Abstract Cysteine desulphydrase (CDes) can degrade cysteine (Cys) to form hydrogen sulfide (H2S). In this study, L-cysteine desulphydrase TaLCD from wheat (Triticum aestivum) was cloned and overexpressed in Arabidopsis thaliana. The effects of TaLCD on seed germination and root growth under osmotic stress in TaLCD overexpressing plants were examined, and the response to drought stress was also investigated. The results showed that, the seed germination rate of TaLCD overexpressing plants under salt treatment was remarkably higher than the wild type (WT), and the root length of TaLCD overexpressing plants was obviously longer when exposed to mannitol. Moreover, overexpression of TaLCD distinctly increased plant drought resistance. In addition, TaLCD overexpressing plants were more sensitive to ABA, with decreased seed germination rate and root length under ABA treatment. Furthermore, the expression of stress-responsive genes (COR47, RD29A, RAB18 and RD22) and ABA signaling pathway-related genes (NCED3, HAB1, HAB2, ABI1, ABI2 and ABF2) was significantly higher in TaLCD overexpressing plants than that in WT under drought stress. These results suggest that TaLCD enhances the drought and salt tolerance of plants possibly by ABA signaling pathway. Keywords:TaLCD;osmotic stress;ABA;germination and growth
PDF (20596KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 张扬, 刘华杰, 薛瑞丽, 李海霞, 李华. 小麦TaLCD基因的克隆及其对渗透胁迫的调节作用. 植物学报, 2020, 55(2): 137-146 doi:10.11983/CBB19109 Zhang Yang, Liu Huajie, Xue Ruili, Li Haixia, Li Hua. Cloning of Wheat TaLCD Gene and Its Regulation on Osmotic Stress. Chinese Bulletin of Botany, 2020, 55(2): 137-146 doi:10.11983/CBB19109
植物体内源H2S主要由半胱氨酸脱巯基酶(cysteine desulphydrase, CDes)催化降解半胱氨酸(Cys)生成。CDes在植物中主要有2种类型: 以L-Cys为底物的L-半胱氨酸脱巯基酶(LCD)和以D-Cys为底物的D-半胱氨酸脱巯基酶(DCD)。LCD和DCD均能催化降解半胱氨酸产生H2S, DCD主要定位在细胞质, LCD存在于叶绿体和线粒体中(Guo et al., 2016)。CDes对植物生长发育及非生物胁迫响应具有重要调节作用。例如, 在大肠杆菌(Escherichia coli)BL21中过表达拟南芥D/LCDes基因, 能增强BL21对Cd的耐受性, 减少H2O2和丙二醛(MDA)的产生(Shen et al., 2012)。拟南芥双突变体lcd/dcd比野生型对干旱更敏感(Jin et al., 2013; Shi et al., 2015), 而AtD/LCDes过表达植株具有较强的抗旱性(Shi et al., 2015)。LCD基因诱导表达可增强拟南芥对Cr6+胁迫的抗性; 相反, lcd突变体对Cr6+更敏感(Fang et al., 2017)。
众多研究显示, H2S参与ABA信号转导途径。在蚕豆(Vicia faba)、拟南芥及洋凤仙(Impatiens walleriana)中的研究表明, H2S促进干旱胁迫下的气孔关闭过程可能通过调节ABC转运体参与依赖ABA的信号途径。在aba3和abi1突变体中, AtLCD基因表达水平降低, 内源H2S含量减少, 而ABA受体基因在atlcd突变体中上调表达, 在NaHS (H2S供体)处理后下调表达(Jin et al., 2013)。拟南芥突变体lcd气孔开度显著大于野生型, 对干旱胁迫更加敏感。而NaHS可促进aba3和abi1突变体的气孔关闭, ABA诱导的气孔关闭在lcd突变体中有所减弱(Jin et al., 2013)。H2S对小麦干旱胁迫的缓解作用也部分依赖于ABA信号途径, 干旱胁迫下, NaHS可调节ABA信号途径相关基因表达(Li et al., 2017), 诱导小麦叶片中ABA合成及活化相关基因表达, 根中ABA合成及代谢相关基因上调表达(Ma et al., 2016)。
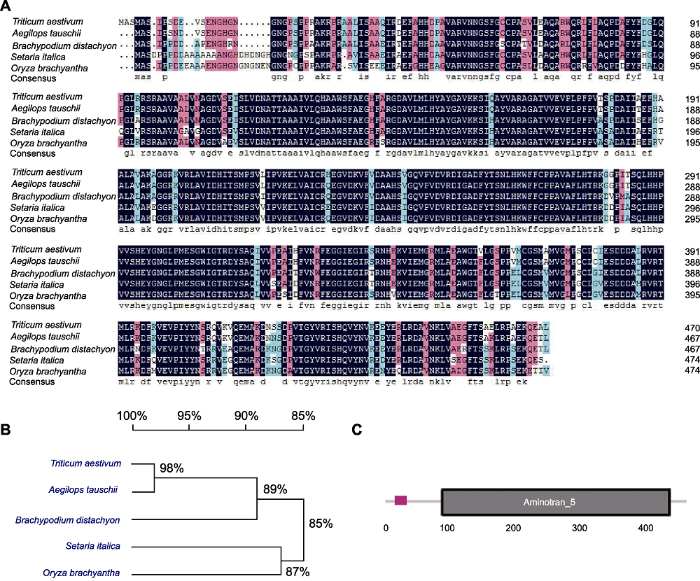
(A) Comparison of the derived amino acid sequences of TaLCD in Triticum aestivum with other 4 species (Aegilops tauschii, Brachypodium distachyon, Setaria italica, and Oryza brachyantha); (B) Phylogenetic analysis of LCD in five species; (C) Schematic structures of TaLCD, black box indicates Aminotran_5 domain.
(A) NaCl处理下TaLCD的表达; (B) PEG处理下TaLCD的表达; (C) NaCl处理下TaLCD酶活变化; (D) PEG处理下TaLCD酶活变化。* 表示差异显著(P<0.05)。 Figure 2The expression of TaLCD and enzyme activity of TaLCD in wheat under NaCl and PEG treatments
(A) The expression of TaLCD under NaCl treatment; (B) The expression of TaLCD under PEG treatment; (C) The enzyme activity of TaLCD under NaCl treatment; (D) The enzyme activity of TaLCD under PEG treatment. * indicate significant differences (P<0.05).
(A) 生长10天后的表型(Bars=1 cm); (B) 培养4天后的种子萌发率; (C) 生长10天后的根长表型(Bars=0.4 cm); (D) 生长10天后的根长。WT: 野生型; ABA: 脱落酸; * 表示差异显著(P<0.05)。 Figure 7Seed germination and growth of Arabidopsis lines expressing TaLCD under different concentrations of ABA treatment
(A) Phenotype on the 10th day (Bars=1 cm); (B) Seed germination rate after 4 d incubation; (C) Phenotype of root growth on the 10th day (Bars=0.4 cm); (D) Root length on the 10th day. WT: Wild type; ABA: Abscisic acid; * indicate significant differences (P<0.05).
(A)-(D) 胁迫响应基因; (E)-(J) ABA信号途径相关基因。WT: 野生型; OX-LCD-2: 转基因株系 Figure 8The expression of stress response genes and ABA signaling related genes in Arabidopsis lines expressing TaLCD under drought stress
(A)-(D) Stress response genes; (E)-(J) ABA signaling related genes. WT: Wild type; OX-LCD-2: Transgenic lines
ChristouA, ManganarisGA, PapadopoulosI, FotopoulosV ( 2013). Hydrogen sulfide induces systemic tolerance to salinity and non-ionic osmotic stress in strawberry plants through modification of reactive species biosynthesis and transcriptional regulation of multiple defence pathways J Exp Bot 64, 1953-1966. [本文引用: 1]
DooleyFD, NairSP, WardPD ( 2013). Increased growth and germination success in plants following hydrogen sulfide administration PLoS One 8, e62048. [本文引用: 1]
FangHH, LiuZQ, LongYP, LiangYL, JinZP, ZhangLP, LiuDM, LiH, ZhaiJX, PeiYX ( 2017). The Ca 2+/CaM2- binding transcription factor TGA3 elevates LCD expression and H2S production to bolster Cr 6+ tolerance in Arabidopsis Plant J 91, 1038-1050. [本文引用: 1]
García-MataC, LamattinaL ( 2010). Hydrogen sulphide, a novel gasotransmitter involved in guard cell signaling New Phytol 188, 977-984. [本文引用: 1]
GuoHM, XiaoTY, ZhouH, XieYJ, ShenWB ( 2016). Hydrogen sulfide: a versatile regulator of environmental stress in plants Acta Physiol Plant 38, 16. [本文引用: 1]
JiaHL, HuYF, FanTT, LiJS ( 2015). Hydrogen sulfide modulates actin-dependent auxin transport via regulating ABPs results in changing of root development in Arabidopsis Sci Rep 5, 8251. [本文引用: 1]
JinZP, XueSW, LuoYA, TianBH, FangHH, LiH, PeiYX ( 2013). Hydrogen sulfide interacting with abscisic acid in stomatal regulation responses to drought stress in Arabidopsis Plant Physiol Biochem 62, 41-46. [本文引用: 5]
LetunicI, DoerksT, BorkP ( 2015). SMART: recent updates, new developments and status in 2015 Nucleic Acids Res 43, D257-D260. [本文引用: 1]
LiCJ, LiuZJ, ZhangQR, WangRZ, XiaoLT, MaH, ChongK, XuYY ( 2012). SKP1 is involved in abscisic acid signaling to regulate seed germination, stomatal opening and root growth in Arabidopsis thaliana Plant Cell Environ 35, 952-965. [本文引用: 1]
LiH, GaoMQ, XueRL, WangD, ZhaoHJ ( 2015). Effect of hydrogen sulfide on D1 protein in wheat under drought stress Acta Physiol Plant 37, 225. [本文引用: 4]
LiH, LiM, WeiXL, ZhangX, XueRL, ZhaoYD, ZhaoHJ ( 2017). Transcriptome analysis of drought-responsive genes regulated by hydrogen sulfide in wheat ( Triticum aestivum L.) leaves Mol Genet Genomics 292, 1091-1110. [本文引用: 3]
MaDY, DingHN, WangCY, QinHX, HanQX, HouJF, LuHF, XieYX, GuoTC ( 2016). Alleviation of drought stress by hydrogen sulfide is partially related to the abscisic acid signaling pathway in wheat PLoS One 11, e0163082. [本文引用: 2]
PapenbrockJ, RiemenschneiderA, KampA, Schulz- VogtHN, SchmidtA ( 2007). Characterization of cysteine-degrading and H2S-releasing enzymes of higher plants—from the field to the test tube and back Plant Biol 9, 582-588. [本文引用: 1]
ShenJJ, QiaoZJ, XingTJ, ZhangLP, LiangYL, JinZP, YangGD, PeiYX ( 2012). Cadmium toxicity is alleviated by AtLCD and AtDCD in Escherichia coli J Appl Microbiol 113, 1130-1138. [本文引用: 1]
... 人类认识和研究硫化氢(H2S)已有300多年的历史.长期以来, 高浓度H2S常因抑制线粒体细胞色素氧化酶C的活性被认为是毒性物质.但近年来的研究显示, H2S同一氧化氮(NO)和一氧化碳(CO)一样可作为信号分子调节植物的生长发育.例如, 外源H2S可促进气孔关闭(García-Mata and Lamattina, 2010; 侯智慧等, 2011); 提高大豆(Glycine max)、玉米(Zea mays)、小麦(Triticum aestivum)和豌豆(Pisum sativum)的发芽率与生长速率(Dooley et al., 2013); 调节根系发育(Jia et al., 2015); 介导植物衰老(Zhang et al., 2011).此外, H2S还能响应各种生物和非生物胁迫.例如, NaHS (H2S供体)可缓解铜、铝和铬等重金属胁迫对小麦种子萌发和幼苗生长的抑制效应(Zhang et al., 2008, 2010a, 2010b); 缓解大麦(Hordeum vulgare)铝中毒现象(Chen et al., 2013); 增强烟草(Nicotiana tabacum)悬浮细胞在高温胁迫下的存活率及生长能力(Li et al., 2012); 提高苜蓿(Medicago sativa)种子萌发时的耐盐性(Wang et al., 2012); 增强狗牙根(Cynodon dactylon)对盐胁迫、渗透胁迫及低温的抗性(Shi et al., 2013); 缓解小麦干旱胁迫(Li et al., 2015). ...
H2S介导ABA诱导蚕豆气孔运动的生理机制研究 1 2011
... ABA可调节种子萌发和根系生长, 并响应多种逆境胁迫.大量研究显示, H2S介导ABA信号途径.H2S的合成抑制剂可显著抑制ABA诱导的蚕豆气孔关闭(刘菁等, 2011).NaHS处理可以减小aba3和abi1突变体的气孔开度, 而ABA对lcd突变体气孔关闭的作用减弱(Jin et al., 2013).NaHS预处理可调节小麦ABA信号途径中相关基因的表达(Li et al., 2017).ABA受体相关基因在小麦叶片和根中均受H2S诱导上调表达(Ma et al., 2016).本研究显示, TaLCD过表达增强了对ABA的敏感性, ABA处理下, TaLCD过表达植株的种子萌发率及根长均显著低于野生型(图7).由此表明, TaLCD参与调控ABA信号途径.ABA被公认为一种胁迫激素, 在植物干旱及盐胁迫中发挥重要作用.而TaLCD过表达可显著提高植株的抗旱及抗盐性, 且TaLCD过表达对ABA更敏感.干旱胁迫下, 胁迫响应基因及ABA信号途径相关基因在TaLCD过表达植株中都显著高于野生型(图8).由此推测, TaLCD可能通过介导ABA信号途径调节植株的抗旱和抗盐性, 但具体机制有待深入研究. ...
外源硫化氢对水分胁迫下玉米种子萌发和生长的影响 1 2011
... H2S作为气体信号分子, 在植物响应非生物胁迫中发挥重要调控作用.研究显示, NaHS (H2S供体)可促进盐胁迫下苜蓿种子的萌发(Wang et al., 2012), 促进水分胁迫下玉米种子的萌发和生长(单长卷和周岩, 2011), 缓解干旱对小麦的氧化伤害(Li et al., 2015), 提高草莓(Fragaria ananassa)和狗牙根对渗透胁迫的耐受性(Christou et al., 2013; Shi et al., 2013).此外, 拟南芥中内源H2S合成基因(AtLCD和AtDCD)的表达受干旱胁迫诱导, 其表达模式与干旱相关的转录因子(如DREB2A、DREB2B、CBF4和RD29A)非常相似(Jin et al., 2011), 且AtLCD和AtDCD过表达植株对干旱和盐胁迫的抗性均显著高于野生型(Shi et al., 2015).由此表明, H2S可显著调节植物的渗透胁迫. ...
Hydrogen sulfide alleviates aluminum toxicity in barley seedlings 1 2013
... 人类认识和研究硫化氢(H2S)已有300多年的历史.长期以来, 高浓度H2S常因抑制线粒体细胞色素氧化酶C的活性被认为是毒性物质.但近年来的研究显示, H2S同一氧化氮(NO)和一氧化碳(CO)一样可作为信号分子调节植物的生长发育.例如, 外源H2S可促进气孔关闭(García-Mata and Lamattina, 2010; 侯智慧等, 2011); 提高大豆(Glycine max)、玉米(Zea mays)、小麦(Triticum aestivum)和豌豆(Pisum sativum)的发芽率与生长速率(Dooley et al., 2013); 调节根系发育(Jia et al., 2015); 介导植物衰老(Zhang et al., 2011).此外, H2S还能响应各种生物和非生物胁迫.例如, NaHS (H2S供体)可缓解铜、铝和铬等重金属胁迫对小麦种子萌发和幼苗生长的抑制效应(Zhang et al., 2008, 2010a, 2010b); 缓解大麦(Hordeum vulgare)铝中毒现象(Chen et al., 2013); 增强烟草(Nicotiana tabacum)悬浮细胞在高温胁迫下的存活率及生长能力(Li et al., 2012); 提高苜蓿(Medicago sativa)种子萌发时的耐盐性(Wang et al., 2012); 增强狗牙根(Cynodon dactylon)对盐胁迫、渗透胁迫及低温的抗性(Shi et al., 2013); 缓解小麦干旱胁迫(Li et al., 2015). ...
Hydrogen sulfide induces systemic tolerance to salinity and non-ionic osmotic stress in strawberry plants through modification of reactive species biosynthesis and transcriptional regulation of multiple defence pathways 1 2013
... H2S作为气体信号分子, 在植物响应非生物胁迫中发挥重要调控作用.研究显示, NaHS (H2S供体)可促进盐胁迫下苜蓿种子的萌发(Wang et al., 2012), 促进水分胁迫下玉米种子的萌发和生长(单长卷和周岩, 2011), 缓解干旱对小麦的氧化伤害(Li et al., 2015), 提高草莓(Fragaria ananassa)和狗牙根对渗透胁迫的耐受性(Christou et al., 2013; Shi et al., 2013).此外, 拟南芥中内源H2S合成基因(AtLCD和AtDCD)的表达受干旱胁迫诱导, 其表达模式与干旱相关的转录因子(如DREB2A、DREB2B、CBF4和RD29A)非常相似(Jin et al., 2011), 且AtLCD和AtDCD过表达植株对干旱和盐胁迫的抗性均显著高于野生型(Shi et al., 2015).由此表明, H2S可显著调节植物的渗透胁迫. ...
Increased growth and germination success in plants following hydrogen sulfide administration 1 2013
... 人类认识和研究硫化氢(H2S)已有300多年的历史.长期以来, 高浓度H2S常因抑制线粒体细胞色素氧化酶C的活性被认为是毒性物质.但近年来的研究显示, H2S同一氧化氮(NO)和一氧化碳(CO)一样可作为信号分子调节植物的生长发育.例如, 外源H2S可促进气孔关闭(García-Mata and Lamattina, 2010; 侯智慧等, 2011); 提高大豆(Glycine max)、玉米(Zea mays)、小麦(Triticum aestivum)和豌豆(Pisum sativum)的发芽率与生长速率(Dooley et al., 2013); 调节根系发育(Jia et al., 2015); 介导植物衰老(Zhang et al., 2011).此外, H2S还能响应各种生物和非生物胁迫.例如, NaHS (H2S供体)可缓解铜、铝和铬等重金属胁迫对小麦种子萌发和幼苗生长的抑制效应(Zhang et al., 2008, 2010a, 2010b); 缓解大麦(Hordeum vulgare)铝中毒现象(Chen et al., 2013); 增强烟草(Nicotiana tabacum)悬浮细胞在高温胁迫下的存活率及生长能力(Li et al., 2012); 提高苜蓿(Medicago sativa)种子萌发时的耐盐性(Wang et al., 2012); 增强狗牙根(Cynodon dactylon)对盐胁迫、渗透胁迫及低温的抗性(Shi et al., 2013); 缓解小麦干旱胁迫(Li et al., 2015). ...
The Ca 2+/CaM2- binding transcription factor TGA3 elevates LCD expression and H2S production to bolster Cr 6+ tolerance in Arabidopsis 1 2017
... 植物体内源H2S主要由半胱氨酸脱巯基酶(cysteine desulphydrase, CDes)催化降解半胱氨酸(Cys)生成.CDes在植物中主要有2种类型: 以L-Cys为底物的L-半胱氨酸脱巯基酶(LCD)和以D-Cys为底物的D-半胱氨酸脱巯基酶(DCD).LCD和DCD均能催化降解半胱氨酸产生H2S, DCD主要定位在细胞质, LCD存在于叶绿体和线粒体中(Guo et al., 2016).CDes对植物生长发育及非生物胁迫响应具有重要调节作用.例如, 在大肠杆菌(Escherichia coli)BL21中过表达拟南芥D/LCDes基因, 能增强BL21对Cd的耐受性, 减少H2O2和丙二醛(MDA)的产生(Shen et al., 2012).拟南芥双突变体lcd/dcd比野生型对干旱更敏感(Jin et al., 2013; Shi et al., 2015), 而AtD/LCDes过表达植株具有较强的抗旱性(Shi et al., 2015).LCD基因诱导表达可增强拟南芥对Cr6+胁迫的抗性; 相反, lcd突变体对Cr6+更敏感(Fang et al., 2017). ...
Hydrogen sulphide, a novel gasotransmitter involved in guard cell signaling 1 2010
... 人类认识和研究硫化氢(H2S)已有300多年的历史.长期以来, 高浓度H2S常因抑制线粒体细胞色素氧化酶C的活性被认为是毒性物质.但近年来的研究显示, H2S同一氧化氮(NO)和一氧化碳(CO)一样可作为信号分子调节植物的生长发育.例如, 外源H2S可促进气孔关闭(García-Mata and Lamattina, 2010; 侯智慧等, 2011); 提高大豆(Glycine max)、玉米(Zea mays)、小麦(Triticum aestivum)和豌豆(Pisum sativum)的发芽率与生长速率(Dooley et al., 2013); 调节根系发育(Jia et al., 2015); 介导植物衰老(Zhang et al., 2011).此外, H2S还能响应各种生物和非生物胁迫.例如, NaHS (H2S供体)可缓解铜、铝和铬等重金属胁迫对小麦种子萌发和幼苗生长的抑制效应(Zhang et al., 2008, 2010a, 2010b); 缓解大麦(Hordeum vulgare)铝中毒现象(Chen et al., 2013); 增强烟草(Nicotiana tabacum)悬浮细胞在高温胁迫下的存活率及生长能力(Li et al., 2012); 提高苜蓿(Medicago sativa)种子萌发时的耐盐性(Wang et al., 2012); 增强狗牙根(Cynodon dactylon)对盐胁迫、渗透胁迫及低温的抗性(Shi et al., 2013); 缓解小麦干旱胁迫(Li et al., 2015). ...
Hydrogen sulfide: a versatile regulator of environmental stress in plants 1 2016
... 植物体内源H2S主要由半胱氨酸脱巯基酶(cysteine desulphydrase, CDes)催化降解半胱氨酸(Cys)生成.CDes在植物中主要有2种类型: 以L-Cys为底物的L-半胱氨酸脱巯基酶(LCD)和以D-Cys为底物的D-半胱氨酸脱巯基酶(DCD).LCD和DCD均能催化降解半胱氨酸产生H2S, DCD主要定位在细胞质, LCD存在于叶绿体和线粒体中(Guo et al., 2016).CDes对植物生长发育及非生物胁迫响应具有重要调节作用.例如, 在大肠杆菌(Escherichia coli)BL21中过表达拟南芥D/LCDes基因, 能增强BL21对Cd的耐受性, 减少H2O2和丙二醛(MDA)的产生(Shen et al., 2012).拟南芥双突变体lcd/dcd比野生型对干旱更敏感(Jin et al., 2013; Shi et al., 2015), 而AtD/LCDes过表达植株具有较强的抗旱性(Shi et al., 2015).LCD基因诱导表达可增强拟南芥对Cr6+胁迫的抗性; 相反, lcd突变体对Cr6+更敏感(Fang et al., 2017). ...
Hydrogen sulfide modulates actin-dependent auxin transport via regulating ABPs results in changing of root development in Arabidopsis 1 2015
... 人类认识和研究硫化氢(H2S)已有300多年的历史.长期以来, 高浓度H2S常因抑制线粒体细胞色素氧化酶C的活性被认为是毒性物质.但近年来的研究显示, H2S同一氧化氮(NO)和一氧化碳(CO)一样可作为信号分子调节植物的生长发育.例如, 外源H2S可促进气孔关闭(García-Mata and Lamattina, 2010; 侯智慧等, 2011); 提高大豆(Glycine max)、玉米(Zea mays)、小麦(Triticum aestivum)和豌豆(Pisum sativum)的发芽率与生长速率(Dooley et al., 2013); 调节根系发育(Jia et al., 2015); 介导植物衰老(Zhang et al., 2011).此外, H2S还能响应各种生物和非生物胁迫.例如, NaHS (H2S供体)可缓解铜、铝和铬等重金属胁迫对小麦种子萌发和幼苗生长的抑制效应(Zhang et al., 2008, 2010a, 2010b); 缓解大麦(Hordeum vulgare)铝中毒现象(Chen et al., 2013); 增强烟草(Nicotiana tabacum)悬浮细胞在高温胁迫下的存活率及生长能力(Li et al., 2012); 提高苜蓿(Medicago sativa)种子萌发时的耐盐性(Wang et al., 2012); 增强狗牙根(Cynodon dactylon)对盐胁迫、渗透胁迫及低温的抗性(Shi et al., 2013); 缓解小麦干旱胁迫(Li et al., 2015). ...
Hydrogen sulfide improves drought resistance in Arabidopsis thaliana 1 2011
... H2S作为气体信号分子, 在植物响应非生物胁迫中发挥重要调控作用.研究显示, NaHS (H2S供体)可促进盐胁迫下苜蓿种子的萌发(Wang et al., 2012), 促进水分胁迫下玉米种子的萌发和生长(单长卷和周岩, 2011), 缓解干旱对小麦的氧化伤害(Li et al., 2015), 提高草莓(Fragaria ananassa)和狗牙根对渗透胁迫的耐受性(Christou et al., 2013; Shi et al., 2013).此外, 拟南芥中内源H2S合成基因(AtLCD和AtDCD)的表达受干旱胁迫诱导, 其表达模式与干旱相关的转录因子(如DREB2A、DREB2B、CBF4和RD29A)非常相似(Jin et al., 2011), 且AtLCD和AtDCD过表达植株对干旱和盐胁迫的抗性均显著高于野生型(Shi et al., 2015).由此表明, H2S可显著调节植物的渗透胁迫. ...
Hydrogen sulfide interacting with abscisic acid in stomatal regulation responses to drought stress in Arabidopsis 5 2013
... 植物体内源H2S主要由半胱氨酸脱巯基酶(cysteine desulphydrase, CDes)催化降解半胱氨酸(Cys)生成.CDes在植物中主要有2种类型: 以L-Cys为底物的L-半胱氨酸脱巯基酶(LCD)和以D-Cys为底物的D-半胱氨酸脱巯基酶(DCD).LCD和DCD均能催化降解半胱氨酸产生H2S, DCD主要定位在细胞质, LCD存在于叶绿体和线粒体中(Guo et al., 2016).CDes对植物生长发育及非生物胁迫响应具有重要调节作用.例如, 在大肠杆菌(Escherichia coli)BL21中过表达拟南芥D/LCDes基因, 能增强BL21对Cd的耐受性, 减少H2O2和丙二醛(MDA)的产生(Shen et al., 2012).拟南芥双突变体lcd/dcd比野生型对干旱更敏感(Jin et al., 2013; Shi et al., 2015), 而AtD/LCDes过表达植株具有较强的抗旱性(Shi et al., 2015).LCD基因诱导表达可增强拟南芥对Cr6+胁迫的抗性; 相反, lcd突变体对Cr6+更敏感(Fang et al., 2017). ...
... 众多研究显示, H2S参与ABA信号转导途径.在蚕豆(Vicia faba)、拟南芥及洋凤仙(Impatiens walleriana)中的研究表明, H2S促进干旱胁迫下的气孔关闭过程可能通过调节ABC转运体参与依赖ABA的信号途径.在aba3和abi1突变体中, AtLCD基因表达水平降低, 内源H2S含量减少, 而ABA受体基因在atlcd突变体中上调表达, 在NaHS (H2S供体)处理后下调表达(Jin et al., 2013).拟南芥突变体lcd气孔开度显著大于野生型, 对干旱胁迫更加敏感.而NaHS可促进aba3和abi1突变体的气孔关闭, ABA诱导的气孔关闭在lcd突变体中有所减弱(Jin et al., 2013).H2S对小麦干旱胁迫的缓解作用也部分依赖于ABA信号途径, 干旱胁迫下, NaHS可调节ABA信号途径相关基因表达(Li et al., 2017), 诱导小麦叶片中ABA合成及活化相关基因表达, 根中ABA合成及代谢相关基因上调表达(Ma et al., 2016). ...
... 突变体中有所减弱(Jin et al., 2013).H2S对小麦干旱胁迫的缓解作用也部分依赖于ABA信号途径, 干旱胁迫下, NaHS可调节ABA信号途径相关基因表达(Li et al., 2017), 诱导小麦叶片中ABA合成及活化相关基因表达, 根中ABA合成及代谢相关基因上调表达(Ma et al., 2016). ...
... H2S作为气体信号分子, 在植物响应非生物胁迫中发挥重要调控作用.研究显示, NaHS (H2S供体)可促进盐胁迫下苜蓿种子的萌发(Wang et al., 2012), 促进水分胁迫下玉米种子的萌发和生长(单长卷和周岩, 2011), 缓解干旱对小麦的氧化伤害(Li et al., 2015), 提高草莓(Fragaria ananassa)和狗牙根对渗透胁迫的耐受性(Christou et al., 2013; Shi et al., 2013).此外, 拟南芥中内源H2S合成基因(AtLCD和AtDCD)的表达受干旱胁迫诱导, 其表达模式与干旱相关的转录因子(如DREB2A、DREB2B、CBF4和RD29A)非常相似(Jin et al., 2011), 且AtLCD和AtDCD过表达植株对干旱和盐胁迫的抗性均显著高于野生型(Shi et al., 2015).由此表明, H2S可显著调节植物的渗透胁迫. ...
Cadmium toxicity is alleviated by AtLCD and AtDCD in Escherichia coli 1 2012
... 植物体内源H2S主要由半胱氨酸脱巯基酶(cysteine desulphydrase, CDes)催化降解半胱氨酸(Cys)生成.CDes在植物中主要有2种类型: 以L-Cys为底物的L-半胱氨酸脱巯基酶(LCD)和以D-Cys为底物的D-半胱氨酸脱巯基酶(DCD).LCD和DCD均能催化降解半胱氨酸产生H2S, DCD主要定位在细胞质, LCD存在于叶绿体和线粒体中(Guo et al., 2016).CDes对植物生长发育及非生物胁迫响应具有重要调节作用.例如, 在大肠杆菌(Escherichia coli)BL21中过表达拟南芥D/LCDes基因, 能增强BL21对Cd的耐受性, 减少H2O2和丙二醛(MDA)的产生(Shen et al., 2012).拟南芥双突变体lcd/dcd比野生型对干旱更敏感(Jin et al., 2013; Shi et al., 2015), 而AtD/LCDes过表达植株具有较强的抗旱性(Shi et al., 2015).LCD基因诱导表达可增强拟南芥对Cr6+胁迫的抗性; 相反, lcd突变体对Cr6+更敏感(Fang et al., 2017). ...
Exogenous application of hydrogen sulfide donor sodium hydrosulfide enhanced multiple abiotic stress tolerance in bermudagrass (Cynodon dactylon(L). Pers.) 2 2013
... 人类认识和研究硫化氢(H2S)已有300多年的历史.长期以来, 高浓度H2S常因抑制线粒体细胞色素氧化酶C的活性被认为是毒性物质.但近年来的研究显示, H2S同一氧化氮(NO)和一氧化碳(CO)一样可作为信号分子调节植物的生长发育.例如, 外源H2S可促进气孔关闭(García-Mata and Lamattina, 2010; 侯智慧等, 2011); 提高大豆(Glycine max)、玉米(Zea mays)、小麦(Triticum aestivum)和豌豆(Pisum sativum)的发芽率与生长速率(Dooley et al., 2013); 调节根系发育(Jia et al., 2015); 介导植物衰老(Zhang et al., 2011).此外, H2S还能响应各种生物和非生物胁迫.例如, NaHS (H2S供体)可缓解铜、铝和铬等重金属胁迫对小麦种子萌发和幼苗生长的抑制效应(Zhang et al., 2008, 2010a, 2010b); 缓解大麦(Hordeum vulgare)铝中毒现象(Chen et al., 2013); 增强烟草(Nicotiana tabacum)悬浮细胞在高温胁迫下的存活率及生长能力(Li et al., 2012); 提高苜蓿(Medicago sativa)种子萌发时的耐盐性(Wang et al., 2012); 增强狗牙根(Cynodon dactylon)对盐胁迫、渗透胁迫及低温的抗性(Shi et al., 2013); 缓解小麦干旱胁迫(Li et al., 2015). ...
... H2S作为气体信号分子, 在植物响应非生物胁迫中发挥重要调控作用.研究显示, NaHS (H2S供体)可促进盐胁迫下苜蓿种子的萌发(Wang et al., 2012), 促进水分胁迫下玉米种子的萌发和生长(单长卷和周岩, 2011), 缓解干旱对小麦的氧化伤害(Li et al., 2015), 提高草莓(Fragaria ananassa)和狗牙根对渗透胁迫的耐受性(Christou et al., 2013; Shi et al., 2013).此外, 拟南芥中内源H2S合成基因(AtLCD和AtDCD)的表达受干旱胁迫诱导, 其表达模式与干旱相关的转录因子(如DREB2A、DREB2B、CBF4和RD29A)非常相似(Jin et al., 2011), 且AtLCD和AtDCD过表达植株对干旱和盐胁迫的抗性均显著高于野生型(Shi et al., 2015).由此表明, H2S可显著调节植物的渗透胁迫. ...
Hydrogen sulfide regulates abiotic stress tolerance and biotic stress resistance in Arabidopsis 3 2015
... 植物体内源H2S主要由半胱氨酸脱巯基酶(cysteine desulphydrase, CDes)催化降解半胱氨酸(Cys)生成.CDes在植物中主要有2种类型: 以L-Cys为底物的L-半胱氨酸脱巯基酶(LCD)和以D-Cys为底物的D-半胱氨酸脱巯基酶(DCD).LCD和DCD均能催化降解半胱氨酸产生H2S, DCD主要定位在细胞质, LCD存在于叶绿体和线粒体中(Guo et al., 2016).CDes对植物生长发育及非生物胁迫响应具有重要调节作用.例如, 在大肠杆菌(Escherichia coli)BL21中过表达拟南芥D/LCDes基因, 能增强BL21对Cd的耐受性, 减少H2O2和丙二醛(MDA)的产生(Shen et al., 2012).拟南芥双突变体lcd/dcd比野生型对干旱更敏感(Jin et al., 2013; Shi et al., 2015), 而AtD/LCDes过表达植株具有较强的抗旱性(Shi et al., 2015).LCD基因诱导表达可增强拟南芥对Cr6+胁迫的抗性; 相反, lcd突变体对Cr6+更敏感(Fang et al., 2017). ...
... 过表达植株具有较强的抗旱性(Shi et al., 2015).LCD基因诱导表达可增强拟南芥对Cr6+胁迫的抗性; 相反, lcd突变体对Cr6+更敏感(Fang et al., 2017). ...
... H2S作为气体信号分子, 在植物响应非生物胁迫中发挥重要调控作用.研究显示, NaHS (H2S供体)可促进盐胁迫下苜蓿种子的萌发(Wang et al., 2012), 促进水分胁迫下玉米种子的萌发和生长(单长卷和周岩, 2011), 缓解干旱对小麦的氧化伤害(Li et al., 2015), 提高草莓(Fragaria ananassa)和狗牙根对渗透胁迫的耐受性(Christou et al., 2013; Shi et al., 2013).此外, 拟南芥中内源H2S合成基因(AtLCD和AtDCD)的表达受干旱胁迫诱导, 其表达模式与干旱相关的转录因子(如DREB2A、DREB2B、CBF4和RD29A)非常相似(Jin et al., 2011), 且AtLCD和AtDCD过表达植株对干旱和盐胁迫的抗性均显著高于野生型(Shi et al., 2015).由此表明, H2S可显著调节植物的渗透胁迫. ...
MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods 1 2011
Hydrogen sulfide enhances alfalfa ( Medicago sativa) tolerance against salinity during seed germination by nitric oxide pathway 2 2012
... 人类认识和研究硫化氢(H2S)已有300多年的历史.长期以来, 高浓度H2S常因抑制线粒体细胞色素氧化酶C的活性被认为是毒性物质.但近年来的研究显示, H2S同一氧化氮(NO)和一氧化碳(CO)一样可作为信号分子调节植物的生长发育.例如, 外源H2S可促进气孔关闭(García-Mata and Lamattina, 2010; 侯智慧等, 2011); 提高大豆(Glycine max)、玉米(Zea mays)、小麦(Triticum aestivum)和豌豆(Pisum sativum)的发芽率与生长速率(Dooley et al., 2013); 调节根系发育(Jia et al., 2015); 介导植物衰老(Zhang et al., 2011).此外, H2S还能响应各种生物和非生物胁迫.例如, NaHS (H2S供体)可缓解铜、铝和铬等重金属胁迫对小麦种子萌发和幼苗生长的抑制效应(Zhang et al., 2008, 2010a, 2010b); 缓解大麦(Hordeum vulgare)铝中毒现象(Chen et al., 2013); 增强烟草(Nicotiana tabacum)悬浮细胞在高温胁迫下的存活率及生长能力(Li et al., 2012); 提高苜蓿(Medicago sativa)种子萌发时的耐盐性(Wang et al., 2012); 增强狗牙根(Cynodon dactylon)对盐胁迫、渗透胁迫及低温的抗性(Shi et al., 2013); 缓解小麦干旱胁迫(Li et al., 2015). ...
... H2S作为气体信号分子, 在植物响应非生物胁迫中发挥重要调控作用.研究显示, NaHS (H2S供体)可促进盐胁迫下苜蓿种子的萌发(Wang et al., 2012), 促进水分胁迫下玉米种子的萌发和生长(单长卷和周岩, 2011), 缓解干旱对小麦的氧化伤害(Li et al., 2015), 提高草莓(Fragaria ananassa)和狗牙根对渗透胁迫的耐受性(Christou et al., 2013; Shi et al., 2013).此外, 拟南芥中内源H2S合成基因(AtLCD和AtDCD)的表达受干旱胁迫诱导, 其表达模式与干旱相关的转录因子(如DREB2A、DREB2B、CBF4和RD29A)非常相似(Jin et al., 2011), 且AtLCD和AtDCD过表达植株对干旱和盐胁迫的抗性均显著高于野生型(Shi et al., 2015).由此表明, H2S可显著调节植物的渗透胁迫. ...
Molecular cloning, characterization, and expression analysis of a novel gene encoding L-cysteine desulfhydrase from Brassica napus 1 2013
 ,*河南农业大学生命科学学院, 郑州 450002
,*河南农业大学生命科学学院, 郑州 450002
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT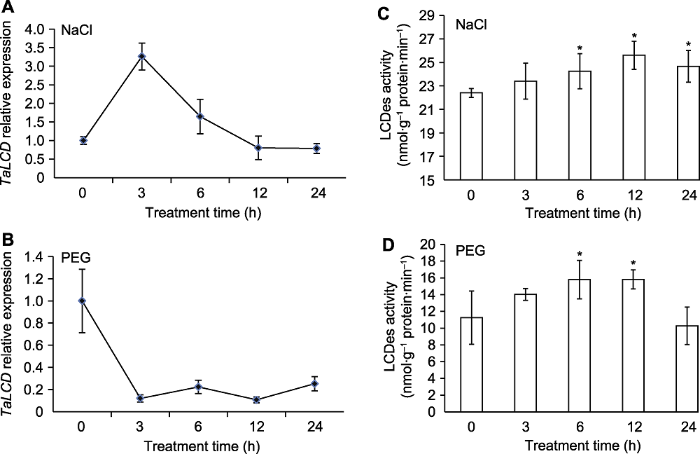 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT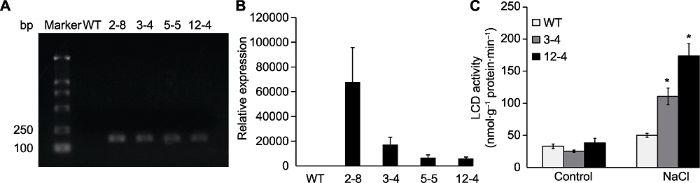 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT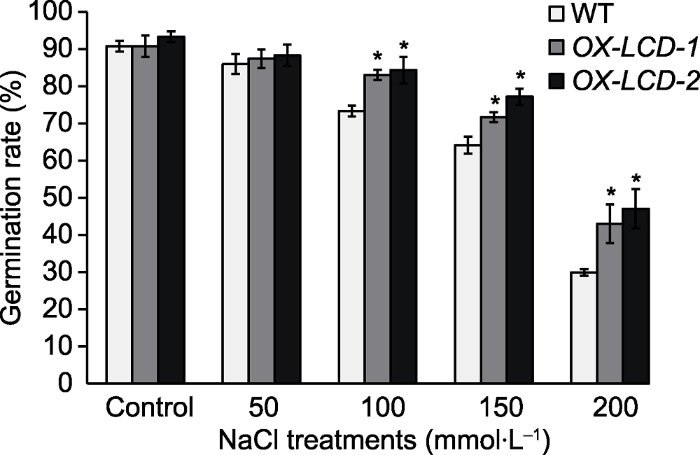 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT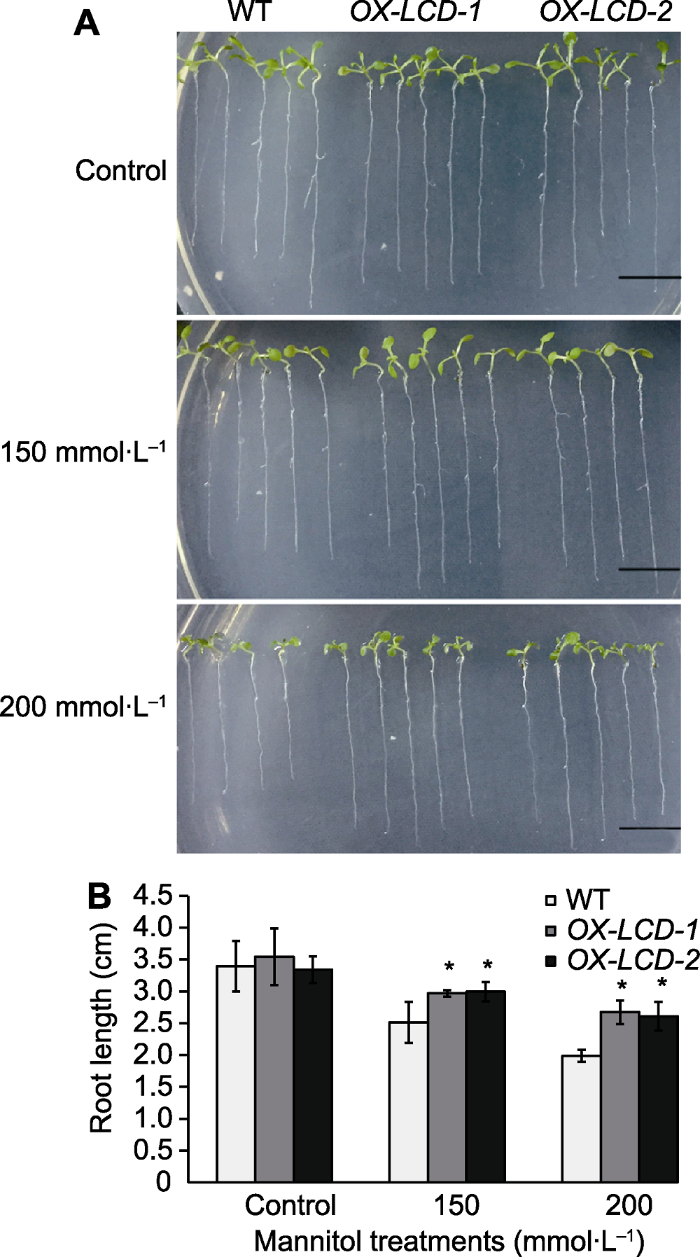 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT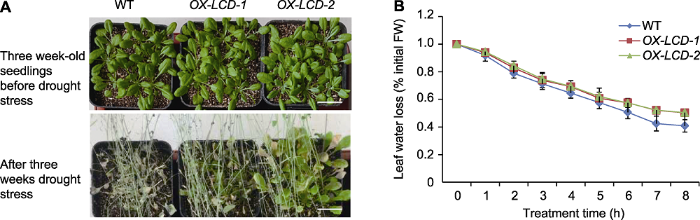 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT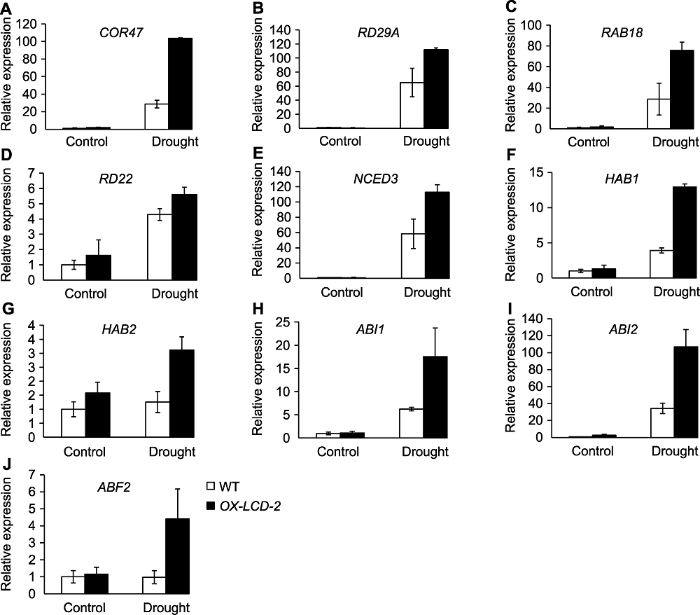 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}