 ,*中国农业科学院蔬菜花卉研究所, 北京 100081
,*中国农业科学院蔬菜花卉研究所, 北京 100081Recent Advances in Molecular Mechanisms of Plant Graft Healing Process
Lulu Xie, Qingqing Cui, Chunjuan Dong, Qingmao Shang,*Institute of Vegetables and Flowers, Chinese Academy of Agricultural Sciences, Beijing 100081, China通讯作者: E-mail:shanglab211@126.com
责任编辑: 朱亚娜
收稿日期:2020-04-9接受日期:2020-06-28网络出版日期:2020-09-01
| 基金资助: |
Corresponding authors: E-mail:shanglab211@126.com
Received:2020-04-9Accepted:2020-06-28Online:2020-09-01

摘要
嫁接能显著改良单一品种的产量、品质和抗逆等性状, 已广泛应用于农业生产。促进砧木和接穗在嫁接面的快速愈合有利于提高嫁接效率。目前对嫁接愈合调控机制尚了解不足, 因此短时间内难以进行有效的技术改良。嫁接愈合过程包括先后发生的创伤应激响应、愈伤组织形成、砧穗细胞通讯以及砧穗再生重连等生理事件, 均涉及复杂而交联的激素应答及基因调控模式。近年来, 相关领域的研究成果为综合解析嫁接愈合的调控机制奠定了基础。该文综述了在嫁接愈合过程中发挥核心作用的植物激素及其应答方式, 以及激素依赖或非依赖的基因表达调控模式, 以期为深入揭示嫁接愈合分子机制提供参考。
关键词:
Abstract
Grafting can significantly improve target traits such as yield, quality, and resistance of vegetable varieties, and is widely applied in agricultural practice. Prompting graft healing between scion and stock at the graft interface is necessary to improve graft efficiency. Currently the improvement on the technology is hampered by our lack of understanding of regulatory mechanisms of graft healing. The graft healing process involves complicated and cross-linked physiological events, including wounding stress response, callus formation, cell communication between scion and stock, and the regeneration and reunion of scion and stock. Recent research has provided a good foundation for our understanding the molecular mechanisms of graft healing. In this review, we summarize the central roles of phytohormones in each of the physiological events, and the phytohormone-dependent and -independent gene regulatory networks in graft healing, to provide a reference for further studying graft healing-related molecular mechanisms.
Keywords:
PDF (867KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
谢露露, 崔青青, 董春娟, 尚庆茂. 植物嫁接愈合分子机制研究进展. 植物学报, 2020, 55(5): 634-643 doi:10.11983/CBB20061
Xie Lulu, Cui Qingqing, Dong Chunjuan, Shang Qingmao.

嫁接是指将一个植株带有芽或枝的接穗(scion)与另一个植株带有根系的砧木(stock)通过机械固定使切割产生的嫁接面(graft interface)紧贴, 培养至砧、穗在嫁接结合部相互愈合, 从而获得嫁接体植株(grafting plant)的技术。优良的嫁接体植株因结合了接穗和砧木各自的优势而具有一种或多种显著的增益效果, 如生物或非生物胁迫抗性增强、产量提高、品质改良、开花或块茎化习性优化以及株型矮化(Goldschmidt, 2014; Wang et al., 2017)。目前, 嫁接技术已作为一种常规手段应用于果树和蔬菜种苗的繁育, 极大地促进了农业生产和农民增收(Lee et al., 2010; Huang et al., 2015)。
砧穗嫁接面能够完全愈合是嫁接成功的关键, 而愈合程度具有很大的不确定性, 受遗传背景、发育阶段和栽培条件等因素的制约(Kawaguchi et al., 2008; Johkan et al., 2009; Vu et al., 2014; 赵渊渊等, 2015)。可通过有针对性地筛选砧穗组合、合理地选择发育阶段及精准地设置环境参数等手段提高嫁接的成功率和愈合速率, 这均需要基础理论作为指导。研究初期以表型和显微结构观测为依据, 将嫁接愈合的进程描述为隔离层的形成和初始粘连(嫁接后0-1天(0-1 days after grafting, DAG))、愈伤组织和胞间连丝的形成(2-3 DAG)以及砧穗维管束的重连(4-8 DAG) (Moore, 1982, 1984; 卢善发等, 1995; 王幼群, 2011)。随后研究发现, 生理和分子水平的变化与表型变化大致契合(Goldschmidt, 2014; 苗丽等, 2017)。多种植物激素参与了砧穗愈合且作用显著(Nanda and Melnyk, 2018)。然而, 嫁接愈合过程中植物激素发挥作用的时机和方式非常复杂, 亟须归纳和梳理。本文根据嫁接愈合进程依次发生的创伤应激响应(wounding stress responses)、愈伤组织形成(callus formation)、砧穗细胞通讯(cell communication between scion and stock)和砧穗再生重连(regeneration and reunion of scion and stock)四个阶段性的生理学事件, 对已证实或推测参与其中的内源性植物激素及基因表达调控机制进行总结和论述(图1)。
图1
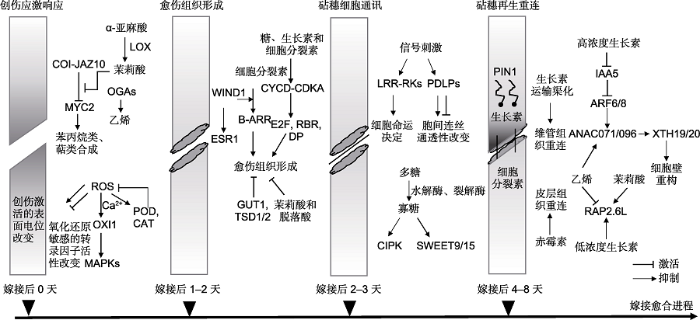 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1嫁接愈合4个生理学事件中的植物激素应答和基因表达调控
LOX: 脂肪氧化酶; ROS: 活性氧; POD: 过氧化物酶; CAT: 过氧化氢酶
Figure 1Phytohormone responses and gene expression regulations in the four physiological events during graft healing process
LOX: Lipoxygenase; ROS: Reactive oxygen species; POD: Peroxidase; CAT: Catalase
1 创伤应激响应
1.1 茉莉酸依赖的创伤应激响应
切割造成嫁接面时, 创伤应激响应是首先发生的事件。茉莉酸信号途径是其中较为重要的调控路径。在番茄(Solanum lycopersicum)中的研究证实, 创伤发生后的短时间内, 内源性植物激素茉莉酸(jasmonic acid, JA)及其活性衍生物, 如茉莉酸甲酯(methyl jasmonate, MeJA)、茉莉酸-异亮氨酸复合物(jasmonoylisoleucine, JA-Ile)以及顺式茉莉酸(cis-jasmone, CJ) 被迅速合成(Howe, 2010)。茉莉酸类激素(JAs)在创伤局部大量积累, 激活一系列防御反应, 且经维管系统运输至其它部位, 运输期间伴随着JAs在维管细胞内的二次合成(Ruan et al., 2019)。在拟南芥(Arabidopsis thaliana)中的研究表明, JAs在创伤部位的积累和扩散发生在数秒或数分钟内(Glauser et al., 2008, 2009)。此外, 还发现JAZ10 (jasmonate ZIM-domain 10)所指示的茉莉酸诱导信号响应与创伤激活的表面电位改变(wound-activated surface potential changes, WASPs)在空间上的扩散完全一致。细胞膜定位的类谷氨酸盐受体(glutamate receptor-like, GLR)蛋白参与维持WASPs, 进而影响茉莉酸信号转导(Mousavi et al., 2013)。COI1-JAZ复合体是JAs的高亲和受体(Sheard et al., 2010)。COI1 (coronatine insensitive 1)是具有F-box结构域的E3泛素化连接酶; JAZ蛋白通过JAs结构域与COI1结合, 通过ZIM结构域与下游bHLH类转录因子MYC2结合。感知JAs信号后, JAZ解除对MYC2的抑制, 后者结合在JAs诱导基因的启动子区, 激活其转录(Sheard et al., 2010)。JA/MeJA能够诱导苯丙素(phenylpropanoids)、生物碱(alkaloids)和萜类(terpenoids)等次生代谢物的合成(Howe, 2010)。
1.2 其它创伤应激响应
除茉莉酸信号途径以外, 还存在其它创伤应激响应机制。使用coi1/jaz突变体筛选出多种参与创伤响应的非JAs依赖的效应物, 包括乙烯(ethylene, ETH)、脱落酸(abscisic acid, ABA)、活性氧(reactive oxygen species, ROS)、寡聚半乳糖醛酸(oligogalacturonides, OGAs)、一氧化氮(nitric oxide, NO)和脂肪酸氨基酸复合物(fatty acid-amino acid conjugates, FACs)等(Schilmiller and Howe, 2005)。其中, OGAs可诱导ETH的合成, ETH又能够抑制JAs响应基因的表达(Rojo et al., 1999)。此外, 水杨酸(salisylic acid, SA)是JAs的拮抗激素, 外施SA能够抑制JAs的信号响应, 且具有明显的时效性, 抑制过程伴随谷氧还蛋白(gluataredoxin) GRX480的显著上调, 被认为与氧化还原反应密切相关(Koornneef et al., 2008)。活性氧(包括超氧阴离子自由基(O2.-)和过氧化氢(H2O2))在创伤后1小时内开始积累(Orozco-Cardenas and Ryan, 1999)。ROS可作为第二信使参与调控诸多生物学过程, 如细胞程序性死亡、细胞周期、生物或非生物胁迫和胚发生(Elhiti and Stasolla, 2015)。ROS通过其受体改变Ca2+流, 促进丝/苏氨酸蛋白激酶(serine/threonine protein kinase) OXI1 (oxidative-signal inducible 1)的表达, 后者是介导重要级联信号转导通路的促分裂素原活化蛋白激酶(mitogen-activated-protein kinases, MAPKs)的直接诱导因子(Mittler et al., 2004)。此外, 由ROS引起的氧化还原电位改变常与双组分信号系统(two-com- ponent signaling system, TCS)整合, 改变一些氧化还原态敏感的转录因子活性(Elhiti and Stasolla, 2015)。研究表明, 番茄嫁接若干天后, 其过氧化物酶(peroxidase, POD)和过氧化氢酶(catalase, CAT)在切口附近积累(Fernández-García et al., 2004)。甜椒(Capsicum annuum)嫁接过程中外施抗坏血酸(ascorbic acid, AA)对嫁接体形成具有促进作用(Johkan et al., 2008)。此外, 番茄嫁接与不嫁接茎段对比显示, ROS清除相关途径基因在嫁接愈合发生时表达量更高(Xie et al., 2019), 表明ROS先升高再降低的调控机制与嫁接愈合密切相关。
2 愈伤组织形成
2.1 愈伤组织形成的诱导
愈伤组织在嫁接面处积累, 填充砧穗之间的空隙, 其中的一些薄壁细胞后续分化为新的维管组织。由于植物组织的再生不需要干细胞龛(stem cell niche)来维持分化, 因而具有更灵活的脱分化(dedifferentiation)和再分化(redifferentiation)能力(Sena et al., 2009)。虽然均能通过诱导产生愈伤组织, 但创伤引起的愈伤组织从生理和分子性质上与植物离体培养所形成的愈伤组织不同。在以适当比例添加生长素(auxin)和细胞分裂素(cytokinins, CTKs)的培养基上诱导产生的愈伤组织中, 分生组织分子标识SCR和WOX5以及根中柱鞘分子标识J0121显著表达(Sugimoto et al., 2010)。然而, 这些分子标识在创伤诱导的愈伤组织中均不表达(Iwase et al., 2011)。AP2/ERF类转录因子WIND1 (wound induced dedifferentiation 1)及其同源蛋白是创伤后组织发育命运的重要调控因子, 其高丰度表达从创伤后数小时一直持续至愈合完全(Iwase et al., 2011; Melnyk et al., 2015)。此外, AP2/ERF类转录因子ESR1 (enhancer of shoot regeneration 1)能够被WIND1激活, 也在愈伤组织诱导形成中发挥作用(Iwase et al., 2017)。WIND1基因过表达株系表现出对CTKs异常敏感, 通过B类ARR (Arabidopsis response regulator)因子介导的信号途径促进细胞的脱分化(Iwase et al., 2011)。CTKs是愈伤组织形成所必需的植物激素。CTKs响应启动子元件连接绿色荧光蛋白报告基因(pTCSn::GFP)在去除根的下胚轴外植体上依然能被激活, 说明外植体形成愈伤组织的过程中存在CTKs的从头合成(Ikeuchi et al., 2017)。愈伤组织的特征是细胞进入快速分裂周期。当受到糖(sucrose)、生长素和细胞分裂素等物质的诱导时, D类细胞周期蛋白(D-type cyclin, CYCD)与A类细胞周期蛋白依赖性蛋白激酶(A-type cyclin-dependent kinase, CDKA)结合, 形成激活态CYCD-CDKA复合体(den Boer and Murray, 2000), 继而调控下游的3个蛋白E2F (E2 promoter binding factor)、RBR (retinoblastoma-related)和DP (dimerization partner), 使细胞进入S期(Inzé and Veylder, 2006)。拟南芥下胚轴中, CDKA在创口附近的表达起始于切割后4天, 可以在维管组织和中柱鞘检测到CDKA启动子活性, 而在内皮层、皮层和表皮中检测不到(Ikeuchi et al., 2017)。在番茄茎中, 愈伤组织在切口上方的维管组织附近形成, 同时在切口上方检测到一些细胞周期蛋白依赖性蛋白激酶编码基因的活跃转录(Xie et al., 2019)。
2.2 愈伤组织形成的抑制
在创伤应激响应中合成量升高的JAs和ABA等植物激素, 在愈伤组织形成中不是必需的。使用JAs或ABA的合成和信号转导突变体进行研究, 发现突变体愈伤组织形成量较野生型轻微但显著增多, 表明JAs和ABA对愈伤组织的形成具有轻微抑制作用(Ikeuchi et al., 2017)。此外, 还有细胞壁框架产生的物理抑制。细胞壁组分如纤维素(cellulose)、半纤维素(hemicellulose)和果胶(pectin)的有序积累, 是建立和维持细胞分化形态的关键步骤(Ikeuchi et al., 2013)。细胞壁形成缺陷突变通常会导致愈伤组织异常形成。例如, 参与果胶合成的GUT1 (glucuronyltransferase 1)基因突变后, 导致愈伤组织堆积于地上部顶端; 参与纤维素合成的TSD1/KOR1/RSW2基因突变后, 导致地上和地下部细胞排列混乱; 被推测为编码高尔基体定位的甲基化转移酶基因TSD2/QUA2/OSU1突变, 导致细胞间不能相互黏连的严重缺陷(Ikeuchi et al., 2013)。因此, 细胞壁组分的正常沉积有助于避免体细胞过分增殖。
3 砧穗细胞通讯
3.1 胞间连丝介导的发育决定和免疫应答
胞间连丝(plasmodesmata, PD)由外膜和内膜2层膜结构组成, 分别为质膜和内质网膜的延续, 但理化性质稍有差别(Tilsner et al., 2011)。借助于膜锚定的受体激酶(receptor kinases, RKs)和受体蛋白(receptor proteins, RPs), 胞间连丝能够对胞外环境变化做出响应, 整合质外体(apoplastic)和共质体(symplastic)途径的信号转导, 从而驱动发育决定和免疫应答, 或通透性调控等(Stahl and Faulkner, 2016)。这一机制在砧、穗细胞之间的初始粘连和通讯中扮演重要角色。在有些发育进程中, 质膜或PD上的RKs和RPs通过非细胞自主性信号途径调控组织形态建成或干细胞分化命运。例如, 通过PD特异性蛋白质组鉴定到的SUB (strubbelig)属于非典型的富含亮氨酸重复受体激酶(LRR-RK), 它通过与另一个具有C2结构域的PD锚定蛋白QKY (quirky)互作, 将信号传递至下游(Vaddepalli et al., 2014)。PD锚定蛋白ACR4 (arabidopsis crinkly 4)以同源复合体或与LRR-RK CLV1 (clavata 1)形成异源复合体的形式定位于PD, 小分子多肽可作为配体(ligand)与ACR4和CLV1的胞外结构域结合。结构上同源的配体多肽CLV3和CLE40经证实分别在茎端及根尖分生组织中促进干细胞的分化(Ohyama et al., 2009; Stahl et al., 2009)。当砧穗嫁接面紧贴时, PD及其上锚定的RKs和RPs的存在很可能为砧穗间初始物质交流提供桥梁。由于嫁接面形成的愈伤组织将面临不同的分化命运, 故PD通过不同受体对发育信号进行承接和传递是重要的因素之一。
PD上的RKs和RPs还可作为模式识别受体(pattern recognition receptors)结合病原衍生配体(pathogen-derived ligand), 如真菌细胞壁的几丁质(chitin)或细菌的鞭毛蛋白(falgellin), 可激活胞内免疫应答(Stahl and Faulkner, 2016)。例如, PD锚定蛋白FLS2 (flagellin sensing 2)和LYM2 (lysin motif domain-containing glycosylphosphatidylinositol-anchored protein 2) (Lee, 2015), 在嫁接愈合初期, 嫁接面细胞壁不健全, 病原菌入侵是大概率事件。作为共质体运输的门户, 胞间连丝能够通过外膜和细胞壁之间拐角处填充的胼胝质(callose)来调节自身通透性(permeability)。胼胝质的积累量随环境或发育状况而变化, 与PD的通透性呈负相关(Lee, 2015)。具有胞外DUF26结构域的PD定位受体蛋白(PD-located proteins, PDLPs)是控制胼胝质积累量和PD通透性的上游因子。通常认为, PDLPs接收本地ROS信号, 激活调控通路, 最终使胼胝质积累量增加及PD关闭(Stahl and Faulkner, 2016)。其中, PDLP5以SA依赖的方式调控细菌性病原菌免疫应答, SA促进PDLP5的转录, 而PDLP5的上调又促进SA的积累(Lee et al., 2011)。重要的是, PDLP蛋白与嫁接处理相关, PDLP1A在嫁接部位高表达(Yin et al., 2012)。
胞间连丝和膜锚定受体一方面需要通过感知嫁接搭档组织或自身组织中的发育信号, 达到加强砧穗交流的目的; 另一方面需要响应创伤应激, 通过免疫应答阻断细胞与外界的联系。在这种情况下, 膜受体(如LRR-RK家族成员)以网络协作的形式对各种胞外信号进行平衡(Smakowska-Luzan et al., 2018)。类似调控机制很可能存在于嫁接愈合的砧穗胞间通讯中。
3.2 寡糖信号途径与胞间通讯
寡糖(oligosaccharides)是细胞的能量来源, 同时也是一类信号分子, 在质膜上有专门的受体蛋白激酶(Chikano et al., 2001)。在嫁接面处, 质膜上的蔗糖转运蛋白SWEET (sugars will eventually be exported transporters) 9和15上调表达, 同时CIPK (calcineurin B-like-interacting protein kinase)等蛋白激酶受寡糖调节。由于同时上调的还有裂解酶(lyase)和水解酶(hydrolase)等, 因此推测由死亡细胞残留细胞壁分解而来的寡糖类物质可能构成砧穗胞间通讯的一种途径(Yin et al., 2012)。4 砧穗再生重连
砧穗组织重连是嫁接愈合完成的标志性事件, 包括维管组织和其它组织的重连等。已开发出一些模式化的切割或嫁接系统用于组织重连研究, 如拟南芥下胚轴微嫁接(micrograft)或改良后的微嫁接(Turnbull et al., 2002; Yin et al., 2012)、黄瓜(Cucumis sativus)或番茄下胚轴和拟南芥花序茎的半切(one-half cut)重连试验(Asahina et al., 2002, 2011)等。这些模式系统为探讨内源性植物激素以及基因调控网络在砧穗再生重连中的作用提供了便利。4.1 生长素和细胞分裂素在维管组织非对称性重连中的作用
生长素在植物多个发育进程中与维管组织的形成密切相关(Ca?o-Delgado et al., 2010)。根据生长素的“渠化假说(canalization hypothesis)”, 生长素极性运输和由此形成的浓度差异可能为维管束的分化和重连提供空间极性信息(Mazur et al., 2016)。叶脉形成研究揭示了若干以反馈调节形式调控叶片维管束形成的参与因子, 包括生长素运输蛋白PIN1 (PIN- FORMED 1)、生长素信号响应因子MP (MONOPTEROS)和ATHB8等(Wenzel et al., 2007; Donner et al., 2009)。生长素在拟南芥花序茎半切损伤愈合中也会产生渠化效应, 由PIN1融合报告基因标识出的生长素运输轨迹先于新维管组织的形成(Mazur et al., 2016)。由于生长素和细胞分裂素的极性运输, 及其运输流被切割所阻断, 嫁接面上下组织中的调控模式呈现非对称的特点。借助携带生长素响应元件的DR5启动子和β-葡萄糖苷酸酶(β-glucuronidase, GUS)报告基因的转基因拟南芥, 观察微嫁接后或非嫁接的GUS染色部位, 发现当转基因拟南芥作为接穗, 野生型拟南芥作为砧木时, 在接穗嫁接面维管束附近部位有明显的染色, 1 DAG时开始出现, 3 DAG时弥漫至整个嫁接面; 而当转基因拟南芥作为砧木时, 嫁接面附近在1-3 DAG期间均未观察到明显染色(Yin et al., 2012)。进一步研究发现, 维管束韧皮部和木质部的重连在时间上是分离的(Melnyk et al., 2015)。韧皮部重连发生时间是3-4 DAG, 而木质部重连发生时间是6-7 DAG。凯氏带合成基因CASP1 (casparian strip membrane domain proteins 1)、指示细胞分裂S期的组蛋白H4编码基因和创伤响应基因WIND1等均呈现出在嫁接面上部表达先于下部的模式。采用生长素响应启动子元件连接报告基因(pDR5::GFP)和细胞分裂素响应启动子元件连接报告基因(pARR5:: GFP和pTCSn::GFP)验证两类植物激素的作用, 结果表明, 对生长素的响应由接穗起始并随之扩散至嫁接结合部, 对细胞分裂素的响应激活由砧木起始并随之扩散至嫁接结合部(Melnyk et al., 2015)。通过突变体筛选得出, 生长素响应蛋白ALF4 (aberrant lateral root formation 4)作为承接生长素信号的关键因子, 其正常功能在韧皮部重连过程中是必需的(Melnyk et al., 2015)。高通量测序研究表明, 生长素应答基因网络在嫁接后的切口上、下组织, 以及砧穗分开放置的切口以上组织中被激活, 而在砧穗分开放置的切口以下组织中不被激活(Melnyk et al., 2018)。上述结果表明, 生长素和细胞分裂素(尤其是生长素)的运输以及信号响应在维管组织重连中的作用显著。
4.2 赤霉素在幼苗期皮层组织重连中的作用
在幼苗期, 赤霉素(gibberellin, GA)的生物合成影响皮层的组织重连。当切除黄瓜和番茄幼苗下胚轴直径的一半长度后, 皮层的细胞分裂起始于切割后3天, 7天后重连。去除子叶后重连被抑制。在地上部尖端施加GA能够逆转抑制作用, 而施加GA抑制剂, 以及GA缺陷突变体(gib-1)呈现与去除子叶类似的表型(Asahina et al., 2002)。此外, 去除芽的顶端生长点, 或者施用生长素极性运输抑制剂(2,3,5-triiodobenzoic acid, TIBA)并不影响重连, 而施用吲哚-3-乙酸(indole-3-acetic acid, IAA)不能弥补去除子叶的重连抑制, 研究表明顶端生长点产生的生长素对去除子叶的黄瓜和番茄幼苗组织重连的影响并不大(Asahina et al., 2002)。继而检测幼苗内源性GA, 发现具有生物活性的GA1/GA4及其前体在去除子叶的幼苗中含量降低。推测子叶维持了GA浓度, 并且GA在黄瓜和番茄幼苗组织重连中具有重要作用(Asahina et al., 2007)。将拟南芥下胚轴进行全切和嫁接, 保留子叶的下胚轴愈合过程伴随GA合成基因(GA20ox1、GA2ox和GA3ox1)的上调, 去除子叶或TIBA处理后上调表达被抑制, 且赤霉素缺陷突变体cps出现维管束正常重连但皮层不能重连的表型, 表明皮层组织重连需要GA的生物合成, 同时受到源于子叶的生长素的控制(Matsuoka et al., 2016)。对拟南芥花序茎半切处理后3天, 细胞分裂主要发生在髓部。去除花序形态学上端、抑制生长素极性运输以及使用生长素运输突变体(pin1), 均会抑制重连过程。而GA缺陷突变体(ga3ox1/ga3ox2)能够正常重连。推测在拟南芥花序茎组织重连的过程中, 生长素极性运输的作用显著而GA的作用不显著(Asahina et al., 2011)。4.3 多激素交联作用下的基因调控
转录因子在组织重连过程中作用显著, 其表达受到生长素、JAs和ETH的交联调控。生长素促进转录因子ANAC071 (NAC domain containing protein 71)和ANAC096的表达, 生长素响应因子IAA5 (indole-3- acetic acid inducible 5)、ARF6 (auxin response factor 6)和ARF8居于级联调控的中部, 推测其在嫁接面以上发挥作用(Pitaksaringkarn et al., 2014a; Matsuoka et al., 2016)。AP2/ERF类转录因子的另1个成员RAP2.6L (related to AP2.6L)、JAs和低浓度生长素可促进其表达, 推测其在嫁接面以下发挥作用(Pitaksaringkarn et al., 2014a)。此外, ETH能够促进ANAC071及抑制RAP2.6L的表达(Asahina et al., 2011)。在乙烯不敏感突变体ein2中, 细胞分裂仅发生在嫁接面的皮层, 与野生型的髓部不同。当ANAC071和RAP2.6L的表达被抑制后, 重连不能正常进行。虽然这2个转录因子的作用尚不完全明晰, 但它们均受到生长素浓度的影响, 并被创伤应激激素JAs和ETH微调(Asahina et al., 2011)。根据表达模式的相似性和功能验证, 参与细胞壁重构过程的木葡聚糖内转糖苷酶/水解酶(xyloglucan endotransglucosylase/hydrolase)编码基因(XTH19和XTH20)受到ANAC071的直接调控(Pitaksaringkarn et al., 2014b)。5 木本和草本植物嫁接愈合的比较
嫁接技术最早应用于多年生果树、林木和观赏花卉等木本植物, 20世纪初期开始大量应用于茄果类和瓜果类蔬菜等草本植物, 近年来有关嫁接生理的研究大多集中在蔬菜嫁接(Goldschmidt, 2014)。模式植物拟南芥的引入也为嫁接愈合分子机制的研究提供了更多便利(Turnbull et al., 2002)。从嫁接时间和部位上来看, 草本嫁接采用幼苗期的胚轴或幼嫩茎, 而木本嫁接通常采用越冬后即将打破休眠时期的枝干和发芽枝条(Gautier et al., 2019)。此外, 愈合期持续时间的长短具有明显差异。例如, 拟南芥和番茄等物种在7-10 DAG就能够完全愈合(Melnyk et al., 2015; Xie et al., 2019); 而葡萄(Vitis vinifera)和美洲山核桃(Carya illinoinensis)等嫁接愈合期则需要数十天(Cookson et al., 2013; Mo et al., 2018)。木本植物嫁接部位的再生能力弱是造成愈合缓慢的可能原因。活跃的维管形成层是木本和草本植物嫁接部位愈合的必要条件。这与大部分双子叶植物、基部被子植物以及裸子植物可以嫁接, 而维管束散生和缺少维管形成层的单子叶植物很难嫁接相互印证(Melnyk and Meyerowitz, 2015)。
由于培养周期长和技术原因, 木本嫁接的调控机制难以采用以草本植物为材料的类似方法进行探究。可通过转录组分析获得的嫁接特异性富集路径对草本和木本愈合过程进行比较。对于拟南芥、番茄、葡萄和美洲山核桃等不同物种, 嫁接后呈现出差异表达的基因参与相似的路径, 包括茉莉酸合成、ROS消解、类黄酮代谢、细胞壁重构、细胞周期、膜锚定受体介导的细胞通讯、生长素运输和响应及维管组织分化(Yin et al., 2012; Cookson et al., 2013; Melnyk et al., 2015; Mo et al., 2018; Xie et al., 2019)。由此推测, 木本和草本嫁接的愈合过程本质上一致, 通过草本嫁接体系研究得出的调控模式在很大程度上也适用于木本嫁接。
6 问题和展望
前人的研究在一定程度上揭示了内源性植物激素和基因表达调控在嫁接愈合中的作用机制。与嫁接愈合形态变化相对应的生理学事件具有特征性的激素应答和基因表达模式。在整个嫁接愈合进程中, 激素含量的变化、基因调控网络的转换以及组织间信号的交叠使愈合机制的解析更加复杂。目前仍存在以下问题: 首先, 物种特异性和共有性植物激素的种类和作用模式还有待进一步澄清; 其次, 模式物种中构成基因调控网络的成员和表达模式需要进一步揭示; 最后, 在嫁接应用最多的茄果类和瓜果类蔬菜中, 未能对模式物种中的研究成果及时加以利用。针对以上问题, 今后可从以下几个方面进行深入研究。(1) 选择不同类群的物种, 通过检测内源性植物激素的种类和含量, 以及施加外源或内源激素抑制剂的方式, 确定各类植物激素的作用; (2) 在目前已被揭示的基因调控网络的基础上, 利用组学分析推测嫁接愈合过程中各层面的调控机制; (3) 选择嫁接应用价值大的作物, 如茄果类或瓜果类蔬菜, 对模式物种中的分子机制进行验证或扩展, 并与生产实践相结合。期待在不久的将来, 研究者对嫁接愈合机制的深入理解能够为精准调控嫁接愈合提供依据, 从而充分发挥嫁接技术的增益潜力。
(责任编辑: 朱亚娜)
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1073/pnas.1110443108URLPMID:21911380 [本文引用: 4]

When grafting or wounding disconnects stem tissues, new tissues are generated to restore the lost connection. In this study, the molecular mechanism of such healing was elucidated in injured stems of Arabidopsis. Soon after the inflorescence stems were incised, the pith cells started to divide. This process was strongly inhibited by the elimination of cauline leaves, shoot apices, or lateral buds that reduced the indole-3-acetic acid supply. Microarray and quantitative RT-PCR analyses revealed that genes related to cell division, phytohormones, and transcription factors were expressed because of incision. Among them, two plant-specific transcription factor genes, ANAC071 and RAP2.6L, were abundantly expressed. ANAC071 was expressed at 1-3 d after cutting exclusively in the upper region of the cut gap, with concomitant accumulation of indole-3-acetic acid. In contrast, RAP2.6L was expressed at 1 d after cutting exclusively in the lower region, with concomitant deprivation of indole-3-acetic acid. The expression of ANAC071 and RAP2.6L were also promoted by ethylene and jasmonic acid, respectively. In transformants suppressing the function of RAP2.6L or ANAC071, the division of pith cells was inhibited. Furthermore, the ethylene signaling-defective ein2 mutant showed incomplete healing. Hence, plant-specific transcription factors differentially expressed around the cut position were essential for tissue reunion of Arabidopsis wounded flowering stems and were under opposite control by polar-transported auxin, with modification by the ethylene and jasmonic acid wound-inducible hormones.
DOI:10.1104/pp.010886URLPMID:12011351 [本文引用: 3]
Cucumber (Cucumis sativus) hypocotyls were cut to one-half of their diameter transversely, and morphological and histochemical analyses of the process of tissue reunion in the cortex were performed. Cell division in the cortex commenced 3 d after cutting, and the cortex was nearly fully united within 7 d. 4',6-Diamidino-2-phenylindole staining and 5-bromo-2'-deoxyuridine labeling experiments indicate that nDNA synthesis occurred during this process. In addition, specific accumulation of pectic substances was observed in the cell wall of attached cells in the reunion region of the cortex. Cell division during tissue reunion was strongly inhibited when the cotyledon was removed. This inhibition was reversed by applying gibberellin (GA, 10(-4) M GA3) to the apical tip of the cotyledon-less plant. Supporting this observation, cell division in the cortex was inhibited by treatment of the cotyledon with 10(-4) M uniconazole-P (an inhibitor of GA biosynthesis), and this inhibition was also reversed by simultaneous application of GA. In contrast to the essential role of cotyledon, normal tissue reunion in cut hypocotyls was still observed when the shoot apex was removed. The requirement of GA for tissue reunion in cut hypocotyls was also evident in the GA-deficient gib-1 mutant of tomato (Lycopersicon esculentum). Our results suggest that GA, possibly produced in cotyledons, is essential for cell division in reuniting cortex of cut hypocotyls.
DOI:10.5511/plantbiotechnology.24.99URL [本文引用: 1]
DOI:10.1146/annurev-cellbio-100109-104107URLPMID:20590454 [本文引用: 1]
Plant vascular tissues, the conduits of water, nutrients, and small molecules, play important roles in plant growth and development. Vascular tissues have allowed plants to successfully adapt to various environmental conditions since they evolved 450 Mya. The majority of plant biomass, an important source of renewable energy, comes from the xylem of the vascular tissues. Efforts have been made to identify the underlying mechanisms of cell specification and patterning of plant vascular tissues and their proliferation. The formation of the plant vascular system is a complex process that integrates signaling and gene regulation at transcriptional and posttranscriptional levels. Recently, a wealth of molecular genetic studies and the advent of cell biology and genomic tools have enabled important progress toward understanding its underlying mechanisms. Here, we provide a comprehensive review of the cell and developmental processes of plant vascular tissue and resources recently available for studying them that will enable the discovery of new ways to develop sustainable energy using plant biomass.
DOI:10.1007/s004380000354URLPMID:11212922 [本文引用: 1]
We searched for genes encoding members of the group-3 SNF1-related protein kinase (SnRK3) family in the Arabidopsis thaliana database, and seven independent sequences were identified. Transcripts of two of them were found to accumulate differentially upon treatment with light, cytokinins and sugars. Full-length cDNAs were isolated and designated as AtSR1 and AtSR2; they encode polypeptides of 442 and 429 amino acids with relative molecular masses of 50.3 kDa and 48.2 kDa, respectively. In etiolated seedlings, no transcripts of either gene were observed. However, upon exposure to light or cytokinins, transcripts of AtSR1 but not AtSR2 began to accumulate. The induction with light was greatly reduced in the presence of a cytokinin antagonist, suggesting that cytokinins are involved in light-signaling pathways. In contrast, transcription of AtSR2, but not of AtSR1, was greatly increased by sucrose, as well as glucose and fructose. AtSR2 expressed in E. coli efficiently phosphorylated sucrose synthase in the presence of manganese ions. These results suggest that, although SnRK3 proteins may generally be involved in sugar metabolism, expression of AtSR1 and AtSR2 is differentially and distinctly regulated by various external signals, and AtSR2 may function in the regulation of sucrose synthase by specific phosphorylation.
DOI:10.1093/jxb/ert144URLPMID:23698628 [本文引用: 2]
Grafting is particularly important to the cultivation of perennial crops such as grapevine (Vitis vinifera) because rootstocks can provide resistance to soil-borne pests and diseases as well as improve tolerance to some abiotic stresses. Successful grafting is a complex biochemical and structural process beginning with the adhesion of the two grafted partners, followed by callus formation and the establishment of a functional vascular system. At the molecular level, the sequence of events underlying graft union formation remains largely uncharacterized. The present study investigates the transcriptome of grapevine rootstock and graft interface tissues sampled 3 d and 28 d after grafting of over-wintering stems in the spring. Many genes were differentially expressed over time, from 3 d to 28 d after grafting, which could be related to the activation of stem growth and metabolic activity in the spring. This hypothesis is supported by the up-regulation of many genes associated with cell wall synthesis, and phloem and xylem development. Generally, there was an up-regulation of gene expression in the graft interface tissue compared with the rootstock, particularly genes involved in cell wall synthesis, secondary metabolism, and signalling. Although there was overlap between the genes differentially expressed over time (from 3 d to 28 d after grafting) with the gene differentially expressed between the rootstock and the graft interface, numerous graft interface-specific genes were identified.
DOI:10.1016/s0962-8924(00)01765-7URLPMID:10802540 [本文引用: 1]
In essence, the mitotic cell cycle in eukaryotes involves the duplication and separation of chromosomes, coupled to the process of dividing one cell into two. Cytokinesis is therefore the culmination of a series of events that were triggered during G1 phase, and brings the daughter cells back to the starting position in G1 for another possible round of division. In all eukaryotes, progression through the cell cycle is controlled by cyclin-dependent kinases that bind to positive regulators called cyclins. This review explores some of the pathways that trigger the plant cell cycle, with emphasis on the G1 phase. Examples include signalling pathways involving glutathione and cellular redox potential, the possible existence of a G1 DNA-damage checkpoint, and the plant hormones auxin and cytokinin. Progress in understanding the link between cell proliferation, cell differentiation and the cell-cycle machinery in a developmental context is discussed.
DOI:10.1242/dev.037028URLPMID:19710171 [本文引用: 1]
The principles underlying the formation of veins in the leaf have long intrigued developmental biologists. In Arabidopsis leaves, files of anatomically inconspicuous subepidermal cells that will elongate into vein-forming procambial cells selectively activate ATHB8 gene expression. The biological role of ATHB8 in vein formation and the molecular events that culminate in acquisition of the ATHB8 preprocambial cell state are unknown, but intertwined pathways of auxin transport and signal transduction have been implicated in defining paths of vascular strand differentiation. Here we show that ATHB8 is required to stabilize preprocambial cell specification against auxin transport perturbations, to restrict preprocambial cell state acquisition to narrow fields and to coordinate procambium formation within and between veins. We further show that ATHB8 expression at preprocambial stages is directly and positively controlled by the auxin-response transcription factor MONOPTEROS (MP) through an auxin-response element in the ATHB8 promoter. We finally show that the consequences of loss of ATHB8 function for vein formation are masked by MP activity. Our observations define, at the molecular level, patterning inputs of auxin signaling in vein formation.
[本文引用: 2]
DOI:10.1093/aob/mch014URLPMID:14630693 [本文引用: 1]
BACKGROUND AND AIMS: The use of grafted plants in vegetable crop production is now being expanded greatly. However, few data are available on the formation of graft unions in vegetables. In this work, the structural development of the graft union formation in tomato plants is studied, together with the possible relationship with activities of peroxidases and catalases. METHODS: Tomato (Lycopersicon esculentum Mill.) seedlings of cultivar Fanny were grafted on the rootstock of cultivar AR-9704 using the 'tongue approach grafting' method, and were grown in a crop chamber. A study of the structural development of the graft union and the involvement of peroxidases and catalases in the process of graft formation was carried out during the first stages of the graft union (4, 8 and 15 d after grafting). KEY RESULTS: Observation of the structure of the graft union showed formation of xylem and phloem vessels through the graft union 8 d after grafting. In addition, root hydraulic conductance, L0, indicate that the graft union is fully functional 8 d after grafting, which coincided with an increase of peroxidase and catalase activities. CONCLUSIONS: These results suggest that increased peroxidase and catalase activities might be implicated in graft development in tomato plants.
DOI:10.1093/jxb/ery422URLPMID:30481315 [本文引用: 1]
Grafting has been utilised for at least the past 7000 years. Historically, grafting has been developed by growers without particular interest beyond the agronomical and ornamental effects, and thus knowledge about grafting has remained largely empirical. Much of the commercial production of fruit, and increasingly vegetables, relies upon grafting with rootstocks to provide resistance to soil-borne pathogens and abiotic stresses as well as to influence scion growth and performance. Although there is considerable agronomic knowledge about the use and selection of rootstocks for many species, we know little of the molecular mechanisms underlying rootstock adaptation to different soil environments and rootstock-conferred modifications of scion phenotypes. Furthermore, the processes involved in the formation of the graft union and graft compatibility are poorly understood despite over a hundred years of scientific study. In this paper, we provide an overview of what is known about grafting and the mechanisms underlying rootstock-scion interactions. We highlight recent studies that have advanced our understanding of graft union formation and outline subjects that require further development.
DOI:10.1074/jbc.M109.061432URLPMID:19846562 [本文引用: 1]
The wound response prohormone jasmonic acid (JA) accumulates rapidly in tissues both proximal and distal to injury sites in plants. Using quantitative liquid chromatography-mass spectrometry after flash freezing of tissues, we found that JA accumulated within 30 s of injury in wounded Arabidopsis leaves (p = 3.5 e(-7)). JA augmentation distal to wounds was strongest in unwounded leaves with direct vascular connections to wounded leaves wherein JA levels increased significantly within 120 s of wounding (p = 0.00027). This gave conservative and statistically robust temporal boundaries for the average velocity of the long distance signal leading to distal JA accumulation in unwounded leaves of 3.4-4.5 cm min(-1). Like JA, transcripts of the JA synthesis gene LIPOXYGENASE2 (LOX2) and the jasmonate response gene JAZ10.3 also accumulated to higher levels in directly interconnected leaves than in indirectly connected leaves. JA accumulation in a lox2-1 mutant plant was initiated rapidly after wounding then slowed progressively compared with the wild type (WT). Despite this, JAZ10.3 expression in the two genotypes was similar. Free cyclopentenone jasmonate levels were similar in both resting WT and lox2-1. In contrast, bound cyclopentenone jasmonates (arabidopsides) were far lower in lox2-1 than in the WT. The major roles of LOX2 are to generate arabidopsides and the large levels of JA that accumulate proximal to the wound. LOX2 is not essential for some of the most rapid events elicited by wounding.
DOI:10.1074/jbc.M801760200URLPMID:18400744 [本文引用: 1]
A new metabolite profiling approach combined with an ultrarapid sample preparation procedure was used to study the temporal and spatial dynamics of the wound-induced accumulation of jasmonic acid (JA) and its oxygenated derivatives in Arabidopsis thaliana. In addition to well known jasmonates, including hydroxyjasmonates (HOJAs), jasmonoyl-isoleucine (JA-Ile), and its 12-hydroxy derivative (12-HOJA-Ile), a new wound-induced dicarboxyjasmonate, 12-carboxyjasmonoyl-l-isoleucine (12-HOOCJA-Ile) was discovered. HOJAs and 12-HOOCJA-Ile were enriched in the midveins of wounded leaves, strongly differentiating them from the other jasmonate metabolites studied. The polarity of these oxylipins at physiological pH correlated with their appearance in midveins. When the time points of accumulation of different jasmonates were determined, JA levels were found to increase within 2-5 min of wounding. Remarkably, these changes occurred throughout the plant and were not restricted to wounded leaves. The speed of the stimulus leading to JA accumulation in leaves distal to a wound is at least 3 cm/min. The data give new insights into the spatial and temporal accumulation of jasmonates and have implications in the understanding of long-distance wound signaling in plants.
DOI:10.3389/fpls.2014.00727URLPMID:25566298 [本文引用: 3]
Grafting, an old plant propagation practice, is still widely used with fruit trees and in recent decades also with vegetables. Taxonomic proximity is a general prerequisite for successful graft-take and long-term survival of the grafted, composite plant. However, the mechanisms underlying interspecific graft incompatibility are as yet insufficiently understood. Hormonal signals, auxin in particular, are believed to play an important role in the wound healing and vascular regeneration within the graft union zone. Incomplete and convoluted vascular connections impede the vital upward and downward whole plant transfer routes. Long-distance protein, mRNA and small RNA graft-transmissible signals currently emerge as novel mechanisms which regulate nutritional and developmental root/top relations and may play a pivotal role in grafting physiology. Grafting also has significant pathogenic projections. On one hand, stock to scion mechanical contact enables the spread of diseases, even without a complete graft union. But, on the other hand, grafting onto resistant rootstocks serves as a principal tool in the management of fruit tree plagues and vegetable soil-borne diseases. The 'graft hybrid' historic controversy has not yet been resolved. Recent evidence suggests that epigenetic modification of DNA-methylation patterns may account for certain graft-transformation phenomena. Root grafting is a wide spread natural phenomenon; both intraspecific and interspecific root grafts have been recorded. Root grafts have an evolutionary role in the survival of storm-hit forest stands as well as in the spread of devastating diseases. A more fundamental evolutionary role is hinted by recent findings that demonstrate plastid and nuclear genome transfer between distinct Nicotiana species in the graft union zone, within a tissue culture system. This has led to the formation of alloploid cells that, under laboratory conditions, gave rise to a novel, alloploid Nicotiana species, indicating that natural grafts may play a role in plant speciation, under certain circumstances.
[本文引用: 2]
[本文引用: 1]
DOI:10.1104/pp.17.01035URLPMID:28904073 [本文引用: 3]
Wounding is a primary trigger of organ regeneration, but how wound stress reactivates cell proliferation and promotes cellular reprogramming remains elusive. In this study, we combined transcriptome analysis with quantitative hormonal analysis to investigate how wounding induces callus formation in Arabidopsis (Arabidopsis thaliana). Our time course RNA-seq analysis revealed that wounding induces dynamic transcriptional changes, starting from rapid stress responses followed by the activation of metabolic processes and protein synthesis and subsequent activation of cell cycle regulators. Gene ontology analyses further uncovered that wounding modifies the expression of hormone biosynthesis and response genes, and quantitative analysis of endogenous plant hormones revealed accumulation of cytokinin prior to callus formation. Mutants defective in cytokinin synthesis and signaling display reduced efficiency in callus formation, indicating that de novo synthesis of cytokinin is critical for wound-induced callus formation. We further demonstrate that type-B ARABIDOPSIS RESPONSE REGULATOR-mediated cytokinin signaling regulates the expression of CYCLIN D3;1 (CYCD3;1) and that mutations in CYCD3;1 and its homologs CYCD3;2 and 3 cause defects in callus formation. In addition to these hormone-mediated changes, our transcriptome data uncovered that wounding activates multiple developmental regulators, and we found novel roles of ETHYLENE RESPONSE FACTOR 115 and PLETHORA3 (PLT3), PLT5, and PLT7 in callus generation. All together, these results provide novel mechanistic insights into how wounding reactivates cell proliferation during callus formation.
DOI:10.1105/tpc.113.116053URLPMID:24076977 [本文引用: 2]
Plants develop unorganized cell masses like callus and tumors in response to various biotic and abiotic stimuli. Since the historical discovery that the combination of two growth-promoting hormones, auxin and cytokinin, induces callus from plant explants in vitro, this experimental system has been used extensively in both basic research and horticultural applications. The molecular basis of callus formation has long been obscure, but we are finally beginning to understand how unscheduled cell proliferation is suppressed during normal plant development and how genetic and environmental cues override these repressions to induce callus formation. In this review, we will first provide a brief overview of callus development in nature and in vitro and then describe our current knowledge of genetic and epigenetic mechanisms underlying callus formation.
DOI:10.1146/annurev.genet.40.110405.090431URLPMID:17094738 [本文引用: 1]
Cell cycle regulation is of pivotal importance for plant growth and development. Although plant cell division shares basic mechanisms with all eukaryotes, plants have evolved novel molecules orchestrating the cell cycle. Some regulatory proteins, such as cyclins and inhibitors of cyclin-dependent kinases, are particularly numerous in plants, possibly reflecting the remarkable ability of plants to modulate their postembryonic development. Many plant cells also can continue DNA replication in the absence of mitosis, a process known as endoreduplication, causing polyploidy. Here, we review the molecular mechanisms that regulate cell division and endoreduplication and we discuss our understanding, albeit very limited, on how the cell cycle is integrated with plant development.
URLPMID:28011694 [本文引用: 1]
DOI:10.1016/j.cub.2011.02.020URLPMID:21396822 [本文引用: 3]
Many multicellular organisms have remarkable capability to regenerate new organs after wounding. As a first step of organ regeneration, adult somatic cells often dedifferentiate to reacquire cell proliferation potential, but mechanisms underlying this process remain unknown in plants. Here we show that an AP2/ERF transcription factor, WOUND INDUCED DEDIFFERENTIATION 1 (WIND1), is involved in the control of cell dedifferentiation in Arabidopsis. WIND1 is rapidly induced at the wound site, and it promotes cell dedifferentiation and subsequent cell proliferation to form a mass of pluripotent cells termed callus. We further demonstrate that ectopic overexpression of WIND1 is sufficient to establish and maintain the dedifferentiated status of somatic cells without exogenous auxin and cytokinin, two plant hormones that are normally required for cell dedifferentiation. In vivo imaging of a synthetic cytokinin reporter reveals that wounding upregulates the B-type ARABIDOPSIS RESPONSE REGULATOR (ARR)-mediated cytokinin response and that WIND1 acts via the ARR-dependent signaling pathway to promote cell dedifferentiation. This study provides novel molecular insights into how plants control cell dedifferentiation in response to wounding.
DOI:10.1016/j.scienta.2008.07.015URL [本文引用: 1]
DOI:10.1016/j.scienta.2008.02.004URL [本文引用: 1]
DOI:10.1080/14620316.2008.11512427URL [本文引用: 1]
DOI:10.1104/pp.108.121392URLPMID:18539774 [本文引用: 1]
Cross talk between salicylic acid (SA) and jasmonic acid (JA) signaling pathways plays an important role in the regulation and fine tuning of induced defenses that are activated upon pathogen or insect attack. Pharmacological experiments revealed that transcription of JA-responsive marker genes, such as PDF1.2 and VSP2, is highly sensitive to suppression by SA. This antagonistic effect of SA on JA signaling was also observed when the JA pathway was biologically activated by necrotrophic pathogens or insect herbivores, and when the SA pathway was triggered by a biotrophic pathogen. Furthermore, all 18 Arabidopsis (Arabidopsis thaliana) accessions tested displayed SA-mediated suppression of JA-responsive gene expression, highlighting the potential significance of this phenomenon in induced plant defenses in nature. During plant-attacker interactions, the kinetics of SA and JA signaling are highly dynamic. Mimicking this dynamic response by applying SA and methyl jasmonate (MeJA) at different concentrations and time intervals revealed that PDF1.2 transcription is readily suppressed when the SA response was activated at or after the onset of the JA response, and that this SA-JA antagonism is long lasting. However, when SA was applied more than 30 h prior to the onset of the JA response, the suppressive effect of SA was completely absent. The window of opportunity of SA to suppress MeJA-induced PDF1.2 transcription coincided with a transient increase in glutathione levels. The glutathione biosynthesis inhibitor l-buthionine-sulfoximine strongly reduced PDF1.2 suppression by SA, suggesting that SA-mediated redox modulation plays an important role in the SA-mediated attenuation of the JA signaling pathway.
DOI:10.1016/j.scienta.2010.08.003URL [本文引用: 1]
DOI:10.1105/tpc.111.087742URLPMID:21934146 [本文引用: 1]
Plasmodesmata (PD) are thought to play a fundamental role in almost every aspect of plant life, including normal growth, physiology, and developmental responses. However, how specific signaling pathways integrate PD-mediated cell-to-cell communication is not well understood. Here, we present experimental evidence showing that the Arabidopsis thaliana plasmodesmata-located protein 5 (PDLP5; also known as HOPW1-1-INDUCED GENE1) mediates crosstalk between PD regulation and salicylic acid-dependent defense responses. PDLP5 was found to localize at the central region of PD channels and associate with PD pit fields, acting as an inhibitor to PD trafficking, potentially through its capacity to modulate PD callose deposition. As a regulator of PD, PDLP5 was also essential for conferring enhanced innate immunity against bacterial pathogens in a salicylic acid-dependent manner. Based on these findings, a model is proposed illustrating that the regulation of PD closure mediated by PDLP5 constitutes a crucial part of coordinated control of cell-to-cell communication and defense signaling.
DOI:10.1016/j.pbi.2015.06.019URLPMID:26247123 [本文引用: 2]
Effective intercellular communication is crucial for the survival of plants. Because plant cells are encased in rigid cell walls, direct cell-to-cell exchange of cytoplasmic content is only possible through plasmodesmata (PD), membrane-lined nanotubes that connect the cytoplasm of adjacent cells. PD are highly dynamic communication channels that can undergo various structural and functional modifications. Recent findings in the field suggest that defense signaling pathways are tightly linked to the regulation of PD, and the restriction of PD-mediated cell-to-cell communication is an essential innate immune response to microbial pathogens. Moreover, several plasma membrane-bound signaling components, including receptor-like kinases that are known to have non-cell autonomous function or pathogen perception at the cell periphery, are found to also partition to PD. These findings hint at the novel role of PD as a signaling hub for both symplasmic and cross-membrane pathways.
DOI:10.1093/pcp/pcw177URLPMID:27986917 [本文引用: 2]
When wounding or grafting interrupts the original connection of plant tissue, cell proliferation is induced and the divided tissue is reunited. Previous studies suggested that gibberellin derived from the cotyledon is required for tissue reunion in cucumber and tomato incised hypocotyls, and tissue reunion of Arabidopsis incised flowering stems is controlled by auxin. Differences in the hormone requirements of the tissue reunion process between Arabidopsis and cucumber might be due to differences in organs or species. In this study, we performed morphological and gene expression analyses of graft union in Arabidopsis hypocotyl. We found that removal of the cotyledon and treatment of the cotyledon with the auxin transport inhibitor triiodobenzoic acid (TIBA) suppressed cell proliferation of vascular tissue during graft union formation. These treatments also suppressed expression of IAA5, ANAC071, ANAC096 and CYCB1;1. ANAC071 is involved in the tissue reunion process. The anac071 anac096 double mutant suppressed cell proliferation more so than either of the single mutants. On the other hand, paclobutrazol treatment or deficiency of gibberellin biosynthesis genes suppressed expansion of cortex cells, and exogenous gibberellin treatment or rga/gai mutations that lack the negative regulator of gibberellin reversed this inhibition. The up-regulation of the key gibberellin biosynthesis gene GA20ox1 during graft union formation was prevented by cotyledon removal or TIBA treatment. These data suggest that auxin regulates cell proliferation of vascular tissue and expansion of cortex cells by promoting gibberellin biosynthesis during graft attachment. We hypothesize that the cotyledon-derived phytohormones are essential for graft reunion of the hypocotyl, processed in a cell type-specific manner, in Arabidopsis.
DOI:10.1038/srep33754URLPMID:27649687 [本文引用: 2]
Synchronized tissue polarization during regeneration or de novo vascular tissue formation is a plant-specific example of intercellular communication and coordinated development. According to the canalization hypothesis, the plant hormone auxin serves as polarizing signal that mediates directional channel formation underlying the spatio-temporal vasculature patterning. A necessary part of canalization is a positive feedback between auxin signaling and polarity of the intercellular auxin flow. The cellular and molecular mechanisms of this process are still poorly understood, not the least, because of a lack of a suitable model system. We show that the main genetic model plant, Arabidopsis (Arabidopsis thaliana) can be used to study the canalization during vascular cambium regeneration and new vasculature formation. We monitored localized auxin responses, directional auxin-transport channels formation, and establishment of new vascular cambium polarity during regenerative processes after stem wounding. The increased auxin response above and around the wound preceded the formation of PIN1 auxin transporter-marked channels from the primarily homogenous tissue and the transient, gradual changes in PIN1 localization preceded the polarity of newly formed vascular tissue. Thus, Arabidopsis is a useful model for studies of coordinated tissue polarization and vasculature formation after wounding allowing for genetic and mechanistic dissection of the canalization hypothesis.
DOI:10.1073/pnas.1718263115URLPMID:29440499 [本文引用: 1]
The ability for cut tissues to join and form a chimeric organism is a remarkable property of many plants; however, grafting is poorly characterized at the molecular level. To better understand this process, we monitored genome-wide gene expression changes in grafted Arabidopsis thaliana hypocotyls. We observed a sequential activation of genes associated with cambium, phloem, and xylem formation. Tissues above and below the graft rapidly developed an asymmetry such that many genes were more highly expressed on one side than on the other. This asymmetry correlated with sugar-responsive genes, and we observed an accumulation of starch above the graft junction. This accumulation decreased along with asymmetry once the sugar-transporting vascular tissues reconnected. Despite the initial starvation response below the graft, many genes associated with vascular formation were rapidly activated in grafted tissues but not in cut and separated tissues, indicating that a recognition mechanism was activated independently of functional vascular connections. Auxin, which is transported cell to cell, had a rapidly elevated response that was symmetric, suggesting that auxin was perceived by the root within hours of tissue attachment to activate the vascular regeneration process. A subset of genes was expressed only in grafted tissues, indicating that wound healing proceeded via different mechanisms depending on the presence or absence of adjoining tissues. Such a recognition process could have broader relevance for tissue regeneration, intertissue communication, and tissue fusion events.
DOI:10.1016/j.cub.2015.01.029URLPMID:25734263 [本文引用: 7]
Since ancient times, people have cut and joined together plants of different varieties or species so they would grow as a single plant - a process known as grafting (Figures 1 and 2). References to grafting appear in the Bible, ancient Greek and ancient Chinese texts, indicating that grafting was practised in Europe, the Middle East and Asia by at least the 5(th) century BCE. It is unknown where or how grafting was first discovered, but it is likely that natural grafting, the process by which two plants touch and fuse limbs or roots in the absence of human interference (Figure 3), influenced people's thinking. Such natural grafts are generally uncommon, but are seen in certain species, including English ivy. Parasitic plants, such as mistletoe, that grow and feed on often unrelated species may have also contributed to the development of grafting as a technique, as people would have observed mistletoe growing on trees such as apples or poplars.
DOI:10.1016/j.cub.2015.03.032URLPMID:25891401
Plant grafting is a biologically important phenomenon involving the physical joining of two plants to generate a chimeric organism. It is widely practiced in horticulture and used in science to study the long-distance movement of molecules. Despite its widespread use, the mechanism of graft formation and vascular reconnection is not well understood. Here, we study the dynamics and mechanisms of vascular regeneration in Arabidopsis thaliana during graft formation when the vascular strands are severed and reconnected. We demonstrate a temporal separation between tissue attachment, phloem connection, root growth, and xylem connection. By analyzing cell division patterns and hormone responses at the graft junction, we found that tissues initially show an asymmetry in cell division, cell differentiation, and gene expression and, through contact with the opposing tissue, lose this asymmetry and reform the vascular connection. In addition, we identified genes involved in vascular reconnection at the graft junction and demonstrate that these auxin response genes are required below the graft junction. We propose an inter-tissue communication process that occurs at the graft junction and promotes vascular connection by tissue-specific auxin responses involving ABERRANT LATERAL ROOT FORMATION 4 (ALF4). Our study has implications for phenomena where forming vascular connections are important including graft formation, parasitic plant infection, and wound healing.
DOI:10.1016/j.tplants.2004.08.009URLPMID:15465684 [本文引用: 1]
DOI:10.3390/genes9020071URL [本文引用: 2]
DOI:10.1093/jxb/33.3.533URL [本文引用: 1]
DOI:10.1007/BF00270192URLPMID:24253508 [本文引用: 1]
Three phases of cohesion were observable during the development of compatible autografts in Solanum pennellii. Phase I cohesion 1) lasted 4-5 d after grafting, 2) was characterized by an average increase in tensile strength of 4 g breaking weight (BW) mm(-2) graft area (GA) d(-1), and 3) correlated positively with cellular interdigitation at the graft interface. The fresh weight of the scion increased by approximately 5% d(-1) during the first 2 d after grafting. Phase II cohesion occurred 5-15 d after grafting, during which time 1) the tensile strength of the graft union increased by 14 g BW mm(-2) GA d(-1), 2) vascular differentiation across the graft interface was completed, and 3) the fresh weight of the scion increased by 9% d(-1). Phase III cohesion occurred subsequent to 15 d after grafting, during which time 1) the tensile strength of the graft union leveled off at a value similar to that of an ungrafted internode, and 2) the fresh weight of the scion increased by 14% d(-1). These results are discussed relative to mechanisms underlying the formation of compatible grafts.
DOI:10.1038/nature12478URLPMID:23969459 [本文引用: 1]
Wounded leaves communicate their damage status to one another through a poorly understood process of long-distance signalling. This stimulates the distal production of jasmonates, potent regulators of defence responses. Using non-invasive electrodes we mapped surface potential changes in Arabidopsis thaliana after wounding leaf eight and found that membrane depolarizations correlated with jasmonate signalling domains in undamaged leaves. Furthermore, current injection elicited jasmonoyl-isoleucine accumulation, resulting in a transcriptome enriched in RNAs encoding key jasmonate signalling regulators. From among 34 screened membrane protein mutant lines, mutations in several clade 3 GLUTAMATE RECEPTOR-LIKE genes (GLRs 3.2, 3.3 and 3.6) attenuated wound-induced surface potential changes. Jasmonate-response gene expression in leaves distal to wounds was reduced in a glr3.3 glr3.6 double mutant. This work provides a genetic basis for investigating mechanisms of long-distance wound signalling in plants and indicates that plant genes related to those important for synaptic activity in animals function in organ-to-organ wound signalling.
DOI:10.1007/s10265-017-0994-5URLPMID:29181647 [本文引用: 1]
For millennia, people have cut and joined different plant tissues together through a process known as grafting. By creating a chimeric organism, desirable properties from two plants combine to enhance disease resistance, abiotic stress tolerance, vigour or facilitate the asexual propagation of plants. In addition, grafting has been extremely informative in science for studying and identifying the long-distance movement of molecules. Despite its increasing use in horticulture and science, how plants undertake the process of grafting remains elusive. Here, we discuss specifically the role of eight major plant hormones during the wound healing and vascular formation process, two phenomena involved in grafting. We furthermore present the roles of these hormones during graft formation and highlight knowledge gaps and future areas of interest for the field of grafting biology.
DOI:10.1038/nchembio.182URLPMID:19525968 [本文引用: 1]
The secreted peptide gene CLAVATA3 (CLV3) regulates stem cell fate in the shoot apical meristem in Arabidopsis thaliana plants, but the molecular structure of the active mature CLV3 peptide is controversial. Here, using nano-LC-MS/MS analysis of apoplastic peptides of A. thaliana plants overexpressing CLV3, we show that CLV3 is a 13-amino-acid arabinosylated glycopeptide. Post-translational arabinosylation of CLV3 is critical for its biological activity and high-affinity binding to its receptor CLV1.
DOI:10.1073/pnas.96.11.6553URLPMID:10339626 [本文引用: 1]
Hydrogen peroxide (H2O2) generated in response to wounding can be detected at wound sites and in distal leaf veins within 1 hr after wounding. The response is systemic and maximizes at about 4-6 hr in both wounded and unwounded leaves, and then declines. The timing of the response corresponds with an increase in wound-inducible polygalacturonase (PG) mRNA and enzyme activity previously reported, suggesting that oligogalacturonic acid (OGA) fragments produced by PG are triggering the H2O2 response. Systemin, OGA, chitosan, and methyl jasmonate (MJ) all induce the accumulation of H2O2 in leaves. Tomato plants transformed with an antisense prosystemin gene produce neither PG activity or H2O2 in leaves in response to wounding, implicating systemin as a primary wound signal. The antisense plants do produce both PG activity and H2O2 when supplied with systemin, OGA, chitosan, or MJ. A mutant tomato line compromised in the octadecanoid pathway does not exhibit PG activity or H2O2 in response to wounding, systemin, OGA, or chitosan, but does respond to MJ, indicating that the generation of H2O2 requires a functional octadecanoid signaling pathway. Among 18 plant species from six families that were assayed for wound-inducible PG activity and H2O2 generation, 14 species exhibited both wound-inducible PG activity and the generation of H2O2. Four species, all from the Fabaceae family, exhibited little or no wound-inducible PG activity and did not generate H2O2. The time course of wound-inducible PG activity and H2O2 in Arabidopsis thaliana leaves was similar to that found in tomato. The cumulative data suggest that systemic wound signals that induce PG activity and H2O2 are widespread in the plant kingdom and that the response may be associated with the defense of plants against both herbivores and pathogens.
DOI:10.5511/plantbiotechnology.13.1028bURL [本文引用: 2]
DOI:10.1111/tpj.12654URLPMID:25182467 [本文引用: 1]
One week after partial incision of Arabidopsis inflorescence stems, the repair process in damaged tissue includes pith cell proliferation. Auxin is a key factor driving this process, and ANAC071, a transcription factor gene, is upregulated in the distal region of the incised stem. Here we show that XTH20 and the closely related XTH19, members of xyloglucan endotransglucosylase/hydrolases family catalyzing molecular grafting and/or hydrolysis of cell wall xyloglucans, were also upregulated in the distal part of the incised stem, similar to ANAC071. XTH19 was expressed in the proximal incision region after 3 days or after auxin application to the decapitated stem. Horizontal positioning of the plant with the incised side up resulted in decreased ProDR 5 :GUS, ANAC071, XTH20, and XTH19 expression and reduced pith cell proliferation. In incised stems of Pro35S :ANAC071-SRDX plants, expression of XTH20 and XTH19 was substantially and moderately decreased, respectively. XTH20 and XTH19 expression and pith cell proliferation were suppressed in anac071 plants and were increased in Pro35S :ANAC071 plants. Pith cell proliferation was also inhibited in the xth20xth19 double mutant. Furthermore, ANAC071 bound to the XTH20 and XTH19 promoters to induce their expression. This study revealed XTH20 and XTH19 induction by auxin via ANAC071 in the distal part of an incised stem and their involvement in cell proliferation in the tissue reunion process.
DOI:10.1046/j.1365-313x.1999.00570.xURL [本文引用: 1]
DOI:10.3390/ijms20102479URL [本文引用: 1]
DOI:10.1016/j.pbi.2005.05.008URLPMID:15939667 [本文引用: 1]
In many plants, localized tissue damage elicits an array of systemic defense responses against herbivore attack. Progress in our understanding of the long-distance signaling events that control these responses has been aided by the identification of mutants that fail to mount systemic defenses in response to wounding. Grafting experiments conducted with various mutants of tomato indicate that systemic signaling requires both the biosynthesis of jasmonic acid at the site of wounding and the ability to perceive a jasmonate signal in remote tissues. These and other studies support the hypothesis that jasmonic acid regulates the production of, or acts as, a mobile wound signal. Following its synthesis in peroxisomes, further metabolism of jasmonic acid might enhance its stability, transport, or action in remote tissues. Recent studies in tomato suggest that the peptide signal systemin promotes long-distance defense responses by amplifying jasmonate production in vascular tissues.
DOI:10.1038/nature07597URLPMID:19182776 [本文引用: 1]
Plants rely on the maintenance of stem cell niches at their apices for the continuous growth of roots and shoots. However, although the developmental plasticity of plant cells has been demonstrated, it is not known whether the stem cell niche is required for organogenesis. Here we explore the capacity of a broad range of differentiating cells to regenerate an organ without the activity of a stem cell niche. Using a root-tip regeneration system in Arabidopsis thaliana to track the molecular and functional recovery of cell fates, we show that re-specification of lost cell identities begins within hours of excision and that the function of specialized cells is restored within one day. Critically, regeneration proceeds in plants with mutations that fail to maintain the stem cell niche. These results show that stem-cell-like properties that mediate complete organ regeneration are dispersed in plant meristems and are not restricted to niches, which nonetheless seem to be necessary for indeterminate growth. This regenerative reprogramming of an entire organ without transition to a stereotypical stem cell environment has intriguing parallels to recent reports of induced transdifferentiation of specific cell types in the adult organs of animals.
DOI:10.1038/nature09430URLPMID:20927106 [本文引用: 2]
Jasmonates are a family of plant hormones that regulate plant growth, development and responses to stress. The F-box protein CORONATINE INSENSITIVE 1 (COI1) mediates jasmonate signalling by promoting hormone-dependent ubiquitylation and degradation of transcriptional repressor JAZ proteins. Despite its importance, the mechanism of jasmonate perception remains unclear. Here we present structural and pharmacological data to show that the true Arabidopsis jasmonate receptor is a complex of both COI1 and JAZ. COI1 contains an open pocket that recognizes the bioactive hormone (3R,7S)-jasmonoyl-l-isoleucine (JA-Ile) with high specificity. High-affinity hormone binding requires a bipartite JAZ degron sequence consisting of a conserved alpha-helix for COI1 docking and a loop region to trap the hormone in its binding pocket. In addition, we identify a third critical component of the jasmonate co-receptor complex, inositol pentakisphosphate, which interacts with both COI1 and JAZ adjacent to the ligand. Our results unravel the mechanism of jasmonate perception and highlight the ability of F-box proteins to evolve as multi-component signalling hubs.
DOI:10.1038/nature25184URLPMID:29320478 [本文引用: 1]
The cells of multicellular organisms receive extracellular signals using surface receptors. The extracellular domains (ECDs) of cell surface receptors function as interaction platforms, and as regulatory modules of receptor activation. Understanding how interactions between ECDs produce signal-competent receptor complexes is challenging because of their low biochemical tractability. In plants, the discovery of ECD interactions is complicated by the massive expansion of receptor families, which creates tremendous potential for changeover in receptor interactions. The largest of these families in Arabidopsis thaliana consists of 225 evolutionarily related leucine-rich repeat receptor kinases (LRR-RKs), which function in the sensing of microorganisms, cell expansion, stomata development and stem-cell maintenance. Although the principles that govern LRR-RK signalling activation are emerging, the systems-level organization of this family of proteins is unknown. Here, to address this, we investigated 40,000 potential ECD interactions using a sensitized high-throughput interaction assay, and produced an LRR-based cell surface interaction network (CSI(LRR)) that consists of 567 interactions. To demonstrate the power of CSI(LRR) for detecting biologically relevant interactions, we predicted and validated the functions of uncharacterized LRR-RKs in plant growth and immunity. In addition, we show that CSI(LRR) operates as a unified regulatory network in which the LRR-RKs most crucial for its overall structure are required to prevent the aberrant signalling of receptors that are several network-steps away. Thus, plants have evolved LRR-RK networks to process extracellular signals into carefully balanced responses.
DOI:10.1016/j.tplants.2015.11.002URLPMID:26655263 [本文引用: 3]
Plant receptor kinases (RKs) and receptor proteins (RPs) are involved in a plethora of cellular processes, including developmental decisions and immune responses. There is increasing evidence that plasmodesmata (PD)-localized RKs and RPs act as nexuses that perceive extracellular signals and convey them into intra- and intercellular responses by regulating the exchange of molecules through PD. How RK/RP complexes regulate the specific and nonspecific traffic of molecules through PD, and how these receptors are specifically targeted to PD, have been elusive but underpin comprehensive understanding of the function and regulation of the symplast. In this review we gather the current knowledge of RK/RP complex function at PD and how they might regulate intercellular traffic.
DOI:10.1016/j.cub.2009.03.060URLPMID:19398337 [本文引用: 1]
The niches of the Arabidopsis shoot and root meristems, the organizing center (OC) and the quiescent center (QC), orchestrate the fine balance of stem cell maintenance and the provision of differentiating descendants. They express the functionally related homeobox genes WUSCHEL (WUS) and WOX5, respectively, that promote stem cell fate in adjacent cells. Shoot stem cells signal back to the OC by secreting the CLAVATA3 (CLV3) dodecapeptide, which represses WUS expression. However, the signals controlling homeostasis of the root stem cell system are not identified to date. Here we show that the differentiating descendants of distal root stem cells express CLE40, a peptide closely related to CLV3. Reducing CLE40 levels delays differentiation and allows stem cell proliferation. Conversely, increased CLE40 levels drastically alter the expression domain of WOX5 and promote stem cell differentiation. We report that the receptor kinase ACR4, previously shown to control cell proliferation, is an essential component, and also a target, of CLE40 signaling. Our results reveal how, in contrast to the shoot system, signals originating from differentiated cells, but not the stem cells, determine the size and position of the root niche.
DOI:10.1016/j.devcel.2010.02.004URLPMID:20230752 [本文引用: 1]
Unlike most animal cells, plant cells can easily regenerate new tissues from a wide variety of organs when properly cultured. The common elements that provide varied plant cells with their remarkable regeneration ability are still largely unknown. Here we describe the initial process of Arabidopsis in vitro regeneration, where a pluripotent cell mass termed callus is induced. We demonstrate that callus resembles the tip of a root meristem, even if it is derived from aerial organs such as petals, which clearly shows that callus formation is not a simple reprogramming process backward to an undifferentiated state as widely believed. Furthermore, callus formation in roots, cotyledons, and petals is blocked in mutant plants incapable of lateral root initiation. It thus appears that the ectopic activation of a lateral root development program is a common mechanism in callus formation from multiple organs.
DOI:10.1007/s00709-010-0217-6URLPMID:20938697 [本文引用: 1]
A significant amount of work has been expended to identify the elusive components of plasmodesmata (PD) to help understand their structure, as well as how proteins are targeted to them. This review focuses on the role that lipid membranes may play in defining PD both structurally and as subcellular targeting addresses. Parallels are drawn to findings in other areas of research which focus on the lateral segregation of membrane domains and the generation of three-dimensional organellar shapes from flat lipid bilayers. We conclude that consideration of the protein-lipid interactions in cell biological studies of PD components and PD-targeted proteins may yield new insights into some of the many open questions regarding these unique structures.
DOI:10.1046/j.1365-313x.2002.01419.xURLPMID:12383090 [本文引用: 2]
Grafting in species other than Arabidopsis has generated persuasive evidence for long-distance signals involved in many plant processes, including regulation of flowering time and shoot branching. Hitherto, such approaches in Arabidopsis have been hampered by the lack of suitable grafting techniques. Here, a range of micrografting methods for young Arabidopsis seedlings are described. The simplest configuration was a single-hypocotyl graft, constructed with or without a supporting collar, allowing tests of root-shoot communication. More complex two-shoot grafts were also constructed, enabling tests of shoot-shoot communication. Integrity of grafts and absence of adventitious roots on scions were assessed using plants constitutively expressing a GUS gene as one graft partner. Using the max1 (more axillary growth) and max3 increased branching mutants, it was shown that a wild-type (WT) rootstock was able to inhibit rosette branching of mutant shoots. In two-shoot grafts with max1 and WT shoots on a max1 rootstock, the mutant shoot branched profusely, but the WT one did not. In two-shoot grafts with max1 and WT shoots on a WT rootstock, neither shoot exhibited increased branching. The results mirror those previously demonstrated in equivalent grafting experiments with the ramosus mutants in pea, and are consistent with the concept that a branching signal is capable of moving from root to shoot, but not from shoot to shoot. These grafting procedures will be valuable for revealing genes associated with many other long-distance signalling pathways, including flowering, systemic resistance and abiotic stress responses.
DOI:10.1242/dev.113878URLPMID:25256344 [本文引用: 1]
Tissue morphogenesis in plants requires communication between cells, a process involving the trafficking of molecules through plasmodesmata (PD). PD conductivity is regulated by endogenous and exogenous signals. However, the underlying signaling mechanisms remain enigmatic. In Arabidopsis, signal transduction mediated by the receptor-like kinase STRUBBELIG (SUB) contributes to inter-cell layer signaling during tissue morphogenesis. Previous analysis has revealed that SUB acts non-cell-autonomously suggesting that SUB controls tissue morphogenesis by participating in the formation or propagation of a downstream mobile signal. A genetic screen identified QUIRKY (QKY), encoding a predicted membrane-anchored C2-domain protein, as a component of SUB signaling. Here, we provide further insight into the role of QKY in this process. We show that like SUB, QKY exhibits non-cell-autonomy when expressed in a tissue-specific manner and that non-autonomy of QKY extends across several cells. In addition, we report on localization studies indicating that QKY and SUB localize to PD but independently of each other. FRET-FLIM analysis suggests that SUB and QKY are in close contact at PD in vivo. We propose a model where SUB and QKY interact at PD to promote tissue morphogenesis, thereby linking RLK-dependent signal transduction and intercellular communication mediated by PD.
[本文引用: 1]
DOI:10.1111/nph.14383URLPMID:27991666 [本文引用: 1]
Grafting has been widely used to improve horticultural traits. It has also served increasingly as a tool to investigate the long-distance transport of molecules that is an essential part for key biological processes. Many studies have revealed the molecular mechanisms of graft-induced phenotypic variation in anatomy, morphology and production. Here, we review the phenomena and their underlying mechanisms by which macromolecules, including RNA, protein, and even DNA, are transported between scions and rootstocks via vascular tissues. We further propose a conceptual framework that characterizes and quantifies the driving mechanisms of scion-rootstock interactions toward vascular reconnection and regeneration.
DOI:10.1111/j.1365-313X.2006.02977.xURLPMID:17217464 [本文引用: 1]
Genetic evidence links the Arabidopsis MONOPTEROS (MP) and PIN-FORMED1 (PIN1) genes to the patterning of leaf veins. To elucidate their potential functions and interactions in this process, we have assessed the dynamics of MP and PIN1 expression during vascular patterning in Arabidopsis leaf primordia. Both genes undergo a dynamic process of gradual refinement of expression into files one to two cells wide before overt vascular differentiation. The subcellular distribution of PIN1 is also gradually refined from a non-polar distribution in isodiametric cells to strongly polarized in elongated procambial cells and provides an indication of overall directions of auxin flow. We found evidence that MP expression can be activated by auxin exposure and that PIN1 as well as DR5::GUS expression is defective in mp mutant leaves. Taken together the results suggest a feedback regulatory loop that involves auxin, MP and PIN1 and provide novel experimental support for the canalization-of-auxin-flow hypothesis.
DOI:10.1186/s12870-019-1976-7URLPMID:31445524 [本文引用: 4]
BACKGROUND: The ability of severed rootstocks and shoots to re-establish vascular connections is used to generate grafted plants that combine desirable traits from both scions and rootstocks. Clarifying the mechanisms of graft healing is essential for its further application. We performed RNA sequencing of internodes near the cut position, making a distinction between separated or grafted tissues above and below the cut, in order to obtain a genetic description of graft union formation. RESULTS: Using weighted gene co-expression analysis, variable transcripts were clustered into 10 distinct co-expression networks (modules) based on expression profiles, and genes with the most
DOI:10.1093/jxb/ers109URLPMID:22511803 [本文引用: 5]
Grafting is an ancient cloning method that has been used widely for thousands of years in agricultural practices. Graft-union development is also an intricate process that involves substantial changes such as organ regeneration and genetic material exchange. However, the molecular mechanisms for graft-union development are still largely unknown. Here, a micrografting method that has been used widely in Arabidopsis was improved to adapt it a smooth procedure to facilitate sample analysis and to allow it to easily be applied to various dicotyledonous plants. The developmental stage of the graft union was characterized based on this method. Histological analysis suggested that the transport activities of vasculature were recovered at 3 days after grafting (dag) and that auxin modulated the vascular reconnection at 2 dag. Microarray data revealed a signal-exchange process between cells of the scion and stock at 1 dag, which re-established the communication network in the graft union. This process was concomitant with the clearing of cell debris, and both processes were initiated by a wound-induced programme. The results demonstrate the feasibility and potential power of investigating various plant developmental processes by this method, and represent a primary and significant step in interpretation of the molecular mechanisms underlying graft-union development.
嫁接植株形成过程中接合部组织学和生长素含量的变化
1
1995
... 砧穗嫁接面能够完全愈合是嫁接成功的关键, 而愈合程度具有很大的不确定性, 受遗传背景、发育阶段和栽培条件等因素的制约(
植物嫁接体接口愈合机制的研究进展
1
2017
... 砧穗嫁接面能够完全愈合是嫁接成功的关键, 而愈合程度具有很大的不确定性, 受遗传背景、发育阶段和栽培条件等因素的制约(
植物嫁接系统及其在植物生命科学研究中的应用
1
2011
... 砧穗嫁接面能够完全愈合是嫁接成功的关键, 而愈合程度具有很大的不确定性, 受遗传背景、发育阶段和栽培条件等因素的制约(
夜温对辣椒套管嫁接苗砧穗愈合的影响
1
2015
... 砧穗嫁接面能够完全愈合是嫁接成功的关键, 而愈合程度具有很大的不确定性, 受遗传背景、发育阶段和栽培条件等因素的制约(
Spatially selective hormonal control of RAP2.6L and ANAC071 transcription factors involved in tissue reunion in Arabidopsis
4
2011
... 砧穗组织重连是嫁接愈合完成的标志性事件, 包括维管组织和其它组织的重连等.已开发出一些模式化的切割或嫁接系统用于组织重连研究, 如拟南芥下胚轴微嫁接(micrograft)或改良后的微嫁接(
... 在幼苗期, 赤霉素(gibberellin, GA)的生物合成影响皮层的组织重连.当切除黄瓜和番茄幼苗下胚轴直径的一半长度后, 皮层的细胞分裂起始于切割后3天, 7天后重连.去除子叶后重连被抑制.在地上部尖端施加GA能够逆转抑制作用, 而施加GA抑制剂, 以及GA缺陷突变体(gib-1)呈现与去除子叶类似的表型(
... 转录因子在组织重连过程中作用显著, 其表达受到生长素、JAs和ETH的交联调控.生长素促进转录因子ANAC071 (NAC domain containing protein 71)和ANAC096的表达, 生长素响应因子IAA5 (indole-3- acetic acid inducible 5)、ARF6 (auxin response factor 6)和ARF8居于级联调控的中部, 推测其在嫁接面以上发挥作用(
... 的表达被抑制后, 重连不能正常进行.虽然这2个转录因子的作用尚不完全明晰, 但它们均受到生长素浓度的影响, 并被创伤应激激素JAs和ETH微调(
Gibberellin produced in the cotyledon is required for cell division during tissue reunion in the cortex of cut cucumber and tomato hypocotyls
3
2002
... 砧穗组织重连是嫁接愈合完成的标志性事件, 包括维管组织和其它组织的重连等.已开发出一些模式化的切割或嫁接系统用于组织重连研究, 如拟南芥下胚轴微嫁接(micrograft)或改良后的微嫁接(
... 在幼苗期, 赤霉素(gibberellin, GA)的生物合成影响皮层的组织重连.当切除黄瓜和番茄幼苗下胚轴直径的一半长度后, 皮层的细胞分裂起始于切割后3天, 7天后重连.去除子叶后重连被抑制.在地上部尖端施加GA能够逆转抑制作用, 而施加GA抑制剂, 以及GA缺陷突变体(gib-1)呈现与去除子叶类似的表型(
... ).此外, 去除芽的顶端生长点, 或者施用生长素极性运输抑制剂(2,3,5-triiodobenzoic acid, TIBA)并不影响重连, 而施用吲哚-3-乙酸(indole-3-acetic acid, IAA)不能弥补去除子叶的重连抑制, 研究表明顶端生长点产生的生长素对去除子叶的黄瓜和番茄幼苗组织重连的影响并不大(
Effects of the removal of cotyledons on endogenous gibberellin levels in hypocotyls of young cucumber and tomato seedlings
1
2007
... 在幼苗期, 赤霉素(gibberellin, GA)的生物合成影响皮层的组织重连.当切除黄瓜和番茄幼苗下胚轴直径的一半长度后, 皮层的细胞分裂起始于切割后3天, 7天后重连.去除子叶后重连被抑制.在地上部尖端施加GA能够逆转抑制作用, 而施加GA抑制剂, 以及GA缺陷突变体(gib-1)呈现与去除子叶类似的表型(
Regulatory mechanisms for specification and patterning of plant vascular tissues
1
2010
... 生长素在植物多个发育进程中与维管组织的形成密切相关(
Two novel genes encoding SNF1-related protein kinases from Arabidopsis thaliana: differential accumulation of AtSR1 and AtSR2 transcripts in response to cytokinins and sugars, and phosphorylation of sucrose synthase by AtSR2
1
2001
... 寡糖(oligosaccharides)是细胞的能量来源, 同时也是一类信号分子, 在质膜上有专门的受体蛋白激酶(
Graft union formation in grapevine induces transcriptional changes related to cell wall modification, wounding, hormone signaling, and secondary metabolism
2
2013
... 从嫁接时间和部位上来看, 草本嫁接采用幼苗期的胚轴或幼嫩茎, 而木本嫁接通常采用越冬后即将打破休眠时期的枝干和发芽枝条(
... 由于培养周期长和技术原因, 木本嫁接的调控机制难以采用以草本植物为材料的类似方法进行探究.可通过转录组分析获得的嫁接特异性富集路径对草本和木本愈合过程进行比较.对于拟南芥、番茄、葡萄和美洲山核桃等不同物种, 嫁接后呈现出差异表达的基因参与相似的路径, 包括茉莉酸合成、ROS消解、类黄酮代谢、细胞壁重构、细胞周期、膜锚定受体介导的细胞通讯、生长素运输和响应及维管组织分化(
Triggering the cell cycle in plants
1
2000
... 愈伤组织的特征是细胞进入快速分裂周期.当受到糖(sucrose)、生长素和细胞分裂素等物质的诱导时, D类细胞周期蛋白(D-type cyclin, CYCD)与A类细胞周期蛋白依赖性蛋白激酶(A-type cyclin-dependent kinase, CDKA)结合, 形成激活态CYCD-CDKA复合体(
Regulation of preprocambial cell state acquisition by auxin signaling in Arabidopsis leaves
1
2009
... 生长素在植物多个发育进程中与维管组织的形成密切相关(
2
2015
... 活性氧(包括超氧阴离子自由基(O2.-)和过氧化氢(H2O2))在创伤后1小时内开始积累(
... ).此外, 由ROS引起的氧化还原电位改变常与双组分信号系统(two-com- ponent signaling system, TCS)整合, 改变一些氧化还原态敏感的转录因子活性(
Graft union formation in tomato plants: peroxidase and catalase involvement
1
2004
... 活性氧(包括超氧阴离子自由基(O2.-)和过氧化氢(H2O2))在创伤后1小时内开始积累(
Merging genotypes: graft union formation and scion-rootstock interactions
1
2019
... 从嫁接时间和部位上来看, 草本嫁接采用幼苗期的胚轴或幼嫩茎, 而木本嫁接通常采用越冬后即将打破休眠时期的枝干和发芽枝条(
Velocity estimates for signal propagation leading to systemic jasmonic acid accumulation in wounded Arabidopsis
1
2009
... 切割造成嫁接面时, 创伤应激响应是首先发生的事件.茉莉酸信号途径是其中较为重要的调控路径.在番茄(Solanum lycopersicum)中的研究证实, 创伤发生后的短时间内, 内源性植物激素茉莉酸(jasmonic acid, JA)及其活性衍生物, 如茉莉酸甲酯(methyl jasmonate, MeJA)、茉莉酸-异亮氨酸复合物(jasmonoylisoleucine, JA-Ile)以及顺式茉莉酸(cis-jasmone, CJ) 被迅速合成(
Spatial and temporal dynamics of jasmonate synthesis and accumulation in Arabidopsis in response to wounding
1
2008
... 切割造成嫁接面时, 创伤应激响应是首先发生的事件.茉莉酸信号途径是其中较为重要的调控路径.在番茄(Solanum lycopersicum)中的研究证实, 创伤发生后的短时间内, 内源性植物激素茉莉酸(jasmonic acid, JA)及其活性衍生物, 如茉莉酸甲酯(methyl jasmonate, MeJA)、茉莉酸-异亮氨酸复合物(jasmonoylisoleucine, JA-Ile)以及顺式茉莉酸(cis-jasmone, CJ) 被迅速合成(
Plant grafting: new mechanisms, evolutionary implications
3
2014
... 嫁接是指将一个植株带有芽或枝的接穗(scion)与另一个植株带有根系的砧木(stock)通过机械固定使切割产生的嫁接面(graft interface)紧贴, 培养至砧、穗在嫁接结合部相互愈合, 从而获得嫁接体植株(grafting plant)的技术.优良的嫁接体植株因结合了接穗和砧木各自的优势而具有一种或多种显著的增益效果, 如生物或非生物胁迫抗性增强、产量提高、品质改良、开花或块茎化习性优化以及株型矮化(
... 砧穗嫁接面能够完全愈合是嫁接成功的关键, 而愈合程度具有很大的不确定性, 受遗传背景、发育阶段和栽培条件等因素的制约(
... 嫁接技术最早应用于多年生果树、林木和观赏花卉等木本植物, 20世纪初期开始大量应用于茄果类和瓜果类蔬菜等草本植物, 近年来有关嫁接生理的研究大多集中在蔬菜嫁接(
The roles of hormones in defense against insects and disease. Plant hormones.
2
2010
... 切割造成嫁接面时, 创伤应激响应是首先发生的事件.茉莉酸信号途径是其中较为重要的调控路径.在番茄(Solanum lycopersicum)中的研究证实, 创伤发生后的短时间内, 内源性植物激素茉莉酸(jasmonic acid, JA)及其活性衍生物, 如茉莉酸甲酯(methyl jasmonate, MeJA)、茉莉酸-异亮氨酸复合物(jasmonoylisoleucine, JA-Ile)以及顺式茉莉酸(cis-jasmone, CJ) 被迅速合成(
... COI1-JAZ复合体是JAs的高亲和受体(
The history, current status and future prospects of vegetable grafting in China
1
2015
... 嫁接是指将一个植株带有芽或枝的接穗(scion)与另一个植株带有根系的砧木(stock)通过机械固定使切割产生的嫁接面(graft interface)紧贴, 培养至砧、穗在嫁接结合部相互愈合, 从而获得嫁接体植株(grafting plant)的技术.优良的嫁接体植株因结合了接穗和砧木各自的优势而具有一种或多种显著的增益效果, 如生物或非生物胁迫抗性增强、产量提高、品质改良、开花或块茎化习性优化以及株型矮化(
Wounding triggers callus formation via dynamic hormonal and transcriptional changes
3
2017
... 愈伤组织在嫁接面处积累, 填充砧穗之间的空隙, 其中的一些薄壁细胞后续分化为新的维管组织.由于植物组织的再生不需要干细胞龛(stem cell niche)来维持分化, 因而具有更灵活的脱分化(dedifferentiation)和再分化(redifferentiation)能力(
... 愈伤组织的特征是细胞进入快速分裂周期.当受到糖(sucrose)、生长素和细胞分裂素等物质的诱导时, D类细胞周期蛋白(D-type cyclin, CYCD)与A类细胞周期蛋白依赖性蛋白激酶(A-type cyclin-dependent kinase, CDKA)结合, 形成激活态CYCD-CDKA复合体(
... 在创伤应激响应中合成量升高的JAs和ABA等植物激素, 在愈伤组织形成中不是必需的.使用JAs或ABA的合成和信号转导突变体进行研究, 发现突变体愈伤组织形成量较野生型轻微但显著增多, 表明JAs和ABA对愈伤组织的形成具有轻微抑制作用(
Plant callus: mechanisms of induction and repression
2
2013
... 此外, 还有细胞壁框架产生的物理抑制.细胞壁组分如纤维素(cellulose)、半纤维素(hemicellulose)和果胶(pectin)的有序积累, 是建立和维持细胞分化形态的关键步骤(
... 突变, 导致细胞间不能相互黏连的严重缺陷(
Cell cycle regulation in plant development
1
2006
... 愈伤组织的特征是细胞进入快速分裂周期.当受到糖(sucrose)、生长素和细胞分裂素等物质的诱导时, D类细胞周期蛋白(D-type cyclin, CYCD)与A类细胞周期蛋白依赖性蛋白激酶(A-type cyclin-dependent kinase, CDKA)结合, 形成激活态CYCD-CDKA复合体(
WIND1 promotes shoot regeneration through transcriptional activation of ENHANCER OF SHOOT REGENERATION1 in Arabidopsis
1
2017
... 愈伤组织在嫁接面处积累, 填充砧穗之间的空隙, 其中的一些薄壁细胞后续分化为新的维管组织.由于植物组织的再生不需要干细胞龛(stem cell niche)来维持分化, 因而具有更灵活的脱分化(dedifferentiation)和再分化(redifferentiation)能力(
The AP2/ERF transcription factor WIND1 controls cell dedifferentiation in Arabidopsis
3
2011
... 愈伤组织在嫁接面处积累, 填充砧穗之间的空隙, 其中的一些薄壁细胞后续分化为新的维管组织.由于植物组织的再生不需要干细胞龛(stem cell niche)来维持分化, 因而具有更灵活的脱分化(dedifferentiation)和再分化(redifferentiation)能力(
... ).AP2/ERF类转录因子WIND1 (wound induced dedifferentiation 1)及其同源蛋白是创伤后组织发育命运的重要调控因子, 其高丰度表达从创伤后数小时一直持续至愈合完全(
... response regulator)因子介导的信号途径促进细胞的脱分化(
Causes of defoliation and low survival rate of grafted sweet pepper plants
1
2009
... 砧穗嫁接面能够完全愈合是嫁接成功的关键, 而愈合程度具有很大的不确定性, 受遗传背景、发育阶段和栽培条件等因素的制约(
Ascorbic acid promotes graft-take in sweet pepper plants (Capsicum annuum L.)
1
2008
... 活性氧(包括超氧阴离子自由基(O2.-)和过氧化氢(H2O2))在创伤后1小时内开始积累(
Anatomy and physiology of graft incompatibility in solanaceous plants
1
2008
... 砧穗嫁接面能够完全愈合是嫁接成功的关键, 而愈合程度具有很大的不确定性, 受遗传背景、发育阶段和栽培条件等因素的制约(
Kinetics of salicylate-mediated suppression of jasmonate signaling reveal a role for redox modulation
1
2008
... 除茉莉酸信号途径以外, 还存在其它创伤应激响应机制.使用coi1/jaz突变体筛选出多种参与创伤响应的非JAs依赖的效应物, 包括乙烯(ethylene, ETH)、脱落酸(abscisic acid, ABA)、活性氧(reactive oxygen species, ROS)、寡聚半乳糖醛酸(oligogalacturonides, OGAs)、一氧化氮(nitric oxide, NO)和脂肪酸氨基酸复合物(fatty acid-amino acid conjugates, FACs)等(
Current status of vegetable grafting: diffusion, grafting techniques, automation
1
2010
... 嫁接是指将一个植株带有芽或枝的接穗(scion)与另一个植株带有根系的砧木(stock)通过机械固定使切割产生的嫁接面(graft interface)紧贴, 培养至砧、穗在嫁接结合部相互愈合, 从而获得嫁接体植株(grafting plant)的技术.优良的嫁接体植株因结合了接穗和砧木各自的优势而具有一种或多种显著的增益效果, 如生物或非生物胁迫抗性增强、产量提高、品质改良、开花或块茎化习性优化以及株型矮化(
A plasmodesmata-localized protein mediates crosstalk between cell-to-cell communication and innate immunity in Arabidopsis
1
2011
... PD上的RKs和RPs还可作为模式识别受体(pattern recognition receptors)结合病原衍生配体(pathogen-derived ligand), 如真菌细胞壁的几丁质(chitin)或细菌的鞭毛蛋白(falgellin), 可激活胞内免疫应答(
Plasmodesmata: a signaling hub at the cellular boundary
2
2015
... PD上的RKs和RPs还可作为模式识别受体(pattern recognition receptors)结合病原衍生配体(pathogen-derived ligand), 如真菌细胞壁的几丁质(chitin)或细菌的鞭毛蛋白(falgellin), 可激活胞内免疫应答(
... ), 在嫁接愈合初期, 嫁接面细胞壁不健全, 病原菌入侵是大概率事件.作为共质体运输的门户, 胞间连丝能够通过外膜和细胞壁之间拐角处填充的胼胝质(callose)来调节自身通透性(permeability).胼胝质的积累量随环境或发育状况而变化, 与PD的通透性呈负相关(
Differential cellular control by cotyledon-derived phytohormones involved in graft reunion of Arabidopsis hypocotyls
2
2016
... 在幼苗期, 赤霉素(gibberellin, GA)的生物合成影响皮层的组织重连.当切除黄瓜和番茄幼苗下胚轴直径的一半长度后, 皮层的细胞分裂起始于切割后3天, 7天后重连.去除子叶后重连被抑制.在地上部尖端施加GA能够逆转抑制作用, 而施加GA抑制剂, 以及GA缺陷突变体(gib-1)呈现与去除子叶类似的表型(
... 转录因子在组织重连过程中作用显著, 其表达受到生长素、JAs和ETH的交联调控.生长素促进转录因子ANAC071 (NAC domain containing protein 71)和ANAC096的表达, 生长素响应因子IAA5 (indole-3- acetic acid inducible 5)、ARF6 (auxin response factor 6)和ARF8居于级联调控的中部, 推测其在嫁接面以上发挥作用(
Vascular cambium regeneration and vessel formation in wounded inflorescence stems of Arabidopsis
2
2016
... 生长素在植物多个发育进程中与维管组织的形成密切相关(
... ).生长素在拟南芥花序茎半切损伤愈合中也会产生渠化效应, 由PIN1融合报告基因标识出的生长素运输轨迹先于新维管组织的形成(
Transcriptome dynamics at Arabidopsis graft junctions reveal an intertissue recognition mechanism that activates vascular regeneration
1
2018
... 由于生长素和细胞分裂素的极性运输, 及其运输流被切割所阻断, 嫁接面上下组织中的调控模式呈现非对称的特点.借助携带生长素响应元件的DR5启动子和β-葡萄糖苷酸酶(β-glucuronidase, GUS)报告基因的转基因拟南芥, 观察微嫁接后或非嫁接的GUS染色部位, 发现当转基因拟南芥作为接穗, 野生型拟南芥作为砧木时, 在接穗嫁接面维管束附近部位有明显的染色, 1 DAG时开始出现, 3 DAG时弥漫至整个嫁接面; 而当转基因拟南芥作为砧木时, 嫁接面附近在1-3 DAG期间均未观察到明显染色(
Plant grafting
7
2015
... 愈伤组织在嫁接面处积累, 填充砧穗之间的空隙, 其中的一些薄壁细胞后续分化为新的维管组织.由于植物组织的再生不需要干细胞龛(stem cell niche)来维持分化, 因而具有更灵活的脱分化(dedifferentiation)和再分化(redifferentiation)能力(
... 由于生长素和细胞分裂素的极性运输, 及其运输流被切割所阻断, 嫁接面上下组织中的调控模式呈现非对称的特点.借助携带生长素响应元件的DR5启动子和β-葡萄糖苷酸酶(β-glucuronidase, GUS)报告基因的转基因拟南芥, 观察微嫁接后或非嫁接的GUS染色部位, 发现当转基因拟南芥作为接穗, 野生型拟南芥作为砧木时, 在接穗嫁接面维管束附近部位有明显的染色, 1 DAG时开始出现, 3 DAG时弥漫至整个嫁接面; 而当转基因拟南芥作为砧木时, 嫁接面附近在1-3 DAG期间均未观察到明显染色(
... )验证两类植物激素的作用, 结果表明, 对生长素的响应由接穗起始并随之扩散至嫁接结合部, 对细胞分裂素的响应激活由砧木起始并随之扩散至嫁接结合部(
... ).通过突变体筛选得出, 生长素响应蛋白ALF4 (aberrant lateral root formation 4)作为承接生长素信号的关键因子, 其正常功能在韧皮部重连过程中是必需的(
... 从嫁接时间和部位上来看, 草本嫁接采用幼苗期的胚轴或幼嫩茎, 而木本嫁接通常采用越冬后即将打破休眠时期的枝干和发芽枝条(
... ).木本植物嫁接部位的再生能力弱是造成愈合缓慢的可能原因.活跃的维管形成层是木本和草本植物嫁接部位愈合的必要条件.这与大部分双子叶植物、基部被子植物以及裸子植物可以嫁接, 而维管束散生和缺少维管形成层的单子叶植物很难嫁接相互印证(
... 由于培养周期长和技术原因, 木本嫁接的调控机制难以采用以草本植物为材料的类似方法进行探究.可通过转录组分析获得的嫁接特异性富集路径对草本和木本愈合过程进行比较.对于拟南芥、番茄、葡萄和美洲山核桃等不同物种, 嫁接后呈现出差异表达的基因参与相似的路径, 包括茉莉酸合成、ROS消解、类黄酮代谢、细胞壁重构、细胞周期、膜锚定受体介导的细胞通讯、生长素运输和响应及维管组织分化(
A developmental framework for graft formation and vascular reconnection in Arabidopsis thaliana
0
2015
Reactive oxygen gene network of plants
1
2004
... 活性氧(包括超氧阴离子自由基(O2.-)和过氧化氢(H2O2))在创伤后1小时内开始积累(
Transcriptomic analysis provides insights into grafting union development in pecan ( Carya illinoinensis)
2
2018
... 从嫁接时间和部位上来看, 草本嫁接采用幼苗期的胚轴或幼嫩茎, 而木本嫁接通常采用越冬后即将打破休眠时期的枝干和发芽枝条(
... 由于培养周期长和技术原因, 木本嫁接的调控机制难以采用以草本植物为材料的类似方法进行探究.可通过转录组分析获得的嫁接特异性富集路径对草本和木本愈合过程进行比较.对于拟南芥、番茄、葡萄和美洲山核桃等不同物种, 嫁接后呈现出差异表达的基因参与相似的路径, 包括茉莉酸合成、ROS消解、类黄酮代谢、细胞壁重构、细胞周期、膜锚定受体介导的细胞通讯、生长素运输和响应及维管组织分化(
Graft formation in Kalanchoe blossfeldiana
1
1982
... 砧穗嫁接面能够完全愈合是嫁接成功的关键, 而愈合程度具有很大的不确定性, 受遗传背景、发育阶段和栽培条件等因素的制约(
Graft formation in Solanum pennellii (Solanaceae)
1
1984
... 砧穗嫁接面能够完全愈合是嫁接成功的关键, 而愈合程度具有很大的不确定性, 受遗传背景、发育阶段和栽培条件等因素的制约(
Glutamate receptor-like genes mediate leaf-to-leaf wound signaling
1
2013
... 切割造成嫁接面时, 创伤应激响应是首先发生的事件.茉莉酸信号途径是其中较为重要的调控路径.在番茄(Solanum lycopersicum)中的研究证实, 创伤发生后的短时间内, 内源性植物激素茉莉酸(jasmonic acid, JA)及其活性衍生物, 如茉莉酸甲酯(methyl jasmonate, MeJA)、茉莉酸-异亮氨酸复合物(jasmonoylisoleucine, JA-Ile)以及顺式茉莉酸(cis-jasmone, CJ) 被迅速合成(
The role of plant hormones during grafting
1
2018
... 砧穗嫁接面能够完全愈合是嫁接成功的关键, 而愈合程度具有很大的不确定性, 受遗传背景、发育阶段和栽培条件等因素的制约(
A glycopeptide regulating stem cell fate in Arabidopsis thaliana
1
2009
... 在有些发育进程中, 质膜或PD上的RKs和RPs通过非细胞自主性信号途径调控组织形态建成或干细胞分化命运.例如, 通过PD特异性蛋白质组鉴定到的SUB (strubbelig)属于非典型的富含亮氨酸重复受体激酶(LRR-RK), 它通过与另一个具有C2结构域的PD锚定蛋白QKY (quirky)互作, 将信号传递至下游(
Hydrogen peroxide is generated systemically in plant leaves by wounding and systemin via the octadecanoid pathway
1
1999
... 活性氧(包括超氧阴离子自由基(O2.-)和过氧化氢(H2O2))在创伤后1小时内开始积累(
ARF6 and ARF8 contribute to tissue reunion in incised Arabidopsis inflorescence stems
2
2014
... 转录因子在组织重连过程中作用显著, 其表达受到生长素、JAs和ETH的交联调控.生长素促进转录因子ANAC071 (NAC domain containing protein 71)和ANAC096的表达, 生长素响应因子IAA5 (indole-3- acetic acid inducible 5)、ARF6 (auxin response factor 6)和ARF8居于级联调控的中部, 推测其在嫁接面以上发挥作用(
... ).AP2/ERF类转录因子的另1个成员RAP2.6L (related to AP2.6L)、JAs和低浓度生长素可促进其表达, 推测其在嫁接面以下发挥作用(
XTH20 and XTH19 regulated by ANAC071 under auxin flow are involved in cell proliferation in incised Arabidopsis inflorescence stems
1
2014
... 转录因子在组织重连过程中作用显著, 其表达受到生长素、JAs和ETH的交联调控.生长素促进转录因子ANAC071 (NAC domain containing protein 71)和ANAC096的表达, 生长素响应因子IAA5 (indole-3- acetic acid inducible 5)、ARF6 (auxin response factor 6)和ARF8居于级联调控的中部, 推测其在嫁接面以上发挥作用(
Cross-talk between wound signaling pathways determines local versus systemic gene expression in Arabidopsis thaliana
1
1999
... 除茉莉酸信号途径以外, 还存在其它创伤应激响应机制.使用coi1/jaz突变体筛选出多种参与创伤响应的非JAs依赖的效应物, 包括乙烯(ethylene, ETH)、脱落酸(abscisic acid, ABA)、活性氧(reactive oxygen species, ROS)、寡聚半乳糖醛酸(oligogalacturonides, OGAs)、一氧化氮(nitric oxide, NO)和脂肪酸氨基酸复合物(fatty acid-amino acid conjugates, FACs)等(
Jasmonic acid signaling pathway in plants
1
2019
... 切割造成嫁接面时, 创伤应激响应是首先发生的事件.茉莉酸信号途径是其中较为重要的调控路径.在番茄(Solanum lycopersicum)中的研究证实, 创伤发生后的短时间内, 内源性植物激素茉莉酸(jasmonic acid, JA)及其活性衍生物, 如茉莉酸甲酯(methyl jasmonate, MeJA)、茉莉酸-异亮氨酸复合物(jasmonoylisoleucine, JA-Ile)以及顺式茉莉酸(cis-jasmone, CJ) 被迅速合成(
Systemic signaling in the wound response
1
2005
... 除茉莉酸信号途径以外, 还存在其它创伤应激响应机制.使用coi1/jaz突变体筛选出多种参与创伤响应的非JAs依赖的效应物, 包括乙烯(ethylene, ETH)、脱落酸(abscisic acid, ABA)、活性氧(reactive oxygen species, ROS)、寡聚半乳糖醛酸(oligogalacturonides, OGAs)、一氧化氮(nitric oxide, NO)和脂肪酸氨基酸复合物(fatty acid-amino acid conjugates, FACs)等(
Organ regeneration does not require a functional stem cell niche in plants
1
2009
... 愈伤组织在嫁接面处积累, 填充砧穗之间的空隙, 其中的一些薄壁细胞后续分化为新的维管组织.由于植物组织的再生不需要干细胞龛(stem cell niche)来维持分化, 因而具有更灵活的脱分化(dedifferentiation)和再分化(redifferentiation)能力(
Jasmonate perception by inositol-phosphate-potentiated COI1-JAZ co-receptor
2
2010
... COI1-JAZ复合体是JAs的高亲和受体(
... ).COI1 (coronatine insensitive 1)是具有F-box结构域的E3泛素化连接酶; JAZ蛋白通过JAs结构域与COI1结合, 通过ZIM结构域与下游bHLH类转录因子MYC2结合.感知JAs信号后, JAZ解除对MYC2的抑制, 后者结合在JAs诱导基因的启动子区, 激活其转录(
An extracellular network of Arabidopsis leucine-rich repeat receptor kinases
1
2018
... 胞间连丝和膜锚定受体一方面需要通过感知嫁接搭档组织或自身组织中的发育信号, 达到加强砧穗交流的目的; 另一方面需要响应创伤应激, 通过免疫应答阻断细胞与外界的联系.在这种情况下, 膜受体(如LRR-RK家族成员)以网络协作的形式对各种胞外信号进行平衡(
Receptor complex mediated regulation of symplastic traffic
3
2016
... 胞间连丝(plasmodesmata, PD)由外膜和内膜2层膜结构组成, 分别为质膜和内质网膜的延续, 但理化性质稍有差别(
... PD上的RKs和RPs还可作为模式识别受体(pattern recognition receptors)结合病原衍生配体(pathogen-derived ligand), 如真菌细胞壁的几丁质(chitin)或细菌的鞭毛蛋白(falgellin), 可激活胞内免疫应答(
... ).具有胞外DUF26结构域的PD定位受体蛋白(PD-located proteins, PDLPs)是控制胼胝质积累量和PD通透性的上游因子.通常认为, PDLPs接收本地ROS信号, 激活调控通路, 最终使胼胝质积累量增加及PD关闭(
A signaling module controlling the stem cell niche in Arabidopsis root meristems
1
2009
... 在有些发育进程中, 质膜或PD上的RKs和RPs通过非细胞自主性信号途径调控组织形态建成或干细胞分化命运.例如, 通过PD特异性蛋白质组鉴定到的SUB (strubbelig)属于非典型的富含亮氨酸重复受体激酶(LRR-RK), 它通过与另一个具有C2结构域的PD锚定蛋白QKY (quirky)互作, 将信号传递至下游(
Arabidopsis regeneration from multiple tissues occurs via a root development pathway
1
2010
... 愈伤组织在嫁接面处积累, 填充砧穗之间的空隙, 其中的一些薄壁细胞后续分化为新的维管组织.由于植物组织的再生不需要干细胞龛(stem cell niche)来维持分化, 因而具有更灵活的脱分化(dedifferentiation)和再分化(redifferentiation)能力(
Plasmodesmata viewed as specialised membrane adhesion sites
1
2011
... 胞间连丝(plasmodesmata, PD)由外膜和内膜2层膜结构组成, 分别为质膜和内质网膜的延续, 但理化性质稍有差别(
Micrografting techniques for testing long-distance signaling in Arabidopsis
2
2002
... 砧穗组织重连是嫁接愈合完成的标志性事件, 包括维管组织和其它组织的重连等.已开发出一些模式化的切割或嫁接系统用于组织重连研究, 如拟南芥下胚轴微嫁接(micrograft)或改良后的微嫁接(
... 嫁接技术最早应用于多年生果树、林木和观赏花卉等木本植物, 20世纪初期开始大量应用于茄果类和瓜果类蔬菜等草本植物, 近年来有关嫁接生理的研究大多集中在蔬菜嫁接(
The C2-domain protein QUIRKY and the receptor-like kinase STRUBBELIG localize to plasmodesmata and mediate tissue morphogenesis in Arabidopsis thaliana
1
2014
... 在有些发育进程中, 质膜或PD上的RKs和RPs通过非细胞自主性信号途径调控组织形态建成或干细胞分化命运.例如, 通过PD特异性蛋白质组鉴定到的SUB (strubbelig)属于非典型的富含亮氨酸重复受体激酶(LRR-RK), 它通过与另一个具有C2结构域的PD锚定蛋白QKY (quirky)互作, 将信号传递至下游(
Effect of nursery environmental condition and different cultivars on survival rate of grafted tomato seedling
1
2014
... 砧穗嫁接面能够完全愈合是嫁接成功的关键, 而愈合程度具有很大的不确定性, 受遗传背景、发育阶段和栽培条件等因素的制约(
Plant grafting: how genetic exchange promotes vascular reconnection
1
2017
... 嫁接是指将一个植株带有芽或枝的接穗(scion)与另一个植株带有根系的砧木(stock)通过机械固定使切割产生的嫁接面(graft interface)紧贴, 培养至砧、穗在嫁接结合部相互愈合, 从而获得嫁接体植株(grafting plant)的技术.优良的嫁接体植株因结合了接穗和砧木各自的优势而具有一种或多种显著的增益效果, 如生物或非生物胁迫抗性增强、产量提高、品质改良、开花或块茎化习性优化以及株型矮化(
Dynamics of MONOPTEROS and PIN-FORMED1 expression during leaf vein pattern formation in Arabidopsis thaliana
1
2007
... 生长素在植物多个发育进程中与维管组织的形成密切相关(
Gene co-expression network analysis reveals pathways associated with graft healing by asymmetric profiling in tomato
4
2019
... 活性氧(包括超氧阴离子自由基(O2.-)和过氧化氢(H2O2))在创伤后1小时内开始积累(
... 愈伤组织的特征是细胞进入快速分裂周期.当受到糖(sucrose)、生长素和细胞分裂素等物质的诱导时, D类细胞周期蛋白(D-type cyclin, CYCD)与A类细胞周期蛋白依赖性蛋白激酶(A-type cyclin-dependent kinase, CDKA)结合, 形成激活态CYCD-CDKA复合体(
... 从嫁接时间和部位上来看, 草本嫁接采用幼苗期的胚轴或幼嫩茎, 而木本嫁接通常采用越冬后即将打破休眠时期的枝干和发芽枝条(
... 由于培养周期长和技术原因, 木本嫁接的调控机制难以采用以草本植物为材料的类似方法进行探究.可通过转录组分析获得的嫁接特异性富集路径对草本和木本愈合过程进行比较.对于拟南芥、番茄、葡萄和美洲山核桃等不同物种, 嫁接后呈现出差异表达的基因参与相似的路径, 包括茉莉酸合成、ROS消解、类黄酮代谢、细胞壁重构、细胞周期、膜锚定受体介导的细胞通讯、生长素运输和响应及维管组织分化(
Graft-union development: a delicate process that involves cell-cell communication between scion and stock for local auxin accumulation
5
2012
... PD上的RKs和RPs还可作为模式识别受体(pattern recognition receptors)结合病原衍生配体(pathogen-derived ligand), 如真菌细胞壁的几丁质(chitin)或细菌的鞭毛蛋白(falgellin), 可激活胞内免疫应答(
... 寡糖(oligosaccharides)是细胞的能量来源, 同时也是一类信号分子, 在质膜上有专门的受体蛋白激酶(
... 砧穗组织重连是嫁接愈合完成的标志性事件, 包括维管组织和其它组织的重连等.已开发出一些模式化的切割或嫁接系统用于组织重连研究, 如拟南芥下胚轴微嫁接(micrograft)或改良后的微嫁接(
... 由于生长素和细胞分裂素的极性运输, 及其运输流被切割所阻断, 嫁接面上下组织中的调控模式呈现非对称的特点.借助携带生长素响应元件的DR5启动子和β-葡萄糖苷酸酶(β-glucuronidase, GUS)报告基因的转基因拟南芥, 观察微嫁接后或非嫁接的GUS染色部位, 发现当转基因拟南芥作为接穗, 野生型拟南芥作为砧木时, 在接穗嫁接面维管束附近部位有明显的染色, 1 DAG时开始出现, 3 DAG时弥漫至整个嫁接面; 而当转基因拟南芥作为砧木时, 嫁接面附近在1-3 DAG期间均未观察到明显染色(
... 由于培养周期长和技术原因, 木本嫁接的调控机制难以采用以草本植物为材料的类似方法进行探究.可通过转录组分析获得的嫁接特异性富集路径对草本和木本愈合过程进行比较.对于拟南芥、番茄、葡萄和美洲山核桃等不同物种, 嫁接后呈现出差异表达的基因参与相似的路径, 包括茉莉酸合成、ROS消解、类黄酮代谢、细胞壁重构、细胞周期、膜锚定受体介导的细胞通讯、生长素运输和响应及维管组织分化(

{kind=link}
{kind=link}