Expression Patterns and Bioinformatic Analyses of Salt Stress Responsive Gene IbMYB3 in Ipomoea batatas
Ge Li, Xiaoqing Meng, Zongyun Li, Mingku Zhu,*Key Lab of Biotechnology for Medicinal Plants of Jiangsu Province/School of Life Science, Jiangsu Normal University, Xuzhou 221116, China
Abstract MYB transcription factors have multiple biological functions and play important roles in mediating plant responses to biotic and abiotic stresses. In this paper, two MYB genes, named IbMYB3 and IbMYB4, which were significantly induced by salt stress, were screened from RNA-seq data of salt-stressed sweetpotato (Ipomoea batatas) plantlets. Gene expression analysis showed that the expression of IbMYB3 was significantly up-regulated by various abiotic stresses and plant growth substances treatments, suggesting that IbMYB3 might be involved in the abiotic stress responses of sweetpotato. Further bioinformatic analysis showed that the open reading frame of IbMYB3 is 1 059 bp in length, encoding 353 amino acids, with a predicted molecular weight of 39.41 kDa and the theoretical isoelectric point (PI) of 5.26, which is an acidic negatively charged hydrophilic protein. Subcellular localization showed that the IbMYB3 protein localizes to the nucleus, and has strong transcriptional activation activity. Taken together, these results demonstrated that the IbMYB3 transcription factor might play an important role in regulating the abiotic stress responses of sweetpotato. This study thus lays the foundation for further illustration of IbMYB3 function. Keywords:bioinformatic analysis;expressional patterns;IbMYB3; Ipomoea batatas;transcriptional activation
PDF (8533KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 李格, 孟小庆, 李宗芸, 朱明库. 甘薯盐胁迫响应基因IbMYB3的表达特征及生物信息学分析. 植物学报, 2020, 55(1): 38-48 doi:10.11983/CBB19094 Li Ge, Meng Xiaoqing, Li Zongyun, Zhu Mingku. Expression Patterns and Bioinformatic Analyses of Salt Stress Responsive Gene IbMYB3 in Ipomoea batatas. Chinese Bulletin of Botany, 2020, 55(1): 38-48 doi:10.11983/CBB19094
甘薯(Ipomoea batatas)是世界第七、中国第四大重要粮食作物, 主要用于粮食、饲料、淀粉制造和乙醇生产等(Liu, 2017)。随着人口的增加和资源需求的增长, 甘薯研究逐渐被重视, 如国家种质甘薯试管苗库的建立、甘薯种质资源中心及各级甘薯研究平台的成立。通过传统杂交育种进行甘薯改良已不能满足现代分子育种的要求, 通过基因工程手段辅助甘薯品种定向改良, 培育优质、高产甘薯新品种对甘薯种植和深化应用具有重要意义(杨俊等, 2011; 李志亮等, 2013)。Liu等(2015)从耐盐甘薯品种ND98中分离出1个新的盐诱导的S-腺苷甲硫氨酸依赖性甲基转移酶基因, 命名为IbSIMT1。盐胁迫和ABA处理能够诱导IbSIMT1基因上调表达, 与野生型相比过表达IbSIMT1的甘薯耐盐性明显增强。甘薯中肌醇-磷酸合成酶基因IbMIPS1的表达受NaCl、聚乙二醇(PEG)和ABA显著诱导, 在甘薯中过表达IbMIPS1可明显增强转基因甘薯的耐盐性和抗旱性(Zhai et al., 2016)。当前对IbMYB1的研究比较广泛, 已证实其具有多种功能, 如异源超表达IbMYB1的马铃薯(Solanum tuberosum)盐胁迫耐受性增强(Cheng et al., 2013); Park等(2015)在甘薯中超表达IbMYB1可提高甘薯的抗氧化能力; 在拟南芥(Arabidopsis thaliana)和烟草(Nicotiana tabacum)中异源超表达IbMYB1均可促进花青素积累(An et al., 2015)。
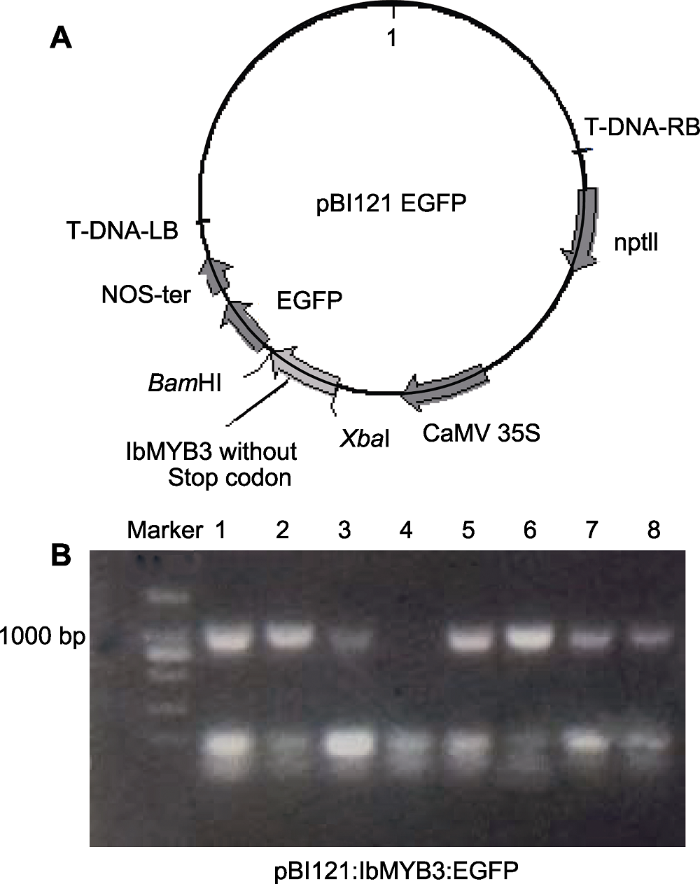
(A) Vector map of pBI121:IbMYB3:EGFP fusion protein construction; (B) Detection in Agrobacterium (Lanes 1-8 represent the PCR identification results after the vector was transferred into Agrobacterium, respectively).
Figure 2Construction of pGBKT7:IbMYB3 vector for transcriptional activation
1.2.7 IbMYB3与其它物种中盐胁迫相关MYB蛋白的氨基酸序列比对分析
从NCBI (National Center for Biotechnology Information, https://www.ncbi.nlm.nih.gov/)和TAIR (The Arabidopsis Information Resource, http://www.arabidopsis.org/index.jsp)数据库中分别下载不同物种中已报道或已被证实正调控盐胁迫耐受的MYB氨基酸序列, 通过DNAMAN和MEGA软件分别进行多重序列比对和系统发生树分析。
(A) IbMYB3与EGFP融合蛋白在洋葱下表皮细胞中的亚细胞定位; (B) IbMYB3的转录激活活性分析。Bars=4 μm Figure 6Subcellular localization in onion epidermal cells and transcriptional activation activity analysis of IbMYB3 protein
(A) Subcellular localization of IbMYB3 and EGFP fusion protein in the onion lower epidermis cells; (B) Transcriptional activation activity analysis of IbMYB3 protein. Bars=4 μm
2.5 蛋白序列比对及系统发生树分析
IbMYB3蛋白具有MYB蛋白家族R2R3亚家族成员典型的2个保守结构域(图7A), 且Xu22中IbMYB3蛋白与甘薯近缘野生种I. triloba中的ItMYB41和牵牛花(Ipomoea nil)中的InMYB330序列相似性最高(分别达98.6%和93.4%) (图7B)。此外, IbMYB3与拟南芥中一些已知功能的MYB转录因子序列也具有很高的相似性。例如, 与拟南芥AtMYB41蛋白的相似性达62.6%, 具有较高的同源性。此外, 我们进一步对比了IbMYB3与其它物种MYB蛋白中R2R3结构域的保守性, 结果显示IbMYB3与ItMYB41、InMYB330、AtMYB41、AtMYB74和AtMYB102中的R2R3结构域的相似性均很高(分别达99.1%、97.2%、91.7%、92.6%和90.7%), 表明负责与DNA结合的R2R3结构域在不同物种中均具有很高的保守性, 暗示IbMYB3可能与已报道的几个拟南芥MYB基因具有类似的功能。已有研究表明, AtMYB41基因能够通过调控基因转录及物质代谢来响应渗透胁迫, 从而应对盐、冷和干旱(Lippold et al., 2009); AtMYB41可在多个物种或细胞类型中激活软木脂的合成, 而软木脂在调节植物水分和保护植物免受生物和非生物胁迫方面发挥关键作用(Kosma et al., 2014); 超表达AtMYB41基因则能调控细胞扩增和角质层沉积以响应非生物胁迫(Cominelli et al., 2008)。此外, 研究表明AtMYB102能响应渗透胁迫(Denekamp and Smeekens, 2003)以及AtMYB74能够被盐诱导(Xu et al., 2015)。综上表明, IbMYB3基因可能参与甘薯非生物胁迫响应, 但需要进一步实验验证。
(A) IbMYB3与其它非甘薯物种中盐胁迫耐受相关MYB蛋白序列比对; (B) IbMYB3与其它物种中盐胁迫耐受相关MYB蛋白的系统发生树 Figure 7MYB proteins sequence alignment and phylogenetic tree
(A) Proteins sequence alignment between IbMYB3 and the MYB proteins associated with salt stress tolerance in species other than sweetpotato; (B) Phylogenetic tree of IbMYB3 and the MYB proteins associated with salt stress tolerance in other species
2.6 讨论
甘薯具有耐盐碱、抗旱和耐贫瘠等特性, 可种植于滩涂瘠薄土地, 不存在“与粮争地”的问题, 且单位面积、单位时间内生产的乙醇比水稻(Oryza sativa)和小麦等主要粮食作物多, 因此成为适宜用于挖掘盐碱地生产潜力的能源作物之一(朱明库等, 2017)。相比水稻和小麦等大作物, 目前甘薯耐盐等非生物胁迫的分子生物学研究还有很大差距(Liu, 2017)。甘薯采用薯苗繁殖, 作为幼嫩组织, 薯苗对土壤盐碱胁迫较敏感, 给甘薯生产带来一定困难(代红军等, 2001)。因此, 深入研究甘薯的耐盐特性, 对甘薯种植和深化应用以及我国生物能源发展具有重要意义。Yang等(2017)通过半单倍型基因组测序法首次成功解析了六倍体栽培种甘薯泰中6号的全基因组信息, 为甘薯中重要基因克隆和功能解析提供了极大便利。过去几十年, 甘薯基因工程已经取得重要进展, 已建立根癌农杆菌介导的转化系统并成功用于多种甘薯基因型(Liu, 2017)。目前已从甘薯中分离和鉴定了许多参与非生物胁迫耐受性(Bian et al., 2014; Wang et al., 2016)、病虫害抗性(柏洁, 2014)、淀粉合成(李丰, 2017)及花青素合成(An et al., 2015)等相关基因。
本研究通过RNA-seq分析筛选出2个受盐胁迫显著诱导的甘薯MYB转录因子(IbMYB3和IbMYB4)。进一步检测2个基因在多种非生物胁迫和植物生长物质处理下的表达水平, 确定对盐胁迫下被显著诱导的IbMYB3进行深入研究。生物信息学分析表明, IbMYB3蛋白属于R2R3-MYB亚家族成员, 在其N端含有高度保守的R2、R3重复单元。IbMYB3蛋白的亚细胞定位以及激活活性结果表明, IbMYB3定位于细胞核, 且具有转录激活活性。基于同源比对及系统进化树分析, 我们发现IbMYB3与已有研究的AtMYB41有较高相似性, 推测其可能具有与AtMYB41类似的功能。前人的研究表明, AtMYB41能够通过调控基因转录及物质代谢来响应渗透胁迫, 从而应对盐、冷和干旱胁迫(Cominelli et al., 2008; Lippold et al., 2009)。此外, 与IbMYB3亲缘关系较近的AtMYB74 (Xu et al., 2015)和AtMYB102 (Denekamp and Smeekens, 2003)均被报道具有增强盐耐受性的功能, 因此推测IbMYB3蛋白可能也参与甘薯逆境响应过程。
由于甘薯遗传转化周期较长, 因此许多研究者将甘薯相关基因转化到其它模式物种中开展相关研究, 如拟南芥(Chu et al., 2013; Kang et al., 2018)、烟草(An et al., 2015)和马铃薯(Cheng et al., 2013)。本研究后期已将IbMYB3在拟南芥中进行异源超表达并鉴定其盐胁迫耐受性。初步结果显示, 与野生型相比, 转基因拟南芥叶的长、宽和面积均大于野生型。植物叶片作为营养器官, 是植物进行光合作用的主要场所, 叶面积大表明转基因株系在单位时间内能够产生更多的生物量, 从而促进植物的生长发育。此外, 盐处理条件下, IbMYB3异源超表达拟南芥植株的萌发率高于野生型(待发表), 表明IbMYB3基因确实参与了对盐胁迫的响应。IbMYB3基因异源超表达增强了拟南芥对盐胁迫的耐受性, 表明该基因具有增强甘薯盐耐受的潜能, 后续可进一步探究其在甘薯响应盐胁迫中的功能。
AmbawatS, SharmaP, YadavNR, YadavRC (2013). MYB transcription factor genes as regulators for plant responses: an overview Physiol Mol Biol Plants 19, 307-321. [本文引用: 1]
AnCH, LeeKW, LeeSH, JeongYJ, WooSG, ChunH, ParkYI, KwakSS, KimCY (2015). Heterologous expression of IbMYB1a by different promoters exhibits different patterns of anthocyanin accumulation in tobacco Plant Physiol Biochem 89, 1-10. [本文引用: 3]
BaumannK, Perez-RodriguezM, BradleyD, VenailJ, BaileyP, JinHL, KoesR, RobertsK, MartinC (2007). Control of cell and petal morphogenesis by R2R3 MYB transcription factors Development 134, 1691-1701. [本文引用: 1]
BianXF, XieYZ, GuoXD, JiaZD, MaPY (2014). Research advance on molecular mechanism of abiotic and biotic stress resistance in sweet potato Agric Sci Technol 15, 901-906, 941. [本文引用: 1]
ChengYJ, KimMD, DengXP, KwakSS, ChenW (2013). Enhanced salt stress tolerance in transgenic potato plants expressing IbMYB1, a sweet potato transcription factor J Microbiol Biotechnol 23, 1737-1746. [本文引用: 2]
ChuH, JeongJC, KimWJ, ChungDM, JeonHK, AhnYO, KimSH, LeeHS, KwakSS, KimCY (2013). Expression of the sweetpotato R2R3-type IbMYB1a gene induces anthocyanin accumulation in Arabidopsis Physiol Plant 148, 189-199. [本文引用: 1]
CominelliE, SalaT, CalviD, GusmaroliG, TonelliC (2008). Over-expression of the Arabidopsis AtMYB41 gene alters cell expansion and leaf surface permeability Plant J 53, 53-64. [本文引用: 2]
DenekampM, SmeekensSC (2003). Integration of wounding and osmotic stress signals determines the expression of the AtMYB102 transcription factor gene Plant Physiol 132, 1415-1423. [本文引用: 2]
DuH, YangSS, LiangZ, FengBR, LiuL, HuangYB, TangYX (2012). Genome-wide analysis of the MYB transcription factor superfamily in soybean BMC Plant Biol 12, 106. [本文引用: 1]
GatesDJ, StricklerSR, MuellerLA, OlsonBJSC, SmithSD (2016). Diversification of R2R3-MYB transcription factors in the tomato family Solanaceae J Mol Evol 83, 26-37. [本文引用: 1]
JiaN, LiuJQ, SunYF, TanPH, CaoH, XieYY, WenBT, GuTY, LiuJM, LiMM, HuangYT, LuJ, JinN, SunLC, XinFJ, FanB (2018). Citrus sinensis MYB transcription factors CsMYB330 and CsMYB308 regulate fruit juice sac lignification through fine-tuning expression of the Cs4CL1 gene Plant Sci 277, 334-343. [本文引用: 1]
JungC, SeoJS, HanSW, KooYJ, KimCH, SongSI, NahmBH, ChoiYD, CheongJJ (2008). Overexpression of AtMYB44 enhances stomatal closure to confer abiotic stress tolerance in transgenic Arabidopsis Plant Physiol 146, 623-635. [本文引用: 1]
KangC, HeSZ, ZhaiH, LiRJ, ZhaoN, LiuQC (2018). A sweetpotato auxin response factor gene (IbARF5) is involved in carotenoid biosynthesis and salt and drought tolerance in transgenic Arabidopsis Front Plant Sci 9, 1307. [本文引用: 1]
KelleyLA, MezulisS, YatesCM, WassMN, SternbergMJE (2015). The Phyre2 web portal for protein modeling, prediction and analysis Nat Protoc 10, 845-858. [本文引用: 2]
KosmaDK, MurmuJ, RazeqFM, SantosP, BourgaultR, MolinaI, RowlandO (2014). AtMYB41 activates ectopic suberin synthesis and assembly in multiple plant species and cell types Plant J 80, 216-229. [本文引用: 1]
LippoldF, SanchezDH, MusialakM, SchlerethA, ScheibleWR, HinchaDK, UdvardiMK (2009). AtMyb41 regulates transcriptional and metabolic responses to osmotic stress in Arabidopsis Plant Physiol 149, 1761-1772. [本文引用: 2]
LiuCG, JunJH, DixonRA (2014). MYB5 and MYB14 play pivotal roles in seed coat polymer biosynthesis in Medicago truncatula Plant Physiol 165, 1424-1439. [本文引用: 1]
LiuDG, HeSZ, SongXJ, ZhaiH, LiuN, ZhangDD, RenZT, LiuQC (2015). IbSIMT1, a novel salt-induced methyltransferase gene from Ipomoea batatas, is involved in salt tolerance Plant Cell Tiss Organ Cult 120, 701-715. [本文引用: 1]
LiuQC (2017). Improvement for agronomically important traits by gene engineering in sweetpotato Breed Sci 67, 15-26. [本文引用: 3]
LotkowskaME, TohgeT, FernieAR, XueGP, BalazadehS, Mueller-RoeberB (2015). The Arabidopsis transcription factor MYB112 promotes anthocyanin formation during salinity and under high light stress Plant Physiol 169, 1862-1880. [本文引用: 1]
MuRL, CaoYR, LiuYF, LeiG, ZouHF, LiaoY, WangHW, ZhangWK, MaB, DuJZ, YuanM, ZhangJS, ChenSY (2009). An R2R3-type transcription factor gene AtMYB59 regulates root growth and cell cycle progression in Arabidopsis Cell Res 19, 1291-1304. [本文引用: 1]
NairR, RostB (2003). Better prediction of sub-cellular localization by combining evolutionary and structural information Proteins 53, 917-930. [本文引用: 1]
ParkSC, KimYH, JiCY, ParkS, JeongJC, LeeHS, KwakSS (2012). Stable internal reference genes for the normalization of Real-Time PCR in different sweetpotato cultivars subjected to abiotic stress conditions PLoS One 7, e51502. [本文引用: 1]
ParkSC, KimYH, KimSH, JeongYJ, KimCY, LeeJS, BaeJY, AhnMJ, JeongJC, LeeHS, KwakSS (2015). Overexpression of the IbMYB1 gene in an orange-fleshed sweet potato cultivar produces a dual-pigmented transgenic sweet potato with improved antioxidant activity Physiol Plant 153, 525-537. [本文引用: 1]
StrackeR, WerberM, WeisshaarB (2001). The R2R3- MYB gene family in Arabidopsis thaliana Curr Opin Plant Biol 4, 447-456. [本文引用: 1]
SunW, CaoZY, LiY, ZhaoYX, ZhangH (2007). A simple and effective method for protein subcellular localization using Agrobacterium-mediated transformation of onion epidermal cells Biologia 62, 529-532. [本文引用: 1]
WangB, ZhaiH, HeSZ, ZhangH, RenZT, ZhangDD, LiuQC (2016). A vacuolar Na+/H+ antiporter gene, IbNHX2, enhances salt and drought tolerance in transgenic sweetpotato Sci Hortic 201, 153-166. [本文引用: 1]
WuPP, PengMS, LiZG, YuanN, HuQ, FosterCE, SaskiC, WuGH, SunDF, LuoH (2019). DRMY1, a Myb-like protein, regulates cell expansion and seed production in Arabidopsis thaliana Plant Cell Physiol 60, 285-302. [本文引用: 1]
XuR, WangYH, ZhengH, LuW, WuCG, HuangJG, YanK, YangGD, ZhengCC (2015). Salt-induced transcription factor MYB74 is regulated by the RNA-directed DNA methylation pathway in Arabidopsis J Exp Bot 66, 5997-6008. [本文引用: 2]
ZhaiH, WangFB, SiZZ, HuoJX, XingL, AnYY, HeSZ, LiuQC (2016). A myo-inositol-1-phosphate synthase gene, IbMIPS1, enhances salt and drought tolerance and stem nematode resistance in transgenic sweet potato Plant Biotechnol J 14, 592-602. [本文引用: 1]
ZhangTQ, ZhaoYL, WangYC, LiuZY, GaoCQ (2018). Comprehensive analysis of MYB gene family and their expressions under abiotic stresses and hormone treatments in Tamarix hispida Front Plant Sci 9, 1303. [本文引用: 1]
ZhaoY, ChengXY, LiuXD, WuHF, BiHH, XuHX (2018). The wheat MYB transcription factor TaMYB 31 is involved in drought stress responses in Arabidopsis Front Plant Sci 9, 1426. [本文引用: 1]
ZhuZG, LiGR, LiuL, ZhangQT, HanZ, ChenXS, LiB (2019). A R2R3-MYB transcription factor, VvMYBC2L2, functions as a transcriptional repressor of anthocyanin biosynthesis in grapevine (Vitis vinifera L.) Molecules 24, 92. [本文引用: 1]
甘薯抗病相关基因SGT1的克隆与表达研究 1 2014
... 甘薯具有耐盐碱、抗旱和耐贫瘠等特性, 可种植于滩涂瘠薄土地, 不存在“与粮争地”的问题, 且单位面积、单位时间内生产的乙醇比水稻(Oryza sativa)和小麦等主要粮食作物多, 因此成为适宜用于挖掘盐碱地生产潜力的能源作物之一(朱明库等, 2017).相比水稻和小麦等大作物, 目前甘薯耐盐等非生物胁迫的分子生物学研究还有很大差距(Liu, 2017).甘薯采用薯苗繁殖, 作为幼嫩组织, 薯苗对土壤盐碱胁迫较敏感, 给甘薯生产带来一定困难(代红军等, 2001).因此, 深入研究甘薯的耐盐特性, 对甘薯种植和深化应用以及我国生物能源发展具有重要意义.Yang等(2017)通过半单倍型基因组测序法首次成功解析了六倍体栽培种甘薯泰中6号的全基因组信息, 为甘薯中重要基因克隆和功能解析提供了极大便利.过去几十年, 甘薯基因工程已经取得重要进展, 已建立根癌农杆菌介导的转化系统并成功用于多种甘薯基因型(Liu, 2017).目前已从甘薯中分离和鉴定了许多参与非生物胁迫耐受性(Bian et al., 2014; Wang et al., 2016)、病虫害抗性(柏洁, 2014)、淀粉合成(李丰, 2017)及花青素合成(An et al., 2015)等相关基因. ...
NACl胁迫下甘薯苗期叶片活性氧代谢与甘薯耐盐性的关系 1 2001
... 甘薯具有耐盐碱、抗旱和耐贫瘠等特性, 可种植于滩涂瘠薄土地, 不存在“与粮争地”的问题, 且单位面积、单位时间内生产的乙醇比水稻(Oryza sativa)和小麦等主要粮食作物多, 因此成为适宜用于挖掘盐碱地生产潜力的能源作物之一(朱明库等, 2017).相比水稻和小麦等大作物, 目前甘薯耐盐等非生物胁迫的分子生物学研究还有很大差距(Liu, 2017).甘薯采用薯苗繁殖, 作为幼嫩组织, 薯苗对土壤盐碱胁迫较敏感, 给甘薯生产带来一定困难(代红军等, 2001).因此, 深入研究甘薯的耐盐特性, 对甘薯种植和深化应用以及我国生物能源发展具有重要意义.Yang等(2017)通过半单倍型基因组测序法首次成功解析了六倍体栽培种甘薯泰中6号的全基因组信息, 为甘薯中重要基因克隆和功能解析提供了极大便利.过去几十年, 甘薯基因工程已经取得重要进展, 已建立根癌农杆菌介导的转化系统并成功用于多种甘薯基因型(Liu, 2017).目前已从甘薯中分离和鉴定了许多参与非生物胁迫耐受性(Bian et al., 2014; Wang et al., 2016)、病虫害抗性(柏洁, 2014)、淀粉合成(李丰, 2017)及花青素合成(An et al., 2015)等相关基因. ...
... 由于甘薯遗传转化周期较长, 因此许多研究者将甘薯相关基因转化到其它模式物种中开展相关研究, 如拟南芥(Chu et al., 2013; Kang et al., 2018)、烟草(An et al., 2015)和马铃薯(Cheng et al., 2013).本研究后期已将IbMYB3在拟南芥中进行异源超表达并鉴定其盐胁迫耐受性.初步结果显示, 与野生型相比, 转基因拟南芥叶的长、宽和面积均大于野生型.植物叶片作为营养器官, 是植物进行光合作用的主要场所, 叶面积大表明转基因株系在单位时间内能够产生更多的生物量, 从而促进植物的生长发育.此外, 盐处理条件下, IbMYB3异源超表达拟南芥植株的萌发率高于野生型(待发表), 表明IbMYB3基因确实参与了对盐胁迫的响应.IbMYB3基因异源超表达增强了拟南芥对盐胁迫的耐受性, 表明该基因具有增强甘薯盐耐受的潜能, 后续可进一步探究其在甘薯响应盐胁迫中的功能. ...
Expression of the sweetpotato R2R3-type IbMYB1a gene induces anthocyanin accumulation in Arabidopsis 1 2013
... 由于甘薯遗传转化周期较长, 因此许多研究者将甘薯相关基因转化到其它模式物种中开展相关研究, 如拟南芥(Chu et al., 2013; Kang et al., 2018)、烟草(An et al., 2015)和马铃薯(Cheng et al., 2013).本研究后期已将IbMYB3在拟南芥中进行异源超表达并鉴定其盐胁迫耐受性.初步结果显示, 与野生型相比, 转基因拟南芥叶的长、宽和面积均大于野生型.植物叶片作为营养器官, 是植物进行光合作用的主要场所, 叶面积大表明转基因株系在单位时间内能够产生更多的生物量, 从而促进植物的生长发育.此外, 盐处理条件下, IbMYB3异源超表达拟南芥植株的萌发率高于野生型(待发表), 表明IbMYB3基因确实参与了对盐胁迫的响应.IbMYB3基因异源超表达增强了拟南芥对盐胁迫的耐受性, 表明该基因具有增强甘薯盐耐受的潜能, 后续可进一步探究其在甘薯响应盐胁迫中的功能. ...
Over-expression of the Arabidopsis AtMYB41 gene alters cell expansion and leaf surface permeability 2 2008
... IbMYB3蛋白具有MYB蛋白家族R2R3亚家族成员典型的2个保守结构域(图7A), 且Xu22中IbMYB3蛋白与甘薯近缘野生种I. triloba中的ItMYB41和牵牛花(Ipomoea nil)中的InMYB330序列相似性最高(分别达98.6%和93.4%) (图7B).此外, IbMYB3与拟南芥中一些已知功能的MYB转录因子序列也具有很高的相似性.例如, 与拟南芥AtMYB41蛋白的相似性达62.6%, 具有较高的同源性.此外, 我们进一步对比了IbMYB3与其它物种MYB蛋白中R2R3结构域的保守性, 结果显示IbMYB3与ItMYB41、InMYB330、AtMYB41、AtMYB74和AtMYB102中的R2R3结构域的相似性均很高(分别达99.1%、97.2%、91.7%、92.6%和90.7%), 表明负责与DNA结合的R2R3结构域在不同物种中均具有很高的保守性, 暗示IbMYB3可能与已报道的几个拟南芥MYB基因具有类似的功能.已有研究表明, AtMYB41基因能够通过调控基因转录及物质代谢来响应渗透胁迫, 从而应对盐、冷和干旱(Lippold et al., 2009); AtMYB41可在多个物种或细胞类型中激活软木脂的合成, 而软木脂在调节植物水分和保护植物免受生物和非生物胁迫方面发挥关键作用(Kosma et al., 2014); 超表达AtMYB41基因则能调控细胞扩增和角质层沉积以响应非生物胁迫(Cominelli et al., 2008).此外, 研究表明AtMYB102能响应渗透胁迫(Denekamp and Smeekens, 2003)以及AtMYB74能够被盐诱导(Xu et al., 2015).综上表明, IbMYB3基因可能参与甘薯非生物胁迫响应, 但需要进一步实验验证. ...
... 本研究通过RNA-seq分析筛选出2个受盐胁迫显著诱导的甘薯MYB转录因子(IbMYB3和IbMYB4).进一步检测2个基因在多种非生物胁迫和植物生长物质处理下的表达水平, 确定对盐胁迫下被显著诱导的IbMYB3进行深入研究.生物信息学分析表明, IbMYB3蛋白属于R2R3-MYB亚家族成员, 在其N端含有高度保守的R2、R3重复单元.IbMYB3蛋白的亚细胞定位以及激活活性结果表明, IbMYB3定位于细胞核, 且具有转录激活活性.基于同源比对及系统进化树分析, 我们发现IbMYB3与已有研究的AtMYB41有较高相似性, 推测其可能具有与AtMYB41类似的功能.前人的研究表明, AtMYB41能够通过调控基因转录及物质代谢来响应渗透胁迫, 从而应对盐、冷和干旱胁迫(Cominelli et al., 2008; Lippold et al., 2009).此外, 与IbMYB3亲缘关系较近的AtMYB74 (Xu et al., 2015)和AtMYB102 (Denekamp and Smeekens, 2003)均被报道具有增强盐耐受性的功能, 因此推测IbMYB3蛋白可能也参与甘薯逆境响应过程. ...
Integration of wounding and osmotic stress signals determines the expression of the AtMYB102 transcription factor gene 2 2003
... IbMYB3蛋白具有MYB蛋白家族R2R3亚家族成员典型的2个保守结构域(图7A), 且Xu22中IbMYB3蛋白与甘薯近缘野生种I. triloba中的ItMYB41和牵牛花(Ipomoea nil)中的InMYB330序列相似性最高(分别达98.6%和93.4%) (图7B).此外, IbMYB3与拟南芥中一些已知功能的MYB转录因子序列也具有很高的相似性.例如, 与拟南芥AtMYB41蛋白的相似性达62.6%, 具有较高的同源性.此外, 我们进一步对比了IbMYB3与其它物种MYB蛋白中R2R3结构域的保守性, 结果显示IbMYB3与ItMYB41、InMYB330、AtMYB41、AtMYB74和AtMYB102中的R2R3结构域的相似性均很高(分别达99.1%、97.2%、91.7%、92.6%和90.7%), 表明负责与DNA结合的R2R3结构域在不同物种中均具有很高的保守性, 暗示IbMYB3可能与已报道的几个拟南芥MYB基因具有类似的功能.已有研究表明, AtMYB41基因能够通过调控基因转录及物质代谢来响应渗透胁迫, 从而应对盐、冷和干旱(Lippold et al., 2009); AtMYB41可在多个物种或细胞类型中激活软木脂的合成, 而软木脂在调节植物水分和保护植物免受生物和非生物胁迫方面发挥关键作用(Kosma et al., 2014); 超表达AtMYB41基因则能调控细胞扩增和角质层沉积以响应非生物胁迫(Cominelli et al., 2008).此外, 研究表明AtMYB102能响应渗透胁迫(Denekamp and Smeekens, 2003)以及AtMYB74能够被盐诱导(Xu et al., 2015).综上表明, IbMYB3基因可能参与甘薯非生物胁迫响应, 但需要进一步实验验证. ...
... 本研究通过RNA-seq分析筛选出2个受盐胁迫显著诱导的甘薯MYB转录因子(IbMYB3和IbMYB4).进一步检测2个基因在多种非生物胁迫和植物生长物质处理下的表达水平, 确定对盐胁迫下被显著诱导的IbMYB3进行深入研究.生物信息学分析表明, IbMYB3蛋白属于R2R3-MYB亚家族成员, 在其N端含有高度保守的R2、R3重复单元.IbMYB3蛋白的亚细胞定位以及激活活性结果表明, IbMYB3定位于细胞核, 且具有转录激活活性.基于同源比对及系统进化树分析, 我们发现IbMYB3与已有研究的AtMYB41有较高相似性, 推测其可能具有与AtMYB41类似的功能.前人的研究表明, AtMYB41能够通过调控基因转录及物质代谢来响应渗透胁迫, 从而应对盐、冷和干旱胁迫(Cominelli et al., 2008; Lippold et al., 2009).此外, 与IbMYB3亲缘关系较近的AtMYB74 (Xu et al., 2015)和AtMYB102 (Denekamp and Smeekens, 2003)均被报道具有增强盐耐受性的功能, 因此推测IbMYB3蛋白可能也参与甘薯逆境响应过程. ...
Genome-wide analysis of the MYB transcription factor superfamily in soybean 1 2012
... MYB类转录因子在植物体内具有广泛的生物学功能, 主要包括以下方面: (1) 对植物初生代谢及次生代谢的调控, 如参与调控花青素的合成(Lotkowska et al., 2015; 祝志欣和鲁迎青, 2016; Zhu et al., 2019)及麻风树(Jatropha curcas)中脂质的合成(Khan et al., 2019); (2) 控制细胞的生长和分化, 如由MYB控制的细胞和花瓣形态的发生(Baumann et al., 2007)、拟南芥中AtMYB59调控根系生长和细胞周期进程(Mu et al., 2009)以及Liu等(2014)证实MYB5和MYB14在紫花苜蓿(Medicago sativa)种皮聚合物的生物合成中发挥关键作用; (3) 调控植物发育, 如甜橙(Citrus sinensis)中MYB转录因子CsMYB330和CsMYB308通过微调Cs4CL1基因的表达来调节果汁囊木质化(Jia et al., 2019), 拟南芥中发育相关基因DRMY1调控细胞扩增和种子产量(Wu et al., 2018); (4) 调控植物对生物和非生物胁迫的响应, 如在拟南芥中超表达AtMYB44可促进气孔关闭进而提升非生物胁迫耐受性(Jung et al., 2008).而小麦(Triticum aestivum) TaMYB31则参与拟南芥的干旱胁迫响应(Zhao et al., 2018).目前, 研究人员已在多个物种中完成MYB基因家族的系统分析, 如在番茄(S. lycopersicom)中对R2R3-MYB多样化的讨论(Gates et al., 2016)、在拟南芥中对MYB家族的分析(Stracke et al., 2001; Dubos et al., 2010)、在大豆(Glycine max)基因组中对MYB超家族的分析(Du et al., 2012)、在刚毛柽柳(Tamarix hispida)中对MYB基因家族及其在非生物胁迫和植物生长物质处理下的表达分析(Zhang et al., 2018)以及MYB转录因子作为植物胁迫响应调控因子的相关综述(Ambawat et al., 2013). ...
MYB transcription factors in Arabidopsis 1 2010
... MYB类转录因子在植物体内具有广泛的生物学功能, 主要包括以下方面: (1) 对植物初生代谢及次生代谢的调控, 如参与调控花青素的合成(Lotkowska et al., 2015; 祝志欣和鲁迎青, 2016; Zhu et al., 2019)及麻风树(Jatropha curcas)中脂质的合成(Khan et al., 2019); (2) 控制细胞的生长和分化, 如由MYB控制的细胞和花瓣形态的发生(Baumann et al., 2007)、拟南芥中AtMYB59调控根系生长和细胞周期进程(Mu et al., 2009)以及Liu等(2014)证实MYB5和MYB14在紫花苜蓿(Medicago sativa)种皮聚合物的生物合成中发挥关键作用; (3) 调控植物发育, 如甜橙(Citrus sinensis)中MYB转录因子CsMYB330和CsMYB308通过微调Cs4CL1基因的表达来调节果汁囊木质化(Jia et al., 2019), 拟南芥中发育相关基因DRMY1调控细胞扩增和种子产量(Wu et al., 2018); (4) 调控植物对生物和非生物胁迫的响应, 如在拟南芥中超表达AtMYB44可促进气孔关闭进而提升非生物胁迫耐受性(Jung et al., 2008).而小麦(Triticum aestivum) TaMYB31则参与拟南芥的干旱胁迫响应(Zhao et al., 2018).目前, 研究人员已在多个物种中完成MYB基因家族的系统分析, 如在番茄(S. lycopersicom)中对R2R3-MYB多样化的讨论(Gates et al., 2016)、在拟南芥中对MYB家族的分析(Stracke et al., 2001; Dubos et al., 2010)、在大豆(Glycine max)基因组中对MYB超家族的分析(Du et al., 2012)、在刚毛柽柳(Tamarix hispida)中对MYB基因家族及其在非生物胁迫和植物生长物质处理下的表达分析(Zhang et al., 2018)以及MYB转录因子作为植物胁迫响应调控因子的相关综述(Ambawat et al., 2013). ...
Diversification of R2R3-MYB transcription factors in the tomato family Solanaceae 1 2016
... MYB类转录因子在植物体内具有广泛的生物学功能, 主要包括以下方面: (1) 对植物初生代谢及次生代谢的调控, 如参与调控花青素的合成(Lotkowska et al., 2015; 祝志欣和鲁迎青, 2016; Zhu et al., 2019)及麻风树(Jatropha curcas)中脂质的合成(Khan et al., 2019); (2) 控制细胞的生长和分化, 如由MYB控制的细胞和花瓣形态的发生(Baumann et al., 2007)、拟南芥中AtMYB59调控根系生长和细胞周期进程(Mu et al., 2009)以及Liu等(2014)证实MYB5和MYB14在紫花苜蓿(Medicago sativa)种皮聚合物的生物合成中发挥关键作用; (3) 调控植物发育, 如甜橙(Citrus sinensis)中MYB转录因子CsMYB330和CsMYB308通过微调Cs4CL1基因的表达来调节果汁囊木质化(Jia et al., 2019), 拟南芥中发育相关基因DRMY1调控细胞扩增和种子产量(Wu et al., 2018); (4) 调控植物对生物和非生物胁迫的响应, 如在拟南芥中超表达AtMYB44可促进气孔关闭进而提升非生物胁迫耐受性(Jung et al., 2008).而小麦(Triticum aestivum) TaMYB31则参与拟南芥的干旱胁迫响应(Zhao et al., 2018).目前, 研究人员已在多个物种中完成MYB基因家族的系统分析, 如在番茄(S. lycopersicom)中对R2R3-MYB多样化的讨论(Gates et al., 2016)、在拟南芥中对MYB家族的分析(Stracke et al., 2001; Dubos et al., 2010)、在大豆(Glycine max)基因组中对MYB超家族的分析(Du et al., 2012)、在刚毛柽柳(Tamarix hispida)中对MYB基因家族及其在非生物胁迫和植物生长物质处理下的表达分析(Zhang et al., 2018)以及MYB转录因子作为植物胁迫响应调控因子的相关综述(Ambawat et al., 2013). ...
Citrus sinensis MYB transcription factors CsMYB330 and CsMYB308 regulate fruit juice sac lignification through fine-tuning expression of the Cs4CL1 gene 1 2018
... MYB类转录因子在植物体内具有广泛的生物学功能, 主要包括以下方面: (1) 对植物初生代谢及次生代谢的调控, 如参与调控花青素的合成(Lotkowska et al., 2015; 祝志欣和鲁迎青, 2016; Zhu et al., 2019)及麻风树(Jatropha curcas)中脂质的合成(Khan et al., 2019); (2) 控制细胞的生长和分化, 如由MYB控制的细胞和花瓣形态的发生(Baumann et al., 2007)、拟南芥中AtMYB59调控根系生长和细胞周期进程(Mu et al., 2009)以及Liu等(2014)证实MYB5和MYB14在紫花苜蓿(Medicago sativa)种皮聚合物的生物合成中发挥关键作用; (3) 调控植物发育, 如甜橙(Citrus sinensis)中MYB转录因子CsMYB330和CsMYB308通过微调Cs4CL1基因的表达来调节果汁囊木质化(Jia et al., 2019), 拟南芥中发育相关基因DRMY1调控细胞扩增和种子产量(Wu et al., 2018); (4) 调控植物对生物和非生物胁迫的响应, 如在拟南芥中超表达AtMYB44可促进气孔关闭进而提升非生物胁迫耐受性(Jung et al., 2008).而小麦(Triticum aestivum) TaMYB31则参与拟南芥的干旱胁迫响应(Zhao et al., 2018).目前, 研究人员已在多个物种中完成MYB基因家族的系统分析, 如在番茄(S. lycopersicom)中对R2R3-MYB多样化的讨论(Gates et al., 2016)、在拟南芥中对MYB家族的分析(Stracke et al., 2001; Dubos et al., 2010)、在大豆(Glycine max)基因组中对MYB超家族的分析(Du et al., 2012)、在刚毛柽柳(Tamarix hispida)中对MYB基因家族及其在非生物胁迫和植物生长物质处理下的表达分析(Zhang et al., 2018)以及MYB转录因子作为植物胁迫响应调控因子的相关综述(Ambawat et al., 2013). ...
Overexpression of AtMYB44 enhances stomatal closure to confer abiotic stress tolerance in transgenic Arabidopsis 1 2008
... MYB类转录因子在植物体内具有广泛的生物学功能, 主要包括以下方面: (1) 对植物初生代谢及次生代谢的调控, 如参与调控花青素的合成(Lotkowska et al., 2015; 祝志欣和鲁迎青, 2016; Zhu et al., 2019)及麻风树(Jatropha curcas)中脂质的合成(Khan et al., 2019); (2) 控制细胞的生长和分化, 如由MYB控制的细胞和花瓣形态的发生(Baumann et al., 2007)、拟南芥中AtMYB59调控根系生长和细胞周期进程(Mu et al., 2009)以及Liu等(2014)证实MYB5和MYB14在紫花苜蓿(Medicago sativa)种皮聚合物的生物合成中发挥关键作用; (3) 调控植物发育, 如甜橙(Citrus sinensis)中MYB转录因子CsMYB330和CsMYB308通过微调Cs4CL1基因的表达来调节果汁囊木质化(Jia et al., 2019), 拟南芥中发育相关基因DRMY1调控细胞扩增和种子产量(Wu et al., 2018); (4) 调控植物对生物和非生物胁迫的响应, 如在拟南芥中超表达AtMYB44可促进气孔关闭进而提升非生物胁迫耐受性(Jung et al., 2008).而小麦(Triticum aestivum) TaMYB31则参与拟南芥的干旱胁迫响应(Zhao et al., 2018).目前, 研究人员已在多个物种中完成MYB基因家族的系统分析, 如在番茄(S. lycopersicom)中对R2R3-MYB多样化的讨论(Gates et al., 2016)、在拟南芥中对MYB家族的分析(Stracke et al., 2001; Dubos et al., 2010)、在大豆(Glycine max)基因组中对MYB超家族的分析(Du et al., 2012)、在刚毛柽柳(Tamarix hispida)中对MYB基因家族及其在非生物胁迫和植物生长物质处理下的表达分析(Zhang et al., 2018)以及MYB转录因子作为植物胁迫响应调控因子的相关综述(Ambawat et al., 2013). ...
A sweetpotato auxin response factor gene (IbARF5) is involved in carotenoid biosynthesis and salt and drought tolerance in transgenic Arabidopsis 1 2018
... 由于甘薯遗传转化周期较长, 因此许多研究者将甘薯相关基因转化到其它模式物种中开展相关研究, 如拟南芥(Chu et al., 2013; Kang et al., 2018)、烟草(An et al., 2015)和马铃薯(Cheng et al., 2013).本研究后期已将IbMYB3在拟南芥中进行异源超表达并鉴定其盐胁迫耐受性.初步结果显示, 与野生型相比, 转基因拟南芥叶的长、宽和面积均大于野生型.植物叶片作为营养器官, 是植物进行光合作用的主要场所, 叶面积大表明转基因株系在单位时间内能够产生更多的生物量, 从而促进植物的生长发育.此外, 盐处理条件下, IbMYB3异源超表达拟南芥植株的萌发率高于野生型(待发表), 表明IbMYB3基因确实参与了对盐胁迫的响应.IbMYB3基因异源超表达增强了拟南芥对盐胁迫的耐受性, 表明该基因具有增强甘薯盐耐受的潜能, 后续可进一步探究其在甘薯响应盐胁迫中的功能. ...
The Phyre2 web portal for protein modeling, prediction and analysis 2 2015
... 甘薯具有耐盐碱、抗旱和耐贫瘠等特性, 可种植于滩涂瘠薄土地, 不存在“与粮争地”的问题, 且单位面积、单位时间内生产的乙醇比水稻(Oryza sativa)和小麦等主要粮食作物多, 因此成为适宜用于挖掘盐碱地生产潜力的能源作物之一(朱明库等, 2017).相比水稻和小麦等大作物, 目前甘薯耐盐等非生物胁迫的分子生物学研究还有很大差距(Liu, 2017).甘薯采用薯苗繁殖, 作为幼嫩组织, 薯苗对土壤盐碱胁迫较敏感, 给甘薯生产带来一定困难(代红军等, 2001).因此, 深入研究甘薯的耐盐特性, 对甘薯种植和深化应用以及我国生物能源发展具有重要意义.Yang等(2017)通过半单倍型基因组测序法首次成功解析了六倍体栽培种甘薯泰中6号的全基因组信息, 为甘薯中重要基因克隆和功能解析提供了极大便利.过去几十年, 甘薯基因工程已经取得重要进展, 已建立根癌农杆菌介导的转化系统并成功用于多种甘薯基因型(Liu, 2017).目前已从甘薯中分离和鉴定了许多参与非生物胁迫耐受性(Bian et al., 2014; Wang et al., 2016)、病虫害抗性(柏洁, 2014)、淀粉合成(李丰, 2017)及花青素合成(An et al., 2015)等相关基因. ...
... 通过半单倍型基因组测序法首次成功解析了六倍体栽培种甘薯泰中6号的全基因组信息, 为甘薯中重要基因克隆和功能解析提供了极大便利.过去几十年, 甘薯基因工程已经取得重要进展, 已建立根癌农杆菌介导的转化系统并成功用于多种甘薯基因型(Liu, 2017).目前已从甘薯中分离和鉴定了许多参与非生物胁迫耐受性(Bian et al., 2014; Wang et al., 2016)、病虫害抗性(柏洁, 2014)、淀粉合成(李丰, 2017)及花青素合成(An et al., 2015)等相关基因. ...
The Arabidopsis transcription factor MYB112 promotes anthocyanin formation during salinity and under high light stress 1 2015
... MYB类转录因子在植物体内具有广泛的生物学功能, 主要包括以下方面: (1) 对植物初生代谢及次生代谢的调控, 如参与调控花青素的合成(Lotkowska et al., 2015; 祝志欣和鲁迎青, 2016; Zhu et al., 2019)及麻风树(Jatropha curcas)中脂质的合成(Khan et al., 2019); (2) 控制细胞的生长和分化, 如由MYB控制的细胞和花瓣形态的发生(Baumann et al., 2007)、拟南芥中AtMYB59调控根系生长和细胞周期进程(Mu et al., 2009)以及Liu等(2014)证实MYB5和MYB14在紫花苜蓿(Medicago sativa)种皮聚合物的生物合成中发挥关键作用; (3) 调控植物发育, 如甜橙(Citrus sinensis)中MYB转录因子CsMYB330和CsMYB308通过微调Cs4CL1基因的表达来调节果汁囊木质化(Jia et al., 2019), 拟南芥中发育相关基因DRMY1调控细胞扩增和种子产量(Wu et al., 2018); (4) 调控植物对生物和非生物胁迫的响应, 如在拟南芥中超表达AtMYB44可促进气孔关闭进而提升非生物胁迫耐受性(Jung et al., 2008).而小麦(Triticum aestivum) TaMYB31则参与拟南芥的干旱胁迫响应(Zhao et al., 2018).目前, 研究人员已在多个物种中完成MYB基因家族的系统分析, 如在番茄(S. lycopersicom)中对R2R3-MYB多样化的讨论(Gates et al., 2016)、在拟南芥中对MYB家族的分析(Stracke et al., 2001; Dubos et al., 2010)、在大豆(Glycine max)基因组中对MYB超家族的分析(Du et al., 2012)、在刚毛柽柳(Tamarix hispida)中对MYB基因家族及其在非生物胁迫和植物生长物质处理下的表达分析(Zhang et al., 2018)以及MYB转录因子作为植物胁迫响应调控因子的相关综述(Ambawat et al., 2013). ...
An R2R3-type transcription factor gene AtMYB59 regulates root growth and cell cycle progression in Arabidopsis 1 2009
... MYB类转录因子在植物体内具有广泛的生物学功能, 主要包括以下方面: (1) 对植物初生代谢及次生代谢的调控, 如参与调控花青素的合成(Lotkowska et al., 2015; 祝志欣和鲁迎青, 2016; Zhu et al., 2019)及麻风树(Jatropha curcas)中脂质的合成(Khan et al., 2019); (2) 控制细胞的生长和分化, 如由MYB控制的细胞和花瓣形态的发生(Baumann et al., 2007)、拟南芥中AtMYB59调控根系生长和细胞周期进程(Mu et al., 2009)以及Liu等(2014)证实MYB5和MYB14在紫花苜蓿(Medicago sativa)种皮聚合物的生物合成中发挥关键作用; (3) 调控植物发育, 如甜橙(Citrus sinensis)中MYB转录因子CsMYB330和CsMYB308通过微调Cs4CL1基因的表达来调节果汁囊木质化(Jia et al., 2019), 拟南芥中发育相关基因DRMY1调控细胞扩增和种子产量(Wu et al., 2018); (4) 调控植物对生物和非生物胁迫的响应, 如在拟南芥中超表达AtMYB44可促进气孔关闭进而提升非生物胁迫耐受性(Jung et al., 2008).而小麦(Triticum aestivum) TaMYB31则参与拟南芥的干旱胁迫响应(Zhao et al., 2018).目前, 研究人员已在多个物种中完成MYB基因家族的系统分析, 如在番茄(S. lycopersicom)中对R2R3-MYB多样化的讨论(Gates et al., 2016)、在拟南芥中对MYB家族的分析(Stracke et al., 2001; Dubos et al., 2010)、在大豆(Glycine max)基因组中对MYB超家族的分析(Du et al., 2012)、在刚毛柽柳(Tamarix hispida)中对MYB基因家族及其在非生物胁迫和植物生长物质处理下的表达分析(Zhang et al., 2018)以及MYB转录因子作为植物胁迫响应调控因子的相关综述(Ambawat et al., 2013). ...
Better prediction of sub-cellular localization by combining evolutionary and structural information 1 2003
... 首先, 通过亚细胞定位预测程序PSORT (Protein Subcellular Localization Prediction Tool, https://www.genscript.com/psort.html)在线对IbMYB3氨基酸序列进行亚细胞定位预测.然后通过与增强的绿色荧光蛋白(Enhanced Green Fluorescent Protein, EGFP)形成融合蛋白(Nair and Rost, 2003)进一步验证IbMYB3蛋白的亚细胞定位.通过引物IbMYB3-EGFP扩增得到含有双酶切位点XbaI和BamHI但无终止密码子的IbMYB3全长CDS片段, 经过双酶切后连接到双元载体pBI121:EGFP上.经过测序比对, 将成功构建的质粒pBI121:IbMYB3:EGFP (图1A)与pBI121:EGFP载体转化到农杆菌LBA4404中, 并进行菌落PCR检测(图1B).挑选阳性克隆, 28°C、每分钟180转暗培养至菌液OD600约为1.0, 经离心重悬后将菌液在无菌条件下侵染洋葱下表皮细胞.将培养2天的洋葱下表皮分别在荧光显微镜(德国莱卡, DM6000B)的明场及绿色荧光通道下观察, 确认绿色荧光蛋白及融合蛋白在细胞中的表达位置(Sun et al., 2007). ...
Stable internal reference genes for the normalization of Real-Time PCR in different sweetpotato cultivars subjected to abiotic stress conditions 1 2012
Overexpression of the IbMYB1 gene in an orange-fleshed sweet potato cultivar produces a dual-pigmented transgenic sweet potato with improved antioxidant activity 1 2015
Comprehensive analysis of MYB gene family and their expressions under abiotic stresses and hormone treatments in Tamarix hispida 1 2018
... MYB类转录因子在植物体内具有广泛的生物学功能, 主要包括以下方面: (1) 对植物初生代谢及次生代谢的调控, 如参与调控花青素的合成(Lotkowska et al., 2015; 祝志欣和鲁迎青, 2016; Zhu et al., 2019)及麻风树(Jatropha curcas)中脂质的合成(Khan et al., 2019); (2) 控制细胞的生长和分化, 如由MYB控制的细胞和花瓣形态的发生(Baumann et al., 2007)、拟南芥中AtMYB59调控根系生长和细胞周期进程(Mu et al., 2009)以及Liu等(2014)证实MYB5和MYB14在紫花苜蓿(Medicago sativa)种皮聚合物的生物合成中发挥关键作用; (3) 调控植物发育, 如甜橙(Citrus sinensis)中MYB转录因子CsMYB330和CsMYB308通过微调Cs4CL1基因的表达来调节果汁囊木质化(Jia et al., 2019), 拟南芥中发育相关基因DRMY1调控细胞扩增和种子产量(Wu et al., 2018); (4) 调控植物对生物和非生物胁迫的响应, 如在拟南芥中超表达AtMYB44可促进气孔关闭进而提升非生物胁迫耐受性(Jung et al., 2008).而小麦(Triticum aestivum) TaMYB31则参与拟南芥的干旱胁迫响应(Zhao et al., 2018).目前, 研究人员已在多个物种中完成MYB基因家族的系统分析, 如在番茄(S. lycopersicom)中对R2R3-MYB多样化的讨论(Gates et al., 2016)、在拟南芥中对MYB家族的分析(Stracke et al., 2001; Dubos et al., 2010)、在大豆(Glycine max)基因组中对MYB超家族的分析(Du et al., 2012)、在刚毛柽柳(Tamarix hispida)中对MYB基因家族及其在非生物胁迫和植物生长物质处理下的表达分析(Zhang et al., 2018)以及MYB转录因子作为植物胁迫响应调控因子的相关综述(Ambawat et al., 2013). ...
The wheat MYB transcription factor TaMYB 31 is involved in drought stress responses in Arabidopsis 1 2018
... MYB类转录因子在植物体内具有广泛的生物学功能, 主要包括以下方面: (1) 对植物初生代谢及次生代谢的调控, 如参与调控花青素的合成(Lotkowska et al., 2015; 祝志欣和鲁迎青, 2016; Zhu et al., 2019)及麻风树(Jatropha curcas)中脂质的合成(Khan et al., 2019); (2) 控制细胞的生长和分化, 如由MYB控制的细胞和花瓣形态的发生(Baumann et al., 2007)、拟南芥中AtMYB59调控根系生长和细胞周期进程(Mu et al., 2009)以及Liu等(2014)证实MYB5和MYB14在紫花苜蓿(Medicago sativa)种皮聚合物的生物合成中发挥关键作用; (3) 调控植物发育, 如甜橙(Citrus sinensis)中MYB转录因子CsMYB330和CsMYB308通过微调Cs4CL1基因的表达来调节果汁囊木质化(Jia et al., 2019), 拟南芥中发育相关基因DRMY1调控细胞扩增和种子产量(Wu et al., 2018); (4) 调控植物对生物和非生物胁迫的响应, 如在拟南芥中超表达AtMYB44可促进气孔关闭进而提升非生物胁迫耐受性(Jung et al., 2008).而小麦(Triticum aestivum) TaMYB31则参与拟南芥的干旱胁迫响应(Zhao et al., 2018).目前, 研究人员已在多个物种中完成MYB基因家族的系统分析, 如在番茄(S. lycopersicom)中对R2R3-MYB多样化的讨论(Gates et al., 2016)、在拟南芥中对MYB家族的分析(Stracke et al., 2001; Dubos et al., 2010)、在大豆(Glycine max)基因组中对MYB超家族的分析(Du et al., 2012)、在刚毛柽柳(Tamarix hispida)中对MYB基因家族及其在非生物胁迫和植物生长物质处理下的表达分析(Zhang et al., 2018)以及MYB转录因子作为植物胁迫响应调控因子的相关综述(Ambawat et al., 2013). ...
A R2R3-MYB transcription factor, VvMYBC2L2, functions as a transcriptional repressor of anthocyanin biosynthesis in grapevine (Vitis vinifera L.) 1 2019
... MYB类转录因子在植物体内具有广泛的生物学功能, 主要包括以下方面: (1) 对植物初生代谢及次生代谢的调控, 如参与调控花青素的合成(Lotkowska et al., 2015; 祝志欣和鲁迎青, 2016; Zhu et al., 2019)及麻风树(Jatropha curcas)中脂质的合成(Khan et al., 2019); (2) 控制细胞的生长和分化, 如由MYB控制的细胞和花瓣形态的发生(Baumann et al., 2007)、拟南芥中AtMYB59调控根系生长和细胞周期进程(Mu et al., 2009)以及Liu等(2014)证实MYB5和MYB14在紫花苜蓿(Medicago sativa)种皮聚合物的生物合成中发挥关键作用; (3) 调控植物发育, 如甜橙(Citrus sinensis)中MYB转录因子CsMYB330和CsMYB308通过微调Cs4CL1基因的表达来调节果汁囊木质化(Jia et al., 2019), 拟南芥中发育相关基因DRMY1调控细胞扩增和种子产量(Wu et al., 2018); (4) 调控植物对生物和非生物胁迫的响应, 如在拟南芥中超表达AtMYB44可促进气孔关闭进而提升非生物胁迫耐受性(Jung et al., 2008).而小麦(Triticum aestivum) TaMYB31则参与拟南芥的干旱胁迫响应(Zhao et al., 2018).目前, 研究人员已在多个物种中完成MYB基因家族的系统分析, 如在番茄(S. lycopersicom)中对R2R3-MYB多样化的讨论(Gates et al., 2016)、在拟南芥中对MYB家族的分析(Stracke et al., 2001; Dubos et al., 2010)、在大豆(Glycine max)基因组中对MYB超家族的分析(Du et al., 2012)、在刚毛柽柳(Tamarix hispida)中对MYB基因家族及其在非生物胁迫和植物生长物质处理下的表达分析(Zhang et al., 2018)以及MYB转录因子作为植物胁迫响应调控因子的相关综述(Ambawat et al., 2013). ...
 ,*江苏师范大学生命科学学院/江苏省药用植物生物技术重点实验室, 徐州 221116
,*江苏师范大学生命科学学院/江苏省药用植物生物技术重点实验室, 徐州 221116
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT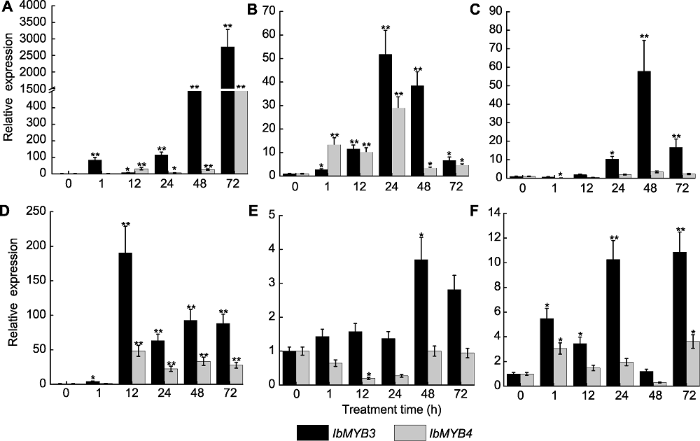 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT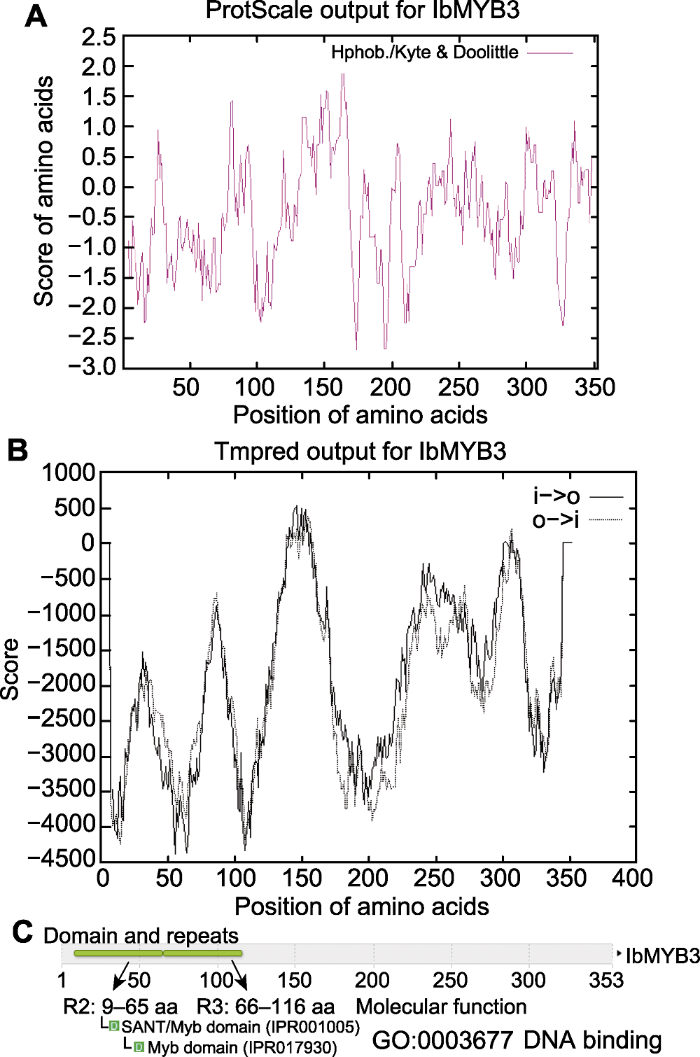 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT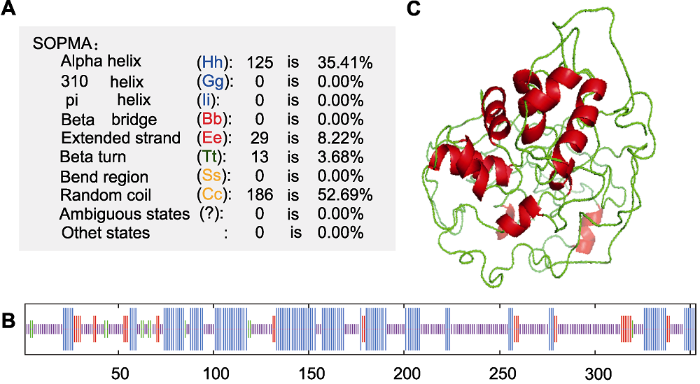 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT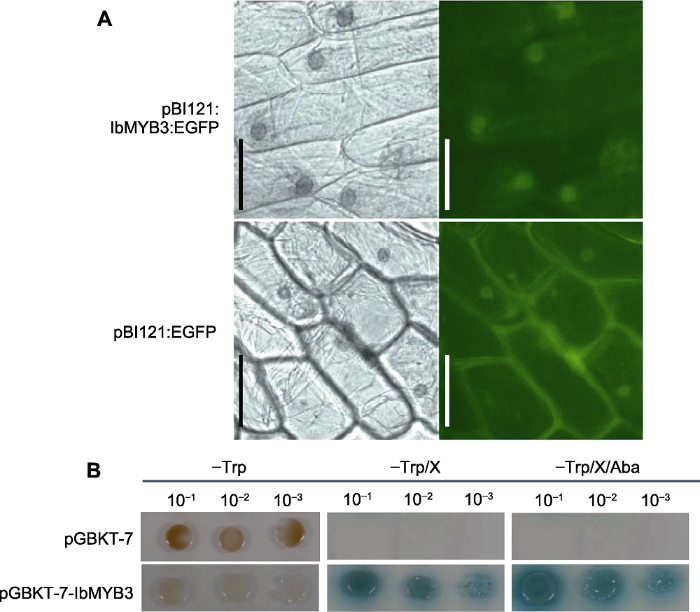 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT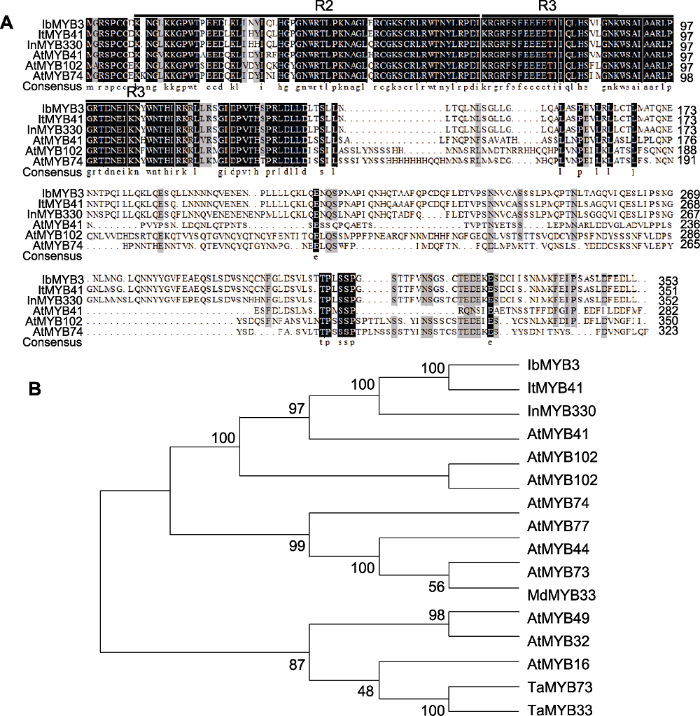 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}