Nitrogen Utilization Mechanism in C3 and C4 Plants
Lu Zhang1,2, Xinhua He,2,3,*1Maize Research Institute of Shanxi Academy of Agricultural Sciences, Xinzhou 034000, China 2College of Resources and Environment, Southwest University, Chongqing 400716, China 3School of Plant Biology, University of Western Australia, Perth 6009, Australia
Abstract Improving nitrogen use efficiency (NUE) of plants is not only an important approach to ensure global food security, but also to achieve sustainable agricultural development. In the past half century, great progress has been made in the study of nitrogen utilization mechanism, but the regulatory mechanism of NUE is not clear and the improvement of NUE is still extremely limited. Photosynthetic carbon assimilation and nitrogen assimilation are integrated in a plant, and only by coordinating carbon and nitrogen metabolism can the carbon/nitrogen balance be maintained and the normal growth and development of plants be regulated. Due to the differences in the photosynthetic nitrogen utilization rate (PNUE) between C3 and C4 plants, there will also be differences in the utilization efficiency of nitrogen. In order to improve crop NUE, it is necessary to understand the functions and regulatory mechanisms of key factors involved in nitrogen absorption, transport, assimilation and signal transduction of C3 and C4 plants. In addition, studies on carbon and nitrogen assimilation and their mechanisms under the conditions of increasing atmospheric CO2 concentration and global warming should not be ignored. This paper reviews the differences of key factors on nitrogen use between C3 and C4 plants and the regulatory mechanisms, and proposes possible ways to improve NUE of C3 crops by using genetic approaches. Keywords:photosynthesis;nitrogen metabolism;nitrogen use efficiency;regulatory mechanisms;genetic improvement
PDF (990KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 张璐, 何新华. C3和C4植物的氮素利用机制. 植物学报, 2020, 55(2): 228-239 doi:10.11983/CBB19113 Zhang Lu, He Xinhua. Nitrogen Utilization Mechanism in C3 and C4 Plants. Chinese Bulletin of Botany, 2020, 55(2): 228-239 doi:10.11983/CBB19113
氮素是维持植物生长发育最主要的元素之一, 提高作物的氮素利用效率(nitrogen use efficiency, NUE)是保障粮食安全, 实现“资源节约和环境友好”的重要途径(徐晓鹏等, 2016)。根据叶片解剖结构、植物碳代谢方式以及生物化学代谢差异, Brown (1978)提出C4比C3植物氮素利用效率高的假说。20世纪80-90年代在室内人工控制条件下佐证了这一假说的正确性(Sage and Pearcy, 1987; Oaks, 1994; He et al., 1994; 何新华等, 1995)。除氮素利用效率在碳代谢上的差异外, NUE差异的关键在于氮代谢特征, 即氮素还原、同化酶类在细胞内发挥功能的位置 以及对氮素吸收、还原、同化的机制不同。何新华等(1995)认为, C3和C4植物氮素利用效率差异由特殊基因控制, 是长期进化的结果。然而, 目前对C3和C4植物氮素利用效率差异机制的进展尚未见系统梳理。本文综述C3和C4植物对氮素利用效率的差异机制、成因及C3作物氮素利用效率遗传改良技术, 旨在为合理利用氮素, 减少农业面源污染提供科学依据。
1 C3和C4植物氮素同化酶类的空间差异
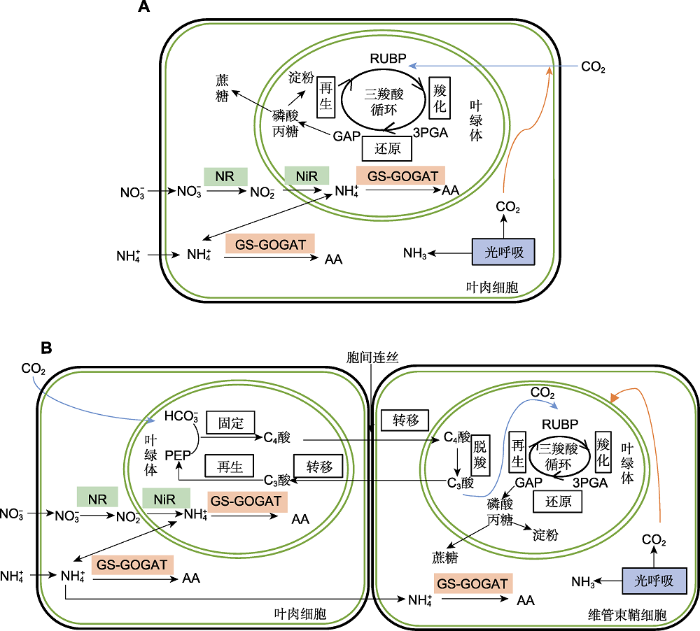
C4植物氮素还原同化酶的空间协同效应是C4比C3植物氮素利用效率高的主要原因(图1A, B)。C4植物在结构上区别于C3植物的典型特征是拥有Kranz结构的维管束鞘细胞(巩玥等, 2014)。C4植物既可以在叶肉细胞, 也可在维管束鞘细胞进行NH4+的同化, 合成氨基酸和蛋白质(何新华等, 1995); 而C3植物氮素还原只在叶肉细胞进行, 导致其NUE低。采用蛋白质组学研究玉米(Zea mays)叶肉细胞和维管束鞘细胞叶绿体(膜)上蛋白质表达差异, 结果表明, 在C4植物中氮同化(如氨基酸合成)在叶肉和维管束鞘细胞之间是被分隔开的(Majeran and Wijk, 2009), 与氮吸收相关酶类主要存在于叶肉细胞的叶绿体中, 而淀粉合成相关酶类主要存在于维管束鞘细胞的叶绿体中(Majeran et al., 2005; Suzuki et al., 2007)。
向上(绿色)和向下(红色)箭头分别表示需提高和限制的目标路径和酶活性。Rubisco: 核酮糖-1,5-二磷酸羧化酶/加氧酶; 2-PG: 磷酸乙醇酸; CCM: CO2浓缩机制 Figure 2Strategies for improving nitrogen use efficiency (NUE) of C3 plants (modified from Nowicka et al., 2018)
Pathways/enzymes for activation are marked in upwards arrows (green), while those for repression are marked in downwards arrow (red). Rubisco: Ribulose bisphosphate carboxylase oxygenase; 2-PG: 2-phosphoglycolate; CCM: CO2 concentrating mechanism
利用DNA重组改良C3作物的技术体系已逐渐成熟。目前, 已从玉米和高粱(Sorghum bicolor)等C4植物中克隆到与C4光合相关的关键酶基因, 如NADP- ME、PCK、PEPC和PPDK, 且这些基因已成功转入水稻(Bandyopadhyay et al., 2007)和小麦(张庆琛等, 2010)等C3作物中。将水稻Cab启动子控制的玉米NADP-ME cDNA转入水稻, 使得水稻叶片NADP- ME活性增加了30-70倍, 但NADP-ME蛋白量也随之增加(Tsuchida et al., 2001)。转玉米PEPC基因的水稻PEPC活性远高于野生型水稻, 与叶片光合速率和叶绿素均呈正相关(李霞等, 2008)。转入外源玉米PEPC基因的水稻植株含氮量显著增加, 并在低氮条件下影响氨基酸的合成和有机酸含量, 促进氮代谢和氮同化产物的积累和植株生长(陈冉冉, 2012)。转入碳相关基因, 对水稻的氮代谢相关酶有显著影响, 这为氮高效利用提供了新的思路(Yanagisawa et al., 2004; Kurai et al., 2011)。研究结果与Melzer和O'Leary (1987)提出的将PEPC基因转入C3植物后可能存在代谢通路相一致, 即PEPC催化PEP与CO2反应形成草酰乙酸(oxaloacetic acid, OAA), 一部分OAA经苹果酸脱氢酶(MDH)催化脱氢产生的苹果酸和OAA进入三羧酸循环(tricarboxylic acid cycle, TCA), 增强呼吸作用的同时为氨基酸合成提供丰富的有机酸和碳骨架; 另一部分OAA经转氨酶作用形成天门冬氨酸。而PEPC催化CO2与PEP反应生成的OAA进入TCA, OAA与苹果酸相互转化, 及时补充合成氨基酸消耗的碳骨架, 进而促进氨基酸的合成, 提高氮素利用率(Ku et al., 1996; Miyao and Fukayama, 2003; Suzuki et al., 2006; Radchuk et al., 2007)。
5.2 其它改良途径及改良限制因素
除C4核心酶CA、NADP-MDH、NADP-ME、PEPC和PPDK外, C4途径还需要将草酸、苹果酸、三磷酸甘油酸和丙酮酸的代谢物转运体插入C3植物内, 以增加C4循环中间体的运输能力, 使卡尔文循环能够在维管束鞘细胞中有效运作(Weber and von Caemmerer, 2010)。由于C3植物Rubisco具有高含量、低催化速率的特点, 因此通过在植物体内低量表达外源高催化速率Rubisco既可以提高光合作用效率也可增加氮素利用效率(Spreitzer and Salvucci, 2002; Whitney et al., 2011)。国际水稻联盟也在研究与Kranz解剖结构有关的基因(Wang et al., 2013), 发现减少叶肉细胞光呼吸也是提高氮素利用率的途径之一(图2)。例如, 降低叶肉细胞中的甘氨酸脱羧酶(Glycine decarboxylase complex, GDC)蛋白, 并限制其在维管束鞘细胞中的积累, 使甘氨酸脱羧完全发生在维管束鞘细胞中, 类似于C3-C4中间体, 从而维持较高的CO2浓度以降低光呼吸(Monson and Rawsthorne, 2000)。Kajala等(2011)通过人工microRNAs技术下调水稻中Rubisco小亚基(RbcS)和GDC-H亚基基因的表达, 使叶肉细胞中Rubisco和甘氨酸脱羧酶(GDC)得到下调。通过对玉米的维管束鞘细胞和叶肉细胞进行蛋白质组学鉴定, 发现在C4代谢途径中过表达转运蛋白, 如2-氧戊二酸/苹果酸转运体(OMT1)、二羧酸转运体1 (DiT1)、二羧酸转运体2 (DiT2)、PEP/磷酸盐转运蛋白(PPT1)、叶肉包膜蛋白(MEP)和三糖-磷酸盐转运体(TPT), 转入水稻后均得到特异性表达(Karki et al., 2013)。对氮素循环再利用也是提高NUE的有效途径。例如, 过表达自噬核心基因OsATG8b能够显著增强缺氮胁迫下转基因拟南芥叶片中的自噬活性, 从而提高氮素再利用效率(甄晓溪等, 2019)。
光合作用是一个经过数十亿年演变而来的复杂代谢过程, 在整个碳代谢过程中与氮代谢紧密联系, 高度整合。光呼吸中间产物对硝酸盐同化等看似与光合作用无关的代谢过程也有影响(Betti et al., 2016)。要充分发挥C4作物高光合效率及氮素利用潜力, 提高C3作物氮素利用率。此外, 编码光合特性的基因通常和氮还原同化的基因同时存在于细胞核和质体基因组中, 需要探索有效方法来同时转化这2个基因组(Ort et al., 2015), 但目前谷物的质体转化尚未成功。即使成功导入和表达了这些基因, 其产物也需要正确地折叠、运输, 并组装成功能复合物, 才能作用于特定部位, 提高氮素利用效率。
AppenrothKJ, Me?oR, JourdanV, LilloC ( 2000). Phytochrome and post-translational regulation of nitrate reductase in higher plants Plant Sci 159, 51-56. [本文引用: 1]
AubryS, BrownNJ, HibberdJM ( 2011). The role of proteins in C3 plants prior to their recruitment into the C4 pathway J Exp Bot 62, 3049-3059. [本文引用: 1]
BasraAS, DhawanAK, GoyalSS ( 2002). Dcmu inhibits in vivo nitrate reduction in illuminated barley (C3) leaves but not in maize (C4): a new mechanism for the role of light? Planta 215, 855-861. [本文引用: 1]
BettiM, BauwH, BuschFA, FernieAR, KeechO, LeveyM, Ort DonaldR, Parry MartinAJ, SageR, TimmS, WalkerB, WeberAPM ( 2016). Manipulating photorespiration to increase plant productivity: recent advances and perspectives for crop improvement J Exp Bot 67, 2977-2988. [本文引用: 2]
BloomAJ, AsensioJSR, RandallL, RachmilevitchS, CousinsAB, CarlisleEA ( 2012). CO2 enrichment inhibits shoot nitrate assimilation in C3 but not C4 plants and slows growth under nitrate in C3 plants Ecology 93, 355-367. [本文引用: 1]
Br?utigamA, KajalaK, WullenweberJ, SommerM, GagneulD, WeberKL, CarrKM, GowikU, MassJ, LercherMJ, WesthoffP, HibberdJM, WeberAP ( 2011). An mRNA blueprint for C4 photosynthesis derived from comparative transcriptomics of closely related C3 and C4 species Plant Physiol 155, 142-156. [本文引用: 1]
Br?utigamA, SchlieskyS, KülahogluC, OsborneCP, WeberAPM ( 2014). Towards an integrative model of C4 photosynthetic subtypes: insights from comparative transcriptome analysis of NAD-ME, NADP-ME, and PEP-CK C4 species J Exp Bot 65, 3579-3593. [本文引用: 2]
BrownRH ( 1978). A difference in N use efficiency in C3 and C4 plants and its implications in adaptation and evolution Crop Sci 18, 93-98. [本文引用: 3]
BuschFA, SageTL, CousinsAB, SageRF ( 2013). C3 plants enhance rates of photosynthesis by reassimilating photorespired and respired CO2 Plant Cell Environ 36, 200-212. [本文引用: 1]
CampbellWH ( 1999). Nitrate reductase structure, function and regulation: bridging the gap between biochemistry and physiology Annu Rev Plant Physiol Plant Mol Biol 50, 277-303. [本文引用: 1]
Carmo-SilvaE, ScalesJC, MadgwickPJ, ParryMAJ ( 2015). Optimizing Rubisco and its regulation for greater resource use efficiency Plant Cell Environ 38, 1817-1832. [本文引用: 1]
FurbankRT ( 2011). Evolution of the C4 photosynthetic mechanism: are there really three C4 acid decarboxylation types? J Exp Bot 62, 3103-3108. [本文引用: 1]
GowikU, Br?utigamA, WeberKL, WeberAP, WesthoffP ( 2011). Evolution of C4 photosynthesis in the genus flaveria: how many and which genes does it take to make C4? Plant Cell 23, 2087-2105. [本文引用: 1]
HagemannM, BauweH ( 2016). Photorespiration and the potential to improve photosynthesis Curr Opin Chem Biol 35, 109-116. [本文引用: 3]
HeXH, OaksA, LiM ( 1994). Nitrogen use efficiency in C3 and C4 plants Agro’s Ann Rev Plant Physiol 1, 147-188. [本文引用: 1]
HibberdJM, SheehyJE, LangdaleJA ( 2008). Using C4 photosynthesis to increase the yield of rice-rationale and feasibility Curr Opin Plant Biol 11, 228-231. [本文引用: 1]
HikosakaK, ShigenoA ( 2009). the role of Rubisco and cell walls in the interspecific variation in photosynthetic capacity Oecologia 160, 443-451. [本文引用: 1]
IPCC ( 2013). Climate change 2013: the physical science basis In: Stocker TF, Qin D, Plattner GK, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, Midgley PM, eds. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press. pp.119-158. [本文引用: 2]
JiangZC, HullRJ, SullivanWM ( 2002). Nitrate uptake and reduction in C3 and C4 grasses J Plant Nutr 25, 1303-1314. [本文引用: 2]
KajalaK, CovshoffS, KarkiS, WoodfieldH, TolleyBJ, DionoraMJA, MogulRT, MabilanganAE, DanilaFR, HibberdJM, QuickWP ( 2011). Strategies for engineering a two-celled C4 photosynthetic pathway into rice J Exp Bot 62, 3001-3010. [本文引用: 1]
KarkiS, RizalG, QuickWP ( 2013). Improvement of photosynthesis in rice ( Oryza sativa L.) by inserting the C4 pathway Rice 6, 28. [本文引用: 1]
KuMSB, Kano-MurakamiY, MatsuokaM ( 1996). Evolution and expression of C4 photosynthesis genes Plant Physiol 111, 949-957. [本文引用: 2]
KülahogluC, DentonAK, SommerM, Ma?J, SchlieskyS, WrobelTJ, BerckmansB, Gongora-CastilloE, BuellCR, SimonR, De VeylderL, Br?utigamA, WeberAPM ( 2014). Comparative transcriptome atlases reveal altered gene expression modules between two cleomaceae C3 and C4 plant species Plant Cell 26, 3243-3260. [本文引用: 1]
KuraiT, WakayamaM, AbikoT, YanagisawaS, AokiN, OhsugiR ( 2011). Introduction of the ZmDof1 gene into rice enhances carbon and nitrogen assimilation under low- nitrogen conditions Plant Biotechnol J 9, 826-837. [本文引用: 1]
LangdaleJA ( 2011). C4 cycles: past, present, and future research on C4 photosynthesis Plant Cell 23, 3879-3892. [本文引用: 1]
LiMR, WedinDA, TieszenLL ( 1999). C3 and C4 photosynthesis in Cyperus( Cyperaceae) in temperate eastern North America Can J Bot 77, 209-218. [本文引用: 1]
LongSP, AinsworthEA, LeakeyADB, N?sbergerJ, OrtDR ( 2006). Food for thought: lower-than-expected crop yield stimulation with rising CO2 concentrations Science 312, 1918-1921. [本文引用: 1]
MajeranW, CaiY, SunQ, van WijkKJ ( 2005). Functional differentiation of bundle sheath and mesophyll maize chloroplasts determined by comparative proteomics Plant Cell 17, 3111-3140. [本文引用: 1]
MajeranW, van WijkKJ ( 2009). Cell-type-specific differentiation of chloroplasts in C4 plants Trends Plant Sci 14, 100-109. [本文引用: 1]
MelzerE, O'LearyMH ( 1987). Anapleurotic CO2 fixation by phosphoenolpyruvate carboxylase in C3 plants Plant Physiol 84, 58-60. [本文引用: 1]
MiyaoM, FukayamaH ( 2003). Metabolic consequences of overproduction of phosphoenolpyruvate carboxylase in C3 plants Arch Biochem Biophys 414, 197-203. [本文引用: 1]
MonsonRK, RawsthorneS ( 2000). CO2 assimilation in C3-C4 intermediate plants In: Leegood RC, Sharkey TD, von Caemmerer S, eds. Photosynthesis: Physiology and Metabolism. Dordrecht: Springer. pp. 533-550. [本文引用: 1]
NASA ( 2014). Global climate change: vital signs of the planet available at: http://climate.nasa.gov/400ppmquotes . 2018-12-30. URL [本文引用: 1]
NelsonT ( 2011). The grass leaf developmental gradient as a platform for a systems understanding of the anatomical specialization of C4 leaves J Exp Bot 62, 3039-3048. [本文引用: 1]
OaksA ( 1994). Efficiency of nitrogen utilization in C3 and C4 cereals Plant Physiol 106, 407-414. [本文引用: 5]
OrtDR, MerchantSS, AlricJ, BarkanA, BlankenshipRE, BockR, CroceR, HansonMR, HibberdJM, LongSP, MooreTA, MoroneyJ, NiyogiKK, ParryMAJ, Peralta-YahyaPP, PrinceRC, ReddingKE, SpaldingMH, van WijkKJ, VermaasWFJ, von CaemmererS, WeberAPM, YeatesTO, YuanJS, ZhuXG ( 2015). Redesigning photosynthesis to sustainably meet global food and bioenergy demand Proc Natl Acad Sci USA 112, 8529-8536. [本文引用: 1]
PattersonDT, FlintEP ( 1980). Potential effects of global atmospheric CO2 enrichment on the growth and competitiveness of C3 and C4 weed and crop plants Weed Sci 28, 71-75. [本文引用: 1]
PeterhanselC, KrauseK, BraunHP, EspieGS, FernieAR, HansonDT, KeechO, MaurinoVG, MielewczikM, SageRF ( 2013). Engineering photorespiration: current state and future possibilities Plant Biol 15, 754-758. [本文引用: 2]
PintoH, SharwoodRE, TissueDT, GhannoumO ( 2014). Photosynthesis of C3, C3-C4, and C4 grasses at glacial CO2 J Exp Bot 65, 3669-3681. [本文引用: 1]
RadchukR, RadchukV, G?tzKP, WeichertH, RichterA, EmeryRJN, WeschkeW, WeberH ( 2007). Ectopic expression of phosphoenolpyruvate carboxylase in Vicia narbonensis seeds: effects of improved nutrient status on seed maturation and transcriptional regulatory networks Plant J 51, 819-839. [本文引用: 1]
ReichPB, HobbieSE, LeeTD, PastoreMA ( 2018). Unexpected reversal of C3 versus C4 grass response to elevated CO2 during a 20-year field experiment Science 360, 317-320. [本文引用: 1]
RipleyBS, CunniffJ, OsborneCP ( 2013). Photosynthetic acclimation and resource use by the C3 and C4 subspecies of Alloteropsis semialata in low CO2 atmospheres Global Change Biol 19, 900-910. [本文引用: 1]
RuanCJ, ShaoHB, dan SilvaJAT ( 2012). A critical review on the improvement of photosynthetic carbon assimilation in C3 plants using genetic engineering Crit Rev Biotechnol 32, 1-21. [本文引用: 3]
SageRF, ChristinPA, EdwardsEJ ( 2011). The C4 plant lineages of planet earth J Exp Bot 62, 3155-3169. [本文引用: 2]
SageRF, PearcyRW ( 1987). The nitrogen use efficiency of C3 and C4 plants. I. Leaf nitrogen, growth, and biomass partitioning in Chenopodium album(L.) and Amaranthus retroflexus Plant Physiol 84, 954-958. [本文引用: 2]
SageRF, SageTL, KocacinarF ( 2012). Photorespiration and the evolution of C4 photosynthesis Annu Rev Plant Biol 63, 19-47. [本文引用: 2]
SageRF, WedinDA, LiMR ( 1999). The biogeography of C4 photosynthesis: patterns and controlling factors In: Sage RF, Monson RK, eds. C4 Plant Biology. San Diego: Academic Press. pp.313-375. [本文引用: 2]
SantosPM, ThorntonB, CorsiM ( 2002). Nitrogen dynamics in the intact grasses poa trivialis and panicum maximum receiving contrasting supplies of nitrogen J Exp Bot 53, 2167-2176. [本文引用: 1]
SehtiyaHL, GoyalSS ( 2000). Comparative uptake of nitrate by intact seedlings of C3 (barley) and C4 (corn) plants: effect of light and exogenously supplied sucrose Plant Soil 227, 185-190. [本文引用: 2]
SpreitzerRJ, SalvucciME ( 2002). Rubisco: structure, regulatory interactions, and possibilities for a better enzyme Annu Rev Plant Biol 53, 449-475. [本文引用: 2]
SuzukiY, MiyamotoT, YoshizawaR, MaeT, MakinoA ( 2009). Rubisco content and photosynthesis of leaves at different positions in transgenic rice with an overexpression of RBCS Plant Cell Environ 32, 417-427. [本文引用: 1]
TamoiM, ShigeokaS ( 2005). Improvement of photosynthesis in higher plants In: Omasa K, Nouchi I, De Kok LJ, eds. Plant Responses to Air Pollution and Global Change. Tokyo: Springer. pp. 141-147. [本文引用: 1]
TaylorSH, HulmeSP, ReesM, RipleyBS, Ian WoodwardF, OsborneCP ( 2010). Ecophysiological traits in C3 and C4 grasses: a phylogenetically controlled screening experiment New Phytol 185, 780-791. [本文引用: 1]
TissueDT, DipperyJK, ThomasRB, StrainBR ( 1995). Effects of low and elevated CO2 on C3 and C4 annuals Oecologia 101, 21-28. [本文引用: 1]
TsuchidaH, TamaiT, FukayamaH, AgarieS, NomuraM, OnoderaH, OnoK, NishizawaY, LeeBH, HiroseS, TokiS, KuMSB, MatsuokaM, MiyaoM ( 2001). High level expression of C4-specific NADP-malic enzyme in leaves and impairment of photoautotrophic growth in a C3 plant, rice Plant Cell Physiol 42, 138-145. [本文引用: 1]
VoganPJ, SageRF ( 2012). Effects of low atmospheric CO2 and elevated temperature during growth on the gas exchange responses of C3, C3-C4 intermediate, and C4 species from three evolutionary lineages of C4 photosynthesis Oecologia 169, 341-352. [本文引用: 1]
VoznesenskayaEV, FranceschiVR, KiiratsO, ArtyushevaEG, FreitagH, EdwardsGE ( 2002). Proof of C4 photosynthesis without Kranz anatomy in Bienertia cycloptera(Chenopodiaceae) Plant J 31, 649-662. [本文引用: 1]
VoznesenskayaEV, FranceschiVR, KiiratsO, FreitagH, EdwardsGE ( 2001). Kranz anatomy is not essential for terrestrial C4 plant photosynthesis Nature 414, 543-546. [本文引用: 1]
WalkerBJ, VanLoockeA, BernacchiCJ, OrtDR ( 2016). The costs of photorespiration to food production now and in the future Annu Rev Plant Biol 67, 107-129. [本文引用: 1]
WangP, KellyS, FouracreJP, LangdaleJA ( 2013). Genome-wide transcript analysis of early maize leaf development reveals gene cohorts associated with the differentiation of C4 Kranz anatomy Plant J 75, 656-670. [本文引用: 1]
WardJK, TissueDT, ThomasBR, StrainBDR ( 1999). Comparative responses of model C3 and C4 plants to drought in low and elevated CO2 Global Change Biol 5, 857-867. [本文引用: 1]
WeberAPM, von CaemmererS ( 2010). Plastid transport and metabolism of C3 and C4 plants-comparative analysis and possible biotechnological exploitation Curr Opin Plant Biol 13, 256-264. [本文引用: 1]
WessingerME, EdwardsGE, KuMSB ( 1989). Quantity and kinetic properties of ribulose 1,5-bisphosphate carboxylase in C3, C4, and C3-C4 intermediate species of flaveria (asteraceae) Plant Cell Physiol 30, 665-671. [本文引用: 1]
WhitneySM, SharwoodRE, OrrD, WhiteSJ, AlonsoH, GalmésJ ( 2011). Isoleucine 309 acts as a C4 catalytic switch that increases ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) carboxylation rate in flaveria Proc Natl Acad Sci USA 108, 14688-14693. [本文引用: 1]
YanagisawaS, AkiyamaA, KisakaH, UchimiyaH, MiwaT ( 2004). Metabolic engineering with dof1 transcription factor in plants: improved nitrogen assimilation and growth under low-nitrogen conditions Proc Natl Acad Sci USA 101, 7833-7838. [本文引用: 1]
YinZH, RavenJA ( 1997). A comparison of the impacts of various nitrogen sources on acid-base balance in C3Triticum aestivum L.and C4Zea mays L. plants J Exp Bot 48, 315-324. [本文引用: 1]
YinZH, RavenJA ( 1998). Influences of different nitrogen sources on nitrogen- and water-use efficiency, and carbon isotope discrimination, in C3Triticum aestivum L. and C4Zea mays L. plants. Planta 205, 574-580. [本文引用: 3]
ZengQ, LiuB, GilnaB, ZhangYL, ZhuCW, MaHL, PangJ, ChenGP, ZhuJG ( 2011). Elevated CO2 effects on nutrient competition between a C3 crop (Oryza sativa L.) and a C4 weed(Echinochloa crusgalli L.) Nutr Cycl Agroecosys 89, 93-104. [本文引用: 1]
ZhengZL ( 2009). Carbon and nitrogen nutrient balance signaling in plants Plant Signal Behav 4, 584-591. [本文引用: 1]
不同氮素水平对转玉米PEPC基因水稻碳、氮代谢的影响 1 2012
... 利用DNA重组改良C3作物的技术体系已逐渐成熟.目前, 已从玉米和高粱(Sorghum bicolor)等C4植物中克隆到与C4光合相关的关键酶基因, 如NADP- ME、PCK、PEPC和PPDK, 且这些基因已成功转入水稻(Bandyopadhyay et al., 2007)和小麦(张庆琛等, 2010)等C3作物中.将水稻Cab启动子控制的玉米NADP-ME cDNA转入水稻, 使得水稻叶片NADP- ME活性增加了30-70倍, 但NADP-ME蛋白量也随之增加(Tsuchida et al., 2001).转玉米PEPC基因的水稻PEPC活性远高于野生型水稻, 与叶片光合速率和叶绿素均呈正相关(李霞等, 2008).转入外源玉米PEPC基因的水稻植株含氮量显著增加, 并在低氮条件下影响氨基酸的合成和有机酸含量, 促进氮代谢和氮同化产物的积累和植株生长(陈冉冉, 2012).转入碳相关基因, 对水稻的氮代谢相关酶有显著影响, 这为氮高效利用提供了新的思路(Yanagisawa et al., 2004; Kurai et al., 2011).研究结果与Melzer和O'Leary (1987)提出的将PEPC基因转入C3植物后可能存在代谢通路相一致, 即PEPC催化PEP与CO2反应形成草酰乙酸(oxaloacetic acid, OAA), 一部分OAA经苹果酸脱氢酶(MDH)催化脱氢产生的苹果酸和OAA进入三羧酸循环(tricarboxylic acid cycle, TCA), 增强呼吸作用的同时为氨基酸合成提供丰富的有机酸和碳骨架; 另一部分OAA经转氨酶作用形成天门冬氨酸.而PEPC催化CO2与PEP反应生成的OAA进入TCA, OAA与苹果酸相互转化, 及时补充合成氨基酸消耗的碳骨架, 进而促进氨基酸的合成, 提高氮素利用率(Ku et al., 1996; Miyao and Fukayama, 2003; Suzuki et al., 2006; Radchuk et al., 2007). ...
Photorespiration connects C3 and C4 photosynthesis 1 2016
... 与C3植物相比, C4植物蛋白质合成相关基因的表达也相应较低(Bräutigam et al., 2011, 2014; Gowik et al., 2011).Bräutigam和Gowik (2016)认为, 基因表达量降低和叶片蛋白质含量减少使一些C4植物具有更高的氮素利用效率, 其原因是Rubisco在光合作用和氮素利用与平衡方面发挥重要作用(Spreitzer and Salvucci, 2002; Nowicka et al., 2018).其一, 核酮糖-1,5-二磷酸(RuBP)的氧化反应生成3-磷酸甘油酸(3-PGA)和2-磷酸甘油酸(2-PG), 光呼吸将2个2-PGA分子循环为1个3-PGA分子, 因此有25%的有机碳以CO2的形式流失(Sage et al., 2012; Betti et al., 2016; Hagemann and Bauwe, 2016).在主要C3作物(如小麦)中, 每3-4分子RuBP中就有1个被氧化为3-PGA和2-PG (Hagemann and Bauwe, 2016), 而2-PG是光合碳代谢酶抑制剂; C4植物的Rubisco加氧酶活性较低, Rubisco羧化酶代谢循环具有高效率优势(Sage, 2001).光呼吸需要大量的能量输入, 导致同化碳和氮分别以CO2和NH3形式流失(Peterhansel et al., 2013).其二, 制约Rubisco缓慢的羧化反应, 其周转率常在1-10次·s-1之间, 而其它中央代谢酶的周转频率在50-100次·s-1之间(Bar-Even et al., 2011).为了弥补Rubisco的低活性, C3植物产生大量Rubisco, 占光合生物可溶性蛋白质的50% (Erb and Zarzycki, 2016).而高光合效率所需的Rubisco更少, 通常C4植物的Rubisco含量占可溶性蛋白总量的10%-25% (Carmo-Silva et al., 2015).C3植物对Rubisco的高氮投入会导致其氮素利用效率(生物产量与投入氮的比值)较低.因此, 提高Rubisco利用效率(RBUE)是C3植物增强光合氮素利用效率的主要驱动因素(Hikosaka and Shigeno, 2009). ...
An mRNA blueprint for C4 photosynthesis derived from comparative transcriptomics of closely related C3 and C4 species 1 2011
... 与C3植物相比, C4植物蛋白质合成相关基因的表达也相应较低(Bräutigam et al., 2011, 2014; Gowik et al., 2011).Bräutigam和Gowik (2016)认为, 基因表达量降低和叶片蛋白质含量减少使一些C4植物具有更高的氮素利用效率, 其原因是Rubisco在光合作用和氮素利用与平衡方面发挥重要作用(Spreitzer and Salvucci, 2002; Nowicka et al., 2018).其一, 核酮糖-1,5-二磷酸(RuBP)的氧化反应生成3-磷酸甘油酸(3-PGA)和2-磷酸甘油酸(2-PG), 光呼吸将2个2-PGA分子循环为1个3-PGA分子, 因此有25%的有机碳以CO2的形式流失(Sage et al., 2012; Betti et al., 2016; Hagemann and Bauwe, 2016).在主要C3作物(如小麦)中, 每3-4分子RuBP中就有1个被氧化为3-PGA和2-PG (Hagemann and Bauwe, 2016), 而2-PG是光合碳代谢酶抑制剂; C4植物的Rubisco加氧酶活性较低, Rubisco羧化酶代谢循环具有高效率优势(Sage, 2001).光呼吸需要大量的能量输入, 导致同化碳和氮分别以CO2和NH3形式流失(Peterhansel et al., 2013).其二, 制约Rubisco缓慢的羧化反应, 其周转率常在1-10次·s-1之间, 而其它中央代谢酶的周转频率在50-100次·s-1之间(Bar-Even et al., 2011).为了弥补Rubisco的低活性, C3植物产生大量Rubisco, 占光合生物可溶性蛋白质的50% (Erb and Zarzycki, 2016).而高光合效率所需的Rubisco更少, 通常C4植物的Rubisco含量占可溶性蛋白总量的10%-25% (Carmo-Silva et al., 2015).C3植物对Rubisco的高氮投入会导致其氮素利用效率(生物产量与投入氮的比值)较低.因此, 提高Rubisco利用效率(RBUE)是C3植物增强光合氮素利用效率的主要驱动因素(Hikosaka and Shigeno, 2009). ...
Towards an integrative model of C4 photosynthetic subtypes: insights from comparative transcriptome analysis of NAD-ME, NADP-ME, and PEP-CK C4 species 2 2014
... 随着人们对光合途径认识的不断深入, 对C3与C4植物光合类型划分界限也越来越不明显.C4途径的关键酶包括磷酸烯醇式丙酮酸羧化酶(phosphoenolpyruvate carboxylase, PEPC)、丙酮酸磷酸双激酶(pyruvate orthophosphate dikinase, PPDK)、NAD (P)-苹果酸酶(NAD (P) malic enzyme, NAD(P)-ME)、NAD (P)-苹果酸脱氢酶(NAD(P)-malic dehydrogenase, NAD (P)-MDH)及碳酸酐酶(carbonic anhydrase, CA)等.研究表明, C4循环中所有酶促反应都存在于C3植物中(Aubry et al., 2011), 并且C4循环酶的大部分基因能够在C3叶片中表达(Christin et al., 2013; Bräutigam et al., 2014).C4光合作用中有3种类型的C4酸脱羧酶, 即需NADP的苹果酸酶(NADP- ME)、需NAD的苹果酸酶(NAD-ME)和磷酸烯醇式丙酮酸羧激酶(phosphoenolpyruvate carboxylase, PEPCK) (Furbank, 2011). ...
... 与C3植物相比, C4植物蛋白质合成相关基因的表达也相应较低(Bräutigam et al., 2011, 2014; Gowik et al., 2011).Bräutigam和Gowik (2016)认为, 基因表达量降低和叶片蛋白质含量减少使一些C4植物具有更高的氮素利用效率, 其原因是Rubisco在光合作用和氮素利用与平衡方面发挥重要作用(Spreitzer and Salvucci, 2002; Nowicka et al., 2018).其一, 核酮糖-1,5-二磷酸(RuBP)的氧化反应生成3-磷酸甘油酸(3-PGA)和2-磷酸甘油酸(2-PG), 光呼吸将2个2-PGA分子循环为1个3-PGA分子, 因此有25%的有机碳以CO2的形式流失(Sage et al., 2012; Betti et al., 2016; Hagemann and Bauwe, 2016).在主要C3作物(如小麦)中, 每3-4分子RuBP中就有1个被氧化为3-PGA和2-PG (Hagemann and Bauwe, 2016), 而2-PG是光合碳代谢酶抑制剂; C4植物的Rubisco加氧酶活性较低, Rubisco羧化酶代谢循环具有高效率优势(Sage, 2001).光呼吸需要大量的能量输入, 导致同化碳和氮分别以CO2和NH3形式流失(Peterhansel et al., 2013).其二, 制约Rubisco缓慢的羧化反应, 其周转率常在1-10次·s-1之间, 而其它中央代谢酶的周转频率在50-100次·s-1之间(Bar-Even et al., 2011).为了弥补Rubisco的低活性, C3植物产生大量Rubisco, 占光合生物可溶性蛋白质的50% (Erb and Zarzycki, 2016).而高光合效率所需的Rubisco更少, 通常C4植物的Rubisco含量占可溶性蛋白总量的10%-25% (Carmo-Silva et al., 2015).C3植物对Rubisco的高氮投入会导致其氮素利用效率(生物产量与投入氮的比值)较低.因此, 提高Rubisco利用效率(RBUE)是C3植物增强光合氮素利用效率的主要驱动因素(Hikosaka and Shigeno, 2009). ...
A difference in N use efficiency in C3 and C4 plants and its implications in adaptation and evolution 3 1978
... 氮素是维持植物生长发育最主要的元素之一, 提高作物的氮素利用效率(nitrogen use efficiency, NUE)是保障粮食安全, 实现“资源节约和环境友好”的重要途径(徐晓鹏等, 2016).根据叶片解剖结构、植物碳代谢方式以及生物化学代谢差异, Brown (1978)提出C4比C3植物氮素利用效率高的假说.20世纪80-90年代在室内人工控制条件下佐证了这一假说的正确性(Sage and Pearcy, 1987; Oaks, 1994; He et al., 1994; 何新华等, 1995).除氮素利用效率在碳代谢上的差异外, NUE差异的关键在于氮代谢特征, 即氮素还原、同化酶类在细胞内发挥功能的位置 以及对氮素吸收、还原、同化的机制不同.何新华等(1995)认为, C3和C4植物氮素利用效率差异由特殊基因控制, 是长期进化的结果.然而, 目前对C3和C4植物氮素利用效率差异机制的进展尚未见系统梳理.本文综述C3和C4植物对氮素利用效率的差异机制、成因及C3作物氮素利用效率遗传改良技术, 旨在为合理利用氮素, 减少农业面源污染提供科学依据. ...
... C4植物光合作用机制是被子植物长期适应环境进化而成(Sage et al., 2011).与C3光合机理相比, C4光合作用进化出了复杂的叶片结构和生化适应机制(Christin and Osborne, 2014).近7 500种C4植物隶属于62个不同进化区系(Sage et al., 2011), 虽然C4植物在数量上只占陆地植物的3%, 却完成了陆生植物25%的光合作用(Edwards et al., 2010).C4植物光合优势在于其气孔数量多, CO2补偿点低, 高温或高辐照条件下较低的光呼吸(Li et al., 1999)以及高氮素利用效率(Brown, 1978; Sage and Pearcy, 1987; Oaks 1994; 何新华等, 1995).例如, 莎草属(Cyperus)植物高NUE的原因在于其高效率的Rubisco光合酶(Sage et al., 1999).C4植物总Rubisco基因表达较C3植物大幅下调, 导致Rubisco蛋白及Calvin-Benson循环酶含量减少(不包括将3-PGA还原为磷酸丙糖酶) (Wessinger et al., 1989). ...
... 在当前CO2浓度条件下, C4植物比C3植物有更高的NUE (Brown, 1978; Taylor et al., 2010).一般而言, 低浓度CO2会降低C3植物的光合速率, 使气孔导度和叶片氮浓度增加(Tissue et al., 1995; Ward et al., 1999; Ripley et al., 2013).Pinto等(2014)研究了C3和C4植物在冰川CO2 浓度为180 mmol∙L-1及环境CO2浓度为400 mmol∙L-1下的光合作用, 结果表明, 与C3植物相比, 冰川较低CO2浓度(180 mmol∙L-1)导致C4植物的光合作用降低幅度较小, 气孔导度增加较大.C3植物通过增加Rubisco活性来适应低浓度CO2, C4植物在低CO2浓度下有较高的气孔导度, 但由于光合速率较高, C4植物有较高的PNUE.Vogan和Sage (2012)研究了黄花菊属(Flaveria)、天芥菜属(Heliotropium)和蓬子草属(Alternanthera)中近缘C3 (2种)、C3-C4 (9种)和C4 (3种)植物对低浓度CO2的响应, 发现在380和180 mmol∙L-1 CO2下, 昼/夜气温为37/29°C时, CO2对任何一种植物的光合氮素利用率、Rubisco含量和电子传递均无显著影响. ...
C3 plants enhance rates of photosynthesis by reassimilating photorespired and respired CO2 1 2013
Strategies for improving nitrogen use efficiency (NUE) of C<sub>3 </sub>plants (modified from <xref ref-type="bibr" rid="b61">Nowicka et al., 2018</xref>) Pathways/enzymes for activation are marked in upwards arrows (green), while those for repression are marked in downwards arrow (red). Rubisco: Ribulose bisphosphate carboxylase oxygenase; 2-PG: 2-phosphoglycolate; CCM: CO2 concentrating mechanism ...
Efficiency of nitrogen utilization in C3 and C4 cereals 5 1994
... 氮素是维持植物生长发育最主要的元素之一, 提高作物的氮素利用效率(nitrogen use efficiency, NUE)是保障粮食安全, 实现“资源节约和环境友好”的重要途径(徐晓鹏等, 2016).根据叶片解剖结构、植物碳代谢方式以及生物化学代谢差异, Brown (1978)提出C4比C3植物氮素利用效率高的假说.20世纪80-90年代在室内人工控制条件下佐证了这一假说的正确性(Sage and Pearcy, 1987; Oaks, 1994; He et al., 1994; 何新华等, 1995).除氮素利用效率在碳代谢上的差异外, NUE差异的关键在于氮代谢特征, 即氮素还原、同化酶类在细胞内发挥功能的位置 以及对氮素吸收、还原、同化的机制不同.何新华等(1995)认为, C3和C4植物氮素利用效率差异由特殊基因控制, 是长期进化的结果.然而, 目前对C3和C4植物氮素利用效率差异机制的进展尚未见系统梳理.本文综述C3和C4植物对氮素利用效率的差异机制、成因及C3作物氮素利用效率遗传改良技术, 旨在为合理利用氮素, 减少农业面源污染提供科学依据. ...
... C4植物光合作用机制是被子植物长期适应环境进化而成(Sage et al., 2011).与C3光合机理相比, C4光合作用进化出了复杂的叶片结构和生化适应机制(Christin and Osborne, 2014).近7 500种C4植物隶属于62个不同进化区系(Sage et al., 2011), 虽然C4植物在数量上只占陆地植物的3%, 却完成了陆生植物25%的光合作用(Edwards et al., 2010).C4植物光合优势在于其气孔数量多, CO2补偿点低, 高温或高辐照条件下较低的光呼吸(Li et al., 1999)以及高氮素利用效率(Brown, 1978; Sage and Pearcy, 1987; Oaks 1994; 何新华等, 1995).例如, 莎草属(Cyperus)植物高NUE的原因在于其高效率的Rubisco光合酶(Sage et al., 1999).C4植物总Rubisco基因表达较C3植物大幅下调, 导致Rubisco蛋白及Calvin-Benson循环酶含量减少(不包括将3-PGA还原为磷酸丙糖酶) (Wessinger et al., 1989). ...
Engineering photorespiration: current state and future possibilities 2 2013
... 与C3植物相比, C4植物蛋白质合成相关基因的表达也相应较低(Bräutigam et al., 2011, 2014; Gowik et al., 2011).Bräutigam和Gowik (2016)认为, 基因表达量降低和叶片蛋白质含量减少使一些C4植物具有更高的氮素利用效率, 其原因是Rubisco在光合作用和氮素利用与平衡方面发挥重要作用(Spreitzer and Salvucci, 2002; Nowicka et al., 2018).其一, 核酮糖-1,5-二磷酸(RuBP)的氧化反应生成3-磷酸甘油酸(3-PGA)和2-磷酸甘油酸(2-PG), 光呼吸将2个2-PGA分子循环为1个3-PGA分子, 因此有25%的有机碳以CO2的形式流失(Sage et al., 2012; Betti et al., 2016; Hagemann and Bauwe, 2016).在主要C3作物(如小麦)中, 每3-4分子RuBP中就有1个被氧化为3-PGA和2-PG (Hagemann and Bauwe, 2016), 而2-PG是光合碳代谢酶抑制剂; C4植物的Rubisco加氧酶活性较低, Rubisco羧化酶代谢循环具有高效率优势(Sage, 2001).光呼吸需要大量的能量输入, 导致同化碳和氮分别以CO2和NH3形式流失(Peterhansel et al., 2013).其二, 制约Rubisco缓慢的羧化反应, 其周转率常在1-10次·s-1之间, 而其它中央代谢酶的周转频率在50-100次·s-1之间(Bar-Even et al., 2011).为了弥补Rubisco的低活性, C3植物产生大量Rubisco, 占光合生物可溶性蛋白质的50% (Erb and Zarzycki, 2016).而高光合效率所需的Rubisco更少, 通常C4植物的Rubisco含量占可溶性蛋白总量的10%-25% (Carmo-Silva et al., 2015).C3植物对Rubisco的高氮投入会导致其氮素利用效率(生物产量与投入氮的比值)较低.因此, 提高Rubisco利用效率(RBUE)是C3植物增强光合氮素利用效率的主要驱动因素(Hikosaka and Shigeno, 2009). ...
... 通过遗传或基因工程技术向C3作物转入C4作物高光合效率相关基因是提高作物产量和保障粮食安全的有效途径之一(Hibberd et al., 2008; Peterhansel et al., 2013).利用基因工程和转基因技术提高Rubisco羧化酶反应效率(Ku et al., 1996, 1999; Tamoi and Shigeoka, 2005; Suzuki et al., 2009), 将C4途径相关酶基因转入C3植物, 通过超量表达C4途径相关核心酶, 在C3植物内建立类似C4植物特性的CCM (CO2 concentrating mechanism)单细胞循环等方面已进行了大量研究(陈太钰, 2012; Ruan et al., 2012; Hagemann and Bauwe, 2016). ...
Photosynthesis of C3, C3-C4, and C4 grasses at glacial CO2 1 2014
... 在当前CO2浓度条件下, C4植物比C3植物有更高的NUE (Brown, 1978; Taylor et al., 2010).一般而言, 低浓度CO2会降低C3植物的光合速率, 使气孔导度和叶片氮浓度增加(Tissue et al., 1995; Ward et al., 1999; Ripley et al., 2013).Pinto等(2014)研究了C3和C4植物在冰川CO2 浓度为180 mmol∙L-1及环境CO2浓度为400 mmol∙L-1下的光合作用, 结果表明, 与C3植物相比, 冰川较低CO2浓度(180 mmol∙L-1)导致C4植物的光合作用降低幅度较小, 气孔导度增加较大.C3植物通过增加Rubisco活性来适应低浓度CO2, C4植物在低CO2浓度下有较高的气孔导度, 但由于光合速率较高, C4植物有较高的PNUE.Vogan和Sage (2012)研究了黄花菊属(Flaveria)、天芥菜属(Heliotropium)和蓬子草属(Alternanthera)中近缘C3 (2种)、C3-C4 (9种)和C4 (3种)植物对低浓度CO2的响应, 发现在380和180 mmol∙L-1 CO2下, 昼/夜气温为37/29°C时, CO2对任何一种植物的光合氮素利用率、Rubisco含量和电子传递均无显著影响. ...
Ectopic expression of phosphoenolpyruvate carboxylase in Vicia narbonensis seeds: effects of improved nutrient status on seed maturation and transcriptional regulatory networks 1 2007
... 利用DNA重组改良C3作物的技术体系已逐渐成熟.目前, 已从玉米和高粱(Sorghum bicolor)等C4植物中克隆到与C4光合相关的关键酶基因, 如NADP- ME、PCK、PEPC和PPDK, 且这些基因已成功转入水稻(Bandyopadhyay et al., 2007)和小麦(张庆琛等, 2010)等C3作物中.将水稻Cab启动子控制的玉米NADP-ME cDNA转入水稻, 使得水稻叶片NADP- ME活性增加了30-70倍, 但NADP-ME蛋白量也随之增加(Tsuchida et al., 2001).转玉米PEPC基因的水稻PEPC活性远高于野生型水稻, 与叶片光合速率和叶绿素均呈正相关(李霞等, 2008).转入外源玉米PEPC基因的水稻植株含氮量显著增加, 并在低氮条件下影响氨基酸的合成和有机酸含量, 促进氮代谢和氮同化产物的积累和植株生长(陈冉冉, 2012).转入碳相关基因, 对水稻的氮代谢相关酶有显著影响, 这为氮高效利用提供了新的思路(Yanagisawa et al., 2004; Kurai et al., 2011).研究结果与Melzer和O'Leary (1987)提出的将PEPC基因转入C3植物后可能存在代谢通路相一致, 即PEPC催化PEP与CO2反应形成草酰乙酸(oxaloacetic acid, OAA), 一部分OAA经苹果酸脱氢酶(MDH)催化脱氢产生的苹果酸和OAA进入三羧酸循环(tricarboxylic acid cycle, TCA), 增强呼吸作用的同时为氨基酸合成提供丰富的有机酸和碳骨架; 另一部分OAA经转氨酶作用形成天门冬氨酸.而PEPC催化CO2与PEP反应生成的OAA进入TCA, OAA与苹果酸相互转化, 及时补充合成氨基酸消耗的碳骨架, 进而促进氨基酸的合成, 提高氮素利用率(Ku et al., 1996; Miyao and Fukayama, 2003; Suzuki et al., 2006; Radchuk et al., 2007). ...
Unexpected reversal of C3 versus C4 grass response to elevated CO2 during a 20-year field experiment 1 2018
... 在正常生长条件下, 光呼吸损失的CO2占植物固定CO2总量的25% (Sage et al., 2012).Walker等(2016)利用模型分析显示, 在未来大气CO2浓度达到预测最高水平时, 植物光呼吸缺失下其光合速率可上调12%-55%.大气中CO2浓度升高导致C3植物光呼吸速率和NO3-同化速率同时下降, 吸收的NO3-不能及时被同化利用, 导致作物品质下降(Long et al., 2006; Bloom et al., 2010, 2014), 这也是植物在CO2浓度升高的条件下生长速率增幅较低的原因(Bloom et al., 2012).光合暗反应中, 光呼吸降低使ATP对NADPH的需求比例也降低.基质中CO2的增加导致NADP/ NADPH升高, 从而限制了NO3-代谢的还原能力.在高浓度CO2下, 当氮素供应不足时, C3植物(小麦和拟南芥(Arabidopsis thaliana))地上部分氮素含量的下降比氮素供应充足时更为显著(Bloom et al., 2010).Zeng等(2011)在白天CO2浓度为(374±21) mmol∙L-1和(568±39) mmol∙L-1, 夜间为(418±29) mmol∙L-1和(623±51) mmol∙L-1的农业FACE (free air CO2 enrichment)系统下研究CO2升高对C3植物水稻与C4植物稗草(Echinochloa crusgalli)氮素利用的影响, 发现其NUE增加, 在有限氮素下高浓度CO2使水稻比稗草具有更强的竞争优势.Reich等(2018)在长达20年的野外试验中发现, C3和C4植物在前12年对高浓度CO2的反应与前人研究结果一致, 在随后的8年里发生了意想不到的逆转.CO2浓度升高背景下, C4草地净氮矿化呈现随时间变化而逐渐增大的正响应, 而C3草地呈逐渐减小的负响应; 生物量对CO2升高的响应均与净氮矿化速率呈正相关. ...
Photosynthetic acclimation and resource use by the C3 and C4 subspecies of Alloteropsis semialata in low CO2 atmospheres 1 2013
... 在当前CO2浓度条件下, C4植物比C3植物有更高的NUE (Brown, 1978; Taylor et al., 2010).一般而言, 低浓度CO2会降低C3植物的光合速率, 使气孔导度和叶片氮浓度增加(Tissue et al., 1995; Ward et al., 1999; Ripley et al., 2013).Pinto等(2014)研究了C3和C4植物在冰川CO2 浓度为180 mmol∙L-1及环境CO2浓度为400 mmol∙L-1下的光合作用, 结果表明, 与C3植物相比, 冰川较低CO2浓度(180 mmol∙L-1)导致C4植物的光合作用降低幅度较小, 气孔导度增加较大.C3植物通过增加Rubisco活性来适应低浓度CO2, C4植物在低CO2浓度下有较高的气孔导度, 但由于光合速率较高, C4植物有较高的PNUE.Vogan和Sage (2012)研究了黄花菊属(Flaveria)、天芥菜属(Heliotropium)和蓬子草属(Alternanthera)中近缘C3 (2种)、C3-C4 (9种)和C4 (3种)植物对低浓度CO2的响应, 发现在380和180 mmol∙L-1 CO2下, 昼/夜气温为37/29°C时, CO2对任何一种植物的光合氮素利用率、Rubisco含量和电子传递均无显著影响. ...
A critical review on the improvement of photosynthetic carbon assimilation in C3 plants using genetic engineering 3 2012
... C4植物光合作用机制是被子植物长期适应环境进化而成(Sage et al., 2011).与C3光合机理相比, C4光合作用进化出了复杂的叶片结构和生化适应机制(Christin and Osborne, 2014).近7 500种C4植物隶属于62个不同进化区系(Sage et al., 2011), 虽然C4植物在数量上只占陆地植物的3%, 却完成了陆生植物25%的光合作用(Edwards et al., 2010).C4植物光合优势在于其气孔数量多, CO2补偿点低, 高温或高辐照条件下较低的光呼吸(Li et al., 1999)以及高氮素利用效率(Brown, 1978; Sage and Pearcy, 1987; Oaks 1994; 何新华等, 1995).例如, 莎草属(Cyperus)植物高NUE的原因在于其高效率的Rubisco光合酶(Sage et al., 1999).C4植物总Rubisco基因表达较C3植物大幅下调, 导致Rubisco蛋白及Calvin-Benson循环酶含量减少(不包括将3-PGA还原为磷酸丙糖酶) (Wessinger et al., 1989). ...
... 植物隶属于62个不同进化区系(Sage et al., 2011), 虽然C4植物在数量上只占陆地植物的3%, 却完成了陆生植物25%的光合作用(Edwards et al., 2010).C4植物光合优势在于其气孔数量多, CO2补偿点低, 高温或高辐照条件下较低的光呼吸(Li et al., 1999)以及高氮素利用效率(Brown, 1978; Sage and Pearcy, 1987; Oaks 1994; 何新华等, 1995).例如, 莎草属(Cyperus)植物高NUE的原因在于其高效率的Rubisco光合酶(Sage et al., 1999).C4植物总Rubisco基因表达较C3植物大幅下调, 导致Rubisco蛋白及Calvin-Benson循环酶含量减少(不包括将3-PGA还原为磷酸丙糖酶) (Wessinger et al., 1989). ...
... 通过遗传或基因工程技术向C3作物转入C4作物高光合效率相关基因是提高作物产量和保障粮食安全的有效途径之一(Hibberd et al., 2008; Peterhansel et al., 2013).利用基因工程和转基因技术提高Rubisco羧化酶反应效率(Ku et al., 1996, 1999; Tamoi and Shigeoka, 2005; Suzuki et al., 2009), 将C4途径相关酶基因转入C3植物, 通过超量表达C4途径相关核心酶, 在C3植物内建立类似C4植物特性的CCM (CO2 concentrating mechanism)单细胞循环等方面已进行了大量研究(陈太钰, 2012; Ruan et al., 2012; Hagemann and Bauwe, 2016). ...
C4 plants 1 2001
... 与C3植物相比, C4植物蛋白质合成相关基因的表达也相应较低(Bräutigam et al., 2011, 2014; Gowik et al., 2011).Bräutigam和Gowik (2016)认为, 基因表达量降低和叶片蛋白质含量减少使一些C4植物具有更高的氮素利用效率, 其原因是Rubisco在光合作用和氮素利用与平衡方面发挥重要作用(Spreitzer and Salvucci, 2002; Nowicka et al., 2018).其一, 核酮糖-1,5-二磷酸(RuBP)的氧化反应生成3-磷酸甘油酸(3-PGA)和2-磷酸甘油酸(2-PG), 光呼吸将2个2-PGA分子循环为1个3-PGA分子, 因此有25%的有机碳以CO2的形式流失(Sage et al., 2012; Betti et al., 2016; Hagemann and Bauwe, 2016).在主要C3作物(如小麦)中, 每3-4分子RuBP中就有1个被氧化为3-PGA和2-PG (Hagemann and Bauwe, 2016), 而2-PG是光合碳代谢酶抑制剂; C4植物的Rubisco加氧酶活性较低, Rubisco羧化酶代谢循环具有高效率优势(Sage, 2001).光呼吸需要大量的能量输入, 导致同化碳和氮分别以CO2和NH3形式流失(Peterhansel et al., 2013).其二, 制约Rubisco缓慢的羧化反应, 其周转率常在1-10次·s-1之间, 而其它中央代谢酶的周转频率在50-100次·s-1之间(Bar-Even et al., 2011).为了弥补Rubisco的低活性, C3植物产生大量Rubisco, 占光合生物可溶性蛋白质的50% (Erb and Zarzycki, 2016).而高光合效率所需的Rubisco更少, 通常C4植物的Rubisco含量占可溶性蛋白总量的10%-25% (Carmo-Silva et al., 2015).C3植物对Rubisco的高氮投入会导致其氮素利用效率(生物产量与投入氮的比值)较低.因此, 提高Rubisco利用效率(RBUE)是C3植物增强光合氮素利用效率的主要驱动因素(Hikosaka and Shigeno, 2009). ...
The C4 plant lineages of planet earth 2 2011
... 与C3植物相比, C4植物蛋白质合成相关基因的表达也相应较低(Bräutigam et al., 2011, 2014; Gowik et al., 2011).Bräutigam和Gowik (2016)认为, 基因表达量降低和叶片蛋白质含量减少使一些C4植物具有更高的氮素利用效率, 其原因是Rubisco在光合作用和氮素利用与平衡方面发挥重要作用(Spreitzer and Salvucci, 2002; Nowicka et al., 2018).其一, 核酮糖-1,5-二磷酸(RuBP)的氧化反应生成3-磷酸甘油酸(3-PGA)和2-磷酸甘油酸(2-PG), 光呼吸将2个2-PGA分子循环为1个3-PGA分子, 因此有25%的有机碳以CO2的形式流失(Sage et al., 2012; Betti et al., 2016; Hagemann and Bauwe, 2016).在主要C3作物(如小麦)中, 每3-4分子RuBP中就有1个被氧化为3-PGA和2-PG (Hagemann and Bauwe, 2016), 而2-PG是光合碳代谢酶抑制剂; C4植物的Rubisco加氧酶活性较低, Rubisco羧化酶代谢循环具有高效率优势(Sage, 2001).光呼吸需要大量的能量输入, 导致同化碳和氮分别以CO2和NH3形式流失(Peterhansel et al., 2013).其二, 制约Rubisco缓慢的羧化反应, 其周转率常在1-10次·s-1之间, 而其它中央代谢酶的周转频率在50-100次·s-1之间(Bar-Even et al., 2011).为了弥补Rubisco的低活性, C3植物产生大量Rubisco, 占光合生物可溶性蛋白质的50% (Erb and Zarzycki, 2016).而高光合效率所需的Rubisco更少, 通常C4植物的Rubisco含量占可溶性蛋白总量的10%-25% (Carmo-Silva et al., 2015).C3植物对Rubisco的高氮投入会导致其氮素利用效率(生物产量与投入氮的比值)较低.因此, 提高Rubisco利用效率(RBUE)是C3植物增强光合氮素利用效率的主要驱动因素(Hikosaka and Shigeno, 2009). ...
... 在正常生长条件下, 光呼吸损失的CO2占植物固定CO2总量的25% (Sage et al., 2012).Walker等(2016)利用模型分析显示, 在未来大气CO2浓度达到预测最高水平时, 植物光呼吸缺失下其光合速率可上调12%-55%.大气中CO2浓度升高导致C3植物光呼吸速率和NO3-同化速率同时下降, 吸收的NO3-不能及时被同化利用, 导致作物品质下降(Long et al., 2006; Bloom et al., 2010, 2014), 这也是植物在CO2浓度升高的条件下生长速率增幅较低的原因(Bloom et al., 2012).光合暗反应中, 光呼吸降低使ATP对NADPH的需求比例也降低.基质中CO2的增加导致NADP/ NADPH升高, 从而限制了NO3-代谢的还原能力.在高浓度CO2下, 当氮素供应不足时, C3植物(小麦和拟南芥(Arabidopsis thaliana))地上部分氮素含量的下降比氮素供应充足时更为显著(Bloom et al., 2010).Zeng等(2011)在白天CO2浓度为(374±21) mmol∙L-1和(568±39) mmol∙L-1, 夜间为(418±29) mmol∙L-1和(623±51) mmol∙L-1的农业FACE (free air CO2 enrichment)系统下研究CO2升高对C3植物水稻与C4植物稗草(Echinochloa crusgalli)氮素利用的影响, 发现其NUE增加, 在有限氮素下高浓度CO2使水稻比稗草具有更强的竞争优势.Reich等(2018)在长达20年的野外试验中发现, C3和C4植物在前12年对高浓度CO2的反应与前人研究结果一致, 在随后的8年里发生了意想不到的逆转.CO2浓度升高背景下, C4草地净氮矿化呈现随时间变化而逐渐增大的正响应, 而C3草地呈逐渐减小的负响应; 生物量对CO2升高的响应均与净氮矿化速率呈正相关. ...
The nitrogen use efficiency of C3 and C4 plants. I. Leaf nitrogen, growth, and biomass partitioning in Chenopodium album(L.) and Amaranthus retroflexus 2 1987
... 氮素是维持植物生长发育最主要的元素之一, 提高作物的氮素利用效率(nitrogen use efficiency, NUE)是保障粮食安全, 实现“资源节约和环境友好”的重要途径(徐晓鹏等, 2016).根据叶片解剖结构、植物碳代谢方式以及生物化学代谢差异, Brown (1978)提出C4比C3植物氮素利用效率高的假说.20世纪80-90年代在室内人工控制条件下佐证了这一假说的正确性(Sage and Pearcy, 1987; Oaks, 1994; He et al., 1994; 何新华等, 1995).除氮素利用效率在碳代谢上的差异外, NUE差异的关键在于氮代谢特征, 即氮素还原、同化酶类在细胞内发挥功能的位置 以及对氮素吸收、还原、同化的机制不同.何新华等(1995)认为, C3和C4植物氮素利用效率差异由特殊基因控制, 是长期进化的结果.然而, 目前对C3和C4植物氮素利用效率差异机制的进展尚未见系统梳理.本文综述C3和C4植物对氮素利用效率的差异机制、成因及C3作物氮素利用效率遗传改良技术, 旨在为合理利用氮素, 减少农业面源污染提供科学依据. ...
... C4植物光合作用机制是被子植物长期适应环境进化而成(Sage et al., 2011).与C3光合机理相比, C4光合作用进化出了复杂的叶片结构和生化适应机制(Christin and Osborne, 2014).近7 500种C4植物隶属于62个不同进化区系(Sage et al., 2011), 虽然C4植物在数量上只占陆地植物的3%, 却完成了陆生植物25%的光合作用(Edwards et al., 2010).C4植物光合优势在于其气孔数量多, CO2补偿点低, 高温或高辐照条件下较低的光呼吸(Li et al., 1999)以及高氮素利用效率(Brown, 1978; Sage and Pearcy, 1987; Oaks 1994; 何新华等, 1995).例如, 莎草属(Cyperus)植物高NUE的原因在于其高效率的Rubisco光合酶(Sage et al., 1999).C4植物总Rubisco基因表达较C3植物大幅下调, 导致Rubisco蛋白及Calvin-Benson循环酶含量减少(不包括将3-PGA还原为磷酸丙糖酶) (Wessinger et al., 1989). ...
Photorespiration and the evolution of C4 photosynthesis 2 2012
Rubisco: structure, regulatory interactions, and possibilities for a better enzyme 2 2002
... 与C3植物相比, C4植物蛋白质合成相关基因的表达也相应较低(Bräutigam et al., 2011, 2014; Gowik et al., 2011).Bräutigam和Gowik (2016)认为, 基因表达量降低和叶片蛋白质含量减少使一些C4植物具有更高的氮素利用效率, 其原因是Rubisco在光合作用和氮素利用与平衡方面发挥重要作用(Spreitzer and Salvucci, 2002; Nowicka et al., 2018).其一, 核酮糖-1,5-二磷酸(RuBP)的氧化反应生成3-磷酸甘油酸(3-PGA)和2-磷酸甘油酸(2-PG), 光呼吸将2个2-PGA分子循环为1个3-PGA分子, 因此有25%的有机碳以CO2的形式流失(Sage et al., 2012; Betti et al., 2016; Hagemann and Bauwe, 2016).在主要C3作物(如小麦)中, 每3-4分子RuBP中就有1个被氧化为3-PGA和2-PG (Hagemann and Bauwe, 2016), 而2-PG是光合碳代谢酶抑制剂; C4植物的Rubisco加氧酶活性较低, Rubisco羧化酶代谢循环具有高效率优势(Sage, 2001).光呼吸需要大量的能量输入, 导致同化碳和氮分别以CO2和NH3形式流失(Peterhansel et al., 2013).其二, 制约Rubisco缓慢的羧化反应, 其周转率常在1-10次·s-1之间, 而其它中央代谢酶的周转频率在50-100次·s-1之间(Bar-Even et al., 2011).为了弥补Rubisco的低活性, C3植物产生大量Rubisco, 占光合生物可溶性蛋白质的50% (Erb and Zarzycki, 2016).而高光合效率所需的Rubisco更少, 通常C4植物的Rubisco含量占可溶性蛋白总量的10%-25% (Carmo-Silva et al., 2015).C3植物对Rubisco的高氮投入会导致其氮素利用效率(生物产量与投入氮的比值)较低.因此, 提高Rubisco利用效率(RBUE)是C3植物增强光合氮素利用效率的主要驱动因素(Hikosaka and Shigeno, 2009). ...
... 除C4核心酶CA、NADP-MDH、NADP-ME、PEPC和PPDK外, C4途径还需要将草酸、苹果酸、三磷酸甘油酸和丙酮酸的代谢物转运体插入C3植物内, 以增加C4循环中间体的运输能力, 使卡尔文循环能够在维管束鞘细胞中有效运作(Weber and von Caemmerer, 2010).由于C3植物Rubisco具有高含量、低催化速率的特点, 因此通过在植物体内低量表达外源高催化速率Rubisco既可以提高光合作用效率也可增加氮素利用效率(Spreitzer and Salvucci, 2002; Whitney et al., 2011).国际水稻联盟也在研究与Kranz解剖结构有关的基因(Wang et al., 2013), 发现减少叶肉细胞光呼吸也是提高氮素利用率的途径之一(图2).例如, 降低叶肉细胞中的甘氨酸脱羧酶(Glycine decarboxylase complex, GDC)蛋白, 并限制其在维管束鞘细胞中的积累, 使甘氨酸脱羧完全发生在维管束鞘细胞中, 类似于C3-C4中间体, 从而维持较高的CO2浓度以降低光呼吸(Monson and Rawsthorne, 2000).Kajala等(2011)通过人工microRNAs技术下调水稻中Rubisco小亚基(RbcS)和GDC-H亚基基因的表达, 使叶肉细胞中Rubisco和甘氨酸脱羧酶(GDC)得到下调.通过对玉米的维管束鞘细胞和叶肉细胞进行蛋白质组学鉴定, 发现在C4代谢途径中过表达转运蛋白, 如2-氧戊二酸/苹果酸转运体(OMT1)、二羧酸转运体1 (DiT1)、二羧酸转运体2 (DiT2)、PEP/磷酸盐转运蛋白(PPT1)、叶肉包膜蛋白(MEP)和三糖-磷酸盐转运体(TPT), 转入水稻后均得到特异性表达(Karki et al., 2013).对氮素循环再利用也是提高NUE的有效途径.例如, 过表达自噬核心基因OsATG8b能够显著增强缺氮胁迫下转基因拟南芥叶片中的自噬活性, 从而提高氮素再利用效率(甄晓溪等, 2019). ...
Carbon metabolism in transgenic rice plants that express phospho enolpyruvate carboxylase and/or phosphoenolpyruvate carboxykinase 1 2006
... 利用DNA重组改良C3作物的技术体系已逐渐成熟.目前, 已从玉米和高粱(Sorghum bicolor)等C4植物中克隆到与C4光合相关的关键酶基因, 如NADP- ME、PCK、PEPC和PPDK, 且这些基因已成功转入水稻(Bandyopadhyay et al., 2007)和小麦(张庆琛等, 2010)等C3作物中.将水稻Cab启动子控制的玉米NADP-ME cDNA转入水稻, 使得水稻叶片NADP- ME活性增加了30-70倍, 但NADP-ME蛋白量也随之增加(Tsuchida et al., 2001).转玉米PEPC基因的水稻PEPC活性远高于野生型水稻, 与叶片光合速率和叶绿素均呈正相关(李霞等, 2008).转入外源玉米PEPC基因的水稻植株含氮量显著增加, 并在低氮条件下影响氨基酸的合成和有机酸含量, 促进氮代谢和氮同化产物的积累和植株生长(陈冉冉, 2012).转入碳相关基因, 对水稻的氮代谢相关酶有显著影响, 这为氮高效利用提供了新的思路(Yanagisawa et al., 2004; Kurai et al., 2011).研究结果与Melzer和O'Leary (1987)提出的将PEPC基因转入C3植物后可能存在代谢通路相一致, 即PEPC催化PEP与CO2反应形成草酰乙酸(oxaloacetic acid, OAA), 一部分OAA经苹果酸脱氢酶(MDH)催化脱氢产生的苹果酸和OAA进入三羧酸循环(tricarboxylic acid cycle, TCA), 增强呼吸作用的同时为氨基酸合成提供丰富的有机酸和碳骨架; 另一部分OAA经转氨酶作用形成天门冬氨酸.而PEPC催化CO2与PEP反应生成的OAA进入TCA, OAA与苹果酸相互转化, 及时补充合成氨基酸消耗的碳骨架, 进而促进氨基酸的合成, 提高氮素利用率(Ku et al., 1996; Miyao and Fukayama, 2003; Suzuki et al., 2006; Radchuk et al., 2007). ...
Rubisco content and photosynthesis of leaves at different positions in transgenic rice with an overexpression of RBCS 1 2009
... 通过遗传或基因工程技术向C3作物转入C4作物高光合效率相关基因是提高作物产量和保障粮食安全的有效途径之一(Hibberd et al., 2008; Peterhansel et al., 2013).利用基因工程和转基因技术提高Rubisco羧化酶反应效率(Ku et al., 1996, 1999; Tamoi and Shigeoka, 2005; Suzuki et al., 2009), 将C4途径相关酶基因转入C3植物, 通过超量表达C4途径相关核心酶, 在C3植物内建立类似C4植物特性的CCM (CO2 concentrating mechanism)单细胞循环等方面已进行了大量研究(陈太钰, 2012; Ruan et al., 2012; Hagemann and Bauwe, 2016). ...
Increased Rubisco content in transgenic rice transformed with the 'Sense' rbcS gene 1 2007
... C4植物氮素还原同化酶的空间协同效应是C4比C3植物氮素利用效率高的主要原因(图1A, B).C4植物在结构上区别于C3植物的典型特征是拥有Kranz结构的维管束鞘细胞(巩玥等, 2014).C4植物既可以在叶肉细胞, 也可在维管束鞘细胞进行NH4+的同化, 合成氨基酸和蛋白质(何新华等, 1995); 而C3植物氮素还原只在叶肉细胞进行, 导致其NUE低.采用蛋白质组学研究玉米(Zea mays)叶肉细胞和维管束鞘细胞叶绿体(膜)上蛋白质表达差异, 结果表明, 在C4植物中氮同化(如氨基酸合成)在叶肉和维管束鞘细胞之间是被分隔开的(Majeran and Wijk, 2009), 与氮吸收相关酶类主要存在于叶肉细胞的叶绿体中, 而淀粉合成相关酶类主要存在于维管束鞘细胞的叶绿体中(Majeran et al., 2005; Suzuki et al., 2007). ...
Improvement of photosynthesis in higher plants 1 2005
... 通过遗传或基因工程技术向C3作物转入C4作物高光合效率相关基因是提高作物产量和保障粮食安全的有效途径之一(Hibberd et al., 2008; Peterhansel et al., 2013).利用基因工程和转基因技术提高Rubisco羧化酶反应效率(Ku et al., 1996, 1999; Tamoi and Shigeoka, 2005; Suzuki et al., 2009), 将C4途径相关酶基因转入C3植物, 通过超量表达C4途径相关核心酶, 在C3植物内建立类似C4植物特性的CCM (CO2 concentrating mechanism)单细胞循环等方面已进行了大量研究(陈太钰, 2012; Ruan et al., 2012; Hagemann and Bauwe, 2016). ...
Ecophysiological traits in C3 and C4 grasses: a phylogenetically controlled screening experiment 1 2010
... 在当前CO2浓度条件下, C4植物比C3植物有更高的NUE (Brown, 1978; Taylor et al., 2010).一般而言, 低浓度CO2会降低C3植物的光合速率, 使气孔导度和叶片氮浓度增加(Tissue et al., 1995; Ward et al., 1999; Ripley et al., 2013).Pinto等(2014)研究了C3和C4植物在冰川CO2 浓度为180 mmol∙L-1及环境CO2浓度为400 mmol∙L-1下的光合作用, 结果表明, 与C3植物相比, 冰川较低CO2浓度(180 mmol∙L-1)导致C4植物的光合作用降低幅度较小, 气孔导度增加较大.C3植物通过增加Rubisco活性来适应低浓度CO2, C4植物在低CO2浓度下有较高的气孔导度, 但由于光合速率较高, C4植物有较高的PNUE.Vogan和Sage (2012)研究了黄花菊属(Flaveria)、天芥菜属(Heliotropium)和蓬子草属(Alternanthera)中近缘C3 (2种)、C3-C4 (9种)和C4 (3种)植物对低浓度CO2的响应, 发现在380和180 mmol∙L-1 CO2下, 昼/夜气温为37/29°C时, CO2对任何一种植物的光合氮素利用率、Rubisco含量和电子传递均无显著影响. ...
Effects of low and elevated CO2 on C3 and C4 annuals 1 1995
... 在当前CO2浓度条件下, C4植物比C3植物有更高的NUE (Brown, 1978; Taylor et al., 2010).一般而言, 低浓度CO2会降低C3植物的光合速率, 使气孔导度和叶片氮浓度增加(Tissue et al., 1995; Ward et al., 1999; Ripley et al., 2013).Pinto等(2014)研究了C3和C4植物在冰川CO2 浓度为180 mmol∙L-1及环境CO2浓度为400 mmol∙L-1下的光合作用, 结果表明, 与C3植物相比, 冰川较低CO2浓度(180 mmol∙L-1)导致C4植物的光合作用降低幅度较小, 气孔导度增加较大.C3植物通过增加Rubisco活性来适应低浓度CO2, C4植物在低CO2浓度下有较高的气孔导度, 但由于光合速率较高, C4植物有较高的PNUE.Vogan和Sage (2012)研究了黄花菊属(Flaveria)、天芥菜属(Heliotropium)和蓬子草属(Alternanthera)中近缘C3 (2种)、C3-C4 (9种)和C4 (3种)植物对低浓度CO2的响应, 发现在380和180 mmol∙L-1 CO2下, 昼/夜气温为37/29°C时, CO2对任何一种植物的光合氮素利用率、Rubisco含量和电子传递均无显著影响. ...
High level expression of C4-specific NADP-malic enzyme in leaves and impairment of photoautotrophic growth in a C3 plant, rice 1 2001
... 利用DNA重组改良C3作物的技术体系已逐渐成熟.目前, 已从玉米和高粱(Sorghum bicolor)等C4植物中克隆到与C4光合相关的关键酶基因, 如NADP- ME、PCK、PEPC和PPDK, 且这些基因已成功转入水稻(Bandyopadhyay et al., 2007)和小麦(张庆琛等, 2010)等C3作物中.将水稻Cab启动子控制的玉米NADP-ME cDNA转入水稻, 使得水稻叶片NADP- ME活性增加了30-70倍, 但NADP-ME蛋白量也随之增加(Tsuchida et al., 2001).转玉米PEPC基因的水稻PEPC活性远高于野生型水稻, 与叶片光合速率和叶绿素均呈正相关(李霞等, 2008).转入外源玉米PEPC基因的水稻植株含氮量显著增加, 并在低氮条件下影响氨基酸的合成和有机酸含量, 促进氮代谢和氮同化产物的积累和植株生长(陈冉冉, 2012).转入碳相关基因, 对水稻的氮代谢相关酶有显著影响, 这为氮高效利用提供了新的思路(Yanagisawa et al., 2004; Kurai et al., 2011).研究结果与Melzer和O'Leary (1987)提出的将PEPC基因转入C3植物后可能存在代谢通路相一致, 即PEPC催化PEP与CO2反应形成草酰乙酸(oxaloacetic acid, OAA), 一部分OAA经苹果酸脱氢酶(MDH)催化脱氢产生的苹果酸和OAA进入三羧酸循环(tricarboxylic acid cycle, TCA), 增强呼吸作用的同时为氨基酸合成提供丰富的有机酸和碳骨架; 另一部分OAA经转氨酶作用形成天门冬氨酸.而PEPC催化CO2与PEP反应生成的OAA进入TCA, OAA与苹果酸相互转化, 及时补充合成氨基酸消耗的碳骨架, 进而促进氨基酸的合成, 提高氮素利用率(Ku et al., 1996; Miyao and Fukayama, 2003; Suzuki et al., 2006; Radchuk et al., 2007). ...
Effects of low atmospheric CO2 and elevated temperature during growth on the gas exchange responses of C3, C3-C4 intermediate, and C4 species from three evolutionary lineages of C4 photosynthesis 1 2012
... 在当前CO2浓度条件下, C4植物比C3植物有更高的NUE (Brown, 1978; Taylor et al., 2010).一般而言, 低浓度CO2会降低C3植物的光合速率, 使气孔导度和叶片氮浓度增加(Tissue et al., 1995; Ward et al., 1999; Ripley et al., 2013).Pinto等(2014)研究了C3和C4植物在冰川CO2 浓度为180 mmol∙L-1及环境CO2浓度为400 mmol∙L-1下的光合作用, 结果表明, 与C3植物相比, 冰川较低CO2浓度(180 mmol∙L-1)导致C4植物的光合作用降低幅度较小, 气孔导度增加较大.C3植物通过增加Rubisco活性来适应低浓度CO2, C4植物在低CO2浓度下有较高的气孔导度, 但由于光合速率较高, C4植物有较高的PNUE.Vogan和Sage (2012)研究了黄花菊属(Flaveria)、天芥菜属(Heliotropium)和蓬子草属(Alternanthera)中近缘C3 (2种)、C3-C4 (9种)和C4 (3种)植物对低浓度CO2的响应, 发现在380和180 mmol∙L-1 CO2下, 昼/夜气温为37/29°C时, CO2对任何一种植物的光合氮素利用率、Rubisco含量和电子传递均无显著影响. ...
Proof of C4 photosynthesis without Kranz anatomy in Bienertia cycloptera(Chenopodiaceae) 1 2002
Kranz anatomy is not essential for terrestrial C4 plant photosynthesis 1 2001
... 氮素的吸收、同化和再分配在一定程度上受光合速率的调节和控制(Zheng, 2009), 因此, 除非光合速率也提高, 否则NUE会趋于平稳.之前研究认为, 实现C4高光效要具有Kranz结构(Sage and Monson, 1999), 但C4植物Kranz结构并非C4光合所必需(Voznesenskaya et al., 2001).因此研究自然条件下植物光合途径的转变机制应侧重光合酶活性, 而不是Kranz结构的改变(图2). ...
The costs of photorespiration to food production now and in the future 1 2016
... 在正常生长条件下, 光呼吸损失的CO2占植物固定CO2总量的25% (Sage et al., 2012).Walker等(2016)利用模型分析显示, 在未来大气CO2浓度达到预测最高水平时, 植物光呼吸缺失下其光合速率可上调12%-55%.大气中CO2浓度升高导致C3植物光呼吸速率和NO3-同化速率同时下降, 吸收的NO3-不能及时被同化利用, 导致作物品质下降(Long et al., 2006; Bloom et al., 2010, 2014), 这也是植物在CO2浓度升高的条件下生长速率增幅较低的原因(Bloom et al., 2012).光合暗反应中, 光呼吸降低使ATP对NADPH的需求比例也降低.基质中CO2的增加导致NADP/ NADPH升高, 从而限制了NO3-代谢的还原能力.在高浓度CO2下, 当氮素供应不足时, C3植物(小麦和拟南芥(Arabidopsis thaliana))地上部分氮素含量的下降比氮素供应充足时更为显著(Bloom et al., 2010).Zeng等(2011)在白天CO2浓度为(374±21) mmol∙L-1和(568±39) mmol∙L-1, 夜间为(418±29) mmol∙L-1和(623±51) mmol∙L-1的农业FACE (free air CO2 enrichment)系统下研究CO2升高对C3植物水稻与C4植物稗草(Echinochloa crusgalli)氮素利用的影响, 发现其NUE增加, 在有限氮素下高浓度CO2使水稻比稗草具有更强的竞争优势.Reich等(2018)在长达20年的野外试验中发现, C3和C4植物在前12年对高浓度CO2的反应与前人研究结果一致, 在随后的8年里发生了意想不到的逆转.CO2浓度升高背景下, C4草地净氮矿化呈现随时间变化而逐渐增大的正响应, 而C3草地呈逐渐减小的负响应; 生物量对CO2升高的响应均与净氮矿化速率呈正相关. ...
Genome-wide transcript analysis of early maize leaf development reveals gene cohorts associated with the differentiation of C4 Kranz anatomy 1 2013
... 除C4核心酶CA、NADP-MDH、NADP-ME、PEPC和PPDK外, C4途径还需要将草酸、苹果酸、三磷酸甘油酸和丙酮酸的代谢物转运体插入C3植物内, 以增加C4循环中间体的运输能力, 使卡尔文循环能够在维管束鞘细胞中有效运作(Weber and von Caemmerer, 2010).由于C3植物Rubisco具有高含量、低催化速率的特点, 因此通过在植物体内低量表达外源高催化速率Rubisco既可以提高光合作用效率也可增加氮素利用效率(Spreitzer and Salvucci, 2002; Whitney et al., 2011).国际水稻联盟也在研究与Kranz解剖结构有关的基因(Wang et al., 2013), 发现减少叶肉细胞光呼吸也是提高氮素利用率的途径之一(图2).例如, 降低叶肉细胞中的甘氨酸脱羧酶(Glycine decarboxylase complex, GDC)蛋白, 并限制其在维管束鞘细胞中的积累, 使甘氨酸脱羧完全发生在维管束鞘细胞中, 类似于C3-C4中间体, 从而维持较高的CO2浓度以降低光呼吸(Monson and Rawsthorne, 2000).Kajala等(2011)通过人工microRNAs技术下调水稻中Rubisco小亚基(RbcS)和GDC-H亚基基因的表达, 使叶肉细胞中Rubisco和甘氨酸脱羧酶(GDC)得到下调.通过对玉米的维管束鞘细胞和叶肉细胞进行蛋白质组学鉴定, 发现在C4代谢途径中过表达转运蛋白, 如2-氧戊二酸/苹果酸转运体(OMT1)、二羧酸转运体1 (DiT1)、二羧酸转运体2 (DiT2)、PEP/磷酸盐转运蛋白(PPT1)、叶肉包膜蛋白(MEP)和三糖-磷酸盐转运体(TPT), 转入水稻后均得到特异性表达(Karki et al., 2013).对氮素循环再利用也是提高NUE的有效途径.例如, 过表达自噬核心基因OsATG8b能够显著增强缺氮胁迫下转基因拟南芥叶片中的自噬活性, 从而提高氮素再利用效率(甄晓溪等, 2019). ...
Comparative responses of model C3 and C4 plants to drought in low and elevated CO2 1 1999
... 在当前CO2浓度条件下, C4植物比C3植物有更高的NUE (Brown, 1978; Taylor et al., 2010).一般而言, 低浓度CO2会降低C3植物的光合速率, 使气孔导度和叶片氮浓度增加(Tissue et al., 1995; Ward et al., 1999; Ripley et al., 2013).Pinto等(2014)研究了C3和C4植物在冰川CO2 浓度为180 mmol∙L-1及环境CO2浓度为400 mmol∙L-1下的光合作用, 结果表明, 与C3植物相比, 冰川较低CO2浓度(180 mmol∙L-1)导致C4植物的光合作用降低幅度较小, 气孔导度增加较大.C3植物通过增加Rubisco活性来适应低浓度CO2, C4植物在低CO2浓度下有较高的气孔导度, 但由于光合速率较高, C4植物有较高的PNUE.Vogan和Sage (2012)研究了黄花菊属(Flaveria)、天芥菜属(Heliotropium)和蓬子草属(Alternanthera)中近缘C3 (2种)、C3-C4 (9种)和C4 (3种)植物对低浓度CO2的响应, 发现在380和180 mmol∙L-1 CO2下, 昼/夜气温为37/29°C时, CO2对任何一种植物的光合氮素利用率、Rubisco含量和电子传递均无显著影响. ...
Plastid transport and metabolism of C3 and C4 plants-comparative analysis and possible biotechnological exploitation 1 2010
... 除C4核心酶CA、NADP-MDH、NADP-ME、PEPC和PPDK外, C4途径还需要将草酸、苹果酸、三磷酸甘油酸和丙酮酸的代谢物转运体插入C3植物内, 以增加C4循环中间体的运输能力, 使卡尔文循环能够在维管束鞘细胞中有效运作(Weber and von Caemmerer, 2010).由于C3植物Rubisco具有高含量、低催化速率的特点, 因此通过在植物体内低量表达外源高催化速率Rubisco既可以提高光合作用效率也可增加氮素利用效率(Spreitzer and Salvucci, 2002; Whitney et al., 2011).国际水稻联盟也在研究与Kranz解剖结构有关的基因(Wang et al., 2013), 发现减少叶肉细胞光呼吸也是提高氮素利用率的途径之一(图2).例如, 降低叶肉细胞中的甘氨酸脱羧酶(Glycine decarboxylase complex, GDC)蛋白, 并限制其在维管束鞘细胞中的积累, 使甘氨酸脱羧完全发生在维管束鞘细胞中, 类似于C3-C4中间体, 从而维持较高的CO2浓度以降低光呼吸(Monson and Rawsthorne, 2000).Kajala等(2011)通过人工microRNAs技术下调水稻中Rubisco小亚基(RbcS)和GDC-H亚基基因的表达, 使叶肉细胞中Rubisco和甘氨酸脱羧酶(GDC)得到下调.通过对玉米的维管束鞘细胞和叶肉细胞进行蛋白质组学鉴定, 发现在C4代谢途径中过表达转运蛋白, 如2-氧戊二酸/苹果酸转运体(OMT1)、二羧酸转运体1 (DiT1)、二羧酸转运体2 (DiT2)、PEP/磷酸盐转运蛋白(PPT1)、叶肉包膜蛋白(MEP)和三糖-磷酸盐转运体(TPT), 转入水稻后均得到特异性表达(Karki et al., 2013).对氮素循环再利用也是提高NUE的有效途径.例如, 过表达自噬核心基因OsATG8b能够显著增强缺氮胁迫下转基因拟南芥叶片中的自噬活性, 从而提高氮素再利用效率(甄晓溪等, 2019). ...
Quantity and kinetic properties of ribulose 1,5-bisphosphate carboxylase in C3, C4, and C3-C4 intermediate species of flaveria (asteraceae) 1 1989
... C4植物光合作用机制是被子植物长期适应环境进化而成(Sage et al., 2011).与C3光合机理相比, C4光合作用进化出了复杂的叶片结构和生化适应机制(Christin and Osborne, 2014).近7 500种C4植物隶属于62个不同进化区系(Sage et al., 2011), 虽然C4植物在数量上只占陆地植物的3%, 却完成了陆生植物25%的光合作用(Edwards et al., 2010).C4植物光合优势在于其气孔数量多, CO2补偿点低, 高温或高辐照条件下较低的光呼吸(Li et al., 1999)以及高氮素利用效率(Brown, 1978; Sage and Pearcy, 1987; Oaks 1994; 何新华等, 1995).例如, 莎草属(Cyperus)植物高NUE的原因在于其高效率的Rubisco光合酶(Sage et al., 1999).C4植物总Rubisco基因表达较C3植物大幅下调, 导致Rubisco蛋白及Calvin-Benson循环酶含量减少(不包括将3-PGA还原为磷酸丙糖酶) (Wessinger et al., 1989). ...
Isoleucine 309 acts as a C4 catalytic switch that increases ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) carboxylation rate in flaveria 1 2011
... 除C4核心酶CA、NADP-MDH、NADP-ME、PEPC和PPDK外, C4途径还需要将草酸、苹果酸、三磷酸甘油酸和丙酮酸的代谢物转运体插入C3植物内, 以增加C4循环中间体的运输能力, 使卡尔文循环能够在维管束鞘细胞中有效运作(Weber and von Caemmerer, 2010).由于C3植物Rubisco具有高含量、低催化速率的特点, 因此通过在植物体内低量表达外源高催化速率Rubisco既可以提高光合作用效率也可增加氮素利用效率(Spreitzer and Salvucci, 2002; Whitney et al., 2011).国际水稻联盟也在研究与Kranz解剖结构有关的基因(Wang et al., 2013), 发现减少叶肉细胞光呼吸也是提高氮素利用率的途径之一(图2).例如, 降低叶肉细胞中的甘氨酸脱羧酶(Glycine decarboxylase complex, GDC)蛋白, 并限制其在维管束鞘细胞中的积累, 使甘氨酸脱羧完全发生在维管束鞘细胞中, 类似于C3-C4中间体, 从而维持较高的CO2浓度以降低光呼吸(Monson and Rawsthorne, 2000).Kajala等(2011)通过人工microRNAs技术下调水稻中Rubisco小亚基(RbcS)和GDC-H亚基基因的表达, 使叶肉细胞中Rubisco和甘氨酸脱羧酶(GDC)得到下调.通过对玉米的维管束鞘细胞和叶肉细胞进行蛋白质组学鉴定, 发现在C4代谢途径中过表达转运蛋白, 如2-氧戊二酸/苹果酸转运体(OMT1)、二羧酸转运体1 (DiT1)、二羧酸转运体2 (DiT2)、PEP/磷酸盐转运蛋白(PPT1)、叶肉包膜蛋白(MEP)和三糖-磷酸盐转运体(TPT), 转入水稻后均得到特异性表达(Karki et al., 2013).对氮素循环再利用也是提高NUE的有效途径.例如, 过表达自噬核心基因OsATG8b能够显著增强缺氮胁迫下转基因拟南芥叶片中的自噬活性, 从而提高氮素再利用效率(甄晓溪等, 2019). ...
Metabolic engineering with dof1 transcription factor in plants: improved nitrogen assimilation and growth under low-nitrogen conditions 1 2004
... 利用DNA重组改良C3作物的技术体系已逐渐成熟.目前, 已从玉米和高粱(Sorghum bicolor)等C4植物中克隆到与C4光合相关的关键酶基因, 如NADP- ME、PCK、PEPC和PPDK, 且这些基因已成功转入水稻(Bandyopadhyay et al., 2007)和小麦(张庆琛等, 2010)等C3作物中.将水稻Cab启动子控制的玉米NADP-ME cDNA转入水稻, 使得水稻叶片NADP- ME活性增加了30-70倍, 但NADP-ME蛋白量也随之增加(Tsuchida et al., 2001).转玉米PEPC基因的水稻PEPC活性远高于野生型水稻, 与叶片光合速率和叶绿素均呈正相关(李霞等, 2008).转入外源玉米PEPC基因的水稻植株含氮量显著增加, 并在低氮条件下影响氨基酸的合成和有机酸含量, 促进氮代谢和氮同化产物的积累和植株生长(陈冉冉, 2012).转入碳相关基因, 对水稻的氮代谢相关酶有显著影响, 这为氮高效利用提供了新的思路(Yanagisawa et al., 2004; Kurai et al., 2011).研究结果与Melzer和O'Leary (1987)提出的将PEPC基因转入C3植物后可能存在代谢通路相一致, 即PEPC催化PEP与CO2反应形成草酰乙酸(oxaloacetic acid, OAA), 一部分OAA经苹果酸脱氢酶(MDH)催化脱氢产生的苹果酸和OAA进入三羧酸循环(tricarboxylic acid cycle, TCA), 增强呼吸作用的同时为氨基酸合成提供丰富的有机酸和碳骨架; 另一部分OAA经转氨酶作用形成天门冬氨酸.而PEPC催化CO2与PEP反应生成的OAA进入TCA, OAA与苹果酸相互转化, 及时补充合成氨基酸消耗的碳骨架, 进而促进氨基酸的合成, 提高氮素利用率(Ku et al., 1996; Miyao and Fukayama, 2003; Suzuki et al., 2006; Radchuk et al., 2007). ...
A comparison of the impacts of various nitrogen sources on acid-base balance in C3Triticum aestivum L.and C4Zea mays L. plants 1 1997
Influences of different nitrogen sources on nitrogen- and water-use efficiency, and carbon isotope discrimination, in C3Triticum aestivum L. and C4Zea mays L. plants. 3 1998
 ,2,3,*
,2,3,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT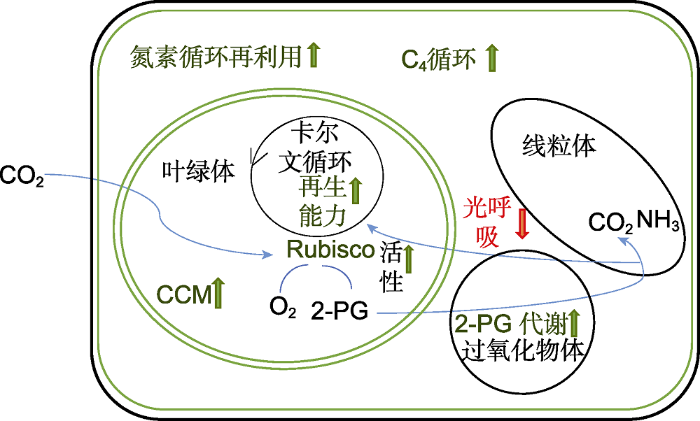 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}