Abstract Upon blue light-triggered autophosphorylation, the activated PHOT1 and PHOT2 photoreceptors function solely or redundantly to regulate diverse responses, such as stomatal opening, chloroplast movements, phototropism, and leaf positioning and flattening in Arabidopsis thaliana. Members of NPH3/RPT2-Like family (NRL) are necessary to elicit several photo-responses. NPH3 is required for phototropism, leaf expansion and positioning. RPT2 regulates chloroplast accumulation movement besides NPH3-mediated responses. NCH1 was recently identified as a Phot1-interacting protein that functions redundantly with RPT2 to mediate chloroplast accumulation movement, but not chloroplast avoidance movement. This article summarized the function of NRL family members in phototropin-mediated signaling and prospected further research, thus providing clues for fully revealing the function of the NRL family proteins. Keywords:Arabidopsis;blue light;NPH3/RPT2-Like family protein;phototropin
PDF (853KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 赵青平, 马世凡, 李芮茜, 王田雨, 赵翔. 拟南芥NPH3/RPT2-Like (NRL)家族蛋白在向光素信号转导通路中的作用研究进展. 植物学报, 2020, 55(2): 240-253 doi:10.11983/CBB19187 Zhao Qingping, Ma Shifan, Li Ruixi, Wang Tianyu, Zhao Xiang. Advances of NPH3/RPT2-Like (NRL) Family Proteins in Phototropin-mediated Signaling in Arabidopsis thaliana. Chinese Bulletin of Botany, 2020, 55(2): 240-253 doi:10.11983/CBB19187
进化树主要参考Suetsugu等(2016)和Christie等(2018)已经报道的进化树, 但分支结构略有不同。6个主要分支颜色的调整以及命名的加入采用Adobe illustrator软件。 Figure 1Bayesian phylogenetic tree of the NPH3/RPT2-Like (NRL) family protein in Arabidopsis
Construction of this phylogenetic tree was based on Suetsugu et al. (2016) and Christie et al. (2018) with slight modifications. The colors and names of six major clades of NRL proteins in Arabidopsis were constructed using the Adobe illustrator software.
NPH3和RPT2是来自2个不同分支的NRL家族成员, 通过与向光素相互作用调节蓝光诱导下胚轴向光性、叶绿体聚光运动、气孔开放以及叶片伸展与定位(Christie et al., 2015, 2018)。NRL8是AGC激酶(cAMP依赖的蛋白激酶A, cGMP依赖的蛋白激酶G和磷脂依赖的蛋白激酶C)活性依赖的花粉管生长调节因子(Lalanne et al., 2004; Zhang et al., 2008), 该蛋白与RPT2来自分支II。与NPH3类似, SR1IP1 (AtSR1 INTERACTING PROTEIN 1)也称NRL31, 可以与CRL3 ( CUL3 RING E3 UBIQUITIN LIGASE)形成复合体, 调节底物的泛素化(Zhang et al., 2014)。SR1IP1通过调节拟南芥中Ca2+/钙调素结合的转录因子AtSR1 (SIGNAL RESPONSIVE 1)的泛素化降解, 增强植物对细菌性病原体的抗性(Zhang et al., 2014)。但系统进化研究发现, SR1IP1也来自分支II, 且比NRL8与RPT2亲缘关系更近, 与NPH3的进化明显不同。Suetsugu等(2016)在筛选参与调节叶绿体运动的NRL家族蛋白时, 发现NRL31基因突变抑制了光诱导的叶绿体运动, 且蛋白NRL31可以与PHOT1发生体内互作, 因此将其重命名为NCH1 (NRL PROTEIN FORCHLOROPLASTMOVEMENT 1)。DEFECTIVELY ORGANIZED TRIBUTARIES 3/NRL23是与NPH3来自同一个分支的NRL家族蛋白, 主要参与维管组织发育以及叶脉结构调节(Petricka et al., 2008)。虽然NRL23与NPH3的亲缘关系较近, 但是该蛋白是否在向光素介导的信号途径中发挥作用尚未见报道。NRL5、NRL6、NRL7、NRL15、NRL20、NRL21、NRL26和NRL30是分支III中的NPY (NAKED PINS IN YUCCA)家族成员, 其中NRL6/NPY2、NRL7/NPY4、NRL20/NPY1、NRL21/NPY5和NRL30/NPY3以功能冗余的方式与AGCVIII激酶共同调节器官发育以及向重力性反应中生长素的运输(Cheng et al., 2008; Li et al., 2011)。通过分析NPYs基因对应的双突变体、三突变体、四突变体以及五突变体根的向重力性表型, 发现双突变体和三突变体向重力性表型缺失不明显; 四突变体表现不同程度的向重力性缺失表型, 其中npy1/npy2/npy4/npy5和npy2/npy3/npy4/npy5严重缺失向重力性; 五突变体的根几乎完全缺失向重力性, 在21°到150°之间随机生长(Li et al., 2011)。
作为植物进行光合作用的细胞器, 叶绿体响应外界光照刺激而发生的位置变化称为叶绿体光下重定位。弱光照射下, 为最大程度地吸收光能, 叶绿体沿细胞平周壁方向聚集, 这种现象为聚光运动(accumulation movement)。强光条件下, 为避免过强的光对自身造成伤害, 叶绿体沿细胞垂周壁聚集, 使之与光平行分布, 这种现象称为避光运动(avoidance movement) (Kasahara et al., 2004)。叶绿体运动受蓝光调节, 蓝光受体向光素PHOT1和PHOT2共同调控叶绿体的聚光反应, 而PHOT2单独调节叶绿体的避光反应(Suetsugu and Wada, 2017)。与向光素调节的其它多细胞协调性反应(如向光性、叶片的伸展与定位)不同, 叶绿体运动是细胞自主性反应, 不受基因表达(Kong and Wada, 2016; Wada, 2016)和生长素运输的调节(Christie et al., 2018), 最终由叶绿体肌动蛋白响应入射光的强度与方向而发生快速重组介导(Kadota et al., 2009; Kong and Wada, 2014)。叶绿体肌动蛋白特异定位于细胞质膜和叶绿体之间, 在叶绿体前进方向的后面叶绿体肌动蛋白纤维迅速解聚, 前进方向的前面叶绿体肌动蛋白纤维迅速聚集, 这种不对称的叶绿体肌动蛋白纤维分布最终决定了叶绿体的运动方向和速度(Kong and Wada, 2016)。CHUP1 (CHLOROPLAST UNUSUAL POSITIONING 1)和
KAC (KINESIN-LIKE PROTEIN FOR ACTIN-BASED CHLOROPLAST MOVEMENT)参与调节叶绿体肌动蛋白纤维的聚集(Wada, 2013; Kong and Wada, 2014), WEB1 (WEAK CHLOROPLAST MOVEMENT UNDER BLUE LIGHT 1)和PMI1 (PLASTIDMOVEMENT IMPAIRED1)对叶绿体肌动蛋白纤维的稳定性至关重要(Suetsugu et al., 2015)。
虽然目前人们还不清楚向光素在感受外界光照后如何通过调节CHUP1、PMI1、WEB1以及JAC蛋白活性影响叶绿体的运动, 但越来越多的证据表明, NRL家族蛋白的NCH1和RPT2是该信号途径的重要中间因子(Suetsugu et al., 2016)。rpt2突变体rpt2-3和rpt2-4的聚光反应略微降低, 但是叶绿体的避光反应和暗恢复反应(撤掉强光后, 叶绿体快速恢复, 导致透光率迅速下降的反应称为暗恢复反应)与野生型相比无明显差异(Suetsugu et al., 2016), 这些结果表明RPT2参与调节叶绿体的聚光反应。nch1单突变体的叶绿体聚光反应减弱, 暗恢复反应缺失, 但是避光反应增强(Suetsugu et al., 2016), 证明NCH1参与调节叶绿体的聚光运动。目前已经证实RPT2和NCH1参与调节弱光下PHOT1和PHOT2介导的叶绿体聚光反应, 这种AGC-NRL调节模块不依赖于生长素的转运以及生长素介导的转录过程(Suetsugu et al., 2016), 可能依赖于JAC1 (J-DOMAIN PROTEIN REQUIRED FOR CHLOROPLAST ACCUMULATION RESPONSE 1)以及WEB1/PMI2介导的信号途径(Suetsugu and Wada, 2017)。弱光下, RPT2/ NCH1通过JAC1依赖与JAC1不依赖途径抑制弱光下的叶绿体避光反应。强光下, WEB1/PMI2复合体通过抑制RPT2/NCH1和JAC1介导的聚光反应, 有效促进PHOT2介导的叶绿体避光反应(Suetsugu and Wada, 2017), 但是同一家族的NPH3并不调节叶绿体的运动(Inada et al., 2004)。
AggarwalC, Bana?AK, Kasprowicz-Malu?kiA, BorghettiC, ?abuzJ, DobruckiJ, Gabry?H ( 2014). Blue-lightactivated phototropin 2 trafficking from the cytoplasm to Golgi/post-Golgi vesicles J Exp Bot 65, 3263-3276. [本文引用: 1]
AssmannSM, SimonciniL, SchroederJI ( 1985). Blue light activates electrogenic ion pumping in guard cell protoplasts of Vicia faba Nature 318, 285-287. [本文引用: 1]
BlakesleeJJ, BandyopadhyayA, PeerWA, MakamSN, MurphyAS ( 2004). Relocalization of the PIN1 auxin efflux facilitator plays a role in phototropic responses Plant Physiol 134, 28-31. [本文引用: 1]
Boex-FontvieilleE, DavantureM, JossierM, ZivyM, HodgesM, TcherkezG ( 2014). Photosynthetic activity influences cellulose biosynthesis and phosphorylation of proteins involved therein in Arabidopsis leaves J Exp Bot 65, 4997-5010. [本文引用: 2]
ChristieJM, MurphyAS ( 2013). Shoot phototropism in higher plants: new light through old concepts Am J Bot 100, 35-46. [本文引用: 2]
ChristieJM, ReymondP, PowellGK, BernasconiP, RaibekasAA, LiscumE, BriggsWR ( 1998). Arabidopsis NPH1: a flavoprotein with the properties of a photoreceptor for phototropism Science 282, 1698-1701. [本文引用: 1]
ChristieJM, SuetsuguN, SullivanS, WadaM ( 2018). Shining light on the function of NPH3/RPT2-like proteins in phototropin signaling Plant Physiol 176, 1015-1024. [本文引用: 9]
ChristieJM, YangHB, RichterGL, SullivanS, ThomsonCE, LinJS, TitapiwatanakunB, EnnisM, KaiserliE, LeeOR, AdamecJ, PeerWA, MurphyAS ( 2011). phot1 inhibition of ABCB19 primes lateral auxin fluxes in the shoot apex required for phototropism PLoS Biol 9, e1001076. [本文引用: 5]
de CarbonnelM, DavisP, RoelfsemaMR, InoueS, SchepensI, LariguetP, GeislerM, ShimazakiK, HangarterR, FankhauserC ( 2010). The Arabidopsis PHYTOCHROME KINASE SUBSTRATE 2 protein is a phototropin signaling element that regulates leaf flattening and leaf positioning Plant Physiol 152, 1391-1405. [本文引用: 5]
DemarsyE, FankhauserC ( 2009). Higher plants use LOV to perceive blue light Curr Opin Plant Biol 12, 69-74. [本文引用: 1]
DemarsyE, SchepensI, OkajimaK, HerschM, BergmannS, ChristieJ, ShimazakiK, TokutomiS, FankhauserC ( 2012). Phytochrome Kinase Substrate 4 is phosphorylated by the phototropin 1 photoreceptor EMBO J 31, 3457-3467. [本文引用: 2]
DengZP, Oses-PrietoJA, KutscheraU, TsengTS, HaoLZ, BurlingameAL, WangZY, BriggsWR ( 2014). Blue light-induced proteomic changes in etiolated Arabidopsis seedlings J Proteome Res 13, 2524-2533. [本文引用: 1]
DingZJ, Galván-AmpudiaCS, DemarsyE, ?angowskiL, Kleine-VehnJ, FanYW, MoritaMT, TasakaM, FankhauserC, OffringaR, FrimlJ ( 2011). Light-mediated polarization of the PIN3 auxin transporter for the phototropic response in Arabidopsis Nat Cell Biol 13, 447-452. [本文引用: 4]
FigueroaP, GusmaroliG, SerinoG, HabashiJ, MaLG, ShenYP, FengSH, BostickM, CallisJ, HellmannH, DengXW ( 2005). Arabidopsis has two redundant Cullin3 proteins that are essential for embryo development and that interact with RBX1 and BTB proteins to form multisubunit E3 ubiquitin ligase complexes in vivo Plant Cell 17, 1180-1195. [本文引用: 1]
FoltaKM, KaufmanLS ( 2003). Phototropin 1 is required for high-fluence blue-light-mediated mRNA destabilization Plant Mol Biol 51, 609-618. [本文引用: 1]
FoltaKM, LiegEJ, DurhamT, SpaldingEP ( 2003). Primary inhibition of hypocotyl growth and phototropism depend differently on phototropin-mediated increases in cytoplasmic calcium induced by blue light Plant Physiol 133, 1464-1470. [本文引用: 1]
GingerichDJ, GagneJM, SalterDW, HellmannH, EstelleM, MaLG, VierstraRD ( 2005). Cullins 3a and 3b assemble with members of the broad complex/tramtrack/bric-a- brac (BTB) protein family to form essential ubiquitin-protein ligases (E3s) in Arabidopsis J Biol Chem 280, 18810-18821. [本文引用: 1]
HagaK, Tsuchida-MayamaT, YamadaM, SakaiT ( 2015). Arabidopsis ROOT PHOTOTROPISM2 contributes to the adaptation to high-intensity light in phototropic responses Plant Cell 27, 1098-1112. [本文引用: 7]
HaradaA, TakemiyaA, InoueSI, SakaiT, ShimazakiK ( 2013). Role of RPT2 in leaf positioning and flattening and a possible inhibition of phot2 signaling by phot1 Plant Cell Physiol 54, 36-47. [本文引用: 9]
HetheringtonAM, WoodwardFI ( 2003). The role of stomata in sensing and driving environmental change Nature 424, 901-908. [本文引用: 1]
HiyamaA, TakemiyaA, MunemasaS, OkumaE, SugiyamaN, TadaY, MurataY, ShimazakiKI ( 2017). Blue light and CO2 signals converge to regulate light-induced stomatal opening Nat Commun 8, 1284. [本文引用: 1]
InadaS, OhgishiM, MayamaT, OkadaK, SakaiT ( 2004). RPT2 is a signal transducer involved in phototropic response and stomatal opening by association with phototropin 1 in Arabidopsis thaliana Plant Cell 16, 887-896. [本文引用: 11]
InoueSI, KinoshitaT ( 2017). Blue light regulation of stomatal opening and the plasma membrane H+-ATPase Plant Physiol 174, 531-538. [本文引用: 2]
InoueSI, KinoshitaT, MatsumotoM, NakayamaKI, DoiM, ShimazakiK (2008a). Blue light-induced autophosphorylation of phototropin is a primary step for signaling Proc Natl Acad Sci USA 105, 5626-5631. [本文引用: 1]
InoueSI, KinoshitaT, TakemiyaA, DoiM, ShimazakiK (2008b). Leaf positioning of Arabidopsis in response to blue light Mol Plant 1, 15-26. [本文引用: 2]
InoueSI, MatsushitaT, TomokiyoY, MatsumotoM, NakayamaKI, KinoshitaT, ShimazakiK ( 2011). Functional analyses of the activation loop of phototropin2 in Arabidopsis Plant Physiol 156, 117-128. [本文引用: 2]
JeongRD, Chandra-ShekaraAC, BarmanSR, NavarreD, KlessigDF, KachrooA, KachrooP ( 2010). Cryptochrome 2 and phototropin 2 regulate resistance protein-mediated viral defense by negatively regulating an E3 ubiquitin ligase Proc Natl Acad Sci USA 107, 13538-13543. [本文引用: 1]
KadotaA, YamadaN, SuetsuguN, HiroseM, SaitoC, ShodaK, IchikawaS, KagawaT, NakanoA, WadaM ( 2009). Short actin-based mechanism for light-directed chloroplast movement in Arabidopsis Proc Natl Acad Sci USA 106, 13106-13111. [本文引用: 1]
KinoshitaT, DoiM, SuetsuguN, KagawaT, WadaM, ShimazakiK ( 2001) Phot1 and phot2 mediate blue light regulation of stomatal opening Nature 414, 656-660. [本文引用: 3]
KongSG, WadaM ( 2014). Recent advances in understanding the molecular mechanism of chloroplast photorelocation movement Biochim Biophys Acta 1837, 522-530. [本文引用: 2]
KongSG, WadaM ( 2016). Molecular basis of chloroplast photorelocation movement J Plant Res 129, 159-166. [本文引用: 2]
LalanneE, HonysD, JohnsonA, BornerGHH, LilleyKS, DupreeP, GrossniklausU, TwellD ( 2004). SETH1 and SETH2, two components of the glycosylphosphatidylinositol anchor biosynthetic pathway, are required for pollen germination and tube growth in Arabidopsis Plant Cell 16, 229-240. [本文引用: 1]
LariguetP, SchepensI, HodgsonD, PedmaleUV, TrevisanM, KamiC, de CarbonnelM, AlonsoJM, EckerJR, LiscumE, FankhauserC ( 2006). PHYTOCHROME KINASE SUBSTRATE 1 is a phototropin 1 binding protein required for phototropism Proc Natl Acad Sci USA 103, 10134-10139. [本文引用: 1]
LiYT, DaiXH, ChengYF, ZhaoYD ( 2011). NPY genes play an essential role in root gravitropic responses in Arabidopsis Mol Plant 4, 171-179. [本文引用: 2]
LiscumE, AskinosieSK, LeuchtmanDL, MorrowJ, WillenburgKT, CoatsDR ( 2014). Phototropism: growing towards an understanding of plant movement Plant Cell 26, 38-55. [本文引用: 2]
LiscumE, BriggsWR ( 1995). Mutations in the NPH1 locus of Arabidopsis disrupt the perception of phototropic stimuli Plant Cell 7, 473-485. [本文引用: 1]
LiscumE, BriggsWR ( 1996). Mutations of Arabidopsis in potential transduction and response components of the phototropic signaling pathway Plant Physiol 112, 291-296. [本文引用: 1]
MartenI, DeekenR, HedrichR, RoelfsemaMR ( 2010). Light-induced modification of plant plasma membrane ion transport Plant Biol 12, 64-79. [本文引用: 1]
MoniA, LeeAY, BriggsWR, HanIS ( 2015). The blue light receptor Phototropin 1 suppresses lateral root growth by controlling cell elongation Plant Biol 17, 34-40. [本文引用: 1]
MorrowJ, WillenburgKT, LiscumE ( 2018). Phototropism in land plants: molecules and mechanism from light perception to response Front Biol 5, 342-357. [本文引用: 1]
MotchoulskiA, LiscumE ( 1999). Arabidopsis NPH3: a NPH1 photoreceptor-linteracting protein essential for phototropism Science 286, 961-964. [本文引用: 4]
MottKA, SibbernsenED, ShopeJC ( 2008). The role of the mesophyll in stomatal responses to light and CO2 Plant Cell Environ 31, 1299-1306. [本文引用: 1]
PedmaleUV, CelayaRB, LiscumE ( 2010). Phototropism: mechanism and outcomes Arabidopsis Book 8, e0125. [本文引用: 1]
PedmaleUV, LiscumE ( 2007). Regulation of phototropic signaling in Arabidopsis via phosphorylation state changes in the phototropin 1-interacting protein NPH3 J Biol Chem 282, 19992-20001. [本文引用: 2]
PetrickaJJ, ClayNK, NelsonTM ( 2008). Vein patterning screens and the defectively organized tributaries mutants in Arabidopsis thaliana Plant J 56, 251-263. [本文引用: 1]
PfeiferA, MathesT, LuYH, HegemannP, KottkeT ( 2010). Blue light induces global and localized conformational changes in the kinase domain of full-length phototropin Biochemistry 49, 1024-1032. [本文引用: 1]
RobertsD, PedmaleUV, MorrowJ, SachdevS, LechnerE, TangXB, ZhengN, HanninkM, GenschikP, LiscumE ( 2011). Modulation of phototropic responsiveness in Arabidopsis through ubiquitination of phototropin 1 by the CUL3-ring E3 ubiquitin ligase CRL3 NPH3. Plant Cell 23, 3627-3640. [本文引用: 4]
RoelfsemaMRG, HedrichR ( 2005). In the light of stomatal opening: new insights into 'the Watergate' New Phytol 167, 665-691. [本文引用: 1]
SakaiT, KagawaT, KasaharaM, SwartzTE, ChristieJM, BriggsWR, WadaM, OkadaK ( 2001). Arabidopsis nph1 and npl1: blue light receptors that mediate both phototropism and chloroplast relocation Proc Natl Acad Sci USA 98, 6969-6974. [本文引用: 4]
SakaiT, WadaT, IshiguroS, OkadaK ( 2000). RPT2: a signal transducer of the phototropic response in Arabidopsis Plant Cell 12, 225-236. [本文引用: 2]
SakamotoK, BriggsWR ( 2002). Cellular and subcellular localization of phototropin 1 Plant Cell 14, 1723-1735. [本文引用: 1]
SalomonM, KniebE, von ZeppelinT, RüdigerW ( 2003). Mapping of low- and high-fluence autophosphorylation sites in phototropin 1 Biochemistry 42, 4217-4225. [本文引用: 1]
ShangBS, ZangYH, ZhaoX, ZhuJD, FanC, GuoXN, ZhangX ( 2019). Functional characterization of GhPHOT2 in chloroplast avoidance of Gossypium hirsutum Plant Physiol Biochem 135, 51-60. [本文引用: 1]
ShimazakiK, DoiM, AssmannSM, KinoshitaT, ( 2007). Light regulation of stomatal movement Annu Rev Plant Biol 58, 219-247. [本文引用: 2]
StoneBB, Stowe-EvansEL, HarperRM, CelayaRB, LjungK, SandbergG, LiscumE ( 2008). Disruptions in AUX1-dependent auxin influx alter hypocotyl phototropism in Arabidopsis Mol Plant 1, 129-144. [本文引用: 1]
SuetsuguN, HigaT, KongSG, WadaM ( 2015). PLASTID MOVEMENT IMPAIRED 1 and PLASTID MOVEMENT IMPAIRED1-RELATED 1 mediate photorelocation movements of both chloroplasts and nuclei Plant Physiol 169, 1155-1167. [本文引用: 1]
SuetsuguN, TakamiT, EbisuY, WatanabeH, IiboshiC, DoiM, ShimazakiK ( 2014). Guard cell chloroplasts are essential for blue light-dependent stomatal opening in Arabidopsis PLoS One 9, e108374. [本文引用: 1]
SuetsuguN, TakemiyaA, KongSG, HigaT, KomatsuA, ShimazakiK, KohchiT, WadaM ( 2016). RPT2/NCH1 subfamily of NPH3-like proteins is essential for the chloroplast accumulation response in land plants Proc Natl Acad Sci USA 113, 10424-10429. [本文引用: 17]
SuetsuguN, WadaM ( 2017). Two coiled-coil proteins, WEB1 and PMI2, suppress the signaling pathway of chloroplast accumulation response that is mediated by two phototropin-interacting proteins, RPT2 and NCH1, in seed plants Int J Mol Sci 18, 1469. [本文引用: 4]
SullivanS, ThomsonCE, KaiserliE, ChristieJM ( 2009). Interaction specificity of Arabidopsis 14-3-3 proteins with phototropin receptor kinases FEBS Lett 583, 2187-2193. [本文引用: 2]
SunJQ, QiLL, LiYN, ZhaiQZ, LiCY ( 2013). PIF4 and PIF5 transcription factors link blue light and auxin to regulate the phototropic response in Arabidopsis Plant Cell 25, 2102-2114. [本文引用: 1]
TakemiyaA, ShimazakiK ( 2016). Arabidopsis phot1 and phot2 phosphorylate BLUS1 kinase with different efficiencies in stomatal opening J Plant Res 129, 167-174. [本文引用: 1]
TsengTS, BriggsWR ( 2010). The Arabidopsis rcn1-1 mutation impairs dephosphorylation of Phot2, resulting in enhanced blue light responses Plant Cell 22, 392-402. [本文引用: 1]
Tsuchida-MayamaT, NakanoM, UeharaY, SanoM, FujisawaN, OkadaK, SakaiT ( 2008). Mapping of the phosphorylation sites on the phototropic signal transducer, NPH3 Plant Sci 174, 626-633. [本文引用: 1]
Tsuchida-MayamaT, SakaiT, HanadaA, UeharaY, AsamiT, YamaguchiS ( 2010). Role of the phytochrome and cryptochrome signaling pathways in hypocotyl phototropism Plant J 62, 653-662. [本文引用: 3]
TsutsumiT, TakemiyaA, HaradaA, ShimazakiK ( 2013). Disruption of ROOT PHOTOTROPISM 2 gene does not affect phototropin-mediated stomatal opening Plant Sci 201-202, 93-97. [本文引用: 4]
WadaM ( 2016). Chloroplast and nuclear photorelocation movements Proc Japan Acad Ser B 92, 387-411. [本文引用: 1]
WanYL, EisingerW, EhrhardtD, KubitscheckU, BaluskaF, BriggsW ( 2008). The subcellular localization and blue-light-induced movement of phototropin 1-GFP in etiolated seedlings of Arabidopsis thaliana Mol Plant 1, 103-117. [本文引用: 1]
WanYL, JasikJ, WangL, HaoHQ, VolkmannD, MenzelD, MancusoS, Balu?kaF, LinJX ( 2012). The signal transducer NPH3 integrates the phototropin1 photosensor with PIN2-based polar auxin transport in Arabidopsis root phototropism Plant Cell 24, 551-565. [本文引用: 3]
WilligeBC, AhlersS, ZourelidouM, BarbosaICR, DemarsyE, TrevisanM, DavisPA, RoelfsemaMRG, HangarterR, FankhauserC, SchwechheimerC ( 2013). D6PK AGCVIII kinases are required for auxin transport and phototropic hypocotyl bending in Arabidopsis Plant Cell 25, 1674-1688. [本文引用: 2]
YamauchiS, TakemiyaA, SakamotoT, KurataT, TsutsumiT, KinoshitaT, ShimazakiK ( 2016). The plasma membrane H+-ATPase AHA1 plays a major role in stomatal opening in response to blue light Plant Physiol 171, 2731-2743. [本文引用: 1]
... 早在达尔文(1880年)时期, 人们就已发现植物向光性, 但是直到向光素PHOT1和PHOT2先后被鉴定(Liscum and Briggs, 1995; Christie et al., 1998), 植物的向光性机理研究才取得突飞猛进的发展.研究发现, PHOT1调节弱蓝光(0.01-1 μmol·m-2·s-1)和强蓝光(>1 μmol·m-2·s-1)诱导的下胚轴的向光性; PHOT2仅在强蓝光下起作用, 且与PHOT1功能冗余(Sakai et al., 2001; Briggs and Christie, 2002).由于强蓝光下PHOT1和PHOT2的功能冗余, 目前向光性机制研究主要集中在PHOT1介导的弱蓝光反应(Motchoulski and Liscum, 1999; Haga et al., 2015), 仅有少量研究通过突变体背景下的抑制子筛选寻找参与强蓝光信号途径的作用因子(赵翔等, 2014).单侧蓝光照射下, 自磷酸化激活的向光素蛋白将信号传递给NPH3、RPT2以及PKS等下游因子(Motchoulski and Liscum, 1999; Inada et al., 2004), 调节生长素运输载体的活性与定位(Ding et al., 2011).例如, 引起内皮层细胞背光侧PIN3 (PIN-FORMED3)积累(刘广超和丁兆军, 2018), 促进背光侧生长素含量增加、细胞伸长速率加快, 最终引起植物弯向光生长(Christie and Murphy, 2013).下胚轴弯曲过程中生长素的不对称分布受各类生长素运输载体的共同调控, 如PINs (PIN3、PIN4和PIN7)和ABCB19 (Christie et al., 2011).蓝光下, PIN3的磷酸化受抑制, 促进PIN3的背光侧定位, 诱导背光侧生长素积累以及下胚轴的向光性生长(Ding et al., 2011).PIN3的磷酸化以及极性定位受AGC激酶VIII亚族成员PID (PINOID)以及D6PK (D6 PROTEIN KINASE)的调节(Ding et al., 2011; Willige et al., 2013).ABCB19是生长素外流载体, 被蓝光激活的PHOT1磷酸化后, ABCB19的生长素转运活性受抑制, 增加拟南芥下胚轴顶端生长素含量并抑制其垂直生长, 同时通过PIN3蛋白促进生长素在下胚轴中侧向移动(Christie et al., 2011).背光侧生长素的大量积累促进了背光侧IAA19的降解, 释放出有活性的ARF7 (Sun et al., 2013), 不对称激活与生长相关基因的表达促进了下胚轴的向光性反应(刘广超和丁兆军, 2018). ...
向光素调节植物向光性及其与光敏色素/隐花色素的相互关系 1 2015
... 光敏色素(phytochrome, phy)和隐花色素(cryptochrome, cry)通过影响RPT2的表达调节向光性反应(Tsuchida-Mayama et al., 2010; 赵翔等, 2015; Zhao et al., 2019).强蓝光处理3小时显著诱导拟南芥野生型中RPT2基因的表达, PHYA、CRY1或CRY2基因突变可部分抑制蓝光诱导的RPT2表达, 强蓝光处理几乎不能诱导phya/cry1/cry2三突变体中RPT2表达(Tsuchida-Mayama et al., 2010).蓝光处理延长至24小时后, phya/cry1/cry2三突变体中RPT2的基因表达被轻微诱导, 且下胚轴能发生轻微的弯曲, 而phya/phyb/cry1/cry2四突变体中几乎检测不到RPT2蛋白的表达, 其下胚轴缺失向光弯曲反应(Tsuchida-Mayama et al., 2010).上述结果表明, 蓝光诱导RPT2的表达需要PHYs或者CRYs参与, PHYs和CRYs通过调节RPT2的表达参与调节蓝光诱导的下胚轴向光弯曲反应. ...
Blue-lightactivated phototropin 2 trafficking from the cytoplasm to Golgi/post-Golgi vesicles 1 2014
... 蓝光照射会诱导PHOT1和PHOT2亚细胞定位改变, PHOT1从细胞质膜脱离后向胞质移动(Kaiserli et al., 2009), 而PHOT2从细胞质膜脱离后向高尔基体转移, 最终与高尔基体偶联(Aggarwal et al., 2014).虽然向光素蛋白亚细胞定位受光照影响的生物学意义还不清楚, 但已知其亚细胞定位变化受PHOT1和PHOT2的激酶活性调节, 且蓝光诱导PHOT1的胞内定位变化(intracelluar position)参与下胚轴向光性调节(Wan et al., 2008; Zhao et al., 2018), 暗示向光素的亚细胞定位变化影响了向光素途径的信号传递, PHOT1和PHOT2亚细胞定位变化的不同可能是2种向光素受体差异调节植物蓝光反应的重要原因. ...
Blue light activates electrogenic ion pumping in guard cell protoplasts of Vicia faba 1 1985
... 拟南芥气孔由1对保卫细胞构成, 气孔通过调节其开闭程度控制着与环境的气体交换来响应外界的刺激信号(Hetherington and Woodward, 2003; Roelfsema and Hedrich, 2005).红光和蓝光都能诱导气孔快速张开(Shimazaki et al., 2007).红光通过影响叶肉细胞和保卫细胞中叶绿体的光合作用调节气孔开放(Mott et al., 2008; Suetsugu et al., 2014).蓝光作为信号通过激活保卫细胞质膜H+-ATPase诱导气孔开放(Assmann et al., 1985; Kinoshita et al., 2001).蓝光诱导的气孔开放受蓝光受体向光素(PHOT1和PHOT2)调节(Kinoshita et al., 2001).目前人们对向光素介导的气孔开放机制研究较为清楚(Inoue et al., 2008b; Inoue and Kinoshita, 2017).蓝光照射下, 自磷酸化激活的PHOT1直接磷酸化蛋白激酶BLUS1 (LIGHT SIGNALING1), 磷酸化的BLUS1间接将信号传递给I型蛋白磷酸酶(PPI)以及该酶的亚基PRSL1 (Takemiya and Shimazaki, 2016).PPI进一步将信号传递给质膜AHA1型H+-ATPase, 促进H+-ATPase的激活.H+-ATPase的激活需要其C末端倒数第2个苏氨酸的磷酸化以及与14-3-3蛋白的结合(Yamauchi et al., 2016).活化的H+-ATPase通过驱动保卫细胞内H+的跨膜输出诱导保卫细胞质膜的超极化(Shimazaki et al., 2007; Marten et al., 2010).超极化的质膜激活内向K+通道, 促进K+在保卫细胞胞质中积累.为了维持胞质的电荷平衡, Cl-和malate2-随之大量增加, 增加的K+、Cl-和malate2-进一步被转入液泡, 使保卫细胞水势降低, 外界水分大量进入保卫细胞, 促进保卫细胞体积膨胀, 气孔张开(Inoue and Kinoshita, 2017).保卫细胞胞质中Cl-的增加主要是由于向光素抑制了S型阴离子外流蛋白的活性(Sussmilch et al., 2019).向光素对S型阴离子外流蛋白活性的抑制需要2个蛋白激酶CBC1/2 (CONVERGENCE OF BLUE LIGHT (BL) and CO21/2)参与(Hiyama et al., 2017; Sussmilch et al., 2019). ...
Relocalization of the PIN1 auxin efflux facilitator plays a role in phototropic responses 1 2004
... 拟南芥中ABCB19 (ATP-BINDING CAS-SETTE B 19)和光敏色素激酶底物(PHYTOCHROME KINASE SUBSTRATE 4, PKS4)参与调节生长素的转运, 是下胚轴向光弯曲的负调节因子(Christie et al., 2011; Demarsy et al., 2012).自磷酸化激活的PHOT1通过直接磷酸化ABCB19和PKS4, 抑制ABCB19和PKS4的生长素转运活性, 促进下胚轴向光弯曲反应(Christie et al., 2011; Demarsy et al., 2012).PHOT1通过磷酸化修饰调节ABCB19的活性, 但是PHOT2并不影响ABCB19的活性(Christie et al., 2011).蛋白磷酸酶2A (PP2A)的A1亚基对应的蛋白RCN1 (Root Curling in N-naphthylphthalamic acid 1)与PHOT2蛋白发生体外互作, 负调节PHOT2介导的向光弯曲, 但RCN1并不与PHOT1直接相互作用(Tseng and Briggs, 2010).NPH3 (NONPHOTOTROPIC HYPOCOTYL 3)和RPT2 (ROOT PHOTOTROPISM 2)是通过筛选拟南芥下胚轴以及根的向光性缺失突变体获得(Motchoulski and Liscum, 1999; Sakai et al., 2000).随后的研究发现, NPH3和RPT2是向光素途径的重要调控因子, 均可与PHOT1蛋白直接相互作用(Blakeslee et al., 2004; Stone et al., 2008).虽然NPH3和RPT2也参与PHOT2信号转导途径, 但目前并无RPT2与PHOT2蛋白之间直接相互作用的报道(Inada et al., 2004; Lariguet et al., 2006).上述结果暗示PHOT1和PHOT2可能通过与不同的下游互作因子差异调节蓝光反应. ...
Photosynthetic activity influences cellulose biosynthesis and phosphorylation of proteins involved therein in Arabidopsis leaves 2 2014
... 拟南芥PHOT1至少有21个氨基酸位点可以发生自磷酸化(Boex-Fontvieille et al., 2014; Deng et al., 2014), PHOT2有29个磷酸化位点(Inoue et al., 2011; Boex-Fontvieille et al., 2014), 这些位点的具体分布参见Christie等(2015)的报道.其中, C端2个保守位点的磷酸化(PHOT1中S849和S851, PHOT2中S761 和S763)对向光素受体的信号传递至关重要(Inoue et al., 2008a; Inoue et al., 2011), 其余的磷酸化位点大部分位于N端LOV1的上游以及LOV1与LOV2中间的连接区域(Christie et al., 2015).虽然这些上游区域磷酸化位点的生物学意义目前还不清楚, 但这些位点的磷酸化程度受光照强度调节(Salomon et al., 2003), 暗示PHOT1和PHOT2可能通过磷酸化程度差异响应不同的光照强度, 从而参与植物多种生理活动的调节. ...
... ; Boex-Fontvieille et al., 2014), 这些位点的具体分布参见Christie等(2015)的报道.其中, C端2个保守位点的磷酸化(PHOT1中S849和S851, PHOT2中S761 和S763)对向光素受体的信号传递至关重要(Inoue et al., 2008a; Inoue et al., 2011), 其余的磷酸化位点大部分位于N端LOV1的上游以及LOV1与LOV2中间的连接区域(Christie et al., 2015).虽然这些上游区域磷酸化位点的生物学意义目前还不清楚, 但这些位点的磷酸化程度受光照强度调节(Salomon et al., 2003), 暗示PHOT1和PHOT2可能通过磷酸化程度差异响应不同的光照强度, 从而参与植物多种生理活动的调节. ...
Phototropins 1 and 2: versatile plant blue-light receptors 1 2002
... 早在达尔文(1880年)时期, 人们就已发现植物向光性, 但是直到向光素PHOT1和PHOT2先后被鉴定(Liscum and Briggs, 1995; Christie et al., 1998), 植物的向光性机理研究才取得突飞猛进的发展.研究发现, PHOT1调节弱蓝光(0.01-1 μmol·m-2·s-1)和强蓝光(>1 μmol·m-2·s-1)诱导的下胚轴的向光性; PHOT2仅在强蓝光下起作用, 且与PHOT1功能冗余(Sakai et al., 2001; Briggs and Christie, 2002).由于强蓝光下PHOT1和PHOT2的功能冗余, 目前向光性机制研究主要集中在PHOT1介导的弱蓝光反应(Motchoulski and Liscum, 1999; Haga et al., 2015), 仅有少量研究通过突变体背景下的抑制子筛选寻找参与强蓝光信号途径的作用因子(赵翔等, 2014).单侧蓝光照射下, 自磷酸化激活的向光素蛋白将信号传递给NPH3、RPT2以及PKS等下游因子(Motchoulski and Liscum, 1999; Inada et al., 2004), 调节生长素运输载体的活性与定位(Ding et al., 2011).例如, 引起内皮层细胞背光侧PIN3 (PIN-FORMED3)积累(刘广超和丁兆军, 2018), 促进背光侧生长素含量增加、细胞伸长速率加快, 最终引起植物弯向光生长(Christie and Murphy, 2013).下胚轴弯曲过程中生长素的不对称分布受各类生长素运输载体的共同调控, 如PINs (PIN3、PIN4和PIN7)和ABCB19 (Christie et al., 2011).蓝光下, PIN3的磷酸化受抑制, 促进PIN3的背光侧定位, 诱导背光侧生长素积累以及下胚轴的向光性生长(Ding et al., 2011).PIN3的磷酸化以及极性定位受AGC激酶VIII亚族成员PID (PINOID)以及D6PK (D6 PROTEIN KINASE)的调节(Ding et al., 2011; Willige et al., 2013).ABCB19是生长素外流载体, 被蓝光激活的PHOT1磷酸化后, ABCB19的生长素转运活性受抑制, 增加拟南芥下胚轴顶端生长素含量并抑制其垂直生长, 同时通过PIN3蛋白促进生长素在下胚轴中侧向移动(Christie et al., 2011).背光侧生长素的大量积累促进了背光侧IAA19的降解, 释放出有活性的ARF7 (Sun et al., 2013), 不对称激活与生长相关基因的表达促进了下胚轴的向光性反应(刘广超和丁兆军, 2018). ...
NPY genes and AGC kinases define two key steps in auxin-mediated organogenesis in Arabidopsis 1 2008
... NPH3和RPT2是来自2个不同分支的NRL家族成员, 通过与向光素相互作用调节蓝光诱导下胚轴向光性、叶绿体聚光运动、气孔开放以及叶片伸展与定位(Christie et al., 2015, 2018).NRL8是AGC激酶(cAMP依赖的蛋白激酶A, cGMP依赖的蛋白激酶G和磷脂依赖的蛋白激酶C)活性依赖的花粉管生长调节因子(Lalanne et al., 2004; Zhang et al., 2008), 该蛋白与RPT2来自分支II.与NPH3类似, SR1IP1 (AtSR1 INTERACTING PROTEIN 1)也称NRL31, 可以与CRL3 ( CUL3 RING E3 UBIQUITIN LIGASE)形成复合体, 调节底物的泛素化(Zhang et al., 2014).SR1IP1通过调节拟南芥中Ca2+/钙调素结合的转录因子AtSR1 (SIGNAL RESPONSIVE 1)的泛素化降解, 增强植物对细菌性病原体的抗性(Zhang et al., 2014).但系统进化研究发现, SR1IP1也来自分支II, 且比NRL8与RPT2亲缘关系更近, 与NPH3的进化明显不同.Suetsugu等(2016)在筛选参与调节叶绿体运动的NRL家族蛋白时, 发现NRL31基因突变抑制了光诱导的叶绿体运动, 且蛋白NRL31可以与PHOT1发生体内互作, 因此将其重命名为NCH1 (NRL PROTEIN FORCHLOROPLASTMOVEMENT 1).DEFECTIVELY ORGANIZED TRIBUTARIES 3/NRL23是与NPH3来自同一个分支的NRL家族蛋白, 主要参与维管组织发育以及叶脉结构调节(Petricka et al., 2008).虽然NRL23与NPH3的亲缘关系较近, 但是该蛋白是否在向光素介导的信号途径中发挥作用尚未见报道.NRL5、NRL6、NRL7、NRL15、NRL20、NRL21、NRL26和NRL30是分支III中的NPY (NAKED PINS IN YUCCA)家族成员, 其中NRL6/NPY2、NRL7/NPY4、NRL20/NPY1、NRL21/NPY5和NRL30/NPY3以功能冗余的方式与AGCVIII激酶共同调节器官发育以及向重力性反应中生长素的运输(Cheng et al., 2008; Li et al., 2011).通过分析NPYs基因对应的双突变体、三突变体、四突变体以及五突变体根的向重力性表型, 发现双突变体和三突变体向重力性表型缺失不明显; 四突变体表现不同程度的向重力性缺失表型, 其中npy1/npy2/npy4/npy5和npy2/npy3/npy4/npy5严重缺失向重力性; 五突变体的根几乎完全缺失向重力性, 在21°到150°之间随机生长(Li et al., 2011). ...
Plant flavoprotein photoreceptors 6 2015
... 植物向光素PHOT1 (PHOTOTROPIN1)和PHOT2是一类细胞质膜结合的Ser/Thr蛋白激酶, 其C端为激酶结构域, N端含有2个光、氧气和电压敏感的LOV (LIGHT, OXYGEN, OR VOLTAGE SENSING)结构域, 分别命名为LOV1和LOV2 (Christie et al., 2015).蓝光照射后诱导向光素蛋白发生构象改变及自我磷酸化激活(Pfeifer et al., 2010), 活化的向光素蛋白进而调节下胚轴向光弯曲反应(Sakai et al., 2001)、叶绿体运动(Kagawa et al., 2001)、气孔开放(Kinoshita et al., 2001)以及叶片的伸展与定位等(Sakamoto and Briggs, 2002; Inoue et al., 2008b), 从而使植物或叶片处于最有利于吸收光的位置, 优化植物的光合作用(Christie et al., 2015). ...
... ), 从而使植物或叶片处于最有利于吸收光的位置, 优化植物的光合作用(Christie et al., 2015). ...
... 拟南芥(Arabidopsis thaliana) PHOT1和PHOT2以功能冗余的方式调节下胚轴向光性和根负向光性、叶绿体聚光运动、气孔开放以及叶片伸展与定位等(Christie et al., 2015).PHOT1特异性抑制光下黄化幼苗下胚轴的快速伸长(Folta et al., 2003)、子叶变绿过程中叶绿素结合蛋白mRNA转录的稳定性(Folta and Kaufman, 2003)以及侧根的快速生长(Moni et al., 2015).PHOT2不仅介导强蓝光诱导的下胚轴向光弯曲(Sakai et al., 2001), 而且也特异调节叶绿体和细胞核的避光运动(Higa et al., 2014)以及抗性蛋白介导的病毒防御反应(Jeong et al., 2010).PHOT1和PHOT2的功能差异可能与磷酸化修饰位点、磷酸化程度、亚细胞定位变化以及下游相互作用蛋白种类不同有关(Zhao et al., 2013; Morrow et al., 2018; Liscum et al., 2020). ...
... 拟南芥PHOT1至少有21个氨基酸位点可以发生自磷酸化(Boex-Fontvieille et al., 2014; Deng et al., 2014), PHOT2有29个磷酸化位点(Inoue et al., 2011; Boex-Fontvieille et al., 2014), 这些位点的具体分布参见Christie等(2015)的报道.其中, C端2个保守位点的磷酸化(PHOT1中S849和S851, PHOT2中S761 和S763)对向光素受体的信号传递至关重要(Inoue et al., 2008a; Inoue et al., 2011), 其余的磷酸化位点大部分位于N端LOV1的上游以及LOV1与LOV2中间的连接区域(Christie et al., 2015).虽然这些上游区域磷酸化位点的生物学意义目前还不清楚, 但这些位点的磷酸化程度受光照强度调节(Salomon et al., 2003), 暗示PHOT1和PHOT2可能通过磷酸化程度差异响应不同的光照强度, 从而参与植物多种生理活动的调节. ...
... ), 其余的磷酸化位点大部分位于N端LOV1的上游以及LOV1与LOV2中间的连接区域(Christie et al., 2015).虽然这些上游区域磷酸化位点的生物学意义目前还不清楚, 但这些位点的磷酸化程度受光照强度调节(Salomon et al., 2003), 暗示PHOT1和PHOT2可能通过磷酸化程度差异响应不同的光照强度, 从而参与植物多种生理活动的调节. ...
... NPH3和RPT2是来自2个不同分支的NRL家族成员, 通过与向光素相互作用调节蓝光诱导下胚轴向光性、叶绿体聚光运动、气孔开放以及叶片伸展与定位(Christie et al., 2015, 2018).NRL8是AGC激酶(cAMP依赖的蛋白激酶A, cGMP依赖的蛋白激酶G和磷脂依赖的蛋白激酶C)活性依赖的花粉管生长调节因子(Lalanne et al., 2004; Zhang et al., 2008), 该蛋白与RPT2来自分支II.与NPH3类似, SR1IP1 (AtSR1 INTERACTING PROTEIN 1)也称NRL31, 可以与CRL3 ( CUL3 RING E3 UBIQUITIN LIGASE)形成复合体, 调节底物的泛素化(Zhang et al., 2014).SR1IP1通过调节拟南芥中Ca2+/钙调素结合的转录因子AtSR1 (SIGNAL RESPONSIVE 1)的泛素化降解, 增强植物对细菌性病原体的抗性(Zhang et al., 2014).但系统进化研究发现, SR1IP1也来自分支II, 且比NRL8与RPT2亲缘关系更近, 与NPH3的进化明显不同.Suetsugu等(2016)在筛选参与调节叶绿体运动的NRL家族蛋白时, 发现NRL31基因突变抑制了光诱导的叶绿体运动, 且蛋白NRL31可以与PHOT1发生体内互作, 因此将其重命名为NCH1 (NRL PROTEIN FORCHLOROPLASTMOVEMENT 1).DEFECTIVELY ORGANIZED TRIBUTARIES 3/NRL23是与NPH3来自同一个分支的NRL家族蛋白, 主要参与维管组织发育以及叶脉结构调节(Petricka et al., 2008).虽然NRL23与NPH3的亲缘关系较近, 但是该蛋白是否在向光素介导的信号途径中发挥作用尚未见报道.NRL5、NRL6、NRL7、NRL15、NRL20、NRL21、NRL26和NRL30是分支III中的NPY (NAKED PINS IN YUCCA)家族成员, 其中NRL6/NPY2、NRL7/NPY4、NRL20/NPY1、NRL21/NPY5和NRL30/NPY3以功能冗余的方式与AGCVIII激酶共同调节器官发育以及向重力性反应中生长素的运输(Cheng et al., 2008; Li et al., 2011).通过分析NPYs基因对应的双突变体、三突变体、四突变体以及五突变体根的向重力性表型, 发现双突变体和三突变体向重力性表型缺失不明显; 四突变体表现不同程度的向重力性缺失表型, 其中npy1/npy2/npy4/npy5和npy2/npy3/npy4/npy5严重缺失向重力性; 五突变体的根几乎完全缺失向重力性, 在21°到150°之间随机生长(Li et al., 2011). ...
Shoot phototropism in higher plants: new light through old concepts 2 2013
... 早在达尔文(1880年)时期, 人们就已发现植物向光性, 但是直到向光素PHOT1和PHOT2先后被鉴定(Liscum and Briggs, 1995; Christie et al., 1998), 植物的向光性机理研究才取得突飞猛进的发展.研究发现, PHOT1调节弱蓝光(0.01-1 μmol·m-2·s-1)和强蓝光(>1 μmol·m-2·s-1)诱导的下胚轴的向光性; PHOT2仅在强蓝光下起作用, 且与PHOT1功能冗余(Sakai et al., 2001; Briggs and Christie, 2002).由于强蓝光下PHOT1和PHOT2的功能冗余, 目前向光性机制研究主要集中在PHOT1介导的弱蓝光反应(Motchoulski and Liscum, 1999; Haga et al., 2015), 仅有少量研究通过突变体背景下的抑制子筛选寻找参与强蓝光信号途径的作用因子(赵翔等, 2014).单侧蓝光照射下, 自磷酸化激活的向光素蛋白将信号传递给NPH3、RPT2以及PKS等下游因子(Motchoulski and Liscum, 1999; Inada et al., 2004), 调节生长素运输载体的活性与定位(Ding et al., 2011).例如, 引起内皮层细胞背光侧PIN3 (PIN-FORMED3)积累(刘广超和丁兆军, 2018), 促进背光侧生长素含量增加、细胞伸长速率加快, 最终引起植物弯向光生长(Christie and Murphy, 2013).下胚轴弯曲过程中生长素的不对称分布受各类生长素运输载体的共同调控, 如PINs (PIN3、PIN4和PIN7)和ABCB19 (Christie et al., 2011).蓝光下, PIN3的磷酸化受抑制, 促进PIN3的背光侧定位, 诱导背光侧生长素积累以及下胚轴的向光性生长(Ding et al., 2011).PIN3的磷酸化以及极性定位受AGC激酶VIII亚族成员PID (PINOID)以及D6PK (D6 PROTEIN KINASE)的调节(Ding et al., 2011; Willige et al., 2013).ABCB19是生长素外流载体, 被蓝光激活的PHOT1磷酸化后, ABCB19的生长素转运活性受抑制, 增加拟南芥下胚轴顶端生长素含量并抑制其垂直生长, 同时通过PIN3蛋白促进生长素在下胚轴中侧向移动(Christie et al., 2011).背光侧生长素的大量积累促进了背光侧IAA19的降解, 释放出有活性的ARF7 (Sun et al., 2013), 不对称激活与生长相关基因的表达促进了下胚轴的向光性反应(刘广超和丁兆军, 2018). ...
... 拟南芥nph3突变体在任何强度的单侧蓝光下均表现缺失下胚轴向光性(Liscum and Briggs, 1996; Liscum et al., 2014; Fankhauser and Christie, 2015), 表明NPH3调节PHOT1和PHOT2介导蓝光诱导的下胚轴弯曲反应.黑暗条件下, 磷酸化的NPH3蛋白与PHOT1结合, 定位于细胞膜, 单侧弱蓝光照射下, NPH3迅速发生PHOT1依赖的去磷酸化反应, 从细胞膜上脱离(Haga et al., 2015; Zhao et al., 2018).目前磷酸化NPH3的激酶还不清楚, 但NPH3去磷酸化主要由I型蛋白磷酸酶介导(Pedmale and Liscum, 2007).去磷酸化后NPH3从细胞膜向胞质的转移降低了其介导向光弯曲反应的能力(Haga et al., 2015; Sullivan et al., 2019).随着光照强度的增加, 下胚轴的弯曲度呈强度依赖的钟形曲线(Christie and Murphy, 2013).随着光照时间的增加, RPT2蛋白含量增加, 与去磷酸化的NPH3结合形成RPT2-NPH3复合物并与PHOT1蛋白结合, 促进NPH3的磷酸化以及从胞质到细胞膜的重定位, 开启第二次向光弯曲(Haga et al., 2015; Zhao et al., 2018).虽然PHOT1- NPH3复合体在细胞膜的重定位对持续光照介导的二次向光弯曲至关重要(Haga et al., 2015; Christie et al., 2018; Zhao et al., 2018), 但其生物学意义还不清楚, 人们认为这种变化有可能对调节生长素的不对称分布有重要影响(Pedmale and Liscum, 2007; Haga et al., 2015). ...
Arabidopsis NPH1: a flavoprotein with the properties of a photoreceptor for phototropism 1 1998
... 早在达尔文(1880年)时期, 人们就已发现植物向光性, 但是直到向光素PHOT1和PHOT2先后被鉴定(Liscum and Briggs, 1995; Christie et al., 1998), 植物的向光性机理研究才取得突飞猛进的发展.研究发现, PHOT1调节弱蓝光(0.01-1 μmol·m-2·s-1)和强蓝光(>1 μmol·m-2·s-1)诱导的下胚轴的向光性; PHOT2仅在强蓝光下起作用, 且与PHOT1功能冗余(Sakai et al., 2001; Briggs and Christie, 2002).由于强蓝光下PHOT1和PHOT2的功能冗余, 目前向光性机制研究主要集中在PHOT1介导的弱蓝光反应(Motchoulski and Liscum, 1999; Haga et al., 2015), 仅有少量研究通过突变体背景下的抑制子筛选寻找参与强蓝光信号途径的作用因子(赵翔等, 2014).单侧蓝光照射下, 自磷酸化激活的向光素蛋白将信号传递给NPH3、RPT2以及PKS等下游因子(Motchoulski and Liscum, 1999; Inada et al., 2004), 调节生长素运输载体的活性与定位(Ding et al., 2011).例如, 引起内皮层细胞背光侧PIN3 (PIN-FORMED3)积累(刘广超和丁兆军, 2018), 促进背光侧生长素含量增加、细胞伸长速率加快, 最终引起植物弯向光生长(Christie and Murphy, 2013).下胚轴弯曲过程中生长素的不对称分布受各类生长素运输载体的共同调控, 如PINs (PIN3、PIN4和PIN7)和ABCB19 (Christie et al., 2011).蓝光下, PIN3的磷酸化受抑制, 促进PIN3的背光侧定位, 诱导背光侧生长素积累以及下胚轴的向光性生长(Ding et al., 2011).PIN3的磷酸化以及极性定位受AGC激酶VIII亚族成员PID (PINOID)以及D6PK (D6 PROTEIN KINASE)的调节(Ding et al., 2011; Willige et al., 2013).ABCB19是生长素外流载体, 被蓝光激活的PHOT1磷酸化后, ABCB19的生长素转运活性受抑制, 增加拟南芥下胚轴顶端生长素含量并抑制其垂直生长, 同时通过PIN3蛋白促进生长素在下胚轴中侧向移动(Christie et al., 2011).背光侧生长素的大量积累促进了背光侧IAA19的降解, 释放出有活性的ARF7 (Sun et al., 2013), 不对称激活与生长相关基因的表达促进了下胚轴的向光性反应(刘广超和丁兆军, 2018). ...
Shining light on the function of NPH3/RPT2-like proteins in phototropin signaling 9 2018
... NRL (NPH3/RPT2-Like)蛋白家族是植物特有的, 其成员含有3个保守结构域: N端的BTB (Bric àbrac, Tramtrack, and Broad complex)结构域、中间的NPH3结构域以及C端的螺旋-螺旋结构域(Liscum et al., 2014).其中, BTB结构域主要起E3连接酶的作用, 与CUL3发生相互作用(Gingerich et al., 2005; Figueroa et al., 2005), C端的螺旋-螺旋结构域主要介导蛋白之间的相互作用(Inada et al., 2004).拟南芥中33个NRL家族成员都含有NPH3结构域, 10个缺少C端的螺旋-螺旋结构域, 2个缺少N端的BTB结构域(Pedmale et al., 2010; Christie et al., 2018).根据NRL家族成员间的进化关系, 拟南芥中的33个NRL家族成员可分为6个分支(图1) (Suetsugu et al., 2016; Christie et al., 2018).目前, 来自2个分支的10个成员的功能已得到鉴定(de Carbonnel et al., 2010; Christie et al., 2018) (图1). ...
... ; Christie et al., 2018).目前, 来自2个分支的10个成员的功能已得到鉴定(de Carbonnel et al., 2010; Christie et al., 2018) (图1). ...
... Construction of this phylogenetic tree was based on Suetsugu et al. (2016) and Christie et al. (2018) with slight modifications. The colors and names of six major clades of NRL proteins in Arabidopsis were constructed using the Adobe illustrator software. ...
... NPH3和RPT2是来自2个不同分支的NRL家族成员, 通过与向光素相互作用调节蓝光诱导下胚轴向光性、叶绿体聚光运动、气孔开放以及叶片伸展与定位(Christie et al., 2015, 2018).NRL8是AGC激酶(cAMP依赖的蛋白激酶A, cGMP依赖的蛋白激酶G和磷脂依赖的蛋白激酶C)活性依赖的花粉管生长调节因子(Lalanne et al., 2004; Zhang et al., 2008), 该蛋白与RPT2来自分支II.与NPH3类似, SR1IP1 (AtSR1 INTERACTING PROTEIN 1)也称NRL31, 可以与CRL3 ( CUL3 RING E3 UBIQUITIN LIGASE)形成复合体, 调节底物的泛素化(Zhang et al., 2014).SR1IP1通过调节拟南芥中Ca2+/钙调素结合的转录因子AtSR1 (SIGNAL RESPONSIVE 1)的泛素化降解, 增强植物对细菌性病原体的抗性(Zhang et al., 2014).但系统进化研究发现, SR1IP1也来自分支II, 且比NRL8与RPT2亲缘关系更近, 与NPH3的进化明显不同.Suetsugu等(2016)在筛选参与调节叶绿体运动的NRL家族蛋白时, 发现NRL31基因突变抑制了光诱导的叶绿体运动, 且蛋白NRL31可以与PHOT1发生体内互作, 因此将其重命名为NCH1 (NRL PROTEIN FORCHLOROPLASTMOVEMENT 1).DEFECTIVELY ORGANIZED TRIBUTARIES 3/NRL23是与NPH3来自同一个分支的NRL家族蛋白, 主要参与维管组织发育以及叶脉结构调节(Petricka et al., 2008).虽然NRL23与NPH3的亲缘关系较近, 但是该蛋白是否在向光素介导的信号途径中发挥作用尚未见报道.NRL5、NRL6、NRL7、NRL15、NRL20、NRL21、NRL26和NRL30是分支III中的NPY (NAKED PINS IN YUCCA)家族成员, 其中NRL6/NPY2、NRL7/NPY4、NRL20/NPY1、NRL21/NPY5和NRL30/NPY3以功能冗余的方式与AGCVIII激酶共同调节器官发育以及向重力性反应中生长素的运输(Cheng et al., 2008; Li et al., 2011).通过分析NPYs基因对应的双突变体、三突变体、四突变体以及五突变体根的向重力性表型, 发现双突变体和三突变体向重力性表型缺失不明显; 四突变体表现不同程度的向重力性缺失表型, 其中npy1/npy2/npy4/npy5和npy2/npy3/npy4/npy5严重缺失向重力性; 五突变体的根几乎完全缺失向重力性, 在21°到150°之间随机生长(Li et al., 2011). ...
... 拟南芥nph3突变体在任何强度的单侧蓝光下均表现缺失下胚轴向光性(Liscum and Briggs, 1996; Liscum et al., 2014; Fankhauser and Christie, 2015), 表明NPH3调节PHOT1和PHOT2介导蓝光诱导的下胚轴弯曲反应.黑暗条件下, 磷酸化的NPH3蛋白与PHOT1结合, 定位于细胞膜, 单侧弱蓝光照射下, NPH3迅速发生PHOT1依赖的去磷酸化反应, 从细胞膜上脱离(Haga et al., 2015; Zhao et al., 2018).目前磷酸化NPH3的激酶还不清楚, 但NPH3去磷酸化主要由I型蛋白磷酸酶介导(Pedmale and Liscum, 2007).去磷酸化后NPH3从细胞膜向胞质的转移降低了其介导向光弯曲反应的能力(Haga et al., 2015; Sullivan et al., 2019).随着光照强度的增加, 下胚轴的弯曲度呈强度依赖的钟形曲线(Christie and Murphy, 2013).随着光照时间的增加, RPT2蛋白含量增加, 与去磷酸化的NPH3结合形成RPT2-NPH3复合物并与PHOT1蛋白结合, 促进NPH3的磷酸化以及从胞质到细胞膜的重定位, 开启第二次向光弯曲(Haga et al., 2015; Zhao et al., 2018).虽然PHOT1- NPH3复合体在细胞膜的重定位对持续光照介导的二次向光弯曲至关重要(Haga et al., 2015; Christie et al., 2018; Zhao et al., 2018), 但其生物学意义还不清楚, 人们认为这种变化有可能对调节生长素的不对称分布有重要影响(Pedmale and Liscum, 2007; Haga et al., 2015). ...
... 作为植物进行光合作用的细胞器, 叶绿体响应外界光照刺激而发生的位置变化称为叶绿体光下重定位.弱光照射下, 为最大程度地吸收光能, 叶绿体沿细胞平周壁方向聚集, 这种现象为聚光运动(accumulation movement).强光条件下, 为避免过强的光对自身造成伤害, 叶绿体沿细胞垂周壁聚集, 使之与光平行分布, 这种现象称为避光运动(avoidance movement) (Kasahara et al., 2004).叶绿体运动受蓝光调节, 蓝光受体向光素PHOT1和PHOT2共同调控叶绿体的聚光反应, 而PHOT2单独调节叶绿体的避光反应(Suetsugu and Wada, 2017).与向光素调节的其它多细胞协调性反应(如向光性、叶片的伸展与定位)不同, 叶绿体运动是细胞自主性反应, 不受基因表达(Kong and Wada, 2016; Wada, 2016)和生长素运输的调节(Christie et al., 2018), 最终由叶绿体肌动蛋白响应入射光的强度与方向而发生快速重组介导(Kadota et al., 2009; Kong and Wada, 2014).叶绿体肌动蛋白特异定位于细胞质膜和叶绿体之间, 在叶绿体前进方向的后面叶绿体肌动蛋白纤维迅速解聚, 前进方向的前面叶绿体肌动蛋白纤维迅速聚集, 这种不对称的叶绿体肌动蛋白纤维分布最终决定了叶绿体的运动方向和速度(Kong and Wada, 2016).CHUP1 (CHLOROPLAST UNUSUAL POSITIONING 1)和 ...
... 近年来, 科学家研究发现, 拟南芥NRL家族蛋白在向光素介导的蓝光信号转导通路中起重要作用.拟南芥NRL家族6个分支中的3个分支(II、III和IV) (图1)蛋白目前已经被报道参与向光素调节的反应, 或者与向光素蛋白存在直接相互作用, 暗示这3个分支的其它成员可能参与向光素信号途径的调节.RPT2与NCH1都来自分支II, 既参与生长素依赖的向光性调节, 又参与非生长素依赖的叶绿体聚光运动调节(Suetsugu et al., 2016; Suetsugu and Wada, 2017).目前RPT2和NCH1如何区分向光素介导的2种信号途径并不清楚.考虑到RPT2和NCH1蛋白所在分支与其余2个分支(III和IV)的亲缘关系较远(Christie et al., 2018), 暗示该分支其它成员也可能同时参与生长素依赖和非依赖性的向光素反应.RPT2可与PHOT1的LOV1区相互作用并抑制PHOT1的自磷酸化, 以降低PHOT1的光敏感性.尽管RPT2也可以与PHOT2的LOV1区域相互作用, 但是RPT2不抑制PHOT2的活性(Kimura et al., 2020).可能存在NRL家族其它蛋白与PHOT2相互作用, 调节PHOT2介导的蓝光信号转导.NPH3主要调节生长素依赖的向光性反应以及叶片的伸展与定位(Harada et al., 2013; Zhao et al., 2018), 但NPH3激活后调节生长素不对称分布的机制并不清楚.NPH3的磷酸化状态与其生理活性以及在细胞中的定位密切相关(Haga et al., 2015; Zhao et al., 2018).PHOT1介导的NPH3去磷酸化在弱蓝光诱导的下胚轴向光弯曲反应中发挥重要作用, 强蓝光下NPH3并未发生去磷酸化反应(Tsuchida-Mayama et al., 2008).那么NPH3如何调节强蓝光诱导的下胚轴向光弯曲反应, 是否与其E3泛素化连接酶活性有关? NPY1与PID互作, 通过影响生长素信号调节植物器官的形成以及根的向重力性反应(Suetsugu et al., 2016).生长素不对称分布决定不对称生长, PID- NPY1与PHOT-NPH3作用方式又相似, 那么NPY分支成员是否参与调节向光素介导的植物向性运动以及叶片的伸展与定位? PID和PHOT都属于AGC激酶VIII亚族成员(Ding et al., 2011; Willige et al., 2013), NRL家族蛋白是否可以与AGC激酶VIII亚家族其它成员相互作用, 以调节生长素介导的其它生理反应? 来自分支IV的NRL2也可以与PHOT1发生互作(Sullivan et al., 2009), 暗示该分支的蛋白也可能在向光素介导的蓝光信号途径中发挥重要作用.综上, NRL家族蛋白功能的陆续鉴定, 有望解析向光素PHOT1和PHOT2感受蓝光信号差异, 进而调节不同生理反应的可能机制. ...
phot1 inhibition of ABCB19 primes lateral auxin fluxes in the shoot apex required for phototropism 5 2011
... 拟南芥中ABCB19 (ATP-BINDING CAS-SETTE B 19)和光敏色素激酶底物(PHYTOCHROME KINASE SUBSTRATE 4, PKS4)参与调节生长素的转运, 是下胚轴向光弯曲的负调节因子(Christie et al., 2011; Demarsy et al., 2012).自磷酸化激活的PHOT1通过直接磷酸化ABCB19和PKS4, 抑制ABCB19和PKS4的生长素转运活性, 促进下胚轴向光弯曲反应(Christie et al., 2011; Demarsy et al., 2012).PHOT1通过磷酸化修饰调节ABCB19的活性, 但是PHOT2并不影响ABCB19的活性(Christie et al., 2011).蛋白磷酸酶2A (PP2A)的A1亚基对应的蛋白RCN1 (Root Curling in N-naphthylphthalamic acid 1)与PHOT2蛋白发生体外互作, 负调节PHOT2介导的向光弯曲, 但RCN1并不与PHOT1直接相互作用(Tseng and Briggs, 2010).NPH3 (NONPHOTOTROPIC HYPOCOTYL 3)和RPT2 (ROOT PHOTOTROPISM 2)是通过筛选拟南芥下胚轴以及根的向光性缺失突变体获得(Motchoulski and Liscum, 1999; Sakai et al., 2000).随后的研究发现, NPH3和RPT2是向光素途径的重要调控因子, 均可与PHOT1蛋白直接相互作用(Blakeslee et al., 2004; Stone et al., 2008).虽然NPH3和RPT2也参与PHOT2信号转导途径, 但目前并无RPT2与PHOT2蛋白之间直接相互作用的报道(Inada et al., 2004; Lariguet et al., 2006).上述结果暗示PHOT1和PHOT2可能通过与不同的下游互作因子差异调节蓝光反应. ...
... ).自磷酸化激活的PHOT1通过直接磷酸化ABCB19和PKS4, 抑制ABCB19和PKS4的生长素转运活性, 促进下胚轴向光弯曲反应(Christie et al., 2011; Demarsy et al., 2012).PHOT1通过磷酸化修饰调节ABCB19的活性, 但是PHOT2并不影响ABCB19的活性(Christie et al., 2011).蛋白磷酸酶2A (PP2A)的A1亚基对应的蛋白RCN1 (Root Curling in N-naphthylphthalamic acid 1)与PHOT2蛋白发生体外互作, 负调节PHOT2介导的向光弯曲, 但RCN1并不与PHOT1直接相互作用(Tseng and Briggs, 2010).NPH3 (NONPHOTOTROPIC HYPOCOTYL 3)和RPT2 (ROOT PHOTOTROPISM 2)是通过筛选拟南芥下胚轴以及根的向光性缺失突变体获得(Motchoulski and Liscum, 1999; Sakai et al., 2000).随后的研究发现, NPH3和RPT2是向光素途径的重要调控因子, 均可与PHOT1蛋白直接相互作用(Blakeslee et al., 2004; Stone et al., 2008).虽然NPH3和RPT2也参与PHOT2信号转导途径, 但目前并无RPT2与PHOT2蛋白之间直接相互作用的报道(Inada et al., 2004; Lariguet et al., 2006).上述结果暗示PHOT1和PHOT2可能通过与不同的下游互作因子差异调节蓝光反应. ...
... ).PHOT1通过磷酸化修饰调节ABCB19的活性, 但是PHOT2并不影响ABCB19的活性(Christie et al., 2011).蛋白磷酸酶2A (PP2A)的A1亚基对应的蛋白RCN1 (Root Curling in N-naphthylphthalamic acid 1)与PHOT2蛋白发生体外互作, 负调节PHOT2介导的向光弯曲, 但RCN1并不与PHOT1直接相互作用(Tseng and Briggs, 2010).NPH3 (NONPHOTOTROPIC HYPOCOTYL 3)和RPT2 (ROOT PHOTOTROPISM 2)是通过筛选拟南芥下胚轴以及根的向光性缺失突变体获得(Motchoulski and Liscum, 1999; Sakai et al., 2000).随后的研究发现, NPH3和RPT2是向光素途径的重要调控因子, 均可与PHOT1蛋白直接相互作用(Blakeslee et al., 2004; Stone et al., 2008).虽然NPH3和RPT2也参与PHOT2信号转导途径, 但目前并无RPT2与PHOT2蛋白之间直接相互作用的报道(Inada et al., 2004; Lariguet et al., 2006).上述结果暗示PHOT1和PHOT2可能通过与不同的下游互作因子差异调节蓝光反应. ...
... 早在达尔文(1880年)时期, 人们就已发现植物向光性, 但是直到向光素PHOT1和PHOT2先后被鉴定(Liscum and Briggs, 1995; Christie et al., 1998), 植物的向光性机理研究才取得突飞猛进的发展.研究发现, PHOT1调节弱蓝光(0.01-1 μmol·m-2·s-1)和强蓝光(>1 μmol·m-2·s-1)诱导的下胚轴的向光性; PHOT2仅在强蓝光下起作用, 且与PHOT1功能冗余(Sakai et al., 2001; Briggs and Christie, 2002).由于强蓝光下PHOT1和PHOT2的功能冗余, 目前向光性机制研究主要集中在PHOT1介导的弱蓝光反应(Motchoulski and Liscum, 1999; Haga et al., 2015), 仅有少量研究通过突变体背景下的抑制子筛选寻找参与强蓝光信号途径的作用因子(赵翔等, 2014).单侧蓝光照射下, 自磷酸化激活的向光素蛋白将信号传递给NPH3、RPT2以及PKS等下游因子(Motchoulski and Liscum, 1999; Inada et al., 2004), 调节生长素运输载体的活性与定位(Ding et al., 2011).例如, 引起内皮层细胞背光侧PIN3 (PIN-FORMED3)积累(刘广超和丁兆军, 2018), 促进背光侧生长素含量增加、细胞伸长速率加快, 最终引起植物弯向光生长(Christie and Murphy, 2013).下胚轴弯曲过程中生长素的不对称分布受各类生长素运输载体的共同调控, 如PINs (PIN3、PIN4和PIN7)和ABCB19 (Christie et al., 2011).蓝光下, PIN3的磷酸化受抑制, 促进PIN3的背光侧定位, 诱导背光侧生长素积累以及下胚轴的向光性生长(Ding et al., 2011).PIN3的磷酸化以及极性定位受AGC激酶VIII亚族成员PID (PINOID)以及D6PK (D6 PROTEIN KINASE)的调节(Ding et al., 2011; Willige et al., 2013).ABCB19是生长素外流载体, 被蓝光激活的PHOT1磷酸化后, ABCB19的生长素转运活性受抑制, 增加拟南芥下胚轴顶端生长素含量并抑制其垂直生长, 同时通过PIN3蛋白促进生长素在下胚轴中侧向移动(Christie et al., 2011).背光侧生长素的大量积累促进了背光侧IAA19的降解, 释放出有活性的ARF7 (Sun et al., 2013), 不对称激活与生长相关基因的表达促进了下胚轴的向光性反应(刘广超和丁兆军, 2018). ...
... ).ABCB19是生长素外流载体, 被蓝光激活的PHOT1磷酸化后, ABCB19的生长素转运活性受抑制, 增加拟南芥下胚轴顶端生长素含量并抑制其垂直生长, 同时通过PIN3蛋白促进生长素在下胚轴中侧向移动(Christie et al., 2011).背光侧生长素的大量积累促进了背光侧IAA19的降解, 释放出有活性的ARF7 (Sun et al., 2013), 不对称激活与生长相关基因的表达促进了下胚轴的向光性反应(刘广超和丁兆军, 2018). ...
The Arabidopsis PHYTOCHROME KINASE SUBSTRATE 2 protein is a phototropin signaling element that regulates leaf flattening and leaf positioning 5 2010
... NRL (NPH3/RPT2-Like)蛋白家族是植物特有的, 其成员含有3个保守结构域: N端的BTB (Bric àbrac, Tramtrack, and Broad complex)结构域、中间的NPH3结构域以及C端的螺旋-螺旋结构域(Liscum et al., 2014).其中, BTB结构域主要起E3连接酶的作用, 与CUL3发生相互作用(Gingerich et al., 2005; Figueroa et al., 2005), C端的螺旋-螺旋结构域主要介导蛋白之间的相互作用(Inada et al., 2004).拟南芥中33个NRL家族成员都含有NPH3结构域, 10个缺少C端的螺旋-螺旋结构域, 2个缺少N端的BTB结构域(Pedmale et al., 2010; Christie et al., 2018).根据NRL家族成员间的进化关系, 拟南芥中的33个NRL家族成员可分为6个分支(图1) (Suetsugu et al., 2016; Christie et al., 2018).目前, 来自2个分支的10个成员的功能已得到鉴定(de Carbonnel et al., 2010; Christie et al., 2018) (图1). ...
... 拟南芥NPH3基因突变后, 其叶柄与水平面之间的夹角变小, 叶片向背面卷曲生长, 与phot1/phot2双突变体表型类似, 暗示该基因参与向光素介导的叶片伸展与定位(de Carbonnel et al., 2010).nph3/phot1双突变体叶片的卷曲程度比nph3明显, 更接近phot1/ phot2, nph3/phot2双突变体的叶片表型与nph3基本相同, 均表现为叶柄与水平面之间的夹角变小, 叶片向背面卷曲生长(de Carbonnel et al., 2010).该结果表明NPH3同时参与PHOT1和PHOT2介导的叶片伸展与定位, 且在PHOT2介导的途径中起重要作用.在拟南芥中的研究结果表明, PKS2通过影响生长素的运输在叶片的伸展与定位调节中起重要作用, 而与PKS2相比, PKS1的作用相对较弱, PKS4并不参与叶片伸展与定位的调节(de Carbonnel et al., 2010).与phot1/phot2双突变体类似, nph3/pks2双突变体叶柄几乎平行于水平面, 叶片向背面卷曲以近似竖直的角度悬挂于叶柄上, 且该表型受PKS2和NPH3的协同调节(de Carbonnel et al., 2010), 证明NPH3可能通过与PKS2相互作用调节PHOT1和PHOT2介导的叶片伸展与定位. ...
... 基本相同, 均表现为叶柄与水平面之间的夹角变小, 叶片向背面卷曲生长(de Carbonnel et al., 2010).该结果表明NPH3同时参与PHOT1和PHOT2介导的叶片伸展与定位, 且在PHOT2介导的途径中起重要作用.在拟南芥中的研究结果表明, PKS2通过影响生长素的运输在叶片的伸展与定位调节中起重要作用, 而与PKS2相比, PKS1的作用相对较弱, PKS4并不参与叶片伸展与定位的调节(de Carbonnel et al., 2010).与phot1/phot2双突变体类似, nph3/pks2双突变体叶柄几乎平行于水平面, 叶片向背面卷曲以近似竖直的角度悬挂于叶柄上, 且该表型受PKS2和NPH3的协同调节(de Carbonnel et al., 2010), 证明NPH3可能通过与PKS2相互作用调节PHOT1和PHOT2介导的叶片伸展与定位. ...
... ).该结果表明NPH3同时参与PHOT1和PHOT2介导的叶片伸展与定位, 且在PHOT2介导的途径中起重要作用.在拟南芥中的研究结果表明, PKS2通过影响生长素的运输在叶片的伸展与定位调节中起重要作用, 而与PKS2相比, PKS1的作用相对较弱, PKS4并不参与叶片伸展与定位的调节(de Carbonnel et al., 2010).与phot1/phot2双突变体类似, nph3/pks2双突变体叶柄几乎平行于水平面, 叶片向背面卷曲以近似竖直的角度悬挂于叶柄上, 且该表型受PKS2和NPH3的协同调节(de Carbonnel et al., 2010), 证明NPH3可能通过与PKS2相互作用调节PHOT1和PHOT2介导的叶片伸展与定位. ...
... 双突变体叶柄几乎平行于水平面, 叶片向背面卷曲以近似竖直的角度悬挂于叶柄上, 且该表型受PKS2和NPH3的协同调节(de Carbonnel et al., 2010), 证明NPH3可能通过与PKS2相互作用调节PHOT1和PHOT2介导的叶片伸展与定位. ...
Higher plants use LOV to perceive blue light 1 2009
... 叶片是植物进行光合作用的场所.自然光下, 野生型拟南芥叶柄向上倾斜生长, 叶片的叶面平整, 几乎平行于水平方向生长, 该现象被称为叶片的伸展与定位.弱光下, 拟南芥phot2突变体叶片伸展与定位表型几乎与野生型相同, phot1突变体叶片的定位发生变化, 叶柄与水平面夹角减小(Harada et al., 2013), 表明弱光下PHOT1单独调节叶片的伸展与定位(Demarsy and Fankhauser, 2009).强光下, 拟南芥phot1和phot2单突变体叶片伸展与定位的表型与野生型基本一致, phot1/phot2双突变体叶柄略微向下倾斜生长, 叶片向背面卷曲, 以几乎垂直于水平面的角度悬挂于叶柄上, 表明PHOT1和PHOT2以功能冗余的方式调节强光下叶片的伸展与定位(Sakai et al., 2001).叶片的伸展与定位可以确保叶片采取最佳的茎叶角度, 以便最大程度地捕获光源并且趋避强光伤害(Harada et al., 2013; Shang et al., 2019).与此一致, 拟南芥的研究表明: 弱光下, phot2的鲜重与野生型几乎相等, phot1的鲜重是野生型的1/2; 强光下, phot1和phot2单突变体的鲜重与野生型基本一致, 但phot1/phot2双突变体的鲜重大约是野生型的1/3 (Harada et al., 2013). ...
Phytochrome Kinase Substrate 4 is phosphorylated by the phototropin 1 photoreceptor 2 2012
... 拟南芥中ABCB19 (ATP-BINDING CAS-SETTE B 19)和光敏色素激酶底物(PHYTOCHROME KINASE SUBSTRATE 4, PKS4)参与调节生长素的转运, 是下胚轴向光弯曲的负调节因子(Christie et al., 2011; Demarsy et al., 2012).自磷酸化激活的PHOT1通过直接磷酸化ABCB19和PKS4, 抑制ABCB19和PKS4的生长素转运活性, 促进下胚轴向光弯曲反应(Christie et al., 2011; Demarsy et al., 2012).PHOT1通过磷酸化修饰调节ABCB19的活性, 但是PHOT2并不影响ABCB19的活性(Christie et al., 2011).蛋白磷酸酶2A (PP2A)的A1亚基对应的蛋白RCN1 (Root Curling in N-naphthylphthalamic acid 1)与PHOT2蛋白发生体外互作, 负调节PHOT2介导的向光弯曲, 但RCN1并不与PHOT1直接相互作用(Tseng and Briggs, 2010).NPH3 (NONPHOTOTROPIC HYPOCOTYL 3)和RPT2 (ROOT PHOTOTROPISM 2)是通过筛选拟南芥下胚轴以及根的向光性缺失突变体获得(Motchoulski and Liscum, 1999; Sakai et al., 2000).随后的研究发现, NPH3和RPT2是向光素途径的重要调控因子, 均可与PHOT1蛋白直接相互作用(Blakeslee et al., 2004; Stone et al., 2008).虽然NPH3和RPT2也参与PHOT2信号转导途径, 但目前并无RPT2与PHOT2蛋白之间直接相互作用的报道(Inada et al., 2004; Lariguet et al., 2006).上述结果暗示PHOT1和PHOT2可能通过与不同的下游互作因子差异调节蓝光反应. ...
... ; Demarsy et al., 2012).PHOT1通过磷酸化修饰调节ABCB19的活性, 但是PHOT2并不影响ABCB19的活性(Christie et al., 2011).蛋白磷酸酶2A (PP2A)的A1亚基对应的蛋白RCN1 (Root Curling in N-naphthylphthalamic acid 1)与PHOT2蛋白发生体外互作, 负调节PHOT2介导的向光弯曲, 但RCN1并不与PHOT1直接相互作用(Tseng and Briggs, 2010).NPH3 (NONPHOTOTROPIC HYPOCOTYL 3)和RPT2 (ROOT PHOTOTROPISM 2)是通过筛选拟南芥下胚轴以及根的向光性缺失突变体获得(Motchoulski and Liscum, 1999; Sakai et al., 2000).随后的研究发现, NPH3和RPT2是向光素途径的重要调控因子, 均可与PHOT1蛋白直接相互作用(Blakeslee et al., 2004; Stone et al., 2008).虽然NPH3和RPT2也参与PHOT2信号转导途径, 但目前并无RPT2与PHOT2蛋白之间直接相互作用的报道(Inada et al., 2004; Lariguet et al., 2006).上述结果暗示PHOT1和PHOT2可能通过与不同的下游互作因子差异调节蓝光反应. ...
Blue light-induced proteomic changes in etiolated Arabidopsis seedlings 1 2014
... 拟南芥PHOT1至少有21个氨基酸位点可以发生自磷酸化(Boex-Fontvieille et al., 2014; Deng et al., 2014), PHOT2有29个磷酸化位点(Inoue et al., 2011; Boex-Fontvieille et al., 2014), 这些位点的具体分布参见Christie等(2015)的报道.其中, C端2个保守位点的磷酸化(PHOT1中S849和S851, PHOT2中S761 和S763)对向光素受体的信号传递至关重要(Inoue et al., 2008a; Inoue et al., 2011), 其余的磷酸化位点大部分位于N端LOV1的上游以及LOV1与LOV2中间的连接区域(Christie et al., 2015).虽然这些上游区域磷酸化位点的生物学意义目前还不清楚, 但这些位点的磷酸化程度受光照强度调节(Salomon et al., 2003), 暗示PHOT1和PHOT2可能通过磷酸化程度差异响应不同的光照强度, 从而参与植物多种生理活动的调节. ...
Light-mediated polarization of the PIN3 auxin transporter for the phototropic response in Arabidopsis 4 2011
... 早在达尔文(1880年)时期, 人们就已发现植物向光性, 但是直到向光素PHOT1和PHOT2先后被鉴定(Liscum and Briggs, 1995; Christie et al., 1998), 植物的向光性机理研究才取得突飞猛进的发展.研究发现, PHOT1调节弱蓝光(0.01-1 μmol·m-2·s-1)和强蓝光(>1 μmol·m-2·s-1)诱导的下胚轴的向光性; PHOT2仅在强蓝光下起作用, 且与PHOT1功能冗余(Sakai et al., 2001; Briggs and Christie, 2002).由于强蓝光下PHOT1和PHOT2的功能冗余, 目前向光性机制研究主要集中在PHOT1介导的弱蓝光反应(Motchoulski and Liscum, 1999; Haga et al., 2015), 仅有少量研究通过突变体背景下的抑制子筛选寻找参与强蓝光信号途径的作用因子(赵翔等, 2014).单侧蓝光照射下, 自磷酸化激活的向光素蛋白将信号传递给NPH3、RPT2以及PKS等下游因子(Motchoulski and Liscum, 1999; Inada et al., 2004), 调节生长素运输载体的活性与定位(Ding et al., 2011).例如, 引起内皮层细胞背光侧PIN3 (PIN-FORMED3)积累(刘广超和丁兆军, 2018), 促进背光侧生长素含量增加、细胞伸长速率加快, 最终引起植物弯向光生长(Christie and Murphy, 2013).下胚轴弯曲过程中生长素的不对称分布受各类生长素运输载体的共同调控, 如PINs (PIN3、PIN4和PIN7)和ABCB19 (Christie et al., 2011).蓝光下, PIN3的磷酸化受抑制, 促进PIN3的背光侧定位, 诱导背光侧生长素积累以及下胚轴的向光性生长(Ding et al., 2011).PIN3的磷酸化以及极性定位受AGC激酶VIII亚族成员PID (PINOID)以及D6PK (D6 PROTEIN KINASE)的调节(Ding et al., 2011; Willige et al., 2013).ABCB19是生长素外流载体, 被蓝光激活的PHOT1磷酸化后, ABCB19的生长素转运活性受抑制, 增加拟南芥下胚轴顶端生长素含量并抑制其垂直生长, 同时通过PIN3蛋白促进生长素在下胚轴中侧向移动(Christie et al., 2011).背光侧生长素的大量积累促进了背光侧IAA19的降解, 释放出有活性的ARF7 (Sun et al., 2013), 不对称激活与生长相关基因的表达促进了下胚轴的向光性反应(刘广超和丁兆军, 2018). ...
... ).蓝光下, PIN3的磷酸化受抑制, 促进PIN3的背光侧定位, 诱导背光侧生长素积累以及下胚轴的向光性生长(Ding et al., 2011).PIN3的磷酸化以及极性定位受AGC激酶VIII亚族成员PID (PINOID)以及D6PK (D6 PROTEIN KINASE)的调节(Ding et al., 2011; Willige et al., 2013).ABCB19是生长素外流载体, 被蓝光激活的PHOT1磷酸化后, ABCB19的生长素转运活性受抑制, 增加拟南芥下胚轴顶端生长素含量并抑制其垂直生长, 同时通过PIN3蛋白促进生长素在下胚轴中侧向移动(Christie et al., 2011).背光侧生长素的大量积累促进了背光侧IAA19的降解, 释放出有活性的ARF7 (Sun et al., 2013), 不对称激活与生长相关基因的表达促进了下胚轴的向光性反应(刘广超和丁兆军, 2018). ...
... ).PIN3的磷酸化以及极性定位受AGC激酶VIII亚族成员PID (PINOID)以及D6PK (D6 PROTEIN KINASE)的调节(Ding et al., 2011; Willige et al., 2013).ABCB19是生长素外流载体, 被蓝光激活的PHOT1磷酸化后, ABCB19的生长素转运活性受抑制, 增加拟南芥下胚轴顶端生长素含量并抑制其垂直生长, 同时通过PIN3蛋白促进生长素在下胚轴中侧向移动(Christie et al., 2011).背光侧生长素的大量积累促进了背光侧IAA19的降解, 释放出有活性的ARF7 (Sun et al., 2013), 不对称激活与生长相关基因的表达促进了下胚轴的向光性反应(刘广超和丁兆军, 2018). ...
... 近年来, 科学家研究发现, 拟南芥NRL家族蛋白在向光素介导的蓝光信号转导通路中起重要作用.拟南芥NRL家族6个分支中的3个分支(II、III和IV) (图1)蛋白目前已经被报道参与向光素调节的反应, 或者与向光素蛋白存在直接相互作用, 暗示这3个分支的其它成员可能参与向光素信号途径的调节.RPT2与NCH1都来自分支II, 既参与生长素依赖的向光性调节, 又参与非生长素依赖的叶绿体聚光运动调节(Suetsugu et al., 2016; Suetsugu and Wada, 2017).目前RPT2和NCH1如何区分向光素介导的2种信号途径并不清楚.考虑到RPT2和NCH1蛋白所在分支与其余2个分支(III和IV)的亲缘关系较远(Christie et al., 2018), 暗示该分支其它成员也可能同时参与生长素依赖和非依赖性的向光素反应.RPT2可与PHOT1的LOV1区相互作用并抑制PHOT1的自磷酸化, 以降低PHOT1的光敏感性.尽管RPT2也可以与PHOT2的LOV1区域相互作用, 但是RPT2不抑制PHOT2的活性(Kimura et al., 2020).可能存在NRL家族其它蛋白与PHOT2相互作用, 调节PHOT2介导的蓝光信号转导.NPH3主要调节生长素依赖的向光性反应以及叶片的伸展与定位(Harada et al., 2013; Zhao et al., 2018), 但NPH3激活后调节生长素不对称分布的机制并不清楚.NPH3的磷酸化状态与其生理活性以及在细胞中的定位密切相关(Haga et al., 2015; Zhao et al., 2018).PHOT1介导的NPH3去磷酸化在弱蓝光诱导的下胚轴向光弯曲反应中发挥重要作用, 强蓝光下NPH3并未发生去磷酸化反应(Tsuchida-Mayama et al., 2008).那么NPH3如何调节强蓝光诱导的下胚轴向光弯曲反应, 是否与其E3泛素化连接酶活性有关? NPY1与PID互作, 通过影响生长素信号调节植物器官的形成以及根的向重力性反应(Suetsugu et al., 2016).生长素不对称分布决定不对称生长, PID- NPY1与PHOT-NPH3作用方式又相似, 那么NPY分支成员是否参与调节向光素介导的植物向性运动以及叶片的伸展与定位? PID和PHOT都属于AGC激酶VIII亚族成员(Ding et al., 2011; Willige et al., 2013), NRL家族蛋白是否可以与AGC激酶VIII亚家族其它成员相互作用, 以调节生长素介导的其它生理反应? 来自分支IV的NRL2也可以与PHOT1发生互作(Sullivan et al., 2009), 暗示该分支的蛋白也可能在向光素介导的蓝光信号途径中发挥重要作用.综上, NRL家族蛋白功能的陆续鉴定, 有望解析向光素PHOT1和PHOT2感受蓝光信号差异, 进而调节不同生理反应的可能机制. ...
Plant phototropic growth 2 2015
... 拟南芥nph3突变体在任何强度的单侧蓝光下均表现缺失下胚轴向光性(Liscum and Briggs, 1996; Liscum et al., 2014; Fankhauser and Christie, 2015), 表明NPH3调节PHOT1和PHOT2介导蓝光诱导的下胚轴弯曲反应.黑暗条件下, 磷酸化的NPH3蛋白与PHOT1结合, 定位于细胞膜, 单侧弱蓝光照射下, NPH3迅速发生PHOT1依赖的去磷酸化反应, 从细胞膜上脱离(Haga et al., 2015; Zhao et al., 2018).目前磷酸化NPH3的激酶还不清楚, 但NPH3去磷酸化主要由I型蛋白磷酸酶介导(Pedmale and Liscum, 2007).去磷酸化后NPH3从细胞膜向胞质的转移降低了其介导向光弯曲反应的能力(Haga et al., 2015; Sullivan et al., 2019).随着光照强度的增加, 下胚轴的弯曲度呈强度依赖的钟形曲线(Christie and Murphy, 2013).随着光照时间的增加, RPT2蛋白含量增加, 与去磷酸化的NPH3结合形成RPT2-NPH3复合物并与PHOT1蛋白结合, 促进NPH3的磷酸化以及从胞质到细胞膜的重定位, 开启第二次向光弯曲(Haga et al., 2015; Zhao et al., 2018).虽然PHOT1- NPH3复合体在细胞膜的重定位对持续光照介导的二次向光弯曲至关重要(Haga et al., 2015; Christie et al., 2018; Zhao et al., 2018), 但其生物学意义还不清楚, 人们认为这种变化有可能对调节生长素的不对称分布有重要影响(Pedmale and Liscum, 2007; Haga et al., 2015). ...
... NPH3主要通过2种途径影响生长素的不对称分布.一是NPH3通过多泛素化修饰PHOT1促进PHOT1的降解, 间接影响生长素转运蛋白(如生长素外流载体PIN1 (Roberts et al., 2011)和PIN2 (Wan et al., 2012))的重定位.NPH3是以Cullin3为基础的E3泛素化连接酶CRL3NPH3复合体的组成成分, 在该复合体中作为底物连接蛋白发挥作用(Roberts et al., 2011; Wan et al., 2012).强蓝光下, CRL3NPH3复合体直接与PHOT1蛋白相互作用, 促进PHOT1的单/多泛素化修饰, 多泛素化修饰的PHOT1进入细胞质后被26S蛋白酶体识别后降解(Roberts et al., 2011).PHOT1从细胞膜进入细胞质影响了生长素外流载体PIN1 (Roberts et al., 2011)和PIN2 (Wan et al., 2012)的重定位, 但具体作用机制有待深入研究.二是NPH3通过与PKS家族蛋白互作调节生长素的转运(Fankhauser and Christie, 2015).PKS家族成员是向光弯曲反应的正向调节因子, 参与生长素转运调节, 它们可以与向光素和NPH3蛋白直接互作, 形成PHOT1-PKS-NPH3复合体(Kami et al., 2014), 但是复合体的作用机制目前还不清楚. ...
Arabidopsis has two redundant Cullin3 proteins that are essential for embryo development and that interact with RBX1 and BTB proteins to form multisubunit E3 ubiquitin ligase complexes in vivo 1 2005
... NRL (NPH3/RPT2-Like)蛋白家族是植物特有的, 其成员含有3个保守结构域: N端的BTB (Bric àbrac, Tramtrack, and Broad complex)结构域、中间的NPH3结构域以及C端的螺旋-螺旋结构域(Liscum et al., 2014).其中, BTB结构域主要起E3连接酶的作用, 与CUL3发生相互作用(Gingerich et al., 2005; Figueroa et al., 2005), C端的螺旋-螺旋结构域主要介导蛋白之间的相互作用(Inada et al., 2004).拟南芥中33个NRL家族成员都含有NPH3结构域, 10个缺少C端的螺旋-螺旋结构域, 2个缺少N端的BTB结构域(Pedmale et al., 2010; Christie et al., 2018).根据NRL家族成员间的进化关系, 拟南芥中的33个NRL家族成员可分为6个分支(图1) (Suetsugu et al., 2016; Christie et al., 2018).目前, 来自2个分支的10个成员的功能已得到鉴定(de Carbonnel et al., 2010; Christie et al., 2018) (图1). ...
Phototropin 1 is required for high-fluence blue-light-mediated mRNA destabilization 1 2003
... 拟南芥(Arabidopsis thaliana) PHOT1和PHOT2以功能冗余的方式调节下胚轴向光性和根负向光性、叶绿体聚光运动、气孔开放以及叶片伸展与定位等(Christie et al., 2015).PHOT1特异性抑制光下黄化幼苗下胚轴的快速伸长(Folta et al., 2003)、子叶变绿过程中叶绿素结合蛋白mRNA转录的稳定性(Folta and Kaufman, 2003)以及侧根的快速生长(Moni et al., 2015).PHOT2不仅介导强蓝光诱导的下胚轴向光弯曲(Sakai et al., 2001), 而且也特异调节叶绿体和细胞核的避光运动(Higa et al., 2014)以及抗性蛋白介导的病毒防御反应(Jeong et al., 2010).PHOT1和PHOT2的功能差异可能与磷酸化修饰位点、磷酸化程度、亚细胞定位变化以及下游相互作用蛋白种类不同有关(Zhao et al., 2013; Morrow et al., 2018; Liscum et al., 2020). ...
Primary inhibition of hypocotyl growth and phototropism depend differently on phototropin-mediated increases in cytoplasmic calcium induced by blue light 1 2003
... 拟南芥(Arabidopsis thaliana) PHOT1和PHOT2以功能冗余的方式调节下胚轴向光性和根负向光性、叶绿体聚光运动、气孔开放以及叶片伸展与定位等(Christie et al., 2015).PHOT1特异性抑制光下黄化幼苗下胚轴的快速伸长(Folta et al., 2003)、子叶变绿过程中叶绿素结合蛋白mRNA转录的稳定性(Folta and Kaufman, 2003)以及侧根的快速生长(Moni et al., 2015).PHOT2不仅介导强蓝光诱导的下胚轴向光弯曲(Sakai et al., 2001), 而且也特异调节叶绿体和细胞核的避光运动(Higa et al., 2014)以及抗性蛋白介导的病毒防御反应(Jeong et al., 2010).PHOT1和PHOT2的功能差异可能与磷酸化修饰位点、磷酸化程度、亚细胞定位变化以及下游相互作用蛋白种类不同有关(Zhao et al., 2013; Morrow et al., 2018; Liscum et al., 2020). ...
Cullins 3a and 3b assemble with members of the broad complex/tramtrack/bric-a- brac (BTB) protein family to form essential ubiquitin-protein ligases (E3s) in Arabidopsis 1 2005
... NRL (NPH3/RPT2-Like)蛋白家族是植物特有的, 其成员含有3个保守结构域: N端的BTB (Bric àbrac, Tramtrack, and Broad complex)结构域、中间的NPH3结构域以及C端的螺旋-螺旋结构域(Liscum et al., 2014).其中, BTB结构域主要起E3连接酶的作用, 与CUL3发生相互作用(Gingerich et al., 2005; Figueroa et al., 2005), C端的螺旋-螺旋结构域主要介导蛋白之间的相互作用(Inada et al., 2004).拟南芥中33个NRL家族成员都含有NPH3结构域, 10个缺少C端的螺旋-螺旋结构域, 2个缺少N端的BTB结构域(Pedmale et al., 2010; Christie et al., 2018).根据NRL家族成员间的进化关系, 拟南芥中的33个NRL家族成员可分为6个分支(图1) (Suetsugu et al., 2016; Christie et al., 2018).目前, 来自2个分支的10个成员的功能已得到鉴定(de Carbonnel et al., 2010; Christie et al., 2018) (图1). ...
Arabidopsis ROOT PHOTOTROPISM2 contributes to the adaptation to high-intensity light in phototropic responses 7 2015
... 早在达尔文(1880年)时期, 人们就已发现植物向光性, 但是直到向光素PHOT1和PHOT2先后被鉴定(Liscum and Briggs, 1995; Christie et al., 1998), 植物的向光性机理研究才取得突飞猛进的发展.研究发现, PHOT1调节弱蓝光(0.01-1 μmol·m-2·s-1)和强蓝光(>1 μmol·m-2·s-1)诱导的下胚轴的向光性; PHOT2仅在强蓝光下起作用, 且与PHOT1功能冗余(Sakai et al., 2001; Briggs and Christie, 2002).由于强蓝光下PHOT1和PHOT2的功能冗余, 目前向光性机制研究主要集中在PHOT1介导的弱蓝光反应(Motchoulski and Liscum, 1999; Haga et al., 2015), 仅有少量研究通过突变体背景下的抑制子筛选寻找参与强蓝光信号途径的作用因子(赵翔等, 2014).单侧蓝光照射下, 自磷酸化激活的向光素蛋白将信号传递给NPH3、RPT2以及PKS等下游因子(Motchoulski and Liscum, 1999; Inada et al., 2004), 调节生长素运输载体的活性与定位(Ding et al., 2011).例如, 引起内皮层细胞背光侧PIN3 (PIN-FORMED3)积累(刘广超和丁兆军, 2018), 促进背光侧生长素含量增加、细胞伸长速率加快, 最终引起植物弯向光生长(Christie and Murphy, 2013).下胚轴弯曲过程中生长素的不对称分布受各类生长素运输载体的共同调控, 如PINs (PIN3、PIN4和PIN7)和ABCB19 (Christie et al., 2011).蓝光下, PIN3的磷酸化受抑制, 促进PIN3的背光侧定位, 诱导背光侧生长素积累以及下胚轴的向光性生长(Ding et al., 2011).PIN3的磷酸化以及极性定位受AGC激酶VIII亚族成员PID (PINOID)以及D6PK (D6 PROTEIN KINASE)的调节(Ding et al., 2011; Willige et al., 2013).ABCB19是生长素外流载体, 被蓝光激活的PHOT1磷酸化后, ABCB19的生长素转运活性受抑制, 增加拟南芥下胚轴顶端生长素含量并抑制其垂直生长, 同时通过PIN3蛋白促进生长素在下胚轴中侧向移动(Christie et al., 2011).背光侧生长素的大量积累促进了背光侧IAA19的降解, 释放出有活性的ARF7 (Sun et al., 2013), 不对称激活与生长相关基因的表达促进了下胚轴的向光性反应(刘广超和丁兆军, 2018). ...
... 拟南芥nph3突变体在任何强度的单侧蓝光下均表现缺失下胚轴向光性(Liscum and Briggs, 1996; Liscum et al., 2014; Fankhauser and Christie, 2015), 表明NPH3调节PHOT1和PHOT2介导蓝光诱导的下胚轴弯曲反应.黑暗条件下, 磷酸化的NPH3蛋白与PHOT1结合, 定位于细胞膜, 单侧弱蓝光照射下, NPH3迅速发生PHOT1依赖的去磷酸化反应, 从细胞膜上脱离(Haga et al., 2015; Zhao et al., 2018).目前磷酸化NPH3的激酶还不清楚, 但NPH3去磷酸化主要由I型蛋白磷酸酶介导(Pedmale and Liscum, 2007).去磷酸化后NPH3从细胞膜向胞质的转移降低了其介导向光弯曲反应的能力(Haga et al., 2015; Sullivan et al., 2019).随着光照强度的增加, 下胚轴的弯曲度呈强度依赖的钟形曲线(Christie and Murphy, 2013).随着光照时间的增加, RPT2蛋白含量增加, 与去磷酸化的NPH3结合形成RPT2-NPH3复合物并与PHOT1蛋白结合, 促进NPH3的磷酸化以及从胞质到细胞膜的重定位, 开启第二次向光弯曲(Haga et al., 2015; Zhao et al., 2018).虽然PHOT1- NPH3复合体在细胞膜的重定位对持续光照介导的二次向光弯曲至关重要(Haga et al., 2015; Christie et al., 2018; Zhao et al., 2018), 但其生物学意义还不清楚, 人们认为这种变化有可能对调节生长素的不对称分布有重要影响(Pedmale and Liscum, 2007; Haga et al., 2015). ...
... ).去磷酸化后NPH3从细胞膜向胞质的转移降低了其介导向光弯曲反应的能力(Haga et al., 2015; Sullivan et al., 2019).随着光照强度的增加, 下胚轴的弯曲度呈强度依赖的钟形曲线(Christie and Murphy, 2013).随着光照时间的增加, RPT2蛋白含量增加, 与去磷酸化的NPH3结合形成RPT2-NPH3复合物并与PHOT1蛋白结合, 促进NPH3的磷酸化以及从胞质到细胞膜的重定位, 开启第二次向光弯曲(Haga et al., 2015; Zhao et al., 2018).虽然PHOT1- NPH3复合体在细胞膜的重定位对持续光照介导的二次向光弯曲至关重要(Haga et al., 2015; Christie et al., 2018; Zhao et al., 2018), 但其生物学意义还不清楚, 人们认为这种变化有可能对调节生长素的不对称分布有重要影响(Pedmale and Liscum, 2007; Haga et al., 2015). ...
... ).随着光照时间的增加, RPT2蛋白含量增加, 与去磷酸化的NPH3结合形成RPT2-NPH3复合物并与PHOT1蛋白结合, 促进NPH3的磷酸化以及从胞质到细胞膜的重定位, 开启第二次向光弯曲(Haga et al., 2015; Zhao et al., 2018).虽然PHOT1- NPH3复合体在细胞膜的重定位对持续光照介导的二次向光弯曲至关重要(Haga et al., 2015; Christie et al., 2018; Zhao et al., 2018), 但其生物学意义还不清楚, 人们认为这种变化有可能对调节生长素的不对称分布有重要影响(Pedmale and Liscum, 2007; Haga et al., 2015). ...
... ).虽然PHOT1- NPH3复合体在细胞膜的重定位对持续光照介导的二次向光弯曲至关重要(Haga et al., 2015; Christie et al., 2018; Zhao et al., 2018), 但其生物学意义还不清楚, 人们认为这种变化有可能对调节生长素的不对称分布有重要影响(Pedmale and Liscum, 2007; Haga et al., 2015). ...
... ; Haga et al., 2015). ...
... 近年来, 科学家研究发现, 拟南芥NRL家族蛋白在向光素介导的蓝光信号转导通路中起重要作用.拟南芥NRL家族6个分支中的3个分支(II、III和IV) (图1)蛋白目前已经被报道参与向光素调节的反应, 或者与向光素蛋白存在直接相互作用, 暗示这3个分支的其它成员可能参与向光素信号途径的调节.RPT2与NCH1都来自分支II, 既参与生长素依赖的向光性调节, 又参与非生长素依赖的叶绿体聚光运动调节(Suetsugu et al., 2016; Suetsugu and Wada, 2017).目前RPT2和NCH1如何区分向光素介导的2种信号途径并不清楚.考虑到RPT2和NCH1蛋白所在分支与其余2个分支(III和IV)的亲缘关系较远(Christie et al., 2018), 暗示该分支其它成员也可能同时参与生长素依赖和非依赖性的向光素反应.RPT2可与PHOT1的LOV1区相互作用并抑制PHOT1的自磷酸化, 以降低PHOT1的光敏感性.尽管RPT2也可以与PHOT2的LOV1区域相互作用, 但是RPT2不抑制PHOT2的活性(Kimura et al., 2020).可能存在NRL家族其它蛋白与PHOT2相互作用, 调节PHOT2介导的蓝光信号转导.NPH3主要调节生长素依赖的向光性反应以及叶片的伸展与定位(Harada et al., 2013; Zhao et al., 2018), 但NPH3激活后调节生长素不对称分布的机制并不清楚.NPH3的磷酸化状态与其生理活性以及在细胞中的定位密切相关(Haga et al., 2015; Zhao et al., 2018).PHOT1介导的NPH3去磷酸化在弱蓝光诱导的下胚轴向光弯曲反应中发挥重要作用, 强蓝光下NPH3并未发生去磷酸化反应(Tsuchida-Mayama et al., 2008).那么NPH3如何调节强蓝光诱导的下胚轴向光弯曲反应, 是否与其E3泛素化连接酶活性有关? NPY1与PID互作, 通过影响生长素信号调节植物器官的形成以及根的向重力性反应(Suetsugu et al., 2016).生长素不对称分布决定不对称生长, PID- NPY1与PHOT-NPH3作用方式又相似, 那么NPY分支成员是否参与调节向光素介导的植物向性运动以及叶片的伸展与定位? PID和PHOT都属于AGC激酶VIII亚族成员(Ding et al., 2011; Willige et al., 2013), NRL家族蛋白是否可以与AGC激酶VIII亚家族其它成员相互作用, 以调节生长素介导的其它生理反应? 来自分支IV的NRL2也可以与PHOT1发生互作(Sullivan et al., 2009), 暗示该分支的蛋白也可能在向光素介导的蓝光信号途径中发挥重要作用.综上, NRL家族蛋白功能的陆续鉴定, 有望解析向光素PHOT1和PHOT2感受蓝光信号差异, 进而调节不同生理反应的可能机制. ...
Role of RPT2 in leaf positioning and flattening and a possible inhibition of phot2 signaling by phot1 9 2013
... 叶片是植物进行光合作用的场所.自然光下, 野生型拟南芥叶柄向上倾斜生长, 叶片的叶面平整, 几乎平行于水平方向生长, 该现象被称为叶片的伸展与定位.弱光下, 拟南芥phot2突变体叶片伸展与定位表型几乎与野生型相同, phot1突变体叶片的定位发生变化, 叶柄与水平面夹角减小(Harada et al., 2013), 表明弱光下PHOT1单独调节叶片的伸展与定位(Demarsy and Fankhauser, 2009).强光下, 拟南芥phot1和phot2单突变体叶片伸展与定位的表型与野生型基本一致, phot1/phot2双突变体叶柄略微向下倾斜生长, 叶片向背面卷曲, 以几乎垂直于水平面的角度悬挂于叶柄上, 表明PHOT1和PHOT2以功能冗余的方式调节强光下叶片的伸展与定位(Sakai et al., 2001).叶片的伸展与定位可以确保叶片采取最佳的茎叶角度, 以便最大程度地捕获光源并且趋避强光伤害(Harada et al., 2013; Shang et al., 2019).与此一致, 拟南芥的研究表明: 弱光下, phot2的鲜重与野生型几乎相等, phot1的鲜重是野生型的1/2; 强光下, phot1和phot2单突变体的鲜重与野生型基本一致, 但phot1/phot2双突变体的鲜重大约是野生型的1/3 (Harada et al., 2013). ...
... ).叶片的伸展与定位可以确保叶片采取最佳的茎叶角度, 以便最大程度地捕获光源并且趋避强光伤害(Harada et al., 2013; Shang et al., 2019).与此一致, 拟南芥的研究表明: 弱光下, phot2的鲜重与野生型几乎相等, phot1的鲜重是野生型的1/2; 强光下, phot1和phot2单突变体的鲜重与野生型基本一致, 但phot1/phot2双突变体的鲜重大约是野生型的1/3 (Harada et al., 2013). ...
... 双突变体的鲜重大约是野生型的1/3 (Harada et al., 2013). ...
... Harada等(2013)在研究拟南芥叶片的伸展与定位时发现, 无论是在弱光、中强光还是强光条件下, phot1/rpt2双突变体叶片的表型均与phot1单突变体一致, 证明RPT2参与POHT1介导的叶片伸展与定位调节.与此一致, 他们还发现, phot2/rpt2双突变体与phot1/phot2双突变体表型类似(Harada et al., 2013).蛋白互作研究结果表明, PHOT1与RPT2在体内和体外均可发生相互作用, 但是未发现PHOT2和RPT2之间存在相互作用(Inada et al., 2004).此外, 叶片伸展与定位表型研究表明, phot1单突变体表型与phot2/rpt2双突变体不同, 但phot2/rpt2双突变体与phot1/phot2双突变体表型相似(Harada et al., 2013).上述结果充分证明, RPT2是作为PHOT1的下游因子(而不是PHOT2的下游因子)参与叶片伸展与定位调节.有趣的是, rpt2单突变体与phot1/phot2双突变体的表型类似(Harada et al., 2013), 说明在rpt2突变体中, PHOT2信号通路未被启动, 这暗示RPT2蛋白对于维持PHOT2信号通路的激活可能是必需的.当rpt2单突变体中PHOT1基因突变后, phot1/rpt2双突变体的表型与phot1单突变体类似, 说明依赖于PHOT2的调控途径被释放, 突变phot1/rpt2双突变体的PHOT2基因后, PHOT2调控途径随之缺失(Harada et al., 2013).上述结果暗示, RPT2蛋白对于PHOT2信号通路的启动至关重要, 该过程可能被依赖于PHOT1的途径所抑制, 但目前关于RPT2在PHOT2介导的信号途径中的具体作用机制还不清楚. ...
... 双突变体表型类似(Harada et al., 2013).蛋白互作研究结果表明, PHOT1与RPT2在体内和体外均可发生相互作用, 但是未发现PHOT2和RPT2之间存在相互作用(Inada et al., 2004).此外, 叶片伸展与定位表型研究表明, phot1单突变体表型与phot2/rpt2双突变体不同, 但phot2/rpt2双突变体与phot1/phot2双突变体表型相似(Harada et al., 2013).上述结果充分证明, RPT2是作为PHOT1的下游因子(而不是PHOT2的下游因子)参与叶片伸展与定位调节.有趣的是, rpt2单突变体与phot1/phot2双突变体的表型类似(Harada et al., 2013), 说明在rpt2突变体中, PHOT2信号通路未被启动, 这暗示RPT2蛋白对于维持PHOT2信号通路的激活可能是必需的.当rpt2单突变体中PHOT1基因突变后, phot1/rpt2双突变体的表型与phot1单突变体类似, 说明依赖于PHOT2的调控途径被释放, 突变phot1/rpt2双突变体的PHOT2基因后, PHOT2调控途径随之缺失(Harada et al., 2013).上述结果暗示, RPT2蛋白对于PHOT2信号通路的启动至关重要, 该过程可能被依赖于PHOT1的途径所抑制, 但目前关于RPT2在PHOT2介导的信号途径中的具体作用机制还不清楚. ...
... 双突变体表型相似(Harada et al., 2013).上述结果充分证明, RPT2是作为PHOT1的下游因子(而不是PHOT2的下游因子)参与叶片伸展与定位调节.有趣的是, rpt2单突变体与phot1/phot2双突变体的表型类似(Harada et al., 2013), 说明在rpt2突变体中, PHOT2信号通路未被启动, 这暗示RPT2蛋白对于维持PHOT2信号通路的激活可能是必需的.当rpt2单突变体中PHOT1基因突变后, phot1/rpt2双突变体的表型与phot1单突变体类似, 说明依赖于PHOT2的调控途径被释放, 突变phot1/rpt2双突变体的PHOT2基因后, PHOT2调控途径随之缺失(Harada et al., 2013).上述结果暗示, RPT2蛋白对于PHOT2信号通路的启动至关重要, 该过程可能被依赖于PHOT1的途径所抑制, 但目前关于RPT2在PHOT2介导的信号途径中的具体作用机制还不清楚. ...
... 双突变体的表型类似(Harada et al., 2013), 说明在rpt2突变体中, PHOT2信号通路未被启动, 这暗示RPT2蛋白对于维持PHOT2信号通路的激活可能是必需的.当rpt2单突变体中PHOT1基因突变后, phot1/rpt2双突变体的表型与phot1单突变体类似, 说明依赖于PHOT2的调控途径被释放, 突变phot1/rpt2双突变体的PHOT2基因后, PHOT2调控途径随之缺失(Harada et al., 2013).上述结果暗示, RPT2蛋白对于PHOT2信号通路的启动至关重要, 该过程可能被依赖于PHOT1的途径所抑制, 但目前关于RPT2在PHOT2介导的信号途径中的具体作用机制还不清楚. ...
... 基因后, PHOT2调控途径随之缺失(Harada et al., 2013).上述结果暗示, RPT2蛋白对于PHOT2信号通路的启动至关重要, 该过程可能被依赖于PHOT1的途径所抑制, 但目前关于RPT2在PHOT2介导的信号途径中的具体作用机制还不清楚. ...
... 近年来, 科学家研究发现, 拟南芥NRL家族蛋白在向光素介导的蓝光信号转导通路中起重要作用.拟南芥NRL家族6个分支中的3个分支(II、III和IV) (图1)蛋白目前已经被报道参与向光素调节的反应, 或者与向光素蛋白存在直接相互作用, 暗示这3个分支的其它成员可能参与向光素信号途径的调节.RPT2与NCH1都来自分支II, 既参与生长素依赖的向光性调节, 又参与非生长素依赖的叶绿体聚光运动调节(Suetsugu et al., 2016; Suetsugu and Wada, 2017).目前RPT2和NCH1如何区分向光素介导的2种信号途径并不清楚.考虑到RPT2和NCH1蛋白所在分支与其余2个分支(III和IV)的亲缘关系较远(Christie et al., 2018), 暗示该分支其它成员也可能同时参与生长素依赖和非依赖性的向光素反应.RPT2可与PHOT1的LOV1区相互作用并抑制PHOT1的自磷酸化, 以降低PHOT1的光敏感性.尽管RPT2也可以与PHOT2的LOV1区域相互作用, 但是RPT2不抑制PHOT2的活性(Kimura et al., 2020).可能存在NRL家族其它蛋白与PHOT2相互作用, 调节PHOT2介导的蓝光信号转导.NPH3主要调节生长素依赖的向光性反应以及叶片的伸展与定位(Harada et al., 2013; Zhao et al., 2018), 但NPH3激活后调节生长素不对称分布的机制并不清楚.NPH3的磷酸化状态与其生理活性以及在细胞中的定位密切相关(Haga et al., 2015; Zhao et al., 2018).PHOT1介导的NPH3去磷酸化在弱蓝光诱导的下胚轴向光弯曲反应中发挥重要作用, 强蓝光下NPH3并未发生去磷酸化反应(Tsuchida-Mayama et al., 2008).那么NPH3如何调节强蓝光诱导的下胚轴向光弯曲反应, 是否与其E3泛素化连接酶活性有关? NPY1与PID互作, 通过影响生长素信号调节植物器官的形成以及根的向重力性反应(Suetsugu et al., 2016).生长素不对称分布决定不对称生长, PID- NPY1与PHOT-NPH3作用方式又相似, 那么NPY分支成员是否参与调节向光素介导的植物向性运动以及叶片的伸展与定位? PID和PHOT都属于AGC激酶VIII亚族成员(Ding et al., 2011; Willige et al., 2013), NRL家族蛋白是否可以与AGC激酶VIII亚家族其它成员相互作用, 以调节生长素介导的其它生理反应? 来自分支IV的NRL2也可以与PHOT1发生互作(Sullivan et al., 2009), 暗示该分支的蛋白也可能在向光素介导的蓝光信号途径中发挥重要作用.综上, NRL家族蛋白功能的陆续鉴定, 有望解析向光素PHOT1和PHOT2感受蓝光信号差异, 进而调节不同生理反应的可能机制. ...
The role of stomata in sensing and driving environmental change 1 2003
... 拟南芥气孔由1对保卫细胞构成, 气孔通过调节其开闭程度控制着与环境的气体交换来响应外界的刺激信号(Hetherington and Woodward, 2003; Roelfsema and Hedrich, 2005).红光和蓝光都能诱导气孔快速张开(Shimazaki et al., 2007).红光通过影响叶肉细胞和保卫细胞中叶绿体的光合作用调节气孔开放(Mott et al., 2008; Suetsugu et al., 2014).蓝光作为信号通过激活保卫细胞质膜H+-ATPase诱导气孔开放(Assmann et al., 1985; Kinoshita et al., 2001).蓝光诱导的气孔开放受蓝光受体向光素(PHOT1和PHOT2)调节(Kinoshita et al., 2001).目前人们对向光素介导的气孔开放机制研究较为清楚(Inoue et al., 2008b; Inoue and Kinoshita, 2017).蓝光照射下, 自磷酸化激活的PHOT1直接磷酸化蛋白激酶BLUS1 (LIGHT SIGNALING1), 磷酸化的BLUS1间接将信号传递给I型蛋白磷酸酶(PPI)以及该酶的亚基PRSL1 (Takemiya and Shimazaki, 2016).PPI进一步将信号传递给质膜AHA1型H+-ATPase, 促进H+-ATPase的激活.H+-ATPase的激活需要其C末端倒数第2个苏氨酸的磷酸化以及与14-3-3蛋白的结合(Yamauchi et al., 2016).活化的H+-ATPase通过驱动保卫细胞内H+的跨膜输出诱导保卫细胞质膜的超极化(Shimazaki et al., 2007; Marten et al., 2010).超极化的质膜激活内向K+通道, 促进K+在保卫细胞胞质中积累.为了维持胞质的电荷平衡, Cl-和malate2-随之大量增加, 增加的K+、Cl-和malate2-进一步被转入液泡, 使保卫细胞水势降低, 外界水分大量进入保卫细胞, 促进保卫细胞体积膨胀, 气孔张开(Inoue and Kinoshita, 2017).保卫细胞胞质中Cl-的增加主要是由于向光素抑制了S型阴离子外流蛋白的活性(Sussmilch et al., 2019).向光素对S型阴离子外流蛋白活性的抑制需要2个蛋白激酶CBC1/2 (CONVERGENCE OF BLUE LIGHT (BL) and CO21/2)参与(Hiyama et al., 2017; Sussmilch et al., 2019). ...
Plant nuclear photorelocation movement 1 2014
... 拟南芥(Arabidopsis thaliana) PHOT1和PHOT2以功能冗余的方式调节下胚轴向光性和根负向光性、叶绿体聚光运动、气孔开放以及叶片伸展与定位等(Christie et al., 2015).PHOT1特异性抑制光下黄化幼苗下胚轴的快速伸长(Folta et al., 2003)、子叶变绿过程中叶绿素结合蛋白mRNA转录的稳定性(Folta and Kaufman, 2003)以及侧根的快速生长(Moni et al., 2015).PHOT2不仅介导强蓝光诱导的下胚轴向光弯曲(Sakai et al., 2001), 而且也特异调节叶绿体和细胞核的避光运动(Higa et al., 2014)以及抗性蛋白介导的病毒防御反应(Jeong et al., 2010).PHOT1和PHOT2的功能差异可能与磷酸化修饰位点、磷酸化程度、亚细胞定位变化以及下游相互作用蛋白种类不同有关(Zhao et al., 2013; Morrow et al., 2018; Liscum et al., 2020). ...
Blue light and CO2 signals converge to regulate light-induced stomatal opening 1 2017
... 拟南芥气孔由1对保卫细胞构成, 气孔通过调节其开闭程度控制着与环境的气体交换来响应外界的刺激信号(Hetherington and Woodward, 2003; Roelfsema and Hedrich, 2005).红光和蓝光都能诱导气孔快速张开(Shimazaki et al., 2007).红光通过影响叶肉细胞和保卫细胞中叶绿体的光合作用调节气孔开放(Mott et al., 2008; Suetsugu et al., 2014).蓝光作为信号通过激活保卫细胞质膜H+-ATPase诱导气孔开放(Assmann et al., 1985; Kinoshita et al., 2001).蓝光诱导的气孔开放受蓝光受体向光素(PHOT1和PHOT2)调节(Kinoshita et al., 2001).目前人们对向光素介导的气孔开放机制研究较为清楚(Inoue et al., 2008b; Inoue and Kinoshita, 2017).蓝光照射下, 自磷酸化激活的PHOT1直接磷酸化蛋白激酶BLUS1 (LIGHT SIGNALING1), 磷酸化的BLUS1间接将信号传递给I型蛋白磷酸酶(PPI)以及该酶的亚基PRSL1 (Takemiya and Shimazaki, 2016).PPI进一步将信号传递给质膜AHA1型H+-ATPase, 促进H+-ATPase的激活.H+-ATPase的激活需要其C末端倒数第2个苏氨酸的磷酸化以及与14-3-3蛋白的结合(Yamauchi et al., 2016).活化的H+-ATPase通过驱动保卫细胞内H+的跨膜输出诱导保卫细胞质膜的超极化(Shimazaki et al., 2007; Marten et al., 2010).超极化的质膜激活内向K+通道, 促进K+在保卫细胞胞质中积累.为了维持胞质的电荷平衡, Cl-和malate2-随之大量增加, 增加的K+、Cl-和malate2-进一步被转入液泡, 使保卫细胞水势降低, 外界水分大量进入保卫细胞, 促进保卫细胞体积膨胀, 气孔张开(Inoue and Kinoshita, 2017).保卫细胞胞质中Cl-的增加主要是由于向光素抑制了S型阴离子外流蛋白的活性(Sussmilch et al., 2019).向光素对S型阴离子外流蛋白活性的抑制需要2个蛋白激酶CBC1/2 (CONVERGENCE OF BLUE LIGHT (BL) and CO21/2)参与(Hiyama et al., 2017; Sussmilch et al., 2019). ...
RPT2 is a signal transducer involved in phototropic response and stomatal opening by association with phototropin 1 in Arabidopsis thaliana 11 2004
... 拟南芥中ABCB19 (ATP-BINDING CAS-SETTE B 19)和光敏色素激酶底物(PHYTOCHROME KINASE SUBSTRATE 4, PKS4)参与调节生长素的转运, 是下胚轴向光弯曲的负调节因子(Christie et al., 2011; Demarsy et al., 2012).自磷酸化激活的PHOT1通过直接磷酸化ABCB19和PKS4, 抑制ABCB19和PKS4的生长素转运活性, 促进下胚轴向光弯曲反应(Christie et al., 2011; Demarsy et al., 2012).PHOT1通过磷酸化修饰调节ABCB19的活性, 但是PHOT2并不影响ABCB19的活性(Christie et al., 2011).蛋白磷酸酶2A (PP2A)的A1亚基对应的蛋白RCN1 (Root Curling in N-naphthylphthalamic acid 1)与PHOT2蛋白发生体外互作, 负调节PHOT2介导的向光弯曲, 但RCN1并不与PHOT1直接相互作用(Tseng and Briggs, 2010).NPH3 (NONPHOTOTROPIC HYPOCOTYL 3)和RPT2 (ROOT PHOTOTROPISM 2)是通过筛选拟南芥下胚轴以及根的向光性缺失突变体获得(Motchoulski and Liscum, 1999; Sakai et al., 2000).随后的研究发现, NPH3和RPT2是向光素途径的重要调控因子, 均可与PHOT1蛋白直接相互作用(Blakeslee et al., 2004; Stone et al., 2008).虽然NPH3和RPT2也参与PHOT2信号转导途径, 但目前并无RPT2与PHOT2蛋白之间直接相互作用的报道(Inada et al., 2004; Lariguet et al., 2006).上述结果暗示PHOT1和PHOT2可能通过与不同的下游互作因子差异调节蓝光反应. ...
... NRL (NPH3/RPT2-Like)蛋白家族是植物特有的, 其成员含有3个保守结构域: N端的BTB (Bric àbrac, Tramtrack, and Broad complex)结构域、中间的NPH3结构域以及C端的螺旋-螺旋结构域(Liscum et al., 2014).其中, BTB结构域主要起E3连接酶的作用, 与CUL3发生相互作用(Gingerich et al., 2005; Figueroa et al., 2005), C端的螺旋-螺旋结构域主要介导蛋白之间的相互作用(Inada et al., 2004).拟南芥中33个NRL家族成员都含有NPH3结构域, 10个缺少C端的螺旋-螺旋结构域, 2个缺少N端的BTB结构域(Pedmale et al., 2010; Christie et al., 2018).根据NRL家族成员间的进化关系, 拟南芥中的33个NRL家族成员可分为6个分支(图1) (Suetsugu et al., 2016; Christie et al., 2018).目前, 来自2个分支的10个成员的功能已得到鉴定(de Carbonnel et al., 2010; Christie et al., 2018) (图1). ...
... 目前已经鉴定的10个有功能的NRL家族蛋白中有3个(NPH3、RPT2和NCH1)可与向光素发生直接相互作用(Motchoulski and Liscum, 1999; Inada et al., 2004; Suetsugu et al., 2016).此外, 酵母双杂交筛选发现NRL2可以与PHOT1发生相互作用(Sullivan et al., 2009), 但是NRL2的功能及其在向光素介导的信号途径中的作用目前还不清楚.虽然大部分NRL家族成员的功能未知, 但已有的实验结果表明, NRL家族成员在向光素信号途径中起重要作用, 因此我们总结NRL家族蛋白在向光素介导的信号途径中的作用, 旨在为全面揭示NPH3/RPT2-Like家族成员的功能提供线索. ...
... 早在达尔文(1880年)时期, 人们就已发现植物向光性, 但是直到向光素PHOT1和PHOT2先后被鉴定(Liscum and Briggs, 1995; Christie et al., 1998), 植物的向光性机理研究才取得突飞猛进的发展.研究发现, PHOT1调节弱蓝光(0.01-1 μmol·m-2·s-1)和强蓝光(>1 μmol·m-2·s-1)诱导的下胚轴的向光性; PHOT2仅在强蓝光下起作用, 且与PHOT1功能冗余(Sakai et al., 2001; Briggs and Christie, 2002).由于强蓝光下PHOT1和PHOT2的功能冗余, 目前向光性机制研究主要集中在PHOT1介导的弱蓝光反应(Motchoulski and Liscum, 1999; Haga et al., 2015), 仅有少量研究通过突变体背景下的抑制子筛选寻找参与强蓝光信号途径的作用因子(赵翔等, 2014).单侧蓝光照射下, 自磷酸化激活的向光素蛋白将信号传递给NPH3、RPT2以及PKS等下游因子(Motchoulski and Liscum, 1999; Inada et al., 2004), 调节生长素运输载体的活性与定位(Ding et al., 2011).例如, 引起内皮层细胞背光侧PIN3 (PIN-FORMED3)积累(刘广超和丁兆军, 2018), 促进背光侧生长素含量增加、细胞伸长速率加快, 最终引起植物弯向光生长(Christie and Murphy, 2013).下胚轴弯曲过程中生长素的不对称分布受各类生长素运输载体的共同调控, 如PINs (PIN3、PIN4和PIN7)和ABCB19 (Christie et al., 2011).蓝光下, PIN3的磷酸化受抑制, 促进PIN3的背光侧定位, 诱导背光侧生长素积累以及下胚轴的向光性生长(Ding et al., 2011).PIN3的磷酸化以及极性定位受AGC激酶VIII亚族成员PID (PINOID)以及D6PK (D6 PROTEIN KINASE)的调节(Ding et al., 2011; Willige et al., 2013).ABCB19是生长素外流载体, 被蓝光激活的PHOT1磷酸化后, ABCB19的生长素转运活性受抑制, 增加拟南芥下胚轴顶端生长素含量并抑制其垂直生长, 同时通过PIN3蛋白促进生长素在下胚轴中侧向移动(Christie et al., 2011).背光侧生长素的大量积累促进了背光侧IAA19的降解, 释放出有活性的ARF7 (Sun et al., 2013), 不对称激活与生长相关基因的表达促进了下胚轴的向光性反应(刘广超和丁兆军, 2018). ...
... 拟南芥rpt2单突变体在极弱光下(≤0.01 μmol·m-2·s-1)下胚轴向光弯曲反应正常, 随着光照强度的增大弯曲度逐渐降低(Inada et al., 2004), 表明RPT2仅参与调节强蓝光诱导的下胚轴向光弯曲反应.phot1/rpt2双突变体与phot1单突变体表型类似, 在10和100 μmol·m-2·s-1的较强蓝光下可以发生下胚轴向光弯曲反应, 但在0.01-1 μmol·m-2·s-1的弱蓝光下则缺失向光弯曲(Sakai et al., 2000), 暗示RPT2作为PHOT1的下游因子调节强蓝光诱导的下胚轴向光弯曲反应.phot2/rpt2双突变体与phot1/phot2双突变体的表型一致, 缺失强蓝光诱导的下胚轴向光弯曲反应(Inada et al., 2004; Zhao et al., 2018), 且PHOT1和RPT2可以直接相互作用.上述结果进一步证明, RPT2确实参与调节PHOT1介导的下胚轴向光弯曲反应.如果RPT2仅作为PHOT1的下游因子发挥作用, 那么rpt2单突变体应该与phot1单突变体表型一致.表型分析结果显示, rpt2单突变体与phot1/phot2双突变体的表型一致, 即缺失强蓝光诱导的下胚轴向光弯曲反应(Inada et al., 2004), 证明RPT2在PHOT2介导的下胚轴向光弯曲反应中也发挥重要作用.Harada等(2013)认为, RPT2在PHOT1抑制PHOT2途径的过程中发挥作用, 阻碍PHOT1对PHOT2途径的抑制.最近研究表明, RPT2可与PHOT1的LOV1区相互作用并抑制PHOT1的自磷酸化, 以降低PHOT1的光敏感性, 从而行使分子变阻器(molecular rheostat)的功能, 将不同强度蓝光下PHOT1的活性维持在适当的水平.尽管RPT2也可以与PHOT2的LOV1区域相互作用, 但是RPT2不抑制PHOT2的活性(Kimura et al., 2020). ...
... 双突变体的表型一致, 缺失强蓝光诱导的下胚轴向光弯曲反应(Inada et al., 2004; Zhao et al., 2018), 且PHOT1和RPT2可以直接相互作用.上述结果进一步证明, RPT2确实参与调节PHOT1介导的下胚轴向光弯曲反应.如果RPT2仅作为PHOT1的下游因子发挥作用, 那么rpt2单突变体应该与phot1单突变体表型一致.表型分析结果显示, rpt2单突变体与phot1/phot2双突变体的表型一致, 即缺失强蓝光诱导的下胚轴向光弯曲反应(Inada et al., 2004), 证明RPT2在PHOT2介导的下胚轴向光弯曲反应中也发挥重要作用.Harada等(2013)认为, RPT2在PHOT1抑制PHOT2途径的过程中发挥作用, 阻碍PHOT1对PHOT2途径的抑制.最近研究表明, RPT2可与PHOT1的LOV1区相互作用并抑制PHOT1的自磷酸化, 以降低PHOT1的光敏感性, 从而行使分子变阻器(molecular rheostat)的功能, 将不同强度蓝光下PHOT1的活性维持在适当的水平.尽管RPT2也可以与PHOT2的LOV1区域相互作用, 但是RPT2不抑制PHOT2的活性(Kimura et al., 2020). ...
... 双突变体的表型一致, 即缺失强蓝光诱导的下胚轴向光弯曲反应(Inada et al., 2004), 证明RPT2在PHOT2介导的下胚轴向光弯曲反应中也发挥重要作用.Harada等(2013)认为, RPT2在PHOT1抑制PHOT2途径的过程中发挥作用, 阻碍PHOT1对PHOT2途径的抑制.最近研究表明, RPT2可与PHOT1的LOV1区相互作用并抑制PHOT1的自磷酸化, 以降低PHOT1的光敏感性, 从而行使分子变阻器(molecular rheostat)的功能, 将不同强度蓝光下PHOT1的活性维持在适当的水平.尽管RPT2也可以与PHOT2的LOV1区域相互作用, 但是RPT2不抑制PHOT2的活性(Kimura et al., 2020). ...
... Harada等(2013)在研究拟南芥叶片的伸展与定位时发现, 无论是在弱光、中强光还是强光条件下, phot1/rpt2双突变体叶片的表型均与phot1单突变体一致, 证明RPT2参与POHT1介导的叶片伸展与定位调节.与此一致, 他们还发现, phot2/rpt2双突变体与phot1/phot2双突变体表型类似(Harada et al., 2013).蛋白互作研究结果表明, PHOT1与RPT2在体内和体外均可发生相互作用, 但是未发现PHOT2和RPT2之间存在相互作用(Inada et al., 2004).此外, 叶片伸展与定位表型研究表明, phot1单突变体表型与phot2/rpt2双突变体不同, 但phot2/rpt2双突变体与phot1/phot2双突变体表型相似(Harada et al., 2013).上述结果充分证明, RPT2是作为PHOT1的下游因子(而不是PHOT2的下游因子)参与叶片伸展与定位调节.有趣的是, rpt2单突变体与phot1/phot2双突变体的表型类似(Harada et al., 2013), 说明在rpt2突变体中, PHOT2信号通路未被启动, 这暗示RPT2蛋白对于维持PHOT2信号通路的激活可能是必需的.当rpt2单突变体中PHOT1基因突变后, phot1/rpt2双突变体的表型与phot1单突变体类似, 说明依赖于PHOT2的调控途径被释放, 突变phot1/rpt2双突变体的PHOT2基因后, PHOT2调控途径随之缺失(Harada et al., 2013).上述结果暗示, RPT2蛋白对于PHOT2信号通路的启动至关重要, 该过程可能被依赖于PHOT1的途径所抑制, 但目前关于RPT2在PHOT2介导的信号途径中的具体作用机制还不清楚. ...
... 虽然目前人们还不清楚向光素在感受外界光照后如何通过调节CHUP1、PMI1、WEB1以及JAC蛋白活性影响叶绿体的运动, 但越来越多的证据表明, NRL家族蛋白的NCH1和RPT2是该信号途径的重要中间因子(Suetsugu et al., 2016).rpt2突变体rpt2-3和rpt2-4的聚光反应略微降低, 但是叶绿体的避光反应和暗恢复反应(撤掉强光后, 叶绿体快速恢复, 导致透光率迅速下降的反应称为暗恢复反应)与野生型相比无明显差异(Suetsugu et al., 2016), 这些结果表明RPT2参与调节叶绿体的聚光反应.nch1单突变体的叶绿体聚光反应减弱, 暗恢复反应缺失, 但是避光反应增强(Suetsugu et al., 2016), 证明NCH1参与调节叶绿体的聚光运动.目前已经证实RPT2和NCH1参与调节弱光下PHOT1和PHOT2介导的叶绿体聚光反应, 这种AGC-NRL调节模块不依赖于生长素的转运以及生长素介导的转录过程(Suetsugu et al., 2016), 可能依赖于JAC1 (J-DOMAIN PROTEIN REQUIRED FOR CHLOROPLAST ACCUMULATION RESPONSE 1)以及WEB1/PMI2介导的信号途径(Suetsugu and Wada, 2017).弱光下, RPT2/ NCH1通过JAC1依赖与JAC1不依赖途径抑制弱光下的叶绿体避光反应.强光下, WEB1/PMI2复合体通过抑制RPT2/NCH1和JAC1介导的聚光反应, 有效促进PHOT2介导的叶绿体避光反应(Suetsugu and Wada, 2017), 但是同一家族的NPH3并不调节叶绿体的运动(Inada et al., 2004). ...
Blue light regulation of stomatal opening and the plasma membrane H+-ATPase 2 2017
... 拟南芥气孔由1对保卫细胞构成, 气孔通过调节其开闭程度控制着与环境的气体交换来响应外界的刺激信号(Hetherington and Woodward, 2003; Roelfsema and Hedrich, 2005).红光和蓝光都能诱导气孔快速张开(Shimazaki et al., 2007).红光通过影响叶肉细胞和保卫细胞中叶绿体的光合作用调节气孔开放(Mott et al., 2008; Suetsugu et al., 2014).蓝光作为信号通过激活保卫细胞质膜H+-ATPase诱导气孔开放(Assmann et al., 1985; Kinoshita et al., 2001).蓝光诱导的气孔开放受蓝光受体向光素(PHOT1和PHOT2)调节(Kinoshita et al., 2001).目前人们对向光素介导的气孔开放机制研究较为清楚(Inoue et al., 2008b; Inoue and Kinoshita, 2017).蓝光照射下, 自磷酸化激活的PHOT1直接磷酸化蛋白激酶BLUS1 (LIGHT SIGNALING1), 磷酸化的BLUS1间接将信号传递给I型蛋白磷酸酶(PPI)以及该酶的亚基PRSL1 (Takemiya and Shimazaki, 2016).PPI进一步将信号传递给质膜AHA1型H+-ATPase, 促进H+-ATPase的激活.H+-ATPase的激活需要其C末端倒数第2个苏氨酸的磷酸化以及与14-3-3蛋白的结合(Yamauchi et al., 2016).活化的H+-ATPase通过驱动保卫细胞内H+的跨膜输出诱导保卫细胞质膜的超极化(Shimazaki et al., 2007; Marten et al., 2010).超极化的质膜激活内向K+通道, 促进K+在保卫细胞胞质中积累.为了维持胞质的电荷平衡, Cl-和malate2-随之大量增加, 增加的K+、Cl-和malate2-进一步被转入液泡, 使保卫细胞水势降低, 外界水分大量进入保卫细胞, 促进保卫细胞体积膨胀, 气孔张开(Inoue and Kinoshita, 2017).保卫细胞胞质中Cl-的增加主要是由于向光素抑制了S型阴离子外流蛋白的活性(Sussmilch et al., 2019).向光素对S型阴离子外流蛋白活性的抑制需要2个蛋白激酶CBC1/2 (CONVERGENCE OF BLUE LIGHT (BL) and CO21/2)参与(Hiyama et al., 2017; Sussmilch et al., 2019). ...
... 进一步被转入液泡, 使保卫细胞水势降低, 外界水分大量进入保卫细胞, 促进保卫细胞体积膨胀, 气孔张开(Inoue and Kinoshita, 2017).保卫细胞胞质中Cl-的增加主要是由于向光素抑制了S型阴离子外流蛋白的活性(Sussmilch et al., 2019).向光素对S型阴离子外流蛋白活性的抑制需要2个蛋白激酶CBC1/2 (CONVERGENCE OF BLUE LIGHT (BL) and CO21/2)参与(Hiyama et al., 2017; Sussmilch et al., 2019). ...
Blue light-induced autophosphorylation of phototropin is a primary step for signaling 1 2008
... 拟南芥PHOT1至少有21个氨基酸位点可以发生自磷酸化(Boex-Fontvieille et al., 2014; Deng et al., 2014), PHOT2有29个磷酸化位点(Inoue et al., 2011; Boex-Fontvieille et al., 2014), 这些位点的具体分布参见Christie等(2015)的报道.其中, C端2个保守位点的磷酸化(PHOT1中S849和S851, PHOT2中S761 和S763)对向光素受体的信号传递至关重要(Inoue et al., 2008a; Inoue et al., 2011), 其余的磷酸化位点大部分位于N端LOV1的上游以及LOV1与LOV2中间的连接区域(Christie et al., 2015).虽然这些上游区域磷酸化位点的生物学意义目前还不清楚, 但这些位点的磷酸化程度受光照强度调节(Salomon et al., 2003), 暗示PHOT1和PHOT2可能通过磷酸化程度差异响应不同的光照强度, 从而参与植物多种生理活动的调节. ...
Leaf positioning of Arabidopsis in response to blue light 2 2008
... 植物向光素PHOT1 (PHOTOTROPIN1)和PHOT2是一类细胞质膜结合的Ser/Thr蛋白激酶, 其C端为激酶结构域, N端含有2个光、氧气和电压敏感的LOV (LIGHT, OXYGEN, OR VOLTAGE SENSING)结构域, 分别命名为LOV1和LOV2 (Christie et al., 2015).蓝光照射后诱导向光素蛋白发生构象改变及自我磷酸化激活(Pfeifer et al., 2010), 活化的向光素蛋白进而调节下胚轴向光弯曲反应(Sakai et al., 2001)、叶绿体运动(Kagawa et al., 2001)、气孔开放(Kinoshita et al., 2001)以及叶片的伸展与定位等(Sakamoto and Briggs, 2002; Inoue et al., 2008b), 从而使植物或叶片处于最有利于吸收光的位置, 优化植物的光合作用(Christie et al., 2015). ...
... 拟南芥气孔由1对保卫细胞构成, 气孔通过调节其开闭程度控制着与环境的气体交换来响应外界的刺激信号(Hetherington and Woodward, 2003; Roelfsema and Hedrich, 2005).红光和蓝光都能诱导气孔快速张开(Shimazaki et al., 2007).红光通过影响叶肉细胞和保卫细胞中叶绿体的光合作用调节气孔开放(Mott et al., 2008; Suetsugu et al., 2014).蓝光作为信号通过激活保卫细胞质膜H+-ATPase诱导气孔开放(Assmann et al., 1985; Kinoshita et al., 2001).蓝光诱导的气孔开放受蓝光受体向光素(PHOT1和PHOT2)调节(Kinoshita et al., 2001).目前人们对向光素介导的气孔开放机制研究较为清楚(Inoue et al., 2008b; Inoue and Kinoshita, 2017).蓝光照射下, 自磷酸化激活的PHOT1直接磷酸化蛋白激酶BLUS1 (LIGHT SIGNALING1), 磷酸化的BLUS1间接将信号传递给I型蛋白磷酸酶(PPI)以及该酶的亚基PRSL1 (Takemiya and Shimazaki, 2016).PPI进一步将信号传递给质膜AHA1型H+-ATPase, 促进H+-ATPase的激活.H+-ATPase的激活需要其C末端倒数第2个苏氨酸的磷酸化以及与14-3-3蛋白的结合(Yamauchi et al., 2016).活化的H+-ATPase通过驱动保卫细胞内H+的跨膜输出诱导保卫细胞质膜的超极化(Shimazaki et al., 2007; Marten et al., 2010).超极化的质膜激活内向K+通道, 促进K+在保卫细胞胞质中积累.为了维持胞质的电荷平衡, Cl-和malate2-随之大量增加, 增加的K+、Cl-和malate2-进一步被转入液泡, 使保卫细胞水势降低, 外界水分大量进入保卫细胞, 促进保卫细胞体积膨胀, 气孔张开(Inoue and Kinoshita, 2017).保卫细胞胞质中Cl-的增加主要是由于向光素抑制了S型阴离子外流蛋白的活性(Sussmilch et al., 2019).向光素对S型阴离子外流蛋白活性的抑制需要2个蛋白激酶CBC1/2 (CONVERGENCE OF BLUE LIGHT (BL) and CO21/2)参与(Hiyama et al., 2017; Sussmilch et al., 2019). ...
Functional analyses of the activation loop of phototropin2 in Arabidopsis 2 2011
... 拟南芥PHOT1至少有21个氨基酸位点可以发生自磷酸化(Boex-Fontvieille et al., 2014; Deng et al., 2014), PHOT2有29个磷酸化位点(Inoue et al., 2011; Boex-Fontvieille et al., 2014), 这些位点的具体分布参见Christie等(2015)的报道.其中, C端2个保守位点的磷酸化(PHOT1中S849和S851, PHOT2中S761 和S763)对向光素受体的信号传递至关重要(Inoue et al., 2008a; Inoue et al., 2011), 其余的磷酸化位点大部分位于N端LOV1的上游以及LOV1与LOV2中间的连接区域(Christie et al., 2015).虽然这些上游区域磷酸化位点的生物学意义目前还不清楚, 但这些位点的磷酸化程度受光照强度调节(Salomon et al., 2003), 暗示PHOT1和PHOT2可能通过磷酸化程度差异响应不同的光照强度, 从而参与植物多种生理活动的调节. ...
... ; Inoue et al., 2011), 其余的磷酸化位点大部分位于N端LOV1的上游以及LOV1与LOV2中间的连接区域(Christie et al., 2015).虽然这些上游区域磷酸化位点的生物学意义目前还不清楚, 但这些位点的磷酸化程度受光照强度调节(Salomon et al., 2003), 暗示PHOT1和PHOT2可能通过磷酸化程度差异响应不同的光照强度, 从而参与植物多种生理活动的调节. ...
Cryptochrome 2 and phototropin 2 regulate resistance protein-mediated viral defense by negatively regulating an E3 ubiquitin ligase 1 2010
... 拟南芥(Arabidopsis thaliana) PHOT1和PHOT2以功能冗余的方式调节下胚轴向光性和根负向光性、叶绿体聚光运动、气孔开放以及叶片伸展与定位等(Christie et al., 2015).PHOT1特异性抑制光下黄化幼苗下胚轴的快速伸长(Folta et al., 2003)、子叶变绿过程中叶绿素结合蛋白mRNA转录的稳定性(Folta and Kaufman, 2003)以及侧根的快速生长(Moni et al., 2015).PHOT2不仅介导强蓝光诱导的下胚轴向光弯曲(Sakai et al., 2001), 而且也特异调节叶绿体和细胞核的避光运动(Higa et al., 2014)以及抗性蛋白介导的病毒防御反应(Jeong et al., 2010).PHOT1和PHOT2的功能差异可能与磷酸化修饰位点、磷酸化程度、亚细胞定位变化以及下游相互作用蛋白种类不同有关(Zhao et al., 2013; Morrow et al., 2018; Liscum et al., 2020). ...
Short actin-based mechanism for light-directed chloroplast movement in Arabidopsis 1 2009
... 作为植物进行光合作用的细胞器, 叶绿体响应外界光照刺激而发生的位置变化称为叶绿体光下重定位.弱光照射下, 为最大程度地吸收光能, 叶绿体沿细胞平周壁方向聚集, 这种现象为聚光运动(accumulation movement).强光条件下, 为避免过强的光对自身造成伤害, 叶绿体沿细胞垂周壁聚集, 使之与光平行分布, 这种现象称为避光运动(avoidance movement) (Kasahara et al., 2004).叶绿体运动受蓝光调节, 蓝光受体向光素PHOT1和PHOT2共同调控叶绿体的聚光反应, 而PHOT2单独调节叶绿体的避光反应(Suetsugu and Wada, 2017).与向光素调节的其它多细胞协调性反应(如向光性、叶片的伸展与定位)不同, 叶绿体运动是细胞自主性反应, 不受基因表达(Kong and Wada, 2016; Wada, 2016)和生长素运输的调节(Christie et al., 2018), 最终由叶绿体肌动蛋白响应入射光的强度与方向而发生快速重组介导(Kadota et al., 2009; Kong and Wada, 2014).叶绿体肌动蛋白特异定位于细胞质膜和叶绿体之间, 在叶绿体前进方向的后面叶绿体肌动蛋白纤维迅速解聚, 前进方向的前面叶绿体肌动蛋白纤维迅速聚集, 这种不对称的叶绿体肌动蛋白纤维分布最终决定了叶绿体的运动方向和速度(Kong and Wada, 2016).CHUP1 (CHLOROPLAST UNUSUAL POSITIONING 1)和 ...
Arabidopsis NPL1: a phototropin homolog controlling the chloroplast high-light avoidance response 1 2001
... 植物向光素PHOT1 (PHOTOTROPIN1)和PHOT2是一类细胞质膜结合的Ser/Thr蛋白激酶, 其C端为激酶结构域, N端含有2个光、氧气和电压敏感的LOV (LIGHT, OXYGEN, OR VOLTAGE SENSING)结构域, 分别命名为LOV1和LOV2 (Christie et al., 2015).蓝光照射后诱导向光素蛋白发生构象改变及自我磷酸化激活(Pfeifer et al., 2010), 活化的向光素蛋白进而调节下胚轴向光弯曲反应(Sakai et al., 2001)、叶绿体运动(Kagawa et al., 2001)、气孔开放(Kinoshita et al., 2001)以及叶片的伸展与定位等(Sakamoto and Briggs, 2002; Inoue et al., 2008b), 从而使植物或叶片处于最有利于吸收光的位置, 优化植物的光合作用(Christie et al., 2015). ...
Domain swapping to assess the mechanistic basis of Arabidopsis phototropin 1 receptor kinase activation and endocytosis by blue light 1 2009
... 蓝光照射会诱导PHOT1和PHOT2亚细胞定位改变, PHOT1从细胞质膜脱离后向胞质移动(Kaiserli et al., 2009), 而PHOT2从细胞质膜脱离后向高尔基体转移, 最终与高尔基体偶联(Aggarwal et al., 2014).虽然向光素蛋白亚细胞定位受光照影响的生物学意义还不清楚, 但已知其亚细胞定位变化受PHOT1和PHOT2的激酶活性调节, 且蓝光诱导PHOT1的胞内定位变化(intracelluar position)参与下胚轴向光性调节(Wan et al., 2008; Zhao et al., 2018), 暗示向光素的亚细胞定位变化影响了向光素途径的信号传递, PHOT1和PHOT2亚细胞定位变化的不同可能是2种向光素受体差异调节植物蓝光反应的重要原因. ...
Reduced phototropism in pks mutants may be due to altered auxin-regulated gene expression or reduced lateral auxin transport 1 2014
... NPH3主要通过2种途径影响生长素的不对称分布.一是NPH3通过多泛素化修饰PHOT1促进PHOT1的降解, 间接影响生长素转运蛋白(如生长素外流载体PIN1 (Roberts et al., 2011)和PIN2 (Wan et al., 2012))的重定位.NPH3是以Cullin3为基础的E3泛素化连接酶CRL3NPH3复合体的组成成分, 在该复合体中作为底物连接蛋白发挥作用(Roberts et al., 2011; Wan et al., 2012).强蓝光下, CRL3NPH3复合体直接与PHOT1蛋白相互作用, 促进PHOT1的单/多泛素化修饰, 多泛素化修饰的PHOT1进入细胞质后被26S蛋白酶体识别后降解(Roberts et al., 2011).PHOT1从细胞膜进入细胞质影响了生长素外流载体PIN1 (Roberts et al., 2011)和PIN2 (Wan et al., 2012)的重定位, 但具体作用机制有待深入研究.二是NPH3通过与PKS家族蛋白互作调节生长素的转运(Fankhauser and Christie, 2015).PKS家族成员是向光弯曲反应的正向调节因子, 参与生长素转运调节, 它们可以与向光素和NPH3蛋白直接互作, 形成PHOT1-PKS-NPH3复合体(Kami et al., 2014), 但是复合体的作用机制目前还不清楚. ...
Phototropins mediate blue and red light-induced chloroplast movements in Physcomitrella patens 1 2004
... 作为植物进行光合作用的细胞器, 叶绿体响应外界光照刺激而发生的位置变化称为叶绿体光下重定位.弱光照射下, 为最大程度地吸收光能, 叶绿体沿细胞平周壁方向聚集, 这种现象为聚光运动(accumulation movement).强光条件下, 为避免过强的光对自身造成伤害, 叶绿体沿细胞垂周壁聚集, 使之与光平行分布, 这种现象称为避光运动(avoidance movement) (Kasahara et al., 2004).叶绿体运动受蓝光调节, 蓝光受体向光素PHOT1和PHOT2共同调控叶绿体的聚光反应, 而PHOT2单独调节叶绿体的避光反应(Suetsugu and Wada, 2017).与向光素调节的其它多细胞协调性反应(如向光性、叶片的伸展与定位)不同, 叶绿体运动是细胞自主性反应, 不受基因表达(Kong and Wada, 2016; Wada, 2016)和生长素运输的调节(Christie et al., 2018), 最终由叶绿体肌动蛋白响应入射光的强度与方向而发生快速重组介导(Kadota et al., 2009; Kong and Wada, 2014).叶绿体肌动蛋白特异定位于细胞质膜和叶绿体之间, 在叶绿体前进方向的后面叶绿体肌动蛋白纤维迅速解聚, 前进方向的前面叶绿体肌动蛋白纤维迅速聚集, 这种不对称的叶绿体肌动蛋白纤维分布最终决定了叶绿体的运动方向和速度(Kong and Wada, 2016).CHUP1 (CHLOROPLAST UNUSUAL POSITIONING 1)和 ...
Arabidopsis ROOT PHOTOTROPISM2 is a light-dependent dynamic modulator of phototropin 1 2 2020
... 拟南芥rpt2单突变体在极弱光下(≤0.01 μmol·m-2·s-1)下胚轴向光弯曲反应正常, 随着光照强度的增大弯曲度逐渐降低(Inada et al., 2004), 表明RPT2仅参与调节强蓝光诱导的下胚轴向光弯曲反应.phot1/rpt2双突变体与phot1单突变体表型类似, 在10和100 μmol·m-2·s-1的较强蓝光下可以发生下胚轴向光弯曲反应, 但在0.01-1 μmol·m-2·s-1的弱蓝光下则缺失向光弯曲(Sakai et al., 2000), 暗示RPT2作为PHOT1的下游因子调节强蓝光诱导的下胚轴向光弯曲反应.phot2/rpt2双突变体与phot1/phot2双突变体的表型一致, 缺失强蓝光诱导的下胚轴向光弯曲反应(Inada et al., 2004; Zhao et al., 2018), 且PHOT1和RPT2可以直接相互作用.上述结果进一步证明, RPT2确实参与调节PHOT1介导的下胚轴向光弯曲反应.如果RPT2仅作为PHOT1的下游因子发挥作用, 那么rpt2单突变体应该与phot1单突变体表型一致.表型分析结果显示, rpt2单突变体与phot1/phot2双突变体的表型一致, 即缺失强蓝光诱导的下胚轴向光弯曲反应(Inada et al., 2004), 证明RPT2在PHOT2介导的下胚轴向光弯曲反应中也发挥重要作用.Harada等(2013)认为, RPT2在PHOT1抑制PHOT2途径的过程中发挥作用, 阻碍PHOT1对PHOT2途径的抑制.最近研究表明, RPT2可与PHOT1的LOV1区相互作用并抑制PHOT1的自磷酸化, 以降低PHOT1的光敏感性, 从而行使分子变阻器(molecular rheostat)的功能, 将不同强度蓝光下PHOT1的活性维持在适当的水平.尽管RPT2也可以与PHOT2的LOV1区域相互作用, 但是RPT2不抑制PHOT2的活性(Kimura et al., 2020). ...
... 近年来, 科学家研究发现, 拟南芥NRL家族蛋白在向光素介导的蓝光信号转导通路中起重要作用.拟南芥NRL家族6个分支中的3个分支(II、III和IV) (图1)蛋白目前已经被报道参与向光素调节的反应, 或者与向光素蛋白存在直接相互作用, 暗示这3个分支的其它成员可能参与向光素信号途径的调节.RPT2与NCH1都来自分支II, 既参与生长素依赖的向光性调节, 又参与非生长素依赖的叶绿体聚光运动调节(Suetsugu et al., 2016; Suetsugu and Wada, 2017).目前RPT2和NCH1如何区分向光素介导的2种信号途径并不清楚.考虑到RPT2和NCH1蛋白所在分支与其余2个分支(III和IV)的亲缘关系较远(Christie et al., 2018), 暗示该分支其它成员也可能同时参与生长素依赖和非依赖性的向光素反应.RPT2可与PHOT1的LOV1区相互作用并抑制PHOT1的自磷酸化, 以降低PHOT1的光敏感性.尽管RPT2也可以与PHOT2的LOV1区域相互作用, 但是RPT2不抑制PHOT2的活性(Kimura et al., 2020).可能存在NRL家族其它蛋白与PHOT2相互作用, 调节PHOT2介导的蓝光信号转导.NPH3主要调节生长素依赖的向光性反应以及叶片的伸展与定位(Harada et al., 2013; Zhao et al., 2018), 但NPH3激活后调节生长素不对称分布的机制并不清楚.NPH3的磷酸化状态与其生理活性以及在细胞中的定位密切相关(Haga et al., 2015; Zhao et al., 2018).PHOT1介导的NPH3去磷酸化在弱蓝光诱导的下胚轴向光弯曲反应中发挥重要作用, 强蓝光下NPH3并未发生去磷酸化反应(Tsuchida-Mayama et al., 2008).那么NPH3如何调节强蓝光诱导的下胚轴向光弯曲反应, 是否与其E3泛素化连接酶活性有关? NPY1与PID互作, 通过影响生长素信号调节植物器官的形成以及根的向重力性反应(Suetsugu et al., 2016).生长素不对称分布决定不对称生长, PID- NPY1与PHOT-NPH3作用方式又相似, 那么NPY分支成员是否参与调节向光素介导的植物向性运动以及叶片的伸展与定位? PID和PHOT都属于AGC激酶VIII亚族成员(Ding et al., 2011; Willige et al., 2013), NRL家族蛋白是否可以与AGC激酶VIII亚家族其它成员相互作用, 以调节生长素介导的其它生理反应? 来自分支IV的NRL2也可以与PHOT1发生互作(Sullivan et al., 2009), 暗示该分支的蛋白也可能在向光素介导的蓝光信号途径中发挥重要作用.综上, NRL家族蛋白功能的陆续鉴定, 有望解析向光素PHOT1和PHOT2感受蓝光信号差异, 进而调节不同生理反应的可能机制. ...
Phot1 and phot2 mediate blue light regulation of stomatal opening 3 2001
... 植物向光素PHOT1 (PHOTOTROPIN1)和PHOT2是一类细胞质膜结合的Ser/Thr蛋白激酶, 其C端为激酶结构域, N端含有2个光、氧气和电压敏感的LOV (LIGHT, OXYGEN, OR VOLTAGE SENSING)结构域, 分别命名为LOV1和LOV2 (Christie et al., 2015).蓝光照射后诱导向光素蛋白发生构象改变及自我磷酸化激活(Pfeifer et al., 2010), 活化的向光素蛋白进而调节下胚轴向光弯曲反应(Sakai et al., 2001)、叶绿体运动(Kagawa et al., 2001)、气孔开放(Kinoshita et al., 2001)以及叶片的伸展与定位等(Sakamoto and Briggs, 2002; Inoue et al., 2008b), 从而使植物或叶片处于最有利于吸收光的位置, 优化植物的光合作用(Christie et al., 2015). ...
... 拟南芥气孔由1对保卫细胞构成, 气孔通过调节其开闭程度控制着与环境的气体交换来响应外界的刺激信号(Hetherington and Woodward, 2003; Roelfsema and Hedrich, 2005).红光和蓝光都能诱导气孔快速张开(Shimazaki et al., 2007).红光通过影响叶肉细胞和保卫细胞中叶绿体的光合作用调节气孔开放(Mott et al., 2008; Suetsugu et al., 2014).蓝光作为信号通过激活保卫细胞质膜H+-ATPase诱导气孔开放(Assmann et al., 1985; Kinoshita et al., 2001).蓝光诱导的气孔开放受蓝光受体向光素(PHOT1和PHOT2)调节(Kinoshita et al., 2001).目前人们对向光素介导的气孔开放机制研究较为清楚(Inoue et al., 2008b; Inoue and Kinoshita, 2017).蓝光照射下, 自磷酸化激活的PHOT1直接磷酸化蛋白激酶BLUS1 (LIGHT SIGNALING1), 磷酸化的BLUS1间接将信号传递给I型蛋白磷酸酶(PPI)以及该酶的亚基PRSL1 (Takemiya and Shimazaki, 2016).PPI进一步将信号传递给质膜AHA1型H+-ATPase, 促进H+-ATPase的激活.H+-ATPase的激活需要其C末端倒数第2个苏氨酸的磷酸化以及与14-3-3蛋白的结合(Yamauchi et al., 2016).活化的H+-ATPase通过驱动保卫细胞内H+的跨膜输出诱导保卫细胞质膜的超极化(Shimazaki et al., 2007; Marten et al., 2010).超极化的质膜激活内向K+通道, 促进K+在保卫细胞胞质中积累.为了维持胞质的电荷平衡, Cl-和malate2-随之大量增加, 增加的K+、Cl-和malate2-进一步被转入液泡, 使保卫细胞水势降低, 外界水分大量进入保卫细胞, 促进保卫细胞体积膨胀, 气孔张开(Inoue and Kinoshita, 2017).保卫细胞胞质中Cl-的增加主要是由于向光素抑制了S型阴离子外流蛋白的活性(Sussmilch et al., 2019).向光素对S型阴离子外流蛋白活性的抑制需要2个蛋白激酶CBC1/2 (CONVERGENCE OF BLUE LIGHT (BL) and CO21/2)参与(Hiyama et al., 2017; Sussmilch et al., 2019). ...
... ).蓝光诱导的气孔开放受蓝光受体向光素(PHOT1和PHOT2)调节(Kinoshita et al., 2001).目前人们对向光素介导的气孔开放机制研究较为清楚(Inoue et al., 2008b; Inoue and Kinoshita, 2017).蓝光照射下, 自磷酸化激活的PHOT1直接磷酸化蛋白激酶BLUS1 (LIGHT SIGNALING1), 磷酸化的BLUS1间接将信号传递给I型蛋白磷酸酶(PPI)以及该酶的亚基PRSL1 (Takemiya and Shimazaki, 2016).PPI进一步将信号传递给质膜AHA1型H+-ATPase, 促进H+-ATPase的激活.H+-ATPase的激活需要其C末端倒数第2个苏氨酸的磷酸化以及与14-3-3蛋白的结合(Yamauchi et al., 2016).活化的H+-ATPase通过驱动保卫细胞内H+的跨膜输出诱导保卫细胞质膜的超极化(Shimazaki et al., 2007; Marten et al., 2010).超极化的质膜激活内向K+通道, 促进K+在保卫细胞胞质中积累.为了维持胞质的电荷平衡, Cl-和malate2-随之大量增加, 增加的K+、Cl-和malate2-进一步被转入液泡, 使保卫细胞水势降低, 外界水分大量进入保卫细胞, 促进保卫细胞体积膨胀, 气孔张开(Inoue and Kinoshita, 2017).保卫细胞胞质中Cl-的增加主要是由于向光素抑制了S型阴离子外流蛋白的活性(Sussmilch et al., 2019).向光素对S型阴离子外流蛋白活性的抑制需要2个蛋白激酶CBC1/2 (CONVERGENCE OF BLUE LIGHT (BL) and CO21/2)参与(Hiyama et al., 2017; Sussmilch et al., 2019). ...
Recent advances in understanding the molecular mechanism of chloroplast photorelocation movement 2 2014
... 作为植物进行光合作用的细胞器, 叶绿体响应外界光照刺激而发生的位置变化称为叶绿体光下重定位.弱光照射下, 为最大程度地吸收光能, 叶绿体沿细胞平周壁方向聚集, 这种现象为聚光运动(accumulation movement).强光条件下, 为避免过强的光对自身造成伤害, 叶绿体沿细胞垂周壁聚集, 使之与光平行分布, 这种现象称为避光运动(avoidance movement) (Kasahara et al., 2004).叶绿体运动受蓝光调节, 蓝光受体向光素PHOT1和PHOT2共同调控叶绿体的聚光反应, 而PHOT2单独调节叶绿体的避光反应(Suetsugu and Wada, 2017).与向光素调节的其它多细胞协调性反应(如向光性、叶片的伸展与定位)不同, 叶绿体运动是细胞自主性反应, 不受基因表达(Kong and Wada, 2016; Wada, 2016)和生长素运输的调节(Christie et al., 2018), 最终由叶绿体肌动蛋白响应入射光的强度与方向而发生快速重组介导(Kadota et al., 2009; Kong and Wada, 2014).叶绿体肌动蛋白特异定位于细胞质膜和叶绿体之间, 在叶绿体前进方向的后面叶绿体肌动蛋白纤维迅速解聚, 前进方向的前面叶绿体肌动蛋白纤维迅速聚集, 这种不对称的叶绿体肌动蛋白纤维分布最终决定了叶绿体的运动方向和速度(Kong and Wada, 2016).CHUP1 (CHLOROPLAST UNUSUAL POSITIONING 1)和 ...
... KAC (KINESIN-LIKE PROTEIN FOR ACTIN-BASED CHLOROPLAST MOVEMENT)参与调节叶绿体肌动蛋白纤维的聚集(Wada, 2013; Kong and Wada, 2014), WEB1 (WEAK CHLOROPLAST MOVEMENT UNDER BLUE LIGHT 1)和PMI1 (PLASTIDMOVEMENT IMPAIRED1)对叶绿体肌动蛋白纤维的稳定性至关重要(Suetsugu et al., 2015). ...
Molecular basis of chloroplast photorelocation movement 2 2016
... 作为植物进行光合作用的细胞器, 叶绿体响应外界光照刺激而发生的位置变化称为叶绿体光下重定位.弱光照射下, 为最大程度地吸收光能, 叶绿体沿细胞平周壁方向聚集, 这种现象为聚光运动(accumulation movement).强光条件下, 为避免过强的光对自身造成伤害, 叶绿体沿细胞垂周壁聚集, 使之与光平行分布, 这种现象称为避光运动(avoidance movement) (Kasahara et al., 2004).叶绿体运动受蓝光调节, 蓝光受体向光素PHOT1和PHOT2共同调控叶绿体的聚光反应, 而PHOT2单独调节叶绿体的避光反应(Suetsugu and Wada, 2017).与向光素调节的其它多细胞协调性反应(如向光性、叶片的伸展与定位)不同, 叶绿体运动是细胞自主性反应, 不受基因表达(Kong and Wada, 2016; Wada, 2016)和生长素运输的调节(Christie et al., 2018), 最终由叶绿体肌动蛋白响应入射光的强度与方向而发生快速重组介导(Kadota et al., 2009; Kong and Wada, 2014).叶绿体肌动蛋白特异定位于细胞质膜和叶绿体之间, 在叶绿体前进方向的后面叶绿体肌动蛋白纤维迅速解聚, 前进方向的前面叶绿体肌动蛋白纤维迅速聚集, 这种不对称的叶绿体肌动蛋白纤维分布最终决定了叶绿体的运动方向和速度(Kong and Wada, 2016).CHUP1 (CHLOROPLAST UNUSUAL POSITIONING 1)和 ...
SETH1 and SETH2, two components of the glycosylphosphatidylinositol anchor biosynthetic pathway, are required for pollen germination and tube growth in Arabidopsis 1 2004
... NPH3和RPT2是来自2个不同分支的NRL家族成员, 通过与向光素相互作用调节蓝光诱导下胚轴向光性、叶绿体聚光运动、气孔开放以及叶片伸展与定位(Christie et al., 2015, 2018).NRL8是AGC激酶(cAMP依赖的蛋白激酶A, cGMP依赖的蛋白激酶G和磷脂依赖的蛋白激酶C)活性依赖的花粉管生长调节因子(Lalanne et al., 2004; Zhang et al., 2008), 该蛋白与RPT2来自分支II.与NPH3类似, SR1IP1 (AtSR1 INTERACTING PROTEIN 1)也称NRL31, 可以与CRL3 ( CUL3 RING E3 UBIQUITIN LIGASE)形成复合体, 调节底物的泛素化(Zhang et al., 2014).SR1IP1通过调节拟南芥中Ca2+/钙调素结合的转录因子AtSR1 (SIGNAL RESPONSIVE 1)的泛素化降解, 增强植物对细菌性病原体的抗性(Zhang et al., 2014).但系统进化研究发现, SR1IP1也来自分支II, 且比NRL8与RPT2亲缘关系更近, 与NPH3的进化明显不同.Suetsugu等(2016)在筛选参与调节叶绿体运动的NRL家族蛋白时, 发现NRL31基因突变抑制了光诱导的叶绿体运动, 且蛋白NRL31可以与PHOT1发生体内互作, 因此将其重命名为NCH1 (NRL PROTEIN FORCHLOROPLASTMOVEMENT 1).DEFECTIVELY ORGANIZED TRIBUTARIES 3/NRL23是与NPH3来自同一个分支的NRL家族蛋白, 主要参与维管组织发育以及叶脉结构调节(Petricka et al., 2008).虽然NRL23与NPH3的亲缘关系较近, 但是该蛋白是否在向光素介导的信号途径中发挥作用尚未见报道.NRL5、NRL6、NRL7、NRL15、NRL20、NRL21、NRL26和NRL30是分支III中的NPY (NAKED PINS IN YUCCA)家族成员, 其中NRL6/NPY2、NRL7/NPY4、NRL20/NPY1、NRL21/NPY5和NRL30/NPY3以功能冗余的方式与AGCVIII激酶共同调节器官发育以及向重力性反应中生长素的运输(Cheng et al., 2008; Li et al., 2011).通过分析NPYs基因对应的双突变体、三突变体、四突变体以及五突变体根的向重力性表型, 发现双突变体和三突变体向重力性表型缺失不明显; 四突变体表现不同程度的向重力性缺失表型, 其中npy1/npy2/npy4/npy5和npy2/npy3/npy4/npy5严重缺失向重力性; 五突变体的根几乎完全缺失向重力性, 在21°到150°之间随机生长(Li et al., 2011). ...
PHYTOCHROME KINASE SUBSTRATE 1 is a phototropin 1 binding protein required for phototropism 1 2006
... 拟南芥中ABCB19 (ATP-BINDING CAS-SETTE B 19)和光敏色素激酶底物(PHYTOCHROME KINASE SUBSTRATE 4, PKS4)参与调节生长素的转运, 是下胚轴向光弯曲的负调节因子(Christie et al., 2011; Demarsy et al., 2012).自磷酸化激活的PHOT1通过直接磷酸化ABCB19和PKS4, 抑制ABCB19和PKS4的生长素转运活性, 促进下胚轴向光弯曲反应(Christie et al., 2011; Demarsy et al., 2012).PHOT1通过磷酸化修饰调节ABCB19的活性, 但是PHOT2并不影响ABCB19的活性(Christie et al., 2011).蛋白磷酸酶2A (PP2A)的A1亚基对应的蛋白RCN1 (Root Curling in N-naphthylphthalamic acid 1)与PHOT2蛋白发生体外互作, 负调节PHOT2介导的向光弯曲, 但RCN1并不与PHOT1直接相互作用(Tseng and Briggs, 2010).NPH3 (NONPHOTOTROPIC HYPOCOTYL 3)和RPT2 (ROOT PHOTOTROPISM 2)是通过筛选拟南芥下胚轴以及根的向光性缺失突变体获得(Motchoulski and Liscum, 1999; Sakai et al., 2000).随后的研究发现, NPH3和RPT2是向光素途径的重要调控因子, 均可与PHOT1蛋白直接相互作用(Blakeslee et al., 2004; Stone et al., 2008).虽然NPH3和RPT2也参与PHOT2信号转导途径, 但目前并无RPT2与PHOT2蛋白之间直接相互作用的报道(Inada et al., 2004; Lariguet et al., 2006).上述结果暗示PHOT1和PHOT2可能通过与不同的下游互作因子差异调节蓝光反应. ...
NPY genes play an essential role in root gravitropic responses in Arabidopsis 2 2011
... NPH3和RPT2是来自2个不同分支的NRL家族成员, 通过与向光素相互作用调节蓝光诱导下胚轴向光性、叶绿体聚光运动、气孔开放以及叶片伸展与定位(Christie et al., 2015, 2018).NRL8是AGC激酶(cAMP依赖的蛋白激酶A, cGMP依赖的蛋白激酶G和磷脂依赖的蛋白激酶C)活性依赖的花粉管生长调节因子(Lalanne et al., 2004; Zhang et al., 2008), 该蛋白与RPT2来自分支II.与NPH3类似, SR1IP1 (AtSR1 INTERACTING PROTEIN 1)也称NRL31, 可以与CRL3 ( CUL3 RING E3 UBIQUITIN LIGASE)形成复合体, 调节底物的泛素化(Zhang et al., 2014).SR1IP1通过调节拟南芥中Ca2+/钙调素结合的转录因子AtSR1 (SIGNAL RESPONSIVE 1)的泛素化降解, 增强植物对细菌性病原体的抗性(Zhang et al., 2014).但系统进化研究发现, SR1IP1也来自分支II, 且比NRL8与RPT2亲缘关系更近, 与NPH3的进化明显不同.Suetsugu等(2016)在筛选参与调节叶绿体运动的NRL家族蛋白时, 发现NRL31基因突变抑制了光诱导的叶绿体运动, 且蛋白NRL31可以与PHOT1发生体内互作, 因此将其重命名为NCH1 (NRL PROTEIN FORCHLOROPLASTMOVEMENT 1).DEFECTIVELY ORGANIZED TRIBUTARIES 3/NRL23是与NPH3来自同一个分支的NRL家族蛋白, 主要参与维管组织发育以及叶脉结构调节(Petricka et al., 2008).虽然NRL23与NPH3的亲缘关系较近, 但是该蛋白是否在向光素介导的信号途径中发挥作用尚未见报道.NRL5、NRL6、NRL7、NRL15、NRL20、NRL21、NRL26和NRL30是分支III中的NPY (NAKED PINS IN YUCCA)家族成员, 其中NRL6/NPY2、NRL7/NPY4、NRL20/NPY1、NRL21/NPY5和NRL30/NPY3以功能冗余的方式与AGCVIII激酶共同调节器官发育以及向重力性反应中生长素的运输(Cheng et al., 2008; Li et al., 2011).通过分析NPYs基因对应的双突变体、三突变体、四突变体以及五突变体根的向重力性表型, 发现双突变体和三突变体向重力性表型缺失不明显; 四突变体表现不同程度的向重力性缺失表型, 其中npy1/npy2/npy4/npy5和npy2/npy3/npy4/npy5严重缺失向重力性; 五突变体的根几乎完全缺失向重力性, 在21°到150°之间随机生长(Li et al., 2011). ...
... 严重缺失向重力性; 五突变体的根几乎完全缺失向重力性, 在21°到150°之间随机生长(Li et al., 2011). ...
Phototropism: growing towards an understanding of plant movement 2 2014
... NRL (NPH3/RPT2-Like)蛋白家族是植物特有的, 其成员含有3个保守结构域: N端的BTB (Bric àbrac, Tramtrack, and Broad complex)结构域、中间的NPH3结构域以及C端的螺旋-螺旋结构域(Liscum et al., 2014).其中, BTB结构域主要起E3连接酶的作用, 与CUL3发生相互作用(Gingerich et al., 2005; Figueroa et al., 2005), C端的螺旋-螺旋结构域主要介导蛋白之间的相互作用(Inada et al., 2004).拟南芥中33个NRL家族成员都含有NPH3结构域, 10个缺少C端的螺旋-螺旋结构域, 2个缺少N端的BTB结构域(Pedmale et al., 2010; Christie et al., 2018).根据NRL家族成员间的进化关系, 拟南芥中的33个NRL家族成员可分为6个分支(图1) (Suetsugu et al., 2016; Christie et al., 2018).目前, 来自2个分支的10个成员的功能已得到鉴定(de Carbonnel et al., 2010; Christie et al., 2018) (图1). ...
... 拟南芥nph3突变体在任何强度的单侧蓝光下均表现缺失下胚轴向光性(Liscum and Briggs, 1996; Liscum et al., 2014; Fankhauser and Christie, 2015), 表明NPH3调节PHOT1和PHOT2介导蓝光诱导的下胚轴弯曲反应.黑暗条件下, 磷酸化的NPH3蛋白与PHOT1结合, 定位于细胞膜, 单侧弱蓝光照射下, NPH3迅速发生PHOT1依赖的去磷酸化反应, 从细胞膜上脱离(Haga et al., 2015; Zhao et al., 2018).目前磷酸化NPH3的激酶还不清楚, 但NPH3去磷酸化主要由I型蛋白磷酸酶介导(Pedmale and Liscum, 2007).去磷酸化后NPH3从细胞膜向胞质的转移降低了其介导向光弯曲反应的能力(Haga et al., 2015; Sullivan et al., 2019).随着光照强度的增加, 下胚轴的弯曲度呈强度依赖的钟形曲线(Christie and Murphy, 2013).随着光照时间的增加, RPT2蛋白含量增加, 与去磷酸化的NPH3结合形成RPT2-NPH3复合物并与PHOT1蛋白结合, 促进NPH3的磷酸化以及从胞质到细胞膜的重定位, 开启第二次向光弯曲(Haga et al., 2015; Zhao et al., 2018).虽然PHOT1- NPH3复合体在细胞膜的重定位对持续光照介导的二次向光弯曲至关重要(Haga et al., 2015; Christie et al., 2018; Zhao et al., 2018), 但其生物学意义还不清楚, 人们认为这种变化有可能对调节生长素的不对称分布有重要影响(Pedmale and Liscum, 2007; Haga et al., 2015). ...
Mutations in the NPH1 locus of Arabidopsis disrupt the perception of phototropic stimuli 1 1995
... 早在达尔文(1880年)时期, 人们就已发现植物向光性, 但是直到向光素PHOT1和PHOT2先后被鉴定(Liscum and Briggs, 1995; Christie et al., 1998), 植物的向光性机理研究才取得突飞猛进的发展.研究发现, PHOT1调节弱蓝光(0.01-1 μmol·m-2·s-1)和强蓝光(>1 μmol·m-2·s-1)诱导的下胚轴的向光性; PHOT2仅在强蓝光下起作用, 且与PHOT1功能冗余(Sakai et al., 2001; Briggs and Christie, 2002).由于强蓝光下PHOT1和PHOT2的功能冗余, 目前向光性机制研究主要集中在PHOT1介导的弱蓝光反应(Motchoulski and Liscum, 1999; Haga et al., 2015), 仅有少量研究通过突变体背景下的抑制子筛选寻找参与强蓝光信号途径的作用因子(赵翔等, 2014).单侧蓝光照射下, 自磷酸化激活的向光素蛋白将信号传递给NPH3、RPT2以及PKS等下游因子(Motchoulski and Liscum, 1999; Inada et al., 2004), 调节生长素运输载体的活性与定位(Ding et al., 2011).例如, 引起内皮层细胞背光侧PIN3 (PIN-FORMED3)积累(刘广超和丁兆军, 2018), 促进背光侧生长素含量增加、细胞伸长速率加快, 最终引起植物弯向光生长(Christie and Murphy, 2013).下胚轴弯曲过程中生长素的不对称分布受各类生长素运输载体的共同调控, 如PINs (PIN3、PIN4和PIN7)和ABCB19 (Christie et al., 2011).蓝光下, PIN3的磷酸化受抑制, 促进PIN3的背光侧定位, 诱导背光侧生长素积累以及下胚轴的向光性生长(Ding et al., 2011).PIN3的磷酸化以及极性定位受AGC激酶VIII亚族成员PID (PINOID)以及D6PK (D6 PROTEIN KINASE)的调节(Ding et al., 2011; Willige et al., 2013).ABCB19是生长素外流载体, 被蓝光激活的PHOT1磷酸化后, ABCB19的生长素转运活性受抑制, 增加拟南芥下胚轴顶端生长素含量并抑制其垂直生长, 同时通过PIN3蛋白促进生长素在下胚轴中侧向移动(Christie et al., 2011).背光侧生长素的大量积累促进了背光侧IAA19的降解, 释放出有活性的ARF7 (Sun et al., 2013), 不对称激活与生长相关基因的表达促进了下胚轴的向光性反应(刘广超和丁兆军, 2018). ...
Mutations of Arabidopsis in potential transduction and response components of the phototropic signaling pathway 1 1996
... 拟南芥nph3突变体在任何强度的单侧蓝光下均表现缺失下胚轴向光性(Liscum and Briggs, 1996; Liscum et al., 2014; Fankhauser and Christie, 2015), 表明NPH3调节PHOT1和PHOT2介导蓝光诱导的下胚轴弯曲反应.黑暗条件下, 磷酸化的NPH3蛋白与PHOT1结合, 定位于细胞膜, 单侧弱蓝光照射下, NPH3迅速发生PHOT1依赖的去磷酸化反应, 从细胞膜上脱离(Haga et al., 2015; Zhao et al., 2018).目前磷酸化NPH3的激酶还不清楚, 但NPH3去磷酸化主要由I型蛋白磷酸酶介导(Pedmale and Liscum, 2007).去磷酸化后NPH3从细胞膜向胞质的转移降低了其介导向光弯曲反应的能力(Haga et al., 2015; Sullivan et al., 2019).随着光照强度的增加, 下胚轴的弯曲度呈强度依赖的钟形曲线(Christie and Murphy, 2013).随着光照时间的增加, RPT2蛋白含量增加, 与去磷酸化的NPH3结合形成RPT2-NPH3复合物并与PHOT1蛋白结合, 促进NPH3的磷酸化以及从胞质到细胞膜的重定位, 开启第二次向光弯曲(Haga et al., 2015; Zhao et al., 2018).虽然PHOT1- NPH3复合体在细胞膜的重定位对持续光照介导的二次向光弯曲至关重要(Haga et al., 2015; Christie et al., 2018; Zhao et al., 2018), 但其生物学意义还不清楚, 人们认为这种变化有可能对调节生长素的不对称分布有重要影响(Pedmale and Liscum, 2007; Haga et al., 2015). ...
The continuing arc toward phototropic enlightenment 1 2020
... 拟南芥(Arabidopsis thaliana) PHOT1和PHOT2以功能冗余的方式调节下胚轴向光性和根负向光性、叶绿体聚光运动、气孔开放以及叶片伸展与定位等(Christie et al., 2015).PHOT1特异性抑制光下黄化幼苗下胚轴的快速伸长(Folta et al., 2003)、子叶变绿过程中叶绿素结合蛋白mRNA转录的稳定性(Folta and Kaufman, 2003)以及侧根的快速生长(Moni et al., 2015).PHOT2不仅介导强蓝光诱导的下胚轴向光弯曲(Sakai et al., 2001), 而且也特异调节叶绿体和细胞核的避光运动(Higa et al., 2014)以及抗性蛋白介导的病毒防御反应(Jeong et al., 2010).PHOT1和PHOT2的功能差异可能与磷酸化修饰位点、磷酸化程度、亚细胞定位变化以及下游相互作用蛋白种类不同有关(Zhao et al., 2013; Morrow et al., 2018; Liscum et al., 2020). ...
Light-induced modification of plant plasma membrane ion transport 1 2010
... 拟南芥气孔由1对保卫细胞构成, 气孔通过调节其开闭程度控制着与环境的气体交换来响应外界的刺激信号(Hetherington and Woodward, 2003; Roelfsema and Hedrich, 2005).红光和蓝光都能诱导气孔快速张开(Shimazaki et al., 2007).红光通过影响叶肉细胞和保卫细胞中叶绿体的光合作用调节气孔开放(Mott et al., 2008; Suetsugu et al., 2014).蓝光作为信号通过激活保卫细胞质膜H+-ATPase诱导气孔开放(Assmann et al., 1985; Kinoshita et al., 2001).蓝光诱导的气孔开放受蓝光受体向光素(PHOT1和PHOT2)调节(Kinoshita et al., 2001).目前人们对向光素介导的气孔开放机制研究较为清楚(Inoue et al., 2008b; Inoue and Kinoshita, 2017).蓝光照射下, 自磷酸化激活的PHOT1直接磷酸化蛋白激酶BLUS1 (LIGHT SIGNALING1), 磷酸化的BLUS1间接将信号传递给I型蛋白磷酸酶(PPI)以及该酶的亚基PRSL1 (Takemiya and Shimazaki, 2016).PPI进一步将信号传递给质膜AHA1型H+-ATPase, 促进H+-ATPase的激活.H+-ATPase的激活需要其C末端倒数第2个苏氨酸的磷酸化以及与14-3-3蛋白的结合(Yamauchi et al., 2016).活化的H+-ATPase通过驱动保卫细胞内H+的跨膜输出诱导保卫细胞质膜的超极化(Shimazaki et al., 2007; Marten et al., 2010).超极化的质膜激活内向K+通道, 促进K+在保卫细胞胞质中积累.为了维持胞质的电荷平衡, Cl-和malate2-随之大量增加, 增加的K+、Cl-和malate2-进一步被转入液泡, 使保卫细胞水势降低, 外界水分大量进入保卫细胞, 促进保卫细胞体积膨胀, 气孔张开(Inoue and Kinoshita, 2017).保卫细胞胞质中Cl-的增加主要是由于向光素抑制了S型阴离子外流蛋白的活性(Sussmilch et al., 2019).向光素对S型阴离子外流蛋白活性的抑制需要2个蛋白激酶CBC1/2 (CONVERGENCE OF BLUE LIGHT (BL) and CO21/2)参与(Hiyama et al., 2017; Sussmilch et al., 2019). ...
The blue light receptor Phototropin 1 suppresses lateral root growth by controlling cell elongation 1 2015
... 拟南芥(Arabidopsis thaliana) PHOT1和PHOT2以功能冗余的方式调节下胚轴向光性和根负向光性、叶绿体聚光运动、气孔开放以及叶片伸展与定位等(Christie et al., 2015).PHOT1特异性抑制光下黄化幼苗下胚轴的快速伸长(Folta et al., 2003)、子叶变绿过程中叶绿素结合蛋白mRNA转录的稳定性(Folta and Kaufman, 2003)以及侧根的快速生长(Moni et al., 2015).PHOT2不仅介导强蓝光诱导的下胚轴向光弯曲(Sakai et al., 2001), 而且也特异调节叶绿体和细胞核的避光运动(Higa et al., 2014)以及抗性蛋白介导的病毒防御反应(Jeong et al., 2010).PHOT1和PHOT2的功能差异可能与磷酸化修饰位点、磷酸化程度、亚细胞定位变化以及下游相互作用蛋白种类不同有关(Zhao et al., 2013; Morrow et al., 2018; Liscum et al., 2020). ...
Phototropism in land plants: molecules and mechanism from light perception to response 1 2018
... 拟南芥(Arabidopsis thaliana) PHOT1和PHOT2以功能冗余的方式调节下胚轴向光性和根负向光性、叶绿体聚光运动、气孔开放以及叶片伸展与定位等(Christie et al., 2015).PHOT1特异性抑制光下黄化幼苗下胚轴的快速伸长(Folta et al., 2003)、子叶变绿过程中叶绿素结合蛋白mRNA转录的稳定性(Folta and Kaufman, 2003)以及侧根的快速生长(Moni et al., 2015).PHOT2不仅介导强蓝光诱导的下胚轴向光弯曲(Sakai et al., 2001), 而且也特异调节叶绿体和细胞核的避光运动(Higa et al., 2014)以及抗性蛋白介导的病毒防御反应(Jeong et al., 2010).PHOT1和PHOT2的功能差异可能与磷酸化修饰位点、磷酸化程度、亚细胞定位变化以及下游相互作用蛋白种类不同有关(Zhao et al., 2013; Morrow et al., 2018; Liscum et al., 2020). ...
Arabidopsis NPH3: a NPH1 photoreceptor-linteracting protein essential for phototropism 4 1999
... 拟南芥中ABCB19 (ATP-BINDING CAS-SETTE B 19)和光敏色素激酶底物(PHYTOCHROME KINASE SUBSTRATE 4, PKS4)参与调节生长素的转运, 是下胚轴向光弯曲的负调节因子(Christie et al., 2011; Demarsy et al., 2012).自磷酸化激活的PHOT1通过直接磷酸化ABCB19和PKS4, 抑制ABCB19和PKS4的生长素转运活性, 促进下胚轴向光弯曲反应(Christie et al., 2011; Demarsy et al., 2012).PHOT1通过磷酸化修饰调节ABCB19的活性, 但是PHOT2并不影响ABCB19的活性(Christie et al., 2011).蛋白磷酸酶2A (PP2A)的A1亚基对应的蛋白RCN1 (Root Curling in N-naphthylphthalamic acid 1)与PHOT2蛋白发生体外互作, 负调节PHOT2介导的向光弯曲, 但RCN1并不与PHOT1直接相互作用(Tseng and Briggs, 2010).NPH3 (NONPHOTOTROPIC HYPOCOTYL 3)和RPT2 (ROOT PHOTOTROPISM 2)是通过筛选拟南芥下胚轴以及根的向光性缺失突变体获得(Motchoulski and Liscum, 1999; Sakai et al., 2000).随后的研究发现, NPH3和RPT2是向光素途径的重要调控因子, 均可与PHOT1蛋白直接相互作用(Blakeslee et al., 2004; Stone et al., 2008).虽然NPH3和RPT2也参与PHOT2信号转导途径, 但目前并无RPT2与PHOT2蛋白之间直接相互作用的报道(Inada et al., 2004; Lariguet et al., 2006).上述结果暗示PHOT1和PHOT2可能通过与不同的下游互作因子差异调节蓝光反应. ...
... 目前已经鉴定的10个有功能的NRL家族蛋白中有3个(NPH3、RPT2和NCH1)可与向光素发生直接相互作用(Motchoulski and Liscum, 1999; Inada et al., 2004; Suetsugu et al., 2016).此外, 酵母双杂交筛选发现NRL2可以与PHOT1发生相互作用(Sullivan et al., 2009), 但是NRL2的功能及其在向光素介导的信号途径中的作用目前还不清楚.虽然大部分NRL家族成员的功能未知, 但已有的实验结果表明, NRL家族成员在向光素信号途径中起重要作用, 因此我们总结NRL家族蛋白在向光素介导的信号途径中的作用, 旨在为全面揭示NPH3/RPT2-Like家族成员的功能提供线索. ...
... 早在达尔文(1880年)时期, 人们就已发现植物向光性, 但是直到向光素PHOT1和PHOT2先后被鉴定(Liscum and Briggs, 1995; Christie et al., 1998), 植物的向光性机理研究才取得突飞猛进的发展.研究发现, PHOT1调节弱蓝光(0.01-1 μmol·m-2·s-1)和强蓝光(>1 μmol·m-2·s-1)诱导的下胚轴的向光性; PHOT2仅在强蓝光下起作用, 且与PHOT1功能冗余(Sakai et al., 2001; Briggs and Christie, 2002).由于强蓝光下PHOT1和PHOT2的功能冗余, 目前向光性机制研究主要集中在PHOT1介导的弱蓝光反应(Motchoulski and Liscum, 1999; Haga et al., 2015), 仅有少量研究通过突变体背景下的抑制子筛选寻找参与强蓝光信号途径的作用因子(赵翔等, 2014).单侧蓝光照射下, 自磷酸化激活的向光素蛋白将信号传递给NPH3、RPT2以及PKS等下游因子(Motchoulski and Liscum, 1999; Inada et al., 2004), 调节生长素运输载体的活性与定位(Ding et al., 2011).例如, 引起内皮层细胞背光侧PIN3 (PIN-FORMED3)积累(刘广超和丁兆军, 2018), 促进背光侧生长素含量增加、细胞伸长速率加快, 最终引起植物弯向光生长(Christie and Murphy, 2013).下胚轴弯曲过程中生长素的不对称分布受各类生长素运输载体的共同调控, 如PINs (PIN3、PIN4和PIN7)和ABCB19 (Christie et al., 2011).蓝光下, PIN3的磷酸化受抑制, 促进PIN3的背光侧定位, 诱导背光侧生长素积累以及下胚轴的向光性生长(Ding et al., 2011).PIN3的磷酸化以及极性定位受AGC激酶VIII亚族成员PID (PINOID)以及D6PK (D6 PROTEIN KINASE)的调节(Ding et al., 2011; Willige et al., 2013).ABCB19是生长素外流载体, 被蓝光激活的PHOT1磷酸化后, ABCB19的生长素转运活性受抑制, 增加拟南芥下胚轴顶端生长素含量并抑制其垂直生长, 同时通过PIN3蛋白促进生长素在下胚轴中侧向移动(Christie et al., 2011).背光侧生长素的大量积累促进了背光侧IAA19的降解, 释放出有活性的ARF7 (Sun et al., 2013), 不对称激活与生长相关基因的表达促进了下胚轴的向光性反应(刘广超和丁兆军, 2018). ...
... ).单侧蓝光照射下, 自磷酸化激活的向光素蛋白将信号传递给NPH3、RPT2以及PKS等下游因子(Motchoulski and Liscum, 1999; Inada et al., 2004), 调节生长素运输载体的活性与定位(Ding et al., 2011).例如, 引起内皮层细胞背光侧PIN3 (PIN-FORMED3)积累(刘广超和丁兆军, 2018), 促进背光侧生长素含量增加、细胞伸长速率加快, 最终引起植物弯向光生长(Christie and Murphy, 2013).下胚轴弯曲过程中生长素的不对称分布受各类生长素运输载体的共同调控, 如PINs (PIN3、PIN4和PIN7)和ABCB19 (Christie et al., 2011).蓝光下, PIN3的磷酸化受抑制, 促进PIN3的背光侧定位, 诱导背光侧生长素积累以及下胚轴的向光性生长(Ding et al., 2011).PIN3的磷酸化以及极性定位受AGC激酶VIII亚族成员PID (PINOID)以及D6PK (D6 PROTEIN KINASE)的调节(Ding et al., 2011; Willige et al., 2013).ABCB19是生长素外流载体, 被蓝光激活的PHOT1磷酸化后, ABCB19的生长素转运活性受抑制, 增加拟南芥下胚轴顶端生长素含量并抑制其垂直生长, 同时通过PIN3蛋白促进生长素在下胚轴中侧向移动(Christie et al., 2011).背光侧生长素的大量积累促进了背光侧IAA19的降解, 释放出有活性的ARF7 (Sun et al., 2013), 不对称激活与生长相关基因的表达促进了下胚轴的向光性反应(刘广超和丁兆军, 2018). ...
The role of the mesophyll in stomatal responses to light and CO2 1 2008
... 拟南芥气孔由1对保卫细胞构成, 气孔通过调节其开闭程度控制着与环境的气体交换来响应外界的刺激信号(Hetherington and Woodward, 2003; Roelfsema and Hedrich, 2005).红光和蓝光都能诱导气孔快速张开(Shimazaki et al., 2007).红光通过影响叶肉细胞和保卫细胞中叶绿体的光合作用调节气孔开放(Mott et al., 2008; Suetsugu et al., 2014).蓝光作为信号通过激活保卫细胞质膜H+-ATPase诱导气孔开放(Assmann et al., 1985; Kinoshita et al., 2001).蓝光诱导的气孔开放受蓝光受体向光素(PHOT1和PHOT2)调节(Kinoshita et al., 2001).目前人们对向光素介导的气孔开放机制研究较为清楚(Inoue et al., 2008b; Inoue and Kinoshita, 2017).蓝光照射下, 自磷酸化激活的PHOT1直接磷酸化蛋白激酶BLUS1 (LIGHT SIGNALING1), 磷酸化的BLUS1间接将信号传递给I型蛋白磷酸酶(PPI)以及该酶的亚基PRSL1 (Takemiya and Shimazaki, 2016).PPI进一步将信号传递给质膜AHA1型H+-ATPase, 促进H+-ATPase的激活.H+-ATPase的激活需要其C末端倒数第2个苏氨酸的磷酸化以及与14-3-3蛋白的结合(Yamauchi et al., 2016).活化的H+-ATPase通过驱动保卫细胞内H+的跨膜输出诱导保卫细胞质膜的超极化(Shimazaki et al., 2007; Marten et al., 2010).超极化的质膜激活内向K+通道, 促进K+在保卫细胞胞质中积累.为了维持胞质的电荷平衡, Cl-和malate2-随之大量增加, 增加的K+、Cl-和malate2-进一步被转入液泡, 使保卫细胞水势降低, 外界水分大量进入保卫细胞, 促进保卫细胞体积膨胀, 气孔张开(Inoue and Kinoshita, 2017).保卫细胞胞质中Cl-的增加主要是由于向光素抑制了S型阴离子外流蛋白的活性(Sussmilch et al., 2019).向光素对S型阴离子外流蛋白活性的抑制需要2个蛋白激酶CBC1/2 (CONVERGENCE OF BLUE LIGHT (BL) and CO21/2)参与(Hiyama et al., 2017; Sussmilch et al., 2019). ...
Phototropism: mechanism and outcomes 1 2010
... NRL (NPH3/RPT2-Like)蛋白家族是植物特有的, 其成员含有3个保守结构域: N端的BTB (Bric àbrac, Tramtrack, and Broad complex)结构域、中间的NPH3结构域以及C端的螺旋-螺旋结构域(Liscum et al., 2014).其中, BTB结构域主要起E3连接酶的作用, 与CUL3发生相互作用(Gingerich et al., 2005; Figueroa et al., 2005), C端的螺旋-螺旋结构域主要介导蛋白之间的相互作用(Inada et al., 2004).拟南芥中33个NRL家族成员都含有NPH3结构域, 10个缺少C端的螺旋-螺旋结构域, 2个缺少N端的BTB结构域(Pedmale et al., 2010; Christie et al., 2018).根据NRL家族成员间的进化关系, 拟南芥中的33个NRL家族成员可分为6个分支(图1) (Suetsugu et al., 2016; Christie et al., 2018).目前, 来自2个分支的10个成员的功能已得到鉴定(de Carbonnel et al., 2010; Christie et al., 2018) (图1). ...
Regulation of phototropic signaling in Arabidopsis via phosphorylation state changes in the phototropin 1-interacting protein NPH3 2 2007
... 拟南芥nph3突变体在任何强度的单侧蓝光下均表现缺失下胚轴向光性(Liscum and Briggs, 1996; Liscum et al., 2014; Fankhauser and Christie, 2015), 表明NPH3调节PHOT1和PHOT2介导蓝光诱导的下胚轴弯曲反应.黑暗条件下, 磷酸化的NPH3蛋白与PHOT1结合, 定位于细胞膜, 单侧弱蓝光照射下, NPH3迅速发生PHOT1依赖的去磷酸化反应, 从细胞膜上脱离(Haga et al., 2015; Zhao et al., 2018).目前磷酸化NPH3的激酶还不清楚, 但NPH3去磷酸化主要由I型蛋白磷酸酶介导(Pedmale and Liscum, 2007).去磷酸化后NPH3从细胞膜向胞质的转移降低了其介导向光弯曲反应的能力(Haga et al., 2015; Sullivan et al., 2019).随着光照强度的增加, 下胚轴的弯曲度呈强度依赖的钟形曲线(Christie and Murphy, 2013).随着光照时间的增加, RPT2蛋白含量增加, 与去磷酸化的NPH3结合形成RPT2-NPH3复合物并与PHOT1蛋白结合, 促进NPH3的磷酸化以及从胞质到细胞膜的重定位, 开启第二次向光弯曲(Haga et al., 2015; Zhao et al., 2018).虽然PHOT1- NPH3复合体在细胞膜的重定位对持续光照介导的二次向光弯曲至关重要(Haga et al., 2015; Christie et al., 2018; Zhao et al., 2018), 但其生物学意义还不清楚, 人们认为这种变化有可能对调节生长素的不对称分布有重要影响(Pedmale and Liscum, 2007; Haga et al., 2015). ...
... ), 但其生物学意义还不清楚, 人们认为这种变化有可能对调节生长素的不对称分布有重要影响(Pedmale and Liscum, 2007; Haga et al., 2015). ...
Vein patterning screens and the defectively organized tributaries mutants in Arabidopsis thaliana 1 2008
... NPH3和RPT2是来自2个不同分支的NRL家族成员, 通过与向光素相互作用调节蓝光诱导下胚轴向光性、叶绿体聚光运动、气孔开放以及叶片伸展与定位(Christie et al., 2015, 2018).NRL8是AGC激酶(cAMP依赖的蛋白激酶A, cGMP依赖的蛋白激酶G和磷脂依赖的蛋白激酶C)活性依赖的花粉管生长调节因子(Lalanne et al., 2004; Zhang et al., 2008), 该蛋白与RPT2来自分支II.与NPH3类似, SR1IP1 (AtSR1 INTERACTING PROTEIN 1)也称NRL31, 可以与CRL3 ( CUL3 RING E3 UBIQUITIN LIGASE)形成复合体, 调节底物的泛素化(Zhang et al., 2014).SR1IP1通过调节拟南芥中Ca2+/钙调素结合的转录因子AtSR1 (SIGNAL RESPONSIVE 1)的泛素化降解, 增强植物对细菌性病原体的抗性(Zhang et al., 2014).但系统进化研究发现, SR1IP1也来自分支II, 且比NRL8与RPT2亲缘关系更近, 与NPH3的进化明显不同.Suetsugu等(2016)在筛选参与调节叶绿体运动的NRL家族蛋白时, 发现NRL31基因突变抑制了光诱导的叶绿体运动, 且蛋白NRL31可以与PHOT1发生体内互作, 因此将其重命名为NCH1 (NRL PROTEIN FORCHLOROPLASTMOVEMENT 1).DEFECTIVELY ORGANIZED TRIBUTARIES 3/NRL23是与NPH3来自同一个分支的NRL家族蛋白, 主要参与维管组织发育以及叶脉结构调节(Petricka et al., 2008).虽然NRL23与NPH3的亲缘关系较近, 但是该蛋白是否在向光素介导的信号途径中发挥作用尚未见报道.NRL5、NRL6、NRL7、NRL15、NRL20、NRL21、NRL26和NRL30是分支III中的NPY (NAKED PINS IN YUCCA)家族成员, 其中NRL6/NPY2、NRL7/NPY4、NRL20/NPY1、NRL21/NPY5和NRL30/NPY3以功能冗余的方式与AGCVIII激酶共同调节器官发育以及向重力性反应中生长素的运输(Cheng et al., 2008; Li et al., 2011).通过分析NPYs基因对应的双突变体、三突变体、四突变体以及五突变体根的向重力性表型, 发现双突变体和三突变体向重力性表型缺失不明显; 四突变体表现不同程度的向重力性缺失表型, 其中npy1/npy2/npy4/npy5和npy2/npy3/npy4/npy5严重缺失向重力性; 五突变体的根几乎完全缺失向重力性, 在21°到150°之间随机生长(Li et al., 2011). ...
Blue light induces global and localized conformational changes in the kinase domain of full-length phototropin 1 2010
... 植物向光素PHOT1 (PHOTOTROPIN1)和PHOT2是一类细胞质膜结合的Ser/Thr蛋白激酶, 其C端为激酶结构域, N端含有2个光、氧气和电压敏感的LOV (LIGHT, OXYGEN, OR VOLTAGE SENSING)结构域, 分别命名为LOV1和LOV2 (Christie et al., 2015).蓝光照射后诱导向光素蛋白发生构象改变及自我磷酸化激活(Pfeifer et al., 2010), 活化的向光素蛋白进而调节下胚轴向光弯曲反应(Sakai et al., 2001)、叶绿体运动(Kagawa et al., 2001)、气孔开放(Kinoshita et al., 2001)以及叶片的伸展与定位等(Sakamoto and Briggs, 2002; Inoue et al., 2008b), 从而使植物或叶片处于最有利于吸收光的位置, 优化植物的光合作用(Christie et al., 2015). ...
Modulation of phototropic responsiveness in Arabidopsis through ubiquitination of phototropin 1 by the CUL3-ring E3 ubiquitin ligase CRL3 NPH3. 4 2011
... NPH3主要通过2种途径影响生长素的不对称分布.一是NPH3通过多泛素化修饰PHOT1促进PHOT1的降解, 间接影响生长素转运蛋白(如生长素外流载体PIN1 (Roberts et al., 2011)和PIN2 (Wan et al., 2012))的重定位.NPH3是以Cullin3为基础的E3泛素化连接酶CRL3NPH3复合体的组成成分, 在该复合体中作为底物连接蛋白发挥作用(Roberts et al., 2011; Wan et al., 2012).强蓝光下, CRL3NPH3复合体直接与PHOT1蛋白相互作用, 促进PHOT1的单/多泛素化修饰, 多泛素化修饰的PHOT1进入细胞质后被26S蛋白酶体识别后降解(Roberts et al., 2011).PHOT1从细胞膜进入细胞质影响了生长素外流载体PIN1 (Roberts et al., 2011)和PIN2 (Wan et al., 2012)的重定位, 但具体作用机制有待深入研究.二是NPH3通过与PKS家族蛋白互作调节生长素的转运(Fankhauser and Christie, 2015).PKS家族成员是向光弯曲反应的正向调节因子, 参与生长素转运调节, 它们可以与向光素和NPH3蛋白直接互作, 形成PHOT1-PKS-NPH3复合体(Kami et al., 2014), 但是复合体的作用机制目前还不清楚. ...
... 复合体的组成成分, 在该复合体中作为底物连接蛋白发挥作用(Roberts et al., 2011; Wan et al., 2012).强蓝光下, CRL3NPH3复合体直接与PHOT1蛋白相互作用, 促进PHOT1的单/多泛素化修饰, 多泛素化修饰的PHOT1进入细胞质后被26S蛋白酶体识别后降解(Roberts et al., 2011).PHOT1从细胞膜进入细胞质影响了生长素外流载体PIN1 (Roberts et al., 2011)和PIN2 (Wan et al., 2012)的重定位, 但具体作用机制有待深入研究.二是NPH3通过与PKS家族蛋白互作调节生长素的转运(Fankhauser and Christie, 2015).PKS家族成员是向光弯曲反应的正向调节因子, 参与生长素转运调节, 它们可以与向光素和NPH3蛋白直接互作, 形成PHOT1-PKS-NPH3复合体(Kami et al., 2014), 但是复合体的作用机制目前还不清楚. ...
... 复合体直接与PHOT1蛋白相互作用, 促进PHOT1的单/多泛素化修饰, 多泛素化修饰的PHOT1进入细胞质后被26S蛋白酶体识别后降解(Roberts et al., 2011).PHOT1从细胞膜进入细胞质影响了生长素外流载体PIN1 (Roberts et al., 2011)和PIN2 (Wan et al., 2012)的重定位, 但具体作用机制有待深入研究.二是NPH3通过与PKS家族蛋白互作调节生长素的转运(Fankhauser and Christie, 2015).PKS家族成员是向光弯曲反应的正向调节因子, 参与生长素转运调节, 它们可以与向光素和NPH3蛋白直接互作, 形成PHOT1-PKS-NPH3复合体(Kami et al., 2014), 但是复合体的作用机制目前还不清楚. ...
... ).PHOT1从细胞膜进入细胞质影响了生长素外流载体PIN1 (Roberts et al., 2011)和PIN2 (Wan et al., 2012)的重定位, 但具体作用机制有待深入研究.二是NPH3通过与PKS家族蛋白互作调节生长素的转运(Fankhauser and Christie, 2015).PKS家族成员是向光弯曲反应的正向调节因子, 参与生长素转运调节, 它们可以与向光素和NPH3蛋白直接互作, 形成PHOT1-PKS-NPH3复合体(Kami et al., 2014), 但是复合体的作用机制目前还不清楚. ...
In the light of stomatal opening: new insights into 'the Watergate' 1 2005
... 拟南芥气孔由1对保卫细胞构成, 气孔通过调节其开闭程度控制着与环境的气体交换来响应外界的刺激信号(Hetherington and Woodward, 2003; Roelfsema and Hedrich, 2005).红光和蓝光都能诱导气孔快速张开(Shimazaki et al., 2007).红光通过影响叶肉细胞和保卫细胞中叶绿体的光合作用调节气孔开放(Mott et al., 2008; Suetsugu et al., 2014).蓝光作为信号通过激活保卫细胞质膜H+-ATPase诱导气孔开放(Assmann et al., 1985; Kinoshita et al., 2001).蓝光诱导的气孔开放受蓝光受体向光素(PHOT1和PHOT2)调节(Kinoshita et al., 2001).目前人们对向光素介导的气孔开放机制研究较为清楚(Inoue et al., 2008b; Inoue and Kinoshita, 2017).蓝光照射下, 自磷酸化激活的PHOT1直接磷酸化蛋白激酶BLUS1 (LIGHT SIGNALING1), 磷酸化的BLUS1间接将信号传递给I型蛋白磷酸酶(PPI)以及该酶的亚基PRSL1 (Takemiya and Shimazaki, 2016).PPI进一步将信号传递给质膜AHA1型H+-ATPase, 促进H+-ATPase的激活.H+-ATPase的激活需要其C末端倒数第2个苏氨酸的磷酸化以及与14-3-3蛋白的结合(Yamauchi et al., 2016).活化的H+-ATPase通过驱动保卫细胞内H+的跨膜输出诱导保卫细胞质膜的超极化(Shimazaki et al., 2007; Marten et al., 2010).超极化的质膜激活内向K+通道, 促进K+在保卫细胞胞质中积累.为了维持胞质的电荷平衡, Cl-和malate2-随之大量增加, 增加的K+、Cl-和malate2-进一步被转入液泡, 使保卫细胞水势降低, 外界水分大量进入保卫细胞, 促进保卫细胞体积膨胀, 气孔张开(Inoue and Kinoshita, 2017).保卫细胞胞质中Cl-的增加主要是由于向光素抑制了S型阴离子外流蛋白的活性(Sussmilch et al., 2019).向光素对S型阴离子外流蛋白活性的抑制需要2个蛋白激酶CBC1/2 (CONVERGENCE OF BLUE LIGHT (BL) and CO21/2)参与(Hiyama et al., 2017; Sussmilch et al., 2019). ...
Arabidopsis nph1 and npl1: blue light receptors that mediate both phototropism and chloroplast relocation 4 2001
... 植物向光素PHOT1 (PHOTOTROPIN1)和PHOT2是一类细胞质膜结合的Ser/Thr蛋白激酶, 其C端为激酶结构域, N端含有2个光、氧气和电压敏感的LOV (LIGHT, OXYGEN, OR VOLTAGE SENSING)结构域, 分别命名为LOV1和LOV2 (Christie et al., 2015).蓝光照射后诱导向光素蛋白发生构象改变及自我磷酸化激活(Pfeifer et al., 2010), 活化的向光素蛋白进而调节下胚轴向光弯曲反应(Sakai et al., 2001)、叶绿体运动(Kagawa et al., 2001)、气孔开放(Kinoshita et al., 2001)以及叶片的伸展与定位等(Sakamoto and Briggs, 2002; Inoue et al., 2008b), 从而使植物或叶片处于最有利于吸收光的位置, 优化植物的光合作用(Christie et al., 2015). ...
... 拟南芥(Arabidopsis thaliana) PHOT1和PHOT2以功能冗余的方式调节下胚轴向光性和根负向光性、叶绿体聚光运动、气孔开放以及叶片伸展与定位等(Christie et al., 2015).PHOT1特异性抑制光下黄化幼苗下胚轴的快速伸长(Folta et al., 2003)、子叶变绿过程中叶绿素结合蛋白mRNA转录的稳定性(Folta and Kaufman, 2003)以及侧根的快速生长(Moni et al., 2015).PHOT2不仅介导强蓝光诱导的下胚轴向光弯曲(Sakai et al., 2001), 而且也特异调节叶绿体和细胞核的避光运动(Higa et al., 2014)以及抗性蛋白介导的病毒防御反应(Jeong et al., 2010).PHOT1和PHOT2的功能差异可能与磷酸化修饰位点、磷酸化程度、亚细胞定位变化以及下游相互作用蛋白种类不同有关(Zhao et al., 2013; Morrow et al., 2018; Liscum et al., 2020). ...
... 早在达尔文(1880年)时期, 人们就已发现植物向光性, 但是直到向光素PHOT1和PHOT2先后被鉴定(Liscum and Briggs, 1995; Christie et al., 1998), 植物的向光性机理研究才取得突飞猛进的发展.研究发现, PHOT1调节弱蓝光(0.01-1 μmol·m-2·s-1)和强蓝光(>1 μmol·m-2·s-1)诱导的下胚轴的向光性; PHOT2仅在强蓝光下起作用, 且与PHOT1功能冗余(Sakai et al., 2001; Briggs and Christie, 2002).由于强蓝光下PHOT1和PHOT2的功能冗余, 目前向光性机制研究主要集中在PHOT1介导的弱蓝光反应(Motchoulski and Liscum, 1999; Haga et al., 2015), 仅有少量研究通过突变体背景下的抑制子筛选寻找参与强蓝光信号途径的作用因子(赵翔等, 2014).单侧蓝光照射下, 自磷酸化激活的向光素蛋白将信号传递给NPH3、RPT2以及PKS等下游因子(Motchoulski and Liscum, 1999; Inada et al., 2004), 调节生长素运输载体的活性与定位(Ding et al., 2011).例如, 引起内皮层细胞背光侧PIN3 (PIN-FORMED3)积累(刘广超和丁兆军, 2018), 促进背光侧生长素含量增加、细胞伸长速率加快, 最终引起植物弯向光生长(Christie and Murphy, 2013).下胚轴弯曲过程中生长素的不对称分布受各类生长素运输载体的共同调控, 如PINs (PIN3、PIN4和PIN7)和ABCB19 (Christie et al., 2011).蓝光下, PIN3的磷酸化受抑制, 促进PIN3的背光侧定位, 诱导背光侧生长素积累以及下胚轴的向光性生长(Ding et al., 2011).PIN3的磷酸化以及极性定位受AGC激酶VIII亚族成员PID (PINOID)以及D6PK (D6 PROTEIN KINASE)的调节(Ding et al., 2011; Willige et al., 2013).ABCB19是生长素外流载体, 被蓝光激活的PHOT1磷酸化后, ABCB19的生长素转运活性受抑制, 增加拟南芥下胚轴顶端生长素含量并抑制其垂直生长, 同时通过PIN3蛋白促进生长素在下胚轴中侧向移动(Christie et al., 2011).背光侧生长素的大量积累促进了背光侧IAA19的降解, 释放出有活性的ARF7 (Sun et al., 2013), 不对称激活与生长相关基因的表达促进了下胚轴的向光性反应(刘广超和丁兆军, 2018). ...
... 叶片是植物进行光合作用的场所.自然光下, 野生型拟南芥叶柄向上倾斜生长, 叶片的叶面平整, 几乎平行于水平方向生长, 该现象被称为叶片的伸展与定位.弱光下, 拟南芥phot2突变体叶片伸展与定位表型几乎与野生型相同, phot1突变体叶片的定位发生变化, 叶柄与水平面夹角减小(Harada et al., 2013), 表明弱光下PHOT1单独调节叶片的伸展与定位(Demarsy and Fankhauser, 2009).强光下, 拟南芥phot1和phot2单突变体叶片伸展与定位的表型与野生型基本一致, phot1/phot2双突变体叶柄略微向下倾斜生长, 叶片向背面卷曲, 以几乎垂直于水平面的角度悬挂于叶柄上, 表明PHOT1和PHOT2以功能冗余的方式调节强光下叶片的伸展与定位(Sakai et al., 2001).叶片的伸展与定位可以确保叶片采取最佳的茎叶角度, 以便最大程度地捕获光源并且趋避强光伤害(Harada et al., 2013; Shang et al., 2019).与此一致, 拟南芥的研究表明: 弱光下, phot2的鲜重与野生型几乎相等, phot1的鲜重是野生型的1/2; 强光下, phot1和phot2单突变体的鲜重与野生型基本一致, 但phot1/phot2双突变体的鲜重大约是野生型的1/3 (Harada et al., 2013). ...
RPT2: a signal transducer of the phototropic response in Arabidopsis 2 2000
... 拟南芥中ABCB19 (ATP-BINDING CAS-SETTE B 19)和光敏色素激酶底物(PHYTOCHROME KINASE SUBSTRATE 4, PKS4)参与调节生长素的转运, 是下胚轴向光弯曲的负调节因子(Christie et al., 2011; Demarsy et al., 2012).自磷酸化激活的PHOT1通过直接磷酸化ABCB19和PKS4, 抑制ABCB19和PKS4的生长素转运活性, 促进下胚轴向光弯曲反应(Christie et al., 2011; Demarsy et al., 2012).PHOT1通过磷酸化修饰调节ABCB19的活性, 但是PHOT2并不影响ABCB19的活性(Christie et al., 2011).蛋白磷酸酶2A (PP2A)的A1亚基对应的蛋白RCN1 (Root Curling in N-naphthylphthalamic acid 1)与PHOT2蛋白发生体外互作, 负调节PHOT2介导的向光弯曲, 但RCN1并不与PHOT1直接相互作用(Tseng and Briggs, 2010).NPH3 (NONPHOTOTROPIC HYPOCOTYL 3)和RPT2 (ROOT PHOTOTROPISM 2)是通过筛选拟南芥下胚轴以及根的向光性缺失突变体获得(Motchoulski and Liscum, 1999; Sakai et al., 2000).随后的研究发现, NPH3和RPT2是向光素途径的重要调控因子, 均可与PHOT1蛋白直接相互作用(Blakeslee et al., 2004; Stone et al., 2008).虽然NPH3和RPT2也参与PHOT2信号转导途径, 但目前并无RPT2与PHOT2蛋白之间直接相互作用的报道(Inada et al., 2004; Lariguet et al., 2006).上述结果暗示PHOT1和PHOT2可能通过与不同的下游互作因子差异调节蓝光反应. ...
... 拟南芥rpt2单突变体在极弱光下(≤0.01 μmol·m-2·s-1)下胚轴向光弯曲反应正常, 随着光照强度的增大弯曲度逐渐降低(Inada et al., 2004), 表明RPT2仅参与调节强蓝光诱导的下胚轴向光弯曲反应.phot1/rpt2双突变体与phot1单突变体表型类似, 在10和100 μmol·m-2·s-1的较强蓝光下可以发生下胚轴向光弯曲反应, 但在0.01-1 μmol·m-2·s-1的弱蓝光下则缺失向光弯曲(Sakai et al., 2000), 暗示RPT2作为PHOT1的下游因子调节强蓝光诱导的下胚轴向光弯曲反应.phot2/rpt2双突变体与phot1/phot2双突变体的表型一致, 缺失强蓝光诱导的下胚轴向光弯曲反应(Inada et al., 2004; Zhao et al., 2018), 且PHOT1和RPT2可以直接相互作用.上述结果进一步证明, RPT2确实参与调节PHOT1介导的下胚轴向光弯曲反应.如果RPT2仅作为PHOT1的下游因子发挥作用, 那么rpt2单突变体应该与phot1单突变体表型一致.表型分析结果显示, rpt2单突变体与phot1/phot2双突变体的表型一致, 即缺失强蓝光诱导的下胚轴向光弯曲反应(Inada et al., 2004), 证明RPT2在PHOT2介导的下胚轴向光弯曲反应中也发挥重要作用.Harada等(2013)认为, RPT2在PHOT1抑制PHOT2途径的过程中发挥作用, 阻碍PHOT1对PHOT2途径的抑制.最近研究表明, RPT2可与PHOT1的LOV1区相互作用并抑制PHOT1的自磷酸化, 以降低PHOT1的光敏感性, 从而行使分子变阻器(molecular rheostat)的功能, 将不同强度蓝光下PHOT1的活性维持在适当的水平.尽管RPT2也可以与PHOT2的LOV1区域相互作用, 但是RPT2不抑制PHOT2的活性(Kimura et al., 2020). ...
Cellular and subcellular localization of phototropin 1 1 2002
... 植物向光素PHOT1 (PHOTOTROPIN1)和PHOT2是一类细胞质膜结合的Ser/Thr蛋白激酶, 其C端为激酶结构域, N端含有2个光、氧气和电压敏感的LOV (LIGHT, OXYGEN, OR VOLTAGE SENSING)结构域, 分别命名为LOV1和LOV2 (Christie et al., 2015).蓝光照射后诱导向光素蛋白发生构象改变及自我磷酸化激活(Pfeifer et al., 2010), 活化的向光素蛋白进而调节下胚轴向光弯曲反应(Sakai et al., 2001)、叶绿体运动(Kagawa et al., 2001)、气孔开放(Kinoshita et al., 2001)以及叶片的伸展与定位等(Sakamoto and Briggs, 2002; Inoue et al., 2008b), 从而使植物或叶片处于最有利于吸收光的位置, 优化植物的光合作用(Christie et al., 2015). ...
Mapping of low- and high-fluence autophosphorylation sites in phototropin 1 1 2003
... 拟南芥PHOT1至少有21个氨基酸位点可以发生自磷酸化(Boex-Fontvieille et al., 2014; Deng et al., 2014), PHOT2有29个磷酸化位点(Inoue et al., 2011; Boex-Fontvieille et al., 2014), 这些位点的具体分布参见Christie等(2015)的报道.其中, C端2个保守位点的磷酸化(PHOT1中S849和S851, PHOT2中S761 和S763)对向光素受体的信号传递至关重要(Inoue et al., 2008a; Inoue et al., 2011), 其余的磷酸化位点大部分位于N端LOV1的上游以及LOV1与LOV2中间的连接区域(Christie et al., 2015).虽然这些上游区域磷酸化位点的生物学意义目前还不清楚, 但这些位点的磷酸化程度受光照强度调节(Salomon et al., 2003), 暗示PHOT1和PHOT2可能通过磷酸化程度差异响应不同的光照强度, 从而参与植物多种生理活动的调节. ...
Functional characterization of GhPHOT2 in chloroplast avoidance of Gossypium hirsutum 1 2019
... 叶片是植物进行光合作用的场所.自然光下, 野生型拟南芥叶柄向上倾斜生长, 叶片的叶面平整, 几乎平行于水平方向生长, 该现象被称为叶片的伸展与定位.弱光下, 拟南芥phot2突变体叶片伸展与定位表型几乎与野生型相同, phot1突变体叶片的定位发生变化, 叶柄与水平面夹角减小(Harada et al., 2013), 表明弱光下PHOT1单独调节叶片的伸展与定位(Demarsy and Fankhauser, 2009).强光下, 拟南芥phot1和phot2单突变体叶片伸展与定位的表型与野生型基本一致, phot1/phot2双突变体叶柄略微向下倾斜生长, 叶片向背面卷曲, 以几乎垂直于水平面的角度悬挂于叶柄上, 表明PHOT1和PHOT2以功能冗余的方式调节强光下叶片的伸展与定位(Sakai et al., 2001).叶片的伸展与定位可以确保叶片采取最佳的茎叶角度, 以便最大程度地捕获光源并且趋避强光伤害(Harada et al., 2013; Shang et al., 2019).与此一致, 拟南芥的研究表明: 弱光下, phot2的鲜重与野生型几乎相等, phot1的鲜重是野生型的1/2; 强光下, phot1和phot2单突变体的鲜重与野生型基本一致, 但phot1/phot2双突变体的鲜重大约是野生型的1/3 (Harada et al., 2013). ...
Light regulation of stomatal movement 2 2007
... 拟南芥气孔由1对保卫细胞构成, 气孔通过调节其开闭程度控制着与环境的气体交换来响应外界的刺激信号(Hetherington and Woodward, 2003; Roelfsema and Hedrich, 2005).红光和蓝光都能诱导气孔快速张开(Shimazaki et al., 2007).红光通过影响叶肉细胞和保卫细胞中叶绿体的光合作用调节气孔开放(Mott et al., 2008; Suetsugu et al., 2014).蓝光作为信号通过激活保卫细胞质膜H+-ATPase诱导气孔开放(Assmann et al., 1985; Kinoshita et al., 2001).蓝光诱导的气孔开放受蓝光受体向光素(PHOT1和PHOT2)调节(Kinoshita et al., 2001).目前人们对向光素介导的气孔开放机制研究较为清楚(Inoue et al., 2008b; Inoue and Kinoshita, 2017).蓝光照射下, 自磷酸化激活的PHOT1直接磷酸化蛋白激酶BLUS1 (LIGHT SIGNALING1), 磷酸化的BLUS1间接将信号传递给I型蛋白磷酸酶(PPI)以及该酶的亚基PRSL1 (Takemiya and Shimazaki, 2016).PPI进一步将信号传递给质膜AHA1型H+-ATPase, 促进H+-ATPase的激活.H+-ATPase的激活需要其C末端倒数第2个苏氨酸的磷酸化以及与14-3-3蛋白的结合(Yamauchi et al., 2016).活化的H+-ATPase通过驱动保卫细胞内H+的跨膜输出诱导保卫细胞质膜的超极化(Shimazaki et al., 2007; Marten et al., 2010).超极化的质膜激活内向K+通道, 促进K+在保卫细胞胞质中积累.为了维持胞质的电荷平衡, Cl-和malate2-随之大量增加, 增加的K+、Cl-和malate2-进一步被转入液泡, 使保卫细胞水势降低, 外界水分大量进入保卫细胞, 促进保卫细胞体积膨胀, 气孔张开(Inoue and Kinoshita, 2017).保卫细胞胞质中Cl-的增加主要是由于向光素抑制了S型阴离子外流蛋白的活性(Sussmilch et al., 2019).向光素对S型阴离子外流蛋白活性的抑制需要2个蛋白激酶CBC1/2 (CONVERGENCE OF BLUE LIGHT (BL) and CO21/2)参与(Hiyama et al., 2017; Sussmilch et al., 2019). ...
... 的跨膜输出诱导保卫细胞质膜的超极化(Shimazaki et al., 2007; Marten et al., 2010).超极化的质膜激活内向K+通道, 促进K+在保卫细胞胞质中积累.为了维持胞质的电荷平衡, Cl-和malate2-随之大量增加, 增加的K+、Cl-和malate2-进一步被转入液泡, 使保卫细胞水势降低, 外界水分大量进入保卫细胞, 促进保卫细胞体积膨胀, 气孔张开(Inoue and Kinoshita, 2017).保卫细胞胞质中Cl-的增加主要是由于向光素抑制了S型阴离子外流蛋白的活性(Sussmilch et al., 2019).向光素对S型阴离子外流蛋白活性的抑制需要2个蛋白激酶CBC1/2 (CONVERGENCE OF BLUE LIGHT (BL) and CO21/2)参与(Hiyama et al., 2017; Sussmilch et al., 2019). ...
Disruptions in AUX1-dependent auxin influx alter hypocotyl phototropism in Arabidopsis 1 2008
... 拟南芥中ABCB19 (ATP-BINDING CAS-SETTE B 19)和光敏色素激酶底物(PHYTOCHROME KINASE SUBSTRATE 4, PKS4)参与调节生长素的转运, 是下胚轴向光弯曲的负调节因子(Christie et al., 2011; Demarsy et al., 2012).自磷酸化激活的PHOT1通过直接磷酸化ABCB19和PKS4, 抑制ABCB19和PKS4的生长素转运活性, 促进下胚轴向光弯曲反应(Christie et al., 2011; Demarsy et al., 2012).PHOT1通过磷酸化修饰调节ABCB19的活性, 但是PHOT2并不影响ABCB19的活性(Christie et al., 2011).蛋白磷酸酶2A (PP2A)的A1亚基对应的蛋白RCN1 (Root Curling in N-naphthylphthalamic acid 1)与PHOT2蛋白发生体外互作, 负调节PHOT2介导的向光弯曲, 但RCN1并不与PHOT1直接相互作用(Tseng and Briggs, 2010).NPH3 (NONPHOTOTROPIC HYPOCOTYL 3)和RPT2 (ROOT PHOTOTROPISM 2)是通过筛选拟南芥下胚轴以及根的向光性缺失突变体获得(Motchoulski and Liscum, 1999; Sakai et al., 2000).随后的研究发现, NPH3和RPT2是向光素途径的重要调控因子, 均可与PHOT1蛋白直接相互作用(Blakeslee et al., 2004; Stone et al., 2008).虽然NPH3和RPT2也参与PHOT2信号转导途径, 但目前并无RPT2与PHOT2蛋白之间直接相互作用的报道(Inada et al., 2004; Lariguet et al., 2006).上述结果暗示PHOT1和PHOT2可能通过与不同的下游互作因子差异调节蓝光反应. ...
PLASTID MOVEMENT IMPAIRED 1 and PLASTID MOVEMENT IMPAIRED1-RELATED 1 mediate photorelocation movements of both chloroplasts and nuclei 1 2015
... KAC (KINESIN-LIKE PROTEIN FOR ACTIN-BASED CHLOROPLAST MOVEMENT)参与调节叶绿体肌动蛋白纤维的聚集(Wada, 2013; Kong and Wada, 2014), WEB1 (WEAK CHLOROPLAST MOVEMENT UNDER BLUE LIGHT 1)和PMI1 (PLASTIDMOVEMENT IMPAIRED1)对叶绿体肌动蛋白纤维的稳定性至关重要(Suetsugu et al., 2015). ...
Guard cell chloroplasts are essential for blue light-dependent stomatal opening in Arabidopsis 1 2014
... 拟南芥气孔由1对保卫细胞构成, 气孔通过调节其开闭程度控制着与环境的气体交换来响应外界的刺激信号(Hetherington and Woodward, 2003; Roelfsema and Hedrich, 2005).红光和蓝光都能诱导气孔快速张开(Shimazaki et al., 2007).红光通过影响叶肉细胞和保卫细胞中叶绿体的光合作用调节气孔开放(Mott et al., 2008; Suetsugu et al., 2014).蓝光作为信号通过激活保卫细胞质膜H+-ATPase诱导气孔开放(Assmann et al., 1985; Kinoshita et al., 2001).蓝光诱导的气孔开放受蓝光受体向光素(PHOT1和PHOT2)调节(Kinoshita et al., 2001).目前人们对向光素介导的气孔开放机制研究较为清楚(Inoue et al., 2008b; Inoue and Kinoshita, 2017).蓝光照射下, 自磷酸化激活的PHOT1直接磷酸化蛋白激酶BLUS1 (LIGHT SIGNALING1), 磷酸化的BLUS1间接将信号传递给I型蛋白磷酸酶(PPI)以及该酶的亚基PRSL1 (Takemiya and Shimazaki, 2016).PPI进一步将信号传递给质膜AHA1型H+-ATPase, 促进H+-ATPase的激活.H+-ATPase的激活需要其C末端倒数第2个苏氨酸的磷酸化以及与14-3-3蛋白的结合(Yamauchi et al., 2016).活化的H+-ATPase通过驱动保卫细胞内H+的跨膜输出诱导保卫细胞质膜的超极化(Shimazaki et al., 2007; Marten et al., 2010).超极化的质膜激活内向K+通道, 促进K+在保卫细胞胞质中积累.为了维持胞质的电荷平衡, Cl-和malate2-随之大量增加, 增加的K+、Cl-和malate2-进一步被转入液泡, 使保卫细胞水势降低, 外界水分大量进入保卫细胞, 促进保卫细胞体积膨胀, 气孔张开(Inoue and Kinoshita, 2017).保卫细胞胞质中Cl-的增加主要是由于向光素抑制了S型阴离子外流蛋白的活性(Sussmilch et al., 2019).向光素对S型阴离子外流蛋白活性的抑制需要2个蛋白激酶CBC1/2 (CONVERGENCE OF BLUE LIGHT (BL) and CO21/2)参与(Hiyama et al., 2017; Sussmilch et al., 2019). ...
RPT2/NCH1 subfamily of NPH3-like proteins is essential for the chloroplast accumulation response in land plants 17 2016
... NRL (NPH3/RPT2-Like)蛋白家族是植物特有的, 其成员含有3个保守结构域: N端的BTB (Bric àbrac, Tramtrack, and Broad complex)结构域、中间的NPH3结构域以及C端的螺旋-螺旋结构域(Liscum et al., 2014).其中, BTB结构域主要起E3连接酶的作用, 与CUL3发生相互作用(Gingerich et al., 2005; Figueroa et al., 2005), C端的螺旋-螺旋结构域主要介导蛋白之间的相互作用(Inada et al., 2004).拟南芥中33个NRL家族成员都含有NPH3结构域, 10个缺少C端的螺旋-螺旋结构域, 2个缺少N端的BTB结构域(Pedmale et al., 2010; Christie et al., 2018).根据NRL家族成员间的进化关系, 拟南芥中的33个NRL家族成员可分为6个分支(图1) (Suetsugu et al., 2016; Christie et al., 2018).目前, 来自2个分支的10个成员的功能已得到鉴定(de Carbonnel et al., 2010; Christie et al., 2018) (图1). ...
... Construction of this phylogenetic tree was based on Suetsugu et al. (2016) and Christie et al. (2018) with slight modifications. The colors and names of six major clades of NRL proteins in Arabidopsis were constructed using the Adobe illustrator software. ...
... NPH3和RPT2是来自2个不同分支的NRL家族成员, 通过与向光素相互作用调节蓝光诱导下胚轴向光性、叶绿体聚光运动、气孔开放以及叶片伸展与定位(Christie et al., 2015, 2018).NRL8是AGC激酶(cAMP依赖的蛋白激酶A, cGMP依赖的蛋白激酶G和磷脂依赖的蛋白激酶C)活性依赖的花粉管生长调节因子(Lalanne et al., 2004; Zhang et al., 2008), 该蛋白与RPT2来自分支II.与NPH3类似, SR1IP1 (AtSR1 INTERACTING PROTEIN 1)也称NRL31, 可以与CRL3 ( CUL3 RING E3 UBIQUITIN LIGASE)形成复合体, 调节底物的泛素化(Zhang et al., 2014).SR1IP1通过调节拟南芥中Ca2+/钙调素结合的转录因子AtSR1 (SIGNAL RESPONSIVE 1)的泛素化降解, 增强植物对细菌性病原体的抗性(Zhang et al., 2014).但系统进化研究发现, SR1IP1也来自分支II, 且比NRL8与RPT2亲缘关系更近, 与NPH3的进化明显不同.Suetsugu等(2016)在筛选参与调节叶绿体运动的NRL家族蛋白时, 发现NRL31基因突变抑制了光诱导的叶绿体运动, 且蛋白NRL31可以与PHOT1发生体内互作, 因此将其重命名为NCH1 (NRL PROTEIN FORCHLOROPLASTMOVEMENT 1).DEFECTIVELY ORGANIZED TRIBUTARIES 3/NRL23是与NPH3来自同一个分支的NRL家族蛋白, 主要参与维管组织发育以及叶脉结构调节(Petricka et al., 2008).虽然NRL23与NPH3的亲缘关系较近, 但是该蛋白是否在向光素介导的信号途径中发挥作用尚未见报道.NRL5、NRL6、NRL7、NRL15、NRL20、NRL21、NRL26和NRL30是分支III中的NPY (NAKED PINS IN YUCCA)家族成员, 其中NRL6/NPY2、NRL7/NPY4、NRL20/NPY1、NRL21/NPY5和NRL30/NPY3以功能冗余的方式与AGCVIII激酶共同调节器官发育以及向重力性反应中生长素的运输(Cheng et al., 2008; Li et al., 2011).通过分析NPYs基因对应的双突变体、三突变体、四突变体以及五突变体根的向重力性表型, 发现双突变体和三突变体向重力性表型缺失不明显; 四突变体表现不同程度的向重力性缺失表型, 其中npy1/npy2/npy4/npy5和npy2/npy3/npy4/npy5严重缺失向重力性; 五突变体的根几乎完全缺失向重力性, 在21°到150°之间随机生长(Li et al., 2011). ...
... 目前已经鉴定的10个有功能的NRL家族蛋白中有3个(NPH3、RPT2和NCH1)可与向光素发生直接相互作用(Motchoulski and Liscum, 1999; Inada et al., 2004; Suetsugu et al., 2016).此外, 酵母双杂交筛选发现NRL2可以与PHOT1发生相互作用(Sullivan et al., 2009), 但是NRL2的功能及其在向光素介导的信号途径中的作用目前还不清楚.虽然大部分NRL家族成员的功能未知, 但已有的实验结果表明, NRL家族成员在向光素信号途径中起重要作用, 因此我们总结NRL家族蛋白在向光素介导的信号途径中的作用, 旨在为全面揭示NPH3/RPT2-Like家族成员的功能提供线索. ...
... NCH1是近年来鉴定到的1个NRL家族蛋白, 定位于质膜.体外Pull-down实验证明, NCH1可以与PHOT1、NPH3以及RPT2的N末端和C末端分别互作(Suetsugu et al., 2016).与nph3和rpt2单突变体类似, 蓝光诱导的PHOT1自磷酸化在nch1单突变体中能正常发生(Suetsugu et al., 2016), 表明NCH1功能类似于NPH3和RPT2, 也是1个与向光素相互作用的下游信号分子(Suetsugu et al., 2016).检测NCH1在下胚轴弯曲中的作用, 发现虽然nch1突变体的向光弯曲度仅稍微大于野生型, 但是在4和40 μmol·m-2·s-1的光下, nch1突变体与野生型的弯曲度存在明显差异.此外, rpt2/nch1双突变体的表型与rpt2单突变体相似(Suetsugu et al., 2016), 暗示NCH1可能不是向光素介导的向光性反应的正调节因子.综上, NRL家族成员NPH3、RPT2和NCH1参与下胚轴弯曲的调节, 其它成员是否调节向光素介导的下胚轴向光弯曲需深入研究. ...
... 单突变体中能正常发生(Suetsugu et al., 2016), 表明NCH1功能类似于NPH3和RPT2, 也是1个与向光素相互作用的下游信号分子(Suetsugu et al., 2016).检测NCH1在下胚轴弯曲中的作用, 发现虽然nch1突变体的向光弯曲度仅稍微大于野生型, 但是在4和40 μmol·m-2·s-1的光下, nch1突变体与野生型的弯曲度存在明显差异.此外, rpt2/nch1双突变体的表型与rpt2单突变体相似(Suetsugu et al., 2016), 暗示NCH1可能不是向光素介导的向光性反应的正调节因子.综上, NRL家族成员NPH3、RPT2和NCH1参与下胚轴弯曲的调节, 其它成员是否调节向光素介导的下胚轴向光弯曲需深入研究. ...
... ), 表明NCH1功能类似于NPH3和RPT2, 也是1个与向光素相互作用的下游信号分子(Suetsugu et al., 2016).检测NCH1在下胚轴弯曲中的作用, 发现虽然nch1突变体的向光弯曲度仅稍微大于野生型, 但是在4和40 μmol·m-2·s-1的光下, nch1突变体与野生型的弯曲度存在明显差异.此外, rpt2/nch1双突变体的表型与rpt2单突变体相似(Suetsugu et al., 2016), 暗示NCH1可能不是向光素介导的向光性反应的正调节因子.综上, NRL家族成员NPH3、RPT2和NCH1参与下胚轴弯曲的调节, 其它成员是否调节向光素介导的下胚轴向光弯曲需深入研究. ...
... 单突变体相似(Suetsugu et al., 2016), 暗示NCH1可能不是向光素介导的向光性反应的正调节因子.综上, NRL家族成员NPH3、RPT2和NCH1参与下胚轴弯曲的调节, 其它成员是否调节向光素介导的下胚轴向光弯曲需深入研究. ...
... 近年来, 科学家研究发现, 拟南芥NRL家族蛋白在向光素介导的蓝光信号转导通路中起重要作用.拟南芥NRL家族6个分支中的3个分支(II、III和IV) (图1)蛋白目前已经被报道参与向光素调节的反应, 或者与向光素蛋白存在直接相互作用, 暗示这3个分支的其它成员可能参与向光素信号途径的调节.RPT2与NCH1都来自分支II, 既参与生长素依赖的向光性调节, 又参与非生长素依赖的叶绿体聚光运动调节(Suetsugu et al., 2016; Suetsugu and Wada, 2017).目前RPT2和NCH1如何区分向光素介导的2种信号途径并不清楚.考虑到RPT2和NCH1蛋白所在分支与其余2个分支(III和IV)的亲缘关系较远(Christie et al., 2018), 暗示该分支其它成员也可能同时参与生长素依赖和非依赖性的向光素反应.RPT2可与PHOT1的LOV1区相互作用并抑制PHOT1的自磷酸化, 以降低PHOT1的光敏感性.尽管RPT2也可以与PHOT2的LOV1区域相互作用, 但是RPT2不抑制PHOT2的活性(Kimura et al., 2020).可能存在NRL家族其它蛋白与PHOT2相互作用, 调节PHOT2介导的蓝光信号转导.NPH3主要调节生长素依赖的向光性反应以及叶片的伸展与定位(Harada et al., 2013; Zhao et al., 2018), 但NPH3激活后调节生长素不对称分布的机制并不清楚.NPH3的磷酸化状态与其生理活性以及在细胞中的定位密切相关(Haga et al., 2015; Zhao et al., 2018).PHOT1介导的NPH3去磷酸化在弱蓝光诱导的下胚轴向光弯曲反应中发挥重要作用, 强蓝光下NPH3并未发生去磷酸化反应(Tsuchida-Mayama et al., 2008).那么NPH3如何调节强蓝光诱导的下胚轴向光弯曲反应, 是否与其E3泛素化连接酶活性有关? NPY1与PID互作, 通过影响生长素信号调节植物器官的形成以及根的向重力性反应(Suetsugu et al., 2016).生长素不对称分布决定不对称生长, PID- NPY1与PHOT-NPH3作用方式又相似, 那么NPY分支成员是否参与调节向光素介导的植物向性运动以及叶片的伸展与定位? PID和PHOT都属于AGC激酶VIII亚族成员(Ding et al., 2011; Willige et al., 2013), NRL家族蛋白是否可以与AGC激酶VIII亚家族其它成员相互作用, 以调节生长素介导的其它生理反应? 来自分支IV的NRL2也可以与PHOT1发生互作(Sullivan et al., 2009), 暗示该分支的蛋白也可能在向光素介导的蓝光信号途径中发挥重要作用.综上, NRL家族蛋白功能的陆续鉴定, 有望解析向光素PHOT1和PHOT2感受蓝光信号差异, 进而调节不同生理反应的可能机制. ...
... ).那么NPH3如何调节强蓝光诱导的下胚轴向光弯曲反应, 是否与其E3泛素化连接酶活性有关? NPY1与PID互作, 通过影响生长素信号调节植物器官的形成以及根的向重力性反应(Suetsugu et al., 2016).生长素不对称分布决定不对称生长, PID- NPY1与PHOT-NPH3作用方式又相似, 那么NPY分支成员是否参与调节向光素介导的植物向性运动以及叶片的伸展与定位? PID和PHOT都属于AGC激酶VIII亚族成员(Ding et al., 2011; Willige et al., 2013), NRL家族蛋白是否可以与AGC激酶VIII亚家族其它成员相互作用, 以调节生长素介导的其它生理反应? 来自分支IV的NRL2也可以与PHOT1发生互作(Sullivan et al., 2009), 暗示该分支的蛋白也可能在向光素介导的蓝光信号途径中发挥重要作用.综上, NRL家族蛋白功能的陆续鉴定, 有望解析向光素PHOT1和PHOT2感受蓝光信号差异, 进而调节不同生理反应的可能机制. ...
Two coiled-coil proteins, WEB1 and PMI2, suppress the signaling pathway of chloroplast accumulation response that is mediated by two phototropin-interacting proteins, RPT2 and NCH1, in seed plants 4 2017
... 作为植物进行光合作用的细胞器, 叶绿体响应外界光照刺激而发生的位置变化称为叶绿体光下重定位.弱光照射下, 为最大程度地吸收光能, 叶绿体沿细胞平周壁方向聚集, 这种现象为聚光运动(accumulation movement).强光条件下, 为避免过强的光对自身造成伤害, 叶绿体沿细胞垂周壁聚集, 使之与光平行分布, 这种现象称为避光运动(avoidance movement) (Kasahara et al., 2004).叶绿体运动受蓝光调节, 蓝光受体向光素PHOT1和PHOT2共同调控叶绿体的聚光反应, 而PHOT2单独调节叶绿体的避光反应(Suetsugu and Wada, 2017).与向光素调节的其它多细胞协调性反应(如向光性、叶片的伸展与定位)不同, 叶绿体运动是细胞自主性反应, 不受基因表达(Kong and Wada, 2016; Wada, 2016)和生长素运输的调节(Christie et al., 2018), 最终由叶绿体肌动蛋白响应入射光的强度与方向而发生快速重组介导(Kadota et al., 2009; Kong and Wada, 2014).叶绿体肌动蛋白特异定位于细胞质膜和叶绿体之间, 在叶绿体前进方向的后面叶绿体肌动蛋白纤维迅速解聚, 前进方向的前面叶绿体肌动蛋白纤维迅速聚集, 这种不对称的叶绿体肌动蛋白纤维分布最终决定了叶绿体的运动方向和速度(Kong and Wada, 2016).CHUP1 (CHLOROPLAST UNUSUAL POSITIONING 1)和 ...
... 虽然目前人们还不清楚向光素在感受外界光照后如何通过调节CHUP1、PMI1、WEB1以及JAC蛋白活性影响叶绿体的运动, 但越来越多的证据表明, NRL家族蛋白的NCH1和RPT2是该信号途径的重要中间因子(Suetsugu et al., 2016).rpt2突变体rpt2-3和rpt2-4的聚光反应略微降低, 但是叶绿体的避光反应和暗恢复反应(撤掉强光后, 叶绿体快速恢复, 导致透光率迅速下降的反应称为暗恢复反应)与野生型相比无明显差异(Suetsugu et al., 2016), 这些结果表明RPT2参与调节叶绿体的聚光反应.nch1单突变体的叶绿体聚光反应减弱, 暗恢复反应缺失, 但是避光反应增强(Suetsugu et al., 2016), 证明NCH1参与调节叶绿体的聚光运动.目前已经证实RPT2和NCH1参与调节弱光下PHOT1和PHOT2介导的叶绿体聚光反应, 这种AGC-NRL调节模块不依赖于生长素的转运以及生长素介导的转录过程(Suetsugu et al., 2016), 可能依赖于JAC1 (J-DOMAIN PROTEIN REQUIRED FOR CHLOROPLAST ACCUMULATION RESPONSE 1)以及WEB1/PMI2介导的信号途径(Suetsugu and Wada, 2017).弱光下, RPT2/ NCH1通过JAC1依赖与JAC1不依赖途径抑制弱光下的叶绿体避光反应.强光下, WEB1/PMI2复合体通过抑制RPT2/NCH1和JAC1介导的聚光反应, 有效促进PHOT2介导的叶绿体避光反应(Suetsugu and Wada, 2017), 但是同一家族的NPH3并不调节叶绿体的运动(Inada et al., 2004). ...
... ).弱光下, RPT2/ NCH1通过JAC1依赖与JAC1不依赖途径抑制弱光下的叶绿体避光反应.强光下, WEB1/PMI2复合体通过抑制RPT2/NCH1和JAC1介导的聚光反应, 有效促进PHOT2介导的叶绿体避光反应(Suetsugu and Wada, 2017), 但是同一家族的NPH3并不调节叶绿体的运动(Inada et al., 2004). ...
... 近年来, 科学家研究发现, 拟南芥NRL家族蛋白在向光素介导的蓝光信号转导通路中起重要作用.拟南芥NRL家族6个分支中的3个分支(II、III和IV) (图1)蛋白目前已经被报道参与向光素调节的反应, 或者与向光素蛋白存在直接相互作用, 暗示这3个分支的其它成员可能参与向光素信号途径的调节.RPT2与NCH1都来自分支II, 既参与生长素依赖的向光性调节, 又参与非生长素依赖的叶绿体聚光运动调节(Suetsugu et al., 2016; Suetsugu and Wada, 2017).目前RPT2和NCH1如何区分向光素介导的2种信号途径并不清楚.考虑到RPT2和NCH1蛋白所在分支与其余2个分支(III和IV)的亲缘关系较远(Christie et al., 2018), 暗示该分支其它成员也可能同时参与生长素依赖和非依赖性的向光素反应.RPT2可与PHOT1的LOV1区相互作用并抑制PHOT1的自磷酸化, 以降低PHOT1的光敏感性.尽管RPT2也可以与PHOT2的LOV1区域相互作用, 但是RPT2不抑制PHOT2的活性(Kimura et al., 2020).可能存在NRL家族其它蛋白与PHOT2相互作用, 调节PHOT2介导的蓝光信号转导.NPH3主要调节生长素依赖的向光性反应以及叶片的伸展与定位(Harada et al., 2013; Zhao et al., 2018), 但NPH3激活后调节生长素不对称分布的机制并不清楚.NPH3的磷酸化状态与其生理活性以及在细胞中的定位密切相关(Haga et al., 2015; Zhao et al., 2018).PHOT1介导的NPH3去磷酸化在弱蓝光诱导的下胚轴向光弯曲反应中发挥重要作用, 强蓝光下NPH3并未发生去磷酸化反应(Tsuchida-Mayama et al., 2008).那么NPH3如何调节强蓝光诱导的下胚轴向光弯曲反应, 是否与其E3泛素化连接酶活性有关? NPY1与PID互作, 通过影响生长素信号调节植物器官的形成以及根的向重力性反应(Suetsugu et al., 2016).生长素不对称分布决定不对称生长, PID- NPY1与PHOT-NPH3作用方式又相似, 那么NPY分支成员是否参与调节向光素介导的植物向性运动以及叶片的伸展与定位? PID和PHOT都属于AGC激酶VIII亚族成员(Ding et al., 2011; Willige et al., 2013), NRL家族蛋白是否可以与AGC激酶VIII亚家族其它成员相互作用, 以调节生长素介导的其它生理反应? 来自分支IV的NRL2也可以与PHOT1发生互作(Sullivan et al., 2009), 暗示该分支的蛋白也可能在向光素介导的蓝光信号途径中发挥重要作用.综上, NRL家族蛋白功能的陆续鉴定, 有望解析向光素PHOT1和PHOT2感受蓝光信号差异, 进而调节不同生理反应的可能机制. ...
Deetiolation enhances phototropism by modulating NON-PHOTOTROPIC HYPOCOTYL 3 phosphorylation status 1 2019
... 拟南芥nph3突变体在任何强度的单侧蓝光下均表现缺失下胚轴向光性(Liscum and Briggs, 1996; Liscum et al., 2014; Fankhauser and Christie, 2015), 表明NPH3调节PHOT1和PHOT2介导蓝光诱导的下胚轴弯曲反应.黑暗条件下, 磷酸化的NPH3蛋白与PHOT1结合, 定位于细胞膜, 单侧弱蓝光照射下, NPH3迅速发生PHOT1依赖的去磷酸化反应, 从细胞膜上脱离(Haga et al., 2015; Zhao et al., 2018).目前磷酸化NPH3的激酶还不清楚, 但NPH3去磷酸化主要由I型蛋白磷酸酶介导(Pedmale and Liscum, 2007).去磷酸化后NPH3从细胞膜向胞质的转移降低了其介导向光弯曲反应的能力(Haga et al., 2015; Sullivan et al., 2019).随着光照强度的增加, 下胚轴的弯曲度呈强度依赖的钟形曲线(Christie and Murphy, 2013).随着光照时间的增加, RPT2蛋白含量增加, 与去磷酸化的NPH3结合形成RPT2-NPH3复合物并与PHOT1蛋白结合, 促进NPH3的磷酸化以及从胞质到细胞膜的重定位, 开启第二次向光弯曲(Haga et al., 2015; Zhao et al., 2018).虽然PHOT1- NPH3复合体在细胞膜的重定位对持续光照介导的二次向光弯曲至关重要(Haga et al., 2015; Christie et al., 2018; Zhao et al., 2018), 但其生物学意义还不清楚, 人们认为这种变化有可能对调节生长素的不对称分布有重要影响(Pedmale and Liscum, 2007; Haga et al., 2015). ...
Interaction specificity of Arabidopsis 14-3-3 proteins with phototropin receptor kinases 2 2009
... 目前已经鉴定的10个有功能的NRL家族蛋白中有3个(NPH3、RPT2和NCH1)可与向光素发生直接相互作用(Motchoulski and Liscum, 1999; Inada et al., 2004; Suetsugu et al., 2016).此外, 酵母双杂交筛选发现NRL2可以与PHOT1发生相互作用(Sullivan et al., 2009), 但是NRL2的功能及其在向光素介导的信号途径中的作用目前还不清楚.虽然大部分NRL家族成员的功能未知, 但已有的实验结果表明, NRL家族成员在向光素信号途径中起重要作用, 因此我们总结NRL家族蛋白在向光素介导的信号途径中的作用, 旨在为全面揭示NPH3/RPT2-Like家族成员的功能提供线索. ...
... 近年来, 科学家研究发现, 拟南芥NRL家族蛋白在向光素介导的蓝光信号转导通路中起重要作用.拟南芥NRL家族6个分支中的3个分支(II、III和IV) (图1)蛋白目前已经被报道参与向光素调节的反应, 或者与向光素蛋白存在直接相互作用, 暗示这3个分支的其它成员可能参与向光素信号途径的调节.RPT2与NCH1都来自分支II, 既参与生长素依赖的向光性调节, 又参与非生长素依赖的叶绿体聚光运动调节(Suetsugu et al., 2016; Suetsugu and Wada, 2017).目前RPT2和NCH1如何区分向光素介导的2种信号途径并不清楚.考虑到RPT2和NCH1蛋白所在分支与其余2个分支(III和IV)的亲缘关系较远(Christie et al., 2018), 暗示该分支其它成员也可能同时参与生长素依赖和非依赖性的向光素反应.RPT2可与PHOT1的LOV1区相互作用并抑制PHOT1的自磷酸化, 以降低PHOT1的光敏感性.尽管RPT2也可以与PHOT2的LOV1区域相互作用, 但是RPT2不抑制PHOT2的活性(Kimura et al., 2020).可能存在NRL家族其它蛋白与PHOT2相互作用, 调节PHOT2介导的蓝光信号转导.NPH3主要调节生长素依赖的向光性反应以及叶片的伸展与定位(Harada et al., 2013; Zhao et al., 2018), 但NPH3激活后调节生长素不对称分布的机制并不清楚.NPH3的磷酸化状态与其生理活性以及在细胞中的定位密切相关(Haga et al., 2015; Zhao et al., 2018).PHOT1介导的NPH3去磷酸化在弱蓝光诱导的下胚轴向光弯曲反应中发挥重要作用, 强蓝光下NPH3并未发生去磷酸化反应(Tsuchida-Mayama et al., 2008).那么NPH3如何调节强蓝光诱导的下胚轴向光弯曲反应, 是否与其E3泛素化连接酶活性有关? NPY1与PID互作, 通过影响生长素信号调节植物器官的形成以及根的向重力性反应(Suetsugu et al., 2016).生长素不对称分布决定不对称生长, PID- NPY1与PHOT-NPH3作用方式又相似, 那么NPY分支成员是否参与调节向光素介导的植物向性运动以及叶片的伸展与定位? PID和PHOT都属于AGC激酶VIII亚族成员(Ding et al., 2011; Willige et al., 2013), NRL家族蛋白是否可以与AGC激酶VIII亚家族其它成员相互作用, 以调节生长素介导的其它生理反应? 来自分支IV的NRL2也可以与PHOT1发生互作(Sullivan et al., 2009), 暗示该分支的蛋白也可能在向光素介导的蓝光信号途径中发挥重要作用.综上, NRL家族蛋白功能的陆续鉴定, 有望解析向光素PHOT1和PHOT2感受蓝光信号差异, 进而调节不同生理反应的可能机制. ...
PIF4 and PIF5 transcription factors link blue light and auxin to regulate the phototropic response in Arabidopsis 1 2013
... 早在达尔文(1880年)时期, 人们就已发现植物向光性, 但是直到向光素PHOT1和PHOT2先后被鉴定(Liscum and Briggs, 1995; Christie et al., 1998), 植物的向光性机理研究才取得突飞猛进的发展.研究发现, PHOT1调节弱蓝光(0.01-1 μmol·m-2·s-1)和强蓝光(>1 μmol·m-2·s-1)诱导的下胚轴的向光性; PHOT2仅在强蓝光下起作用, 且与PHOT1功能冗余(Sakai et al., 2001; Briggs and Christie, 2002).由于强蓝光下PHOT1和PHOT2的功能冗余, 目前向光性机制研究主要集中在PHOT1介导的弱蓝光反应(Motchoulski and Liscum, 1999; Haga et al., 2015), 仅有少量研究通过突变体背景下的抑制子筛选寻找参与强蓝光信号途径的作用因子(赵翔等, 2014).单侧蓝光照射下, 自磷酸化激活的向光素蛋白将信号传递给NPH3、RPT2以及PKS等下游因子(Motchoulski and Liscum, 1999; Inada et al., 2004), 调节生长素运输载体的活性与定位(Ding et al., 2011).例如, 引起内皮层细胞背光侧PIN3 (PIN-FORMED3)积累(刘广超和丁兆军, 2018), 促进背光侧生长素含量增加、细胞伸长速率加快, 最终引起植物弯向光生长(Christie and Murphy, 2013).下胚轴弯曲过程中生长素的不对称分布受各类生长素运输载体的共同调控, 如PINs (PIN3、PIN4和PIN7)和ABCB19 (Christie et al., 2011).蓝光下, PIN3的磷酸化受抑制, 促进PIN3的背光侧定位, 诱导背光侧生长素积累以及下胚轴的向光性生长(Ding et al., 2011).PIN3的磷酸化以及极性定位受AGC激酶VIII亚族成员PID (PINOID)以及D6PK (D6 PROTEIN KINASE)的调节(Ding et al., 2011; Willige et al., 2013).ABCB19是生长素外流载体, 被蓝光激活的PHOT1磷酸化后, ABCB19的生长素转运活性受抑制, 增加拟南芥下胚轴顶端生长素含量并抑制其垂直生长, 同时通过PIN3蛋白促进生长素在下胚轴中侧向移动(Christie et al., 2011).背光侧生长素的大量积累促进了背光侧IAA19的降解, 释放出有活性的ARF7 (Sun et al., 2013), 不对称激活与生长相关基因的表达促进了下胚轴的向光性反应(刘广超和丁兆军, 2018). ...
Acquiring control: the evolution of stomatal signaling pathways 2 2019
... 拟南芥气孔由1对保卫细胞构成, 气孔通过调节其开闭程度控制着与环境的气体交换来响应外界的刺激信号(Hetherington and Woodward, 2003; Roelfsema and Hedrich, 2005).红光和蓝光都能诱导气孔快速张开(Shimazaki et al., 2007).红光通过影响叶肉细胞和保卫细胞中叶绿体的光合作用调节气孔开放(Mott et al., 2008; Suetsugu et al., 2014).蓝光作为信号通过激活保卫细胞质膜H+-ATPase诱导气孔开放(Assmann et al., 1985; Kinoshita et al., 2001).蓝光诱导的气孔开放受蓝光受体向光素(PHOT1和PHOT2)调节(Kinoshita et al., 2001).目前人们对向光素介导的气孔开放机制研究较为清楚(Inoue et al., 2008b; Inoue and Kinoshita, 2017).蓝光照射下, 自磷酸化激活的PHOT1直接磷酸化蛋白激酶BLUS1 (LIGHT SIGNALING1), 磷酸化的BLUS1间接将信号传递给I型蛋白磷酸酶(PPI)以及该酶的亚基PRSL1 (Takemiya and Shimazaki, 2016).PPI进一步将信号传递给质膜AHA1型H+-ATPase, 促进H+-ATPase的激活.H+-ATPase的激活需要其C末端倒数第2个苏氨酸的磷酸化以及与14-3-3蛋白的结合(Yamauchi et al., 2016).活化的H+-ATPase通过驱动保卫细胞内H+的跨膜输出诱导保卫细胞质膜的超极化(Shimazaki et al., 2007; Marten et al., 2010).超极化的质膜激活内向K+通道, 促进K+在保卫细胞胞质中积累.为了维持胞质的电荷平衡, Cl-和malate2-随之大量增加, 增加的K+、Cl-和malate2-进一步被转入液泡, 使保卫细胞水势降低, 外界水分大量进入保卫细胞, 促进保卫细胞体积膨胀, 气孔张开(Inoue and Kinoshita, 2017).保卫细胞胞质中Cl-的增加主要是由于向光素抑制了S型阴离子外流蛋白的活性(Sussmilch et al., 2019).向光素对S型阴离子外流蛋白活性的抑制需要2个蛋白激酶CBC1/2 (CONVERGENCE OF BLUE LIGHT (BL) and CO21/2)参与(Hiyama et al., 2017; Sussmilch et al., 2019). ...
... ; Sussmilch et al., 2019). ...
Arabidopsis phot1 and phot2 phosphorylate BLUS1 kinase with different efficiencies in stomatal opening 1 2016
... 拟南芥气孔由1对保卫细胞构成, 气孔通过调节其开闭程度控制着与环境的气体交换来响应外界的刺激信号(Hetherington and Woodward, 2003; Roelfsema and Hedrich, 2005).红光和蓝光都能诱导气孔快速张开(Shimazaki et al., 2007).红光通过影响叶肉细胞和保卫细胞中叶绿体的光合作用调节气孔开放(Mott et al., 2008; Suetsugu et al., 2014).蓝光作为信号通过激活保卫细胞质膜H+-ATPase诱导气孔开放(Assmann et al., 1985; Kinoshita et al., 2001).蓝光诱导的气孔开放受蓝光受体向光素(PHOT1和PHOT2)调节(Kinoshita et al., 2001).目前人们对向光素介导的气孔开放机制研究较为清楚(Inoue et al., 2008b; Inoue and Kinoshita, 2017).蓝光照射下, 自磷酸化激活的PHOT1直接磷酸化蛋白激酶BLUS1 (LIGHT SIGNALING1), 磷酸化的BLUS1间接将信号传递给I型蛋白磷酸酶(PPI)以及该酶的亚基PRSL1 (Takemiya and Shimazaki, 2016).PPI进一步将信号传递给质膜AHA1型H+-ATPase, 促进H+-ATPase的激活.H+-ATPase的激活需要其C末端倒数第2个苏氨酸的磷酸化以及与14-3-3蛋白的结合(Yamauchi et al., 2016).活化的H+-ATPase通过驱动保卫细胞内H+的跨膜输出诱导保卫细胞质膜的超极化(Shimazaki et al., 2007; Marten et al., 2010).超极化的质膜激活内向K+通道, 促进K+在保卫细胞胞质中积累.为了维持胞质的电荷平衡, Cl-和malate2-随之大量增加, 增加的K+、Cl-和malate2-进一步被转入液泡, 使保卫细胞水势降低, 外界水分大量进入保卫细胞, 促进保卫细胞体积膨胀, 气孔张开(Inoue and Kinoshita, 2017).保卫细胞胞质中Cl-的增加主要是由于向光素抑制了S型阴离子外流蛋白的活性(Sussmilch et al., 2019).向光素对S型阴离子外流蛋白活性的抑制需要2个蛋白激酶CBC1/2 (CONVERGENCE OF BLUE LIGHT (BL) and CO21/2)参与(Hiyama et al., 2017; Sussmilch et al., 2019). ...
The Arabidopsis rcn1-1 mutation impairs dephosphorylation of Phot2, resulting in enhanced blue light responses 1 2010
... 拟南芥中ABCB19 (ATP-BINDING CAS-SETTE B 19)和光敏色素激酶底物(PHYTOCHROME KINASE SUBSTRATE 4, PKS4)参与调节生长素的转运, 是下胚轴向光弯曲的负调节因子(Christie et al., 2011; Demarsy et al., 2012).自磷酸化激活的PHOT1通过直接磷酸化ABCB19和PKS4, 抑制ABCB19和PKS4的生长素转运活性, 促进下胚轴向光弯曲反应(Christie et al., 2011; Demarsy et al., 2012).PHOT1通过磷酸化修饰调节ABCB19的活性, 但是PHOT2并不影响ABCB19的活性(Christie et al., 2011).蛋白磷酸酶2A (PP2A)的A1亚基对应的蛋白RCN1 (Root Curling in N-naphthylphthalamic acid 1)与PHOT2蛋白发生体外互作, 负调节PHOT2介导的向光弯曲, 但RCN1并不与PHOT1直接相互作用(Tseng and Briggs, 2010).NPH3 (NONPHOTOTROPIC HYPOCOTYL 3)和RPT2 (ROOT PHOTOTROPISM 2)是通过筛选拟南芥下胚轴以及根的向光性缺失突变体获得(Motchoulski and Liscum, 1999; Sakai et al., 2000).随后的研究发现, NPH3和RPT2是向光素途径的重要调控因子, 均可与PHOT1蛋白直接相互作用(Blakeslee et al., 2004; Stone et al., 2008).虽然NPH3和RPT2也参与PHOT2信号转导途径, 但目前并无RPT2与PHOT2蛋白之间直接相互作用的报道(Inada et al., 2004; Lariguet et al., 2006).上述结果暗示PHOT1和PHOT2可能通过与不同的下游互作因子差异调节蓝光反应. ...
Mapping of the phosphorylation sites on the phototropic signal transducer, NPH3 1 2008
... 近年来, 科学家研究发现, 拟南芥NRL家族蛋白在向光素介导的蓝光信号转导通路中起重要作用.拟南芥NRL家族6个分支中的3个分支(II、III和IV) (图1)蛋白目前已经被报道参与向光素调节的反应, 或者与向光素蛋白存在直接相互作用, 暗示这3个分支的其它成员可能参与向光素信号途径的调节.RPT2与NCH1都来自分支II, 既参与生长素依赖的向光性调节, 又参与非生长素依赖的叶绿体聚光运动调节(Suetsugu et al., 2016; Suetsugu and Wada, 2017).目前RPT2和NCH1如何区分向光素介导的2种信号途径并不清楚.考虑到RPT2和NCH1蛋白所在分支与其余2个分支(III和IV)的亲缘关系较远(Christie et al., 2018), 暗示该分支其它成员也可能同时参与生长素依赖和非依赖性的向光素反应.RPT2可与PHOT1的LOV1区相互作用并抑制PHOT1的自磷酸化, 以降低PHOT1的光敏感性.尽管RPT2也可以与PHOT2的LOV1区域相互作用, 但是RPT2不抑制PHOT2的活性(Kimura et al., 2020).可能存在NRL家族其它蛋白与PHOT2相互作用, 调节PHOT2介导的蓝光信号转导.NPH3主要调节生长素依赖的向光性反应以及叶片的伸展与定位(Harada et al., 2013; Zhao et al., 2018), 但NPH3激活后调节生长素不对称分布的机制并不清楚.NPH3的磷酸化状态与其生理活性以及在细胞中的定位密切相关(Haga et al., 2015; Zhao et al., 2018).PHOT1介导的NPH3去磷酸化在弱蓝光诱导的下胚轴向光弯曲反应中发挥重要作用, 强蓝光下NPH3并未发生去磷酸化反应(Tsuchida-Mayama et al., 2008).那么NPH3如何调节强蓝光诱导的下胚轴向光弯曲反应, 是否与其E3泛素化连接酶活性有关? NPY1与PID互作, 通过影响生长素信号调节植物器官的形成以及根的向重力性反应(Suetsugu et al., 2016).生长素不对称分布决定不对称生长, PID- NPY1与PHOT-NPH3作用方式又相似, 那么NPY分支成员是否参与调节向光素介导的植物向性运动以及叶片的伸展与定位? PID和PHOT都属于AGC激酶VIII亚族成员(Ding et al., 2011; Willige et al., 2013), NRL家族蛋白是否可以与AGC激酶VIII亚家族其它成员相互作用, 以调节生长素介导的其它生理反应? 来自分支IV的NRL2也可以与PHOT1发生互作(Sullivan et al., 2009), 暗示该分支的蛋白也可能在向光素介导的蓝光信号途径中发挥重要作用.综上, NRL家族蛋白功能的陆续鉴定, 有望解析向光素PHOT1和PHOT2感受蓝光信号差异, 进而调节不同生理反应的可能机制. ...
Role of the phytochrome and cryptochrome signaling pathways in hypocotyl phototropism 3 2010
... 光敏色素(phytochrome, phy)和隐花色素(cryptochrome, cry)通过影响RPT2的表达调节向光性反应(Tsuchida-Mayama et al., 2010; 赵翔等, 2015; Zhao et al., 2019).强蓝光处理3小时显著诱导拟南芥野生型中RPT2基因的表达, PHYA、CRY1或CRY2基因突变可部分抑制蓝光诱导的RPT2表达, 强蓝光处理几乎不能诱导phya/cry1/cry2三突变体中RPT2表达(Tsuchida-Mayama et al., 2010).蓝光处理延长至24小时后, phya/cry1/cry2三突变体中RPT2的基因表达被轻微诱导, 且下胚轴能发生轻微的弯曲, 而phya/phyb/cry1/cry2四突变体中几乎检测不到RPT2蛋白的表达, 其下胚轴缺失向光弯曲反应(Tsuchida-Mayama et al., 2010).上述结果表明, 蓝光诱导RPT2的表达需要PHYs或者CRYs参与, PHYs和CRYs通过调节RPT2的表达参与调节蓝光诱导的下胚轴向光弯曲反应. ...
... 表达(Tsuchida-Mayama et al., 2010).蓝光处理延长至24小时后, phya/cry1/cry2三突变体中RPT2的基因表达被轻微诱导, 且下胚轴能发生轻微的弯曲, 而phya/phyb/cry1/cry2四突变体中几乎检测不到RPT2蛋白的表达, 其下胚轴缺失向光弯曲反应(Tsuchida-Mayama et al., 2010).上述结果表明, 蓝光诱导RPT2的表达需要PHYs或者CRYs参与, PHYs和CRYs通过调节RPT2的表达参与调节蓝光诱导的下胚轴向光弯曲反应. ...
... 四突变体中几乎检测不到RPT2蛋白的表达, 其下胚轴缺失向光弯曲反应(Tsuchida-Mayama et al., 2010).上述结果表明, 蓝光诱导RPT2的表达需要PHYs或者CRYs参与, PHYs和CRYs通过调节RPT2的表达参与调节蓝光诱导的下胚轴向光弯曲反应. ...
Disruption of ROOT PHOTOTROPISM 2 gene does not affect phototropin-mediated stomatal opening 4 2013
... KAC (KINESIN-LIKE PROTEIN FOR ACTIN-BASED CHLOROPLAST MOVEMENT)参与调节叶绿体肌动蛋白纤维的聚集(Wada, 2013; Kong and Wada, 2014), WEB1 (WEAK CHLOROPLAST MOVEMENT UNDER BLUE LIGHT 1)和PMI1 (PLASTIDMOVEMENT IMPAIRED1)对叶绿体肌动蛋白纤维的稳定性至关重要(Suetsugu et al., 2015). ...
Chloroplast and nuclear photorelocation movements 1 2016
... 作为植物进行光合作用的细胞器, 叶绿体响应外界光照刺激而发生的位置变化称为叶绿体光下重定位.弱光照射下, 为最大程度地吸收光能, 叶绿体沿细胞平周壁方向聚集, 这种现象为聚光运动(accumulation movement).强光条件下, 为避免过强的光对自身造成伤害, 叶绿体沿细胞垂周壁聚集, 使之与光平行分布, 这种现象称为避光运动(avoidance movement) (Kasahara et al., 2004).叶绿体运动受蓝光调节, 蓝光受体向光素PHOT1和PHOT2共同调控叶绿体的聚光反应, 而PHOT2单独调节叶绿体的避光反应(Suetsugu and Wada, 2017).与向光素调节的其它多细胞协调性反应(如向光性、叶片的伸展与定位)不同, 叶绿体运动是细胞自主性反应, 不受基因表达(Kong and Wada, 2016; Wada, 2016)和生长素运输的调节(Christie et al., 2018), 最终由叶绿体肌动蛋白响应入射光的强度与方向而发生快速重组介导(Kadota et al., 2009; Kong and Wada, 2014).叶绿体肌动蛋白特异定位于细胞质膜和叶绿体之间, 在叶绿体前进方向的后面叶绿体肌动蛋白纤维迅速解聚, 前进方向的前面叶绿体肌动蛋白纤维迅速聚集, 这种不对称的叶绿体肌动蛋白纤维分布最终决定了叶绿体的运动方向和速度(Kong and Wada, 2016).CHUP1 (CHLOROPLAST UNUSUAL POSITIONING 1)和 ...
The subcellular localization and blue-light-induced movement of phototropin 1-GFP in etiolated seedlings of Arabidopsis thaliana 1 2008
... 蓝光照射会诱导PHOT1和PHOT2亚细胞定位改变, PHOT1从细胞质膜脱离后向胞质移动(Kaiserli et al., 2009), 而PHOT2从细胞质膜脱离后向高尔基体转移, 最终与高尔基体偶联(Aggarwal et al., 2014).虽然向光素蛋白亚细胞定位受光照影响的生物学意义还不清楚, 但已知其亚细胞定位变化受PHOT1和PHOT2的激酶活性调节, 且蓝光诱导PHOT1的胞内定位变化(intracelluar position)参与下胚轴向光性调节(Wan et al., 2008; Zhao et al., 2018), 暗示向光素的亚细胞定位变化影响了向光素途径的信号传递, PHOT1和PHOT2亚细胞定位变化的不同可能是2种向光素受体差异调节植物蓝光反应的重要原因. ...
The signal transducer NPH3 integrates the phototropin1 photosensor with PIN2-based polar auxin transport in Arabidopsis root phototropism 3 2012
... NPH3主要通过2种途径影响生长素的不对称分布.一是NPH3通过多泛素化修饰PHOT1促进PHOT1的降解, 间接影响生长素转运蛋白(如生长素外流载体PIN1 (Roberts et al., 2011)和PIN2 (Wan et al., 2012))的重定位.NPH3是以Cullin3为基础的E3泛素化连接酶CRL3NPH3复合体的组成成分, 在该复合体中作为底物连接蛋白发挥作用(Roberts et al., 2011; Wan et al., 2012).强蓝光下, CRL3NPH3复合体直接与PHOT1蛋白相互作用, 促进PHOT1的单/多泛素化修饰, 多泛素化修饰的PHOT1进入细胞质后被26S蛋白酶体识别后降解(Roberts et al., 2011).PHOT1从细胞膜进入细胞质影响了生长素外流载体PIN1 (Roberts et al., 2011)和PIN2 (Wan et al., 2012)的重定位, 但具体作用机制有待深入研究.二是NPH3通过与PKS家族蛋白互作调节生长素的转运(Fankhauser and Christie, 2015).PKS家族成员是向光弯曲反应的正向调节因子, 参与生长素转运调节, 它们可以与向光素和NPH3蛋白直接互作, 形成PHOT1-PKS-NPH3复合体(Kami et al., 2014), 但是复合体的作用机制目前还不清楚. ...
... ; Wan et al., 2012).强蓝光下, CRL3NPH3复合体直接与PHOT1蛋白相互作用, 促进PHOT1的单/多泛素化修饰, 多泛素化修饰的PHOT1进入细胞质后被26S蛋白酶体识别后降解(Roberts et al., 2011).PHOT1从细胞膜进入细胞质影响了生长素外流载体PIN1 (Roberts et al., 2011)和PIN2 (Wan et al., 2012)的重定位, 但具体作用机制有待深入研究.二是NPH3通过与PKS家族蛋白互作调节生长素的转运(Fankhauser and Christie, 2015).PKS家族成员是向光弯曲反应的正向调节因子, 参与生长素转运调节, 它们可以与向光素和NPH3蛋白直接互作, 形成PHOT1-PKS-NPH3复合体(Kami et al., 2014), 但是复合体的作用机制目前还不清楚. ...
... )和PIN2 (Wan et al., 2012)的重定位, 但具体作用机制有待深入研究.二是NPH3通过与PKS家族蛋白互作调节生长素的转运(Fankhauser and Christie, 2015).PKS家族成员是向光弯曲反应的正向调节因子, 参与生长素转运调节, 它们可以与向光素和NPH3蛋白直接互作, 形成PHOT1-PKS-NPH3复合体(Kami et al., 2014), 但是复合体的作用机制目前还不清楚. ...
D6PK AGCVIII kinases are required for auxin transport and phototropic hypocotyl bending in Arabidopsis 2 2013
... 早在达尔文(1880年)时期, 人们就已发现植物向光性, 但是直到向光素PHOT1和PHOT2先后被鉴定(Liscum and Briggs, 1995; Christie et al., 1998), 植物的向光性机理研究才取得突飞猛进的发展.研究发现, PHOT1调节弱蓝光(0.01-1 μmol·m-2·s-1)和强蓝光(>1 μmol·m-2·s-1)诱导的下胚轴的向光性; PHOT2仅在强蓝光下起作用, 且与PHOT1功能冗余(Sakai et al., 2001; Briggs and Christie, 2002).由于强蓝光下PHOT1和PHOT2的功能冗余, 目前向光性机制研究主要集中在PHOT1介导的弱蓝光反应(Motchoulski and Liscum, 1999; Haga et al., 2015), 仅有少量研究通过突变体背景下的抑制子筛选寻找参与强蓝光信号途径的作用因子(赵翔等, 2014).单侧蓝光照射下, 自磷酸化激活的向光素蛋白将信号传递给NPH3、RPT2以及PKS等下游因子(Motchoulski and Liscum, 1999; Inada et al., 2004), 调节生长素运输载体的活性与定位(Ding et al., 2011).例如, 引起内皮层细胞背光侧PIN3 (PIN-FORMED3)积累(刘广超和丁兆军, 2018), 促进背光侧生长素含量增加、细胞伸长速率加快, 最终引起植物弯向光生长(Christie and Murphy, 2013).下胚轴弯曲过程中生长素的不对称分布受各类生长素运输载体的共同调控, 如PINs (PIN3、PIN4和PIN7)和ABCB19 (Christie et al., 2011).蓝光下, PIN3的磷酸化受抑制, 促进PIN3的背光侧定位, 诱导背光侧生长素积累以及下胚轴的向光性生长(Ding et al., 2011).PIN3的磷酸化以及极性定位受AGC激酶VIII亚族成员PID (PINOID)以及D6PK (D6 PROTEIN KINASE)的调节(Ding et al., 2011; Willige et al., 2013).ABCB19是生长素外流载体, 被蓝光激活的PHOT1磷酸化后, ABCB19的生长素转运活性受抑制, 增加拟南芥下胚轴顶端生长素含量并抑制其垂直生长, 同时通过PIN3蛋白促进生长素在下胚轴中侧向移动(Christie et al., 2011).背光侧生长素的大量积累促进了背光侧IAA19的降解, 释放出有活性的ARF7 (Sun et al., 2013), 不对称激活与生长相关基因的表达促进了下胚轴的向光性反应(刘广超和丁兆军, 2018). ...
... 近年来, 科学家研究发现, 拟南芥NRL家族蛋白在向光素介导的蓝光信号转导通路中起重要作用.拟南芥NRL家族6个分支中的3个分支(II、III和IV) (图1)蛋白目前已经被报道参与向光素调节的反应, 或者与向光素蛋白存在直接相互作用, 暗示这3个分支的其它成员可能参与向光素信号途径的调节.RPT2与NCH1都来自分支II, 既参与生长素依赖的向光性调节, 又参与非生长素依赖的叶绿体聚光运动调节(Suetsugu et al., 2016; Suetsugu and Wada, 2017).目前RPT2和NCH1如何区分向光素介导的2种信号途径并不清楚.考虑到RPT2和NCH1蛋白所在分支与其余2个分支(III和IV)的亲缘关系较远(Christie et al., 2018), 暗示该分支其它成员也可能同时参与生长素依赖和非依赖性的向光素反应.RPT2可与PHOT1的LOV1区相互作用并抑制PHOT1的自磷酸化, 以降低PHOT1的光敏感性.尽管RPT2也可以与PHOT2的LOV1区域相互作用, 但是RPT2不抑制PHOT2的活性(Kimura et al., 2020).可能存在NRL家族其它蛋白与PHOT2相互作用, 调节PHOT2介导的蓝光信号转导.NPH3主要调节生长素依赖的向光性反应以及叶片的伸展与定位(Harada et al., 2013; Zhao et al., 2018), 但NPH3激活后调节生长素不对称分布的机制并不清楚.NPH3的磷酸化状态与其生理活性以及在细胞中的定位密切相关(Haga et al., 2015; Zhao et al., 2018).PHOT1介导的NPH3去磷酸化在弱蓝光诱导的下胚轴向光弯曲反应中发挥重要作用, 强蓝光下NPH3并未发生去磷酸化反应(Tsuchida-Mayama et al., 2008).那么NPH3如何调节强蓝光诱导的下胚轴向光弯曲反应, 是否与其E3泛素化连接酶活性有关? NPY1与PID互作, 通过影响生长素信号调节植物器官的形成以及根的向重力性反应(Suetsugu et al., 2016).生长素不对称分布决定不对称生长, PID- NPY1与PHOT-NPH3作用方式又相似, 那么NPY分支成员是否参与调节向光素介导的植物向性运动以及叶片的伸展与定位? PID和PHOT都属于AGC激酶VIII亚族成员(Ding et al., 2011; Willige et al., 2013), NRL家族蛋白是否可以与AGC激酶VIII亚家族其它成员相互作用, 以调节生长素介导的其它生理反应? 来自分支IV的NRL2也可以与PHOT1发生互作(Sullivan et al., 2009), 暗示该分支的蛋白也可能在向光素介导的蓝光信号途径中发挥重要作用.综上, NRL家族蛋白功能的陆续鉴定, 有望解析向光素PHOT1和PHOT2感受蓝光信号差异, 进而调节不同生理反应的可能机制. ...
The plasma membrane H+-ATPase AHA1 plays a major role in stomatal opening in response to blue light 1 2016
... 拟南芥气孔由1对保卫细胞构成, 气孔通过调节其开闭程度控制着与环境的气体交换来响应外界的刺激信号(Hetherington and Woodward, 2003; Roelfsema and Hedrich, 2005).红光和蓝光都能诱导气孔快速张开(Shimazaki et al., 2007).红光通过影响叶肉细胞和保卫细胞中叶绿体的光合作用调节气孔开放(Mott et al., 2008; Suetsugu et al., 2014).蓝光作为信号通过激活保卫细胞质膜H+-ATPase诱导气孔开放(Assmann et al., 1985; Kinoshita et al., 2001).蓝光诱导的气孔开放受蓝光受体向光素(PHOT1和PHOT2)调节(Kinoshita et al., 2001).目前人们对向光素介导的气孔开放机制研究较为清楚(Inoue et al., 2008b; Inoue and Kinoshita, 2017).蓝光照射下, 自磷酸化激活的PHOT1直接磷酸化蛋白激酶BLUS1 (LIGHT SIGNALING1), 磷酸化的BLUS1间接将信号传递给I型蛋白磷酸酶(PPI)以及该酶的亚基PRSL1 (Takemiya and Shimazaki, 2016).PPI进一步将信号传递给质膜AHA1型H+-ATPase, 促进H+-ATPase的激活.H+-ATPase的激活需要其C末端倒数第2个苏氨酸的磷酸化以及与14-3-3蛋白的结合(Yamauchi et al., 2016).活化的H+-ATPase通过驱动保卫细胞内H+的跨膜输出诱导保卫细胞质膜的超极化(Shimazaki et al., 2007; Marten et al., 2010).超极化的质膜激活内向K+通道, 促进K+在保卫细胞胞质中积累.为了维持胞质的电荷平衡, Cl-和malate2-随之大量增加, 增加的K+、Cl-和malate2-进一步被转入液泡, 使保卫细胞水势降低, 外界水分大量进入保卫细胞, 促进保卫细胞体积膨胀, 气孔张开(Inoue and Kinoshita, 2017).保卫细胞胞质中Cl-的增加主要是由于向光素抑制了S型阴离子外流蛋白的活性(Sussmilch et al., 2019).向光素对S型阴离子外流蛋白活性的抑制需要2个蛋白激酶CBC1/2 (CONVERGENCE OF BLUE LIGHT (BL) and CO21/2)参与(Hiyama et al., 2017; Sussmilch et al., 2019). ...
The pollen receptor kinase LePRK2 mediates growth-promoting signals and positively regulates pollen germination and tube growth 1 2008
... NPH3和RPT2是来自2个不同分支的NRL家族成员, 通过与向光素相互作用调节蓝光诱导下胚轴向光性、叶绿体聚光运动、气孔开放以及叶片伸展与定位(Christie et al., 2015, 2018).NRL8是AGC激酶(cAMP依赖的蛋白激酶A, cGMP依赖的蛋白激酶G和磷脂依赖的蛋白激酶C)活性依赖的花粉管生长调节因子(Lalanne et al., 2004; Zhang et al., 2008), 该蛋白与RPT2来自分支II.与NPH3类似, SR1IP1 (AtSR1 INTERACTING PROTEIN 1)也称NRL31, 可以与CRL3 ( CUL3 RING E3 UBIQUITIN LIGASE)形成复合体, 调节底物的泛素化(Zhang et al., 2014).SR1IP1通过调节拟南芥中Ca2+/钙调素结合的转录因子AtSR1 (SIGNAL RESPONSIVE 1)的泛素化降解, 增强植物对细菌性病原体的抗性(Zhang et al., 2014).但系统进化研究发现, SR1IP1也来自分支II, 且比NRL8与RPT2亲缘关系更近, 与NPH3的进化明显不同.Suetsugu等(2016)在筛选参与调节叶绿体运动的NRL家族蛋白时, 发现NRL31基因突变抑制了光诱导的叶绿体运动, 且蛋白NRL31可以与PHOT1发生体内互作, 因此将其重命名为NCH1 (NRL PROTEIN FORCHLOROPLASTMOVEMENT 1).DEFECTIVELY ORGANIZED TRIBUTARIES 3/NRL23是与NPH3来自同一个分支的NRL家族蛋白, 主要参与维管组织发育以及叶脉结构调节(Petricka et al., 2008).虽然NRL23与NPH3的亲缘关系较近, 但是该蛋白是否在向光素介导的信号途径中发挥作用尚未见报道.NRL5、NRL6、NRL7、NRL15、NRL20、NRL21、NRL26和NRL30是分支III中的NPY (NAKED PINS IN YUCCA)家族成员, 其中NRL6/NPY2、NRL7/NPY4、NRL20/NPY1、NRL21/NPY5和NRL30/NPY3以功能冗余的方式与AGCVIII激酶共同调节器官发育以及向重力性反应中生长素的运输(Cheng et al., 2008; Li et al., 2011).通过分析NPYs基因对应的双突变体、三突变体、四突变体以及五突变体根的向重力性表型, 发现双突变体和三突变体向重力性表型缺失不明显; 四突变体表现不同程度的向重力性缺失表型, 其中npy1/npy2/npy4/npy5和npy2/npy3/npy4/npy5严重缺失向重力性; 五突变体的根几乎完全缺失向重力性, 在21°到150°之间随机生长(Li et al., 2011). ...
Regulation of plant immunity through ubiquitin-mediated modulation of Ca 2+-calmodulin-AtSR1/CAMTA3 signaling 2 2014
... NPH3和RPT2是来自2个不同分支的NRL家族成员, 通过与向光素相互作用调节蓝光诱导下胚轴向光性、叶绿体聚光运动、气孔开放以及叶片伸展与定位(Christie et al., 2015, 2018).NRL8是AGC激酶(cAMP依赖的蛋白激酶A, cGMP依赖的蛋白激酶G和磷脂依赖的蛋白激酶C)活性依赖的花粉管生长调节因子(Lalanne et al., 2004; Zhang et al., 2008), 该蛋白与RPT2来自分支II.与NPH3类似, SR1IP1 (AtSR1 INTERACTING PROTEIN 1)也称NRL31, 可以与CRL3 ( CUL3 RING E3 UBIQUITIN LIGASE)形成复合体, 调节底物的泛素化(Zhang et al., 2014).SR1IP1通过调节拟南芥中Ca2+/钙调素结合的转录因子AtSR1 (SIGNAL RESPONSIVE 1)的泛素化降解, 增强植物对细菌性病原体的抗性(Zhang et al., 2014).但系统进化研究发现, SR1IP1也来自分支II, 且比NRL8与RPT2亲缘关系更近, 与NPH3的进化明显不同.Suetsugu等(2016)在筛选参与调节叶绿体运动的NRL家族蛋白时, 发现NRL31基因突变抑制了光诱导的叶绿体运动, 且蛋白NRL31可以与PHOT1发生体内互作, 因此将其重命名为NCH1 (NRL PROTEIN FORCHLOROPLASTMOVEMENT 1).DEFECTIVELY ORGANIZED TRIBUTARIES 3/NRL23是与NPH3来自同一个分支的NRL家族蛋白, 主要参与维管组织发育以及叶脉结构调节(Petricka et al., 2008).虽然NRL23与NPH3的亲缘关系较近, 但是该蛋白是否在向光素介导的信号途径中发挥作用尚未见报道.NRL5、NRL6、NRL7、NRL15、NRL20、NRL21、NRL26和NRL30是分支III中的NPY (NAKED PINS IN YUCCA)家族成员, 其中NRL6/NPY2、NRL7/NPY4、NRL20/NPY1、NRL21/NPY5和NRL30/NPY3以功能冗余的方式与AGCVIII激酶共同调节器官发育以及向重力性反应中生长素的运输(Cheng et al., 2008; Li et al., 2011).通过分析NPYs基因对应的双突变体、三突变体、四突变体以及五突变体根的向重力性表型, 发现双突变体和三突变体向重力性表型缺失不明显; 四突变体表现不同程度的向重力性缺失表型, 其中npy1/npy2/npy4/npy5和npy2/npy3/npy4/npy5严重缺失向重力性; 五突变体的根几乎完全缺失向重力性, 在21°到150°之间随机生长(Li et al., 2011). ...
... /钙调素结合的转录因子AtSR1 (SIGNAL RESPONSIVE 1)的泛素化降解, 增强植物对细菌性病原体的抗性(Zhang et al., 2014).但系统进化研究发现, SR1IP1也来自分支II, 且比NRL8与RPT2亲缘关系更近, 与NPH3的进化明显不同.Suetsugu等(2016)在筛选参与调节叶绿体运动的NRL家族蛋白时, 发现NRL31基因突变抑制了光诱导的叶绿体运动, 且蛋白NRL31可以与PHOT1发生体内互作, 因此将其重命名为NCH1 (NRL PROTEIN FORCHLOROPLASTMOVEMENT 1).DEFECTIVELY ORGANIZED TRIBUTARIES 3/NRL23是与NPH3来自同一个分支的NRL家族蛋白, 主要参与维管组织发育以及叶脉结构调节(Petricka et al., 2008).虽然NRL23与NPH3的亲缘关系较近, 但是该蛋白是否在向光素介导的信号途径中发挥作用尚未见报道.NRL5、NRL6、NRL7、NRL15、NRL20、NRL21、NRL26和NRL30是分支III中的NPY (NAKED PINS IN YUCCA)家族成员, 其中NRL6/NPY2、NRL7/NPY4、NRL20/NPY1、NRL21/NPY5和NRL30/NPY3以功能冗余的方式与AGCVIII激酶共同调节器官发育以及向重力性反应中生长素的运输(Cheng et al., 2008; Li et al., 2011).通过分析NPYs基因对应的双突变体、三突变体、四突变体以及五突变体根的向重力性表型, 发现双突变体和三突变体向重力性表型缺失不明显; 四突变体表现不同程度的向重力性缺失表型, 其中npy1/npy2/npy4/npy5和npy2/npy3/npy4/npy5严重缺失向重力性; 五突变体的根几乎完全缺失向重力性, 在21°到150°之间随机生长(Li et al., 2011). ...
Cryptochrome-mediated hypocotyl phototropism was regulated antagonistically by gibberellic acid and sucrose in Arabidopsis 1 2019
... 光敏色素(phytochrome, phy)和隐花色素(cryptochrome, cry)通过影响RPT2的表达调节向光性反应(Tsuchida-Mayama et al., 2010; 赵翔等, 2015; Zhao et al., 2019).强蓝光处理3小时显著诱导拟南芥野生型中RPT2基因的表达, PHYA、CRY1或CRY2基因突变可部分抑制蓝光诱导的RPT2表达, 强蓝光处理几乎不能诱导phya/cry1/cry2三突变体中RPT2表达(Tsuchida-Mayama et al., 2010).蓝光处理延长至24小时后, phya/cry1/cry2三突变体中RPT2的基因表达被轻微诱导, 且下胚轴能发生轻微的弯曲, 而phya/phyb/cry1/cry2四突变体中几乎检测不到RPT2蛋白的表达, 其下胚轴缺失向光弯曲反应(Tsuchida-Mayama et al., 2010).上述结果表明, 蓝光诱导RPT2的表达需要PHYs或者CRYs参与, PHYs和CRYs通过调节RPT2的表达参与调节蓝光诱导的下胚轴向光弯曲反应. ...
Phototropins function in high-intensity blue light-induced hypocotyl phototropism in Arabidopsis by altering cytosolic calcium 1 2013
... 拟南芥(Arabidopsis thaliana) PHOT1和PHOT2以功能冗余的方式调节下胚轴向光性和根负向光性、叶绿体聚光运动、气孔开放以及叶片伸展与定位等(Christie et al., 2015).PHOT1特异性抑制光下黄化幼苗下胚轴的快速伸长(Folta et al., 2003)、子叶变绿过程中叶绿素结合蛋白mRNA转录的稳定性(Folta and Kaufman, 2003)以及侧根的快速生长(Moni et al., 2015).PHOT2不仅介导强蓝光诱导的下胚轴向光弯曲(Sakai et al., 2001), 而且也特异调节叶绿体和细胞核的避光运动(Higa et al., 2014)以及抗性蛋白介导的病毒防御反应(Jeong et al., 2010).PHOT1和PHOT2的功能差异可能与磷酸化修饰位点、磷酸化程度、亚细胞定位变化以及下游相互作用蛋白种类不同有关(Zhao et al., 2013; Morrow et al., 2018; Liscum et al., 2020). ...
Phot2-regulated relocation of NPH3 mediates phototropic response to high-intensity blue light in Arabidopsis thaliana 7 2018
... 蓝光照射会诱导PHOT1和PHOT2亚细胞定位改变, PHOT1从细胞质膜脱离后向胞质移动(Kaiserli et al., 2009), 而PHOT2从细胞质膜脱离后向高尔基体转移, 最终与高尔基体偶联(Aggarwal et al., 2014).虽然向光素蛋白亚细胞定位受光照影响的生物学意义还不清楚, 但已知其亚细胞定位变化受PHOT1和PHOT2的激酶活性调节, 且蓝光诱导PHOT1的胞内定位变化(intracelluar position)参与下胚轴向光性调节(Wan et al., 2008; Zhao et al., 2018), 暗示向光素的亚细胞定位变化影响了向光素途径的信号传递, PHOT1和PHOT2亚细胞定位变化的不同可能是2种向光素受体差异调节植物蓝光反应的重要原因. ...
... 拟南芥nph3突变体在任何强度的单侧蓝光下均表现缺失下胚轴向光性(Liscum and Briggs, 1996; Liscum et al., 2014; Fankhauser and Christie, 2015), 表明NPH3调节PHOT1和PHOT2介导蓝光诱导的下胚轴弯曲反应.黑暗条件下, 磷酸化的NPH3蛋白与PHOT1结合, 定位于细胞膜, 单侧弱蓝光照射下, NPH3迅速发生PHOT1依赖的去磷酸化反应, 从细胞膜上脱离(Haga et al., 2015; Zhao et al., 2018).目前磷酸化NPH3的激酶还不清楚, 但NPH3去磷酸化主要由I型蛋白磷酸酶介导(Pedmale and Liscum, 2007).去磷酸化后NPH3从细胞膜向胞质的转移降低了其介导向光弯曲反应的能力(Haga et al., 2015; Sullivan et al., 2019).随着光照强度的增加, 下胚轴的弯曲度呈强度依赖的钟形曲线(Christie and Murphy, 2013).随着光照时间的增加, RPT2蛋白含量增加, 与去磷酸化的NPH3结合形成RPT2-NPH3复合物并与PHOT1蛋白结合, 促进NPH3的磷酸化以及从胞质到细胞膜的重定位, 开启第二次向光弯曲(Haga et al., 2015; Zhao et al., 2018).虽然PHOT1- NPH3复合体在细胞膜的重定位对持续光照介导的二次向光弯曲至关重要(Haga et al., 2015; Christie et al., 2018; Zhao et al., 2018), 但其生物学意义还不清楚, 人们认为这种变化有可能对调节生长素的不对称分布有重要影响(Pedmale and Liscum, 2007; Haga et al., 2015). ...
... ; Zhao et al., 2018).虽然PHOT1- NPH3复合体在细胞膜的重定位对持续光照介导的二次向光弯曲至关重要(Haga et al., 2015; Christie et al., 2018; Zhao et al., 2018), 但其生物学意义还不清楚, 人们认为这种变化有可能对调节生长素的不对称分布有重要影响(Pedmale and Liscum, 2007; Haga et al., 2015). ...
... ; Zhao et al., 2018), 但其生物学意义还不清楚, 人们认为这种变化有可能对调节生长素的不对称分布有重要影响(Pedmale and Liscum, 2007; Haga et al., 2015). ...
... 拟南芥rpt2单突变体在极弱光下(≤0.01 μmol·m-2·s-1)下胚轴向光弯曲反应正常, 随着光照强度的增大弯曲度逐渐降低(Inada et al., 2004), 表明RPT2仅参与调节强蓝光诱导的下胚轴向光弯曲反应.phot1/rpt2双突变体与phot1单突变体表型类似, 在10和100 μmol·m-2·s-1的较强蓝光下可以发生下胚轴向光弯曲反应, 但在0.01-1 μmol·m-2·s-1的弱蓝光下则缺失向光弯曲(Sakai et al., 2000), 暗示RPT2作为PHOT1的下游因子调节强蓝光诱导的下胚轴向光弯曲反应.phot2/rpt2双突变体与phot1/phot2双突变体的表型一致, 缺失强蓝光诱导的下胚轴向光弯曲反应(Inada et al., 2004; Zhao et al., 2018), 且PHOT1和RPT2可以直接相互作用.上述结果进一步证明, RPT2确实参与调节PHOT1介导的下胚轴向光弯曲反应.如果RPT2仅作为PHOT1的下游因子发挥作用, 那么rpt2单突变体应该与phot1单突变体表型一致.表型分析结果显示, rpt2单突变体与phot1/phot2双突变体的表型一致, 即缺失强蓝光诱导的下胚轴向光弯曲反应(Inada et al., 2004), 证明RPT2在PHOT2介导的下胚轴向光弯曲反应中也发挥重要作用.Harada等(2013)认为, RPT2在PHOT1抑制PHOT2途径的过程中发挥作用, 阻碍PHOT1对PHOT2途径的抑制.最近研究表明, RPT2可与PHOT1的LOV1区相互作用并抑制PHOT1的自磷酸化, 以降低PHOT1的光敏感性, 从而行使分子变阻器(molecular rheostat)的功能, 将不同强度蓝光下PHOT1的活性维持在适当的水平.尽管RPT2也可以与PHOT2的LOV1区域相互作用, 但是RPT2不抑制PHOT2的活性(Kimura et al., 2020). ...
... 近年来, 科学家研究发现, 拟南芥NRL家族蛋白在向光素介导的蓝光信号转导通路中起重要作用.拟南芥NRL家族6个分支中的3个分支(II、III和IV) (图1)蛋白目前已经被报道参与向光素调节的反应, 或者与向光素蛋白存在直接相互作用, 暗示这3个分支的其它成员可能参与向光素信号途径的调节.RPT2与NCH1都来自分支II, 既参与生长素依赖的向光性调节, 又参与非生长素依赖的叶绿体聚光运动调节(Suetsugu et al., 2016; Suetsugu and Wada, 2017).目前RPT2和NCH1如何区分向光素介导的2种信号途径并不清楚.考虑到RPT2和NCH1蛋白所在分支与其余2个分支(III和IV)的亲缘关系较远(Christie et al., 2018), 暗示该分支其它成员也可能同时参与生长素依赖和非依赖性的向光素反应.RPT2可与PHOT1的LOV1区相互作用并抑制PHOT1的自磷酸化, 以降低PHOT1的光敏感性.尽管RPT2也可以与PHOT2的LOV1区域相互作用, 但是RPT2不抑制PHOT2的活性(Kimura et al., 2020).可能存在NRL家族其它蛋白与PHOT2相互作用, 调节PHOT2介导的蓝光信号转导.NPH3主要调节生长素依赖的向光性反应以及叶片的伸展与定位(Harada et al., 2013; Zhao et al., 2018), 但NPH3激活后调节生长素不对称分布的机制并不清楚.NPH3的磷酸化状态与其生理活性以及在细胞中的定位密切相关(Haga et al., 2015; Zhao et al., 2018).PHOT1介导的NPH3去磷酸化在弱蓝光诱导的下胚轴向光弯曲反应中发挥重要作用, 强蓝光下NPH3并未发生去磷酸化反应(Tsuchida-Mayama et al., 2008).那么NPH3如何调节强蓝光诱导的下胚轴向光弯曲反应, 是否与其E3泛素化连接酶活性有关? NPY1与PID互作, 通过影响生长素信号调节植物器官的形成以及根的向重力性反应(Suetsugu et al., 2016).生长素不对称分布决定不对称生长, PID- NPY1与PHOT-NPH3作用方式又相似, 那么NPY分支成员是否参与调节向光素介导的植物向性运动以及叶片的伸展与定位? PID和PHOT都属于AGC激酶VIII亚族成员(Ding et al., 2011; Willige et al., 2013), NRL家族蛋白是否可以与AGC激酶VIII亚家族其它成员相互作用, 以调节生长素介导的其它生理反应? 来自分支IV的NRL2也可以与PHOT1发生互作(Sullivan et al., 2009), 暗示该分支的蛋白也可能在向光素介导的蓝光信号途径中发挥重要作用.综上, NRL家族蛋白功能的陆续鉴定, 有望解析向光素PHOT1和PHOT2感受蓝光信号差异, 进而调节不同生理反应的可能机制. ...
... ; Zhao et al., 2018).PHOT1介导的NPH3去磷酸化在弱蓝光诱导的下胚轴向光弯曲反应中发挥重要作用, 强蓝光下NPH3并未发生去磷酸化反应(Tsuchida-Mayama et al., 2008).那么NPH3如何调节强蓝光诱导的下胚轴向光弯曲反应, 是否与其E3泛素化连接酶活性有关? NPY1与PID互作, 通过影响生长素信号调节植物器官的形成以及根的向重力性反应(Suetsugu et al., 2016).生长素不对称分布决定不对称生长, PID- NPY1与PHOT-NPH3作用方式又相似, 那么NPY分支成员是否参与调节向光素介导的植物向性运动以及叶片的伸展与定位? PID和PHOT都属于AGC激酶VIII亚族成员(Ding et al., 2011; Willige et al., 2013), NRL家族蛋白是否可以与AGC激酶VIII亚家族其它成员相互作用, 以调节生长素介导的其它生理反应? 来自分支IV的NRL2也可以与PHOT1发生互作(Sullivan et al., 2009), 暗示该分支的蛋白也可能在向光素介导的蓝光信号途径中发挥重要作用.综上, NRL家族蛋白功能的陆续鉴定, 有望解析向光素PHOT1和PHOT2感受蓝光信号差异, 进而调节不同生理反应的可能机制. ...
 ,*河南大学生命科学学院/棉花生物学国家重点实验室/植物逆境生物学重点实验室, 开封 475004
,*河南大学生命科学学院/棉花生物学国家重点实验室/植物逆境生物学重点实验室, 开封 475004
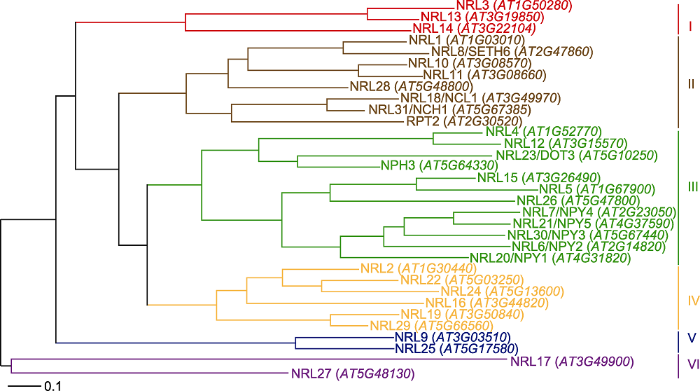 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}