Morphology Characters and Evolution of Ferns Scale Ι: Pteridaceae
Yufeng Gu1,2,3, Dongmei Jin1, Baodong Liu2, Xiling Dai3, Yuehong Yan,1,*1Shanghai Chenshan Plant Science Research Center, Shanghai Chenshan Botanical Garden, Chinese Academy of Sciences, Shanghai 201602, China 2Key Laboratory of Plant Biology, College of Heilongjiang Province, Harbin Normal University, Harbin 150025, China 3College of Life Sciences, Shanghai Normal University, Shanghai 200234, China
Abstract Scales are epidermal appendages on the rhizomes and leaves of many ferns. Features of scales play an important role in classification of ferns. The phylogenetic position and delimitation of Pteridaceae were treated differently by different authors. Here we collected scales of 76 fern species in Pteridaceae, and observed them under a dissecting microscope. By comparing the morphological characters, we found that scales are different among genus and subfamilies. We reconstructed a phylogenetic tree with the plastid rbcL sequence of the species in this study downloaded from GenBank database and reconstruced the ancestral state for two selected characters (margin of the scale and mesh type). The results suggested that homogeneous scale and entire margin were plesiomorphic characters, while non-entire margin and transparent mesh were evolved late in evolutionary process. We also speculated that the formation of transparent or non-transparent mesh may relate to the light intensity in the habitats. Keywords:morphology and anatomy;classification;scales;phylogeny;Pteridaceae
PDF (5022KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 顾钰峰, 金冬梅, 刘保东, 戴锡玲, 严岳鸿. 蕨类植物的鳞片特征及演化I: 凤尾蕨科. 植物学报, 2020, 55(2): 163-176 doi:10.11983/CBB19118 Gu Yufeng, Jin Dongmei, Liu Baodong, Dai Xiling, Yan Yuehong. Morphology Characters and Evolution of Ferns Scale Ι: Pteridaceae. Chinese Bulletin of Botany, 2020, 55(2): 163-176 doi:10.11983/CBB19118
(A) Coniogramme caudiformis; (B) C. robusta; (C) C. emeiensis; (D) C. japonica; (E) C. jinggangshanensis; (F) C. intermedia; (G) C. wilsonii; (H) C. procera; (I) Cryptogramma brunoniana. Bars=0.5 mm
(A) Pteris vittata; (B) Pityrogramme calomelanos; (C) Anogramma microphylla; (D) Taenitis blechnoides; (E) Onychium siliculosum; (F) O. japonicum var. lucidum; (G) O. japonicum; (H) P. wallichiana; (I) P. cretica var. nervosa; (J) P. deltodon; (K) P. henryi; (L) P. gallinopes; (M) P. dactylina; (N) P. dispar; (O) P. amoena; (P) P. decrescens; (Q) P. cadieri; (R) P. insignis; (S) P. excelsa; (T) P. aspericaulis var. tricolor; (U) P. setuloso-costulata; (V) P. aspericaulis; (W) P. plumbea; (X) P. stenophylla. Bars=0.5 mm
翠蕨(Anogramma microphylla)的鳞片(图4C)长约1.0 mm, 呈红棕色; 基部为3-4列细胞, 上半部为单列细胞。
用MrBayes 3.2.7软件基于贝叶斯信息准则(BIC)在jModelTest最优模型进行BI分析。每条链跑样1 000 000代, 每1 000代进行抽样。各分支上数字代表支持率。A: 珠蕨亚科; B: 水蕨亚科; C: 凤尾蕨亚科; D: 碎米蕨亚科; E: 书带蕨亚科 Figure 7Phylogenetic tree of Pteridaceae obtained from the maximum likelihood analysis of the plastid rbcL sequence
The BI analysis was performed in MrBayes 3.2.7 based on the best model Bayesian information criterion (BIC) implemented in jModelTest. Each chains was run for 1 000 000 generations, and was sampled every 1 000 generations. Numbers on branches indicate support values. A: Cryptogrammoideae; B: Ceratopteridoideae; C: Pteridoideae; D: Cheilanthoideae; E: Vittarioideae
系统树中只采用凤尾蕨科的物种。饼状图显示每个节点缺失的百分比和每个节点的状态在所有树中似然的平均值。(L) 筛孔类型; (R) 鳞片边缘。A: 珠蕨亚科; B: 水蕨亚科; C: 凤尾蕨亚科; D: 碎米蕨亚科; E: 书带蕨亚科 Figure 8Ancestral state reconstruction for margin of scales and mesh type characters optimized onto the best tree obtained in the plateau phase of the Bayesian with the maximum likelihood (ML) analysis of plastid gene rbcL sequence
Only species of Pteridaceae were used for the phylogeny. Pie charts show the percentage of node absence and the average likelihood received by each state across all the trees possessing that node. (L) Mesh type; (R) Margin of scales. A: Cryptogrammoideae; B: Ceratopteridoideae; C: Pteridoideae; D: Cheilanthoideae; E: Vittarioideae
ChristenhuszMJM, ChaseMW ( 2014). Trends and concepts in fern classification Ann Bot 113, 571-594. [本文引用: 2]
ChristenhuszMJM, ZhangXC, SchneiderH ( 2011). A linear sequence of extant families and genera of lycophytes and ferns Phytotaxa 19, 7-54. [本文引用: 1]
GreenfieldSS ( 1938). A comparison of the basal scales and Indusia of Dryopteris Goldiana × marginalis with those of its parent species Am Fern J 28, 55-62. [本文引用: 1]
HolttumER ( 1957). The scales of Cyatheaceae (With special reference to the genus Schizocaena J.Sm.) Kew Bull 12, 41-45. [本文引用: 3]
HolttumRE ( 1949). The classification of ferns Biol Rev 24, 267-296. [本文引用: 3]
HoshizakiBJ ( 1970). The rhizome scales of Platycerium Am Fern J 60, 144-160. [本文引用: 4]
KuoLY, LiFW, ChiouWL, WangCN ( 2011). First insights into fern matK phylogeny Mol Phylogenet Evol 59, 556-566. [本文引用: 2]
LabiakPH ( 2003). A new combination in the fern genus Ceradenia(Grammitidaceae) Kew Bull 58, 991-994. [本文引用: 1]
LehtonenS ( 2011). Towards resolving the complete fern tree of life PLoS One 6, e24851. [本文引用: 1]
MaddisonDR, MaddisonWP ( 2018). Mesquite: a modular system for evolutionary analysis Version 3.61.http://www. mesquiteproject.org . URL [本文引用: 1]
NayarBK ( 1970). A phylogenetic classification of the homosporous ferns Taxon 19, 229-236. [本文引用: 1]
PPGI ( 2016). A community-derived classification for extant lycophytes and ferns J Syst Evol 54, 563-603. [本文引用: 5]
PryerKM, SchuettpelzE, WolfPG, SchneiderH, SmithAR, CranfillR ( 2004). Phylogeny and evolution of ferns (monilophytes) with a focus on the early leptosporangiate divergences Am J Bot 91, 1582-1598. [本文引用: 1]
QiXP, KuoLY, GuoCC, LiH, LiZY, QiJ, WangLB, HuY, XiangJY, ZhangCF, GuoJ, HuangCH, MaH ( 2018). A well-resolved fern nuclear phylogeny reveals the evolution history of numerous transcription factor families Mol Phylogenet Evol 127, 961-977. [本文引用: 2]
RaiHS, GrahamSW ( 2010). Utility of a large, multigene plastid data set in inferring higher-order relationships in ferns and relatives (monilophytes) Am J Bot 97, 1444-1456. [本文引用: 1]
SchneiderH ( 2013). Evolutionary morphology of ferns (monilophytes). In: Ambrose BA, Purugganan M, eds Annual Plant Reviews, Vol. 45. The Evolution of Plant Form. Hoboken, New Jersey: John Wiley & Sons, Ltd. pp. 115-140. [本文引用: 1]
SchuettpelzE, SchneiderH, HuietL, WindhamMD, PryerKM ( 2007). A molecular phylogeny of the fern family Pteridaceae: assessing overall relationships and the affinities of previously unsampled genera Mol Phylogenet Evol 44, 1172-1185. [本文引用: 2]
SermolliREGP ( 1977). Tentamen Pteridophytorum genera in taxonomicum ordinem redigendi Webbia 31, 313-512. [本文引用: 1]
SmithAR, PryerKM, SchuettpelzE, KorallP, SchneiderH, WolfPG ( 2006). A classification for extant ferns Taxon 55, 705-731. [本文引用: 2]
TryonRM, TryonAF ( 1982). Ferns and Allied Plants New York: Springer- Verlag. pp. 166-212. [本文引用: 1]
TsutsumiC, KatoM ( 2008). Morphology and evolution of epiphytic Davalliaceae scales Botany 86, 1393-1403. [本文引用: 1]
TsutsumiC, ZhangXC, KatoM ( 2008). Molecular phylogeny of Davalliaceae and implications for generic classification Syst Bot 33, 44-48. [本文引用: 2]
Van den HeedeCJ, VianeRLL, ChaseMW ( 2003). Phylogenetic analysis of Asplenium subgenus Ceterach (Pteridophyta: Aspleniaceae) based on plastid and nuclear ribosomal ITS DNA sequences Am J Bot 90, 481-495. [本文引用: 1]
Watkins JrJE, KawaharaAY, LeichtTA, AuldJR, BickslerAJ, KaiserK ( 2006). Fern laminar scales protect against photoinhibition from excess light Am Fern J 96, 83-92. [本文引用: 1]
WeiR, SchneiderH, ZhangXC ( 2013). Toward a new circumscription of the twinsorus-fern genus (Diplazium Athyriaceae): a molecular phylogeny with morphological implications and infrageneric taxonomy Taxon 62, 441-457. [本文引用: 2]
WolfPG, RobisonTA, JohnsonMG, SundueMA, TestoWL, RothfelsCJ ( 2018). Target sequence capture of nuclear-encoded genes for phylogenetic analysis in ferns Appl Plant Sci 6, e01148. [本文引用: 2]
... 鳞片是大部分蕨类植物非常显著的特征, 是识别蕨类植物及对其进行分类的重要依据.在早期的蕨类植物分类研究中(Holttum, 1949, 1957, 1963; Tryon and Tryon, 1982; 于顺利和林尤兴, 1996; 石雷和张宪春, 1999, 2001; Van den Heede et al., 2003; Labiak, 2003; Tsutsumi et al., 2008; Wei et al., 2013; 顾钰峰等, 2014), 鳞片均被作为重要的分类佐证.然而在分子生物学快速发展的今天, 形态学研究渐被忽视, 蕨类植物的鳞片形态研究少有****涉及.Hoshizaki (1970)对鹿角蕨属(Platycerium)的鳞片形态进行了研究.柯勇男(2002)研究了台湾产广义鳞毛蕨科的鳞片形态.Tsutsumi等(2008)对骨碎补科植物的鳞片形态和进化进行了研究.顾钰峰(2015)对蕨类植物的毛状物和鳞片进行了广泛取材观察.他们的研究均认为鳞片对于蕨类植物的分类学研究有非常重要的意义.然而生物性状的表现往往受环境影响, 决定其物种的关键还是内部的基因结构.因此形态特征仅作为参考, 很难起决定性作用. ...
石松类和蕨类植物研究进展: 兼论国产类群的科级分类系统 1 2008
... 凤尾蕨科(Pteridaceae)植物形态上差异较大, 导致其分类一直存在很大争议(Holttum, 1949; Nayar, 1970; Sermolli, 1977).随着分子生物学的发展, 采用DNA序列构建系统发育树成为研究分类系统的重要手段, 不同****对于凤尾蕨科在蕨类植物分类系统中的位置各持不同观点, 凤尾蕨科所包含的类群也有很大差别(Smith et al., 2006; Schuettpelz et al., 2007; 刘红梅等, 2008; Christenhusz et al., 2011; Lehtonen, 2011; Kuo et al., 2011; 张宪春等, 2013; Christenhusz and Chase, 2014; PPG I, 2016; Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018).Pryer等(2004)认为, 凤尾蕨科与真水龙骨类构成姐妹群, 然后共同与碗蕨科形成姐妹群.Rai和Graham (2010)的研究也得出类似的结果.但是Smith等(2006)和Schneider (2013)的研究结果却相反, 他们认为碗蕨科和真水龙骨类形成姐妹群, 两者共同与凤尾蕨科植物形成姐妹群.在后续研究中, 也有****(Kuo et al., 2011; Christenhusz and Chase, 2014)得出相同的结果.然而, 在2018年发表的系统发育树中, ****们一致认为碗蕨科和真水龙骨类是姐妹群, 两者共同与凤尾蕨科形成姐妹群(Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018). ...
中国蕨类植物科属的系统排列和历史来源 5 1978
... 珠蕨亚科包含凤了蕨属、珠蕨属和Llavea三个属(PPG I, 2016), 中国仅有凤了蕨属和珠蕨属分布(Zhang et al., 2013).凤了蕨属原属于裸子蕨科(Hemionitidaceae), 而珠蕨属曾属于中国蕨科(Sinopteridaceae) (秦仁昌, 1978). ...
... 凤尾蕨科(Pteridaceae)植物形态上差异较大, 导致其分类一直存在很大争议(Holttum, 1949; Nayar, 1970; Sermolli, 1977).随着分子生物学的发展, 采用DNA序列构建系统发育树成为研究分类系统的重要手段, 不同****对于凤尾蕨科在蕨类植物分类系统中的位置各持不同观点, 凤尾蕨科所包含的类群也有很大差别(Smith et al., 2006; Schuettpelz et al., 2007; 刘红梅等, 2008; Christenhusz et al., 2011; Lehtonen, 2011; Kuo et al., 2011; 张宪春等, 2013; Christenhusz and Chase, 2014; PPG I, 2016; Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018).Pryer等(2004)认为, 凤尾蕨科与真水龙骨类构成姐妹群, 然后共同与碗蕨科形成姐妹群.Rai和Graham (2010)的研究也得出类似的结果.但是Smith等(2006)和Schneider (2013)的研究结果却相反, 他们认为碗蕨科和真水龙骨类形成姐妹群, 两者共同与凤尾蕨科植物形成姐妹群.在后续研究中, 也有****(Kuo et al., 2011; Christenhusz and Chase, 2014)得出相同的结果.然而, 在2018年发表的系统发育树中, ****们一致认为碗蕨科和真水龙骨类是姐妹群, 两者共同与凤尾蕨科形成姐妹群(Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018). ...
... 凤尾蕨科(Pteridaceae)植物形态上差异较大, 导致其分类一直存在很大争议(Holttum, 1949; Nayar, 1970; Sermolli, 1977).随着分子生物学的发展, 采用DNA序列构建系统发育树成为研究分类系统的重要手段, 不同****对于凤尾蕨科在蕨类植物分类系统中的位置各持不同观点, 凤尾蕨科所包含的类群也有很大差别(Smith et al., 2006; Schuettpelz et al., 2007; 刘红梅等, 2008; Christenhusz et al., 2011; Lehtonen, 2011; Kuo et al., 2011; 张宪春等, 2013; Christenhusz and Chase, 2014; PPG I, 2016; Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018).Pryer等(2004)认为, 凤尾蕨科与真水龙骨类构成姐妹群, 然后共同与碗蕨科形成姐妹群.Rai和Graham (2010)的研究也得出类似的结果.但是Smith等(2006)和Schneider (2013)的研究结果却相反, 他们认为碗蕨科和真水龙骨类形成姐妹群, 两者共同与凤尾蕨科植物形成姐妹群.在后续研究中, 也有****(Kuo et al., 2011; Christenhusz and Chase, 2014)得出相同的结果.然而, 在2018年发表的系统发育树中, ****们一致认为碗蕨科和真水龙骨类是姐妹群, 两者共同与凤尾蕨科形成姐妹群(Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018). ...
... ; Christenhusz and Chase, 2014)得出相同的结果.然而, 在2018年发表的系统发育树中, ****们一致认为碗蕨科和真水龙骨类是姐妹群, 两者共同与凤尾蕨科形成姐妹群(Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018). ...
A linear sequence of extant families and genera of lycophytes and ferns 1 2011
... 凤尾蕨科(Pteridaceae)植物形态上差异较大, 导致其分类一直存在很大争议(Holttum, 1949; Nayar, 1970; Sermolli, 1977).随着分子生物学的发展, 采用DNA序列构建系统发育树成为研究分类系统的重要手段, 不同****对于凤尾蕨科在蕨类植物分类系统中的位置各持不同观点, 凤尾蕨科所包含的类群也有很大差别(Smith et al., 2006; Schuettpelz et al., 2007; 刘红梅等, 2008; Christenhusz et al., 2011; Lehtonen, 2011; Kuo et al., 2011; 张宪春等, 2013; Christenhusz and Chase, 2014; PPG I, 2016; Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018).Pryer等(2004)认为, 凤尾蕨科与真水龙骨类构成姐妹群, 然后共同与碗蕨科形成姐妹群.Rai和Graham (2010)的研究也得出类似的结果.但是Smith等(2006)和Schneider (2013)的研究结果却相反, 他们认为碗蕨科和真水龙骨类形成姐妹群, 两者共同与凤尾蕨科植物形成姐妹群.在后续研究中, 也有****(Kuo et al., 2011; Christenhusz and Chase, 2014)得出相同的结果.然而, 在2018年发表的系统发育树中, ****们一致认为碗蕨科和真水龙骨类是姐妹群, 两者共同与凤尾蕨科形成姐妹群(Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018). ...
A comparison of the basal scales and Indusia of Dryopteris Goldiana × marginalis with those of its parent species 1 1938
... 凤尾蕨科(Pteridaceae)植物形态上差异较大, 导致其分类一直存在很大争议(Holttum, 1949; Nayar, 1970; Sermolli, 1977).随着分子生物学的发展, 采用DNA序列构建系统发育树成为研究分类系统的重要手段, 不同****对于凤尾蕨科在蕨类植物分类系统中的位置各持不同观点, 凤尾蕨科所包含的类群也有很大差别(Smith et al., 2006; Schuettpelz et al., 2007; 刘红梅等, 2008; Christenhusz et al., 2011; Lehtonen, 2011; Kuo et al., 2011; 张宪春等, 2013; Christenhusz and Chase, 2014; PPG I, 2016; Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018).Pryer等(2004)认为, 凤尾蕨科与真水龙骨类构成姐妹群, 然后共同与碗蕨科形成姐妹群.Rai和Graham (2010)的研究也得出类似的结果.但是Smith等(2006)和Schneider (2013)的研究结果却相反, 他们认为碗蕨科和真水龙骨类形成姐妹群, 两者共同与凤尾蕨科植物形成姐妹群.在后续研究中, 也有****(Kuo et al., 2011; Christenhusz and Chase, 2014)得出相同的结果.然而, 在2018年发表的系统发育树中, ****们一致认为碗蕨科和真水龙骨类是姐妹群, 两者共同与凤尾蕨科形成姐妹群(Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018). ...
... 鳞片是大部分蕨类植物非常显著的特征, 是识别蕨类植物及对其进行分类的重要依据.在早期的蕨类植物分类研究中(Holttum, 1949, 1957, 1963; Tryon and Tryon, 1982; 于顺利和林尤兴, 1996; 石雷和张宪春, 1999, 2001; Van den Heede et al., 2003; Labiak, 2003; Tsutsumi et al., 2008; Wei et al., 2013; 顾钰峰等, 2014), 鳞片均被作为重要的分类佐证.然而在分子生物学快速发展的今天, 形态学研究渐被忽视, 蕨类植物的鳞片形态研究少有****涉及.Hoshizaki (1970)对鹿角蕨属(Platycerium)的鳞片形态进行了研究.柯勇男(2002)研究了台湾产广义鳞毛蕨科的鳞片形态.Tsutsumi等(2008)对骨碎补科植物的鳞片形态和进化进行了研究.顾钰峰(2015)对蕨类植物的毛状物和鳞片进行了广泛取材观察.他们的研究均认为鳞片对于蕨类植物的分类学研究有非常重要的意义.然而生物性状的表现往往受环境影响, 决定其物种的关键还是内部的基因结构.因此形态特征仅作为参考, 很难起决定性作用. ...
... 鳞片是大部分蕨类植物非常显著的特征, 是识别蕨类植物及对其进行分类的重要依据.在早期的蕨类植物分类研究中(Holttum, 1949, 1957, 1963; Tryon and Tryon, 1982; 于顺利和林尤兴, 1996; 石雷和张宪春, 1999, 2001; Van den Heede et al., 2003; Labiak, 2003; Tsutsumi et al., 2008; Wei et al., 2013; 顾钰峰等, 2014), 鳞片均被作为重要的分类佐证.然而在分子生物学快速发展的今天, 形态学研究渐被忽视, 蕨类植物的鳞片形态研究少有****涉及.Hoshizaki (1970)对鹿角蕨属(Platycerium)的鳞片形态进行了研究.柯勇男(2002)研究了台湾产广义鳞毛蕨科的鳞片形态.Tsutsumi等(2008)对骨碎补科植物的鳞片形态和进化进行了研究.顾钰峰(2015)对蕨类植物的毛状物和鳞片进行了广泛取材观察.他们的研究均认为鳞片对于蕨类植物的分类学研究有非常重要的意义.然而生物性状的表现往往受环境影响, 决定其物种的关键还是内部的基因结构.因此形态特征仅作为参考, 很难起决定性作用. ...
First insights into fern matK phylogeny 2 2011
... 凤尾蕨科(Pteridaceae)植物形态上差异较大, 导致其分类一直存在很大争议(Holttum, 1949; Nayar, 1970; Sermolli, 1977).随着分子生物学的发展, 采用DNA序列构建系统发育树成为研究分类系统的重要手段, 不同****对于凤尾蕨科在蕨类植物分类系统中的位置各持不同观点, 凤尾蕨科所包含的类群也有很大差别(Smith et al., 2006; Schuettpelz et al., 2007; 刘红梅等, 2008; Christenhusz et al., 2011; Lehtonen, 2011; Kuo et al., 2011; 张宪春等, 2013; Christenhusz and Chase, 2014; PPG I, 2016; Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018).Pryer等(2004)认为, 凤尾蕨科与真水龙骨类构成姐妹群, 然后共同与碗蕨科形成姐妹群.Rai和Graham (2010)的研究也得出类似的结果.但是Smith等(2006)和Schneider (2013)的研究结果却相反, 他们认为碗蕨科和真水龙骨类形成姐妹群, 两者共同与凤尾蕨科植物形成姐妹群.在后续研究中, 也有****(Kuo et al., 2011; Christenhusz and Chase, 2014)得出相同的结果.然而, 在2018年发表的系统发育树中, ****们一致认为碗蕨科和真水龙骨类是姐妹群, 两者共同与凤尾蕨科形成姐妹群(Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018). ...
... 的研究结果却相反, 他们认为碗蕨科和真水龙骨类形成姐妹群, 两者共同与凤尾蕨科植物形成姐妹群.在后续研究中, 也有****(Kuo et al., 2011; Christenhusz and Chase, 2014)得出相同的结果.然而, 在2018年发表的系统发育树中, ****们一致认为碗蕨科和真水龙骨类是姐妹群, 两者共同与凤尾蕨科形成姐妹群(Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018). ...
A new combination in the fern genus Ceradenia(Grammitidaceae) 1 2003
... 鳞片是大部分蕨类植物非常显著的特征, 是识别蕨类植物及对其进行分类的重要依据.在早期的蕨类植物分类研究中(Holttum, 1949, 1957, 1963; Tryon and Tryon, 1982; 于顺利和林尤兴, 1996; 石雷和张宪春, 1999, 2001; Van den Heede et al., 2003; Labiak, 2003; Tsutsumi et al., 2008; Wei et al., 2013; 顾钰峰等, 2014), 鳞片均被作为重要的分类佐证.然而在分子生物学快速发展的今天, 形态学研究渐被忽视, 蕨类植物的鳞片形态研究少有****涉及.Hoshizaki (1970)对鹿角蕨属(Platycerium)的鳞片形态进行了研究.柯勇男(2002)研究了台湾产广义鳞毛蕨科的鳞片形态.Tsutsumi等(2008)对骨碎补科植物的鳞片形态和进化进行了研究.顾钰峰(2015)对蕨类植物的毛状物和鳞片进行了广泛取材观察.他们的研究均认为鳞片对于蕨类植物的分类学研究有非常重要的意义.然而生物性状的表现往往受环境影响, 决定其物种的关键还是内部的基因结构.因此形态特征仅作为参考, 很难起决定性作用. ...
Towards resolving the complete fern tree of life 1 2011
... 凤尾蕨科(Pteridaceae)植物形态上差异较大, 导致其分类一直存在很大争议(Holttum, 1949; Nayar, 1970; Sermolli, 1977).随着分子生物学的发展, 采用DNA序列构建系统发育树成为研究分类系统的重要手段, 不同****对于凤尾蕨科在蕨类植物分类系统中的位置各持不同观点, 凤尾蕨科所包含的类群也有很大差别(Smith et al., 2006; Schuettpelz et al., 2007; 刘红梅等, 2008; Christenhusz et al., 2011; Lehtonen, 2011; Kuo et al., 2011; 张宪春等, 2013; Christenhusz and Chase, 2014; PPG I, 2016; Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018).Pryer等(2004)认为, 凤尾蕨科与真水龙骨类构成姐妹群, 然后共同与碗蕨科形成姐妹群.Rai和Graham (2010)的研究也得出类似的结果.但是Smith等(2006)和Schneider (2013)的研究结果却相反, 他们认为碗蕨科和真水龙骨类形成姐妹群, 两者共同与凤尾蕨科植物形成姐妹群.在后续研究中, 也有****(Kuo et al., 2011; Christenhusz and Chase, 2014)得出相同的结果.然而, 在2018年发表的系统发育树中, ****们一致认为碗蕨科和真水龙骨类是姐妹群, 两者共同与凤尾蕨科形成姐妹群(Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018). ...
Mesquite: a modular system for evolutionary analysis 1 2018
A phylogenetic classification of the homosporous ferns 1 1970
... 凤尾蕨科(Pteridaceae)植物形态上差异较大, 导致其分类一直存在很大争议(Holttum, 1949; Nayar, 1970; Sermolli, 1977).随着分子生物学的发展, 采用DNA序列构建系统发育树成为研究分类系统的重要手段, 不同****对于凤尾蕨科在蕨类植物分类系统中的位置各持不同观点, 凤尾蕨科所包含的类群也有很大差别(Smith et al., 2006; Schuettpelz et al., 2007; 刘红梅等, 2008; Christenhusz et al., 2011; Lehtonen, 2011; Kuo et al., 2011; 张宪春等, 2013; Christenhusz and Chase, 2014; PPG I, 2016; Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018).Pryer等(2004)认为, 凤尾蕨科与真水龙骨类构成姐妹群, 然后共同与碗蕨科形成姐妹群.Rai和Graham (2010)的研究也得出类似的结果.但是Smith等(2006)和Schneider (2013)的研究结果却相反, 他们认为碗蕨科和真水龙骨类形成姐妹群, 两者共同与凤尾蕨科植物形成姐妹群.在后续研究中, 也有****(Kuo et al., 2011; Christenhusz and Chase, 2014)得出相同的结果.然而, 在2018年发表的系统发育树中, ****们一致认为碗蕨科和真水龙骨类是姐妹群, 两者共同与凤尾蕨科形成姐妹群(Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018). ...
A community-derived classification for extant lycophytes and ferns 5 2016
... 凤尾蕨科(Pteridaceae)植物形态上差异较大, 导致其分类一直存在很大争议(Holttum, 1949; Nayar, 1970; Sermolli, 1977).随着分子生物学的发展, 采用DNA序列构建系统发育树成为研究分类系统的重要手段, 不同****对于凤尾蕨科在蕨类植物分类系统中的位置各持不同观点, 凤尾蕨科所包含的类群也有很大差别(Smith et al., 2006; Schuettpelz et al., 2007; 刘红梅等, 2008; Christenhusz et al., 2011; Lehtonen, 2011; Kuo et al., 2011; 张宪春等, 2013; Christenhusz and Chase, 2014; PPG I, 2016; Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018).Pryer等(2004)认为, 凤尾蕨科与真水龙骨类构成姐妹群, 然后共同与碗蕨科形成姐妹群.Rai和Graham (2010)的研究也得出类似的结果.但是Smith等(2006)和Schneider (2013)的研究结果却相反, 他们认为碗蕨科和真水龙骨类形成姐妹群, 两者共同与凤尾蕨科植物形成姐妹群.在后续研究中, 也有****(Kuo et al., 2011; Christenhusz and Chase, 2014)得出相同的结果.然而, 在2018年发表的系统发育树中, ****们一致认为碗蕨科和真水龙骨类是姐妹群, 两者共同与凤尾蕨科形成姐妹群(Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018). ...
... 珠蕨亚科包含凤了蕨属、珠蕨属和Llavea三个属(PPG I, 2016), 中国仅有凤了蕨属和珠蕨属分布(Zhang et al., 2013).凤了蕨属原属于裸子蕨科(Hemionitidaceae), 而珠蕨属曾属于中国蕨科(Sinopteridaceae) (秦仁昌, 1978). ...
... 碎米蕨亚科包含了秦仁昌分类系统(秦仁昌, 1978)中裸子蕨科的金毛裸蕨属、泽泻蕨属和中国蕨科的粉背蕨属、碎米蕨属、黑心蕨属、隐囊蕨属和旱蕨属, 将薄鳞蕨属(Leptolepidium)归并入粉背蕨属, 新建立了戟叶黑心蕨属(Calciphilopteris) (Zhang et al., 2013; PPG I, 2016). ...
Phylogeny and evolution of ferns (monilophytes) with a focus on the early leptosporangiate divergences 1 2004
... 凤尾蕨科(Pteridaceae)植物形态上差异较大, 导致其分类一直存在很大争议(Holttum, 1949; Nayar, 1970; Sermolli, 1977).随着分子生物学的发展, 采用DNA序列构建系统发育树成为研究分类系统的重要手段, 不同****对于凤尾蕨科在蕨类植物分类系统中的位置各持不同观点, 凤尾蕨科所包含的类群也有很大差别(Smith et al., 2006; Schuettpelz et al., 2007; 刘红梅等, 2008; Christenhusz et al., 2011; Lehtonen, 2011; Kuo et al., 2011; 张宪春等, 2013; Christenhusz and Chase, 2014; PPG I, 2016; Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018).Pryer等(2004)认为, 凤尾蕨科与真水龙骨类构成姐妹群, 然后共同与碗蕨科形成姐妹群.Rai和Graham (2010)的研究也得出类似的结果.但是Smith等(2006)和Schneider (2013)的研究结果却相反, 他们认为碗蕨科和真水龙骨类形成姐妹群, 两者共同与凤尾蕨科植物形成姐妹群.在后续研究中, 也有****(Kuo et al., 2011; Christenhusz and Chase, 2014)得出相同的结果.然而, 在2018年发表的系统发育树中, ****们一致认为碗蕨科和真水龙骨类是姐妹群, 两者共同与凤尾蕨科形成姐妹群(Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018). ...
A well-resolved fern nuclear phylogeny reveals the evolution history of numerous transcription factor families 2 2018
... 凤尾蕨科(Pteridaceae)植物形态上差异较大, 导致其分类一直存在很大争议(Holttum, 1949; Nayar, 1970; Sermolli, 1977).随着分子生物学的发展, 采用DNA序列构建系统发育树成为研究分类系统的重要手段, 不同****对于凤尾蕨科在蕨类植物分类系统中的位置各持不同观点, 凤尾蕨科所包含的类群也有很大差别(Smith et al., 2006; Schuettpelz et al., 2007; 刘红梅等, 2008; Christenhusz et al., 2011; Lehtonen, 2011; Kuo et al., 2011; 张宪春等, 2013; Christenhusz and Chase, 2014; PPG I, 2016; Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018).Pryer等(2004)认为, 凤尾蕨科与真水龙骨类构成姐妹群, 然后共同与碗蕨科形成姐妹群.Rai和Graham (2010)的研究也得出类似的结果.但是Smith等(2006)和Schneider (2013)的研究结果却相反, 他们认为碗蕨科和真水龙骨类形成姐妹群, 两者共同与凤尾蕨科植物形成姐妹群.在后续研究中, 也有****(Kuo et al., 2011; Christenhusz and Chase, 2014)得出相同的结果.然而, 在2018年发表的系统发育树中, ****们一致认为碗蕨科和真水龙骨类是姐妹群, 两者共同与凤尾蕨科形成姐妹群(Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018). ...
... ; Qi et al., 2018). ...
Utility of a large, multigene plastid data set in inferring higher-order relationships in ferns and relatives (monilophytes) 1 2010
... 凤尾蕨科(Pteridaceae)植物形态上差异较大, 导致其分类一直存在很大争议(Holttum, 1949; Nayar, 1970; Sermolli, 1977).随着分子生物学的发展, 采用DNA序列构建系统发育树成为研究分类系统的重要手段, 不同****对于凤尾蕨科在蕨类植物分类系统中的位置各持不同观点, 凤尾蕨科所包含的类群也有很大差别(Smith et al., 2006; Schuettpelz et al., 2007; 刘红梅等, 2008; Christenhusz et al., 2011; Lehtonen, 2011; Kuo et al., 2011; 张宪春等, 2013; Christenhusz and Chase, 2014; PPG I, 2016; Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018).Pryer等(2004)认为, 凤尾蕨科与真水龙骨类构成姐妹群, 然后共同与碗蕨科形成姐妹群.Rai和Graham (2010)的研究也得出类似的结果.但是Smith等(2006)和Schneider (2013)的研究结果却相反, 他们认为碗蕨科和真水龙骨类形成姐妹群, 两者共同与凤尾蕨科植物形成姐妹群.在后续研究中, 也有****(Kuo et al., 2011; Christenhusz and Chase, 2014)得出相同的结果.然而, 在2018年发表的系统发育树中, ****们一致认为碗蕨科和真水龙骨类是姐妹群, 两者共同与凤尾蕨科形成姐妹群(Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018). ...
Evolutionary morphology of ferns (monilophytes). In: Ambrose BA, Purugganan M, eds 1 2013
... 凤尾蕨科(Pteridaceae)植物形态上差异较大, 导致其分类一直存在很大争议(Holttum, 1949; Nayar, 1970; Sermolli, 1977).随着分子生物学的发展, 采用DNA序列构建系统发育树成为研究分类系统的重要手段, 不同****对于凤尾蕨科在蕨类植物分类系统中的位置各持不同观点, 凤尾蕨科所包含的类群也有很大差别(Smith et al., 2006; Schuettpelz et al., 2007; 刘红梅等, 2008; Christenhusz et al., 2011; Lehtonen, 2011; Kuo et al., 2011; 张宪春等, 2013; Christenhusz and Chase, 2014; PPG I, 2016; Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018).Pryer等(2004)认为, 凤尾蕨科与真水龙骨类构成姐妹群, 然后共同与碗蕨科形成姐妹群.Rai和Graham (2010)的研究也得出类似的结果.但是Smith等(2006)和Schneider (2013)的研究结果却相反, 他们认为碗蕨科和真水龙骨类形成姐妹群, 两者共同与凤尾蕨科植物形成姐妹群.在后续研究中, 也有****(Kuo et al., 2011; Christenhusz and Chase, 2014)得出相同的结果.然而, 在2018年发表的系统发育树中, ****们一致认为碗蕨科和真水龙骨类是姐妹群, 两者共同与凤尾蕨科形成姐妹群(Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018). ...
A molecular phylogeny of the fern family Pteridaceae: assessing overall relationships and the affinities of previously unsampled genera 2 2007
... 凤尾蕨科(Pteridaceae)植物形态上差异较大, 导致其分类一直存在很大争议(Holttum, 1949; Nayar, 1970; Sermolli, 1977).随着分子生物学的发展, 采用DNA序列构建系统发育树成为研究分类系统的重要手段, 不同****对于凤尾蕨科在蕨类植物分类系统中的位置各持不同观点, 凤尾蕨科所包含的类群也有很大差别(Smith et al., 2006; Schuettpelz et al., 2007; 刘红梅等, 2008; Christenhusz et al., 2011; Lehtonen, 2011; Kuo et al., 2011; 张宪春等, 2013; Christenhusz and Chase, 2014; PPG I, 2016; Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018).Pryer等(2004)认为, 凤尾蕨科与真水龙骨类构成姐妹群, 然后共同与碗蕨科形成姐妹群.Rai和Graham (2010)的研究也得出类似的结果.但是Smith等(2006)和Schneider (2013)的研究结果却相反, 他们认为碗蕨科和真水龙骨类形成姐妹群, 两者共同与凤尾蕨科植物形成姐妹群.在后续研究中, 也有****(Kuo et al., 2011; Christenhusz and Chase, 2014)得出相同的结果.然而, 在2018年发表的系统发育树中, ****们一致认为碗蕨科和真水龙骨类是姐妹群, 两者共同与凤尾蕨科形成姐妹群(Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018). ...
Tentamen Pteridophytorum genera in taxonomicum ordinem redigendi 1 1977
... 凤尾蕨科(Pteridaceae)植物形态上差异较大, 导致其分类一直存在很大争议(Holttum, 1949; Nayar, 1970; Sermolli, 1977).随着分子生物学的发展, 采用DNA序列构建系统发育树成为研究分类系统的重要手段, 不同****对于凤尾蕨科在蕨类植物分类系统中的位置各持不同观点, 凤尾蕨科所包含的类群也有很大差别(Smith et al., 2006; Schuettpelz et al., 2007; 刘红梅等, 2008; Christenhusz et al., 2011; Lehtonen, 2011; Kuo et al., 2011; 张宪春等, 2013; Christenhusz and Chase, 2014; PPG I, 2016; Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018).Pryer等(2004)认为, 凤尾蕨科与真水龙骨类构成姐妹群, 然后共同与碗蕨科形成姐妹群.Rai和Graham (2010)的研究也得出类似的结果.但是Smith等(2006)和Schneider (2013)的研究结果却相反, 他们认为碗蕨科和真水龙骨类形成姐妹群, 两者共同与凤尾蕨科植物形成姐妹群.在后续研究中, 也有****(Kuo et al., 2011; Christenhusz and Chase, 2014)得出相同的结果.然而, 在2018年发表的系统发育树中, ****们一致认为碗蕨科和真水龙骨类是姐妹群, 两者共同与凤尾蕨科形成姐妹群(Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018). ...
Large-scale phylogenomic analysis resolves a backbone phylogeny in ferns 2 2018
... 凤尾蕨科(Pteridaceae)植物形态上差异较大, 导致其分类一直存在很大争议(Holttum, 1949; Nayar, 1970; Sermolli, 1977).随着分子生物学的发展, 采用DNA序列构建系统发育树成为研究分类系统的重要手段, 不同****对于凤尾蕨科在蕨类植物分类系统中的位置各持不同观点, 凤尾蕨科所包含的类群也有很大差别(Smith et al., 2006; Schuettpelz et al., 2007; 刘红梅等, 2008; Christenhusz et al., 2011; Lehtonen, 2011; Kuo et al., 2011; 张宪春等, 2013; Christenhusz and Chase, 2014; PPG I, 2016; Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018).Pryer等(2004)认为, 凤尾蕨科与真水龙骨类构成姐妹群, 然后共同与碗蕨科形成姐妹群.Rai和Graham (2010)的研究也得出类似的结果.但是Smith等(2006)和Schneider (2013)的研究结果却相反, 他们认为碗蕨科和真水龙骨类形成姐妹群, 两者共同与凤尾蕨科植物形成姐妹群.在后续研究中, 也有****(Kuo et al., 2011; Christenhusz and Chase, 2014)得出相同的结果.然而, 在2018年发表的系统发育树中, ****们一致认为碗蕨科和真水龙骨类是姐妹群, 两者共同与凤尾蕨科形成姐妹群(Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018). ...
... ; Shen et al., 2018; Qi et al., 2018). ...
A classification for extant ferns 2 2006
... 凤尾蕨科(Pteridaceae)植物形态上差异较大, 导致其分类一直存在很大争议(Holttum, 1949; Nayar, 1970; Sermolli, 1977).随着分子生物学的发展, 采用DNA序列构建系统发育树成为研究分类系统的重要手段, 不同****对于凤尾蕨科在蕨类植物分类系统中的位置各持不同观点, 凤尾蕨科所包含的类群也有很大差别(Smith et al., 2006; Schuettpelz et al., 2007; 刘红梅等, 2008; Christenhusz et al., 2011; Lehtonen, 2011; Kuo et al., 2011; 张宪春等, 2013; Christenhusz and Chase, 2014; PPG I, 2016; Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018).Pryer等(2004)认为, 凤尾蕨科与真水龙骨类构成姐妹群, 然后共同与碗蕨科形成姐妹群.Rai和Graham (2010)的研究也得出类似的结果.但是Smith等(2006)和Schneider (2013)的研究结果却相反, 他们认为碗蕨科和真水龙骨类形成姐妹群, 两者共同与凤尾蕨科植物形成姐妹群.在后续研究中, 也有****(Kuo et al., 2011; Christenhusz and Chase, 2014)得出相同的结果.然而, 在2018年发表的系统发育树中, ****们一致认为碗蕨科和真水龙骨类是姐妹群, 两者共同与凤尾蕨科形成姐妹群(Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018). ...
... 的研究也得出类似的结果.但是Smith等(2006)和Schneider (2013)的研究结果却相反, 他们认为碗蕨科和真水龙骨类形成姐妹群, 两者共同与凤尾蕨科植物形成姐妹群.在后续研究中, 也有****(Kuo et al., 2011; Christenhusz and Chase, 2014)得出相同的结果.然而, 在2018年发表的系统发育树中, ****们一致认为碗蕨科和真水龙骨类是姐妹群, 两者共同与凤尾蕨科形成姐妹群(Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018). ...
Ferns and Allied Plants 1 1982
... 鳞片是大部分蕨类植物非常显著的特征, 是识别蕨类植物及对其进行分类的重要依据.在早期的蕨类植物分类研究中(Holttum, 1949, 1957, 1963; Tryon and Tryon, 1982; 于顺利和林尤兴, 1996; 石雷和张宪春, 1999, 2001; Van den Heede et al., 2003; Labiak, 2003; Tsutsumi et al., 2008; Wei et al., 2013; 顾钰峰等, 2014), 鳞片均被作为重要的分类佐证.然而在分子生物学快速发展的今天, 形态学研究渐被忽视, 蕨类植物的鳞片形态研究少有****涉及.Hoshizaki (1970)对鹿角蕨属(Platycerium)的鳞片形态进行了研究.柯勇男(2002)研究了台湾产广义鳞毛蕨科的鳞片形态.Tsutsumi等(2008)对骨碎补科植物的鳞片形态和进化进行了研究.顾钰峰(2015)对蕨类植物的毛状物和鳞片进行了广泛取材观察.他们的研究均认为鳞片对于蕨类植物的分类学研究有非常重要的意义.然而生物性状的表现往往受环境影响, 决定其物种的关键还是内部的基因结构.因此形态特征仅作为参考, 很难起决定性作用. ...
Morphology and evolution of epiphytic Davalliaceae scales 1 2008
Toward a new circumscription of the twinsorus-fern genus (Diplazium Athyriaceae): a molecular phylogeny with morphological implications and infrageneric taxonomy 2 2013
... 鳞片是大部分蕨类植物非常显著的特征, 是识别蕨类植物及对其进行分类的重要依据.在早期的蕨类植物分类研究中(Holttum, 1949, 1957, 1963; Tryon and Tryon, 1982; 于顺利和林尤兴, 1996; 石雷和张宪春, 1999, 2001; Van den Heede et al., 2003; Labiak, 2003; Tsutsumi et al., 2008; Wei et al., 2013; 顾钰峰等, 2014), 鳞片均被作为重要的分类佐证.然而在分子生物学快速发展的今天, 形态学研究渐被忽视, 蕨类植物的鳞片形态研究少有****涉及.Hoshizaki (1970)对鹿角蕨属(Platycerium)的鳞片形态进行了研究.柯勇男(2002)研究了台湾产广义鳞毛蕨科的鳞片形态.Tsutsumi等(2008)对骨碎补科植物的鳞片形态和进化进行了研究.顾钰峰(2015)对蕨类植物的毛状物和鳞片进行了广泛取材观察.他们的研究均认为鳞片对于蕨类植物的分类学研究有非常重要的意义.然而生物性状的表现往往受环境影响, 决定其物种的关键还是内部的基因结构.因此形态特征仅作为参考, 很难起决定性作用. ...
Target sequence capture of nuclear-encoded genes for phylogenetic analysis in ferns 2 2018
... 凤尾蕨科(Pteridaceae)植物形态上差异较大, 导致其分类一直存在很大争议(Holttum, 1949; Nayar, 1970; Sermolli, 1977).随着分子生物学的发展, 采用DNA序列构建系统发育树成为研究分类系统的重要手段, 不同****对于凤尾蕨科在蕨类植物分类系统中的位置各持不同观点, 凤尾蕨科所包含的类群也有很大差别(Smith et al., 2006; Schuettpelz et al., 2007; 刘红梅等, 2008; Christenhusz et al., 2011; Lehtonen, 2011; Kuo et al., 2011; 张宪春等, 2013; Christenhusz and Chase, 2014; PPG I, 2016; Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018).Pryer等(2004)认为, 凤尾蕨科与真水龙骨类构成姐妹群, 然后共同与碗蕨科形成姐妹群.Rai和Graham (2010)的研究也得出类似的结果.但是Smith等(2006)和Schneider (2013)的研究结果却相反, 他们认为碗蕨科和真水龙骨类形成姐妹群, 两者共同与凤尾蕨科植物形成姐妹群.在后续研究中, 也有****(Kuo et al., 2011; Christenhusz and Chase, 2014)得出相同的结果.然而, 在2018年发表的系统发育树中, ****们一致认为碗蕨科和真水龙骨类是姐妹群, 两者共同与凤尾蕨科形成姐妹群(Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018). ...
... )得出相同的结果.然而, 在2018年发表的系统发育树中, ****们一致认为碗蕨科和真水龙骨类是姐妹群, 两者共同与凤尾蕨科形成姐妹群(Wolf et al., 2018; Shen et al., 2018; Qi et al., 2018). ...
 ,1,*
,1,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT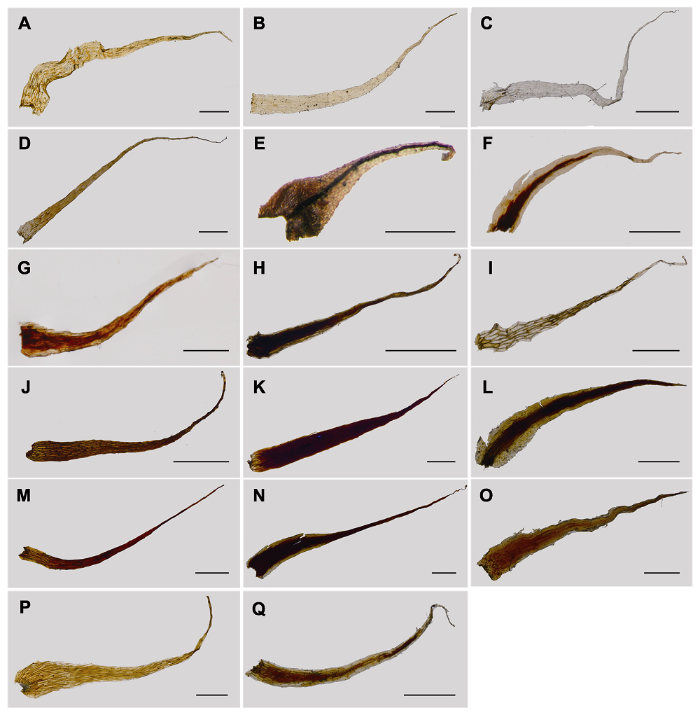 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT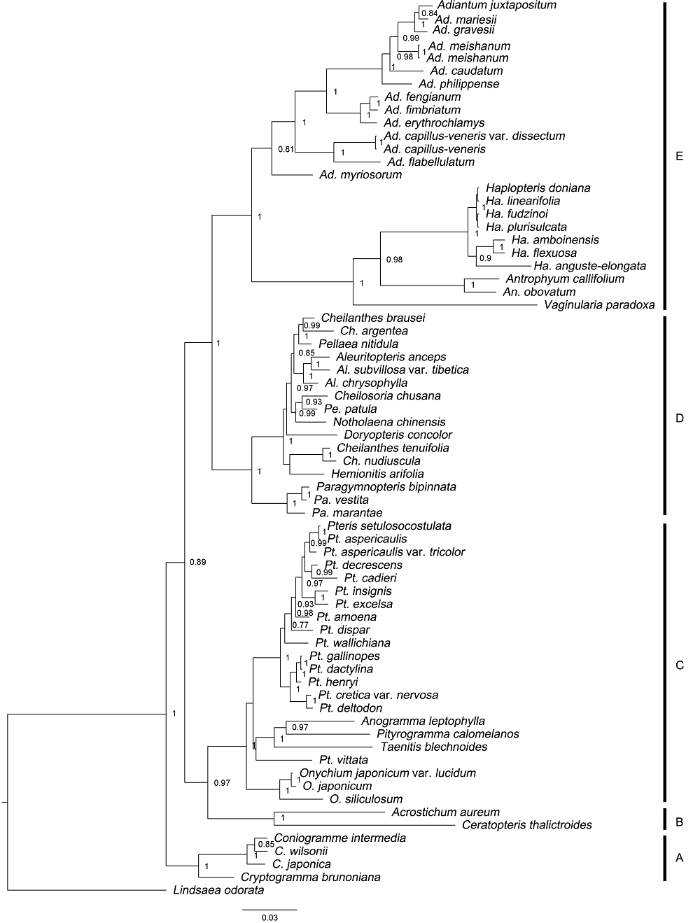 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT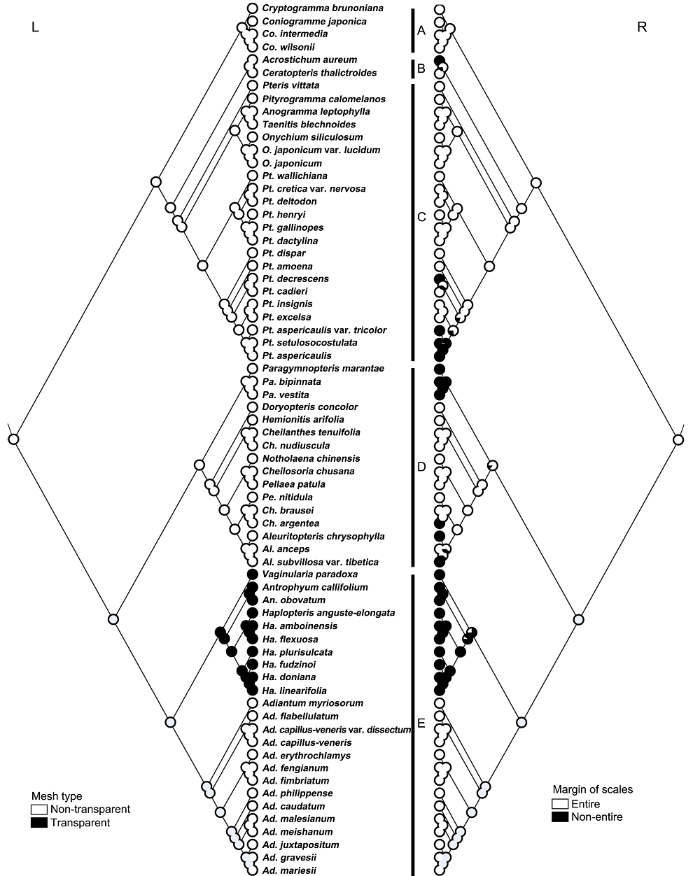 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}