Influence Mechanisms of Nitric Oxide on Nodulation and Nitrogen Fixation in Legumes
Weiqin Zhang1, Hang Zou2,3, Nina Zhang1, Xueyuan Lin1, Juan Chen,1,2,*1State Key Laboratory of Soil Erosion and Dryland Farming on the Loess Plateau, Northwest A&F University, Yangling 712100, China 2State Key Laboratory of Crop Stress Biology in Arid Areas, College of Life Sciences, Northwest A&F University, Yangling 712100, China 3Shaanxi Key Laboratory of Agricultural and Environmental Microbiology, Yangling 712100, China
Abstract Legume-rhizobium symbiosis is genetically co-regulated by the genes of both partners. The symbiosis process involves the formation of special nodule structure where the inert nitrogen (N2) from the atmosphere is converted into ammonia nitrogen that can be directly used by plants. Nodulation and nitrogen fixation are affected by many factors. As a free radical reactive gas signaling molecule, nitric oxide (NO) participates in the regulation of many plant growth and development processes, such as respiration, photomorphogenesis, seed germination, tissue and organ development, aging, and response to various biotic and abiotic stresses. In the legumes, it has been found that NO not only affects the establishment of the symbiotic relationship between the host and the bacteria, but also is involved in regulating the fixation of nitrogen by the rhizobia and increases the efficiency of nitrogen nutrition utilization. Here we review the mechanism of NO regulating nodule formation and symbiotic nitrogen fixation in legume-rhizobium symbiosis system, including the production and degradation of NO in legumes and rhizobia and its effect on nodulation, symbiotic nitrogen fixation and their response to environmental stress. We discuss the prospects and challenges of studying NO signaling molecule in symbiotic nitrogen fixation system of legume-rhizobium. Keywords:nitric oxide (NO);nodule;symbiotic nitrogen fixation;hemoglobins (Hbs)
PDF (1365KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 张卫勤, 邹杭, 张妮娜, 林雪媛, 陈娟. 一氧化氮对豆科植物结瘤及固氮的影响机制. 植物学报, 2020, 55(5): 623-633 doi:10.11983/CBB20034 Zhang Weiqin, Zou Hang, Zhang Nina, Lin Xueyuan, Chen Juan. Influence Mechanisms of Nitric Oxide on Nodulation and Nitrogen Fixation in Legumes. Chinese Bulletin of Botany, 2020, 55(5): 623-633 doi:10.11983/CBB20034
图中包含上下2部分, 分别对应植物和共生体NO的产生与降解。其中, 七角星图示代表氧化途径, 椭圆形图示代表还原途径。实线表示已有研究证实, 虚线表示还有待考证。ETC: 线粒体电子传递链; GSNOR: 亚硝基谷胱甘肽还原酶; Hmp: 黄素血红蛋白; Lb: 豆血红蛋白; NnrS: 含血红素和铜的膜蛋白; Nor: NO还原酶; NOS: NO合酶; ns-Hb: 非共生血红蛋白; NR: 硝酸还原酶; PAOX: 多胺氧化酶; sd-Hb: 单域血红蛋白; Tr-Hb: 截短血红蛋白; TrxR: 硫氧还蛋白还原酶; XOR: 黄嘌呤氧化还原酶 Figure 1The schematic diagram of NO production and degradation in the symbiotic system (modified from Hichri et al., 2016a)
The figure contains the upper and lower parts, which correspond to the production and degradation of NO from plants and symbiotes, respectively. The seven horns star diagrams refer to the oxidation pathway and the oval diagrams refer to the reduction pathway. The lines indicate that studies have been confirmed, and the dashed lines indicate that it is yet to be studied. ETC: Mitochondrial electron transport chain; GSNOR: S-nitrosoglutathione reductase; Hmp: Flavin hemoglobin; Lb: Leghemoglobin; NnrS: Haem- and copper-con- taining membrane protein; Nor: NO reductase; NOS: NO synthase; ns-Hb: Nonsymbiotic hemoglobin; NR: Nitrate reductase; PAOX: Polyamine oxidase; sd-Hb: Single domain hemoglobin; Tr-Hb: Truncated hemoglobin; TrxR: Thioredoxin reduction enzymes; XOR: Xanthine oxidoreductase
Cueto等(1996)在白羽扇豆(Lupinus albus)的根和根瘤中鉴定到NO合酶类似酶(nitric oxide synthase-like enzymatic, NOS-like)。Baudouin等(2006)认为这种酶可能就是根瘤菌侵染细胞时产生NO的主要原因。此外, 有研究表明, NOS-like和多胺氧化酶(polyamine oxidase, PAOX)均可在正常条件下介导成熟根瘤中氧化性NO产生(Mur et al., 2013; Hichri et al., 2015)。上述结果表明, 植物和根瘤菌中产生NO的过程并非完全各自独立, 而是通过相关途径或转导系统相互联系。例如, NR、ETC和NOS是植物及其共生菌中NO产生的共同来源(图1)。
在根瘤菌侵染豆科植物的每个阶段(包括接种4小时后、根毛卷曲、侵染线的产生、根瘤原基的形成和根瘤成熟)都伴有NO的产生(Hichri et al., 2015), 即从豆科植物与根瘤菌的早期共生到根瘤成熟与衰老, NO的产生贯穿始终(Meilhoc et al., 2011)。尽管目前对于NO的产生已经进行了许多研究, 但由于气体分子本身的性质以及与其它体系反应的复杂性, 使人们对植物中NO产生机制的研究受限, 因此关于豆科植物与根瘤菌共生的各个阶段中NO产生的机理还有待深入探索。
1.2 共生体系内NO的降解
豆科植物根瘤中包含一种与其它蛋白性质和功能不同的血红蛋白(hemoglobins, Hbs), 能够清除机体产生或累积的NO (Gupta et al., 2011)。基于其序列同源性和对氧的亲和力, 可将植物体内的Hbs划分成3类: 非共生血红蛋白(non-symbiotic hemoglobins, ns-Hbs, 1类)、豆血红蛋白(leghemoglobins, Lbs, 2类)和截短血红蛋白(truncated hemoglobin, Tr-Hbs, 3类) (Gupta et al., 2011; Hill, 2012)。Lbs作为高等植物中发现的第1种豆血红蛋白, 在豆科植物中主要负责将根瘤中的O2运输至根瘤菌(Trevaskis et al., 1997)。而Hbs对O2和NO的亲和力非常强, 能够有效清除O2和NO并将其转化为硝酸盐。有研究表明, Hbs能够维持植物细胞在缺氧状态下的氧化还原能力和能量状态(Igamberdiev and Hill, 2009)。除植物体蛋白外, 根瘤菌内也含有3类血红蛋白: 黄素血红蛋白(flavohaemoglobins, fHbs/Hmp)、单域血红蛋白(single-domain haemoglobins, sd-Hbs)和截短血红蛋白(Sánchez et al., 2011)。Hmp是蒺藜苜蓿(Medicago truncatula)中主要的NO清除剂, 而NO可诱导hmp基因表达以影响自身生成量(Meilhoc et al., 2011; Cam et al., 2012) (图1)。另有研究表明, 在蒺藜苜蓿hmp突变的根瘤中, NO水平显著高于野生型, 且固氮效率降低和根瘤衰老提前(Cam et al., 2012; Hichri et al., 2016a)。因此, 植物中的Hbs与菌内的Hmp对于维持共生体NO水平至关重要。
一方面, NO抑制固氮和C、N代谢; 另一方面, NO调控细胞氧化还原和保持低氧水平下的能量状态。带+的细线表示NO的活化、诱导和保持效果; 带有-的细线表示NO的抑制作用。椭圆形粗线箭头表示NO主要的代谢途径。爆炸型图示指来自植物和菌共生体的酶, 闪电型图示表示根瘤菌内的基因。ACO: 乌头酸; CS: 柠檬酸合酶; Gln: 谷氨酰胺; Glu: 谷氨酸; GS: 谷氨酰胺合成酶; GSH: 谷胱甘肽; GSHS: 谷胱甘肽合成酶; GSNO: S-亚硝基谷胱甘肽; Hb: 血红蛋白; IDH: 异柠檬酸脱氢酶; MDH: 苹果酸脱氢酶; NH4+: 铵根离子; Nif: 固氮酶; SDH: 琥珀酸脱氢酶; γ-EC: γ-谷氨酰半胱氨酸; γ-ECS: γ-谷氨酰半胱氨酸合成酶 Figure 2Schematic diagram of the role of NO in symbiotic nitrogen fixation (modified from Boscari et al., 2013; Hichri et al., 2015, 2016b)
On the one hand, NO inhibits nitrogen fixation and carbon and nitrogen metabolism; on the other hand, it regulates cellular redox status and maintains the energy state under low oxygen levels. A thin line with + indicate the activation, induction, and retention effects of NO; a thin line with - indicate the inhibition of NO. The oval thick line arrows indicate the main metabolic pathways of NO. Explosive type diagrams refer to enzymes from plants and bacterial symbionts, and lightning type diagram represents genes within rhizobium. ACO: Aconitic acid; CS: Citrate synthase; Gln: Glutamine; Glu: Glutamic acid; GS: Glutamine synthetase; GSH: Glutathione; GSHS: Glutathione synthetase; GSNO: S-nitrosoglu- tathione; Hb: Hemoglobin; IDH: Isocitrate dehydrogenase; MDH: Malate dehydrogenase; NH4+: Ammonium ion; Nif: Nitrogenase; SDH: Succinate dehydrogenase; γ-EC: γ-glu- tamylcysteine; γ-ECS: γ-glutamyl cysteine synthetase
2.3 NO与激素相互作用影响结瘤
脱落酸(abscisic acid, ABA)是植物适应环境胁迫的关键激素(Yoshida et al., 2015), 可通过抑制根毛变形及侵染线的形成影响豆科植物结瘤(Suzuki et al., 2004; Ding et al., 2008)。Tominaga等(2010)在百脉根中发现, ABA不仅可调节根瘤的固氮酶活性, 还可通过影响结瘤过程中NO的产生来影响结瘤。在根瘤形成过程中, NO通过酪氨酸(tyrosine, Tyr)硝化调控ABA信号传递, 而ABA又在信号转导过程中决定NO的产生与积累(Castillo et al., 2015)。Tyr硝化是NO介导的翻译后修饰, 其特征在于将-NO2添加到Tyr残基芳环的等价邻位C上, 产生3-硝基酪氨酸(3-nitrotyrosine, 3-NT), 并由此改变蛋白质构象(Radi, 2004)。作为硝化分子的前体, NO通过Tyr硝化改变细胞溶质谷氨酰胺合成酶(glutamine synthetase 1, GS1)和Lb等关键结瘤蛋白的活性(Melo et al., 2011; Navascués et al., 2012; Li et al., 2014)。豆科植物结瘤时Lb发生Tyr硝化, 可保护共生体免受过氧亚硝酸盐毒害, 因此3-NT的形成既是硝基氧化应激的标志, 也是功能性结瘤活跃代谢的结果(Sainz et al., 2015)。此外, NOS抑制剂N-硝基-L-精氨酸甲酯(NG-nitro- L-arginine methyl ester, L-NAME)和cPTIO处理可减弱豌豆(Pisum sativum)表皮由ABA诱导的气孔关闭(张绪成等, 2005)。
ArjonaD, Wikstr?mM, ?delrothP ( 2015). Nitric oxide is a potent inhibitor of the cbb3-type heme-copper oxidases FEBS Lett 589, 1214-1218. DOI:10.1016/j.febslet.2015.03.033URLPMID:25862499 [本文引用: 1] C-type heme-copper oxidases terminate the respiratory chain in many pathogenic bacteria, and will encounter elevated concentrations of NO produced by the immune defense of the host. Thus, a decreased sensitivity to NO in C-type oxidases would increase the survival of these pathogens. Here we have compared the inhibitory effect of NO in C-type oxidases to that in the mitochondrial A-type. We show that O2-reduction in both the Rhodobacter sphaeroides and Vibrio cholerae C-type oxidases is strongly and reversibly inhibited by submicromolar NO, with an inhibition pattern similar to the A-type. Thus, NO tolerance in pathogens with a C-type terminal oxidase has to rely mainly on other mechanisms.
BartnikasTB, WangYS, BoboT, VeseloA, ScholesCP, ShapleighJP ( 2002). Characterization of a member of the NnrR regulon in Rhodobacter sphaeroides 2.4.3 encoding a haem-copper protein: the GenBank accession number for nnrS is U62403 Microbiology 148, 825-833. DOI:10.1099/00221287-148-3-825URLPMID:11882718 [本文引用: 1] Upstream of the nor and nnrR cluster in Rhodobacter sphaeroides 2.4.3 is a previously uncharacterized gene that has been designated nnrS. nnrS is only expressed when 2.4.3 is grown under denitrifying conditions. Expression of nnrS is dependent on the transcriptional regulator NnrR, which also regulates expression of genes required for the reduction of nitrite to nitrous oxide, including nirK and nor. Deletion analysis indicated the sequence 5'-TTGCG(N4)CACAA-3', which is similar to sequences found in nirK and nor, is required for nnrS expression. Mutation of this sequence to the consensus Fnr-binding sequence by changing two bases in each half site caused nnrS expression to become nitrate independent. Inactivation of nnrS did not affect nitric oxide metabolism, nor did it affect expression of any of the genes involved in nitric oxide metabolism. However, taxis towards nitrate and nitrite was affected by nnrS inactivation. Purification of a histidine-tagged NnrS demonstrated that NnrS is a haem- and copper-containing membrane protein. Genes encoding putative orthologues of NnrS are sometimes but not always found in bacteria encoding nitrite and/or nitric oxide reductase.
BaudouinE, PieuchotL, EnglerG, PaulyN, PuppoA ( 2006). Nitric oxide is formed in Medicago truncatula-Sinorhizobium meliloti functional nodules Mol Plant Microbe Interact 19, 970-975. DOI:10.1094/MPMI-19-0970URLPMID:16941901 [本文引用: 1] Nitric oxide (NO) has recently gained interest as a major signaling molecule during plant development and response to environmental cues. Its role is particularly crucial for plant-pathogen interactions, during which it participates in the control of plant defense response and resistance. Indication for the presence of NO during symbiotic interactions has also been reported. Here, we defined when and where NO is produced during Medicago truncatula-Sinorhizobium meliloti symbiosis. Using the NO-specific fluorescent probe 4,5-diaminofluorescein diacetate, NO production was detected by confocal microscopy in functional nodules. NO production was localized in the bacteroid-containing cells of the nodule fixation zone. The infection of Medicago roots with bacterial strains impaired in nitrogenase or nitrite reductase activities lead to the formation of nodules with an unaffected NO level, indicating that neither nitrogen fixation nor denitrification pathways are required for NO production. On the other hand, the NO synthase inhibitor N-methyl-L-arginine impaired NO detection, suggesting that a NO synthase may participate to NO production in nodules. These data indicate that a NO production occurs in functional nodules. The location of such a production in fully metabolically active cells raises the hypothesis of a new function for NO during this interaction unrelated to defense and cell-death activation.
BergerA, BrouquisseR, PathakPK, HichriI, SinghI, BhatiaS, BoscariA, IgamberdievAU, GuptaKJ ( 2018). Pathways of nitric oxide metabolism and operation of phytoglobins in legume nodules: missing links and future directions Plant Cell Environ 41, 2057-2068. [本文引用: 1]
BethkePC, BadgerMR, JonesRL ( 2004). Apoplastic synthesis of nitric oxide by plant tissues Plant Cell 16, 332-341. DOI:10.1105/tpc.017822URLPMID:14742874 [本文引用: 1] Nitric oxide (NO) is an important signaling molecule in animals and plants. In mammals, NO is produced from Arg by the enzyme NO synthase. In plants, NO synthesis from Arg using an NO synthase-type enzyme and from nitrite using nitrate reductase has been demonstrated previously. The data presented in this report strongly support the hypothesis that plant tissues also synthesize NO via the nonenzymatic reduction of apoplastic nitrite. As measured by mass spectrometry or an NO-reactive fluorescent probe, Hordeum vulgare (barley) aleurone layers produce NO rapidly when nitrite is added to the medium in which they are incubated. NO production requires an acid apoplast and is accompanied by a loss of nitrite from the medium. Phenolic compounds in the medium can increase the rate of NO production. The possible significance of apoplastic NO production for germinating grain and for plant roots is discussed.
BlanquetP, SilvaL, CatriceO, BruandC, CarvalhoH, MeilhocE ( 2015). Sinorhizobium meliloti controls nitric oxide-mediated post-translational modification of a Medicago truncatula nodule protein Mol Plant Microbe Interact 28, 1353-1363. DOI:10.1094/MPMI-05-15-0118-RURLPMID:26422404 [本文引用: 6] Nitric oxide (NO) is involved in various plant-microbe interactions. In the symbiosis between soil bacterium Sinorhizobium meliloti and model legume Medicago truncatula, NO is required for an optimal establishment of the interaction but is also a signal for nodule senescence. Little is known about the molecular mechanisms responsible for NO effects in the legume-rhizobium interaction. Here, we investigate the contribution of the bacterial NO response to the modulation of a plant protein post-translational modification in nitrogen-fixing nodules. We made use of different bacterial mutants to finely modulate NO levels inside M. truncatula root nodules and to examine the consequence on tyrosine nitration of the plant glutamine synthetase, a protein responsible for assimilation of the ammonia released by nitrogen fixation. Our results reveal that S. meliloti possesses several proteins that limit inactivation of plant enzyme activity via NO-mediated post-translational modifications. This is the first demonstration that rhizobia can impact the course of nitrogen fixation by modulating the activity of a plant protein.
BoscariA, del GiudiceJ, FerrariniA, VenturiniL, ZaffiniAL, DelledonneM, PuppoA ( 2013). Expression dynamics of the Medicago truncatula transcriptome during the symbiotic interaction with Sinorhizobium meliloti: which role for nitric oxide Plant Physiol 161, 425-439. DOI:10.1104/pp.112.208538URLPMID:23136381 [本文引用: 8] Medicago truncatula is one of the most studied model plants. Nevertheless, the genome of this legume remains incompletely determined. We used RNA-Seq to characterize the transcriptome during the early organogenesis of the nodule and during its functioning. We detected 37,333 expressed transcription units; to our knowledge, 1,670 had never been described before and were functionally annotated. We identified 7,595 new transcribed regions, mostly corresponding to 5' and 3' untranslated region extensions and new exons associated with 5,264 previously annotated genes. We also inferred 23,165 putative transcript isoforms from 6,587 genes and measured the abundance of transcripts for each isoform, which suggests an important role for alternative splicing in the generation of proteome diversity in M. truncatula. Finally, we carried out a differential expression analysis, which provided a comprehensive view of transcriptional reprogramming during nodulation. In particular, depletion of nitric oxide in roots inoculated with Sinorhizobium meliloti greatly increased our understanding of the role of this reactive species in the optimal establishment of the symbiotic interaction, revealing differential patterns of expression for 2,030 genes and pointing to the inhibition of the expression of defense genes.
BreakspearA, LiuCW, RoyS, StaceyN, RogersC, TrickM, MorieriG, MysoreKS, WenJQ, OldroydGED, DownieJA, MurrayJD ( 2014). The root hair "infectome" of Medicago truncatula uncovers changes in cell cycle genes and reveals a requirement for auxin signaling in rhizobial infection Plant Cell 26, 4680-4701. DOI:10.1105/tpc.114.133496URLPMID:25527707 [本文引用: 1] Nitrogen-fixing rhizobia colonize legume roots via plant-made intracellular infection threads. Genetics has identified some genes involved but has not provided sufficient detail to understand requirements for infection thread development. Therefore, we transcriptionally profiled Medicago truncatula root hairs prior to and during the initial stages of infection. This revealed changes in the responses to plant hormones, most notably auxin, strigolactone, gibberellic acid, and brassinosteroids. Several auxin responsive genes, including the ortholog of Arabidopsis thaliana Auxin Response Factor 16, were induced at infection sites and in nodule primordia, and mutation of ARF16a reduced rhizobial infection. Associated with the induction of auxin signaling genes, there was increased expression of cell cycle genes including an A-type cyclin and a subunit of the anaphase promoting complex. There was also induction of several chalcone O-methyltransferases involved in the synthesis of an inducer of Sinorhizobium meliloti nod genes, as well as a gene associated with Nod factor degradation, suggesting both positive and negative feedback loops that control Nod factor levels during rhizobial infection. We conclude that the onset of infection is associated with reactivation of the cell cycle as well as increased expression of genes required for hormone and flavonoid biosynthesis and that the regulation of auxin signaling is necessary for initiation of rhizobial infection threads.
Calvo-BegueriaL, RubioMC, MartínezJI, Pérez-Ron- toméC, DelgadoMJ, BedmarEJ, BecanaM ( 2018). Redefining nitric oxide production in legume nodules through complementary insights from electron paramagnetic resonance spectroscopy and specific fluorescent probes J Exp Bot 69, 3703-3714. DOI:10.1093/jxb/ery159URLPMID:29701804 [本文引用: 2] Nitric oxide (NO) is a signaling molecule with multiple functions in plants. Given its critical importance and reactivity as a gaseous free radical, we have examined NO production in legume nodules using electron paramagnetic resonance (EPR) spectroscopy and the specific fluorescent dye 4,5-diaminofluorescein diacetate. Also, in this context, we critically assess previous and current views of NO production and detection in nodules. EPR of intact nodules revealed that nitrosyl-leghemoglobin (Lb2+NO) was absent from bean or soybean nodules regardless of nitrate supply, but accumulated in soybean nodules treated with nitrate that were defective in nitrite or nitric oxide reductases or that were exposed to ambient temperature. Consequently, bacteroids are a major source of NO, denitrification enzymes are required for NO homeostasis, and Lb2+NO is not responsible for the inhibition of nitrogen fixation by nitrate. Further, we noted that Lb2+NO is artifactually generated in nodule extracts or in intact nodules not analyzed immediately after detachment. The fluorescent probe detected NO formation in bean and soybean nodule infected cells and in soybean nodule parenchyma. The NO signal was slightly decreased by inhibitors of nitrate reductase but not by those of nitric oxide synthase, which could indicate a minor contribution of plant nitrate reductase and supports the existence of nitrate- and arginine-independent pathways for NO production. Together, our data indicate that EPR and fluorometric methods are complementary to draw reliable conclusions about NO production in plants.
CamY, PierreO, BoncompagniE, HérouartD, MeilhocE, BruandC ( 2012). Nitric oxide (NO): a key player in the senescence of Medicago truncatula root nodules New Phytol 196, 548-560. DOI:10.1111/j.1469-8137.2012.04282.xURLPMID:22937888 [本文引用: 12] Nitric oxide (NO) is a signalling and defence molecule involved in diverse plant developmental processes, as well as in the plant response to pathogens. NO has also been detected at different steps of the symbiosis between legumes and rhizobia. NO is required for an optimal establishment of the Medicago truncatula-Sinorhizobium meliloti symbiotic interaction, but little is known about the role of NO in mature nodules. Here, we investigate the role of NO in the late steps of symbiosis. Genetic and pharmacological approaches were conducted to modulate the NO level inside root nodules, and their effects on nitrogen fixation and root nodule senescence were monitored. An increase in endogenous NO levels led to a decrease in nitrogen fixation and early nodule senescence, characterized by cytological modifications of the nodule structure and the early expression of a specific senescence marker. By contrast, a decrease in NO levels led to a delay in nodule senescence. Together, our results strongly suggest that NO is a signal in developmental as well as stress-induced nodule senescence. In addition, this work demonstrates the pivotal role of the bacterial NO detoxification response in the prevention of early nodule senescence, and hence the maintenance of efficient symbiosis.
CastilloMC, Lozano-JusteJ, González-GuzmánM, RodriguezL, RodriguezPL, LeónJ ( 2015). Inactivation of PYR/PYL/RCAR ABA receptors by tyrosine nitration may enable rapid inhibition of ABA signaling by nitric oxide in plants Sci Signal 8, ra89. DOI:10.1126/scisignal.aad5111URLPMID:26577921 [本文引用: 1] Pharmacological inhibitors against the PI3K-AKT-mTOR (phosphatidylinositol 3-kinase-AKT-mammalian target of rapamycin) pathway, a frequently deregulated signaling pathway in cancer, are clinically promising, but the development of drug resistance is a major limitation. We found that 4EBP1, the central inhibitor of cap-dependent translation, was a critical regulator of both prostate cancer initiation and maintenance downstream of mTOR signaling in a genetic mouse model. 4EBP1 abundance was distinctly different between the epithelial cell types of the normal prostate. Of tumor-prone prostate epithelial cell types, luminal epithelial cells exhibited the highest transcript and protein abundance of 4EBP1 and the lowest protein synthesis rates, which mediated resistance to both pharmacologic and genetic inhibition of the PI3K-AKT-mTOR signaling pathway. Decreasing total 4EBP1 abundance reversed resistance in drug-insensitive cells. Increased 4EBP1 abundance was a common feature in prostate cancer patients who had been treated with the PI3K pathway inhibitor BKM120; thus, 4EBP1 may be associated with drug resistance in human tumors. Our findings reveal a molecular program controlling cell type-specific 4EBP1 abundance coupled to the regulation of global protein synthesis rates that renders each epithelial cell type of the prostate uniquely sensitive or resistant to inhibitors of the PI3K-AKT-mTOR signaling pathway.
ChadhaN, MishraM, RajpalK, BajajR, ChoudharyDK, VarmaA ( 2015). An ecological role of fungal endophytes to ameliorate plants under biotic stress Arch Microbiol 197, 869-881. DOI:10.1007/s00203-015-1130-3URLPMID:26123239 [本文引用: 1] It is our consensus that plants survive and flourish in stressed ecosystems because of endosymbiotic organisms that have co-evolved and were essential for their adaptation to changing environments. Some of these microbial components are noncultivable and vertically transmitted from generation to generation. They represent a vast reservoir of heritable DNA that can enhance plant performance in changing environments and add genetic flexibility to adaptation of long-lived plants. If such endophytes can be identified that not only persist in progeny of novel hosts, but can confer benefits in mechanized, agricultural systems, they would be increasingly important in agricultural production and lead to a rapid and economical method of providing novel germplasms of native and crop plants. In the present review, authors advocate the deployment of fungal diversity and its role to overcome the biotic stress in plants. Endophytic fungal association with plants helps it to protect from various pathogen and pests and adapt to survive in harsh biotic and abiotic stress condition.
ChakiM, KovacsI, SpannaglM, LindermayrC ( 2014). Computational prediction of candidate proteins for S-nitrosylation in Arabidopsis thaliana PLoS One 9, e110232. DOI:10.1371/journal.pone.0110232URLPMID:25333472 [本文引用: 1] Nitric oxide (NO) is an important signaling molecule that regulates many physiological processes in plants. One of the most important regulatory mechanisms of NO is S-nitrosylation-the covalent attachment of NO to cysteine residues. Although the involvement of cysteine S-nitrosylation in the regulation of protein functions is well established, its substrate specificity remains unknown. Identification of candidates for S-nitrosylation and their target cysteine residues is fundamental for studying the molecular mechanisms and regulatory roles of S-nitrosylation in plants. Several experimental methods that are based on the biotin switch have been developed to identify target proteins for S-nitrosylation. However, these methods have their limits. Thus, computational methods are attracting considerable attention for the identification of modification sites in proteins. Using GPS-SNO version 1.0, a recently developed S-nitrosylation site-prediction program, a set of 16,610 candidate proteins for S-nitrosylation containing 31,900 S-nitrosylation sites was isolated from the entire Arabidopsis proteome using the medium threshold. In the compartments
CuetoM, Hernández-PereraO, MartínR, BenturaML, RodrigoJ, LamasS, GolvanoMP ( 1996). Presence of nitric oxide synthase activity in roots and nodules of Lupinus albus FEBS Lett 398, 159-164. DOI:10.1016/s0014-5793(96)01232-xURLPMID:8977098 [本文引用: 1] NO is a widespread messenger molecule in physiology. We were interested in investigating whether an NO-generating system could be present in plants. NO and L-[14C]citrulline were synthesized by roots and nodules of Lupinus albus in an L-arginine-dependent manner. L-[14C]Citrulline production was inhibited by N(G)-monomethyl-L-arginine, a nitric oxide synthase antagonist, in a competitive way. NADPH-diaphorase activity was localized in the vascular bundles in root and nodules, and also in the nodule infected zone. This staining was significantly reduced in the presence of N(G)-monomethyl-L-arginine. These results indicate the presence of a putative nitric oxide synthase in plants.
De BruijnFJ, RossbachS, BruanC, ParrishJR ( 2006). A highly conserved Sinorhizobium meliloti operon is induced microaerobically via the FixLJ system and by nitric oxide (NO) via NnrR Environ Microbiol 8, 1371-1381. DOI:10.1111/j.1462-2920.2006.01030.xURLPMID:16872401 [本文引用: 2] A previously generated collection of 11 Tn5-luxAB insertion mutants of Sinorhizobium meliloti harbouring lux reporter gene fusions induced under microaerobic (1% O2) conditions was further characterized and mapped on the sequenced S. meliloti genome. One highly induced gene fusion from this collection (loe-7) was found to be located in the intergenic region between sma1292, encoding a putative protease/collagenase, and a gene of unknown function (sma1294). The loe-7 fusion had been shown previously to be partially controlled by the oxygen sensor/regulator FixLJ system, but significant ( approximately 40%) Lux activity remained in a fixLJ mutant background. Therefore, a secondary Tn1721 mutagenesis of the loe-7 strain was carried out. Nine Tn1721 ('dark') insertions completely abolishing the Lux activity of the loe-7 fusion under microaerobic conditions were isolated. Surprisingly, five dark insertions mapped in denitrification genes [napA, napC, nirK--two insertions--and sma1245 encoding a NnrR-like transcriptional regulator controlling denitrification in response to nitric oxide (NO)]; Tn1721 insertions in the respiration genes fixG and fixP resulted in a reduced expression of the loe-7-lux fusion, and insertions in the regulatory genes fixJ and fixK1 resulted in low, but still detectable Lux activity. On the contrary, insertions in the norD or norQ genes resulted in constitutive Lux activity. In these mutant strains, NO would be expected to accumulate under microaerobic conditions. NO was found to be able to strongly induce the loe-7-luxAB fusion under microaerobic and aerobic conditions, but only in the presence of the functional nnrR-like gene (sma1245). These results suggest that NO, via the NnrR regulator, can serve as a signal molecule to induce the loe-7-luxAB fusion in concert with the FixLJ system.
DeanJV, HarperJE ( 1988). The conversion of nitrite to nitrogen oxide(s) by the constitutive NAD(P)H-nitrate reductase enzyme from soybean Plant Physiol 88, 389-395. DOI:10.1104/pp.88.2.389URLPMID:16666314 [本文引用: 1] A two-step purification protocol was used in an attempt to separate the constitutive NAD(P)H-nitrate reductase [NAD(P)H-NR, pH 6.5; EC 1.6.6.2] activity from the nitric oxide and nitrogen dioxide (NO((x))) evolution activity extracted from soybean (Glycine max [L.] Merr.) leaflets. Both of these activities were eluted with NADPH from Blue Sepharose columns loaded with extracts from either wild-type or LNR-5 and LNR-6 (lack constitutive NADH-NR [pH 6.5]) mutant soybean plants regardless of nutrient growth conditions. Fast protein liquid chromatography-anion exchange (Mono Q column) chromatography following Blue Sepharose affinity chromatography was also unable to separate the two activities. These data provide strong evidence that the constitutive NAD(P)H-NR (pH 6.5) in soybean is the enzyme responsible for NO((x)) formation. The Blue Sepharose-purified soybean enzyme has a pH optimum of 6.75, an apparent K(m) for nitrite of 0.49 millimolar, and an apparent K(m) for NADPH and NADH of 7.2 and 7.4 micromolar, respectively, for the NO((x)) evolution activity. In addition to NAD(P)H, reduced flavin mononucleotide (FMNH(2)) and reduced methyl viologen (MV) can serve as electron donors for NO((x)) evolution activity. The NADPH-, FMNH(2)-, and reduced MV-NO((x)) evolution activities were all inhibited by cyanide. The NADPH activity was also inhibited by p-hydroxymer-curibenzoate, whereas, the FMNH(2) and MV activities were relatively insensitive to inhibition. These data indicate that the terminal molybdenum-containing portion of the enzyme is involved in the reduction of nitrite to NO((x)). NADPH eluted both NR and NO((x)) evolution activities from Blue Sepharose columns loaded with extracts of either nitrate- or zero N-grown winged bean (Psophocarpus tetragonolobus [L.]), whereas NADH did not elute either type of activity. Winged bean appears to contain only one type of NR enzyme that is similar to the constitutive NAD(P)H-NR (pH 6.5) enzyme of soybean.
del GiudiceJ, CamY, DamianiI, Fung-ChatF, MeilhocE, BruandC, BrouquisseR, PuppoA, BoscariA ( 2011). Nitric oxide is required for an optimal establishment of the Medicago truncatula-Sinorhizobium meliloti symbiosis New Phytol 191, 405-417. DOI:10.1111/j.1469-8137.2011.03693.xURLPMID:21457261 [本文引用: 5] Nitric oxide (NO) is a gaseous molecule that participates in numerous plant signalling pathways. It is involved in plant responses to pathogens and development processes such as seed germination, flowering and stomatal closure. Using a permeable NO-specific fluorescent probe and a bacterial reporter strain expressing the lacZ gene under the control of a NO-responsive promoter, we detected NO production in the first steps, during infection threads growth, of the Medicago truncatula-Sinorhizobium meliloti symbiotic interaction. Nitric oxide was also detected, by confocal microscopy, in nodule primordia. Depletion of NO caused by cPTIO (2-(4-carboxyphenyl)-4,4,5,5-tetramethyl imidazoline-1-oxyl-3-oxide), an NO scavenger, resulted in a significant delay in nodule appearance. The overexpression of a bacterial hmp gene, encoding a flavohaemoglobin able to scavenge NO, under the control of a nodule-specific promoter (pENOD20) in transgenic roots, led to the same phenotype. The NO scavenging resulting from these approaches provoked the downregulation of plant genes involved in nodule development, such as MtCRE1 and MtCCS52A. Furthermore, an Hmp-overexpressing S. meliloti mutant strain was found to be less competitive than the wild type in the nodulation process. Taken together, these results indicate that NO is required for an optimal establishment of the M. truncatula-S. meliloti symbiotic interaction.
DesalvoMK, SunagawaS, VoolstraCR, MedinaM ( 2010). Transcriptomic responses to heat stress and bleaching in the Elkhorn coral Acropora palmata Mar Ecol Prog Ser 402, 97-113. DOI:10.3354/meps08372URL [本文引用: 1]
DingYL, KaloP, YendrekC, SunJ, LiangY, MarshJF, HarrisJM, OldroydGED ( 2008). Abscisic acid coordinates nod factor and cytokinin signaling during the regulation of nodulation in Medicago truncatula Plant Cell 20, 2681-2695. DOI:10.1105/tpc.108.061739URLPMID:18931020 [本文引用: 1] Nodulation is tightly regulated in legumes to ensure appropriate levels of nitrogen fixation without excessive depletion of carbon reserves. This balance is maintained by intimately linking nodulation and its regulation with plant hormones. It has previously been shown that ethylene and jasmonic acid (JA) are able to regulate nodulation and Nod factor signal transduction. Here, we characterize the nature of abscisic acid (ABA) regulation of nodulation. We show that application of ABA inhibits nodulation, bacterial infection, and nodulin gene expression in Medicago truncatula. ABA acts in a similar manner as JA and ethylene, regulating Nod factor signaling and affecting the nature of Nod factor-induced calcium spiking. However, this action is independent of the ethylene signal transduction pathway. We show that genetic inhibition of ABA signaling through the use of a dominant-negative allele of ABSCISIC ACID INSENSITIVE1 leads to a hypernodulation phenotype. In addition, we characterize a novel locus of M. truncatula, SENSITIVITY TO ABA, that dictates the sensitivity of the plant to ABA and, as such, impacts the regulation of nodulation. We show that ABA can suppress Nod factor signal transduction in the epidermis and can regulate cytokinin induction of the nodule primordium in the root cortex. Therefore, ABA is capable of coordinately regulating the diverse developmental pathways associated with nodule formation and can intimately dictate the nature of the plants' response to the symbiotic bacteria.
EscuredoPR, MinchinFR, GogorcenaY, Iturbe-Or- maetxeI, KlucasRV, BecanaM ( 1996). Involvement of activated oxygen in nitrate-induced senescence of pea root nodules Plant Physiol 110, 1187-1195. DOI:10.1104/pp.110.4.1187URLPMID:12226252 [本文引用: 1] The effect of short-term nitrate application (10 mM, 0-4 d) on nitrogenase (N2ase) activity, antioxidant defenses, and related parameters was investigated in pea (Pisum sativum L. cv Frilene) nodules. The response of nodules to nitrate comprised two stages. In the first stage (0-2 d), there were major decreases in N2ase activity and N2ase-linked respiration and concomitant increases in carbon cost of N2ase and oxygen diffusion resistance of nodules. There was no apparent oxidative damage, and the decline in N2ase activity was, to a certain extent, reversible. The second stage (>2 d) was typical of a senescent, essentially irreversible process. It was characterized by moderate increases in oxidized proteins and catalytic Fe and by major decreases in antioxidant enzymes and metabolites. The restriction in oxygen supply to bacteroids may explain the initial decline in N2ase activity. The decrease in antioxidant protection is not involved in this process and is not specifically caused by nitrate, since it also occurs with drought stress. However, comparison of nitrate- and drought-induced senescence shows an important difference: there is no lipid degradation or lipid peroxide accumulation with nitrate, indicating that lipid peroxidation is not necessarily involved in nodule senescence.
FerrariniA, De StefanoM, BaudouinE, PucciarielloC, PolverariA, PuppoA, DelledonneM ( 2008). Expression of Medicago truncatula genes responsive to nitric oxide in pathogenic and symbiotic conditions Mol Plant Microbe Interact 21, 781-790. URLPMID:18624641 [本文引用: 1]
GargN, Geetanjali , (2007). Symbiotic nitrogen fixation in legume nodules: process and signaling. A review. In: Lichtfouse E, Navarrete M, Debaeke P, Véronique S, Alberola C, eds. Sustainable Agriculture. Dordrecht: Springer. pp. 519-531. [本文引用: 1]
Gonzalez-RizzoS, CrespiM, FrugierF ( 2006). The Medicago truncatula CRE1 cytokinin receptor regulates lateral root development and early symbiotic interaction with Sinorhizobium meliloti Plant Cell 18, 2680-2693. DOI:10.1105/tpc.106.043778URLPMID:17028204 [本文引用: 1] Legumes develop different types of lateral organs from their primary root, lateral roots and nodules, the latter depending on a symbiotic interaction with Sinorhizobium meliloti. Phytohormones have been shown to function in the control of these organogeneses. However, related signaling pathways have not been identified in legumes. We cloned and characterized the expression of Medicago truncatula genes encoding members of cytokinin signaling pathways. RNA interference of the cytokinin receptor homolog Cytokinin Response1 (Mt CRE1) led to cytokinin-insensitive roots, which showed an increased number of lateral roots and a strong reduction in nodulation. Both the progression of S. meliloti infection and nodule primordia formation were affected. We also identified two cytokinin signaling response regulator genes, Mt RR1 and Mt RR4, which are induced early during the symbiotic interaction. Induction of these genes by S. meliloti infection is altered in mutants affected in the Nod factor signaling pathway; conversely, cytokinin regulation of the early nodulin Nodule Inception1 (Mt NIN) depends on Mt CRE1. Hence, cytokinin signaling mediated by a single receptor, Mt CRE1, leads to an opposite control of symbiotic nodule and lateral root organogenesis. Mt NIN, Mt RR1, and Mt RR4 define a common pathway activated during early S. meliloti interaction, allowing crosstalk between plant cytokinins and bacterial Nod factors signals.
GuptaKJ, HebelstrupKH, MurLAJ, IgamberdievAU ( 2011). Plant hemoglobins: important players at the crossroads between oxygen and nitric oxide FEBS Lett 585, 3843-3849. DOI:10.1016/j.febslet.2011.10.036URLPMID:22036787 [本文引用: 6] Plant hemoglobins constitute a diverse group of hemeproteins and evolutionarily belong to three different classes. Class 1 hemoglobins possess an extremely high affinity to oxygen and their main function consists in scavenging of nitric oxide (NO) at very low oxygen levels. Class 2 hemoglobins have a lower oxygen affinity and they facilitate oxygen supply to developing tissues. Symbiotic hemoglobins in nodules have mostly evolved from class 2 hemoglobins. Class 3 hemoglobins are truncated and represent a clade with a very low similarity to class 1 and 2 hemoglobins. They may regulate oxygen delivery at high O(2) concentrations. Depending on their physical properties, hemoglobins belong either to hexacoordinate non-symbiotic or pentacoordinate symbiotic groups. Plant hemoglobins are plausible targets for improving resistance to multiple stresses.
HawkinsTD, KruegerT, BeckerS, FisherPL, DavySK ( 2014). Differential nitric oxide synthesis and host apoptotic events correlate with bleaching susceptibility in reef corals Coral Reefs 33, 141-153. DOI:10.1007/s00338-013-1103-4URL [本文引用: 1]
HeroldS, PuppoA ( 2005). Oxyleghemoglobin scavenges nitrogen monoxide and peroxynitrite: a possible role in functioning nodules J Biol Inorg Chem 10, 935-945. DOI:10.1007/s00775-005-0046-9URLPMID:16267661 [本文引用: 1] It has been demonstrated that the NO* produced by nitric oxide synthase or by the reduction of nitrite by nitrate reductase plays an important role in plants' defense against microbial pathogens. The detection of nitrosyl Lb in nodules strongly suggests that NO* is also formed in functional nodules. Moreover, NO* may react with superoxide (which has been shown to be produced in nodules by various processes), leading to the formation of peroxynitrite. We have determined the second-order rate constants of the reactions of soybean oxyleghemoglobin with nitrogen monoxide and peroxynitrite. At pH 7.3 and 20 degrees C, the values are on the order of 10(8) and 10(4) M-1 s-1, respectively. In the presence of physiological amounts of CO2 (1.2 mM), the second-order rate constant of the reaction of oxyleghemoglobin peroxynitrite is even larger (10(5) M-1 s-1). The results presented here clearly show that oxyleghemoglobin is able to scavenge any NO* and peroxynitrite formed in functional nodules. This may help to stop NO* triggering a plant defense reaction.
HichriI, BoscariA, CastellaC, RovereM, PuppoA, BrouquisseR ( 2015). Nitric oxide: a multifaceted regulator of the nitrogen-fixing symbiosis J Exp Bot 66, 2877-2887. DOI:10.1093/jxb/erv051URLPMID:25732535 [本文引用: 9] The specific interaction between legumes and Rhizobium-type bacteria leads to the establishment of a symbiotic relationship characterized by the formation of new differentiated organs named nodules, which provide a niche for bacterial nitrogen (N2) fixation. In the nodules, bacteria differentiate into bacteroids with the ability to fix atmospheric N2 via nitrogenase activity. As nitrogenase is strongly inhibited by oxygen, N2 fixation is made possible by the microaerophilic conditions prevailing in the nodules. Increasing evidence has shown the presence of NO during symbiosis, from early interaction steps between the plant and the bacterial partners to N2-fixing and senescence steps in mature nodules. Both the plant and the bacterial partners participate in NO synthesis. NO was found to be required for the optimal establishment of the symbiotic interaction. Transcriptomic analysis at an early stage of the symbiosis showed that NO is potentially involved in the repression of plant defence reactions, favouring the establishment of the plant-microbe interaction. In mature nodules, NO was shown to inhibit N2 fixation, but it was also demonstrated to have a regulatory role in nitrogen metabolism, to play a beneficial metabolic function for the maintenance of the energy status under hypoxic conditions, and to trigger nodule senescence. The present review provides an overview of NO sources and multifaceted effects from the early steps of the interaction to the senescence of the nodule, and presents several approaches which appear to be particularly promising in deciphering the roles of NO in N2-fixing symbioses.
HichriI, BoscariA, MeilhocE, CataláM, BarrenoE, BruandC, LanfrancoL, BrouquisseR (2016a). Nitric oxide: a multitask player in plant-microorganism symbioses. In: Lamattina L, García-Mata C, eds. Gasotransmitters in Plants: The Rise of a New Paradigm in Cell Signaling. Cham: Springer. pp. 239-268. [本文引用: 3]
HichriI, MeilhocE, BoscariA, BruandC, FrendoP, BrouquisseR ( 2016b). Nitric oxide: jack-of-all-trades of the nitrogen-fixing symbiosis? Adv Bot Res 77, 193-218. [本文引用: 8]
HillRD ( 2012). Non-symbiotic haemoglobins-what's happening beyond nitric oxide scavenging AoB Plants2012, pls004. DOI:10.1093/aobpla/plaa028URLPMID:32765824 [本文引用: 1] The intraspecific variations of leaf structure and anatomy in rice leaves and their impacts on gas diffusion are still unknown. Researches about the tradeoff between structural compositions and intracellular chemical components within rice leaves are still lacking. The objectives of the present study were to investigate the varietal differences in leaf structure and leaf chemical compositions, and the tradeoff between leaf structural tissues and intracellular chemical components in rice leaves. Leaf structure, leaf anatomy, leaf chemical composition concentrations and gas exchange parameters were measured on eight Oryza sativa L. genotypes to investigate the intraspecific variations in leaf structure and leaf anatomy and their impacts on gas exchange parameters, and to study the tradeoff between leaf structural compositions (cell wall compounds) and intracellular chemical components (non-structural carbohydrates, nitrogen, chlorophyll). Leaf thickness increased with leaf mass per area (LMA), while leaf density did not correlate with LMA. Mesophyll cell surface area exposed to intercellular airspace (IAS) per leaf area, the surface area of chloroplasts exposed to IAS and cell wall thickness increased with LMA. Cell wall compounds accounted for 71.5 % of leaf dry mass, while mass-based nitrogen and chlorophyll concentrations decreased with LMA. Mesophyll conductance was negatively correlated with LMA and cell wall thickness. High LMA rice genotypes invest more leaf mass to cell wall and possess a low mesophyll conductance.
HorchaniF, PrévotM, BoscariA, EvangelistiE, MeilhocE, BruandC, RaymondP, BoncompagniE, Aschi- SmitiS, PuppoA, BrouquisseR ( 2011). Both plant and bacterial nitrate reductases contribute to nitric oxide production in Medicago truncatula nitrogen-fixing nodules Plant Physiol 15, 1023-1036. [本文引用: 9]
HuJL, HuangXH, ChenLC, SunXW, LuCM, ZhangLX, WangYC, ZuoJR ( 2015). Site-speci?c nitrosoproteomic identi?cation of endogenously S-nitrosylated proteins in Arabidopsis Plant Physiol 167, 1731-1746. DOI:10.1104/pp.15.00026URLPMID:25699590 [本文引用: 1] Nitric oxide (NO) regulates multiple developmental events and stress responses in plants. A major biologically active species of NO is S-nitrosoglutathione (GSNO), which is irreversibly degraded by GSNO reductase (GSNOR). The major physiological effect of NO is protein S-nitrosylation, a redox-based posttranslational modification mechanism by covalently linking an NO molecule to a cysteine thiol. However, little is known about the mechanisms of S-nitrosylation-regulated signaling, partly due to limited S-nitrosylated proteins being identified. In this study, we identified 1,195 endogenously S-nitrosylated peptides in 926 proteins from the Arabidopsis (Arabidopsis thaliana) by a site-specific nitrosoproteomic approach, which, to date, is the largest data set of S-nitrosylated proteins among all organisms. Consensus sequence analysis of these peptides identified several motifs that contain acidic, but not basic, amino acid residues flanking the S-nitrosylated cysteine residues. These S-nitrosylated proteins are involved in a wide range of biological processes and are significantly enriched in chlorophyll metabolism, photosynthesis, carbohydrate metabolism, and stress responses. Consistently, the gsnor1-3 mutant shows the decreased chlorophyll content and altered photosynthetic properties, suggesting that S-nitrosylation is an important regulatory mechanism in these processes. These results have provided valuable resources and new clues to the studies on S-nitrosylation-regulated signaling in plants.
IarullinaDR, AsafovaEV, KartunovaIE, ZiiatdinovaGK, Il'inskaiaON ( 2014). Probiotics for plants: NO-producing lactobacilli protect plants from drought Prikl Biokhim Mikrobiol 50, 189-192. URLPMID:25272737 [本文引用: 1] After the inoculation of wheat roots with a suspension of the bacterium Lactobacillus plantarum, leveling of oxidative stress detected by the accumulation of H2O2 and MDA was found in leaves. Activation of catalase and increased integral antioxidant capacity in seedlings treated with NO-producing lactobacilli were detected during the determination of the contribution of bacterial NO to the plant stress reaction. Thus, for the first time, we have demonstrated that lactobacilli affect plant adaptive responses to stress by the involvement of nitric oxide.
IgamberdievAU, HillRD ( 2004). Nitrate, NO and haemoglobin in plant adaptation to hypoxia: an alternative to classic fermentation pathways J Exp Bot 55, 2473-2482. URLPMID:15448180 [本文引用: 1]
IgamberdievAU, HillRD ( 2009). Plant mitochondrial function during anaerobiosis Ann Bot 103, 259-268. DOI:10.1093/aob/mcn100URLPMID:18586697 [本文引用: 2] BACKGROUND: Under hypoxic conditions, plant mitochondria preserve the capacity to oxidize external NADH, NADPH and tricarboxylic acid cycle substrates. Nitrite serves as an alternative electron acceptor at the level of cytochrome oxidase, with possibly complex III and the alternative oxidase also being involved. Nitric oxide is a significant product of the reaction, which has a high affinity for cytochrome c oxidase, inhibiting it. The excess NO is scavenged by hypoxically induced class 1 haemoglobin in the reaction involving ascorbate. SCOPE: By using nitrite, mitochondria retain a limited capacity for ATP synthesis. NADH, produced from glycolysis during anaerobiosis and oxidized in the mitochondrial electron transport chain, should shift the composition of metabolites formed during anaerobiosis with increased conversion of pyruvate to alanine and greater involvement of other transamination reactions, such as those involving gamma-aminobutyric acid formation. CONCLUSIONS: Anaerobic mitochondrial metabolism may have a more significant role than previously thought in alleviating the effects of anoxia on plant cells. There is a need to re-examine mitochondrial carbon and nitrogen metabolism under anoxia to establish the extent of this involvement.
IgamberdievAU, RatcliffeRG, GuptaKJ ( 2014). Plant mitochondria: source and target for nitric oxide Mitochondrion 19, 329-333. DOI:10.1016/j.mito.2014.02.003URLPMID:24561220 [本文引用: 3] Plant mitochondria generate nitric oxide (NO) under anoxia through the action of cytochrome c oxidase and other electron transport chain components on nitrite. This reductive mechanism operates under aerobic conditions at high electron transport rates. Indirect evidence also indicates that the oxidative pathway of NO production may be associated with mitochondria. We review the consequences of mitochondrial NO production, including the inhibition of oxygen uptake by cytochrome c oxidase, the inhibition of aconitase and succinate dehydrogenase, the induction of alternative oxidase, and the nitrosylation of several proteins, including glycine decarboxylase. The importance of these events in adaptation to abiotic and biotic stresses is discussed.
InnocentiG, PucciarielloC, Le GleuherM, HopkinsJ, de StefanoM, DelledonneM, PuppoA, BaudouinE, FrendoP ( 2007). Glutathione synthesis is regulated by nitric oxide in Medicago truncatula roots Planta 225, 1597-1602. URLPMID:17195940 [本文引用: 2]
KatoK, KanahamaK, KanayamaY ( 2010). Involvement of nitric oxide in the inhibition of nitrogenase activity by nitrate in Lotus root nodules J Plant Physiol 167, 238-241. DOI:10.1016/j.jplph.2009.08.006URLPMID:19733934 [本文引用: 5] Nitrogenase activity, as acetylene-reduction activity (ARA), in Lotus root nodules was clearly inhibited 27h after the addition of nitrate. Nitric oxide (NO) production was detected at that time in nitrate-supplied root nodules using the NO-reactive fluorescent probe diaminofluorescein-2 diacetate. The involvement of NO production in the inhibition of nitrogenase activity by nitrate was investigated using the NO donor sodium nitroprusside (SNP) and the NO scavenger 2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide (c-PTIO). SNP inhibited ARA at 1mM, and c-PTIO suppressed the inhibition of ARA by nitrate. These results suggest that NO is involved in the inhibition of nitrogenase activity by nitrate in Lotus root nodules.
LeeHW, HitchcocTM, ParkSH, MiR, KraftJD, LuoJ, CaoWG ( 2010). Involvement of thioredoxin domain-containing 5 in resistance to nitrosative stress Free Radic Biol Med 49, 872-880. DOI:10.1016/j.freeradbiomed.2010.06.012URLPMID:20550962 [本文引用: 1] Living organisms are exposed to nitrosative stress mediated by nitric oxide (NO) and its derivatives. Multiple cellular mechanisms may be needed to cope with nitrosative stress. This work takes advantage of a hypersensitive Escherichia coli genetic system to identify genes involved in resistance to nitrosative stress in mouse lungs. Mouse thioredoxin domain-containing 5 (mTrx 5) was identified as one of the candidate genes. Its ability to complement the hypersensitive phenotype in an E. coli mutant strain was confirmed by genetic analysis. Purified recombinant mouse thioredoxin domain-containing 5 protein reduced DNA damage that is sensitive to cleavage by the deamination repair enzyme endonuclease V, indicating that mTrx 5 may play a role in scavenging the reactive nitrogen species. E. coli thioredoxin 1 and thioredoxin 2 proteins also reduced the DNA damage in a similar manner. Deletion of trxA (encodes thioredoxin 1) or trxC (encodes thioredoxin 2) in E. coli resulted in a slightly higher sensitivity to nitrosative stress. On the other hand, deletion of both trxA and trxC greatly increased its sensitivity to nitrosative stress. Complementation with the mTrx 5 gene rescued the sensitive phenotype of the double deletion mutant. The potential roles that mTrx 5 may play in coping with nitrosative stress are discussed.
LiBH, LiGJ, KronzuckerHJ, Balu?kaF, ShiWM ( 2014). Ammonium stress in Arabidopsis: signaling, genetic loci, and physiological targets Trends Plant Sci 19, 107-114. DOI:10.1016/j.tplants.2013.09.004URLPMID:24126103 [本文引用: 3] Ammonium (NH4(+)) toxicity is a significant ecological and agricultural issue, and an important phenomenon in cell biology. As a result of increasing soil nitrogen input and atmospheric deposition, plants have to deal with unprecedented NH4(+) stress from sources below and above ground. In this review, we describe recent advances in elucidating the signaling pathways and identifying the main physiological targets and genetic loci involved in the effects of NH4(+) stress in the roots and shoots of Arabidopsis thaliana. We outline new experimental approaches that are being used to study NH4(+) toxicity in Arabidopsis and propose an integrated view of behavior and signaling in response to NH4(+) stress in the Arabidopsis system.
LinAH, WangYQ, TangJY, XueP, LiCL, LiuLC, HuB, YangFQ, LoakeGJ, ChuCC ( 2012). Nitric oxide and protein S-nitrosylation are integral to hydrogen peroxide-induced leaf cell death in rice Plant Physiol 158, 451-464. DOI:10.1104/pp.111.184531URLPMID:22106097 [本文引用: 1] Nitric oxide (NO) is a key redox-active, small molecule involved in various aspects of plant growth and development. Here, we report the identification of an NO accumulation mutant, nitric oxide excess1 (noe1), in rice (Oryza sativa), the isolation of the corresponding gene, and the analysis of its role in NO-mediated leaf cell death. Map-based cloning revealed that NOE1 encoded a rice catalase, OsCATC. Furthermore, noe1 resulted in an increase of hydrogen peroxide (H(2)O(2)) in the leaves, which consequently promoted NO production via the activation of nitrate reductase. The removal of excess NO reduced cell death in both leaves and suspension cultures derived from noe1 plants, implicating NO as an important endogenous mediator of H(2)O(2)-induced leaf cell death. Reduction of intracellular S-nitrosothiol (SNO) levels, generated by overexpression of rice S-nitrosoglutathione reductase gene (GSNOR1), which regulates global levels of protein S-nitrosylation, alleviated leaf cell death in noe1 plants. Thus, S-nitrosylation was also involved in light-dependent leaf cell death in noe1. Utilizing the biotin-switch assay, nanoliquid chromatography, and tandem mass spectrometry, S-nitrosylated proteins were identified in both wild-type and noe1 plants. NO targets identified only in noe1 plants included glyceraldehyde 3-phosphate dehydrogenase and thioredoxin, which have been reported to be involved in S-nitrosylation-regulated cell death in animals. Collectively, our data suggest that both NO and SNOs are important mediators in the process of H(2)O(2)-induced leaf cell death in rice.
LoscosJ, MatamorosMA, BecanaM ( 2008). Ascorbate and homoglutathione metabolism in common bean nodules under stress conditions and during natural senescence Plant Physiol 146, 1282-1292. DOI:10.1104/pp.107.114066URLPMID:18218966 [本文引用: 1] Ascorbate and glutathione are major antioxidants and redox buffers in plant cells but also play key functions in growth, development, and stress responses. We have studied the regulation of ascorbate and homoglutathione biosynthesis in common bean (Phaseolus vulgaris) nodules under stress conditions and during aging. The expression of five genes of the major ascorbate biosynthetic pathway was analyzed in nodules, and evidence was found that L-galactono-1,4-lactone dehydrogenase, the last committed step of the pathway, is posttranscriptionally regulated. Also, in nodules under stress conditions, gamma-glutamylcysteine synthetase was translationally regulated, but homoglutathione synthetase (mRNA and activity) and homoglutathione (content and redox state) were not affected. Most interestingly, in nodules exposed to jasmonic acid, dehydroascorbate reductase activity was posttranslationally suppressed, ascorbate oxidase showed strong transcriptional up-regulation, and dehydroascorbate content increased moderately. These changes were not due to a direct effect of jasmonic acid on the enzyme activities but might be part of the signaling pathway in the response of nodules to stress. We determined ascorbate, homoglutathione, and ascorbate-glutathione pathway enzyme activities in two senescing stages of nodules undergoing oxidative stress. When all parameters were expressed on a nodule fresh weight basis, we found that in the first stage ascorbate decreased by 60% and homoglutathione and antioxidant activities remained fairly constant, whereas in the second stage ascorbate and homoglutathione, their redox states, and their associated enzyme activities significantly decreased. The coexistence in the same plants of nodules at different senescence stages, with different ascorbate concentrations and redox states, indicates that the life span of nodules is in part controlled by endogenous factors and points to ascorbate as one of the key players.
MatamorosMA, MoranJF, Iturbe-OrmaetxeI, RubioMC, BecanaM ( 1999). Glutathione and homoglutathione synthesis in legume root nodules Plant Physiol 121, 879-888. DOI:10.1104/pp.121.3.879URLPMID:10557236 [本文引用: 1] High-performance liquid chromatography (HPLC) with fluorescence detection was used to study thiol metabolism in legume nodules. Glutathione (GSH) was the major non-protein thiol in all indeterminate nodules examined, as well as in the determinate nodules of cowpea (Vigna unguiculata), whereas homoglutathione (hGSH) predominated in soybean (Glycine max), bean (Phaseolus vulgaris), and mungbean (Vigna radiata) nodules. All nodules had greater thiol concentrations than the leaves and roots of the same plants because of active thiol synthesis in nodule tissue. The correlation between thiol tripeptides and the activities of glutathione synthetase (GSHS) and homoglutathione synthetase (hGSHS) in the nodules of eight legumes, and the contrasting thiol contents and activities in alfalfa (Medicago sativa) leaves (98% hGSH, 100% hGSHS) and nodules (72% GSH, 80% GSHS) indicated that the distribution of GSH and hGSH is determined by specific synthetases. Thiol contents and synthesis decreased with both natural and induced nodule senescence, and were also reduced in the senescent zone of indeterminate nodules. Thiols and GSHS were especially abundant in the meristematic and infected zones of pea (Pisum sativum) nodules. Thiols and gamma-glutamylcysteinyl synthetase were also more abundant in the infected zone of bean nodules, but hGSHS was predominant in the cortex. Isolation of full-length cDNA sequences coding for gamma-glutamylcysteinyl synthetase from legume nodules revealed that they are highly homologous to those from other higher plants.
MatamorosMA, SaizA, Pe?uelasM, Bustos-SanmamedP, MuletJM, BarjaMV, RouhierN, MooreM, JamesEK, DietzKJ, BecanaM ( 2015). Function of glutathione peroxidases in legume root nodules J Exp Bot 66, 2979-2990. DOI:10.1093/jxb/erv066URLPMID:25740929 [本文引用: 2] Glutathione peroxidases (Gpxs) are antioxidant enzymes not studied so far in legume nodules, despite the fact that reactive oxygen species are produced at different steps of the symbiosis. The function of two Gpxs that are highly expressed in nodules of the model legume Lotus japonicus was examined. Gene expression analysis, enzymatic and nitrosylation assays, yeast cell complementation, in situ mRNA hybridization, immunoelectron microscopy, and LjGpx-green fluorescent protein (GFP) fusions were used to characterize the enzymes and to localize each transcript and isoform in nodules. The LjGpx1 and LjGpx3 genes encode thioredoxin-dependent phospholipid hydroperoxidases and are differentially regulated in response to nitric oxide (NO) and hormones. LjGpx1 and LjGpx3 are nitrosylated in vitro or in plants treated with S-nitrosoglutathione (GSNO). Consistent with the modification of the peroxidatic cysteine of LjGpx3, in vitro assays demonstrated that this modification results in enzyme inhibition. The enzymes are highly expressed in the infected zone, but the LjGpx3 mRNA is also detected in the cortex and vascular bundles. LjGpx1 is localized to the plastids and nuclei, and LjGpx3 to the cytosol and endoplasmic reticulum. Based on yeast complementation experiments, both enzymes protect against oxidative stress, salt stress, and membrane damage. It is concluded that both LjGpxs perform major antioxidative functions in nodules, preventing lipid peroxidation and other oxidative processes at different subcellular sites of vascular and infected cells. The enzymes are probably involved in hormone and NO signalling, and may be regulated through nitrosylation of the peroxidatic cysteine essential for catalytic function.
MeakinGE, BuenoE, JepsonB, BedmarEJ, RichardsonDJ, DelgadoMJ ( 2007). The contribution of bacteroidal nitrate and nitrite reduction to the formation of nitrosylleghaemoglobin complexes in soybean root nodules Microbiology 153, 411-419. DOI:10.1099/mic.0.2006/000059-0URLPMID:17259612 [本文引用: 2] It is becoming recognized that leghaemoglobin constitutes an important buffer for the cytotoxic nitric oxide radical (NO(*)) in root nodules, although the sources of this NO(*) within nodules are unclear. In Bradyrhizobium japonicum bacteroids, NO(*) can be produced through the denitrification process, during which nitrate is reduced to nitrite by the periplasmic nitrate reductase Nap, and nitrite is reduced to NO(*) by the respiratory nitrite reductase NirK. To assess the contribution of bacteroidal denitrification to the NO(*) within nitrate-treated soybean nodules, electron paramagnetic resonance and UV-visible spectroscopy were employed to study the presence of nitrosylleghaemoglobin (LbNO) within nodules from plants inoculated with wild-type, napA or nirK B. japonicum strains. Since it has been found that hypoxia induces NO(*) production in plant root tissue, and that plant roots can be subjected to hypoxic stress during drought and flooding, the effect of hypoxic stress on the formation of LbNO complexes within nodules was also investigated. Maximal levels of LbNO were observed in nodules from plants treated with nitrate and subjected to hypoxic conditions. It is shown that, in the presence of nitrate, all of the LbNO within normoxic nodules arises from nitrate reduction by the bacteroidal periplasmic nitrate reductase, whereas Nap activity is only responsible for half of the LbNO within hypoxic nodules. In contrast to Nap, NirK is not essential for LbNO formation under any condition tested.
MeilhocE, BlanquetP, CamY, BruandC ( 2013). Control of NO level in rhizobium-legume root nodules: not only a plant globin story Plant Signal Behav 8, e25923. DOI:10.4161/psb.25923URL [本文引用: 2]
MeilhocE, BoscariA, BruandC, PuppoA, BrouquisseR ( 2011). Nitric oxide in legume-rhizobium symbiosis Plant Sci 181, 573-581. DOI:10.1016/j.plantsci.2011.04.007URLPMID:21893254 [本文引用: 5] Nitric oxide (NO) is a gaseous signaling molecule with a broad spectrum of regulatory functions in plant growth and development. NO has been found to be involved in various pathogenic or symbiotic plant-microbe interactions. During the last decade, increasing evidence of the occurrence of NO during legume-rhizobium symbioses has been reported, from early steps of plant-bacteria interaction, to the nitrogen-fixing step in mature nodules. This review focuses on recent advances on NO production and function in nitrogen-fixing symbiosis. First, the potential plant and bacterial sources of NO, including NO synthase-like, nitrate reductase or electron transfer chains of both partners, are presented. Then responses of plant and bacterial cells to the presence of NO are presented in the context of the N(2)-fixing symbiosis. Finally, the roles of NO as either a regulatory signal of development, or a toxic compound with inhibitory effects on nitrogen fixation, or an intermediate involved in energy metabolism, during symbiosis establishment and nodule functioning are discussed.
MeloPM, SilvaLS, RibeiroI, SeabraAR, CarvalhoHG ( 2011). Glutamine synthetase is a molecular target of nitric oxide in root nodules of Medicago truncatula and is regulated by tyrosine nitration Plant Physiol 157, 1505-1517. DOI:10.1104/pp.111.186056URLPMID:21914816 [本文引用: 2] Nitric oxide (NO) is emerging as an important regulatory player in the Rhizobium-legume symbiosis, but its biological role in nodule functioning is still far from being understood. To unravel the signal transduction cascade and ultimately NO function, it is necessary to identify its molecular targets. This study provides evidence that glutamine synthetase (GS), a key enzyme for root nodule metabolism, is a molecular target of NO in root nodules of Medicago truncatula, being regulated by tyrosine (Tyr) nitration in relation to active nitrogen fixation. In vitro studies, using purified recombinant enzymes produced in Escherichia coli, demonstrated that the M. truncatula nodule GS isoenzyme (MtGS1a) is subjected to NO-mediated inactivation through Tyr nitration and identified Tyr-167 as the regulatory nitration site crucial for enzyme inactivation. Using a sandwich enzyme-linked immunosorbent assay, it is shown that GS is nitrated in planta and that its nitration status changes in relation to active nitrogen fixation. In ineffective nodules and in nodules fed with nitrate, two conditions in which nitrogen fixation is impaired and GS activity is reduced, a significant increase in nodule GS nitration levels was observed. Furthermore, treatment of root nodules with the NO donor sodium nitroprusside resulted in increased in vivo GS nitration accompanied by a reduction in GS activity. Our results support a role of NO in the regulation of nitrogen metabolism in root nodules and places GS as an important player in the process. We propose that the NO-mediated GS posttranslational inactivation is related to metabolite channeling to boost the nodule antioxidant defenses in response to NO.
MoreauM, LindermayrC, DurnerJ, KlessigDF ( 2010). NO synthesis and signaling in plants-where do we stand? Physiol Plant 138, 372-383. DOI:10.1111/j.1399-3054.2009.01308.xURLPMID:19912564 [本文引用: 1] Over the past 20 years, nitric oxide (NO) research has generated a lot of interest in various aspects of plant biology. It is now clear that NO plays a role in a wide range of physiological processes in plants. However, in spite of the significant progress that has been made in understanding NO biosynthesis and signaling in planta, several crucial questions remain unanswered. Here we highlight several challenges in NO plant research by summarizing the latest knowledge of NO synthesis and by focusing on the potential NO source(s) and players involved. Our goal is also to provide an overview of how our understanding of NO signaling has been enhanced by the identification of array of genes and proteins regulated by NO.
MurLAJ, PratsE, PierreS, HallMA, HebelstrupKH ( 2013). Integrating nitric oxide into salicylic acid and jasmonic acid/ethylene plant defense pathways Front Plant Sci 4, 215. DOI:10.3389/fpls.2013.00215URLPMID:23818890 [本文引用: 3] Plant defense against pests and pathogens is known to be conferred by either salicylic acid (SA) or jasmonic acid (JA)/ethylene (ET) pathways, depending on infection or herbivore-grazing strategy. It is well attested that SA and JA/ET pathways are mutually antagonistic allowing defense responses to be tailored to particular biotic stresses. Nitric oxide (NO) has emerged as a major signal influencing resistance mediated by both signaling pathways but no attempt has been made to integrate NO into established SA/JA/ET interactions. NO has been shown to act as an inducer or suppressor of signaling along each pathway. NO will initiate SA biosynthesis and nitrosylate key cysteines on TGA-class transcription factors to aid in the initiation of SA-dependent gene expression. Against this, S-nitrosylation of NONEXPRESSOR OF PATHOGENESIS-RELATED PROTEINS1 (NPR1) will promote the NPR1 oligomerization within the cytoplasm to reduce TGA activation. In JA biosynthesis, NO will initiate the expression of JA biosynthetic enzymes, presumably to over-come any antagonistic effects of SA on JA-mediated transcription. NO will also initiate the expression of ET biosynthetic genes but a suppressive role is also observed in the S-nitrosylation and inhibition of S-adenosylmethionine transferases which provides methyl groups for ET production. Based on these data a model for NO action is proposed but we have also highlighted the need to understand when and how inductive and suppressive steps are used.
MurakamiEI, NagataM, ShimodaY, KuchoKI, HigashiS, AbeM, HashimotoM, UchiumiT ( 2011). Nitric oxide production induced in roots of Lotus japonicus by lipopolysaccharide from Mesorhizobium loti Plant Cell Physiol 52, 610-617. DOI:10.1093/pcp/pcr020URLPMID:21330297 [本文引用: 1] Lipopolysaccharide (LPS) is a bacterial molecule that induces nitric oxide (NO) production and triggers defense systems in plant-pathogen interactions. NO production is induced in the roots of Lotus japonicus after inoculation of the roots with its microsymbiont Mesorhizobium loti. However, the rhizobial molecule that induces NO production has not yet been identified. We investigated NO production in the roots of L. japonicus by treatment with LPS of M. loti. LPS was prepared by phenol-hot water extraction and separated into several fractions: polysaccharide, lipooligosaccharide, oligosaccharide and lipid A. In the roots of L. japonicus, NO production was observed with an NO-specific fluorescent dye 4, 10 and 24 h after treatment with each fraction of LPS. NO production was detected 4 h after treatment with all fractions. NO production was also detectable 24 h after treatment, except after treatment with the polysaccharide and oligosaccharide fractions. Expression of a class 1 hemoglobin gene and application of an NO scavenger showed that the treatment with LPS and LOS induced a similar response to inoculation with M. loti. These data suggest that LPS of M. loti induces NO production after inoculation with M. loti.
NagataM, MurakamiEI, ShimodaY, Shimoda-SasakuraF, KuchoKI, SuzukiA, AbeM, HigashiS, UchiumiT ( 2008). Expression of a class 1 hemoglobin gene and production of nitric oxide in response to symbiotic and pathogenic bacteria in Lotus japonicus Mol Plant Microbe Interact 21, 1175-1183. DOI:10.1094/MPMI-21-9-1175URLPMID:18700822 [本文引用: 1] Symbiotic nitrogen fixation by the collaboration between leguminous plants and rhizobia is an important system in the global nitrogen cycle, and some molecular aspects during the early stage of host-symbiont recognition have been revealed. To understand the responses of a host plant against various bacteria, we examined expression of hemoglobin (Hb) genes and production of nitric oxide (NO) in Lotus japonicus after inoculation with rhizobia or plant pathogens. When the symbiotic rhizobium Mesorhizobium loti was inoculated, expression of LjHb1 and NO production were induced transiently in the roots at 4 h after inoculation. In contrast, inoculation with the nonsymbiotic rhizobia Sinorhizobium meliloti and Bradyrhizobium japonicum induced neither expression of LjHb1 nor NO production. When L. japonicus was inoculated with plant pathogens (Ralstonia solanacearum or Pseudomonas syringae), continuous NO production was observed in roots but induction of LjHb1 did not occur. These results suggest that modulation of NO levels and expression of class 1 Hb are involved in the establishment of the symbiosis.
NavascuésJ, Pérez-RontoméC, GayM, MarcosM, YangF, WalkerFA, DesboisA, AbiánJ, BecanaM ( 2012). Leghemoglobin green derivatives with nitrated hemes evidence production of highly reactive nitrogen species during aging of legume nodules Proc Natl Acad Sci USA 109, 2660-2665. URLPMID:22308405 [本文引用: 1]
NeillS, BarrosR, BrightJ, DesikanR, HancockJ, HarrisonJ, MorrisP, RibeiroD, WilsonI ( 2008). Nitric oxide, stomatal closure, and abiotic stress J Exp Bot 59, 165-176. DOI:10.1093/jxb/erm293URLPMID:18332225 [本文引用: 1] Various data indicate that nitric oxide (NO) is an endogenous signal in plants that mediates responses to several stimuli. Experimental evidence in support of such signalling roles for NO has been obtained via the application of NO, usually in the form of NO donors, via the measurement of endogenous NO, and through the manipulation of endogenous NO content by chemical and genetic means. Stomatal closure, initiated by abscisic acid (ABA), is effected through a complex symphony of intracellular signalling in which NO appears to be one component. Exogenous NO induces stomatal closure, ABA triggers NO generation, removal of NO by scavengers inhibits stomatal closure in response to ABA, and ABA-induced stomatal closure is reduced in mutants that are impaired in NO generation. The data indicate that ABA-induced guard cell NO generation requires both nitric oxide synthase-like activity and, in Arabidopsis, the NIA1 isoform of nitrate reductase (NR). NO stimulates mitogen-activated protein kinase (MAPK) activity and cGMP production. Both these NO-stimulated events are required for ABA-induced stomatal closure. ABA also stimulates the generation of H2O2 in guard cells, and pharmacological and genetic data demonstrate that NO accumulation in these cells is dependent on such production. Recent data have extended this model to maize mesophyll cells where the induction of antioxidant defences by water stress and ABA required the generation of H2O2 and NO and the activation of a MAPK. Published data suggest that drought and salinity induce NO generation which activates cellular processes that afford some protection against the oxidative stress associated with these conditions. Exogenous NO can also protect cells against oxidative stress. Thus, the data suggest an emerging model of stress responses in which ABA has several ameliorative functions. These include the rapid induction of stomatal closure to reduce transpirational water loss and the activation of antioxidant defences to combat oxidative stress. These are two processes that both involve NO as a key signalling intermediate.
PalmieriMC, SellS, HuangX, ScherfM, WernerT, DurnerJ, LindermayrC ( 2008). Nitric oxide-responsive genes and promoters in Arabidopsis thaliana: a bioinformatics approach J Exp Bot 59, 177-186. DOI:10.1093/jxb/erm345URLPMID:18272923 [本文引用: 1] Due to its high reactivity and its ability to diffuse and permeate the cell membrane, nitric oxide (NO) and its exchangeable redox-activated species are unique biological messengers in animals and in plants. Although an increasing number of reports indicate that NO is an essential molecule in several physiological processes, there is not a clear picture of its method of action. Studies on the transcriptional changes induced by NO permitted identification of genes involved in different functional processes such as signal transduction, defence and cell death, transport, basic metabolism, and reactive oxygen species (ROS) production and degradation. The co-expression of these genes can be explained by the co-operation of a set of transcription factors that bind a common region in the promoter of the regulated genes. The present report describes the search for a common transcription factor-binding site (TFBS) in promoter regions of NO-regulated genes, based on microarray analyses. Using Genomatix Gene2Promotor and MatInspector, eight families of TFBSs were found to occur at least 15% more often in the promoter regions of the responsive genes in comparison with the promoter regions of 28,447 Arabidopsis control genes. Most of these TFBSs, such as ocs element-like sequences and WRKY, have already been reported to be involved in particular stress responses. Furthermore, the promoter regions of genes involved in jasmonic acid (JA) biosynthesis were analysed for a common TFBS module, since some genes responsible for JA biosynthesis are induced by NO, and an interaction between NO and JA signalling has already been described.
Pérez-ChacaMV, Rodríguez-SerranoM, MolinaAS, PedranzaniHE, ZirulnikF, SandalioLM, Romero- PuertasMC ( 2014). Cadmium induces two waves of reactive oxygen species in Glycine max (L.) roots Plant Cell Environ 37, 1672-1687. DOI:10.1111/pce.12280URLPMID:24433233 [本文引用: 1] Cadmium (Cd) is a non-essential heavy metal that may be toxic or even lethal to plants as it can be easily taken up by the roots and loaded into the xylem to the leaves. Using soybean roots (Glycine max L.) DM 4800, we have analysed various parameters related to reactive oxygen metabolism and nitric oxide (NO) during a 6 day Cd exposure. A rise in H(2)O(2) and NO, and to a lesser extent O(2)(.-) content was observed after 6 h exposure with a concomitant increase in lipid peroxidation and carbonyl group content. Both oxidative markers were significantly reduced after 24 h. A second, higher wave of O(2)(.-) production was also observed after 72 h of exposure followed by a reduction until the end of the treatment. NOX and glicolate oxidase activity might be involved in the initial Cd-induced reactive oxygen species (ROS) production and it appears that other sources may also participate. The analysis of antioxidative enzymes showed an increase in glutathione-S-transferase activity and in transcript levels and activity of enzymes involved in the ascorbate-glutathione cycle and the NADPH-generating enzymes. These results suggest that soybean is able to respond rapidly to oxidative stress imposed by Cd by improving the availability of NADPH necessary for the ascorbate-glutathione cycle.
Pérez GuerraJC, CoussensG, De KeyserA, De RyckeR, De BodtS, Van De VeldeW, GoormachtigS, HolstersM ( 2010). Comparison of developmental and stress-indu- ced nodule senescence in Medicago truncatula Plant Physiol 152, 1574-1584. DOI:10.1104/pp.109.151399URLPMID:20081044 [本文引用: 2] Mature indeterminate Medicago truncatula nodules are zonated with an apical meristem, an infection zone, a fixation zone with nitrogen-fixing bacteroids, and a
ProcházkováD, WilhelmováN ( 2011). Nitric oxide, reactive nitrogen species and associated enzymes during plant senescence Nitric Oxide 24, 61-65. DOI:10.1016/j.niox.2011.01.005URLPMID:21256240 [本文引用: 1] Leaf senescence is often associated with increased oxidative damage to cellular macromolecules by reactive oxygen species. However, very little is known about other radicals: gaseous free radical nitric oxide and related molecules--reactive nitrogen species. This review brings a short survey of the questions.
PuppoA, PaulyN, BoscariA, MandonK, BrouquisseR ( 2013). Hydrogen peroxide and nitric oxide: key regulators of the legume- Rhizobium and mycorrhizal symbioses Antioxid Redox Signal 18, 2202-2219. URLPMID:23249379 [本文引用: 2]
RadiR ( 2004). Nitric oxide, oxidants, and protein tyrosine nitration Proc Natl Acad Sci USA 101, 4003-4008. DOI:10.1073/pnas.0307446101URLPMID:15020765 [本文引用: 1] The occurrence of protein tyrosine nitration under disease conditions is now firmly established and represents a shift from the signal transducing physiological actions of (.)NO to oxidative and potentially pathogenic pathways. Tyrosine nitration is mediated by reactive nitrogen species such as peroxynitrite anion (ONOO(-)) and nitrogen dioxide ((.)NO2), formed as secondary products of (.)NO metabolism in the presence of oxidants including superoxide radicals (O2(.-)), hydrogen peroxide (H2O2), and transition metal centers. The precise interplay between (.)NO and oxidants and the identification of the proximal intermediate(s) responsible for nitration in vivo have been under controversy. Despite the capacity of peroxynitrite to mediate tyrosine nitration in vitro, its role on nitration in vivo has been questioned, and alternative pathways, including the nitrite/H2O2/hemeperoxidase and transition metal-dependent mechanisms, have been proposed. A balanced analysis of existing evidence indicates that (i) different nitration pathways can contribute to tyrosine nitration in vivo, and (ii) most, if not all, nitration pathways involve free radical biochemistry with carbonate radicals (CO3(.-)) and/or oxo-metal complexes oxidizing tyrosine to tyrosyl radical followed by the diffusion-controlled reaction with (.)NO2 to yield 3-nitrotyrosine. Although protein tyrosine nitration is a low-yield process in vivo, 3-nitrotyrosine has been revealed as a relevant biomarker of (.)NO-dependent oxidative stress; additionally, site-specific nitration focused on particular protein tyrosines may result in modification of function and promote a biological effect. Tissue distribution and quantitation of protein 3-nitrotyrosine, recognition of the predominant nitration pathways and individual identification of nitrated proteins in disease states open new avenues for the understanding and treatment of human pathologies.
RomanovVI, FedulovaNG, TchermenskayaIE, ShramkoVI, MolchanovMI, KretovichWL ( 1980). Metabolism of poly-hydroxybutyric acid in bacteroids of Rhizobium lupini in connection with nitrogen ?xation and photosynthesis Plant Soil 56, 379-390. DOI:10.1007/BF02143032URL [本文引用: 1]
SainzM, Calvo-BegueriaL, Pérez-RontoméC, WienkoopS, AbiánJ, StaudingerC, BartesaghiS, RadiR, BecanaM ( 2015). Leghemoglobin is nitrated in functional legume nodules in a tyrosine residue within the heme cavity by a nitrite/peroxide-dependent mechanism Plant J 81, 723-735. DOI:10.1111/tpj.12762URLPMID:25603991 [本文引用: 2] Protein tyrosine (Tyr) nitration is a post-translational modification yielding 3-nitrotyrosine (NO2 -Tyr). Formation of NO2 -Tyr is generally considered as a marker of nitro-oxidative stress and is involved in some human pathophysiological disorders, but has been poorly studied in plants. Leghemoglobin (Lb) is an abundant hemeprotein of legume nodules that plays an essential role as an O2 transporter. Liquid chromatography coupled to tandem mass spectrometry was used for a targeted search and quantification of NO2 -Tyr in Lb. For all Lbs examined, Tyr30, located in the distal heme pocket, is the major target of nitration. Lower amounts were found for NO2 -Tyr25 and NO2 -Tyr133. Nitrated Lb and other as yet unidentified nitrated proteins were also detected in nodules of plants not receiving NO3- and were found to decrease during senescence. This demonstrates formation of nitric oxide ( NO) and NO2- by alternative means to nitrate reductase, probably via a NO synthase-like enzyme, and strongly suggests that nitrated proteins perform biological functions and are not merely metabolic byproducts. In vitro assays with purified Lb revealed that Tyr nitration requires NO2- + H2 O2 and that peroxynitrite is not an efficient inducer of nitration, probably because Lb isomerizes it to NO3-. Nitrated Lb is formed via oxoferryl Lb, which generates nitrogen dioxide and tyrosyl radicals. This mechanism is distinctly different from that involved in heme nitration. Formation of NO2 -Tyr in Lb is a consequence of active metabolism in functional nodules, where Lb may act as a sink of toxic peroxynitrite and may play a protective role in the symbiosis.
SánchezC, CabreraJJ, GatesAJ, BedmarEJ, RichardsonDJ, DelgadoMJ ( 2011). Nitric oxide detoxification in the rhizobia-legume symbiosis Biochem Soc Trans 39, 184-188. DOI:10.1042/BST0390184URLPMID:21265770 [本文引用: 4] NO (nitric oxide) is a signal molecule involved in diverse physiological processes in cells which can become very toxic under certain conditions determined by its rate of production and diffusion. Several studies have clearly shown the production of NO in early stages of rhizobia-legume symbiosis and in mature nodules. In functioning nodules, it has been demonstrated that NO, which has been reported as a potent inhibitor of nitrogenase activity, can bind Lb (leghaemoglobin) to form LbNOs (nitrosyl-leghaemoglobin complexes). These observations have led to the question of how nodules overcome the toxicity of NO. On the bacterial side, one candidate for NO detoxification in nodules is the respiratory Nor (NO reductase) that catalyses the reduction of NO to nitrous oxide. In addition, rhizobial fHbs (flavohaemoglobins) and single-domain Hbs which dioxygenate NO to form nitrate are candidates to detoxify NO under free-living and symbiotic conditions. On the plant side, sHbs (symbiotic Hbs) (Lb) and nsHbs (non-symbiotic Hbs) have been proposed to play important roles as modulators of NO levels in the rhizobia-legume symbiosis. In the present review, current knowledge of NO detoxification by legume-associated endosymbiotic bacteria is summarized.
SánchezC, GatesAJ, MeakinGE, UchiumiT, GirardL, RichardsonDJ, BedmarEJ, DelgadoMJ ( 2010). Production of nitric oxide and nitrosylleghemoglobin complexes in soybean nodules in response to flooding Mol Plant Microbe Interact 23, 702-711. DOI:10.1094/MPMI-23-5-0702URLPMID:20367476 [本文引用: 5] Nitric oxide (NO) has gained interest as a major signaling molecule during plant development and in response to environmental cues. Formation of NO during symbiotic interactions has been reported, but the role and sources of NO in nodules remain unclear. In this work, the involvement of denitrification, performed by the symbiont Bradyrhizobium japonicum, in NO formation in soybean nodules in response to flooding conditions has been investigated by inoculating plants with napA-, nirK-, or norC-deficient mutants. Levels of nitrosylleghemoglobin (LbNO) in flooded nirK and norC nodules were significantly higher than those observed in wild-type nodules. In addition, nirK and norC nodules accumulated more nitrite and NO, respectively, than wild-type nodules. By contrast, levels of LbNO, nitrite, and NO in flooded napA nodules were lower than in wild-type nodules. These results suggest that LbNO formation in soybean nodules in response to flooding conditions is caused by nitrite and NO generated from periplasmic nitrate reductase (Nap) and also containing nitrite reductase (NirK) denitrification enzymes. Flooding caused a decrease of nifH expression and nitrogenase activity in wild-type and norC nodules but not in napA or nirK nodules. Incubation of wild-type and norC nodules with a NO scavenger counteracted the effect of flooding. Under free-living conditions, beta-galactosidase activity from a nifD'-'lacZ fusion decreased in a norC mutant, which also accumulated NO in the medium. These results suggest that NO formed by Cu-containing nitrite reductase in soybean nodules in response to flooding has a negative effect on expression of nitrogenase. We propose that Lb has a major role in detoxifying NO and nitrite produced by bacteroidal denitrification in response to flooding conditions.
SheXP, SongXG, HeJM ( 2004). Role and relationship of nitric oxide and hydrogen peroxide in light/dark-regulated stomatal movement in Vicia faba Acta Bot Sin 46, 1292-1300. [本文引用: 1]
ShimodaY, NagataM, SuzukiA, AbeM, SatoS, KatoT, TabataS, HigashiS, UchiumiT ( 2005). Symbiotic rhizobium and nitric oxide induce gene expression of non- symbiotic hemoglobin in Lotus japonicus Plant Cell Physiol 46, 99-107. DOI:10.1093/pci/pci001URLPMID:15668209 [本文引用: 4] We characterized the expression profiles of LjHb1 and LjHb2, non-symbiotic hemoglobin (non-sym-Hb) genes of Lotus japonicus. Although LjHb1 and LjHb2 showed 77% homology in their cDNA sequences, LjHb2 is located in a unique position in the phylogenetic tree of plant Hbs. The 5'-upstream regions of both genes contain the motif AAAGGG at a position similar to that in promoters of other non-sym-Hb genes. Expression profiles obtained by using quantitative RT-PCR showed that LjHb1 and LjHb2 were expressed in all tissues of mature plants, and expression was enhanced in mature root nodules. LjHb1 was strongly induced under both hypoxic and cold conditions, and by the application of nitric oxide (NO) donor, whereas LjHb2 was induced only by the application of sucrose. LjHb1 was also induced transiently by the inoculation with the symbiotic rhizobium Mesorhizobium loti MAFF303099. Observations using fluorescence microscopy revealed the induction of LjHb1 expression corresponded to the generation of NO. These results suggest that non-sym-Hb and NO have important roles in stress adaptation and in the early stage of legume-rhizobium symbiosis.
SinghVP, SinghS, KumarJ, PrasadSM ( 2015). Hydrogen sulfide alleviates toxic effects of arsenate in pea seedlings through up-regulation of the ascorbate-glutathione cycle: possible involvement of nitric oxide J Plant Physiol 181, 20-29. URLPMID:25974366 [本文引用: 1]
SuzukiA, AkuneM, KogisoM, ImagamaY, OsukK, UchiumiT, HigashiS, HanSY, YoshidaS, AsamiT, AbeM ( 2004). Control of nodule number by the phytohormone abscisic acid in the roots of two leguminous species Plant Cell Physiol 45, 914-922. DOI:10.1093/pcp/pch107URLPMID:15295075 [本文引用: 1] The effects of the phytohormone abscisic acid (ABA) on plant growth and root nodule formation were analyzed in Trifolium repense (white clover) and Lotus japonicus, which form indeterminate and determinate nodules, respectively. In T. repense, although the number of nodules formed after inoculation with Rhizobium leguminosarum bv. trifolii strain 4S (wild type) was slightly affected by exogenous ABA, those formed by strain H1(pC4S8), which forms ineffective nodules, were dramatically reduced 28 days after inoculation (DAI). At 14 and 21 DAI, the number of nodules formed with the wild-type strain was decreased by exogenous ABA. In L. japonicus, the number of nodules was also reduced by ABA treatment. Thus, exogenous ABA inhibits root nodule formation after inoculation with rhizobia. Observation of root hair deformation revealed that ABA blocked the step between root hair swelling and curling. When the ABA concentration in plants was decreased by using abamine, a specific inhibitor of 9-cis-epoxycarotenoid dioxygenase, the number of nodules on lateral roots of abamine-treated L. japonicus increased dramatically, indicating that lower-than-normal concentrations of endogenous ABA enhance nodule formation. We hypothesize that the ABA concentration controls the number of root nodules.
SwarajK, SheokandS, Fernandez-PascualMM, de FelipeMR ( 2001). Dark-induced changes in legume nodule functioning Aust J Plant Physiol 28, 429-438. [本文引用: 1]
TominagaA, NagataM, FutsukiK, AbeH, UchiumiT, AbeM, KuchoKI, HashiguchiM, AkashiR, HirschA, ArimaS, SuzukiA ( 2010). Effect of abscisic acid on symbiotic nitrogen fixation activity in the root nodules of Lotus japonicus Plant Signal Behav 5, 440-443. DOI:10.4161/psb.5.4.10849URLPMID:20118670 [本文引用: 1] The phytohormone abscisic acid (ABA) is known to be a negative regulator of legume root nodule formation. By screening Lotus japonicus seedlings for survival on an agar medium containing 70 muM ABA, we obtained mutants that not only showed increased root nodule number, but also enhanced nitrogen fixation. The mutant was designated enf1 (enhanced nitrogen fixation 1) and was confirmed to be monogenic and incompletely dominant. In long-term growth experiments with M. loti, although some yield parameters were the same for both enf1 and wild-type plants, both the dry weight and N content of 100 seeds and entire enf1 plants were significantly larger compared than those traits in wild-type seeds and plants. The augmentation of the weight and N content of the enf1 plants most likely reflects the increased N supplied by the additional enf1 nodules and the concomitant increase in N fixation activity. We determined that the endogenous ABA concentration and the sensitivity to ABA of enf1 were lower than that of wild-type seedlings. When wild-type plants were treated with abamine, a specific inhibitor of 9-cis-epoxycarotenoid dioxygenase (NCED), which results in reduced ABA content, the N fixation activity of abamine-treated plants was elevated to the same levels as enf1. We also determined that production of nitric oxide (NO) in enf1 nodules was decreased. We conclude that endogenous ABA concentration not only regulates nodulation, but also nitrogen fixation activity by decreasing NO production in nodules.
TrevaskisB, WattsRA, AnderssonCR, LlewellynDJ, HargroveMS, OlsonJS, DennisES, PeacockWJ ( 1997). Two hemoglobin genes in Arabidopsis thaliana: the evolutionary origins of leghemoglobins Proc Natl Acad Sci USA 94, 12230-12234. DOI:10.1073/pnas.94.22.12230URLPMID:9342391 [本文引用: 1] We cloned two hemoglobin genes from Arabidopsis thaliana. One gene, AHB1, is related in sequence to the family of nonsymbiotic hemoglobin genes previously identified in a number of plant species (class 1). The second hemoglobin gene, AHB2, represents a class of nonsymbiotic hemoglobin (class 2) related in sequence to the symbiotic hemoglobin genes of legumes and Casuarina. The properties of these two hemoglobins suggest that the two families of nonsymbiotic hemoglobins may differ in function from each other and from the symbiotic hemoglobins. AHB1 is induced, in both roots and rosette leaves, by low oxygen levels. Recombinant AHB1 has an oxygen affinity so high as to make it unlikely to function as an oxygen transporter. AHB2 is expressed at a low level in rosette leaves and is low temperature-inducible. AHB2 protein has a lower affinity for oxygen than AHB1 but is similar to AHB1 in having an unusually low, pH-sensitive oxygen off-rate.
Van de VeldeW, GuerraJCP, De KeyserA, De RyckeR, RombautsS, MaunouryN, MergaertP, KondorosiE, HolstersM, GoormachtigS ( 2006). Aging in legume symbiosis. A molecular view on nodule senescence in Medicago truncatula Plant Physiol 141, 711-720. DOI:10.1104/pp.106.078691URLPMID:16648219 [本文引用: 1] Rhizobia reside as symbiosomes in the infected cells of legume nodules to fix atmospheric nitrogen. The symbiotic relation is strictly controlled, lasts for some time, but eventually leads to nodule senescence. We present a comprehensive transcriptomics study to understand the onset of nodule senescence in the legume Medicago truncatula. Distinct developmental stages with characteristic gene expression were delineated during which the two symbiotic partners were degraded consecutively, marking the switch in nodule tissue status from carbon sink to general nutrient source. Cluster analysis discriminated an early expression group that harbored regulatory genes that might be primary tools to interfere with pod filling-related or stress-induced nodule senescence, ultimately causing prolonged nitrogen fixation. Interestingly, the transcriptomes of nodule and leaf senescence had a high degree of overlap, arguing for the recruitment of similar pathways.
VinardellJM, FedorovaE, CebollaA, KeveiZ, HorvathG, KelemenZ, TarayreS, RoudierF, MergaertP, KondorosiA, KondorosiE ( 2003). Endoreduplication mediated by the anaphase-promoting complex activator CCS52A is required for symbiotic cell differentiation in Medicago truncatula nodules Plant Cell 15, 2093-2105. DOI:10.1105/tpc.014373URLPMID:12953113 [本文引用: 1] In Medicago nodules, endoreduplication cycles and ploidy-dependent cell enlargement occur during the differentiation of bacteroid-containing nitrogen-fixing symbiotic cells. These events are accompanied by the expression of ccs52A, a plant ortholog of the yeast and animal cdh1/srw1/fzr genes, acting as a substrate-specific activator of the anaphase-promoting complex (APC) ubiquitin ligase. Because CCS52A is involved in the transition of mitotic cycles to endoreduplication cycles, we investigated the importance of somatic endoploidy and the role of the M. truncatula ccs52A gene in symbiotic cell differentiation. Transcription analysis and ccs52A promoter-driven beta-glucuronidase activity in transgenic plants showed that ccs52A was dispensable for the mitotic cycles and nodule primordium formation, whereas it was induced before nodule differentiation. The CCS52A protein was present in the nucleus of endoreduplication-competent cells, indicating that it may activate APC constitutively during the endoreduplication cycles. Downregulation of ccs52A in transgenic M. truncatula plants drastically affected nodule development, resulting in lower ploidy, reduced cell size, inefficient invasion, and the maturation of symbiotic cells, accompanied by early senescence and finally the death of both the bacterium and plant cells. Thus, ccs52A expression is essential for the formation of large highly polyploid symbiotic cells, and endoreduplication is an integral part of normal nodule development.
WallyOSD, MiraMM, HillRD, StasollaC ( 2013). Hemoglobin regulation of plant embryogenesis and plant pathogen interaction Plant Signal Behav 8, e25264. DOI:10.4161/psb.25264URLPMID:23759548 [本文引用: 1] Plant hemoglobins are ubiquitous molecules involved in several aspects of plant development and stress responses. Studies on the functional aspects of plant hemoglobins at the cellular level in these processes are limited, despite their ability to scavenge nitric oxide (NO), an important signal molecule interfering with hormone synthesis and sensitivity. This mini-review summarizes current knowledge on plant hemoglobins, analyzes their participation in plant pathogen interaction and embryogenesis and proposes a possible model centering on jasmonic acid (JA) as a downstream component of hemoglobin responses.
WodalaB, DeákZ, VassI, ErdeiL, AltorjayI, HorváthF ( 2008). In vivo target sites of nitric oxide in photosynthetic electron transport as studied by chlorophyll ?uorescence in pea leaves Plant Physiol 146, 1920-1927. DOI:10.1104/pp.107.110205URLPMID:18245457 [本文引用: 1] The role of nitric oxide (NO) in photosynthesis is poorly understood as indicated by a number of studies in this field with often conflicting results. As various NO donors may be the primary source of discrepancies, the aim of this study was to apply a set of NO donors and its scavengers, and examine the effect of exogenous NO on photosynthetic electron transport in vivo as determined by chlorophyll fluorescence of pea (Pisum sativum) leaves. Sodium nitroprusside-induced changes were shown to be mediated partly by cyanide, and S-nitroso-N-acetylpenicillinamine provided low yields of NO. However, the effects of S-nitrosoglutathione are inferred exclusively by NO, which made it an ideal choice for this study. Q(A)(-) reoxidation kinetics show that NO slows down electron transfer between Q(A) and Q(B), and inhibits charge recombination reactions of Q(A)(-) with the S(2) state of the water-oxidizing complex in photosystem II. Consistent with these results, chlorophyll fluorescence induction suggests that NO also inhibits steady-state photochemical and nonphotochemical quenching processes. NO also appears to modulate reaction-center-associated nonphotochemical quenching.
YoshidaT, MogamiJ, Yamaguchi-ShinozakiK ( 2015). Omics approaches toward defining the comprehensive abscisic acid signaling network in plants Plant Cell Physiol 56, 1043-1052. DOI:10.1093/pcp/pcv060URLPMID:25917608 [本文引用: 1] ABA is a plant hormone that plays crucial roles in controlling cellular and physiological responses to osmotic stress and in developmental processes. Endogenous ABA levels are increased in response to a decrease in water availability in cells, and ABA sensing and signaling are thought to be mediated according to the current model established in Arabidopsis thaliana, which involves pyrabactin resistance 1 (PYR)/PYR1-like (PYL)/regulatory components of ABA receptor (RCAR), protein phosphatase 2C (PP2C) and sucrose non-fermenting-1 (SNF1)-related protein kinase 2 (SnRK2). These core components of ABA signaling have a pivotal role in stress-responsive gene expression and stomatal regulation. However, because a limited number of their upstream and downstream factors have been characterized, it is still difficult to define the comprehensive network of ABA signaling in plants. This review focuses on current progress in the study of PYR/PYL/RCARs, PP2Cs and SnRK2s, with particular emphasis on omics approaches, such as interactome and phosphoproteome studies. Moreover, the role of ABA in plant growth and development is discussed based on recent metabolomic profiling studies.
Zimmer-PradosLM, MoreiraASFP, MagalhaesJR, Fran?aMGC ( 2014). Nitric oxide increases tolerance responses to moderate water deficit in leaves of Phaseolus vulgaris and Vigna unguiculata bean species Physiol Mol Biol Plants 20, 295-301. DOI:10.1007/s12298-014-0239-1URLPMID:25049456 [本文引用: 1] Drought stress is one of the most intensively studied and widespread constraints, and nitric oxide (NO) is a key signaling molecule involved in the mediation of abiotic stresses in plants. We demonstrated that a sprayed solution of NO from donor sodium nitroprusside increased drought stress tolerance responses in both sensitive (Phaseolus vulgaris) and tolerant (Vigna unguiculata) beans. In intact plants subjected to halting irrigation, NO increased the leaf relative water content and stomatal conductance in both species. After cutting leaf discs and washing them, NO induced increased electrolyte leakage, which was more evident in the tolerant species. These leaf discs were then subjected to different water deficits, simulating moderate and severe drought stress conditions through polyethylene glycol solutions. NO supplied at moderate drought stress revealed a reduced membrane injury index in sensitive species. In hydrated discs and at this level of water deficit, NO increased the electron transport rate in both species, and a reduction of these rates was observed at severe stress levels. Taken together, it can be shown that NO has an effective role in ameliorating drought stress effects, activating tolerance responses at moderate water deficit levels and in both bean species which present differential drought tolerance. 豆科植物早期共生信号转导的研究进展 1 2013
... NO不仅能调节植物生长发育, 还可参与植物对非生物胁迫的响应(Moreau et al., 2010).目前的研究发现, 在不同非生物胁迫(如干旱和缺氧)下, 共生有机体中的NO均会产生响应(Desalvo et al., 2010; Sánchez et al., 2011; Hawkins et al., 2014; Iarullina et al., 2014; Zimmer-Prados et al., 2014).同时, 在豆科植物中NO还可能参与调控机体对金属元素镉(Cd)和铝(Al)的胁迫反应以及应对非金属元素砷(As)的胁迫(Singh et al., 2015; 尚玉婷等, 2018), 但都缺乏在结瘤和共生固氮体系中的实验证据(Pérez- Chaca et al., 2014).Meilhoc等(2011)提出, 宿主中任何应激反应下NO的产生都会直接影响共生, 在成熟根瘤中, 除了NO专用调节器NnrR外, 多数根瘤菌的基因通过双组分系统(FixLJ)来调节NO, 其中FixLJ是O2限制反应的主要调节因子.nnrR是Nor结构基因norCB的上游基因, 可防止NO这种高活性氮氧化物的积累, 其插入失活可抑制亚硝酸盐的产生以及亚硝酸盐和NO的还原(De Bruijn et al., 2006).NO还能够可逆地结合光系统II (photosystem II, PSII)并调节电子的转移和猝灭, 对光合作用至关重要(Wodala et al., 2008).从农业和进化的角度来看, NO在植物与根瘤菌共生体系中的调控作用, 可以改善植物对病原体的防御状态、保护植物免受虫害以及增强植物对非生物胁迫的适应性(Chadha et al., 2015). ...
NO对植物生长发育的调控机制 1 2005
... 脱落酸(abscisic acid, ABA)是植物适应环境胁迫的关键激素(Yoshida et al., 2015), 可通过抑制根毛变形及侵染线的形成影响豆科植物结瘤(Suzuki et al., 2004; Ding et al., 2008).Tominaga等(2010)在百脉根中发现, ABA不仅可调节根瘤的固氮酶活性, 还可通过影响结瘤过程中NO的产生来影响结瘤.在根瘤形成过程中, NO通过酪氨酸(tyrosine, Tyr)硝化调控ABA信号传递, 而ABA又在信号转导过程中决定NO的产生与积累(Castillo et al., 2015).Tyr硝化是NO介导的翻译后修饰, 其特征在于将-NO2添加到Tyr残基芳环的等价邻位C上, 产生3-硝基酪氨酸(3-nitrotyrosine, 3-NT), 并由此改变蛋白质构象(Radi, 2004).作为硝化分子的前体, NO通过Tyr硝化改变细胞溶质谷氨酰胺合成酶(glutamine synthetase 1, GS1)和Lb等关键结瘤蛋白的活性(Melo et al., 2011; Navascués et al., 2012; Li et al., 2014).豆科植物结瘤时Lb发生Tyr硝化, 可保护共生体免受过氧亚硝酸盐毒害, 因此3-NT的形成既是硝基氧化应激的标志, 也是功能性结瘤活跃代谢的结果(Sainz et al., 2015).此外, NOS抑制剂N-硝基-L-精氨酸甲酯(NG-nitro- L-arginine methyl ester, L-NAME)和cPTIO处理可减弱豌豆(Pisum sativum)表皮由ABA诱导的气孔关闭(张绪成等, 2005). ...
Nitric oxide is a potent inhibitor of the cbb3-type heme-copper oxidases 1 2015
... 尽管目前对于豆科植物与根瘤菌共生固氮的初始过程已有较多研究, 但因发育或应激引起根瘤衰老的分子机制却报道较少.根瘤衰老区域的表型是由粉红色变为绿色, 其实质源于植物Hbs的降解; 在转录水平上, 衰老表现为编码蛋白质降解, 核酸、膜脂和糖类的相关基因被激活; 同时伴有衰老根瘤中氧化还原平衡的改变, 以及抗氧化防御的整体减弱等现象(Van de Velde et al., 2006; Loscos et al., 2008).NO在植物的衰老过程中具有重要作用(Procházková and Wilhelmová, 2011).例如, NO是水稻(Oryza sativa)光依赖性叶细胞死亡的关键介质(Lin et al., 2012); 蒺藜苜蓿根瘤内NO含量的增加会导致根瘤过早衰老, 且衰老程度与NO水平相关(Cam et al., 2012; Meilhoc et al., 2013; Blanquet et al., 2015), 表明NO对于维持共生体系和避免根瘤过早衰老极为重要.此外, 施加硝酸盐可导致豌豆表现出典型的衰老特征(Escuredo et al., 1996).对蒺藜苜蓿添加硝酸盐后, 其自身也可通过还原硝酸盐促使根瘤中生成NO, 然而硝酸盐引起的根瘤衰老是否依赖于NO目前尚不清楚(Horchani et al., 2011).此外, NO也是一种有效的呼吸抑制剂, 能够抑制线粒体和细菌末端的呼吸氧化酶, 并增加活性氧(reactive oxygen species, ROS)和活性氮(reactive nitrogen species, RNS)的积累, 而NO和ROS均为共生发育所必需(Shimoda et al., 2005; Cam et al., 2012; Igamberdiev et al., 2014; Arjona et al., 2015), 因此衰老也可能是固氮时呼吸抑制的间接结果. ...
Characterization of a member of the NnrR regulon in Rhodobacter sphaeroides 2.4.3 encoding a haem-copper protein: the GenBank accession number for nnrS is U62403 1 2002
... 共生体内NO的降解还受相关基因及其它蛋白调控.苜蓿中华根瘤菌(Sinorhizobium meliloti)中包含1个与短链脱氢酶相关且位于nnrR下游的nnrS基因家族, nnrS的表达依赖于转录调节器NnrR (De Bruijn et al., 2006).当nnrS1突变时, 根瘤内NO的水平升高(Blanquet et al., 2015).NnrS是一种haeme-Cu膜蛋白(Bartnikas et al., 2002), NnrS家族的2种蛋白(NnrS1和NnrS2)和Nor能够直接或间接参与NO的降解并有效地维持共生固氮(Meilhoc et al., 2013; Blanquet et al., 2015). ...
Nitric oxide is formed in Medicago truncatula-Sinorhizobium meliloti functional nodules 1 2006
Pathways of nitric oxide metabolism and operation of phytoglobins in legume nodules: missing links and future directions 1 2018
... 植物和根瘤菌内NO的来源丰富, 主要通过硝酸还原酶(nitrate reductase, NR)和一氧化氮合酶(nitric oxide synthase, NOS)介导以及线粒体电子传递链(electron transport chain, ETC)的还原产生.在植物体内, NO既可由质外体中的亚硝酸根离子(NO2-)转化(Bethke et al., 2004), 又可通过以精氨酸(arginine)、多胺(polyamines)或羟胺(hydroxylamine)作为底物的氧化途径产生(Hichri et al., 2015).在低氧环境下, 还可在还原途径中通过质膜结合亚硝酸盐, 然后通过NO还原酶(nitric oxide reductase, Nor)和黄嘌呤氧化还原酶(xanthine oxidoreductase, XOR)或利用ETC的作用将NO2-还原为NO (Gupta et al., 2011; Mur et al., 2013; Hichri et al., 2015).而Horchani等(2011)发现, 植物及其共生菌中的NR和ETC均可产生NO, 其中NR在植物体内的主要功能是将硝酸盐转化为亚硝酸盐以同化N2 (Neill et al., 2008).但早期的研究发现, 在NAD(P)H作为电子供体时, 大豆(Glycine max)中的NR可将亚硝酸盐转化为NO (Dean and Harper, 1988).且NR抑制剂钨酸盐(tungstate, Tg)也能够抑制NO的产生, 表明NO的合成依赖于NR活性, 但并不是由其直接产生(Horchani et al., 2011).在类菌体中, NO则主要产生于反硝化途径(Hichri et al., 2015), 该途径涉及NR、亚硝酸还原酶(nitrite reductase, NiR)、Nor和NOS (Horchani et al., 2011; Sánchez et al., 2011; Calvo-Begueria et al., 2018) (图1).Berger等(2018)发现NOS也是植物NO的来源, 但目前关于共生菌内NO产生途径的机制研究仍较少. ...
Apoplastic synthesis of nitric oxide by plant tissues 1 2004
... 植物和根瘤菌内NO的来源丰富, 主要通过硝酸还原酶(nitrate reductase, NR)和一氧化氮合酶(nitric oxide synthase, NOS)介导以及线粒体电子传递链(electron transport chain, ETC)的还原产生.在植物体内, NO既可由质外体中的亚硝酸根离子(NO2-)转化(Bethke et al., 2004), 又可通过以精氨酸(arginine)、多胺(polyamines)或羟胺(hydroxylamine)作为底物的氧化途径产生(Hichri et al., 2015).在低氧环境下, 还可在还原途径中通过质膜结合亚硝酸盐, 然后通过NO还原酶(nitric oxide reductase, Nor)和黄嘌呤氧化还原酶(xanthine oxidoreductase, XOR)或利用ETC的作用将NO2-还原为NO (Gupta et al., 2011; Mur et al., 2013; Hichri et al., 2015).而Horchani等(2011)发现, 植物及其共生菌中的NR和ETC均可产生NO, 其中NR在植物体内的主要功能是将硝酸盐转化为亚硝酸盐以同化N2 (Neill et al., 2008).但早期的研究发现, 在NAD(P)H作为电子供体时, 大豆(Glycine max)中的NR可将亚硝酸盐转化为NO (Dean and Harper, 1988).且NR抑制剂钨酸盐(tungstate, Tg)也能够抑制NO的产生, 表明NO的合成依赖于NR活性, 但并不是由其直接产生(Horchani et al., 2011).在类菌体中, NO则主要产生于反硝化途径(Hichri et al., 2015), 该途径涉及NR、亚硝酸还原酶(nitrite reductase, NiR)、Nor和NOS (Horchani et al., 2011; Sánchez et al., 2011; Calvo-Begueria et al., 2018) (图1).Berger等(2018)发现NOS也是植物NO的来源, 但目前关于共生菌内NO产生途径的机制研究仍较少. ...
Sinorhizobium meliloti controls nitric oxide-mediated post-translational modification of a Medicago truncatula nodule protein 6 2015
... 豆科植物根瘤菌共生体系受多种信号分子和转录因子调控, 其中一氧化氮(nitric oxide, NO)信号分子在豆科植物微生物互作过程中发挥重要作用.共生体早期的转录组学分析表明, NO可调节细胞脱分化和器官发生(Ferrarini et al., 2008; Boscari et al., 2013), 并抑制植物防御反应(Gonzalez-Rizzo et al., 2006), 在建立植物与菌共生关系中发挥重要调控作用(Hichri et al., 2016b).而在成熟根瘤中, NO既可抑制植物体固氮(Shimoda et al., 2005; Kato et al., 2010; Cam et al., 2012), 又可诱发根瘤衰老(Horchani et al., 2011; Cam et al., 2012; Blanquet et al., 2015), 亦可作为能量代谢调节器维持低氧(oxygen, O2)状态下的能量平衡(Kato et al., 2010; Hichri et al., 2016b).尽管目前关于NO在植物体内的生理功能已进行了充分研究, 但国内对于其在豆科植物根瘤菌共生体系中的作用机制并未进行系统的阐述.本文综述了豆科植物根瘤菌共生体系中NO的产生和降解, 及其对根瘤形成与共生固氮影响的研究进展, 并对NO在共生体系中对非生物胁迫的响应及其与激素的相互作用进行了阐述. ...
... 共生体内NO的降解还受相关基因及其它蛋白调控.苜蓿中华根瘤菌(Sinorhizobium meliloti)中包含1个与短链脱氢酶相关且位于nnrR下游的nnrS基因家族, nnrS的表达依赖于转录调节器NnrR (De Bruijn et al., 2006).当nnrS1突变时, 根瘤内NO的水平升高(Blanquet et al., 2015).NnrS是一种haeme-Cu膜蛋白(Bartnikas et al., 2002), NnrS家族的2种蛋白(NnrS1和NnrS2)和Nor能够直接或间接参与NO的降解并有效地维持共生固氮(Meilhoc et al., 2013; Blanquet et al., 2015). ...
... ; Blanquet et al., 2015). ...
... 除根瘤菌内的血红蛋白以外, 还有其它细菌蛋白调控根瘤中的NO水平.例如, 硫氧还蛋白(thioredoxin reductase, TrxR)或亚硝基谷胱甘肽还原酶(S-nitroglutathione reductase, GSNOR)都可能间接参与NO的降解和信号传递过程(Lee et al., 2010).而细菌的NO降解蛋白不仅能保护其自身蛋白免受NO的负面影响, 还能使共生植物蛋白免受NO介导的失活(Blanquet et al., 2015; Hichri et al., 2016b).综上, 鉴于NO的影响, 共生菌可能需要更多不同的系统来降低自身的NO水平. ...
... NO在影响相关基因的表达、调控豆科植物防御反应进而促进共生和结瘤中发挥重要作用.转录组分析表明, NO影响多种植物基因的表达, 如编码富含半胱氨酸蛋白(cysteine-rich proteins, NCRs)、核糖体蛋白、肽酶、结瘤发育细胞周期蛋白及细胞周期转换蛋白的基因(Vinardell et al., 2003; del Giudice et al., 2011; Boscari et al., 2013), 进而调控细胞脱分化、诱导组织形成及抑制植物防御反应(Hichri et al., 2016b).LjHb1是百脉根(Lotus japonicus) ns-Hbs的编码基因, 外源添加NO、缺氧和低温均可诱导其表达(Shimoda et al., 2005).在根瘤形成的早期阶段, NO的产生可以上调LjHb1的表达, 反之LjHb1的表达又下调NO的水平, 以降低植物防御反应, 促进根瘤菌和宿主植物建立共生关系(Nagata et al., 2008; Murakami et al., 2011).植物也可通过降低Hb的表达促进内源NO产生, 调控自身机体内相关防御基因的表达(Wally et al., 2013) (图2).而NO的清除会引起基因MtCRE1和MtCCS52A的下调表达, 从而延缓结瘤(del Giudice et al., 2011).通过对接种中华根瘤菌的蒺藜苜蓿外源添加NO清除剂(2-(4-carboxyphenyl)- 4,4,5,5-tetramethyl imidazoline-1-oxyl-3-oxide, cPTIO), 发现受NO调节的3种基因(Contig525、Contig3307和Medtr5g010350)表达下调(Boscari et al., 2013).其中, 前2种基因分别与ns-Hbs和茉莉酮酸代谢途径(Palmieri et al., 2008)相关, 而最后1种基因可调控还原型谷胱甘肽(glutathione, GSH)的合成(Innocenti et al., 2007).转录组分析显示, 蒺藜苜蓿接种中华根瘤菌后, 外源添加cPTIO或NADPH氧化酶抑制剂二苯烯碘铵(diphenylene iodide, DPI), 导致细胞壁形成和发育过程中相关基因下调表达, 而植物防御和二次代谢相关基因上调表达(Puppo et al., 2013).hmp的过表达和外源添加cPTIO都可降低蒺藜苜蓿的结瘤效率(Kato et al., 2010; del Giudice et al., 2011), NO则通过调节上述基因的表达, 抑制相关防御反应从而促进共生关系的建立.此外, 在根瘤菌内也鉴定出数百个依赖NO表达的基因, 如中华根瘤菌中编码Hmp的hmp基因(del Giudice et al., 2011; Cam et al., 2012)、编码haeme-Cu蛋白的nnrSI和nnrS2基因(Blanquet et al., 2015)及编码NiR的nirKV基因(Meilhoc et al., 2011), 但关于根瘤菌内受NO影响且与共生结瘤相关的基因报道很少. ...
... 尽管目前对于豆科植物与根瘤菌共生固氮的初始过程已有较多研究, 但因发育或应激引起根瘤衰老的分子机制却报道较少.根瘤衰老区域的表型是由粉红色变为绿色, 其实质源于植物Hbs的降解; 在转录水平上, 衰老表现为编码蛋白质降解, 核酸、膜脂和糖类的相关基因被激活; 同时伴有衰老根瘤中氧化还原平衡的改变, 以及抗氧化防御的整体减弱等现象(Van de Velde et al., 2006; Loscos et al., 2008).NO在植物的衰老过程中具有重要作用(Procházková and Wilhelmová, 2011).例如, NO是水稻(Oryza sativa)光依赖性叶细胞死亡的关键介质(Lin et al., 2012); 蒺藜苜蓿根瘤内NO含量的增加会导致根瘤过早衰老, 且衰老程度与NO水平相关(Cam et al., 2012; Meilhoc et al., 2013; Blanquet et al., 2015), 表明NO对于维持共生体系和避免根瘤过早衰老极为重要.此外, 施加硝酸盐可导致豌豆表现出典型的衰老特征(Escuredo et al., 1996).对蒺藜苜蓿添加硝酸盐后, 其自身也可通过还原硝酸盐促使根瘤中生成NO, 然而硝酸盐引起的根瘤衰老是否依赖于NO目前尚不清楚(Horchani et al., 2011).此外, NO也是一种有效的呼吸抑制剂, 能够抑制线粒体和细菌末端的呼吸氧化酶, 并增加活性氧(reactive oxygen species, ROS)和活性氮(reactive nitrogen species, RNS)的积累, 而NO和ROS均为共生发育所必需(Shimoda et al., 2005; Cam et al., 2012; Igamberdiev et al., 2014; Arjona et al., 2015), 因此衰老也可能是固氮时呼吸抑制的间接结果. ...
Expression dynamics of the Medicago truncatula transcriptome during the symbiotic interaction with Sinorhizobium meliloti: which role for nitric oxide 8 2013
... 豆科植物根瘤菌共生体系受多种信号分子和转录因子调控, 其中一氧化氮(nitric oxide, NO)信号分子在豆科植物微生物互作过程中发挥重要作用.共生体早期的转录组学分析表明, NO可调节细胞脱分化和器官发生(Ferrarini et al., 2008; Boscari et al., 2013), 并抑制植物防御反应(Gonzalez-Rizzo et al., 2006), 在建立植物与菌共生关系中发挥重要调控作用(Hichri et al., 2016b).而在成熟根瘤中, NO既可抑制植物体固氮(Shimoda et al., 2005; Kato et al., 2010; Cam et al., 2012), 又可诱发根瘤衰老(Horchani et al., 2011; Cam et al., 2012; Blanquet et al., 2015), 亦可作为能量代谢调节器维持低氧(oxygen, O2)状态下的能量平衡(Kato et al., 2010; Hichri et al., 2016b).尽管目前关于NO在植物体内的生理功能已进行了充分研究, 但国内对于其在豆科植物根瘤菌共生体系中的作用机制并未进行系统的阐述.本文综述了豆科植物根瘤菌共生体系中NO的产生和降解, 及其对根瘤形成与共生固氮影响的研究进展, 并对NO在共生体系中对非生物胁迫的响应及其与激素的相互作用进行了阐述. ...
... NO在影响相关基因的表达、调控豆科植物防御反应进而促进共生和结瘤中发挥重要作用.转录组分析表明, NO影响多种植物基因的表达, 如编码富含半胱氨酸蛋白(cysteine-rich proteins, NCRs)、核糖体蛋白、肽酶、结瘤发育细胞周期蛋白及细胞周期转换蛋白的基因(Vinardell et al., 2003; del Giudice et al., 2011; Boscari et al., 2013), 进而调控细胞脱分化、诱导组织形成及抑制植物防御反应(Hichri et al., 2016b).LjHb1是百脉根(Lotus japonicus) ns-Hbs的编码基因, 外源添加NO、缺氧和低温均可诱导其表达(Shimoda et al., 2005).在根瘤形成的早期阶段, NO的产生可以上调LjHb1的表达, 反之LjHb1的表达又下调NO的水平, 以降低植物防御反应, 促进根瘤菌和宿主植物建立共生关系(Nagata et al., 2008; Murakami et al., 2011).植物也可通过降低Hb的表达促进内源NO产生, 调控自身机体内相关防御基因的表达(Wally et al., 2013) (图2).而NO的清除会引起基因MtCRE1和MtCCS52A的下调表达, 从而延缓结瘤(del Giudice et al., 2011).通过对接种中华根瘤菌的蒺藜苜蓿外源添加NO清除剂(2-(4-carboxyphenyl)- 4,4,5,5-tetramethyl imidazoline-1-oxyl-3-oxide, cPTIO), 发现受NO调节的3种基因(Contig525、Contig3307和Medtr5g010350)表达下调(Boscari et al., 2013).其中, 前2种基因分别与ns-Hbs和茉莉酮酸代谢途径(Palmieri et al., 2008)相关, 而最后1种基因可调控还原型谷胱甘肽(glutathione, GSH)的合成(Innocenti et al., 2007).转录组分析显示, 蒺藜苜蓿接种中华根瘤菌后, 外源添加cPTIO或NADPH氧化酶抑制剂二苯烯碘铵(diphenylene iodide, DPI), 导致细胞壁形成和发育过程中相关基因下调表达, 而植物防御和二次代谢相关基因上调表达(Puppo et al., 2013).hmp的过表达和外源添加cPTIO都可降低蒺藜苜蓿的结瘤效率(Kato et al., 2010; del Giudice et al., 2011), NO则通过调节上述基因的表达, 抑制相关防御反应从而促进共生关系的建立.此外, 在根瘤菌内也鉴定出数百个依赖NO表达的基因, 如中华根瘤菌中编码Hmp的hmp基因(del Giudice et al., 2011; Cam et al., 2012)、编码haeme-Cu蛋白的nnrSI和nnrS2基因(Blanquet et al., 2015)及编码NiR的nirKV基因(Meilhoc et al., 2011), 但关于根瘤菌内受NO影响且与共生结瘤相关的基因报道很少. ...
... )表达下调(Boscari et al., 2013).其中, 前2种基因分别与ns-Hbs和茉莉酮酸代谢途径(Palmieri et al., 2008)相关, 而最后1种基因可调控还原型谷胱甘肽(glutathione, GSH)的合成(Innocenti et al., 2007).转录组分析显示, 蒺藜苜蓿接种中华根瘤菌后, 外源添加cPTIO或NADPH氧化酶抑制剂二苯烯碘铵(diphenylene iodide, DPI), 导致细胞壁形成和发育过程中相关基因下调表达, 而植物防御和二次代谢相关基因上调表达(Puppo et al., 2013).hmp的过表达和外源添加cPTIO都可降低蒺藜苜蓿的结瘤效率(Kato et al., 2010; del Giudice et al., 2011), NO则通过调节上述基因的表达, 抑制相关防御反应从而促进共生关系的建立.此外, 在根瘤菌内也鉴定出数百个依赖NO表达的基因, 如中华根瘤菌中编码Hmp的hmp基因(del Giudice et al., 2011; Cam et al., 2012)、编码haeme-Cu蛋白的nnrSI和nnrS2基因(Blanquet et al., 2015)及编码NiR的nirKV基因(Meilhoc et al., 2011), 但关于根瘤菌内受NO影响且与共生结瘤相关的基因报道很少. ...
... 一方面, NO抑制固氮和C、N代谢; 另一方面, NO调控细胞氧化还原和保持低氧水平下的能量状态.带+的细线表示NO的活化、诱导和保持效果; 带有-的细线表示NO的抑制作用.椭圆形粗线箭头表示NO主要的代谢途径.爆炸型图示指来自植物和菌共生体的酶, 闪电型图示表示根瘤菌内的基因.ACO: 乌头酸; CS: 柠檬酸合酶; Gln: 谷氨酰胺; Glu: 谷氨酸; GS: 谷氨酰胺合成酶; GSH: 谷胱甘肽; GSHS: 谷胱甘肽合成酶; GSNO: S-亚硝基谷胱甘肽; Hb: 血红蛋白; IDH: 异柠檬酸脱氢酶; MDH: 苹果酸脱氢酶; NH4+: 铵根离子; Nif: 固氮酶; SDH: 琥珀酸脱氢酶; γ-EC: γ-谷氨酰半胱氨酸; γ-ECS: γ-谷氨酰半胱氨酸合成酶 Schematic diagram of the role of NO in symbiotic nitrogen fixation (modified from <xref ref-type="bibr" rid="b11">Boscari et al., 2013</xref>; <xref ref-type="bibr" rid="b31">Hichri et al., 2015</xref>, <xref ref-type="bibr" rid="b33">2016b</xref>)On the one hand, NO inhibits nitrogen fixation and carbon and nitrogen metabolism; on the other hand, it regulates cellular redox status and maintains the energy state under low oxygen levels. A thin line with + indicate the activation, induction, and retention effects of NO; a thin line with - indicate the inhibition of NO. The oval thick line arrows indicate the main metabolic pathways of NO. Explosive type diagrams refer to enzymes from plants and bacterial symbionts, and lightning type diagram represents genes within rhizobium. ACO: Aconitic acid; CS: Citrate synthase; Gln: Glutamine; Glu: Glutamic acid; GS: Glutamine synthetase; GSH: Glutathione; GSHS: Glutathione synthetase; GSNO: S-nitrosoglu- tathione; Hb: Hemoglobin; IDH: Isocitrate dehydrogenase; MDH: Malate dehydrogenase; NH4+: Ammonium ion; Nif: Nitrogenase; SDH: Succinate dehydrogenase; γ-EC: γ-glu- tamylcysteine; γ-ECS: γ-glutamyl cysteine synthetase ...
... 研究表明, 外源添加NO可抑制大豆与百脉根根瘤的固氮酶活性(Kato et al., 2010; Cam et al., 2012; Boscari et al., 2013).Cam等(2012)报道, 成熟根瘤中NO的积累不仅可降低固氮效率, 还会影响植物的生长适应性, 而通过降解NO可增强植物体固氮能力和延长植物整体固氮周期.研究发现, hmp的过表达降低了根瘤中NO的含量, 却增强了乙炔还原活性(acetylene-reducing activity, ARA)和延迟根瘤衰老(Meilhoc et al., 2011; Cam et al., 2012).但NO的毒性、信号功能或代谢效应取决于其所在作用部位的NO浓度(Boscari et al., 2013; Mur et al., 2013).例如, 在百脉根中, 高浓度的NO可抑制氮的固定, 适当低浓度的NO则可增强固氮作用(Kato et al., 2010).因此, 较低且稳定浓度的NO是维持豆科植物根瘤形成和发育的重要因素(Shimoda et al., 2005; Cam et al., 2012; Calvo-Begueria et al., 2018). ...
... ).但NO的毒性、信号功能或代谢效应取决于其所在作用部位的NO浓度(Boscari et al., 2013; Mur et al., 2013).例如, 在百脉根中, 高浓度的NO可抑制氮的固定, 适当低浓度的NO则可增强固氮作用(Kato et al., 2010).因此, 较低且稳定浓度的NO是维持豆科植物根瘤形成和发育的重要因素(Shimoda et al., 2005; Cam et al., 2012; Calvo-Begueria et al., 2018). ...
... NO可通过调节共生体的能量代谢影响N2固定.固氮过程中会产生铵根离子(ammonium ion, NH4+), 当其达到一定浓度时即具有毒性并损害植物的生长(Li et al., 2014).GS可同化NH4+, 解除这种离子胁迫, 而NO通过Tyr硝化改变其活性, 因此在N代谢中NO与GS起着至关重要的作用(Melo et al., 2011; Sainz et al., 2015).S-亚硝基化也是植物体内NO信号转导的关键机制.研究发现蒺藜苜蓿成熟根瘤中约有80种S-亚硝基化蛋白, 且多数与C、N代谢及固氮酶有关, 如果糖激酶、固氮酶钼铁(MoFe)蛋白nifK与铁(Fe)蛋白nifH (Puppo et al., 2013).NO可能通过影响根瘤中与S-亚硝基化蛋白相关的酶活性, 利用三羧酸循环(tricarboxylic acid cycle, TCA cycle)和糖酵解(glycolysis)等共生体的能量代谢过程调控固氮酶活性(Igamberdiev and Hill, 2009; Boscari et al., 2013; Li et al., 2014; Igamberdiev et al., 2014) (图2).但对于NO究竟是直接抑制固氮酶活性还是通过间接调节C、N代谢影响豆科植物体固氮, 目前尚不清楚(Hichri et al., 2016b). ...
The root hair "infectome" of Medicago truncatula uncovers changes in cell cycle genes and reveals a requirement for auxin signaling in rhizobial infection 1 2014
Redefining nitric oxide production in legume nodules through complementary insights from electron paramagnetic resonance spectroscopy and specific fluorescent probes 2 2018
... 植物和根瘤菌内NO的来源丰富, 主要通过硝酸还原酶(nitrate reductase, NR)和一氧化氮合酶(nitric oxide synthase, NOS)介导以及线粒体电子传递链(electron transport chain, ETC)的还原产生.在植物体内, NO既可由质外体中的亚硝酸根离子(NO2-)转化(Bethke et al., 2004), 又可通过以精氨酸(arginine)、多胺(polyamines)或羟胺(hydroxylamine)作为底物的氧化途径产生(Hichri et al., 2015).在低氧环境下, 还可在还原途径中通过质膜结合亚硝酸盐, 然后通过NO还原酶(nitric oxide reductase, Nor)和黄嘌呤氧化还原酶(xanthine oxidoreductase, XOR)或利用ETC的作用将NO2-还原为NO (Gupta et al., 2011; Mur et al., 2013; Hichri et al., 2015).而Horchani等(2011)发现, 植物及其共生菌中的NR和ETC均可产生NO, 其中NR在植物体内的主要功能是将硝酸盐转化为亚硝酸盐以同化N2 (Neill et al., 2008).但早期的研究发现, 在NAD(P)H作为电子供体时, 大豆(Glycine max)中的NR可将亚硝酸盐转化为NO (Dean and Harper, 1988).且NR抑制剂钨酸盐(tungstate, Tg)也能够抑制NO的产生, 表明NO的合成依赖于NR活性, 但并不是由其直接产生(Horchani et al., 2011).在类菌体中, NO则主要产生于反硝化途径(Hichri et al., 2015), 该途径涉及NR、亚硝酸还原酶(nitrite reductase, NiR)、Nor和NOS (Horchani et al., 2011; Sánchez et al., 2011; Calvo-Begueria et al., 2018) (图1).Berger等(2018)发现NOS也是植物NO的来源, 但目前关于共生菌内NO产生途径的机制研究仍较少. ...
... 研究表明, 外源添加NO可抑制大豆与百脉根根瘤的固氮酶活性(Kato et al., 2010; Cam et al., 2012; Boscari et al., 2013).Cam等(2012)报道, 成熟根瘤中NO的积累不仅可降低固氮效率, 还会影响植物的生长适应性, 而通过降解NO可增强植物体固氮能力和延长植物整体固氮周期.研究发现, hmp的过表达降低了根瘤中NO的含量, 却增强了乙炔还原活性(acetylene-reducing activity, ARA)和延迟根瘤衰老(Meilhoc et al., 2011; Cam et al., 2012).但NO的毒性、信号功能或代谢效应取决于其所在作用部位的NO浓度(Boscari et al., 2013; Mur et al., 2013).例如, 在百脉根中, 高浓度的NO可抑制氮的固定, 适当低浓度的NO则可增强固氮作用(Kato et al., 2010).因此, 较低且稳定浓度的NO是维持豆科植物根瘤形成和发育的重要因素(Shimoda et al., 2005; Cam et al., 2012; Calvo-Begueria et al., 2018). ...
Nitric oxide (NO): a key player in the senescence of Medicago truncatula root nodules 12 2012
... 豆科植物根瘤菌共生体系受多种信号分子和转录因子调控, 其中一氧化氮(nitric oxide, NO)信号分子在豆科植物微生物互作过程中发挥重要作用.共生体早期的转录组学分析表明, NO可调节细胞脱分化和器官发生(Ferrarini et al., 2008; Boscari et al., 2013), 并抑制植物防御反应(Gonzalez-Rizzo et al., 2006), 在建立植物与菌共生关系中发挥重要调控作用(Hichri et al., 2016b).而在成熟根瘤中, NO既可抑制植物体固氮(Shimoda et al., 2005; Kato et al., 2010; Cam et al., 2012), 又可诱发根瘤衰老(Horchani et al., 2011; Cam et al., 2012; Blanquet et al., 2015), 亦可作为能量代谢调节器维持低氧(oxygen, O2)状态下的能量平衡(Kato et al., 2010; Hichri et al., 2016b).尽管目前关于NO在植物体内的生理功能已进行了充分研究, 但国内对于其在豆科植物根瘤菌共生体系中的作用机制并未进行系统的阐述.本文综述了豆科植物根瘤菌共生体系中NO的产生和降解, 及其对根瘤形成与共生固氮影响的研究进展, 并对NO在共生体系中对非生物胁迫的响应及其与激素的相互作用进行了阐述. ...
... ; Cam et al., 2012; Blanquet et al., 2015), 亦可作为能量代谢调节器维持低氧(oxygen, O2)状态下的能量平衡(Kato et al., 2010; Hichri et al., 2016b).尽管目前关于NO在植物体内的生理功能已进行了充分研究, 但国内对于其在豆科植物根瘤菌共生体系中的作用机制并未进行系统的阐述.本文综述了豆科植物根瘤菌共生体系中NO的产生和降解, 及其对根瘤形成与共生固氮影响的研究进展, 并对NO在共生体系中对非生物胁迫的响应及其与激素的相互作用进行了阐述. ...
... 豆科植物根瘤中包含一种与其它蛋白性质和功能不同的血红蛋白(hemoglobins, Hbs), 能够清除机体产生或累积的NO (Gupta et al., 2011).基于其序列同源性和对氧的亲和力, 可将植物体内的Hbs划分成3类: 非共生血红蛋白(non-symbiotic hemoglobins, ns-Hbs, 1类)、豆血红蛋白(leghemoglobins, Lbs, 2类)和截短血红蛋白(truncated hemoglobin, Tr-Hbs, 3类) (Gupta et al., 2011; Hill, 2012).Lbs作为高等植物中发现的第1种豆血红蛋白, 在豆科植物中主要负责将根瘤中的O2运输至根瘤菌(Trevaskis et al., 1997).而Hbs对O2和NO的亲和力非常强, 能够有效清除O2和NO并将其转化为硝酸盐.有研究表明, Hbs能够维持植物细胞在缺氧状态下的氧化还原能力和能量状态(Igamberdiev and Hill, 2009).除植物体蛋白外, 根瘤菌内也含有3类血红蛋白: 黄素血红蛋白(flavohaemoglobins, fHbs/Hmp)、单域血红蛋白(single-domain haemoglobins, sd-Hbs)和截短血红蛋白(Sánchez et al., 2011).Hmp是蒺藜苜蓿(Medicago truncatula)中主要的NO清除剂, 而NO可诱导hmp基因表达以影响自身生成量(Meilhoc et al., 2011; Cam et al., 2012) (图1).另有研究表明, 在蒺藜苜蓿hmp突变的根瘤中, NO水平显著高于野生型, 且固氮效率降低和根瘤衰老提前(Cam et al., 2012; Hichri et al., 2016a).因此, 植物中的Hbs与菌内的Hmp对于维持共生体NO水平至关重要. ...
... 突变的根瘤中, NO水平显著高于野生型, 且固氮效率降低和根瘤衰老提前(Cam et al., 2012; Hichri et al., 2016a).因此, 植物中的Hbs与菌内的Hmp对于维持共生体NO水平至关重要. ...
... NO在影响相关基因的表达、调控豆科植物防御反应进而促进共生和结瘤中发挥重要作用.转录组分析表明, NO影响多种植物基因的表达, 如编码富含半胱氨酸蛋白(cysteine-rich proteins, NCRs)、核糖体蛋白、肽酶、结瘤发育细胞周期蛋白及细胞周期转换蛋白的基因(Vinardell et al., 2003; del Giudice et al., 2011; Boscari et al., 2013), 进而调控细胞脱分化、诱导组织形成及抑制植物防御反应(Hichri et al., 2016b).LjHb1是百脉根(Lotus japonicus) ns-Hbs的编码基因, 外源添加NO、缺氧和低温均可诱导其表达(Shimoda et al., 2005).在根瘤形成的早期阶段, NO的产生可以上调LjHb1的表达, 反之LjHb1的表达又下调NO的水平, 以降低植物防御反应, 促进根瘤菌和宿主植物建立共生关系(Nagata et al., 2008; Murakami et al., 2011).植物也可通过降低Hb的表达促进内源NO产生, 调控自身机体内相关防御基因的表达(Wally et al., 2013) (图2).而NO的清除会引起基因MtCRE1和MtCCS52A的下调表达, 从而延缓结瘤(del Giudice et al., 2011).通过对接种中华根瘤菌的蒺藜苜蓿外源添加NO清除剂(2-(4-carboxyphenyl)- 4,4,5,5-tetramethyl imidazoline-1-oxyl-3-oxide, cPTIO), 发现受NO调节的3种基因(Contig525、Contig3307和Medtr5g010350)表达下调(Boscari et al., 2013).其中, 前2种基因分别与ns-Hbs和茉莉酮酸代谢途径(Palmieri et al., 2008)相关, 而最后1种基因可调控还原型谷胱甘肽(glutathione, GSH)的合成(Innocenti et al., 2007).转录组分析显示, 蒺藜苜蓿接种中华根瘤菌后, 外源添加cPTIO或NADPH氧化酶抑制剂二苯烯碘铵(diphenylene iodide, DPI), 导致细胞壁形成和发育过程中相关基因下调表达, 而植物防御和二次代谢相关基因上调表达(Puppo et al., 2013).hmp的过表达和外源添加cPTIO都可降低蒺藜苜蓿的结瘤效率(Kato et al., 2010; del Giudice et al., 2011), NO则通过调节上述基因的表达, 抑制相关防御反应从而促进共生关系的建立.此外, 在根瘤菌内也鉴定出数百个依赖NO表达的基因, 如中华根瘤菌中编码Hmp的hmp基因(del Giudice et al., 2011; Cam et al., 2012)、编码haeme-Cu蛋白的nnrSI和nnrS2基因(Blanquet et al., 2015)及编码NiR的nirKV基因(Meilhoc et al., 2011), 但关于根瘤菌内受NO影响且与共生结瘤相关的基因报道很少. ...
... 研究表明, 外源添加NO可抑制大豆与百脉根根瘤的固氮酶活性(Kato et al., 2010; Cam et al., 2012; Boscari et al., 2013).Cam等(2012)报道, 成熟根瘤中NO的积累不仅可降低固氮效率, 还会影响植物的生长适应性, 而通过降解NO可增强植物体固氮能力和延长植物整体固氮周期.研究发现, hmp的过表达降低了根瘤中NO的含量, 却增强了乙炔还原活性(acetylene-reducing activity, ARA)和延迟根瘤衰老(Meilhoc et al., 2011; Cam et al., 2012).但NO的毒性、信号功能或代谢效应取决于其所在作用部位的NO浓度(Boscari et al., 2013; Mur et al., 2013).例如, 在百脉根中, 高浓度的NO可抑制氮的固定, 适当低浓度的NO则可增强固氮作用(Kato et al., 2010).因此, 较低且稳定浓度的NO是维持豆科植物根瘤形成和发育的重要因素(Shimoda et al., 2005; Cam et al., 2012; Calvo-Begueria et al., 2018). ...
... ).Cam等(2012)报道, 成熟根瘤中NO的积累不仅可降低固氮效率, 还会影响植物的生长适应性, 而通过降解NO可增强植物体固氮能力和延长植物整体固氮周期.研究发现, hmp的过表达降低了根瘤中NO的含量, 却增强了乙炔还原活性(acetylene-reducing activity, ARA)和延迟根瘤衰老(Meilhoc et al., 2011; Cam et al., 2012).但NO的毒性、信号功能或代谢效应取决于其所在作用部位的NO浓度(Boscari et al., 2013; Mur et al., 2013).例如, 在百脉根中, 高浓度的NO可抑制氮的固定, 适当低浓度的NO则可增强固氮作用(Kato et al., 2010).因此, 较低且稳定浓度的NO是维持豆科植物根瘤形成和发育的重要因素(Shimoda et al., 2005; Cam et al., 2012; Calvo-Begueria et al., 2018). ...
... ; Cam et al., 2012).但NO的毒性、信号功能或代谢效应取决于其所在作用部位的NO浓度(Boscari et al., 2013; Mur et al., 2013).例如, 在百脉根中, 高浓度的NO可抑制氮的固定, 适当低浓度的NO则可增强固氮作用(Kato et al., 2010).因此, 较低且稳定浓度的NO是维持豆科植物根瘤形成和发育的重要因素(Shimoda et al., 2005; Cam et al., 2012; Calvo-Begueria et al., 2018). ...
... ; Cam et al., 2012; Calvo-Begueria et al., 2018). ...
... 尽管目前对于豆科植物与根瘤菌共生固氮的初始过程已有较多研究, 但因发育或应激引起根瘤衰老的分子机制却报道较少.根瘤衰老区域的表型是由粉红色变为绿色, 其实质源于植物Hbs的降解; 在转录水平上, 衰老表现为编码蛋白质降解, 核酸、膜脂和糖类的相关基因被激活; 同时伴有衰老根瘤中氧化还原平衡的改变, 以及抗氧化防御的整体减弱等现象(Van de Velde et al., 2006; Loscos et al., 2008).NO在植物的衰老过程中具有重要作用(Procházková and Wilhelmová, 2011).例如, NO是水稻(Oryza sativa)光依赖性叶细胞死亡的关键介质(Lin et al., 2012); 蒺藜苜蓿根瘤内NO含量的增加会导致根瘤过早衰老, 且衰老程度与NO水平相关(Cam et al., 2012; Meilhoc et al., 2013; Blanquet et al., 2015), 表明NO对于维持共生体系和避免根瘤过早衰老极为重要.此外, 施加硝酸盐可导致豌豆表现出典型的衰老特征(Escuredo et al., 1996).对蒺藜苜蓿添加硝酸盐后, 其自身也可通过还原硝酸盐促使根瘤中生成NO, 然而硝酸盐引起的根瘤衰老是否依赖于NO目前尚不清楚(Horchani et al., 2011).此外, NO也是一种有效的呼吸抑制剂, 能够抑制线粒体和细菌末端的呼吸氧化酶, 并增加活性氧(reactive oxygen species, ROS)和活性氮(reactive nitrogen species, RNS)的积累, 而NO和ROS均为共生发育所必需(Shimoda et al., 2005; Cam et al., 2012; Igamberdiev et al., 2014; Arjona et al., 2015), 因此衰老也可能是固氮时呼吸抑制的间接结果. ...
... ; Cam et al., 2012; Igamberdiev et al., 2014; Arjona et al., 2015), 因此衰老也可能是固氮时呼吸抑制的间接结果. ...
... 研究表明, 长期黑暗处理能显著诱导根瘤衰老, 同时降低固氮酶活性和Hbs水平(Romanov et al., 1980; Matamoros et al., 1999; Swaraj et al., 2001; Pérez Guerra et al., 2010).持续的暗处理会干扰根瘤功能, 导致根瘤完全衰老(Pérez-Guerra et al., 2010), 而通过降解根瘤中的NO可以延缓黑暗诱导的衰老, 但关于NO在根瘤衰老过程中的作用机制还需进一步研究(Cam et al., 2012).She等(2004)发现在光/暗条件下,对蚕豆(Vicia faba)外源添加cPTIO和L-NAME可减弱H2O2对NO产生的诱导效应和改变人为光照引起的气孔关闭.气孔关闭由多种生物和非生物因素引起, 如渗透胁迫、黑暗、高浓度的CO2和机械压力, 其中光是调节气孔运动最重要的环境因素之一(Zeiger, 1983; Kearns and Assmann, 1993; Herold and Puppo, 2005; Sánchez et al., 2010).但关于黑暗条件下NO调节豆科植物共生固氮的作用机制还未见报道. ...
Inactivation of PYR/PYL/RCAR ABA receptors by tyrosine nitration may enable rapid inhibition of ABA signaling by nitric oxide in plants 1 2015
... 脱落酸(abscisic acid, ABA)是植物适应环境胁迫的关键激素(Yoshida et al., 2015), 可通过抑制根毛变形及侵染线的形成影响豆科植物结瘤(Suzuki et al., 2004; Ding et al., 2008).Tominaga等(2010)在百脉根中发现, ABA不仅可调节根瘤的固氮酶活性, 还可通过影响结瘤过程中NO的产生来影响结瘤.在根瘤形成过程中, NO通过酪氨酸(tyrosine, Tyr)硝化调控ABA信号传递, 而ABA又在信号转导过程中决定NO的产生与积累(Castillo et al., 2015).Tyr硝化是NO介导的翻译后修饰, 其特征在于将-NO2添加到Tyr残基芳环的等价邻位C上, 产生3-硝基酪氨酸(3-nitrotyrosine, 3-NT), 并由此改变蛋白质构象(Radi, 2004).作为硝化分子的前体, NO通过Tyr硝化改变细胞溶质谷氨酰胺合成酶(glutamine synthetase 1, GS1)和Lb等关键结瘤蛋白的活性(Melo et al., 2011; Navascués et al., 2012; Li et al., 2014).豆科植物结瘤时Lb发生Tyr硝化, 可保护共生体免受过氧亚硝酸盐毒害, 因此3-NT的形成既是硝基氧化应激的标志, 也是功能性结瘤活跃代谢的结果(Sainz et al., 2015).此外, NOS抑制剂N-硝基-L-精氨酸甲酯(NG-nitro- L-arginine methyl ester, L-NAME)和cPTIO处理可减弱豌豆(Pisum sativum)表皮由ABA诱导的气孔关闭(张绪成等, 2005). ...
An ecological role of fungal endophytes to ameliorate plants under biotic stress 1 2015
... NO不仅能调节植物生长发育, 还可参与植物对非生物胁迫的响应(Moreau et al., 2010).目前的研究发现, 在不同非生物胁迫(如干旱和缺氧)下, 共生有机体中的NO均会产生响应(Desalvo et al., 2010; Sánchez et al., 2011; Hawkins et al., 2014; Iarullina et al., 2014; Zimmer-Prados et al., 2014).同时, 在豆科植物中NO还可能参与调控机体对金属元素镉(Cd)和铝(Al)的胁迫反应以及应对非金属元素砷(As)的胁迫(Singh et al., 2015; 尚玉婷等, 2018), 但都缺乏在结瘤和共生固氮体系中的实验证据(Pérez- Chaca et al., 2014).Meilhoc等(2011)提出, 宿主中任何应激反应下NO的产生都会直接影响共生, 在成熟根瘤中, 除了NO专用调节器NnrR外, 多数根瘤菌的基因通过双组分系统(FixLJ)来调节NO, 其中FixLJ是O2限制反应的主要调节因子.nnrR是Nor结构基因norCB的上游基因, 可防止NO这种高活性氮氧化物的积累, 其插入失活可抑制亚硝酸盐的产生以及亚硝酸盐和NO的还原(De Bruijn et al., 2006).NO还能够可逆地结合光系统II (photosystem II, PSII)并调节电子的转移和猝灭, 对光合作用至关重要(Wodala et al., 2008).从农业和进化的角度来看, NO在植物与根瘤菌共生体系中的调控作用, 可以改善植物对病原体的防御状态、保护植物免受虫害以及增强植物对非生物胁迫的适应性(Chadha et al., 2015). ...
Computational prediction of candidate proteins for S-nitrosylation in Arabidopsis thaliana 1 2014
... 根瘤中的NO可通过调控酶的翻译后修饰调节碳(C)、氮(N)及能量代谢(Chaki et al., 2014; Hu et al., 2015).NO的产生是植物应对缺氧胁迫的一种响应, 与呼吸循环过程相关联, 形成“Hb/NO”循环.缺氧条件下, Hb/NO循环可维持细胞内的氧化还原和能量平衡状态(Igamberdiev and Hill, 2004; Gupta et al., 2011).Hb/NO呼吸循环包括4个步骤: (1) 硝酸根离子(nitrate ion, NO3-)被NR还原为NO2-; (2) NO2-从细胞质转移到线粒体基质; (3) 通过线粒体ETC将NO2-还原为NO, 使ATP再生; (4) 最终NO通过被动扩散到胞质溶胶中, 由Hb将其氧化成硝酸盐(Gupta et al., 2011; Horchani et al., 2011; Hichri et al., 2015).因此, 在低氧环境下, 植物线粒体可以保持其氧化NADH的能力, 从而合成ATP.相关研究也表明, 缺氧环境下, 大豆和苜蓿的成熟根瘤通过硝酸还原途径和细菌反硝化途径均能产生NO, 参与Hb/NO呼吸循环, 为机体提供能量(Meakin et al., 2007; Horchani et al., 2011; Sánchez et al., 2011).此外, 在缺氧条件下, 结瘤的能量状态几乎全部取决于NR的功能(Horchani et al., 2011).值得注意的是, 在Hb/NO循环过程中, 由于多数情况下处于缺氧状态, 因此最终的电子受体是亚硝酸盐而非O2 (Gupta et al., 2011; Igamberdiev et al., 2014).Hichri等(2015)发现, NO在结瘤过程中具有双重作用: 一方面作为维持基础能量代谢所必需的中间物; 另一方面作为C、N代谢的调节剂, 降低微氧环境下的能量需求.由于缺氧对于植物的生长发育影响很大, 因此当O2不足时, NO对维持豆科植物与根瘤菌共生结瘤的氧化还原和能量平衡状态至关重要. ...
Presence of nitric oxide synthase activity in roots and nodules of Lupinus albus 1 1996
A highly conserved Sinorhizobium meliloti operon is induced microaerobically via the FixLJ system and by nitric oxide (NO) via NnrR 2 2006
... 共生体内NO的降解还受相关基因及其它蛋白调控.苜蓿中华根瘤菌(Sinorhizobium meliloti)中包含1个与短链脱氢酶相关且位于nnrR下游的nnrS基因家族, nnrS的表达依赖于转录调节器NnrR (De Bruijn et al., 2006).当nnrS1突变时, 根瘤内NO的水平升高(Blanquet et al., 2015).NnrS是一种haeme-Cu膜蛋白(Bartnikas et al., 2002), NnrS家族的2种蛋白(NnrS1和NnrS2)和Nor能够直接或间接参与NO的降解并有效地维持共生固氮(Meilhoc et al., 2013; Blanquet et al., 2015). ...
... NO不仅能调节植物生长发育, 还可参与植物对非生物胁迫的响应(Moreau et al., 2010).目前的研究发现, 在不同非生物胁迫(如干旱和缺氧)下, 共生有机体中的NO均会产生响应(Desalvo et al., 2010; Sánchez et al., 2011; Hawkins et al., 2014; Iarullina et al., 2014; Zimmer-Prados et al., 2014).同时, 在豆科植物中NO还可能参与调控机体对金属元素镉(Cd)和铝(Al)的胁迫反应以及应对非金属元素砷(As)的胁迫(Singh et al., 2015; 尚玉婷等, 2018), 但都缺乏在结瘤和共生固氮体系中的实验证据(Pérez- Chaca et al., 2014).Meilhoc等(2011)提出, 宿主中任何应激反应下NO的产生都会直接影响共生, 在成熟根瘤中, 除了NO专用调节器NnrR外, 多数根瘤菌的基因通过双组分系统(FixLJ)来调节NO, 其中FixLJ是O2限制反应的主要调节因子.nnrR是Nor结构基因norCB的上游基因, 可防止NO这种高活性氮氧化物的积累, 其插入失活可抑制亚硝酸盐的产生以及亚硝酸盐和NO的还原(De Bruijn et al., 2006).NO还能够可逆地结合光系统II (photosystem II, PSII)并调节电子的转移和猝灭, 对光合作用至关重要(Wodala et al., 2008).从农业和进化的角度来看, NO在植物与根瘤菌共生体系中的调控作用, 可以改善植物对病原体的防御状态、保护植物免受虫害以及增强植物对非生物胁迫的适应性(Chadha et al., 2015). ...
The conversion of nitrite to nitrogen oxide(s) by the constitutive NAD(P)H-nitrate reductase enzyme from soybean 1 1988
... 植物和根瘤菌内NO的来源丰富, 主要通过硝酸还原酶(nitrate reductase, NR)和一氧化氮合酶(nitric oxide synthase, NOS)介导以及线粒体电子传递链(electron transport chain, ETC)的还原产生.在植物体内, NO既可由质外体中的亚硝酸根离子(NO2-)转化(Bethke et al., 2004), 又可通过以精氨酸(arginine)、多胺(polyamines)或羟胺(hydroxylamine)作为底物的氧化途径产生(Hichri et al., 2015).在低氧环境下, 还可在还原途径中通过质膜结合亚硝酸盐, 然后通过NO还原酶(nitric oxide reductase, Nor)和黄嘌呤氧化还原酶(xanthine oxidoreductase, XOR)或利用ETC的作用将NO2-还原为NO (Gupta et al., 2011; Mur et al., 2013; Hichri et al., 2015).而Horchani等(2011)发现, 植物及其共生菌中的NR和ETC均可产生NO, 其中NR在植物体内的主要功能是将硝酸盐转化为亚硝酸盐以同化N2 (Neill et al., 2008).但早期的研究发现, 在NAD(P)H作为电子供体时, 大豆(Glycine max)中的NR可将亚硝酸盐转化为NO (Dean and Harper, 1988).且NR抑制剂钨酸盐(tungstate, Tg)也能够抑制NO的产生, 表明NO的合成依赖于NR活性, 但并不是由其直接产生(Horchani et al., 2011).在类菌体中, NO则主要产生于反硝化途径(Hichri et al., 2015), 该途径涉及NR、亚硝酸还原酶(nitrite reductase, NiR)、Nor和NOS (Horchani et al., 2011; Sánchez et al., 2011; Calvo-Begueria et al., 2018) (图1).Berger等(2018)发现NOS也是植物NO的来源, 但目前关于共生菌内NO产生途径的机制研究仍较少. ...
Nitric oxide is required for an optimal establishment of the Medicago truncatula-Sinorhizobium meliloti symbiosis 5 2011
... NO在影响相关基因的表达、调控豆科植物防御反应进而促进共生和结瘤中发挥重要作用.转录组分析表明, NO影响多种植物基因的表达, 如编码富含半胱氨酸蛋白(cysteine-rich proteins, NCRs)、核糖体蛋白、肽酶、结瘤发育细胞周期蛋白及细胞周期转换蛋白的基因(Vinardell et al., 2003; del Giudice et al., 2011; Boscari et al., 2013), 进而调控细胞脱分化、诱导组织形成及抑制植物防御反应(Hichri et al., 2016b).LjHb1是百脉根(Lotus japonicus) ns-Hbs的编码基因, 外源添加NO、缺氧和低温均可诱导其表达(Shimoda et al., 2005).在根瘤形成的早期阶段, NO的产生可以上调LjHb1的表达, 反之LjHb1的表达又下调NO的水平, 以降低植物防御反应, 促进根瘤菌和宿主植物建立共生关系(Nagata et al., 2008; Murakami et al., 2011).植物也可通过降低Hb的表达促进内源NO产生, 调控自身机体内相关防御基因的表达(Wally et al., 2013) (图2).而NO的清除会引起基因MtCRE1和MtCCS52A的下调表达, 从而延缓结瘤(del Giudice et al., 2011).通过对接种中华根瘤菌的蒺藜苜蓿外源添加NO清除剂(2-(4-carboxyphenyl)- 4,4,5,5-tetramethyl imidazoline-1-oxyl-3-oxide, cPTIO), 发现受NO调节的3种基因(Contig525、Contig3307和Medtr5g010350)表达下调(Boscari et al., 2013).其中, 前2种基因分别与ns-Hbs和茉莉酮酸代谢途径(Palmieri et al., 2008)相关, 而最后1种基因可调控还原型谷胱甘肽(glutathione, GSH)的合成(Innocenti et al., 2007).转录组分析显示, 蒺藜苜蓿接种中华根瘤菌后, 外源添加cPTIO或NADPH氧化酶抑制剂二苯烯碘铵(diphenylene iodide, DPI), 导致细胞壁形成和发育过程中相关基因下调表达, 而植物防御和二次代谢相关基因上调表达(Puppo et al., 2013).hmp的过表达和外源添加cPTIO都可降低蒺藜苜蓿的结瘤效率(Kato et al., 2010; del Giudice et al., 2011), NO则通过调节上述基因的表达, 抑制相关防御反应从而促进共生关系的建立.此外, 在根瘤菌内也鉴定出数百个依赖NO表达的基因, 如中华根瘤菌中编码Hmp的hmp基因(del Giudice et al., 2011; Cam et al., 2012)、编码haeme-Cu蛋白的nnrSI和nnrS2基因(Blanquet et al., 2015)及编码NiR的nirKV基因(Meilhoc et al., 2011), 但关于根瘤菌内受NO影响且与共生结瘤相关的基因报道很少. ...
... 的下调表达, 从而延缓结瘤(del Giudice et al., 2011).通过对接种中华根瘤菌的蒺藜苜蓿外源添加NO清除剂(2-(4-carboxyphenyl)- 4,4,5,5-tetramethyl imidazoline-1-oxyl-3-oxide, cPTIO), 发现受NO调节的3种基因(Contig525、Contig3307和Medtr5g010350)表达下调(Boscari et al., 2013).其中, 前2种基因分别与ns-Hbs和茉莉酮酸代谢途径(Palmieri et al., 2008)相关, 而最后1种基因可调控还原型谷胱甘肽(glutathione, GSH)的合成(Innocenti et al., 2007).转录组分析显示, 蒺藜苜蓿接种中华根瘤菌后, 外源添加cPTIO或NADPH氧化酶抑制剂二苯烯碘铵(diphenylene iodide, DPI), 导致细胞壁形成和发育过程中相关基因下调表达, 而植物防御和二次代谢相关基因上调表达(Puppo et al., 2013).hmp的过表达和外源添加cPTIO都可降低蒺藜苜蓿的结瘤效率(Kato et al., 2010; del Giudice et al., 2011), NO则通过调节上述基因的表达, 抑制相关防御反应从而促进共生关系的建立.此外, 在根瘤菌内也鉴定出数百个依赖NO表达的基因, 如中华根瘤菌中编码Hmp的hmp基因(del Giudice et al., 2011; Cam et al., 2012)、编码haeme-Cu蛋白的nnrSI和nnrS2基因(Blanquet et al., 2015)及编码NiR的nirKV基因(Meilhoc et al., 2011), 但关于根瘤菌内受NO影响且与共生结瘤相关的基因报道很少. ...
... ; del Giudice et al., 2011), NO则通过调节上述基因的表达, 抑制相关防御反应从而促进共生关系的建立.此外, 在根瘤菌内也鉴定出数百个依赖NO表达的基因, 如中华根瘤菌中编码Hmp的hmp基因(del Giudice et al., 2011; Cam et al., 2012)、编码haeme-Cu蛋白的nnrSI和nnrS2基因(Blanquet et al., 2015)及编码NiR的nirKV基因(Meilhoc et al., 2011), 但关于根瘤菌内受NO影响且与共生结瘤相关的基因报道很少. ...
... 基因(del Giudice et al., 2011; Cam et al., 2012)、编码haeme-Cu蛋白的nnrSI和nnrS2基因(Blanquet et al., 2015)及编码NiR的nirKV基因(Meilhoc et al., 2011), 但关于根瘤菌内受NO影响且与共生结瘤相关的基因报道很少. ...
Transcriptomic responses to heat stress and bleaching in the Elkhorn coral Acropora palmata 1 2010
... NO不仅能调节植物生长发育, 还可参与植物对非生物胁迫的响应(Moreau et al., 2010).目前的研究发现, 在不同非生物胁迫(如干旱和缺氧)下, 共生有机体中的NO均会产生响应(Desalvo et al., 2010; Sánchez et al., 2011; Hawkins et al., 2014; Iarullina et al., 2014; Zimmer-Prados et al., 2014).同时, 在豆科植物中NO还可能参与调控机体对金属元素镉(Cd)和铝(Al)的胁迫反应以及应对非金属元素砷(As)的胁迫(Singh et al., 2015; 尚玉婷等, 2018), 但都缺乏在结瘤和共生固氮体系中的实验证据(Pérez- Chaca et al., 2014).Meilhoc等(2011)提出, 宿主中任何应激反应下NO的产生都会直接影响共生, 在成熟根瘤中, 除了NO专用调节器NnrR外, 多数根瘤菌的基因通过双组分系统(FixLJ)来调节NO, 其中FixLJ是O2限制反应的主要调节因子.nnrR是Nor结构基因norCB的上游基因, 可防止NO这种高活性氮氧化物的积累, 其插入失活可抑制亚硝酸盐的产生以及亚硝酸盐和NO的还原(De Bruijn et al., 2006).NO还能够可逆地结合光系统II (photosystem II, PSII)并调节电子的转移和猝灭, 对光合作用至关重要(Wodala et al., 2008).从农业和进化的角度来看, NO在植物与根瘤菌共生体系中的调控作用, 可以改善植物对病原体的防御状态、保护植物免受虫害以及增强植物对非生物胁迫的适应性(Chadha et al., 2015). ...
Abscisic acid coordinates nod factor and cytokinin signaling during the regulation of nodulation in Medicago truncatula 1 2008
... 脱落酸(abscisic acid, ABA)是植物适应环境胁迫的关键激素(Yoshida et al., 2015), 可通过抑制根毛变形及侵染线的形成影响豆科植物结瘤(Suzuki et al., 2004; Ding et al., 2008).Tominaga等(2010)在百脉根中发现, ABA不仅可调节根瘤的固氮酶活性, 还可通过影响结瘤过程中NO的产生来影响结瘤.在根瘤形成过程中, NO通过酪氨酸(tyrosine, Tyr)硝化调控ABA信号传递, 而ABA又在信号转导过程中决定NO的产生与积累(Castillo et al., 2015).Tyr硝化是NO介导的翻译后修饰, 其特征在于将-NO2添加到Tyr残基芳环的等价邻位C上, 产生3-硝基酪氨酸(3-nitrotyrosine, 3-NT), 并由此改变蛋白质构象(Radi, 2004).作为硝化分子的前体, NO通过Tyr硝化改变细胞溶质谷氨酰胺合成酶(glutamine synthetase 1, GS1)和Lb等关键结瘤蛋白的活性(Melo et al., 2011; Navascués et al., 2012; Li et al., 2014).豆科植物结瘤时Lb发生Tyr硝化, 可保护共生体免受过氧亚硝酸盐毒害, 因此3-NT的形成既是硝基氧化应激的标志, 也是功能性结瘤活跃代谢的结果(Sainz et al., 2015).此外, NOS抑制剂N-硝基-L-精氨酸甲酯(NG-nitro- L-arginine methyl ester, L-NAME)和cPTIO处理可减弱豌豆(Pisum sativum)表皮由ABA诱导的气孔关闭(张绪成等, 2005). ...
Involvement of activated oxygen in nitrate-induced senescence of pea root nodules 1 1996
... 尽管目前对于豆科植物与根瘤菌共生固氮的初始过程已有较多研究, 但因发育或应激引起根瘤衰老的分子机制却报道较少.根瘤衰老区域的表型是由粉红色变为绿色, 其实质源于植物Hbs的降解; 在转录水平上, 衰老表现为编码蛋白质降解, 核酸、膜脂和糖类的相关基因被激活; 同时伴有衰老根瘤中氧化还原平衡的改变, 以及抗氧化防御的整体减弱等现象(Van de Velde et al., 2006; Loscos et al., 2008).NO在植物的衰老过程中具有重要作用(Procházková and Wilhelmová, 2011).例如, NO是水稻(Oryza sativa)光依赖性叶细胞死亡的关键介质(Lin et al., 2012); 蒺藜苜蓿根瘤内NO含量的增加会导致根瘤过早衰老, 且衰老程度与NO水平相关(Cam et al., 2012; Meilhoc et al., 2013; Blanquet et al., 2015), 表明NO对于维持共生体系和避免根瘤过早衰老极为重要.此外, 施加硝酸盐可导致豌豆表现出典型的衰老特征(Escuredo et al., 1996).对蒺藜苜蓿添加硝酸盐后, 其自身也可通过还原硝酸盐促使根瘤中生成NO, 然而硝酸盐引起的根瘤衰老是否依赖于NO目前尚不清楚(Horchani et al., 2011).此外, NO也是一种有效的呼吸抑制剂, 能够抑制线粒体和细菌末端的呼吸氧化酶, 并增加活性氧(reactive oxygen species, ROS)和活性氮(reactive nitrogen species, RNS)的积累, 而NO和ROS均为共生发育所必需(Shimoda et al., 2005; Cam et al., 2012; Igamberdiev et al., 2014; Arjona et al., 2015), 因此衰老也可能是固氮时呼吸抑制的间接结果. ...
Expression of Medicago truncatula genes responsive to nitric oxide in pathogenic and symbiotic conditions 1 2008
... 豆科植物根瘤菌共生体系受多种信号分子和转录因子调控, 其中一氧化氮(nitric oxide, NO)信号分子在豆科植物微生物互作过程中发挥重要作用.共生体早期的转录组学分析表明, NO可调节细胞脱分化和器官发生(Ferrarini et al., 2008; Boscari et al., 2013), 并抑制植物防御反应(Gonzalez-Rizzo et al., 2006), 在建立植物与菌共生关系中发挥重要调控作用(Hichri et al., 2016b).而在成熟根瘤中, NO既可抑制植物体固氮(Shimoda et al., 2005; Kato et al., 2010; Cam et al., 2012), 又可诱发根瘤衰老(Horchani et al., 2011; Cam et al., 2012; Blanquet et al., 2015), 亦可作为能量代谢调节器维持低氧(oxygen, O2)状态下的能量平衡(Kato et al., 2010; Hichri et al., 2016b).尽管目前关于NO在植物体内的生理功能已进行了充分研究, 但国内对于其在豆科植物根瘤菌共生体系中的作用机制并未进行系统的阐述.本文综述了豆科植物根瘤菌共生体系中NO的产生和降解, 及其对根瘤形成与共生固氮影响的研究进展, 并对NO在共生体系中对非生物胁迫的响应及其与激素的相互作用进行了阐述. ...
The Medicago truncatula CRE1 cytokinin receptor regulates lateral root development and early symbiotic interaction with Sinorhizobium meliloti 1 2006
... 豆科植物根瘤菌共生体系受多种信号分子和转录因子调控, 其中一氧化氮(nitric oxide, NO)信号分子在豆科植物微生物互作过程中发挥重要作用.共生体早期的转录组学分析表明, NO可调节细胞脱分化和器官发生(Ferrarini et al., 2008; Boscari et al., 2013), 并抑制植物防御反应(Gonzalez-Rizzo et al., 2006), 在建立植物与菌共生关系中发挥重要调控作用(Hichri et al., 2016b).而在成熟根瘤中, NO既可抑制植物体固氮(Shimoda et al., 2005; Kato et al., 2010; Cam et al., 2012), 又可诱发根瘤衰老(Horchani et al., 2011; Cam et al., 2012; Blanquet et al., 2015), 亦可作为能量代谢调节器维持低氧(oxygen, O2)状态下的能量平衡(Kato et al., 2010; Hichri et al., 2016b).尽管目前关于NO在植物体内的生理功能已进行了充分研究, 但国内对于其在豆科植物根瘤菌共生体系中的作用机制并未进行系统的阐述.本文综述了豆科植物根瘤菌共生体系中NO的产生和降解, 及其对根瘤形成与共生固氮影响的研究进展, 并对NO在共生体系中对非生物胁迫的响应及其与激素的相互作用进行了阐述. ...
Plant hemoglobins: important players at the crossroads between oxygen and nitric oxide 6 2011
... 植物和根瘤菌内NO的来源丰富, 主要通过硝酸还原酶(nitrate reductase, NR)和一氧化氮合酶(nitric oxide synthase, NOS)介导以及线粒体电子传递链(electron transport chain, ETC)的还原产生.在植物体内, NO既可由质外体中的亚硝酸根离子(NO2-)转化(Bethke et al., 2004), 又可通过以精氨酸(arginine)、多胺(polyamines)或羟胺(hydroxylamine)作为底物的氧化途径产生(Hichri et al., 2015).在低氧环境下, 还可在还原途径中通过质膜结合亚硝酸盐, 然后通过NO还原酶(nitric oxide reductase, Nor)和黄嘌呤氧化还原酶(xanthine oxidoreductase, XOR)或利用ETC的作用将NO2-还原为NO (Gupta et al., 2011; Mur et al., 2013; Hichri et al., 2015).而Horchani等(2011)发现, 植物及其共生菌中的NR和ETC均可产生NO, 其中NR在植物体内的主要功能是将硝酸盐转化为亚硝酸盐以同化N2 (Neill et al., 2008).但早期的研究发现, 在NAD(P)H作为电子供体时, 大豆(Glycine max)中的NR可将亚硝酸盐转化为NO (Dean and Harper, 1988).且NR抑制剂钨酸盐(tungstate, Tg)也能够抑制NO的产生, 表明NO的合成依赖于NR活性, 但并不是由其直接产生(Horchani et al., 2011).在类菌体中, NO则主要产生于反硝化途径(Hichri et al., 2015), 该途径涉及NR、亚硝酸还原酶(nitrite reductase, NiR)、Nor和NOS (Horchani et al., 2011; Sánchez et al., 2011; Calvo-Begueria et al., 2018) (图1).Berger等(2018)发现NOS也是植物NO的来源, 但目前关于共生菌内NO产生途径的机制研究仍较少. ...
... 豆科植物根瘤中包含一种与其它蛋白性质和功能不同的血红蛋白(hemoglobins, Hbs), 能够清除机体产生或累积的NO (Gupta et al., 2011).基于其序列同源性和对氧的亲和力, 可将植物体内的Hbs划分成3类: 非共生血红蛋白(non-symbiotic hemoglobins, ns-Hbs, 1类)、豆血红蛋白(leghemoglobins, Lbs, 2类)和截短血红蛋白(truncated hemoglobin, Tr-Hbs, 3类) (Gupta et al., 2011; Hill, 2012).Lbs作为高等植物中发现的第1种豆血红蛋白, 在豆科植物中主要负责将根瘤中的O2运输至根瘤菌(Trevaskis et al., 1997).而Hbs对O2和NO的亲和力非常强, 能够有效清除O2和NO并将其转化为硝酸盐.有研究表明, Hbs能够维持植物细胞在缺氧状态下的氧化还原能力和能量状态(Igamberdiev and Hill, 2009).除植物体蛋白外, 根瘤菌内也含有3类血红蛋白: 黄素血红蛋白(flavohaemoglobins, fHbs/Hmp)、单域血红蛋白(single-domain haemoglobins, sd-Hbs)和截短血红蛋白(Sánchez et al., 2011).Hmp是蒺藜苜蓿(Medicago truncatula)中主要的NO清除剂, 而NO可诱导hmp基因表达以影响自身生成量(Meilhoc et al., 2011; Cam et al., 2012) (图1).另有研究表明, 在蒺藜苜蓿hmp突变的根瘤中, NO水平显著高于野生型, 且固氮效率降低和根瘤衰老提前(Cam et al., 2012; Hichri et al., 2016a).因此, 植物中的Hbs与菌内的Hmp对于维持共生体NO水平至关重要. ...
... ).基于其序列同源性和对氧的亲和力, 可将植物体内的Hbs划分成3类: 非共生血红蛋白(non-symbiotic hemoglobins, ns-Hbs, 1类)、豆血红蛋白(leghemoglobins, Lbs, 2类)和截短血红蛋白(truncated hemoglobin, Tr-Hbs, 3类) (Gupta et al., 2011; Hill, 2012).Lbs作为高等植物中发现的第1种豆血红蛋白, 在豆科植物中主要负责将根瘤中的O2运输至根瘤菌(Trevaskis et al., 1997).而Hbs对O2和NO的亲和力非常强, 能够有效清除O2和NO并将其转化为硝酸盐.有研究表明, Hbs能够维持植物细胞在缺氧状态下的氧化还原能力和能量状态(Igamberdiev and Hill, 2009).除植物体蛋白外, 根瘤菌内也含有3类血红蛋白: 黄素血红蛋白(flavohaemoglobins, fHbs/Hmp)、单域血红蛋白(single-domain haemoglobins, sd-Hbs)和截短血红蛋白(Sánchez et al., 2011).Hmp是蒺藜苜蓿(Medicago truncatula)中主要的NO清除剂, 而NO可诱导hmp基因表达以影响自身生成量(Meilhoc et al., 2011; Cam et al., 2012) (图1).另有研究表明, 在蒺藜苜蓿hmp突变的根瘤中, NO水平显著高于野生型, 且固氮效率降低和根瘤衰老提前(Cam et al., 2012; Hichri et al., 2016a).因此, 植物中的Hbs与菌内的Hmp对于维持共生体NO水平至关重要. ...
... 根瘤中的NO可通过调控酶的翻译后修饰调节碳(C)、氮(N)及能量代谢(Chaki et al., 2014; Hu et al., 2015).NO的产生是植物应对缺氧胁迫的一种响应, 与呼吸循环过程相关联, 形成“Hb/NO”循环.缺氧条件下, Hb/NO循环可维持细胞内的氧化还原和能量平衡状态(Igamberdiev and Hill, 2004; Gupta et al., 2011).Hb/NO呼吸循环包括4个步骤: (1) 硝酸根离子(nitrate ion, NO3-)被NR还原为NO2-; (2) NO2-从细胞质转移到线粒体基质; (3) 通过线粒体ETC将NO2-还原为NO, 使ATP再生; (4) 最终NO通过被动扩散到胞质溶胶中, 由Hb将其氧化成硝酸盐(Gupta et al., 2011; Horchani et al., 2011; Hichri et al., 2015).因此, 在低氧环境下, 植物线粒体可以保持其氧化NADH的能力, 从而合成ATP.相关研究也表明, 缺氧环境下, 大豆和苜蓿的成熟根瘤通过硝酸还原途径和细菌反硝化途径均能产生NO, 参与Hb/NO呼吸循环, 为机体提供能量(Meakin et al., 2007; Horchani et al., 2011; Sánchez et al., 2011).此外, 在缺氧条件下, 结瘤的能量状态几乎全部取决于NR的功能(Horchani et al., 2011).值得注意的是, 在Hb/NO循环过程中, 由于多数情况下处于缺氧状态, 因此最终的电子受体是亚硝酸盐而非O2 (Gupta et al., 2011; Igamberdiev et al., 2014).Hichri等(2015)发现, NO在结瘤过程中具有双重作用: 一方面作为维持基础能量代谢所必需的中间物; 另一方面作为C、N代谢的调节剂, 降低微氧环境下的能量需求.由于缺氧对于植物的生长发育影响很大, 因此当O2不足时, NO对维持豆科植物与根瘤菌共生结瘤的氧化还原和能量平衡状态至关重要. ...
... 还原为NO, 使ATP再生; (4) 最终NO通过被动扩散到胞质溶胶中, 由Hb将其氧化成硝酸盐(Gupta et al., 2011; Horchani et al., 2011; Hichri et al., 2015).因此, 在低氧环境下, 植物线粒体可以保持其氧化NADH的能力, 从而合成ATP.相关研究也表明, 缺氧环境下, 大豆和苜蓿的成熟根瘤通过硝酸还原途径和细菌反硝化途径均能产生NO, 参与Hb/NO呼吸循环, 为机体提供能量(Meakin et al., 2007; Horchani et al., 2011; Sánchez et al., 2011).此外, 在缺氧条件下, 结瘤的能量状态几乎全部取决于NR的功能(Horchani et al., 2011).值得注意的是, 在Hb/NO循环过程中, 由于多数情况下处于缺氧状态, 因此最终的电子受体是亚硝酸盐而非O2 (Gupta et al., 2011; Igamberdiev et al., 2014).Hichri等(2015)发现, NO在结瘤过程中具有双重作用: 一方面作为维持基础能量代谢所必需的中间物; 另一方面作为C、N代谢的调节剂, 降低微氧环境下的能量需求.由于缺氧对于植物的生长发育影响很大, 因此当O2不足时, NO对维持豆科植物与根瘤菌共生结瘤的氧化还原和能量平衡状态至关重要. ...
... (Gupta et al., 2011; Igamberdiev et al., 2014).Hichri等(2015)发现, NO在结瘤过程中具有双重作用: 一方面作为维持基础能量代谢所必需的中间物; 另一方面作为C、N代谢的调节剂, 降低微氧环境下的能量需求.由于缺氧对于植物的生长发育影响很大, 因此当O2不足时, NO对维持豆科植物与根瘤菌共生结瘤的氧化还原和能量平衡状态至关重要. ...
Differential nitric oxide synthesis and host apoptotic events correlate with bleaching susceptibility in reef corals 1 2014
... NO不仅能调节植物生长发育, 还可参与植物对非生物胁迫的响应(Moreau et al., 2010).目前的研究发现, 在不同非生物胁迫(如干旱和缺氧)下, 共生有机体中的NO均会产生响应(Desalvo et al., 2010; Sánchez et al., 2011; Hawkins et al., 2014; Iarullina et al., 2014; Zimmer-Prados et al., 2014).同时, 在豆科植物中NO还可能参与调控机体对金属元素镉(Cd)和铝(Al)的胁迫反应以及应对非金属元素砷(As)的胁迫(Singh et al., 2015; 尚玉婷等, 2018), 但都缺乏在结瘤和共生固氮体系中的实验证据(Pérez- Chaca et al., 2014).Meilhoc等(2011)提出, 宿主中任何应激反应下NO的产生都会直接影响共生, 在成熟根瘤中, 除了NO专用调节器NnrR外, 多数根瘤菌的基因通过双组分系统(FixLJ)来调节NO, 其中FixLJ是O2限制反应的主要调节因子.nnrR是Nor结构基因norCB的上游基因, 可防止NO这种高活性氮氧化物的积累, 其插入失活可抑制亚硝酸盐的产生以及亚硝酸盐和NO的还原(De Bruijn et al., 2006).NO还能够可逆地结合光系统II (photosystem II, PSII)并调节电子的转移和猝灭, 对光合作用至关重要(Wodala et al., 2008).从农业和进化的角度来看, NO在植物与根瘤菌共生体系中的调控作用, 可以改善植物对病原体的防御状态、保护植物免受虫害以及增强植物对非生物胁迫的适应性(Chadha et al., 2015). ...
Oxyleghemoglobin scavenges nitrogen monoxide and peroxynitrite: a possible role in functioning nodules 1 2005
... 研究表明, 长期黑暗处理能显著诱导根瘤衰老, 同时降低固氮酶活性和Hbs水平(Romanov et al., 1980; Matamoros et al., 1999; Swaraj et al., 2001; Pérez Guerra et al., 2010).持续的暗处理会干扰根瘤功能, 导致根瘤完全衰老(Pérez-Guerra et al., 2010), 而通过降解根瘤中的NO可以延缓黑暗诱导的衰老, 但关于NO在根瘤衰老过程中的作用机制还需进一步研究(Cam et al., 2012).She等(2004)发现在光/暗条件下,对蚕豆(Vicia faba)外源添加cPTIO和L-NAME可减弱H2O2对NO产生的诱导效应和改变人为光照引起的气孔关闭.气孔关闭由多种生物和非生物因素引起, 如渗透胁迫、黑暗、高浓度的CO2和机械压力, 其中光是调节气孔运动最重要的环境因素之一(Zeiger, 1983; Kearns and Assmann, 1993; Herold and Puppo, 2005; Sánchez et al., 2010).但关于黑暗条件下NO调节豆科植物共生固氮的作用机制还未见报道. ...
Nitric oxide: a multifaceted regulator of the nitrogen-fixing symbiosis 9 2015
... 植物和根瘤菌内NO的来源丰富, 主要通过硝酸还原酶(nitrate reductase, NR)和一氧化氮合酶(nitric oxide synthase, NOS)介导以及线粒体电子传递链(electron transport chain, ETC)的还原产生.在植物体内, NO既可由质外体中的亚硝酸根离子(NO2-)转化(Bethke et al., 2004), 又可通过以精氨酸(arginine)、多胺(polyamines)或羟胺(hydroxylamine)作为底物的氧化途径产生(Hichri et al., 2015).在低氧环境下, 还可在还原途径中通过质膜结合亚硝酸盐, 然后通过NO还原酶(nitric oxide reductase, Nor)和黄嘌呤氧化还原酶(xanthine oxidoreductase, XOR)或利用ETC的作用将NO2-还原为NO (Gupta et al., 2011; Mur et al., 2013; Hichri et al., 2015).而Horchani等(2011)发现, 植物及其共生菌中的NR和ETC均可产生NO, 其中NR在植物体内的主要功能是将硝酸盐转化为亚硝酸盐以同化N2 (Neill et al., 2008).但早期的研究发现, 在NAD(P)H作为电子供体时, 大豆(Glycine max)中的NR可将亚硝酸盐转化为NO (Dean and Harper, 1988).且NR抑制剂钨酸盐(tungstate, Tg)也能够抑制NO的产生, 表明NO的合成依赖于NR活性, 但并不是由其直接产生(Horchani et al., 2011).在类菌体中, NO则主要产生于反硝化途径(Hichri et al., 2015), 该途径涉及NR、亚硝酸还原酶(nitrite reductase, NiR)、Nor和NOS (Horchani et al., 2011; Sánchez et al., 2011; Calvo-Begueria et al., 2018) (图1).Berger等(2018)发现NOS也是植物NO的来源, 但目前关于共生菌内NO产生途径的机制研究仍较少. ...
... ; Hichri et al., 2015).而Horchani等(2011)发现, 植物及其共生菌中的NR和ETC均可产生NO, 其中NR在植物体内的主要功能是将硝酸盐转化为亚硝酸盐以同化N2 (Neill et al., 2008).但早期的研究发现, 在NAD(P)H作为电子供体时, 大豆(Glycine max)中的NR可将亚硝酸盐转化为NO (Dean and Harper, 1988).且NR抑制剂钨酸盐(tungstate, Tg)也能够抑制NO的产生, 表明NO的合成依赖于NR活性, 但并不是由其直接产生(Horchani et al., 2011).在类菌体中, NO则主要产生于反硝化途径(Hichri et al., 2015), 该途径涉及NR、亚硝酸还原酶(nitrite reductase, NiR)、Nor和NOS (Horchani et al., 2011; Sánchez et al., 2011; Calvo-Begueria et al., 2018) (图1).Berger等(2018)发现NOS也是植物NO的来源, 但目前关于共生菌内NO产生途径的机制研究仍较少. ...
... ).在类菌体中, NO则主要产生于反硝化途径(Hichri et al., 2015), 该途径涉及NR、亚硝酸还原酶(nitrite reductase, NiR)、Nor和NOS (Horchani et al., 2011; Sánchez et al., 2011; Calvo-Begueria et al., 2018) (图1).Berger等(2018)发现NOS也是植物NO的来源, 但目前关于共生菌内NO产生途径的机制研究仍较少. ...
... NO在影响相关基因的表达、调控豆科植物防御反应进而促进共生和结瘤中发挥重要作用.转录组分析表明, NO影响多种植物基因的表达, 如编码富含半胱氨酸蛋白(cysteine-rich proteins, NCRs)、核糖体蛋白、肽酶、结瘤发育细胞周期蛋白及细胞周期转换蛋白的基因(Vinardell et al., 2003; del Giudice et al., 2011; Boscari et al., 2013), 进而调控细胞脱分化、诱导组织形成及抑制植物防御反应(Hichri et al., 2016b).LjHb1是百脉根(Lotus japonicus) ns-Hbs的编码基因, 外源添加NO、缺氧和低温均可诱导其表达(Shimoda et al., 2005).在根瘤形成的早期阶段, NO的产生可以上调LjHb1的表达, 反之LjHb1的表达又下调NO的水平, 以降低植物防御反应, 促进根瘤菌和宿主植物建立共生关系(Nagata et al., 2008; Murakami et al., 2011).植物也可通过降低Hb的表达促进内源NO产生, 调控自身机体内相关防御基因的表达(Wally et al., 2013) (图2).而NO的清除会引起基因MtCRE1和MtCCS52A的下调表达, 从而延缓结瘤(del Giudice et al., 2011).通过对接种中华根瘤菌的蒺藜苜蓿外源添加NO清除剂(2-(4-carboxyphenyl)- 4,4,5,5-tetramethyl imidazoline-1-oxyl-3-oxide, cPTIO), 发现受NO调节的3种基因(Contig525、Contig3307和Medtr5g010350)表达下调(Boscari et al., 2013).其中, 前2种基因分别与ns-Hbs和茉莉酮酸代谢途径(Palmieri et al., 2008)相关, 而最后1种基因可调控还原型谷胱甘肽(glutathione, GSH)的合成(Innocenti et al., 2007).转录组分析显示, 蒺藜苜蓿接种中华根瘤菌后, 外源添加cPTIO或NADPH氧化酶抑制剂二苯烯碘铵(diphenylene iodide, DPI), 导致细胞壁形成和发育过程中相关基因下调表达, 而植物防御和二次代谢相关基因上调表达(Puppo et al., 2013).hmp的过表达和外源添加cPTIO都可降低蒺藜苜蓿的结瘤效率(Kato et al., 2010; del Giudice et al., 2011), NO则通过调节上述基因的表达, 抑制相关防御反应从而促进共生关系的建立.此外, 在根瘤菌内也鉴定出数百个依赖NO表达的基因, 如中华根瘤菌中编码Hmp的hmp基因(del Giudice et al., 2011; Cam et al., 2012)、编码haeme-Cu蛋白的nnrSI和nnrS2基因(Blanquet et al., 2015)及编码NiR的nirKV基因(Meilhoc et al., 2011), 但关于根瘤菌内受NO影响且与共生结瘤相关的基因报道很少. 10.11983/CBB20034.F002
Schematic diagram of the role of NO in symbiotic nitrogen fixation (modified from <xref ref-type="bibr" rid="b11">Boscari et al., 2013</xref>; <xref ref-type="bibr" rid="b31">Hichri et al., 2015</xref>, <xref ref-type="bibr" rid="b33">2016b</xref>)On the one hand, NO inhibits nitrogen fixation and carbon and nitrogen metabolism; on the other hand, it regulates cellular redox status and maintains the energy state under low oxygen levels. A thin line with + indicate the activation, induction, and retention effects of NO; a thin line with - indicate the inhibition of NO. The oval thick line arrows indicate the main metabolic pathways of NO. Explosive type diagrams refer to enzymes from plants and bacterial symbionts, and lightning type diagram represents genes within rhizobium. ACO: Aconitic acid; CS: Citrate synthase; Gln: Glutamine; Glu: Glutamic acid; GS: Glutamine synthetase; GSH: Glutathione; GSHS: Glutathione synthetase; GSNO: S-nitrosoglu- tathione; Hb: Hemoglobin; IDH: Isocitrate dehydrogenase; MDH: Malate dehydrogenase; NH4+: Ammonium ion; Nif: Nitrogenase; SDH: Succinate dehydrogenase; γ-EC: γ-glu- tamylcysteine; γ-ECS: γ-glutamyl cysteine synthetase ...
3 2016
... 植物和根瘤菌内NO的来源丰富, 主要通过硝酸还原酶(nitrate reductase, NR)和一氧化氮合酶(nitric oxide synthase, NOS)介导以及线粒体电子传递链(electron transport chain, ETC)的还原产生.在植物体内, NO既可由质外体中的亚硝酸根离子(NO2-)转化(Bethke et al., 2004), 又可通过以精氨酸(arginine)、多胺(polyamines)或羟胺(hydroxylamine)作为底物的氧化途径产生(Hichri et al., 2015).在低氧环境下, 还可在还原途径中通过质膜结合亚硝酸盐, 然后通过NO还原酶(nitric oxide reductase, Nor)和黄嘌呤氧化还原酶(xanthine oxidoreductase, XOR)或利用ETC的作用将NO2-还原为NO (Gupta et al., 2011; Mur et al., 2013; Hichri et al., 2015).而Horchani等(2011)发现, 植物及其共生菌中的NR和ETC均可产生NO, 其中NR在植物体内的主要功能是将硝酸盐转化为亚硝酸盐以同化N2 (Neill et al., 2008).但早期的研究发现, 在NAD(P)H作为电子供体时, 大豆(Glycine max)中的NR可将亚硝酸盐转化为NO (Dean and Harper, 1988).且NR抑制剂钨酸盐(tungstate, Tg)也能够抑制NO的产生, 表明NO的合成依赖于NR活性, 但并不是由其直接产生(Horchani et al., 2011).在类菌体中, NO则主要产生于反硝化途径(Hichri et al., 2015), 该途径涉及NR、亚硝酸还原酶(nitrite reductase, NiR)、Nor和NOS (Horchani et al., 2011; Sánchez et al., 2011; Calvo-Begueria et al., 2018) (图1).Berger等(2018)发现NOS也是植物NO的来源, 但目前关于共生菌内NO产生途径的机制研究仍较少. 10.11983/CBB20034.F001
The schematic diagram of NO production and degradation in the symbiotic system (modified from <xref ref-type="bibr" rid="b32">Hichri et al., 2016a</xref>)The figure contains the upper and lower parts, which correspond to the production and degradation of NO from plants and symbiotes, respectively. The seven horns star diagrams refer to the oxidation pathway and the oval diagrams refer to the reduction pathway. The lines indicate that studies have been confirmed, and the dashed lines indicate that it is yet to be studied. ETC: Mitochondrial electron transport chain; GSNOR: S-nitrosoglutathione reductase; Hmp: Flavin hemoglobin; Lb: Leghemoglobin; NnrS: Haem- and copper-con- taining membrane protein; Nor: NO reductase; NOS: NO synthase; ns-Hb: Nonsymbiotic hemoglobin; NR: Nitrate reductase; PAOX: Polyamine oxidase; sd-Hb: Single domain hemoglobin; Tr-Hb: Truncated hemoglobin; TrxR: Thioredoxin reduction enzymes; XOR: Xanthine oxidoreductase ...
... 豆科植物根瘤中包含一种与其它蛋白性质和功能不同的血红蛋白(hemoglobins, Hbs), 能够清除机体产生或累积的NO (Gupta et al., 2011).基于其序列同源性和对氧的亲和力, 可将植物体内的Hbs划分成3类: 非共生血红蛋白(non-symbiotic hemoglobins, ns-Hbs, 1类)、豆血红蛋白(leghemoglobins, Lbs, 2类)和截短血红蛋白(truncated hemoglobin, Tr-Hbs, 3类) (Gupta et al., 2011; Hill, 2012).Lbs作为高等植物中发现的第1种豆血红蛋白, 在豆科植物中主要负责将根瘤中的O2运输至根瘤菌(Trevaskis et al., 1997).而Hbs对O2和NO的亲和力非常强, 能够有效清除O2和NO并将其转化为硝酸盐.有研究表明, Hbs能够维持植物细胞在缺氧状态下的氧化还原能力和能量状态(Igamberdiev and Hill, 2009).除植物体蛋白外, 根瘤菌内也含有3类血红蛋白: 黄素血红蛋白(flavohaemoglobins, fHbs/Hmp)、单域血红蛋白(single-domain haemoglobins, sd-Hbs)和截短血红蛋白(Sánchez et al., 2011).Hmp是蒺藜苜蓿(Medicago truncatula)中主要的NO清除剂, 而NO可诱导hmp基因表达以影响自身生成量(Meilhoc et al., 2011; Cam et al., 2012) (图1).另有研究表明, 在蒺藜苜蓿hmp突变的根瘤中, NO水平显著高于野生型, 且固氮效率降低和根瘤衰老提前(Cam et al., 2012; Hichri et al., 2016a).因此, 植物中的Hbs与菌内的Hmp对于维持共生体NO水平至关重要. ...
Nitric oxide: jack-of-all-trades of the nitrogen-fixing symbiosis? 8 2016
... 豆科植物根瘤菌共生体系受多种信号分子和转录因子调控, 其中一氧化氮(nitric oxide, NO)信号分子在豆科植物微生物互作过程中发挥重要作用.共生体早期的转录组学分析表明, NO可调节细胞脱分化和器官发生(Ferrarini et al., 2008; Boscari et al., 2013), 并抑制植物防御反应(Gonzalez-Rizzo et al., 2006), 在建立植物与菌共生关系中发挥重要调控作用(Hichri et al., 2016b).而在成熟根瘤中, NO既可抑制植物体固氮(Shimoda et al., 2005; Kato et al., 2010; Cam et al., 2012), 又可诱发根瘤衰老(Horchani et al., 2011; Cam et al., 2012; Blanquet et al., 2015), 亦可作为能量代谢调节器维持低氧(oxygen, O2)状态下的能量平衡(Kato et al., 2010; Hichri et al., 2016b).尽管目前关于NO在植物体内的生理功能已进行了充分研究, 但国内对于其在豆科植物根瘤菌共生体系中的作用机制并未进行系统的阐述.本文综述了豆科植物根瘤菌共生体系中NO的产生和降解, 及其对根瘤形成与共生固氮影响的研究进展, 并对NO在共生体系中对非生物胁迫的响应及其与激素的相互作用进行了阐述. ...
... ; Hichri et al., 2016b).尽管目前关于NO在植物体内的生理功能已进行了充分研究, 但国内对于其在豆科植物根瘤菌共生体系中的作用机制并未进行系统的阐述.本文综述了豆科植物根瘤菌共生体系中NO的产生和降解, 及其对根瘤形成与共生固氮影响的研究进展, 并对NO在共生体系中对非生物胁迫的响应及其与激素的相互作用进行了阐述. ...
... 除根瘤菌内的血红蛋白以外, 还有其它细菌蛋白调控根瘤中的NO水平.例如, 硫氧还蛋白(thioredoxin reductase, TrxR)或亚硝基谷胱甘肽还原酶(S-nitroglutathione reductase, GSNOR)都可能间接参与NO的降解和信号传递过程(Lee et al., 2010).而细菌的NO降解蛋白不仅能保护其自身蛋白免受NO的负面影响, 还能使共生植物蛋白免受NO介导的失活(Blanquet et al., 2015; Hichri et al., 2016b).综上, 鉴于NO的影响, 共生菌可能需要更多不同的系统来降低自身的NO水平. ...
... NO在影响相关基因的表达、调控豆科植物防御反应进而促进共生和结瘤中发挥重要作用.转录组分析表明, NO影响多种植物基因的表达, 如编码富含半胱氨酸蛋白(cysteine-rich proteins, NCRs)、核糖体蛋白、肽酶、结瘤发育细胞周期蛋白及细胞周期转换蛋白的基因(Vinardell et al., 2003; del Giudice et al., 2011; Boscari et al., 2013), 进而调控细胞脱分化、诱导组织形成及抑制植物防御反应(Hichri et al., 2016b).LjHb1是百脉根(Lotus japonicus) ns-Hbs的编码基因, 外源添加NO、缺氧和低温均可诱导其表达(Shimoda et al., 2005).在根瘤形成的早期阶段, NO的产生可以上调LjHb1的表达, 反之LjHb1的表达又下调NO的水平, 以降低植物防御反应, 促进根瘤菌和宿主植物建立共生关系(Nagata et al., 2008; Murakami et al., 2011).植物也可通过降低Hb的表达促进内源NO产生, 调控自身机体内相关防御基因的表达(Wally et al., 2013) (图2).而NO的清除会引起基因MtCRE1和MtCCS52A的下调表达, 从而延缓结瘤(del Giudice et al., 2011).通过对接种中华根瘤菌的蒺藜苜蓿外源添加NO清除剂(2-(4-carboxyphenyl)- 4,4,5,5-tetramethyl imidazoline-1-oxyl-3-oxide, cPTIO), 发现受NO调节的3种基因(Contig525、Contig3307和Medtr5g010350)表达下调(Boscari et al., 2013).其中, 前2种基因分别与ns-Hbs和茉莉酮酸代谢途径(Palmieri et al., 2008)相关, 而最后1种基因可调控还原型谷胱甘肽(glutathione, GSH)的合成(Innocenti et al., 2007).转录组分析显示, 蒺藜苜蓿接种中华根瘤菌后, 外源添加cPTIO或NADPH氧化酶抑制剂二苯烯碘铵(diphenylene iodide, DPI), 导致细胞壁形成和发育过程中相关基因下调表达, 而植物防御和二次代谢相关基因上调表达(Puppo et al., 2013).hmp的过表达和外源添加cPTIO都可降低蒺藜苜蓿的结瘤效率(Kato et al., 2010; del Giudice et al., 2011), NO则通过调节上述基因的表达, 抑制相关防御反应从而促进共生关系的建立.此外, 在根瘤菌内也鉴定出数百个依赖NO表达的基因, 如中华根瘤菌中编码Hmp的hmp基因(del Giudice et al., 2011; Cam et al., 2012)、编码haeme-Cu蛋白的nnrSI和nnrS2基因(Blanquet et al., 2015)及编码NiR的nirKV基因(Meilhoc et al., 2011), 但关于根瘤菌内受NO影响且与共生结瘤相关的基因报道很少. ...
... 一方面, NO抑制固氮和C、N代谢; 另一方面, NO调控细胞氧化还原和保持低氧水平下的能量状态.带+的细线表示NO的活化、诱导和保持效果; 带有-的细线表示NO的抑制作用.椭圆形粗线箭头表示NO主要的代谢途径.爆炸型图示指来自植物和菌共生体的酶, 闪电型图示表示根瘤菌内的基因.ACO: 乌头酸; CS: 柠檬酸合酶; Gln: 谷氨酰胺; Glu: 谷氨酸; GS: 谷氨酰胺合成酶; GSH: 谷胱甘肽; GSHS: 谷胱甘肽合成酶; GSNO: S-亚硝基谷胱甘肽; Hb: 血红蛋白; IDH: 异柠檬酸脱氢酶; MDH: 苹果酸脱氢酶; NH4+: 铵根离子; Nif: 固氮酶; SDH: 琥珀酸脱氢酶; γ-EC: γ-谷氨酰半胱氨酸; γ-ECS: γ-谷氨酰半胱氨酸合成酶 Schematic diagram of the role of NO in symbiotic nitrogen fixation (modified from <xref ref-type="bibr" rid="b11">Boscari et al., 2013</xref>; <xref ref-type="bibr" rid="b31">Hichri et al., 2015</xref>, <xref ref-type="bibr" rid="b33">2016b</xref>)On the one hand, NO inhibits nitrogen fixation and carbon and nitrogen metabolism; on the other hand, it regulates cellular redox status and maintains the energy state under low oxygen levels. A thin line with + indicate the activation, induction, and retention effects of NO; a thin line with - indicate the inhibition of NO. The oval thick line arrows indicate the main metabolic pathways of NO. Explosive type diagrams refer to enzymes from plants and bacterial symbionts, and lightning type diagram represents genes within rhizobium. ACO: Aconitic acid; CS: Citrate synthase; Gln: Glutamine; Glu: Glutamic acid; GS: Glutamine synthetase; GSH: Glutathione; GSHS: Glutathione synthetase; GSNO: S-nitrosoglu- tathione; Hb: Hemoglobin; IDH: Isocitrate dehydrogenase; MDH: Malate dehydrogenase; NH4+: Ammonium ion; Nif: Nitrogenase; SDH: Succinate dehydrogenase; γ-EC: γ-glu- tamylcysteine; γ-ECS: γ-glutamyl cysteine synthetase ...
... NO可通过调节共生体的能量代谢影响N2固定.固氮过程中会产生铵根离子(ammonium ion, NH4+), 当其达到一定浓度时即具有毒性并损害植物的生长(Li et al., 2014).GS可同化NH4+, 解除这种离子胁迫, 而NO通过Tyr硝化改变其活性, 因此在N代谢中NO与GS起着至关重要的作用(Melo et al., 2011; Sainz et al., 2015).S-亚硝基化也是植物体内NO信号转导的关键机制.研究发现蒺藜苜蓿成熟根瘤中约有80种S-亚硝基化蛋白, 且多数与C、N代谢及固氮酶有关, 如果糖激酶、固氮酶钼铁(MoFe)蛋白nifK与铁(Fe)蛋白nifH (Puppo et al., 2013).NO可能通过影响根瘤中与S-亚硝基化蛋白相关的酶活性, 利用三羧酸循环(tricarboxylic acid cycle, TCA cycle)和糖酵解(glycolysis)等共生体的能量代谢过程调控固氮酶活性(Igamberdiev and Hill, 2009; Boscari et al., 2013; Li et al., 2014; Igamberdiev et al., 2014) (图2).但对于NO究竟是直接抑制固氮酶活性还是通过间接调节C、N代谢影响豆科植物体固氮, 目前尚不清楚(Hichri et al., 2016b). ...
... 豆科植物根瘤中包含一种与其它蛋白性质和功能不同的血红蛋白(hemoglobins, Hbs), 能够清除机体产生或累积的NO (Gupta et al., 2011).基于其序列同源性和对氧的亲和力, 可将植物体内的Hbs划分成3类: 非共生血红蛋白(non-symbiotic hemoglobins, ns-Hbs, 1类)、豆血红蛋白(leghemoglobins, Lbs, 2类)和截短血红蛋白(truncated hemoglobin, Tr-Hbs, 3类) (Gupta et al., 2011; Hill, 2012).Lbs作为高等植物中发现的第1种豆血红蛋白, 在豆科植物中主要负责将根瘤中的O2运输至根瘤菌(Trevaskis et al., 1997).而Hbs对O2和NO的亲和力非常强, 能够有效清除O2和NO并将其转化为硝酸盐.有研究表明, Hbs能够维持植物细胞在缺氧状态下的氧化还原能力和能量状态(Igamberdiev and Hill, 2009).除植物体蛋白外, 根瘤菌内也含有3类血红蛋白: 黄素血红蛋白(flavohaemoglobins, fHbs/Hmp)、单域血红蛋白(single-domain haemoglobins, sd-Hbs)和截短血红蛋白(Sánchez et al., 2011).Hmp是蒺藜苜蓿(Medicago truncatula)中主要的NO清除剂, 而NO可诱导hmp基因表达以影响自身生成量(Meilhoc et al., 2011; Cam et al., 2012) (图1).另有研究表明, 在蒺藜苜蓿hmp突变的根瘤中, NO水平显著高于野生型, 且固氮效率降低和根瘤衰老提前(Cam et al., 2012; Hichri et al., 2016a).因此, 植物中的Hbs与菌内的Hmp对于维持共生体NO水平至关重要. ...
Both plant and bacterial nitrate reductases contribute to nitric oxide production in Medicago truncatula nitrogen-fixing nodules 9 2011
... 豆科植物根瘤菌共生体系受多种信号分子和转录因子调控, 其中一氧化氮(nitric oxide, NO)信号分子在豆科植物微生物互作过程中发挥重要作用.共生体早期的转录组学分析表明, NO可调节细胞脱分化和器官发生(Ferrarini et al., 2008; Boscari et al., 2013), 并抑制植物防御反应(Gonzalez-Rizzo et al., 2006), 在建立植物与菌共生关系中发挥重要调控作用(Hichri et al., 2016b).而在成熟根瘤中, NO既可抑制植物体固氮(Shimoda et al., 2005; Kato et al., 2010; Cam et al., 2012), 又可诱发根瘤衰老(Horchani et al., 2011; Cam et al., 2012; Blanquet et al., 2015), 亦可作为能量代谢调节器维持低氧(oxygen, O2)状态下的能量平衡(Kato et al., 2010; Hichri et al., 2016b).尽管目前关于NO在植物体内的生理功能已进行了充分研究, 但国内对于其在豆科植物根瘤菌共生体系中的作用机制并未进行系统的阐述.本文综述了豆科植物根瘤菌共生体系中NO的产生和降解, 及其对根瘤形成与共生固氮影响的研究进展, 并对NO在共生体系中对非生物胁迫的响应及其与激素的相互作用进行了阐述. ...
... 植物和根瘤菌内NO的来源丰富, 主要通过硝酸还原酶(nitrate reductase, NR)和一氧化氮合酶(nitric oxide synthase, NOS)介导以及线粒体电子传递链(electron transport chain, ETC)的还原产生.在植物体内, NO既可由质外体中的亚硝酸根离子(NO2-)转化(Bethke et al., 2004), 又可通过以精氨酸(arginine)、多胺(polyamines)或羟胺(hydroxylamine)作为底物的氧化途径产生(Hichri et al., 2015).在低氧环境下, 还可在还原途径中通过质膜结合亚硝酸盐, 然后通过NO还原酶(nitric oxide reductase, Nor)和黄嘌呤氧化还原酶(xanthine oxidoreductase, XOR)或利用ETC的作用将NO2-还原为NO (Gupta et al., 2011; Mur et al., 2013; Hichri et al., 2015).而Horchani等(2011)发现, 植物及其共生菌中的NR和ETC均可产生NO, 其中NR在植物体内的主要功能是将硝酸盐转化为亚硝酸盐以同化N2 (Neill et al., 2008).但早期的研究发现, 在NAD(P)H作为电子供体时, 大豆(Glycine max)中的NR可将亚硝酸盐转化为NO (Dean and Harper, 1988).且NR抑制剂钨酸盐(tungstate, Tg)也能够抑制NO的产生, 表明NO的合成依赖于NR活性, 但并不是由其直接产生(Horchani et al., 2011).在类菌体中, NO则主要产生于反硝化途径(Hichri et al., 2015), 该途径涉及NR、亚硝酸还原酶(nitrite reductase, NiR)、Nor和NOS (Horchani et al., 2011; Sánchez et al., 2011; Calvo-Begueria et al., 2018) (图1).Berger等(2018)发现NOS也是植物NO的来源, 但目前关于共生菌内NO产生途径的机制研究仍较少. ...
... ).且NR抑制剂钨酸盐(tungstate, Tg)也能够抑制NO的产生, 表明NO的合成依赖于NR活性, 但并不是由其直接产生(Horchani et al., 2011).在类菌体中, NO则主要产生于反硝化途径(Hichri et al., 2015), 该途径涉及NR、亚硝酸还原酶(nitrite reductase, NiR)、Nor和NOS (Horchani et al., 2011; Sánchez et al., 2011; Calvo-Begueria et al., 2018) (图1).Berger等(2018)发现NOS也是植物NO的来源, 但目前关于共生菌内NO产生途径的机制研究仍较少. ...
... ), 该途径涉及NR、亚硝酸还原酶(nitrite reductase, NiR)、Nor和NOS (Horchani et al., 2011; Sánchez et al., 2011; Calvo-Begueria et al., 2018) (图1).Berger等(2018)发现NOS也是植物NO的来源, 但目前关于共生菌内NO产生途径的机制研究仍较少. ...
... 根瘤中的NO可通过调控酶的翻译后修饰调节碳(C)、氮(N)及能量代谢(Chaki et al., 2014; Hu et al., 2015).NO的产生是植物应对缺氧胁迫的一种响应, 与呼吸循环过程相关联, 形成“Hb/NO”循环.缺氧条件下, Hb/NO循环可维持细胞内的氧化还原和能量平衡状态(Igamberdiev and Hill, 2004; Gupta et al., 2011).Hb/NO呼吸循环包括4个步骤: (1) 硝酸根离子(nitrate ion, NO3-)被NR还原为NO2-; (2) NO2-从细胞质转移到线粒体基质; (3) 通过线粒体ETC将NO2-还原为NO, 使ATP再生; (4) 最终NO通过被动扩散到胞质溶胶中, 由Hb将其氧化成硝酸盐(Gupta et al., 2011; Horchani et al., 2011; Hichri et al., 2015).因此, 在低氧环境下, 植物线粒体可以保持其氧化NADH的能力, 从而合成ATP.相关研究也表明, 缺氧环境下, 大豆和苜蓿的成熟根瘤通过硝酸还原途径和细菌反硝化途径均能产生NO, 参与Hb/NO呼吸循环, 为机体提供能量(Meakin et al., 2007; Horchani et al., 2011; Sánchez et al., 2011).此外, 在缺氧条件下, 结瘤的能量状态几乎全部取决于NR的功能(Horchani et al., 2011).值得注意的是, 在Hb/NO循环过程中, 由于多数情况下处于缺氧状态, 因此最终的电子受体是亚硝酸盐而非O2 (Gupta et al., 2011; Igamberdiev et al., 2014).Hichri等(2015)发现, NO在结瘤过程中具有双重作用: 一方面作为维持基础能量代谢所必需的中间物; 另一方面作为C、N代谢的调节剂, 降低微氧环境下的能量需求.由于缺氧对于植物的生长发育影响很大, 因此当O2不足时, NO对维持豆科植物与根瘤菌共生结瘤的氧化还原和能量平衡状态至关重要. ...
... ; Horchani et al., 2011; Sánchez et al., 2011).此外, 在缺氧条件下, 结瘤的能量状态几乎全部取决于NR的功能(Horchani et al., 2011).值得注意的是, 在Hb/NO循环过程中, 由于多数情况下处于缺氧状态, 因此最终的电子受体是亚硝酸盐而非O2 (Gupta et al., 2011; Igamberdiev et al., 2014).Hichri等(2015)发现, NO在结瘤过程中具有双重作用: 一方面作为维持基础能量代谢所必需的中间物; 另一方面作为C、N代谢的调节剂, 降低微氧环境下的能量需求.由于缺氧对于植物的生长发育影响很大, 因此当O2不足时, NO对维持豆科植物与根瘤菌共生结瘤的氧化还原和能量平衡状态至关重要. ...
... ).此外, 在缺氧条件下, 结瘤的能量状态几乎全部取决于NR的功能(Horchani et al., 2011).值得注意的是, 在Hb/NO循环过程中, 由于多数情况下处于缺氧状态, 因此最终的电子受体是亚硝酸盐而非O2 (Gupta et al., 2011; Igamberdiev et al., 2014).Hichri等(2015)发现, NO在结瘤过程中具有双重作用: 一方面作为维持基础能量代谢所必需的中间物; 另一方面作为C、N代谢的调节剂, 降低微氧环境下的能量需求.由于缺氧对于植物的生长发育影响很大, 因此当O2不足时, NO对维持豆科植物与根瘤菌共生结瘤的氧化还原和能量平衡状态至关重要. ...
... 尽管目前对于豆科植物与根瘤菌共生固氮的初始过程已有较多研究, 但因发育或应激引起根瘤衰老的分子机制却报道较少.根瘤衰老区域的表型是由粉红色变为绿色, 其实质源于植物Hbs的降解; 在转录水平上, 衰老表现为编码蛋白质降解, 核酸、膜脂和糖类的相关基因被激活; 同时伴有衰老根瘤中氧化还原平衡的改变, 以及抗氧化防御的整体减弱等现象(Van de Velde et al., 2006; Loscos et al., 2008).NO在植物的衰老过程中具有重要作用(Procházková and Wilhelmová, 2011).例如, NO是水稻(Oryza sativa)光依赖性叶细胞死亡的关键介质(Lin et al., 2012); 蒺藜苜蓿根瘤内NO含量的增加会导致根瘤过早衰老, 且衰老程度与NO水平相关(Cam et al., 2012; Meilhoc et al., 2013; Blanquet et al., 2015), 表明NO对于维持共生体系和避免根瘤过早衰老极为重要.此外, 施加硝酸盐可导致豌豆表现出典型的衰老特征(Escuredo et al., 1996).对蒺藜苜蓿添加硝酸盐后, 其自身也可通过还原硝酸盐促使根瘤中生成NO, 然而硝酸盐引起的根瘤衰老是否依赖于NO目前尚不清楚(Horchani et al., 2011).此外, NO也是一种有效的呼吸抑制剂, 能够抑制线粒体和细菌末端的呼吸氧化酶, 并增加活性氧(reactive oxygen species, ROS)和活性氮(reactive nitrogen species, RNS)的积累, 而NO和ROS均为共生发育所必需(Shimoda et al., 2005; Cam et al., 2012; Igamberdiev et al., 2014; Arjona et al., 2015), 因此衰老也可能是固氮时呼吸抑制的间接结果. ...
... NO还参与豆科植物对缺氧胁迫的反应.缺氧胁迫可诱导百脉根根瘤产生亚硝酰基豆血红蛋白(nitrosylleghemoglobin, LbNO)以减少根瘤内的硝酸盐, 同时LbNO的形成也有利于解除NO对植物体的毒害(Meakin et al., 2007).根瘤中LbNOs的增加不会抑制植物的固氮作用, 其实质源于Lb在根瘤中既可清除由细菌反硝化作用产生的NO和亚硝酸盐, 又可保护淹水条件下的固氮酶活性(Sánchez et al., 2010).Sánchez等(2010)发现, 在水淹缺氧条件下, 野生型和norC突变体根瘤中编码固氮酶Fe蛋白的nifH基因表达量降低、固氮酶活性下降, 通过清除NO可抵消这种影响.而当从常氧转为缺氧时, 根瘤内NO迅速增加, 表明根瘤中不仅能产生NO, 还具有响应缺氧胁迫而迅速上调表达的基因(Sánchez et al., 2010; Horchani et al., 2011).因此, 无论在转录水平还是翻译水平, 固氮酶都是NO抑制N2固定的关键靶标(Sánchez et al., 2010), 同时也表明在微氧环境中, NO主要通过影响固氮酶活性来调节植物体固氮. ...
Site-speci?c nitrosoproteomic identi?cation of endogenously S-nitrosylated proteins in Arabidopsis 1 2015
... 根瘤中的NO可通过调控酶的翻译后修饰调节碳(C)、氮(N)及能量代谢(Chaki et al., 2014; Hu et al., 2015).NO的产生是植物应对缺氧胁迫的一种响应, 与呼吸循环过程相关联, 形成“Hb/NO”循环.缺氧条件下, Hb/NO循环可维持细胞内的氧化还原和能量平衡状态(Igamberdiev and Hill, 2004; Gupta et al., 2011).Hb/NO呼吸循环包括4个步骤: (1) 硝酸根离子(nitrate ion, NO3-)被NR还原为NO2-; (2) NO2-从细胞质转移到线粒体基质; (3) 通过线粒体ETC将NO2-还原为NO, 使ATP再生; (4) 最终NO通过被动扩散到胞质溶胶中, 由Hb将其氧化成硝酸盐(Gupta et al., 2011; Horchani et al., 2011; Hichri et al., 2015).因此, 在低氧环境下, 植物线粒体可以保持其氧化NADH的能力, 从而合成ATP.相关研究也表明, 缺氧环境下, 大豆和苜蓿的成熟根瘤通过硝酸还原途径和细菌反硝化途径均能产生NO, 参与Hb/NO呼吸循环, 为机体提供能量(Meakin et al., 2007; Horchani et al., 2011; Sánchez et al., 2011).此外, 在缺氧条件下, 结瘤的能量状态几乎全部取决于NR的功能(Horchani et al., 2011).值得注意的是, 在Hb/NO循环过程中, 由于多数情况下处于缺氧状态, 因此最终的电子受体是亚硝酸盐而非O2 (Gupta et al., 2011; Igamberdiev et al., 2014).Hichri等(2015)发现, NO在结瘤过程中具有双重作用: 一方面作为维持基础能量代谢所必需的中间物; 另一方面作为C、N代谢的调节剂, 降低微氧环境下的能量需求.由于缺氧对于植物的生长发育影响很大, 因此当O2不足时, NO对维持豆科植物与根瘤菌共生结瘤的氧化还原和能量平衡状态至关重要. ...
Probiotics for plants: NO-producing lactobacilli protect plants from drought 1 2014
... NO不仅能调节植物生长发育, 还可参与植物对非生物胁迫的响应(Moreau et al., 2010).目前的研究发现, 在不同非生物胁迫(如干旱和缺氧)下, 共生有机体中的NO均会产生响应(Desalvo et al., 2010; Sánchez et al., 2011; Hawkins et al., 2014; Iarullina et al., 2014; Zimmer-Prados et al., 2014).同时, 在豆科植物中NO还可能参与调控机体对金属元素镉(Cd)和铝(Al)的胁迫反应以及应对非金属元素砷(As)的胁迫(Singh et al., 2015; 尚玉婷等, 2018), 但都缺乏在结瘤和共生固氮体系中的实验证据(Pérez- Chaca et al., 2014).Meilhoc等(2011)提出, 宿主中任何应激反应下NO的产生都会直接影响共生, 在成熟根瘤中, 除了NO专用调节器NnrR外, 多数根瘤菌的基因通过双组分系统(FixLJ)来调节NO, 其中FixLJ是O2限制反应的主要调节因子.nnrR是Nor结构基因norCB的上游基因, 可防止NO这种高活性氮氧化物的积累, 其插入失活可抑制亚硝酸盐的产生以及亚硝酸盐和NO的还原(De Bruijn et al., 2006).NO还能够可逆地结合光系统II (photosystem II, PSII)并调节电子的转移和猝灭, 对光合作用至关重要(Wodala et al., 2008).从农业和进化的角度来看, NO在植物与根瘤菌共生体系中的调控作用, 可以改善植物对病原体的防御状态、保护植物免受虫害以及增强植物对非生物胁迫的适应性(Chadha et al., 2015). ...
Nitrate, NO and haemoglobin in plant adaptation to hypoxia: an alternative to classic fermentation pathways 1 2004
... 根瘤中的NO可通过调控酶的翻译后修饰调节碳(C)、氮(N)及能量代谢(Chaki et al., 2014; Hu et al., 2015).NO的产生是植物应对缺氧胁迫的一种响应, 与呼吸循环过程相关联, 形成“Hb/NO”循环.缺氧条件下, Hb/NO循环可维持细胞内的氧化还原和能量平衡状态(Igamberdiev and Hill, 2004; Gupta et al., 2011).Hb/NO呼吸循环包括4个步骤: (1) 硝酸根离子(nitrate ion, NO3-)被NR还原为NO2-; (2) NO2-从细胞质转移到线粒体基质; (3) 通过线粒体ETC将NO2-还原为NO, 使ATP再生; (4) 最终NO通过被动扩散到胞质溶胶中, 由Hb将其氧化成硝酸盐(Gupta et al., 2011; Horchani et al., 2011; Hichri et al., 2015).因此, 在低氧环境下, 植物线粒体可以保持其氧化NADH的能力, 从而合成ATP.相关研究也表明, 缺氧环境下, 大豆和苜蓿的成熟根瘤通过硝酸还原途径和细菌反硝化途径均能产生NO, 参与Hb/NO呼吸循环, 为机体提供能量(Meakin et al., 2007; Horchani et al., 2011; Sánchez et al., 2011).此外, 在缺氧条件下, 结瘤的能量状态几乎全部取决于NR的功能(Horchani et al., 2011).值得注意的是, 在Hb/NO循环过程中, 由于多数情况下处于缺氧状态, 因此最终的电子受体是亚硝酸盐而非O2 (Gupta et al., 2011; Igamberdiev et al., 2014).Hichri等(2015)发现, NO在结瘤过程中具有双重作用: 一方面作为维持基础能量代谢所必需的中间物; 另一方面作为C、N代谢的调节剂, 降低微氧环境下的能量需求.由于缺氧对于植物的生长发育影响很大, 因此当O2不足时, NO对维持豆科植物与根瘤菌共生结瘤的氧化还原和能量平衡状态至关重要. ...
Plant mitochondrial function during anaerobiosis 2 2009
... 豆科植物根瘤中包含一种与其它蛋白性质和功能不同的血红蛋白(hemoglobins, Hbs), 能够清除机体产生或累积的NO (Gupta et al., 2011).基于其序列同源性和对氧的亲和力, 可将植物体内的Hbs划分成3类: 非共生血红蛋白(non-symbiotic hemoglobins, ns-Hbs, 1类)、豆血红蛋白(leghemoglobins, Lbs, 2类)和截短血红蛋白(truncated hemoglobin, Tr-Hbs, 3类) (Gupta et al., 2011; Hill, 2012).Lbs作为高等植物中发现的第1种豆血红蛋白, 在豆科植物中主要负责将根瘤中的O2运输至根瘤菌(Trevaskis et al., 1997).而Hbs对O2和NO的亲和力非常强, 能够有效清除O2和NO并将其转化为硝酸盐.有研究表明, Hbs能够维持植物细胞在缺氧状态下的氧化还原能力和能量状态(Igamberdiev and Hill, 2009).除植物体蛋白外, 根瘤菌内也含有3类血红蛋白: 黄素血红蛋白(flavohaemoglobins, fHbs/Hmp)、单域血红蛋白(single-domain haemoglobins, sd-Hbs)和截短血红蛋白(Sánchez et al., 2011).Hmp是蒺藜苜蓿(Medicago truncatula)中主要的NO清除剂, 而NO可诱导hmp基因表达以影响自身生成量(Meilhoc et al., 2011; Cam et al., 2012) (图1).另有研究表明, 在蒺藜苜蓿hmp突变的根瘤中, NO水平显著高于野生型, 且固氮效率降低和根瘤衰老提前(Cam et al., 2012; Hichri et al., 2016a).因此, 植物中的Hbs与菌内的Hmp对于维持共生体NO水平至关重要. ...
... NO可通过调节共生体的能量代谢影响N2固定.固氮过程中会产生铵根离子(ammonium ion, NH4+), 当其达到一定浓度时即具有毒性并损害植物的生长(Li et al., 2014).GS可同化NH4+, 解除这种离子胁迫, 而NO通过Tyr硝化改变其活性, 因此在N代谢中NO与GS起着至关重要的作用(Melo et al., 2011; Sainz et al., 2015).S-亚硝基化也是植物体内NO信号转导的关键机制.研究发现蒺藜苜蓿成熟根瘤中约有80种S-亚硝基化蛋白, 且多数与C、N代谢及固氮酶有关, 如果糖激酶、固氮酶钼铁(MoFe)蛋白nifK与铁(Fe)蛋白nifH (Puppo et al., 2013).NO可能通过影响根瘤中与S-亚硝基化蛋白相关的酶活性, 利用三羧酸循环(tricarboxylic acid cycle, TCA cycle)和糖酵解(glycolysis)等共生体的能量代谢过程调控固氮酶活性(Igamberdiev and Hill, 2009; Boscari et al., 2013; Li et al., 2014; Igamberdiev et al., 2014) (图2).但对于NO究竟是直接抑制固氮酶活性还是通过间接调节C、N代谢影响豆科植物体固氮, 目前尚不清楚(Hichri et al., 2016b). ...
Plant mitochondria: source and target for nitric oxide 3 2014
... 根瘤中的NO可通过调控酶的翻译后修饰调节碳(C)、氮(N)及能量代谢(Chaki et al., 2014; Hu et al., 2015).NO的产生是植物应对缺氧胁迫的一种响应, 与呼吸循环过程相关联, 形成“Hb/NO”循环.缺氧条件下, Hb/NO循环可维持细胞内的氧化还原和能量平衡状态(Igamberdiev and Hill, 2004; Gupta et al., 2011).Hb/NO呼吸循环包括4个步骤: (1) 硝酸根离子(nitrate ion, NO3-)被NR还原为NO2-; (2) NO2-从细胞质转移到线粒体基质; (3) 通过线粒体ETC将NO2-还原为NO, 使ATP再生; (4) 最终NO通过被动扩散到胞质溶胶中, 由Hb将其氧化成硝酸盐(Gupta et al., 2011; Horchani et al., 2011; Hichri et al., 2015).因此, 在低氧环境下, 植物线粒体可以保持其氧化NADH的能力, 从而合成ATP.相关研究也表明, 缺氧环境下, 大豆和苜蓿的成熟根瘤通过硝酸还原途径和细菌反硝化途径均能产生NO, 参与Hb/NO呼吸循环, 为机体提供能量(Meakin et al., 2007; Horchani et al., 2011; Sánchez et al., 2011).此外, 在缺氧条件下, 结瘤的能量状态几乎全部取决于NR的功能(Horchani et al., 2011).值得注意的是, 在Hb/NO循环过程中, 由于多数情况下处于缺氧状态, 因此最终的电子受体是亚硝酸盐而非O2 (Gupta et al., 2011; Igamberdiev et al., 2014).Hichri等(2015)发现, NO在结瘤过程中具有双重作用: 一方面作为维持基础能量代谢所必需的中间物; 另一方面作为C、N代谢的调节剂, 降低微氧环境下的能量需求.由于缺氧对于植物的生长发育影响很大, 因此当O2不足时, NO对维持豆科植物与根瘤菌共生结瘤的氧化还原和能量平衡状态至关重要. ...
... NO可通过调节共生体的能量代谢影响N2固定.固氮过程中会产生铵根离子(ammonium ion, NH4+), 当其达到一定浓度时即具有毒性并损害植物的生长(Li et al., 2014).GS可同化NH4+, 解除这种离子胁迫, 而NO通过Tyr硝化改变其活性, 因此在N代谢中NO与GS起着至关重要的作用(Melo et al., 2011; Sainz et al., 2015).S-亚硝基化也是植物体内NO信号转导的关键机制.研究发现蒺藜苜蓿成熟根瘤中约有80种S-亚硝基化蛋白, 且多数与C、N代谢及固氮酶有关, 如果糖激酶、固氮酶钼铁(MoFe)蛋白nifK与铁(Fe)蛋白nifH (Puppo et al., 2013).NO可能通过影响根瘤中与S-亚硝基化蛋白相关的酶活性, 利用三羧酸循环(tricarboxylic acid cycle, TCA cycle)和糖酵解(glycolysis)等共生体的能量代谢过程调控固氮酶活性(Igamberdiev and Hill, 2009; Boscari et al., 2013; Li et al., 2014; Igamberdiev et al., 2014) (图2).但对于NO究竟是直接抑制固氮酶活性还是通过间接调节C、N代谢影响豆科植物体固氮, 目前尚不清楚(Hichri et al., 2016b). ...
... 尽管目前对于豆科植物与根瘤菌共生固氮的初始过程已有较多研究, 但因发育或应激引起根瘤衰老的分子机制却报道较少.根瘤衰老区域的表型是由粉红色变为绿色, 其实质源于植物Hbs的降解; 在转录水平上, 衰老表现为编码蛋白质降解, 核酸、膜脂和糖类的相关基因被激活; 同时伴有衰老根瘤中氧化还原平衡的改变, 以及抗氧化防御的整体减弱等现象(Van de Velde et al., 2006; Loscos et al., 2008).NO在植物的衰老过程中具有重要作用(Procházková and Wilhelmová, 2011).例如, NO是水稻(Oryza sativa)光依赖性叶细胞死亡的关键介质(Lin et al., 2012); 蒺藜苜蓿根瘤内NO含量的增加会导致根瘤过早衰老, 且衰老程度与NO水平相关(Cam et al., 2012; Meilhoc et al., 2013; Blanquet et al., 2015), 表明NO对于维持共生体系和避免根瘤过早衰老极为重要.此外, 施加硝酸盐可导致豌豆表现出典型的衰老特征(Escuredo et al., 1996).对蒺藜苜蓿添加硝酸盐后, 其自身也可通过还原硝酸盐促使根瘤中生成NO, 然而硝酸盐引起的根瘤衰老是否依赖于NO目前尚不清楚(Horchani et al., 2011).此外, NO也是一种有效的呼吸抑制剂, 能够抑制线粒体和细菌末端的呼吸氧化酶, 并增加活性氧(reactive oxygen species, ROS)和活性氮(reactive nitrogen species, RNS)的积累, 而NO和ROS均为共生发育所必需(Shimoda et al., 2005; Cam et al., 2012; Igamberdiev et al., 2014; Arjona et al., 2015), 因此衰老也可能是固氮时呼吸抑制的间接结果. ...
Glutathione synthesis is regulated by nitric oxide in Medicago truncatula roots 2 2007
... NO在影响相关基因的表达、调控豆科植物防御反应进而促进共生和结瘤中发挥重要作用.转录组分析表明, NO影响多种植物基因的表达, 如编码富含半胱氨酸蛋白(cysteine-rich proteins, NCRs)、核糖体蛋白、肽酶、结瘤发育细胞周期蛋白及细胞周期转换蛋白的基因(Vinardell et al., 2003; del Giudice et al., 2011; Boscari et al., 2013), 进而调控细胞脱分化、诱导组织形成及抑制植物防御反应(Hichri et al., 2016b).LjHb1是百脉根(Lotus japonicus) ns-Hbs的编码基因, 外源添加NO、缺氧和低温均可诱导其表达(Shimoda et al., 2005).在根瘤形成的早期阶段, NO的产生可以上调LjHb1的表达, 反之LjHb1的表达又下调NO的水平, 以降低植物防御反应, 促进根瘤菌和宿主植物建立共生关系(Nagata et al., 2008; Murakami et al., 2011).植物也可通过降低Hb的表达促进内源NO产生, 调控自身机体内相关防御基因的表达(Wally et al., 2013) (图2).而NO的清除会引起基因MtCRE1和MtCCS52A的下调表达, 从而延缓结瘤(del Giudice et al., 2011).通过对接种中华根瘤菌的蒺藜苜蓿外源添加NO清除剂(2-(4-carboxyphenyl)- 4,4,5,5-tetramethyl imidazoline-1-oxyl-3-oxide, cPTIO), 发现受NO调节的3种基因(Contig525、Contig3307和Medtr5g010350)表达下调(Boscari et al., 2013).其中, 前2种基因分别与ns-Hbs和茉莉酮酸代谢途径(Palmieri et al., 2008)相关, 而最后1种基因可调控还原型谷胱甘肽(glutathione, GSH)的合成(Innocenti et al., 2007).转录组分析显示, 蒺藜苜蓿接种中华根瘤菌后, 外源添加cPTIO或NADPH氧化酶抑制剂二苯烯碘铵(diphenylene iodide, DPI), 导致细胞壁形成和发育过程中相关基因下调表达, 而植物防御和二次代谢相关基因上调表达(Puppo et al., 2013).hmp的过表达和外源添加cPTIO都可降低蒺藜苜蓿的结瘤效率(Kato et al., 2010; del Giudice et al., 2011), NO则通过调节上述基因的表达, 抑制相关防御反应从而促进共生关系的建立.此外, 在根瘤菌内也鉴定出数百个依赖NO表达的基因, 如中华根瘤菌中编码Hmp的hmp基因(del Giudice et al., 2011; Cam et al., 2012)、编码haeme-Cu蛋白的nnrSI和nnrS2基因(Blanquet et al., 2015)及编码NiR的nirKV基因(Meilhoc et al., 2011), 但关于根瘤菌内受NO影响且与共生结瘤相关的基因报道很少. ...
Involvement of nitric oxide in the inhibition of nitrogenase activity by nitrate in Lotus root nodules 5 2010
... 豆科植物根瘤菌共生体系受多种信号分子和转录因子调控, 其中一氧化氮(nitric oxide, NO)信号分子在豆科植物微生物互作过程中发挥重要作用.共生体早期的转录组学分析表明, NO可调节细胞脱分化和器官发生(Ferrarini et al., 2008; Boscari et al., 2013), 并抑制植物防御反应(Gonzalez-Rizzo et al., 2006), 在建立植物与菌共生关系中发挥重要调控作用(Hichri et al., 2016b).而在成熟根瘤中, NO既可抑制植物体固氮(Shimoda et al., 2005; Kato et al., 2010; Cam et al., 2012), 又可诱发根瘤衰老(Horchani et al., 2011; Cam et al., 2012; Blanquet et al., 2015), 亦可作为能量代谢调节器维持低氧(oxygen, O2)状态下的能量平衡(Kato et al., 2010; Hichri et al., 2016b).尽管目前关于NO在植物体内的生理功能已进行了充分研究, 但国内对于其在豆科植物根瘤菌共生体系中的作用机制并未进行系统的阐述.本文综述了豆科植物根瘤菌共生体系中NO的产生和降解, 及其对根瘤形成与共生固氮影响的研究进展, 并对NO在共生体系中对非生物胁迫的响应及其与激素的相互作用进行了阐述. ...
... )状态下的能量平衡(Kato et al., 2010; Hichri et al., 2016b).尽管目前关于NO在植物体内的生理功能已进行了充分研究, 但国内对于其在豆科植物根瘤菌共生体系中的作用机制并未进行系统的阐述.本文综述了豆科植物根瘤菌共生体系中NO的产生和降解, 及其对根瘤形成与共生固氮影响的研究进展, 并对NO在共生体系中对非生物胁迫的响应及其与激素的相互作用进行了阐述. ...
... NO在影响相关基因的表达、调控豆科植物防御反应进而促进共生和结瘤中发挥重要作用.转录组分析表明, NO影响多种植物基因的表达, 如编码富含半胱氨酸蛋白(cysteine-rich proteins, NCRs)、核糖体蛋白、肽酶、结瘤发育细胞周期蛋白及细胞周期转换蛋白的基因(Vinardell et al., 2003; del Giudice et al., 2011; Boscari et al., 2013), 进而调控细胞脱分化、诱导组织形成及抑制植物防御反应(Hichri et al., 2016b).LjHb1是百脉根(Lotus japonicus) ns-Hbs的编码基因, 外源添加NO、缺氧和低温均可诱导其表达(Shimoda et al., 2005).在根瘤形成的早期阶段, NO的产生可以上调LjHb1的表达, 反之LjHb1的表达又下调NO的水平, 以降低植物防御反应, 促进根瘤菌和宿主植物建立共生关系(Nagata et al., 2008; Murakami et al., 2011).植物也可通过降低Hb的表达促进内源NO产生, 调控自身机体内相关防御基因的表达(Wally et al., 2013) (图2).而NO的清除会引起基因MtCRE1和MtCCS52A的下调表达, 从而延缓结瘤(del Giudice et al., 2011).通过对接种中华根瘤菌的蒺藜苜蓿外源添加NO清除剂(2-(4-carboxyphenyl)- 4,4,5,5-tetramethyl imidazoline-1-oxyl-3-oxide, cPTIO), 发现受NO调节的3种基因(Contig525、Contig3307和Medtr5g010350)表达下调(Boscari et al., 2013).其中, 前2种基因分别与ns-Hbs和茉莉酮酸代谢途径(Palmieri et al., 2008)相关, 而最后1种基因可调控还原型谷胱甘肽(glutathione, GSH)的合成(Innocenti et al., 2007).转录组分析显示, 蒺藜苜蓿接种中华根瘤菌后, 外源添加cPTIO或NADPH氧化酶抑制剂二苯烯碘铵(diphenylene iodide, DPI), 导致细胞壁形成和发育过程中相关基因下调表达, 而植物防御和二次代谢相关基因上调表达(Puppo et al., 2013).hmp的过表达和外源添加cPTIO都可降低蒺藜苜蓿的结瘤效率(Kato et al., 2010; del Giudice et al., 2011), NO则通过调节上述基因的表达, 抑制相关防御反应从而促进共生关系的建立.此外, 在根瘤菌内也鉴定出数百个依赖NO表达的基因, 如中华根瘤菌中编码Hmp的hmp基因(del Giudice et al., 2011; Cam et al., 2012)、编码haeme-Cu蛋白的nnrSI和nnrS2基因(Blanquet et al., 2015)及编码NiR的nirKV基因(Meilhoc et al., 2011), 但关于根瘤菌内受NO影响且与共生结瘤相关的基因报道很少. ...
... 研究表明, 外源添加NO可抑制大豆与百脉根根瘤的固氮酶活性(Kato et al., 2010; Cam et al., 2012; Boscari et al., 2013).Cam等(2012)报道, 成熟根瘤中NO的积累不仅可降低固氮效率, 还会影响植物的生长适应性, 而通过降解NO可增强植物体固氮能力和延长植物整体固氮周期.研究发现, hmp的过表达降低了根瘤中NO的含量, 却增强了乙炔还原活性(acetylene-reducing activity, ARA)和延迟根瘤衰老(Meilhoc et al., 2011; Cam et al., 2012).但NO的毒性、信号功能或代谢效应取决于其所在作用部位的NO浓度(Boscari et al., 2013; Mur et al., 2013).例如, 在百脉根中, 高浓度的NO可抑制氮的固定, 适当低浓度的NO则可增强固氮作用(Kato et al., 2010).因此, 较低且稳定浓度的NO是维持豆科植物根瘤形成和发育的重要因素(Shimoda et al., 2005; Cam et al., 2012; Calvo-Begueria et al., 2018). ...
... ).例如, 在百脉根中, 高浓度的NO可抑制氮的固定, 适当低浓度的NO则可增强固氮作用(Kato et al., 2010).因此, 较低且稳定浓度的NO是维持豆科植物根瘤形成和发育的重要因素(Shimoda et al., 2005; Cam et al., 2012; Calvo-Begueria et al., 2018). ...
The guard cell-environment connection 1 1993
... 研究表明, 长期黑暗处理能显著诱导根瘤衰老, 同时降低固氮酶活性和Hbs水平(Romanov et al., 1980; Matamoros et al., 1999; Swaraj et al., 2001; Pérez Guerra et al., 2010).持续的暗处理会干扰根瘤功能, 导致根瘤完全衰老(Pérez-Guerra et al., 2010), 而通过降解根瘤中的NO可以延缓黑暗诱导的衰老, 但关于NO在根瘤衰老过程中的作用机制还需进一步研究(Cam et al., 2012).She等(2004)发现在光/暗条件下,对蚕豆(Vicia faba)外源添加cPTIO和L-NAME可减弱H2O2对NO产生的诱导效应和改变人为光照引起的气孔关闭.气孔关闭由多种生物和非生物因素引起, 如渗透胁迫、黑暗、高浓度的CO2和机械压力, 其中光是调节气孔运动最重要的环境因素之一(Zeiger, 1983; Kearns and Assmann, 1993; Herold and Puppo, 2005; Sánchez et al., 2010).但关于黑暗条件下NO调节豆科植物共生固氮的作用机制还未见报道. ...
Involvement of thioredoxin domain-containing 5 in resistance to nitrosative stress 1 2010
... 除根瘤菌内的血红蛋白以外, 还有其它细菌蛋白调控根瘤中的NO水平.例如, 硫氧还蛋白(thioredoxin reductase, TrxR)或亚硝基谷胱甘肽还原酶(S-nitroglutathione reductase, GSNOR)都可能间接参与NO的降解和信号传递过程(Lee et al., 2010).而细菌的NO降解蛋白不仅能保护其自身蛋白免受NO的负面影响, 还能使共生植物蛋白免受NO介导的失活(Blanquet et al., 2015; Hichri et al., 2016b).综上, 鉴于NO的影响, 共生菌可能需要更多不同的系统来降低自身的NO水平. ...
Ammonium stress in Arabidopsis: signaling, genetic loci, and physiological targets 3 2014
... 脱落酸(abscisic acid, ABA)是植物适应环境胁迫的关键激素(Yoshida et al., 2015), 可通过抑制根毛变形及侵染线的形成影响豆科植物结瘤(Suzuki et al., 2004; Ding et al., 2008).Tominaga等(2010)在百脉根中发现, ABA不仅可调节根瘤的固氮酶活性, 还可通过影响结瘤过程中NO的产生来影响结瘤.在根瘤形成过程中, NO通过酪氨酸(tyrosine, Tyr)硝化调控ABA信号传递, 而ABA又在信号转导过程中决定NO的产生与积累(Castillo et al., 2015).Tyr硝化是NO介导的翻译后修饰, 其特征在于将-NO2添加到Tyr残基芳环的等价邻位C上, 产生3-硝基酪氨酸(3-nitrotyrosine, 3-NT), 并由此改变蛋白质构象(Radi, 2004).作为硝化分子的前体, NO通过Tyr硝化改变细胞溶质谷氨酰胺合成酶(glutamine synthetase 1, GS1)和Lb等关键结瘤蛋白的活性(Melo et al., 2011; Navascués et al., 2012; Li et al., 2014).豆科植物结瘤时Lb发生Tyr硝化, 可保护共生体免受过氧亚硝酸盐毒害, 因此3-NT的形成既是硝基氧化应激的标志, 也是功能性结瘤活跃代谢的结果(Sainz et al., 2015).此外, NOS抑制剂N-硝基-L-精氨酸甲酯(NG-nitro- L-arginine methyl ester, L-NAME)和cPTIO处理可减弱豌豆(Pisum sativum)表皮由ABA诱导的气孔关闭(张绪成等, 2005). ...
... NO可通过调节共生体的能量代谢影响N2固定.固氮过程中会产生铵根离子(ammonium ion, NH4+), 当其达到一定浓度时即具有毒性并损害植物的生长(Li et al., 2014).GS可同化NH4+, 解除这种离子胁迫, 而NO通过Tyr硝化改变其活性, 因此在N代谢中NO与GS起着至关重要的作用(Melo et al., 2011; Sainz et al., 2015).S-亚硝基化也是植物体内NO信号转导的关键机制.研究发现蒺藜苜蓿成熟根瘤中约有80种S-亚硝基化蛋白, 且多数与C、N代谢及固氮酶有关, 如果糖激酶、固氮酶钼铁(MoFe)蛋白nifK与铁(Fe)蛋白nifH (Puppo et al., 2013).NO可能通过影响根瘤中与S-亚硝基化蛋白相关的酶活性, 利用三羧酸循环(tricarboxylic acid cycle, TCA cycle)和糖酵解(glycolysis)等共生体的能量代谢过程调控固氮酶活性(Igamberdiev and Hill, 2009; Boscari et al., 2013; Li et al., 2014; Igamberdiev et al., 2014) (图2).但对于NO究竟是直接抑制固氮酶活性还是通过间接调节C、N代谢影响豆科植物体固氮, 目前尚不清楚(Hichri et al., 2016b). ...
... ; Li et al., 2014; Igamberdiev et al., 2014) (图2).但对于NO究竟是直接抑制固氮酶活性还是通过间接调节C、N代谢影响豆科植物体固氮, 目前尚不清楚(Hichri et al., 2016b). ...
Nitric oxide and protein S-nitrosylation are integral to hydrogen peroxide-induced leaf cell death in rice 1 2012
... 尽管目前对于豆科植物与根瘤菌共生固氮的初始过程已有较多研究, 但因发育或应激引起根瘤衰老的分子机制却报道较少.根瘤衰老区域的表型是由粉红色变为绿色, 其实质源于植物Hbs的降解; 在转录水平上, 衰老表现为编码蛋白质降解, 核酸、膜脂和糖类的相关基因被激活; 同时伴有衰老根瘤中氧化还原平衡的改变, 以及抗氧化防御的整体减弱等现象(Van de Velde et al., 2006; Loscos et al., 2008).NO在植物的衰老过程中具有重要作用(Procházková and Wilhelmová, 2011).例如, NO是水稻(Oryza sativa)光依赖性叶细胞死亡的关键介质(Lin et al., 2012); 蒺藜苜蓿根瘤内NO含量的增加会导致根瘤过早衰老, 且衰老程度与NO水平相关(Cam et al., 2012; Meilhoc et al., 2013; Blanquet et al., 2015), 表明NO对于维持共生体系和避免根瘤过早衰老极为重要.此外, 施加硝酸盐可导致豌豆表现出典型的衰老特征(Escuredo et al., 1996).对蒺藜苜蓿添加硝酸盐后, 其自身也可通过还原硝酸盐促使根瘤中生成NO, 然而硝酸盐引起的根瘤衰老是否依赖于NO目前尚不清楚(Horchani et al., 2011).此外, NO也是一种有效的呼吸抑制剂, 能够抑制线粒体和细菌末端的呼吸氧化酶, 并增加活性氧(reactive oxygen species, ROS)和活性氮(reactive nitrogen species, RNS)的积累, 而NO和ROS均为共生发育所必需(Shimoda et al., 2005; Cam et al., 2012; Igamberdiev et al., 2014; Arjona et al., 2015), 因此衰老也可能是固氮时呼吸抑制的间接结果. ...
Ascorbate and homoglutathione metabolism in common bean nodules under stress conditions and during natural senescence 1 2008
... 尽管目前对于豆科植物与根瘤菌共生固氮的初始过程已有较多研究, 但因发育或应激引起根瘤衰老的分子机制却报道较少.根瘤衰老区域的表型是由粉红色变为绿色, 其实质源于植物Hbs的降解; 在转录水平上, 衰老表现为编码蛋白质降解, 核酸、膜脂和糖类的相关基因被激活; 同时伴有衰老根瘤中氧化还原平衡的改变, 以及抗氧化防御的整体减弱等现象(Van de Velde et al., 2006; Loscos et al., 2008).NO在植物的衰老过程中具有重要作用(Procházková and Wilhelmová, 2011).例如, NO是水稻(Oryza sativa)光依赖性叶细胞死亡的关键介质(Lin et al., 2012); 蒺藜苜蓿根瘤内NO含量的增加会导致根瘤过早衰老, 且衰老程度与NO水平相关(Cam et al., 2012; Meilhoc et al., 2013; Blanquet et al., 2015), 表明NO对于维持共生体系和避免根瘤过早衰老极为重要.此外, 施加硝酸盐可导致豌豆表现出典型的衰老特征(Escuredo et al., 1996).对蒺藜苜蓿添加硝酸盐后, 其自身也可通过还原硝酸盐促使根瘤中生成NO, 然而硝酸盐引起的根瘤衰老是否依赖于NO目前尚不清楚(Horchani et al., 2011).此外, NO也是一种有效的呼吸抑制剂, 能够抑制线粒体和细菌末端的呼吸氧化酶, 并增加活性氧(reactive oxygen species, ROS)和活性氮(reactive nitrogen species, RNS)的积累, 而NO和ROS均为共生发育所必需(Shimoda et al., 2005; Cam et al., 2012; Igamberdiev et al., 2014; Arjona et al., 2015), 因此衰老也可能是固氮时呼吸抑制的间接结果. ...
Glutathione and homoglutathione synthesis in legume root nodules 1 1999
... 研究表明, 长期黑暗处理能显著诱导根瘤衰老, 同时降低固氮酶活性和Hbs水平(Romanov et al., 1980; Matamoros et al., 1999; Swaraj et al., 2001; Pérez Guerra et al., 2010).持续的暗处理会干扰根瘤功能, 导致根瘤完全衰老(Pérez-Guerra et al., 2010), 而通过降解根瘤中的NO可以延缓黑暗诱导的衰老, 但关于NO在根瘤衰老过程中的作用机制还需进一步研究(Cam et al., 2012).She等(2004)发现在光/暗条件下,对蚕豆(Vicia faba)外源添加cPTIO和L-NAME可减弱H2O2对NO产生的诱导效应和改变人为光照引起的气孔关闭.气孔关闭由多种生物和非生物因素引起, 如渗透胁迫、黑暗、高浓度的CO2和机械压力, 其中光是调节气孔运动最重要的环境因素之一(Zeiger, 1983; Kearns and Assmann, 1993; Herold and Puppo, 2005; Sánchez et al., 2010).但关于黑暗条件下NO调节豆科植物共生固氮的作用机制还未见报道. ...
Function of glutathione peroxidases in legume root nodules 2 2015
The contribution of bacteroidal nitrate and nitrite reduction to the formation of nitrosylleghaemoglobin complexes in soybean root nodules 2 2007
... 根瘤中的NO可通过调控酶的翻译后修饰调节碳(C)、氮(N)及能量代谢(Chaki et al., 2014; Hu et al., 2015).NO的产生是植物应对缺氧胁迫的一种响应, 与呼吸循环过程相关联, 形成“Hb/NO”循环.缺氧条件下, Hb/NO循环可维持细胞内的氧化还原和能量平衡状态(Igamberdiev and Hill, 2004; Gupta et al., 2011).Hb/NO呼吸循环包括4个步骤: (1) 硝酸根离子(nitrate ion, NO3-)被NR还原为NO2-; (2) NO2-从细胞质转移到线粒体基质; (3) 通过线粒体ETC将NO2-还原为NO, 使ATP再生; (4) 最终NO通过被动扩散到胞质溶胶中, 由Hb将其氧化成硝酸盐(Gupta et al., 2011; Horchani et al., 2011; Hichri et al., 2015).因此, 在低氧环境下, 植物线粒体可以保持其氧化NADH的能力, 从而合成ATP.相关研究也表明, 缺氧环境下, 大豆和苜蓿的成熟根瘤通过硝酸还原途径和细菌反硝化途径均能产生NO, 参与Hb/NO呼吸循环, 为机体提供能量(Meakin et al., 2007; Horchani et al., 2011; Sánchez et al., 2011).此外, 在缺氧条件下, 结瘤的能量状态几乎全部取决于NR的功能(Horchani et al., 2011).值得注意的是, 在Hb/NO循环过程中, 由于多数情况下处于缺氧状态, 因此最终的电子受体是亚硝酸盐而非O2 (Gupta et al., 2011; Igamberdiev et al., 2014).Hichri等(2015)发现, NO在结瘤过程中具有双重作用: 一方面作为维持基础能量代谢所必需的中间物; 另一方面作为C、N代谢的调节剂, 降低微氧环境下的能量需求.由于缺氧对于植物的生长发育影响很大, 因此当O2不足时, NO对维持豆科植物与根瘤菌共生结瘤的氧化还原和能量平衡状态至关重要. ...
... NO还参与豆科植物对缺氧胁迫的反应.缺氧胁迫可诱导百脉根根瘤产生亚硝酰基豆血红蛋白(nitrosylleghemoglobin, LbNO)以减少根瘤内的硝酸盐, 同时LbNO的形成也有利于解除NO对植物体的毒害(Meakin et al., 2007).根瘤中LbNOs的增加不会抑制植物的固氮作用, 其实质源于Lb在根瘤中既可清除由细菌反硝化作用产生的NO和亚硝酸盐, 又可保护淹水条件下的固氮酶活性(Sánchez et al., 2010).Sánchez等(2010)发现, 在水淹缺氧条件下, 野生型和norC突变体根瘤中编码固氮酶Fe蛋白的nifH基因表达量降低、固氮酶活性下降, 通过清除NO可抵消这种影响.而当从常氧转为缺氧时, 根瘤内NO迅速增加, 表明根瘤中不仅能产生NO, 还具有响应缺氧胁迫而迅速上调表达的基因(Sánchez et al., 2010; Horchani et al., 2011).因此, 无论在转录水平还是翻译水平, 固氮酶都是NO抑制N2固定的关键靶标(Sánchez et al., 2010), 同时也表明在微氧环境中, NO主要通过影响固氮酶活性来调节植物体固氮. ...
Control of NO level in rhizobium-legume root nodules: not only a plant globin story 2 2013
... 共生体内NO的降解还受相关基因及其它蛋白调控.苜蓿中华根瘤菌(Sinorhizobium meliloti)中包含1个与短链脱氢酶相关且位于nnrR下游的nnrS基因家族, nnrS的表达依赖于转录调节器NnrR (De Bruijn et al., 2006).当nnrS1突变时, 根瘤内NO的水平升高(Blanquet et al., 2015).NnrS是一种haeme-Cu膜蛋白(Bartnikas et al., 2002), NnrS家族的2种蛋白(NnrS1和NnrS2)和Nor能够直接或间接参与NO的降解并有效地维持共生固氮(Meilhoc et al., 2013; Blanquet et al., 2015). ...
... 尽管目前对于豆科植物与根瘤菌共生固氮的初始过程已有较多研究, 但因发育或应激引起根瘤衰老的分子机制却报道较少.根瘤衰老区域的表型是由粉红色变为绿色, 其实质源于植物Hbs的降解; 在转录水平上, 衰老表现为编码蛋白质降解, 核酸、膜脂和糖类的相关基因被激活; 同时伴有衰老根瘤中氧化还原平衡的改变, 以及抗氧化防御的整体减弱等现象(Van de Velde et al., 2006; Loscos et al., 2008).NO在植物的衰老过程中具有重要作用(Procházková and Wilhelmová, 2011).例如, NO是水稻(Oryza sativa)光依赖性叶细胞死亡的关键介质(Lin et al., 2012); 蒺藜苜蓿根瘤内NO含量的增加会导致根瘤过早衰老, 且衰老程度与NO水平相关(Cam et al., 2012; Meilhoc et al., 2013; Blanquet et al., 2015), 表明NO对于维持共生体系和避免根瘤过早衰老极为重要.此外, 施加硝酸盐可导致豌豆表现出典型的衰老特征(Escuredo et al., 1996).对蒺藜苜蓿添加硝酸盐后, 其自身也可通过还原硝酸盐促使根瘤中生成NO, 然而硝酸盐引起的根瘤衰老是否依赖于NO目前尚不清楚(Horchani et al., 2011).此外, NO也是一种有效的呼吸抑制剂, 能够抑制线粒体和细菌末端的呼吸氧化酶, 并增加活性氧(reactive oxygen species, ROS)和活性氮(reactive nitrogen species, RNS)的积累, 而NO和ROS均为共生发育所必需(Shimoda et al., 2005; Cam et al., 2012; Igamberdiev et al., 2014; Arjona et al., 2015), 因此衰老也可能是固氮时呼吸抑制的间接结果. ...
Nitric oxide in legume-rhizobium symbiosis 5 2011
... 在根瘤菌侵染豆科植物的每个阶段(包括接种4小时后、根毛卷曲、侵染线的产生、根瘤原基的形成和根瘤成熟)都伴有NO的产生(Hichri et al., 2015), 即从豆科植物与根瘤菌的早期共生到根瘤成熟与衰老, NO的产生贯穿始终(Meilhoc et al., 2011).尽管目前对于NO的产生已经进行了许多研究, 但由于气体分子本身的性质以及与其它体系反应的复杂性, 使人们对植物中NO产生机制的研究受限, 因此关于豆科植物与根瘤菌共生的各个阶段中NO产生的机理还有待深入探索. ...
... 豆科植物根瘤中包含一种与其它蛋白性质和功能不同的血红蛋白(hemoglobins, Hbs), 能够清除机体产生或累积的NO (Gupta et al., 2011).基于其序列同源性和对氧的亲和力, 可将植物体内的Hbs划分成3类: 非共生血红蛋白(non-symbiotic hemoglobins, ns-Hbs, 1类)、豆血红蛋白(leghemoglobins, Lbs, 2类)和截短血红蛋白(truncated hemoglobin, Tr-Hbs, 3类) (Gupta et al., 2011; Hill, 2012).Lbs作为高等植物中发现的第1种豆血红蛋白, 在豆科植物中主要负责将根瘤中的O2运输至根瘤菌(Trevaskis et al., 1997).而Hbs对O2和NO的亲和力非常强, 能够有效清除O2和NO并将其转化为硝酸盐.有研究表明, Hbs能够维持植物细胞在缺氧状态下的氧化还原能力和能量状态(Igamberdiev and Hill, 2009).除植物体蛋白外, 根瘤菌内也含有3类血红蛋白: 黄素血红蛋白(flavohaemoglobins, fHbs/Hmp)、单域血红蛋白(single-domain haemoglobins, sd-Hbs)和截短血红蛋白(Sánchez et al., 2011).Hmp是蒺藜苜蓿(Medicago truncatula)中主要的NO清除剂, 而NO可诱导hmp基因表达以影响自身生成量(Meilhoc et al., 2011; Cam et al., 2012) (图1).另有研究表明, 在蒺藜苜蓿hmp突变的根瘤中, NO水平显著高于野生型, 且固氮效率降低和根瘤衰老提前(Cam et al., 2012; Hichri et al., 2016a).因此, 植物中的Hbs与菌内的Hmp对于维持共生体NO水平至关重要. ...
... NO在影响相关基因的表达、调控豆科植物防御反应进而促进共生和结瘤中发挥重要作用.转录组分析表明, NO影响多种植物基因的表达, 如编码富含半胱氨酸蛋白(cysteine-rich proteins, NCRs)、核糖体蛋白、肽酶、结瘤发育细胞周期蛋白及细胞周期转换蛋白的基因(Vinardell et al., 2003; del Giudice et al., 2011; Boscari et al., 2013), 进而调控细胞脱分化、诱导组织形成及抑制植物防御反应(Hichri et al., 2016b).LjHb1是百脉根(Lotus japonicus) ns-Hbs的编码基因, 外源添加NO、缺氧和低温均可诱导其表达(Shimoda et al., 2005).在根瘤形成的早期阶段, NO的产生可以上调LjHb1的表达, 反之LjHb1的表达又下调NO的水平, 以降低植物防御反应, 促进根瘤菌和宿主植物建立共生关系(Nagata et al., 2008; Murakami et al., 2011).植物也可通过降低Hb的表达促进内源NO产生, 调控自身机体内相关防御基因的表达(Wally et al., 2013) (图2).而NO的清除会引起基因MtCRE1和MtCCS52A的下调表达, 从而延缓结瘤(del Giudice et al., 2011).通过对接种中华根瘤菌的蒺藜苜蓿外源添加NO清除剂(2-(4-carboxyphenyl)- 4,4,5,5-tetramethyl imidazoline-1-oxyl-3-oxide, cPTIO), 发现受NO调节的3种基因(Contig525、Contig3307和Medtr5g010350)表达下调(Boscari et al., 2013).其中, 前2种基因分别与ns-Hbs和茉莉酮酸代谢途径(Palmieri et al., 2008)相关, 而最后1种基因可调控还原型谷胱甘肽(glutathione, GSH)的合成(Innocenti et al., 2007).转录组分析显示, 蒺藜苜蓿接种中华根瘤菌后, 外源添加cPTIO或NADPH氧化酶抑制剂二苯烯碘铵(diphenylene iodide, DPI), 导致细胞壁形成和发育过程中相关基因下调表达, 而植物防御和二次代谢相关基因上调表达(Puppo et al., 2013).hmp的过表达和外源添加cPTIO都可降低蒺藜苜蓿的结瘤效率(Kato et al., 2010; del Giudice et al., 2011), NO则通过调节上述基因的表达, 抑制相关防御反应从而促进共生关系的建立.此外, 在根瘤菌内也鉴定出数百个依赖NO表达的基因, 如中华根瘤菌中编码Hmp的hmp基因(del Giudice et al., 2011; Cam et al., 2012)、编码haeme-Cu蛋白的nnrSI和nnrS2基因(Blanquet et al., 2015)及编码NiR的nirKV基因(Meilhoc et al., 2011), 但关于根瘤菌内受NO影响且与共生结瘤相关的基因报道很少. ...
... 研究表明, 外源添加NO可抑制大豆与百脉根根瘤的固氮酶活性(Kato et al., 2010; Cam et al., 2012; Boscari et al., 2013).Cam等(2012)报道, 成熟根瘤中NO的积累不仅可降低固氮效率, 还会影响植物的生长适应性, 而通过降解NO可增强植物体固氮能力和延长植物整体固氮周期.研究发现, hmp的过表达降低了根瘤中NO的含量, 却增强了乙炔还原活性(acetylene-reducing activity, ARA)和延迟根瘤衰老(Meilhoc et al., 2011; Cam et al., 2012).但NO的毒性、信号功能或代谢效应取决于其所在作用部位的NO浓度(Boscari et al., 2013; Mur et al., 2013).例如, 在百脉根中, 高浓度的NO可抑制氮的固定, 适当低浓度的NO则可增强固氮作用(Kato et al., 2010).因此, 较低且稳定浓度的NO是维持豆科植物根瘤形成和发育的重要因素(Shimoda et al., 2005; Cam et al., 2012; Calvo-Begueria et al., 2018). ...
... NO不仅能调节植物生长发育, 还可参与植物对非生物胁迫的响应(Moreau et al., 2010).目前的研究发现, 在不同非生物胁迫(如干旱和缺氧)下, 共生有机体中的NO均会产生响应(Desalvo et al., 2010; Sánchez et al., 2011; Hawkins et al., 2014; Iarullina et al., 2014; Zimmer-Prados et al., 2014).同时, 在豆科植物中NO还可能参与调控机体对金属元素镉(Cd)和铝(Al)的胁迫反应以及应对非金属元素砷(As)的胁迫(Singh et al., 2015; 尚玉婷等, 2018), 但都缺乏在结瘤和共生固氮体系中的实验证据(Pérez- Chaca et al., 2014).Meilhoc等(2011)提出, 宿主中任何应激反应下NO的产生都会直接影响共生, 在成熟根瘤中, 除了NO专用调节器NnrR外, 多数根瘤菌的基因通过双组分系统(FixLJ)来调节NO, 其中FixLJ是O2限制反应的主要调节因子.nnrR是Nor结构基因norCB的上游基因, 可防止NO这种高活性氮氧化物的积累, 其插入失活可抑制亚硝酸盐的产生以及亚硝酸盐和NO的还原(De Bruijn et al., 2006).NO还能够可逆地结合光系统II (photosystem II, PSII)并调节电子的转移和猝灭, 对光合作用至关重要(Wodala et al., 2008).从农业和进化的角度来看, NO在植物与根瘤菌共生体系中的调控作用, 可以改善植物对病原体的防御状态、保护植物免受虫害以及增强植物对非生物胁迫的适应性(Chadha et al., 2015). ...
Glutamine synthetase is a molecular target of nitric oxide in root nodules of Medicago truncatula and is regulated by tyrosine nitration 2 2011
... 脱落酸(abscisic acid, ABA)是植物适应环境胁迫的关键激素(Yoshida et al., 2015), 可通过抑制根毛变形及侵染线的形成影响豆科植物结瘤(Suzuki et al., 2004; Ding et al., 2008).Tominaga等(2010)在百脉根中发现, ABA不仅可调节根瘤的固氮酶活性, 还可通过影响结瘤过程中NO的产生来影响结瘤.在根瘤形成过程中, NO通过酪氨酸(tyrosine, Tyr)硝化调控ABA信号传递, 而ABA又在信号转导过程中决定NO的产生与积累(Castillo et al., 2015).Tyr硝化是NO介导的翻译后修饰, 其特征在于将-NO2添加到Tyr残基芳环的等价邻位C上, 产生3-硝基酪氨酸(3-nitrotyrosine, 3-NT), 并由此改变蛋白质构象(Radi, 2004).作为硝化分子的前体, NO通过Tyr硝化改变细胞溶质谷氨酰胺合成酶(glutamine synthetase 1, GS1)和Lb等关键结瘤蛋白的活性(Melo et al., 2011; Navascués et al., 2012; Li et al., 2014).豆科植物结瘤时Lb发生Tyr硝化, 可保护共生体免受过氧亚硝酸盐毒害, 因此3-NT的形成既是硝基氧化应激的标志, 也是功能性结瘤活跃代谢的结果(Sainz et al., 2015).此外, NOS抑制剂N-硝基-L-精氨酸甲酯(NG-nitro- L-arginine methyl ester, L-NAME)和cPTIO处理可减弱豌豆(Pisum sativum)表皮由ABA诱导的气孔关闭(张绪成等, 2005). ...
... NO可通过调节共生体的能量代谢影响N2固定.固氮过程中会产生铵根离子(ammonium ion, NH4+), 当其达到一定浓度时即具有毒性并损害植物的生长(Li et al., 2014).GS可同化NH4+, 解除这种离子胁迫, 而NO通过Tyr硝化改变其活性, 因此在N代谢中NO与GS起着至关重要的作用(Melo et al., 2011; Sainz et al., 2015).S-亚硝基化也是植物体内NO信号转导的关键机制.研究发现蒺藜苜蓿成熟根瘤中约有80种S-亚硝基化蛋白, 且多数与C、N代谢及固氮酶有关, 如果糖激酶、固氮酶钼铁(MoFe)蛋白nifK与铁(Fe)蛋白nifH (Puppo et al., 2013).NO可能通过影响根瘤中与S-亚硝基化蛋白相关的酶活性, 利用三羧酸循环(tricarboxylic acid cycle, TCA cycle)和糖酵解(glycolysis)等共生体的能量代谢过程调控固氮酶活性(Igamberdiev and Hill, 2009; Boscari et al., 2013; Li et al., 2014; Igamberdiev et al., 2014) (图2).但对于NO究竟是直接抑制固氮酶活性还是通过间接调节C、N代谢影响豆科植物体固氮, 目前尚不清楚(Hichri et al., 2016b). ...
NO synthesis and signaling in plants-where do we stand? 1 2010
... NO不仅能调节植物生长发育, 还可参与植物对非生物胁迫的响应(Moreau et al., 2010).目前的研究发现, 在不同非生物胁迫(如干旱和缺氧)下, 共生有机体中的NO均会产生响应(Desalvo et al., 2010; Sánchez et al., 2011; Hawkins et al., 2014; Iarullina et al., 2014; Zimmer-Prados et al., 2014).同时, 在豆科植物中NO还可能参与调控机体对金属元素镉(Cd)和铝(Al)的胁迫反应以及应对非金属元素砷(As)的胁迫(Singh et al., 2015; 尚玉婷等, 2018), 但都缺乏在结瘤和共生固氮体系中的实验证据(Pérez- Chaca et al., 2014).Meilhoc等(2011)提出, 宿主中任何应激反应下NO的产生都会直接影响共生, 在成熟根瘤中, 除了NO专用调节器NnrR外, 多数根瘤菌的基因通过双组分系统(FixLJ)来调节NO, 其中FixLJ是O2限制反应的主要调节因子.nnrR是Nor结构基因norCB的上游基因, 可防止NO这种高活性氮氧化物的积累, 其插入失活可抑制亚硝酸盐的产生以及亚硝酸盐和NO的还原(De Bruijn et al., 2006).NO还能够可逆地结合光系统II (photosystem II, PSII)并调节电子的转移和猝灭, 对光合作用至关重要(Wodala et al., 2008).从农业和进化的角度来看, NO在植物与根瘤菌共生体系中的调控作用, 可以改善植物对病原体的防御状态、保护植物免受虫害以及增强植物对非生物胁迫的适应性(Chadha et al., 2015). ...
Integrating nitric oxide into salicylic acid and jasmonic acid/ethylene plant defense pathways 3 2013
... 植物和根瘤菌内NO的来源丰富, 主要通过硝酸还原酶(nitrate reductase, NR)和一氧化氮合酶(nitric oxide synthase, NOS)介导以及线粒体电子传递链(electron transport chain, ETC)的还原产生.在植物体内, NO既可由质外体中的亚硝酸根离子(NO2-)转化(Bethke et al., 2004), 又可通过以精氨酸(arginine)、多胺(polyamines)或羟胺(hydroxylamine)作为底物的氧化途径产生(Hichri et al., 2015).在低氧环境下, 还可在还原途径中通过质膜结合亚硝酸盐, 然后通过NO还原酶(nitric oxide reductase, Nor)和黄嘌呤氧化还原酶(xanthine oxidoreductase, XOR)或利用ETC的作用将NO2-还原为NO (Gupta et al., 2011; Mur et al., 2013; Hichri et al., 2015).而Horchani等(2011)发现, 植物及其共生菌中的NR和ETC均可产生NO, 其中NR在植物体内的主要功能是将硝酸盐转化为亚硝酸盐以同化N2 (Neill et al., 2008).但早期的研究发现, 在NAD(P)H作为电子供体时, 大豆(Glycine max)中的NR可将亚硝酸盐转化为NO (Dean and Harper, 1988).且NR抑制剂钨酸盐(tungstate, Tg)也能够抑制NO的产生, 表明NO的合成依赖于NR活性, 但并不是由其直接产生(Horchani et al., 2011).在类菌体中, NO则主要产生于反硝化途径(Hichri et al., 2015), 该途径涉及NR、亚硝酸还原酶(nitrite reductase, NiR)、Nor和NOS (Horchani et al., 2011; Sánchez et al., 2011; Calvo-Begueria et al., 2018) (图1).Berger等(2018)发现NOS也是植物NO的来源, 但目前关于共生菌内NO产生途径的机制研究仍较少. ...
... 研究表明, 外源添加NO可抑制大豆与百脉根根瘤的固氮酶活性(Kato et al., 2010; Cam et al., 2012; Boscari et al., 2013).Cam等(2012)报道, 成熟根瘤中NO的积累不仅可降低固氮效率, 还会影响植物的生长适应性, 而通过降解NO可增强植物体固氮能力和延长植物整体固氮周期.研究发现, hmp的过表达降低了根瘤中NO的含量, 却增强了乙炔还原活性(acetylene-reducing activity, ARA)和延迟根瘤衰老(Meilhoc et al., 2011; Cam et al., 2012).但NO的毒性、信号功能或代谢效应取决于其所在作用部位的NO浓度(Boscari et al., 2013; Mur et al., 2013).例如, 在百脉根中, 高浓度的NO可抑制氮的固定, 适当低浓度的NO则可增强固氮作用(Kato et al., 2010).因此, 较低且稳定浓度的NO是维持豆科植物根瘤形成和发育的重要因素(Shimoda et al., 2005; Cam et al., 2012; Calvo-Begueria et al., 2018). ...
Nitric oxide production induced in roots of Lotus japonicus by lipopolysaccharide from Mesorhizobium loti 1 2011
... NO在影响相关基因的表达、调控豆科植物防御反应进而促进共生和结瘤中发挥重要作用.转录组分析表明, NO影响多种植物基因的表达, 如编码富含半胱氨酸蛋白(cysteine-rich proteins, NCRs)、核糖体蛋白、肽酶、结瘤发育细胞周期蛋白及细胞周期转换蛋白的基因(Vinardell et al., 2003; del Giudice et al., 2011; Boscari et al., 2013), 进而调控细胞脱分化、诱导组织形成及抑制植物防御反应(Hichri et al., 2016b).LjHb1是百脉根(Lotus japonicus) ns-Hbs的编码基因, 外源添加NO、缺氧和低温均可诱导其表达(Shimoda et al., 2005).在根瘤形成的早期阶段, NO的产生可以上调LjHb1的表达, 反之LjHb1的表达又下调NO的水平, 以降低植物防御反应, 促进根瘤菌和宿主植物建立共生关系(Nagata et al., 2008; Murakami et al., 2011).植物也可通过降低Hb的表达促进内源NO产生, 调控自身机体内相关防御基因的表达(Wally et al., 2013) (图2).而NO的清除会引起基因MtCRE1和MtCCS52A的下调表达, 从而延缓结瘤(del Giudice et al., 2011).通过对接种中华根瘤菌的蒺藜苜蓿外源添加NO清除剂(2-(4-carboxyphenyl)- 4,4,5,5-tetramethyl imidazoline-1-oxyl-3-oxide, cPTIO), 发现受NO调节的3种基因(Contig525、Contig3307和Medtr5g010350)表达下调(Boscari et al., 2013).其中, 前2种基因分别与ns-Hbs和茉莉酮酸代谢途径(Palmieri et al., 2008)相关, 而最后1种基因可调控还原型谷胱甘肽(glutathione, GSH)的合成(Innocenti et al., 2007).转录组分析显示, 蒺藜苜蓿接种中华根瘤菌后, 外源添加cPTIO或NADPH氧化酶抑制剂二苯烯碘铵(diphenylene iodide, DPI), 导致细胞壁形成和发育过程中相关基因下调表达, 而植物防御和二次代谢相关基因上调表达(Puppo et al., 2013).hmp的过表达和外源添加cPTIO都可降低蒺藜苜蓿的结瘤效率(Kato et al., 2010; del Giudice et al., 2011), NO则通过调节上述基因的表达, 抑制相关防御反应从而促进共生关系的建立.此外, 在根瘤菌内也鉴定出数百个依赖NO表达的基因, 如中华根瘤菌中编码Hmp的hmp基因(del Giudice et al., 2011; Cam et al., 2012)、编码haeme-Cu蛋白的nnrSI和nnrS2基因(Blanquet et al., 2015)及编码NiR的nirKV基因(Meilhoc et al., 2011), 但关于根瘤菌内受NO影响且与共生结瘤相关的基因报道很少. ...
Expression of a class 1 hemoglobin gene and production of nitric oxide in response to symbiotic and pathogenic bacteria in Lotus japonicus 1 2008
... NO在影响相关基因的表达、调控豆科植物防御反应进而促进共生和结瘤中发挥重要作用.转录组分析表明, NO影响多种植物基因的表达, 如编码富含半胱氨酸蛋白(cysteine-rich proteins, NCRs)、核糖体蛋白、肽酶、结瘤发育细胞周期蛋白及细胞周期转换蛋白的基因(Vinardell et al., 2003; del Giudice et al., 2011; Boscari et al., 2013), 进而调控细胞脱分化、诱导组织形成及抑制植物防御反应(Hichri et al., 2016b).LjHb1是百脉根(Lotus japonicus) ns-Hbs的编码基因, 外源添加NO、缺氧和低温均可诱导其表达(Shimoda et al., 2005).在根瘤形成的早期阶段, NO的产生可以上调LjHb1的表达, 反之LjHb1的表达又下调NO的水平, 以降低植物防御反应, 促进根瘤菌和宿主植物建立共生关系(Nagata et al., 2008; Murakami et al., 2011).植物也可通过降低Hb的表达促进内源NO产生, 调控自身机体内相关防御基因的表达(Wally et al., 2013) (图2).而NO的清除会引起基因MtCRE1和MtCCS52A的下调表达, 从而延缓结瘤(del Giudice et al., 2011).通过对接种中华根瘤菌的蒺藜苜蓿外源添加NO清除剂(2-(4-carboxyphenyl)- 4,4,5,5-tetramethyl imidazoline-1-oxyl-3-oxide, cPTIO), 发现受NO调节的3种基因(Contig525、Contig3307和Medtr5g010350)表达下调(Boscari et al., 2013).其中, 前2种基因分别与ns-Hbs和茉莉酮酸代谢途径(Palmieri et al., 2008)相关, 而最后1种基因可调控还原型谷胱甘肽(glutathione, GSH)的合成(Innocenti et al., 2007).转录组分析显示, 蒺藜苜蓿接种中华根瘤菌后, 外源添加cPTIO或NADPH氧化酶抑制剂二苯烯碘铵(diphenylene iodide, DPI), 导致细胞壁形成和发育过程中相关基因下调表达, 而植物防御和二次代谢相关基因上调表达(Puppo et al., 2013).hmp的过表达和外源添加cPTIO都可降低蒺藜苜蓿的结瘤效率(Kato et al., 2010; del Giudice et al., 2011), NO则通过调节上述基因的表达, 抑制相关防御反应从而促进共生关系的建立.此外, 在根瘤菌内也鉴定出数百个依赖NO表达的基因, 如中华根瘤菌中编码Hmp的hmp基因(del Giudice et al., 2011; Cam et al., 2012)、编码haeme-Cu蛋白的nnrSI和nnrS2基因(Blanquet et al., 2015)及编码NiR的nirKV基因(Meilhoc et al., 2011), 但关于根瘤菌内受NO影响且与共生结瘤相关的基因报道很少. ...
Leghemoglobin green derivatives with nitrated hemes evidence production of highly reactive nitrogen species during aging of legume nodules 1 2012
... 脱落酸(abscisic acid, ABA)是植物适应环境胁迫的关键激素(Yoshida et al., 2015), 可通过抑制根毛变形及侵染线的形成影响豆科植物结瘤(Suzuki et al., 2004; Ding et al., 2008).Tominaga等(2010)在百脉根中发现, ABA不仅可调节根瘤的固氮酶活性, 还可通过影响结瘤过程中NO的产生来影响结瘤.在根瘤形成过程中, NO通过酪氨酸(tyrosine, Tyr)硝化调控ABA信号传递, 而ABA又在信号转导过程中决定NO的产生与积累(Castillo et al., 2015).Tyr硝化是NO介导的翻译后修饰, 其特征在于将-NO2添加到Tyr残基芳环的等价邻位C上, 产生3-硝基酪氨酸(3-nitrotyrosine, 3-NT), 并由此改变蛋白质构象(Radi, 2004).作为硝化分子的前体, NO通过Tyr硝化改变细胞溶质谷氨酰胺合成酶(glutamine synthetase 1, GS1)和Lb等关键结瘤蛋白的活性(Melo et al., 2011; Navascués et al., 2012; Li et al., 2014).豆科植物结瘤时Lb发生Tyr硝化, 可保护共生体免受过氧亚硝酸盐毒害, 因此3-NT的形成既是硝基氧化应激的标志, 也是功能性结瘤活跃代谢的结果(Sainz et al., 2015).此外, NOS抑制剂N-硝基-L-精氨酸甲酯(NG-nitro- L-arginine methyl ester, L-NAME)和cPTIO处理可减弱豌豆(Pisum sativum)表皮由ABA诱导的气孔关闭(张绪成等, 2005). ...
Nitric oxide, stomatal closure, and abiotic stress 1 2008
... 植物和根瘤菌内NO的来源丰富, 主要通过硝酸还原酶(nitrate reductase, NR)和一氧化氮合酶(nitric oxide synthase, NOS)介导以及线粒体电子传递链(electron transport chain, ETC)的还原产生.在植物体内, NO既可由质外体中的亚硝酸根离子(NO2-)转化(Bethke et al., 2004), 又可通过以精氨酸(arginine)、多胺(polyamines)或羟胺(hydroxylamine)作为底物的氧化途径产生(Hichri et al., 2015).在低氧环境下, 还可在还原途径中通过质膜结合亚硝酸盐, 然后通过NO还原酶(nitric oxide reductase, Nor)和黄嘌呤氧化还原酶(xanthine oxidoreductase, XOR)或利用ETC的作用将NO2-还原为NO (Gupta et al., 2011; Mur et al., 2013; Hichri et al., 2015).而Horchani等(2011)发现, 植物及其共生菌中的NR和ETC均可产生NO, 其中NR在植物体内的主要功能是将硝酸盐转化为亚硝酸盐以同化N2 (Neill et al., 2008).但早期的研究发现, 在NAD(P)H作为电子供体时, 大豆(Glycine max)中的NR可将亚硝酸盐转化为NO (Dean and Harper, 1988).且NR抑制剂钨酸盐(tungstate, Tg)也能够抑制NO的产生, 表明NO的合成依赖于NR活性, 但并不是由其直接产生(Horchani et al., 2011).在类菌体中, NO则主要产生于反硝化途径(Hichri et al., 2015), 该途径涉及NR、亚硝酸还原酶(nitrite reductase, NiR)、Nor和NOS (Horchani et al., 2011; Sánchez et al., 2011; Calvo-Begueria et al., 2018) (图1).Berger等(2018)发现NOS也是植物NO的来源, 但目前关于共生菌内NO产生途径的机制研究仍较少. ...
Nitric oxide-responsive genes and promoters in Arabidopsis thaliana: a bioinformatics approach 1 2008
... NO在影响相关基因的表达、调控豆科植物防御反应进而促进共生和结瘤中发挥重要作用.转录组分析表明, NO影响多种植物基因的表达, 如编码富含半胱氨酸蛋白(cysteine-rich proteins, NCRs)、核糖体蛋白、肽酶、结瘤发育细胞周期蛋白及细胞周期转换蛋白的基因(Vinardell et al., 2003; del Giudice et al., 2011; Boscari et al., 2013), 进而调控细胞脱分化、诱导组织形成及抑制植物防御反应(Hichri et al., 2016b).LjHb1是百脉根(Lotus japonicus) ns-Hbs的编码基因, 外源添加NO、缺氧和低温均可诱导其表达(Shimoda et al., 2005).在根瘤形成的早期阶段, NO的产生可以上调LjHb1的表达, 反之LjHb1的表达又下调NO的水平, 以降低植物防御反应, 促进根瘤菌和宿主植物建立共生关系(Nagata et al., 2008; Murakami et al., 2011).植物也可通过降低Hb的表达促进内源NO产生, 调控自身机体内相关防御基因的表达(Wally et al., 2013) (图2).而NO的清除会引起基因MtCRE1和MtCCS52A的下调表达, 从而延缓结瘤(del Giudice et al., 2011).通过对接种中华根瘤菌的蒺藜苜蓿外源添加NO清除剂(2-(4-carboxyphenyl)- 4,4,5,5-tetramethyl imidazoline-1-oxyl-3-oxide, cPTIO), 发现受NO调节的3种基因(Contig525、Contig3307和Medtr5g010350)表达下调(Boscari et al., 2013).其中, 前2种基因分别与ns-Hbs和茉莉酮酸代谢途径(Palmieri et al., 2008)相关, 而最后1种基因可调控还原型谷胱甘肽(glutathione, GSH)的合成(Innocenti et al., 2007).转录组分析显示, 蒺藜苜蓿接种中华根瘤菌后, 外源添加cPTIO或NADPH氧化酶抑制剂二苯烯碘铵(diphenylene iodide, DPI), 导致细胞壁形成和发育过程中相关基因下调表达, 而植物防御和二次代谢相关基因上调表达(Puppo et al., 2013).hmp的过表达和外源添加cPTIO都可降低蒺藜苜蓿的结瘤效率(Kato et al., 2010; del Giudice et al., 2011), NO则通过调节上述基因的表达, 抑制相关防御反应从而促进共生关系的建立.此外, 在根瘤菌内也鉴定出数百个依赖NO表达的基因, 如中华根瘤菌中编码Hmp的hmp基因(del Giudice et al., 2011; Cam et al., 2012)、编码haeme-Cu蛋白的nnrSI和nnrS2基因(Blanquet et al., 2015)及编码NiR的nirKV基因(Meilhoc et al., 2011), 但关于根瘤菌内受NO影响且与共生结瘤相关的基因报道很少. ...
Cadmium induces two waves of reactive oxygen species in Glycine max (L.) roots 1 2014
... NO不仅能调节植物生长发育, 还可参与植物对非生物胁迫的响应(Moreau et al., 2010).目前的研究发现, 在不同非生物胁迫(如干旱和缺氧)下, 共生有机体中的NO均会产生响应(Desalvo et al., 2010; Sánchez et al., 2011; Hawkins et al., 2014; Iarullina et al., 2014; Zimmer-Prados et al., 2014).同时, 在豆科植物中NO还可能参与调控机体对金属元素镉(Cd)和铝(Al)的胁迫反应以及应对非金属元素砷(As)的胁迫(Singh et al., 2015; 尚玉婷等, 2018), 但都缺乏在结瘤和共生固氮体系中的实验证据(Pérez- Chaca et al., 2014).Meilhoc等(2011)提出, 宿主中任何应激反应下NO的产生都会直接影响共生, 在成熟根瘤中, 除了NO专用调节器NnrR外, 多数根瘤菌的基因通过双组分系统(FixLJ)来调节NO, 其中FixLJ是O2限制反应的主要调节因子.nnrR是Nor结构基因norCB的上游基因, 可防止NO这种高活性氮氧化物的积累, 其插入失活可抑制亚硝酸盐的产生以及亚硝酸盐和NO的还原(De Bruijn et al., 2006).NO还能够可逆地结合光系统II (photosystem II, PSII)并调节电子的转移和猝灭, 对光合作用至关重要(Wodala et al., 2008).从农业和进化的角度来看, NO在植物与根瘤菌共生体系中的调控作用, 可以改善植物对病原体的防御状态、保护植物免受虫害以及增强植物对非生物胁迫的适应性(Chadha et al., 2015). ...
Comparison of developmental and stress-indu- ced nodule senescence in Medicago truncatula 2 2010
... 研究表明, 长期黑暗处理能显著诱导根瘤衰老, 同时降低固氮酶活性和Hbs水平(Romanov et al., 1980; Matamoros et al., 1999; Swaraj et al., 2001; Pérez Guerra et al., 2010).持续的暗处理会干扰根瘤功能, 导致根瘤完全衰老(Pérez-Guerra et al., 2010), 而通过降解根瘤中的NO可以延缓黑暗诱导的衰老, 但关于NO在根瘤衰老过程中的作用机制还需进一步研究(Cam et al., 2012).She等(2004)发现在光/暗条件下,对蚕豆(Vicia faba)外源添加cPTIO和L-NAME可减弱H2O2对NO产生的诱导效应和改变人为光照引起的气孔关闭.气孔关闭由多种生物和非生物因素引起, 如渗透胁迫、黑暗、高浓度的CO2和机械压力, 其中光是调节气孔运动最重要的环境因素之一(Zeiger, 1983; Kearns and Assmann, 1993; Herold and Puppo, 2005; Sánchez et al., 2010).但关于黑暗条件下NO调节豆科植物共生固氮的作用机制还未见报道. ...
... ).持续的暗处理会干扰根瘤功能, 导致根瘤完全衰老(Pérez-Guerra et al., 2010), 而通过降解根瘤中的NO可以延缓黑暗诱导的衰老, 但关于NO在根瘤衰老过程中的作用机制还需进一步研究(Cam et al., 2012).She等(2004)发现在光/暗条件下,对蚕豆(Vicia faba)外源添加cPTIO和L-NAME可减弱H2O2对NO产生的诱导效应和改变人为光照引起的气孔关闭.气孔关闭由多种生物和非生物因素引起, 如渗透胁迫、黑暗、高浓度的CO2和机械压力, 其中光是调节气孔运动最重要的环境因素之一(Zeiger, 1983; Kearns and Assmann, 1993; Herold and Puppo, 2005; Sánchez et al., 2010).但关于黑暗条件下NO调节豆科植物共生固氮的作用机制还未见报道. ...
Auxin and nitric oxide control indeterminate nodule formation 1 2007
Nitric oxide, reactive nitrogen species and associated enzymes during plant senescence 1 2011
... 尽管目前对于豆科植物与根瘤菌共生固氮的初始过程已有较多研究, 但因发育或应激引起根瘤衰老的分子机制却报道较少.根瘤衰老区域的表型是由粉红色变为绿色, 其实质源于植物Hbs的降解; 在转录水平上, 衰老表现为编码蛋白质降解, 核酸、膜脂和糖类的相关基因被激活; 同时伴有衰老根瘤中氧化还原平衡的改变, 以及抗氧化防御的整体减弱等现象(Van de Velde et al., 2006; Loscos et al., 2008).NO在植物的衰老过程中具有重要作用(Procházková and Wilhelmová, 2011).例如, NO是水稻(Oryza sativa)光依赖性叶细胞死亡的关键介质(Lin et al., 2012); 蒺藜苜蓿根瘤内NO含量的增加会导致根瘤过早衰老, 且衰老程度与NO水平相关(Cam et al., 2012; Meilhoc et al., 2013; Blanquet et al., 2015), 表明NO对于维持共生体系和避免根瘤过早衰老极为重要.此外, 施加硝酸盐可导致豌豆表现出典型的衰老特征(Escuredo et al., 1996).对蒺藜苜蓿添加硝酸盐后, 其自身也可通过还原硝酸盐促使根瘤中生成NO, 然而硝酸盐引起的根瘤衰老是否依赖于NO目前尚不清楚(Horchani et al., 2011).此外, NO也是一种有效的呼吸抑制剂, 能够抑制线粒体和细菌末端的呼吸氧化酶, 并增加活性氧(reactive oxygen species, ROS)和活性氮(reactive nitrogen species, RNS)的积累, 而NO和ROS均为共生发育所必需(Shimoda et al., 2005; Cam et al., 2012; Igamberdiev et al., 2014; Arjona et al., 2015), 因此衰老也可能是固氮时呼吸抑制的间接结果. ...
Hydrogen peroxide and nitric oxide: key regulators of the legume- Rhizobium and mycorrhizal symbioses 2 2013
... NO在影响相关基因的表达、调控豆科植物防御反应进而促进共生和结瘤中发挥重要作用.转录组分析表明, NO影响多种植物基因的表达, 如编码富含半胱氨酸蛋白(cysteine-rich proteins, NCRs)、核糖体蛋白、肽酶、结瘤发育细胞周期蛋白及细胞周期转换蛋白的基因(Vinardell et al., 2003; del Giudice et al., 2011; Boscari et al., 2013), 进而调控细胞脱分化、诱导组织形成及抑制植物防御反应(Hichri et al., 2016b).LjHb1是百脉根(Lotus japonicus) ns-Hbs的编码基因, 外源添加NO、缺氧和低温均可诱导其表达(Shimoda et al., 2005).在根瘤形成的早期阶段, NO的产生可以上调LjHb1的表达, 反之LjHb1的表达又下调NO的水平, 以降低植物防御反应, 促进根瘤菌和宿主植物建立共生关系(Nagata et al., 2008; Murakami et al., 2011).植物也可通过降低Hb的表达促进内源NO产生, 调控自身机体内相关防御基因的表达(Wally et al., 2013) (图2).而NO的清除会引起基因MtCRE1和MtCCS52A的下调表达, 从而延缓结瘤(del Giudice et al., 2011).通过对接种中华根瘤菌的蒺藜苜蓿外源添加NO清除剂(2-(4-carboxyphenyl)- 4,4,5,5-tetramethyl imidazoline-1-oxyl-3-oxide, cPTIO), 发现受NO调节的3种基因(Contig525、Contig3307和Medtr5g010350)表达下调(Boscari et al., 2013).其中, 前2种基因分别与ns-Hbs和茉莉酮酸代谢途径(Palmieri et al., 2008)相关, 而最后1种基因可调控还原型谷胱甘肽(glutathione, GSH)的合成(Innocenti et al., 2007).转录组分析显示, 蒺藜苜蓿接种中华根瘤菌后, 外源添加cPTIO或NADPH氧化酶抑制剂二苯烯碘铵(diphenylene iodide, DPI), 导致细胞壁形成和发育过程中相关基因下调表达, 而植物防御和二次代谢相关基因上调表达(Puppo et al., 2013).hmp的过表达和外源添加cPTIO都可降低蒺藜苜蓿的结瘤效率(Kato et al., 2010; del Giudice et al., 2011), NO则通过调节上述基因的表达, 抑制相关防御反应从而促进共生关系的建立.此外, 在根瘤菌内也鉴定出数百个依赖NO表达的基因, 如中华根瘤菌中编码Hmp的hmp基因(del Giudice et al., 2011; Cam et al., 2012)、编码haeme-Cu蛋白的nnrSI和nnrS2基因(Blanquet et al., 2015)及编码NiR的nirKV基因(Meilhoc et al., 2011), 但关于根瘤菌内受NO影响且与共生结瘤相关的基因报道很少. ...
... NO可通过调节共生体的能量代谢影响N2固定.固氮过程中会产生铵根离子(ammonium ion, NH4+), 当其达到一定浓度时即具有毒性并损害植物的生长(Li et al., 2014).GS可同化NH4+, 解除这种离子胁迫, 而NO通过Tyr硝化改变其活性, 因此在N代谢中NO与GS起着至关重要的作用(Melo et al., 2011; Sainz et al., 2015).S-亚硝基化也是植物体内NO信号转导的关键机制.研究发现蒺藜苜蓿成熟根瘤中约有80种S-亚硝基化蛋白, 且多数与C、N代谢及固氮酶有关, 如果糖激酶、固氮酶钼铁(MoFe)蛋白nifK与铁(Fe)蛋白nifH (Puppo et al., 2013).NO可能通过影响根瘤中与S-亚硝基化蛋白相关的酶活性, 利用三羧酸循环(tricarboxylic acid cycle, TCA cycle)和糖酵解(glycolysis)等共生体的能量代谢过程调控固氮酶活性(Igamberdiev and Hill, 2009; Boscari et al., 2013; Li et al., 2014; Igamberdiev et al., 2014) (图2).但对于NO究竟是直接抑制固氮酶活性还是通过间接调节C、N代谢影响豆科植物体固氮, 目前尚不清楚(Hichri et al., 2016b). ...
Nitric oxide, oxidants, and protein tyrosine nitration 1 2004
... 脱落酸(abscisic acid, ABA)是植物适应环境胁迫的关键激素(Yoshida et al., 2015), 可通过抑制根毛变形及侵染线的形成影响豆科植物结瘤(Suzuki et al., 2004; Ding et al., 2008).Tominaga等(2010)在百脉根中发现, ABA不仅可调节根瘤的固氮酶活性, 还可通过影响结瘤过程中NO的产生来影响结瘤.在根瘤形成过程中, NO通过酪氨酸(tyrosine, Tyr)硝化调控ABA信号传递, 而ABA又在信号转导过程中决定NO的产生与积累(Castillo et al., 2015).Tyr硝化是NO介导的翻译后修饰, 其特征在于将-NO2添加到Tyr残基芳环的等价邻位C上, 产生3-硝基酪氨酸(3-nitrotyrosine, 3-NT), 并由此改变蛋白质构象(Radi, 2004).作为硝化分子的前体, NO通过Tyr硝化改变细胞溶质谷氨酰胺合成酶(glutamine synthetase 1, GS1)和Lb等关键结瘤蛋白的活性(Melo et al., 2011; Navascués et al., 2012; Li et al., 2014).豆科植物结瘤时Lb发生Tyr硝化, 可保护共生体免受过氧亚硝酸盐毒害, 因此3-NT的形成既是硝基氧化应激的标志, 也是功能性结瘤活跃代谢的结果(Sainz et al., 2015).此外, NOS抑制剂N-硝基-L-精氨酸甲酯(NG-nitro- L-arginine methyl ester, L-NAME)和cPTIO处理可减弱豌豆(Pisum sativum)表皮由ABA诱导的气孔关闭(张绪成等, 2005). ...
Metabolism of poly-hydroxybutyric acid in bacteroids of Rhizobium lupini in connection with nitrogen ?xation and photosynthesis 1 1980
... 研究表明, 长期黑暗处理能显著诱导根瘤衰老, 同时降低固氮酶活性和Hbs水平(Romanov et al., 1980; Matamoros et al., 1999; Swaraj et al., 2001; Pérez Guerra et al., 2010).持续的暗处理会干扰根瘤功能, 导致根瘤完全衰老(Pérez-Guerra et al., 2010), 而通过降解根瘤中的NO可以延缓黑暗诱导的衰老, 但关于NO在根瘤衰老过程中的作用机制还需进一步研究(Cam et al., 2012).She等(2004)发现在光/暗条件下,对蚕豆(Vicia faba)外源添加cPTIO和L-NAME可减弱H2O2对NO产生的诱导效应和改变人为光照引起的气孔关闭.气孔关闭由多种生物和非生物因素引起, 如渗透胁迫、黑暗、高浓度的CO2和机械压力, 其中光是调节气孔运动最重要的环境因素之一(Zeiger, 1983; Kearns and Assmann, 1993; Herold and Puppo, 2005; Sánchez et al., 2010).但关于黑暗条件下NO调节豆科植物共生固氮的作用机制还未见报道. ...
Leghemoglobin is nitrated in functional legume nodules in a tyrosine residue within the heme cavity by a nitrite/peroxide-dependent mechanism 2 2015
... 脱落酸(abscisic acid, ABA)是植物适应环境胁迫的关键激素(Yoshida et al., 2015), 可通过抑制根毛变形及侵染线的形成影响豆科植物结瘤(Suzuki et al., 2004; Ding et al., 2008).Tominaga等(2010)在百脉根中发现, ABA不仅可调节根瘤的固氮酶活性, 还可通过影响结瘤过程中NO的产生来影响结瘤.在根瘤形成过程中, NO通过酪氨酸(tyrosine, Tyr)硝化调控ABA信号传递, 而ABA又在信号转导过程中决定NO的产生与积累(Castillo et al., 2015).Tyr硝化是NO介导的翻译后修饰, 其特征在于将-NO2添加到Tyr残基芳环的等价邻位C上, 产生3-硝基酪氨酸(3-nitrotyrosine, 3-NT), 并由此改变蛋白质构象(Radi, 2004).作为硝化分子的前体, NO通过Tyr硝化改变细胞溶质谷氨酰胺合成酶(glutamine synthetase 1, GS1)和Lb等关键结瘤蛋白的活性(Melo et al., 2011; Navascués et al., 2012; Li et al., 2014).豆科植物结瘤时Lb发生Tyr硝化, 可保护共生体免受过氧亚硝酸盐毒害, 因此3-NT的形成既是硝基氧化应激的标志, 也是功能性结瘤活跃代谢的结果(Sainz et al., 2015).此外, NOS抑制剂N-硝基-L-精氨酸甲酯(NG-nitro- L-arginine methyl ester, L-NAME)和cPTIO处理可减弱豌豆(Pisum sativum)表皮由ABA诱导的气孔关闭(张绪成等, 2005). ...
... NO可通过调节共生体的能量代谢影响N2固定.固氮过程中会产生铵根离子(ammonium ion, NH4+), 当其达到一定浓度时即具有毒性并损害植物的生长(Li et al., 2014).GS可同化NH4+, 解除这种离子胁迫, 而NO通过Tyr硝化改变其活性, 因此在N代谢中NO与GS起着至关重要的作用(Melo et al., 2011; Sainz et al., 2015).S-亚硝基化也是植物体内NO信号转导的关键机制.研究发现蒺藜苜蓿成熟根瘤中约有80种S-亚硝基化蛋白, 且多数与C、N代谢及固氮酶有关, 如果糖激酶、固氮酶钼铁(MoFe)蛋白nifK与铁(Fe)蛋白nifH (Puppo et al., 2013).NO可能通过影响根瘤中与S-亚硝基化蛋白相关的酶活性, 利用三羧酸循环(tricarboxylic acid cycle, TCA cycle)和糖酵解(glycolysis)等共生体的能量代谢过程调控固氮酶活性(Igamberdiev and Hill, 2009; Boscari et al., 2013; Li et al., 2014; Igamberdiev et al., 2014) (图2).但对于NO究竟是直接抑制固氮酶活性还是通过间接调节C、N代谢影响豆科植物体固氮, 目前尚不清楚(Hichri et al., 2016b). ...
Nitric oxide detoxification in the rhizobia-legume symbiosis 4 2011
... 植物和根瘤菌内NO的来源丰富, 主要通过硝酸还原酶(nitrate reductase, NR)和一氧化氮合酶(nitric oxide synthase, NOS)介导以及线粒体电子传递链(electron transport chain, ETC)的还原产生.在植物体内, NO既可由质外体中的亚硝酸根离子(NO2-)转化(Bethke et al., 2004), 又可通过以精氨酸(arginine)、多胺(polyamines)或羟胺(hydroxylamine)作为底物的氧化途径产生(Hichri et al., 2015).在低氧环境下, 还可在还原途径中通过质膜结合亚硝酸盐, 然后通过NO还原酶(nitric oxide reductase, Nor)和黄嘌呤氧化还原酶(xanthine oxidoreductase, XOR)或利用ETC的作用将NO2-还原为NO (Gupta et al., 2011; Mur et al., 2013; Hichri et al., 2015).而Horchani等(2011)发现, 植物及其共生菌中的NR和ETC均可产生NO, 其中NR在植物体内的主要功能是将硝酸盐转化为亚硝酸盐以同化N2 (Neill et al., 2008).但早期的研究发现, 在NAD(P)H作为电子供体时, 大豆(Glycine max)中的NR可将亚硝酸盐转化为NO (Dean and Harper, 1988).且NR抑制剂钨酸盐(tungstate, Tg)也能够抑制NO的产生, 表明NO的合成依赖于NR活性, 但并不是由其直接产生(Horchani et al., 2011).在类菌体中, NO则主要产生于反硝化途径(Hichri et al., 2015), 该途径涉及NR、亚硝酸还原酶(nitrite reductase, NiR)、Nor和NOS (Horchani et al., 2011; Sánchez et al., 2011; Calvo-Begueria et al., 2018) (图1).Berger等(2018)发现NOS也是植物NO的来源, 但目前关于共生菌内NO产生途径的机制研究仍较少. ...
... 豆科植物根瘤中包含一种与其它蛋白性质和功能不同的血红蛋白(hemoglobins, Hbs), 能够清除机体产生或累积的NO (Gupta et al., 2011).基于其序列同源性和对氧的亲和力, 可将植物体内的Hbs划分成3类: 非共生血红蛋白(non-symbiotic hemoglobins, ns-Hbs, 1类)、豆血红蛋白(leghemoglobins, Lbs, 2类)和截短血红蛋白(truncated hemoglobin, Tr-Hbs, 3类) (Gupta et al., 2011; Hill, 2012).Lbs作为高等植物中发现的第1种豆血红蛋白, 在豆科植物中主要负责将根瘤中的O2运输至根瘤菌(Trevaskis et al., 1997).而Hbs对O2和NO的亲和力非常强, 能够有效清除O2和NO并将其转化为硝酸盐.有研究表明, Hbs能够维持植物细胞在缺氧状态下的氧化还原能力和能量状态(Igamberdiev and Hill, 2009).除植物体蛋白外, 根瘤菌内也含有3类血红蛋白: 黄素血红蛋白(flavohaemoglobins, fHbs/Hmp)、单域血红蛋白(single-domain haemoglobins, sd-Hbs)和截短血红蛋白(Sánchez et al., 2011).Hmp是蒺藜苜蓿(Medicago truncatula)中主要的NO清除剂, 而NO可诱导hmp基因表达以影响自身生成量(Meilhoc et al., 2011; Cam et al., 2012) (图1).另有研究表明, 在蒺藜苜蓿hmp突变的根瘤中, NO水平显著高于野生型, 且固氮效率降低和根瘤衰老提前(Cam et al., 2012; Hichri et al., 2016a).因此, 植物中的Hbs与菌内的Hmp对于维持共生体NO水平至关重要. ...
... 根瘤中的NO可通过调控酶的翻译后修饰调节碳(C)、氮(N)及能量代谢(Chaki et al., 2014; Hu et al., 2015).NO的产生是植物应对缺氧胁迫的一种响应, 与呼吸循环过程相关联, 形成“Hb/NO”循环.缺氧条件下, Hb/NO循环可维持细胞内的氧化还原和能量平衡状态(Igamberdiev and Hill, 2004; Gupta et al., 2011).Hb/NO呼吸循环包括4个步骤: (1) 硝酸根离子(nitrate ion, NO3-)被NR还原为NO2-; (2) NO2-从细胞质转移到线粒体基质; (3) 通过线粒体ETC将NO2-还原为NO, 使ATP再生; (4) 最终NO通过被动扩散到胞质溶胶中, 由Hb将其氧化成硝酸盐(Gupta et al., 2011; Horchani et al., 2011; Hichri et al., 2015).因此, 在低氧环境下, 植物线粒体可以保持其氧化NADH的能力, 从而合成ATP.相关研究也表明, 缺氧环境下, 大豆和苜蓿的成熟根瘤通过硝酸还原途径和细菌反硝化途径均能产生NO, 参与Hb/NO呼吸循环, 为机体提供能量(Meakin et al., 2007; Horchani et al., 2011; Sánchez et al., 2011).此外, 在缺氧条件下, 结瘤的能量状态几乎全部取决于NR的功能(Horchani et al., 2011).值得注意的是, 在Hb/NO循环过程中, 由于多数情况下处于缺氧状态, 因此最终的电子受体是亚硝酸盐而非O2 (Gupta et al., 2011; Igamberdiev et al., 2014).Hichri等(2015)发现, NO在结瘤过程中具有双重作用: 一方面作为维持基础能量代谢所必需的中间物; 另一方面作为C、N代谢的调节剂, 降低微氧环境下的能量需求.由于缺氧对于植物的生长发育影响很大, 因此当O2不足时, NO对维持豆科植物与根瘤菌共生结瘤的氧化还原和能量平衡状态至关重要. ...
... NO不仅能调节植物生长发育, 还可参与植物对非生物胁迫的响应(Moreau et al., 2010).目前的研究发现, 在不同非生物胁迫(如干旱和缺氧)下, 共生有机体中的NO均会产生响应(Desalvo et al., 2010; Sánchez et al., 2011; Hawkins et al., 2014; Iarullina et al., 2014; Zimmer-Prados et al., 2014).同时, 在豆科植物中NO还可能参与调控机体对金属元素镉(Cd)和铝(Al)的胁迫反应以及应对非金属元素砷(As)的胁迫(Singh et al., 2015; 尚玉婷等, 2018), 但都缺乏在结瘤和共生固氮体系中的实验证据(Pérez- Chaca et al., 2014).Meilhoc等(2011)提出, 宿主中任何应激反应下NO的产生都会直接影响共生, 在成熟根瘤中, 除了NO专用调节器NnrR外, 多数根瘤菌的基因通过双组分系统(FixLJ)来调节NO, 其中FixLJ是O2限制反应的主要调节因子.nnrR是Nor结构基因norCB的上游基因, 可防止NO这种高活性氮氧化物的积累, 其插入失活可抑制亚硝酸盐的产生以及亚硝酸盐和NO的还原(De Bruijn et al., 2006).NO还能够可逆地结合光系统II (photosystem II, PSII)并调节电子的转移和猝灭, 对光合作用至关重要(Wodala et al., 2008).从农业和进化的角度来看, NO在植物与根瘤菌共生体系中的调控作用, 可以改善植物对病原体的防御状态、保护植物免受虫害以及增强植物对非生物胁迫的适应性(Chadha et al., 2015). ...
Production of nitric oxide and nitrosylleghemoglobin complexes in soybean nodules in response to flooding 5 2010
... 研究表明, 长期黑暗处理能显著诱导根瘤衰老, 同时降低固氮酶活性和Hbs水平(Romanov et al., 1980; Matamoros et al., 1999; Swaraj et al., 2001; Pérez Guerra et al., 2010).持续的暗处理会干扰根瘤功能, 导致根瘤完全衰老(Pérez-Guerra et al., 2010), 而通过降解根瘤中的NO可以延缓黑暗诱导的衰老, 但关于NO在根瘤衰老过程中的作用机制还需进一步研究(Cam et al., 2012).She等(2004)发现在光/暗条件下,对蚕豆(Vicia faba)外源添加cPTIO和L-NAME可减弱H2O2对NO产生的诱导效应和改变人为光照引起的气孔关闭.气孔关闭由多种生物和非生物因素引起, 如渗透胁迫、黑暗、高浓度的CO2和机械压力, 其中光是调节气孔运动最重要的环境因素之一(Zeiger, 1983; Kearns and Assmann, 1993; Herold and Puppo, 2005; Sánchez et al., 2010).但关于黑暗条件下NO调节豆科植物共生固氮的作用机制还未见报道. ...
... NO还参与豆科植物对缺氧胁迫的反应.缺氧胁迫可诱导百脉根根瘤产生亚硝酰基豆血红蛋白(nitrosylleghemoglobin, LbNO)以减少根瘤内的硝酸盐, 同时LbNO的形成也有利于解除NO对植物体的毒害(Meakin et al., 2007).根瘤中LbNOs的增加不会抑制植物的固氮作用, 其实质源于Lb在根瘤中既可清除由细菌反硝化作用产生的NO和亚硝酸盐, 又可保护淹水条件下的固氮酶活性(Sánchez et al., 2010).Sánchez等(2010)发现, 在水淹缺氧条件下, 野生型和norC突变体根瘤中编码固氮酶Fe蛋白的nifH基因表达量降低、固氮酶活性下降, 通过清除NO可抵消这种影响.而当从常氧转为缺氧时, 根瘤内NO迅速增加, 表明根瘤中不仅能产生NO, 还具有响应缺氧胁迫而迅速上调表达的基因(Sánchez et al., 2010; Horchani et al., 2011).因此, 无论在转录水平还是翻译水平, 固氮酶都是NO抑制N2固定的关键靶标(Sánchez et al., 2010), 同时也表明在微氧环境中, NO主要通过影响固氮酶活性来调节植物体固氮. ...
... ).Sánchez等(2010)发现, 在水淹缺氧条件下, 野生型和norC突变体根瘤中编码固氮酶Fe蛋白的nifH基因表达量降低、固氮酶活性下降, 通过清除NO可抵消这种影响.而当从常氧转为缺氧时, 根瘤内NO迅速增加, 表明根瘤中不仅能产生NO, 还具有响应缺氧胁迫而迅速上调表达的基因(Sánchez et al., 2010; Horchani et al., 2011).因此, 无论在转录水平还是翻译水平, 固氮酶都是NO抑制N2固定的关键靶标(Sánchez et al., 2010), 同时也表明在微氧环境中, NO主要通过影响固氮酶活性来调节植物体固氮. ...
... 基因表达量降低、固氮酶活性下降, 通过清除NO可抵消这种影响.而当从常氧转为缺氧时, 根瘤内NO迅速增加, 表明根瘤中不仅能产生NO, 还具有响应缺氧胁迫而迅速上调表达的基因(Sánchez et al., 2010; Horchani et al., 2011).因此, 无论在转录水平还是翻译水平, 固氮酶都是NO抑制N2固定的关键靶标(Sánchez et al., 2010), 同时也表明在微氧环境中, NO主要通过影响固氮酶活性来调节植物体固氮. ...
... 固定的关键靶标(Sánchez et al., 2010), 同时也表明在微氧环境中, NO主要通过影响固氮酶活性来调节植物体固氮. ...
Role and relationship of nitric oxide and hydrogen peroxide in light/dark-regulated stomatal movement in Vicia faba 1 2004
... 研究表明, 长期黑暗处理能显著诱导根瘤衰老, 同时降低固氮酶活性和Hbs水平(Romanov et al., 1980; Matamoros et al., 1999; Swaraj et al., 2001; Pérez Guerra et al., 2010).持续的暗处理会干扰根瘤功能, 导致根瘤完全衰老(Pérez-Guerra et al., 2010), 而通过降解根瘤中的NO可以延缓黑暗诱导的衰老, 但关于NO在根瘤衰老过程中的作用机制还需进一步研究(Cam et al., 2012).She等(2004)发现在光/暗条件下,对蚕豆(Vicia faba)外源添加cPTIO和L-NAME可减弱H2O2对NO产生的诱导效应和改变人为光照引起的气孔关闭.气孔关闭由多种生物和非生物因素引起, 如渗透胁迫、黑暗、高浓度的CO2和机械压力, 其中光是调节气孔运动最重要的环境因素之一(Zeiger, 1983; Kearns and Assmann, 1993; Herold and Puppo, 2005; Sánchez et al., 2010).但关于黑暗条件下NO调节豆科植物共生固氮的作用机制还未见报道. ...
Symbiotic rhizobium and nitric oxide induce gene expression of non- symbiotic hemoglobin in Lotus japonicus 4 2005
... 豆科植物根瘤菌共生体系受多种信号分子和转录因子调控, 其中一氧化氮(nitric oxide, NO)信号分子在豆科植物微生物互作过程中发挥重要作用.共生体早期的转录组学分析表明, NO可调节细胞脱分化和器官发生(Ferrarini et al., 2008; Boscari et al., 2013), 并抑制植物防御反应(Gonzalez-Rizzo et al., 2006), 在建立植物与菌共生关系中发挥重要调控作用(Hichri et al., 2016b).而在成熟根瘤中, NO既可抑制植物体固氮(Shimoda et al., 2005; Kato et al., 2010; Cam et al., 2012), 又可诱发根瘤衰老(Horchani et al., 2011; Cam et al., 2012; Blanquet et al., 2015), 亦可作为能量代谢调节器维持低氧(oxygen, O2)状态下的能量平衡(Kato et al., 2010; Hichri et al., 2016b).尽管目前关于NO在植物体内的生理功能已进行了充分研究, 但国内对于其在豆科植物根瘤菌共生体系中的作用机制并未进行系统的阐述.本文综述了豆科植物根瘤菌共生体系中NO的产生和降解, 及其对根瘤形成与共生固氮影响的研究进展, 并对NO在共生体系中对非生物胁迫的响应及其与激素的相互作用进行了阐述. ...
... NO在影响相关基因的表达、调控豆科植物防御反应进而促进共生和结瘤中发挥重要作用.转录组分析表明, NO影响多种植物基因的表达, 如编码富含半胱氨酸蛋白(cysteine-rich proteins, NCRs)、核糖体蛋白、肽酶、结瘤发育细胞周期蛋白及细胞周期转换蛋白的基因(Vinardell et al., 2003; del Giudice et al., 2011; Boscari et al., 2013), 进而调控细胞脱分化、诱导组织形成及抑制植物防御反应(Hichri et al., 2016b).LjHb1是百脉根(Lotus japonicus) ns-Hbs的编码基因, 外源添加NO、缺氧和低温均可诱导其表达(Shimoda et al., 2005).在根瘤形成的早期阶段, NO的产生可以上调LjHb1的表达, 反之LjHb1的表达又下调NO的水平, 以降低植物防御反应, 促进根瘤菌和宿主植物建立共生关系(Nagata et al., 2008; Murakami et al., 2011).植物也可通过降低Hb的表达促进内源NO产生, 调控自身机体内相关防御基因的表达(Wally et al., 2013) (图2).而NO的清除会引起基因MtCRE1和MtCCS52A的下调表达, 从而延缓结瘤(del Giudice et al., 2011).通过对接种中华根瘤菌的蒺藜苜蓿外源添加NO清除剂(2-(4-carboxyphenyl)- 4,4,5,5-tetramethyl imidazoline-1-oxyl-3-oxide, cPTIO), 发现受NO调节的3种基因(Contig525、Contig3307和Medtr5g010350)表达下调(Boscari et al., 2013).其中, 前2种基因分别与ns-Hbs和茉莉酮酸代谢途径(Palmieri et al., 2008)相关, 而最后1种基因可调控还原型谷胱甘肽(glutathione, GSH)的合成(Innocenti et al., 2007).转录组分析显示, 蒺藜苜蓿接种中华根瘤菌后, 外源添加cPTIO或NADPH氧化酶抑制剂二苯烯碘铵(diphenylene iodide, DPI), 导致细胞壁形成和发育过程中相关基因下调表达, 而植物防御和二次代谢相关基因上调表达(Puppo et al., 2013).hmp的过表达和外源添加cPTIO都可降低蒺藜苜蓿的结瘤效率(Kato et al., 2010; del Giudice et al., 2011), NO则通过调节上述基因的表达, 抑制相关防御反应从而促进共生关系的建立.此外, 在根瘤菌内也鉴定出数百个依赖NO表达的基因, 如中华根瘤菌中编码Hmp的hmp基因(del Giudice et al., 2011; Cam et al., 2012)、编码haeme-Cu蛋白的nnrSI和nnrS2基因(Blanquet et al., 2015)及编码NiR的nirKV基因(Meilhoc et al., 2011), 但关于根瘤菌内受NO影响且与共生结瘤相关的基因报道很少. ...
... 研究表明, 外源添加NO可抑制大豆与百脉根根瘤的固氮酶活性(Kato et al., 2010; Cam et al., 2012; Boscari et al., 2013).Cam等(2012)报道, 成熟根瘤中NO的积累不仅可降低固氮效率, 还会影响植物的生长适应性, 而通过降解NO可增强植物体固氮能力和延长植物整体固氮周期.研究发现, hmp的过表达降低了根瘤中NO的含量, 却增强了乙炔还原活性(acetylene-reducing activity, ARA)和延迟根瘤衰老(Meilhoc et al., 2011; Cam et al., 2012).但NO的毒性、信号功能或代谢效应取决于其所在作用部位的NO浓度(Boscari et al., 2013; Mur et al., 2013).例如, 在百脉根中, 高浓度的NO可抑制氮的固定, 适当低浓度的NO则可增强固氮作用(Kato et al., 2010).因此, 较低且稳定浓度的NO是维持豆科植物根瘤形成和发育的重要因素(Shimoda et al., 2005; Cam et al., 2012; Calvo-Begueria et al., 2018). ...
... 尽管目前对于豆科植物与根瘤菌共生固氮的初始过程已有较多研究, 但因发育或应激引起根瘤衰老的分子机制却报道较少.根瘤衰老区域的表型是由粉红色变为绿色, 其实质源于植物Hbs的降解; 在转录水平上, 衰老表现为编码蛋白质降解, 核酸、膜脂和糖类的相关基因被激活; 同时伴有衰老根瘤中氧化还原平衡的改变, 以及抗氧化防御的整体减弱等现象(Van de Velde et al., 2006; Loscos et al., 2008).NO在植物的衰老过程中具有重要作用(Procházková and Wilhelmová, 2011).例如, NO是水稻(Oryza sativa)光依赖性叶细胞死亡的关键介质(Lin et al., 2012); 蒺藜苜蓿根瘤内NO含量的增加会导致根瘤过早衰老, 且衰老程度与NO水平相关(Cam et al., 2012; Meilhoc et al., 2013; Blanquet et al., 2015), 表明NO对于维持共生体系和避免根瘤过早衰老极为重要.此外, 施加硝酸盐可导致豌豆表现出典型的衰老特征(Escuredo et al., 1996).对蒺藜苜蓿添加硝酸盐后, 其自身也可通过还原硝酸盐促使根瘤中生成NO, 然而硝酸盐引起的根瘤衰老是否依赖于NO目前尚不清楚(Horchani et al., 2011).此外, NO也是一种有效的呼吸抑制剂, 能够抑制线粒体和细菌末端的呼吸氧化酶, 并增加活性氧(reactive oxygen species, ROS)和活性氮(reactive nitrogen species, RNS)的积累, 而NO和ROS均为共生发育所必需(Shimoda et al., 2005; Cam et al., 2012; Igamberdiev et al., 2014; Arjona et al., 2015), 因此衰老也可能是固氮时呼吸抑制的间接结果. ...
Hydrogen sulfide alleviates toxic effects of arsenate in pea seedlings through up-regulation of the ascorbate-glutathione cycle: possible involvement of nitric oxide 1 2015
... NO不仅能调节植物生长发育, 还可参与植物对非生物胁迫的响应(Moreau et al., 2010).目前的研究发现, 在不同非生物胁迫(如干旱和缺氧)下, 共生有机体中的NO均会产生响应(Desalvo et al., 2010; Sánchez et al., 2011; Hawkins et al., 2014; Iarullina et al., 2014; Zimmer-Prados et al., 2014).同时, 在豆科植物中NO还可能参与调控机体对金属元素镉(Cd)和铝(Al)的胁迫反应以及应对非金属元素砷(As)的胁迫(Singh et al., 2015; 尚玉婷等, 2018), 但都缺乏在结瘤和共生固氮体系中的实验证据(Pérez- Chaca et al., 2014).Meilhoc等(2011)提出, 宿主中任何应激反应下NO的产生都会直接影响共生, 在成熟根瘤中, 除了NO专用调节器NnrR外, 多数根瘤菌的基因通过双组分系统(FixLJ)来调节NO, 其中FixLJ是O2限制反应的主要调节因子.nnrR是Nor结构基因norCB的上游基因, 可防止NO这种高活性氮氧化物的积累, 其插入失活可抑制亚硝酸盐的产生以及亚硝酸盐和NO的还原(De Bruijn et al., 2006).NO还能够可逆地结合光系统II (photosystem II, PSII)并调节电子的转移和猝灭, 对光合作用至关重要(Wodala et al., 2008).从农业和进化的角度来看, NO在植物与根瘤菌共生体系中的调控作用, 可以改善植物对病原体的防御状态、保护植物免受虫害以及增强植物对非生物胁迫的适应性(Chadha et al., 2015). ...
Control of nodule number by the phytohormone abscisic acid in the roots of two leguminous species 1 2004
... 脱落酸(abscisic acid, ABA)是植物适应环境胁迫的关键激素(Yoshida et al., 2015), 可通过抑制根毛变形及侵染线的形成影响豆科植物结瘤(Suzuki et al., 2004; Ding et al., 2008).Tominaga等(2010)在百脉根中发现, ABA不仅可调节根瘤的固氮酶活性, 还可通过影响结瘤过程中NO的产生来影响结瘤.在根瘤形成过程中, NO通过酪氨酸(tyrosine, Tyr)硝化调控ABA信号传递, 而ABA又在信号转导过程中决定NO的产生与积累(Castillo et al., 2015).Tyr硝化是NO介导的翻译后修饰, 其特征在于将-NO2添加到Tyr残基芳环的等价邻位C上, 产生3-硝基酪氨酸(3-nitrotyrosine, 3-NT), 并由此改变蛋白质构象(Radi, 2004).作为硝化分子的前体, NO通过Tyr硝化改变细胞溶质谷氨酰胺合成酶(glutamine synthetase 1, GS1)和Lb等关键结瘤蛋白的活性(Melo et al., 2011; Navascués et al., 2012; Li et al., 2014).豆科植物结瘤时Lb发生Tyr硝化, 可保护共生体免受过氧亚硝酸盐毒害, 因此3-NT的形成既是硝基氧化应激的标志, 也是功能性结瘤活跃代谢的结果(Sainz et al., 2015).此外, NOS抑制剂N-硝基-L-精氨酸甲酯(NG-nitro- L-arginine methyl ester, L-NAME)和cPTIO处理可减弱豌豆(Pisum sativum)表皮由ABA诱导的气孔关闭(张绪成等, 2005). ...
Dark-induced changes in legume nodule functioning 1 2001
... 研究表明, 长期黑暗处理能显著诱导根瘤衰老, 同时降低固氮酶活性和Hbs水平(Romanov et al., 1980; Matamoros et al., 1999; Swaraj et al., 2001; Pérez Guerra et al., 2010).持续的暗处理会干扰根瘤功能, 导致根瘤完全衰老(Pérez-Guerra et al., 2010), 而通过降解根瘤中的NO可以延缓黑暗诱导的衰老, 但关于NO在根瘤衰老过程中的作用机制还需进一步研究(Cam et al., 2012).She等(2004)发现在光/暗条件下,对蚕豆(Vicia faba)外源添加cPTIO和L-NAME可减弱H2O2对NO产生的诱导效应和改变人为光照引起的气孔关闭.气孔关闭由多种生物和非生物因素引起, 如渗透胁迫、黑暗、高浓度的CO2和机械压力, 其中光是调节气孔运动最重要的环境因素之一(Zeiger, 1983; Kearns and Assmann, 1993; Herold and Puppo, 2005; Sánchez et al., 2010).但关于黑暗条件下NO调节豆科植物共生固氮的作用机制还未见报道. ...
Effect of abscisic acid on symbiotic nitrogen fixation activity in the root nodules of Lotus japonicus 1 2010
... 脱落酸(abscisic acid, ABA)是植物适应环境胁迫的关键激素(Yoshida et al., 2015), 可通过抑制根毛变形及侵染线的形成影响豆科植物结瘤(Suzuki et al., 2004; Ding et al., 2008).Tominaga等(2010)在百脉根中发现, ABA不仅可调节根瘤的固氮酶活性, 还可通过影响结瘤过程中NO的产生来影响结瘤.在根瘤形成过程中, NO通过酪氨酸(tyrosine, Tyr)硝化调控ABA信号传递, 而ABA又在信号转导过程中决定NO的产生与积累(Castillo et al., 2015).Tyr硝化是NO介导的翻译后修饰, 其特征在于将-NO2添加到Tyr残基芳环的等价邻位C上, 产生3-硝基酪氨酸(3-nitrotyrosine, 3-NT), 并由此改变蛋白质构象(Radi, 2004).作为硝化分子的前体, NO通过Tyr硝化改变细胞溶质谷氨酰胺合成酶(glutamine synthetase 1, GS1)和Lb等关键结瘤蛋白的活性(Melo et al., 2011; Navascués et al., 2012; Li et al., 2014).豆科植物结瘤时Lb发生Tyr硝化, 可保护共生体免受过氧亚硝酸盐毒害, 因此3-NT的形成既是硝基氧化应激的标志, 也是功能性结瘤活跃代谢的结果(Sainz et al., 2015).此外, NOS抑制剂N-硝基-L-精氨酸甲酯(NG-nitro- L-arginine methyl ester, L-NAME)和cPTIO处理可减弱豌豆(Pisum sativum)表皮由ABA诱导的气孔关闭(张绪成等, 2005). ...
Two hemoglobin genes in Arabidopsis thaliana: the evolutionary origins of leghemoglobins 1 1997
... 豆科植物根瘤中包含一种与其它蛋白性质和功能不同的血红蛋白(hemoglobins, Hbs), 能够清除机体产生或累积的NO (Gupta et al., 2011).基于其序列同源性和对氧的亲和力, 可将植物体内的Hbs划分成3类: 非共生血红蛋白(non-symbiotic hemoglobins, ns-Hbs, 1类)、豆血红蛋白(leghemoglobins, Lbs, 2类)和截短血红蛋白(truncated hemoglobin, Tr-Hbs, 3类) (Gupta et al., 2011; Hill, 2012).Lbs作为高等植物中发现的第1种豆血红蛋白, 在豆科植物中主要负责将根瘤中的O2运输至根瘤菌(Trevaskis et al., 1997).而Hbs对O2和NO的亲和力非常强, 能够有效清除O2和NO并将其转化为硝酸盐.有研究表明, Hbs能够维持植物细胞在缺氧状态下的氧化还原能力和能量状态(Igamberdiev and Hill, 2009).除植物体蛋白外, 根瘤菌内也含有3类血红蛋白: 黄素血红蛋白(flavohaemoglobins, fHbs/Hmp)、单域血红蛋白(single-domain haemoglobins, sd-Hbs)和截短血红蛋白(Sánchez et al., 2011).Hmp是蒺藜苜蓿(Medicago truncatula)中主要的NO清除剂, 而NO可诱导hmp基因表达以影响自身生成量(Meilhoc et al., 2011; Cam et al., 2012) (图1).另有研究表明, 在蒺藜苜蓿hmp突变的根瘤中, NO水平显著高于野生型, 且固氮效率降低和根瘤衰老提前(Cam et al., 2012; Hichri et al., 2016a).因此, 植物中的Hbs与菌内的Hmp对于维持共生体NO水平至关重要. ...
Aging in legume symbiosis. A molecular view on nodule senescence in Medicago truncatula 1 2006
... 尽管目前对于豆科植物与根瘤菌共生固氮的初始过程已有较多研究, 但因发育或应激引起根瘤衰老的分子机制却报道较少.根瘤衰老区域的表型是由粉红色变为绿色, 其实质源于植物Hbs的降解; 在转录水平上, 衰老表现为编码蛋白质降解, 核酸、膜脂和糖类的相关基因被激活; 同时伴有衰老根瘤中氧化还原平衡的改变, 以及抗氧化防御的整体减弱等现象(Van de Velde et al., 2006; Loscos et al., 2008).NO在植物的衰老过程中具有重要作用(Procházková and Wilhelmová, 2011).例如, NO是水稻(Oryza sativa)光依赖性叶细胞死亡的关键介质(Lin et al., 2012); 蒺藜苜蓿根瘤内NO含量的增加会导致根瘤过早衰老, 且衰老程度与NO水平相关(Cam et al., 2012; Meilhoc et al., 2013; Blanquet et al., 2015), 表明NO对于维持共生体系和避免根瘤过早衰老极为重要.此外, 施加硝酸盐可导致豌豆表现出典型的衰老特征(Escuredo et al., 1996).对蒺藜苜蓿添加硝酸盐后, 其自身也可通过还原硝酸盐促使根瘤中生成NO, 然而硝酸盐引起的根瘤衰老是否依赖于NO目前尚不清楚(Horchani et al., 2011).此外, NO也是一种有效的呼吸抑制剂, 能够抑制线粒体和细菌末端的呼吸氧化酶, 并增加活性氧(reactive oxygen species, ROS)和活性氮(reactive nitrogen species, RNS)的积累, 而NO和ROS均为共生发育所必需(Shimoda et al., 2005; Cam et al., 2012; Igamberdiev et al., 2014; Arjona et al., 2015), 因此衰老也可能是固氮时呼吸抑制的间接结果. ...
Endoreduplication mediated by the anaphase-promoting complex activator CCS52A is required for symbiotic cell differentiation in Medicago truncatula nodules 1 2003
... NO在影响相关基因的表达、调控豆科植物防御反应进而促进共生和结瘤中发挥重要作用.转录组分析表明, NO影响多种植物基因的表达, 如编码富含半胱氨酸蛋白(cysteine-rich proteins, NCRs)、核糖体蛋白、肽酶、结瘤发育细胞周期蛋白及细胞周期转换蛋白的基因(Vinardell et al., 2003; del Giudice et al., 2011; Boscari et al., 2013), 进而调控细胞脱分化、诱导组织形成及抑制植物防御反应(Hichri et al., 2016b).LjHb1是百脉根(Lotus japonicus) ns-Hbs的编码基因, 外源添加NO、缺氧和低温均可诱导其表达(Shimoda et al., 2005).在根瘤形成的早期阶段, NO的产生可以上调LjHb1的表达, 反之LjHb1的表达又下调NO的水平, 以降低植物防御反应, 促进根瘤菌和宿主植物建立共生关系(Nagata et al., 2008; Murakami et al., 2011).植物也可通过降低Hb的表达促进内源NO产生, 调控自身机体内相关防御基因的表达(Wally et al., 2013) (图2).而NO的清除会引起基因MtCRE1和MtCCS52A的下调表达, 从而延缓结瘤(del Giudice et al., 2011).通过对接种中华根瘤菌的蒺藜苜蓿外源添加NO清除剂(2-(4-carboxyphenyl)- 4,4,5,5-tetramethyl imidazoline-1-oxyl-3-oxide, cPTIO), 发现受NO调节的3种基因(Contig525、Contig3307和Medtr5g010350)表达下调(Boscari et al., 2013).其中, 前2种基因分别与ns-Hbs和茉莉酮酸代谢途径(Palmieri et al., 2008)相关, 而最后1种基因可调控还原型谷胱甘肽(glutathione, GSH)的合成(Innocenti et al., 2007).转录组分析显示, 蒺藜苜蓿接种中华根瘤菌后, 外源添加cPTIO或NADPH氧化酶抑制剂二苯烯碘铵(diphenylene iodide, DPI), 导致细胞壁形成和发育过程中相关基因下调表达, 而植物防御和二次代谢相关基因上调表达(Puppo et al., 2013).hmp的过表达和外源添加cPTIO都可降低蒺藜苜蓿的结瘤效率(Kato et al., 2010; del Giudice et al., 2011), NO则通过调节上述基因的表达, 抑制相关防御反应从而促进共生关系的建立.此外, 在根瘤菌内也鉴定出数百个依赖NO表达的基因, 如中华根瘤菌中编码Hmp的hmp基因(del Giudice et al., 2011; Cam et al., 2012)、编码haeme-Cu蛋白的nnrSI和nnrS2基因(Blanquet et al., 2015)及编码NiR的nirKV基因(Meilhoc et al., 2011), 但关于根瘤菌内受NO影响且与共生结瘤相关的基因报道很少. ...
Hemoglobin regulation of plant embryogenesis and plant pathogen interaction 1 2013
... NO在影响相关基因的表达、调控豆科植物防御反应进而促进共生和结瘤中发挥重要作用.转录组分析表明, NO影响多种植物基因的表达, 如编码富含半胱氨酸蛋白(cysteine-rich proteins, NCRs)、核糖体蛋白、肽酶、结瘤发育细胞周期蛋白及细胞周期转换蛋白的基因(Vinardell et al., 2003; del Giudice et al., 2011; Boscari et al., 2013), 进而调控细胞脱分化、诱导组织形成及抑制植物防御反应(Hichri et al., 2016b).LjHb1是百脉根(Lotus japonicus) ns-Hbs的编码基因, 外源添加NO、缺氧和低温均可诱导其表达(Shimoda et al., 2005).在根瘤形成的早期阶段, NO的产生可以上调LjHb1的表达, 反之LjHb1的表达又下调NO的水平, 以降低植物防御反应, 促进根瘤菌和宿主植物建立共生关系(Nagata et al., 2008; Murakami et al., 2011).植物也可通过降低Hb的表达促进内源NO产生, 调控自身机体内相关防御基因的表达(Wally et al., 2013) (图2).而NO的清除会引起基因MtCRE1和MtCCS52A的下调表达, 从而延缓结瘤(del Giudice et al., 2011).通过对接种中华根瘤菌的蒺藜苜蓿外源添加NO清除剂(2-(4-carboxyphenyl)- 4,4,5,5-tetramethyl imidazoline-1-oxyl-3-oxide, cPTIO), 发现受NO调节的3种基因(Contig525、Contig3307和Medtr5g010350)表达下调(Boscari et al., 2013).其中, 前2种基因分别与ns-Hbs和茉莉酮酸代谢途径(Palmieri et al., 2008)相关, 而最后1种基因可调控还原型谷胱甘肽(glutathione, GSH)的合成(Innocenti et al., 2007).转录组分析显示, 蒺藜苜蓿接种中华根瘤菌后, 外源添加cPTIO或NADPH氧化酶抑制剂二苯烯碘铵(diphenylene iodide, DPI), 导致细胞壁形成和发育过程中相关基因下调表达, 而植物防御和二次代谢相关基因上调表达(Puppo et al., 2013).hmp的过表达和外源添加cPTIO都可降低蒺藜苜蓿的结瘤效率(Kato et al., 2010; del Giudice et al., 2011), NO则通过调节上述基因的表达, 抑制相关防御反应从而促进共生关系的建立.此外, 在根瘤菌内也鉴定出数百个依赖NO表达的基因, 如中华根瘤菌中编码Hmp的hmp基因(del Giudice et al., 2011; Cam et al., 2012)、编码haeme-Cu蛋白的nnrSI和nnrS2基因(Blanquet et al., 2015)及编码NiR的nirKV基因(Meilhoc et al., 2011), 但关于根瘤菌内受NO影响且与共生结瘤相关的基因报道很少. ...
In vivo target sites of nitric oxide in photosynthetic electron transport as studied by chlorophyll ?uorescence in pea leaves 1 2008
... NO不仅能调节植物生长发育, 还可参与植物对非生物胁迫的响应(Moreau et al., 2010).目前的研究发现, 在不同非生物胁迫(如干旱和缺氧)下, 共生有机体中的NO均会产生响应(Desalvo et al., 2010; Sánchez et al., 2011; Hawkins et al., 2014; Iarullina et al., 2014; Zimmer-Prados et al., 2014).同时, 在豆科植物中NO还可能参与调控机体对金属元素镉(Cd)和铝(Al)的胁迫反应以及应对非金属元素砷(As)的胁迫(Singh et al., 2015; 尚玉婷等, 2018), 但都缺乏在结瘤和共生固氮体系中的实验证据(Pérez- Chaca et al., 2014).Meilhoc等(2011)提出, 宿主中任何应激反应下NO的产生都会直接影响共生, 在成熟根瘤中, 除了NO专用调节器NnrR外, 多数根瘤菌的基因通过双组分系统(FixLJ)来调节NO, 其中FixLJ是O2限制反应的主要调节因子.nnrR是Nor结构基因norCB的上游基因, 可防止NO这种高活性氮氧化物的积累, 其插入失活可抑制亚硝酸盐的产生以及亚硝酸盐和NO的还原(De Bruijn et al., 2006).NO还能够可逆地结合光系统II (photosystem II, PSII)并调节电子的转移和猝灭, 对光合作用至关重要(Wodala et al., 2008).从农业和进化的角度来看, NO在植物与根瘤菌共生体系中的调控作用, 可以改善植物对病原体的防御状态、保护植物免受虫害以及增强植物对非生物胁迫的适应性(Chadha et al., 2015). ...
Omics approaches toward defining the comprehensive abscisic acid signaling network in plants 1 2015
... 脱落酸(abscisic acid, ABA)是植物适应环境胁迫的关键激素(Yoshida et al., 2015), 可通过抑制根毛变形及侵染线的形成影响豆科植物结瘤(Suzuki et al., 2004; Ding et al., 2008).Tominaga等(2010)在百脉根中发现, ABA不仅可调节根瘤的固氮酶活性, 还可通过影响结瘤过程中NO的产生来影响结瘤.在根瘤形成过程中, NO通过酪氨酸(tyrosine, Tyr)硝化调控ABA信号传递, 而ABA又在信号转导过程中决定NO的产生与积累(Castillo et al., 2015).Tyr硝化是NO介导的翻译后修饰, 其特征在于将-NO2添加到Tyr残基芳环的等价邻位C上, 产生3-硝基酪氨酸(3-nitrotyrosine, 3-NT), 并由此改变蛋白质构象(Radi, 2004).作为硝化分子的前体, NO通过Tyr硝化改变细胞溶质谷氨酰胺合成酶(glutamine synthetase 1, GS1)和Lb等关键结瘤蛋白的活性(Melo et al., 2011; Navascués et al., 2012; Li et al., 2014).豆科植物结瘤时Lb发生Tyr硝化, 可保护共生体免受过氧亚硝酸盐毒害, 因此3-NT的形成既是硝基氧化应激的标志, 也是功能性结瘤活跃代谢的结果(Sainz et al., 2015).此外, NOS抑制剂N-硝基-L-精氨酸甲酯(NG-nitro- L-arginine methyl ester, L-NAME)和cPTIO处理可减弱豌豆(Pisum sativum)表皮由ABA诱导的气孔关闭(张绪成等, 2005). ...
The biology of stomatal guard cells 1 1983
... 研究表明, 长期黑暗处理能显著诱导根瘤衰老, 同时降低固氮酶活性和Hbs水平(Romanov et al., 1980; Matamoros et al., 1999; Swaraj et al., 2001; Pérez Guerra et al., 2010).持续的暗处理会干扰根瘤功能, 导致根瘤完全衰老(Pérez-Guerra et al., 2010), 而通过降解根瘤中的NO可以延缓黑暗诱导的衰老, 但关于NO在根瘤衰老过程中的作用机制还需进一步研究(Cam et al., 2012).She等(2004)发现在光/暗条件下,对蚕豆(Vicia faba)外源添加cPTIO和L-NAME可减弱H2O2对NO产生的诱导效应和改变人为光照引起的气孔关闭.气孔关闭由多种生物和非生物因素引起, 如渗透胁迫、黑暗、高浓度的CO2和机械压力, 其中光是调节气孔运动最重要的环境因素之一(Zeiger, 1983; Kearns and Assmann, 1993; Herold and Puppo, 2005; Sánchez et al., 2010).但关于黑暗条件下NO调节豆科植物共生固氮的作用机制还未见报道. ...
Nitric oxide increases tolerance responses to moderate water deficit in leaves of Phaseolus vulgaris and Vigna unguiculata bean species 1 2014
... NO不仅能调节植物生长发育, 还可参与植物对非生物胁迫的响应(Moreau et al., 2010).目前的研究发现, 在不同非生物胁迫(如干旱和缺氧)下, 共生有机体中的NO均会产生响应(Desalvo et al., 2010; Sánchez et al., 2011; Hawkins et al., 2014; Iarullina et al., 2014; Zimmer-Prados et al., 2014).同时, 在豆科植物中NO还可能参与调控机体对金属元素镉(Cd)和铝(Al)的胁迫反应以及应对非金属元素砷(As)的胁迫(Singh et al., 2015; 尚玉婷等, 2018), 但都缺乏在结瘤和共生固氮体系中的实验证据(Pérez- Chaca et al., 2014).Meilhoc等(2011)提出, 宿主中任何应激反应下NO的产生都会直接影响共生, 在成熟根瘤中, 除了NO专用调节器NnrR外, 多数根瘤菌的基因通过双组分系统(FixLJ)来调节NO, 其中FixLJ是O2限制反应的主要调节因子.nnrR是Nor结构基因norCB的上游基因, 可防止NO这种高活性氮氧化物的积累, 其插入失活可抑制亚硝酸盐的产生以及亚硝酸盐和NO的还原(De Bruijn et al., 2006).NO还能够可逆地结合光系统II (photosystem II, PSII)并调节电子的转移和猝灭, 对光合作用至关重要(Wodala et al., 2008).从农业和进化的角度来看, NO在植物与根瘤菌共生体系中的调控作用, 可以改善植物对病原体的防御状态、保护植物免受虫害以及增强植物对非生物胁迫的适应性(Chadha et al., 2015). ...
 ,1,2,*
,1,2,*

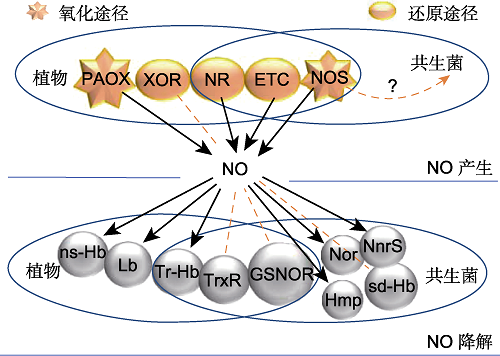 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT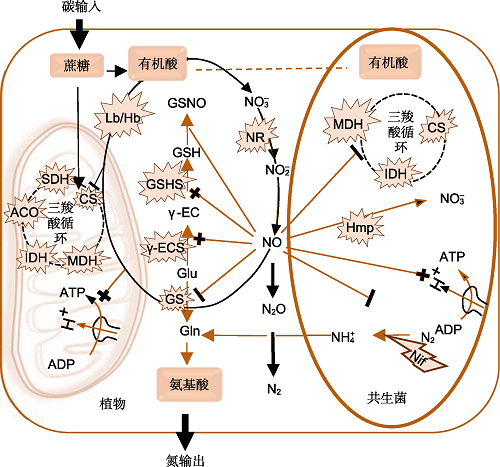 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}