Expression Characterization of Rice OsPR10A and Its Function in Response to Drought Stress
Tong Zhang1, Yalu Guo1,2, Yue Chen1, Jinjiao Ma1, Jinping Lan1,3, Gaowei Yan1, Yuqing Liu1, Shan Xu1, Liyun Li1, Guozhen Liu,1,*, Shijuan Dou,1,*1 College of Life Sciences, Hebei Agricultural University, Baoding 071001, China 2 Agricultural Genomics Institute at Shenzhen, Chinese Academy of Agricultural Sciences, Shenzhen 518116, China 3 Research Center for Life Sciences, Hebei North University, Zhangjiakou 075000, China
Abstract In this study, we surveyed the expression patterns of rice OsPR10A protein by using western blot analysis for protein samples extracted from different tissues at different developmental stages and a variety of abiotic stresses. The expression of OsPR10A was upregulated significantly under drought, salt stress and hormone treatments such as methyl jasmonate and abscisic acid, which suggests that OsPR10A protein plays a role in drought and salt stress response. To test this hypothesis, we constructed a vector for overexpression of OsPR10A protein and obtained homozygous transgenic lines via Agrobacterium-mediated rice transformation. Phenotypic investigation in field experiments revealed shorter plant height and spike length as well as lower seed setting rate in transgenic lines than controls. At seed germination stage, drought stress treatment was applied by using 20% PEG6000. Root and shoot length was significantly longer in OsPR10A overexpression lines than the wild-type control, which supports that overexpression of OsPR10A protein enhanced the drought tolerance of rice during germination. The study increases our understanding of the function of rice OsPR10A protein. Keywords:rice;drought stress;pathogenesis-related protein;western blot;antibody-based proteomics
PDF (1893KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 张彤, 郭亚璐, 陈悦, 马金姣, 兰金苹, 燕高伟, 刘玉晴, 徐珊, 李莉云, 刘国振, 窦世娟. 水稻OsPR10A的表达特征及其在干旱胁迫应答过程中的功能. 植物学报, 2019, 54(6): 711-722 doi:10.11983/CBB19042 Zhang Tong, Guo Yalu, Chen Yue, Ma Jinjiao, Lan Jinping, Yan Gaowei, Liu Yuqing, Xu Shan, Li Liyun, Liu Guozhen, Dou Shijuan. Expression Characterization of Rice OsPR10A and Its Function in Response to Drought Stress. Chinese Bulletin of Botany, 2019, 54(6): 711-722 doi:10.11983/CBB19042
水稻(Oryza sativa)是世界上最重要的粮食作物之一, 其高产和稳产对全球尤其是亚洲地区的粮食安全具有重要意义。水稻生长常受到冷害、高温、水淹、干旱和盐碱等恶劣环境的威胁(Thakur et al., 2010)。 因此, 了解水稻胁迫应答机理对培育耐逆水稻品种具有重要意义。
为应对多变的自然环境, 植物进化出复杂的抗性机制(Nürnberger et al., 2004; Ausubel, 2005)。例如, 在受到病原菌侵害或其它生物胁迫时, 植物会激活自身的防御体系来应对侵害。病程相关(pathogenesis- related, PR)蛋白是一类受病原物诱导的蛋白质。水稻是单子叶植物遗传研究的模式生物, 其基因组包含900多个PR基因(窦世娟等, 2014)。根据蛋白质的结构, 可将PR蛋白分为17个亚类, 不同亚类的PR蛋白生物学功能也各有不同。探究水稻PR基因的功能有助于深入了解水稻的抗逆机理。大量研究表明, 水稻PR基因在抵抗生物及非生物逆境胁迫中发挥重要作用, 调控特定PR基因的表达会影响水稻的抗逆反应, PR蛋白在抗病反应中也被诱导表达(李雪姣等, 2014)。PR1家族的OsPR1A和OsPR1B的转录水平易受病原菌诱导, 可作为抗病反应发生的分子标记(Ponciano et al., 2006)。在水稻与白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)互作反应中, OsPR1A和OsPR1B蛋白质在Xoo侵染水稻叶片144小时后大量表达(Wu et al., 2011)。在水稻幼苗中, OsPR1A和OsPR1B基因的转录与光处理密切相关(Agrawal et al., 2000b), 且外源施加茉莉酸(jasmonic acid, JA)和乙烯(ethylene, ET)能显著增加OsPR1B基因的转录(Luan and Zhou, 2015)。GNS1属于PR2家族, 具有β-1,3-葡聚糖酶活性, 在真菌诱导下, 水稻中GNS1基因的转录和蛋白质表达水平均明显升高; 在黑暗处理时, GNS1基因表达水平也有所上升(Nishizawa et al., 2003)。转PR3基因的水稻对纹枯病的抗性增强(Datta et al., 2001)。超表达OsPR4A的水稻植株在幼苗期和孕穗期抗旱性明显增强(Wang et al., 2011)。水稻PR1A、PR5、PR8和PR16蛋白质随盐胁迫时间的延长下调表达, 而PR2在干旱、高盐及水淹胁迫中均被诱导(高庆华等, 2013)。OsWRKY67与OsPR1A和OsPR10的启动子区结合进而提高对白叶枯病菌的抗性(Liu et al., 2018)。在其它植物中也有大量PR基因的相关报道。拟南芥(Arabidopsis thaliana)受到南方根结线虫(Meloidogyne incognita)侵袭后, PR1、PR2和PR5的表达被大量诱导(Hamamouch et al., 2011)。干旱胁迫下, 拟南芥中PR1、PR2和PR5基因表达升高(Seo et al., 2008)。在烟草(Nicotiana tabacum)中表达OsGLP1基因可增强其对真菌病原物的抗性(Banerjee et al., 2010)。
PR10蛋白结构保守且功能多样, 在植物抗逆方面的功能也有报道(谢纯政等, 2008)。水稻OsPR10A蛋白分子量约为17 kDa, 经生化验证其具有RNA酶活性, 经GUS染色确定OsPR10A的转录受到多种方式的调控, 如生物胁迫Xoo的侵染、非生物胁迫伤害及缺磷。此外, 在叶片的正常衰老过程中OsPR10A也会被诱导表达(Huang et al., 2016)。PR10A (PBZ1)在细胞衰亡的组织中积累, 可诱导水稻细胞死亡, 也能通过其RNA酶活性诱导拟南芥和烟草细胞死亡(Kim et al., 2011)。蛋白质免疫印迹(western blot, WB)分析发现, OsPR10A在水稻受到Xoo侵染后144小时大量表达, 而且不亲和互作中表达量高于亲和互作反应(Wu et al., 2011)。超表达OsPR10A增强了水稻对白叶枯病菌的抗性(Huang et al., 2016)。OsPR10A的上游有转录因子结合位点, 可能受转录因子OsWRKY6的调控, OsWRKY6与类W-box元件1 (WLE1)物理结合, 正向调控OsPR10A的表达(Choi et al., 2015)。序列同源性分析表明, OsPR10A和RSOsPR10高度相似, 具有相似的酶活性质与功能(Hashimoto et al., 2004)。RSOsPR10是水稻根部特有的病程相关基因(Hashimoto et al., 2004), 在干旱和高盐胁迫下均可被诱导表达(Takeuchi et al., 2011); 在根皮细胞中超表达RSOsPR10的水稻植株对干旱胁迫具有更强的耐受性, 但对盐胁迫的耐受性较差(Takeuchi et al., 2016)。超表达JIOsPR10增强了水稻对稻瘟病菌的抗性, 证明其在水稻防卫机制中发挥作用; 半定量RT-PCR分析表明, 高盐和干旱胁迫条件下, 水稻JIOsPR10的表达上调(Wu et al., 2016); WB检测发现, 其在正常组织(如花和根)中都表达, 衰老及伤害条件也能提高其表达水平(Kim et al., 2008)。
蛋白质是生物功能的主要执行者, 了解蛋白质的表达丰度、修饰及其变化是蛋白质组学的主要目标(Pandey and Mann, 2000)。利用蛋白质特异性抗体可对特定的目标蛋白进行靶向研究, 这一理念被称为基于抗体的蛋白质组学(antibody-based proteomics, AbP) (Uhlén et al., 2005)。通过免疫学方法对蛋白质表达特性展开调查, 具有靶向性强、灵敏度高、操作简便和容易对大量样品进行分析等优点。在水稻及其它植物研究中也可借鉴这一思路, 水稻抗体资源库和蛋白质样品资源库的构建工作也已经开展(刘国振等, 2011; 张剑硕等, 2018)。
所用水稻(Oryza sativa L.)品种为TP309, 基因克隆用大肠杆菌(Escherichia coli)菌株DH5α, 重组蛋白质表达菌株为BL21 (DE3) pLysS。蛋白质原核表达载体为pET30a和pGST (Liu et al., 2002)。水稻OsPR10A基因全长cDNA质粒购自日本农业生物资源研究所水稻基因组资源中心(Rice Genome Resource Center, National Institute of Agrobiological Sciences)。质粒pUC57-3HA由南京金斯瑞生物科技有限公司合成, pEASY-T1购自上海生工生物技术有限公司, pUBI-C4300由Pamela Ronald博士(UC Davis, USA)提供。茉莉酸甲酯和脱落酸等试剂购自北京博奥拓达科技有限公司。
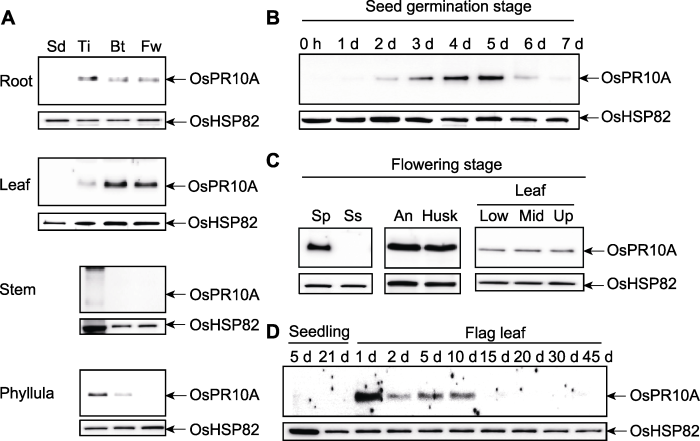
(A) OsPR10A在水稻不同生长时期和组织部位的表达(Sd: 苗期; Ti: 分蘖期; Bt: 孕穗期; Fw: 开花期); (B) OsPR10A在萌发期种子中的表达; (C) OsPR10A在开花期组织中的表达(Sp: 穗; Ss: 穗轴; An: 花药; Husk: 颖壳; Low: 叶下部; Mid: 叶中部; Up: 叶上部); (D) OsPR10A在幼苗和旗叶中的表达 Figure 1Expression of OsPR10A during rice growth and development
(A) Expression of OsPR10A in different growth stages and tissue parts of rice (Sd: Seedling stage; Ti: Tillering stage; Bt: Booting stage; Fw: Flowering stage); (B) Expression of OsPR10A in seeds during germination; (C) Expression of OsPR10A in tissues during flowering stage (Sp: Spike; Ss: Spike-stalk; An: Anther; Husk: Husk; Low: Lower part of leaf; Mid: Middle part of leaf; Up: Upper part of leaf); (D) Expression of OsPR10A in seedling and flag leaf
Table 1 表1 表1OsPR10A基因的转录分析 Table 1Transcriptome analysis of the OsPR10A gene
Library description
FPKM
Shoots
50.1661
Leaves-20 day
144.7290
Pre-emergence inflorescence
4.1989
Post-emergence inflorescence
88.9886
Anther
0.5100
Pistil
2.3483
Seed-5 DAP
77.4876
Seed-10 DAP
0.0000
Embryo-25 DAP
1.5052
Endosperm-25 DAP
2.3371
FPKM: Expected number of fragments per kilobase of transcript sequence per millions base pairs sequenced; DAP: Days after pollination FPKM: 每百万片段中对应来自某一基因的每千碱基的片段数目; DAP: 授粉后天数
(A) OsPR10A在干旱胁迫下的表达; (B) OsPR10A在盐胁迫下的表达; (C) OsPR10A在外源施加茉莉酸甲酯后的表达; (D) OsPR10A在外源施加脱落酸后的表达。MeJA: 茉莉酸甲酯; ABA: 脱落酸 Figure 2Dynamic expression of rice OsPR10A under abiotic stress
(A) Expression of OsPR10A under drought stress; (B) Expression of OsPR10A under salt stress; (C) Expression of OsPR10A modulated by exogenous MeJA; (D) Expression of OsPR10A modulated by exogenous ABA. MeJA: Methyl jasmonate; ABA: Abscisic acid
A721, A726, A728 and A730 represent different transgenic lines, respectively; Lane 1-7 or 1-8 represent different plants of the same transgenic line, respectively; WT: Wildtype
最初报道的PR基因功能大都与抗病相关, 近年来发现PR基因在抵御非生物胁迫中也行使功能。OsPR10A启动子驱动的GUS受磷饥饿、伤害和Xoo侵染的诱导, 超表达OsPR10A的水稻和拟南芥在磷饥饿条件下根长明显长于野生型(Huang et al., 2016)。本研究则表明OsPR10A超表达能增强水稻种子萌发期的耐旱性, 丰富了人们对OsPR10A功能的认识。
本研究还发现, 在离体水稻叶片激素处理过程中, OsPR10A的表达丰度受MeJA和ABA诱导, 表明该蛋白功能的发挥与JA和ABA的信号转导途径有关。与野生型相比, OsJAZ1超表达植株对干旱敏感, 同时对MeJA和ABA表现更为敏感, RNA-seq分析结果表明, 在干旱处理下, JA和ABA信号转导途径中许多基因的表达水平在OsJAZ1超表达植株和野生型之间差异显著, 表明JA和ABA介导的途径参与水稻对干旱胁迫的抵御(Fu et al., 2017)。水稻转录因子OsbZIP23与耐旱性有关, 同时也是ABA信号途径和ABA合成的关键调控子(Zong et al., 2016)。水稻OsASR5与水稻耐旱性有关, 超表达OsASR5使种子萌发期和萌发后期内源ABA水平增高, 对外源施加的ABA也更为敏感(Li et al., 2017)。水稻ABA8ox3是ABA生物合成的关键基因, 其转基因RNAi株系耐旱性更强, 而超表达株系对干旱胁迫更为敏感(Cai et al., 2015)。本研究中, OsPR10A分别受干旱胁迫和激素(MeJA和ABA)的诱导, 推测该蛋白可能通过JA和ABA相关的信号转导途径增强水稻的耐旱性。
研究表明, OsPR10A可受SA、JA、ET和ABA等激素的诱导(Hwang et al., 2008), 且OsPR10A启动子687-637之间的WLE1在SA介导的OsPR10A表达中起重要作用(McGee et al., 2001)。RSOsPR10的表达受生物和非生物逆境诱导, 并可能受JA诱导(Hashimoto et al., 2004)。本研究检测了激素处理条件下OsPR10A蛋白的表达丰度变化, 后续将深入研究JA和ABA等激素如何发挥功能。
AgrawalGK, JwaNS, RakwalR ( 2000a). A novel rice (Oryza sativa L.) acidic PR1 gene highly responsive to cut, phytohormones, and protein phosphatase inhibitors Biochem Biophys Res Commun 274, 157-165. [本文引用: 1]
AgrawalGK, RakwalR, JwaNS ( 2000b). Rice (Oryza sativa L.) OsPR1b gene is phytohormonally regulated in close interaction with light signals Biochem Biophys Res Commun 278, 290-298. [本文引用: 1]
AusubelFM ( 2005). Are innate immune signaling pathways in plants and animals conserved? Nat Immunol 6, 973-979. [本文引用: 1]
BanerjeeJ, DasN, DeyP, MaitiMK ( 2010). Transgenically expressed rice germin-like protein 1 in tobacco causes hyper-accumulation of H2O2 and reinforcement of the cell wall components Biochem Biophys Res Commun 402, 637-643. [本文引用: 1]
CaiSL, JiangGB, YeNH, ChuZZ, XuXZ, ZhangJH, ZhuGH ( 2015). A key ABA catabolic gene, OsABA8ox3 , is involved in drought stress resistance in rice PLoS One 10, e0116646. [本文引用: 1]
ChoiC, HwangSH, FangIR, KwonSI, ParkSR, AhnI, KimJB, HwangDJ ( 2015). Molecular characterization of Oryza sativa WRKY6, which binds to W-box-like element 1 of the Oryza sativa pathogenesis-related (PR) 10a promoter and confers reduced susceptibility to pathogens New Phytol 208, 846-859. [本文引用: 1]
DansanaPK, KothariKS, VijS, TyagiAK ( 2014). OsiSAP1 overexpression improves water-deficit stress tolerance in transgenic rice by affecting expression of endogenous stress-related genes Plant Cell Rep 33, 1425-1440. [本文引用: 1]
DattaK, TuJM, OlivaN, OnaI, VelazhahanR, MewTW, MuthukrishnanS, DattaSK ( 2001). Enhanced resistance to sheath blight by constitutive expression of infection-related rice chitinase in transgenic elite indica rice cultivars Plant Sci 160, 405-414. [本文引用: 1]
DuanYB, ZhaiCG, LiH, LiJ, MeiWQ, GuiHP, NiDH, SongFS, LiL, ZhangWG, YangJB ( 2012). An efficient and high-throughput protocol for Agrobacterium-mediated transformation based on phosphomannose isomerase positive selection in Japonica rice(Oryza sativa L.) Plant Cell Rep 31, 1611-1624. [本文引用: 1]
FuJ, WuH, MaSQ, XiangDH, LiuRY, XiongLZ ( 2017). OsJAZ1 attenuates drought resistance by regulating JA and ABA signaling in rice Front Plant Sci 8, 2108. [本文引用: 1]
HashimotoM, KisselevaL, SawaS, FurukawaT, KomatsuS, KoshibaT ( 2004). A novel rice PR10 protein, RSOsPR10, specifically induced in roots by biotic and abiotic stresses, possibly via the jasmonic acid signaling pathway Plant Cell Physiol 45, 550-559. [本文引用: 3]
HuangLF, LinKH, HeSL, ChenJL, JiangJZ, ChenBH, HouYS, ChenRS, HongCY, HoSL ( 2016). Multiple patterns of regulation and overexpression of a ribonuclease-like pathogenesis-related protein gene,OsPR10a, conferring disease resistance in rice and Arabidopsis PLoS One 11, e0156414. [本文引用: 3]
HwangSH, LeeIA, YieSW, HwangDJ ( 2008). Identification of an OsPR10a promoter region responsive to salicylic acid Planta 227, 1141-1150. [本文引用: 1]
KimSG, KimST, WangYM, YuS, ChoiIS, KimYC, KimWT, AgrawalGK, RakwalR, KangKY ( 2011). The RNase activity of rice probenazole-induced protein1 (PBZ1) plays a key role in cell death in plants Mol Cells 31, 25-31. [本文引用: 1]
KimST, YuS, KangYH, KimSG, KimJY, KimSH, KangKY ( 2008). The rice pathogen-related protein 10 (JIOsPR10) is induced by abiotic and biotic stresses and exhibits ribonuclease activity Plant Cell Rep 27, 593-603. [本文引用: 1]
LiGW, XieXS ( 2011). Central dogma at the single-molecule level in living cells Nature 475, 308-315. [本文引用: 3]
LiJJ, LiY, YinZG, JiangJH, ZhangMH, GuoX, YeZJ, ZhaoY, XiongHY, ZhangZY, ShaoYJ, JiangCH, ZhangHL, AnG, PaekNC, AliJ, LiZC ( 2017). OsASR5 enhances drought tolerance through a stomatal closure pathway associated with ABA and H2O2 signaling in rice Plant Biotechnol J 15, 183-196. [本文引用: 1]
LiXM, BaiH, WangXY, LiLY, CaoYH, WeiJ, LiuYM, LiuLJ, GongXD, WuL, LiuSQ, LiuGZ ( 2011). Identification and validation of rice reference proteins for western blotting J Exp Bot 62, 4763-4772.
LiuGZ, PiLY, WalkerJC, RonaldPC, SongWY ( 2002). Biochemical characterization of the kinase domain of the rice disease resistance receptor-like kinase XA21 J Biol Chem 277, 20264-20269. [本文引用: 1]
LiuQ, LiX, YanSJ, YuT, YangJY, DongJF, ZhangSH, ZhaoJL, YangTF, MaoXX, ZhuXY, LiuB ( 2018). OsWRKY67 positively regulates blast and bacteria blight resistance by direct activation of PR genes in rice BMC Plant Biol 18, 257. [本文引用: 1]
LuanZH, ZhouDW ( 2015). Screening of rice ( Oryza sativa L.) OsPR1b-interacting factors and their roles in resisting bacterial blight Genet Mol Res 14, 1868-1874. [本文引用: 1]
McGeeJD, HamerJE, HodgesTK ( 2001). Characterization of a PR-10 pathogenesis-related gene family induced in rice during infection with Magnaporthe grisea Mol Plant Microbe Interact 14, 877-886. [本文引用: 1]
NürnbergerT, BrunnerF, KemmerlingB, PiaterL ( 2004). Innate immunity in plants and animals: striking similarities and obvious differences Immunol Rev 198, 249-266. [本文引用: 1]
PandeyA, MannM ( 2000). Proteomics to study genes and genomes Nature 405, 837-846. [本文引用: 1]
PoncianoG, YoshikawaM, LeeJL, RonaldPC, WhalenMC ( 2006). Pathogenesis-related gene expression in rice is correlated with developmentally controlled Xa21-mediated resistance against Xanthomonas oryzae pv Oryzae. Physiol Mol Plant Pathol 69, 131-139. [本文引用: 1]
SeoPJ, LeeAK, XiangFN, ParkCM ( 2008). Molecular and functional profiling of Arabidopsis Pathogenesis-related genes: insights into their roles in salt response of seed germination Plant Cell Physiol 49, 334-344. [本文引用: 1]
TakeuchiK, GyohdaA, TominagaM, KawakatsuM, HatakeyamaA, IshiiN, ShimayaK, NishimuraT, RiemannM, NickP, HashimotoM, KomanoT, EndoA, OkamotoT, JikumaruY, KamiyaY, TerakawaT, KoshibaT ( 2011). RSOsPR10 expression in response to environmental stresses is regulated antagonistically by jasmonate/ethylene and salicylic acid signaling pathways in rice roots Plant Cell Physiol 52, 1686-1696. [本文引用: 1]
TakeuchiK, HasegawaH, GyohdaA, KomatsuS, OkamotoT, OkadaK, TerakawaT, KoshibaT ( 2016). Overexpression of RSOsPR10, a root-specific rice PR10 gene, confers tolerance against drought stress in rice and drought and salt stresses in bentgrass Plant Cell Tissue Organ Cult 127, 35-46. [本文引用: 1]
ThakurP, KumarS, MalikJA, BergerJD, NayyarH ( 2010). Cold stress effects on reproductive development in grain crops: an overview Environ Exp Bot 67, 429-443. [本文引用: 1]
UhlénM, Bj?rlingE, AgatonC, SzigyartoCAK, AminiB, AndersenE, AnderssonAC, AngelidouP, AsplundA, AsplundC, BerglundL, Bergstr?mK, BrumerH, CerjanD, Ekstr?mM, ElobeidA, ErikssonC, FagerbergL, FalkR, FallJ, ForsbergM, Bj?rklundMG, GumbelK, HalimiA, HallinI, HamstenC, HanssonM, HedhammarM, HerculesG, KampfC, LarssonK, LindskogM, LodewyckxW, LundJ, LundebergJ, MagnussonK, MalmE, NilssonP, ?dlingJ, OksvoldP, OlssonI, ?sterE, OttossonJ, PaavilainenL, PerssonA, RiminiR, RockbergJ, RunesonM, Sivertsson?, Sk?llermoA, SteenJ, StenvallM, SterkyF, Str?mbergS, SundbergM, TegelH, TourleS, WahlundE, WaldénA, WanJH, WernérusH, WestbergJ, WesterK, WrethagenU, XuLL, HoberS, PonténF ( 2005). A human protein atlas for normal and cancer tissues based on antibody proteomics Mol Cell Proteomics 4, 1920-1932. [本文引用: 1]
WangNL, XiaoBZ, XiongLZ ( 2011). Identification of a cluster of PR4-like genes involved in stress responses in rice J Plant Physiol 168, 2212-2224. [本文引用: 1]
WuJN, KimSG, KangKY, KimJG, ParkSR, GuptaR, KimYH, WangYM, KimST ( 2016). Overexpression of a pathogenesis-related protein 10 enhances biotic and abiotic stress tolerance in rice Plant Pathol J 32, 552-562. [本文引用: 1]
Rice (Oryza sativa L.) OsPR1b gene is phytohormonally regulated in close interaction with light signals 1 2000
... 为应对多变的自然环境, 植物进化出复杂的抗性机制(Nürnberger et al., 2004; Ausubel, 2005).例如, 在受到病原菌侵害或其它生物胁迫时, 植物会激活自身的防御体系来应对侵害.病程相关(pathogenesis- related, PR)蛋白是一类受病原物诱导的蛋白质.水稻是单子叶植物遗传研究的模式生物, 其基因组包含900多个PR基因(窦世娟等, 2014).根据蛋白质的结构, 可将PR蛋白分为17个亚类, 不同亚类的PR蛋白生物学功能也各有不同.探究水稻PR基因的功能有助于深入了解水稻的抗逆机理.大量研究表明, 水稻PR基因在抵抗生物及非生物逆境胁迫中发挥重要作用, 调控特定PR基因的表达会影响水稻的抗逆反应, PR蛋白在抗病反应中也被诱导表达(李雪姣等, 2014).PR1家族的OsPR1A和OsPR1B的转录水平易受病原菌诱导, 可作为抗病反应发生的分子标记(Ponciano et al., 2006).在水稻与白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)互作反应中, OsPR1A和OsPR1B蛋白质在Xoo侵染水稻叶片144小时后大量表达(Wu et al., 2011).在水稻幼苗中, OsPR1A和OsPR1B基因的转录与光处理密切相关(Agrawal et al., 2000b), 且外源施加茉莉酸(jasmonic acid, JA)和乙烯(ethylene, ET)能显著增加OsPR1B基因的转录(Luan and Zhou, 2015).GNS1属于PR2家族, 具有β-1,3-葡聚糖酶活性, 在真菌诱导下, 水稻中GNS1基因的转录和蛋白质表达水平均明显升高; 在黑暗处理时, GNS1基因表达水平也有所上升(Nishizawa et al., 2003).转PR3基因的水稻对纹枯病的抗性增强(Datta et al., 2001).超表达OsPR4A的水稻植株在幼苗期和孕穗期抗旱性明显增强(Wang et al., 2011).水稻PR1A、PR5、PR8和PR16蛋白质随盐胁迫时间的延长下调表达, 而PR2在干旱、高盐及水淹胁迫中均被诱导(高庆华等, 2013).OsWRKY67与OsPR1A和OsPR10的启动子区结合进而提高对白叶枯病菌的抗性(Liu et al., 2018).在其它植物中也有大量PR基因的相关报道.拟南芥(Arabidopsis thaliana)受到南方根结线虫(Meloidogyne incognita)侵袭后, PR1、PR2和PR5的表达被大量诱导(Hamamouch et al., 2011).干旱胁迫下, 拟南芥中PR1、PR2和PR5基因表达升高(Seo et al., 2008).在烟草(Nicotiana tabacum)中表达OsGLP1基因可增强其对真菌病原物的抗性(Banerjee et al., 2010). ...
Are innate immune signaling pathways in plants and animals conserved? 1 2005
... 为应对多变的自然环境, 植物进化出复杂的抗性机制(Nürnberger et al., 2004; Ausubel, 2005).例如, 在受到病原菌侵害或其它生物胁迫时, 植物会激活自身的防御体系来应对侵害.病程相关(pathogenesis- related, PR)蛋白是一类受病原物诱导的蛋白质.水稻是单子叶植物遗传研究的模式生物, 其基因组包含900多个PR基因(窦世娟等, 2014).根据蛋白质的结构, 可将PR蛋白分为17个亚类, 不同亚类的PR蛋白生物学功能也各有不同.探究水稻PR基因的功能有助于深入了解水稻的抗逆机理.大量研究表明, 水稻PR基因在抵抗生物及非生物逆境胁迫中发挥重要作用, 调控特定PR基因的表达会影响水稻的抗逆反应, PR蛋白在抗病反应中也被诱导表达(李雪姣等, 2014).PR1家族的OsPR1A和OsPR1B的转录水平易受病原菌诱导, 可作为抗病反应发生的分子标记(Ponciano et al., 2006).在水稻与白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)互作反应中, OsPR1A和OsPR1B蛋白质在Xoo侵染水稻叶片144小时后大量表达(Wu et al., 2011).在水稻幼苗中, OsPR1A和OsPR1B基因的转录与光处理密切相关(Agrawal et al., 2000b), 且外源施加茉莉酸(jasmonic acid, JA)和乙烯(ethylene, ET)能显著增加OsPR1B基因的转录(Luan and Zhou, 2015).GNS1属于PR2家族, 具有β-1,3-葡聚糖酶活性, 在真菌诱导下, 水稻中GNS1基因的转录和蛋白质表达水平均明显升高; 在黑暗处理时, GNS1基因表达水平也有所上升(Nishizawa et al., 2003).转PR3基因的水稻对纹枯病的抗性增强(Datta et al., 2001).超表达OsPR4A的水稻植株在幼苗期和孕穗期抗旱性明显增强(Wang et al., 2011).水稻PR1A、PR5、PR8和PR16蛋白质随盐胁迫时间的延长下调表达, 而PR2在干旱、高盐及水淹胁迫中均被诱导(高庆华等, 2013).OsWRKY67与OsPR1A和OsPR10的启动子区结合进而提高对白叶枯病菌的抗性(Liu et al., 2018).在其它植物中也有大量PR基因的相关报道.拟南芥(Arabidopsis thaliana)受到南方根结线虫(Meloidogyne incognita)侵袭后, PR1、PR2和PR5的表达被大量诱导(Hamamouch et al., 2011).干旱胁迫下, 拟南芥中PR1、PR2和PR5基因表达升高(Seo et al., 2008).在烟草(Nicotiana tabacum)中表达OsGLP1基因可增强其对真菌病原物的抗性(Banerjee et al., 2010). ...
Transgenically expressed rice germin-like protein 1 in tobacco causes hyper-accumulation of H2O2 and reinforcement of the cell wall components 1 2010
... 为应对多变的自然环境, 植物进化出复杂的抗性机制(Nürnberger et al., 2004; Ausubel, 2005).例如, 在受到病原菌侵害或其它生物胁迫时, 植物会激活自身的防御体系来应对侵害.病程相关(pathogenesis- related, PR)蛋白是一类受病原物诱导的蛋白质.水稻是单子叶植物遗传研究的模式生物, 其基因组包含900多个PR基因(窦世娟等, 2014).根据蛋白质的结构, 可将PR蛋白分为17个亚类, 不同亚类的PR蛋白生物学功能也各有不同.探究水稻PR基因的功能有助于深入了解水稻的抗逆机理.大量研究表明, 水稻PR基因在抵抗生物及非生物逆境胁迫中发挥重要作用, 调控特定PR基因的表达会影响水稻的抗逆反应, PR蛋白在抗病反应中也被诱导表达(李雪姣等, 2014).PR1家族的OsPR1A和OsPR1B的转录水平易受病原菌诱导, 可作为抗病反应发生的分子标记(Ponciano et al., 2006).在水稻与白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)互作反应中, OsPR1A和OsPR1B蛋白质在Xoo侵染水稻叶片144小时后大量表达(Wu et al., 2011).在水稻幼苗中, OsPR1A和OsPR1B基因的转录与光处理密切相关(Agrawal et al., 2000b), 且外源施加茉莉酸(jasmonic acid, JA)和乙烯(ethylene, ET)能显著增加OsPR1B基因的转录(Luan and Zhou, 2015).GNS1属于PR2家族, 具有β-1,3-葡聚糖酶活性, 在真菌诱导下, 水稻中GNS1基因的转录和蛋白质表达水平均明显升高; 在黑暗处理时, GNS1基因表达水平也有所上升(Nishizawa et al., 2003).转PR3基因的水稻对纹枯病的抗性增强(Datta et al., 2001).超表达OsPR4A的水稻植株在幼苗期和孕穗期抗旱性明显增强(Wang et al., 2011).水稻PR1A、PR5、PR8和PR16蛋白质随盐胁迫时间的延长下调表达, 而PR2在干旱、高盐及水淹胁迫中均被诱导(高庆华等, 2013).OsWRKY67与OsPR1A和OsPR10的启动子区结合进而提高对白叶枯病菌的抗性(Liu et al., 2018).在其它植物中也有大量PR基因的相关报道.拟南芥(Arabidopsis thaliana)受到南方根结线虫(Meloidogyne incognita)侵袭后, PR1、PR2和PR5的表达被大量诱导(Hamamouch et al., 2011).干旱胁迫下, 拟南芥中PR1、PR2和PR5基因表达升高(Seo et al., 2008).在烟草(Nicotiana tabacum)中表达OsGLP1基因可增强其对真菌病原物的抗性(Banerjee et al., 2010). ...
A key ABA catabolic gene, OsABA8ox3 , is involved in drought stress resistance in rice 1 2015
... 本研究还发现, 在离体水稻叶片激素处理过程中, OsPR10A的表达丰度受MeJA和ABA诱导, 表明该蛋白功能的发挥与JA和ABA的信号转导途径有关.与野生型相比, OsJAZ1超表达植株对干旱敏感, 同时对MeJA和ABA表现更为敏感, RNA-seq分析结果表明, 在干旱处理下, JA和ABA信号转导途径中许多基因的表达水平在OsJAZ1超表达植株和野生型之间差异显著, 表明JA和ABA介导的途径参与水稻对干旱胁迫的抵御(Fu et al., 2017).水稻转录因子OsbZIP23与耐旱性有关, 同时也是ABA信号途径和ABA合成的关键调控子(Zong et al., 2016).水稻OsASR5与水稻耐旱性有关, 超表达OsASR5使种子萌发期和萌发后期内源ABA水平增高, 对外源施加的ABA也更为敏感(Li et al., 2017).水稻ABA8ox3是ABA生物合成的关键基因, 其转基因RNAi株系耐旱性更强, 而超表达株系对干旱胁迫更为敏感(Cai et al., 2015).本研究中, OsPR10A分别受干旱胁迫和激素(MeJA和ABA)的诱导, 推测该蛋白可能通过JA和ABA相关的信号转导途径增强水稻的耐旱性. ...
Molecular characterization of Oryza sativa WRKY6, which binds to W-box-like element 1 of the Oryza sativa pathogenesis-related (PR) 10a promoter and confers reduced susceptibility to pathogens 1 2015
... PR10蛋白结构保守且功能多样, 在植物抗逆方面的功能也有报道(谢纯政等, 2008).水稻OsPR10A蛋白分子量约为17 kDa, 经生化验证其具有RNA酶活性, 经GUS染色确定OsPR10A的转录受到多种方式的调控, 如生物胁迫Xoo的侵染、非生物胁迫伤害及缺磷.此外, 在叶片的正常衰老过程中OsPR10A也会被诱导表达(Huang et al., 2016).PR10A (PBZ1)在细胞衰亡的组织中积累, 可诱导水稻细胞死亡, 也能通过其RNA酶活性诱导拟南芥和烟草细胞死亡(Kim et al., 2011).蛋白质免疫印迹(western blot, WB)分析发现, OsPR10A在水稻受到Xoo侵染后144小时大量表达, 而且不亲和互作中表达量高于亲和互作反应(Wu et al., 2011).超表达OsPR10A增强了水稻对白叶枯病菌的抗性(Huang et al., 2016).OsPR10A的上游有转录因子结合位点, 可能受转录因子OsWRKY6的调控, OsWRKY6与类W-box元件1 (WLE1)物理结合, 正向调控OsPR10A的表达(Choi et al., 2015).序列同源性分析表明, OsPR10A和RSOsPR10高度相似, 具有相似的酶活性质与功能(Hashimoto et al., 2004).RSOsPR10是水稻根部特有的病程相关基因(Hashimoto et al., 2004), 在干旱和高盐胁迫下均可被诱导表达(Takeuchi et al., 2011); 在根皮细胞中超表达RSOsPR10的水稻植株对干旱胁迫具有更强的耐受性, 但对盐胁迫的耐受性较差(Takeuchi et al., 2016).超表达JIOsPR10增强了水稻对稻瘟病菌的抗性, 证明其在水稻防卫机制中发挥作用; 半定量RT-PCR分析表明, 高盐和干旱胁迫条件下, 水稻JIOsPR10的表达上调(Wu et al., 2016); WB检测发现, 其在正常组织(如花和根)中都表达, 衰老及伤害条件也能提高其表达水平(Kim et al., 2008). ...
OsiSAP1 overexpression improves water-deficit stress tolerance in transgenic rice by affecting expression of endogenous stress-related genes 1 2014
Enhanced resistance to sheath blight by constitutive expression of infection-related rice chitinase in transgenic elite indica rice cultivars 1 2001
... 为应对多变的自然环境, 植物进化出复杂的抗性机制(Nürnberger et al., 2004; Ausubel, 2005).例如, 在受到病原菌侵害或其它生物胁迫时, 植物会激活自身的防御体系来应对侵害.病程相关(pathogenesis- related, PR)蛋白是一类受病原物诱导的蛋白质.水稻是单子叶植物遗传研究的模式生物, 其基因组包含900多个PR基因(窦世娟等, 2014).根据蛋白质的结构, 可将PR蛋白分为17个亚类, 不同亚类的PR蛋白生物学功能也各有不同.探究水稻PR基因的功能有助于深入了解水稻的抗逆机理.大量研究表明, 水稻PR基因在抵抗生物及非生物逆境胁迫中发挥重要作用, 调控特定PR基因的表达会影响水稻的抗逆反应, PR蛋白在抗病反应中也被诱导表达(李雪姣等, 2014).PR1家族的OsPR1A和OsPR1B的转录水平易受病原菌诱导, 可作为抗病反应发生的分子标记(Ponciano et al., 2006).在水稻与白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)互作反应中, OsPR1A和OsPR1B蛋白质在Xoo侵染水稻叶片144小时后大量表达(Wu et al., 2011).在水稻幼苗中, OsPR1A和OsPR1B基因的转录与光处理密切相关(Agrawal et al., 2000b), 且外源施加茉莉酸(jasmonic acid, JA)和乙烯(ethylene, ET)能显著增加OsPR1B基因的转录(Luan and Zhou, 2015).GNS1属于PR2家族, 具有β-1,3-葡聚糖酶活性, 在真菌诱导下, 水稻中GNS1基因的转录和蛋白质表达水平均明显升高; 在黑暗处理时, GNS1基因表达水平也有所上升(Nishizawa et al., 2003).转PR3基因的水稻对纹枯病的抗性增强(Datta et al., 2001).超表达OsPR4A的水稻植株在幼苗期和孕穗期抗旱性明显增强(Wang et al., 2011).水稻PR1A、PR5、PR8和PR16蛋白质随盐胁迫时间的延长下调表达, 而PR2在干旱、高盐及水淹胁迫中均被诱导(高庆华等, 2013).OsWRKY67与OsPR1A和OsPR10的启动子区结合进而提高对白叶枯病菌的抗性(Liu et al., 2018).在其它植物中也有大量PR基因的相关报道.拟南芥(Arabidopsis thaliana)受到南方根结线虫(Meloidogyne incognita)侵袭后, PR1、PR2和PR5的表达被大量诱导(Hamamouch et al., 2011).干旱胁迫下, 拟南芥中PR1、PR2和PR5基因表达升高(Seo et al., 2008).在烟草(Nicotiana tabacum)中表达OsGLP1基因可增强其对真菌病原物的抗性(Banerjee et al., 2010). ...
An efficient and high-throughput protocol for Agrobacterium-mediated transformation based on phosphomannose isomerase positive selection in Japonica rice(Oryza sativa L.) 1 2012
OsJAZ1 attenuates drought resistance by regulating JA and ABA signaling in rice 1 2017
... 本研究还发现, 在离体水稻叶片激素处理过程中, OsPR10A的表达丰度受MeJA和ABA诱导, 表明该蛋白功能的发挥与JA和ABA的信号转导途径有关.与野生型相比, OsJAZ1超表达植株对干旱敏感, 同时对MeJA和ABA表现更为敏感, RNA-seq分析结果表明, 在干旱处理下, JA和ABA信号转导途径中许多基因的表达水平在OsJAZ1超表达植株和野生型之间差异显著, 表明JA和ABA介导的途径参与水稻对干旱胁迫的抵御(Fu et al., 2017).水稻转录因子OsbZIP23与耐旱性有关, 同时也是ABA信号途径和ABA合成的关键调控子(Zong et al., 2016).水稻OsASR5与水稻耐旱性有关, 超表达OsASR5使种子萌发期和萌发后期内源ABA水平增高, 对外源施加的ABA也更为敏感(Li et al., 2017).水稻ABA8ox3是ABA生物合成的关键基因, 其转基因RNAi株系耐旱性更强, 而超表达株系对干旱胁迫更为敏感(Cai et al., 2015).本研究中, OsPR10A分别受干旱胁迫和激素(MeJA和ABA)的诱导, 推测该蛋白可能通过JA和ABA相关的信号转导途径增强水稻的耐旱性. ...
Expression of Arabidopsis pathogenesis-related genes during nematode infection 1 2011
... 为应对多变的自然环境, 植物进化出复杂的抗性机制(Nürnberger et al., 2004; Ausubel, 2005).例如, 在受到病原菌侵害或其它生物胁迫时, 植物会激活自身的防御体系来应对侵害.病程相关(pathogenesis- related, PR)蛋白是一类受病原物诱导的蛋白质.水稻是单子叶植物遗传研究的模式生物, 其基因组包含900多个PR基因(窦世娟等, 2014).根据蛋白质的结构, 可将PR蛋白分为17个亚类, 不同亚类的PR蛋白生物学功能也各有不同.探究水稻PR基因的功能有助于深入了解水稻的抗逆机理.大量研究表明, 水稻PR基因在抵抗生物及非生物逆境胁迫中发挥重要作用, 调控特定PR基因的表达会影响水稻的抗逆反应, PR蛋白在抗病反应中也被诱导表达(李雪姣等, 2014).PR1家族的OsPR1A和OsPR1B的转录水平易受病原菌诱导, 可作为抗病反应发生的分子标记(Ponciano et al., 2006).在水稻与白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)互作反应中, OsPR1A和OsPR1B蛋白质在Xoo侵染水稻叶片144小时后大量表达(Wu et al., 2011).在水稻幼苗中, OsPR1A和OsPR1B基因的转录与光处理密切相关(Agrawal et al., 2000b), 且外源施加茉莉酸(jasmonic acid, JA)和乙烯(ethylene, ET)能显著增加OsPR1B基因的转录(Luan and Zhou, 2015).GNS1属于PR2家族, 具有β-1,3-葡聚糖酶活性, 在真菌诱导下, 水稻中GNS1基因的转录和蛋白质表达水平均明显升高; 在黑暗处理时, GNS1基因表达水平也有所上升(Nishizawa et al., 2003).转PR3基因的水稻对纹枯病的抗性增强(Datta et al., 2001).超表达OsPR4A的水稻植株在幼苗期和孕穗期抗旱性明显增强(Wang et al., 2011).水稻PR1A、PR5、PR8和PR16蛋白质随盐胁迫时间的延长下调表达, 而PR2在干旱、高盐及水淹胁迫中均被诱导(高庆华等, 2013).OsWRKY67与OsPR1A和OsPR10的启动子区结合进而提高对白叶枯病菌的抗性(Liu et al., 2018).在其它植物中也有大量PR基因的相关报道.拟南芥(Arabidopsis thaliana)受到南方根结线虫(Meloidogyne incognita)侵袭后, PR1、PR2和PR5的表达被大量诱导(Hamamouch et al., 2011).干旱胁迫下, 拟南芥中PR1、PR2和PR5基因表达升高(Seo et al., 2008).在烟草(Nicotiana tabacum)中表达OsGLP1基因可增强其对真菌病原物的抗性(Banerjee et al., 2010). ...
A novel rice PR10 protein, RSOsPR10, specifically induced in roots by biotic and abiotic stresses, possibly via the jasmonic acid signaling pathway 3 2004
... PR10蛋白结构保守且功能多样, 在植物抗逆方面的功能也有报道(谢纯政等, 2008).水稻OsPR10A蛋白分子量约为17 kDa, 经生化验证其具有RNA酶活性, 经GUS染色确定OsPR10A的转录受到多种方式的调控, 如生物胁迫Xoo的侵染、非生物胁迫伤害及缺磷.此外, 在叶片的正常衰老过程中OsPR10A也会被诱导表达(Huang et al., 2016).PR10A (PBZ1)在细胞衰亡的组织中积累, 可诱导水稻细胞死亡, 也能通过其RNA酶活性诱导拟南芥和烟草细胞死亡(Kim et al., 2011).蛋白质免疫印迹(western blot, WB)分析发现, OsPR10A在水稻受到Xoo侵染后144小时大量表达, 而且不亲和互作中表达量高于亲和互作反应(Wu et al., 2011).超表达OsPR10A增强了水稻对白叶枯病菌的抗性(Huang et al., 2016).OsPR10A的上游有转录因子结合位点, 可能受转录因子OsWRKY6的调控, OsWRKY6与类W-box元件1 (WLE1)物理结合, 正向调控OsPR10A的表达(Choi et al., 2015).序列同源性分析表明, OsPR10A和RSOsPR10高度相似, 具有相似的酶活性质与功能(Hashimoto et al., 2004).RSOsPR10是水稻根部特有的病程相关基因(Hashimoto et al., 2004), 在干旱和高盐胁迫下均可被诱导表达(Takeuchi et al., 2011); 在根皮细胞中超表达RSOsPR10的水稻植株对干旱胁迫具有更强的耐受性, 但对盐胁迫的耐受性较差(Takeuchi et al., 2016).超表达JIOsPR10增强了水稻对稻瘟病菌的抗性, 证明其在水稻防卫机制中发挥作用; 半定量RT-PCR分析表明, 高盐和干旱胁迫条件下, 水稻JIOsPR10的表达上调(Wu et al., 2016); WB检测发现, 其在正常组织(如花和根)中都表达, 衰老及伤害条件也能提高其表达水平(Kim et al., 2008). ...
... 是水稻根部特有的病程相关基因(Hashimoto et al., 2004), 在干旱和高盐胁迫下均可被诱导表达(Takeuchi et al., 2011); 在根皮细胞中超表达RSOsPR10的水稻植株对干旱胁迫具有更强的耐受性, 但对盐胁迫的耐受性较差(Takeuchi et al., 2016).超表达JIOsPR10增强了水稻对稻瘟病菌的抗性, 证明其在水稻防卫机制中发挥作用; 半定量RT-PCR分析表明, 高盐和干旱胁迫条件下, 水稻JIOsPR10的表达上调(Wu et al., 2016); WB检测发现, 其在正常组织(如花和根)中都表达, 衰老及伤害条件也能提高其表达水平(Kim et al., 2008). ...
... 研究表明, OsPR10A可受SA、JA、ET和ABA等激素的诱导(Hwang et al., 2008), 且OsPR10A启动子687-637之间的WLE1在SA介导的OsPR10A表达中起重要作用(McGee et al., 2001).RSOsPR10的表达受生物和非生物逆境诱导, 并可能受JA诱导(Hashimoto et al., 2004).本研究检测了激素处理条件下OsPR10A蛋白的表达丰度变化, 后续将深入研究JA和ABA等激素如何发挥功能. ...
Multiple patterns of regulation and overexpression of a ribonuclease-like pathogenesis-related protein gene,OsPR10a, conferring disease resistance in rice and Arabidopsis 3 2016
... PR10蛋白结构保守且功能多样, 在植物抗逆方面的功能也有报道(谢纯政等, 2008).水稻OsPR10A蛋白分子量约为17 kDa, 经生化验证其具有RNA酶活性, 经GUS染色确定OsPR10A的转录受到多种方式的调控, 如生物胁迫Xoo的侵染、非生物胁迫伤害及缺磷.此外, 在叶片的正常衰老过程中OsPR10A也会被诱导表达(Huang et al., 2016).PR10A (PBZ1)在细胞衰亡的组织中积累, 可诱导水稻细胞死亡, 也能通过其RNA酶活性诱导拟南芥和烟草细胞死亡(Kim et al., 2011).蛋白质免疫印迹(western blot, WB)分析发现, OsPR10A在水稻受到Xoo侵染后144小时大量表达, 而且不亲和互作中表达量高于亲和互作反应(Wu et al., 2011).超表达OsPR10A增强了水稻对白叶枯病菌的抗性(Huang et al., 2016).OsPR10A的上游有转录因子结合位点, 可能受转录因子OsWRKY6的调控, OsWRKY6与类W-box元件1 (WLE1)物理结合, 正向调控OsPR10A的表达(Choi et al., 2015).序列同源性分析表明, OsPR10A和RSOsPR10高度相似, 具有相似的酶活性质与功能(Hashimoto et al., 2004).RSOsPR10是水稻根部特有的病程相关基因(Hashimoto et al., 2004), 在干旱和高盐胁迫下均可被诱导表达(Takeuchi et al., 2011); 在根皮细胞中超表达RSOsPR10的水稻植株对干旱胁迫具有更强的耐受性, 但对盐胁迫的耐受性较差(Takeuchi et al., 2016).超表达JIOsPR10增强了水稻对稻瘟病菌的抗性, 证明其在水稻防卫机制中发挥作用; 半定量RT-PCR分析表明, 高盐和干旱胁迫条件下, 水稻JIOsPR10的表达上调(Wu et al., 2016); WB检测发现, 其在正常组织(如花和根)中都表达, 衰老及伤害条件也能提高其表达水平(Kim et al., 2008). ...
... ).超表达OsPR10A增强了水稻对白叶枯病菌的抗性(Huang et al., 2016).OsPR10A的上游有转录因子结合位点, 可能受转录因子OsWRKY6的调控, OsWRKY6与类W-box元件1 (WLE1)物理结合, 正向调控OsPR10A的表达(Choi et al., 2015).序列同源性分析表明, OsPR10A和RSOsPR10高度相似, 具有相似的酶活性质与功能(Hashimoto et al., 2004).RSOsPR10是水稻根部特有的病程相关基因(Hashimoto et al., 2004), 在干旱和高盐胁迫下均可被诱导表达(Takeuchi et al., 2011); 在根皮细胞中超表达RSOsPR10的水稻植株对干旱胁迫具有更强的耐受性, 但对盐胁迫的耐受性较差(Takeuchi et al., 2016).超表达JIOsPR10增强了水稻对稻瘟病菌的抗性, 证明其在水稻防卫机制中发挥作用; 半定量RT-PCR分析表明, 高盐和干旱胁迫条件下, 水稻JIOsPR10的表达上调(Wu et al., 2016); WB检测发现, 其在正常组织(如花和根)中都表达, 衰老及伤害条件也能提高其表达水平(Kim et al., 2008). ...
... 最初报道的PR基因功能大都与抗病相关, 近年来发现PR基因在抵御非生物胁迫中也行使功能.OsPR10A启动子驱动的GUS受磷饥饿、伤害和Xoo侵染的诱导, 超表达OsPR10A的水稻和拟南芥在磷饥饿条件下根长明显长于野生型(Huang et al., 2016).本研究则表明OsPR10A超表达能增强水稻种子萌发期的耐旱性, 丰富了人们对OsPR10A功能的认识. ...
Identification of an OsPR10a promoter region responsive to salicylic acid 1 2008
... 研究表明, OsPR10A可受SA、JA、ET和ABA等激素的诱导(Hwang et al., 2008), 且OsPR10A启动子687-637之间的WLE1在SA介导的OsPR10A表达中起重要作用(McGee et al., 2001).RSOsPR10的表达受生物和非生物逆境诱导, 并可能受JA诱导(Hashimoto et al., 2004).本研究检测了激素处理条件下OsPR10A蛋白的表达丰度变化, 后续将深入研究JA和ABA等激素如何发挥功能. ...
The RNase activity of rice probenazole-induced protein1 (PBZ1) plays a key role in cell death in plants 1 2011
... PR10蛋白结构保守且功能多样, 在植物抗逆方面的功能也有报道(谢纯政等, 2008).水稻OsPR10A蛋白分子量约为17 kDa, 经生化验证其具有RNA酶活性, 经GUS染色确定OsPR10A的转录受到多种方式的调控, 如生物胁迫Xoo的侵染、非生物胁迫伤害及缺磷.此外, 在叶片的正常衰老过程中OsPR10A也会被诱导表达(Huang et al., 2016).PR10A (PBZ1)在细胞衰亡的组织中积累, 可诱导水稻细胞死亡, 也能通过其RNA酶活性诱导拟南芥和烟草细胞死亡(Kim et al., 2011).蛋白质免疫印迹(western blot, WB)分析发现, OsPR10A在水稻受到Xoo侵染后144小时大量表达, 而且不亲和互作中表达量高于亲和互作反应(Wu et al., 2011).超表达OsPR10A增强了水稻对白叶枯病菌的抗性(Huang et al., 2016).OsPR10A的上游有转录因子结合位点, 可能受转录因子OsWRKY6的调控, OsWRKY6与类W-box元件1 (WLE1)物理结合, 正向调控OsPR10A的表达(Choi et al., 2015).序列同源性分析表明, OsPR10A和RSOsPR10高度相似, 具有相似的酶活性质与功能(Hashimoto et al., 2004).RSOsPR10是水稻根部特有的病程相关基因(Hashimoto et al., 2004), 在干旱和高盐胁迫下均可被诱导表达(Takeuchi et al., 2011); 在根皮细胞中超表达RSOsPR10的水稻植株对干旱胁迫具有更强的耐受性, 但对盐胁迫的耐受性较差(Takeuchi et al., 2016).超表达JIOsPR10增强了水稻对稻瘟病菌的抗性, 证明其在水稻防卫机制中发挥作用; 半定量RT-PCR分析表明, 高盐和干旱胁迫条件下, 水稻JIOsPR10的表达上调(Wu et al., 2016); WB检测发现, 其在正常组织(如花和根)中都表达, 衰老及伤害条件也能提高其表达水平(Kim et al., 2008). ...
The rice pathogen-related protein 10 (JIOsPR10) is induced by abiotic and biotic stresses and exhibits ribonuclease activity 1 2008
... PR10蛋白结构保守且功能多样, 在植物抗逆方面的功能也有报道(谢纯政等, 2008).水稻OsPR10A蛋白分子量约为17 kDa, 经生化验证其具有RNA酶活性, 经GUS染色确定OsPR10A的转录受到多种方式的调控, 如生物胁迫Xoo的侵染、非生物胁迫伤害及缺磷.此外, 在叶片的正常衰老过程中OsPR10A也会被诱导表达(Huang et al., 2016).PR10A (PBZ1)在细胞衰亡的组织中积累, 可诱导水稻细胞死亡, 也能通过其RNA酶活性诱导拟南芥和烟草细胞死亡(Kim et al., 2011).蛋白质免疫印迹(western blot, WB)分析发现, OsPR10A在水稻受到Xoo侵染后144小时大量表达, 而且不亲和互作中表达量高于亲和互作反应(Wu et al., 2011).超表达OsPR10A增强了水稻对白叶枯病菌的抗性(Huang et al., 2016).OsPR10A的上游有转录因子结合位点, 可能受转录因子OsWRKY6的调控, OsWRKY6与类W-box元件1 (WLE1)物理结合, 正向调控OsPR10A的表达(Choi et al., 2015).序列同源性分析表明, OsPR10A和RSOsPR10高度相似, 具有相似的酶活性质与功能(Hashimoto et al., 2004).RSOsPR10是水稻根部特有的病程相关基因(Hashimoto et al., 2004), 在干旱和高盐胁迫下均可被诱导表达(Takeuchi et al., 2011); 在根皮细胞中超表达RSOsPR10的水稻植株对干旱胁迫具有更强的耐受性, 但对盐胁迫的耐受性较差(Takeuchi et al., 2016).超表达JIOsPR10增强了水稻对稻瘟病菌的抗性, 证明其在水稻防卫机制中发挥作用; 半定量RT-PCR分析表明, 高盐和干旱胁迫条件下, 水稻JIOsPR10的表达上调(Wu et al., 2016); WB检测发现, 其在正常组织(如花和根)中都表达, 衰老及伤害条件也能提高其表达水平(Kim et al., 2008). ...
Central dogma at the single-molecule level in living cells 3 2011
... 用Tricine-SDS-PAGE分离提取的水稻总蛋白质, 上样体积为10 µL, 然后转移到PVDF膜上, 蛋白质免疫印迹一抗为Anti-OsPR10A polyclonal抗体(Wu et al., 2011), 二抗为HRP标记的羊抗兔, 具体操作参见白辉等(2010)所述方法.以HSP抗体检测的信号作为内参(Li et al., 2011), 所用抗体购自北京华大蛋白质研发中心有限公司.用化学发光成像仪(MiniChemi610, 北京赛智创业科技有限公司)检测信号.WB实验至少重复3次. ...
OsASR5 enhances drought tolerance through a stomatal closure pathway associated with ABA and H2O2 signaling in rice 1 2017
... 本研究还发现, 在离体水稻叶片激素处理过程中, OsPR10A的表达丰度受MeJA和ABA诱导, 表明该蛋白功能的发挥与JA和ABA的信号转导途径有关.与野生型相比, OsJAZ1超表达植株对干旱敏感, 同时对MeJA和ABA表现更为敏感, RNA-seq分析结果表明, 在干旱处理下, JA和ABA信号转导途径中许多基因的表达水平在OsJAZ1超表达植株和野生型之间差异显著, 表明JA和ABA介导的途径参与水稻对干旱胁迫的抵御(Fu et al., 2017).水稻转录因子OsbZIP23与耐旱性有关, 同时也是ABA信号途径和ABA合成的关键调控子(Zong et al., 2016).水稻OsASR5与水稻耐旱性有关, 超表达OsASR5使种子萌发期和萌发后期内源ABA水平增高, 对外源施加的ABA也更为敏感(Li et al., 2017).水稻ABA8ox3是ABA生物合成的关键基因, 其转基因RNAi株系耐旱性更强, 而超表达株系对干旱胁迫更为敏感(Cai et al., 2015).本研究中, OsPR10A分别受干旱胁迫和激素(MeJA和ABA)的诱导, 推测该蛋白可能通过JA和ABA相关的信号转导途径增强水稻的耐旱性. ...
Identification and validation of rice reference proteins for western blotting 0 2011
Biochemical characterization of the kinase domain of the rice disease resistance receptor-like kinase XA21 1 2002
... 所用水稻(Oryza sativa L.)品种为TP309, 基因克隆用大肠杆菌(Escherichia coli)菌株DH5α, 重组蛋白质表达菌株为BL21 (DE3) pLysS.蛋白质原核表达载体为pET30a和pGST (Liu et al., 2002).水稻OsPR10A基因全长cDNA质粒购自日本农业生物资源研究所水稻基因组资源中心(Rice Genome Resource Center, National Institute of Agrobiological Sciences).质粒pUC57-3HA由南京金斯瑞生物科技有限公司合成, pEASY-T1购自上海生工生物技术有限公司, pUBI-C4300由Pamela Ronald博士(UC Davis, USA)提供.茉莉酸甲酯和脱落酸等试剂购自北京博奥拓达科技有限公司. ...
OsWRKY67 positively regulates blast and bacteria blight resistance by direct activation of PR genes in rice 1 2018
... 为应对多变的自然环境, 植物进化出复杂的抗性机制(Nürnberger et al., 2004; Ausubel, 2005).例如, 在受到病原菌侵害或其它生物胁迫时, 植物会激活自身的防御体系来应对侵害.病程相关(pathogenesis- related, PR)蛋白是一类受病原物诱导的蛋白质.水稻是单子叶植物遗传研究的模式生物, 其基因组包含900多个PR基因(窦世娟等, 2014).根据蛋白质的结构, 可将PR蛋白分为17个亚类, 不同亚类的PR蛋白生物学功能也各有不同.探究水稻PR基因的功能有助于深入了解水稻的抗逆机理.大量研究表明, 水稻PR基因在抵抗生物及非生物逆境胁迫中发挥重要作用, 调控特定PR基因的表达会影响水稻的抗逆反应, PR蛋白在抗病反应中也被诱导表达(李雪姣等, 2014).PR1家族的OsPR1A和OsPR1B的转录水平易受病原菌诱导, 可作为抗病反应发生的分子标记(Ponciano et al., 2006).在水稻与白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)互作反应中, OsPR1A和OsPR1B蛋白质在Xoo侵染水稻叶片144小时后大量表达(Wu et al., 2011).在水稻幼苗中, OsPR1A和OsPR1B基因的转录与光处理密切相关(Agrawal et al., 2000b), 且外源施加茉莉酸(jasmonic acid, JA)和乙烯(ethylene, ET)能显著增加OsPR1B基因的转录(Luan and Zhou, 2015).GNS1属于PR2家族, 具有β-1,3-葡聚糖酶活性, 在真菌诱导下, 水稻中GNS1基因的转录和蛋白质表达水平均明显升高; 在黑暗处理时, GNS1基因表达水平也有所上升(Nishizawa et al., 2003).转PR3基因的水稻对纹枯病的抗性增强(Datta et al., 2001).超表达OsPR4A的水稻植株在幼苗期和孕穗期抗旱性明显增强(Wang et al., 2011).水稻PR1A、PR5、PR8和PR16蛋白质随盐胁迫时间的延长下调表达, 而PR2在干旱、高盐及水淹胁迫中均被诱导(高庆华等, 2013).OsWRKY67与OsPR1A和OsPR10的启动子区结合进而提高对白叶枯病菌的抗性(Liu et al., 2018).在其它植物中也有大量PR基因的相关报道.拟南芥(Arabidopsis thaliana)受到南方根结线虫(Meloidogyne incognita)侵袭后, PR1、PR2和PR5的表达被大量诱导(Hamamouch et al., 2011).干旱胁迫下, 拟南芥中PR1、PR2和PR5基因表达升高(Seo et al., 2008).在烟草(Nicotiana tabacum)中表达OsGLP1基因可增强其对真菌病原物的抗性(Banerjee et al., 2010). ...
Screening of rice ( Oryza sativa L.) OsPR1b-interacting factors and their roles in resisting bacterial blight 1 2015
... 为应对多变的自然环境, 植物进化出复杂的抗性机制(Nürnberger et al., 2004; Ausubel, 2005).例如, 在受到病原菌侵害或其它生物胁迫时, 植物会激活自身的防御体系来应对侵害.病程相关(pathogenesis- related, PR)蛋白是一类受病原物诱导的蛋白质.水稻是单子叶植物遗传研究的模式生物, 其基因组包含900多个PR基因(窦世娟等, 2014).根据蛋白质的结构, 可将PR蛋白分为17个亚类, 不同亚类的PR蛋白生物学功能也各有不同.探究水稻PR基因的功能有助于深入了解水稻的抗逆机理.大量研究表明, 水稻PR基因在抵抗生物及非生物逆境胁迫中发挥重要作用, 调控特定PR基因的表达会影响水稻的抗逆反应, PR蛋白在抗病反应中也被诱导表达(李雪姣等, 2014).PR1家族的OsPR1A和OsPR1B的转录水平易受病原菌诱导, 可作为抗病反应发生的分子标记(Ponciano et al., 2006).在水稻与白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)互作反应中, OsPR1A和OsPR1B蛋白质在Xoo侵染水稻叶片144小时后大量表达(Wu et al., 2011).在水稻幼苗中, OsPR1A和OsPR1B基因的转录与光处理密切相关(Agrawal et al., 2000b), 且外源施加茉莉酸(jasmonic acid, JA)和乙烯(ethylene, ET)能显著增加OsPR1B基因的转录(Luan and Zhou, 2015).GNS1属于PR2家族, 具有β-1,3-葡聚糖酶活性, 在真菌诱导下, 水稻中GNS1基因的转录和蛋白质表达水平均明显升高; 在黑暗处理时, GNS1基因表达水平也有所上升(Nishizawa et al., 2003).转PR3基因的水稻对纹枯病的抗性增强(Datta et al., 2001).超表达OsPR4A的水稻植株在幼苗期和孕穗期抗旱性明显增强(Wang et al., 2011).水稻PR1A、PR5、PR8和PR16蛋白质随盐胁迫时间的延长下调表达, 而PR2在干旱、高盐及水淹胁迫中均被诱导(高庆华等, 2013).OsWRKY67与OsPR1A和OsPR10的启动子区结合进而提高对白叶枯病菌的抗性(Liu et al., 2018).在其它植物中也有大量PR基因的相关报道.拟南芥(Arabidopsis thaliana)受到南方根结线虫(Meloidogyne incognita)侵袭后, PR1、PR2和PR5的表达被大量诱导(Hamamouch et al., 2011).干旱胁迫下, 拟南芥中PR1、PR2和PR5基因表达升高(Seo et al., 2008).在烟草(Nicotiana tabacum)中表达OsGLP1基因可增强其对真菌病原物的抗性(Banerjee et al., 2010). ...
Characterization of a PR-10 pathogenesis-related gene family induced in rice during infection with Magnaporthe grisea 1 2001
... 研究表明, OsPR10A可受SA、JA、ET和ABA等激素的诱导(Hwang et al., 2008), 且OsPR10A启动子687-637之间的WLE1在SA介导的OsPR10A表达中起重要作用(McGee et al., 2001).RSOsPR10的表达受生物和非生物逆境诱导, 并可能受JA诱导(Hashimoto et al., 2004).本研究检测了激素处理条件下OsPR10A蛋白的表达丰度变化, 后续将深入研究JA和ABA等激素如何发挥功能. ...
Characterization of transgenic rice plants over-expressing the stress-inducible β-glucanase gene Gns1 1 2003
... 为应对多变的自然环境, 植物进化出复杂的抗性机制(Nürnberger et al., 2004; Ausubel, 2005).例如, 在受到病原菌侵害或其它生物胁迫时, 植物会激活自身的防御体系来应对侵害.病程相关(pathogenesis- related, PR)蛋白是一类受病原物诱导的蛋白质.水稻是单子叶植物遗传研究的模式生物, 其基因组包含900多个PR基因(窦世娟等, 2014).根据蛋白质的结构, 可将PR蛋白分为17个亚类, 不同亚类的PR蛋白生物学功能也各有不同.探究水稻PR基因的功能有助于深入了解水稻的抗逆机理.大量研究表明, 水稻PR基因在抵抗生物及非生物逆境胁迫中发挥重要作用, 调控特定PR基因的表达会影响水稻的抗逆反应, PR蛋白在抗病反应中也被诱导表达(李雪姣等, 2014).PR1家族的OsPR1A和OsPR1B的转录水平易受病原菌诱导, 可作为抗病反应发生的分子标记(Ponciano et al., 2006).在水稻与白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)互作反应中, OsPR1A和OsPR1B蛋白质在Xoo侵染水稻叶片144小时后大量表达(Wu et al., 2011).在水稻幼苗中, OsPR1A和OsPR1B基因的转录与光处理密切相关(Agrawal et al., 2000b), 且外源施加茉莉酸(jasmonic acid, JA)和乙烯(ethylene, ET)能显著增加OsPR1B基因的转录(Luan and Zhou, 2015).GNS1属于PR2家族, 具有β-1,3-葡聚糖酶活性, 在真菌诱导下, 水稻中GNS1基因的转录和蛋白质表达水平均明显升高; 在黑暗处理时, GNS1基因表达水平也有所上升(Nishizawa et al., 2003).转PR3基因的水稻对纹枯病的抗性增强(Datta et al., 2001).超表达OsPR4A的水稻植株在幼苗期和孕穗期抗旱性明显增强(Wang et al., 2011).水稻PR1A、PR5、PR8和PR16蛋白质随盐胁迫时间的延长下调表达, 而PR2在干旱、高盐及水淹胁迫中均被诱导(高庆华等, 2013).OsWRKY67与OsPR1A和OsPR10的启动子区结合进而提高对白叶枯病菌的抗性(Liu et al., 2018).在其它植物中也有大量PR基因的相关报道.拟南芥(Arabidopsis thaliana)受到南方根结线虫(Meloidogyne incognita)侵袭后, PR1、PR2和PR5的表达被大量诱导(Hamamouch et al., 2011).干旱胁迫下, 拟南芥中PR1、PR2和PR5基因表达升高(Seo et al., 2008).在烟草(Nicotiana tabacum)中表达OsGLP1基因可增强其对真菌病原物的抗性(Banerjee et al., 2010). ...
Innate immunity in plants and animals: striking similarities and obvious differences 1 2004
... 为应对多变的自然环境, 植物进化出复杂的抗性机制(Nürnberger et al., 2004; Ausubel, 2005).例如, 在受到病原菌侵害或其它生物胁迫时, 植物会激活自身的防御体系来应对侵害.病程相关(pathogenesis- related, PR)蛋白是一类受病原物诱导的蛋白质.水稻是单子叶植物遗传研究的模式生物, 其基因组包含900多个PR基因(窦世娟等, 2014).根据蛋白质的结构, 可将PR蛋白分为17个亚类, 不同亚类的PR蛋白生物学功能也各有不同.探究水稻PR基因的功能有助于深入了解水稻的抗逆机理.大量研究表明, 水稻PR基因在抵抗生物及非生物逆境胁迫中发挥重要作用, 调控特定PR基因的表达会影响水稻的抗逆反应, PR蛋白在抗病反应中也被诱导表达(李雪姣等, 2014).PR1家族的OsPR1A和OsPR1B的转录水平易受病原菌诱导, 可作为抗病反应发生的分子标记(Ponciano et al., 2006).在水稻与白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)互作反应中, OsPR1A和OsPR1B蛋白质在Xoo侵染水稻叶片144小时后大量表达(Wu et al., 2011).在水稻幼苗中, OsPR1A和OsPR1B基因的转录与光处理密切相关(Agrawal et al., 2000b), 且外源施加茉莉酸(jasmonic acid, JA)和乙烯(ethylene, ET)能显著增加OsPR1B基因的转录(Luan and Zhou, 2015).GNS1属于PR2家族, 具有β-1,3-葡聚糖酶活性, 在真菌诱导下, 水稻中GNS1基因的转录和蛋白质表达水平均明显升高; 在黑暗处理时, GNS1基因表达水平也有所上升(Nishizawa et al., 2003).转PR3基因的水稻对纹枯病的抗性增强(Datta et al., 2001).超表达OsPR4A的水稻植株在幼苗期和孕穗期抗旱性明显增强(Wang et al., 2011).水稻PR1A、PR5、PR8和PR16蛋白质随盐胁迫时间的延长下调表达, 而PR2在干旱、高盐及水淹胁迫中均被诱导(高庆华等, 2013).OsWRKY67与OsPR1A和OsPR10的启动子区结合进而提高对白叶枯病菌的抗性(Liu et al., 2018).在其它植物中也有大量PR基因的相关报道.拟南芥(Arabidopsis thaliana)受到南方根结线虫(Meloidogyne incognita)侵袭后, PR1、PR2和PR5的表达被大量诱导(Hamamouch et al., 2011).干旱胁迫下, 拟南芥中PR1、PR2和PR5基因表达升高(Seo et al., 2008).在烟草(Nicotiana tabacum)中表达OsGLP1基因可增强其对真菌病原物的抗性(Banerjee et al., 2010). ...
Proteomics to study genes and genomes 1 2000
... 蛋白质是生物功能的主要执行者, 了解蛋白质的表达丰度、修饰及其变化是蛋白质组学的主要目标(Pandey and Mann, 2000).利用蛋白质特异性抗体可对特定的目标蛋白进行靶向研究, 这一理念被称为基于抗体的蛋白质组学(antibody-based proteomics, AbP) (Uhlén et al., 2005).通过免疫学方法对蛋白质表达特性展开调查, 具有靶向性强、灵敏度高、操作简便和容易对大量样品进行分析等优点.在水稻及其它植物研究中也可借鉴这一思路, 水稻抗体资源库和蛋白质样品资源库的构建工作也已经开展(刘国振等, 2011; 张剑硕等, 2018). ...
Pathogenesis-related gene expression in rice is correlated with developmentally controlled Xa21-mediated resistance against Xanthomonas oryzae pv 1 2006
... 为应对多变的自然环境, 植物进化出复杂的抗性机制(Nürnberger et al., 2004; Ausubel, 2005).例如, 在受到病原菌侵害或其它生物胁迫时, 植物会激活自身的防御体系来应对侵害.病程相关(pathogenesis- related, PR)蛋白是一类受病原物诱导的蛋白质.水稻是单子叶植物遗传研究的模式生物, 其基因组包含900多个PR基因(窦世娟等, 2014).根据蛋白质的结构, 可将PR蛋白分为17个亚类, 不同亚类的PR蛋白生物学功能也各有不同.探究水稻PR基因的功能有助于深入了解水稻的抗逆机理.大量研究表明, 水稻PR基因在抵抗生物及非生物逆境胁迫中发挥重要作用, 调控特定PR基因的表达会影响水稻的抗逆反应, PR蛋白在抗病反应中也被诱导表达(李雪姣等, 2014).PR1家族的OsPR1A和OsPR1B的转录水平易受病原菌诱导, 可作为抗病反应发生的分子标记(Ponciano et al., 2006).在水稻与白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)互作反应中, OsPR1A和OsPR1B蛋白质在Xoo侵染水稻叶片144小时后大量表达(Wu et al., 2011).在水稻幼苗中, OsPR1A和OsPR1B基因的转录与光处理密切相关(Agrawal et al., 2000b), 且外源施加茉莉酸(jasmonic acid, JA)和乙烯(ethylene, ET)能显著增加OsPR1B基因的转录(Luan and Zhou, 2015).GNS1属于PR2家族, 具有β-1,3-葡聚糖酶活性, 在真菌诱导下, 水稻中GNS1基因的转录和蛋白质表达水平均明显升高; 在黑暗处理时, GNS1基因表达水平也有所上升(Nishizawa et al., 2003).转PR3基因的水稻对纹枯病的抗性增强(Datta et al., 2001).超表达OsPR4A的水稻植株在幼苗期和孕穗期抗旱性明显增强(Wang et al., 2011).水稻PR1A、PR5、PR8和PR16蛋白质随盐胁迫时间的延长下调表达, 而PR2在干旱、高盐及水淹胁迫中均被诱导(高庆华等, 2013).OsWRKY67与OsPR1A和OsPR10的启动子区结合进而提高对白叶枯病菌的抗性(Liu et al., 2018).在其它植物中也有大量PR基因的相关报道.拟南芥(Arabidopsis thaliana)受到南方根结线虫(Meloidogyne incognita)侵袭后, PR1、PR2和PR5的表达被大量诱导(Hamamouch et al., 2011).干旱胁迫下, 拟南芥中PR1、PR2和PR5基因表达升高(Seo et al., 2008).在烟草(Nicotiana tabacum)中表达OsGLP1基因可增强其对真菌病原物的抗性(Banerjee et al., 2010). ...
Molecular and functional profiling of Arabidopsis Pathogenesis-related genes: insights into their roles in salt response of seed germination 1 2008
... 为应对多变的自然环境, 植物进化出复杂的抗性机制(Nürnberger et al., 2004; Ausubel, 2005).例如, 在受到病原菌侵害或其它生物胁迫时, 植物会激活自身的防御体系来应对侵害.病程相关(pathogenesis- related, PR)蛋白是一类受病原物诱导的蛋白质.水稻是单子叶植物遗传研究的模式生物, 其基因组包含900多个PR基因(窦世娟等, 2014).根据蛋白质的结构, 可将PR蛋白分为17个亚类, 不同亚类的PR蛋白生物学功能也各有不同.探究水稻PR基因的功能有助于深入了解水稻的抗逆机理.大量研究表明, 水稻PR基因在抵抗生物及非生物逆境胁迫中发挥重要作用, 调控特定PR基因的表达会影响水稻的抗逆反应, PR蛋白在抗病反应中也被诱导表达(李雪姣等, 2014).PR1家族的OsPR1A和OsPR1B的转录水平易受病原菌诱导, 可作为抗病反应发生的分子标记(Ponciano et al., 2006).在水稻与白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)互作反应中, OsPR1A和OsPR1B蛋白质在Xoo侵染水稻叶片144小时后大量表达(Wu et al., 2011).在水稻幼苗中, OsPR1A和OsPR1B基因的转录与光处理密切相关(Agrawal et al., 2000b), 且外源施加茉莉酸(jasmonic acid, JA)和乙烯(ethylene, ET)能显著增加OsPR1B基因的转录(Luan and Zhou, 2015).GNS1属于PR2家族, 具有β-1,3-葡聚糖酶活性, 在真菌诱导下, 水稻中GNS1基因的转录和蛋白质表达水平均明显升高; 在黑暗处理时, GNS1基因表达水平也有所上升(Nishizawa et al., 2003).转PR3基因的水稻对纹枯病的抗性增强(Datta et al., 2001).超表达OsPR4A的水稻植株在幼苗期和孕穗期抗旱性明显增强(Wang et al., 2011).水稻PR1A、PR5、PR8和PR16蛋白质随盐胁迫时间的延长下调表达, 而PR2在干旱、高盐及水淹胁迫中均被诱导(高庆华等, 2013).OsWRKY67与OsPR1A和OsPR10的启动子区结合进而提高对白叶枯病菌的抗性(Liu et al., 2018).在其它植物中也有大量PR基因的相关报道.拟南芥(Arabidopsis thaliana)受到南方根结线虫(Meloidogyne incognita)侵袭后, PR1、PR2和PR5的表达被大量诱导(Hamamouch et al., 2011).干旱胁迫下, 拟南芥中PR1、PR2和PR5基因表达升高(Seo et al., 2008).在烟草(Nicotiana tabacum)中表达OsGLP1基因可增强其对真菌病原物的抗性(Banerjee et al., 2010). ...
RSOsPR10 expression in response to environmental stresses is regulated antagonistically by jasmonate/ethylene and salicylic acid signaling pathways in rice roots 1 2011
... PR10蛋白结构保守且功能多样, 在植物抗逆方面的功能也有报道(谢纯政等, 2008).水稻OsPR10A蛋白分子量约为17 kDa, 经生化验证其具有RNA酶活性, 经GUS染色确定OsPR10A的转录受到多种方式的调控, 如生物胁迫Xoo的侵染、非生物胁迫伤害及缺磷.此外, 在叶片的正常衰老过程中OsPR10A也会被诱导表达(Huang et al., 2016).PR10A (PBZ1)在细胞衰亡的组织中积累, 可诱导水稻细胞死亡, 也能通过其RNA酶活性诱导拟南芥和烟草细胞死亡(Kim et al., 2011).蛋白质免疫印迹(western blot, WB)分析发现, OsPR10A在水稻受到Xoo侵染后144小时大量表达, 而且不亲和互作中表达量高于亲和互作反应(Wu et al., 2011).超表达OsPR10A增强了水稻对白叶枯病菌的抗性(Huang et al., 2016).OsPR10A的上游有转录因子结合位点, 可能受转录因子OsWRKY6的调控, OsWRKY6与类W-box元件1 (WLE1)物理结合, 正向调控OsPR10A的表达(Choi et al., 2015).序列同源性分析表明, OsPR10A和RSOsPR10高度相似, 具有相似的酶活性质与功能(Hashimoto et al., 2004).RSOsPR10是水稻根部特有的病程相关基因(Hashimoto et al., 2004), 在干旱和高盐胁迫下均可被诱导表达(Takeuchi et al., 2011); 在根皮细胞中超表达RSOsPR10的水稻植株对干旱胁迫具有更强的耐受性, 但对盐胁迫的耐受性较差(Takeuchi et al., 2016).超表达JIOsPR10增强了水稻对稻瘟病菌的抗性, 证明其在水稻防卫机制中发挥作用; 半定量RT-PCR分析表明, 高盐和干旱胁迫条件下, 水稻JIOsPR10的表达上调(Wu et al., 2016); WB检测发现, 其在正常组织(如花和根)中都表达, 衰老及伤害条件也能提高其表达水平(Kim et al., 2008). ...
Overexpression of RSOsPR10, a root-specific rice PR10 gene, confers tolerance against drought stress in rice and drought and salt stresses in bentgrass 1 2016
... PR10蛋白结构保守且功能多样, 在植物抗逆方面的功能也有报道(谢纯政等, 2008).水稻OsPR10A蛋白分子量约为17 kDa, 经生化验证其具有RNA酶活性, 经GUS染色确定OsPR10A的转录受到多种方式的调控, 如生物胁迫Xoo的侵染、非生物胁迫伤害及缺磷.此外, 在叶片的正常衰老过程中OsPR10A也会被诱导表达(Huang et al., 2016).PR10A (PBZ1)在细胞衰亡的组织中积累, 可诱导水稻细胞死亡, 也能通过其RNA酶活性诱导拟南芥和烟草细胞死亡(Kim et al., 2011).蛋白质免疫印迹(western blot, WB)分析发现, OsPR10A在水稻受到Xoo侵染后144小时大量表达, 而且不亲和互作中表达量高于亲和互作反应(Wu et al., 2011).超表达OsPR10A增强了水稻对白叶枯病菌的抗性(Huang et al., 2016).OsPR10A的上游有转录因子结合位点, 可能受转录因子OsWRKY6的调控, OsWRKY6与类W-box元件1 (WLE1)物理结合, 正向调控OsPR10A的表达(Choi et al., 2015).序列同源性分析表明, OsPR10A和RSOsPR10高度相似, 具有相似的酶活性质与功能(Hashimoto et al., 2004).RSOsPR10是水稻根部特有的病程相关基因(Hashimoto et al., 2004), 在干旱和高盐胁迫下均可被诱导表达(Takeuchi et al., 2011); 在根皮细胞中超表达RSOsPR10的水稻植株对干旱胁迫具有更强的耐受性, 但对盐胁迫的耐受性较差(Takeuchi et al., 2016).超表达JIOsPR10增强了水稻对稻瘟病菌的抗性, 证明其在水稻防卫机制中发挥作用; 半定量RT-PCR分析表明, 高盐和干旱胁迫条件下, 水稻JIOsPR10的表达上调(Wu et al., 2016); WB检测发现, 其在正常组织(如花和根)中都表达, 衰老及伤害条件也能提高其表达水平(Kim et al., 2008). ...
Cold stress effects on reproductive development in grain crops: an overview 1 2010
... 水稻(Oryza sativa)是世界上最重要的粮食作物之一, 其高产和稳产对全球尤其是亚洲地区的粮食安全具有重要意义.水稻生长常受到冷害、高温、水淹、干旱和盐碱等恶劣环境的威胁(Thakur et al., 2010). 因此, 了解水稻胁迫应答机理对培育耐逆水稻品种具有重要意义. ...
A human protein atlas for normal and cancer tissues based on antibody proteomics 1 2005
... 蛋白质是生物功能的主要执行者, 了解蛋白质的表达丰度、修饰及其变化是蛋白质组学的主要目标(Pandey and Mann, 2000).利用蛋白质特异性抗体可对特定的目标蛋白进行靶向研究, 这一理念被称为基于抗体的蛋白质组学(antibody-based proteomics, AbP) (Uhlén et al., 2005).通过免疫学方法对蛋白质表达特性展开调查, 具有靶向性强、灵敏度高、操作简便和容易对大量样品进行分析等优点.在水稻及其它植物研究中也可借鉴这一思路, 水稻抗体资源库和蛋白质样品资源库的构建工作也已经开展(刘国振等, 2011; 张剑硕等, 2018). ...
Identification of a cluster of PR4-like genes involved in stress responses in rice 1 2011
... 为应对多变的自然环境, 植物进化出复杂的抗性机制(Nürnberger et al., 2004; Ausubel, 2005).例如, 在受到病原菌侵害或其它生物胁迫时, 植物会激活自身的防御体系来应对侵害.病程相关(pathogenesis- related, PR)蛋白是一类受病原物诱导的蛋白质.水稻是单子叶植物遗传研究的模式生物, 其基因组包含900多个PR基因(窦世娟等, 2014).根据蛋白质的结构, 可将PR蛋白分为17个亚类, 不同亚类的PR蛋白生物学功能也各有不同.探究水稻PR基因的功能有助于深入了解水稻的抗逆机理.大量研究表明, 水稻PR基因在抵抗生物及非生物逆境胁迫中发挥重要作用, 调控特定PR基因的表达会影响水稻的抗逆反应, PR蛋白在抗病反应中也被诱导表达(李雪姣等, 2014).PR1家族的OsPR1A和OsPR1B的转录水平易受病原菌诱导, 可作为抗病反应发生的分子标记(Ponciano et al., 2006).在水稻与白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)互作反应中, OsPR1A和OsPR1B蛋白质在Xoo侵染水稻叶片144小时后大量表达(Wu et al., 2011).在水稻幼苗中, OsPR1A和OsPR1B基因的转录与光处理密切相关(Agrawal et al., 2000b), 且外源施加茉莉酸(jasmonic acid, JA)和乙烯(ethylene, ET)能显著增加OsPR1B基因的转录(Luan and Zhou, 2015).GNS1属于PR2家族, 具有β-1,3-葡聚糖酶活性, 在真菌诱导下, 水稻中GNS1基因的转录和蛋白质表达水平均明显升高; 在黑暗处理时, GNS1基因表达水平也有所上升(Nishizawa et al., 2003).转PR3基因的水稻对纹枯病的抗性增强(Datta et al., 2001).超表达OsPR4A的水稻植株在幼苗期和孕穗期抗旱性明显增强(Wang et al., 2011).水稻PR1A、PR5、PR8和PR16蛋白质随盐胁迫时间的延长下调表达, 而PR2在干旱、高盐及水淹胁迫中均被诱导(高庆华等, 2013).OsWRKY67与OsPR1A和OsPR10的启动子区结合进而提高对白叶枯病菌的抗性(Liu et al., 2018).在其它植物中也有大量PR基因的相关报道.拟南芥(Arabidopsis thaliana)受到南方根结线虫(Meloidogyne incognita)侵袭后, PR1、PR2和PR5的表达被大量诱导(Hamamouch et al., 2011).干旱胁迫下, 拟南芥中PR1、PR2和PR5基因表达升高(Seo et al., 2008).在烟草(Nicotiana tabacum)中表达OsGLP1基因可增强其对真菌病原物的抗性(Banerjee et al., 2010). ...
Overexpression of a pathogenesis-related protein 10 enhances biotic and abiotic stress tolerance in rice 1 2016
... PR10蛋白结构保守且功能多样, 在植物抗逆方面的功能也有报道(谢纯政等, 2008).水稻OsPR10A蛋白分子量约为17 kDa, 经生化验证其具有RNA酶活性, 经GUS染色确定OsPR10A的转录受到多种方式的调控, 如生物胁迫Xoo的侵染、非生物胁迫伤害及缺磷.此外, 在叶片的正常衰老过程中OsPR10A也会被诱导表达(Huang et al., 2016).PR10A (PBZ1)在细胞衰亡的组织中积累, 可诱导水稻细胞死亡, 也能通过其RNA酶活性诱导拟南芥和烟草细胞死亡(Kim et al., 2011).蛋白质免疫印迹(western blot, WB)分析发现, OsPR10A在水稻受到Xoo侵染后144小时大量表达, 而且不亲和互作中表达量高于亲和互作反应(Wu et al., 2011).超表达OsPR10A增强了水稻对白叶枯病菌的抗性(Huang et al., 2016).OsPR10A的上游有转录因子结合位点, 可能受转录因子OsWRKY6的调控, OsWRKY6与类W-box元件1 (WLE1)物理结合, 正向调控OsPR10A的表达(Choi et al., 2015).序列同源性分析表明, OsPR10A和RSOsPR10高度相似, 具有相似的酶活性质与功能(Hashimoto et al., 2004).RSOsPR10是水稻根部特有的病程相关基因(Hashimoto et al., 2004), 在干旱和高盐胁迫下均可被诱导表达(Takeuchi et al., 2011); 在根皮细胞中超表达RSOsPR10的水稻植株对干旱胁迫具有更强的耐受性, 但对盐胁迫的耐受性较差(Takeuchi et al., 2016).超表达JIOsPR10增强了水稻对稻瘟病菌的抗性, 证明其在水稻防卫机制中发挥作用; 半定量RT-PCR分析表明, 高盐和干旱胁迫条件下, 水稻JIOsPR10的表达上调(Wu et al., 2016); WB检测发现, 其在正常组织(如花和根)中都表达, 衰老及伤害条件也能提高其表达水平(Kim et al., 2008). ...
Induction of pathogenesis-related proteins in rice bacterial blight resistant gene XA21-mediated interactions with Xanthomonas oryzae pv. oryzae 3 2011
... 为应对多变的自然环境, 植物进化出复杂的抗性机制(Nürnberger et al., 2004; Ausubel, 2005).例如, 在受到病原菌侵害或其它生物胁迫时, 植物会激活自身的防御体系来应对侵害.病程相关(pathogenesis- related, PR)蛋白是一类受病原物诱导的蛋白质.水稻是单子叶植物遗传研究的模式生物, 其基因组包含900多个PR基因(窦世娟等, 2014).根据蛋白质的结构, 可将PR蛋白分为17个亚类, 不同亚类的PR蛋白生物学功能也各有不同.探究水稻PR基因的功能有助于深入了解水稻的抗逆机理.大量研究表明, 水稻PR基因在抵抗生物及非生物逆境胁迫中发挥重要作用, 调控特定PR基因的表达会影响水稻的抗逆反应, PR蛋白在抗病反应中也被诱导表达(李雪姣等, 2014).PR1家族的OsPR1A和OsPR1B的转录水平易受病原菌诱导, 可作为抗病反应发生的分子标记(Ponciano et al., 2006).在水稻与白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)互作反应中, OsPR1A和OsPR1B蛋白质在Xoo侵染水稻叶片144小时后大量表达(Wu et al., 2011).在水稻幼苗中, OsPR1A和OsPR1B基因的转录与光处理密切相关(Agrawal et al., 2000b), 且外源施加茉莉酸(jasmonic acid, JA)和乙烯(ethylene, ET)能显著增加OsPR1B基因的转录(Luan and Zhou, 2015).GNS1属于PR2家族, 具有β-1,3-葡聚糖酶活性, 在真菌诱导下, 水稻中GNS1基因的转录和蛋白质表达水平均明显升高; 在黑暗处理时, GNS1基因表达水平也有所上升(Nishizawa et al., 2003).转PR3基因的水稻对纹枯病的抗性增强(Datta et al., 2001).超表达OsPR4A的水稻植株在幼苗期和孕穗期抗旱性明显增强(Wang et al., 2011).水稻PR1A、PR5、PR8和PR16蛋白质随盐胁迫时间的延长下调表达, 而PR2在干旱、高盐及水淹胁迫中均被诱导(高庆华等, 2013).OsWRKY67与OsPR1A和OsPR10的启动子区结合进而提高对白叶枯病菌的抗性(Liu et al., 2018).在其它植物中也有大量PR基因的相关报道.拟南芥(Arabidopsis thaliana)受到南方根结线虫(Meloidogyne incognita)侵袭后, PR1、PR2和PR5的表达被大量诱导(Hamamouch et al., 2011).干旱胁迫下, 拟南芥中PR1、PR2和PR5基因表达升高(Seo et al., 2008).在烟草(Nicotiana tabacum)中表达OsGLP1基因可增强其对真菌病原物的抗性(Banerjee et al., 2010). ...
... PR10蛋白结构保守且功能多样, 在植物抗逆方面的功能也有报道(谢纯政等, 2008).水稻OsPR10A蛋白分子量约为17 kDa, 经生化验证其具有RNA酶活性, 经GUS染色确定OsPR10A的转录受到多种方式的调控, 如生物胁迫Xoo的侵染、非生物胁迫伤害及缺磷.此外, 在叶片的正常衰老过程中OsPR10A也会被诱导表达(Huang et al., 2016).PR10A (PBZ1)在细胞衰亡的组织中积累, 可诱导水稻细胞死亡, 也能通过其RNA酶活性诱导拟南芥和烟草细胞死亡(Kim et al., 2011).蛋白质免疫印迹(western blot, WB)分析发现, OsPR10A在水稻受到Xoo侵染后144小时大量表达, 而且不亲和互作中表达量高于亲和互作反应(Wu et al., 2011).超表达OsPR10A增强了水稻对白叶枯病菌的抗性(Huang et al., 2016).OsPR10A的上游有转录因子结合位点, 可能受转录因子OsWRKY6的调控, OsWRKY6与类W-box元件1 (WLE1)物理结合, 正向调控OsPR10A的表达(Choi et al., 2015).序列同源性分析表明, OsPR10A和RSOsPR10高度相似, 具有相似的酶活性质与功能(Hashimoto et al., 2004).RSOsPR10是水稻根部特有的病程相关基因(Hashimoto et al., 2004), 在干旱和高盐胁迫下均可被诱导表达(Takeuchi et al., 2011); 在根皮细胞中超表达RSOsPR10的水稻植株对干旱胁迫具有更强的耐受性, 但对盐胁迫的耐受性较差(Takeuchi et al., 2016).超表达JIOsPR10增强了水稻对稻瘟病菌的抗性, 证明其在水稻防卫机制中发挥作用; 半定量RT-PCR分析表明, 高盐和干旱胁迫条件下, 水稻JIOsPR10的表达上调(Wu et al., 2016); WB检测发现, 其在正常组织(如花和根)中都表达, 衰老及伤害条件也能提高其表达水平(Kim et al., 2008). ...
... 用Tricine-SDS-PAGE分离提取的水稻总蛋白质, 上样体积为10 µL, 然后转移到PVDF膜上, 蛋白质免疫印迹一抗为Anti-OsPR10A polyclonal抗体(Wu et al., 2011), 二抗为HRP标记的羊抗兔, 具体操作参见白辉等(2010)所述方法.以HSP抗体检测的信号作为内参(Li et al., 2011), 所用抗体购自北京华大蛋白质研发中心有限公司.用化学发光成像仪(MiniChemi610, 北京赛智创业科技有限公司)检测信号.WB实验至少重复3次. ...
Feedback regulation of ABA signaling and biosynthesis by a bZIP transcription factor targets drought- resistance-related genes 1 2016
... 本研究还发现, 在离体水稻叶片激素处理过程中, OsPR10A的表达丰度受MeJA和ABA诱导, 表明该蛋白功能的发挥与JA和ABA的信号转导途径有关.与野生型相比, OsJAZ1超表达植株对干旱敏感, 同时对MeJA和ABA表现更为敏感, RNA-seq分析结果表明, 在干旱处理下, JA和ABA信号转导途径中许多基因的表达水平在OsJAZ1超表达植株和野生型之间差异显著, 表明JA和ABA介导的途径参与水稻对干旱胁迫的抵御(Fu et al., 2017).水稻转录因子OsbZIP23与耐旱性有关, 同时也是ABA信号途径和ABA合成的关键调控子(Zong et al., 2016).水稻OsASR5与水稻耐旱性有关, 超表达OsASR5使种子萌发期和萌发后期内源ABA水平增高, 对外源施加的ABA也更为敏感(Li et al., 2017).水稻ABA8ox3是ABA生物合成的关键基因, 其转基因RNAi株系耐旱性更强, 而超表达株系对干旱胁迫更为敏感(Cai et al., 2015).本研究中, OsPR10A分别受干旱胁迫和激素(MeJA和ABA)的诱导, 推测该蛋白可能通过JA和ABA相关的信号转导途径增强水稻的耐旱性. ...
 ,1,*, 窦世娟
,1,*, 窦世娟
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT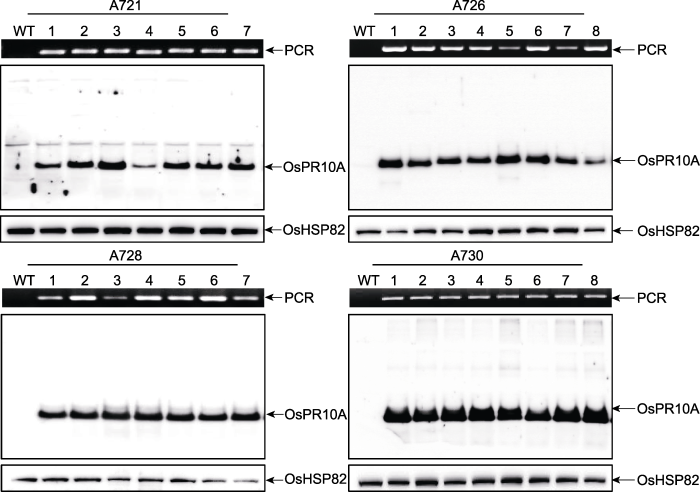 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT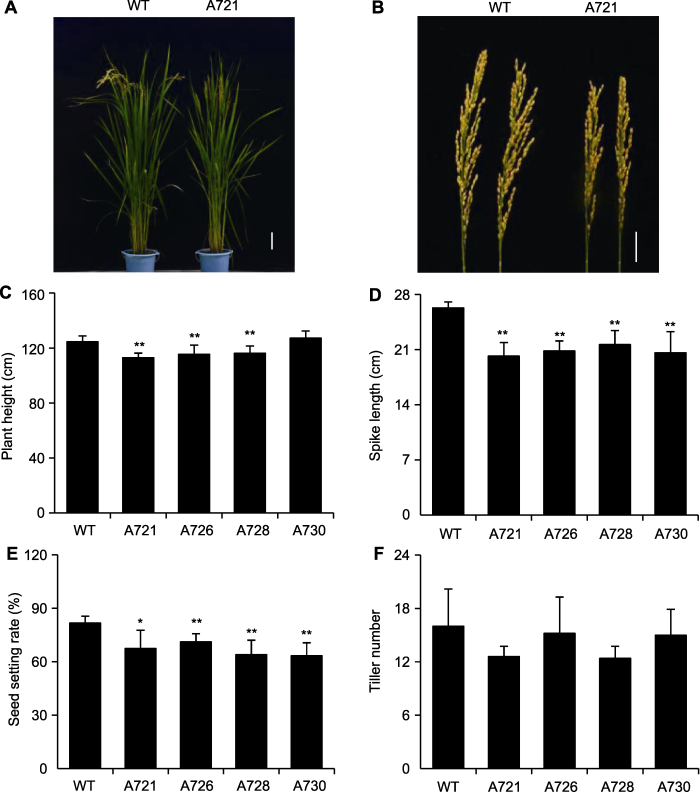 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT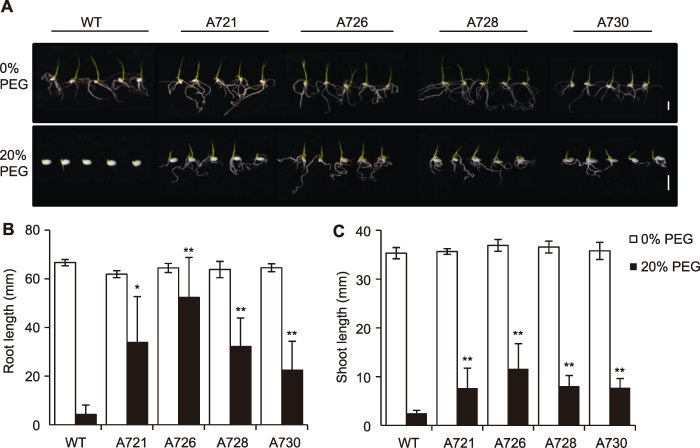 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}