Dan Zhu, Hanwei Cao, Yuan Li, Dongtao Ren,*State Key Laboratory of Plant Physiology and Biochemistry, College of Biological Sciences, China Agricultural University, Beijing 100193, China
Abstract Protein phosphorylation is one of the important protein posttranslational modifications that is involved in the regulation of most cellular processes in plants. Protein kinases catalyze the phosphorylation by transferring the phosphate group in ATP to the substrate proteins. The phosphate is usually covalently linked to the hydroxyl group of specific amino acid residues in the substrates by an ester bond. The mostly studied phosphorylation sites are serine, threonine, and tyrosine residues. Here, we present protocols and related tips for the in vitro and in vivo protein phosphorylation assays. Keywords:protein phosphorylation;protein kinase;plant
PDF (4000KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 朱丹, 曹汉威, 李媛, 任东涛. 植物蛋白磷酸化的检测方法. 植物学报, 2020, 55(1): 76-82 doi:10.11983/CBB19208 Zhu Dan, Cao Hanwei, Li Yuan, Ren Dongtao. Protocols for Analyzing Plant Phospho-proteins. Chinese Bulletin of Botany, 2020, 55(1): 76-82 doi:10.11983/CBB19208
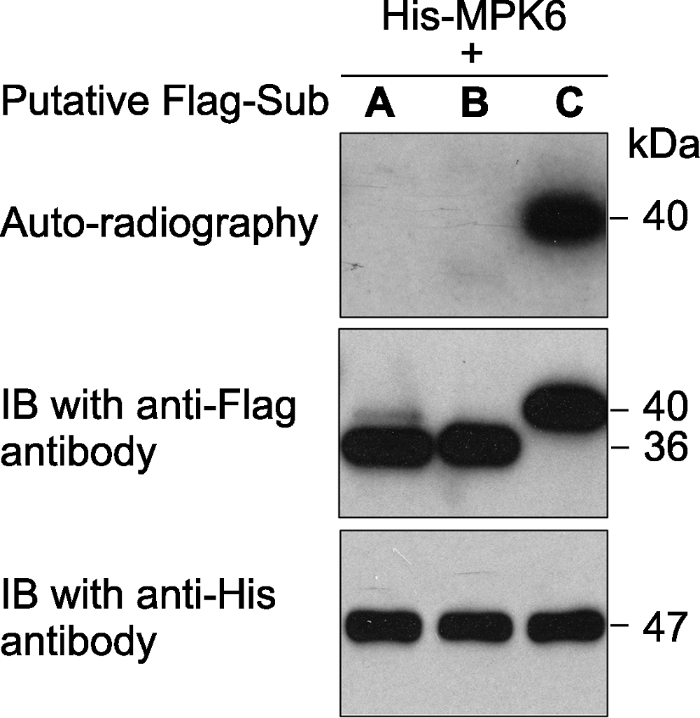
激酶(MPK6)连接有6×His标签、推测的底物蛋白(A、B和C)连接有Flag标签。体外磷酸化反应完成后, 反应体系中的蛋白用SDS-PAGE gel分离。然后进行磷酸化底物的放射自显影分析(上图)。底物蛋白(中图)和激酶(下图)的免疫印迹分析显示底物、激酶各自在磷酸化反应中的蛋白使用量基本一致。 Figure 1An in vitro phosphorylation assay of 3 putative substrate proteins by MPK6 in Arabidopsis thaliana
MPK6 was fused with the 6×His tag and the 3 putative substrates were fused with the Flag tag. After phosphorylation reaction, the proteins in the mixture were separated on a SDS- PAGE gel. The gel was dried and exposed to X-ray film (top). Immunoblotting with anti-His and anti-Flag antibodies were used to show the levels of the substrate (middle) and kinase (bottom) proteins in the reactions.
磷酸酶(如Calf intestine alkaline phosphatase, CIAP)可去除磷酸化蛋白上的磷酸基团。同一样品在CIAP处理前(-)、后(+), 磷酸化和非磷酸化形式的蛋白经Phos-tag gel (上图)和SDS- PAGE gel (下图)电泳后的免疫印迹分析结果。经Phos-tag gel电泳, 未加CIAP的样品中特定蛋白的磷酸化(P-form)和非磷酸化形式(None-P-form)被清晰地分开(上图); 加CIAP后磷酸化形式消失而非磷酸化形式条带增强。经SDS-PAGE gel电泳, 未加和加CIAP处理的样品中, 特定蛋白的磷酸化和非磷酸化形式分开不明显(下图)。 Figure 2Immunoblotting detection of phospho-proteins in samples after Phos-tag gel and SDS-PAGE gel separation
Phospho-proteins can be dephosphorylated by phosphatases (e.g. Calf intestine alkaline phosphatase, CIAP). Protein samples treated with (+) or without (-) CIAP were separated by Phos-tag (top) and SDS-PAGE (bottom) gels, and the specific protein was further detected by immunoblotting. The P-form and None-P-form of the protein were separated clearly by a Phos-tag gel (top), while the two forms were not separated by a SDS-PAGE gel (bottom). The missing of the upper band and the increasing of the bottom band after Phos-tag gel separation indicated that P-form protein was completely dephosphorylated by CIAP treatment.
2周龄拟南芥幼苗总蛋白提取液经双向凝胶电泳分离后, 用Pro-Q Diamond试剂对磷酸化蛋白进行染色(左图)。荧光扫描后的凝胶再进行考马斯亮蓝染色(blue-silver staining) (右图)。 Figure 3Pro-Q staining assay of the phospho-proteins in total proteins extracted from Arabidopsis thaliana seedlings and separated by a 2D gel
Two-week-old Arabidopsis thaliana seedlings were used for total protein extraction. The total proteins were separated by a 2D gel and the phospho-proteins were stained by Pro-Q (left). The gel was scanned with a Typhoon 9410 fluorescence scanner, and then stained with blue-silver to visualize the proteins (right).
AgrawalGK, ThelenJJ (2005). Development of a simplified, economical polyacrylamide gel staining protocol for phosphoproteins Proteomics 5, 4684-4688. [本文引用: 1]
AgrawalGK, ThelenJJ (2006). Large scale identification and quantitative profiling of phosphoproteins expressed during seed filling in oilseed rape Mol Cell Proteomics 5, 2044-2059. [本文引用: 1]
BlomN, Sicheritz-PonténT, GuptaR, GammeltoftS, BrunakS (2004). Prediction of post-translational glycosylation and phosphorylation of proteins from the amino acid sequence Proteomics 4, 1633-1649. [本文引用: 1]
CandianoG, BruschiM, MusanteL, SantucciL, GhiggeriGM, CarnemollaB, OrecchiaP, ZardiL, RighettiPG (2004). Blue silver: a very sensitive colloidal Coomassie G-250 staining for proteome analysis Electrophoresis 25, 1327-1333. [本文引用: 1]
ChaoQ, LiuXY, MeiYC, GaoZF, ChenYB, QianCR, HaoYB, WangBC (2014). Light-regulated phosphorylation of maize phosphoenolpyruvate carboxykinase plays a vital role in its activity Plant Mol Biol 85, 95-105. [本文引用: 1]
ChenMJ, DixonJE, ManningG (2017). Genomics and evolution of protein phosphatases Sci Signal 10, eaag1796. [本文引用: 1]
de la Fuente van BentemS, HirtH (2007). Using phosphoproteomics to reveal signaling dynamics in plants Trends Plant Sci 12, 404-411. [本文引用: 1]
FicarroSB, McClelandML, StukenbergPT, BurkeDJ, RossMM, ShabanowitzJ, HuntDF, WhiteFM (2002). Phosphoproteome analysis by mass spectrometry and its application to Saccharomyces cerevisiae Nat Biotechnol 20, 301-305. [本文引用: 1]
FischerEH, KrebsEG (1955). Conversion of phosphorylase B to phosphorylase A in muscle extracts J Biol Chem 216, 121-132. [本文引用: 1]
FrostDC, LiLJ (2014). Recent advances in mass spectrometry-based glycoproteomics Adv Protein Chem Struct Biol 95, 71-123. [本文引用: 1]
HubbardMJ, CohenP (1993). On target with a new mechanism for the regulation of protein phosphorylation Trends Biochem Sci 18, 172-177. [本文引用: 1]
KeYQ, HanGQ, HeHQ, LiJX (2009). Differential regulation of proteins and phosphoproteins in rice under drought stress Biochem Biophys Res Commun 379, 133-138. [本文引用: 1]
KhanM, TakasakiH, KomatsuS (2005). Comprehensive phosphoproteome analysis in rice and identification of phosphoproteins responsive to different hormones/stresses J Proteome Res 4, 1592-1599. [本文引用: 1]
KimHS, FernandesG, LeeCW (2016). Protein phosphatases involved in regulating mitosis: facts and hypotheses Mol Cell 39, 654-662. [本文引用: 1]
KinoshitaE, Kinoshita-KikutaE, KoikeT (2007). Specific recognition and detection of phosphorylated proteins using characteristics of metal ion Yakugaku Zasshi 127, 1897-1913. [本文引用: 1]
KosakoH, NaganoK (2011). Quantitative phosphoproteomics strategies for understanding protein kinase-mediated signal transduction pathways Expert Rev Proteomics 8, 81-94. [本文引用: 1]
KrupaA, PreethiG, SrinivasanN (2004). Structural modes of stabilization of permissive phosphorylation sites in protein kinases: distinct strategies in Ser/Thr and Tyr kinases J Mol Biol 339, 1025-1039. [本文引用: 1]
LaemmliUK (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4 Nature 227, 680-685. [本文引用: 1]
LaugesenS, BergoinA, RossignolM (2004). Deciphering the plant phosphoproteome: tools and strategies for a challenging task Plant Physiol Biochem 42, 929-936. [本文引用: 1]
PeckSC (2003). Early phosphorylation events in biotic stress Curr Opin Plant Biol 6, 334-338. [本文引用: 1]
PrakS, HemS, BoudetJ, ViennoisG, SommererN, RossignolM, MaurelC, SantoniV (2008). Multiple phosphorylations in the C-terminal tail of plant plasma membrane aquaporins: role in subcellular trafficking of AtPIP2;1 in response to salt stress Mol Cell Proteomics 7, 1019-1030. [本文引用: 1]
WangPC, ZhaoY, LiZP, HsuCC, LiuX, FuLW, HouYJ, DuYY, XieSJ, ZhangCG, GaoJH, CaoMJ, HuangXS, ZhuYF, TangK, WangXG, TaoWA, XiongY, ZhuJK (2018). Reciprocal regulation of the TOR kinase and ABA receptor balances plant growth and stress response Mol Cell 69, 100-112. [本文引用: 1]
WhitemanSA, NühseTS, AshfordDA, SandersD, MaathuisFJ (2008). A proteomic and phosphoproteomic analysis of Oryza sativa plasma membrane and vacuolar membrane Plant J 56, 146-156. [本文引用: 1]
WuCF, WangRN, LiangQJ, LiangJJ, LiWK, JungSY, QinJ, LinSH, KuangJ (2010). Dissecting the M phase- specific phosphorylation of serine-proline or threonine- proline motifs Mol Biol Cell 21, 1470-1481. [本文引用: 1]
YangC, WangZG, ZhuPF (2004). Recent advances of protein phosphorylation in proteome Prog Physiol Sci 35, 119-124. [本文引用: 1]
YinXJ, WangX, KomatsuS (2018). Phosphoproteomics: protein phosphorylation in regulation of seed germination and plant growth Curr Protein Pept Sci 19, 401-412. [本文引用: 1]
Large scale identification and quantitative profiling of phosphoproteins expressed during seed filling in oilseed rape 1 2006
... 研究显示, 真核生物细胞中约有1/3的蛋白被磷酸化修饰, 表明蛋白磷酸化修饰的普遍性和重要性(Hubbard and Cohen, 1993; Ficarro et al., 2002).蛋白磷酸化由蛋白激酶(protein kinase)催化完成.蛋白激酶将供体ATP (少数情况下用GTP)上的γ位磷酸基团以酯键的形式连接到底物蛋白的特定氨基酸侧链的羟基上(Fischer and Krebs, 1955; Krupa et al., 2004).真核生物蛋白被磷酸化的氨基酸主要包括丝氨酸、苏氨酸和酪氨酸.原核生物还可能发生在天冬氨酸、谷氨酸和组氨酸上(Wu et al., 2010).磷酸化蛋白上的磷酸基团可以由磷酸酶(phosphatase)除去.蛋白的磷酸化与去磷酸化的可逆调控参与诸多生命活动过程(Peck, 2003; Yang et al., 2004; Kim et al., 2016; Chen et al., 2017).磷酸化修饰通过改变蛋白结构或局部结构进而影响蛋白与特定蛋白间的互作、蛋白自身的活性和稳定性及蛋白的正常定位等.细胞中蛋白磷酸化水平是动态变化的, 细微变化即可导致细胞代谢及生理状态的变化(Laugesen et al., 2004; Khan et al., 2005; Agrawal and Thelen, 2006; Whiteman et al., 2008; Chao et al., 2014).研究表明, 植物的生长发育和对各种逆境响应的生理及信号调控过程都有蛋白磷酸化修饰的发生(de la Fuente van Bentem and Hirt, 2007; Prak et al., 2008; Ke et al., 2009; Wang et al., 2018; Yin et al., 2018; Zhang et al., 2019).因此, 解析关键蛋白磷酸化水平的变化规律, 可从分子水平揭示植物重要性状和代谢途径的调节机制. ...
Prediction of post-translational glycosylation and phosphorylation of proteins from the amino acid sequence 1 2004
... 蛋白质翻译后修饰(posttranslational modification)是指通过酶促反应将一些化学基团共价连接到底物蛋白质分子上的化学修饰.蛋白质翻译后修饰对多种蛋白质正常行使功能起重要作用.蛋白质翻译后修饰的种类很多, 包括磷酸化、乙酰化、泛素化、糖基化、脂酰化及异戊烯基化等(Blom et al., 2004; Kosako and Nagano, 2011; Frost and Li, 2014; Zhu, 2017). ...
Blue silver: a very sensitive colloidal Coomassie G-250 staining for proteome analysis 1 2004
Light-regulated phosphorylation of maize phosphoenolpyruvate carboxykinase plays a vital role in its activity 1 2014
... 研究显示, 真核生物细胞中约有1/3的蛋白被磷酸化修饰, 表明蛋白磷酸化修饰的普遍性和重要性(Hubbard and Cohen, 1993; Ficarro et al., 2002).蛋白磷酸化由蛋白激酶(protein kinase)催化完成.蛋白激酶将供体ATP (少数情况下用GTP)上的γ位磷酸基团以酯键的形式连接到底物蛋白的特定氨基酸侧链的羟基上(Fischer and Krebs, 1955; Krupa et al., 2004).真核生物蛋白被磷酸化的氨基酸主要包括丝氨酸、苏氨酸和酪氨酸.原核生物还可能发生在天冬氨酸、谷氨酸和组氨酸上(Wu et al., 2010).磷酸化蛋白上的磷酸基团可以由磷酸酶(phosphatase)除去.蛋白的磷酸化与去磷酸化的可逆调控参与诸多生命活动过程(Peck, 2003; Yang et al., 2004; Kim et al., 2016; Chen et al., 2017).磷酸化修饰通过改变蛋白结构或局部结构进而影响蛋白与特定蛋白间的互作、蛋白自身的活性和稳定性及蛋白的正常定位等.细胞中蛋白磷酸化水平是动态变化的, 细微变化即可导致细胞代谢及生理状态的变化(Laugesen et al., 2004; Khan et al., 2005; Agrawal and Thelen, 2006; Whiteman et al., 2008; Chao et al., 2014).研究表明, 植物的生长发育和对各种逆境响应的生理及信号调控过程都有蛋白磷酸化修饰的发生(de la Fuente van Bentem and Hirt, 2007; Prak et al., 2008; Ke et al., 2009; Wang et al., 2018; Yin et al., 2018; Zhang et al., 2019).因此, 解析关键蛋白磷酸化水平的变化规律, 可从分子水平揭示植物重要性状和代谢途径的调节机制. ...
Genomics and evolution of protein phosphatases 1 2017
... 研究显示, 真核生物细胞中约有1/3的蛋白被磷酸化修饰, 表明蛋白磷酸化修饰的普遍性和重要性(Hubbard and Cohen, 1993; Ficarro et al., 2002).蛋白磷酸化由蛋白激酶(protein kinase)催化完成.蛋白激酶将供体ATP (少数情况下用GTP)上的γ位磷酸基团以酯键的形式连接到底物蛋白的特定氨基酸侧链的羟基上(Fischer and Krebs, 1955; Krupa et al., 2004).真核生物蛋白被磷酸化的氨基酸主要包括丝氨酸、苏氨酸和酪氨酸.原核生物还可能发生在天冬氨酸、谷氨酸和组氨酸上(Wu et al., 2010).磷酸化蛋白上的磷酸基团可以由磷酸酶(phosphatase)除去.蛋白的磷酸化与去磷酸化的可逆调控参与诸多生命活动过程(Peck, 2003; Yang et al., 2004; Kim et al., 2016; Chen et al., 2017).磷酸化修饰通过改变蛋白结构或局部结构进而影响蛋白与特定蛋白间的互作、蛋白自身的活性和稳定性及蛋白的正常定位等.细胞中蛋白磷酸化水平是动态变化的, 细微变化即可导致细胞代谢及生理状态的变化(Laugesen et al., 2004; Khan et al., 2005; Agrawal and Thelen, 2006; Whiteman et al., 2008; Chao et al., 2014).研究表明, 植物的生长发育和对各种逆境响应的生理及信号调控过程都有蛋白磷酸化修饰的发生(de la Fuente van Bentem and Hirt, 2007; Prak et al., 2008; Ke et al., 2009; Wang et al., 2018; Yin et al., 2018; Zhang et al., 2019).因此, 解析关键蛋白磷酸化水平的变化规律, 可从分子水平揭示植物重要性状和代谢途径的调节机制. ...
Using phosphoproteomics to reveal signaling dynamics in plants 1 2007
... 研究显示, 真核生物细胞中约有1/3的蛋白被磷酸化修饰, 表明蛋白磷酸化修饰的普遍性和重要性(Hubbard and Cohen, 1993; Ficarro et al., 2002).蛋白磷酸化由蛋白激酶(protein kinase)催化完成.蛋白激酶将供体ATP (少数情况下用GTP)上的γ位磷酸基团以酯键的形式连接到底物蛋白的特定氨基酸侧链的羟基上(Fischer and Krebs, 1955; Krupa et al., 2004).真核生物蛋白被磷酸化的氨基酸主要包括丝氨酸、苏氨酸和酪氨酸.原核生物还可能发生在天冬氨酸、谷氨酸和组氨酸上(Wu et al., 2010).磷酸化蛋白上的磷酸基团可以由磷酸酶(phosphatase)除去.蛋白的磷酸化与去磷酸化的可逆调控参与诸多生命活动过程(Peck, 2003; Yang et al., 2004; Kim et al., 2016; Chen et al., 2017).磷酸化修饰通过改变蛋白结构或局部结构进而影响蛋白与特定蛋白间的互作、蛋白自身的活性和稳定性及蛋白的正常定位等.细胞中蛋白磷酸化水平是动态变化的, 细微变化即可导致细胞代谢及生理状态的变化(Laugesen et al., 2004; Khan et al., 2005; Agrawal and Thelen, 2006; Whiteman et al., 2008; Chao et al., 2014).研究表明, 植物的生长发育和对各种逆境响应的生理及信号调控过程都有蛋白磷酸化修饰的发生(de la Fuente van Bentem and Hirt, 2007; Prak et al., 2008; Ke et al., 2009; Wang et al., 2018; Yin et al., 2018; Zhang et al., 2019).因此, 解析关键蛋白磷酸化水平的变化规律, 可从分子水平揭示植物重要性状和代谢途径的调节机制. ...
Phosphoproteome analysis by mass spectrometry and its application to Saccharomyces cerevisiae 1 2002
... 研究显示, 真核生物细胞中约有1/3的蛋白被磷酸化修饰, 表明蛋白磷酸化修饰的普遍性和重要性(Hubbard and Cohen, 1993; Ficarro et al., 2002).蛋白磷酸化由蛋白激酶(protein kinase)催化完成.蛋白激酶将供体ATP (少数情况下用GTP)上的γ位磷酸基团以酯键的形式连接到底物蛋白的特定氨基酸侧链的羟基上(Fischer and Krebs, 1955; Krupa et al., 2004).真核生物蛋白被磷酸化的氨基酸主要包括丝氨酸、苏氨酸和酪氨酸.原核生物还可能发生在天冬氨酸、谷氨酸和组氨酸上(Wu et al., 2010).磷酸化蛋白上的磷酸基团可以由磷酸酶(phosphatase)除去.蛋白的磷酸化与去磷酸化的可逆调控参与诸多生命活动过程(Peck, 2003; Yang et al., 2004; Kim et al., 2016; Chen et al., 2017).磷酸化修饰通过改变蛋白结构或局部结构进而影响蛋白与特定蛋白间的互作、蛋白自身的活性和稳定性及蛋白的正常定位等.细胞中蛋白磷酸化水平是动态变化的, 细微变化即可导致细胞代谢及生理状态的变化(Laugesen et al., 2004; Khan et al., 2005; Agrawal and Thelen, 2006; Whiteman et al., 2008; Chao et al., 2014).研究表明, 植物的生长发育和对各种逆境响应的生理及信号调控过程都有蛋白磷酸化修饰的发生(de la Fuente van Bentem and Hirt, 2007; Prak et al., 2008; Ke et al., 2009; Wang et al., 2018; Yin et al., 2018; Zhang et al., 2019).因此, 解析关键蛋白磷酸化水平的变化规律, 可从分子水平揭示植物重要性状和代谢途径的调节机制. ...
Conversion of phosphorylase B to phosphorylase A in muscle extracts 1 1955
... 研究显示, 真核生物细胞中约有1/3的蛋白被磷酸化修饰, 表明蛋白磷酸化修饰的普遍性和重要性(Hubbard and Cohen, 1993; Ficarro et al., 2002).蛋白磷酸化由蛋白激酶(protein kinase)催化完成.蛋白激酶将供体ATP (少数情况下用GTP)上的γ位磷酸基团以酯键的形式连接到底物蛋白的特定氨基酸侧链的羟基上(Fischer and Krebs, 1955; Krupa et al., 2004).真核生物蛋白被磷酸化的氨基酸主要包括丝氨酸、苏氨酸和酪氨酸.原核生物还可能发生在天冬氨酸、谷氨酸和组氨酸上(Wu et al., 2010).磷酸化蛋白上的磷酸基团可以由磷酸酶(phosphatase)除去.蛋白的磷酸化与去磷酸化的可逆调控参与诸多生命活动过程(Peck, 2003; Yang et al., 2004; Kim et al., 2016; Chen et al., 2017).磷酸化修饰通过改变蛋白结构或局部结构进而影响蛋白与特定蛋白间的互作、蛋白自身的活性和稳定性及蛋白的正常定位等.细胞中蛋白磷酸化水平是动态变化的, 细微变化即可导致细胞代谢及生理状态的变化(Laugesen et al., 2004; Khan et al., 2005; Agrawal and Thelen, 2006; Whiteman et al., 2008; Chao et al., 2014).研究表明, 植物的生长发育和对各种逆境响应的生理及信号调控过程都有蛋白磷酸化修饰的发生(de la Fuente van Bentem and Hirt, 2007; Prak et al., 2008; Ke et al., 2009; Wang et al., 2018; Yin et al., 2018; Zhang et al., 2019).因此, 解析关键蛋白磷酸化水平的变化规律, 可从分子水平揭示植物重要性状和代谢途径的调节机制. ...
Recent advances in mass spectrometry-based glycoproteomics 1 2014
... 蛋白质翻译后修饰(posttranslational modification)是指通过酶促反应将一些化学基团共价连接到底物蛋白质分子上的化学修饰.蛋白质翻译后修饰对多种蛋白质正常行使功能起重要作用.蛋白质翻译后修饰的种类很多, 包括磷酸化、乙酰化、泛素化、糖基化、脂酰化及异戊烯基化等(Blom et al., 2004; Kosako and Nagano, 2011; Frost and Li, 2014; Zhu, 2017). ...
On target with a new mechanism for the regulation of protein phosphorylation 1 1993
... 研究显示, 真核生物细胞中约有1/3的蛋白被磷酸化修饰, 表明蛋白磷酸化修饰的普遍性和重要性(Hubbard and Cohen, 1993; Ficarro et al., 2002).蛋白磷酸化由蛋白激酶(protein kinase)催化完成.蛋白激酶将供体ATP (少数情况下用GTP)上的γ位磷酸基团以酯键的形式连接到底物蛋白的特定氨基酸侧链的羟基上(Fischer and Krebs, 1955; Krupa et al., 2004).真核生物蛋白被磷酸化的氨基酸主要包括丝氨酸、苏氨酸和酪氨酸.原核生物还可能发生在天冬氨酸、谷氨酸和组氨酸上(Wu et al., 2010).磷酸化蛋白上的磷酸基团可以由磷酸酶(phosphatase)除去.蛋白的磷酸化与去磷酸化的可逆调控参与诸多生命活动过程(Peck, 2003; Yang et al., 2004; Kim et al., 2016; Chen et al., 2017).磷酸化修饰通过改变蛋白结构或局部结构进而影响蛋白与特定蛋白间的互作、蛋白自身的活性和稳定性及蛋白的正常定位等.细胞中蛋白磷酸化水平是动态变化的, 细微变化即可导致细胞代谢及生理状态的变化(Laugesen et al., 2004; Khan et al., 2005; Agrawal and Thelen, 2006; Whiteman et al., 2008; Chao et al., 2014).研究表明, 植物的生长发育和对各种逆境响应的生理及信号调控过程都有蛋白磷酸化修饰的发生(de la Fuente van Bentem and Hirt, 2007; Prak et al., 2008; Ke et al., 2009; Wang et al., 2018; Yin et al., 2018; Zhang et al., 2019).因此, 解析关键蛋白磷酸化水平的变化规律, 可从分子水平揭示植物重要性状和代谢途径的调节机制. ...
Differential regulation of proteins and phosphoproteins in rice under drought stress 1 2009
... 研究显示, 真核生物细胞中约有1/3的蛋白被磷酸化修饰, 表明蛋白磷酸化修饰的普遍性和重要性(Hubbard and Cohen, 1993; Ficarro et al., 2002).蛋白磷酸化由蛋白激酶(protein kinase)催化完成.蛋白激酶将供体ATP (少数情况下用GTP)上的γ位磷酸基团以酯键的形式连接到底物蛋白的特定氨基酸侧链的羟基上(Fischer and Krebs, 1955; Krupa et al., 2004).真核生物蛋白被磷酸化的氨基酸主要包括丝氨酸、苏氨酸和酪氨酸.原核生物还可能发生在天冬氨酸、谷氨酸和组氨酸上(Wu et al., 2010).磷酸化蛋白上的磷酸基团可以由磷酸酶(phosphatase)除去.蛋白的磷酸化与去磷酸化的可逆调控参与诸多生命活动过程(Peck, 2003; Yang et al., 2004; Kim et al., 2016; Chen et al., 2017).磷酸化修饰通过改变蛋白结构或局部结构进而影响蛋白与特定蛋白间的互作、蛋白自身的活性和稳定性及蛋白的正常定位等.细胞中蛋白磷酸化水平是动态变化的, 细微变化即可导致细胞代谢及生理状态的变化(Laugesen et al., 2004; Khan et al., 2005; Agrawal and Thelen, 2006; Whiteman et al., 2008; Chao et al., 2014).研究表明, 植物的生长发育和对各种逆境响应的生理及信号调控过程都有蛋白磷酸化修饰的发生(de la Fuente van Bentem and Hirt, 2007; Prak et al., 2008; Ke et al., 2009; Wang et al., 2018; Yin et al., 2018; Zhang et al., 2019).因此, 解析关键蛋白磷酸化水平的变化规律, 可从分子水平揭示植物重要性状和代谢途径的调节机制. ...
Comprehensive phosphoproteome analysis in rice and identification of phosphoproteins responsive to different hormones/stresses 1 2005
... 研究显示, 真核生物细胞中约有1/3的蛋白被磷酸化修饰, 表明蛋白磷酸化修饰的普遍性和重要性(Hubbard and Cohen, 1993; Ficarro et al., 2002).蛋白磷酸化由蛋白激酶(protein kinase)催化完成.蛋白激酶将供体ATP (少数情况下用GTP)上的γ位磷酸基团以酯键的形式连接到底物蛋白的特定氨基酸侧链的羟基上(Fischer and Krebs, 1955; Krupa et al., 2004).真核生物蛋白被磷酸化的氨基酸主要包括丝氨酸、苏氨酸和酪氨酸.原核生物还可能发生在天冬氨酸、谷氨酸和组氨酸上(Wu et al., 2010).磷酸化蛋白上的磷酸基团可以由磷酸酶(phosphatase)除去.蛋白的磷酸化与去磷酸化的可逆调控参与诸多生命活动过程(Peck, 2003; Yang et al., 2004; Kim et al., 2016; Chen et al., 2017).磷酸化修饰通过改变蛋白结构或局部结构进而影响蛋白与特定蛋白间的互作、蛋白自身的活性和稳定性及蛋白的正常定位等.细胞中蛋白磷酸化水平是动态变化的, 细微变化即可导致细胞代谢及生理状态的变化(Laugesen et al., 2004; Khan et al., 2005; Agrawal and Thelen, 2006; Whiteman et al., 2008; Chao et al., 2014).研究表明, 植物的生长发育和对各种逆境响应的生理及信号调控过程都有蛋白磷酸化修饰的发生(de la Fuente van Bentem and Hirt, 2007; Prak et al., 2008; Ke et al., 2009; Wang et al., 2018; Yin et al., 2018; Zhang et al., 2019).因此, 解析关键蛋白磷酸化水平的变化规律, 可从分子水平揭示植物重要性状和代谢途径的调节机制. ...
Protein phosphatases involved in regulating mitosis: facts and hypotheses 1 2016
... 研究显示, 真核生物细胞中约有1/3的蛋白被磷酸化修饰, 表明蛋白磷酸化修饰的普遍性和重要性(Hubbard and Cohen, 1993; Ficarro et al., 2002).蛋白磷酸化由蛋白激酶(protein kinase)催化完成.蛋白激酶将供体ATP (少数情况下用GTP)上的γ位磷酸基团以酯键的形式连接到底物蛋白的特定氨基酸侧链的羟基上(Fischer and Krebs, 1955; Krupa et al., 2004).真核生物蛋白被磷酸化的氨基酸主要包括丝氨酸、苏氨酸和酪氨酸.原核生物还可能发生在天冬氨酸、谷氨酸和组氨酸上(Wu et al., 2010).磷酸化蛋白上的磷酸基团可以由磷酸酶(phosphatase)除去.蛋白的磷酸化与去磷酸化的可逆调控参与诸多生命活动过程(Peck, 2003; Yang et al., 2004; Kim et al., 2016; Chen et al., 2017).磷酸化修饰通过改变蛋白结构或局部结构进而影响蛋白与特定蛋白间的互作、蛋白自身的活性和稳定性及蛋白的正常定位等.细胞中蛋白磷酸化水平是动态变化的, 细微变化即可导致细胞代谢及生理状态的变化(Laugesen et al., 2004; Khan et al., 2005; Agrawal and Thelen, 2006; Whiteman et al., 2008; Chao et al., 2014).研究表明, 植物的生长发育和对各种逆境响应的生理及信号调控过程都有蛋白磷酸化修饰的发生(de la Fuente van Bentem and Hirt, 2007; Prak et al., 2008; Ke et al., 2009; Wang et al., 2018; Yin et al., 2018; Zhang et al., 2019).因此, 解析关键蛋白磷酸化水平的变化规律, 可从分子水平揭示植物重要性状和代谢途径的调节机制. ...
Specific recognition and detection of phosphorylated proteins using characteristics of metal ion 1 2007
Quantitative phosphoproteomics strategies for understanding protein kinase-mediated signal transduction pathways 1 2011
... 蛋白质翻译后修饰(posttranslational modification)是指通过酶促反应将一些化学基团共价连接到底物蛋白质分子上的化学修饰.蛋白质翻译后修饰对多种蛋白质正常行使功能起重要作用.蛋白质翻译后修饰的种类很多, 包括磷酸化、乙酰化、泛素化、糖基化、脂酰化及异戊烯基化等(Blom et al., 2004; Kosako and Nagano, 2011; Frost and Li, 2014; Zhu, 2017). ...
Structural modes of stabilization of permissive phosphorylation sites in protein kinases: distinct strategies in Ser/Thr and Tyr kinases 1 2004
... 研究显示, 真核生物细胞中约有1/3的蛋白被磷酸化修饰, 表明蛋白磷酸化修饰的普遍性和重要性(Hubbard and Cohen, 1993; Ficarro et al., 2002).蛋白磷酸化由蛋白激酶(protein kinase)催化完成.蛋白激酶将供体ATP (少数情况下用GTP)上的γ位磷酸基团以酯键的形式连接到底物蛋白的特定氨基酸侧链的羟基上(Fischer and Krebs, 1955; Krupa et al., 2004).真核生物蛋白被磷酸化的氨基酸主要包括丝氨酸、苏氨酸和酪氨酸.原核生物还可能发生在天冬氨酸、谷氨酸和组氨酸上(Wu et al., 2010).磷酸化蛋白上的磷酸基团可以由磷酸酶(phosphatase)除去.蛋白的磷酸化与去磷酸化的可逆调控参与诸多生命活动过程(Peck, 2003; Yang et al., 2004; Kim et al., 2016; Chen et al., 2017).磷酸化修饰通过改变蛋白结构或局部结构进而影响蛋白与特定蛋白间的互作、蛋白自身的活性和稳定性及蛋白的正常定位等.细胞中蛋白磷酸化水平是动态变化的, 细微变化即可导致细胞代谢及生理状态的变化(Laugesen et al., 2004; Khan et al., 2005; Agrawal and Thelen, 2006; Whiteman et al., 2008; Chao et al., 2014).研究表明, 植物的生长发育和对各种逆境响应的生理及信号调控过程都有蛋白磷酸化修饰的发生(de la Fuente van Bentem and Hirt, 2007; Prak et al., 2008; Ke et al., 2009; Wang et al., 2018; Yin et al., 2018; Zhang et al., 2019).因此, 解析关键蛋白磷酸化水平的变化规律, 可从分子水平揭示植物重要性状和代谢途径的调节机制. ...
Cleavage of structural proteins during the assembly of the head of bacteriophage T4 1 1970
Deciphering the plant phosphoproteome: tools and strategies for a challenging task 1 2004
... 研究显示, 真核生物细胞中约有1/3的蛋白被磷酸化修饰, 表明蛋白磷酸化修饰的普遍性和重要性(Hubbard and Cohen, 1993; Ficarro et al., 2002).蛋白磷酸化由蛋白激酶(protein kinase)催化完成.蛋白激酶将供体ATP (少数情况下用GTP)上的γ位磷酸基团以酯键的形式连接到底物蛋白的特定氨基酸侧链的羟基上(Fischer and Krebs, 1955; Krupa et al., 2004).真核生物蛋白被磷酸化的氨基酸主要包括丝氨酸、苏氨酸和酪氨酸.原核生物还可能发生在天冬氨酸、谷氨酸和组氨酸上(Wu et al., 2010).磷酸化蛋白上的磷酸基团可以由磷酸酶(phosphatase)除去.蛋白的磷酸化与去磷酸化的可逆调控参与诸多生命活动过程(Peck, 2003; Yang et al., 2004; Kim et al., 2016; Chen et al., 2017).磷酸化修饰通过改变蛋白结构或局部结构进而影响蛋白与特定蛋白间的互作、蛋白自身的活性和稳定性及蛋白的正常定位等.细胞中蛋白磷酸化水平是动态变化的, 细微变化即可导致细胞代谢及生理状态的变化(Laugesen et al., 2004; Khan et al., 2005; Agrawal and Thelen, 2006; Whiteman et al., 2008; Chao et al., 2014).研究表明, 植物的生长发育和对各种逆境响应的生理及信号调控过程都有蛋白磷酸化修饰的发生(de la Fuente van Bentem and Hirt, 2007; Prak et al., 2008; Ke et al., 2009; Wang et al., 2018; Yin et al., 2018; Zhang et al., 2019).因此, 解析关键蛋白磷酸化水平的变化规律, 可从分子水平揭示植物重要性状和代谢途径的调节机制. ...
Early phosphorylation events in biotic stress 1 2003
... 研究显示, 真核生物细胞中约有1/3的蛋白被磷酸化修饰, 表明蛋白磷酸化修饰的普遍性和重要性(Hubbard and Cohen, 1993; Ficarro et al., 2002).蛋白磷酸化由蛋白激酶(protein kinase)催化完成.蛋白激酶将供体ATP (少数情况下用GTP)上的γ位磷酸基团以酯键的形式连接到底物蛋白的特定氨基酸侧链的羟基上(Fischer and Krebs, 1955; Krupa et al., 2004).真核生物蛋白被磷酸化的氨基酸主要包括丝氨酸、苏氨酸和酪氨酸.原核生物还可能发生在天冬氨酸、谷氨酸和组氨酸上(Wu et al., 2010).磷酸化蛋白上的磷酸基团可以由磷酸酶(phosphatase)除去.蛋白的磷酸化与去磷酸化的可逆调控参与诸多生命活动过程(Peck, 2003; Yang et al., 2004; Kim et al., 2016; Chen et al., 2017).磷酸化修饰通过改变蛋白结构或局部结构进而影响蛋白与特定蛋白间的互作、蛋白自身的活性和稳定性及蛋白的正常定位等.细胞中蛋白磷酸化水平是动态变化的, 细微变化即可导致细胞代谢及生理状态的变化(Laugesen et al., 2004; Khan et al., 2005; Agrawal and Thelen, 2006; Whiteman et al., 2008; Chao et al., 2014).研究表明, 植物的生长发育和对各种逆境响应的生理及信号调控过程都有蛋白磷酸化修饰的发生(de la Fuente van Bentem and Hirt, 2007; Prak et al., 2008; Ke et al., 2009; Wang et al., 2018; Yin et al., 2018; Zhang et al., 2019).因此, 解析关键蛋白磷酸化水平的变化规律, 可从分子水平揭示植物重要性状和代谢途径的调节机制. ...
Multiple phosphorylations in the C-terminal tail of plant plasma membrane aquaporins: role in subcellular trafficking of AtPIP2;1 in response to salt stress 1 2008
... 研究显示, 真核生物细胞中约有1/3的蛋白被磷酸化修饰, 表明蛋白磷酸化修饰的普遍性和重要性(Hubbard and Cohen, 1993; Ficarro et al., 2002).蛋白磷酸化由蛋白激酶(protein kinase)催化完成.蛋白激酶将供体ATP (少数情况下用GTP)上的γ位磷酸基团以酯键的形式连接到底物蛋白的特定氨基酸侧链的羟基上(Fischer and Krebs, 1955; Krupa et al., 2004).真核生物蛋白被磷酸化的氨基酸主要包括丝氨酸、苏氨酸和酪氨酸.原核生物还可能发生在天冬氨酸、谷氨酸和组氨酸上(Wu et al., 2010).磷酸化蛋白上的磷酸基团可以由磷酸酶(phosphatase)除去.蛋白的磷酸化与去磷酸化的可逆调控参与诸多生命活动过程(Peck, 2003; Yang et al., 2004; Kim et al., 2016; Chen et al., 2017).磷酸化修饰通过改变蛋白结构或局部结构进而影响蛋白与特定蛋白间的互作、蛋白自身的活性和稳定性及蛋白的正常定位等.细胞中蛋白磷酸化水平是动态变化的, 细微变化即可导致细胞代谢及生理状态的变化(Laugesen et al., 2004; Khan et al., 2005; Agrawal and Thelen, 2006; Whiteman et al., 2008; Chao et al., 2014).研究表明, 植物的生长发育和对各种逆境响应的生理及信号调控过程都有蛋白磷酸化修饰的发生(de la Fuente van Bentem and Hirt, 2007; Prak et al., 2008; Ke et al., 2009; Wang et al., 2018; Yin et al., 2018; Zhang et al., 2019).因此, 解析关键蛋白磷酸化水平的变化规律, 可从分子水平揭示植物重要性状和代谢途径的调节机制. ...
Reciprocal regulation of the TOR kinase and ABA receptor balances plant growth and stress response 1 2018
... 研究显示, 真核生物细胞中约有1/3的蛋白被磷酸化修饰, 表明蛋白磷酸化修饰的普遍性和重要性(Hubbard and Cohen, 1993; Ficarro et al., 2002).蛋白磷酸化由蛋白激酶(protein kinase)催化完成.蛋白激酶将供体ATP (少数情况下用GTP)上的γ位磷酸基团以酯键的形式连接到底物蛋白的特定氨基酸侧链的羟基上(Fischer and Krebs, 1955; Krupa et al., 2004).真核生物蛋白被磷酸化的氨基酸主要包括丝氨酸、苏氨酸和酪氨酸.原核生物还可能发生在天冬氨酸、谷氨酸和组氨酸上(Wu et al., 2010).磷酸化蛋白上的磷酸基团可以由磷酸酶(phosphatase)除去.蛋白的磷酸化与去磷酸化的可逆调控参与诸多生命活动过程(Peck, 2003; Yang et al., 2004; Kim et al., 2016; Chen et al., 2017).磷酸化修饰通过改变蛋白结构或局部结构进而影响蛋白与特定蛋白间的互作、蛋白自身的活性和稳定性及蛋白的正常定位等.细胞中蛋白磷酸化水平是动态变化的, 细微变化即可导致细胞代谢及生理状态的变化(Laugesen et al., 2004; Khan et al., 2005; Agrawal and Thelen, 2006; Whiteman et al., 2008; Chao et al., 2014).研究表明, 植物的生长发育和对各种逆境响应的生理及信号调控过程都有蛋白磷酸化修饰的发生(de la Fuente van Bentem and Hirt, 2007; Prak et al., 2008; Ke et al., 2009; Wang et al., 2018; Yin et al., 2018; Zhang et al., 2019).因此, 解析关键蛋白磷酸化水平的变化规律, 可从分子水平揭示植物重要性状和代谢途径的调节机制. ...
A proteomic and phosphoproteomic analysis of Oryza sativa plasma membrane and vacuolar membrane 1 2008
... 研究显示, 真核生物细胞中约有1/3的蛋白被磷酸化修饰, 表明蛋白磷酸化修饰的普遍性和重要性(Hubbard and Cohen, 1993; Ficarro et al., 2002).蛋白磷酸化由蛋白激酶(protein kinase)催化完成.蛋白激酶将供体ATP (少数情况下用GTP)上的γ位磷酸基团以酯键的形式连接到底物蛋白的特定氨基酸侧链的羟基上(Fischer and Krebs, 1955; Krupa et al., 2004).真核生物蛋白被磷酸化的氨基酸主要包括丝氨酸、苏氨酸和酪氨酸.原核生物还可能发生在天冬氨酸、谷氨酸和组氨酸上(Wu et al., 2010).磷酸化蛋白上的磷酸基团可以由磷酸酶(phosphatase)除去.蛋白的磷酸化与去磷酸化的可逆调控参与诸多生命活动过程(Peck, 2003; Yang et al., 2004; Kim et al., 2016; Chen et al., 2017).磷酸化修饰通过改变蛋白结构或局部结构进而影响蛋白与特定蛋白间的互作、蛋白自身的活性和稳定性及蛋白的正常定位等.细胞中蛋白磷酸化水平是动态变化的, 细微变化即可导致细胞代谢及生理状态的变化(Laugesen et al., 2004; Khan et al., 2005; Agrawal and Thelen, 2006; Whiteman et al., 2008; Chao et al., 2014).研究表明, 植物的生长发育和对各种逆境响应的生理及信号调控过程都有蛋白磷酸化修饰的发生(de la Fuente van Bentem and Hirt, 2007; Prak et al., 2008; Ke et al., 2009; Wang et al., 2018; Yin et al., 2018; Zhang et al., 2019).因此, 解析关键蛋白磷酸化水平的变化规律, 可从分子水平揭示植物重要性状和代谢途径的调节机制. ...
Dissecting the M phase- specific phosphorylation of serine-proline or threonine- proline motifs 1 2010
... 研究显示, 真核生物细胞中约有1/3的蛋白被磷酸化修饰, 表明蛋白磷酸化修饰的普遍性和重要性(Hubbard and Cohen, 1993; Ficarro et al., 2002).蛋白磷酸化由蛋白激酶(protein kinase)催化完成.蛋白激酶将供体ATP (少数情况下用GTP)上的γ位磷酸基团以酯键的形式连接到底物蛋白的特定氨基酸侧链的羟基上(Fischer and Krebs, 1955; Krupa et al., 2004).真核生物蛋白被磷酸化的氨基酸主要包括丝氨酸、苏氨酸和酪氨酸.原核生物还可能发生在天冬氨酸、谷氨酸和组氨酸上(Wu et al., 2010).磷酸化蛋白上的磷酸基团可以由磷酸酶(phosphatase)除去.蛋白的磷酸化与去磷酸化的可逆调控参与诸多生命活动过程(Peck, 2003; Yang et al., 2004; Kim et al., 2016; Chen et al., 2017).磷酸化修饰通过改变蛋白结构或局部结构进而影响蛋白与特定蛋白间的互作、蛋白自身的活性和稳定性及蛋白的正常定位等.细胞中蛋白磷酸化水平是动态变化的, 细微变化即可导致细胞代谢及生理状态的变化(Laugesen et al., 2004; Khan et al., 2005; Agrawal and Thelen, 2006; Whiteman et al., 2008; Chao et al., 2014).研究表明, 植物的生长发育和对各种逆境响应的生理及信号调控过程都有蛋白磷酸化修饰的发生(de la Fuente van Bentem and Hirt, 2007; Prak et al., 2008; Ke et al., 2009; Wang et al., 2018; Yin et al., 2018; Zhang et al., 2019).因此, 解析关键蛋白磷酸化水平的变化规律, 可从分子水平揭示植物重要性状和代谢途径的调节机制. ...
Recent advances of protein phosphorylation in proteome 1 2004
... 研究显示, 真核生物细胞中约有1/3的蛋白被磷酸化修饰, 表明蛋白磷酸化修饰的普遍性和重要性(Hubbard and Cohen, 1993; Ficarro et al., 2002).蛋白磷酸化由蛋白激酶(protein kinase)催化完成.蛋白激酶将供体ATP (少数情况下用GTP)上的γ位磷酸基团以酯键的形式连接到底物蛋白的特定氨基酸侧链的羟基上(Fischer and Krebs, 1955; Krupa et al., 2004).真核生物蛋白被磷酸化的氨基酸主要包括丝氨酸、苏氨酸和酪氨酸.原核生物还可能发生在天冬氨酸、谷氨酸和组氨酸上(Wu et al., 2010).磷酸化蛋白上的磷酸基团可以由磷酸酶(phosphatase)除去.蛋白的磷酸化与去磷酸化的可逆调控参与诸多生命活动过程(Peck, 2003; Yang et al., 2004; Kim et al., 2016; Chen et al., 2017).磷酸化修饰通过改变蛋白结构或局部结构进而影响蛋白与特定蛋白间的互作、蛋白自身的活性和稳定性及蛋白的正常定位等.细胞中蛋白磷酸化水平是动态变化的, 细微变化即可导致细胞代谢及生理状态的变化(Laugesen et al., 2004; Khan et al., 2005; Agrawal and Thelen, 2006; Whiteman et al., 2008; Chao et al., 2014).研究表明, 植物的生长发育和对各种逆境响应的生理及信号调控过程都有蛋白磷酸化修饰的发生(de la Fuente van Bentem and Hirt, 2007; Prak et al., 2008; Ke et al., 2009; Wang et al., 2018; Yin et al., 2018; Zhang et al., 2019).因此, 解析关键蛋白磷酸化水平的变化规律, 可从分子水平揭示植物重要性状和代谢途径的调节机制. ...
Phosphoproteomics: protein phosphorylation in regulation of seed germination and plant growth 1 2018
... 研究显示, 真核生物细胞中约有1/3的蛋白被磷酸化修饰, 表明蛋白磷酸化修饰的普遍性和重要性(Hubbard and Cohen, 1993; Ficarro et al., 2002).蛋白磷酸化由蛋白激酶(protein kinase)催化完成.蛋白激酶将供体ATP (少数情况下用GTP)上的γ位磷酸基团以酯键的形式连接到底物蛋白的特定氨基酸侧链的羟基上(Fischer and Krebs, 1955; Krupa et al., 2004).真核生物蛋白被磷酸化的氨基酸主要包括丝氨酸、苏氨酸和酪氨酸.原核生物还可能发生在天冬氨酸、谷氨酸和组氨酸上(Wu et al., 2010).磷酸化蛋白上的磷酸基团可以由磷酸酶(phosphatase)除去.蛋白的磷酸化与去磷酸化的可逆调控参与诸多生命活动过程(Peck, 2003; Yang et al., 2004; Kim et al., 2016; Chen et al., 2017).磷酸化修饰通过改变蛋白结构或局部结构进而影响蛋白与特定蛋白间的互作、蛋白自身的活性和稳定性及蛋白的正常定位等.细胞中蛋白磷酸化水平是动态变化的, 细微变化即可导致细胞代谢及生理状态的变化(Laugesen et al., 2004; Khan et al., 2005; Agrawal and Thelen, 2006; Whiteman et al., 2008; Chao et al., 2014).研究表明, 植物的生长发育和对各种逆境响应的生理及信号调控过程都有蛋白磷酸化修饰的发生(de la Fuente van Bentem and Hirt, 2007; Prak et al., 2008; Ke et al., 2009; Wang et al., 2018; Yin et al., 2018; Zhang et al., 2019).因此, 解析关键蛋白磷酸化水平的变化规律, 可从分子水平揭示植物重要性状和代谢途径的调节机制. ...
Phosphorylation-mediated dynamics of nitrate transceptor NRT1.1 regulate auxin flux and nitrate signaling in lateral root growth 1 2019
... 研究显示, 真核生物细胞中约有1/3的蛋白被磷酸化修饰, 表明蛋白磷酸化修饰的普遍性和重要性(Hubbard and Cohen, 1993; Ficarro et al., 2002).蛋白磷酸化由蛋白激酶(protein kinase)催化完成.蛋白激酶将供体ATP (少数情况下用GTP)上的γ位磷酸基团以酯键的形式连接到底物蛋白的特定氨基酸侧链的羟基上(Fischer and Krebs, 1955; Krupa et al., 2004).真核生物蛋白被磷酸化的氨基酸主要包括丝氨酸、苏氨酸和酪氨酸.原核生物还可能发生在天冬氨酸、谷氨酸和组氨酸上(Wu et al., 2010).磷酸化蛋白上的磷酸基团可以由磷酸酶(phosphatase)除去.蛋白的磷酸化与去磷酸化的可逆调控参与诸多生命活动过程(Peck, 2003; Yang et al., 2004; Kim et al., 2016; Chen et al., 2017).磷酸化修饰通过改变蛋白结构或局部结构进而影响蛋白与特定蛋白间的互作、蛋白自身的活性和稳定性及蛋白的正常定位等.细胞中蛋白磷酸化水平是动态变化的, 细微变化即可导致细胞代谢及生理状态的变化(Laugesen et al., 2004; Khan et al., 2005; Agrawal and Thelen, 2006; Whiteman et al., 2008; Chao et al., 2014).研究表明, 植物的生长发育和对各种逆境响应的生理及信号调控过程都有蛋白磷酸化修饰的发生(de la Fuente van Bentem and Hirt, 2007; Prak et al., 2008; Ke et al., 2009; Wang et al., 2018; Yin et al., 2018; Zhang et al., 2019).因此, 解析关键蛋白磷酸化水平的变化规律, 可从分子水平揭示植物重要性状和代谢途径的调节机制. ...
Regulation of p53 acetylation 1 2017
... 蛋白质翻译后修饰(posttranslational modification)是指通过酶促反应将一些化学基团共价连接到底物蛋白质分子上的化学修饰.蛋白质翻译后修饰对多种蛋白质正常行使功能起重要作用.蛋白质翻译后修饰的种类很多, 包括磷酸化、乙酰化、泛素化、糖基化、脂酰化及异戊烯基化等(Blom et al., 2004; Kosako and Nagano, 2011; Frost and Li, 2014; Zhu, 2017). ...
 ,*中国农业大学生物学院, 植物生理学与生物化学国家重点实验室, 北京 100193
,*中国农业大学生物学院, 植物生理学与生物化学国家重点实验室, 北京 100193
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT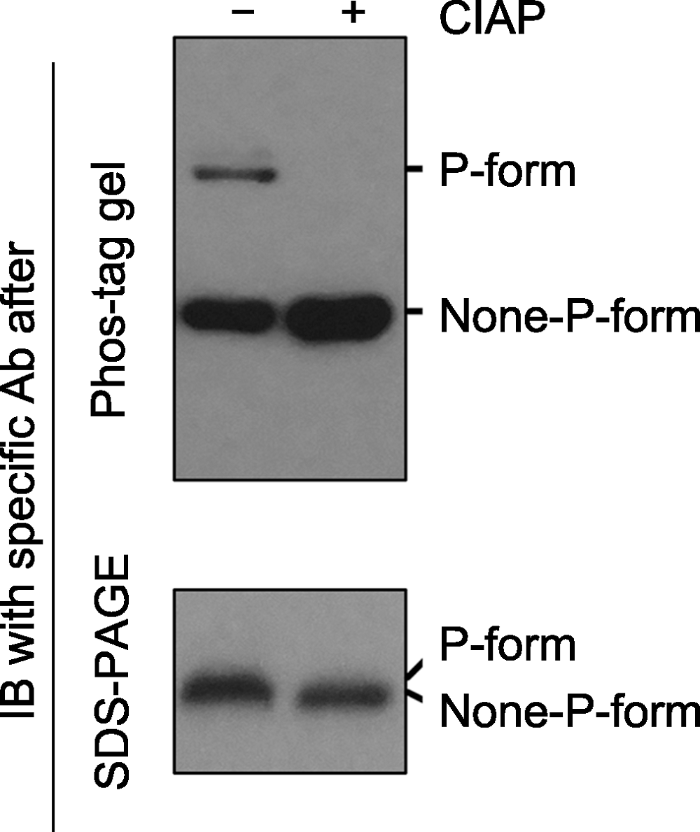 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT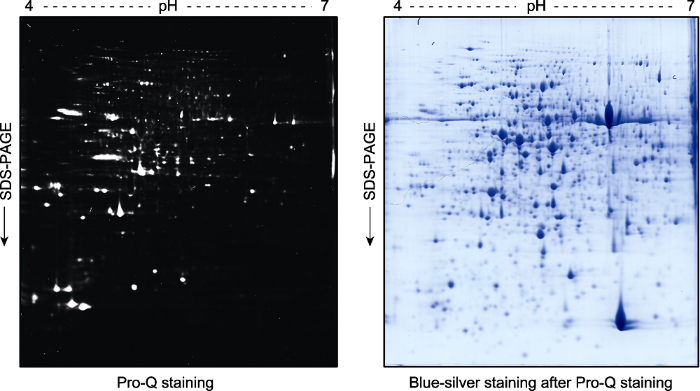 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}