Patterns and Influence Factors of Fine Root Turnover in Forest Ecosystems
Jianing Zhao1, Yun Liang2, Ying Liu3, Yujue Wang1, Qianru Yang1, Chunwang Xiao,1,*1College of Life and Environmental Sciences, Minzu University of China, Beijing 100081, China 2State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 3College of Geomatics, Xi’an University of Science and Technology, Xi’an 710054, China
Abstract Root turnover is a key process in the carbon cycle of terrestrial ecosystems, and plays an important role in studying soil carbon pool changes and global climate change. Various methods for measuring root turnover rate have been described, from which highly variable results are obtained. Moreover, the studies on root turnover in large regional scales are not sufficient, making the patterns of root turnover in global forest ecosystem still not clear. This study integrates the fine root turnover spatial pattern of five forest types in the world by collecting literature data and unifying the calculation method of turnover rate. Combined with soil physical and chemical properties and climate data, several factors driving the fine root turnover of forest ecosystems are obtained. We show that there was a significant difference in the different forest ecosystems and the fine root turnover rate almost decreased with the increase of latitude. The fine root turnover rate of forest ecosystems was positively correlated to the mean annual temperature and the mean annual precipitation. The fine root turnover rate of forest ecosystems was positively correlated to the soil organic carbon content but negatively correlated to soil pH. This study provides a model to study the fine root turnover laws and mechanisms of forest ecosystems. Keywords:fine root turnover;global forest scale;climate factor;soil properties
PDF (965KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 赵佳宁, 梁韵, 柳莹, 王玉珏, 杨倩茹, 肖春旺. 森林生态系统细根周转规律及影响因素. 植物学报, 2020, 55(3): 308-317 doi:10.11983/CBB19231 Zhao Jianing, Liang Yun, Liu Ying, Wang Yujue, Yang Qianru, Xiao Chunwang. Patterns and Influence Factors of Fine Root Turnover in Forest Ecosystems. Chinese Bulletin of Botany, 2020, 55(3): 308-317 doi:10.11983/CBB19231
森林是陆地生态系统的主体, 也是陆地上最庞大、最复杂、多物种、多功能与多效益的生态系统, 在保障国土生态安全、改善生态环境、维持人与生物圈的生态平衡及维护生物多样性等方面起着不可或缺的作用。而研究不同气候带典型森林细根周转对揭示森林生态系统物质循环和能量流动机制有重要意义。目前, 针对森林生态系统细根周转的研究主要集中在某一生态系统季节变化以及环境因子方面(梅莉等, 2004; Wang et al., 2017; Cai et al., 2019), 在全球尺度上的森林生态系统细根周转研究较少。
Table 2 表2 表2不同森林类型的细根周转率 Table 2Fine root turnover rate in different types of forest ecosystem
Forest type
Data number
Means ± SE
Tropical rainforest
11
1.312±0.182 a
Subtropical evergreen broad-leaved forest
9
0.802±0.161 b
Warm temperate deciduous broad-leaved forest
13
0.724±0.859 b
Temperate coniferous and broad-leaved mixed forest
30
0.766±0.995 b
Cold temperate coniferous forest
7
0.602±0.106 b
Different lowercase letters within a soil layer indicate significant differences (P<0.05), according to the Duncan post hoc test. 通过Duncan多重比较检验, 不同小写字母表示差异显著(P< 0.05)。
*表示在0.05水平显著相关。 Figure 2Relationships between fine root turnover rate in forest ecosystems and mean annual temperature (A) and mean annual precipitation (B)
* indicates significant correlation at the 0.05 level.
DiabateB (2016). 松嫩草地地下净生产力和细根动态对不同的管理实践的响应 博士论文. 长春: 东北师范大学. pp. 78-79. [本文引用: 1]
AberJD, MelilloJM, NadelhofferKJ, McClaughertyCA, PastorJ (1985). Fine root turnover in forest ecosystems in relation to quantity and form of nitrogen availability: a comparison of two methods Oecologia 66, 317-321. [本文引用: 1]
ArthurMA, FaheyTJ (1992). Biomass and nutrients in an Engelmann spruce—subalpine fir forest in north central Colorado: pools, annual production, and internal cycling Can J For Res 22, 315-325. [本文引用: 1]
BaiWM, WanSQ, NiuSL, LiuWX, ChenQS, WangQB, ZhangWH, HanXG, LiLH (2010). Increased temperature and precipitation interact to affect root production, mortality, and turnover in a temperate steppe: implications for ecosystem C cycling Glob Chang Biol 16, 1306-1316. [本文引用: 1]
BrunnerI, BakkerMR, Bj?rkRG, HiranoY, LukacM, ArandaX, B?rjaI, EldhusetTD, HelmisaariHS, JourdanC, Kon?pkaB, LópezBC, PérezCM, PerssonH, OstonenI (2013). Fine-root turnover rates of European forests revisited: an analysis of data from sequential coring and ingrowth cores Plant Soil 362, 357-372. [本文引用: 3]
CaiHY, LiFR, JinGZ (2019). Fine root biomass, production and turnover rates in plantations versus natural forests: effects of stand characteristics and soil properties Plant Soil 436, 463-474. [本文引用: 1]
CavelierJ, WrightSJ, SantamariaJ (1999). Effects of irrigation on litterfall, fine root biomass and production in a semideciduous lowland forest in Panama Plant Soil 211, 207-213. [本文引用: 1]
GillRA, JacksonRB (2000). Global patterns of root turnover for terrestrial ecosystems New Phytol 147, 13-31. [本文引用: 4]
HenryAL, ClelandEE, FieldCB, VitousekPM (2005). Interactive effects of elevated CO2, N deposition and climate change on plant litter quality in a California annual grassland Oecologia 142, 465-473. [本文引用: 1]
IversenCM, LedfordJ, NorbyRJ (2008). CO2 enrichment increases carbon and nitrogen input from fine roots in a deciduous forest New Phytol 179, 837-847. [本文引用: 1]
JoslinJD, WolfeMH, HansonPJ (2000). Effects of altered water regimes on forest root systems New Phytol 147, 117-129. [本文引用: 1]
KosolaKR, EissenstatDM, GrahamJH (1995). Root demography of mature citrus trees: the influence of Phytophthora nicotianae Plant Soil 171, 283-288. [本文引用: 1]
LiuL, YangF, WangYJ, ShenX, JanssensIA, GuenetB, XiaoCW (2019). Fine-root turnover, litterfall, and soil microbial community of three mixed coniferous-deciduous forests dominated by Korean pine (Pinus koraiensis) along a latitudinal gradient Front Plant Sci 10, 1298. [本文引用: 2]
LiuW, WangGL, YuKX, LiP, XiaoL, LiuGB (2018). A new method to optimize root order classification based on the diameter interval of fine root Sci Rep 8, 2960. [本文引用: 1]
LuoYQ (2003). Uncertainties in interpretation of isotope signals for estimation of fine root longevity: theoretical considerations Glob Chang Biol 9, 1118-1129. [本文引用: 1]
MajdiH, PregitzerK, MorénAS, NylundJE, ?grenGI (2005). Measuring fine root turnover in forest ecosystems Plant Soil 276, 1-8. [本文引用: 1]
MatamalaR, Gonzàlez-MelerMA, JastrowJD, NorbyRJ, SchlesingerWH (2003). Impacts of fine root turnover on forest NPP and soil C sequestration potential Science 302, 1385-1387. URLPMID:14631037 [本文引用: 1]
PritchardSG, StrandAE (2008). Can you believe what you see? Reconciling minirhizotron and isotopically derived estimates of fine root longevity New Phytol 177, 287-291. [本文引用: 3]
SilverWL, MiyaRK (2001). Global patterns in root decomposition: comparisons of climate and litter quality effects Oecologia 129, 407-419. URLPMID:28547196 [本文引用: 2]
VogtKA, VogtDJ, BloomfieldJ (1998). Analysis of some direct and indirect methods for estimating root biomass and production of forests at an ecosystem level Plant Soil 200, 71-89. [本文引用: 3]
VogtKA, VogtDJ, PalmiottoPA, BoonP, O’HaraJ, AsbjornsenH (1995). Review of root dynamics in forest ecosystems grouped by climate, climatic forest type and species Plant Soil 187, 159-219. [本文引用: 1]
WangAS, AngleJS, ChaneyRL, DelormeTA, McIntoshM (2006). Changes in soil biological activities under reduced soil pH during Thlaspi caerulescens phytoextraction Soil Biol Biochem 38, 1451-1461. [本文引用: 1]
WangSZ, WangZQ, GuJC (2017). Variation patterns of fine root biomass, production and turnover in Chinese forests J For Res 28, 1185-1194. [本文引用: 2]
XiaoCW, JanssensIA, SangWG, WangRZ, XieZQ, PeiZQ, YiY (2010). Belowground carbon pools and dynamics in China’s warm temperate and sub-tropical deciduous forests Biogeosciences 7, 275-287. [本文引用: 2]
XiaoCW, SangWG, WangRZ (2008). Fine root dynamics and turnover rate in an Asia white birch forest of Donglingshan Mountain, China For Ecol Manage 255, 765-773. [本文引用: 2]
YuanZY, ChenHYH (2010). Fine root biomass, production, turnover rates, and nutrient contents in boreal forest ecosystems in relation to species, climate, fertility, and stand age: literature review and meta-analyses CRC Crit Rev Plant Sci 29, 204-221. [本文引用: 2]
ZhouY, SuJQ, JanssensIA, ZhouGS, XiaoCW (2014). Fine root and litterfall dynamics of three Korean pine (Pinus koraiensis) forests along an altitudinal gradient Plant Soil 374, 19-32. [本文引用: 1]
... 森林是陆地生态系统的主体, 也是陆地上最庞大、最复杂、多物种、多功能与多效益的生态系统, 在保障国土生态安全、改善生态环境、维持人与生物圈的生态平衡及维护生物多样性等方面起着不可或缺的作用.而研究不同气候带典型森林细根周转对揭示森林生态系统物质循环和能量流动机制有重要意义.目前, 针对森林生态系统细根周转的研究主要集中在某一生态系统季节变化以及环境因子方面(梅莉等, 2004; Wang et al., 2017; Cai et al., 2019), 在全球尺度上的森林生态系统细根周转研究较少. ...
... 对森林生态系统细根周转率的测量方法多种多样, 过去几十年里产生了许多估算陆地生态系统细根周转的方法(Vogt et al., 1998; Majdi et al., 2005).现今常用的有根钻法(soil coring methods) (Pritchard and Strand, 2008; Xiao et al., 2008; Zhou et al., 2014)、内生长法(ingrowth coring methods) (Xiao et al., 2010; Liu et al., 2019)、微根管法(minirhizotron methods) (Trumbore and Gaudinski, 2003; Iversen et al., 2008; Bai et al., 2010)、碳氮平衡法(C and N budget methods) (Vogt et al., 1998)及同位素14C和13C法(Luo, 2003; Pritchard and Strand, 2008).其中, 根钻法是应用最早的一种方法(Pritchard and Strand, 2008; Xiao et al., 2008), 也是研究根系生物量、生产力和周转率最常用的方法(吴伊波等, 2014).该方法的优点是可相对容易和快速地估计根产量, 且能同时得到较准确的根系生物量数据(肖春旺等, 2017), 在一些需要具体生物量或生产力的研究中有明显优势.Brunner等(2013)对比分析了测量细根周转率的不同方法, 发现在一些浅层土壤及沙石含量较多的土壤和陡峭的斜坡中, 使用根钻法得到的结果更准确且更具可比性.但根钻法需要连续多次测量, 工作量大, 费时费力, 而且取样时可能会错过根系生物量的高峰或低谷(肖春旺等, 2017).由于根钻法计算时要取差值, 有研究认为该方法只适用于细根生物量波动很大的植物根系(Brunner et al., 2013). ...
... 对森林生态系统细根周转率的测量方法多种多样, 过去几十年里产生了许多估算陆地生态系统细根周转的方法(Vogt et al., 1998; Majdi et al., 2005).现今常用的有根钻法(soil coring methods) (Pritchard and Strand, 2008; Xiao et al., 2008; Zhou et al., 2014)、内生长法(ingrowth coring methods) (Xiao et al., 2010; Liu et al., 2019)、微根管法(minirhizotron methods) (Trumbore and Gaudinski, 2003; Iversen et al., 2008; Bai et al., 2010)、碳氮平衡法(C and N budget methods) (Vogt et al., 1998)及同位素14C和13C法(Luo, 2003; Pritchard and Strand, 2008).其中, 根钻法是应用最早的一种方法(Pritchard and Strand, 2008; Xiao et al., 2008), 也是研究根系生物量、生产力和周转率最常用的方法(吴伊波等, 2014).该方法的优点是可相对容易和快速地估计根产量, 且能同时得到较准确的根系生物量数据(肖春旺等, 2017), 在一些需要具体生物量或生产力的研究中有明显优势.Brunner等(2013)对比分析了测量细根周转率的不同方法, 发现在一些浅层土壤及沙石含量较多的土壤和陡峭的斜坡中, 使用根钻法得到的结果更准确且更具可比性.但根钻法需要连续多次测量, 工作量大, 费时费力, 而且取样时可能会错过根系生物量的高峰或低谷(肖春旺等, 2017).由于根钻法计算时要取差值, 有研究认为该方法只适用于细根生物量波动很大的植物根系(Brunner et al., 2013). ...
Increased temperature and precipitation interact to affect root production, mortality, and turnover in a temperate steppe: implications for ecosystem C cycling 1 2010
... 对森林生态系统细根周转率的测量方法多种多样, 过去几十年里产生了许多估算陆地生态系统细根周转的方法(Vogt et al., 1998; Majdi et al., 2005).现今常用的有根钻法(soil coring methods) (Pritchard and Strand, 2008; Xiao et al., 2008; Zhou et al., 2014)、内生长法(ingrowth coring methods) (Xiao et al., 2010; Liu et al., 2019)、微根管法(minirhizotron methods) (Trumbore and Gaudinski, 2003; Iversen et al., 2008; Bai et al., 2010)、碳氮平衡法(C and N budget methods) (Vogt et al., 1998)及同位素14C和13C法(Luo, 2003; Pritchard and Strand, 2008).其中, 根钻法是应用最早的一种方法(Pritchard and Strand, 2008; Xiao et al., 2008), 也是研究根系生物量、生产力和周转率最常用的方法(吴伊波等, 2014).该方法的优点是可相对容易和快速地估计根产量, 且能同时得到较准确的根系生物量数据(肖春旺等, 2017), 在一些需要具体生物量或生产力的研究中有明显优势.Brunner等(2013)对比分析了测量细根周转率的不同方法, 发现在一些浅层土壤及沙石含量较多的土壤和陡峭的斜坡中, 使用根钻法得到的结果更准确且更具可比性.但根钻法需要连续多次测量, 工作量大, 费时费力, 而且取样时可能会错过根系生物量的高峰或低谷(肖春旺等, 2017).由于根钻法计算时要取差值, 有研究认为该方法只适用于细根生物量波动很大的植物根系(Brunner et al., 2013). ...
Fine-root turnover rates of European forests revisited: an analysis of data from sequential coring and ingrowth cores 3 2013
... 对森林生态系统细根周转率的测量方法多种多样, 过去几十年里产生了许多估算陆地生态系统细根周转的方法(Vogt et al., 1998; Majdi et al., 2005).现今常用的有根钻法(soil coring methods) (Pritchard and Strand, 2008; Xiao et al., 2008; Zhou et al., 2014)、内生长法(ingrowth coring methods) (Xiao et al., 2010; Liu et al., 2019)、微根管法(minirhizotron methods) (Trumbore and Gaudinski, 2003; Iversen et al., 2008; Bai et al., 2010)、碳氮平衡法(C and N budget methods) (Vogt et al., 1998)及同位素14C和13C法(Luo, 2003; Pritchard and Strand, 2008).其中, 根钻法是应用最早的一种方法(Pritchard and Strand, 2008; Xiao et al., 2008), 也是研究根系生物量、生产力和周转率最常用的方法(吴伊波等, 2014).该方法的优点是可相对容易和快速地估计根产量, 且能同时得到较准确的根系生物量数据(肖春旺等, 2017), 在一些需要具体生物量或生产力的研究中有明显优势.Brunner等(2013)对比分析了测量细根周转率的不同方法, 发现在一些浅层土壤及沙石含量较多的土壤和陡峭的斜坡中, 使用根钻法得到的结果更准确且更具可比性.但根钻法需要连续多次测量, 工作量大, 费时费力, 而且取样时可能会错过根系生物量的高峰或低谷(肖春旺等, 2017).由于根钻法计算时要取差值, 有研究认为该方法只适用于细根生物量波动很大的植物根系(Brunner et al., 2013). ...
... ).由于根钻法计算时要取差值, 有研究认为该方法只适用于细根生物量波动很大的植物根系(Brunner et al., 2013). ...
Fine root biomass, production and turnover rates in plantations versus natural forests: effects of stand characteristics and soil properties 1 2019
... 森林是陆地生态系统的主体, 也是陆地上最庞大、最复杂、多物种、多功能与多效益的生态系统, 在保障国土生态安全、改善生态环境、维持人与生物圈的生态平衡及维护生物多样性等方面起着不可或缺的作用.而研究不同气候带典型森林细根周转对揭示森林生态系统物质循环和能量流动机制有重要意义.目前, 针对森林生态系统细根周转的研究主要集中在某一生态系统季节变化以及环境因子方面(梅莉等, 2004; Wang et al., 2017; Cai et al., 2019), 在全球尺度上的森林生态系统细根周转研究较少. ...
Effects of irrigation on litterfall, fine root biomass and production in a semideciduous lowland forest in Panama 1 1999
CO2 enrichment increases carbon and nitrogen input from fine roots in a deciduous forest 1 2008
... 对森林生态系统细根周转率的测量方法多种多样, 过去几十年里产生了许多估算陆地生态系统细根周转的方法(Vogt et al., 1998; Majdi et al., 2005).现今常用的有根钻法(soil coring methods) (Pritchard and Strand, 2008; Xiao et al., 2008; Zhou et al., 2014)、内生长法(ingrowth coring methods) (Xiao et al., 2010; Liu et al., 2019)、微根管法(minirhizotron methods) (Trumbore and Gaudinski, 2003; Iversen et al., 2008; Bai et al., 2010)、碳氮平衡法(C and N budget methods) (Vogt et al., 1998)及同位素14C和13C法(Luo, 2003; Pritchard and Strand, 2008).其中, 根钻法是应用最早的一种方法(Pritchard and Strand, 2008; Xiao et al., 2008), 也是研究根系生物量、生产力和周转率最常用的方法(吴伊波等, 2014).该方法的优点是可相对容易和快速地估计根产量, 且能同时得到较准确的根系生物量数据(肖春旺等, 2017), 在一些需要具体生物量或生产力的研究中有明显优势.Brunner等(2013)对比分析了测量细根周转率的不同方法, 发现在一些浅层土壤及沙石含量较多的土壤和陡峭的斜坡中, 使用根钻法得到的结果更准确且更具可比性.但根钻法需要连续多次测量, 工作量大, 费时费力, 而且取样时可能会错过根系生物量的高峰或低谷(肖春旺等, 2017).由于根钻法计算时要取差值, 有研究认为该方法只适用于细根生物量波动很大的植物根系(Brunner et al., 2013). ...
Effects of altered water regimes on forest root systems 1 2000
Fine-root turnover, litterfall, and soil microbial community of three mixed coniferous-deciduous forests dominated by Korean pine (Pinus koraiensis) along a latitudinal gradient 2 2019
... 对森林生态系统细根周转率的测量方法多种多样, 过去几十年里产生了许多估算陆地生态系统细根周转的方法(Vogt et al., 1998; Majdi et al., 2005).现今常用的有根钻法(soil coring methods) (Pritchard and Strand, 2008; Xiao et al., 2008; Zhou et al., 2014)、内生长法(ingrowth coring methods) (Xiao et al., 2010; Liu et al., 2019)、微根管法(minirhizotron methods) (Trumbore and Gaudinski, 2003; Iversen et al., 2008; Bai et al., 2010)、碳氮平衡法(C and N budget methods) (Vogt et al., 1998)及同位素14C和13C法(Luo, 2003; Pritchard and Strand, 2008).其中, 根钻法是应用最早的一种方法(Pritchard and Strand, 2008; Xiao et al., 2008), 也是研究根系生物量、生产力和周转率最常用的方法(吴伊波等, 2014).该方法的优点是可相对容易和快速地估计根产量, 且能同时得到较准确的根系生物量数据(肖春旺等, 2017), 在一些需要具体生物量或生产力的研究中有明显优势.Brunner等(2013)对比分析了测量细根周转率的不同方法, 发现在一些浅层土壤及沙石含量较多的土壤和陡峭的斜坡中, 使用根钻法得到的结果更准确且更具可比性.但根钻法需要连续多次测量, 工作量大, 费时费力, 而且取样时可能会错过根系生物量的高峰或低谷(肖春旺等, 2017).由于根钻法计算时要取差值, 有研究认为该方法只适用于细根生物量波动很大的植物根系(Brunner et al., 2013). ...
Uncertainties in interpretation of isotope signals for estimation of fine root longevity: theoretical considerations 1 2003
... 对森林生态系统细根周转率的测量方法多种多样, 过去几十年里产生了许多估算陆地生态系统细根周转的方法(Vogt et al., 1998; Majdi et al., 2005).现今常用的有根钻法(soil coring methods) (Pritchard and Strand, 2008; Xiao et al., 2008; Zhou et al., 2014)、内生长法(ingrowth coring methods) (Xiao et al., 2010; Liu et al., 2019)、微根管法(minirhizotron methods) (Trumbore and Gaudinski, 2003; Iversen et al., 2008; Bai et al., 2010)、碳氮平衡法(C and N budget methods) (Vogt et al., 1998)及同位素14C和13C法(Luo, 2003; Pritchard and Strand, 2008).其中, 根钻法是应用最早的一种方法(Pritchard and Strand, 2008; Xiao et al., 2008), 也是研究根系生物量、生产力和周转率最常用的方法(吴伊波等, 2014).该方法的优点是可相对容易和快速地估计根产量, 且能同时得到较准确的根系生物量数据(肖春旺等, 2017), 在一些需要具体生物量或生产力的研究中有明显优势.Brunner等(2013)对比分析了测量细根周转率的不同方法, 发现在一些浅层土壤及沙石含量较多的土壤和陡峭的斜坡中, 使用根钻法得到的结果更准确且更具可比性.但根钻法需要连续多次测量, 工作量大, 费时费力, 而且取样时可能会错过根系生物量的高峰或低谷(肖春旺等, 2017).由于根钻法计算时要取差值, 有研究认为该方法只适用于细根生物量波动很大的植物根系(Brunner et al., 2013). ...
Measuring fine root turnover in forest ecosystems 1 2005
... 对森林生态系统细根周转率的测量方法多种多样, 过去几十年里产生了许多估算陆地生态系统细根周转的方法(Vogt et al., 1998; Majdi et al., 2005).现今常用的有根钻法(soil coring methods) (Pritchard and Strand, 2008; Xiao et al., 2008; Zhou et al., 2014)、内生长法(ingrowth coring methods) (Xiao et al., 2010; Liu et al., 2019)、微根管法(minirhizotron methods) (Trumbore and Gaudinski, 2003; Iversen et al., 2008; Bai et al., 2010)、碳氮平衡法(C and N budget methods) (Vogt et al., 1998)及同位素14C和13C法(Luo, 2003; Pritchard and Strand, 2008).其中, 根钻法是应用最早的一种方法(Pritchard and Strand, 2008; Xiao et al., 2008), 也是研究根系生物量、生产力和周转率最常用的方法(吴伊波等, 2014).该方法的优点是可相对容易和快速地估计根产量, 且能同时得到较准确的根系生物量数据(肖春旺等, 2017), 在一些需要具体生物量或生产力的研究中有明显优势.Brunner等(2013)对比分析了测量细根周转率的不同方法, 发现在一些浅层土壤及沙石含量较多的土壤和陡峭的斜坡中, 使用根钻法得到的结果更准确且更具可比性.但根钻法需要连续多次测量, 工作量大, 费时费力, 而且取样时可能会错过根系生物量的高峰或低谷(肖春旺等, 2017).由于根钻法计算时要取差值, 有研究认为该方法只适用于细根生物量波动很大的植物根系(Brunner et al., 2013). ...
Impacts of fine root turnover on forest NPP and soil C sequestration potential 1 2003
Can you believe what you see? Reconciling minirhizotron and isotopically derived estimates of fine root longevity 3 2008
... 对森林生态系统细根周转率的测量方法多种多样, 过去几十年里产生了许多估算陆地生态系统细根周转的方法(Vogt et al., 1998; Majdi et al., 2005).现今常用的有根钻法(soil coring methods) (Pritchard and Strand, 2008; Xiao et al., 2008; Zhou et al., 2014)、内生长法(ingrowth coring methods) (Xiao et al., 2010; Liu et al., 2019)、微根管法(minirhizotron methods) (Trumbore and Gaudinski, 2003; Iversen et al., 2008; Bai et al., 2010)、碳氮平衡法(C and N budget methods) (Vogt et al., 1998)及同位素14C和13C法(Luo, 2003; Pritchard and Strand, 2008).其中, 根钻法是应用最早的一种方法(Pritchard and Strand, 2008; Xiao et al., 2008), 也是研究根系生物量、生产力和周转率最常用的方法(吴伊波等, 2014).该方法的优点是可相对容易和快速地估计根产量, 且能同时得到较准确的根系生物量数据(肖春旺等, 2017), 在一些需要具体生物量或生产力的研究中有明显优势.Brunner等(2013)对比分析了测量细根周转率的不同方法, 发现在一些浅层土壤及沙石含量较多的土壤和陡峭的斜坡中, 使用根钻法得到的结果更准确且更具可比性.但根钻法需要连续多次测量, 工作量大, 费时费力, 而且取样时可能会错过根系生物量的高峰或低谷(肖春旺等, 2017).由于根钻法计算时要取差值, 有研究认为该方法只适用于细根生物量波动很大的植物根系(Brunner et al., 2013). ...
... ; Pritchard and Strand, 2008).其中, 根钻法是应用最早的一种方法(Pritchard and Strand, 2008; Xiao et al., 2008), 也是研究根系生物量、生产力和周转率最常用的方法(吴伊波等, 2014).该方法的优点是可相对容易和快速地估计根产量, 且能同时得到较准确的根系生物量数据(肖春旺等, 2017), 在一些需要具体生物量或生产力的研究中有明显优势.Brunner等(2013)对比分析了测量细根周转率的不同方法, 发现在一些浅层土壤及沙石含量较多的土壤和陡峭的斜坡中, 使用根钻法得到的结果更准确且更具可比性.但根钻法需要连续多次测量, 工作量大, 费时费力, 而且取样时可能会错过根系生物量的高峰或低谷(肖春旺等, 2017).由于根钻法计算时要取差值, 有研究认为该方法只适用于细根生物量波动很大的植物根系(Brunner et al., 2013). ...
... ).其中, 根钻法是应用最早的一种方法(Pritchard and Strand, 2008; Xiao et al., 2008), 也是研究根系生物量、生产力和周转率最常用的方法(吴伊波等, 2014).该方法的优点是可相对容易和快速地估计根产量, 且能同时得到较准确的根系生物量数据(肖春旺等, 2017), 在一些需要具体生物量或生产力的研究中有明显优势.Brunner等(2013)对比分析了测量细根周转率的不同方法, 发现在一些浅层土壤及沙石含量较多的土壤和陡峭的斜坡中, 使用根钻法得到的结果更准确且更具可比性.但根钻法需要连续多次测量, 工作量大, 费时费力, 而且取样时可能会错过根系生物量的高峰或低谷(肖春旺等, 2017).由于根钻法计算时要取差值, 有研究认为该方法只适用于细根生物量波动很大的植物根系(Brunner et al., 2013). ...
Global patterns in root decomposition: comparisons of climate and litter quality effects 2 2001
... 对森林生态系统细根周转率的测量方法多种多样, 过去几十年里产生了许多估算陆地生态系统细根周转的方法(Vogt et al., 1998; Majdi et al., 2005).现今常用的有根钻法(soil coring methods) (Pritchard and Strand, 2008; Xiao et al., 2008; Zhou et al., 2014)、内生长法(ingrowth coring methods) (Xiao et al., 2010; Liu et al., 2019)、微根管法(minirhizotron methods) (Trumbore and Gaudinski, 2003; Iversen et al., 2008; Bai et al., 2010)、碳氮平衡法(C and N budget methods) (Vogt et al., 1998)及同位素14C和13C法(Luo, 2003; Pritchard and Strand, 2008).其中, 根钻法是应用最早的一种方法(Pritchard and Strand, 2008; Xiao et al., 2008), 也是研究根系生物量、生产力和周转率最常用的方法(吴伊波等, 2014).该方法的优点是可相对容易和快速地估计根产量, 且能同时得到较准确的根系生物量数据(肖春旺等, 2017), 在一些需要具体生物量或生产力的研究中有明显优势.Brunner等(2013)对比分析了测量细根周转率的不同方法, 发现在一些浅层土壤及沙石含量较多的土壤和陡峭的斜坡中, 使用根钻法得到的结果更准确且更具可比性.但根钻法需要连续多次测量, 工作量大, 费时费力, 而且取样时可能会错过根系生物量的高峰或低谷(肖春旺等, 2017).由于根钻法计算时要取差值, 有研究认为该方法只适用于细根生物量波动很大的植物根系(Brunner et al., 2013). ...
Analysis of some direct and indirect methods for estimating root biomass and production of forests at an ecosystem level 3 1998
... 对森林生态系统细根周转率的测量方法多种多样, 过去几十年里产生了许多估算陆地生态系统细根周转的方法(Vogt et al., 1998; Majdi et al., 2005).现今常用的有根钻法(soil coring methods) (Pritchard and Strand, 2008; Xiao et al., 2008; Zhou et al., 2014)、内生长法(ingrowth coring methods) (Xiao et al., 2010; Liu et al., 2019)、微根管法(minirhizotron methods) (Trumbore and Gaudinski, 2003; Iversen et al., 2008; Bai et al., 2010)、碳氮平衡法(C and N budget methods) (Vogt et al., 1998)及同位素14C和13C法(Luo, 2003; Pritchard and Strand, 2008).其中, 根钻法是应用最早的一种方法(Pritchard and Strand, 2008; Xiao et al., 2008), 也是研究根系生物量、生产力和周转率最常用的方法(吴伊波等, 2014).该方法的优点是可相对容易和快速地估计根产量, 且能同时得到较准确的根系生物量数据(肖春旺等, 2017), 在一些需要具体生物量或生产力的研究中有明显优势.Brunner等(2013)对比分析了测量细根周转率的不同方法, 发现在一些浅层土壤及沙石含量较多的土壤和陡峭的斜坡中, 使用根钻法得到的结果更准确且更具可比性.但根钻法需要连续多次测量, 工作量大, 费时费力, 而且取样时可能会错过根系生物量的高峰或低谷(肖春旺等, 2017).由于根钻法计算时要取差值, 有研究认为该方法只适用于细根生物量波动很大的植物根系(Brunner et al., 2013). ...
... )、碳氮平衡法(C and N budget methods) (Vogt et al., 1998)及同位素14C和13C法(Luo, 2003; Pritchard and Strand, 2008).其中, 根钻法是应用最早的一种方法(Pritchard and Strand, 2008; Xiao et al., 2008), 也是研究根系生物量、生产力和周转率最常用的方法(吴伊波等, 2014).该方法的优点是可相对容易和快速地估计根产量, 且能同时得到较准确的根系生物量数据(肖春旺等, 2017), 在一些需要具体生物量或生产力的研究中有明显优势.Brunner等(2013)对比分析了测量细根周转率的不同方法, 发现在一些浅层土壤及沙石含量较多的土壤和陡峭的斜坡中, 使用根钻法得到的结果更准确且更具可比性.但根钻法需要连续多次测量, 工作量大, 费时费力, 而且取样时可能会错过根系生物量的高峰或低谷(肖春旺等, 2017).由于根钻法计算时要取差值, 有研究认为该方法只适用于细根生物量波动很大的植物根系(Brunner et al., 2013). ...
... 为避免不同方法测定的周转率差异, 本研究统一采用生物量法计算细根周转率(细根周转率(单位: a-1)=地下净初级生产力(BNPP)/细根生物量平均值(Bmean) (Aber et al., 1985), 其中, BNPP=细根生物量最大值-细根生物量最小值(Vogt et al., 1998)).若文献中细根周转率数值的计算方法与此方法一致, 则予以采用; 若文献中数据与本研究细根周转率的计算方法不同, 则要记录文献中提供的森林细根生物量的季节性变化数据, 提取年生物量的最大值以及最小值得出BNPP, 再根据上述公式计算出细根周转率. ...
Review of root dynamics in forest ecosystems grouped by climate, climatic forest type and species 1 1995
Variation patterns of fine root biomass, production and turnover in Chinese forests 2 2017
... 森林是陆地生态系统的主体, 也是陆地上最庞大、最复杂、多物种、多功能与多效益的生态系统, 在保障国土生态安全、改善生态环境、维持人与生物圈的生态平衡及维护生物多样性等方面起着不可或缺的作用.而研究不同气候带典型森林细根周转对揭示森林生态系统物质循环和能量流动机制有重要意义.目前, 针对森林生态系统细根周转的研究主要集中在某一生态系统季节变化以及环境因子方面(梅莉等, 2004; Wang et al., 2017; Cai et al., 2019), 在全球尺度上的森林生态系统细根周转研究较少. ...
Belowground carbon pools and dynamics in China’s warm temperate and sub-tropical deciduous forests 2 2010
... 对森林生态系统细根周转率的测量方法多种多样, 过去几十年里产生了许多估算陆地生态系统细根周转的方法(Vogt et al., 1998; Majdi et al., 2005).现今常用的有根钻法(soil coring methods) (Pritchard and Strand, 2008; Xiao et al., 2008; Zhou et al., 2014)、内生长法(ingrowth coring methods) (Xiao et al., 2010; Liu et al., 2019)、微根管法(minirhizotron methods) (Trumbore and Gaudinski, 2003; Iversen et al., 2008; Bai et al., 2010)、碳氮平衡法(C and N budget methods) (Vogt et al., 1998)及同位素14C和13C法(Luo, 2003; Pritchard and Strand, 2008).其中, 根钻法是应用最早的一种方法(Pritchard and Strand, 2008; Xiao et al., 2008), 也是研究根系生物量、生产力和周转率最常用的方法(吴伊波等, 2014).该方法的优点是可相对容易和快速地估计根产量, 且能同时得到较准确的根系生物量数据(肖春旺等, 2017), 在一些需要具体生物量或生产力的研究中有明显优势.Brunner等(2013)对比分析了测量细根周转率的不同方法, 发现在一些浅层土壤及沙石含量较多的土壤和陡峭的斜坡中, 使用根钻法得到的结果更准确且更具可比性.但根钻法需要连续多次测量, 工作量大, 费时费力, 而且取样时可能会错过根系生物量的高峰或低谷(肖春旺等, 2017).由于根钻法计算时要取差值, 有研究认为该方法只适用于细根生物量波动很大的植物根系(Brunner et al., 2013). ...
Fine root dynamics and turnover rate in an Asia white birch forest of Donglingshan Mountain, China 2 2008
... 对森林生态系统细根周转率的测量方法多种多样, 过去几十年里产生了许多估算陆地生态系统细根周转的方法(Vogt et al., 1998; Majdi et al., 2005).现今常用的有根钻法(soil coring methods) (Pritchard and Strand, 2008; Xiao et al., 2008; Zhou et al., 2014)、内生长法(ingrowth coring methods) (Xiao et al., 2010; Liu et al., 2019)、微根管法(minirhizotron methods) (Trumbore and Gaudinski, 2003; Iversen et al., 2008; Bai et al., 2010)、碳氮平衡法(C and N budget methods) (Vogt et al., 1998)及同位素14C和13C法(Luo, 2003; Pritchard and Strand, 2008).其中, 根钻法是应用最早的一种方法(Pritchard and Strand, 2008; Xiao et al., 2008), 也是研究根系生物量、生产力和周转率最常用的方法(吴伊波等, 2014).该方法的优点是可相对容易和快速地估计根产量, 且能同时得到较准确的根系生物量数据(肖春旺等, 2017), 在一些需要具体生物量或生产力的研究中有明显优势.Brunner等(2013)对比分析了测量细根周转率的不同方法, 发现在一些浅层土壤及沙石含量较多的土壤和陡峭的斜坡中, 使用根钻法得到的结果更准确且更具可比性.但根钻法需要连续多次测量, 工作量大, 费时费力, 而且取样时可能会错过根系生物量的高峰或低谷(肖春旺等, 2017).由于根钻法计算时要取差值, 有研究认为该方法只适用于细根生物量波动很大的植物根系(Brunner et al., 2013). ...
Fine root biomass, production, turnover rates, and nutrient contents in boreal forest ecosystems in relation to species, climate, fertility, and stand age: literature review and meta-analyses 2 2010
Fine root and litterfall dynamics of three Korean pine (Pinus koraiensis) forests along an altitudinal gradient 1 2014
... 对森林生态系统细根周转率的测量方法多种多样, 过去几十年里产生了许多估算陆地生态系统细根周转的方法(Vogt et al., 1998; Majdi et al., 2005).现今常用的有根钻法(soil coring methods) (Pritchard and Strand, 2008; Xiao et al., 2008; Zhou et al., 2014)、内生长法(ingrowth coring methods) (Xiao et al., 2010; Liu et al., 2019)、微根管法(minirhizotron methods) (Trumbore and Gaudinski, 2003; Iversen et al., 2008; Bai et al., 2010)、碳氮平衡法(C and N budget methods) (Vogt et al., 1998)及同位素14C和13C法(Luo, 2003; Pritchard and Strand, 2008).其中, 根钻法是应用最早的一种方法(Pritchard and Strand, 2008; Xiao et al., 2008), 也是研究根系生物量、生产力和周转率最常用的方法(吴伊波等, 2014).该方法的优点是可相对容易和快速地估计根产量, 且能同时得到较准确的根系生物量数据(肖春旺等, 2017), 在一些需要具体生物量或生产力的研究中有明显优势.Brunner等(2013)对比分析了测量细根周转率的不同方法, 发现在一些浅层土壤及沙石含量较多的土壤和陡峭的斜坡中, 使用根钻法得到的结果更准确且更具可比性.但根钻法需要连续多次测量, 工作量大, 费时费力, 而且取样时可能会错过根系生物量的高峰或低谷(肖春旺等, 2017).由于根钻法计算时要取差值, 有研究认为该方法只适用于细根生物量波动很大的植物根系(Brunner et al., 2013). ...
 ,1,*
,1,*
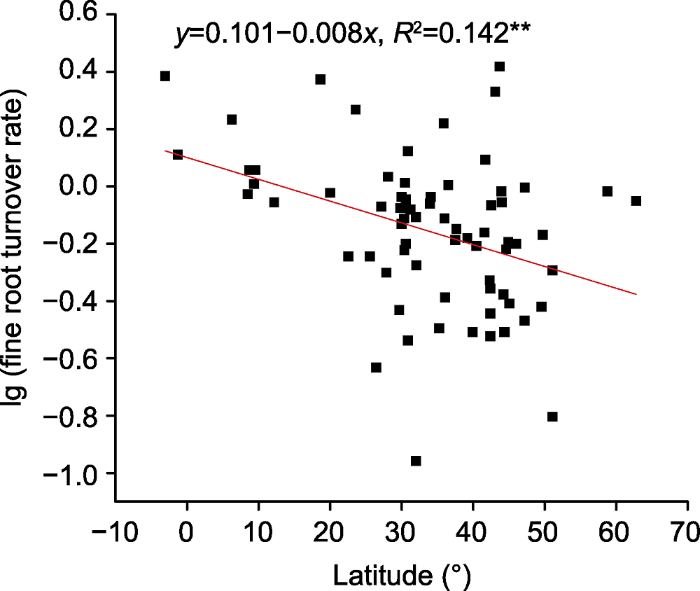 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT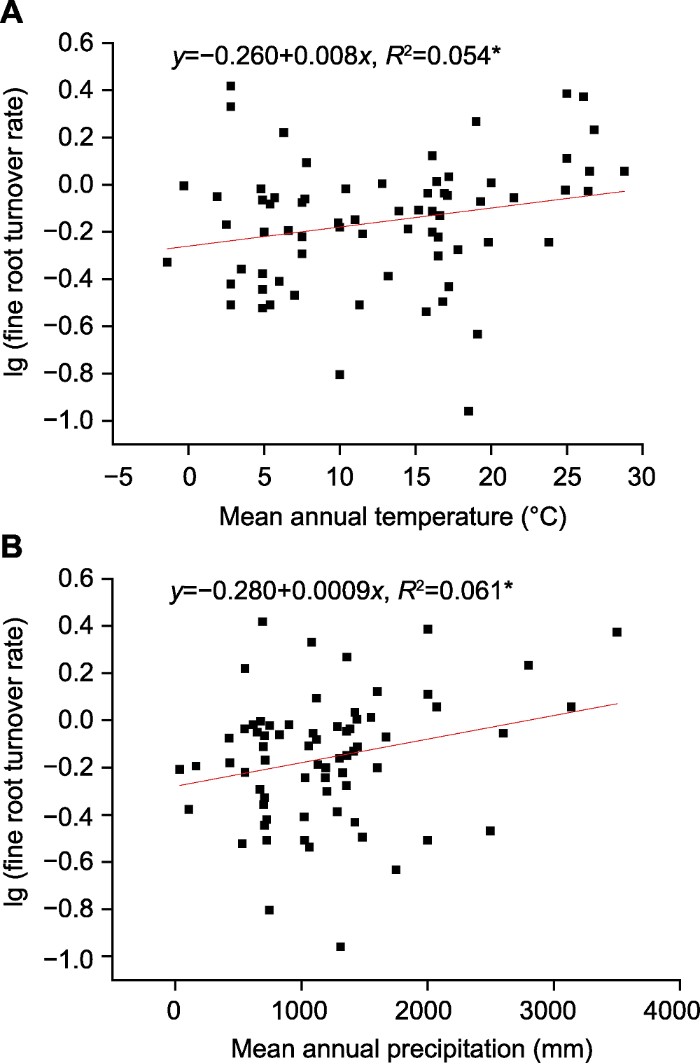 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT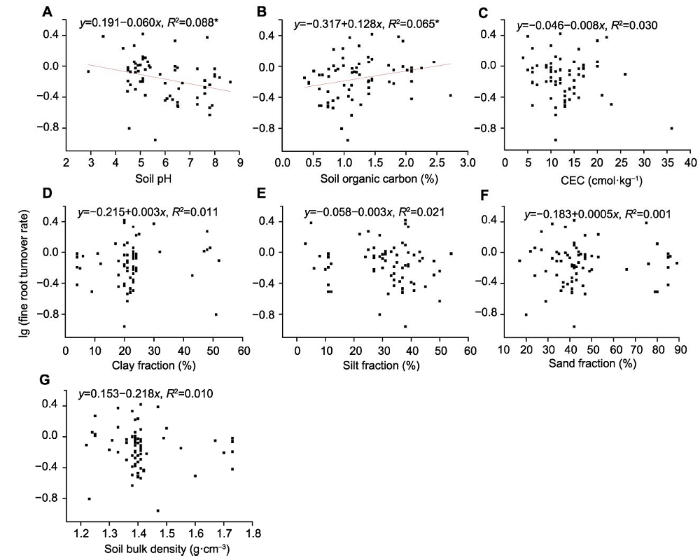 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}