan et al., 2016).bZIP家族转录因子VIP1 (VirE2-interacting protein 1)为渗透胁迫的调节因子, VIP1直接结合AtCYP707A1/3启动子并激活其转录(Tsugama et al., 2012).拟南芥bHLH122是干旱、NaCl和渗透胁迫的正调节因子, bHLH122结合AtCYP707A3启动子G-/E-box元件并抑制其表达, 进而抑制ABA代谢(Liu et al., 2014).拟南芥成花抑制因子SVP (short vegetative phase)直接结合AtCYP707A1/3和AtBG1启动子的CArG Motif元件, 抑制AtCYP707A1/3, 促进AtBG1转录, 表明SVP是干旱胁迫下ABA代谢的重要调控因子(Wang et al., 2018). ...
Overexpression of the AtLOS5 gene increased abscisic acid level and drought tolerance in transgenic cotton 1 2012
... ABA合成最后2步是胞质溶胶中的黄质醛被短链脱氢酶/还原酶(short chain dehydrogenase/reductase-like, SDR1)还原为ABA醛(ABA aldehyde), 继而被ABA醛氧化酶(ABA-aldehyde oxidase, AAO)和钼辅因子硫化酶(molybdenum cofactor sulfurase, MoCo)转化为活性ABA.拟南芥仅有1个AtABA/SDR1 (abscisic acid 2)基因, 在所有器官中组成型表达, 且不受ABA、盐和甘露醇诱导.aba2突变体内源ABA含量减少、种子不休眠、植株呈萎蔫状且主根变短, 外源ABA可恢复其主根根长; 而AtABA2过表达株系的表型与之相反(Lin et al., 2007).拟南芥有4个AAO基因(AtAAO1-4), AtAAO1和AtAAO4分别在干种子和发育中的角果大量表达; AtAAO3是种子和失水叶片中ABA合成关键酶, aao3突变体植株呈萎蔫状、种子休眠程度略有降低(Seo et al., 2004).番茄sitiens突变体不能将ABA醛转化为活性ABA, 导致内源ABA缺失而呈萎蔫状, AtAAO3异位表达能恢复番茄sitiens表型(Harrison et al., 2011).豌豆(Pisum sativum)中有3个PsAO基因, 重组PsAO3蛋白显示AAO酶活性, 干旱使豌豆叶和根的PsAO3基因表达量增加(Zdunek-Zastocka and Sobczak, 2013).拟南芥AtLOS5/ABA3、番茄SiFLACCA和烟草NpABA1/ CKP1基因编码MoCo酶, 可将ABA醛转化为ABA (Endo et al., 2014).AtLOS5/ABA3在所有器官中均表达, 受干旱、盐和ABA诱导, los5突变体内源ABA含量减少、叶片蒸腾速率增加, 抗冷、抗盐及抗旱性减弱, 但AtLOS5过表达可提高转基因棉花(Gossy- pium hirsutum)的内源ABA水平和抗旱性(Xiong et al., 2001; Yue et al., 2012). ...
Expression of Pisum sativum PsAO3 gene, which encodes an aldehyde oxidase utilizing abscisic aldehyde, is induced under progressively but not rapidly imposed drought stress 1 2013
... ABA合成最后2步是胞质溶胶中的黄质醛被短链脱氢酶/还原酶(short chain dehydrogenase/reductase-like, SDR1)还原为ABA醛(ABA aldehyde), 继而被ABA醛氧化酶(ABA-aldehyde oxidase, AAO)和钼辅因子硫化酶(molybdenum cofactor sulfurase, MoCo)转化为活性ABA.拟南芥仅有1个AtABA/SDR1 (abscisic acid 2)基因, 在所有器官中组成型表达, 且不受ABA、盐和甘露醇诱导.aba2突变体内源ABA含量减少、种子不休眠、植株呈萎蔫状且主根变短, 外源ABA可恢复其主根根长; 而AtABA2过表达株系的表型与之相反(Lin et al., 2007).拟南芥有4个AAO基因(AtAAO1-4), AtAAO1和AtAAO4分别在干种子和发育中的角果大量表达; AtAAO3是种子和失水叶片中ABA合成关键酶, aao3突变体植株呈萎蔫状、种子休眠程度略有降低(Seo et al., 2004).番茄sitiens突变体不能将ABA醛转化为活性ABA, 导致内源ABA缺失而呈萎蔫状, AtAAO3异位表达能恢复番茄sitiens表型(Harrison et al., 2011).豌豆(Pisum sativum)中有3个PsAO基因, 重组PsAO3蛋白显示AAO酶活性, 干旱使豌豆叶和根的PsAO3基因表达量增加(Zdunek-Zastocka and Sobczak, 2013).拟南芥AtLOS5/ABA3、番茄SiFLACCA和烟草NpABA1/ CKP1基因编码MoCo酶, 可将ABA醛转化为ABA (Endo et al., 2014).AtLOS5/ABA3在所有器官中均表达, 受干旱、盐和ABA诱导, los5突变体内源ABA含量减少、叶片蒸腾速率增加, 抗冷、抗盐及抗旱性减弱, 但AtLOS5过表达可提高转基因棉花(Gossy- pium hirsutum)的内源ABA水平和抗旱性(Xiong et al., 2001; Yue et al., 2012). ...
A DTX/MATE-type transporter facilitates abscisic acid efflux and modulates ABA sensitivity and drought tolerance in Arabidopsis 1 2014
... AtABCG40/PDR12 (pleiotropic drug resistance 12)定位在保卫细胞和其它细胞的质膜, abcg40突变体原生质体对ABA吸收能力显著下降(Kang et al., 2010).AtABCG25是ABA输出转运蛋白, AtABCG25在维管组织中表达, 过表达植株保卫细胞ABA信号增强、植株水分利用率提高(Kuromori et al., 2010, 2016).AtNRT1.2/AIT1介导质膜ABA输入转运, nrt1.2/ait1突变体对外源ABA敏感性降低(Kanno et al., 2012).AtABCG40和AtNRT1.2/AIT1介导细胞吸收ABA的空间表达谱不同, AtABCG40在保卫细胞中表达, 而AtNRT1.2/AIT1在维管细胞中表达; AtNRT1.2/AIT1与AtABCG25共同调节维管组织向保卫细胞转运ABA, 且AtNRT1.2/AIT1介导ABA进入维管组织可能与植物通过木质部和韧皮部进行ABA再分配相关(Kuromori et al., 2018).拟南芥MATE (DTX/multidrug and toxin compound extrusion)家族成员DTX50蛋白定位在维管细胞和保卫细胞质膜, 介导维管细胞和保卫细胞向外转运ABA; 外源ABA促进DTX50表达, dtx50突变体对ABA敏感、气孔导度较低、抗旱性增强(Zhang et al., 2014).AtABCG22定位于质膜, 具有ABA转运功能, 参与ABA转运、合成和信号转导(Kuromori et al., 2011).此外, 研究表明AtABCG25/31/30/40协同参与种子的ABA转运, 其中胚乳表达的AtABCG25/31为ABA输出转运蛋白, 协同将胚乳细胞合成的ABA转运出去, 而种胚表达的AtABCG30/40为输入转运蛋白, 共同将ABA转运入种胚细胞(Kang et al., 2015). ...
Arabidopsis IQM4, a novel calmodulin-binding protein, is involved with seed dormancy and germination in Arabidopsis 1 2018
... 此外, 硫元素供应、角质层合成基因、钙调素信号蛋白也参与ABA生物合成的调控.叶绿体硫转运蛋白AtSULTR3;1功能缺失显著降低幼苗AAO酶活性和内源ABA水平, 硫元素供应可提高AtNCED3和AtABA3转录水平; ABA促进AtSULTR3;1和其它硫代谢基因转录, 表明硫在植物响应环境胁迫时发挥重要作用(Cao et al., 2014).干旱使木质部汁液硫酸盐含量增加, 进而促进保卫细胞AtNCED3表达和ABA合成, 引起气孔关闭(Malcheska et al., 2017).已知植物角质层作为物理屏障减少水分流失并介导ABA合成和信号转导, 进而调控植物渗透胁迫反应.拟南芥ced1 (9-cis-epoxycarotenoid dioxygenase defective 1)突变体对轻微渗透胁迫极为敏感, CED1/BDG1基因促进ABA合成基因的表达(Wang et al., 2011b).植物钙调素类似蛋白AtCML37 (calmodulin-like 37)和AtCML42拮抗调控干旱胁迫诱导的ABA积累, AtCML37为正调节因子, 而AtCML42为负调节因子(Scholz et al., 2015).我们也发现钙调素结合蛋白AtIQM4能促进种子ABA合成, iqm4突变体种子ABA含量降低, 角果AtNCED6/9表达量显著降低(Zhou et al., 2018). ...
Glucose-induced delay of seed germination in rice is mediated by the suppression of ABA catabolism rather than an enhancement of ABA biosynthesis 1 2009
... Schwartz等(1997)证明重组的VP14蛋白可切割9′-顺式-紫黄质形成黄质醛, 推测VP14具有NCED功能.目前, 已证明植物NCED蛋白是一个多基因家族, 先后从水稻(OsNCED1-5)、拟南芥(AtNCED2/3/5/6/9)、四季豆(Phaseolus vulgaris) (PvNCED1)、鳄梨(Persea americana) (PaNCED1/3)以及菟丝子(Cuscuta reflexa) (CrNCED1/2)中克隆到NCED基因(Endo et al., 2014).水稻OsNCED1在叶片中表达量最高, OsNCED2主要在种子中表达, OsNCED5参与水稻籽粒灌浆期ABA的合成, OsNCED3转基因拟南芥ABA含量增加, 叶片形态发生改变(Zhu et al., 2009).拟南芥基因组有9个VP14同源序列(AtNCED1-9), 其中AtNCED2/3/5/6/9编码蛋白具有NCED酶活性, AtNCED2/3在植物根和叶中表达量较高.干旱迅速诱导AtNCED3基因表达并促进ABA合成, AtNCED3过表达株系的内源ABA水平较高、叶片蒸腾速率较低、抗旱性较强, 且ABA响应基因的转录本增加(任慧波等, 2007; Hao et al., 2009).种子成熟晚期AtNCED5/6/9表达显著增强, AtNCED6在胚乳中表达, AtNCED5/9在胚乳和胚中表达.nced6/nced9双突变体种子内源ABA含量和休眠程度显著降低, 说明AtNCED6/9主要调控种子休眠(Tan et al., 2003; Lefebvre et al., 2006).nced3/nced5双突变体内源ABA含量减少、营养器官变小且植株抗旱性降低, 说明AtNCED3/5参与植物生长发育和胁迫响应(Frey et al., 2012). ...
The dynamic regulation mechanism of the endogenous ABA in plant 0 2019
 ,*广州大学生命科学学院, 植物抗逆基因功能研究广州市重点实验室, 广州 510006
,*广州大学生命科学学院, 植物抗逆基因功能研究广州市重点实验室, 广州 510006
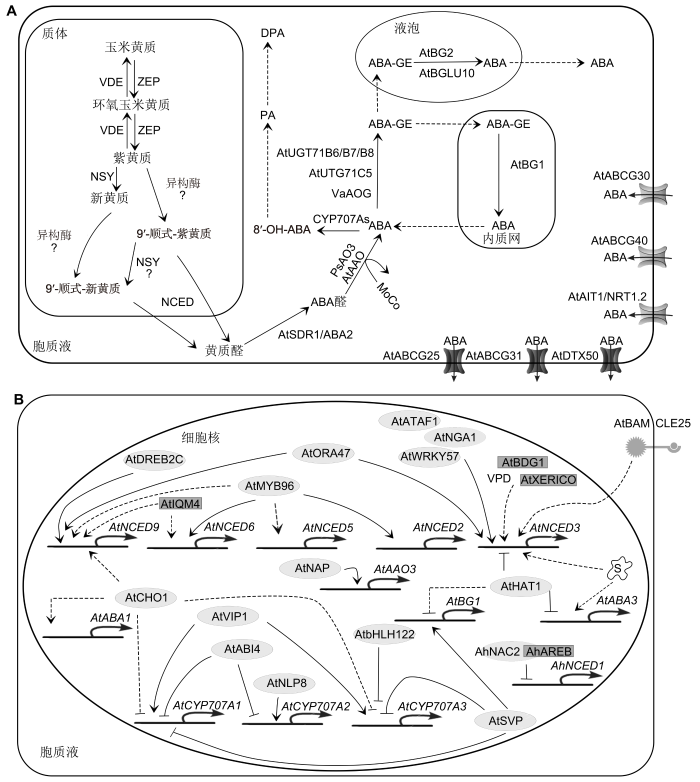 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}