A Non-invasive Method for Measuring and Analyzing Circadian Phenotype in Living Plants
Yingjun Yu1,2, Hang Xu1,2, Lei Wang,1,2,*1Key Laboratory of Plant Molecular Physiology, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 2University of Chinese Academy of Sciences, Beijing 100049, China
Abstract The plant circadian clock is a time-keeping molecular system, with a cycle of ~ 24 h. It was evolved to adapt to the diel rhythmic environmental cues generated by the self-rotation of the Earth. In addition to the time-keeping function, the circadian clock also regulates a plethora of plant growth and development processes by synchronizing the endogenous energy and metabolomic status. By sensing and integrating the dynamics of external environmental cues, circadian clock can coordinate gene expression at multiple levels, thus to increase the fitness of plants. Recently, there is an increasingly demand for measuring and assessing circadian phenotype for many non-circadian research field. Here, we summarized the currently available methods for detecting circadian rhythm, and show one most commonly used standard procedure for evaluating circadian phenotype in plants, which may help to provide the applicable technical assistance for the study of the circadian clock. Keywords:circadian clock;phenotype analysis;FFT-NLLS;BRASS
PDF (824KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 于英俊, 徐航, 王雷. 生物发光成像无损伤研究植物生物钟的方法. 植物学报, 2020, 55(2): 177-181 doi:10.11983/CBB19238 Yu Yingjun, Xu Hang, Wang Lei. A Non-invasive Method for Measuring and Analyzing Circadian Phenotype in Living Plants. Chinese Bulletin of Botany, 2020, 55(2): 177-181 doi:10.11983/CBB19238
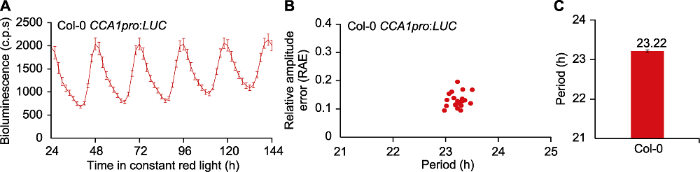
(A) 携带有CCA1pro:LUC报告基因的Col-0野生型拟南芥在持续红光条件下, 从LL24到LL144时间范围内生物发光的荧光节律(c.p.s代表每株植物每秒钟采集到的光子数)。(B) 野生型拟南芥生物钟周期的散点图, X轴表示生物钟周期, Y轴表示相对振幅误差; 根据结果显示每株幼苗周期较为集中, 相对一致, 而且相对振幅误差低于0.5, 结果可靠(一般认为当RAE值大于0.5时, 节律的振荡强度偏低, 不适于计算生物钟周期)。(C) 野生型拟南芥生物钟周期的柱状图; 根据结果显示周期长度在持续红光下为23.22小时。计算方法: FFT-NLLS。数据为21株幼苗的平均值±标准误。 Figure 1Circadian phenotype analysis of Col-0 wild type Arabidopsis thaliana CCA1pro:LUC under continuous red light
(A) Trace plot of bioluminescence in Col-0 wild type Arabidopsis thaliana with CCA1pro:LUC reporter under continuous red light conditions, from LL24 to LL144 (the abbreviation of c.p.s represents the counts of photons per seedling). (B) Scatter plot of circadian period (x-axis) and relative amplitude error (RAE) (y-axis) of Col-0; According to the results, the periods are more concentrated and relatively consistent, and the relative amplitude errors are all lower than 0.5, which indicates that the result is reliable (generally, when the RAE is greater than 0.5, the circadian oscillation intensity is relatively low, which is not suitable for calculating the period). (C) Estimated circadian period of CCA1pro:LUC in Col-0; According to the calculation, circadian period length of Col-0 is 23.22 hours under continuous red light. Calculation was performed using FFT-NLLS method. Data are presented as means±SE (n=21).
DoyleMR, DavisSJ, BastowRM, McWattersHG, Kozma-BognárL, NagyF, MillarAJ, AmasinoRM ( 2002). The ELF4 gene controls circadian rhythms and flowering time in Arabidopsis thaliana Nature 419, 74-77. [本文引用: 2]
EdwardsKD, MillarAJ ( 2007). Analysis of circadian leaf movement rhythms in Arabidopsis thaliana Methods Mol Biol 362, 103-113. [本文引用: 1]
KimJ, SomersDE ( 2010). Rapid assessment of gene function in the circadian clock using artificial microRNA in Arabidopsis mesophyll protoplasts Plant Physiol 154, 611-621. [本文引用: 1]
LiB, WangY, ZhangYY, TianWW, ChongK, JangJC, WangL ( 2019). PRR5, 7 and 9 positively modulate TOR signaling-mediated root cell proliferation by repressing TANDEM ZINC FINGER 1 in Arabidopsis Nucleic Acids Res 47, 5001-5015. [本文引用: 1]
MásP, AlabadiD, YanovskyMJ, OyamaT, KaySA ( 2003). Dual role of TOC1 in the control of circadian and photomorphogenic responses in Arabidopsis Plant Cell 15, 223-236. [本文引用: 1]
MüllerLM, von KorffM, DavisSJ ( 2014). Connections bet- ween circadian clocks and carbon metabolism reveal species-specific effects on growth control J Exp Bot 65, 2915-2923. [本文引用: 1]
MullerNA, Jimenez-GomezJM ( 2016). Analysis of circadian leaf movements. In: Duque P, ed. Environmental Responses in Plants Methods in Molecular Biology, Vol. 1398. New York: Humana Press. pp. 71-79. [本文引用: 1]
NakamichiN, ItoS, OyamaT, YamashinoT, KondoT, MizunoT ( 2004). Characterization of plant circadian rhythms by employing Arabidopsis cultured cells with bioluminescence reporters Plant Cell Physiol 45, 57-67. [本文引用: 1]
NiwaY, YamashinoT, MizunoT ( 2009). The circadian clock regulates the photoperiodic response of hypocotyl elongation through a coincidence mechanism in Arabidopsis thaliana Plant Cell Physiol 50, 838-854. [本文引用: 1]
RonaldJ, DavisSJ ( 2017). Making the clock tick: the transcriptional landscape of the plant circadian clock F1000Res 6, 951. [本文引用: 1]
SaloméPA, MichaelTP, KearnsEV, Fett-NetoAG, SharrockRA, McClungCR ( 2002). The out of phase 1 mutant defines a role for PHYB in circadian phase control in Arabidopsis Plant Physiol 129, 1674-1685. [本文引用: 1]
SanchezA, ShinJ, DavisSJ ( 2011). Abiotic stress and the plant circadian clock Plant Signal Behav 6, 223-231. [本文引用: 1]
SomersDE, WebbAA, PearsonM, KaySA ( 1998). The short-period mutant, toc1-1, alters circadian clock regulation of multiple outputs throughout development in Arabidopsis thaliana Development 125, 485-494. [本文引用: 1]
UehleinN, KaldenhoffR ( 2008). Aquaporins and plant leaf movements Ann Bot 101, 1-4. [本文引用: 1]
WangL, KimJ, SomersDE ( 2013). Transcriptional corepressor TOPLESS complexes with pseudoresponse regulator proteins and histone deacetylases to regulate circadian transcription Proc Natl Acad Sci USA 110, 761-766. [本文引用: 1]
YanovskyMJ, IzaguirreM, WagmaisterJA, GatzC, JacksonSD, ThomasB, CasalJJ ( 2000). Phytochrome A resets the circadian clock and delays tuber formation under long days in potato Plant J 23, 223-232. [本文引用: 1]
ZhangYY, BoCP, WangL ( 2019). Novel crosstalks between circadian clock and jasmonic acid pathway finely coordinate the tradeoff among plant growth, senescence and defense Int J Mol Sci 20, 5254. [本文引用: 1]
ZielinskiT, MooreAM, TroupE, HallidayKJ, MillarAJ ( 2014). Strengths and limitations of period estimation methods for circadian data PLoS One 9, e96462. [本文引用: 1]
植物生物钟及其调控生长发育的研究进展 1 2018
... 地球自转引起光照和温度等诸多环境因子昼夜周期变化.为了更好地应对变化的环境信号, 生物体进化出内源性生物钟系统来协调环境变化与自身生长发育.不同物种中生物钟系统关键组分虽有明显差异, 但均包含一些共同特征: (1) 具有内源性, 在持续光照或恒定温度条件下仍能维持近24小时的节律性; (2) 具有可驯化性, 如改变光周期, 生物钟可被重新设定来适应新的生存环境; (3) 具有温度补偿现象, 即在一定温度范围内, 生物钟系统仍能维持周期基本稳定(不被改变) (谢启光和徐小冬, 2015).植物生物钟参与调控植物多种生命活动, 如激素信号(Covington and Harmer, 2007)、叶片衰老(Zhang et al., 2018, 2019)、下胚轴伸长(Niwa et al., 2009)、根发育(Li et al., 2019)、生长代谢(Müller et al., 2014)、开花过程(Doyle et al., 2002)、光合作用(Dodd et al., 2005)以及对生物和非生物胁迫的响应(Sanchez et al., 2011; Ronald and Davis, 2017), 在植物正常生长发育过程中起着非常重要的作用(魏华等, 2018). ...
植物生物钟与关键农艺性状调控 1 2015
... 地球自转引起光照和温度等诸多环境因子昼夜周期变化.为了更好地应对变化的环境信号, 生物体进化出内源性生物钟系统来协调环境变化与自身生长发育.不同物种中生物钟系统关键组分虽有明显差异, 但均包含一些共同特征: (1) 具有内源性, 在持续光照或恒定温度条件下仍能维持近24小时的节律性; (2) 具有可驯化性, 如改变光周期, 生物钟可被重新设定来适应新的生存环境; (3) 具有温度补偿现象, 即在一定温度范围内, 生物钟系统仍能维持周期基本稳定(不被改变) (谢启光和徐小冬, 2015).植物生物钟参与调控植物多种生命活动, 如激素信号(Covington and Harmer, 2007)、叶片衰老(Zhang et al., 2018, 2019)、下胚轴伸长(Niwa et al., 2009)、根发育(Li et al., 2019)、生长代谢(Müller et al., 2014)、开花过程(Doyle et al., 2002)、光合作用(Dodd et al., 2005)以及对生物和非生物胁迫的响应(Sanchez et al., 2011; Ronald and Davis, 2017), 在植物正常生长发育过程中起着非常重要的作用(魏华等, 2018). ...
Computer-assisted measurements of plant growth with linear variable differential transformer (LVDT) sensors 1 1997
... (2) 叶片运动活体监测与分析方法.分析植物生物钟节律也可通过测量植物生理上的节律变化进行, 如叶片位置(Edwards and Millar, 2007)、下胚轴伸长(Dowson-Day and Millar, 1999)、花序伸长(Agosti et al., 1997)和叶片气孔开合(Somers et al., 1998).例如, 拟南芥(Arabidopsis thaliana)幼苗, 支撑叶片的叶柄正面和背面细胞伸长的反相振荡会导致叶片(尤其是幼叶)高度在白天降低, 而在夜间升高(Uehlein and Kaldenhoff, 2008).根据图像之间叶片位置的差异可测定叶片运动的节律.其原理基于在持续的一段时间里收集连续的幼苗图像, 并测量每个图像中初生叶片的垂直位置.该方法最初是手动收集和测量一系列图像, 后来Edwards和Millar (2007)在拟南芥中开发了自动化系统, 该系统使用1组CCD相机定时捕获幼苗图像并使用图像分析软件进行测量分析.该方法优点在于非损伤检测, 且易于观察, 无须事先对植物进行遗传操作.此后, 基于上述原理人们还开发出了几种类似系统, 可实现多次拍摄多株幼苗(Yanovsky et al., 2000; Salomé et al., 2002; Muller and Jimenezomez, 2016). ...
Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock 1 2001
... (3) 植物活体生物发光方法.利用生物钟关键基因的启动子驱动荧光素酶(luciferase, LUC)表达, 结合生物发光实时采集检测系统, 是目前植物生物钟研究最可靠且最重要的方法, 具有非损伤性、时间精度高且数据采集和分析全自动化等优点.该方法最早使用CAB2 (CHLOROPHYLLA/B BINDING PROTEIN 2)基因启动子融合LUC基因作为报告基因, 实验结果显示CAB2pro:LUC的荧光强度与CAB2自身转录本的表达几乎一致, 并可进行连续检测(Millar et al., 1995).随后, 又出现使用CCR2 (COLD CIRCADIAN RHYTHM RNA BINDING 2) (Más et al., 2003)、CCA1 (CIRCADIAN CLOCK-ASSOCIATED 1)以及TOC1 (TIME OF CAB EXPRESSION 1)等基因启动子驱动LUC基因表达来检测生物钟表型(Alabadi et al., 2001; Doyle et al., 2002).目前, 根据选取材料的不同, 又可细分为原生质体方法、细胞培养方法和整株植物方法.原生质体方法可用来研究突变后可能导致植株致死的关键基因的生物钟表型, 也可对由于基因功能冗余而表现出单个基因表型微弱甚至观察不到表型的基因家族进行研究.此外, 还可结合amiRNA (artificial microRNA)技术在拟南芥原生质体中降低内源靶基因或者基因家族的表达水平, 同时瞬时共转报告基因来快速确定基因的功能(Wang et al., 2013).具体操作可参照Kim和Somers (2010)的方法.细胞培养方法目前应用较少.Naka-michi等(2004)建立了拟南芥营养缺陷型培养细胞系T87, 并且通过农杆菌介导的DNA转化方法构建了带有CCA1pro:LUC以及PRRspro: LUC (PSEUDO- RESPONSE REGULATORspro: LUC)的转基因细胞系.他们将这些细胞系的抗性愈伤组织样细胞转移至装有新鲜CIM琼脂培养基的玻璃瓶中, 驯化培养3天后进行生物发光检测, 结果显示与完整植株的生物钟表型一致.该方法为生物钟表型的快速检测提供了新思路.本文主要对利用整株植物的生物发光监测和生物钟表型计算方法进行详细阐述, 旨在为生物钟表型研究提供参考.此外, 也可为基于水稻(Oryza sativa)愈伤组织以及大豆(Glycine max)发根系统的生物发光成像研究植物生物钟提供借鉴. ...
The circadian clock regulates auxin signaling and responses in Arabidopsis 1 2007
... 地球自转引起光照和温度等诸多环境因子昼夜周期变化.为了更好地应对变化的环境信号, 生物体进化出内源性生物钟系统来协调环境变化与自身生长发育.不同物种中生物钟系统关键组分虽有明显差异, 但均包含一些共同特征: (1) 具有内源性, 在持续光照或恒定温度条件下仍能维持近24小时的节律性; (2) 具有可驯化性, 如改变光周期, 生物钟可被重新设定来适应新的生存环境; (3) 具有温度补偿现象, 即在一定温度范围内, 生物钟系统仍能维持周期基本稳定(不被改变) (谢启光和徐小冬, 2015).植物生物钟参与调控植物多种生命活动, 如激素信号(Covington and Harmer, 2007)、叶片衰老(Zhang et al., 2018, 2019)、下胚轴伸长(Niwa et al., 2009)、根发育(Li et al., 2019)、生长代谢(Müller et al., 2014)、开花过程(Doyle et al., 2002)、光合作用(Dodd et al., 2005)以及对生物和非生物胁迫的响应(Sanchez et al., 2011; Ronald and Davis, 2017), 在植物正常生长发育过程中起着非常重要的作用(魏华等, 2018). ...
... 地球自转引起光照和温度等诸多环境因子昼夜周期变化.为了更好地应对变化的环境信号, 生物体进化出内源性生物钟系统来协调环境变化与自身生长发育.不同物种中生物钟系统关键组分虽有明显差异, 但均包含一些共同特征: (1) 具有内源性, 在持续光照或恒定温度条件下仍能维持近24小时的节律性; (2) 具有可驯化性, 如改变光周期, 生物钟可被重新设定来适应新的生存环境; (3) 具有温度补偿现象, 即在一定温度范围内, 生物钟系统仍能维持周期基本稳定(不被改变) (谢启光和徐小冬, 2015).植物生物钟参与调控植物多种生命活动, 如激素信号(Covington and Harmer, 2007)、叶片衰老(Zhang et al., 2018, 2019)、下胚轴伸长(Niwa et al., 2009)、根发育(Li et al., 2019)、生长代谢(Müller et al., 2014)、开花过程(Doyle et al., 2002)、光合作用(Dodd et al., 2005)以及对生物和非生物胁迫的响应(Sanchez et al., 2011; Ronald and Davis, 2017), 在植物正常生长发育过程中起着非常重要的作用(魏华等, 2018). ...
... (2) 叶片运动活体监测与分析方法.分析植物生物钟节律也可通过测量植物生理上的节律变化进行, 如叶片位置(Edwards and Millar, 2007)、下胚轴伸长(Dowson-Day and Millar, 1999)、花序伸长(Agosti et al., 1997)和叶片气孔开合(Somers et al., 1998).例如, 拟南芥(Arabidopsis thaliana)幼苗, 支撑叶片的叶柄正面和背面细胞伸长的反相振荡会导致叶片(尤其是幼叶)高度在白天降低, 而在夜间升高(Uehlein and Kaldenhoff, 2008).根据图像之间叶片位置的差异可测定叶片运动的节律.其原理基于在持续的一段时间里收集连续的幼苗图像, 并测量每个图像中初生叶片的垂直位置.该方法最初是手动收集和测量一系列图像, 后来Edwards和Millar (2007)在拟南芥中开发了自动化系统, 该系统使用1组CCD相机定时捕获幼苗图像并使用图像分析软件进行测量分析.该方法优点在于非损伤检测, 且易于观察, 无须事先对植物进行遗传操作.此后, 基于上述原理人们还开发出了几种类似系统, 可实现多次拍摄多株幼苗(Yanovsky et al., 2000; Salomé et al., 2002; Muller and Jimenezomez, 2016). ...
The ELF4 gene controls circadian rhythms and flowering time in Arabidopsis thaliana 2 2002
... 地球自转引起光照和温度等诸多环境因子昼夜周期变化.为了更好地应对变化的环境信号, 生物体进化出内源性生物钟系统来协调环境变化与自身生长发育.不同物种中生物钟系统关键组分虽有明显差异, 但均包含一些共同特征: (1) 具有内源性, 在持续光照或恒定温度条件下仍能维持近24小时的节律性; (2) 具有可驯化性, 如改变光周期, 生物钟可被重新设定来适应新的生存环境; (3) 具有温度补偿现象, 即在一定温度范围内, 生物钟系统仍能维持周期基本稳定(不被改变) (谢启光和徐小冬, 2015).植物生物钟参与调控植物多种生命活动, 如激素信号(Covington and Harmer, 2007)、叶片衰老(Zhang et al., 2018, 2019)、下胚轴伸长(Niwa et al., 2009)、根发育(Li et al., 2019)、生长代谢(Müller et al., 2014)、开花过程(Doyle et al., 2002)、光合作用(Dodd et al., 2005)以及对生物和非生物胁迫的响应(Sanchez et al., 2011; Ronald and Davis, 2017), 在植物正常生长发育过程中起着非常重要的作用(魏华等, 2018). ...
... (3) 植物活体生物发光方法.利用生物钟关键基因的启动子驱动荧光素酶(luciferase, LUC)表达, 结合生物发光实时采集检测系统, 是目前植物生物钟研究最可靠且最重要的方法, 具有非损伤性、时间精度高且数据采集和分析全自动化等优点.该方法最早使用CAB2 (CHLOROPHYLLA/B BINDING PROTEIN 2)基因启动子融合LUC基因作为报告基因, 实验结果显示CAB2pro:LUC的荧光强度与CAB2自身转录本的表达几乎一致, 并可进行连续检测(Millar et al., 1995).随后, 又出现使用CCR2 (COLD CIRCADIAN RHYTHM RNA BINDING 2) (Más et al., 2003)、CCA1 (CIRCADIAN CLOCK-ASSOCIATED 1)以及TOC1 (TIME OF CAB EXPRESSION 1)等基因启动子驱动LUC基因表达来检测生物钟表型(Alabadi et al., 2001; Doyle et al., 2002).目前, 根据选取材料的不同, 又可细分为原生质体方法、细胞培养方法和整株植物方法.原生质体方法可用来研究突变后可能导致植株致死的关键基因的生物钟表型, 也可对由于基因功能冗余而表现出单个基因表型微弱甚至观察不到表型的基因家族进行研究.此外, 还可结合amiRNA (artificial microRNA)技术在拟南芥原生质体中降低内源靶基因或者基因家族的表达水平, 同时瞬时共转报告基因来快速确定基因的功能(Wang et al., 2013).具体操作可参照Kim和Somers (2010)的方法.细胞培养方法目前应用较少.Naka-michi等(2004)建立了拟南芥营养缺陷型培养细胞系T87, 并且通过农杆菌介导的DNA转化方法构建了带有CCA1pro:LUC以及PRRspro: LUC (PSEUDO- RESPONSE REGULATORspro: LUC)的转基因细胞系.他们将这些细胞系的抗性愈伤组织样细胞转移至装有新鲜CIM琼脂培养基的玻璃瓶中, 驯化培养3天后进行生物发光检测, 结果显示与完整植株的生物钟表型一致.该方法为生物钟表型的快速检测提供了新思路.本文主要对利用整株植物的生物发光监测和生物钟表型计算方法进行详细阐述, 旨在为生物钟表型研究提供参考.此外, 也可为基于水稻(Oryza sativa)愈伤组织以及大豆(Glycine max)发根系统的生物发光成像研究植物生物钟提供借鉴. ...
Analysis of circadian leaf movement rhythms in Arabidopsis thaliana 1 2007
... (2) 叶片运动活体监测与分析方法.分析植物生物钟节律也可通过测量植物生理上的节律变化进行, 如叶片位置(Edwards and Millar, 2007)、下胚轴伸长(Dowson-Day and Millar, 1999)、花序伸长(Agosti et al., 1997)和叶片气孔开合(Somers et al., 1998).例如, 拟南芥(Arabidopsis thaliana)幼苗, 支撑叶片的叶柄正面和背面细胞伸长的反相振荡会导致叶片(尤其是幼叶)高度在白天降低, 而在夜间升高(Uehlein and Kaldenhoff, 2008).根据图像之间叶片位置的差异可测定叶片运动的节律.其原理基于在持续的一段时间里收集连续的幼苗图像, 并测量每个图像中初生叶片的垂直位置.该方法最初是手动收集和测量一系列图像, 后来Edwards和Millar (2007)在拟南芥中开发了自动化系统, 该系统使用1组CCD相机定时捕获幼苗图像并使用图像分析软件进行测量分析.该方法优点在于非损伤检测, 且易于观察, 无须事先对植物进行遗传操作.此后, 基于上述原理人们还开发出了几种类似系统, 可实现多次拍摄多株幼苗(Yanovsky et al., 2000; Salomé et al., 2002; Muller and Jimenezomez, 2016). ...
Rapid assessment of gene function in the circadian clock using artificial microRNA in Arabidopsis mesophyll protoplasts 1 2010
... (3) 植物活体生物发光方法.利用生物钟关键基因的启动子驱动荧光素酶(luciferase, LUC)表达, 结合生物发光实时采集检测系统, 是目前植物生物钟研究最可靠且最重要的方法, 具有非损伤性、时间精度高且数据采集和分析全自动化等优点.该方法最早使用CAB2 (CHLOROPHYLLA/B BINDING PROTEIN 2)基因启动子融合LUC基因作为报告基因, 实验结果显示CAB2pro:LUC的荧光强度与CAB2自身转录本的表达几乎一致, 并可进行连续检测(Millar et al., 1995).随后, 又出现使用CCR2 (COLD CIRCADIAN RHYTHM RNA BINDING 2) (Más et al., 2003)、CCA1 (CIRCADIAN CLOCK-ASSOCIATED 1)以及TOC1 (TIME OF CAB EXPRESSION 1)等基因启动子驱动LUC基因表达来检测生物钟表型(Alabadi et al., 2001; Doyle et al., 2002).目前, 根据选取材料的不同, 又可细分为原生质体方法、细胞培养方法和整株植物方法.原生质体方法可用来研究突变后可能导致植株致死的关键基因的生物钟表型, 也可对由于基因功能冗余而表现出单个基因表型微弱甚至观察不到表型的基因家族进行研究.此外, 还可结合amiRNA (artificial microRNA)技术在拟南芥原生质体中降低内源靶基因或者基因家族的表达水平, 同时瞬时共转报告基因来快速确定基因的功能(Wang et al., 2013).具体操作可参照Kim和Somers (2010)的方法.细胞培养方法目前应用较少.Naka-michi等(2004)建立了拟南芥营养缺陷型培养细胞系T87, 并且通过农杆菌介导的DNA转化方法构建了带有CCA1pro:LUC以及PRRspro: LUC (PSEUDO- RESPONSE REGULATORspro: LUC)的转基因细胞系.他们将这些细胞系的抗性愈伤组织样细胞转移至装有新鲜CIM琼脂培养基的玻璃瓶中, 驯化培养3天后进行生物发光检测, 结果显示与完整植株的生物钟表型一致.该方法为生物钟表型的快速检测提供了新思路.本文主要对利用整株植物的生物发光监测和生物钟表型计算方法进行详细阐述, 旨在为生物钟表型研究提供参考.此外, 也可为基于水稻(Oryza sativa)愈伤组织以及大豆(Glycine max)发根系统的生物发光成像研究植物生物钟提供借鉴. ...
PRR5, 7 and 9 positively modulate TOR signaling-mediated root cell proliferation by repressing TANDEM ZINC FINGER 1 in Arabidopsis 1 2019
... 地球自转引起光照和温度等诸多环境因子昼夜周期变化.为了更好地应对变化的环境信号, 生物体进化出内源性生物钟系统来协调环境变化与自身生长发育.不同物种中生物钟系统关键组分虽有明显差异, 但均包含一些共同特征: (1) 具有内源性, 在持续光照或恒定温度条件下仍能维持近24小时的节律性; (2) 具有可驯化性, 如改变光周期, 生物钟可被重新设定来适应新的生存环境; (3) 具有温度补偿现象, 即在一定温度范围内, 生物钟系统仍能维持周期基本稳定(不被改变) (谢启光和徐小冬, 2015).植物生物钟参与调控植物多种生命活动, 如激素信号(Covington and Harmer, 2007)、叶片衰老(Zhang et al., 2018, 2019)、下胚轴伸长(Niwa et al., 2009)、根发育(Li et al., 2019)、生长代谢(Müller et al., 2014)、开花过程(Doyle et al., 2002)、光合作用(Dodd et al., 2005)以及对生物和非生物胁迫的响应(Sanchez et al., 2011; Ronald and Davis, 2017), 在植物正常生长发育过程中起着非常重要的作用(魏华等, 2018). ...
Dual role of TOC1 in the control of circadian and photomorphogenic responses in Arabidopsis 1 2003
... (3) 植物活体生物发光方法.利用生物钟关键基因的启动子驱动荧光素酶(luciferase, LUC)表达, 结合生物发光实时采集检测系统, 是目前植物生物钟研究最可靠且最重要的方法, 具有非损伤性、时间精度高且数据采集和分析全自动化等优点.该方法最早使用CAB2 (CHLOROPHYLLA/B BINDING PROTEIN 2)基因启动子融合LUC基因作为报告基因, 实验结果显示CAB2pro:LUC的荧光强度与CAB2自身转录本的表达几乎一致, 并可进行连续检测(Millar et al., 1995).随后, 又出现使用CCR2 (COLD CIRCADIAN RHYTHM RNA BINDING 2) (Más et al., 2003)、CCA1 (CIRCADIAN CLOCK-ASSOCIATED 1)以及TOC1 (TIME OF CAB EXPRESSION 1)等基因启动子驱动LUC基因表达来检测生物钟表型(Alabadi et al., 2001; Doyle et al., 2002).目前, 根据选取材料的不同, 又可细分为原生质体方法、细胞培养方法和整株植物方法.原生质体方法可用来研究突变后可能导致植株致死的关键基因的生物钟表型, 也可对由于基因功能冗余而表现出单个基因表型微弱甚至观察不到表型的基因家族进行研究.此外, 还可结合amiRNA (artificial microRNA)技术在拟南芥原生质体中降低内源靶基因或者基因家族的表达水平, 同时瞬时共转报告基因来快速确定基因的功能(Wang et al., 2013).具体操作可参照Kim和Somers (2010)的方法.细胞培养方法目前应用较少.Naka-michi等(2004)建立了拟南芥营养缺陷型培养细胞系T87, 并且通过农杆菌介导的DNA转化方法构建了带有CCA1pro:LUC以及PRRspro: LUC (PSEUDO- RESPONSE REGULATORspro: LUC)的转基因细胞系.他们将这些细胞系的抗性愈伤组织样细胞转移至装有新鲜CIM琼脂培养基的玻璃瓶中, 驯化培养3天后进行生物发光检测, 结果显示与完整植株的生物钟表型一致.该方法为生物钟表型的快速检测提供了新思路.本文主要对利用整株植物的生物发光监测和生物钟表型计算方法进行详细阐述, 旨在为生物钟表型研究提供参考.此外, 也可为基于水稻(Oryza sativa)愈伤组织以及大豆(Glycine max)发根系统的生物发光成像研究植物生物钟提供借鉴. ...
Circadian clock mutants in Arabidopsis identified by luciferase imaging 1 1995
... (3) 植物活体生物发光方法.利用生物钟关键基因的启动子驱动荧光素酶(luciferase, LUC)表达, 结合生物发光实时采集检测系统, 是目前植物生物钟研究最可靠且最重要的方法, 具有非损伤性、时间精度高且数据采集和分析全自动化等优点.该方法最早使用CAB2 (CHLOROPHYLLA/B BINDING PROTEIN 2)基因启动子融合LUC基因作为报告基因, 实验结果显示CAB2pro:LUC的荧光强度与CAB2自身转录本的表达几乎一致, 并可进行连续检测(Millar et al., 1995).随后, 又出现使用CCR2 (COLD CIRCADIAN RHYTHM RNA BINDING 2) (Más et al., 2003)、CCA1 (CIRCADIAN CLOCK-ASSOCIATED 1)以及TOC1 (TIME OF CAB EXPRESSION 1)等基因启动子驱动LUC基因表达来检测生物钟表型(Alabadi et al., 2001; Doyle et al., 2002).目前, 根据选取材料的不同, 又可细分为原生质体方法、细胞培养方法和整株植物方法.原生质体方法可用来研究突变后可能导致植株致死的关键基因的生物钟表型, 也可对由于基因功能冗余而表现出单个基因表型微弱甚至观察不到表型的基因家族进行研究.此外, 还可结合amiRNA (artificial microRNA)技术在拟南芥原生质体中降低内源靶基因或者基因家族的表达水平, 同时瞬时共转报告基因来快速确定基因的功能(Wang et al., 2013).具体操作可参照Kim和Somers (2010)的方法.细胞培养方法目前应用较少.Naka-michi等(2004)建立了拟南芥营养缺陷型培养细胞系T87, 并且通过农杆菌介导的DNA转化方法构建了带有CCA1pro:LUC以及PRRspro: LUC (PSEUDO- RESPONSE REGULATORspro: LUC)的转基因细胞系.他们将这些细胞系的抗性愈伤组织样细胞转移至装有新鲜CIM琼脂培养基的玻璃瓶中, 驯化培养3天后进行生物发光检测, 结果显示与完整植株的生物钟表型一致.该方法为生物钟表型的快速检测提供了新思路.本文主要对利用整株植物的生物发光监测和生物钟表型计算方法进行详细阐述, 旨在为生物钟表型研究提供参考.此外, 也可为基于水稻(Oryza sativa)愈伤组织以及大豆(Glycine max)发根系统的生物发光成像研究植物生物钟提供借鉴. ...
Connections bet- ween circadian clocks and carbon metabolism reveal species-specific effects on growth control 1 2014
... 地球自转引起光照和温度等诸多环境因子昼夜周期变化.为了更好地应对变化的环境信号, 生物体进化出内源性生物钟系统来协调环境变化与自身生长发育.不同物种中生物钟系统关键组分虽有明显差异, 但均包含一些共同特征: (1) 具有内源性, 在持续光照或恒定温度条件下仍能维持近24小时的节律性; (2) 具有可驯化性, 如改变光周期, 生物钟可被重新设定来适应新的生存环境; (3) 具有温度补偿现象, 即在一定温度范围内, 生物钟系统仍能维持周期基本稳定(不被改变) (谢启光和徐小冬, 2015).植物生物钟参与调控植物多种生命活动, 如激素信号(Covington and Harmer, 2007)、叶片衰老(Zhang et al., 2018, 2019)、下胚轴伸长(Niwa et al., 2009)、根发育(Li et al., 2019)、生长代谢(Müller et al., 2014)、开花过程(Doyle et al., 2002)、光合作用(Dodd et al., 2005)以及对生物和非生物胁迫的响应(Sanchez et al., 2011; Ronald and Davis, 2017), 在植物正常生长发育过程中起着非常重要的作用(魏华等, 2018). ...
Analysis of circadian leaf movements. In: Duque P, ed. Environmental Responses in Plants 1 2016
... (2) 叶片运动活体监测与分析方法.分析植物生物钟节律也可通过测量植物生理上的节律变化进行, 如叶片位置(Edwards and Millar, 2007)、下胚轴伸长(Dowson-Day and Millar, 1999)、花序伸长(Agosti et al., 1997)和叶片气孔开合(Somers et al., 1998).例如, 拟南芥(Arabidopsis thaliana)幼苗, 支撑叶片的叶柄正面和背面细胞伸长的反相振荡会导致叶片(尤其是幼叶)高度在白天降低, 而在夜间升高(Uehlein and Kaldenhoff, 2008).根据图像之间叶片位置的差异可测定叶片运动的节律.其原理基于在持续的一段时间里收集连续的幼苗图像, 并测量每个图像中初生叶片的垂直位置.该方法最初是手动收集和测量一系列图像, 后来Edwards和Millar (2007)在拟南芥中开发了自动化系统, 该系统使用1组CCD相机定时捕获幼苗图像并使用图像分析软件进行测量分析.该方法优点在于非损伤检测, 且易于观察, 无须事先对植物进行遗传操作.此后, 基于上述原理人们还开发出了几种类似系统, 可实现多次拍摄多株幼苗(Yanovsky et al., 2000; Salomé et al., 2002; Muller and Jimenezomez, 2016). ...
Characterization of plant circadian rhythms by employing Arabidopsis cultured cells with bioluminescence reporters 1 2004
... (3) 植物活体生物发光方法.利用生物钟关键基因的启动子驱动荧光素酶(luciferase, LUC)表达, 结合生物发光实时采集检测系统, 是目前植物生物钟研究最可靠且最重要的方法, 具有非损伤性、时间精度高且数据采集和分析全自动化等优点.该方法最早使用CAB2 (CHLOROPHYLLA/B BINDING PROTEIN 2)基因启动子融合LUC基因作为报告基因, 实验结果显示CAB2pro:LUC的荧光强度与CAB2自身转录本的表达几乎一致, 并可进行连续检测(Millar et al., 1995).随后, 又出现使用CCR2 (COLD CIRCADIAN RHYTHM RNA BINDING 2) (Más et al., 2003)、CCA1 (CIRCADIAN CLOCK-ASSOCIATED 1)以及TOC1 (TIME OF CAB EXPRESSION 1)等基因启动子驱动LUC基因表达来检测生物钟表型(Alabadi et al., 2001; Doyle et al., 2002).目前, 根据选取材料的不同, 又可细分为原生质体方法、细胞培养方法和整株植物方法.原生质体方法可用来研究突变后可能导致植株致死的关键基因的生物钟表型, 也可对由于基因功能冗余而表现出单个基因表型微弱甚至观察不到表型的基因家族进行研究.此外, 还可结合amiRNA (artificial microRNA)技术在拟南芥原生质体中降低内源靶基因或者基因家族的表达水平, 同时瞬时共转报告基因来快速确定基因的功能(Wang et al., 2013).具体操作可参照Kim和Somers (2010)的方法.细胞培养方法目前应用较少.Naka-michi等(2004)建立了拟南芥营养缺陷型培养细胞系T87, 并且通过农杆菌介导的DNA转化方法构建了带有CCA1pro:LUC以及PRRspro: LUC (PSEUDO- RESPONSE REGULATORspro: LUC)的转基因细胞系.他们将这些细胞系的抗性愈伤组织样细胞转移至装有新鲜CIM琼脂培养基的玻璃瓶中, 驯化培养3天后进行生物发光检测, 结果显示与完整植株的生物钟表型一致.该方法为生物钟表型的快速检测提供了新思路.本文主要对利用整株植物的生物发光监测和生物钟表型计算方法进行详细阐述, 旨在为生物钟表型研究提供参考.此外, 也可为基于水稻(Oryza sativa)愈伤组织以及大豆(Glycine max)发根系统的生物发光成像研究植物生物钟提供借鉴. ...
The circadian clock regulates the photoperiodic response of hypocotyl elongation through a coincidence mechanism in Arabidopsis thaliana 1 2009
... 地球自转引起光照和温度等诸多环境因子昼夜周期变化.为了更好地应对变化的环境信号, 生物体进化出内源性生物钟系统来协调环境变化与自身生长发育.不同物种中生物钟系统关键组分虽有明显差异, 但均包含一些共同特征: (1) 具有内源性, 在持续光照或恒定温度条件下仍能维持近24小时的节律性; (2) 具有可驯化性, 如改变光周期, 生物钟可被重新设定来适应新的生存环境; (3) 具有温度补偿现象, 即在一定温度范围内, 生物钟系统仍能维持周期基本稳定(不被改变) (谢启光和徐小冬, 2015).植物生物钟参与调控植物多种生命活动, 如激素信号(Covington and Harmer, 2007)、叶片衰老(Zhang et al., 2018, 2019)、下胚轴伸长(Niwa et al., 2009)、根发育(Li et al., 2019)、生长代谢(Müller et al., 2014)、开花过程(Doyle et al., 2002)、光合作用(Dodd et al., 2005)以及对生物和非生物胁迫的响应(Sanchez et al., 2011; Ronald and Davis, 2017), 在植物正常生长发育过程中起着非常重要的作用(魏华等, 2018). ...
Making the clock tick: the transcriptional landscape of the plant circadian clock 1 2017
... 地球自转引起光照和温度等诸多环境因子昼夜周期变化.为了更好地应对变化的环境信号, 生物体进化出内源性生物钟系统来协调环境变化与自身生长发育.不同物种中生物钟系统关键组分虽有明显差异, 但均包含一些共同特征: (1) 具有内源性, 在持续光照或恒定温度条件下仍能维持近24小时的节律性; (2) 具有可驯化性, 如改变光周期, 生物钟可被重新设定来适应新的生存环境; (3) 具有温度补偿现象, 即在一定温度范围内, 生物钟系统仍能维持周期基本稳定(不被改变) (谢启光和徐小冬, 2015).植物生物钟参与调控植物多种生命活动, 如激素信号(Covington and Harmer, 2007)、叶片衰老(Zhang et al., 2018, 2019)、下胚轴伸长(Niwa et al., 2009)、根发育(Li et al., 2019)、生长代谢(Müller et al., 2014)、开花过程(Doyle et al., 2002)、光合作用(Dodd et al., 2005)以及对生物和非生物胁迫的响应(Sanchez et al., 2011; Ronald and Davis, 2017), 在植物正常生长发育过程中起着非常重要的作用(魏华等, 2018). ...
The out of phase 1 mutant defines a role for PHYB in circadian phase control in Arabidopsis 1 2002
... (2) 叶片运动活体监测与分析方法.分析植物生物钟节律也可通过测量植物生理上的节律变化进行, 如叶片位置(Edwards and Millar, 2007)、下胚轴伸长(Dowson-Day and Millar, 1999)、花序伸长(Agosti et al., 1997)和叶片气孔开合(Somers et al., 1998).例如, 拟南芥(Arabidopsis thaliana)幼苗, 支撑叶片的叶柄正面和背面细胞伸长的反相振荡会导致叶片(尤其是幼叶)高度在白天降低, 而在夜间升高(Uehlein and Kaldenhoff, 2008).根据图像之间叶片位置的差异可测定叶片运动的节律.其原理基于在持续的一段时间里收集连续的幼苗图像, 并测量每个图像中初生叶片的垂直位置.该方法最初是手动收集和测量一系列图像, 后来Edwards和Millar (2007)在拟南芥中开发了自动化系统, 该系统使用1组CCD相机定时捕获幼苗图像并使用图像分析软件进行测量分析.该方法优点在于非损伤检测, 且易于观察, 无须事先对植物进行遗传操作.此后, 基于上述原理人们还开发出了几种类似系统, 可实现多次拍摄多株幼苗(Yanovsky et al., 2000; Salomé et al., 2002; Muller and Jimenezomez, 2016). ...
Abiotic stress and the plant circadian clock 1 2011
... 地球自转引起光照和温度等诸多环境因子昼夜周期变化.为了更好地应对变化的环境信号, 生物体进化出内源性生物钟系统来协调环境变化与自身生长发育.不同物种中生物钟系统关键组分虽有明显差异, 但均包含一些共同特征: (1) 具有内源性, 在持续光照或恒定温度条件下仍能维持近24小时的节律性; (2) 具有可驯化性, 如改变光周期, 生物钟可被重新设定来适应新的生存环境; (3) 具有温度补偿现象, 即在一定温度范围内, 生物钟系统仍能维持周期基本稳定(不被改变) (谢启光和徐小冬, 2015).植物生物钟参与调控植物多种生命活动, 如激素信号(Covington and Harmer, 2007)、叶片衰老(Zhang et al., 2018, 2019)、下胚轴伸长(Niwa et al., 2009)、根发育(Li et al., 2019)、生长代谢(Müller et al., 2014)、开花过程(Doyle et al., 2002)、光合作用(Dodd et al., 2005)以及对生物和非生物胁迫的响应(Sanchez et al., 2011; Ronald and Davis, 2017), 在植物正常生长发育过程中起着非常重要的作用(魏华等, 2018). ...
The short-period mutant, toc1-1, alters circadian clock regulation of multiple outputs throughout development in Arabidopsis thaliana 1 1998
... (2) 叶片运动活体监测与分析方法.分析植物生物钟节律也可通过测量植物生理上的节律变化进行, 如叶片位置(Edwards and Millar, 2007)、下胚轴伸长(Dowson-Day and Millar, 1999)、花序伸长(Agosti et al., 1997)和叶片气孔开合(Somers et al., 1998).例如, 拟南芥(Arabidopsis thaliana)幼苗, 支撑叶片的叶柄正面和背面细胞伸长的反相振荡会导致叶片(尤其是幼叶)高度在白天降低, 而在夜间升高(Uehlein and Kaldenhoff, 2008).根据图像之间叶片位置的差异可测定叶片运动的节律.其原理基于在持续的一段时间里收集连续的幼苗图像, 并测量每个图像中初生叶片的垂直位置.该方法最初是手动收集和测量一系列图像, 后来Edwards和Millar (2007)在拟南芥中开发了自动化系统, 该系统使用1组CCD相机定时捕获幼苗图像并使用图像分析软件进行测量分析.该方法优点在于非损伤检测, 且易于观察, 无须事先对植物进行遗传操作.此后, 基于上述原理人们还开发出了几种类似系统, 可实现多次拍摄多株幼苗(Yanovsky et al., 2000; Salomé et al., 2002; Muller and Jimenezomez, 2016). ...
Aquaporins and plant leaf movements 1 2008
... (2) 叶片运动活体监测与分析方法.分析植物生物钟节律也可通过测量植物生理上的节律变化进行, 如叶片位置(Edwards and Millar, 2007)、下胚轴伸长(Dowson-Day and Millar, 1999)、花序伸长(Agosti et al., 1997)和叶片气孔开合(Somers et al., 1998).例如, 拟南芥(Arabidopsis thaliana)幼苗, 支撑叶片的叶柄正面和背面细胞伸长的反相振荡会导致叶片(尤其是幼叶)高度在白天降低, 而在夜间升高(Uehlein and Kaldenhoff, 2008).根据图像之间叶片位置的差异可测定叶片运动的节律.其原理基于在持续的一段时间里收集连续的幼苗图像, 并测量每个图像中初生叶片的垂直位置.该方法最初是手动收集和测量一系列图像, 后来Edwards和Millar (2007)在拟南芥中开发了自动化系统, 该系统使用1组CCD相机定时捕获幼苗图像并使用图像分析软件进行测量分析.该方法优点在于非损伤检测, 且易于观察, 无须事先对植物进行遗传操作.此后, 基于上述原理人们还开发出了几种类似系统, 可实现多次拍摄多株幼苗(Yanovsky et al., 2000; Salomé et al., 2002; Muller and Jimenezomez, 2016). ...
Transcriptional corepressor TOPLESS complexes with pseudoresponse regulator proteins and histone deacetylases to regulate circadian transcription 1 2013
... (3) 植物活体生物发光方法.利用生物钟关键基因的启动子驱动荧光素酶(luciferase, LUC)表达, 结合生物发光实时采集检测系统, 是目前植物生物钟研究最可靠且最重要的方法, 具有非损伤性、时间精度高且数据采集和分析全自动化等优点.该方法最早使用CAB2 (CHLOROPHYLLA/B BINDING PROTEIN 2)基因启动子融合LUC基因作为报告基因, 实验结果显示CAB2pro:LUC的荧光强度与CAB2自身转录本的表达几乎一致, 并可进行连续检测(Millar et al., 1995).随后, 又出现使用CCR2 (COLD CIRCADIAN RHYTHM RNA BINDING 2) (Más et al., 2003)、CCA1 (CIRCADIAN CLOCK-ASSOCIATED 1)以及TOC1 (TIME OF CAB EXPRESSION 1)等基因启动子驱动LUC基因表达来检测生物钟表型(Alabadi et al., 2001; Doyle et al., 2002).目前, 根据选取材料的不同, 又可细分为原生质体方法、细胞培养方法和整株植物方法.原生质体方法可用来研究突变后可能导致植株致死的关键基因的生物钟表型, 也可对由于基因功能冗余而表现出单个基因表型微弱甚至观察不到表型的基因家族进行研究.此外, 还可结合amiRNA (artificial microRNA)技术在拟南芥原生质体中降低内源靶基因或者基因家族的表达水平, 同时瞬时共转报告基因来快速确定基因的功能(Wang et al., 2013).具体操作可参照Kim和Somers (2010)的方法.细胞培养方法目前应用较少.Naka-michi等(2004)建立了拟南芥营养缺陷型培养细胞系T87, 并且通过农杆菌介导的DNA转化方法构建了带有CCA1pro:LUC以及PRRspro: LUC (PSEUDO- RESPONSE REGULATORspro: LUC)的转基因细胞系.他们将这些细胞系的抗性愈伤组织样细胞转移至装有新鲜CIM琼脂培养基的玻璃瓶中, 驯化培养3天后进行生物发光检测, 结果显示与完整植株的生物钟表型一致.该方法为生物钟表型的快速检测提供了新思路.本文主要对利用整株植物的生物发光监测和生物钟表型计算方法进行详细阐述, 旨在为生物钟表型研究提供参考.此外, 也可为基于水稻(Oryza sativa)愈伤组织以及大豆(Glycine max)发根系统的生物发光成像研究植物生物钟提供借鉴. ...
Phytochrome A resets the circadian clock and delays tuber formation under long days in potato 1 2000
... (2) 叶片运动活体监测与分析方法.分析植物生物钟节律也可通过测量植物生理上的节律变化进行, 如叶片位置(Edwards and Millar, 2007)、下胚轴伸长(Dowson-Day and Millar, 1999)、花序伸长(Agosti et al., 1997)和叶片气孔开合(Somers et al., 1998).例如, 拟南芥(Arabidopsis thaliana)幼苗, 支撑叶片的叶柄正面和背面细胞伸长的反相振荡会导致叶片(尤其是幼叶)高度在白天降低, 而在夜间升高(Uehlein and Kaldenhoff, 2008).根据图像之间叶片位置的差异可测定叶片运动的节律.其原理基于在持续的一段时间里收集连续的幼苗图像, 并测量每个图像中初生叶片的垂直位置.该方法最初是手动收集和测量一系列图像, 后来Edwards和Millar (2007)在拟南芥中开发了自动化系统, 该系统使用1组CCD相机定时捕获幼苗图像并使用图像分析软件进行测量分析.该方法优点在于非损伤检测, 且易于观察, 无须事先对植物进行遗传操作.此后, 基于上述原理人们还开发出了几种类似系统, 可实现多次拍摄多株幼苗(Yanovsky et al., 2000; Salomé et al., 2002; Muller and Jimenezomez, 2016). ...
Novel crosstalks between circadian clock and jasmonic acid pathway finely coordinate the tradeoff among plant growth, senescence and defense 1 2019
... 地球自转引起光照和温度等诸多环境因子昼夜周期变化.为了更好地应对变化的环境信号, 生物体进化出内源性生物钟系统来协调环境变化与自身生长发育.不同物种中生物钟系统关键组分虽有明显差异, 但均包含一些共同特征: (1) 具有内源性, 在持续光照或恒定温度条件下仍能维持近24小时的节律性; (2) 具有可驯化性, 如改变光周期, 生物钟可被重新设定来适应新的生存环境; (3) 具有温度补偿现象, 即在一定温度范围内, 生物钟系统仍能维持周期基本稳定(不被改变) (谢启光和徐小冬, 2015).植物生物钟参与调控植物多种生命活动, 如激素信号(Covington and Harmer, 2007)、叶片衰老(Zhang et al., 2018, 2019)、下胚轴伸长(Niwa et al., 2009)、根发育(Li et al., 2019)、生长代谢(Müller et al., 2014)、开花过程(Doyle et al., 2002)、光合作用(Dodd et al., 2005)以及对生物和非生物胁迫的响应(Sanchez et al., 2011; Ronald and Davis, 2017), 在植物正常生长发育过程中起着非常重要的作用(魏华等, 2018). ...
... 地球自转引起光照和温度等诸多环境因子昼夜周期变化.为了更好地应对变化的环境信号, 生物体进化出内源性生物钟系统来协调环境变化与自身生长发育.不同物种中生物钟系统关键组分虽有明显差异, 但均包含一些共同特征: (1) 具有内源性, 在持续光照或恒定温度条件下仍能维持近24小时的节律性; (2) 具有可驯化性, 如改变光周期, 生物钟可被重新设定来适应新的生存环境; (3) 具有温度补偿现象, 即在一定温度范围内, 生物钟系统仍能维持周期基本稳定(不被改变) (谢启光和徐小冬, 2015).植物生物钟参与调控植物多种生命活动, 如激素信号(Covington and Harmer, 2007)、叶片衰老(Zhang et al., 2018, 2019)、下胚轴伸长(Niwa et al., 2009)、根发育(Li et al., 2019)、生长代谢(Müller et al., 2014)、开花过程(Doyle et al., 2002)、光合作用(Dodd et al., 2005)以及对生物和非生物胁迫的响应(Sanchez et al., 2011; Ronald and Davis, 2017), 在植物正常生长发育过程中起着非常重要的作用(魏华等, 2018). ...
Strengths and limitations of period estimation methods for circadian data 1 2014
 ,1,2,*
,1,2,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}