Abstract Previous studies have shown that the mitochondrial alternative oxidase (AOX) respiratory pathway contributes to the photoprotection of photosystem II (PSII) under high light. However, the role of another respiratory pathway in mitochondria, the cytochrome-respiratory (COX) pathway, to photoprotection under high light remains unknown. Here, we studied the contributions of COX and AOX pathways to PSII photoprotection, as well as the relationship between COX and AOX pathways in PSII photoprotection in tobacco leaves under high light using chlorophyll fluorescence transients and chlorophyll fluorescence quenching approaches. After high light treatment, the PSII activity decreased in all leaves. The inhibition of AOX pathway significantly accelerated the decrease of PSII activity in leaves. However, there was no significant difference between the PSII activity in the leaves pretreated with COX pathway inhibitor and water-pretreated control leaves. When both AOX and COX pathways are inhibited in leaves, the decrease in PSII activity was severe, than that of leaves pretreated by only AOX pathway inhibitor. In addition, inhibition of respiratory electron transport resulted in an increase of non-photochemical quenching (NPQ). The up-regulation of NPQ caused by AOX pathway inhibitor was more obviously than that caused by COX pathway inhibitor, and the increase of NPQ was highest when both AOX and COX pathways were inhibited. These results indicate that both COX and AOX pathways contribute to the PSII photoprotection in tobacco leaves under high light. In addition, when COX pathway is inhibited, its photoprotective function can be compensated by AOX pathway and NPQ, however, photoprotection of AOX pathway cannot be completely compensated by COX pathway and NPQ. Keywords:alternative oxidation respiratory pathway;cytochrome respiratory pathway;non-photochemical quenching;photoprotection;PSII photoinhibition
PDF (1401KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 罗蛟, 李玉婷, 张子山, 车兴凯, 梁英, 李月楠, 李滢, 赵世杰, 高辉远. 烟草叶片中呼吸电子传递途径在缓解叶绿体PSII光抑制中的作用. 植物学报, 2020, 55(1): 31-37 doi:10.11983/CBB19117 Luo Jiao, Li Yuting, Zhang Zishan, Che Xingkai, Liang Ying, Li Yuenan, Li Ying, Zhao Shijie, Gao Huiyuan. Effects of the Respiratory Electron Transport Pathways in Relieving Photoinhibition of Chloroplast PSII in Tobacco Leaves. Chinese Bulletin of Botany, 2020, 55(1): 31-37 doi:10.11983/CBB19117
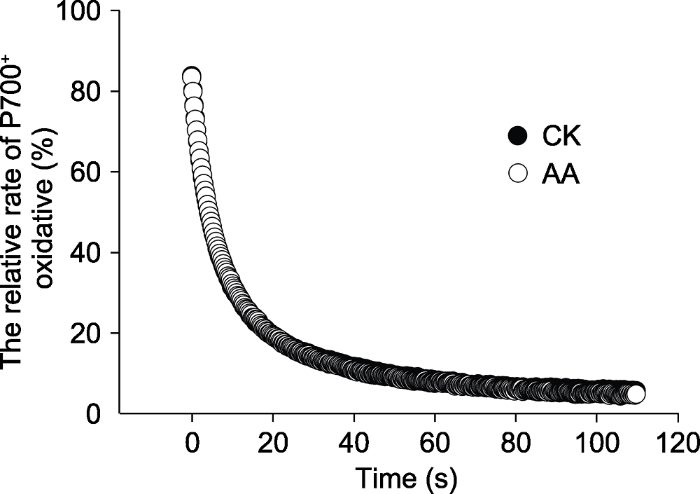
所有数值均为8次重复的平均值。 Figure 1The effect of water (CK) or 2 μmol∙L-1 antimycin A (AA) treatment on the P700+ relaxation curve after far-red light in tobacco leaves
所有数值均为8次重复的平均值。不同小写字母表示差异显著(P<0.05)。 Figure 2The effect of 0 (A) or 1 h (B) high light treatment on the OJIP transients in different pretreated tobacco leaves and the effect of high light treatment on the maximum quantum yield of photosystem II (Fv/Fm) (C) in different pretreated tobacco leaves
All data are the average of 8 replicates. Different lowercase letters indicate significant differences (P<0.05).
所有数值均为8次重复的平均值。不同小写字母表示差异显著(P<0.05)。 Figure 3The effect of high light treatment on the actual PSII photochemical efficiencies (ФPSII ) (A), the efficiency of excitation energy capture by open PSII reaction centres (Fv′/Fm′) (B), the photochemical quenching coefficients (qP) (C) and the non-photochemical quenching (NPQ) (D) in different pretreated tobacco leaves
All data are the average of 8 replicates. Different lowercase letters indicate significant differences (P<0.05).
AroEM, SuorsaM, RokkaA, AllahverdiyevaY, PaakkarinenV, SaleemA, BattchikovaN, Rintam?kiE (2005). Dynamics of photosystem II: a proteomic approach to thylakoid protein complexes J Exp Bot 56, 347-356. [本文引用: 1]
BartoliCG, GomezF, GergoffG, GuiamétJJ, PuntaruloS (2005). Up-regulation of the mitochondrial alternative oxidase pathway enhances photosynthetic electron transport under drought conditions J Exp Bot 53, 1269-1276. [本文引用: 2]
Florez-SarasaI, Ribas-CarboM, Del-SazNF, SchwahnK, NikoloskiZ, FernieAR, FlexasJ (2016). Unravelling the in vivo regulation and metabolic role of the alternative oxidase pathway in C3 species under photoinhibitory conditions New Phytol 212, 66-79. [本文引用: 1]
Jo?tT, CournacL, HorvathEM, MedgyesyP, PeltierG (2011). Increased sensitivity of photosynthesis to antimycin a induced by inactivation of the chloroplast ndhB gene. Evidence for a participation of the NADH-dehydrogenase complex to cyclic electron flow around photosystem I Plant Physiol 125, 1919-1929. [本文引用: 1]
JuszczukIM, RychterAM (2003). Alternative oxidase in higher plants Acta Biochim Pol 50, 1257-1271. [本文引用: 1]
McDonaldAE (2008). Alternative oxidase: an inter-kingdom perspective on the function and regulation of this broadly distributed ‘cyanide-resistant’ terminal oxidase Funct Plant Biol 35, 535-552. [本文引用: 2]
MiyakeC (2010). Alternative electron flows (water-water cycle and cyclic electron flow around PSI) in photosynthesis: molecular mechanisms and physiological functions Plant Cell Physiol 51, 1951-1963. [本文引用: 1]
NishiyamaY, AllakhverdievSI, MurataN (2011). Protein synthesis is the primary target of reactive oxygen species in the photoinhibition of photosystem II Physiol Plant 142, 35-46. [本文引用: 2]
Nunes-NesiA, SweetloveLJ, FernieAR (2007). Operation and function of the tricarboxylic acid cycle in the illuminated leaf Physiol Plant 129, 45-56. [本文引用: 1]
PadmasreeK, RaghavendraAS (1999). Importance of oxidative electron transport over oxidative phosphorylation in optimizing photosynthesis in mesophyll protoplasts of pea ( Pisum sativum L.) Physiol Plant 105, 546-553. [本文引用: 2]
Pospi??ilP (2009). Production of reactive oxygen species by photosystem II Biochim Biophys Acta 1787, 1151-1160. [本文引用: 1]
RubanAV (2016). Nonphotochemical chlorophyll fluorescence quenching: mechanism and effectiveness in protecting plants from photodamage Plant Physiol 170, 1903-1916. [本文引用: 3]
SelinskiJ, ScheibeR, DayDA, WhelanJ (2018). Alternative oxidase is positive for plant performance Trends Plant Sci 23, 588-597. [本文引用: 4]
TakahashiS, MurataN (2008). How do environmental stresses accelerate photoinhibition? Trends Plant Sci 13, 178-182. [本文引用: 1]
TcherkezG, BlignyR, GoutE, MahéA, HodgesM, CornicG (2008). Respiratory metabolism of illuminated leaves depends on CO2 and O2 conditions Proc Natl Acad Sci USA 105, 797-802. [本文引用: 1]
WangP, DuanW, TakabayashiA, EndoT, ShikanaiT, YeJY, MiHL (2006). Chloroplastic NAD(P)H dehydrogenase in tobacco leaves functions in alleviation of oxidative damage caused by temperature stress Plant Physiol 141, 465-474. [本文引用: 1]
WatanabeCKA, YamoriW, TakahashiS, TerashimaI, NoguchiK (2016). Mitochondrial alternative pathway- associated photoprotection of photosystem II is related to the photorespiratory pathway Plant Cell Physiol 57, 1426-1431. [本文引用: 3]
YoshidaK, TerashimaI, NoguchiK (2006). Distinct roles of the cytochrome pathway and alternative oxidase in leaf photosynthesis Plant Cell Physiol 47, 22-31. [本文引用: 4]
YoshidaK, TerashimaI, NoguchiK (2007). Up-regulation of mitochondrial alternative oxidase concomitant with chloroplast over-reduction by excess light Plant Cell Physiol 48, 606-614. [本文引用: 2]
YoshidaK, TerashimaI, NoguchiK (2011). How and why does mitochondrial respiratory chain respond to light? Plant Signal Behav 6, 864-866. [本文引用: 4]
ZhangLT, GaoHY, ZhangZS, XueZC, MengQW (2012a). Multiple effects of inhibition of mitochondrial alternative oxidase pathway on photosynthetic apparatus in Rumex K-1 leaves Biol Plant 56, 365-368. [本文引用: 1]
ZhangLT, ZhangZS, GaoHY, MengXL, YangC, LiuJG, MengQW (2012b). The mitochondrial alternative oxidase pathway protects the photosynthetic apparatus against photodamage in Rumex K-1 leaves BMC Plant Biol 12, 40. [本文引用: 1]
ZhangLT, ZhangZS, GaoHY, XueZC, YangC, MengXL, MengQW (2011a). Mitochondrial alternative oxidase pathway protects plants against photoinhibition by alleviating inhibition of the repair of photodamaged PSII through preventing formation of reactive oxygen species in Rumex K-1 leaves Physiol Plant 143, 396-407. [本文引用: 3]
ZhangZS, JiaYJ, GaoHY, ZhangLT, LiHD, MengQW (2011b). Characterization of PSI recovery after chilling-induced photoinhibition in cucumber (Cucumis sativus L.) leaves Planta 234, 883-889. [本文引用: 1]
ZhangZS, LiuMJ, ScheibeR, SelinskiJ, ZhangLT, YangC, MengXL, GaoHY (2017). Contribution of the alternative respiratory pathway to PSII photoprotection in C3 and C4 plants Mol Plant 10, 131-142. [本文引用: 2]
突变体在植物光破坏防御研究中的应用 1 2003
... 光抑制是指光合放氧及光合机构电子传递效率减弱的现象(陈玮等, 2003; 李天来等, 2016).植物处于常温强光下时, 光合机构中光抑制主要发生在光系统II (PSII) (张子山等, 2009).PSII位于光合电子传递链的上游, 强光产生的过多还原力会造成PSII受体侧电子传递链过还原, 导致过剩激发能增加, 生成活性氧, 进而对PSII造成伤害(Pospi?šil, 2009).PSII光抑制的本质是PSII核心蛋白, 尤其是D1蛋白的净降解.D1蛋白在光下时刻处于降解-合成的循环之中, 被称为D1蛋白周转(Aro et al., 2005; Nishiyama et al., 2011).研究表明, 强光及其它非生物胁迫均通过抑制D1蛋白合成导致D1蛋白净降解, 最终引发PSII光抑制(Takahashi and Murata, 2008; Nishiyama et al., 2011).为了维持D1蛋白的周转和缓解PSII光抑制, 植物进化出许多光保护机制, 如热耗散(Ruban, 2016)、水水循环(Miyake, 2010)及环式电子传递(薛娴等, 2017), 而这些光保护途径均存在于叶绿体内.在叶绿体外也存在光保护途径, 目前研究最多的是线粒体内的交替氧化酶(AOX)途径. ...
... 光抑制是指光合放氧及光合机构电子传递效率减弱的现象(陈玮等, 2003; 李天来等, 2016).植物处于常温强光下时, 光合机构中光抑制主要发生在光系统II (PSII) (张子山等, 2009).PSII位于光合电子传递链的上游, 强光产生的过多还原力会造成PSII受体侧电子传递链过还原, 导致过剩激发能增加, 生成活性氧, 进而对PSII造成伤害(Pospi?šil, 2009).PSII光抑制的本质是PSII核心蛋白, 尤其是D1蛋白的净降解.D1蛋白在光下时刻处于降解-合成的循环之中, 被称为D1蛋白周转(Aro et al., 2005; Nishiyama et al., 2011).研究表明, 强光及其它非生物胁迫均通过抑制D1蛋白合成导致D1蛋白净降解, 最终引发PSII光抑制(Takahashi and Murata, 2008; Nishiyama et al., 2011).为了维持D1蛋白的周转和缓解PSII光抑制, 植物进化出许多光保护机制, 如热耗散(Ruban, 2016)、水水循环(Miyake, 2010)及环式电子传递(薛娴等, 2017), 而这些光保护途径均存在于叶绿体内.在叶绿体外也存在光保护途径, 目前研究最多的是线粒体内的交替氧化酶(AOX)途径. ...
小麦叶片PSI光抑制对光合电子传递链的影响 1 2018
... 本实验以SHAM专一性抑制AOX途径(Padmasree and Raghavendra, 1999; Bartoli et al., 2005), 以2 μmol∙L-1抗霉素A抑制COX途径(Yoshida et al., 2006; Watanabe et al., 2016).较高浓度的抗霉素A可以抑制依赖PGR5的PSI环式电子传递(Joët et al, 2001), PSI环式电子传递受抑后会影响PSII电子传递和光保护(梁英等, 2018).为了验证2 μmol∙L-1抗霉素A对烟草叶片PSI环式电子传递的影响, 我们测定了远红光关闭后P700+的弛豫过程.结果显示, 2 μmol∙L-1抗霉素A处理烟草叶片的P700+弛豫与水处理叶片无明显差异(图1), 表明2 μmol∙L-1抗霉素A处理并未抑制PSI环式电子传递. ...
植物光合作用循环电子传递的研究进展 1 2017
... 光抑制是指光合放氧及光合机构电子传递效率减弱的现象(陈玮等, 2003; 李天来等, 2016).植物处于常温强光下时, 光合机构中光抑制主要发生在光系统II (PSII) (张子山等, 2009).PSII位于光合电子传递链的上游, 强光产生的过多还原力会造成PSII受体侧电子传递链过还原, 导致过剩激发能增加, 生成活性氧, 进而对PSII造成伤害(Pospi?šil, 2009).PSII光抑制的本质是PSII核心蛋白, 尤其是D1蛋白的净降解.D1蛋白在光下时刻处于降解-合成的循环之中, 被称为D1蛋白周转(Aro et al., 2005; Nishiyama et al., 2011).研究表明, 强光及其它非生物胁迫均通过抑制D1蛋白合成导致D1蛋白净降解, 最终引发PSII光抑制(Takahashi and Murata, 2008; Nishiyama et al., 2011).为了维持D1蛋白的周转和缓解PSII光抑制, 植物进化出许多光保护机制, 如热耗散(Ruban, 2016)、水水循环(Miyake, 2010)及环式电子传递(薛娴等, 2017), 而这些光保护途径均存在于叶绿体内.在叶绿体外也存在光保护途径, 目前研究最多的是线粒体内的交替氧化酶(AOX)途径. ...
不同光强与低温交叉胁迫下黄瓜PSI与PSII的光抑制研究 1 2009
... 光抑制是指光合放氧及光合机构电子传递效率减弱的现象(陈玮等, 2003; 李天来等, 2016).植物处于常温强光下时, 光合机构中光抑制主要发生在光系统II (PSII) (张子山等, 2009).PSII位于光合电子传递链的上游, 强光产生的过多还原力会造成PSII受体侧电子传递链过还原, 导致过剩激发能增加, 生成活性氧, 进而对PSII造成伤害(Pospi?šil, 2009).PSII光抑制的本质是PSII核心蛋白, 尤其是D1蛋白的净降解.D1蛋白在光下时刻处于降解-合成的循环之中, 被称为D1蛋白周转(Aro et al., 2005; Nishiyama et al., 2011).研究表明, 强光及其它非生物胁迫均通过抑制D1蛋白合成导致D1蛋白净降解, 最终引发PSII光抑制(Takahashi and Murata, 2008; Nishiyama et al., 2011).为了维持D1蛋白的周转和缓解PSII光抑制, 植物进化出许多光保护机制, 如热耗散(Ruban, 2016)、水水循环(Miyake, 2010)及环式电子传递(薛娴等, 2017), 而这些光保护途径均存在于叶绿体内.在叶绿体外也存在光保护途径, 目前研究最多的是线粒体内的交替氧化酶(AOX)途径. ...
Dynamics of photosystem II: a proteomic approach to thylakoid protein complexes 1 2005
... 光抑制是指光合放氧及光合机构电子传递效率减弱的现象(陈玮等, 2003; 李天来等, 2016).植物处于常温强光下时, 光合机构中光抑制主要发生在光系统II (PSII) (张子山等, 2009).PSII位于光合电子传递链的上游, 强光产生的过多还原力会造成PSII受体侧电子传递链过还原, 导致过剩激发能增加, 生成活性氧, 进而对PSII造成伤害(Pospi?šil, 2009).PSII光抑制的本质是PSII核心蛋白, 尤其是D1蛋白的净降解.D1蛋白在光下时刻处于降解-合成的循环之中, 被称为D1蛋白周转(Aro et al., 2005; Nishiyama et al., 2011).研究表明, 强光及其它非生物胁迫均通过抑制D1蛋白合成导致D1蛋白净降解, 最终引发PSII光抑制(Takahashi and Murata, 2008; Nishiyama et al., 2011).为了维持D1蛋白的周转和缓解PSII光抑制, 植物进化出许多光保护机制, 如热耗散(Ruban, 2016)、水水循环(Miyake, 2010)及环式电子传递(薛娴等, 2017), 而这些光保护途径均存在于叶绿体内.在叶绿体外也存在光保护途径, 目前研究最多的是线粒体内的交替氧化酶(AOX)途径. ...
Up-regulation of the mitochondrial alternative oxidase pathway enhances photosynthetic electron transport under drought conditions 2 2005
... 本研究以烟草(Nicotiana tabacum)叶片为材料, 利用AOX途径的专一性抑制剂水杨基羟肟酸(SHAM) (Padmasree and Raghavendra, 1999; Bartoli et al., 2005)和COX途径的专一性抑制剂抗霉素A (Yoshida et al., 2006)处理烟草叶片, 测定了叶绿素荧光快速诱导动力学曲线并进行叶绿素荧光淬灭分析, 系统研究了AOX和COX途径受抑后叶片PSII电子传递活性和光保护途径的变化, 旨在探讨AOX途径和COX途径对PSII光保护的贡献及二者之间的关系. ...
... 本实验以SHAM专一性抑制AOX途径(Padmasree and Raghavendra, 1999; Bartoli et al., 2005), 以2 μmol∙L-1抗霉素A抑制COX途径(Yoshida et al., 2006; Watanabe et al., 2016).较高浓度的抗霉素A可以抑制依赖PGR5的PSI环式电子传递(Joët et al, 2001), PSI环式电子传递受抑后会影响PSII电子传递和光保护(梁英等, 2018).为了验证2 μmol∙L-1抗霉素A对烟草叶片PSI环式电子传递的影响, 我们测定了远红光关闭后P700+的弛豫过程.结果显示, 2 μmol∙L-1抗霉素A处理烟草叶片的P700+弛豫与水处理叶片无明显差异(图1), 表明2 μmol∙L-1抗霉素A处理并未抑制PSI环式电子传递. ...
Unravelling the in vivo regulation and metabolic role of the alternative oxidase pathway in C3 species under photoinhibitory conditions 1 2016
... NPQ是位于叶绿体内的PSII最重要的光保护途径之一(Ruban, 2016).本研究还表明, 当AOX或COX途径受抑时, NPQ都会明显上调(图3D).AOX和COX途径可以与叶绿体内的光保护途径相互配合, 共同保护PSII, 在AOX和COX途径受抑时可以上调NPQ进行补偿.尤其是在COX途径受抑时, 虽然PSII最大光化学效率和PSII实际光化学效率都没有明显下降, 但NPQ仍明显上调.作为叶绿体外的光保护途径, AOX和COX途径与叶绿体内的光保护途径的配合和调控机制研究较少, 之前有研究针对AOX途径与环式电子传递的关系进行探讨, 但没有得出一致的结论(Yoshida et al., 2011; Florez-Sarasa et al., 2016).本研究为进一步探讨叶绿体内外的光保护途径相互配合机制和调控过程提供了新的切入点. ...
Increased sensitivity of photosynthesis to antimycin a induced by inactivation of the chloroplast ndhB gene. Evidence for a participation of the NADH-dehydrogenase complex to cyclic electron flow around photosystem I 1 2011
... 本实验以SHAM专一性抑制AOX途径(Padmasree and Raghavendra, 1999; Bartoli et al., 2005), 以2 μmol∙L-1抗霉素A抑制COX途径(Yoshida et al., 2006; Watanabe et al., 2016).较高浓度的抗霉素A可以抑制依赖PGR5的PSI环式电子传递(Joët et al, 2001), PSI环式电子传递受抑后会影响PSII电子传递和光保护(梁英等, 2018).为了验证2 μmol∙L-1抗霉素A对烟草叶片PSI环式电子传递的影响, 我们测定了远红光关闭后P700+的弛豫过程.结果显示, 2 μmol∙L-1抗霉素A处理烟草叶片的P700+弛豫与水处理叶片无明显差异(图1), 表明2 μmol∙L-1抗霉素A处理并未抑制PSI环式电子传递. ...
Alternative oxidase in higher plants 1 2003
... 虽然AOX途径的光保护作用已被广泛研究, 但在线粒体中与AOX途径平行的COX途径是否也具有光保护功能尚不清楚.AOX和COX途径都介导从UQ向下游的电子传递, 已有研究表明, 抑制AOX和COX都可能影响线粒体内的活性氧和还原力的含量(Siedow and Umbach, 2000; Juszczuk and Rychter, 2003).然而由于COX途径与跨膜质子梯度的形成及ATP合成相偶联, 其速度会受到跨膜质子梯度和ADP不足的限制, 因此, 推测COX途径不能像AOX途径一样快速消耗还原力, 从而起到明显的光保护作用(Selinski et al., 2018).但是, 上述推测一直没有得到验证.此外, AOX和COX途径是平行的电子传递途径, 在光保护作用中二者之间的关系也不清楚. ...
Alternative oxidase: an inter-kingdom perspective on the function and regulation of this broadly distributed ‘cyanide-resistant’ terminal oxidase 2 2008
... 在植物线粒体中, 还原力NADH和FADH2分别通过NADH脱氢酶(复合体I)和琥珀酸还原酶(复合体II)进入呼吸电子传递链并将电子传递至泛醌(Selinski et al., 2018).泛醌的电子可以通过2条途径传递给氧气生成水(McDonald, 2008; Yoshida et al., 2011): (1) 电子流经复合体III和复合体IV最终通过细胞色素氧化酶(COX)传递给氧气, 期间偶联质子的跨膜运输形成跨膜质子梯度用于ATP的合成; (2) 电子通过AOX途径直接传递给氧气, 期间不偶联跨膜质子梯度和ATP的产生(Yoshida et al., 2011; Selinski et al., 2018).前人研究表明, AOX途径可以消耗强光下从叶绿体转运至线粒体的过剩还原力, 从而缓解PSII光抑制(Yoshida et al., 2006, 2007; Zhang et al., 2011a).AOX途径被抑制后, 苹果酸-草酰乙酸穿梭机制中的NADP-苹果酸还原酶的活性显著下降, 植物细胞内的活性氧含量增加, 光抑制明显加重(Zhang et al., 2011a).另有研究表明, 光呼吸过程中在线粒体内进行甘氨酸向丝氨酸的转化并产生大量还原力, AOX途径通过消耗这些还原力维持光呼吸运转, 避免有毒的光呼吸中间产物积累, 从而保护PSII (Watanabe et al., 2016; Zhang et al., 2017). ...
... 由于人们一直认为COX途径不起光保护作用, 对COX途径参与光保护作用的机理知之甚少.然而, AOX途径和COX途径是介导电子从UQ传递到氧气的2条平行途径(McDonald, 2008; Yoshida et al., 2011), 我们推测AOX和COX途径参与光保护的机制可能类似.COX可能通过2条途径参与PSII光保护: (1) 强光下叶绿体内产生的过剩还原力通过苹果酸-草酰乙酸(Malate-OAA)穿梭(Yoshida et al., 2006, 2007)被转运到线粒体内, COX途径可以通过消耗还原力缓解叶绿体的过还原, 最终缓解PSII光抑制; (2) 光呼吸过程中, 在线粒体内会发生甘氨酸向丝氨酸的转化并伴随着大量还原力的产生, COX途径可以通过消耗还原力维持光呼吸的运转, 避免有毒光呼吸中间代谢产物的积累, 从而缓解PSII光抑制(Zhang et al., 2017). ...
Alternative electron flows (water-water cycle and cyclic electron flow around PSI) in photosynthesis: molecular mechanisms and physiological functions 1 2010
... 光抑制是指光合放氧及光合机构电子传递效率减弱的现象(陈玮等, 2003; 李天来等, 2016).植物处于常温强光下时, 光合机构中光抑制主要发生在光系统II (PSII) (张子山等, 2009).PSII位于光合电子传递链的上游, 强光产生的过多还原力会造成PSII受体侧电子传递链过还原, 导致过剩激发能增加, 生成活性氧, 进而对PSII造成伤害(Pospi?šil, 2009).PSII光抑制的本质是PSII核心蛋白, 尤其是D1蛋白的净降解.D1蛋白在光下时刻处于降解-合成的循环之中, 被称为D1蛋白周转(Aro et al., 2005; Nishiyama et al., 2011).研究表明, 强光及其它非生物胁迫均通过抑制D1蛋白合成导致D1蛋白净降解, 最终引发PSII光抑制(Takahashi and Murata, 2008; Nishiyama et al., 2011).为了维持D1蛋白的周转和缓解PSII光抑制, 植物进化出许多光保护机制, 如热耗散(Ruban, 2016)、水水循环(Miyake, 2010)及环式电子传递(薛娴等, 2017), 而这些光保护途径均存在于叶绿体内.在叶绿体外也存在光保护途径, 目前研究最多的是线粒体内的交替氧化酶(AOX)途径. ...
Protein synthesis is the primary target of reactive oxygen species in the photoinhibition of photosystem II 2 2011
... 光抑制是指光合放氧及光合机构电子传递效率减弱的现象(陈玮等, 2003; 李天来等, 2016).植物处于常温强光下时, 光合机构中光抑制主要发生在光系统II (PSII) (张子山等, 2009).PSII位于光合电子传递链的上游, 强光产生的过多还原力会造成PSII受体侧电子传递链过还原, 导致过剩激发能增加, 生成活性氧, 进而对PSII造成伤害(Pospi?šil, 2009).PSII光抑制的本质是PSII核心蛋白, 尤其是D1蛋白的净降解.D1蛋白在光下时刻处于降解-合成的循环之中, 被称为D1蛋白周转(Aro et al., 2005; Nishiyama et al., 2011).研究表明, 强光及其它非生物胁迫均通过抑制D1蛋白合成导致D1蛋白净降解, 最终引发PSII光抑制(Takahashi and Murata, 2008; Nishiyama et al., 2011).为了维持D1蛋白的周转和缓解PSII光抑制, 植物进化出许多光保护机制, 如热耗散(Ruban, 2016)、水水循环(Miyake, 2010)及环式电子传递(薛娴等, 2017), 而这些光保护途径均存在于叶绿体内.在叶绿体外也存在光保护途径, 目前研究最多的是线粒体内的交替氧化酶(AOX)途径. ...
Operation and function of the tricarboxylic acid cycle in the illuminated leaf 1 2007
... 传统观点认为, COX途径由于受跨膜质子梯度和能荷平衡的限制, 其活性不能通过大幅上调来快速消耗还原力, 因此COX途径不发挥主要的光保护功能(Selinski et al., 2018).然而本研究结果不支持这一猜测.事实上, 当植物进行光合作用时, 线粒体中的糖酵解和TCA循环很弱, 产生的还原力很少(Nunes- Nesi et al., 2007; Tcherkez et al., 2008).此时无论是AOX途径还是COX途径消耗的还原力均来自叶绿体, 所以, 虽然COX途径的活性不像AOX途径那样大幅上调, 但COX途径仍然消耗了可观的还原力, 因此, 也具有一定程度的光保护作用. ...
Importance of oxidative electron transport over oxidative phosphorylation in optimizing photosynthesis in mesophyll protoplasts of pea ( Pisum sativum L.) 2 1999
... 本研究以烟草(Nicotiana tabacum)叶片为材料, 利用AOX途径的专一性抑制剂水杨基羟肟酸(SHAM) (Padmasree and Raghavendra, 1999; Bartoli et al., 2005)和COX途径的专一性抑制剂抗霉素A (Yoshida et al., 2006)处理烟草叶片, 测定了叶绿素荧光快速诱导动力学曲线并进行叶绿素荧光淬灭分析, 系统研究了AOX和COX途径受抑后叶片PSII电子传递活性和光保护途径的变化, 旨在探讨AOX途径和COX途径对PSII光保护的贡献及二者之间的关系. ...
... 本实验以SHAM专一性抑制AOX途径(Padmasree and Raghavendra, 1999; Bartoli et al., 2005), 以2 μmol∙L-1抗霉素A抑制COX途径(Yoshida et al., 2006; Watanabe et al., 2016).较高浓度的抗霉素A可以抑制依赖PGR5的PSI环式电子传递(Joët et al, 2001), PSI环式电子传递受抑后会影响PSII电子传递和光保护(梁英等, 2018).为了验证2 μmol∙L-1抗霉素A对烟草叶片PSI环式电子传递的影响, 我们测定了远红光关闭后P700+的弛豫过程.结果显示, 2 μmol∙L-1抗霉素A处理烟草叶片的P700+弛豫与水处理叶片无明显差异(图1), 表明2 μmol∙L-1抗霉素A处理并未抑制PSI环式电子传递. ...
Production of reactive oxygen species by photosystem II 1 2009
... 光抑制是指光合放氧及光合机构电子传递效率减弱的现象(陈玮等, 2003; 李天来等, 2016).植物处于常温强光下时, 光合机构中光抑制主要发生在光系统II (PSII) (张子山等, 2009).PSII位于光合电子传递链的上游, 强光产生的过多还原力会造成PSII受体侧电子传递链过还原, 导致过剩激发能增加, 生成活性氧, 进而对PSII造成伤害(Pospi?šil, 2009).PSII光抑制的本质是PSII核心蛋白, 尤其是D1蛋白的净降解.D1蛋白在光下时刻处于降解-合成的循环之中, 被称为D1蛋白周转(Aro et al., 2005; Nishiyama et al., 2011).研究表明, 强光及其它非生物胁迫均通过抑制D1蛋白合成导致D1蛋白净降解, 最终引发PSII光抑制(Takahashi and Murata, 2008; Nishiyama et al., 2011).为了维持D1蛋白的周转和缓解PSII光抑制, 植物进化出许多光保护机制, 如热耗散(Ruban, 2016)、水水循环(Miyake, 2010)及环式电子传递(薛娴等, 2017), 而这些光保护途径均存在于叶绿体内.在叶绿体外也存在光保护途径, 目前研究最多的是线粒体内的交替氧化酶(AOX)途径. ...
Nonphotochemical chlorophyll fluorescence quenching: mechanism and effectiveness in protecting plants from photodamage 3 2016
... 光抑制是指光合放氧及光合机构电子传递效率减弱的现象(陈玮等, 2003; 李天来等, 2016).植物处于常温强光下时, 光合机构中光抑制主要发生在光系统II (PSII) (张子山等, 2009).PSII位于光合电子传递链的上游, 强光产生的过多还原力会造成PSII受体侧电子传递链过还原, 导致过剩激发能增加, 生成活性氧, 进而对PSII造成伤害(Pospi?šil, 2009).PSII光抑制的本质是PSII核心蛋白, 尤其是D1蛋白的净降解.D1蛋白在光下时刻处于降解-合成的循环之中, 被称为D1蛋白周转(Aro et al., 2005; Nishiyama et al., 2011).研究表明, 强光及其它非生物胁迫均通过抑制D1蛋白合成导致D1蛋白净降解, 最终引发PSII光抑制(Takahashi and Murata, 2008; Nishiyama et al., 2011).为了维持D1蛋白的周转和缓解PSII光抑制, 植物进化出许多光保护机制, 如热耗散(Ruban, 2016)、水水循环(Miyake, 2010)及环式电子传递(薛娴等, 2017), 而这些光保护途径均存在于叶绿体内.在叶绿体外也存在光保护途径, 目前研究最多的是线粒体内的交替氧化酶(AOX)途径. ...
Mitochondrial alternative oxidase pathway protects plants against photoinhibition by alleviating inhibition of the repair of photodamaged PSII through preventing formation of reactive oxygen species in Rumex K-1 leaves 3 2011a
... 在植物线粒体中, 还原力NADH和FADH2分别通过NADH脱氢酶(复合体I)和琥珀酸还原酶(复合体II)进入呼吸电子传递链并将电子传递至泛醌(Selinski et al., 2018).泛醌的电子可以通过2条途径传递给氧气生成水(McDonald, 2008; Yoshida et al., 2011): (1) 电子流经复合体III和复合体IV最终通过细胞色素氧化酶(COX)传递给氧气, 期间偶联质子的跨膜运输形成跨膜质子梯度用于ATP的合成; (2) 电子通过AOX途径直接传递给氧气, 期间不偶联跨膜质子梯度和ATP的产生(Yoshida et al., 2011; Selinski et al., 2018).前人研究表明, AOX途径可以消耗强光下从叶绿体转运至线粒体的过剩还原力, 从而缓解PSII光抑制(Yoshida et al., 2006, 2007; Zhang et al., 2011a).AOX途径被抑制后, 苹果酸-草酰乙酸穿梭机制中的NADP-苹果酸还原酶的活性显著下降, 植物细胞内的活性氧含量增加, 光抑制明显加重(Zhang et al., 2011a).另有研究表明, 光呼吸过程中在线粒体内进行甘氨酸向丝氨酸的转化并产生大量还原力, AOX途径通过消耗这些还原力维持光呼吸运转, 避免有毒的光呼吸中间产物积累, 从而保护PSII (Watanabe et al., 2016; Zhang et al., 2017). ...
... ).AOX途径被抑制后, 苹果酸-草酰乙酸穿梭机制中的NADP-苹果酸还原酶的活性显著下降, 植物细胞内的活性氧含量增加, 光抑制明显加重(Zhang et al., 2011a).另有研究表明, 光呼吸过程中在线粒体内进行甘氨酸向丝氨酸的转化并产生大量还原力, AOX途径通过消耗这些还原力维持光呼吸运转, 避免有毒的光呼吸中间产物积累, 从而保护PSII (Watanabe et al., 2016; Zhang et al., 2017). ...
 ,*, 车兴凯, 梁英, 李月楠, 李滢, 赵世杰, 高辉远山东农业大学生命科学学院, 作物生物学国家重点实验室, 泰安 271018
,*, 车兴凯, 梁英, 李月楠, 李滢, 赵世杰, 高辉远山东农业大学生命科学学院, 作物生物学国家重点实验室, 泰安 271018
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT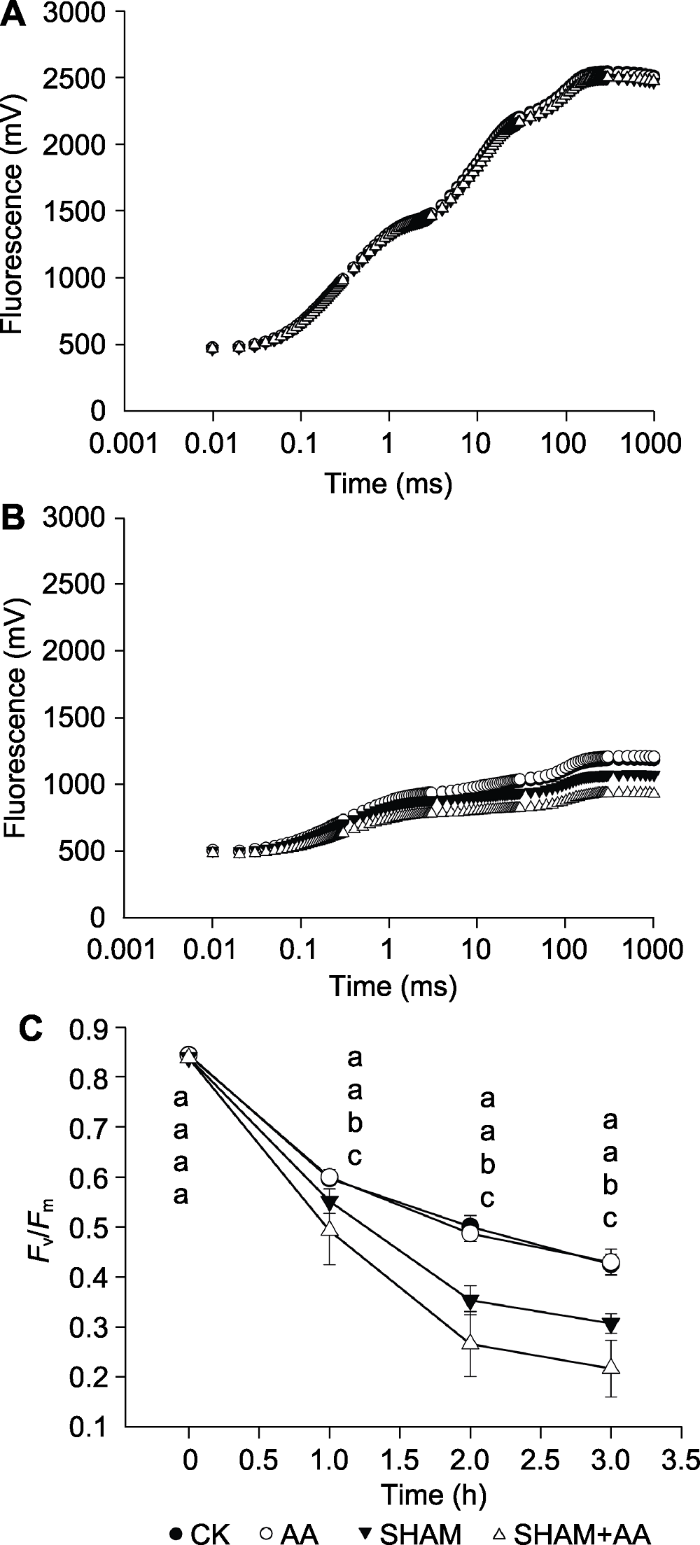 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT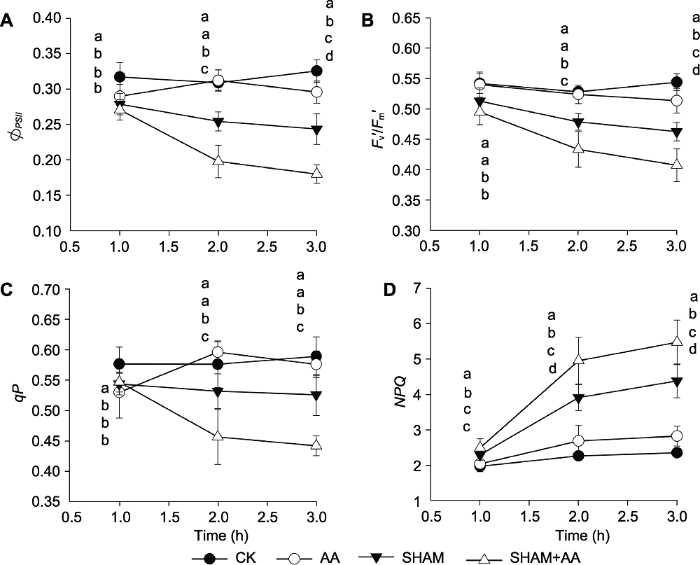 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}