Advances in the Regulation of Plant Growth and Development and Stress Response by miR172-AP2 Module
Jindong Wang, Yu Zhou, Jiawen Yu, Xiaolei Fan, Changquan Zhang, Qianfeng Li,*, Qiaoquan Liu,*Co-innovation Center for Modern Production Technology of Grain Crops/Key Laboratory of Plant Functional Genomics of the Ministry of Education/Jiangsu Key Laboratory of Crop Genomics and Molecular Breeding, College of Agriculture, Yangzhou University, Yangzhou 225009, China
Abstract MicroRNA (miRNA), a kind of regulatory non-coding small RNA, induces degradation of target mRNA or inhibits its translation by specific or non-specific binding, thereby regulating plant growth and development. AP2, the target of miR172, encodes transcription factors that are unique to plants. miR172 regulates the expression of AP2 at the post-transcriptional or translational levels, thus regulating plant floral development, phase transition, spikelet morphology, tuber and fruit development, nodulation in legumes and stress response. Here we summarize the recent advances in the regulation of plant growth and development by miR172-AP2 regulatory module. Keywords:miR172;AP2 transcription factor;expression regulation;plant growth and development
PDF (1293KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 王劲东, 周豫, 余佳雯, 范晓磊, 张昌泉, 李钱峰, 刘巧泉. miR172-AP2模块调控植物生长发育及逆境响应的研究进展. 植物学报, 2020, 55(2): 205-215 doi:10.11983/CBB19177 Wang Jindong, Zhou Yu, Yu Jiawen, Fan Xiaolei, Zhang Changquan, Li Qianfeng, Liu Qiaoquan. Advances in the Regulation of Plant Growth and Development and Stress Response by miR172-AP2 Module. Chinese Bulletin of Botany, 2020, 55(2): 205-215 doi:10.11983/CBB19177
Aguilar-JaramilloAE, Marín-GonzálezE, Matías-HernándezL, OsnatoM, PelazS, Suárez-LópezP ( 2019). TEMPRANILLO is a direct repressor of the microRNA miR172 Plant J 100, 522-535. [本文引用: 1]
AnwarN, OhtaM, YazawaT, SatoY, LiC, TagiriA, SakumaM, NussbaumerT, BregitzerP, PourkheirandishM, WuJZ, KomatsudaT ( 2018). miR172 downregulates the translation of cleistogamy 1 in barley Ann Bot 122, 251-265. [本文引用: 1]
AukermanMJ, SakaiH ( 2003). Regulation of flowering time and floral organ identity by a microRNA and its APETALA2-like target genes Plant Cell 15, 2730-2741. [本文引用: 3]
BartelDP ( 2004). MicroRNAs: genomics, biogenesis, mechanism, and function Cell 116, 281-297. [本文引用: 1]
B?umlerJ, RiberW, KleckerM, MüllerL, DissmeyerN, WeigAR, MustrophA ( 2019). AtERF#111/ABR1 is a transcriptional activator involved in the wounding response Plant J 100, 969-990. [本文引用: 2]
BoualemA, LaporteP, JovanovicM, LaffontC, PletJ, CombierJP, NiebelA, CrespiM, FrugierF ( 2008). MicroRNA166 controls root and nodule development in Medicago truncatula Plant J 54, 876-887. [本文引用: 1]
BoutilierK, OffringaR, SharmaVK, KieftH, OuelletT, ZhangLM, HattoriJ, LiuCM, van LammerenAAM, MikiBLA, CustersJBM, van Lookeren CampagneMM ( 2002). Ectopic expression of BABY BOOM triggers a conversion from vegetative to embryonic growth Plant Cell 14, 1737-1749. [本文引用: 1]
BowmanJL, SmythDR, MeyerowitzEM ( 2012). The ABC model of flower development: then and now Development 139, 4095-4098. [本文引用: 1]
Candar-CakirB, AricanE, ZhangBH ( 2016). Small RNA and degradome deep sequencing reveals drought-and tissue-specific micrornas and their important roles in drought-sensitive and drought-tolerant tomato genotypes Plant Biotechnol J 14, 1727-1746. [本文引用: 1]
CarringtonJC, AmbrosV ( 2003). Role of microRNAs in plant and animal development Science 301, 336-338. [本文引用: 1]
ChenXM ( 2004). A microRNA as a translational repressor of APETALA2 in Arabidopsis flower development Science 303, 2022-2025. [本文引用: 3]
ChuckG, CiganAM, SaeteurnK, HakeS ( 2007). The heterochronic maize mutant Corngrass1 results from overexpression of a tandem microRNA Nat Genet 39, 544-549. [本文引用: 2]
ChuckG, MeeleyR, HakeS ( 2008). Floral meristem initiation and meristem cell fate are regulated by the maize AP2 genes ids1 and sid1 Development 135, 3013-3019. [本文引用: 1]
CombierJP, FrugierF, de BillyF, BoualemA, El-YahyaouiF, MoreauS, VerniéT, OttT, GamasP, CrespiM, NiebelA ( 2006). MtHAP2-1 is a key transcriptional regulator of symbiotic nodule development regulated by microRNA169 in Medicago truncatula Genes Dev 20, 3084-3088. [本文引用: 1]
De LuisA, MarkmannK, CognatV, HoltDB, CharpentierM, ParniskeM, StougaardJ, VoinnetO ( 2012). Two microRNAs linked to nodule infection and nitrogen-fixing ability in the legume Lotus japonicus Plant Physiol 160, 2137-2154. [本文引用: 1]
DubouzetJG, SakumaY, ItoY, KasugaM, DubouzetEG, MiuraS, SekiM, ShinozakiK, Yamaguchi-ShinozakiK ( 2003). OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in drought-, high-salt- and cold-responsive gene expression Plant J 33, 751-763. [本文引用: 2]
ElliottRC, BetznerAS, HuttnerE, OakesMP, TuckerWQ, GerentesD, PerezP, SmythDR ( 1996). AINTEGUMENTA, an APETALA2-like gene of Arabidopsis with pleiotropic roles in ovule development and floral organ growth Plant Cell 8, 155-168. [本文引用: 1]
Fran?oisL, VerdenaudM, FuXP, RulemanD, DuboisA, VandenbusscheM, BendahmaneA, RaymondO, JustJ, BendahmaneM ( 2018). A miR172 target-deficient AP2-like gene correlates with the double flower phenotype in roses Sci Rep 8, 12912. [本文引用: 1]
FrazierTP, SunGL, BurklewCE, ZhangBH ( 2011). Salt and drought stresses induce the aberrant expression of microRNA genes in tobacco Mol Biotechnol 49, 159-165. [本文引用: 1]
FuFF, XueHW ( 2010). Coexpression analysis identifies Rice Starch Regulator1, a rice AP2/EREBP family transcription factor, as a novel rice starch biosynthesis regulator Plant Physiol 154, 927-938. [本文引用: 1]
GlazińskaP, ZienkiewiczA, WojciechowskiW, KopcewiczJ ( 2009). The putative miR172 target gene InAPETALA2-like is involved in the photoperiodic flower induction of Ipomoea nil J Plant Physiol 166, 1801-1813. [本文引用: 1]
HanYY, ZhangX, WangYF, MingF ( 2013). The suppression of WRKY44 by GIGANTEA-miR172 pathway is involved in drought response of Arabidopsis thaliana PLoS One 8, e73541. [本文引用: 1]
HoltDB, GuptaV, MeyerD, AbelNB, AndersenSU, StougaardJ, MarkmannK ( 2015). MicroRNA 172 (miR172) signals epidermal infection and is expressed in cells primed for bacterial invasion in Lotus japonicus roots and nodules New Phytol 208, 241-256. [本文引用: 1]
HoustonK, McKimSM, ComadranJ, BonarN, DrukaI, UzrekN, CirilloE, Guzy-WrobelskaJ, CollinsNC, HalpinC, HanssonM, DockterC, DrukaA, WaughR ( 2013). Variation in the interaction between alleles of HvAPETALA2 and microRNA172 determines the density of grains on the barley inflorescence Proc Natl Acad Sci USA 110, 16675-16680. [本文引用: 1]
HuYX, WangYX, LiuXF, LiJY ( 2004). Arabidopsis RAV1 is down-regulated by brassinosteroid and may act as a negative regulator during plant development Cell Res 14, 8-15. [本文引用: 1]
HutvágnerG, ZamorePD ( 2002). A microRNA in a multiple-turnover RNAi enzyme complex Science 297, 2056-2060. [本文引用: 1]
JiLJ, LiuXG, YanJ, WangWM, YumulRE, KimYJ, DinhTT, LiuJ, CuiX, ZhengBL, AgarwalM, LiuCY, CaoXF, TangGL, ChenXM ( 2011). ARGONAUTE10 and ARGONAUTE1 regulate the termination of floral stem cells through two microRNAs in Arabidopsis PLoS Genet 7, e1001358. [本文引用: 1]
JofukuKD, den BoerBG, Van MontaguM, OkamuroJK ( 1994). Control of Arabidopsis flower and seed development by the homeotic gene APETALA2 Plant Cell 6, 1211-1225. [本文引用: 1]
Jones-RhoadesMW, BartelDP, BartelB ( 2006). MicroRNAs and their regulatory roles in plants Annu Rev Plant Biol 57, 19-53. [本文引用: 1]
José RipollJ, BaileyLJ, MaiQA, WuSL, HonCT, ChapmanEJ, DittaGS, EstelleM, YanofskyMF ( 2015). MicroRNA regulation of fruit growth Nat Plants 1, 15036. [本文引用: 1]
JungJH, SeoYH, SeoPJ, ReyesJL, YunJ, ChuaNH, ParkCM ( 2007). The GIGANTEA-regulated microRNA172 mediates photoperiodic flowering independent of CONSTANS in Arabidopsis Plant Cell 19, 2736-2748. [本文引用: 3]
KhandayI, SkinnerD, YangB, MercierR, SundaresanV ( 2019). A male-expressed rice embryogenic trigger redirected for asexual propagation through seeds Nature 565, 91-95. [本文引用: 1]
KunstL, KlenzJE, Martinez-ZapaterJ, HaughnGW ( 1989). AP2 gene determines the identity of perianth organs in flowers of Arabidopsis thaliana Plant Cell 1, 1195-1208. [本文引用: 1]
KuriharaY, WatanabeY ( 2004). Arabidopsis micro-RNA biogenesis through dicer-like 1 protein functions Proc Natl Acad Sci USA 101, 12753-12758. [本文引用: 1]
LauterN, KampaniA, CarlsonS, GoebelM, MooseSP ( 2005). MicroRNA172 down-regulates glossy15 to promote vegetative phase change in maize Proc Natl Acad Sci USA 102, 9412-9417. [本文引用: 2]
LeeDY, AnG ( 2012). Two AP2 family genes, SUPERNUMERARY BRACT (SNB) and OsINDETERMINATE SPIKELET 1 (OsIDS1), synergistically control inflorescence architecture and floral meristem establishment in rice Plant J 69, 445-461. [本文引用: 1]
LeeYS, LeeDY, ChoLH, AnG ( 2014). Rice miR172 induces flowering by suppressing OsIDS1 and SNB, two AP2 genes that negatively regulate expression of Ehd1 and florigens Rice 7, 31. [本文引用: 1]
LeiM, LiZY, WangJB, FuYL, XuL ( 2019). Ectopic expression of the Aechmea fasciata APETALA2 gene AfAP2-2 reduces seed size and delays flowering in Arabidopsis Plant Physiol Biochem 139, 642-650. [本文引用: 1]
LiJJ, YangZY, YuB, LiuJ, ChenXM ( 2005). Methylation protects miRNAs and siRNAs from a 3°-end uridylation activity in Arabidopsis Curr Biol 15, 1501-1507. [本文引用: 1]
LiJP, ChenFJ, LiYQ, LiPC, WangYQ, MiGH, YuanLX ( 2019a). ZmRAP2.7, an AP2 transcription factor, is involved in maize brace roots development Front Plant Sci 10, 820. [本文引用: 1]
LiXY, GuoF, MaSY, ZhuMY, PanWH, BianHW ( 2019b). Regulation of flowering time via miR172-mediated APETALA2-like expression in ornamental gloxinia (Sinningia speciosa) J Zhejiang Univ Sci B 20, 322-331. [本文引用: 2]
LiuP, LiuJ, DongHX, SunJQ ( 2018). Functional regulation of Q by microRNA172 and transcriptional co-repressor TOPLESS in controlling bread wheat spikelet density Plant Biotechnol J 16, 495-506. [本文引用: 1]
LlaveC, XieZX, KasschauKD, CarringtonJC ( 2002). Cleavage of Scarecrow-like mRNA targets directed by a class of Arabidopsis miRNA Science 297, 2053-2056. [本文引用: 1]
LuanYS, CuiJ, LiJ, JiangN, LiuP, MengJ ( 2018). Effective enhancement of resistance to Phytophthora infestans by overexpression of miR172a and b in Solanum lycopersicum Planta 247, 127-138. [本文引用: 3]
MartinA, AdamH, Díaz-MendozaM, ?urczakM, González-SchainND, Suárez-LópezP ( 2009). Graft-transmissible induction of potato tuberization by the microRNA miR172 Development 136, 2873-2881. [本文引用: 1]
MathieuJ, YantLJ, MürdterF, KüttnerF, SchmidM ( 2009). Repression of flowering by the miR172 target SMZ PLoS Biol 7, e1000148. [本文引用: 1]
MiaoCB, WangD, HeRQ, LiuSK, ZhuJK ( 2020). Mutations in MIR396e and MIR396f increase grain size and modulate shoot architecture in rice Plant Biotechnol J 18, 491-501. [本文引用: 1]
MiaoCB, WangZ, ZhangL, YaoJJ, HuaK, LiuX, ShiHZ, ZhuJK ( 2019). The grain yield modulator miR156 regulates seed dormancy through the gibberellin pathway in rice Nat Commun 10, 3822. [本文引用: 1]
MlotshwaS, YangZY, KimY, ChenXM ( 2006). Floral patterning defects induced by Arabidopsis APETALA2 and microRNA172 expression in Nicotiana benthamiana Plant Mol Biol 61, 781-793. [本文引用: 1]
MooseSP, SiscoPH ( 1994). Glossy15 controls the epidermal juvenile-to-adult phase transition in maize Plant Cell 6, 1343-1355. [本文引用: 1]
MooseSP, SiscoPH ( 1996). Glossy15, an APETALA2-like gene from maize that regulates leaf epidermal cell identity Genes Dev 10, 3018-3027. [本文引用: 1]
NairSK, WangN, TuruspekovY, PourkheirandishM, SinsuwongwatS, ChenGX, SameriM, TagiriA, HondaI, WatanabeY, KanamoriH, WickerT, SteinN, NagamuraY, MatsumotoT, KomatsudaT ( 2009). Cleistogamous flowering in barley arises from the suppression of microRNA-guided HvAP2 mRNA cleavage Proc Natl Acad Sci USA 107, 490-495. [本文引用: 1]
NakanoT, SuzukiK, FujimuraT, ShinshiH ( 2006). Genome-wide analysis of the ERF gene family in Arabidopsis and rice Plant Physiol 140, 411-432. [本文引用: 1]
Nova-FrancoB, í?iguezLP, Valdés-LópezO, Alvarado- AffantrangerX, LeijaA, FuentesSI, RamírezM, PaulS, ReyesJL, GirardL, HernándezG ( 2015). The microRNA172c-APETALA2-1 node as a key regulator of the common bean- Rhizobium etli nitrogen fixation symbiosis Plant Physiol 168, 273-291. [本文引用: 1]
ParkMY, WuG, Gonzalez-SulserA, VaucheretH, PoethigRS ( 2005). Nuclear processing and export of microRNAs in Arabidopsis Proc Natl Acad Sci USA 102, 3691-3696. [本文引用: 1]
ParkW, LiJJ, SongRT, MessingJ, ChenXM ( 2002). CARPEL FACTORY, a dicer homolog, and HEN1, a novel protein, act in microRNA metabolism in Arabidopsis thaliana Curr Biol 12, 1484-1495. [本文引用: 2]
RiechmannJL, MeyerowitzEM ( 1998). The AP2/EREBP family of plant transcription factors Biol Chem 379, 633-646. [本文引用: 1]
RogersK, ChenXM ( 2013). Biogenesis, turnover, and mode of action of plant microRNAs Plant Cell 25, 2383-2399. [本文引用: 1]
SahitoZA, WangLX, SunZX, YanQQ, ZhangXK, JiangQ, UllahI, TangYP, LiX ( 2017). The miR172c-NNC1 module modulates root plastic development in response to salt in soybean BMC Plant Biol 17, 229. [本文引用: 1]
SaminathanT, AlvaradoA, LopezC, ShindeS, GajanayakeB, AbburiVL, VajjaVG, JagadeeswaranG, Raja ReddyK, NimmakayalaP, ReddyUK ( 2019). Elevated carbon dioxide and drought modulate physiology and storage-root development in sweet potato by regulating microRNAs Funct Integr Genomics 19, 171-190. [本文引用: 3]
SchmidM, UhlenhautNH, GodardF, DemarM, BressanR, WeigelD, LohmannJU ( 2003). Dissection of floral induction pathways using global expression analysis Development 130, 6001-6012. [本文引用: 1]
SchwabR, PalatnikJF, RiesterM, SchommerC, SchmidM, WeigelD ( 2005). Specific effects of microRNAs on the plant transcriptome Dev Cell 8, 517-527. [本文引用: 1]
ShannonS, Meeks-WagnerDR ( 1993). Genetic interactions that regulate inflorescence development in Arabidopsis Plant Cell 5, 639-655. [本文引用: 1]
SimonsKJ, FellersJP, TrickHN, ZhangZC, TaiYS, GillBS, FarisJD ( 2005). Molecular characterization of the major wheat domestication gene Q Genetics 172, 547-555. [本文引用: 1]
SohnKH, LeeSC, JungHW, HongJK, HwangBK ( 2006). Expression and functional roles of the pepper pathogen-induced transcription factor RAV1 in bacterial disease resistance, and drought and salt stress tolerance Plant Mol Biol 61, 897-915. [本文引用: 1]
TangMY, BaiX, NiuLJ, ChaiX, ChenMS, XuZF ( 2018). miR172 regulates both vegetative and reproductive development in the perennial woody plant Jatropha curcas Plant Cell Physiol 59, 2549-2563. [本文引用: 1]
WangGD, XuXP, WangH, LiuQ, YangXT, LiaoLX, CaiGH ( 2019a). A tomato transcription factor, SlDREB3 enhances the tolerance to chilling in transgenic tomato Plant Physiol Biochem 142, 254-262. [本文引用: 2]
WangL, SunSY, JinJY, FuDB, YangXF, WengXY, XuCG, LiXH, XiaoJH, ZhangQF ( 2015a). Coordinated regulation of vegetative and reproductive branching in rice Proc Natl Acad Sci USA 112, 15504-11509. [本文引用: 3]
WangTY, PingXK, CaoYR, JianHJ, GaoYM, WangJ, TanYC, XuXF, LuK, LiJN, LiuLZ ( 2019b). Genome-wide exploration and characterization of miR172/ euAP2 genes in Brassica napus L for likely role in flower organ development. BMC Plant Biol 19, 336. [本文引用: 2]
WangY, WangZS, AmyotL, TianLN, XuZQ, GruberMY, HannoufaA ( 2015b). Ectopic expression of miR156 represses nodulation and causes morphological and developmental changes in Lotus japonicus Mol Genet Genomics 290, 471-484. [本文引用: 1]
WangYN, LiKX, ChenL, ZouYM, LiuHP, TianYP, LiDX, WangR, ZhaoF, FergusonBJ, GresshoffPM, LiX (c). 2015 MicroRNA167-directed regulation of the auxin response factors GmARF8a and GmARF8b is required for soybean nodulation and lateral root development Plant Physiol 168, 984-999. [本文引用: 1]
WangYW, LiPC, CaoXF, WangXJ, ZhangAM, LiX ( 2009). Identification and expression analysis of miRNAs from nitrogen-fixing soybean nodules Biochem Biophys Res Commun 378, 799-803. [本文引用: 1]
WollmannH, MicaE, TodescoM, LongJA, WeigelD ( 2010). On reconciling the interactions between APETALA2, miR172 and AGAMOUS with the ABC model of flower development Development 137, 3633-3642. [本文引用: 1]
WuG, ParkMY, ConwaySR, WangJW, WeigelD, PoethigRS ( 2009). The sequential action of miR156 and miR172 regulates developmental timing in Arabidopsis Cell 138, 750-759. [本文引用: 3]
XieZL, NolanTM, JiangH, YinYH ( 2019). AP2/ERF transcription factor regulatory networks in hormone and abiotic stress responses in Arabidopsis Front Plant Sci 10, 228. [本文引用: 1]
YangJW, ZhangN, MaCY, QuY, SiHJ, WangD ( 2013). Prediction and verification of microRNAs related to proline accumulation under drought stress in potato Comput Biol Chem 46, 48-54. [本文引用: 1]
ZhangKX, ZhaoL, YangX, LiMM, SunJZ, WangK, LiYH, ZhengYH, YaoYH, LiWB ( 2019). GmRAV1 regulates regeneration of roots and adventitious buds by the cytokinin signaling pathway in Arabidopsis and soybean Physiol Plant 165, 814-829. [本文引用: 1]
ZhaoXH, CaoD, HuangZJ, WangJL, LuSJ, XuY, LiuBH, KongFJ, YuanXH ( 2015). Dual functions of GmTOE4a in the regulation of photoperiod-mediated flowering and plant morphology in soybean Plant Mol Biol 88, 343-355. [本文引用: 1]
ZhaoXY, QiCH, JiangH, YouCX, GuanQM, MaFW, LiYY, HaoYJ ( 2019). The MdWRKY31 transcription factor binds to the MdRAV1 promoter to mediate ABA sensitivity Hortic Res 6, 66. [本文引用: 1]
ZhouXF, WangGD, SutohK, ZhuJK, ZhangWX ( 2008). Identification of cold-inducible microRNAs in plants by transcriptome analysis Biochim Biophys Acta 1779, 780-788. [本文引用: 1]
ZhuQH, HelliwellCA ( 2011). Regulation of flowering time and floral patterning by miR172 J Exp Bot 62, 487-495. [本文引用: 2]
ZhuQH, UpadhyayaNM, GublerF, HelliwellCA ( 2009). Over-expression of miR172 causes loss of spikelet determinacy and floral organ abnormalities in rice ( Oryza sativa) BMC Plant Biol 9, 149. [本文引用: 4]
ZouYM, WangYN, WangLX, YangL, WangR, LiX ( 2013). miR172b controls the transition to autotrophic development inhibited by ABA in Arabidopsis PLoS One 8, e64770. [本文引用: 2]
油菜miRNA研究现状与展望 1 2018
... MicroRNA (miRNA)是广泛存在于真核生物中、由20-24个核苷酸组成的非编码RNA, 在转录后水平调控靶基因表达(Carrington and Ambros, 2003; Bartel, 2004).miRNA在调控植物种子休眠、粒形和株型等生长发育过程及逆境响应中发挥重要作用(王幼宁等, 2016; Saminathan et al., 2019; Miao et al., 2019, 2020).植物中miRNA生成过程与编码蛋白基因类似, 起初由RNA聚合酶II转录形成一段称作初级转录物的pri-miRNA的核苷酸序列, 长度几百到几千个碱基不等, 且在5′和3′端分别带有甲基鸟苷帽子和polyA尾巴(Rogers and Chen, 2013).pri-miRNA根据其自身序列的互补性可形成1至数个发夹茎环结构, 进而在细胞核内被Dicer-like DCL1酶、双链结合蛋白辅因子HYL1以及锌指蛋白SE切割成具有茎环结构的前体miRNA (pre-miRNA).pre-miRNA很不稳定, 很快又被DCL1、HYL1和SE进一步剪切成约22 nt的miRNA-miRNA*二聚体(Kurihara and Watanabe, 2004; Jones-Rhoades et al., 2006; 陈丽等, 2018), 该二聚体首先被HEN1 (HUAENHAN-CER1)甲基化来增强其稳定性, 随后在转运蛋白HST (HASTY)的作用下进入沉默复合体(RISC)状态, 且其中一条成熟单链miRNA保留在复合体中, 与互补的靶标mRNA特异性结合来调控其表达水平(Li et al., 2005; Park et al., 2005; Brodersen et al., 2008; 张俊红等, 2014). ...
降解组测序技术在植物miRNA研究中的应用 1 2013
... 目前, 常用的miRNA靶基因鉴定方法包括生物信息学预测、AGO蛋白免疫共沉淀、双荧光素酶和降解组测序等(董淼等, 2013).miRNA主要通过剪切靶基因mRNA和抑制靶基因翻译2种途径调控靶基因, 但具体方式取决于miRNA与其靶基因mRNA的互补程度及作用位置(Hutvágner and Zamore, 2002; Llave et al., 2002; Schwab et al., 2005; Jung et al., 2007; Zhu et al., 2009).miRNA172与AP2家族基因的互补区域通常位于靠近3°UTR的CDS区, 主要调控方式为抑制AP2靶基因翻译, 但也存在部分与靶基因mRNA完全互补从而对其进行剪切调控的情况(Aukerman and Sakai, 2003; Schmid et al., 2003; Chen, 2004; Mathieu et al., 2009).当miRNA通过抑制翻译对靶基因进行调控时, 两者将表现出协同表达模式(Zhu et al., 2009).例如, 水稻中miRNA172及其靶基因OsIDS1和RSR1在包括胚和胚乳在内的大多数组织中表达模式一致; 同样, 过量表达miR172也会导致各发育阶段穗子中的OsIDS1 (OsINDETERMINATE SPIKELET1)和RSR1 (Rice Starch Regulator1)表达呈现出升高或降低的动态变化模式, 表明miR172对靶基因的调控具有时空特异性.在玉米(Zea mays)中, miR172通过剪切靶基因GL15的mRNA来调控玉米从营养生长向生殖生长的过渡(Lauter et al., 2005).此外, 在大豆(Glycine max)、马铃薯(Solanum tuberosum)和拟南芥中也有类似报道, 即miR172通过切割其靶基因的mRNA来调控植物的生长发育(Wang et al., 2014). ...
植物miR172家族成员进化与分子特性分析 3 2018
... miRNA家族中, miR172是较早被发现同时也是被研究最透彻的成员之一(Park et al., 2002), 其靶基因主要编码AP2/ERF (APETALA2/ethylene responsive factor)类转录因子.AP2/ERF转录因子为植物所特有, 且几乎参与植物生长发育调控的各个环节.miRNA172通过特异或非特异性结合AP2/ERF的mRNA, 以mRNA剪切和翻译抑制2种方式调控AP2/ERF靶基因的表达, 从而调控植物众多生长发育过程(Jung et al., 2007; Zhu et al., 2009; 刘炜婳等, 2018).近期, 研究者在miRNA172-AP2模块调控植物花器官发育、时序转换、块茎及果实发育、根瘤形成和胁迫响应等方面取得一系列研究进展(王幼宁等, 2016; 赵晓晖等, 2017; Luan et al., 2018; Saminathan et al., 2019).本文以miRNA172-AP2模块为中心, 在简要介绍miR172和AP2的结构、功能及互作模式基础上, 重点论述miRNA172-AP2模块参与的植物生长发育及胁迫响应调控, 深入解析其生物学效应和分子机制, 并初步建立由miR172-AP2模块介导的植物生长发育分子调控网络. ...
... 虽然miR172家族成员成熟miRNA的序列高度保守, 但功能上并非简单重复.研究表明, miR172各成员启动子序列及其包含的顺式作用元件并不相同, 从而决定了它们表达的时空特异性(Zhu and Helliwell, 2011; 刘炜婳等, 2018).双子叶模式植物拟南芥(Arabidopsis thaliana)中存在5个miR172成员, 即miR172a-1、miR172a-2、miR172b-1、miR172b-2和miR172c (Chen, 2004).虽然拟南芥miR172各成员的表达模式不尽相同, 但根据其表达量和表达模式总体可分为2类: 一类包括miR172b-2和miR172c, 它们在拟南芥生长发育过程中表达量一直很低; 另一类包括miR172a-1、miR172a-2和miR172b-1, 它们的表达量随着拟南芥的生长发育逐渐升高, 到生殖生长阶段达到峰值(Jung et al., 2007).在表达的组织特异性方面, 除了在植物根中未检测到miR172的表达, 在其它包括花、叶片、茎和角果等组织中均检测到其表达(赵晓晖等, 2017).单子叶模式植物水稻(Oryza sativa)中存在4个miR172成员(miR172a-miR172d), 它们在苗期表达均较强, 但同样各成员表达也具有一定的组织特异性.例如, miR172c在水稻籽粒中不表达(Zhu et al., 2009).综上, miR172在植物中的表达具有明显的时空特异性(Zhu and Helliwell, 2011; 赵晓晖等, 2017). ...
... 植物不像动物可以躲避逆境, 因此其进化出特有的复杂且精细的分子调控机制来应对各种环境胁迫.许多研究表明, miRNA在植物响应各种生物和非生物胁迫过程中具有重要作用.miR172作为miRNA家族中非常保守的一员, 其自身表达受低温、盐碱和干旱等多种胁迫诱导, 同时它也可通过调控AP2类转录因子参与植物响应各种环境胁迫(Zhou et al., 2008; Frazier et al., 2011; Yang et al., 2013; Candar-Cakir et al., 2016).当然, miR172-AP2的具体调控模式还与植物种类及其响应的逆境因子密切相关.拟南芥中, ABA及渗透胁迫可诱导miR172与其靶基因SNZ共表达.miR172b过表达或snz突变体对ABA及渗透胁迫的敏感性增强, 证明miR172通过控制其靶基因SNZ来介导拟南芥对渗透胁迫的响应(Zou et al., 2013).干旱胁迫可诱导miR172e的表达, 同时过表达miR172e又可显著增强拟南芥对干旱的抗性.进一步研究表明, 干旱通过诱导GI (GIGANTEA)表达促进miR172e的积累以抑制下游靶基因TOE1的表达, 进而影响TOE1蛋白与WRKY44的互作, 最终实现对植物抗旱能力的调控(Han et al., 2013).在大豆中过表达miRNA172c或敲除GmNNC1均可增强大豆的耐盐性; 反之, 干扰miR172的表达或过表达GmNNC1均增强了大豆的盐敏感性(Sahito et al., 2017).此外, 在拟南芥中过表达大豆miR172c也可显著增强拟南芥对干旱及盐胁迫的耐受性(Zou et al., 2013).尽管目前人们对拟南芥和大豆中miR172介导的植物响应逆境胁迫机制有了较清晰的认识, 但在其它植物特别是水稻和小麦等重要作物中的相关研究仍停留在全基因组表达分析层面, 还未涉及具体分子机制的解析与调控网络的构建.例如, 香蕉(Musa nana)遇到冷胁迫后miR172的表达会显著上调, 初步推测miR172可能在冷应激反应中起中心协调作用(刘炜婳等, 2018).此外, 在番茄(Lycopersicon esculentum)中过表达miR172a和miR172b显著增强了其对致病疫霉的抗性, 潜在分子机理也与以miR172-AP2模块为核心的表达调控路径相关(Luan et al., 2018). ...
microRNA172参与植物生长发育及逆境响应的研究进展 3 2016
... MicroRNA (miRNA)是广泛存在于真核生物中、由20-24个核苷酸组成的非编码RNA, 在转录后水平调控靶基因表达(Carrington and Ambros, 2003; Bartel, 2004).miRNA在调控植物种子休眠、粒形和株型等生长发育过程及逆境响应中发挥重要作用(王幼宁等, 2016; Saminathan et al., 2019; Miao et al., 2019, 2020).植物中miRNA生成过程与编码蛋白基因类似, 起初由RNA聚合酶II转录形成一段称作初级转录物的pri-miRNA的核苷酸序列, 长度几百到几千个碱基不等, 且在5′和3′端分别带有甲基鸟苷帽子和polyA尾巴(Rogers and Chen, 2013).pri-miRNA根据其自身序列的互补性可形成1至数个发夹茎环结构, 进而在细胞核内被Dicer-like DCL1酶、双链结合蛋白辅因子HYL1以及锌指蛋白SE切割成具有茎环结构的前体miRNA (pre-miRNA).pre-miRNA很不稳定, 很快又被DCL1、HYL1和SE进一步剪切成约22 nt的miRNA-miRNA*二聚体(Kurihara and Watanabe, 2004; Jones-Rhoades et al., 2006; 陈丽等, 2018), 该二聚体首先被HEN1 (HUAENHAN-CER1)甲基化来增强其稳定性, 随后在转运蛋白HST (HASTY)的作用下进入沉默复合体(RISC)状态, 且其中一条成熟单链miRNA保留在复合体中, 与互补的靶标mRNA特异性结合来调控其表达水平(Li et al., 2005; Park et al., 2005; Brodersen et al., 2008; 张俊红等, 2014). ...
... miRNA家族中, miR172是较早被发现同时也是被研究最透彻的成员之一(Park et al., 2002), 其靶基因主要编码AP2/ERF (APETALA2/ethylene responsive factor)类转录因子.AP2/ERF转录因子为植物所特有, 且几乎参与植物生长发育调控的各个环节.miRNA172通过特异或非特异性结合AP2/ERF的mRNA, 以mRNA剪切和翻译抑制2种方式调控AP2/ERF靶基因的表达, 从而调控植物众多生长发育过程(Jung et al., 2007; Zhu et al., 2009; 刘炜婳等, 2018).近期, 研究者在miRNA172-AP2模块调控植物花器官发育、时序转换、块茎及果实发育、根瘤形成和胁迫响应等方面取得一系列研究进展(王幼宁等, 2016; 赵晓晖等, 2017; Luan et al., 2018; Saminathan et al., 2019).本文以miRNA172-AP2模块为中心, 在简要介绍miR172和AP2的结构、功能及互作模式基础上, 重点论述miRNA172-AP2模块参与的植物生长发育及胁迫响应调控, 深入解析其生物学效应和分子机制, 并初步建立由miR172-AP2模块介导的植物生长发育分子调控网络. ...
... miR172在植物的生殖转换、花器官形成及胁迫响应等方面具有重要调控作用(Wu et al., 2009; 王幼宁等, 2016; 赵晓晖等, 2017; Luan et al., 2018; Saminathan et al., 2019).miR172在高等植物中高度保守, 虽然不同物种中miR172成员的数目和长度可能不同, 但在亲缘关系较近的物种间序列高度保守, 暗示其在不同物种中可能具有相似的调控方式和生物学功能. ...
植物成熟microRNA转录后修饰与降解的研究进展 1 2014
... MicroRNA (miRNA)是广泛存在于真核生物中、由20-24个核苷酸组成的非编码RNA, 在转录后水平调控靶基因表达(Carrington and Ambros, 2003; Bartel, 2004).miRNA在调控植物种子休眠、粒形和株型等生长发育过程及逆境响应中发挥重要作用(王幼宁等, 2016; Saminathan et al., 2019; Miao et al., 2019, 2020).植物中miRNA生成过程与编码蛋白基因类似, 起初由RNA聚合酶II转录形成一段称作初级转录物的pri-miRNA的核苷酸序列, 长度几百到几千个碱基不等, 且在5′和3′端分别带有甲基鸟苷帽子和polyA尾巴(Rogers and Chen, 2013).pri-miRNA根据其自身序列的互补性可形成1至数个发夹茎环结构, 进而在细胞核内被Dicer-like DCL1酶、双链结合蛋白辅因子HYL1以及锌指蛋白SE切割成具有茎环结构的前体miRNA (pre-miRNA).pre-miRNA很不稳定, 很快又被DCL1、HYL1和SE进一步剪切成约22 nt的miRNA-miRNA*二聚体(Kurihara and Watanabe, 2004; Jones-Rhoades et al., 2006; 陈丽等, 2018), 该二聚体首先被HEN1 (HUAENHAN-CER1)甲基化来增强其稳定性, 随后在转运蛋白HST (HASTY)的作用下进入沉默复合体(RISC)状态, 且其中一条成熟单链miRNA保留在复合体中, 与互补的靶标mRNA特异性结合来调控其表达水平(Li et al., 2005; Park et al., 2005; Brodersen et al., 2008; 张俊红等, 2014). ...
植物miR172及其靶基因调控开花与发育的研究进展 4 2017
... miRNA家族中, miR172是较早被发现同时也是被研究最透彻的成员之一(Park et al., 2002), 其靶基因主要编码AP2/ERF (APETALA2/ethylene responsive factor)类转录因子.AP2/ERF转录因子为植物所特有, 且几乎参与植物生长发育调控的各个环节.miRNA172通过特异或非特异性结合AP2/ERF的mRNA, 以mRNA剪切和翻译抑制2种方式调控AP2/ERF靶基因的表达, 从而调控植物众多生长发育过程(Jung et al., 2007; Zhu et al., 2009; 刘炜婳等, 2018).近期, 研究者在miRNA172-AP2模块调控植物花器官发育、时序转换、块茎及果实发育、根瘤形成和胁迫响应等方面取得一系列研究进展(王幼宁等, 2016; 赵晓晖等, 2017; Luan et al., 2018; Saminathan et al., 2019).本文以miRNA172-AP2模块为中心, 在简要介绍miR172和AP2的结构、功能及互作模式基础上, 重点论述miRNA172-AP2模块参与的植物生长发育及胁迫响应调控, 深入解析其生物学效应和分子机制, 并初步建立由miR172-AP2模块介导的植物生长发育分子调控网络. ...
... miR172在植物的生殖转换、花器官形成及胁迫响应等方面具有重要调控作用(Wu et al., 2009; 王幼宁等, 2016; 赵晓晖等, 2017; Luan et al., 2018; Saminathan et al., 2019).miR172在高等植物中高度保守, 虽然不同物种中miR172成员的数目和长度可能不同, 但在亲缘关系较近的物种间序列高度保守, 暗示其在不同物种中可能具有相似的调控方式和生物学功能. ...
... 虽然miR172家族成员成熟miRNA的序列高度保守, 但功能上并非简单重复.研究表明, miR172各成员启动子序列及其包含的顺式作用元件并不相同, 从而决定了它们表达的时空特异性(Zhu and Helliwell, 2011; 刘炜婳等, 2018).双子叶模式植物拟南芥(Arabidopsis thaliana)中存在5个miR172成员, 即miR172a-1、miR172a-2、miR172b-1、miR172b-2和miR172c (Chen, 2004).虽然拟南芥miR172各成员的表达模式不尽相同, 但根据其表达量和表达模式总体可分为2类: 一类包括miR172b-2和miR172c, 它们在拟南芥生长发育过程中表达量一直很低; 另一类包括miR172a-1、miR172a-2和miR172b-1, 它们的表达量随着拟南芥的生长发育逐渐升高, 到生殖生长阶段达到峰值(Jung et al., 2007).在表达的组织特异性方面, 除了在植物根中未检测到miR172的表达, 在其它包括花、叶片、茎和角果等组织中均检测到其表达(赵晓晖等, 2017).单子叶模式植物水稻(Oryza sativa)中存在4个miR172成员(miR172a-miR172d), 它们在苗期表达均较强, 但同样各成员表达也具有一定的组织特异性.例如, miR172c在水稻籽粒中不表达(Zhu et al., 2009).综上, miR172在植物中的表达具有明显的时空特异性(Zhu and Helliwell, 2011; 赵晓晖等, 2017). ...
... ; 赵晓晖等, 2017). ...
tasselseed1 is a lipoxygenase affecting jasmonic acid signaling in sex determination of maize 1 2009
... miR172-AP2模块除了在拟南芥中参与花器官发育调控, 在烟草(Nicotiana tabacum)中也有类似作用.将拟南芥miR172在烟草中过表达同样会导致烟草花结构发育异常, 与ap2突变体类似(Mlotshwa et al., 2006), 表明miR172-AP2模块在花器官发育调控方面具保守性.最新研究表明, miR172-AP2模块在油菜(Brassica campestris)、大岩桐(Sinningia speciosa)和玫瑰(Rosa rugosa)等双子叶植物中也参与花器官发育的调控(François et al., 2018; Li et al., 2019b; Wang et al., 2019b).此外, 该调控模式也存在于单子叶植物中.例如, 水稻AP2家族成员OsSNB和OsIDS1在小穗向小花转变及花器官形态建成过程中发挥关键作用.OsSNB突变会导致花发育异常.例如, 小穗向小花转变显著延迟, 产生额外颖苞, 花中浆片异常膨大进而发育成内外稃等; 在OsSNB单突变基础上同时突变OsIDS1则会进一步增强相关表型.水稻中过表达miR172也会导致花器官发育异常, 形成类似Ossnb突变体的表型(Lee and An, 2012; Wang et al., 2015a), 暗示miR172可能通过调控AP2基因OsSNB和OsIDS1的表达来影响水稻花器官的发育.另一禾本科单子叶植物玉米, 其花为单性花, 含有雄穗和雌穗两种花序.玉米的ts (tasselseed)突变体由于雄穗中的雌蕊不退化从而形成两性花, 导致雄穗雌化并进而开花结实(Chuck et al., 2007; Acosta et al., 2009).利用具有雄穗雌化现象的玉米突变体ts4和ts6开展目标基因克隆, 结果显示这2种突变体分别是由于miR172及其靶基因IDS1突变所致, 表明miR172-AP2模块在玉米性别决定中同样具有关键作用(Chuck et al., 2007).而IDS1的异位表达会推迟小穗向花转变, 从而使单个小穗产生多个小花(大于2朵).IDS1同源基因SID1 (SISTER OF INDETERMINATE SPIKELET 1)突变会进一步增强ids1突变体的表型, 表明这2个基因在玉米花器官发育过程中的功能具有累加作用(Chuck et al., 2008).禾本科植物的浆片在开花过程中具有重要作用, 即在特定时期吸水膨胀, 从而撑开颖壳, 完成开花过程.已知大麦(Hordeum vulgare)的闭花受精特性由AP2家族的Cly1 (Cleistogamy 1)基因控制, 该基因表达也受到miR172调控; 当Cly1序列中与miR172结合的位点发生突变时, 会干扰其mRNA与miR172结合, 从而减弱miR172的翻译抑制作用, 进而产生非常小的浆片而无法开颖, 最终发生闭花受精(Nair et al., 2009; Anwar et al., 2018).可见, miR172-AP2模块在不同植物中的功能非常保守, 且对植物花器官发育调控极为重要, miR172通过抑制AP2家族成员的表达, 进而影响AP2基因与B类和C类基因间的互作, 最终调控花器官发育、雌雄分化和开花等过程. ...
Overexpression of PgDREB2A transcription factor enhances abiotic stress tolerance and activates downstream stress-responsive genes 1 2010
... 各AP2/ERF亚家族转录因子所识别的顺式作用元件具有一定的特异性.例如, AP2亚家族可与启动子中gCAC(A/G)N(A/T)TcCC(a/g)ANG(c/t)顺式作用元件结合来调节靶基因的表达, 进而调控植物的花器官发育、开花时间及时序转换等(Boutilier et al., 2002; Lei et al., 2019; Wang et al., 2019b).RVA亚家族由于含有AP2/ERF和B3结构域, 因此可识别CAACA和CACCTG基序.ERF亚家族主要识别启动子区域的GCC-box, DREB亚家族则可与干旱胁迫响应和冷诱导响应元件(A/GCCGAC)结合, 从而调节植物的非生物胁迫响应(Dubouzet et al., 2003; Agarwal et al., 2010; Mizoi et al., 2012; Bäumler et al., 2019; Wang et al., 2019a). ...
TEMPRANILLO is a direct repressor of the microRNA miR172 1 2019
MicroRNAs: genomics, biogenesis, mechanism, and function 1 2004
... MicroRNA (miRNA)是广泛存在于真核生物中、由20-24个核苷酸组成的非编码RNA, 在转录后水平调控靶基因表达(Carrington and Ambros, 2003; Bartel, 2004).miRNA在调控植物种子休眠、粒形和株型等生长发育过程及逆境响应中发挥重要作用(王幼宁等, 2016; Saminathan et al., 2019; Miao et al., 2019, 2020).植物中miRNA生成过程与编码蛋白基因类似, 起初由RNA聚合酶II转录形成一段称作初级转录物的pri-miRNA的核苷酸序列, 长度几百到几千个碱基不等, 且在5′和3′端分别带有甲基鸟苷帽子和polyA尾巴(Rogers and Chen, 2013).pri-miRNA根据其自身序列的互补性可形成1至数个发夹茎环结构, 进而在细胞核内被Dicer-like DCL1酶、双链结合蛋白辅因子HYL1以及锌指蛋白SE切割成具有茎环结构的前体miRNA (pre-miRNA).pre-miRNA很不稳定, 很快又被DCL1、HYL1和SE进一步剪切成约22 nt的miRNA-miRNA*二聚体(Kurihara and Watanabe, 2004; Jones-Rhoades et al., 2006; 陈丽等, 2018), 该二聚体首先被HEN1 (HUAENHAN-CER1)甲基化来增强其稳定性, 随后在转运蛋白HST (HASTY)的作用下进入沉默复合体(RISC)状态, 且其中一条成熟单链miRNA保留在复合体中, 与互补的靶标mRNA特异性结合来调控其表达水平(Li et al., 2005; Park et al., 2005; Brodersen et al., 2008; 张俊红等, 2014). ...
AtERF#111/ABR1 is a transcriptional activator involved in the wounding response 2 2019
... AP2/ERF类转录因子均含有1个由60-70个氨基酸残基组成的高度保守的AP2/ERF结构域(Riechmann and Meyerowitz, 1998).该结构域包含3个反向平行的β折叠和1个几乎与β折叠平行的α螺旋.β折叠在识别目标基因的顺式作用元件上具有重要作用, α螺旋可能介导与其它转录因子的互作(Nakano et al., 2006).根据保守结构域差异, 可将AP2/ERF转录因子进一步分为4个亚家族, 即AP2、ERF、DREB (dehydration-responsive element binding protein)和RAV (related to ABI3/VP1) (Xie et al., 2019).AP2亚家族含有2个AP2/ERF结构域, 主要参与调控植物花器官发育; 另外, 最新研究表明, 该亚家族还参与调控植物根系发育和孤雌生殖(Khanday et al., 2019; Li et al., 2019a).RAV亚家族含有1个AP2/ERF及1个B3结构域(Swaminathan et al., 2008), 该亚家族成员参与植物激素及胁迫响应(Hu et al., 2004; Sohn et al., 2006; Zhang et al., 2019; Zhao et al., 2019).ERF和DREB亚家族均仅包含1个AP2/ERF结构域, 两者的区别在于AP2/ERF结构域的第14和19位氨基酸不同, DREB亚家族这2个位置分别是缬氨酸(Val)和谷氨酸(Glu), 而ERF亚家族分别为丙氨酸(Ala)和天冬氨酸(Asp), 功能上两亚家族均参与植物响应生物和非生物胁迫(Dubouzet et al., 2003; Bäumler et al., 2019; He et al., 2019; Wang et al., 2019a). ...
... 各AP2/ERF亚家族转录因子所识别的顺式作用元件具有一定的特异性.例如, AP2亚家族可与启动子中gCAC(A/G)N(A/T)TcCC(a/g)ANG(c/t)顺式作用元件结合来调节靶基因的表达, 进而调控植物的花器官发育、开花时间及时序转换等(Boutilier et al., 2002; Lei et al., 2019; Wang et al., 2019b).RVA亚家族由于含有AP2/ERF和B3结构域, 因此可识别CAACA和CACCTG基序.ERF亚家族主要识别启动子区域的GCC-box, DREB亚家族则可与干旱胁迫响应和冷诱导响应元件(A/GCCGAC)结合, 从而调节植物的非生物胁迫响应(Dubouzet et al., 2003; Agarwal et al., 2010; Mizoi et al., 2012; Bäumler et al., 2019; Wang et al., 2019a). ...
MicroRNA166 controls root and nodule development in Medicago truncatula 1 2008
... 根瘤菌不仅能感染豆科植物根系形成根瘤, 还可与其共生固氮.在苜蓿(Medicago sativa)中首先证实miR166和miR169参与根瘤发育的调控(Combier et al., 2006; Boualem et al., 2008).百脉根(Lotus corniculatus)中的研究也表明, miR171、miR397和miR156与根瘤的发育、病毒感染和固氮能力密切相关(De Luis et al., 2012; Wang et al., 2015b).在大豆中过表达miR482、miR1512、miR1515、miR156及miR167均可以显著改变大豆根瘤的数量(Li et al., 2010; Wang et al., 2015c).因此, miRNA在豆科植物结瘤和共生固氮过程中发挥重要作用. ...
Ectopic expression of BABY BOOM triggers a conversion from vegetative to embryonic growth 1 2002
... 各AP2/ERF亚家族转录因子所识别的顺式作用元件具有一定的特异性.例如, AP2亚家族可与启动子中gCAC(A/G)N(A/T)TcCC(a/g)ANG(c/t)顺式作用元件结合来调节靶基因的表达, 进而调控植物的花器官发育、开花时间及时序转换等(Boutilier et al., 2002; Lei et al., 2019; Wang et al., 2019b).RVA亚家族由于含有AP2/ERF和B3结构域, 因此可识别CAACA和CACCTG基序.ERF亚家族主要识别启动子区域的GCC-box, DREB亚家族则可与干旱胁迫响应和冷诱导响应元件(A/GCCGAC)结合, 从而调节植物的非生物胁迫响应(Dubouzet et al., 2003; Agarwal et al., 2010; Mizoi et al., 2012; Bäumler et al., 2019; Wang et al., 2019a). ...
The ABC model of flower development: then and now 1 2012
... AP2在模式植物拟南芥中被发现时便认为与植物花器官特征调控相关(Kunst et al., 1989).拟南芥的花包括花萼、花瓣、雄蕊和心皮4轮器官, 其特征由A、B和C三类基因控制(Bowman et al., 2012).AP2属于A类基因, 不仅参与第1轮花萼特征的调控, 还与B类基因AP3和PI (PISTILLATA)共同控制第2轮花瓣的形成; 并可通过拮抗C类基因AG (AGAMOUS)的作用抑制雄蕊和心皮的发育(Shannon and Meeks- Wagner, 1993; Elliott et al., 1996).miR172在拟南芥花器官中具有优势表达, 且miR172过表达植株花器官发育异常, 与ap2突变体类似(Park et al., 2002), 主要原因在于miR172从翻译水平抑制了AP2的表达(Aukerman and Sakai, 2003; Chen, 2004).时空表达分析结果表明, AP2表达产物主要积累于花器官外周, 与miR172信号在第2或第3轮花器官处有瞬时重叠(Wollmann et al., 2010).尽管miR172和AP2共表达时间非常短暂, 但两者共同调控花分生细胞的时序进程, 在花器官发育中发挥决定性作用(Ji et al., 2011). ...
Widespread translational inhibition by plant miRNAs and siRNAs 1 2008
... MicroRNA (miRNA)是广泛存在于真核生物中、由20-24个核苷酸组成的非编码RNA, 在转录后水平调控靶基因表达(Carrington and Ambros, 2003; Bartel, 2004).miRNA在调控植物种子休眠、粒形和株型等生长发育过程及逆境响应中发挥重要作用(王幼宁等, 2016; Saminathan et al., 2019; Miao et al., 2019, 2020).植物中miRNA生成过程与编码蛋白基因类似, 起初由RNA聚合酶II转录形成一段称作初级转录物的pri-miRNA的核苷酸序列, 长度几百到几千个碱基不等, 且在5′和3′端分别带有甲基鸟苷帽子和polyA尾巴(Rogers and Chen, 2013).pri-miRNA根据其自身序列的互补性可形成1至数个发夹茎环结构, 进而在细胞核内被Dicer-like DCL1酶、双链结合蛋白辅因子HYL1以及锌指蛋白SE切割成具有茎环结构的前体miRNA (pre-miRNA).pre-miRNA很不稳定, 很快又被DCL1、HYL1和SE进一步剪切成约22 nt的miRNA-miRNA*二聚体(Kurihara and Watanabe, 2004; Jones-Rhoades et al., 2006; 陈丽等, 2018), 该二聚体首先被HEN1 (HUAENHAN-CER1)甲基化来增强其稳定性, 随后在转运蛋白HST (HASTY)的作用下进入沉默复合体(RISC)状态, 且其中一条成熟单链miRNA保留在复合体中, 与互补的靶标mRNA特异性结合来调控其表达水平(Li et al., 2005; Park et al., 2005; Brodersen et al., 2008; 张俊红等, 2014). ...
Small RNA and degradome deep sequencing reveals drought-and tissue-specific micrornas and their important roles in drought-sensitive and drought-tolerant tomato genotypes 1 2016
... 植物不像动物可以躲避逆境, 因此其进化出特有的复杂且精细的分子调控机制来应对各种环境胁迫.许多研究表明, miRNA在植物响应各种生物和非生物胁迫过程中具有重要作用.miR172作为miRNA家族中非常保守的一员, 其自身表达受低温、盐碱和干旱等多种胁迫诱导, 同时它也可通过调控AP2类转录因子参与植物响应各种环境胁迫(Zhou et al., 2008; Frazier et al., 2011; Yang et al., 2013; Candar-Cakir et al., 2016).当然, miR172-AP2的具体调控模式还与植物种类及其响应的逆境因子密切相关.拟南芥中, ABA及渗透胁迫可诱导miR172与其靶基因SNZ共表达.miR172b过表达或snz突变体对ABA及渗透胁迫的敏感性增强, 证明miR172通过控制其靶基因SNZ来介导拟南芥对渗透胁迫的响应(Zou et al., 2013).干旱胁迫可诱导miR172e的表达, 同时过表达miR172e又可显著增强拟南芥对干旱的抗性.进一步研究表明, 干旱通过诱导GI (GIGANTEA)表达促进miR172e的积累以抑制下游靶基因TOE1的表达, 进而影响TOE1蛋白与WRKY44的互作, 最终实现对植物抗旱能力的调控(Han et al., 2013).在大豆中过表达miRNA172c或敲除GmNNC1均可增强大豆的耐盐性; 反之, 干扰miR172的表达或过表达GmNNC1均增强了大豆的盐敏感性(Sahito et al., 2017).此外, 在拟南芥中过表达大豆miR172c也可显著增强拟南芥对干旱及盐胁迫的耐受性(Zou et al., 2013).尽管目前人们对拟南芥和大豆中miR172介导的植物响应逆境胁迫机制有了较清晰的认识, 但在其它植物特别是水稻和小麦等重要作物中的相关研究仍停留在全基因组表达分析层面, 还未涉及具体分子机制的解析与调控网络的构建.例如, 香蕉(Musa nana)遇到冷胁迫后miR172的表达会显著上调, 初步推测miR172可能在冷应激反应中起中心协调作用(刘炜婳等, 2018).此外, 在番茄(Lycopersicon esculentum)中过表达miR172a和miR172b显著增强了其对致病疫霉的抗性, 潜在分子机理也与以miR172-AP2模块为核心的表达调控路径相关(Luan et al., 2018). ...
Role of microRNAs in plant and animal development 1 2003
... MicroRNA (miRNA)是广泛存在于真核生物中、由20-24个核苷酸组成的非编码RNA, 在转录后水平调控靶基因表达(Carrington and Ambros, 2003; Bartel, 2004).miRNA在调控植物种子休眠、粒形和株型等生长发育过程及逆境响应中发挥重要作用(王幼宁等, 2016; Saminathan et al., 2019; Miao et al., 2019, 2020).植物中miRNA生成过程与编码蛋白基因类似, 起初由RNA聚合酶II转录形成一段称作初级转录物的pri-miRNA的核苷酸序列, 长度几百到几千个碱基不等, 且在5′和3′端分别带有甲基鸟苷帽子和polyA尾巴(Rogers and Chen, 2013).pri-miRNA根据其自身序列的互补性可形成1至数个发夹茎环结构, 进而在细胞核内被Dicer-like DCL1酶、双链结合蛋白辅因子HYL1以及锌指蛋白SE切割成具有茎环结构的前体miRNA (pre-miRNA).pre-miRNA很不稳定, 很快又被DCL1、HYL1和SE进一步剪切成约22 nt的miRNA-miRNA*二聚体(Kurihara and Watanabe, 2004; Jones-Rhoades et al., 2006; 陈丽等, 2018), 该二聚体首先被HEN1 (HUAENHAN-CER1)甲基化来增强其稳定性, 随后在转运蛋白HST (HASTY)的作用下进入沉默复合体(RISC)状态, 且其中一条成熟单链miRNA保留在复合体中, 与互补的靶标mRNA特异性结合来调控其表达水平(Li et al., 2005; Park et al., 2005; Brodersen et al., 2008; 张俊红等, 2014). ...
A microRNA as a translational repressor of APETALA2 in Arabidopsis flower development 3 2004
... 虽然miR172家族成员成熟miRNA的序列高度保守, 但功能上并非简单重复.研究表明, miR172各成员启动子序列及其包含的顺式作用元件并不相同, 从而决定了它们表达的时空特异性(Zhu and Helliwell, 2011; 刘炜婳等, 2018).双子叶模式植物拟南芥(Arabidopsis thaliana)中存在5个miR172成员, 即miR172a-1、miR172a-2、miR172b-1、miR172b-2和miR172c (Chen, 2004).虽然拟南芥miR172各成员的表达模式不尽相同, 但根据其表达量和表达模式总体可分为2类: 一类包括miR172b-2和miR172c, 它们在拟南芥生长发育过程中表达量一直很低; 另一类包括miR172a-1、miR172a-2和miR172b-1, 它们的表达量随着拟南芥的生长发育逐渐升高, 到生殖生长阶段达到峰值(Jung et al., 2007).在表达的组织特异性方面, 除了在植物根中未检测到miR172的表达, 在其它包括花、叶片、茎和角果等组织中均检测到其表达(赵晓晖等, 2017).单子叶模式植物水稻(Oryza sativa)中存在4个miR172成员(miR172a-miR172d), 它们在苗期表达均较强, 但同样各成员表达也具有一定的组织特异性.例如, miR172c在水稻籽粒中不表达(Zhu et al., 2009).综上, miR172在植物中的表达具有明显的时空特异性(Zhu and Helliwell, 2011; 赵晓晖等, 2017). ...
... 目前, 常用的miRNA靶基因鉴定方法包括生物信息学预测、AGO蛋白免疫共沉淀、双荧光素酶和降解组测序等(董淼等, 2013).miRNA主要通过剪切靶基因mRNA和抑制靶基因翻译2种途径调控靶基因, 但具体方式取决于miRNA与其靶基因mRNA的互补程度及作用位置(Hutvágner and Zamore, 2002; Llave et al., 2002; Schwab et al., 2005; Jung et al., 2007; Zhu et al., 2009).miRNA172与AP2家族基因的互补区域通常位于靠近3°UTR的CDS区, 主要调控方式为抑制AP2靶基因翻译, 但也存在部分与靶基因mRNA完全互补从而对其进行剪切调控的情况(Aukerman and Sakai, 2003; Schmid et al., 2003; Chen, 2004; Mathieu et al., 2009).当miRNA通过抑制翻译对靶基因进行调控时, 两者将表现出协同表达模式(Zhu et al., 2009).例如, 水稻中miRNA172及其靶基因OsIDS1和RSR1在包括胚和胚乳在内的大多数组织中表达模式一致; 同样, 过量表达miR172也会导致各发育阶段穗子中的OsIDS1 (OsINDETERMINATE SPIKELET1)和RSR1 (Rice Starch Regulator1)表达呈现出升高或降低的动态变化模式, 表明miR172对靶基因的调控具有时空特异性.在玉米(Zea mays)中, miR172通过剪切靶基因GL15的mRNA来调控玉米从营养生长向生殖生长的过渡(Lauter et al., 2005).此外, 在大豆(Glycine max)、马铃薯(Solanum tuberosum)和拟南芥中也有类似报道, 即miR172通过切割其靶基因的mRNA来调控植物的生长发育(Wang et al., 2014). ...
... AP2在模式植物拟南芥中被发现时便认为与植物花器官特征调控相关(Kunst et al., 1989).拟南芥的花包括花萼、花瓣、雄蕊和心皮4轮器官, 其特征由A、B和C三类基因控制(Bowman et al., 2012).AP2属于A类基因, 不仅参与第1轮花萼特征的调控, 还与B类基因AP3和PI (PISTILLATA)共同控制第2轮花瓣的形成; 并可通过拮抗C类基因AG (AGAMOUS)的作用抑制雄蕊和心皮的发育(Shannon and Meeks- Wagner, 1993; Elliott et al., 1996).miR172在拟南芥花器官中具有优势表达, 且miR172过表达植株花器官发育异常, 与ap2突变体类似(Park et al., 2002), 主要原因在于miR172从翻译水平抑制了AP2的表达(Aukerman and Sakai, 2003; Chen, 2004).时空表达分析结果表明, AP2表达产物主要积累于花器官外周, 与miR172信号在第2或第3轮花器官处有瞬时重叠(Wollmann et al., 2010).尽管miR172和AP2共表达时间非常短暂, 但两者共同调控花分生细胞的时序进程, 在花器官发育中发挥决定性作用(Ji et al., 2011). ...
The heterochronic maize mutant Corngrass1 results from overexpression of a tandem microRNA 2 2007
... miR172-AP2模块除了在拟南芥中参与花器官发育调控, 在烟草(Nicotiana tabacum)中也有类似作用.将拟南芥miR172在烟草中过表达同样会导致烟草花结构发育异常, 与ap2突变体类似(Mlotshwa et al., 2006), 表明miR172-AP2模块在花器官发育调控方面具保守性.最新研究表明, miR172-AP2模块在油菜(Brassica campestris)、大岩桐(Sinningia speciosa)和玫瑰(Rosa rugosa)等双子叶植物中也参与花器官发育的调控(François et al., 2018; Li et al., 2019b; Wang et al., 2019b).此外, 该调控模式也存在于单子叶植物中.例如, 水稻AP2家族成员OsSNB和OsIDS1在小穗向小花转变及花器官形态建成过程中发挥关键作用.OsSNB突变会导致花发育异常.例如, 小穗向小花转变显著延迟, 产生额外颖苞, 花中浆片异常膨大进而发育成内外稃等; 在OsSNB单突变基础上同时突变OsIDS1则会进一步增强相关表型.水稻中过表达miR172也会导致花器官发育异常, 形成类似Ossnb突变体的表型(Lee and An, 2012; Wang et al., 2015a), 暗示miR172可能通过调控AP2基因OsSNB和OsIDS1的表达来影响水稻花器官的发育.另一禾本科单子叶植物玉米, 其花为单性花, 含有雄穗和雌穗两种花序.玉米的ts (tasselseed)突变体由于雄穗中的雌蕊不退化从而形成两性花, 导致雄穗雌化并进而开花结实(Chuck et al., 2007; Acosta et al., 2009).利用具有雄穗雌化现象的玉米突变体ts4和ts6开展目标基因克隆, 结果显示这2种突变体分别是由于miR172及其靶基因IDS1突变所致, 表明miR172-AP2模块在玉米性别决定中同样具有关键作用(Chuck et al., 2007).而IDS1的异位表达会推迟小穗向花转变, 从而使单个小穗产生多个小花(大于2朵).IDS1同源基因SID1 (SISTER OF INDETERMINATE SPIKELET 1)突变会进一步增强ids1突变体的表型, 表明这2个基因在玉米花器官发育过程中的功能具有累加作用(Chuck et al., 2008).禾本科植物的浆片在开花过程中具有重要作用, 即在特定时期吸水膨胀, 从而撑开颖壳, 完成开花过程.已知大麦(Hordeum vulgare)的闭花受精特性由AP2家族的Cly1 (Cleistogamy 1)基因控制, 该基因表达也受到miR172调控; 当Cly1序列中与miR172结合的位点发生突变时, 会干扰其mRNA与miR172结合, 从而减弱miR172的翻译抑制作用, 进而产生非常小的浆片而无法开颖, 最终发生闭花受精(Nair et al., 2009; Anwar et al., 2018).可见, miR172-AP2模块在不同植物中的功能非常保守, 且对植物花器官发育调控极为重要, miR172通过抑制AP2家族成员的表达, 进而影响AP2基因与B类和C类基因间的互作, 最终调控花器官发育、雌雄分化和开花等过程. ...
... 突变所致, 表明miR172-AP2模块在玉米性别决定中同样具有关键作用(Chuck et al., 2007).而IDS1的异位表达会推迟小穗向花转变, 从而使单个小穗产生多个小花(大于2朵).IDS1同源基因SID1 (SISTER OF INDETERMINATE SPIKELET 1)突变会进一步增强ids1突变体的表型, 表明这2个基因在玉米花器官发育过程中的功能具有累加作用(Chuck et al., 2008).禾本科植物的浆片在开花过程中具有重要作用, 即在特定时期吸水膨胀, 从而撑开颖壳, 完成开花过程.已知大麦(Hordeum vulgare)的闭花受精特性由AP2家族的Cly1 (Cleistogamy 1)基因控制, 该基因表达也受到miR172调控; 当Cly1序列中与miR172结合的位点发生突变时, 会干扰其mRNA与miR172结合, 从而减弱miR172的翻译抑制作用, 进而产生非常小的浆片而无法开颖, 最终发生闭花受精(Nair et al., 2009; Anwar et al., 2018).可见, miR172-AP2模块在不同植物中的功能非常保守, 且对植物花器官发育调控极为重要, miR172通过抑制AP2家族成员的表达, 进而影响AP2基因与B类和C类基因间的互作, 最终调控花器官发育、雌雄分化和开花等过程. ...
Floral meristem initiation and meristem cell fate are regulated by the maize AP2 genes ids1 and sid1 1 2008
... miR172-AP2模块除了在拟南芥中参与花器官发育调控, 在烟草(Nicotiana tabacum)中也有类似作用.将拟南芥miR172在烟草中过表达同样会导致烟草花结构发育异常, 与ap2突变体类似(Mlotshwa et al., 2006), 表明miR172-AP2模块在花器官发育调控方面具保守性.最新研究表明, miR172-AP2模块在油菜(Brassica campestris)、大岩桐(Sinningia speciosa)和玫瑰(Rosa rugosa)等双子叶植物中也参与花器官发育的调控(François et al., 2018; Li et al., 2019b; Wang et al., 2019b).此外, 该调控模式也存在于单子叶植物中.例如, 水稻AP2家族成员OsSNB和OsIDS1在小穗向小花转变及花器官形态建成过程中发挥关键作用.OsSNB突变会导致花发育异常.例如, 小穗向小花转变显著延迟, 产生额外颖苞, 花中浆片异常膨大进而发育成内外稃等; 在OsSNB单突变基础上同时突变OsIDS1则会进一步增强相关表型.水稻中过表达miR172也会导致花器官发育异常, 形成类似Ossnb突变体的表型(Lee and An, 2012; Wang et al., 2015a), 暗示miR172可能通过调控AP2基因OsSNB和OsIDS1的表达来影响水稻花器官的发育.另一禾本科单子叶植物玉米, 其花为单性花, 含有雄穗和雌穗两种花序.玉米的ts (tasselseed)突变体由于雄穗中的雌蕊不退化从而形成两性花, 导致雄穗雌化并进而开花结实(Chuck et al., 2007; Acosta et al., 2009).利用具有雄穗雌化现象的玉米突变体ts4和ts6开展目标基因克隆, 结果显示这2种突变体分别是由于miR172及其靶基因IDS1突变所致, 表明miR172-AP2模块在玉米性别决定中同样具有关键作用(Chuck et al., 2007).而IDS1的异位表达会推迟小穗向花转变, 从而使单个小穗产生多个小花(大于2朵).IDS1同源基因SID1 (SISTER OF INDETERMINATE SPIKELET 1)突变会进一步增强ids1突变体的表型, 表明这2个基因在玉米花器官发育过程中的功能具有累加作用(Chuck et al., 2008).禾本科植物的浆片在开花过程中具有重要作用, 即在特定时期吸水膨胀, 从而撑开颖壳, 完成开花过程.已知大麦(Hordeum vulgare)的闭花受精特性由AP2家族的Cly1 (Cleistogamy 1)基因控制, 该基因表达也受到miR172调控; 当Cly1序列中与miR172结合的位点发生突变时, 会干扰其mRNA与miR172结合, 从而减弱miR172的翻译抑制作用, 进而产生非常小的浆片而无法开颖, 最终发生闭花受精(Nair et al., 2009; Anwar et al., 2018).可见, miR172-AP2模块在不同植物中的功能非常保守, 且对植物花器官发育调控极为重要, miR172通过抑制AP2家族成员的表达, 进而影响AP2基因与B类和C类基因间的互作, 最终调控花器官发育、雌雄分化和开花等过程. ...
MtHAP2-1 is a key transcriptional regulator of symbiotic nodule development regulated by microRNA169 in Medicago truncatula 1 2006
... 根瘤菌不仅能感染豆科植物根系形成根瘤, 还可与其共生固氮.在苜蓿(Medicago sativa)中首先证实miR166和miR169参与根瘤发育的调控(Combier et al., 2006; Boualem et al., 2008).百脉根(Lotus corniculatus)中的研究也表明, miR171、miR397和miR156与根瘤的发育、病毒感染和固氮能力密切相关(De Luis et al., 2012; Wang et al., 2015b).在大豆中过表达miR482、miR1512、miR1515、miR156及miR167均可以显著改变大豆根瘤的数量(Li et al., 2010; Wang et al., 2015c).因此, miRNA在豆科植物结瘤和共生固氮过程中发挥重要作用. ...
Two microRNAs linked to nodule infection and nitrogen-fixing ability in the legume Lotus japonicus 1 2012
... 根瘤菌不仅能感染豆科植物根系形成根瘤, 还可与其共生固氮.在苜蓿(Medicago sativa)中首先证实miR166和miR169参与根瘤发育的调控(Combier et al., 2006; Boualem et al., 2008).百脉根(Lotus corniculatus)中的研究也表明, miR171、miR397和miR156与根瘤的发育、病毒感染和固氮能力密切相关(De Luis et al., 2012; Wang et al., 2015b).在大豆中过表达miR482、miR1512、miR1515、miR156及miR167均可以显著改变大豆根瘤的数量(Li et al., 2010; Wang et al., 2015c).因此, miRNA在豆科植物结瘤和共生固氮过程中发挥重要作用. ...
OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in drought-, high-salt- and cold-responsive gene expression 2 2003
... AP2/ERF类转录因子均含有1个由60-70个氨基酸残基组成的高度保守的AP2/ERF结构域(Riechmann and Meyerowitz, 1998).该结构域包含3个反向平行的β折叠和1个几乎与β折叠平行的α螺旋.β折叠在识别目标基因的顺式作用元件上具有重要作用, α螺旋可能介导与其它转录因子的互作(Nakano et al., 2006).根据保守结构域差异, 可将AP2/ERF转录因子进一步分为4个亚家族, 即AP2、ERF、DREB (dehydration-responsive element binding protein)和RAV (related to ABI3/VP1) (Xie et al., 2019).AP2亚家族含有2个AP2/ERF结构域, 主要参与调控植物花器官发育; 另外, 最新研究表明, 该亚家族还参与调控植物根系发育和孤雌生殖(Khanday et al., 2019; Li et al., 2019a).RAV亚家族含有1个AP2/ERF及1个B3结构域(Swaminathan et al., 2008), 该亚家族成员参与植物激素及胁迫响应(Hu et al., 2004; Sohn et al., 2006; Zhang et al., 2019; Zhao et al., 2019).ERF和DREB亚家族均仅包含1个AP2/ERF结构域, 两者的区别在于AP2/ERF结构域的第14和19位氨基酸不同, DREB亚家族这2个位置分别是缬氨酸(Val)和谷氨酸(Glu), 而ERF亚家族分别为丙氨酸(Ala)和天冬氨酸(Asp), 功能上两亚家族均参与植物响应生物和非生物胁迫(Dubouzet et al., 2003; Bäumler et al., 2019; He et al., 2019; Wang et al., 2019a). ...
... 各AP2/ERF亚家族转录因子所识别的顺式作用元件具有一定的特异性.例如, AP2亚家族可与启动子中gCAC(A/G)N(A/T)TcCC(a/g)ANG(c/t)顺式作用元件结合来调节靶基因的表达, 进而调控植物的花器官发育、开花时间及时序转换等(Boutilier et al., 2002; Lei et al., 2019; Wang et al., 2019b).RVA亚家族由于含有AP2/ERF和B3结构域, 因此可识别CAACA和CACCTG基序.ERF亚家族主要识别启动子区域的GCC-box, DREB亚家族则可与干旱胁迫响应和冷诱导响应元件(A/GCCGAC)结合, 从而调节植物的非生物胁迫响应(Dubouzet et al., 2003; Agarwal et al., 2010; Mizoi et al., 2012; Bäumler et al., 2019; Wang et al., 2019a). ...
AINTEGUMENTA, an APETALA2-like gene of Arabidopsis with pleiotropic roles in ovule development and floral organ growth 1 1996
... AP2在模式植物拟南芥中被发现时便认为与植物花器官特征调控相关(Kunst et al., 1989).拟南芥的花包括花萼、花瓣、雄蕊和心皮4轮器官, 其特征由A、B和C三类基因控制(Bowman et al., 2012).AP2属于A类基因, 不仅参与第1轮花萼特征的调控, 还与B类基因AP3和PI (PISTILLATA)共同控制第2轮花瓣的形成; 并可通过拮抗C类基因AG (AGAMOUS)的作用抑制雄蕊和心皮的发育(Shannon and Meeks- Wagner, 1993; Elliott et al., 1996).miR172在拟南芥花器官中具有优势表达, 且miR172过表达植株花器官发育异常, 与ap2突变体类似(Park et al., 2002), 主要原因在于miR172从翻译水平抑制了AP2的表达(Aukerman and Sakai, 2003; Chen, 2004).时空表达分析结果表明, AP2表达产物主要积累于花器官外周, 与miR172信号在第2或第3轮花器官处有瞬时重叠(Wollmann et al., 2010).尽管miR172和AP2共表达时间非常短暂, 但两者共同调控花分生细胞的时序进程, 在花器官发育中发挥决定性作用(Ji et al., 2011). ...
A miR172 target-deficient AP2-like gene correlates with the double flower phenotype in roses 1 2018
... miR172-AP2模块除了在拟南芥中参与花器官发育调控, 在烟草(Nicotiana tabacum)中也有类似作用.将拟南芥miR172在烟草中过表达同样会导致烟草花结构发育异常, 与ap2突变体类似(Mlotshwa et al., 2006), 表明miR172-AP2模块在花器官发育调控方面具保守性.最新研究表明, miR172-AP2模块在油菜(Brassica campestris)、大岩桐(Sinningia speciosa)和玫瑰(Rosa rugosa)等双子叶植物中也参与花器官发育的调控(François et al., 2018; Li et al., 2019b; Wang et al., 2019b).此外, 该调控模式也存在于单子叶植物中.例如, 水稻AP2家族成员OsSNB和OsIDS1在小穗向小花转变及花器官形态建成过程中发挥关键作用.OsSNB突变会导致花发育异常.例如, 小穗向小花转变显著延迟, 产生额外颖苞, 花中浆片异常膨大进而发育成内外稃等; 在OsSNB单突变基础上同时突变OsIDS1则会进一步增强相关表型.水稻中过表达miR172也会导致花器官发育异常, 形成类似Ossnb突变体的表型(Lee and An, 2012; Wang et al., 2015a), 暗示miR172可能通过调控AP2基因OsSNB和OsIDS1的表达来影响水稻花器官的发育.另一禾本科单子叶植物玉米, 其花为单性花, 含有雄穗和雌穗两种花序.玉米的ts (tasselseed)突变体由于雄穗中的雌蕊不退化从而形成两性花, 导致雄穗雌化并进而开花结实(Chuck et al., 2007; Acosta et al., 2009).利用具有雄穗雌化现象的玉米突变体ts4和ts6开展目标基因克隆, 结果显示这2种突变体分别是由于miR172及其靶基因IDS1突变所致, 表明miR172-AP2模块在玉米性别决定中同样具有关键作用(Chuck et al., 2007).而IDS1的异位表达会推迟小穗向花转变, 从而使单个小穗产生多个小花(大于2朵).IDS1同源基因SID1 (SISTER OF INDETERMINATE SPIKELET 1)突变会进一步增强ids1突变体的表型, 表明这2个基因在玉米花器官发育过程中的功能具有累加作用(Chuck et al., 2008).禾本科植物的浆片在开花过程中具有重要作用, 即在特定时期吸水膨胀, 从而撑开颖壳, 完成开花过程.已知大麦(Hordeum vulgare)的闭花受精特性由AP2家族的Cly1 (Cleistogamy 1)基因控制, 该基因表达也受到miR172调控; 当Cly1序列中与miR172结合的位点发生突变时, 会干扰其mRNA与miR172结合, 从而减弱miR172的翻译抑制作用, 进而产生非常小的浆片而无法开颖, 最终发生闭花受精(Nair et al., 2009; Anwar et al., 2018).可见, miR172-AP2模块在不同植物中的功能非常保守, 且对植物花器官发育调控极为重要, miR172通过抑制AP2家族成员的表达, 进而影响AP2基因与B类和C类基因间的互作, 最终调控花器官发育、雌雄分化和开花等过程. ...
Salt and drought stresses induce the aberrant expression of microRNA genes in tobacco 1 2011
... 植物不像动物可以躲避逆境, 因此其进化出特有的复杂且精细的分子调控机制来应对各种环境胁迫.许多研究表明, miRNA在植物响应各种生物和非生物胁迫过程中具有重要作用.miR172作为miRNA家族中非常保守的一员, 其自身表达受低温、盐碱和干旱等多种胁迫诱导, 同时它也可通过调控AP2类转录因子参与植物响应各种环境胁迫(Zhou et al., 2008; Frazier et al., 2011; Yang et al., 2013; Candar-Cakir et al., 2016).当然, miR172-AP2的具体调控模式还与植物种类及其响应的逆境因子密切相关.拟南芥中, ABA及渗透胁迫可诱导miR172与其靶基因SNZ共表达.miR172b过表达或snz突变体对ABA及渗透胁迫的敏感性增强, 证明miR172通过控制其靶基因SNZ来介导拟南芥对渗透胁迫的响应(Zou et al., 2013).干旱胁迫可诱导miR172e的表达, 同时过表达miR172e又可显著增强拟南芥对干旱的抗性.进一步研究表明, 干旱通过诱导GI (GIGANTEA)表达促进miR172e的积累以抑制下游靶基因TOE1的表达, 进而影响TOE1蛋白与WRKY44的互作, 最终实现对植物抗旱能力的调控(Han et al., 2013).在大豆中过表达miRNA172c或敲除GmNNC1均可增强大豆的耐盐性; 反之, 干扰miR172的表达或过表达GmNNC1均增强了大豆的盐敏感性(Sahito et al., 2017).此外, 在拟南芥中过表达大豆miR172c也可显著增强拟南芥对干旱及盐胁迫的耐受性(Zou et al., 2013).尽管目前人们对拟南芥和大豆中miR172介导的植物响应逆境胁迫机制有了较清晰的认识, 但在其它植物特别是水稻和小麦等重要作物中的相关研究仍停留在全基因组表达分析层面, 还未涉及具体分子机制的解析与调控网络的构建.例如, 香蕉(Musa nana)遇到冷胁迫后miR172的表达会显著上调, 初步推测miR172可能在冷应激反应中起中心协调作用(刘炜婳等, 2018).此外, 在番茄(Lycopersicon esculentum)中过表达miR172a和miR172b显著增强了其对致病疫霉的抗性, 潜在分子机理也与以miR172-AP2模块为核心的表达调控路径相关(Luan et al., 2018). ...
Coexpression analysis identifies Rice Starch Regulator1, a rice AP2/EREBP family transcription factor, as a novel rice starch biosynthesis regulator 1 2010
... miR172-AP2模块对果实发育的调控至关重要.拟南芥中, miR172-AP2模块可通过调节果荚发育来影响其果实发育.同时, 蛋白FUL与ARF (Auxin Response Factor)可直接互作并结合到miR172前体编码基因的启动子区来激活miR172表达, 进而抑制其下游靶基因AP2及TOE3的表达, 最终促进果实发育(José Ripoll et al., 2015).水稻AP2基因RSR1突变导致其籽粒变大、千粒重及稻米直链淀粉含量增加(Fu and Xue, 2010).进一步研究表明, RSR1正是受miR172调控的靶基因(Wang et al., 2015a), 但RSR1调控下游靶基因导致水稻籽粒变化的分子路径尚无报道.已知RSR1在水稻籽粒中不表达, 暗示miR172-RSR1模块对稻米粒形和品质的影响可能为间接作用, 即通过调控其它组织的生长发育来影响果实发育.综上, miR172-AP2模块对单子叶和双子叶植物果实发育均具有调控作用. ...
The putative miR172 target gene InAPETALA2-like is involved in the photoperiodic flower induction of Ipomoea nil 1 2009
... 水稻中, miR172通过抑制AP2基因OsIDS1和SNB的表达来解除它们对靶基因Ehd1 (Early hea- ding date1)的抑制, 从而诱导开花(Lee et al., 2014).最近, 大岩桐中也有类似报道, 即过表达miR172抑制SsAP2表达从而诱导早花(Li et al., 2019b).此外, 在大豆、牵牛花(Pharbitis nil)和麻风树(Jatropha curcas)中也有miR172-AP2模块参与开花时间调控的报道(Glazińska et al., 2009; Zhao et al., 2015; Tang et al., 2018).因此, 我们推测植物主要通过miR172-AP2分子模块调节开花时间来实现其营养生长向生殖生长的过渡. ...
The suppression of WRKY44 by GIGANTEA-miR172 pathway is involved in drought response of Arabidopsis thaliana 1 2013
... 植物不像动物可以躲避逆境, 因此其进化出特有的复杂且精细的分子调控机制来应对各种环境胁迫.许多研究表明, miRNA在植物响应各种生物和非生物胁迫过程中具有重要作用.miR172作为miRNA家族中非常保守的一员, 其自身表达受低温、盐碱和干旱等多种胁迫诱导, 同时它也可通过调控AP2类转录因子参与植物响应各种环境胁迫(Zhou et al., 2008; Frazier et al., 2011; Yang et al., 2013; Candar-Cakir et al., 2016).当然, miR172-AP2的具体调控模式还与植物种类及其响应的逆境因子密切相关.拟南芥中, ABA及渗透胁迫可诱导miR172与其靶基因SNZ共表达.miR172b过表达或snz突变体对ABA及渗透胁迫的敏感性增强, 证明miR172通过控制其靶基因SNZ来介导拟南芥对渗透胁迫的响应(Zou et al., 2013).干旱胁迫可诱导miR172e的表达, 同时过表达miR172e又可显著增强拟南芥对干旱的抗性.进一步研究表明, 干旱通过诱导GI (GIGANTEA)表达促进miR172e的积累以抑制下游靶基因TOE1的表达, 进而影响TOE1蛋白与WRKY44的互作, 最终实现对植物抗旱能力的调控(Han et al., 2013).在大豆中过表达miRNA172c或敲除GmNNC1均可增强大豆的耐盐性; 反之, 干扰miR172的表达或过表达GmNNC1均增强了大豆的盐敏感性(Sahito et al., 2017).此外, 在拟南芥中过表达大豆miR172c也可显著增强拟南芥对干旱及盐胁迫的耐受性(Zou et al., 2013).尽管目前人们对拟南芥和大豆中miR172介导的植物响应逆境胁迫机制有了较清晰的认识, 但在其它植物特别是水稻和小麦等重要作物中的相关研究仍停留在全基因组表达分析层面, 还未涉及具体分子机制的解析与调控网络的构建.例如, 香蕉(Musa nana)遇到冷胁迫后miR172的表达会显著上调, 初步推测miR172可能在冷应激反应中起中心协调作用(刘炜婳等, 2018).此外, 在番茄(Lycopersicon esculentum)中过表达miR172a和miR172b显著增强了其对致病疫霉的抗性, 潜在分子机理也与以miR172-AP2模块为核心的表达调控路径相关(Luan et al., 2018). ...
Functional analysis of citrus AP2 transcription factors identified CsAP2-09 involved in citrus canker disease response and tolerance 1 2019
... AP2/ERF类转录因子均含有1个由60-70个氨基酸残基组成的高度保守的AP2/ERF结构域(Riechmann and Meyerowitz, 1998).该结构域包含3个反向平行的β折叠和1个几乎与β折叠平行的α螺旋.β折叠在识别目标基因的顺式作用元件上具有重要作用, α螺旋可能介导与其它转录因子的互作(Nakano et al., 2006).根据保守结构域差异, 可将AP2/ERF转录因子进一步分为4个亚家族, 即AP2、ERF、DREB (dehydration-responsive element binding protein)和RAV (related to ABI3/VP1) (Xie et al., 2019).AP2亚家族含有2个AP2/ERF结构域, 主要参与调控植物花器官发育; 另外, 最新研究表明, 该亚家族还参与调控植物根系发育和孤雌生殖(Khanday et al., 2019; Li et al., 2019a).RAV亚家族含有1个AP2/ERF及1个B3结构域(Swaminathan et al., 2008), 该亚家族成员参与植物激素及胁迫响应(Hu et al., 2004; Sohn et al., 2006; Zhang et al., 2019; Zhao et al., 2019).ERF和DREB亚家族均仅包含1个AP2/ERF结构域, 两者的区别在于AP2/ERF结构域的第14和19位氨基酸不同, DREB亚家族这2个位置分别是缬氨酸(Val)和谷氨酸(Glu), 而ERF亚家族分别为丙氨酸(Ala)和天冬氨酸(Asp), 功能上两亚家族均参与植物响应生物和非生物胁迫(Dubouzet et al., 2003; Bäumler et al., 2019; He et al., 2019; Wang et al., 2019a). ...
MicroRNA 172 (miR172) signals epidermal infection and is expressed in cells primed for bacterial invasion in Lotus japonicus roots and nodules 1 2015
... 之前的研究表明, 根瘤菌处理3小时的大豆根中miR172表达显著上调(Subramanian et al., 2008), 且在成熟的根瘤中miR172c高度表达(Wang et al., 2009).而在大豆中过表达miR172c可促进根瘤菌的感染和根瘤的形态发生, 从而增加根瘤数量(Yan et al., 2013).此外, miR172还可调控结瘤因子(NF)的表达(Martin et al., 2009).例如, miR172的靶基因GmNNC1编码1个AP2家族转录抑制子, 可直接结合关键早期结瘤因子基因ENOD40的启动子并调节其表达, 从而调控结瘤数目(Wang et al., 2014).此外, 考虑到miR172自身表达受到1个AON (autoregulation of nodulation)结瘤自调控信号的抑制, 表明miR172-AP2模块可能是整合NF和AON信号通路的关键节点.后续研究表明, miR172在菜豆(Phaseolus vulgaris)和百脉根等豆科植物根瘤形成过程中同样发挥重要作用(Holt et al., 2015; Nova-Franco et al., 2015), 表明miR172-AP2模块在调控豆科植物结瘤及共生固氮方面的功能相对保守.进一步探讨miR172- AP2模块调控豆科植物结瘤固氮的机理, 将会促进豆科植物结瘤和固氮过程遗传机制的解析与调控网络的完善, 并有助于将其更好地应用于作物育种实践. ...
Variation in the interaction between alleles of HvAPETALA2 and microRNA172 determines the density of grains on the barley inflorescence 1 2013
... 利用全基因组关联分析结合遗传定位克隆到1个控制大麦穗粒密度的基因Zeo (ZEOCRITON), 即大麦AP2的直系同源基因HvAP2.大麦密穗表型的产生是由于HvAP2基因的miR172识别位点发生突变, 从而减弱miR172对其表达的调控, 造成穗轴延伸受阻, 最终影响小穗在花序上的分布所致(Houston et al., 2013).小麦的Q基因是1个典型的驯化基因, 控制小麦脱粒性、粒形、穗长和株高等多个驯化相关性状.基因克隆结果表明, Q基因属于AP2家族, 且其表达在mRNA水平受miR172的剪切调控(Simons et al., 2005; Liu et al., 2018).miR172-AP2模块在禾本科植物中参与穗型的调控, 表明其功能的保守性.深入研究该模块控制作物穗部形态的分子机理, 并结合目前已获解析的多个粒形调控基因信息, 可为今后通过分子设计育种优化穗部和粒形性状, 培育高产优质作物新品种奠定基础. ...
Arabidopsis RAV1 is down-regulated by brassinosteroid and may act as a negative regulator during plant development 1 2004
... AP2/ERF类转录因子均含有1个由60-70个氨基酸残基组成的高度保守的AP2/ERF结构域(Riechmann and Meyerowitz, 1998).该结构域包含3个反向平行的β折叠和1个几乎与β折叠平行的α螺旋.β折叠在识别目标基因的顺式作用元件上具有重要作用, α螺旋可能介导与其它转录因子的互作(Nakano et al., 2006).根据保守结构域差异, 可将AP2/ERF转录因子进一步分为4个亚家族, 即AP2、ERF、DREB (dehydration-responsive element binding protein)和RAV (related to ABI3/VP1) (Xie et al., 2019).AP2亚家族含有2个AP2/ERF结构域, 主要参与调控植物花器官发育; 另外, 最新研究表明, 该亚家族还参与调控植物根系发育和孤雌生殖(Khanday et al., 2019; Li et al., 2019a).RAV亚家族含有1个AP2/ERF及1个B3结构域(Swaminathan et al., 2008), 该亚家族成员参与植物激素及胁迫响应(Hu et al., 2004; Sohn et al., 2006; Zhang et al., 2019; Zhao et al., 2019).ERF和DREB亚家族均仅包含1个AP2/ERF结构域, 两者的区别在于AP2/ERF结构域的第14和19位氨基酸不同, DREB亚家族这2个位置分别是缬氨酸(Val)和谷氨酸(Glu), 而ERF亚家族分别为丙氨酸(Ala)和天冬氨酸(Asp), 功能上两亚家族均参与植物响应生物和非生物胁迫(Dubouzet et al., 2003; Bäumler et al., 2019; He et al., 2019; Wang et al., 2019a). ...
A microRNA in a multiple-turnover RNAi enzyme complex 1 2002
... 目前, 常用的miRNA靶基因鉴定方法包括生物信息学预测、AGO蛋白免疫共沉淀、双荧光素酶和降解组测序等(董淼等, 2013).miRNA主要通过剪切靶基因mRNA和抑制靶基因翻译2种途径调控靶基因, 但具体方式取决于miRNA与其靶基因mRNA的互补程度及作用位置(Hutvágner and Zamore, 2002; Llave et al., 2002; Schwab et al., 2005; Jung et al., 2007; Zhu et al., 2009).miRNA172与AP2家族基因的互补区域通常位于靠近3°UTR的CDS区, 主要调控方式为抑制AP2靶基因翻译, 但也存在部分与靶基因mRNA完全互补从而对其进行剪切调控的情况(Aukerman and Sakai, 2003; Schmid et al., 2003; Chen, 2004; Mathieu et al., 2009).当miRNA通过抑制翻译对靶基因进行调控时, 两者将表现出协同表达模式(Zhu et al., 2009).例如, 水稻中miRNA172及其靶基因OsIDS1和RSR1在包括胚和胚乳在内的大多数组织中表达模式一致; 同样, 过量表达miR172也会导致各发育阶段穗子中的OsIDS1 (OsINDETERMINATE SPIKELET1)和RSR1 (Rice Starch Regulator1)表达呈现出升高或降低的动态变化模式, 表明miR172对靶基因的调控具有时空特异性.在玉米(Zea mays)中, miR172通过剪切靶基因GL15的mRNA来调控玉米从营养生长向生殖生长的过渡(Lauter et al., 2005).此外, 在大豆(Glycine max)、马铃薯(Solanum tuberosum)和拟南芥中也有类似报道, 即miR172通过切割其靶基因的mRNA来调控植物的生长发育(Wang et al., 2014). ...
ARGONAUTE10 and ARGONAUTE1 regulate the termination of floral stem cells through two microRNAs in Arabidopsis 1 2011
... AP2在模式植物拟南芥中被发现时便认为与植物花器官特征调控相关(Kunst et al., 1989).拟南芥的花包括花萼、花瓣、雄蕊和心皮4轮器官, 其特征由A、B和C三类基因控制(Bowman et al., 2012).AP2属于A类基因, 不仅参与第1轮花萼特征的调控, 还与B类基因AP3和PI (PISTILLATA)共同控制第2轮花瓣的形成; 并可通过拮抗C类基因AG (AGAMOUS)的作用抑制雄蕊和心皮的发育(Shannon and Meeks- Wagner, 1993; Elliott et al., 1996).miR172在拟南芥花器官中具有优势表达, 且miR172过表达植株花器官发育异常, 与ap2突变体类似(Park et al., 2002), 主要原因在于miR172从翻译水平抑制了AP2的表达(Aukerman and Sakai, 2003; Chen, 2004).时空表达分析结果表明, AP2表达产物主要积累于花器官外周, 与miR172信号在第2或第3轮花器官处有瞬时重叠(Wollmann et al., 2010).尽管miR172和AP2共表达时间非常短暂, 但两者共同调控花分生细胞的时序进程, 在花器官发育中发挥决定性作用(Ji et al., 2011). ...
Control of Arabidopsis flower and seed development by the homeotic gene APETALA2 1 1994
... 转录因子的最重要特征是可通过与靶基因启动子区的特定顺式作用元件结合来激活或抑制靶基因转录, 从而参与植物的生长发育及其与环境的互作调控.AP2/ERF转录因子很早便被发现与拟南芥的花发育有关(Jofuku et al., 1994).随着研究的不断深入, 其在许多植物生长发育过程中的重要性开始显现. ...
MicroRNAs and their regulatory roles in plants 1 2006
... MicroRNA (miRNA)是广泛存在于真核生物中、由20-24个核苷酸组成的非编码RNA, 在转录后水平调控靶基因表达(Carrington and Ambros, 2003; Bartel, 2004).miRNA在调控植物种子休眠、粒形和株型等生长发育过程及逆境响应中发挥重要作用(王幼宁等, 2016; Saminathan et al., 2019; Miao et al., 2019, 2020).植物中miRNA生成过程与编码蛋白基因类似, 起初由RNA聚合酶II转录形成一段称作初级转录物的pri-miRNA的核苷酸序列, 长度几百到几千个碱基不等, 且在5′和3′端分别带有甲基鸟苷帽子和polyA尾巴(Rogers and Chen, 2013).pri-miRNA根据其自身序列的互补性可形成1至数个发夹茎环结构, 进而在细胞核内被Dicer-like DCL1酶、双链结合蛋白辅因子HYL1以及锌指蛋白SE切割成具有茎环结构的前体miRNA (pre-miRNA).pre-miRNA很不稳定, 很快又被DCL1、HYL1和SE进一步剪切成约22 nt的miRNA-miRNA*二聚体(Kurihara and Watanabe, 2004; Jones-Rhoades et al., 2006; 陈丽等, 2018), 该二聚体首先被HEN1 (HUAENHAN-CER1)甲基化来增强其稳定性, 随后在转运蛋白HST (HASTY)的作用下进入沉默复合体(RISC)状态, 且其中一条成熟单链miRNA保留在复合体中, 与互补的靶标mRNA特异性结合来调控其表达水平(Li et al., 2005; Park et al., 2005; Brodersen et al., 2008; 张俊红等, 2014). ...
MicroRNA regulation of fruit growth 1 2015
... miR172-AP2模块对果实发育的调控至关重要.拟南芥中, miR172-AP2模块可通过调节果荚发育来影响其果实发育.同时, 蛋白FUL与ARF (Auxin Response Factor)可直接互作并结合到miR172前体编码基因的启动子区来激活miR172表达, 进而抑制其下游靶基因AP2及TOE3的表达, 最终促进果实发育(José Ripoll et al., 2015).水稻AP2基因RSR1突变导致其籽粒变大、千粒重及稻米直链淀粉含量增加(Fu and Xue, 2010).进一步研究表明, RSR1正是受miR172调控的靶基因(Wang et al., 2015a), 但RSR1调控下游靶基因导致水稻籽粒变化的分子路径尚无报道.已知RSR1在水稻籽粒中不表达, 暗示miR172-RSR1模块对稻米粒形和品质的影响可能为间接作用, 即通过调控其它组织的生长发育来影响果实发育.综上, miR172-AP2模块对单子叶和双子叶植物果实发育均具有调控作用. ...
The GIGANTEA-regulated microRNA172 mediates photoperiodic flowering independent of CONSTANS in Arabidopsis 3 2007
... miRNA家族中, miR172是较早被发现同时也是被研究最透彻的成员之一(Park et al., 2002), 其靶基因主要编码AP2/ERF (APETALA2/ethylene responsive factor)类转录因子.AP2/ERF转录因子为植物所特有, 且几乎参与植物生长发育调控的各个环节.miRNA172通过特异或非特异性结合AP2/ERF的mRNA, 以mRNA剪切和翻译抑制2种方式调控AP2/ERF靶基因的表达, 从而调控植物众多生长发育过程(Jung et al., 2007; Zhu et al., 2009; 刘炜婳等, 2018).近期, 研究者在miRNA172-AP2模块调控植物花器官发育、时序转换、块茎及果实发育、根瘤形成和胁迫响应等方面取得一系列研究进展(王幼宁等, 2016; 赵晓晖等, 2017; Luan et al., 2018; Saminathan et al., 2019).本文以miRNA172-AP2模块为中心, 在简要介绍miR172和AP2的结构、功能及互作模式基础上, 重点论述miRNA172-AP2模块参与的植物生长发育及胁迫响应调控, 深入解析其生物学效应和分子机制, 并初步建立由miR172-AP2模块介导的植物生长发育分子调控网络. ...
... 虽然miR172家族成员成熟miRNA的序列高度保守, 但功能上并非简单重复.研究表明, miR172各成员启动子序列及其包含的顺式作用元件并不相同, 从而决定了它们表达的时空特异性(Zhu and Helliwell, 2011; 刘炜婳等, 2018).双子叶模式植物拟南芥(Arabidopsis thaliana)中存在5个miR172成员, 即miR172a-1、miR172a-2、miR172b-1、miR172b-2和miR172c (Chen, 2004).虽然拟南芥miR172各成员的表达模式不尽相同, 但根据其表达量和表达模式总体可分为2类: 一类包括miR172b-2和miR172c, 它们在拟南芥生长发育过程中表达量一直很低; 另一类包括miR172a-1、miR172a-2和miR172b-1, 它们的表达量随着拟南芥的生长发育逐渐升高, 到生殖生长阶段达到峰值(Jung et al., 2007).在表达的组织特异性方面, 除了在植物根中未检测到miR172的表达, 在其它包括花、叶片、茎和角果等组织中均检测到其表达(赵晓晖等, 2017).单子叶模式植物水稻(Oryza sativa)中存在4个miR172成员(miR172a-miR172d), 它们在苗期表达均较强, 但同样各成员表达也具有一定的组织特异性.例如, miR172c在水稻籽粒中不表达(Zhu et al., 2009).综上, miR172在植物中的表达具有明显的时空特异性(Zhu and Helliwell, 2011; 赵晓晖等, 2017). ...
... 目前, 常用的miRNA靶基因鉴定方法包括生物信息学预测、AGO蛋白免疫共沉淀、双荧光素酶和降解组测序等(董淼等, 2013).miRNA主要通过剪切靶基因mRNA和抑制靶基因翻译2种途径调控靶基因, 但具体方式取决于miRNA与其靶基因mRNA的互补程度及作用位置(Hutvágner and Zamore, 2002; Llave et al., 2002; Schwab et al., 2005; Jung et al., 2007; Zhu et al., 2009).miRNA172与AP2家族基因的互补区域通常位于靠近3°UTR的CDS区, 主要调控方式为抑制AP2靶基因翻译, 但也存在部分与靶基因mRNA完全互补从而对其进行剪切调控的情况(Aukerman and Sakai, 2003; Schmid et al., 2003; Chen, 2004; Mathieu et al., 2009).当miRNA通过抑制翻译对靶基因进行调控时, 两者将表现出协同表达模式(Zhu et al., 2009).例如, 水稻中miRNA172及其靶基因OsIDS1和RSR1在包括胚和胚乳在内的大多数组织中表达模式一致; 同样, 过量表达miR172也会导致各发育阶段穗子中的OsIDS1 (OsINDETERMINATE SPIKELET1)和RSR1 (Rice Starch Regulator1)表达呈现出升高或降低的动态变化模式, 表明miR172对靶基因的调控具有时空特异性.在玉米(Zea mays)中, miR172通过剪切靶基因GL15的mRNA来调控玉米从营养生长向生殖生长的过渡(Lauter et al., 2005).此外, 在大豆(Glycine max)、马铃薯(Solanum tuberosum)和拟南芥中也有类似报道, 即miR172通过切割其靶基因的mRNA来调控植物的生长发育(Wang et al., 2014). ...
A male-expressed rice embryogenic trigger redirected for asexual propagation through seeds 1 2019
... AP2/ERF类转录因子均含有1个由60-70个氨基酸残基组成的高度保守的AP2/ERF结构域(Riechmann and Meyerowitz, 1998).该结构域包含3个反向平行的β折叠和1个几乎与β折叠平行的α螺旋.β折叠在识别目标基因的顺式作用元件上具有重要作用, α螺旋可能介导与其它转录因子的互作(Nakano et al., 2006).根据保守结构域差异, 可将AP2/ERF转录因子进一步分为4个亚家族, 即AP2、ERF、DREB (dehydration-responsive element binding protein)和RAV (related to ABI3/VP1) (Xie et al., 2019).AP2亚家族含有2个AP2/ERF结构域, 主要参与调控植物花器官发育; 另外, 最新研究表明, 该亚家族还参与调控植物根系发育和孤雌生殖(Khanday et al., 2019; Li et al., 2019a).RAV亚家族含有1个AP2/ERF及1个B3结构域(Swaminathan et al., 2008), 该亚家族成员参与植物激素及胁迫响应(Hu et al., 2004; Sohn et al., 2006; Zhang et al., 2019; Zhao et al., 2019).ERF和DREB亚家族均仅包含1个AP2/ERF结构域, 两者的区别在于AP2/ERF结构域的第14和19位氨基酸不同, DREB亚家族这2个位置分别是缬氨酸(Val)和谷氨酸(Glu), 而ERF亚家族分别为丙氨酸(Ala)和天冬氨酸(Asp), 功能上两亚家族均参与植物响应生物和非生物胁迫(Dubouzet et al., 2003; Bäumler et al., 2019; He et al., 2019; Wang et al., 2019a). ...
AP2 gene determines the identity of perianth organs in flowers of Arabidopsis thaliana 1 1989
... AP2在模式植物拟南芥中被发现时便认为与植物花器官特征调控相关(Kunst et al., 1989).拟南芥的花包括花萼、花瓣、雄蕊和心皮4轮器官, 其特征由A、B和C三类基因控制(Bowman et al., 2012).AP2属于A类基因, 不仅参与第1轮花萼特征的调控, 还与B类基因AP3和PI (PISTILLATA)共同控制第2轮花瓣的形成; 并可通过拮抗C类基因AG (AGAMOUS)的作用抑制雄蕊和心皮的发育(Shannon and Meeks- Wagner, 1993; Elliott et al., 1996).miR172在拟南芥花器官中具有优势表达, 且miR172过表达植株花器官发育异常, 与ap2突变体类似(Park et al., 2002), 主要原因在于miR172从翻译水平抑制了AP2的表达(Aukerman and Sakai, 2003; Chen, 2004).时空表达分析结果表明, AP2表达产物主要积累于花器官外周, 与miR172信号在第2或第3轮花器官处有瞬时重叠(Wollmann et al., 2010).尽管miR172和AP2共表达时间非常短暂, 但两者共同调控花分生细胞的时序进程, 在花器官发育中发挥决定性作用(Ji et al., 2011). ...
Arabidopsis micro-RNA biogenesis through dicer-like 1 protein functions 1 2004
... MicroRNA (miRNA)是广泛存在于真核生物中、由20-24个核苷酸组成的非编码RNA, 在转录后水平调控靶基因表达(Carrington and Ambros, 2003; Bartel, 2004).miRNA在调控植物种子休眠、粒形和株型等生长发育过程及逆境响应中发挥重要作用(王幼宁等, 2016; Saminathan et al., 2019; Miao et al., 2019, 2020).植物中miRNA生成过程与编码蛋白基因类似, 起初由RNA聚合酶II转录形成一段称作初级转录物的pri-miRNA的核苷酸序列, 长度几百到几千个碱基不等, 且在5′和3′端分别带有甲基鸟苷帽子和polyA尾巴(Rogers and Chen, 2013).pri-miRNA根据其自身序列的互补性可形成1至数个发夹茎环结构, 进而在细胞核内被Dicer-like DCL1酶、双链结合蛋白辅因子HYL1以及锌指蛋白SE切割成具有茎环结构的前体miRNA (pre-miRNA).pre-miRNA很不稳定, 很快又被DCL1、HYL1和SE进一步剪切成约22 nt的miRNA-miRNA*二聚体(Kurihara and Watanabe, 2004; Jones-Rhoades et al., 2006; 陈丽等, 2018), 该二聚体首先被HEN1 (HUAENHAN-CER1)甲基化来增强其稳定性, 随后在转运蛋白HST (HASTY)的作用下进入沉默复合体(RISC)状态, 且其中一条成熟单链miRNA保留在复合体中, 与互补的靶标mRNA特异性结合来调控其表达水平(Li et al., 2005; Park et al., 2005; Brodersen et al., 2008; 张俊红等, 2014). ...
MicroRNA172 down-regulates glossy15 to promote vegetative phase change in maize 2 2005
... 目前, 常用的miRNA靶基因鉴定方法包括生物信息学预测、AGO蛋白免疫共沉淀、双荧光素酶和降解组测序等(董淼等, 2013).miRNA主要通过剪切靶基因mRNA和抑制靶基因翻译2种途径调控靶基因, 但具体方式取决于miRNA与其靶基因mRNA的互补程度及作用位置(Hutvágner and Zamore, 2002; Llave et al., 2002; Schwab et al., 2005; Jung et al., 2007; Zhu et al., 2009).miRNA172与AP2家族基因的互补区域通常位于靠近3°UTR的CDS区, 主要调控方式为抑制AP2靶基因翻译, 但也存在部分与靶基因mRNA完全互补从而对其进行剪切调控的情况(Aukerman and Sakai, 2003; Schmid et al., 2003; Chen, 2004; Mathieu et al., 2009).当miRNA通过抑制翻译对靶基因进行调控时, 两者将表现出协同表达模式(Zhu et al., 2009).例如, 水稻中miRNA172及其靶基因OsIDS1和RSR1在包括胚和胚乳在内的大多数组织中表达模式一致; 同样, 过量表达miR172也会导致各发育阶段穗子中的OsIDS1 (OsINDETERMINATE SPIKELET1)和RSR1 (Rice Starch Regulator1)表达呈现出升高或降低的动态变化模式, 表明miR172对靶基因的调控具有时空特异性.在玉米(Zea mays)中, miR172通过剪切靶基因GL15的mRNA来调控玉米从营养生长向生殖生长的过渡(Lauter et al., 2005).此外, 在大豆(Glycine max)、马铃薯(Solanum tuberosum)和拟南芥中也有类似报道, 即miR172通过切割其靶基因的mRNA来调控植物的生长发育(Wang et al., 2014). ...
... 此外, miR172-AP2模块在植物营养生长期不同阶段转变过程中也发挥重要作用(Wu et al., 2009).例如, GL15 (Glossy 15 )是1个控制玉米幼苗向成熟期转变时间的AP2基因, 其表达同样受到miR172调控(Moose and Sisco, 1994, 1996; Lauter et al., 2005).可见, miR172-AP2模块在植物时序转换调控中也发挥关键作用.AP2家族成员众多, 在功能上兼具共性和特异性, 解析其分子机制可为生产实践中利用特定的miR172-AP2互作模块开展分子设计育种提供可能, 从而实现真正精准育种. ...
Two AP2 family genes, SUPERNUMERARY BRACT (SNB) and OsINDETERMINATE SPIKELET 1 (OsIDS1), synergistically control inflorescence architecture and floral meristem establishment in rice 1 2012
... miR172-AP2模块除了在拟南芥中参与花器官发育调控, 在烟草(Nicotiana tabacum)中也有类似作用.将拟南芥miR172在烟草中过表达同样会导致烟草花结构发育异常, 与ap2突变体类似(Mlotshwa et al., 2006), 表明miR172-AP2模块在花器官发育调控方面具保守性.最新研究表明, miR172-AP2模块在油菜(Brassica campestris)、大岩桐(Sinningia speciosa)和玫瑰(Rosa rugosa)等双子叶植物中也参与花器官发育的调控(François et al., 2018; Li et al., 2019b; Wang et al., 2019b).此外, 该调控模式也存在于单子叶植物中.例如, 水稻AP2家族成员OsSNB和OsIDS1在小穗向小花转变及花器官形态建成过程中发挥关键作用.OsSNB突变会导致花发育异常.例如, 小穗向小花转变显著延迟, 产生额外颖苞, 花中浆片异常膨大进而发育成内外稃等; 在OsSNB单突变基础上同时突变OsIDS1则会进一步增强相关表型.水稻中过表达miR172也会导致花器官发育异常, 形成类似Ossnb突变体的表型(Lee and An, 2012; Wang et al., 2015a), 暗示miR172可能通过调控AP2基因OsSNB和OsIDS1的表达来影响水稻花器官的发育.另一禾本科单子叶植物玉米, 其花为单性花, 含有雄穗和雌穗两种花序.玉米的ts (tasselseed)突变体由于雄穗中的雌蕊不退化从而形成两性花, 导致雄穗雌化并进而开花结实(Chuck et al., 2007; Acosta et al., 2009).利用具有雄穗雌化现象的玉米突变体ts4和ts6开展目标基因克隆, 结果显示这2种突变体分别是由于miR172及其靶基因IDS1突变所致, 表明miR172-AP2模块在玉米性别决定中同样具有关键作用(Chuck et al., 2007).而IDS1的异位表达会推迟小穗向花转变, 从而使单个小穗产生多个小花(大于2朵).IDS1同源基因SID1 (SISTER OF INDETERMINATE SPIKELET 1)突变会进一步增强ids1突变体的表型, 表明这2个基因在玉米花器官发育过程中的功能具有累加作用(Chuck et al., 2008).禾本科植物的浆片在开花过程中具有重要作用, 即在特定时期吸水膨胀, 从而撑开颖壳, 完成开花过程.已知大麦(Hordeum vulgare)的闭花受精特性由AP2家族的Cly1 (Cleistogamy 1)基因控制, 该基因表达也受到miR172调控; 当Cly1序列中与miR172结合的位点发生突变时, 会干扰其mRNA与miR172结合, 从而减弱miR172的翻译抑制作用, 进而产生非常小的浆片而无法开颖, 最终发生闭花受精(Nair et al., 2009; Anwar et al., 2018).可见, miR172-AP2模块在不同植物中的功能非常保守, 且对植物花器官发育调控极为重要, miR172通过抑制AP2家族成员的表达, 进而影响AP2基因与B类和C类基因间的互作, 最终调控花器官发育、雌雄分化和开花等过程. ...
Rice miR172 induces flowering by suppressing OsIDS1 and SNB, two AP2 genes that negatively regulate expression of Ehd1 and florigens 1 2014
... 水稻中, miR172通过抑制AP2基因OsIDS1和SNB的表达来解除它们对靶基因Ehd1 (Early hea- ding date1)的抑制, 从而诱导开花(Lee et al., 2014).最近, 大岩桐中也有类似报道, 即过表达miR172抑制SsAP2表达从而诱导早花(Li et al., 2019b).此外, 在大豆、牵牛花(Pharbitis nil)和麻风树(Jatropha curcas)中也有miR172-AP2模块参与开花时间调控的报道(Glazińska et al., 2009; Zhao et al., 2015; Tang et al., 2018).因此, 我们推测植物主要通过miR172-AP2分子模块调节开花时间来实现其营养生长向生殖生长的过渡. ...
Ectopic expression of the Aechmea fasciata APETALA2 gene AfAP2-2 reduces seed size and delays flowering in Arabidopsis 1 2019
... 各AP2/ERF亚家族转录因子所识别的顺式作用元件具有一定的特异性.例如, AP2亚家族可与启动子中gCAC(A/G)N(A/T)TcCC(a/g)ANG(c/t)顺式作用元件结合来调节靶基因的表达, 进而调控植物的花器官发育、开花时间及时序转换等(Boutilier et al., 2002; Lei et al., 2019; Wang et al., 2019b).RVA亚家族由于含有AP2/ERF和B3结构域, 因此可识别CAACA和CACCTG基序.ERF亚家族主要识别启动子区域的GCC-box, DREB亚家族则可与干旱胁迫响应和冷诱导响应元件(A/GCCGAC)结合, 从而调节植物的非生物胁迫响应(Dubouzet et al., 2003; Agarwal et al., 2010; Mizoi et al., 2012; Bäumler et al., 2019; Wang et al., 2019a). ...
Misexpression of miR482, miR1512, and miR1515 increases soybean nodulation 1 2010
... 根瘤菌不仅能感染豆科植物根系形成根瘤, 还可与其共生固氮.在苜蓿(Medicago sativa)中首先证实miR166和miR169参与根瘤发育的调控(Combier et al., 2006; Boualem et al., 2008).百脉根(Lotus corniculatus)中的研究也表明, miR171、miR397和miR156与根瘤的发育、病毒感染和固氮能力密切相关(De Luis et al., 2012; Wang et al., 2015b).在大豆中过表达miR482、miR1512、miR1515、miR156及miR167均可以显著改变大豆根瘤的数量(Li et al., 2010; Wang et al., 2015c).因此, miRNA在豆科植物结瘤和共生固氮过程中发挥重要作用. ...
Methylation protects miRNAs and siRNAs from a 3°-end uridylation activity in Arabidopsis 1 2005
... MicroRNA (miRNA)是广泛存在于真核生物中、由20-24个核苷酸组成的非编码RNA, 在转录后水平调控靶基因表达(Carrington and Ambros, 2003; Bartel, 2004).miRNA在调控植物种子休眠、粒形和株型等生长发育过程及逆境响应中发挥重要作用(王幼宁等, 2016; Saminathan et al., 2019; Miao et al., 2019, 2020).植物中miRNA生成过程与编码蛋白基因类似, 起初由RNA聚合酶II转录形成一段称作初级转录物的pri-miRNA的核苷酸序列, 长度几百到几千个碱基不等, 且在5′和3′端分别带有甲基鸟苷帽子和polyA尾巴(Rogers and Chen, 2013).pri-miRNA根据其自身序列的互补性可形成1至数个发夹茎环结构, 进而在细胞核内被Dicer-like DCL1酶、双链结合蛋白辅因子HYL1以及锌指蛋白SE切割成具有茎环结构的前体miRNA (pre-miRNA).pre-miRNA很不稳定, 很快又被DCL1、HYL1和SE进一步剪切成约22 nt的miRNA-miRNA*二聚体(Kurihara and Watanabe, 2004; Jones-Rhoades et al., 2006; 陈丽等, 2018), 该二聚体首先被HEN1 (HUAENHAN-CER1)甲基化来增强其稳定性, 随后在转运蛋白HST (HASTY)的作用下进入沉默复合体(RISC)状态, 且其中一条成熟单链miRNA保留在复合体中, 与互补的靶标mRNA特异性结合来调控其表达水平(Li et al., 2005; Park et al., 2005; Brodersen et al., 2008; 张俊红等, 2014). ...
ZmRAP2.7, an AP2 transcription factor, is involved in maize brace roots development 1 2019
... AP2/ERF类转录因子均含有1个由60-70个氨基酸残基组成的高度保守的AP2/ERF结构域(Riechmann and Meyerowitz, 1998).该结构域包含3个反向平行的β折叠和1个几乎与β折叠平行的α螺旋.β折叠在识别目标基因的顺式作用元件上具有重要作用, α螺旋可能介导与其它转录因子的互作(Nakano et al., 2006).根据保守结构域差异, 可将AP2/ERF转录因子进一步分为4个亚家族, 即AP2、ERF、DREB (dehydration-responsive element binding protein)和RAV (related to ABI3/VP1) (Xie et al., 2019).AP2亚家族含有2个AP2/ERF结构域, 主要参与调控植物花器官发育; 另外, 最新研究表明, 该亚家族还参与调控植物根系发育和孤雌生殖(Khanday et al., 2019; Li et al., 2019a).RAV亚家族含有1个AP2/ERF及1个B3结构域(Swaminathan et al., 2008), 该亚家族成员参与植物激素及胁迫响应(Hu et al., 2004; Sohn et al., 2006; Zhang et al., 2019; Zhao et al., 2019).ERF和DREB亚家族均仅包含1个AP2/ERF结构域, 两者的区别在于AP2/ERF结构域的第14和19位氨基酸不同, DREB亚家族这2个位置分别是缬氨酸(Val)和谷氨酸(Glu), 而ERF亚家族分别为丙氨酸(Ala)和天冬氨酸(Asp), 功能上两亚家族均参与植物响应生物和非生物胁迫(Dubouzet et al., 2003; Bäumler et al., 2019; He et al., 2019; Wang et al., 2019a). ...
Regulation of flowering time via miR172-mediated APETALA2-like expression in ornamental gloxinia (Sinningia speciosa) 2 2019
... miR172-AP2模块除了在拟南芥中参与花器官发育调控, 在烟草(Nicotiana tabacum)中也有类似作用.将拟南芥miR172在烟草中过表达同样会导致烟草花结构发育异常, 与ap2突变体类似(Mlotshwa et al., 2006), 表明miR172-AP2模块在花器官发育调控方面具保守性.最新研究表明, miR172-AP2模块在油菜(Brassica campestris)、大岩桐(Sinningia speciosa)和玫瑰(Rosa rugosa)等双子叶植物中也参与花器官发育的调控(François et al., 2018; Li et al., 2019b; Wang et al., 2019b).此外, 该调控模式也存在于单子叶植物中.例如, 水稻AP2家族成员OsSNB和OsIDS1在小穗向小花转变及花器官形态建成过程中发挥关键作用.OsSNB突变会导致花发育异常.例如, 小穗向小花转变显著延迟, 产生额外颖苞, 花中浆片异常膨大进而发育成内外稃等; 在OsSNB单突变基础上同时突变OsIDS1则会进一步增强相关表型.水稻中过表达miR172也会导致花器官发育异常, 形成类似Ossnb突变体的表型(Lee and An, 2012; Wang et al., 2015a), 暗示miR172可能通过调控AP2基因OsSNB和OsIDS1的表达来影响水稻花器官的发育.另一禾本科单子叶植物玉米, 其花为单性花, 含有雄穗和雌穗两种花序.玉米的ts (tasselseed)突变体由于雄穗中的雌蕊不退化从而形成两性花, 导致雄穗雌化并进而开花结实(Chuck et al., 2007; Acosta et al., 2009).利用具有雄穗雌化现象的玉米突变体ts4和ts6开展目标基因克隆, 结果显示这2种突变体分别是由于miR172及其靶基因IDS1突变所致, 表明miR172-AP2模块在玉米性别决定中同样具有关键作用(Chuck et al., 2007).而IDS1的异位表达会推迟小穗向花转变, 从而使单个小穗产生多个小花(大于2朵).IDS1同源基因SID1 (SISTER OF INDETERMINATE SPIKELET 1)突变会进一步增强ids1突变体的表型, 表明这2个基因在玉米花器官发育过程中的功能具有累加作用(Chuck et al., 2008).禾本科植物的浆片在开花过程中具有重要作用, 即在特定时期吸水膨胀, 从而撑开颖壳, 完成开花过程.已知大麦(Hordeum vulgare)的闭花受精特性由AP2家族的Cly1 (Cleistogamy 1)基因控制, 该基因表达也受到miR172调控; 当Cly1序列中与miR172结合的位点发生突变时, 会干扰其mRNA与miR172结合, 从而减弱miR172的翻译抑制作用, 进而产生非常小的浆片而无法开颖, 最终发生闭花受精(Nair et al., 2009; Anwar et al., 2018).可见, miR172-AP2模块在不同植物中的功能非常保守, 且对植物花器官发育调控极为重要, miR172通过抑制AP2家族成员的表达, 进而影响AP2基因与B类和C类基因间的互作, 最终调控花器官发育、雌雄分化和开花等过程. ...
... 水稻中, miR172通过抑制AP2基因OsIDS1和SNB的表达来解除它们对靶基因Ehd1 (Early hea- ding date1)的抑制, 从而诱导开花(Lee et al., 2014).最近, 大岩桐中也有类似报道, 即过表达miR172抑制SsAP2表达从而诱导早花(Li et al., 2019b).此外, 在大豆、牵牛花(Pharbitis nil)和麻风树(Jatropha curcas)中也有miR172-AP2模块参与开花时间调控的报道(Glazińska et al., 2009; Zhao et al., 2015; Tang et al., 2018).因此, 我们推测植物主要通过miR172-AP2分子模块调节开花时间来实现其营养生长向生殖生长的过渡. ...
Functional regulation of Q by microRNA172 and transcriptional co-repressor TOPLESS in controlling bread wheat spikelet density 1 2018
... 利用全基因组关联分析结合遗传定位克隆到1个控制大麦穗粒密度的基因Zeo (ZEOCRITON), 即大麦AP2的直系同源基因HvAP2.大麦密穗表型的产生是由于HvAP2基因的miR172识别位点发生突变, 从而减弱miR172对其表达的调控, 造成穗轴延伸受阻, 最终影响小穗在花序上的分布所致(Houston et al., 2013).小麦的Q基因是1个典型的驯化基因, 控制小麦脱粒性、粒形、穗长和株高等多个驯化相关性状.基因克隆结果表明, Q基因属于AP2家族, 且其表达在mRNA水平受miR172的剪切调控(Simons et al., 2005; Liu et al., 2018).miR172-AP2模块在禾本科植物中参与穗型的调控, 表明其功能的保守性.深入研究该模块控制作物穗部形态的分子机理, 并结合目前已获解析的多个粒形调控基因信息, 可为今后通过分子设计育种优化穗部和粒形性状, 培育高产优质作物新品种奠定基础. ...
Cleavage of Scarecrow-like mRNA targets directed by a class of Arabidopsis miRNA 1 2002
... 目前, 常用的miRNA靶基因鉴定方法包括生物信息学预测、AGO蛋白免疫共沉淀、双荧光素酶和降解组测序等(董淼等, 2013).miRNA主要通过剪切靶基因mRNA和抑制靶基因翻译2种途径调控靶基因, 但具体方式取决于miRNA与其靶基因mRNA的互补程度及作用位置(Hutvágner and Zamore, 2002; Llave et al., 2002; Schwab et al., 2005; Jung et al., 2007; Zhu et al., 2009).miRNA172与AP2家族基因的互补区域通常位于靠近3°UTR的CDS区, 主要调控方式为抑制AP2靶基因翻译, 但也存在部分与靶基因mRNA完全互补从而对其进行剪切调控的情况(Aukerman and Sakai, 2003; Schmid et al., 2003; Chen, 2004; Mathieu et al., 2009).当miRNA通过抑制翻译对靶基因进行调控时, 两者将表现出协同表达模式(Zhu et al., 2009).例如, 水稻中miRNA172及其靶基因OsIDS1和RSR1在包括胚和胚乳在内的大多数组织中表达模式一致; 同样, 过量表达miR172也会导致各发育阶段穗子中的OsIDS1 (OsINDETERMINATE SPIKELET1)和RSR1 (Rice Starch Regulator1)表达呈现出升高或降低的动态变化模式, 表明miR172对靶基因的调控具有时空特异性.在玉米(Zea mays)中, miR172通过剪切靶基因GL15的mRNA来调控玉米从营养生长向生殖生长的过渡(Lauter et al., 2005).此外, 在大豆(Glycine max)、马铃薯(Solanum tuberosum)和拟南芥中也有类似报道, 即miR172通过切割其靶基因的mRNA来调控植物的生长发育(Wang et al., 2014). ...
Effective enhancement of resistance to Phytophthora infestans by overexpression of miR172a and b in Solanum lycopersicum 3 2018
... miRNA家族中, miR172是较早被发现同时也是被研究最透彻的成员之一(Park et al., 2002), 其靶基因主要编码AP2/ERF (APETALA2/ethylene responsive factor)类转录因子.AP2/ERF转录因子为植物所特有, 且几乎参与植物生长发育调控的各个环节.miRNA172通过特异或非特异性结合AP2/ERF的mRNA, 以mRNA剪切和翻译抑制2种方式调控AP2/ERF靶基因的表达, 从而调控植物众多生长发育过程(Jung et al., 2007; Zhu et al., 2009; 刘炜婳等, 2018).近期, 研究者在miRNA172-AP2模块调控植物花器官发育、时序转换、块茎及果实发育、根瘤形成和胁迫响应等方面取得一系列研究进展(王幼宁等, 2016; 赵晓晖等, 2017; Luan et al., 2018; Saminathan et al., 2019).本文以miRNA172-AP2模块为中心, 在简要介绍miR172和AP2的结构、功能及互作模式基础上, 重点论述miRNA172-AP2模块参与的植物生长发育及胁迫响应调控, 深入解析其生物学效应和分子机制, 并初步建立由miR172-AP2模块介导的植物生长发育分子调控网络. ...
... miR172在植物的生殖转换、花器官形成及胁迫响应等方面具有重要调控作用(Wu et al., 2009; 王幼宁等, 2016; 赵晓晖等, 2017; Luan et al., 2018; Saminathan et al., 2019).miR172在高等植物中高度保守, 虽然不同物种中miR172成员的数目和长度可能不同, 但在亲缘关系较近的物种间序列高度保守, 暗示其在不同物种中可能具有相似的调控方式和生物学功能. ...
... 植物不像动物可以躲避逆境, 因此其进化出特有的复杂且精细的分子调控机制来应对各种环境胁迫.许多研究表明, miRNA在植物响应各种生物和非生物胁迫过程中具有重要作用.miR172作为miRNA家族中非常保守的一员, 其自身表达受低温、盐碱和干旱等多种胁迫诱导, 同时它也可通过调控AP2类转录因子参与植物响应各种环境胁迫(Zhou et al., 2008; Frazier et al., 2011; Yang et al., 2013; Candar-Cakir et al., 2016).当然, miR172-AP2的具体调控模式还与植物种类及其响应的逆境因子密切相关.拟南芥中, ABA及渗透胁迫可诱导miR172与其靶基因SNZ共表达.miR172b过表达或snz突变体对ABA及渗透胁迫的敏感性增强, 证明miR172通过控制其靶基因SNZ来介导拟南芥对渗透胁迫的响应(Zou et al., 2013).干旱胁迫可诱导miR172e的表达, 同时过表达miR172e又可显著增强拟南芥对干旱的抗性.进一步研究表明, 干旱通过诱导GI (GIGANTEA)表达促进miR172e的积累以抑制下游靶基因TOE1的表达, 进而影响TOE1蛋白与WRKY44的互作, 最终实现对植物抗旱能力的调控(Han et al., 2013).在大豆中过表达miRNA172c或敲除GmNNC1均可增强大豆的耐盐性; 反之, 干扰miR172的表达或过表达GmNNC1均增强了大豆的盐敏感性(Sahito et al., 2017).此外, 在拟南芥中过表达大豆miR172c也可显著增强拟南芥对干旱及盐胁迫的耐受性(Zou et al., 2013).尽管目前人们对拟南芥和大豆中miR172介导的植物响应逆境胁迫机制有了较清晰的认识, 但在其它植物特别是水稻和小麦等重要作物中的相关研究仍停留在全基因组表达分析层面, 还未涉及具体分子机制的解析与调控网络的构建.例如, 香蕉(Musa nana)遇到冷胁迫后miR172的表达会显著上调, 初步推测miR172可能在冷应激反应中起中心协调作用(刘炜婳等, 2018).此外, 在番茄(Lycopersicon esculentum)中过表达miR172a和miR172b显著增强了其对致病疫霉的抗性, 潜在分子机理也与以miR172-AP2模块为核心的表达调控路径相关(Luan et al., 2018). ...
Graft-transmissible induction of potato tuberization by the microRNA miR172 1 2009
... 之前的研究表明, 根瘤菌处理3小时的大豆根中miR172表达显著上调(Subramanian et al., 2008), 且在成熟的根瘤中miR172c高度表达(Wang et al., 2009).而在大豆中过表达miR172c可促进根瘤菌的感染和根瘤的形态发生, 从而增加根瘤数量(Yan et al., 2013).此外, miR172还可调控结瘤因子(NF)的表达(Martin et al., 2009).例如, miR172的靶基因GmNNC1编码1个AP2家族转录抑制子, 可直接结合关键早期结瘤因子基因ENOD40的启动子并调节其表达, 从而调控结瘤数目(Wang et al., 2014).此外, 考虑到miR172自身表达受到1个AON (autoregulation of nodulation)结瘤自调控信号的抑制, 表明miR172-AP2模块可能是整合NF和AON信号通路的关键节点.后续研究表明, miR172在菜豆(Phaseolus vulgaris)和百脉根等豆科植物根瘤形成过程中同样发挥重要作用(Holt et al., 2015; Nova-Franco et al., 2015), 表明miR172-AP2模块在调控豆科植物结瘤及共生固氮方面的功能相对保守.进一步探讨miR172- AP2模块调控豆科植物结瘤固氮的机理, 将会促进豆科植物结瘤和固氮过程遗传机制的解析与调控网络的完善, 并有助于将其更好地应用于作物育种实践. ...
Repression of flowering by the miR172 target SMZ 1 2009
... 目前, 常用的miRNA靶基因鉴定方法包括生物信息学预测、AGO蛋白免疫共沉淀、双荧光素酶和降解组测序等(董淼等, 2013).miRNA主要通过剪切靶基因mRNA和抑制靶基因翻译2种途径调控靶基因, 但具体方式取决于miRNA与其靶基因mRNA的互补程度及作用位置(Hutvágner and Zamore, 2002; Llave et al., 2002; Schwab et al., 2005; Jung et al., 2007; Zhu et al., 2009).miRNA172与AP2家族基因的互补区域通常位于靠近3°UTR的CDS区, 主要调控方式为抑制AP2靶基因翻译, 但也存在部分与靶基因mRNA完全互补从而对其进行剪切调控的情况(Aukerman and Sakai, 2003; Schmid et al., 2003; Chen, 2004; Mathieu et al., 2009).当miRNA通过抑制翻译对靶基因进行调控时, 两者将表现出协同表达模式(Zhu et al., 2009).例如, 水稻中miRNA172及其靶基因OsIDS1和RSR1在包括胚和胚乳在内的大多数组织中表达模式一致; 同样, 过量表达miR172也会导致各发育阶段穗子中的OsIDS1 (OsINDETERMINATE SPIKELET1)和RSR1 (Rice Starch Regulator1)表达呈现出升高或降低的动态变化模式, 表明miR172对靶基因的调控具有时空特异性.在玉米(Zea mays)中, miR172通过剪切靶基因GL15的mRNA来调控玉米从营养生长向生殖生长的过渡(Lauter et al., 2005).此外, 在大豆(Glycine max)、马铃薯(Solanum tuberosum)和拟南芥中也有类似报道, 即miR172通过切割其靶基因的mRNA来调控植物的生长发育(Wang et al., 2014). ...
Mutations in MIR396e and MIR396f increase grain size and modulate shoot architecture in rice 1 2020
... MicroRNA (miRNA)是广泛存在于真核生物中、由20-24个核苷酸组成的非编码RNA, 在转录后水平调控靶基因表达(Carrington and Ambros, 2003; Bartel, 2004).miRNA在调控植物种子休眠、粒形和株型等生长发育过程及逆境响应中发挥重要作用(王幼宁等, 2016; Saminathan et al., 2019; Miao et al., 2019, 2020).植物中miRNA生成过程与编码蛋白基因类似, 起初由RNA聚合酶II转录形成一段称作初级转录物的pri-miRNA的核苷酸序列, 长度几百到几千个碱基不等, 且在5′和3′端分别带有甲基鸟苷帽子和polyA尾巴(Rogers and Chen, 2013).pri-miRNA根据其自身序列的互补性可形成1至数个发夹茎环结构, 进而在细胞核内被Dicer-like DCL1酶、双链结合蛋白辅因子HYL1以及锌指蛋白SE切割成具有茎环结构的前体miRNA (pre-miRNA).pre-miRNA很不稳定, 很快又被DCL1、HYL1和SE进一步剪切成约22 nt的miRNA-miRNA*二聚体(Kurihara and Watanabe, 2004; Jones-Rhoades et al., 2006; 陈丽等, 2018), 该二聚体首先被HEN1 (HUAENHAN-CER1)甲基化来增强其稳定性, 随后在转运蛋白HST (HASTY)的作用下进入沉默复合体(RISC)状态, 且其中一条成熟单链miRNA保留在复合体中, 与互补的靶标mRNA特异性结合来调控其表达水平(Li et al., 2005; Park et al., 2005; Brodersen et al., 2008; 张俊红等, 2014). ...
The grain yield modulator miR156 regulates seed dormancy through the gibberellin pathway in rice 1 2019
... MicroRNA (miRNA)是广泛存在于真核生物中、由20-24个核苷酸组成的非编码RNA, 在转录后水平调控靶基因表达(Carrington and Ambros, 2003; Bartel, 2004).miRNA在调控植物种子休眠、粒形和株型等生长发育过程及逆境响应中发挥重要作用(王幼宁等, 2016; Saminathan et al., 2019; Miao et al., 2019, 2020).植物中miRNA生成过程与编码蛋白基因类似, 起初由RNA聚合酶II转录形成一段称作初级转录物的pri-miRNA的核苷酸序列, 长度几百到几千个碱基不等, 且在5′和3′端分别带有甲基鸟苷帽子和polyA尾巴(Rogers and Chen, 2013).pri-miRNA根据其自身序列的互补性可形成1至数个发夹茎环结构, 进而在细胞核内被Dicer-like DCL1酶、双链结合蛋白辅因子HYL1以及锌指蛋白SE切割成具有茎环结构的前体miRNA (pre-miRNA).pre-miRNA很不稳定, 很快又被DCL1、HYL1和SE进一步剪切成约22 nt的miRNA-miRNA*二聚体(Kurihara and Watanabe, 2004; Jones-Rhoades et al., 2006; 陈丽等, 2018), 该二聚体首先被HEN1 (HUAENHAN-CER1)甲基化来增强其稳定性, 随后在转运蛋白HST (HASTY)的作用下进入沉默复合体(RISC)状态, 且其中一条成熟单链miRNA保留在复合体中, 与互补的靶标mRNA特异性结合来调控其表达水平(Li et al., 2005; Park et al., 2005; Brodersen et al., 2008; 张俊红等, 2014). ...
AP2/ERF family transcription factors in plant abiotic stress responses 1 2012
... 各AP2/ERF亚家族转录因子所识别的顺式作用元件具有一定的特异性.例如, AP2亚家族可与启动子中gCAC(A/G)N(A/T)TcCC(a/g)ANG(c/t)顺式作用元件结合来调节靶基因的表达, 进而调控植物的花器官发育、开花时间及时序转换等(Boutilier et al., 2002; Lei et al., 2019; Wang et al., 2019b).RVA亚家族由于含有AP2/ERF和B3结构域, 因此可识别CAACA和CACCTG基序.ERF亚家族主要识别启动子区域的GCC-box, DREB亚家族则可与干旱胁迫响应和冷诱导响应元件(A/GCCGAC)结合, 从而调节植物的非生物胁迫响应(Dubouzet et al., 2003; Agarwal et al., 2010; Mizoi et al., 2012; Bäumler et al., 2019; Wang et al., 2019a). ...
Floral patterning defects induced by Arabidopsis APETALA2 and microRNA172 expression in Nicotiana benthamiana 1 2006
... miR172-AP2模块除了在拟南芥中参与花器官发育调控, 在烟草(Nicotiana tabacum)中也有类似作用.将拟南芥miR172在烟草中过表达同样会导致烟草花结构发育异常, 与ap2突变体类似(Mlotshwa et al., 2006), 表明miR172-AP2模块在花器官发育调控方面具保守性.最新研究表明, miR172-AP2模块在油菜(Brassica campestris)、大岩桐(Sinningia speciosa)和玫瑰(Rosa rugosa)等双子叶植物中也参与花器官发育的调控(François et al., 2018; Li et al., 2019b; Wang et al., 2019b).此外, 该调控模式也存在于单子叶植物中.例如, 水稻AP2家族成员OsSNB和OsIDS1在小穗向小花转变及花器官形态建成过程中发挥关键作用.OsSNB突变会导致花发育异常.例如, 小穗向小花转变显著延迟, 产生额外颖苞, 花中浆片异常膨大进而发育成内外稃等; 在OsSNB单突变基础上同时突变OsIDS1则会进一步增强相关表型.水稻中过表达miR172也会导致花器官发育异常, 形成类似Ossnb突变体的表型(Lee and An, 2012; Wang et al., 2015a), 暗示miR172可能通过调控AP2基因OsSNB和OsIDS1的表达来影响水稻花器官的发育.另一禾本科单子叶植物玉米, 其花为单性花, 含有雄穗和雌穗两种花序.玉米的ts (tasselseed)突变体由于雄穗中的雌蕊不退化从而形成两性花, 导致雄穗雌化并进而开花结实(Chuck et al., 2007; Acosta et al., 2009).利用具有雄穗雌化现象的玉米突变体ts4和ts6开展目标基因克隆, 结果显示这2种突变体分别是由于miR172及其靶基因IDS1突变所致, 表明miR172-AP2模块在玉米性别决定中同样具有关键作用(Chuck et al., 2007).而IDS1的异位表达会推迟小穗向花转变, 从而使单个小穗产生多个小花(大于2朵).IDS1同源基因SID1 (SISTER OF INDETERMINATE SPIKELET 1)突变会进一步增强ids1突变体的表型, 表明这2个基因在玉米花器官发育过程中的功能具有累加作用(Chuck et al., 2008).禾本科植物的浆片在开花过程中具有重要作用, 即在特定时期吸水膨胀, 从而撑开颖壳, 完成开花过程.已知大麦(Hordeum vulgare)的闭花受精特性由AP2家族的Cly1 (Cleistogamy 1)基因控制, 该基因表达也受到miR172调控; 当Cly1序列中与miR172结合的位点发生突变时, 会干扰其mRNA与miR172结合, 从而减弱miR172的翻译抑制作用, 进而产生非常小的浆片而无法开颖, 最终发生闭花受精(Nair et al., 2009; Anwar et al., 2018).可见, miR172-AP2模块在不同植物中的功能非常保守, 且对植物花器官发育调控极为重要, miR172通过抑制AP2家族成员的表达, 进而影响AP2基因与B类和C类基因间的互作, 最终调控花器官发育、雌雄分化和开花等过程. ...
Glossy15 controls the epidermal juvenile-to-adult phase transition in maize 1 1994
... 此外, miR172-AP2模块在植物营养生长期不同阶段转变过程中也发挥重要作用(Wu et al., 2009).例如, GL15 (Glossy 15 )是1个控制玉米幼苗向成熟期转变时间的AP2基因, 其表达同样受到miR172调控(Moose and Sisco, 1994, 1996; Lauter et al., 2005).可见, miR172-AP2模块在植物时序转换调控中也发挥关键作用.AP2家族成员众多, 在功能上兼具共性和特异性, 解析其分子机制可为生产实践中利用特定的miR172-AP2互作模块开展分子设计育种提供可能, 从而实现真正精准育种. ...
Glossy15, an APETALA2-like gene from maize that regulates leaf epidermal cell identity 1 1996
... 此外, miR172-AP2模块在植物营养生长期不同阶段转变过程中也发挥重要作用(Wu et al., 2009).例如, GL15 (Glossy 15 )是1个控制玉米幼苗向成熟期转变时间的AP2基因, 其表达同样受到miR172调控(Moose and Sisco, 1994, 1996; Lauter et al., 2005).可见, miR172-AP2模块在植物时序转换调控中也发挥关键作用.AP2家族成员众多, 在功能上兼具共性和特异性, 解析其分子机制可为生产实践中利用特定的miR172-AP2互作模块开展分子设计育种提供可能, 从而实现真正精准育种. ...
Cleistogamous flowering in barley arises from the suppression of microRNA-guided HvAP2 mRNA cleavage 1 2009
... miR172-AP2模块除了在拟南芥中参与花器官发育调控, 在烟草(Nicotiana tabacum)中也有类似作用.将拟南芥miR172在烟草中过表达同样会导致烟草花结构发育异常, 与ap2突变体类似(Mlotshwa et al., 2006), 表明miR172-AP2模块在花器官发育调控方面具保守性.最新研究表明, miR172-AP2模块在油菜(Brassica campestris)、大岩桐(Sinningia speciosa)和玫瑰(Rosa rugosa)等双子叶植物中也参与花器官发育的调控(François et al., 2018; Li et al., 2019b; Wang et al., 2019b).此外, 该调控模式也存在于单子叶植物中.例如, 水稻AP2家族成员OsSNB和OsIDS1在小穗向小花转变及花器官形态建成过程中发挥关键作用.OsSNB突变会导致花发育异常.例如, 小穗向小花转变显著延迟, 产生额外颖苞, 花中浆片异常膨大进而发育成内外稃等; 在OsSNB单突变基础上同时突变OsIDS1则会进一步增强相关表型.水稻中过表达miR172也会导致花器官发育异常, 形成类似Ossnb突变体的表型(Lee and An, 2012; Wang et al., 2015a), 暗示miR172可能通过调控AP2基因OsSNB和OsIDS1的表达来影响水稻花器官的发育.另一禾本科单子叶植物玉米, 其花为单性花, 含有雄穗和雌穗两种花序.玉米的ts (tasselseed)突变体由于雄穗中的雌蕊不退化从而形成两性花, 导致雄穗雌化并进而开花结实(Chuck et al., 2007; Acosta et al., 2009).利用具有雄穗雌化现象的玉米突变体ts4和ts6开展目标基因克隆, 结果显示这2种突变体分别是由于miR172及其靶基因IDS1突变所致, 表明miR172-AP2模块在玉米性别决定中同样具有关键作用(Chuck et al., 2007).而IDS1的异位表达会推迟小穗向花转变, 从而使单个小穗产生多个小花(大于2朵).IDS1同源基因SID1 (SISTER OF INDETERMINATE SPIKELET 1)突变会进一步增强ids1突变体的表型, 表明这2个基因在玉米花器官发育过程中的功能具有累加作用(Chuck et al., 2008).禾本科植物的浆片在开花过程中具有重要作用, 即在特定时期吸水膨胀, 从而撑开颖壳, 完成开花过程.已知大麦(Hordeum vulgare)的闭花受精特性由AP2家族的Cly1 (Cleistogamy 1)基因控制, 该基因表达也受到miR172调控; 当Cly1序列中与miR172结合的位点发生突变时, 会干扰其mRNA与miR172结合, 从而减弱miR172的翻译抑制作用, 进而产生非常小的浆片而无法开颖, 最终发生闭花受精(Nair et al., 2009; Anwar et al., 2018).可见, miR172-AP2模块在不同植物中的功能非常保守, 且对植物花器官发育调控极为重要, miR172通过抑制AP2家族成员的表达, 进而影响AP2基因与B类和C类基因间的互作, 最终调控花器官发育、雌雄分化和开花等过程. ...
Genome-wide analysis of the ERF gene family in Arabidopsis and rice 1 2006
... AP2/ERF类转录因子均含有1个由60-70个氨基酸残基组成的高度保守的AP2/ERF结构域(Riechmann and Meyerowitz, 1998).该结构域包含3个反向平行的β折叠和1个几乎与β折叠平行的α螺旋.β折叠在识别目标基因的顺式作用元件上具有重要作用, α螺旋可能介导与其它转录因子的互作(Nakano et al., 2006).根据保守结构域差异, 可将AP2/ERF转录因子进一步分为4个亚家族, 即AP2、ERF、DREB (dehydration-responsive element binding protein)和RAV (related to ABI3/VP1) (Xie et al., 2019).AP2亚家族含有2个AP2/ERF结构域, 主要参与调控植物花器官发育; 另外, 最新研究表明, 该亚家族还参与调控植物根系发育和孤雌生殖(Khanday et al., 2019; Li et al., 2019a).RAV亚家族含有1个AP2/ERF及1个B3结构域(Swaminathan et al., 2008), 该亚家族成员参与植物激素及胁迫响应(Hu et al., 2004; Sohn et al., 2006; Zhang et al., 2019; Zhao et al., 2019).ERF和DREB亚家族均仅包含1个AP2/ERF结构域, 两者的区别在于AP2/ERF结构域的第14和19位氨基酸不同, DREB亚家族这2个位置分别是缬氨酸(Val)和谷氨酸(Glu), 而ERF亚家族分别为丙氨酸(Ala)和天冬氨酸(Asp), 功能上两亚家族均参与植物响应生物和非生物胁迫(Dubouzet et al., 2003; Bäumler et al., 2019; He et al., 2019; Wang et al., 2019a). ...
The microRNA172c-APETALA2-1 node as a key regulator of the common bean- Rhizobium etli nitrogen fixation symbiosis 1 2015
... 之前的研究表明, 根瘤菌处理3小时的大豆根中miR172表达显著上调(Subramanian et al., 2008), 且在成熟的根瘤中miR172c高度表达(Wang et al., 2009).而在大豆中过表达miR172c可促进根瘤菌的感染和根瘤的形态发生, 从而增加根瘤数量(Yan et al., 2013).此外, miR172还可调控结瘤因子(NF)的表达(Martin et al., 2009).例如, miR172的靶基因GmNNC1编码1个AP2家族转录抑制子, 可直接结合关键早期结瘤因子基因ENOD40的启动子并调节其表达, 从而调控结瘤数目(Wang et al., 2014).此外, 考虑到miR172自身表达受到1个AON (autoregulation of nodulation)结瘤自调控信号的抑制, 表明miR172-AP2模块可能是整合NF和AON信号通路的关键节点.后续研究表明, miR172在菜豆(Phaseolus vulgaris)和百脉根等豆科植物根瘤形成过程中同样发挥重要作用(Holt et al., 2015; Nova-Franco et al., 2015), 表明miR172-AP2模块在调控豆科植物结瘤及共生固氮方面的功能相对保守.进一步探讨miR172- AP2模块调控豆科植物结瘤固氮的机理, 将会促进豆科植物结瘤和固氮过程遗传机制的解析与调控网络的完善, 并有助于将其更好地应用于作物育种实践. ...
Nuclear processing and export of microRNAs in Arabidopsis 1 2005
... MicroRNA (miRNA)是广泛存在于真核生物中、由20-24个核苷酸组成的非编码RNA, 在转录后水平调控靶基因表达(Carrington and Ambros, 2003; Bartel, 2004).miRNA在调控植物种子休眠、粒形和株型等生长发育过程及逆境响应中发挥重要作用(王幼宁等, 2016; Saminathan et al., 2019; Miao et al., 2019, 2020).植物中miRNA生成过程与编码蛋白基因类似, 起初由RNA聚合酶II转录形成一段称作初级转录物的pri-miRNA的核苷酸序列, 长度几百到几千个碱基不等, 且在5′和3′端分别带有甲基鸟苷帽子和polyA尾巴(Rogers and Chen, 2013).pri-miRNA根据其自身序列的互补性可形成1至数个发夹茎环结构, 进而在细胞核内被Dicer-like DCL1酶、双链结合蛋白辅因子HYL1以及锌指蛋白SE切割成具有茎环结构的前体miRNA (pre-miRNA).pre-miRNA很不稳定, 很快又被DCL1、HYL1和SE进一步剪切成约22 nt的miRNA-miRNA*二聚体(Kurihara and Watanabe, 2004; Jones-Rhoades et al., 2006; 陈丽等, 2018), 该二聚体首先被HEN1 (HUAENHAN-CER1)甲基化来增强其稳定性, 随后在转运蛋白HST (HASTY)的作用下进入沉默复合体(RISC)状态, 且其中一条成熟单链miRNA保留在复合体中, 与互补的靶标mRNA特异性结合来调控其表达水平(Li et al., 2005; Park et al., 2005; Brodersen et al., 2008; 张俊红等, 2014). ...
CARPEL FACTORY, a dicer homolog, and HEN1, a novel protein, act in microRNA metabolism in Arabidopsis thaliana 2 2002
... miRNA家族中, miR172是较早被发现同时也是被研究最透彻的成员之一(Park et al., 2002), 其靶基因主要编码AP2/ERF (APETALA2/ethylene responsive factor)类转录因子.AP2/ERF转录因子为植物所特有, 且几乎参与植物生长发育调控的各个环节.miRNA172通过特异或非特异性结合AP2/ERF的mRNA, 以mRNA剪切和翻译抑制2种方式调控AP2/ERF靶基因的表达, 从而调控植物众多生长发育过程(Jung et al., 2007; Zhu et al., 2009; 刘炜婳等, 2018).近期, 研究者在miRNA172-AP2模块调控植物花器官发育、时序转换、块茎及果实发育、根瘤形成和胁迫响应等方面取得一系列研究进展(王幼宁等, 2016; 赵晓晖等, 2017; Luan et al., 2018; Saminathan et al., 2019).本文以miRNA172-AP2模块为中心, 在简要介绍miR172和AP2的结构、功能及互作模式基础上, 重点论述miRNA172-AP2模块参与的植物生长发育及胁迫响应调控, 深入解析其生物学效应和分子机制, 并初步建立由miR172-AP2模块介导的植物生长发育分子调控网络. ...
... AP2在模式植物拟南芥中被发现时便认为与植物花器官特征调控相关(Kunst et al., 1989).拟南芥的花包括花萼、花瓣、雄蕊和心皮4轮器官, 其特征由A、B和C三类基因控制(Bowman et al., 2012).AP2属于A类基因, 不仅参与第1轮花萼特征的调控, 还与B类基因AP3和PI (PISTILLATA)共同控制第2轮花瓣的形成; 并可通过拮抗C类基因AG (AGAMOUS)的作用抑制雄蕊和心皮的发育(Shannon and Meeks- Wagner, 1993; Elliott et al., 1996).miR172在拟南芥花器官中具有优势表达, 且miR172过表达植株花器官发育异常, 与ap2突变体类似(Park et al., 2002), 主要原因在于miR172从翻译水平抑制了AP2的表达(Aukerman and Sakai, 2003; Chen, 2004).时空表达分析结果表明, AP2表达产物主要积累于花器官外周, 与miR172信号在第2或第3轮花器官处有瞬时重叠(Wollmann et al., 2010).尽管miR172和AP2共表达时间非常短暂, 但两者共同调控花分生细胞的时序进程, 在花器官发育中发挥决定性作用(Ji et al., 2011). ...
The AP2/EREBP family of plant transcription factors 1 1998
... AP2/ERF类转录因子均含有1个由60-70个氨基酸残基组成的高度保守的AP2/ERF结构域(Riechmann and Meyerowitz, 1998).该结构域包含3个反向平行的β折叠和1个几乎与β折叠平行的α螺旋.β折叠在识别目标基因的顺式作用元件上具有重要作用, α螺旋可能介导与其它转录因子的互作(Nakano et al., 2006).根据保守结构域差异, 可将AP2/ERF转录因子进一步分为4个亚家族, 即AP2、ERF、DREB (dehydration-responsive element binding protein)和RAV (related to ABI3/VP1) (Xie et al., 2019).AP2亚家族含有2个AP2/ERF结构域, 主要参与调控植物花器官发育; 另外, 最新研究表明, 该亚家族还参与调控植物根系发育和孤雌生殖(Khanday et al., 2019; Li et al., 2019a).RAV亚家族含有1个AP2/ERF及1个B3结构域(Swaminathan et al., 2008), 该亚家族成员参与植物激素及胁迫响应(Hu et al., 2004; Sohn et al., 2006; Zhang et al., 2019; Zhao et al., 2019).ERF和DREB亚家族均仅包含1个AP2/ERF结构域, 两者的区别在于AP2/ERF结构域的第14和19位氨基酸不同, DREB亚家族这2个位置分别是缬氨酸(Val)和谷氨酸(Glu), 而ERF亚家族分别为丙氨酸(Ala)和天冬氨酸(Asp), 功能上两亚家族均参与植物响应生物和非生物胁迫(Dubouzet et al., 2003; Bäumler et al., 2019; He et al., 2019; Wang et al., 2019a). ...
Biogenesis, turnover, and mode of action of plant microRNAs 1 2013
... MicroRNA (miRNA)是广泛存在于真核生物中、由20-24个核苷酸组成的非编码RNA, 在转录后水平调控靶基因表达(Carrington and Ambros, 2003; Bartel, 2004).miRNA在调控植物种子休眠、粒形和株型等生长发育过程及逆境响应中发挥重要作用(王幼宁等, 2016; Saminathan et al., 2019; Miao et al., 2019, 2020).植物中miRNA生成过程与编码蛋白基因类似, 起初由RNA聚合酶II转录形成一段称作初级转录物的pri-miRNA的核苷酸序列, 长度几百到几千个碱基不等, 且在5′和3′端分别带有甲基鸟苷帽子和polyA尾巴(Rogers and Chen, 2013).pri-miRNA根据其自身序列的互补性可形成1至数个发夹茎环结构, 进而在细胞核内被Dicer-like DCL1酶、双链结合蛋白辅因子HYL1以及锌指蛋白SE切割成具有茎环结构的前体miRNA (pre-miRNA).pre-miRNA很不稳定, 很快又被DCL1、HYL1和SE进一步剪切成约22 nt的miRNA-miRNA*二聚体(Kurihara and Watanabe, 2004; Jones-Rhoades et al., 2006; 陈丽等, 2018), 该二聚体首先被HEN1 (HUAENHAN-CER1)甲基化来增强其稳定性, 随后在转运蛋白HST (HASTY)的作用下进入沉默复合体(RISC)状态, 且其中一条成熟单链miRNA保留在复合体中, 与互补的靶标mRNA特异性结合来调控其表达水平(Li et al., 2005; Park et al., 2005; Brodersen et al., 2008; 张俊红等, 2014). ...
The miR172c-NNC1 module modulates root plastic development in response to salt in soybean 1 2017
... 植物不像动物可以躲避逆境, 因此其进化出特有的复杂且精细的分子调控机制来应对各种环境胁迫.许多研究表明, miRNA在植物响应各种生物和非生物胁迫过程中具有重要作用.miR172作为miRNA家族中非常保守的一员, 其自身表达受低温、盐碱和干旱等多种胁迫诱导, 同时它也可通过调控AP2类转录因子参与植物响应各种环境胁迫(Zhou et al., 2008; Frazier et al., 2011; Yang et al., 2013; Candar-Cakir et al., 2016).当然, miR172-AP2的具体调控模式还与植物种类及其响应的逆境因子密切相关.拟南芥中, ABA及渗透胁迫可诱导miR172与其靶基因SNZ共表达.miR172b过表达或snz突变体对ABA及渗透胁迫的敏感性增强, 证明miR172通过控制其靶基因SNZ来介导拟南芥对渗透胁迫的响应(Zou et al., 2013).干旱胁迫可诱导miR172e的表达, 同时过表达miR172e又可显著增强拟南芥对干旱的抗性.进一步研究表明, 干旱通过诱导GI (GIGANTEA)表达促进miR172e的积累以抑制下游靶基因TOE1的表达, 进而影响TOE1蛋白与WRKY44的互作, 最终实现对植物抗旱能力的调控(Han et al., 2013).在大豆中过表达miRNA172c或敲除GmNNC1均可增强大豆的耐盐性; 反之, 干扰miR172的表达或过表达GmNNC1均增强了大豆的盐敏感性(Sahito et al., 2017).此外, 在拟南芥中过表达大豆miR172c也可显著增强拟南芥对干旱及盐胁迫的耐受性(Zou et al., 2013).尽管目前人们对拟南芥和大豆中miR172介导的植物响应逆境胁迫机制有了较清晰的认识, 但在其它植物特别是水稻和小麦等重要作物中的相关研究仍停留在全基因组表达分析层面, 还未涉及具体分子机制的解析与调控网络的构建.例如, 香蕉(Musa nana)遇到冷胁迫后miR172的表达会显著上调, 初步推测miR172可能在冷应激反应中起中心协调作用(刘炜婳等, 2018).此外, 在番茄(Lycopersicon esculentum)中过表达miR172a和miR172b显著增强了其对致病疫霉的抗性, 潜在分子机理也与以miR172-AP2模块为核心的表达调控路径相关(Luan et al., 2018). ...
Elevated carbon dioxide and drought modulate physiology and storage-root development in sweet potato by regulating microRNAs 3 2019
... MicroRNA (miRNA)是广泛存在于真核生物中、由20-24个核苷酸组成的非编码RNA, 在转录后水平调控靶基因表达(Carrington and Ambros, 2003; Bartel, 2004).miRNA在调控植物种子休眠、粒形和株型等生长发育过程及逆境响应中发挥重要作用(王幼宁等, 2016; Saminathan et al., 2019; Miao et al., 2019, 2020).植物中miRNA生成过程与编码蛋白基因类似, 起初由RNA聚合酶II转录形成一段称作初级转录物的pri-miRNA的核苷酸序列, 长度几百到几千个碱基不等, 且在5′和3′端分别带有甲基鸟苷帽子和polyA尾巴(Rogers and Chen, 2013).pri-miRNA根据其自身序列的互补性可形成1至数个发夹茎环结构, 进而在细胞核内被Dicer-like DCL1酶、双链结合蛋白辅因子HYL1以及锌指蛋白SE切割成具有茎环结构的前体miRNA (pre-miRNA).pre-miRNA很不稳定, 很快又被DCL1、HYL1和SE进一步剪切成约22 nt的miRNA-miRNA*二聚体(Kurihara and Watanabe, 2004; Jones-Rhoades et al., 2006; 陈丽等, 2018), 该二聚体首先被HEN1 (HUAENHAN-CER1)甲基化来增强其稳定性, 随后在转运蛋白HST (HASTY)的作用下进入沉默复合体(RISC)状态, 且其中一条成熟单链miRNA保留在复合体中, 与互补的靶标mRNA特异性结合来调控其表达水平(Li et al., 2005; Park et al., 2005; Brodersen et al., 2008; 张俊红等, 2014). ...
... miRNA家族中, miR172是较早被发现同时也是被研究最透彻的成员之一(Park et al., 2002), 其靶基因主要编码AP2/ERF (APETALA2/ethylene responsive factor)类转录因子.AP2/ERF转录因子为植物所特有, 且几乎参与植物生长发育调控的各个环节.miRNA172通过特异或非特异性结合AP2/ERF的mRNA, 以mRNA剪切和翻译抑制2种方式调控AP2/ERF靶基因的表达, 从而调控植物众多生长发育过程(Jung et al., 2007; Zhu et al., 2009; 刘炜婳等, 2018).近期, 研究者在miRNA172-AP2模块调控植物花器官发育、时序转换、块茎及果实发育、根瘤形成和胁迫响应等方面取得一系列研究进展(王幼宁等, 2016; 赵晓晖等, 2017; Luan et al., 2018; Saminathan et al., 2019).本文以miRNA172-AP2模块为中心, 在简要介绍miR172和AP2的结构、功能及互作模式基础上, 重点论述miRNA172-AP2模块参与的植物生长发育及胁迫响应调控, 深入解析其生物学效应和分子机制, 并初步建立由miR172-AP2模块介导的植物生长发育分子调控网络. ...
... miR172在植物的生殖转换、花器官形成及胁迫响应等方面具有重要调控作用(Wu et al., 2009; 王幼宁等, 2016; 赵晓晖等, 2017; Luan et al., 2018; Saminathan et al., 2019).miR172在高等植物中高度保守, 虽然不同物种中miR172成员的数目和长度可能不同, 但在亲缘关系较近的物种间序列高度保守, 暗示其在不同物种中可能具有相似的调控方式和生物学功能. ...
Dissection of floral induction pathways using global expression analysis 1 2003
... 目前, 常用的miRNA靶基因鉴定方法包括生物信息学预测、AGO蛋白免疫共沉淀、双荧光素酶和降解组测序等(董淼等, 2013).miRNA主要通过剪切靶基因mRNA和抑制靶基因翻译2种途径调控靶基因, 但具体方式取决于miRNA与其靶基因mRNA的互补程度及作用位置(Hutvágner and Zamore, 2002; Llave et al., 2002; Schwab et al., 2005; Jung et al., 2007; Zhu et al., 2009).miRNA172与AP2家族基因的互补区域通常位于靠近3°UTR的CDS区, 主要调控方式为抑制AP2靶基因翻译, 但也存在部分与靶基因mRNA完全互补从而对其进行剪切调控的情况(Aukerman and Sakai, 2003; Schmid et al., 2003; Chen, 2004; Mathieu et al., 2009).当miRNA通过抑制翻译对靶基因进行调控时, 两者将表现出协同表达模式(Zhu et al., 2009).例如, 水稻中miRNA172及其靶基因OsIDS1和RSR1在包括胚和胚乳在内的大多数组织中表达模式一致; 同样, 过量表达miR172也会导致各发育阶段穗子中的OsIDS1 (OsINDETERMINATE SPIKELET1)和RSR1 (Rice Starch Regulator1)表达呈现出升高或降低的动态变化模式, 表明miR172对靶基因的调控具有时空特异性.在玉米(Zea mays)中, miR172通过剪切靶基因GL15的mRNA来调控玉米从营养生长向生殖生长的过渡(Lauter et al., 2005).此外, 在大豆(Glycine max)、马铃薯(Solanum tuberosum)和拟南芥中也有类似报道, 即miR172通过切割其靶基因的mRNA来调控植物的生长发育(Wang et al., 2014). ...
Specific effects of microRNAs on the plant transcriptome 1 2005
... 目前, 常用的miRNA靶基因鉴定方法包括生物信息学预测、AGO蛋白免疫共沉淀、双荧光素酶和降解组测序等(董淼等, 2013).miRNA主要通过剪切靶基因mRNA和抑制靶基因翻译2种途径调控靶基因, 但具体方式取决于miRNA与其靶基因mRNA的互补程度及作用位置(Hutvágner and Zamore, 2002; Llave et al., 2002; Schwab et al., 2005; Jung et al., 2007; Zhu et al., 2009).miRNA172与AP2家族基因的互补区域通常位于靠近3°UTR的CDS区, 主要调控方式为抑制AP2靶基因翻译, 但也存在部分与靶基因mRNA完全互补从而对其进行剪切调控的情况(Aukerman and Sakai, 2003; Schmid et al., 2003; Chen, 2004; Mathieu et al., 2009).当miRNA通过抑制翻译对靶基因进行调控时, 两者将表现出协同表达模式(Zhu et al., 2009).例如, 水稻中miRNA172及其靶基因OsIDS1和RSR1在包括胚和胚乳在内的大多数组织中表达模式一致; 同样, 过量表达miR172也会导致各发育阶段穗子中的OsIDS1 (OsINDETERMINATE SPIKELET1)和RSR1 (Rice Starch Regulator1)表达呈现出升高或降低的动态变化模式, 表明miR172对靶基因的调控具有时空特异性.在玉米(Zea mays)中, miR172通过剪切靶基因GL15的mRNA来调控玉米从营养生长向生殖生长的过渡(Lauter et al., 2005).此外, 在大豆(Glycine max)、马铃薯(Solanum tuberosum)和拟南芥中也有类似报道, 即miR172通过切割其靶基因的mRNA来调控植物的生长发育(Wang et al., 2014). ...
Genetic interactions that regulate inflorescence development in Arabidopsis 1 1993
... AP2在模式植物拟南芥中被发现时便认为与植物花器官特征调控相关(Kunst et al., 1989).拟南芥的花包括花萼、花瓣、雄蕊和心皮4轮器官, 其特征由A、B和C三类基因控制(Bowman et al., 2012).AP2属于A类基因, 不仅参与第1轮花萼特征的调控, 还与B类基因AP3和PI (PISTILLATA)共同控制第2轮花瓣的形成; 并可通过拮抗C类基因AG (AGAMOUS)的作用抑制雄蕊和心皮的发育(Shannon and Meeks- Wagner, 1993; Elliott et al., 1996).miR172在拟南芥花器官中具有优势表达, 且miR172过表达植株花器官发育异常, 与ap2突变体类似(Park et al., 2002), 主要原因在于miR172从翻译水平抑制了AP2的表达(Aukerman and Sakai, 2003; Chen, 2004).时空表达分析结果表明, AP2表达产物主要积累于花器官外周, 与miR172信号在第2或第3轮花器官处有瞬时重叠(Wollmann et al., 2010).尽管miR172和AP2共表达时间非常短暂, 但两者共同调控花分生细胞的时序进程, 在花器官发育中发挥决定性作用(Ji et al., 2011). ...
Molecular characterization of the major wheat domestication gene Q 1 2005
... 利用全基因组关联分析结合遗传定位克隆到1个控制大麦穗粒密度的基因Zeo (ZEOCRITON), 即大麦AP2的直系同源基因HvAP2.大麦密穗表型的产生是由于HvAP2基因的miR172识别位点发生突变, 从而减弱miR172对其表达的调控, 造成穗轴延伸受阻, 最终影响小穗在花序上的分布所致(Houston et al., 2013).小麦的Q基因是1个典型的驯化基因, 控制小麦脱粒性、粒形、穗长和株高等多个驯化相关性状.基因克隆结果表明, Q基因属于AP2家族, 且其表达在mRNA水平受miR172的剪切调控(Simons et al., 2005; Liu et al., 2018).miR172-AP2模块在禾本科植物中参与穗型的调控, 表明其功能的保守性.深入研究该模块控制作物穗部形态的分子机理, 并结合目前已获解析的多个粒形调控基因信息, 可为今后通过分子设计育种优化穗部和粒形性状, 培育高产优质作物新品种奠定基础. ...
Expression and functional roles of the pepper pathogen-induced transcription factor RAV1 in bacterial disease resistance, and drought and salt stress tolerance 1 2006
... AP2/ERF类转录因子均含有1个由60-70个氨基酸残基组成的高度保守的AP2/ERF结构域(Riechmann and Meyerowitz, 1998).该结构域包含3个反向平行的β折叠和1个几乎与β折叠平行的α螺旋.β折叠在识别目标基因的顺式作用元件上具有重要作用, α螺旋可能介导与其它转录因子的互作(Nakano et al., 2006).根据保守结构域差异, 可将AP2/ERF转录因子进一步分为4个亚家族, 即AP2、ERF、DREB (dehydration-responsive element binding protein)和RAV (related to ABI3/VP1) (Xie et al., 2019).AP2亚家族含有2个AP2/ERF结构域, 主要参与调控植物花器官发育; 另外, 最新研究表明, 该亚家族还参与调控植物根系发育和孤雌生殖(Khanday et al., 2019; Li et al., 2019a).RAV亚家族含有1个AP2/ERF及1个B3结构域(Swaminathan et al., 2008), 该亚家族成员参与植物激素及胁迫响应(Hu et al., 2004; Sohn et al., 2006; Zhang et al., 2019; Zhao et al., 2019).ERF和DREB亚家族均仅包含1个AP2/ERF结构域, 两者的区别在于AP2/ERF结构域的第14和19位氨基酸不同, DREB亚家族这2个位置分别是缬氨酸(Val)和谷氨酸(Glu), 而ERF亚家族分别为丙氨酸(Ala)和天冬氨酸(Asp), 功能上两亚家族均参与植物响应生物和非生物胁迫(Dubouzet et al., 2003; Bäumler et al., 2019; He et al., 2019; Wang et al., 2019a). ...
Novel and nodulation-regulated microRNAs in soybean roots 1 2008
... 之前的研究表明, 根瘤菌处理3小时的大豆根中miR172表达显著上调(Subramanian et al., 2008), 且在成熟的根瘤中miR172c高度表达(Wang et al., 2009).而在大豆中过表达miR172c可促进根瘤菌的感染和根瘤的形态发生, 从而增加根瘤数量(Yan et al., 2013).此外, miR172还可调控结瘤因子(NF)的表达(Martin et al., 2009).例如, miR172的靶基因GmNNC1编码1个AP2家族转录抑制子, 可直接结合关键早期结瘤因子基因ENOD40的启动子并调节其表达, 从而调控结瘤数目(Wang et al., 2014).此外, 考虑到miR172自身表达受到1个AON (autoregulation of nodulation)结瘤自调控信号的抑制, 表明miR172-AP2模块可能是整合NF和AON信号通路的关键节点.后续研究表明, miR172在菜豆(Phaseolus vulgaris)和百脉根等豆科植物根瘤形成过程中同样发挥重要作用(Holt et al., 2015; Nova-Franco et al., 2015), 表明miR172-AP2模块在调控豆科植物结瘤及共生固氮方面的功能相对保守.进一步探讨miR172- AP2模块调控豆科植物结瘤固氮的机理, 将会促进豆科植物结瘤和固氮过程遗传机制的解析与调控网络的完善, 并有助于将其更好地应用于作物育种实践. ...
The plant B3 superfamily 1 2008
... AP2/ERF类转录因子均含有1个由60-70个氨基酸残基组成的高度保守的AP2/ERF结构域(Riechmann and Meyerowitz, 1998).该结构域包含3个反向平行的β折叠和1个几乎与β折叠平行的α螺旋.β折叠在识别目标基因的顺式作用元件上具有重要作用, α螺旋可能介导与其它转录因子的互作(Nakano et al., 2006).根据保守结构域差异, 可将AP2/ERF转录因子进一步分为4个亚家族, 即AP2、ERF、DREB (dehydration-responsive element binding protein)和RAV (related to ABI3/VP1) (Xie et al., 2019).AP2亚家族含有2个AP2/ERF结构域, 主要参与调控植物花器官发育; 另外, 最新研究表明, 该亚家族还参与调控植物根系发育和孤雌生殖(Khanday et al., 2019; Li et al., 2019a).RAV亚家族含有1个AP2/ERF及1个B3结构域(Swaminathan et al., 2008), 该亚家族成员参与植物激素及胁迫响应(Hu et al., 2004; Sohn et al., 2006; Zhang et al., 2019; Zhao et al., 2019).ERF和DREB亚家族均仅包含1个AP2/ERF结构域, 两者的区别在于AP2/ERF结构域的第14和19位氨基酸不同, DREB亚家族这2个位置分别是缬氨酸(Val)和谷氨酸(Glu), 而ERF亚家族分别为丙氨酸(Ala)和天冬氨酸(Asp), 功能上两亚家族均参与植物响应生物和非生物胁迫(Dubouzet et al., 2003; Bäumler et al., 2019; He et al., 2019; Wang et al., 2019a). ...
miR172 regulates both vegetative and reproductive development in the perennial woody plant Jatropha curcas 1 2018
... 水稻中, miR172通过抑制AP2基因OsIDS1和SNB的表达来解除它们对靶基因Ehd1 (Early hea- ding date1)的抑制, 从而诱导开花(Lee et al., 2014).最近, 大岩桐中也有类似报道, 即过表达miR172抑制SsAP2表达从而诱导早花(Li et al., 2019b).此外, 在大豆、牵牛花(Pharbitis nil)和麻风树(Jatropha curcas)中也有miR172-AP2模块参与开花时间调控的报道(Glazińska et al., 2009; Zhao et al., 2015; Tang et al., 2018).因此, 我们推测植物主要通过miR172-AP2分子模块调节开花时间来实现其营养生长向生殖生长的过渡. ...
A tomato transcription factor, SlDREB3 enhances the tolerance to chilling in transgenic tomato 2 2019
... AP2/ERF类转录因子均含有1个由60-70个氨基酸残基组成的高度保守的AP2/ERF结构域(Riechmann and Meyerowitz, 1998).该结构域包含3个反向平行的β折叠和1个几乎与β折叠平行的α螺旋.β折叠在识别目标基因的顺式作用元件上具有重要作用, α螺旋可能介导与其它转录因子的互作(Nakano et al., 2006).根据保守结构域差异, 可将AP2/ERF转录因子进一步分为4个亚家族, 即AP2、ERF、DREB (dehydration-responsive element binding protein)和RAV (related to ABI3/VP1) (Xie et al., 2019).AP2亚家族含有2个AP2/ERF结构域, 主要参与调控植物花器官发育; 另外, 最新研究表明, 该亚家族还参与调控植物根系发育和孤雌生殖(Khanday et al., 2019; Li et al., 2019a).RAV亚家族含有1个AP2/ERF及1个B3结构域(Swaminathan et al., 2008), 该亚家族成员参与植物激素及胁迫响应(Hu et al., 2004; Sohn et al., 2006; Zhang et al., 2019; Zhao et al., 2019).ERF和DREB亚家族均仅包含1个AP2/ERF结构域, 两者的区别在于AP2/ERF结构域的第14和19位氨基酸不同, DREB亚家族这2个位置分别是缬氨酸(Val)和谷氨酸(Glu), 而ERF亚家族分别为丙氨酸(Ala)和天冬氨酸(Asp), 功能上两亚家族均参与植物响应生物和非生物胁迫(Dubouzet et al., 2003; Bäumler et al., 2019; He et al., 2019; Wang et al., 2019a). ...
... 各AP2/ERF亚家族转录因子所识别的顺式作用元件具有一定的特异性.例如, AP2亚家族可与启动子中gCAC(A/G)N(A/T)TcCC(a/g)ANG(c/t)顺式作用元件结合来调节靶基因的表达, 进而调控植物的花器官发育、开花时间及时序转换等(Boutilier et al., 2002; Lei et al., 2019; Wang et al., 2019b).RVA亚家族由于含有AP2/ERF和B3结构域, 因此可识别CAACA和CACCTG基序.ERF亚家族主要识别启动子区域的GCC-box, DREB亚家族则可与干旱胁迫响应和冷诱导响应元件(A/GCCGAC)结合, 从而调节植物的非生物胁迫响应(Dubouzet et al., 2003; Agarwal et al., 2010; Mizoi et al., 2012; Bäumler et al., 2019; Wang et al., 2019a). ...
Coordinated regulation of vegetative and reproductive branching in rice 3 2015
... miR172-AP2模块除了在拟南芥中参与花器官发育调控, 在烟草(Nicotiana tabacum)中也有类似作用.将拟南芥miR172在烟草中过表达同样会导致烟草花结构发育异常, 与ap2突变体类似(Mlotshwa et al., 2006), 表明miR172-AP2模块在花器官发育调控方面具保守性.最新研究表明, miR172-AP2模块在油菜(Brassica campestris)、大岩桐(Sinningia speciosa)和玫瑰(Rosa rugosa)等双子叶植物中也参与花器官发育的调控(François et al., 2018; Li et al., 2019b; Wang et al., 2019b).此外, 该调控模式也存在于单子叶植物中.例如, 水稻AP2家族成员OsSNB和OsIDS1在小穗向小花转变及花器官形态建成过程中发挥关键作用.OsSNB突变会导致花发育异常.例如, 小穗向小花转变显著延迟, 产生额外颖苞, 花中浆片异常膨大进而发育成内外稃等; 在OsSNB单突变基础上同时突变OsIDS1则会进一步增强相关表型.水稻中过表达miR172也会导致花器官发育异常, 形成类似Ossnb突变体的表型(Lee and An, 2012; Wang et al., 2015a), 暗示miR172可能通过调控AP2基因OsSNB和OsIDS1的表达来影响水稻花器官的发育.另一禾本科单子叶植物玉米, 其花为单性花, 含有雄穗和雌穗两种花序.玉米的ts (tasselseed)突变体由于雄穗中的雌蕊不退化从而形成两性花, 导致雄穗雌化并进而开花结实(Chuck et al., 2007; Acosta et al., 2009).利用具有雄穗雌化现象的玉米突变体ts4和ts6开展目标基因克隆, 结果显示这2种突变体分别是由于miR172及其靶基因IDS1突变所致, 表明miR172-AP2模块在玉米性别决定中同样具有关键作用(Chuck et al., 2007).而IDS1的异位表达会推迟小穗向花转变, 从而使单个小穗产生多个小花(大于2朵).IDS1同源基因SID1 (SISTER OF INDETERMINATE SPIKELET 1)突变会进一步增强ids1突变体的表型, 表明这2个基因在玉米花器官发育过程中的功能具有累加作用(Chuck et al., 2008).禾本科植物的浆片在开花过程中具有重要作用, 即在特定时期吸水膨胀, 从而撑开颖壳, 完成开花过程.已知大麦(Hordeum vulgare)的闭花受精特性由AP2家族的Cly1 (Cleistogamy 1)基因控制, 该基因表达也受到miR172调控; 当Cly1序列中与miR172结合的位点发生突变时, 会干扰其mRNA与miR172结合, 从而减弱miR172的翻译抑制作用, 进而产生非常小的浆片而无法开颖, 最终发生闭花受精(Nair et al., 2009; Anwar et al., 2018).可见, miR172-AP2模块在不同植物中的功能非常保守, 且对植物花器官发育调控极为重要, miR172通过抑制AP2家族成员的表达, 进而影响AP2基因与B类和C类基因间的互作, 最终调控花器官发育、雌雄分化和开花等过程. ...
... miR172-AP2模块对果实发育的调控至关重要.拟南芥中, miR172-AP2模块可通过调节果荚发育来影响其果实发育.同时, 蛋白FUL与ARF (Auxin Response Factor)可直接互作并结合到miR172前体编码基因的启动子区来激活miR172表达, 进而抑制其下游靶基因AP2及TOE3的表达, 最终促进果实发育(José Ripoll et al., 2015).水稻AP2基因RSR1突变导致其籽粒变大、千粒重及稻米直链淀粉含量增加(Fu and Xue, 2010).进一步研究表明, RSR1正是受miR172调控的靶基因(Wang et al., 2015a), 但RSR1调控下游靶基因导致水稻籽粒变化的分子路径尚无报道.已知RSR1在水稻籽粒中不表达, 暗示miR172-RSR1模块对稻米粒形和品质的影响可能为间接作用, 即通过调控其它组织的生长发育来影响果实发育.综上, miR172-AP2模块对单子叶和双子叶植物果实发育均具有调控作用. ...
Genome-wide exploration and characterization of miR172/ euAP2 genes in Brassica napus L 2 2019
... 各AP2/ERF亚家族转录因子所识别的顺式作用元件具有一定的特异性.例如, AP2亚家族可与启动子中gCAC(A/G)N(A/T)TcCC(a/g)ANG(c/t)顺式作用元件结合来调节靶基因的表达, 进而调控植物的花器官发育、开花时间及时序转换等(Boutilier et al., 2002; Lei et al., 2019; Wang et al., 2019b).RVA亚家族由于含有AP2/ERF和B3结构域, 因此可识别CAACA和CACCTG基序.ERF亚家族主要识别启动子区域的GCC-box, DREB亚家族则可与干旱胁迫响应和冷诱导响应元件(A/GCCGAC)结合, 从而调节植物的非生物胁迫响应(Dubouzet et al., 2003; Agarwal et al., 2010; Mizoi et al., 2012; Bäumler et al., 2019; Wang et al., 2019a). ...
... miR172-AP2模块除了在拟南芥中参与花器官发育调控, 在烟草(Nicotiana tabacum)中也有类似作用.将拟南芥miR172在烟草中过表达同样会导致烟草花结构发育异常, 与ap2突变体类似(Mlotshwa et al., 2006), 表明miR172-AP2模块在花器官发育调控方面具保守性.最新研究表明, miR172-AP2模块在油菜(Brassica campestris)、大岩桐(Sinningia speciosa)和玫瑰(Rosa rugosa)等双子叶植物中也参与花器官发育的调控(François et al., 2018; Li et al., 2019b; Wang et al., 2019b).此外, 该调控模式也存在于单子叶植物中.例如, 水稻AP2家族成员OsSNB和OsIDS1在小穗向小花转变及花器官形态建成过程中发挥关键作用.OsSNB突变会导致花发育异常.例如, 小穗向小花转变显著延迟, 产生额外颖苞, 花中浆片异常膨大进而发育成内外稃等; 在OsSNB单突变基础上同时突变OsIDS1则会进一步增强相关表型.水稻中过表达miR172也会导致花器官发育异常, 形成类似Ossnb突变体的表型(Lee and An, 2012; Wang et al., 2015a), 暗示miR172可能通过调控AP2基因OsSNB和OsIDS1的表达来影响水稻花器官的发育.另一禾本科单子叶植物玉米, 其花为单性花, 含有雄穗和雌穗两种花序.玉米的ts (tasselseed)突变体由于雄穗中的雌蕊不退化从而形成两性花, 导致雄穗雌化并进而开花结实(Chuck et al., 2007; Acosta et al., 2009).利用具有雄穗雌化现象的玉米突变体ts4和ts6开展目标基因克隆, 结果显示这2种突变体分别是由于miR172及其靶基因IDS1突变所致, 表明miR172-AP2模块在玉米性别决定中同样具有关键作用(Chuck et al., 2007).而IDS1的异位表达会推迟小穗向花转变, 从而使单个小穗产生多个小花(大于2朵).IDS1同源基因SID1 (SISTER OF INDETERMINATE SPIKELET 1)突变会进一步增强ids1突变体的表型, 表明这2个基因在玉米花器官发育过程中的功能具有累加作用(Chuck et al., 2008).禾本科植物的浆片在开花过程中具有重要作用, 即在特定时期吸水膨胀, 从而撑开颖壳, 完成开花过程.已知大麦(Hordeum vulgare)的闭花受精特性由AP2家族的Cly1 (Cleistogamy 1)基因控制, 该基因表达也受到miR172调控; 当Cly1序列中与miR172结合的位点发生突变时, 会干扰其mRNA与miR172结合, 从而减弱miR172的翻译抑制作用, 进而产生非常小的浆片而无法开颖, 最终发生闭花受精(Nair et al., 2009; Anwar et al., 2018).可见, miR172-AP2模块在不同植物中的功能非常保守, 且对植物花器官发育调控极为重要, miR172通过抑制AP2家族成员的表达, 进而影响AP2基因与B类和C类基因间的互作, 最终调控花器官发育、雌雄分化和开花等过程. ...
Ectopic expression of miR156 represses nodulation and causes morphological and developmental changes in Lotus japonicus 1 2015
... 根瘤菌不仅能感染豆科植物根系形成根瘤, 还可与其共生固氮.在苜蓿(Medicago sativa)中首先证实miR166和miR169参与根瘤发育的调控(Combier et al., 2006; Boualem et al., 2008).百脉根(Lotus corniculatus)中的研究也表明, miR171、miR397和miR156与根瘤的发育、病毒感染和固氮能力密切相关(De Luis et al., 2012; Wang et al., 2015b).在大豆中过表达miR482、miR1512、miR1515、miR156及miR167均可以显著改变大豆根瘤的数量(Li et al., 2010; Wang et al., 2015c).因此, miRNA在豆科植物结瘤和共生固氮过程中发挥重要作用. ...
MicroRNA167-directed regulation of the auxin response factors GmARF8a and GmARF8b is required for soybean nodulation and lateral root development 1 2015
... 根瘤菌不仅能感染豆科植物根系形成根瘤, 还可与其共生固氮.在苜蓿(Medicago sativa)中首先证实miR166和miR169参与根瘤发育的调控(Combier et al., 2006; Boualem et al., 2008).百脉根(Lotus corniculatus)中的研究也表明, miR171、miR397和miR156与根瘤的发育、病毒感染和固氮能力密切相关(De Luis et al., 2012; Wang et al., 2015b).在大豆中过表达miR482、miR1512、miR1515、miR156及miR167均可以显著改变大豆根瘤的数量(Li et al., 2010; Wang et al., 2015c).因此, miRNA在豆科植物结瘤和共生固氮过程中发挥重要作用. ...
Soybean miR172c targets the repressive AP2 transcription factor NNC1 to activate ENOD40 expression and regulate nodule initiation 2 2014
... 目前, 常用的miRNA靶基因鉴定方法包括生物信息学预测、AGO蛋白免疫共沉淀、双荧光素酶和降解组测序等(董淼等, 2013).miRNA主要通过剪切靶基因mRNA和抑制靶基因翻译2种途径调控靶基因, 但具体方式取决于miRNA与其靶基因mRNA的互补程度及作用位置(Hutvágner and Zamore, 2002; Llave et al., 2002; Schwab et al., 2005; Jung et al., 2007; Zhu et al., 2009).miRNA172与AP2家族基因的互补区域通常位于靠近3°UTR的CDS区, 主要调控方式为抑制AP2靶基因翻译, 但也存在部分与靶基因mRNA完全互补从而对其进行剪切调控的情况(Aukerman and Sakai, 2003; Schmid et al., 2003; Chen, 2004; Mathieu et al., 2009).当miRNA通过抑制翻译对靶基因进行调控时, 两者将表现出协同表达模式(Zhu et al., 2009).例如, 水稻中miRNA172及其靶基因OsIDS1和RSR1在包括胚和胚乳在内的大多数组织中表达模式一致; 同样, 过量表达miR172也会导致各发育阶段穗子中的OsIDS1 (OsINDETERMINATE SPIKELET1)和RSR1 (Rice Starch Regulator1)表达呈现出升高或降低的动态变化模式, 表明miR172对靶基因的调控具有时空特异性.在玉米(Zea mays)中, miR172通过剪切靶基因GL15的mRNA来调控玉米从营养生长向生殖生长的过渡(Lauter et al., 2005).此外, 在大豆(Glycine max)、马铃薯(Solanum tuberosum)和拟南芥中也有类似报道, 即miR172通过切割其靶基因的mRNA来调控植物的生长发育(Wang et al., 2014). ...
... 之前的研究表明, 根瘤菌处理3小时的大豆根中miR172表达显著上调(Subramanian et al., 2008), 且在成熟的根瘤中miR172c高度表达(Wang et al., 2009).而在大豆中过表达miR172c可促进根瘤菌的感染和根瘤的形态发生, 从而增加根瘤数量(Yan et al., 2013).此外, miR172还可调控结瘤因子(NF)的表达(Martin et al., 2009).例如, miR172的靶基因GmNNC1编码1个AP2家族转录抑制子, 可直接结合关键早期结瘤因子基因ENOD40的启动子并调节其表达, 从而调控结瘤数目(Wang et al., 2014).此外, 考虑到miR172自身表达受到1个AON (autoregulation of nodulation)结瘤自调控信号的抑制, 表明miR172-AP2模块可能是整合NF和AON信号通路的关键节点.后续研究表明, miR172在菜豆(Phaseolus vulgaris)和百脉根等豆科植物根瘤形成过程中同样发挥重要作用(Holt et al., 2015; Nova-Franco et al., 2015), 表明miR172-AP2模块在调控豆科植物结瘤及共生固氮方面的功能相对保守.进一步探讨miR172- AP2模块调控豆科植物结瘤固氮的机理, 将会促进豆科植物结瘤和固氮过程遗传机制的解析与调控网络的完善, 并有助于将其更好地应用于作物育种实践. ...
Identification and expression analysis of miRNAs from nitrogen-fixing soybean nodules 1 2009
... 之前的研究表明, 根瘤菌处理3小时的大豆根中miR172表达显著上调(Subramanian et al., 2008), 且在成熟的根瘤中miR172c高度表达(Wang et al., 2009).而在大豆中过表达miR172c可促进根瘤菌的感染和根瘤的形态发生, 从而增加根瘤数量(Yan et al., 2013).此外, miR172还可调控结瘤因子(NF)的表达(Martin et al., 2009).例如, miR172的靶基因GmNNC1编码1个AP2家族转录抑制子, 可直接结合关键早期结瘤因子基因ENOD40的启动子并调节其表达, 从而调控结瘤数目(Wang et al., 2014).此外, 考虑到miR172自身表达受到1个AON (autoregulation of nodulation)结瘤自调控信号的抑制, 表明miR172-AP2模块可能是整合NF和AON信号通路的关键节点.后续研究表明, miR172在菜豆(Phaseolus vulgaris)和百脉根等豆科植物根瘤形成过程中同样发挥重要作用(Holt et al., 2015; Nova-Franco et al., 2015), 表明miR172-AP2模块在调控豆科植物结瘤及共生固氮方面的功能相对保守.进一步探讨miR172- AP2模块调控豆科植物结瘤固氮的机理, 将会促进豆科植物结瘤和固氮过程遗传机制的解析与调控网络的完善, 并有助于将其更好地应用于作物育种实践. ...
On reconciling the interactions between APETALA2, miR172 and AGAMOUS with the ABC model of flower development 1 2010
... AP2在模式植物拟南芥中被发现时便认为与植物花器官特征调控相关(Kunst et al., 1989).拟南芥的花包括花萼、花瓣、雄蕊和心皮4轮器官, 其特征由A、B和C三类基因控制(Bowman et al., 2012).AP2属于A类基因, 不仅参与第1轮花萼特征的调控, 还与B类基因AP3和PI (PISTILLATA)共同控制第2轮花瓣的形成; 并可通过拮抗C类基因AG (AGAMOUS)的作用抑制雄蕊和心皮的发育(Shannon and Meeks- Wagner, 1993; Elliott et al., 1996).miR172在拟南芥花器官中具有优势表达, 且miR172过表达植株花器官发育异常, 与ap2突变体类似(Park et al., 2002), 主要原因在于miR172从翻译水平抑制了AP2的表达(Aukerman and Sakai, 2003; Chen, 2004).时空表达分析结果表明, AP2表达产物主要积累于花器官外周, 与miR172信号在第2或第3轮花器官处有瞬时重叠(Wollmann et al., 2010).尽管miR172和AP2共表达时间非常短暂, 但两者共同调控花分生细胞的时序进程, 在花器官发育中发挥决定性作用(Ji et al., 2011). ...
The sequential action of miR156 and miR172 regulates developmental timing in Arabidopsis 3 2009
... miR172在植物的生殖转换、花器官形成及胁迫响应等方面具有重要调控作用(Wu et al., 2009; 王幼宁等, 2016; 赵晓晖等, 2017; Luan et al., 2018; Saminathan et al., 2019).miR172在高等植物中高度保守, 虽然不同物种中miR172成员的数目和长度可能不同, 但在亲缘关系较近的物种间序列高度保守, 暗示其在不同物种中可能具有相似的调控方式和生物学功能. ...
... 此外, miR172-AP2模块在植物营养生长期不同阶段转变过程中也发挥重要作用(Wu et al., 2009).例如, GL15 (Glossy 15 )是1个控制玉米幼苗向成熟期转变时间的AP2基因, 其表达同样受到miR172调控(Moose and Sisco, 1994, 1996; Lauter et al., 2005).可见, miR172-AP2模块在植物时序转换调控中也发挥关键作用.AP2家族成员众多, 在功能上兼具共性和特异性, 解析其分子机制可为生产实践中利用特定的miR172-AP2互作模块开展分子设计育种提供可能, 从而实现真正精准育种. ...
AP2/ERF transcription factor regulatory networks in hormone and abiotic stress responses in Arabidopsis 1 2019
... AP2/ERF类转录因子均含有1个由60-70个氨基酸残基组成的高度保守的AP2/ERF结构域(Riechmann and Meyerowitz, 1998).该结构域包含3个反向平行的β折叠和1个几乎与β折叠平行的α螺旋.β折叠在识别目标基因的顺式作用元件上具有重要作用, α螺旋可能介导与其它转录因子的互作(Nakano et al., 2006).根据保守结构域差异, 可将AP2/ERF转录因子进一步分为4个亚家族, 即AP2、ERF、DREB (dehydration-responsive element binding protein)和RAV (related to ABI3/VP1) (Xie et al., 2019).AP2亚家族含有2个AP2/ERF结构域, 主要参与调控植物花器官发育; 另外, 最新研究表明, 该亚家族还参与调控植物根系发育和孤雌生殖(Khanday et al., 2019; Li et al., 2019a).RAV亚家族含有1个AP2/ERF及1个B3结构域(Swaminathan et al., 2008), 该亚家族成员参与植物激素及胁迫响应(Hu et al., 2004; Sohn et al., 2006; Zhang et al., 2019; Zhao et al., 2019).ERF和DREB亚家族均仅包含1个AP2/ERF结构域, 两者的区别在于AP2/ERF结构域的第14和19位氨基酸不同, DREB亚家族这2个位置分别是缬氨酸(Val)和谷氨酸(Glu), 而ERF亚家族分别为丙氨酸(Ala)和天冬氨酸(Asp), 功能上两亚家族均参与植物响应生物和非生物胁迫(Dubouzet et al., 2003; Bäumler et al., 2019; He et al., 2019; Wang et al., 2019a). ...
miR172 regulates soybean nodulation 1 2013
... 之前的研究表明, 根瘤菌处理3小时的大豆根中miR172表达显著上调(Subramanian et al., 2008), 且在成熟的根瘤中miR172c高度表达(Wang et al., 2009).而在大豆中过表达miR172c可促进根瘤菌的感染和根瘤的形态发生, 从而增加根瘤数量(Yan et al., 2013).此外, miR172还可调控结瘤因子(NF)的表达(Martin et al., 2009).例如, miR172的靶基因GmNNC1编码1个AP2家族转录抑制子, 可直接结合关键早期结瘤因子基因ENOD40的启动子并调节其表达, 从而调控结瘤数目(Wang et al., 2014).此外, 考虑到miR172自身表达受到1个AON (autoregulation of nodulation)结瘤自调控信号的抑制, 表明miR172-AP2模块可能是整合NF和AON信号通路的关键节点.后续研究表明, miR172在菜豆(Phaseolus vulgaris)和百脉根等豆科植物根瘤形成过程中同样发挥重要作用(Holt et al., 2015; Nova-Franco et al., 2015), 表明miR172-AP2模块在调控豆科植物结瘤及共生固氮方面的功能相对保守.进一步探讨miR172- AP2模块调控豆科植物结瘤固氮的机理, 将会促进豆科植物结瘤和固氮过程遗传机制的解析与调控网络的完善, 并有助于将其更好地应用于作物育种实践. ...
Prediction and verification of microRNAs related to proline accumulation under drought stress in potato 1 2013
... 植物不像动物可以躲避逆境, 因此其进化出特有的复杂且精细的分子调控机制来应对各种环境胁迫.许多研究表明, miRNA在植物响应各种生物和非生物胁迫过程中具有重要作用.miR172作为miRNA家族中非常保守的一员, 其自身表达受低温、盐碱和干旱等多种胁迫诱导, 同时它也可通过调控AP2类转录因子参与植物响应各种环境胁迫(Zhou et al., 2008; Frazier et al., 2011; Yang et al., 2013; Candar-Cakir et al., 2016).当然, miR172-AP2的具体调控模式还与植物种类及其响应的逆境因子密切相关.拟南芥中, ABA及渗透胁迫可诱导miR172与其靶基因SNZ共表达.miR172b过表达或snz突变体对ABA及渗透胁迫的敏感性增强, 证明miR172通过控制其靶基因SNZ来介导拟南芥对渗透胁迫的响应(Zou et al., 2013).干旱胁迫可诱导miR172e的表达, 同时过表达miR172e又可显著增强拟南芥对干旱的抗性.进一步研究表明, 干旱通过诱导GI (GIGANTEA)表达促进miR172e的积累以抑制下游靶基因TOE1的表达, 进而影响TOE1蛋白与WRKY44的互作, 最终实现对植物抗旱能力的调控(Han et al., 2013).在大豆中过表达miRNA172c或敲除GmNNC1均可增强大豆的耐盐性; 反之, 干扰miR172的表达或过表达GmNNC1均增强了大豆的盐敏感性(Sahito et al., 2017).此外, 在拟南芥中过表达大豆miR172c也可显著增强拟南芥对干旱及盐胁迫的耐受性(Zou et al., 2013).尽管目前人们对拟南芥和大豆中miR172介导的植物响应逆境胁迫机制有了较清晰的认识, 但在其它植物特别是水稻和小麦等重要作物中的相关研究仍停留在全基因组表达分析层面, 还未涉及具体分子机制的解析与调控网络的构建.例如, 香蕉(Musa nana)遇到冷胁迫后miR172的表达会显著上调, 初步推测miR172可能在冷应激反应中起中心协调作用(刘炜婳等, 2018).此外, 在番茄(Lycopersicon esculentum)中过表达miR172a和miR172b显著增强了其对致病疫霉的抗性, 潜在分子机理也与以miR172-AP2模块为核心的表达调控路径相关(Luan et al., 2018). ...
GmRAV1 regulates regeneration of roots and adventitious buds by the cytokinin signaling pathway in Arabidopsis and soybean 1 2019
... AP2/ERF类转录因子均含有1个由60-70个氨基酸残基组成的高度保守的AP2/ERF结构域(Riechmann and Meyerowitz, 1998).该结构域包含3个反向平行的β折叠和1个几乎与β折叠平行的α螺旋.β折叠在识别目标基因的顺式作用元件上具有重要作用, α螺旋可能介导与其它转录因子的互作(Nakano et al., 2006).根据保守结构域差异, 可将AP2/ERF转录因子进一步分为4个亚家族, 即AP2、ERF、DREB (dehydration-responsive element binding protein)和RAV (related to ABI3/VP1) (Xie et al., 2019).AP2亚家族含有2个AP2/ERF结构域, 主要参与调控植物花器官发育; 另外, 最新研究表明, 该亚家族还参与调控植物根系发育和孤雌生殖(Khanday et al., 2019; Li et al., 2019a).RAV亚家族含有1个AP2/ERF及1个B3结构域(Swaminathan et al., 2008), 该亚家族成员参与植物激素及胁迫响应(Hu et al., 2004; Sohn et al., 2006; Zhang et al., 2019; Zhao et al., 2019).ERF和DREB亚家族均仅包含1个AP2/ERF结构域, 两者的区别在于AP2/ERF结构域的第14和19位氨基酸不同, DREB亚家族这2个位置分别是缬氨酸(Val)和谷氨酸(Glu), 而ERF亚家族分别为丙氨酸(Ala)和天冬氨酸(Asp), 功能上两亚家族均参与植物响应生物和非生物胁迫(Dubouzet et al., 2003; Bäumler et al., 2019; He et al., 2019; Wang et al., 2019a). ...
Dual functions of GmTOE4a in the regulation of photoperiod-mediated flowering and plant morphology in soybean 1 2015
... 水稻中, miR172通过抑制AP2基因OsIDS1和SNB的表达来解除它们对靶基因Ehd1 (Early hea- ding date1)的抑制, 从而诱导开花(Lee et al., 2014).最近, 大岩桐中也有类似报道, 即过表达miR172抑制SsAP2表达从而诱导早花(Li et al., 2019b).此外, 在大豆、牵牛花(Pharbitis nil)和麻风树(Jatropha curcas)中也有miR172-AP2模块参与开花时间调控的报道(Glazińska et al., 2009; Zhao et al., 2015; Tang et al., 2018).因此, 我们推测植物主要通过miR172-AP2分子模块调节开花时间来实现其营养生长向生殖生长的过渡. ...
The MdWRKY31 transcription factor binds to the MdRAV1 promoter to mediate ABA sensitivity 1 2019
... AP2/ERF类转录因子均含有1个由60-70个氨基酸残基组成的高度保守的AP2/ERF结构域(Riechmann and Meyerowitz, 1998).该结构域包含3个反向平行的β折叠和1个几乎与β折叠平行的α螺旋.β折叠在识别目标基因的顺式作用元件上具有重要作用, α螺旋可能介导与其它转录因子的互作(Nakano et al., 2006).根据保守结构域差异, 可将AP2/ERF转录因子进一步分为4个亚家族, 即AP2、ERF、DREB (dehydration-responsive element binding protein)和RAV (related to ABI3/VP1) (Xie et al., 2019).AP2亚家族含有2个AP2/ERF结构域, 主要参与调控植物花器官发育; 另外, 最新研究表明, 该亚家族还参与调控植物根系发育和孤雌生殖(Khanday et al., 2019; Li et al., 2019a).RAV亚家族含有1个AP2/ERF及1个B3结构域(Swaminathan et al., 2008), 该亚家族成员参与植物激素及胁迫响应(Hu et al., 2004; Sohn et al., 2006; Zhang et al., 2019; Zhao et al., 2019).ERF和DREB亚家族均仅包含1个AP2/ERF结构域, 两者的区别在于AP2/ERF结构域的第14和19位氨基酸不同, DREB亚家族这2个位置分别是缬氨酸(Val)和谷氨酸(Glu), 而ERF亚家族分别为丙氨酸(Ala)和天冬氨酸(Asp), 功能上两亚家族均参与植物响应生物和非生物胁迫(Dubouzet et al., 2003; Bäumler et al., 2019; He et al., 2019; Wang et al., 2019a). ...
Identification of cold-inducible microRNAs in plants by transcriptome analysis 1 2008
... 植物不像动物可以躲避逆境, 因此其进化出特有的复杂且精细的分子调控机制来应对各种环境胁迫.许多研究表明, miRNA在植物响应各种生物和非生物胁迫过程中具有重要作用.miR172作为miRNA家族中非常保守的一员, 其自身表达受低温、盐碱和干旱等多种胁迫诱导, 同时它也可通过调控AP2类转录因子参与植物响应各种环境胁迫(Zhou et al., 2008; Frazier et al., 2011; Yang et al., 2013; Candar-Cakir et al., 2016).当然, miR172-AP2的具体调控模式还与植物种类及其响应的逆境因子密切相关.拟南芥中, ABA及渗透胁迫可诱导miR172与其靶基因SNZ共表达.miR172b过表达或snz突变体对ABA及渗透胁迫的敏感性增强, 证明miR172通过控制其靶基因SNZ来介导拟南芥对渗透胁迫的响应(Zou et al., 2013).干旱胁迫可诱导miR172e的表达, 同时过表达miR172e又可显著增强拟南芥对干旱的抗性.进一步研究表明, 干旱通过诱导GI (GIGANTEA)表达促进miR172e的积累以抑制下游靶基因TOE1的表达, 进而影响TOE1蛋白与WRKY44的互作, 最终实现对植物抗旱能力的调控(Han et al., 2013).在大豆中过表达miRNA172c或敲除GmNNC1均可增强大豆的耐盐性; 反之, 干扰miR172的表达或过表达GmNNC1均增强了大豆的盐敏感性(Sahito et al., 2017).此外, 在拟南芥中过表达大豆miR172c也可显著增强拟南芥对干旱及盐胁迫的耐受性(Zou et al., 2013).尽管目前人们对拟南芥和大豆中miR172介导的植物响应逆境胁迫机制有了较清晰的认识, 但在其它植物特别是水稻和小麦等重要作物中的相关研究仍停留在全基因组表达分析层面, 还未涉及具体分子机制的解析与调控网络的构建.例如, 香蕉(Musa nana)遇到冷胁迫后miR172的表达会显著上调, 初步推测miR172可能在冷应激反应中起中心协调作用(刘炜婳等, 2018).此外, 在番茄(Lycopersicon esculentum)中过表达miR172a和miR172b显著增强了其对致病疫霉的抗性, 潜在分子机理也与以miR172-AP2模块为核心的表达调控路径相关(Luan et al., 2018). ...
Regulation of flowering time and floral patterning by miR172 2 2011
... 虽然miR172家族成员成熟miRNA的序列高度保守, 但功能上并非简单重复.研究表明, miR172各成员启动子序列及其包含的顺式作用元件并不相同, 从而决定了它们表达的时空特异性(Zhu and Helliwell, 2011; 刘炜婳等, 2018).双子叶模式植物拟南芥(Arabidopsis thaliana)中存在5个miR172成员, 即miR172a-1、miR172a-2、miR172b-1、miR172b-2和miR172c (Chen, 2004).虽然拟南芥miR172各成员的表达模式不尽相同, 但根据其表达量和表达模式总体可分为2类: 一类包括miR172b-2和miR172c, 它们在拟南芥生长发育过程中表达量一直很低; 另一类包括miR172a-1、miR172a-2和miR172b-1, 它们的表达量随着拟南芥的生长发育逐渐升高, 到生殖生长阶段达到峰值(Jung et al., 2007).在表达的组织特异性方面, 除了在植物根中未检测到miR172的表达, 在其它包括花、叶片、茎和角果等组织中均检测到其表达(赵晓晖等, 2017).单子叶模式植物水稻(Oryza sativa)中存在4个miR172成员(miR172a-miR172d), 它们在苗期表达均较强, 但同样各成员表达也具有一定的组织特异性.例如, miR172c在水稻籽粒中不表达(Zhu et al., 2009).综上, miR172在植物中的表达具有明显的时空特异性(Zhu and Helliwell, 2011; 赵晓晖等, 2017). ...
... ).综上, miR172在植物中的表达具有明显的时空特异性(Zhu and Helliwell, 2011; 赵晓晖等, 2017). ...
Over-expression of miR172 causes loss of spikelet determinacy and floral organ abnormalities in rice ( Oryza sativa) 4 2009
... miRNA家族中, miR172是较早被发现同时也是被研究最透彻的成员之一(Park et al., 2002), 其靶基因主要编码AP2/ERF (APETALA2/ethylene responsive factor)类转录因子.AP2/ERF转录因子为植物所特有, 且几乎参与植物生长发育调控的各个环节.miRNA172通过特异或非特异性结合AP2/ERF的mRNA, 以mRNA剪切和翻译抑制2种方式调控AP2/ERF靶基因的表达, 从而调控植物众多生长发育过程(Jung et al., 2007; Zhu et al., 2009; 刘炜婳等, 2018).近期, 研究者在miRNA172-AP2模块调控植物花器官发育、时序转换、块茎及果实发育、根瘤形成和胁迫响应等方面取得一系列研究进展(王幼宁等, 2016; 赵晓晖等, 2017; Luan et al., 2018; Saminathan et al., 2019).本文以miRNA172-AP2模块为中心, 在简要介绍miR172和AP2的结构、功能及互作模式基础上, 重点论述miRNA172-AP2模块参与的植物生长发育及胁迫响应调控, 深入解析其生物学效应和分子机制, 并初步建立由miR172-AP2模块介导的植物生长发育分子调控网络. ...
... 虽然miR172家族成员成熟miRNA的序列高度保守, 但功能上并非简单重复.研究表明, miR172各成员启动子序列及其包含的顺式作用元件并不相同, 从而决定了它们表达的时空特异性(Zhu and Helliwell, 2011; 刘炜婳等, 2018).双子叶模式植物拟南芥(Arabidopsis thaliana)中存在5个miR172成员, 即miR172a-1、miR172a-2、miR172b-1、miR172b-2和miR172c (Chen, 2004).虽然拟南芥miR172各成员的表达模式不尽相同, 但根据其表达量和表达模式总体可分为2类: 一类包括miR172b-2和miR172c, 它们在拟南芥生长发育过程中表达量一直很低; 另一类包括miR172a-1、miR172a-2和miR172b-1, 它们的表达量随着拟南芥的生长发育逐渐升高, 到生殖生长阶段达到峰值(Jung et al., 2007).在表达的组织特异性方面, 除了在植物根中未检测到miR172的表达, 在其它包括花、叶片、茎和角果等组织中均检测到其表达(赵晓晖等, 2017).单子叶模式植物水稻(Oryza sativa)中存在4个miR172成员(miR172a-miR172d), 它们在苗期表达均较强, 但同样各成员表达也具有一定的组织特异性.例如, miR172c在水稻籽粒中不表达(Zhu et al., 2009).综上, miR172在植物中的表达具有明显的时空特异性(Zhu and Helliwell, 2011; 赵晓晖等, 2017). ...
... 目前, 常用的miRNA靶基因鉴定方法包括生物信息学预测、AGO蛋白免疫共沉淀、双荧光素酶和降解组测序等(董淼等, 2013).miRNA主要通过剪切靶基因mRNA和抑制靶基因翻译2种途径调控靶基因, 但具体方式取决于miRNA与其靶基因mRNA的互补程度及作用位置(Hutvágner and Zamore, 2002; Llave et al., 2002; Schwab et al., 2005; Jung et al., 2007; Zhu et al., 2009).miRNA172与AP2家族基因的互补区域通常位于靠近3°UTR的CDS区, 主要调控方式为抑制AP2靶基因翻译, 但也存在部分与靶基因mRNA完全互补从而对其进行剪切调控的情况(Aukerman and Sakai, 2003; Schmid et al., 2003; Chen, 2004; Mathieu et al., 2009).当miRNA通过抑制翻译对靶基因进行调控时, 两者将表现出协同表达模式(Zhu et al., 2009).例如, 水稻中miRNA172及其靶基因OsIDS1和RSR1在包括胚和胚乳在内的大多数组织中表达模式一致; 同样, 过量表达miR172也会导致各发育阶段穗子中的OsIDS1 (OsINDETERMINATE SPIKELET1)和RSR1 (Rice Starch Regulator1)表达呈现出升高或降低的动态变化模式, 表明miR172对靶基因的调控具有时空特异性.在玉米(Zea mays)中, miR172通过剪切靶基因GL15的mRNA来调控玉米从营养生长向生殖生长的过渡(Lauter et al., 2005).此外, 在大豆(Glycine max)、马铃薯(Solanum tuberosum)和拟南芥中也有类似报道, 即miR172通过切割其靶基因的mRNA来调控植物的生长发育(Wang et al., 2014). ...
... ).当miRNA通过抑制翻译对靶基因进行调控时, 两者将表现出协同表达模式(Zhu et al., 2009).例如, 水稻中miRNA172及其靶基因OsIDS1和RSR1在包括胚和胚乳在内的大多数组织中表达模式一致; 同样, 过量表达miR172也会导致各发育阶段穗子中的OsIDS1 (OsINDETERMINATE SPIKELET1)和RSR1 (Rice Starch Regulator1)表达呈现出升高或降低的动态变化模式, 表明miR172对靶基因的调控具有时空特异性.在玉米(Zea mays)中, miR172通过剪切靶基因GL15的mRNA来调控玉米从营养生长向生殖生长的过渡(Lauter et al., 2005).此外, 在大豆(Glycine max)、马铃薯(Solanum tuberosum)和拟南芥中也有类似报道, 即miR172通过切割其靶基因的mRNA来调控植物的生长发育(Wang et al., 2014). ...
miR172b controls the transition to autotrophic development inhibited by ABA in Arabidopsis 2 2013
... 植物不像动物可以躲避逆境, 因此其进化出特有的复杂且精细的分子调控机制来应对各种环境胁迫.许多研究表明, miRNA在植物响应各种生物和非生物胁迫过程中具有重要作用.miR172作为miRNA家族中非常保守的一员, 其自身表达受低温、盐碱和干旱等多种胁迫诱导, 同时它也可通过调控AP2类转录因子参与植物响应各种环境胁迫(Zhou et al., 2008; Frazier et al., 2011; Yang et al., 2013; Candar-Cakir et al., 2016).当然, miR172-AP2的具体调控模式还与植物种类及其响应的逆境因子密切相关.拟南芥中, ABA及渗透胁迫可诱导miR172与其靶基因SNZ共表达.miR172b过表达或snz突变体对ABA及渗透胁迫的敏感性增强, 证明miR172通过控制其靶基因SNZ来介导拟南芥对渗透胁迫的响应(Zou et al., 2013).干旱胁迫可诱导miR172e的表达, 同时过表达miR172e又可显著增强拟南芥对干旱的抗性.进一步研究表明, 干旱通过诱导GI (GIGANTEA)表达促进miR172e的积累以抑制下游靶基因TOE1的表达, 进而影响TOE1蛋白与WRKY44的互作, 最终实现对植物抗旱能力的调控(Han et al., 2013).在大豆中过表达miRNA172c或敲除GmNNC1均可增强大豆的耐盐性; 反之, 干扰miR172的表达或过表达GmNNC1均增强了大豆的盐敏感性(Sahito et al., 2017).此外, 在拟南芥中过表达大豆miR172c也可显著增强拟南芥对干旱及盐胁迫的耐受性(Zou et al., 2013).尽管目前人们对拟南芥和大豆中miR172介导的植物响应逆境胁迫机制有了较清晰的认识, 但在其它植物特别是水稻和小麦等重要作物中的相关研究仍停留在全基因组表达分析层面, 还未涉及具体分子机制的解析与调控网络的构建.例如, 香蕉(Musa nana)遇到冷胁迫后miR172的表达会显著上调, 初步推测miR172可能在冷应激反应中起中心协调作用(刘炜婳等, 2018).此外, 在番茄(Lycopersicon esculentum)中过表达miR172a和miR172b显著增强了其对致病疫霉的抗性, 潜在分子机理也与以miR172-AP2模块为核心的表达调控路径相关(Luan et al., 2018). ...
... ).此外, 在拟南芥中过表达大豆miR172c也可显著增强拟南芥对干旱及盐胁迫的耐受性(Zou et al., 2013).尽管目前人们对拟南芥和大豆中miR172介导的植物响应逆境胁迫机制有了较清晰的认识, 但在其它植物特别是水稻和小麦等重要作物中的相关研究仍停留在全基因组表达分析层面, 还未涉及具体分子机制的解析与调控网络的构建.例如, 香蕉(Musa nana)遇到冷胁迫后miR172的表达会显著上调, 初步推测miR172可能在冷应激反应中起中心协调作用(刘炜婳等, 2018).此外, 在番茄(Lycopersicon esculentum)中过表达miR172a和miR172b显著增强了其对致病疫霉的抗性, 潜在分子机理也与以miR172-AP2模块为核心的表达调控路径相关(Luan et al., 2018). ...
 ,*, 刘巧泉
,*, 刘巧泉
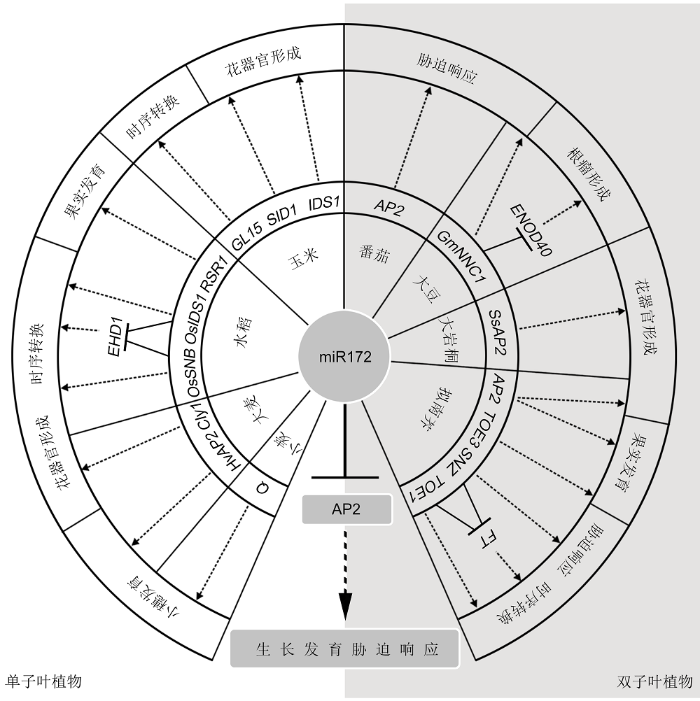 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}