Xyloglucan and the Advances in Its Roles in Plant Tolerance to Stresses
Yingyan Xiao?, Weina Yuan, Jing Liu, Jian Meng, Qiming Sheng, Yehuan Tan, Chunxiang Xu*College of Horticulture, South China Agricultural University, Guangzhou 510642, China通讯作者: *E-mail:chxxu@scau.edu.cn
责任编辑: 白羽红
收稿日期:2020-02-10接受日期:2020-05-8网络出版日期:2020-11-01
| 基金资助: |
Corresponding authors: *E-mail:chxxu@scau.edu.cn
Received:2020-02-10Accepted:2020-05-8Online:2020-11-01

摘要
木葡聚糖(XyG)是一种存在于所有陆生植物细胞壁中的基质多糖, 是双子叶植物初生细胞壁中含量(20%-25%, w/w)最丰富的半纤维素成分。作为细胞壁的组分, XyG不仅与植物的生长发育密切相关, 还在植物抵抗各种生物和非生物逆境过程中发挥重要作用。XyG代谢相关基因主要通过改变植物细胞壁的组成以及对细胞壁进行重排进而改变细胞壁的弹性/硬度等特性, 影响植物的抗逆性。XyG及其寡糖也可能作为信号分子, 或与其它信号分子协同作用应对逆境胁迫。该文概述了XyG的结构与类型及参与XyG生物合成与降解的相关基因, 重点阐述XyG相关基因应答生物和非生物胁迫的作用机制。
关键词:
Abstract
Xyloglucan (XyG) is a matrix polysaccharide present in the cell wall of all land plants. It is the most abundant hemicellulose in the primary cell walls of dicots (20%-25%, w/w). As a very important plant cell wall component, XyG is not only involved in plant growth and development, but also plays important roles in responses of plants to various abiotic and biotic stresses. The use of genes involved in XyG biosynthesis and degradation possibly improve the tolerance of plants to stresses through influencing the cell wall structure (remodelling) and compositions. In addition, XyG and XyG oligosaccharides likely act as signaling molecules or cooperate with other signaling molecules to induce plant resistance. Here, we review the structure and variety of XyG, the genes involved in XyG biosynthesis and degradation, and advances in potential roles of XyG and XyG-related genes in responses to biotic and abiotic stresses.
Keywords:
PDF (1008KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
肖银燕, 袁伟娜, 刘静, 孟建, 盛奇明, 谭烨欢, 徐春香. 木葡聚糖及其在植物抗逆过程中的功能研究进展. 植物学报, 2020, 55(6): 777-787 doi:10.11983/CBB20020
Xiao Yingyan, Yuan Weina, Liu Jing, Meng Jian, Sheng Qiming, Tan Yehuan, Xu Chunxiang.

植物细胞壁是围绕在植物原生质体外的一种细胞结构, 由初生细胞壁和次生细胞壁组成。初生细胞壁位于外层, 是由纤维素、半纤维素和果胶等多糖及结构蛋白组成的一种复杂网络结构。次生细胞壁的结构与组成均与初生细胞壁有所不同, 除了纤维素和半纤维素外, 还含有木质素。植物细胞壁除了作为细胞的主要结构成分, 承担机械支撑、维持和决定细胞形态等功能外, 还在植物的生长发育(陈昆松等, 1999)及抵抗各种生物和非生物逆境过程中发挥重要作用(Houston et al., 2016; Vaahtera et al., 2019), 并影响作物的农艺性状(Hu et al., 2017)。
木葡聚糖(XyG)是双子叶植物初生壁中含量最为丰富的半纤维素成分, 约占细胞壁重量的20%-25%, 而禾本科植物中一般仅为2%-10% (Scheller and Ulvskov, 2010)。前人对XyG的结构、分类及参与XyG代谢过程的相关酶基因进行了详细介绍(李雄彪和张金忠, 1994; Del Bem and Vincentz, 2010; 解敏敏等, 2015; Pauly and Keegstra, 2016)。近年来, 随着分子生物学技术的发展, 包括半纤维素在内的细胞壁
成分代谢及其在植物生长发育和抗逆过程中的功能研究取得了令人鼓舞的进展。然而, 国内外尚未见关于XyG在植物抗逆过程中的功能及相关机理研究的综述。本文概述XyG结构和类型及参与XyG代谢的相关基因, 并重点介绍这些基因及其产物在植物抗逆过程中的功能及抗性机理。
1 XyG的结构与分类
XyG是一种由D-吡喃葡萄糖残基以β-1→4键相连构成主链的杂多糖, 葡萄糖残基在O-6位上可被D-吡喃木糖残基所取代(Kiefer et al., 1989)。主链上的葡萄糖残基和侧链上的木糖残基都能以特定连接方式在特定位置被各种糖残基取代(Scheller and Ulvskov, 2010)。根据XyG取代基及其连接方式, 目前已鉴定出24种独特结构(Pauly and Keegstra, 2016)。在标准命名法中分别用不同的单个字母代码来命名和区分不同类型的XyG: 如字母G代表主链上的葡萄糖残基未被任何其它糖/非糖基团取代, X代表葡萄糖残基O-6位被α-D-吡喃木糖基团取代, L代表木糖残基的O-2位被半乳糖基团取代, F代表半乳糖残基的O-2位被岩藻糖基团取代(Fry et al., 1993)。XyG可能含有O-乙酰基取代基, 发生乙酰化(Kiefer et al., 1989)。乙酰基团既可修饰主链的葡萄糖残基, 也可修饰支链半乳糖残基或其它糖残基。这些结构分别用相应字母加下划线表示。例如, XLFG型XyG表示每4个葡糖残基中前3个被木糖基化, 其中第2个木糖残基被半乳糖基团取代, 第3个被半乳糖基团取代后又进一步被岩藻糖基团修饰, 且半乳糖残基发生乙酰化(图1)。图1
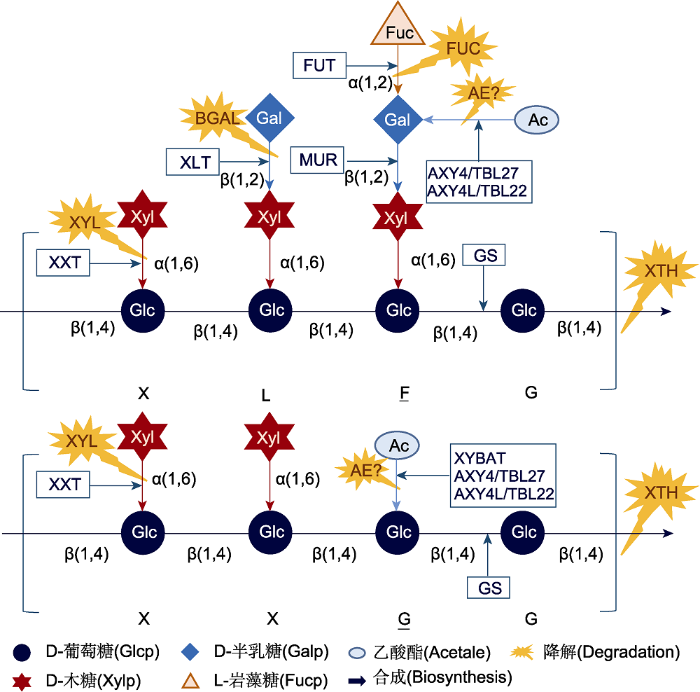 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1XLFG型和XXGG型木葡聚糖(XyG)的生物合成与降解示意图
木葡聚糖生物合成酶有FUT (XyG特异岩藻糖基转移酶)、GS (β-1,4-葡聚糖合成酶)、MUR/XLT (XyG特异半乳糖基转移酶)、AXY4L/TBL22和AXY4/TBL27 (XyG特异乙酰基转移酶)、XXT (XyG特异木糖基转移酶)以及XYBAT (XyG葡聚糖骨架乙酰转移酶)。XyG降解酶类有AE (乙酰酯酶)、BGAL (β-半乳糖苷酶)、FUC (α-岩藻糖苷酶)、XTH (XyG内转糖苷酶/水解酶)和XYL (α-木糖苷酶)。图下方为根据XyG命名法获得的相应单字母代码。G: XyG主链葡萄糖残基未连接其它任何糖残基或非糖基团; G: XyG主链葡萄糖残基发生乙酰化; X: XyG主链葡萄糖残基O-6位连接木糖基团; L: X型基团的木糖残基O-2位连接半乳糖基团; F: L型基团的半乳糖残基O-2位连接岩藻糖基团, 且半乳糖残基发生乙酰化。
Figure 1Schematic of the biosynthesis and degradation of XLFG-type and XXGG-type xyloglucan (XyG)
Enzymes involved in XyG biosynthesis are FUT (XyG: fucosyltransferase), GS (β-1,4-glucan synthase), MUR/XLT (XyG: galactosyltransferase), AXY4L/TBL22 and AXY4/TBL27 (XyG: acetyltransferases), XXT (XyG: xylosyltransferase), and XYBAT (XyG backbone acetyltransferase). Enzymes involved in XyG degradation are AE (acetylesterases), BGAL (β-galactosidase), FUC (α-fucosidase), XTH (XyG endotransglycosylase/hydrolases), and XYL (α-xylosidase). The corresponding one-letter codes from the XyG nomenclature have shown below the pictograms. G: Unsubstituted glucosyl residue of the backbone glucose of XyG; G: Backbone glucose of XyG carries an O-acetyl substituent; X: Xylosyl residue is attached to the glucan backbone of XyG at O-6; L: Galactosyl residue is attached to the xylosyl residue of X group at O-2; F: Fucosyl residue is attached to the galactosyl residue of L group at O-2, and galactosyl residue carries an O-acetyl substituent.
根据寡糖的结构可将XyG分成XXXG和XXGG两种主要类型。双子叶植物的XyG为XXXG型, 而禾本科和茄科植物的XyG为XXGG型。与XXXG型XyG相比, XXGG型XyG的木糖基化程度相对较低, 通常不发生岩藻糖基化(Pauly and Keegstra, 2016)。植物的XyG类型不仅具有物种特异性, 还因不同组织器官及发育阶段而异(Liu et al., 2015)。
XyG在结构和生物合成方面都是研究得最清楚的植物细胞壁多糖之一。许多XyG生物合成相关酶已被鉴定。与大多数复杂碳水化合物相同, XyG也在高尔基体中合成。
2 XyG代谢
2.1 XyG的生物合成
2.1.1 主链生物合成XyG主链骨架的生物合成由XyG特异葡聚糖合成酶, 即β-1,4-葡聚糖合成酶(β-1,4-glucan synthase, GS)负责(Cocuron et al., 2007), 该酶由类纤维素合成酶基因4 (cellulose synthase-like C4, CSLC4)编码, 后者属于碳水化合物活性酶(carbohydrate-active enzymes, CAZy)糖基转移酶2 (glycosyltransferase 2, GT2)家族。
2.1.2 骨架木糖化
所有类型的XyG葡聚糖主链都含有木糖基取代基。XyG特异木糖基转移酶(XyG: xylosyltransferase, XXT)负责将UDP-木糖添加到主链葡萄糖残基O-6位上(Faik et al., 2002)。目前, 已在拟南芥(Arabidopsis thaliana)中发现多个GT34基因家族成员编码XXTs (Cavalier et al., 2008)。此外, 在水稻(Oryza sativa)、番茄(Solanum lycopersicum)和旱金莲(Tropaeolum majus)的种子(Wang et al., 2014; Mansoori et al., 2015)中也发现GT34家族的XXT成员。
2.1.3 半乳糖苷化和其它类型的糖基化
大多数XyG木糖残基在GT47家族的一些酶作用下被其它糖基进一步取代, 其中最常见的是D-半乳糖。UDP-半乳糖在β-半乳糖基转移酶(β-galactosyltransferases, β-GAL)的作用下掺入XyG。例如, MUR3专一性负责XXXG第3位木糖残基的半乳糖化, 将XXXG变成XXLG (Kong et al., 2015; Xu et al., 2018), 而XLT2负责XXXG第2位木糖残基的半乳糖化, 将XXXG变成XLXG (Jensen et al., 2012)。此外, 木糖残基的O-2位也可能被其它糖残基修饰。近年来, 相继鉴定到一些负责糖基转移的酶, 如拟南芥XyG特异半乳糖醛酸基转移酶1 (XyG: galacturonosyltransferase 1, XUT1) (Pe?a et al., 2012)和番茄XyG特异阿拉伯呋喃糖基转移酶(SlXST1和SlXST2) (Schultink et al., 2013)。
2.1.4 岩藻糖基化
XXLG型XyG的半乳糖残基通常会被岩藻糖基进一步取代, 形成XXFG。XyG特异岩藻糖基转移酶(XyG: fucosyltransferases 1, FUT1)负责将GDP-岩藻糖添加到XXLG或XLLG的L上(Vanzin et al., 2002)。FUT1属于CAZy家族GT37, 是拟南芥基因组编码的唯一具有FUT活性的酶(Liang et al., 2013)。
2.1.5 骨架及侧链O-乙酰化
XyG的乙酰化主要发生在XXGG型XyG主链的葡萄糖残基及XXXG型XyG侧链的半乳糖残基上(Gille et al., 2011)。植物共有3个蛋白家族参与XyG的乙酰化, 即TBL (trichome birefringence-like)、AXY9 (ALTERED XYLOGLUCAN 9)和RWA (reduced wall acetylation) (Pawar et al., 2017; Pauly and Ramírez, 2018)。植物RWA被认为是乙酰基供体转运蛋白, AXY9蛋白起产生乙酰化中间体的作用(Schultink et al., 2015), 而TBL蛋白是多糖特异性的乙酰基转移酶(Gille et al., 2011; Manabe et al., 2011)。例如, 由TBL基因家族成员编码的AXY4/TBL27及AXY4-like (AXY4L/ TBL22)特异性地乙酰化种子中的XyG (Gille et al., 2011)。在二穗短柄草(Brachypodium distachyon)中也鉴定出1种XyG骨架乙酰转移酶(XyG glucan backbone acetyltransferase, XYBAT) (Liu et al., 2016)。
2.1.6 质外沉积与成熟
XyG在高尔基体中合成后, 被囊泡运输到细胞壁。迄今为止, 人们对XyG掺入现有细胞壁的过程知之甚少。已有研究表明, XyG可以通过非酶自组装的方式与现有纤维素微纤维网络结合(Hayashi et al., 1987)。XyG一旦沉积在壁中, 其结构保持动态, 原因是相关酶的靶标结构域会经历结构成熟和由质外体植物酶降解的周转(Franková and Fry, 2013)。
2.2 XyG的降解
2.2.1 主链降解与重排植物XyG内转糖苷酶/水解酶(endotransglycosylase/ hydrolases, XTHs)基因属于CAZy糖基水解酶16 (glycoside hydrolase 16, GH16)家族基因, 编码的蛋白可能具有2种不同的催化活性, 对XyG的影响不同。其中XyG特异内水解酶(XyG: endohydrolase, XEH)活性在形式上是XyG特异的内切葡聚糖酶(xyloglucan-specific endo-1,4-glucanases, XEGs), 降解XyG主链骨架(Hayashi et al., 1984); 而XyG特异内转糖基酶(XyG: endotransglycosylase, XET)活性则负责XyG网络重塑, 即XyG被降解后再重新连接到其它XyG链上, 从而改变XyG的长度(Franková and Fry, 2013)。XTHs编码的某些蛋白仅具有XEH活性, 从还原端将XyG水解为XyG寡糖(XyG oligosaccharides, XGO) (Shinohara et al., 2017)。此外, Sampedro等(2017)证明, 拟南芥有2个来自GH3家族的β-葡萄糖苷酶(β-GLU)能从非还原端裂解XyG主链骨架未被取代的Glc基团。
2.2.2 支链降解
参与XyG支链降解的糖苷酶主要包括α-木糖苷酶(如AtXYL1/AXY3) (Shigeyama et al., 2016)、β-半乳糖苷酶(如AtBGAL10)和α-岩藻糖苷酶(如AtFUC95A和AXY8) (Günl et al., 2011; Sampedro et al., 2012), 分别降解相应的糖残基。
3 XyG在植物抗逆中的功能
植物在整个生命周期中经常遭受各种生物和非生物逆境胁迫。相应地, 植物采取多种防御策略来应对逆境, 包括改变细胞壁结构与组成、表达包括与木葡聚糖代谢相关基因在内的抗逆基因(Xie et al., 2011; Ma et al., 2013; Yan et al., 2015; Wu et al., 2017; Bacete et al., 2018; Niu et al., 2018), 还可能通过启动抗性相关信号来应对逆境(Engelsdorf et al., 2018)。3.1 非生物胁迫
3.1.1 低温胁迫早在20多年前, 拟南芥XyG特异糖基化基因TCH4 (即AtXTH22)就已被证明非常耐(12-18)°C低温(Purugganan et al., 1997)。近年来, 大量组学研究结果表明多种植物的XyG代谢相关基因(如XTHs)响应低温胁迫, 并可能与植物的抗寒性密切相关(刘静, 2018; Takahashi et al., 2019)。推测低温下植物XTHs等基因表达量的改变影响XyG之间的共价交联以及XyG与纤维素微丝之间的联系, 从而改变细胞壁特性, 增强其抗冻性(Rao and Dixon, 2017)。此外, 研究发现sfr2基因编码的β-GLU是植物抗冻性必不可少的组成型表达蛋白(Thorlby et al., 2004)。鹰嘴豆(Cicer arietinum)在冷驯化过程中, 脱水蛋白和β-glu的表达量以及H2O2和丙二醛的含量显著升高, 耐冷能力增强(Khazaei et al., 2015)。
3.1.2 旱涝胁迫
研究表明, 植物XyG降解相关基因参与抗旱/涝过程。Kumar等(2019)发现在聚乙二醇模拟的干旱条件下, 2个耐旱类型鹰嘴豆中的XTH23基因均下调表达, 推测植物通过维持足够的木葡聚糖和纤维素含量来确保保卫细胞对气孔的控制(Rui and Anderson, 2016), 从而抵御逆境。此外, 在涝害条件下10个大豆(Glycine max) GmXTHs的表达量发生显著变化, 过表达拟南芥AtXTH31的大豆耐涝性增强(Song et al., 2018)。
3.1.3 盐胁迫
植物XyG代谢相关基因与其抗盐性的研究多见于XTH基因家族。例如, 转辣椒(Capsicum annuum) CaXTH3的拟南芥耐盐性增强, 推测其通过调节气孔关闭来防止过度失水, 从而提高耐盐性(Cho et al., 2006)。与此类似, 在长期的盐胁迫下, 耐盐植物胡杨(Populus euphratica)叶子发育成肉质叶从而抵御逆境。过表达胡杨PeXTH的烟草(Nicotiana tabacum)耐盐性增强, 叶片肉质化但不膨胀(Han et al., 2013)。相反, 在盐胁迫下, 拟南芥AtXTH30表达量大幅上升, 敲除该基因后其耐盐性增强, 而过表达该基因增加其盐敏感性。盐胁迫引起拟南芥xth30突变体中XGO含量的上升较野生型少, 根部Na+积累量和H2O2含量减少。上述结果表明, XTH30通过调控XyG的侧链合成、改变XLFG的含量和纤维素合成以及皮层维管稳定性负调控植物的耐盐性(Yan et al., 2019)。导致相反结果的可能原因是XTHs的双重酶活性(Rose et al., 2002)。
3.1.4 铝胁迫
研究表明, 铝毒是由于Al3+与植物的细胞壁结合从而影响根系的伸长, 而XyG被认为是Al主要的结合位点(Zhu et al., 2012)。拟南芥XyG的乙酰基转移酶基因TBL27缺失的突变体比野生型植株对Al更敏感, 细胞壁半纤维素中Al积累更多。此外, Al处理可显著降低XyG的O-乙酰化水平。研究表明, XyG通过调节O-乙酰化水平影响半纤维素中的Al结合能力, 进而影响拟南芥对Al的敏感性(Zhu et al., 2017)。
3.1.5 多重逆境
近年来, 越来越多的研究表明, XyG代谢相关基因能够响应多种逆境或增强植物对多种逆境的抗性。例如, PeXTH基因同时具有提高胡杨耐盐及重金属的功能(Han et al., 2013, 2014)。水稻OsXET9能够被冷、热和干旱等多种逆境高度诱导表达, 是一个逆境响应标记基因(Dong et al., 2011)。类似地, 辣椒CaXTH3也受多种非生物逆境诱导表达, 过表达该基因的拟南芥和番茄表现出对盐和干旱胁迫的耐受性增强(Cho et al., 2006; Choi et al., 2011)。NtEXGT是烟草中1个编码XTHs的基因, 该基因不仅受生长素等多种植物生长调节剂诱导, 还在盐、干旱、冷和重金属(如镉)胁迫下上调表达, 但是在低温(0°C)和较高浓度(100 μmol·L-1) ABA处理后表达量却下降, 这表明NtEXGT受ABA调控, 在逆境条件下依赖于ABA信号途径。过表达NtEXGT烟草植株具有更强的耐寒性和耐热性(Kuluev et al., 2017)。Iurlaro等(2016)系统研究了硬粒小麦(Triticum durum) XTH基因家族对脱水和热胁迫的响应, 发现两种逆境对TdXTHs表达量的影响因品种的抗逆能力、器官及根的不同区域而异, 脱水时根尖TdXTHs表达量及XEG酶活性升高而热处理的效果相反。烟草中的12个NtXTHs对盐、碱和冷、热胁迫响应不同(Wang et al., 2018)。上述结果表明, XyG代谢相关基因对逆境的响应可能具有物种、器官及逆境特异性。
3.2 生物胁迫
3.2.1 病害降解植物XyG是病原菌成功侵染寄主的必要条件(Choi et al., 2013)。在侵染过程中, 病原菌通过产生各种酶来降解植物细胞壁XyG (DeBoy et al., 2008)。为了应对病原菌的侵染, 植物采取多种措施进行防卫。例如, 表达相应的抑制蛋白抵御病原菌, 植物XyG的降解产物也能诱导抗病性。
3.2.1.1 XyG降解相关基因
质外体组织是植物抵御病原菌的关键战场。病原菌分泌质外体XEGs降解植物XyG的主链, 相应地, 植物通过诱导表达其抑制蛋白(xyloglucan-speci?c endo- 1,4-glucanase inhibitor protein, XEGIP)的方式进行抵抗。这在苹果(Malus x domestica)和大豆等多种植物中得到证实。植物XEGIP通过与病原菌XEG结合从而使其失活, 进而抑制病原菌对植物细胞壁XyG的降解(Bai et al., 2015; Ma et al., 2017)。此外, XEGIP还可通过过敏性坏死(Jones et al., 2006)和诱导抗性(Choi et al., 2013)等途径增强植物的抗性。另外, 植物XEGs也可能与感病性有关(Karczmarek et al., 2008)。Li等(2019)发现在接种柑橘溃疡病病原菌(Xanthomonas citri subsp. citri, Xcc)后, CsXTH04/ 21在感病品种中上调表达而抗病品种正好相反, 并受外源水杨酸和茉莉酸甲酯诱导, 且过表达CsXTH04的甜橙(Citrus sinensis)表现感病, 沉默该基因则使甜橙抗病, 表明该基因可能与柑橘对溃疡病的感病性相关。相反, Sharmin等(2012)发现在遭受茎/芽腐病病原菌(Macrophomina phaseolina)侵染后, 抗病黄麻(Corchorus capsularis)野生种CtXTH1基因上调表达而感病栽培种CoXTH1下调表达, 表明黄麻XTHs表达量可能与其抗病性呈正相关。类似地, MaXTH23基因在接种香蕉枯萎病菌(Fusarium oxysporum f. sp. cubense)前后在抗病品种中的表达量均显著高于感病品种, 表明该基因可能与香蕉对枯萎病的抗性有关(Niu et al., 2018)。马铃薯(Solanum tuberosum) XTH-Xet5在诱导抗病品种对马铃薯Y病毒(PVY)的过敏性坏死反应过程中起联接不同多糖的作用, 可通过增强细胞壁强度来提高其抗病性(Otulak- Kozie? et al., 2018)。
3.2.1.2 XyG乙酰化
有研究表明, XyG乙酰化程度与植物的抗逆性相关。例如, 将真菌的鼠李糖半乳糖醛酸乙酰酯酶转入拟南芥后, 植株XyG的乙酰化程度显著降低, 转基因植株对灰葡萄孢菌(Botrytis cinerea)等真菌的抗性增强, 但不能增强对病原细菌的抗性(Pogorelko et al., 2013)。拟南芥细胞壁乙酰化程度降低20%的突变体rwa2对灰葡萄孢菌的抗性较野生型高(Manabe et al., 2011)。与野生型相比, rwa2抗氧化系统酶基因等与植物抗性相关的基因上调表达, 表明降低细胞壁乙酰化程度会导致拟南芥广泛的抗性反应(Nafifisi et al., 2015)。多糖的乙酰化修饰可以保护其不受特定的糖基水解酶水解, 且可与控制细胞延展性的细胞壁成分交联, 从而进一步阻止病原菌的入侵(Gille and Pauly, 2012)。
3.2.1.3 XyG及其降解产物
早期人们发现, XyG降解产生的寡糖片段XGO可抑制烟草坏死病毒, 具有诱导抗性的功能(Subíkova et al., 1994)。后来发现用XGO处理烟草BY-2细胞培养物可上调与抗逆相关基因的表达(González-Pérez et al., 2014)。最近, XyG又被证明具有启动葡萄(Vitis vinifera)和拟南芥等植物免疫反应的功能(Claverie et al., 2018), 及诱导葡萄细胞悬浮系产生白藜芦醇和拟南芥积累胼胝质等抗性物质。此外, 研究发现纯化的CaXEGIP1蛋白能显著抑制病原菌XEG酶的水解活性, 从而减轻病害(Choi et al., 2013)。Niu等(2018)发现抗枯萎病香蕉品种根毛中识别XXXG型XyG的LM15抗体荧光信号强于感病品种, 且病原菌侵染仅导致抗病品种信号增强, 表明LM15抗原可能与香蕉对枯萎病的抗性呈正相关。
3.2.2 虫害
目前, XyG代谢与植物抗虫性的关系研究较少。芹菜(Apium graveolens) AgXTH1在桃蚜(Myzus persicae)侵染后系统性地在韧皮部中上调表达, 拟南芥中该基因的同源基因AtXTH33的突变体比野生型更吸引蚜虫, 表明AtXTH33调控拟南芥对蚜虫的抗性(Divol et al., 2007)。Mageroy等(2015)发现, 白云杉(Picea glauca) Pgβglu-1在抗性品种中的表达量比感病品种高约1 000倍。Pgβglu-1表达量、PgβGLU-1酶活性及苯乙酮的含量可作为云杉对食心虫的抗性标记, 而PgβGLU-1酶催化底物后释放的苷元物质对云杉食心虫具有抗性(Delvas et al., 2011)。
4 总结与展望
XyG是双子叶植物初生细胞壁中重要的半纤维素, 根据寡糖的结构可将XyG分成XXXG和XXGG两种主要类型。参与XyG生物合成的主要酶包括GS、XXT、β-GAL、FUT、XUT和TBL, 参与XyG降解的主要酶有XTH、β-GLU、α-木糖苷酶、β-半乳糖苷酶和α-岩藻糖苷酶。虽然近年来有关木葡聚糖代谢方面的研究进展显著, 但有关木葡聚糖的侧链修饰以及参与合成的酶蛋白复合体的组成等仍需进一步探讨。XyG代谢相关基因在植物抵抗各种逆境过程中具有重要作用, 尤其是XyG降解相关基因。关于XyG在植物抗逆过程中的作用机理, 目前普遍认为XyG代谢相关基因通过改变细胞壁的组成及XyG之间或XyG与其它细胞壁成分之间的交联从而改变细胞壁的弹性或硬度, 进而影响植物对非生物胁迫的抗性。也有证据表明, XyG和XGO可作为植物信号分子激发植物的防卫反应, 还可能与其它信号分子协同作用, 共同应对逆境胁迫。随着遗传学、生物化学和分子生物学等多种技术的发展及先进仪器设备的应用, 人们将最终解析XyG在植物抗逆过程中的功能及作用机理。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:29266460 [本文引用: 1]
[本文引用: 1]
URLPMID:18544630 [本文引用: 1]
URLPMID:16684525 [本文引用: 2]
URLPMID:23093361 [本文引用: 3]
DOI:10.1007/s00299-010-0989-3URLPMID:21207033 [本文引用: 1]

The hot pepper xyloglucan endo-trans-gluco-sylase/hydrolase (CaXTH3) gene that was inducible by a broad spectrum of abiotic stresses in hot pepper has been reported to enhance tolerance to drought and high salinity in transgenic Arabidopsis. To assess whether CaXTH3 is a practically useful target gene for improving the stress tolerance of crop plants, we ectopically over-expressed the full-length CaXTH3 cDNA in tomato (Solanum lycopersicum cv. Dotaerang) and found that the 35S:CaXTH3 transgenic tomato plants exhibited a markedly increased tolerance to salt and drought stresses. Transgenic tomato plants exposed to a salt stress of 100 mM NaCl retained the chlorophyll in their leaves and showed normal root elongation. They also remained green and unwithered following exposure to 2 weeks of dehydration. A high proportion of stomatal closures in 35S:CaXTH3 was likely to be conferred by increased cell-wall remodeling activity of CaXTH3 in guard cell, which may reduce transpirational water loss in response to dehydration stress. Despite this increased stress tolerance, the transgenic tomato plants showed no detectable phenotype defects, such as abnormal morphology and growth retardation, under normal growth conditions. These results raise the possibility that CaXTH3 gene is appropriate for application in genetic engineering strategies aimed at improving abiotic stress tolerance in agriculturally and economically valuable crop plants.
[本文引用: 1]
DOI:10.1073/pnas.0703133104URLPMID:17488821 [本文引用: 1]
Despite the central role of xyloglucan (XyG) in plant cell wall structure and function, important details of its biosynthesis are not understood. To identify the gene(s) responsible for synthesizing the beta-1,4 glucan backbone of XyG, we exploited a property of nasturtium (Tropaeolum majus) seed development. During the last stages of nasturtium seed maturation, a large amount of XyG is deposited as a reserve polysaccharide. A cDNA library was produced from mRNA isolated during the deposition of XyG, and partial sequences of 10,000 cDNA clones were determined. A single member of the C subfamily from the large family of cellulose synthase-like (CSL) genes was found to be overrepresented in the cDNA library. Heterologous expression of this gene in the yeast Pichia pastoris resulted in the production of a beta-1,4 glucan, confirming that the CSLC protein has glucan synthase activity. The Arabidopsis CSLC4 gene, which is the gene with the highest sequence similarity to the nasturtium CSL gene, is coordinately expressed with other genes involved in XyG biosynthesis. These and other observations provide a compelling case that the CSLC gene family encode proteins that synthesize the XyG backbone.
URLPMID:18556790 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:17238910 [本文引用: 1]
[本文引用: 1]
DOI:10.1126/scisignal.aao3070URLPMID:29945884 [本文引用: 1]
Cell walls surround all plant cells, and their composition and structure are modified in a tightly controlled, adaptive manner to meet sometimes opposing functional requirements during growth and development. The plant cell wall integrity (CWI) maintenance mechanism controls these functional modifications, as well as responses to cell wall damage (CWD). We investigated how the CWI system mediates responses to CWD in Arabidopsis thaliana CWD induced by cell wall-degrading enzymes or an inhibitor of cellulose biosynthesis elicited similar, turgor-sensitive stress responses. Phenotypic clustering with 27 genotypes identified a core group of receptor-like kinases (RLKs) and ion channels required for the activation of CWD responses. A genetic analysis showed that the RLK FEI2 and the plasma membrane-localized mechanosensitive Ca(2+) channel MCA1 functioned downstream of the RLK THE1 in CWD perception. In contrast, pattern-triggered immunity (PTI) signaling components, including the receptors for plant elicitor peptides (AtPeps) PEPR1 and PEPR2, repressed responses to CWD. CWD induced the expression of PROPEP1 and PROPEP3, which encode the precursors of AtPep1 and AtPep3, and the release of PROPEP3 into the growth medium. Application of AtPep1 and AtPep3 repressed CWD-induced phytohormone accumulation in a concentration-dependent manner. These results suggest that AtPep-mediated signaling suppresses CWD-induced defense responses controlled by the CWI mechanism. This suppression was alleviated when PTI signaling downstream of PEPR1 and PEPR2 was impaired. Defense responses controlled by the CWI maintenance mechanism might thus compensate to some extent for the loss of PTI signaling elements.
URLPMID:12032363 [本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 3]
DOI:10.3389/fpls.2012.00012URLPMID:22639638 [本文引用: 1]
Plant cell walls are composed of structurally diverse polymers, many of which are O-acetylated. How plants O-acetylate wall polymers and what its function is remained elusive until recently, when two protein families were identified in the model plant Arabidopsis that are involved in the O-acetylation of wall polysaccharides - the reduced wall acetylation (RWA) and the trichome birefringence-like (TBL) proteins. This review discusses the role of these two protein families in polysaccharide O-acetylation and outlines the differences and similarities of polymer acetylation mechanisms in plants, fungi, bacteria, and mammals. Members of the TBL protein family had been shown to impact pathogen resistance, freezing tolerance, and cellulose biosynthesis. The connection of TBLs to polysaccharide O-acetylation thus gives crucial leads into the biological function of wall polymer O-acetylation. From a biotechnological point understanding the O-acetylation mechanism is important as acetyl-substituents inhibit the enzymatic degradation of wall polymers and released acetate can be a potent inhibitor in microbial fermentations, thus impacting the economic viability of, e.g., lignocellulosic based biofuel production.
DOI:10.1007/s11033-014-3566-yURLPMID:25008996 [本文引用: 1]
Xyloglucan oligosaccharides (XGOs) are breakdown products of XGs, the most abundant hemicelluloses of the primary cell walls of non-Poalean species. Treatment of cell cultures or whole plants with XGOs results in accelerated cell elongation and cell division, changes in primary root growth, and a stimulation of defence responses. They may therefore act as signalling molecules regulating plant growth and development. Previous work suggests an interaction with auxins and effects on cell wall loosening, however their mode of action is not fully understood. The effect of an XGO extract from tamarind (Tamarindus indica) on global gene expression was therefore investigated in tobacco BY-2 cells using microarrays. Over 500 genes were differentially regulated with similar numbers and functional classes of genes up- and down-regulated, indicating a complex interaction with the cellular machinery. Up-regulation of a putative XG endotransglycosylase/hydrolase-related (XTH) gene supports the mechanism of XGO action through cell wall loosening. Differential expression of defence-related genes supports a role for XGOs as elicitors. Changes in the expression of genes related to mitotic control and differentiation also support previous work showing that XGOs are mitotic inducers. XGOs also affected expression of several receptor-like kinase genes and transcription factors. Hence, XGOs have significant effects on expression of genes related to cell wall metabolism, signalling, stress responses, cell division and transcriptional control.
DOI:10.1105/tpc.111.089193URLPMID:22080600 [本文引用: 1]
An Arabidopsis thaliana mutant with an altered structure of its hemicellulose xyloglucan (XyG; axy-8) identified by a forward genetic screen facilitating oligosaccharide mass profiling was characterized. axy8 exhibits increased XyG fucosylation and the occurrence of XyG fragments not present in the wild-type plant. AXY8 was identified to encode an alpha-fucosidase acting on XyG that was previously designated FUC95A. Green fluorescent protein fusion localization studies and analysis of nascent XyG in microsomal preparations demonstrated that this glycosylhydrolase acts mainly on XyG in the apoplast. Detailed structural analysis of XyG in axy8 gave unique insights into the role of the fucosidase in XyG metabolism in vivo. The genetic evidence indicates that the activity of glycosylhydrolases in the apoplast plays a major role in generating the heterogeneity of XyG side chains in the wall. Furthermore, without the dominant apoplastic glycosylhydrolases, the XyG structure in the wall is mainly composed of XXXG and XXFG subunits.
DOI:10.1016/j.envexpbot.2013.12.021URL [本文引用: 1]
Cadmium (Cd2+) is a toxic heavy metal impairing plant growth and development. Xyloglucan endotransglucosylase/hydrolase gene (XTH) is involved in the plant response to heavy metal toxicity, in addition to controlling cell wall extensibility. However, the link between XTH and Cd2+ stress has not yet been established in higher plants. PeXTH expression was up-regulated by 1.2-2.1-fold in Populus euphratica roots and leaves upon Cd2+ exposure (40-80 mu M CdCl2). Cellular Cd2+ analysis and flux data showed that the cadmium-elicited expression of PeXTH markedly restricted Cd2+ uptake and accumulation in P. euphratica roots. Moreover, tobacco plants overexpressing PeXTH were more tolerant to Cd2+ stress (80 mu M CdCl2) than wild-type tobacco in terms of root and shoot growth. Transgenic lines accumulated 49-58% less Cd2+ in root apical and mature regions, as compared to the wild type. The less buildup of Cd2+ in roots of transgenic lines was the result of lower influx of Cd2+ under Cd2+ stress. It is noting that transgenic plants displayed 56-87% higher xyloglucan degradation activity (XDA) than the wild type, leading to a 25-27% decline of xyloglucan content in the root cell walls. Therefore, overexpression of PeXTH increased the activity of XDA in transgenic plants, which enhanced the degradation of xyloglucan in the wall. The down-regulated amount of xyloglucan led to less binding sites for Cd2+ and thus reduced the root Cd2+ uptake and buildup in transgenic plants. Consequently, the Cd2+ toxicity was eventually alleviated in transgenic tobacco. (C) 2014 Elsevier B.V.
URLPMID:24085577 [本文引用: 2]
DOI:10.1104/pp.83.2.384URLPMID:16665254 [本文引用: 1]
Since xyloglucan is believed to bind to cellulose microfibrils in the primary cell walls of higher plants and, when isolated from the walls, can also bind to cellulose in vitro, the binding mechanism of xyloglucan to cellulose was further investigated using radioiodinated pea xyloglucan. A time course for the binding showed that the radioiodinated xyloglucan continued to be bound for at least 4 hours at 40 degrees C. Binding was inhibited above pH 6. Binding capacity was shown to vary for celluloses of different origin and was directly related to the relative surface area of the microfibrils. The binding of xyloglucan to cellulose was very specific and was not affected by the presence of a 10-fold excess of (1-->2)-beta-glucan, (1-->3)-beta-glucan, (1-->6)-beta-glucan, (1-->3, 1-->4)-beta-glucan, arabinogalactan, or pectin. When xyloglucan (0.1%) was added to a cellulose-forming culture of Acetobacter xylinum, cellulose ribbon structure was partially disrupted indicating an association of xyloglucan with cellulose at the time of synthesis. Such a result suggests that the small size of primary wall microfibrils in higher plants may well be due to the binding of xyloglucan to cellulose during synthesis which prevents fasciation of small fibrils into larger bundles. Fluorescent xyloglucan was used to stain pea cell wall ghosts prepared to contain only the native xyloglucan:cellulose network or only cellulose. Ghosts containing only cellulose showed strong fluorescence when prepared before or after elongation; as predicted, the presence of native xyloglucan in the ghosts repressed binding of added fluorescent xyloglucan. Such ghosts, prepared after elongation when the ratio of native xyloglucan:cellulose is substantially reduced, still showed only faint fluorescence, indicating that microfibrils continue to be coated with xyloglucan throughout the growth period.
DOI:10.1104/pp.75.3.605URLPMID:16663673 [本文引用: 1]
Two auxin-induced endo-1,4-beta-glucanases (EC 3.2.1.4) were purified from pea (Pisum sativum L. var. Alaska) epicotyls and used to degrade purified pea xyloglucan. Hydrolysis yielded nonasaccharide (glucose/xylose/galactose/fucose, 4:3:1:1) and heptasaccharide (glucose/xylose, 4:3) as the products. The progress of hydrolysis, as monitored viscometrically (with amyloid xyloglucan) and by determination of residual xyloglucan-iodine complex (pea) confirmed that both pea glucanases acted as endohydrolases versus xyloglucan. K(m) values for amyloid and pea xyloglucans were approximately the same as those for cellulose derivatives, but V(max) values were lower for the xyloglucans. Auxin treatment of epicotyls in vivo resulted in increases in net deposits of xyloglucan and cellulose in spite of a great increase (induction) of endogenous 1,4-beta-glucanase activity. However, the average degree of polymerization of the resulting xyloglucan was much lower than in controls, and the amount of soluble xyloglucan increased. When macromolecular complexes of xyloglucan and cellulose (cell wall ghosts) were treated in vitro with pea 1,4-beta-glucanase, the xyloglucan component was preferentially hydrolyzed and solubilized. It is concluded that xyloglucan is the main cell wall substrate for pea endo-1,4-beta-glucanase in growing tissue.
DOI:10.3389/fpls.2016.00984URLPMID:27559336 [本文引用: 1]
The plant cell wall has a diversity of functions. It provides a structural framework to support plant growth and acts as the first line of defense when the plant encounters pathogens. The cell wall must also retain some flexibility, such that when subjected to developmental, biotic, or abiotic stimuli it can be rapidly remodeled in response. Genes encoding enzymes capable of synthesizing or hydrolyzing components of the plant cell wall show differential expression when subjected to different stresses, suggesting they may facilitate stress tolerance through changes in cell wall composition. In this review we summarize recent genetic and transcriptomic data from the literature supporting a role for specific cell wall-related genes in stress responses, in both dicot and monocot systems. These studies highlight that the molecular signatures of cell wall modification are often complex and dynamic, with multiple genes appearing to respond to a given stimulus. Despite this, comparisons between publically available datasets indicate that in many instances cell wall-related genes respond similarly to different pathogens and abiotic stresses, even across the monocot-dicot boundary. We propose that the emerging picture of cell wall remodeling during stress is one that utilizes a common toolkit of cell wall-related genes, multiple modifications to cell wall structure, and a defined set of stress-responsive transcription factors that regulate them.
DOI:10.1038/nplants.2017.9URLPMID:28211849 [本文引用: 1]
The major disease resistance gene Xa4 confers race-specific durable resistance against Xanthomonas oryzae pv. oryzae, which causes the most damaging bacterial disease in rice worldwide. Although Xa4 has been one of the most widely exploited resistance genes in rice production worldwide, its molecular nature remains unknown. Here we show that Xa4, encoding a cell wall-associated kinase, improves multiple traits of agronomic importance without compromising grain yield by strengthening the cell wall via promoting cellulose synthesis and suppressing cell wall loosening. Strengthening of the cell wall by Xa4 enhances resistance to bacterial infection, and also increases mechanical strength of the culm with slightly reduced plant height, which may improve lodging resistance of the rice plant. The simultaneous improvement of multiple agronomic traits conferred by Xa4 may account for its widespread and lasting utilization in rice breeding programmes globally.
DOI:10.3389/fpls.2016.01686URLPMID:27891140 [本文引用: 1]
Heat and drought stress have emerged as major constraints for durum wheat production. In the Mediterranean area, their negative effect on crop productivity is expected to be exacerbated by the occurring climate change. Xyloglucan endotransglucosylase/hydrolases (XTHs) are chief enzymes in cell wall remodeling, whose relevance in cell expansion and morphogenesis suggests a central role in stress responses. In this work the potential role of XTHs in abiotic stress tolerance was investigated in durum wheat. The separate effects of dehydration and heat exposure on XTH expression and its endotransglucosylase (XET) in vitro activity and in vivo action have been monitored, up to 24 h, in the apical and sub-apical root regions and shoots excised from 3-day-old seedlings of durum wheat cultivars differing in stress susceptibility/tolerance. Dehydration and heat stress differentially influence the XTH expression profiles and the activity and action of XET in the wheat seedlings, depending on the degree of susceptibility/tolerance of the cultivars, the organ, the topological region of the root and, within the root, on the gradient of cell differentiation. The root apical region was the zone mainly affected by both treatments in all assayed cultivars, while no change in XET activity was observed at shoot level, irrespective of susceptibility/tolerance, confirming the pivotal role of the root in stress perception, signaling, and response. Conflicting effects were observed depending on stress type: dehydration evoked an overall increase, at least in the apical region of the root, of XET activity and action, while a significant inhibition was caused by heat treatment in most cultivars. The data suggest that differential changes in XET action in defined portions of the root of young durum wheat seedlings may have a role as a response to drought and heat stress, thus contributing to seedling survival and crop establishment. A thorough understanding of the mechanisms underlying these variations could represent the theoretical basis for implementing breeding strategies to develop new highly productive hybrids adapted to future climate scenarios.
DOI:10.1093/mp/sss032URLPMID:22474179 [本文引用: 1]
A deep-sequencing approach was pursued utilizing 454 and Illumina sequencing methods to discover new genes involved in xyloglucan biosynthesis. cDNA sequences were generated from developing nasturtium (Tropaeolum majus) seeds, which produce large amounts of non-fucosylated xyloglucan as a seed storage polymer. In addition to known xyloglucan biosynthetic genes, a previously uncharacterized putative xyloglucan galactosyltransferase was identified. Analysis of an Arabidopsis thaliana mutant line defective in the corresponding ortholog (AT5G62220) revealed that this gene shows no redundancy with the previously characterized xyloglucan galactosyltransferase, MUR3, but is required for galactosyl-substitution of xyloglucan at a different position. The gene was termed XLT2 for Xyloglucan L-side chain galactosylTransferase position 2. It represents an enzyme in the same subclade of glycosyltransferase family 47 as MUR3. A double mutant defective in both MUR3 (mur3.1) and XLT2 led to an Arabidopsis plant with xyloglucan that consists essentially of only xylosylated glucosyl units, with no further substitutions.
DOI:10.1007/BF02869608URL [本文引用: 1]
Durable resistance to late blight is believed to be dependent on genes other than R genes. Reduced rates of lesion expansion, as found in the germplasm B 0767-2, can provide durable resistance by limiting tissue damage and reducing pathogen populations. We have tested the role of the proteinaceous xyloglucan-specific endoglucanase inhibitor (XEIP) in limiting late blight lesion expansion in B 0767-2. Plant material was silenced for the XEIP using localized infiltration ofAgrobacterium tumefaciens harboring pCAMBIA 1302 containing an antisense copy of XEIP. Plant cell transformations with this construct induced effective silencing as indicated by a reduction of up to 80% in XEIP transcript levels. Silencing of XEIP resulted in a marked increase in lesion size and water soaking, indicating that the inhibitor protein plays a role in limiting lesion expansion in B 0767-2.]]>
DOI:10.1094/MPMI-21-6-0791URLPMID:18624642 [本文引用: 1]
For the proliferation of their feeding sites (syncytia), the potato cyst nematode Globodera rostochiensis is thought to recruit plant endo-beta-1,4-glucanases (EGases, EC. 3.2.1.4). Reverse-transcription polymerase chain reaction experiments on tomato (Solanum lycopersicum) indicated that the expression of two out of the at least eight EGases, namely Sl-cel7 and Sl-cel9C1, is specifically upregulated during syncytium formation. In situ hybridization and immunodetection studies demonstrated that both EGases are specifically expressed inside and adjacent to proliferating syncytia. To assess the importance of Sl-cel7 and Sl-cel9C1 for nematode development, we decided to knock them out individually. Sl-cel9C1 probably is the only class C EGase in tomato, and we were unable to regenerate Sl-cel9C1-silenced plants. Potato (S. tuberosum), a close relative of tomato, harbors at least two class C EGases, and St-cel7-or St-cel9C1-silenced potato plants showed no obvious aberrant phenotype. Infection with potato cyst nematodes resulted in a severe reduction of the number of adult females (up to 60%) and a sharp increase in the fraction of females without eggs (up to 89%). Hence, the recruitment of CEL7, an enzyme that uses xyloglucan and noncrystalline cellulose as natural substrates, and CEL9C1, an enzyme that uses crystalline cellulose, is essential for growth and development of potato cyst nematodes.
[本文引用: 1]
[本文引用: 2]
DOI:10.1104/pp.114.255943URLPMID:25673778 [本文引用: 1]
Xyloglucan is a polysaccharide that has important roles in the formation and function of the walls that surround growing land plant cells. Many of these plants synthesize xyloglucan that contains galactose in two different side chains (L and F), which exist in distinct molecular environments. However, little is known about the contribution of these side chains to xyloglucan function. Here, we show that Arabidopsis (Arabidopsis thaliana) mutants devoid of the F side chain galactosyltransferase MURUS3 (MUR3) form xyloglucan that lacks F side chains and contains much less galactosylated xylose than its wild-type counterpart. The galactose-depleted xyloglucan is dysfunctional, as it leads to mutants that are dwarfed with curled rosette leaves, short petioles, and short inflorescence stems. Moreover, cell wall matrix polysaccharides, including xyloglucan and pectin, are not properly secreted and instead accumulate within intracellular aggregates. Near-normal growth is restored by generating mur3 mutants that produce no detectable amounts of xyloglucan. Thus, cellular processes are affected more by the presence of the dysfunctional xyloglucan than by eliminating xyloglucan altogether. To identify structural features responsible for xyloglucan dysfunction, xyloglucan structure was modified in situ by generating mur3 mutants that lack specific xyloglucan xylosyltransferases (XXTs) or that overexpress the XYLOGLUCAN L-SIDE CHAIN GALACTOSYLTRANSFERASE2 (XLT2) gene. Normal growth was restored in the mur3-3 mutant overexpressing XLT2 and in mur3-3 xxt double mutants when the dysfunctional xyloglucan was modified by doubling the amounts of galactosylated side chains. Our study assigns a role for galactosylation in normal xyloglucan function and demonstrates that altering xyloglucan side chain structure disturbs diverse cellular and physiological processes.
DOI:10.1016/j.plaphy.2016.12.005URLPMID:27940271 [本文引用: 1]
Despite the intensive study of xyloglucan endotransglucosylases/hydrolases, their multifaceted role in plant growth regulation in changing environmental conditions is not yet clarified. The functional role of the large number of genes encoding this group of enzymes is also still unclear. NtEXGT gene encodes one of xyloglucan endotransglucosylases/hydrolases (XTHs) of Nicotiana tabacum L. The highest level of NtEXGT gene expression was detected in young flowers and leaves near the shoot apex. Expression of the NtEXGT gene in leaves was induced by cytokinins, auxins, brassinosteroids and gibberellins. NtEXGT gene was also up-regulated by salinity, drought, cold, cadmium and 10 muM abscisic acid treatments and down-regulated in response to 0 degrees C and 100 muM abscisic acid. Pretreatment of leaves with fluridone contributed to smaller increase in the level of NtEXGT transcripts in response to drought stress. These data suggest that NtEXGT gene is ABA-regulated and probably implicated in ABA-dependent signaling in response to stress factors. 35S::NtEXGT plants of tobacco showed higher rate of root growth under salt-stress conditions, greater frost and heat tolerance as compared with the wild type tobacco plants.
[本文引用: 1]
DOI:10.3389/fpls.2019.01109URLPMID:31611887 [本文引用: 1]
In this study, we performed a comprehensive survey of xyloglucan endotransglucosylase/hydrolase (XTH) and a functional validation of Citrus sinensis (Cs) XTH genes to provide new insights into the involvement of XTHs in Xanthomonas citri subsp. citri (Xcc) infection. From the genome of sweet orange, 34 CsXTH genes with XTH characteristic domains were identified and clustered into groups I/II, IIIA, and IIIB. Except for chromosome 9, the CsXTH genes were unevenly distributed and duplicated among all chromosomes, identifying a CsXTH duplication hot spot on chromosome 4. With Xcc induction, a group of citrus canker-related CsXTHs were detected. CsXTH04 was identified as a putative candidate gene, which is up-regulated in citrus bacterial canker (CBC)-resistant varieties and induced by exogenous treatment with salicylic acid (SA) and methyl jasmonate (MeJA). CsXTH04 overexpression conferred CBC susceptibility to transgenic citrus, while CsXTH04 silencing conferred CBC resistance. Taken together, the annotation of the CsXTH family provides an initial basis for the functional and evolutionary study of this family as potential CBC-susceptible genes. CsXTH04, validated in this study, can be used in citrus breeding to improve CBC resistance.
DOI:10.1093/jxb/ert321URLPMID:24127514 [本文引用: 1]
Arabinogalactan-proteins (AGPs) are highly glycosylated hydroxyproline-rich glycoproteins present in plant cell walls. AGPs are characterized by arabinose-/galactose-rich side chains, which define their interactive molecular surface. Fucose residues are found in some dicotyledon AGPs, and AGP fucosylation is developmentally regulated. We previously identified Arabidopsis thaliana FUT4 and FUT6 genes as AGP-specific fucosyltransferases (FUTs) based on their enzymatic activities when heterologously expressed in tobacco (Nicotiana tabacum) BY2 suspension-cultured cells. Here, the functions of FUT4 and FUT6 and the physiological roles of fucosylated AGPs were further investigated using Arabidopsis fut4, fut6, and fut4/fut6 mutant plants. All mutant plants showed no phenotypic differences compared to wild-type plants under physiological conditions, but showed reduced root growth in the presence of elevated NaCl. However, roots of wild-type and fut4 mutant plants contained terminal fucose epitopes, which were absent in fut6 and fut4/fut6 mutant plants as indicated by eel lectin staining. Monosaccharide analysis showed fucose was present in wild-type leaf and root AGPs, but absent in fut4 leaf AGPs and in fut4/fut6 double mutant leaf and root AGPs, indicating that FUT4 was required for fucosylation of leaf AGPs while both FUT4 and FUT6 contributed to fucosylation of root AGPs. Glycome profiling of cell wall fractions from mutant roots and leaves showed distinct glycome profiles compared to wild-type plants, indicating that fucosyl residues on AGPs may regulate intermolecular interactions between AGPs and other wall components. The current work exemplifies the possibilities of refinement of cell wall structures by manipulation of a single or a few cell wall biosynthetic genes.
DOI:10.1016/j.molp.2015.11.004URLPMID:26589447 [本文引用: 1]
DOI:10.1104/pp.15.00441URLPMID:25869654 [本文引用: 1]
The predominant structure of the hemicellulose xyloglucan (XyG) found in the cell walls of dicots is a fucogalactoXyG with an XXXG core motif, whereas in the Poaceae (grasses and cereals), the structure of XyG is less xylosylated (XXGGn core motif) and lacks fucosyl residues. However, specialized tissues of rice (Oryza sativa) also contain fucogalactoXyG. Orthologous genes of the fucogalactoXyG biosynthetic machinery of Arabidopsis (Arabidopsis thaliana) are present in the rice genome. Expression of these rice genes, including fucosyl-, galactosyl-, and acetyltransferases, in the corresponding Arabidopsis mutants confirmed their activity and substrate specificity, indicating that plants in the Poaceae family have the ability to synthesize fucogalactoXyG in vivo. The data presented here provide support for a functional conservation of XyG structure in higher plants.
DOI:10.1093/jxb/ert088URLPMID:23580752 [本文引用: 1]
Recent studies suggest that plant pectin methylesterases (PMEs) are directly involved in plant defence besides their roles in plant development. However, the molecular mechanisms of PME action on pectins are not well understood. In order to understand how PMEs modify pectins during banana (Musa spp.)-Fusarium interaction, the expression and enzyme activities of PMEs in two banana cultivars, highly resistant or susceptible to Fusarium, were compared with each other. Furthermore, the spatial distribution of PMEs and their effect on pectin methylesterification of 10 individual homogalacturonan (HG) epitopes with different degrees of methylesterification (DMs) were also examined. The results showed that, before pathogen treatment, the resistant cultivar displayed higher PME activity than the susceptible cultivar, corresponding well to the lower level of pectin DM. A significant increase in PME expression and activity and a decrease in pectin DM were observed in the susceptible cultivar but not in the resistant cultivar when plants were wounded, which was necessary for successful infection. With the increase of PME in the wounded susceptible cultivar, the JIM5 antigen (low methyestrified HGs) increased. Forty-eight hours after pathogen infection, the PME activity and expression in the susceptible cultivar were higher than those in the resistant cultivar, while the DM was lower. In conclusion, the resistant and the susceptible cultivars differ significantly in their response to wounding. Increased PMEs and thereafter decreased DMs acompanied by increased low methylesterified HGs in the root vascular cylinder appear to play a key role in determination of banana susceptibility to Fusarium.
DOI:10.1126/science.aai7919URLPMID:28082413 [本文引用: 1]
The extracellular space (apoplast) of plant tissue represents a critical battleground between plants and attacking microbes. Here we show that a pathogen-secreted apoplastic xyloglucan-specific endoglucanase, PsXEG1, is a focus of this struggle in the Phytophthora sojae-soybean interaction. We show that soybean produces an apoplastic glucanase inhibitor protein, GmGIP1, that binds to PsXEG1 to block its contribution to virulence. P. sojae, however, secretes a paralogous PsXEG1-like protein, PsXLP1, that has lost enzyme activity but binds to GmGIP1 more tightly than does PsXEG1, thus freeing PsXEG1 to support P. sojae infection. The gene pair encoding PsXEG1 and PsXLP1 is conserved in many Phytophthora species, and the P. parasitica orthologs PpXEG1 and PpXLP1 have similar functions. Thus, this apoplastic decoy strategy may be widely used in Phytophthora pathosystems.
DOI:10.1111/tpj.12699URLPMID:25302566 [本文引用: 1]
Periodic outbreaks of spruce budworm (SBW) affect large areas of ecologically and economically important conifer forests in North America, causing tree mortality and reduced forest productivity. Host resistance against SBW has been linked to growth phenology and the chemical composition of foliage, but the underlying molecular mechanisms and population variation are largely unknown. Using a genomics approach, we discovered a beta-glucosidase gene, Pgbetaglu-1, whose expression levels and function underpin natural resistance to SBW in mature white spruce (Picea glauca) trees. In phenotypically resistant trees, Pgbetaglu-1 transcripts were up to 1000 times more abundant than in non-resistant trees and were highly enriched in foliage. The encoded PgbetaGLU-1 enzyme catalysed the cleavage of acetophenone sugar conjugates to release the aglycons piceol and pungenol. These aglycons were previously shown to be active against SBW. Levels of Pgbetaglu-1 transcripts and biologically active acetophenone aglycons were substantially different between resistant and non-resistant trees over time, were positively correlated with each other and were highly variable in a natural white spruce population. These results suggest that expression of Pgbetaglu-1 and accumulation of acetophenone aglycons is a constitutive defence mechanism in white spruce. The progeny of resistant trees had higher Pgbetaglu-1 gene expression than non-resistant progeny, indicating that the trait is heritable. With reported increases in the intensity of SBW outbreaks, influenced by climate, variation of Pgbetaglu-1 transcript expression, PgbetaGLU-1 enzyme activity and acetophenone accumulation may serve as resistance markers to better predict impacts of SBW in both managed and wild spruce populations.
DOI:10.1104/pp.110.168989URLPMID:21212300 [本文引用: 2]
Nearly all polysaccharides in plant cell walls are O-acetylated, including the various pectic polysaccharides and the hemicelluloses xylan, mannan, and xyloglucan. However, the enzymes involved in the polysaccharide acetylation have not been identified. While the role of polysaccharide acetylation in vivo is unclear, it is known to reduce biofuel yield from lignocellulosic biomass by the inhibition of microorganisms used for fermentation. We have analyzed four Arabidopsis (Arabidopsis thaliana) homologs of the protein Cas1p known to be involved in polysaccharide O-acetylation in Cryptococcus neoformans. Loss-of-function mutants in one of the genes, designated REDUCED WALL ACETYLATION2 (RWA2), had decreased levels of acetylated cell wall polymers. Cell wall material isolated from mutant leaves and treated with alkali released about 20% lower amounts of acetic acid when compared with the wild type. The same level of acetate deficiency was found in several pectic polymers and in xyloglucan. Thus, the rwa2 mutations affect different polymers to the same extent. There were no obvious morphological or growth differences observed between the wild type and rwa2 mutants. However, both alleles of rwa2 displayed increased tolerance toward the necrotrophic fungal pathogen Botrytis cinerea.
DOI:10.1007/s00425-015-2243-2URLPMID:25604050 [本文引用: 1]
MAIN CONCLUSION: Putative XyG xylosyltransferases from Tropaeolum majus (nasturtium) and Solanum lycopersicum (tomato) homologous to characterized Arabidopsis genes were identified and shown to functionally complement Arabidopsis mutants lacking xyloglucan demonstrating they represent xyloglucan xylosyltransferases. Xyloglucan is a major hemicellulose in the plant cell wall and is important for the structural organization of the wall. The fine structure of xyloglucan can vary dependent on plant species and tissue type. Most vascular seed-bearing plants including Arabidopsis thaliana and nasturtium (Tropaeolum majus) have a xyloglucan structure, in which three out of four backbone glucosyl-residues are substituted with xylosyl-residues. In contrast, the xyloglucan found in plants of the Solanaceae family, which includes tomato (Solanum lycopersicum), is typically less xylosylated with only two of the four backbone glucosyl-residues substituted with xylosyl-residues. To investigate the genetics of xyloglucan xylosylation, candidate xyloglucan xylosyltransferase genes (XXTs) homologous to known A. thaliana XXTs were cloned from nasturtium and tomato. These candidate XXTs were expressed in the A. thaliana xxt1/2 double and xxt1/2/5 triple mutant, whose walls lack detectable xyloglucan. Expression of the orthologs of XXT5 resulted in no detectable xyloglucan in the transgenic A. thaliana plants, consistent with a lack of xyloglucan in the A. thaliana xxt1/2 double mutant. However, transformation of both the tomato and nasturtium orthologs of AtXXT1 and AtXXT2 resulted in the production of xyloglucan with a xylosylation pattern similar to wild type A. thaliana indicating that both SlXXT2 and TmXXT2 likely have xylosyltransferase activity. As the expression of the SlXXT2 did not result in xyloglucan with a decreased xylosylation frequency found in tomato, this gene is not responsible for the unique xylosylation pattern found in the solanaceous plants.
DOI:10.3389/fpls.2015.00550URLPMID:26257757 [本文引用: 1]
The epidermis on leaves protects plants from pathogen invasion and provides a waterproof barrier. It consists of a layer of cells that is surrounded by thick cell walls, which are partially impregnated by highly hydrophobic cuticular components. We show that the Arabidopsis T-DNA insertion mutants of REDUCED WALL ACETYLATION 2 (rwa2), previously identified as having reduced O-acetylation of both pectins and hemicelluloses, exhibit pleiotrophic phenotype on the leaf surface. The cuticle layer appeared diffused and was significantly thicker and underneath cell wall layer was interspersed with electron-dense deposits. A large number of trichomes were collapsed and surface permeability of the leaves was enhanced in rwa2 as compared to the wild type. A massive reprogramming of the transcriptome was observed in rwa2 as compared to the wild type, including a coordinated up-regulation of genes involved in responses to abiotic stress, particularly detoxification of reactive oxygen species and defense against microbial pathogens (e.g., lipid transfer proteins, peroxidases). In accordance, peroxidase activities were found to be elevated in rwa2 as compared to the wild type. These results indicate that cell wall acetylation is essential for maintaining the structural integrity of leaf epidermis, and that reduction of cell wall acetylation leads to global stress responses in Arabidopsis.
DOI:10.3390/ijms19020350URL [本文引用: 3]
DOI:10.3390/ijms19082287URL [本文引用: 1]
DOI:10.1146/annurev-arplant-043015-112222URLPMID:26927904 [本文引用: 3]
Xyloglucan (XyG) is a matrix polysaccharide that is present in the cell walls of all land plants. It consists of a beta-1,4-linked glucan backbone that is further substituted with xylosyl residues. These xylosyl residues can be further substituted with other glycosyl and nonglycosyl substituents that vary depending on the plant family and specific tissue. Advances in plant mutant isolation and characterization, functional genomics, and DNA sequencing have led to the identification of nearly all transferases and synthases necessary to synthesize XyG. Thus, in terms of the molecular mechanisms of plant cell wall polysaccharide biosynthesis, XyG is the most well understood. However, much remains to be learned about the molecular mechanisms of polysaccharide assembly and the regulation of these processes. Knowledge of the XyG biosynthetic machinery allows the XyG structure to be tailored in planta to ascertain the functions of this polysaccharide and its substituents in plant growth and interactions with the environment.
[本文引用: 1]
DOI:10.1111/nph.14489URLPMID:28257170 [本文引用: 1]
High acetylation of angiosperm wood hinders its conversion to sugars by glycoside hydrolases, subsequent ethanol fermentation and (hence) its use for biofuel production. We studied the REDUCED WALL ACETYLATION (RWA) gene family of the hardwood model Populus to evaluate its potential for improving saccharification. The family has two clades, AB and CD, containing two genes each. All four genes are expressed in developing wood but only RWA-A and -B are activated by master switches of the secondary cell wall PtNST1 and PtMYB21. Histochemical analysis of promoter::GUS lines in hybrid aspen (Populus tremula x tremuloides) showed activation of RWA-A and -B promoters in the secondary wall formation zone, while RWA-C and -D promoter activity was diffuse. Ectopic downregulation of either clade reduced wood xylan and xyloglucan acetylation. Suppressing both clades simultaneously using the wood-specific promoter reduced wood acetylation by 25% and decreased acetylation at position 2 of Xylp in the dimethyl sulfoxide-extracted xylan. This did not affect plant growth but decreased xylose and increased glucose contents in the noncellulosic monosaccharide fraction, and increased glucose and xylose yields of wood enzymatic hydrolysis without pretreatment. Both RWA clades regulate wood xylan acetylation in aspen and are promising targets to improve wood saccharification.
DOI:10.1105/tpc.112.103390URLPMID:23175743 [本文引用: 1]
Root hairs provide a model system to study plant cell growth, yet little is known about the polysaccharide compositions of their walls or the role of these polysaccharides in wall expansion. We report that Arabidopsis thaliana root hair walls contain a previously unidentified xyloglucan that is composed of both neutral and galacturonic acid-containing subunits, the latter containing the beta-D-galactosyluronic acid-(1 -> 2)-alpha-D-xylosyl-(1 -> and/or alpha-L-fucosyl-(1 -> 2)-beta-D-galactosyluronic acid-(1 -> 2)-alpha-D-xylosyl-(1 ->) side chains. Arabidopsis mutants lacking root hairs have no acidic xyloglucan. A loss-of-function mutation in At1g63450, a root hair-specific gene encoding a family GT47 glycosyltransferase, results in the synthesis of xyloglucan that lacks galacturonic acid. The root hairs of this mutant are shorter than those of the wild type. This mutant phenotype and the absence of galacturonic acid in the root xyloglucan are complemented by At1g63450. The leaf and stem cell walls of wild-type Arabidopsis contain no acidic xyloglucan. However, overexpression of At1g63450 led to the synthesis of galacturonic acid-containing xyloglucan in these tissues. We propose that At1g63450 encodes XYLOGLUCAN-SPECIFIC GALACTURONOSYLTRANSFERASE1, which catalyzes the formation of the galactosyluronic acid-(1 -> 2)-alpha-D-xylopyranosyl linkage and that the acidic xyloglucan is present only in root hair cell walls. The role of the acidic xyloglucan in root hair tip growth is discussed.
DOI:10.1104/pp.113.214460URLPMID:23463782 [本文引用: 1]
The plant cell wall has many significant structural and physiological roles, but the contributions of the various components to these roles remain unclear. Modification of cell wall properties can affect key agronomic traits such as disease resistance and plant growth. The plant cell wall is composed of diverse polysaccharides often decorated with methyl, acetyl, and feruloyl groups linked to the sugar subunits. In this study, we examined the effect of perturbing cell wall acetylation by making transgenic Arabidopsis (Arabidopsis thaliana) and Brachypodium (Brachypodium distachyon) plants expressing hemicellulose-and pectin-specific fungal acetylesterases. All transgenic plants carried highly expressed active Aspergillus nidulans acetylesterases localized to the apoplast and had significant reduction of cell wall acetylation compared with wild-type plants. Partial deacetylation of polysaccharides caused compensatory up-regulation of three known acetyltransferases and increased polysaccharide accessibility to glycosyl hydrolases. Transgenic plants showed increased resistance to the fungal pathogens Botrytis cinerea and Bipolaris sorokiniana but not to the bacterial pathogens Pseudomonas syringae and Xanthomonas oryzae. These results demonstrate a role, in both monocot and dicot plants, of hemicellulose and pectin acetylation in plant defense against fungal pathogens.
DOI:10.1104/pp.115.1.181URLPMID:9306698 [本文引用: 1]
Xyloglucan endotransglycosylases (XETs) modify a major component of the plant cell wall and therefore may play critical roles in generating tissue properties and influencing morphogenesis. An XET-related gene family exists in Arabidopsis thaliana, the members of which show differential regulation of expression. TCH4 expression is rapidly regulated by mechanical stimuli, temperature shifts, light, and hormones. As a first step in determining whether Arabidopsis XET-related proteins have distinct properties, we produced recombinant TCH4 protein in bacteria and determined its enzymatic characteristics. TCH4 specifically transglycosylates only xyloglucan. The enzyme prefers to transfer a portion of a donor polymer onto another xyloglucan polymer (acceptor); TCH4 will also utilize xyloglucan-derived oligosaccharides as acceptors but discriminates between differentially fucosylated oligosaccharides. TCH4 is most active at pH 6.0 to 6.5 and is surprisingly cold-tolerant with an optimum of 12 to 18 degrees C. TCH4 activity is enhanced by urea and bovine serum albumin, but nor cations, reducing agents, or carboxymethylcellulose. These studies indicate that TCH4 is specific for xyloglucan, but that the molecular mass and the fucosyl content of the substrates influence enzymatic reaction rates. TCH4 is unlikely to play a role in acid-induced wall loosening but may function in cold acclimation or cold-tolerant growth.
DOI:10.3389/fpls.2017.00806URLPMID:28567047 [本文引用: 1]
Unlike animals, plants, being sessile, cannot escape from exposure to severe abiotic stresses such as extreme temperature and water deficit. The dynamic structure of plant cell wall enables them to undergo compensatory changes, as well as maintain physical strength, with changing environments. Plant hormones known as brassinosteroids (BRs) play a key role in determining cell wall expansion during stress responses. Cell wall deposition differs between grasses (Poaceae) and dicots. Grass species include many important food, fiber, and biofuel crops. In this article, we focus on recent advances in BR-regulated cell wall biosynthesis and remodeling in response to stresses, comparing our understanding of the mechanisms in grass species with those in the more studied dicots. A more comprehensive understanding of BR-mediated changes in cell wall integrity in grass species will benefit the development of genetic tools to improve crop productivity, fiber quality and plant biomass recalcitrance.
DOI:10.1093/pcp/pcf171URLPMID:12514239 [本文引用: 1]
The polysaccharide xyloglucan is thought to play an important structural role in the primary cell wall of dicotyledons. Accordingly, there is considerable interest in understanding the biochemical basis and regulation of xyloglucan metabolism, and research over the last 16 years has identified a large family of cell wall proteins that specifically catalyze xyloglucan endohydrolysis and/or endotransglucosylation. However, a confusing and contradictory series of nomenclatures has emerged in the literature, of which xyloglucan endotransglycosylases (XETs) and endoxyloglucan transferases (EXGTs) are just two examples, to describe members of essentially the same class of genes/proteins. The completion of the first plant genome sequencing projects has revealed the full extent of this gene family and so this is an opportune time to resolve the many discrepancies in the database that include different names being assigned to the same gene. Following consultation with members of the scientific community involved in plant cell wall research, we propose a new unifying nomenclature that conveys an accurate description of the spectrum of biochemical activities that cumulative research has shown are catalyzed by these enzymes. Thus, a member of this class of genes/proteins will be referred to as a xyloglucan endotransglucosylase/hydrolase (XTH). The two known activities of XTH proteins are referred to enzymologically as xyloglucan endotransglucosylase (XET, which is hereby re-defined) activity and xyloglucan endohydrolase (XEH) activity. This review provides a summary of the biochemical and functional diversity of XTHs, including an overview of the structure and organization of the Arabidopsis XTH gene family, and highlights the potentially important roles that XTHs appear to play in numerous examples of plant growth and development.
DOI:10.1104/pp.15.01066URLPMID:26729799 [本文引用: 1]
Stomatal guard cells are pairs of specialized epidermal cells that control water and CO2 exchange between the plant and the environment. To fulfill the functions of stomatal opening and closure that are driven by changes in turgor pressure, guard cell walls must be both strong and flexible, but how the structure and dynamics of guard cell walls enable stomatal function remains poorly understood. To address this question, we applied cell biological and genetic analyses to investigate guard cell walls and their relationship to stomatal function in Arabidopsis (Arabidopsis thaliana). Using live-cell spinning disk confocal microscopy, we measured the motility of cellulose synthase (CESA)-containing complexes labeled by green fluorescent protein (GFP)-CESA3 and observed a reduced proportion of GFP-CESA3 particles colocalizing with microtubules upon stomatal closure. Imaging cellulose organization in guard cells revealed a relatively uniform distribution of cellulose in the open state and a more fibrillar pattern in the closed state, indicating that cellulose microfibrils undergo dynamic reorganization during stomatal movements. In cesa3(je5) mutants defective in cellulose synthesis and xxt1 xxt2 mutants lacking the hemicellulose xyloglucan, stomatal apertures, changes in guard cell length, and cellulose reorganization were aberrant during fusicoccin-induced stomatal opening or abscisic acid-induced stomatal closure, indicating that sufficient cellulose and xyloglucan are required for normal guard cell dynamics. Together, these results provide new insights into how guard cell walls allow stomata to function as responsive mediators of gas exchange at the plant surface.
DOI:10.1104/pp.111.192195URLPMID:22267505 [本文引用: 1]
In growing cells, xyloglucan is thought to connect cellulose microfibrils and regulate their separation during wall extension. In Arabidopsis (Arabidopsis thaliana), a significant proportion of xyloglucan side chains contain beta-galactose linked to alpha-xylose at O2. In this work, we identified AtBGAL10 (At5g63810) as the gene responsible for the majority of beta-galactosidase activity against xyloglucan. Xyloglucan from bgal10 insertional mutants was found to contain a large proportion of unusual subunits, such as GLG and GLLG. These subunits were not detected in a bgal10 xyl1 double mutant, deficient in both beta-galactosidase and alpha-xylosidase. Xyloglucan from bgal10 xyl1 plants was enriched instead in XXLG/XLXG and XLLG subunits. In both cases, changes in xyloglucan composition were larger in the endoglucanase-accessible fraction. These results suggest that glycosidases acting on nonreducing ends digest large amounts of xyloglucan in wild-type plants, while plants deficient in any of these activities accumulate partly digested subunits. In both bgal10 and bgal10 xyl1, siliques and sepals were shorter, a phenotype that could be explained by an excess of nonreducing ends leading to a reinforced xyloglucan network. Additionally, AtBGAL10 expression was examined with a promoter-reporter construct. Expression was high in many cell types undergoing wall extension or remodeling, such as young stems, abscission zones, or developing vasculature, showing good correlation with alpha-xylosidase expression.
DOI:10.1104/pp.16.01713URLPMID:27956490
In many flowering plants, xyloglucan is a major component of primary cell walls, where it plays an important role in growth regulation. Xyloglucan can be degraded by a suite of exoglycosidases that remove specific sugars. In this work, we show that the xyloglucan backbone, formed by (1-->4)-linked beta-d-glucopyranosyl residues, can be attacked by two different Arabidopsis (Arabidopsis thaliana) beta-glucosidases from glycoside hydrolase family 3. While BGLC1 (At5g20950; for beta-glucosidase active against xyloglucan 1) is responsible for all or most of the soluble activity, BGLC3 (At5g04885) is usually a membrane-anchored protein. Mutations in these two genes, whether on their own or combined with mutations in other exoglycosidase genes, resulted in the accumulation of partially digested xyloglucan subunits, such as GXXG, GXLG, or GXFG. While a mutation in BGLC1 had significant effects on its own, lack of BGLC3 had only minor effects. On the other hand, double bglc1 bglc3 mutants revealed a synergistic interaction that supports a role for membrane-bound BGLC3 in xyloglucan metabolism. In addition, bglc1 bglc3 was complemented by overexpression of either BGLC1 or BGLC3 In overexpression lines, BGLC3 activity was concentrated in a microsome-enriched fraction but also was present in soluble form. Finally, both genes were generally expressed in the same cell types, although, in some cases, BGLC3 was expressed at earlier stages than BGLC1 We propose that functional specialization could explain the separate localization of both enzymes, as a membrane-bound beta-glucosidase could specifically digest soluble xyloglucan without affecting the wall-bound polymer.
DOI:10.1146/annurev-arplant-042809-112315URLPMID:20192742 [本文引用: 2]
Hemicelluloses are polysaccharides in plant cell walls that have beta-(1-->4)-linked backbones with an equatorial configuration. Hemicelluloses include xyloglucans, xylans, mannans and glucomannans, and beta-(1-->3,1-->4)-glucans. These types of hemicelluloses are present in the cell walls of all terrestrial plants, except for beta-(1-->3,1-->4)-glucans, which are restricted to Poales and a few other groups. The detailed structure of the hemicelluloses and their abundance vary widely between different species and cell types. The most important biological role of hemicelluloses is their contribution to strengthening the cell wall by interaction with cellulose and, in some walls, with lignin. These features are discussed in relation to widely accepted models of the primary wall. Hemicelluloses are synthesized by glycosyltransferases located in the Golgi membranes. Many glycosyltransferases needed for biosynthesis of xyloglucans and mannans are known. In contrast, the biosynthesis of xylans and beta-(1-->3,1-->4)-glucans remains very elusive, and recent studies have led to more questions than answers.
DOI:10.1104/pp.113.221788URLPMID:23893172 [本文引用: 1]
Xyloglucan (XyG) is the dominant hemicellulose present in the primary cell walls of dicotyledonous plants. Unlike Arabidopsis (Arabidopsis thaliana) XyG, which contains galactosyl and fucosyl substituents, tomato (Solanum lycopersicum) XyG contains arabinofuranosyl residues. To investigate the biological function of these differing substituents, we used a functional complementation approach. Candidate glycosyltransferases were identified from tomato by using comparative genomics with known XyG galactosyltransferase genes from Arabidopsis. These candidate genes were expressed in an Arabidopsis mutant lacking XyG galactosylation, and two of them resulted in the production of arabinosylated XyG, a structure not previously found in this plant species. These genes may therefore encode XyG arabinofuranosyltransferases. Moreover, the addition of arabinofuranosyl residues to the XyG of this Arabidopsis mutant rescued a growth and cell wall biomechanics phenotype, demonstrating that the function of XyG in plant growth, development, and mechanics has considerable flexibility in terms of the specific residues in the side chains. These experiments also highlight the potential of reengineering the sugar substituents on plant wall polysaccharides without compromising growth or viability.
DOI:10.1104/pp.114.256479URLPMID:25681330 [本文引用: 1]
A mutation in the ALTERED XYLOGLUCAN9 (AXY9) gene was found to be causative for the decreased xyloglucan acetylation phenotype of the axy9.1 mutant, which was identified in a forward genetic screen for Arabidopsis (Arabidopsis thaliana) mutants. The axy9.1 mutant also exhibits decreased O-acetylation of xylan, implying that the AXY9 protein has a broad role in polysaccharide acetylation. An axy9 insertional mutant exhibits severe growth defects and collapsed xylem, demonstrating the importance of wall polysaccharide O-acetylation for normal plant growth and development. Localization and topological experiments indicate that the active site of the AXY9 protein resides within the Golgi lumen. The AXY9 protein appears to be a component of the plant cell wall polysaccharide acetylation pathway, which also includes the REDUCED WALL ACETYLATION and TRICHOME BIREFRINGENCE-LIKE proteins. The AXY9 protein is distinct from the TRICHOME BIREFRINGENCE-LIKE proteins, reported to be polysaccharide acetyltransferases, but does share homology with them and other acetyltransferases, suggesting that the AXY9 protein may act to produce an acetylated intermediate that is part of the O-acetylation pathway.
DOI:10.4161/cib.21422URLPMID:23336031 [本文引用: 1]
Two of the most widely and intensively cultivated jute species, Corchorus capsularis and Corchorus olitorius, suffer severely from a stem rot disease caused by the fungus Macrophomina phaseolina. Wild jute species, C. trilocularis, shows resistance to this pathogenic fungus. In this study, the technique of differential display was applied to identify genes which are differentially expressed, under both infected and un-infected conditions, between C. trilocularis and C. olitorius var O-72. Two xyloglucan endotransglycosylase/hydrolase (XTH) genes designated CoXTH1 (from Corchorus olitorius) and CtXTH1 (from C.trilocularis) were identified from each of the two species which show different expression patterns upon fungal infection. A steady rise in the expression of CtXTH1 in response to infection was observed by quantitative real time PCR whereas the expression of CoXTH1 was found to be downregulated. Full length sequences of these two genes were determined using primer based gene walking and RACE PCR. This study confirms the involvement of XTH in molecular interactions between M. phaseolina and jute. However, it remains to be explored whether XTH is an essential component of the signaling pathway involved in plant-fungal interaction.
DOI:10.1093/jxb/erw321URLPMID:27605715 [本文引用: 1]
Regulation and maintenance of cell wall physical properties are crucial for plant growth and environmental response. In the germination process, hypocotyl cell expansion and endosperm weakening are prerequisites for dicot seeds to complete germination. We have identified the Arabidopsis mutant thermoinhibition-resistant germination 1 (trg1), which has reduced seed dormancy and insensitivity to unfavourable conditions for germination owing to a loss-of-function mutation of TRG1/XYL1, which encodes an alpha-xylosidase. Compared to those of wild type, the elongating stem of trg1 showed significantly lower viscoelasticity, and the fruit epidermal cells were longitudinally shorter and horizontally enlarged. Actively growing tissues of trg1 over-accumulated free xyloglucan oligosaccharides (XGOs), and the seed cell wall had xyloglucan with a greatly reduced molecular weight. These observations suggest that XGOs reduce xyloglucan size by serving as an acceptor in transglycosylation and eventually enhancing cell wall loosening. TRG1/XYL1 gene expression was abundant in growing wild-type organs and tissues but relatively low in cells at most actively elongating part of the tissues, suggesting that alpha-xylosidase contributes to maintaining the mechanical integrity of the primary cell wall in the growing and pre-growing tissues. In germinating seeds of trg1, expression of genes encoding specific abscisic acid and gibberellin metabolism enzymes was altered in accordance with the aberrant germination phenotype. Thus, cell wall integrity could affect seed germination not only directly through the physical properties of the cell wall but also indirectly through the regulation of hormone gene expression.
DOI:10.1038/srep46099URLPMID:28443615 [本文引用: 1]
Cellulose is an economically important material, but routes of its industrial processing have not been fully explored. The plant cell wall - the major source of cellulose - harbours enzymes of the xyloglucan endotransglucosylase/hydrolase (XTH) family. This class of enzymes is unique in that it is capable of elongating polysaccharide chains without the requirement for activated nucleotide sugars (e.g., UDP-glucose) and in seamlessly splitting and reconnecting chains of xyloglucan, a naturally occurring soluble analogue of cellulose. Here, we show that a recombinant version of AtXTH3, a thus far uncharacterized member of the Arabidopsis XTH family, catalysed the transglycosylation between cellulose and cello-oligosaccharide, between cellulose and xyloglucan-oligosaccharide, and between xyloglucan and xyloglucan-oligosaccharide, with the highest reaction rate observed for the latter reaction. In addition, this enzyme formed cellulose-like insoluble material from a soluble cello-oligosaccharide in the absence of additional substrates. This newly found activity (designated
[本文引用: 1]
[本文引用: 1]
DOI:10.1038/s41598-019-38688-3URLPMID:30783145 [本文引用: 1]
Cold acclimation (CA) leads to increased plant freezing tolerance during exposure to low, non-freezing temperatures as a result of many physiological, biochemical and molecular changes that have been extensively investigated. In addition, many plant species, such as Arabidopsis thaliana, respond to a subsequent exposure to mild, non-damaging freezing temperatures with an additional increase in freezing tolerance referred to as sub-zero acclimation (SZA). There is comparatively little information available about the molecular basis of SZA. However, previous transcriptomic studies indicated that cell wall modification may play an important role during SZA. Here we show that CA and SZA are accompanied by extensive changes in cell wall amount, composition and structure. While CA leads to a significant increase in cell wall amount, the relative proportions of pectin, hemicellulose and cellulose remained unaltered during both CA and SZA. However, both treatments resulted in more subtle changes in structure as determined by infrared spectroscopy and monosaccharide composition as determined by gas chromatography-mass spectrometry. These differences could be related through a proteomic approach to the accumulation of cell wall modifying enzymes such as pectin methylesterases, pectin methylesterase inhibitors and xyloglucan endotransglucosylases/hydrolases in the extracellular matrix.
DOI:10.1105/tpc.104.024018URLPMID:15258268 [本文引用: 1]
The sensitive to freezing2-1 (sfr2-1) mutation causes freezing sensitivity in Arabidopsis thaliana. By mapping, transgenic complementation, and sequencing, sfr2-1 was revealed to be a mutation in gene At3g06510. A new knockout allele was obtained, and its identical freezing-sensitive phenotype confirmed that the SFR2 gene product is essential for freezing tolerance. Transcription of SFR2 was observed to be constitutive rather than stress inducible and was distributed throughout most aerial tissues. SFR2 encodes a protein homologous to family 1 glycosyl hydrolases (beta-glycosidases), but the predicted AtSFR2 protein is divergent from all other family 1 beta-glycosidases of Arabidopsis, showing closer homology to the sequences of several beta-glycosidases from thermophilic archea and bacteria. After purification from a heterologous expression system, AtSFR2 displayed a specific hydrolytic activity against beta-d-glucosides.
DOI:10.1038/s41477-019-0502-0URLPMID:31506641 [本文引用: 1]
Cell walls are highly dynamic structures that provide mechanical support for plant cells during growth, development and adaptation to a changing environment. Thus, it is important for plants to monitor the state of their cell walls and ensure their functional integrity at all times. This monitoring involves perception of physical forces at the cell wall-plasma membrane interphase. These forces are altered during cell division and morphogenesis, as well as in response to various abiotic and biotic stresses. Mechanisms responsible for the perception of physical stimuli involved in these processes have been difficult to separate from other regulatory mechanisms perceiving chemical signals such as hormones, peptides or cell wall fragments. However, recently developed technologies in combination with more established genetic and biochemical approaches are beginning to open up this exciting field of study. Here, we will review our current knowledge of plant cell wall integrity signalling using selected recent findings and highlight how the cell wall-plasma membrane interphase can act as a venue for sensing changes in the physical forces affecting plant development and stress responses. More importantly, we discuss how these signals may be integrated with chemical signals derived from established signalling cascades to control specific adaptive responses during exposure to biotic and abiotic stresses.
DOI:10.1073/pnas.052450699URLPMID:11854459 [本文引用: 1]
Cell walls of the Arabidopsis mutant mur2 contain less than 2% of the wild-type amount of fucosylated xyloglucan because of a point mutation in the fucosyltransferase AtFUT1. The mur2 mutation eliminates xyloglucan fucosylation in all major plant organs, indicating that Arabidopsis thaliana fucosyltransferase 1 (AtFUT1) accounts for all of the xyloglucan fucosyltransferase activity in Arabidopsis. Despite this alteration in structure, mur2 plants show a normal growth habit and wall strength. In contrast, Arabidopsis mur1 mutants that are defective in the de novo synthesis of l-fucose exhibit a dwarfed growth habit and decreased wall strength [Reiter, W. D., Chapple, C. & Somerville, C. R. (1993) Science 261, 1032-1035]. Because the mur1 mutation affects several cell wall polysaccharides, whereas the mur2 mutation is specific to xyloglucan, the phenotypes of mur1 plants appear to be caused by structural changes in fucosylated pectic components such as rhamnogalacturonan-II. The normal growth habit and wall strength of mur2 plants casts doubt on hypotheses regarding roles of xyloglucan fucosylation in facilitating xyloglucan-cellulose interactions or in modulating growth regulator activity.
DOI:10.1093/jxb/eru189URL [本文引用: 1]
Root hairs are important for nutrient uptake, anchorage, and plant-microbe interactions. From a population of rice (Oryza sativa) mutagenized by ethyl methanesulfonate (EMS), a short root hair2 (srh2) mutant was identified. In hydroponic culture, srh2 seedlings were significantly reduced in root hair length. Bubble-like extrusions and irregular epidermal cells were observed at the tips of srh2 root hairs when grown under acidic conditions, suggesting the possible reduction of the tensile strength of the cell wall in this mutant. Map-based cloning identified a mutation in the gene encoding xyloglucan (XyG) 6-xylosyltransferase (OsXXT1). OsXXT1 displays more than 70% amino acid sequence identity with the previously characterized Arabidopsis thaliana XYG XYLOSYL TRANSFERASE 1 (AtXXT1) and XYG XYLOSYL TRANSFERASE 2 (AtXXT2), which catalyse the transfer of xylose onto beta-1,4-glucan chains. Furthermore, expression of the full-length coding sequence of OsXXT1 could complement the root hair defect, and slow growth and XyG synthesis in the Arabidopsis xxt1 xxt2 double mutant. Transgenic plants expressing the beta-glucuronidase (GUS) reporter under the control of the OsXXT1 promoter displayed GUS expression in multiple tissues, most prominently in root epidermal cells. These results demonstrate the importance of OsXXT1 in maintaining cell wall structure and tensile strength in rice, a typical grass species that contains relatively low XyG content in cell walls.
DOI:10.3390/genes9060273URL [本文引用: 1]
DOI:10.1038/srep42400URLPMID:28218299 [本文引用: 1]
Banana Fusarium wilt caused by Fusarium oxysporum f. sp. cubense (Foc) is soil-borne disease of banana (Musa spp.) causing significant economic losses. Extensins and arabinogalactan proteins (AGPs) are cell wall components important for pathogen defence. Their significance for Foc resistance in banana was not reported so far. In this study, two banana cultivars differing in Foc sensitivity were used to monitor the changes in transcript levels, abundance and distribution of extensins and AGPs after wounding and Foc inoculation. Extensins mainly appeared in the root cap and meristematic cells. AGPs recognized by JIM13, JIM8, PN16.4B4 and CCRC-M134 antibodies located in root hairs, xylem and root cap. Individual AGPs and extensins showed specific radial distribution in banana roots. At the transcript level, seven extensins and 23 AGPs were differentially expressed between two banana cultivars before and after treatments. Two extensins and five AGPs responded to the treatments at the protein level. Most extensins and AGPs were up-regulated by wounding and pathogen inoculation of intact plants but down-regulated by pathogen attack of wounded plants. Main components responsible for the resistance of banana were MaELP-2 and MaPELP-2. Our data revealed that AGPs and extensins represent dynamic cell wall components involved in wounding and Foc resistance.
[本文引用: 1]
DOI:10.3389/fpls.2018.01773URLPMID:30619385 [本文引用: 1]
Sorghum (Sorghum bicolor) is an important bioenergy crop. Its biomass mainly consists of the cellulosic and non-cellulosic polysaccharides, both which can be converted to biofuels. The biosynthesis of non-cellulosic polysaccharides involves several glycosyltransferases (GT) families including GT47. However, there was no systemic study on GT47 family in sorghum to date. Here, we identified 39 sorghum GT47 family members and showed the functional divergences of MURUS3 (MUR3) homologs. Sorghum GT47 proteins were phylogenetically clustered into four distinct subfamilies. Within each subfamily, gene structure was relatively conserved between the members. Ten gene pairs were identified from the 39 GT47 genes, of which two pairs might be originated from tandem duplication. 25.6% (10/39) of sorghum GT47 genes were homologous to Arabidopsis MUR3, a xyloglucan biosynthesis gene in primary cell walls. SbGT47_2, SbGT47_7, and SbGT47_8, three most homologous genes of MUR3, exhibited different tissue expression patterns and were selected for complementation into Arabidopsis mur3-3. Physiological and cell wall analyses showed that SbGT47_2 and SbGT47_7 may be two functional xyloglucan galactosyltransferases in sorghum. Further studies found that MUR3-like genes are widely present in the seed plants but not in the chlorophytic alga Chlamydomonas reinhardtii. Our results provide novel information for evolutionary analysis and functional dissection of sorghum GT47 family members.
DOI:10.1093/jxb/erz311URLPMID:31257449 [本文引用: 1]
Plants have evolved various strategies to sense and respond to saline environments, which severely reduce plant growth and limit agricultural productivity. Alteration to the cell wall is one strategy that helps plants adapt to salt stress. However, the physiological mechanism of how the cell wall components respond to salt stress is not fully understood. Here, we show that expression of XTH30, encoding xyloglucan endotransglucosylase-hydrolase30, is strongly up-regulated in response to salt stress in Arabidopsis. Loss-of-function of XTH30 leads to increased salt tolerance and overexpression of XTH30 results in salt hypersensitivity. XTH30 is located in the plasma membrane and is highly expressed in the root, flower, stem, and etiolated hypocotyl. The NaCl-induced increase in xyloglucan (XyG)-derived oligosaccharide (XLFG) of the wild type is partly blocked in xth30 mutants. Loss-of-function of XTH30 slows down the decrease of crystalline cellulose content and the depolymerization of microtubules caused by salt stress. Moreover, lower Na+ accumulation in shoot and lower H2O2 content are found in xth30 mutants in response to salt stress. Taken together, these results indicate that XTH30 modulates XyG side chains, altered abundance of XLFG, cellulose synthesis, and cortical microtubule stability, and negatively affecting salt tolerance.
DOI:10.3389/fpls.2015.00353URLPMID:26074928 [本文引用: 1]
Information on the spatial distribution of arabinogalactan proteins (AGPs) in plant organs and tissues during plant reactions to low temperature (LT) is limited. In this study, the extracellular distribution of AGPs in banana leaves and roots, and their changes under LT stress were investigated in two genotypes differing in chilling tolerance, by immuno-techniques using 17 monoclonal antibodies against different AGP epitopes. Changes in total classical AGPs in banana leaves were also tested. The results showed that AGP epitopes recognized by JIM4, JIM14, JIM16, and CCRC-M32 antibodies were primarily distributed in leaf veins, while those recognized by JIM8, JIM13, JIM15, and PN16.4B4 antibodies exhibited predominant sclerenchymal localization. Epitopes recognized by LM2, LM14, and MAC207 antibodies were distributed in both epidermal and mesophyll cells. Both genotypes accumulated classical AGPs in leaves under LT treatment, and the chilling tolerant genotype contained higher classical AGPs at each temperature treatment. The abundance of JIM4 and JIM16 epitopes in the chilling-sensitive genotype decreased slightly after LT treatment, and this trend was opposite for the tolerant one. LT induced accumulation of LM2- and LM14-immunoreactive AGPs in the tolerant genotype compared to the sensitive one, especially in phloem and mesophyll cells. These epitopes thus might play important roles in banana LT tolerance. Different AGP components also showed differential distribution patterns in banana roots. In general, banana roots started to accumulate AGPs under LT treatment earlier than leaves. The levels of AGPs recognized by MAC207 and JIM13 antibodies in the control roots of the tolerant genotype were higher than in the chilling sensitive one. Furthermore, the chilling tolerant genotype showed high immuno-reactivity against JIM13 antibody. These results indicate that several AGPs are likely involved in banana tolerance to chilling injury.
DOI:10.1104/pp.17.00375URLPMID:28710128 [本文引用: 1]
In search of Botrytis cinerea cell death-inducing proteins, we found a xyloglucanase (BcXYG1) that induced strong necrosis and a resistance response in dicot plants. Expression of the BcXYG1 gene was strongly induced during the first 12 h post inoculation, and analysis of disease dynamics using PathTrack showed that a B. cinerea strain overexpressing BcXYG1 produced early local necrosis, supporting a role of BcXYG1 as an early cell death-inducing factor. The xyloglucanase activity of BcXYG1 was not necessary for the induction of necrosis and plant resistance, as a mutant of BcXYG1 lacking the xyloglucanase enzymatic activity retained both functions. Residues in two exposed loops on the surface of BcXYG1 were found to be necessary for the induction of cell death but not to induce plant resistance. Further analyses showed that BcXYG1 is apoplastic and possibly interacts with the proteins of the plant cell membrane and also that the BcXYG1 cell death-promoting signal is mediated by the leucine-rich repeat receptor-like kinases BAK1 and SOBIR1. Our findings support the role of cell death-inducing proteins in establishing the infection of necrotrophic pathogens and highlight the recognition of fungal apoplastic proteins by the plant immune system as an important mechanism of resistance against this class of pathogens.
DOI:10.1105/tpc.112.106039URLPMID:23204407 [本文引用: 1]
Xyloglucan endohydrolase (XEH) and xyloglucan endotransglucosylase (XET) activities, encoded by xyloglucan endotransglucosylase-hydrolase (XTH) genes, are involved in cell wall extension by cutting or cutting and rejoining xyloglucan chains, respectively. However, the physiological significance of this biochemical activity remains incompletely understood. Here, we find that an XTH31 T-DNA insertion mutant, xth31, is more Al resistant than the wild type. XTH31 is bound to the plasma membrane and the encoding gene is expressed in the root elongation zone and in nascent leaves, suggesting a role in cell expansion. XTH31 transcript accumulation is strongly downregulated by Al treatment. XTH31 expression in yeast yields a protein with an in vitro XEH: XET activity ratio of >5000:1. xth31 accumulates significantly less Al in the root apex and cell wall, shows remarkably lower in vivo XET action and extractable XET activity, has a lower xyloglucan content, and exhibits slower elongation. An exogenous supply of xyloglucan significantly ameliorates Al toxicity by reducing Al accumulation in the roots, owing to the formation of an Al-xyloglucan complex in the medium, as verified by an obvious change in chemical shift of Al-27-NMR. Taken together, the data indicate that XTH31 affects Al sensitivity by modulating cell wall xyloglucan content and Al binding capacity.
猕猴桃果实成熟进程中木葡聚糖内糖基转移酶mRNA水平的变化
1
1999
... 植物细胞壁是围绕在植物原生质体外的一种细胞结构, 由初生细胞壁和次生细胞壁组成.初生细胞壁位于外层, 是由纤维素、半纤维素和果胶等多糖及结构蛋白组成的一种复杂网络结构.次生细胞壁的结构与组成均与初生细胞壁有所不同, 除了纤维素和半纤维素外, 还含有木质素.植物细胞壁除了作为细胞的主要结构成分, 承担机械支撑、维持和决定细胞形态等功能外, 还在植物的生长发育(
半纤维素的化学结构和生理功能
1
1994
... 木葡聚糖(XyG)是双子叶植物初生壁中含量最为丰富的半纤维素成分, 约占细胞壁重量的20%-25%, 而禾本科植物中一般仅为2%-10% (
低温胁迫对香蕉(Musa spp.)细胞壁半纤维素代谢的影响
1
2018
... 早在20多年前, 拟南芥XyG特异糖基化基因TCH4 (即AtXTH22)就已被证明非常耐(12-18)°C低温(
参与木葡聚糖合成的糖基转移酶基因研究进展
1
2015
... 木葡聚糖(XyG)是双子叶植物初生壁中含量最为丰富的半纤维素成分, 约占细胞壁重量的20%-25%, 而禾本科植物中一般仅为2%-10% (
Plant cell wall-mediated immunity: cell wall changes trigger disease resistance responses
1
2018
... 植物在整个生命周期中经常遭受各种生物和非生物逆境胁迫.相应地, 植物采取多种防御策略来应对逆境, 包括改变细胞壁结构与组成、表达包括与木葡聚糖代谢相关基因在内的抗逆基因(
Identification of a xyloglucan-specific endo-(1-4)-beta-D-glucanase inhibitor protein from apple ( Malus × domestica Borkh.) as a potential defense gene against Botryos- phaeria dothidea
1
2015
... 质外体组织是植物抵御病原菌的关键战场.病原菌分泌质外体XEGs降解植物XyG的主链, 相应地, 植物通过诱导表达其抑制蛋白(xyloglucan-speci?c endo- 1,4-glucanase inhibitor protein, XEGIP)的方式进行抵抗.这在苹果(Malus x domestica)和大豆等多种植物中得到证实.植物XEGIP通过与病原菌XEG结合从而使其失活, 进而抑制病原菌对植物细胞壁XyG的降解(
Disrupting two Arabidopsis thaliana xylosyltransferase genes results in plants deficient in xyloglucan, a major primary cell wall component
1
2008
... 所有类型的XyG葡聚糖主链都含有木糖基取代基.XyG特异木糖基转移酶(XyG: xylosyltransferase, XXT)负责将UDP-木糖添加到主链葡萄糖残基O-6位上(
Constitutive expression of abiotic stress-inducible hot pepper CaXTH3, which encodes a xyloglucan endotransglucosylase/hydrolase homolog, improves drought and salt tolerance in transgenic Arabidopsis plants
2
2006
... 植物XyG代谢相关基因与其抗盐性的研究多见于XTH基因家族.例如, 转辣椒(Capsicum annuum) CaXTH3的拟南芥耐盐性增强, 推测其通过调节气孔关闭来防止过度失水, 从而提高耐盐性(
... 近年来, 越来越多的研究表明, XyG代谢相关基因能够响应多种逆境或增强植物对多种逆境的抗性.例如, PeXTH基因同时具有提高胡杨耐盐及重金属的功能(
The pepper extracellular xyloglucan-specific endo-β-1,4-glucanase inhibitor protein gene, CaXEGIP1, is required for plant cell death and defense responses
3
2013
... 降解植物XyG是病原菌成功侵染寄主的必要条件(
... 质外体组织是植物抵御病原菌的关键战场.病原菌分泌质外体XEGs降解植物XyG的主链, 相应地, 植物通过诱导表达其抑制蛋白(xyloglucan-speci?c endo- 1,4-glucanase inhibitor protein, XEGIP)的方式进行抵抗.这在苹果(Malus x domestica)和大豆等多种植物中得到证实.植物XEGIP通过与病原菌XEG结合从而使其失活, 进而抑制病原菌对植物细胞壁XyG的降解(
... 早期人们发现, XyG降解产生的寡糖片段XGO可抑制烟草坏死病毒, 具有诱导抗性的功能(
Constitutive expression of CaXTH3, a hot pepper xyloglucan endotransglucosylase/hydrolase, enhanced tolerance to salt and drought stresses without phenotypic defects in tomato plants ( Solanum lycopersicum cv. Dotaerang)
1
2011
... 近年来, 越来越多的研究表明, XyG代谢相关基因能够响应多种逆境或增强植物对多种逆境的抗性.例如, PeXTH基因同时具有提高胡杨耐盐及重金属的功能(
The cell wall-derived xyloglucan is a new DAMP triggering plant immunity in Vitis vinifera and Arabidopsis thaliana
1
2018
... 早期人们发现, XyG降解产生的寡糖片段XGO可抑制烟草坏死病毒, 具有诱导抗性的功能(
A gene from the cellulose synthase-like C family encodes a β-1,4-glucan synthase
1
2007
... XyG主链骨架的生物合成由XyG特异葡聚糖合成酶, 即β-1,4-葡聚糖合成酶(β-1,4-glucan synthase, GS)负责(
Insights into plant cell wall degradation from the genome sequence of the soil bacterium Cellvibrio japonicus
1
2008
... 降解植物XyG是病原菌成功侵染寄主的必要条件(
Evolution of xyloglucan-related genes in green plants
1
2010
... 木葡聚糖(XyG)是双子叶植物初生壁中含量最为丰富的半纤维素成分, 约占细胞壁重量的20%-25%, 而禾本科植物中一般仅为2%-10% (
Phenolic compounds that confer resistance to spruce budworm
1
2011
... 目前, XyG代谢与植物抗虫性的关系研究较少.芹菜(Apium graveolens) AgXTH1在桃蚜(Myzus persicae)侵染后系统性地在韧皮部中上调表达, 拟南芥中该基因的同源基因AtXTH33的突变体比野生型更吸引蚜虫, 表明AtXTH33调控拟南芥对蚜虫的抗性(
Involvement of the xyloglucan endotransglycosylase/hydrolases encoded by celery XTH1 and Arabidopsis XTH33 in the phloem response to aphids
1
2007
... 目前, XyG代谢与植物抗虫性的关系研究较少.芹菜(Apium graveolens) AgXTH1在桃蚜(Myzus persicae)侵染后系统性地在韧皮部中上调表达, 拟南芥中该基因的同源基因AtXTH33的突变体比野生型更吸引蚜虫, 表明AtXTH33调控拟南芥对蚜虫的抗性(
Isolation of a novel xyloglucan endotransglucosylase ( OsXET9) gene from rice and analysis of the response of this gene to abiotic stresses
1
2011
... 近年来, 越来越多的研究表明, XyG代谢相关基因能够响应多种逆境或增强植物对多种逆境的抗性.例如, PeXTH基因同时具有提高胡杨耐盐及重金属的功能(
The plant cell wall integrity maintenance and immune signaling systems cooperate to control stress responses in Arabidopsis thaliana
1
2018
... 植物在整个生命周期中经常遭受各种生物和非生物逆境胁迫.相应地, 植物采取多种防御策略来应对逆境, 包括改变细胞壁结构与组成、表达包括与木葡聚糖代谢相关基因在内的抗逆基因(
An Arabidopsis gene encoding an α-xylosyltransferase involved in xyloglucan biosynthesis
1
2002
... 所有类型的XyG葡聚糖主链都含有木糖基取代基.XyG特异木糖基转移酶(XyG: xylosyltransferase, XXT)负责将UDP-木糖添加到主链葡萄糖残基O-6位上(
Biochemistry and physiological roles of enzymes that ‘cut and paste’ plant cell-wall polysaccharides
2
2013
... XyG在高尔基体中合成后, 被囊泡运输到细胞壁.迄今为止, 人们对XyG掺入现有细胞壁的过程知之甚少.已有研究表明, XyG可以通过非酶自组装的方式与现有纤维素微纤维网络结合(
... 植物XyG内转糖苷酶/水解酶(endotransglycosylase/ hydrolases, XTHs)基因属于CAZy糖基水解酶16 (glycoside hydrolase 16, GH16)家族基因, 编码的蛋白可能具有2种不同的催化活性, 对XyG的影响不同.其中XyG特异内水解酶(XyG: endohydrolase, XEH)活性在形式上是XyG特异的内切葡聚糖酶(xyloglucan-specific endo-1,4-glucanases, XEGs), 降解XyG主链骨架(
An unambiguous nomenclature for xyloglucan-derived oligosaccharides
1
1993
... XyG是一种由D-吡喃葡萄糖残基以β-1→4键相连构成主链的杂多糖, 葡萄糖残基在O-6位上可被D-吡喃木糖残基所取代(
O-acetylation of Arabidopsis hemicellulose xyloglucan requires AXY4 or AXY4L, proteins with a TBL and DUF231 domain.
3
2011
... XyG的乙酰化主要发生在XXGG型XyG主链的葡萄糖残基及XXXG型XyG侧链的半乳糖残基上(
... ), 而TBL蛋白是多糖特异性的乙酰基转移酶(
... ).例如, 由TBL基因家族成员编码的AXY4/TBL27及AXY4-like (AXY4L/ TBL22)特异性地乙酰化种子中的XyG (
O-acetylation of plant cell wall polysaccharides
1
2012
... 有研究表明, XyG乙酰化程度与植物的抗逆性相关.例如, 将真菌的鼠李糖半乳糖醛酸乙酰酯酶转入拟南芥后, 植株XyG的乙酰化程度显著降低, 转基因植株对灰葡萄孢菌(Botrytis cinerea)等真菌的抗性增强, 但不能增强对病原细菌的抗性(
In tobacco BY-2 cells xyloglucan oligosaccharides alter the expression of genes involved in cell wall metabolism, signaling, stress responses, cell division and transcriptional control
1
2014
... 早期人们发现, XyG降解产生的寡糖片段XGO可抑制烟草坏死病毒, 具有诱导抗性的功能(
AXY8 encodes an α-fucosidase, underscoring the importance of apoplastic metabolism on the fine structure of Arabidopsis cell wall polysaccharides
1
2011
... 参与XyG支链降解的糖苷酶主要包括α-木糖苷酶(如AtXYL1/AXY3) (
Overexpression of Populus euphratica xyloglucan endotransglucosylase/hydrolase gene confers enhanced cadmium tolerance by the restriction of root cadmium uptake in transgenic tobacco
1
2014
... 近年来, 越来越多的研究表明, XyG代谢相关基因能够响应多种逆境或增强植物对多种逆境的抗性.例如, PeXTH基因同时具有提高胡杨耐盐及重金属的功能(
Populus euphratica XTH overexpression enhances salinity tolerance by the development of leaf succulence in transgenic tobacco plants
2
2013
... 植物XyG代谢相关基因与其抗盐性的研究多见于XTH基因家族.例如, 转辣椒(Capsicum annuum) CaXTH3的拟南芥耐盐性增强, 推测其通过调节气孔关闭来防止过度失水, 从而提高耐盐性(
... 近年来, 越来越多的研究表明, XyG代谢相关基因能够响应多种逆境或增强植物对多种逆境的抗性.例如, PeXTH基因同时具有提高胡杨耐盐及重金属的功能(
Pea xyloglucan and cellulose: VI. Xyloglucan-cellulose interactions in vitro and in vivo
1
1987
... XyG在高尔基体中合成后, 被囊泡运输到细胞壁.迄今为止, 人们对XyG掺入现有细胞壁的过程知之甚少.已有研究表明, XyG可以通过非酶自组装的方式与现有纤维素微纤维网络结合(
Pea xyloglucan and cellulose: II. Hydrolysis by pea endo-1,4-β-glucanases
1
1984
... 植物XyG内转糖苷酶/水解酶(endotransglycosylase/ hydrolases, XTHs)基因属于CAZy糖基水解酶16 (glycoside hydrolase 16, GH16)家族基因, 编码的蛋白可能具有2种不同的催化活性, 对XyG的影响不同.其中XyG特异内水解酶(XyG: endohydrolase, XEH)活性在形式上是XyG特异的内切葡聚糖酶(xyloglucan-specific endo-1,4-glucanases, XEGs), 降解XyG主链骨架(
The plant cell wall: a complex and dynamic structure as revealed by the responses of genes under stress conditions
1
2016
... 植物细胞壁是围绕在植物原生质体外的一种细胞结构, 由初生细胞壁和次生细胞壁组成.初生细胞壁位于外层, 是由纤维素、半纤维素和果胶等多糖及结构蛋白组成的一种复杂网络结构.次生细胞壁的结构与组成均与初生细胞壁有所不同, 除了纤维素和半纤维素外, 还含有木质素.植物细胞壁除了作为细胞的主要结构成分, 承担机械支撑、维持和决定细胞形态等功能外, 还在植物的生长发育(
Improvement of multiple agronomic traits by a disease resistance gene via cell wall reinforcement
1
2017
... 植物细胞壁是围绕在植物原生质体外的一种细胞结构, 由初生细胞壁和次生细胞壁组成.初生细胞壁位于外层, 是由纤维素、半纤维素和果胶等多糖及结构蛋白组成的一种复杂网络结构.次生细胞壁的结构与组成均与初生细胞壁有所不同, 除了纤维素和半纤维素外, 还含有木质素.植物细胞壁除了作为细胞的主要结构成分, 承担机械支撑、维持和决定细胞形态等功能外, 还在植物的生长发育(
Drought and heat differentially affect XTH expression and XET activity and action in 3-day-old seedlings of durum wheat cultivars with different stress susceptibility
1
2016
... 近年来, 越来越多的研究表明, XyG代谢相关基因能够响应多种逆境或增强植物对多种逆境的抗性.例如, PeXTH基因同时具有提高胡杨耐盐及重金属的功能(
RNA-Seq analysis of developing nasturtium seeds ( Tropaeolum majus): identification and characterization of an additional galactosyltransferase involved in xyloglucan biosynthesis
1
2012
... 大多数XyG木糖残基在GT47家族的一些酶作用下被其它糖基进一步取代, 其中最常见的是D-半乳糖.UDP-半乳糖在β-半乳糖基转移酶(β-galactosyltransferases, β-GAL)的作用下掺入XyG.例如, MUR3专一性负责XXXG第3位木糖残基的半乳糖化, 将XXXG变成XXLG (
Gene silencing indicates a role for potato endoglucanase inhibitor protein in germplasm resistance to late blight
1
2006
... 质外体组织是植物抵御病原菌的关键战场.病原菌分泌质外体XEGs降解植物XyG的主链, 相应地, 植物通过诱导表达其抑制蛋白(xyloglucan-speci?c endo- 1,4-glucanase inhibitor protein, XEGIP)的方式进行抵抗.这在苹果(Malus x domestica)和大豆等多种植物中得到证实.植物XEGIP通过与病原菌XEG结合从而使其失活, 进而抑制病原菌对植物细胞壁XyG的降解(
Expression of two functionally distinct plant endo-β-1,4-glucanases is essential for the compatible interaction between potato cyst nematode and its hosts
1
2008
... 质外体组织是植物抵御病原菌的关键战场.病原菌分泌质外体XEGs降解植物XyG的主链, 相应地, 植物通过诱导表达其抑制蛋白(xyloglucan-speci?c endo- 1,4-glucanase inhibitor protein, XEGIP)的方式进行抵抗.这在苹果(Malus x domestica)和大豆等多种植物中得到证实.植物XEGIP通过与病原菌XEG结合从而使其失活, 进而抑制病原菌对植物细胞壁XyG的降解(
Differential transcript accumulation of dhydrin and beta-glucosidase genes to cold-induced oxidative stress in chickpea
1
2015
... 早在20多年前, 拟南芥XyG特异糖基化基因TCH4 (即AtXTH22)就已被证明非常耐(12-18)°C低温(
Xyloglucan isolated from suspension-cultured sycamore cell walls is O-acetylated
2
1989
... XyG是一种由D-吡喃葡萄糖残基以β-1→4键相连构成主链的杂多糖, 葡萄糖残基在O-6位上可被D-吡喃木糖残基所取代(
... -乙酰基取代基, 发生乙酰化(
Galactose-depleted xyloglucan is dysfunctional and leads to dwarfism in Arabidopsis
1
2015
... 大多数XyG木糖残基在GT47家族的一些酶作用下被其它糖基进一步取代, 其中最常见的是D-半乳糖.UDP-半乳糖在β-半乳糖基转移酶(β-galactosyltransferases, β-GAL)的作用下掺入XyG.例如, MUR3专一性负责XXXG第3位木糖残基的半乳糖化, 将XXXG变成XXLG (
Expression profiles and hormonal regulation of tobacco NtEXGT gene and its involvement in abiotic stress response
1
2017
... 近年来, 越来越多的研究表明, XyG代谢相关基因能够响应多种逆境或增强植物对多种逆境的抗性.例如, PeXTH基因同时具有提高胡杨耐盐及重金属的功能(
Transcriptome sequencing of chickpea ( Cicer arietinum L.) genotypes for identification of drought-responsive genes under drought stress condition
1
2019
... 研究表明, 植物XyG降解相关基因参与抗旱/涝过程.
Systematic analysis and functional validation of citrus XTH genes reveal the role of Csxth04 in citrus bacterial canker resistance and tolerance
1
2019
... 质外体组织是植物抵御病原菌的关键战场.病原菌分泌质外体XEGs降解植物XyG的主链, 相应地, 植物通过诱导表达其抑制蛋白(xyloglucan-speci?c endo- 1,4-glucanase inhibitor protein, XEGIP)的方式进行抵抗.这在苹果(Malus x domestica)和大豆等多种植物中得到证实.植物XEGIP通过与病原菌XEG结合从而使其失活, 进而抑制病原菌对植物细胞壁XyG的降解(
Biochemical and physiological characterization of fut4 and fut6 mutants defective in arabinogalactan-protein fucosylation in Arabidopsis
1
2013
... XXLG型XyG的半乳糖残基通常会被岩藻糖基进一步取代, 形成XXFG.XyG特异岩藻糖基转移酶(XyG: fucosyltransferases 1, FUT1)负责将GDP-岩藻糖添加到XXLG或XLLG的L上(
A xyloglucan backbone 6- O-acetyltransferase from Brachypodium distachyon modulates xyloglucan xylosylation
1
2016
... XyG的乙酰化主要发生在XXGG型XyG主链的葡萄糖残基及XXXG型XyG侧链的半乳糖残基上(
The presence of fucogalactoxyloglucan and its synthesis in rice indicates conserved functional importance in plants
1
2015
... 根据寡糖的结构可将XyG分成XXXG和XXGG两种主要类型.双子叶植物的XyG为XXXG型, 而禾本科和茄科植物的XyG为XXGG型.与XXXG型XyG相比, XXGG型XyG的木糖基化程度相对较低, 通常不发生岩藻糖基化(
Wound-induced pectin methylesterases enhance banana ( Musa spp. AAA) susceptibility to Fusarium oxysporum f. sp cubense
1
2013
... 植物在整个生命周期中经常遭受各种生物和非生物逆境胁迫.相应地, 植物采取多种防御策略来应对逆境, 包括改变细胞壁结构与组成、表达包括与木葡聚糖代谢相关基因在内的抗逆基因(
A paralogous decoy protects Phytophthora sojae apoplastic effector PsXEG1 from a host inhibitor
1
2017
... 质外体组织是植物抵御病原菌的关键战场.病原菌分泌质外体XEGs降解植物XyG的主链, 相应地, 植物通过诱导表达其抑制蛋白(xyloglucan-speci?c endo- 1,4-glucanase inhibitor protein, XEGIP)的方式进行抵抗.这在苹果(Malus x domestica)和大豆等多种植物中得到证实.植物XEGIP通过与病原菌XEG结合从而使其失活, 进而抑制病原菌对植物细胞壁XyG的降解(
Expression of the β-glucosidase gene Pgβglu-1 underpins natural resistance of white spruce against spruce budworm
1
2015
... 目前, XyG代谢与植物抗虫性的关系研究较少.芹菜(Apium graveolens) AgXTH1在桃蚜(Myzus persicae)侵染后系统性地在韧皮部中上调表达, 拟南芥中该基因的同源基因AtXTH33的突变体比野生型更吸引蚜虫, 表明AtXTH33调控拟南芥对蚜虫的抗性(
Loss-of-function mutation of REDUCED WALL ACETYLATION 2 in Arabidopsis leads to reduced cell wall acetylation and increased resistance to Botrytis cinerea
2
2011
... XyG的乙酰化主要发生在XXGG型XyG主链的葡萄糖残基及XXXG型XyG侧链的半乳糖残基上(
... 有研究表明, XyG乙酰化程度与植物的抗逆性相关.例如, 将真菌的鼠李糖半乳糖醛酸乙酰酯酶转入拟南芥后, 植株XyG的乙酰化程度显著降低, 转基因植株对灰葡萄孢菌(Botrytis cinerea)等真菌的抗性增强, 但不能增强对病原细菌的抗性(
Expression of heterologous xyloglucan xylosyltransferases in Arabidopsis to investigate their role in determi- ning xyloglucan xylosylation substitution patterns
1
2015
... 所有类型的XyG葡聚糖主链都含有木糖基取代基.XyG特异木糖基转移酶(XyG: xylosyltransferase, XXT)负责将UDP-木糖添加到主链葡萄糖残基O-6位上(
Acetylation of cell wall is required for structural integrity of the leaf surface and exerts a global impact on plant stress responses
1
2015
... 有研究表明, XyG乙酰化程度与植物的抗逆性相关.例如, 将真菌的鼠李糖半乳糖醛酸乙酰酯酶转入拟南芥后, 植株XyG的乙酰化程度显著降低, 转基因植株对灰葡萄孢菌(Botrytis cinerea)等真菌的抗性增强, 但不能增强对病原细菌的抗性(
Comparative digital gene expression analysis of tissue-cultured plantlets of highly resistant and susceptible banana cultivars in response to Fusarium oxysporum
3
2018
... 植物在整个生命周期中经常遭受各种生物和非生物逆境胁迫.相应地, 植物采取多种防御策略来应对逆境, 包括改变细胞壁结构与组成、表达包括与木葡聚糖代谢相关基因在内的抗逆基因(
... 质外体组织是植物抵御病原菌的关键战场.病原菌分泌质外体XEGs降解植物XyG的主链, 相应地, 植物通过诱导表达其抑制蛋白(xyloglucan-speci?c endo- 1,4-glucanase inhibitor protein, XEGIP)的方式进行抵抗.这在苹果(Malus x domestica)和大豆等多种植物中得到证实.植物XEGIP通过与病原菌XEG结合从而使其失活, 进而抑制病原菌对植物细胞壁XyG的降解(
... 早期人们发现, XyG降解产生的寡糖片段XGO可抑制烟草坏死病毒, 具有诱导抗性的功能(
Spatiotemporal changes in xylan-1/xyloglucan and xyloglucan xyloglucosyl transferase (XTH-Xet5) as a step-in of ultrastructural cell wall remodelling in potato-potato virus Y (PVYNTN) hypersensitive and susceptible reaction
1
2018
... 质外体组织是植物抵御病原菌的关键战场.病原菌分泌质外体XEGs降解植物XyG的主链, 相应地, 植物通过诱导表达其抑制蛋白(xyloglucan-speci?c endo- 1,4-glucanase inhibitor protein, XEGIP)的方式进行抵抗.这在苹果(Malus x domestica)和大豆等多种植物中得到证实.植物XEGIP通过与病原菌XEG结合从而使其失活, 进而抑制病原菌对植物细胞壁XyG的降解(
Biosynthesis of the plant cell wall matrix polysaccharide xyloglucan
3
2016
... 木葡聚糖(XyG)是双子叶植物初生壁中含量最为丰富的半纤维素成分, 约占细胞壁重量的20%-25%, 而禾本科植物中一般仅为2%-10% (
... XyG是一种由D-吡喃葡萄糖残基以β-1→4键相连构成主链的杂多糖, 葡萄糖残基在O-6位上可被D-吡喃木糖残基所取代(
... 根据寡糖的结构可将XyG分成XXXG和XXGG两种主要类型.双子叶植物的XyG为XXXG型, 而禾本科和茄科植物的XyG为XXGG型.与XXXG型XyG相比, XXGG型XyG的木糖基化程度相对较低, 通常不发生岩藻糖基化(
New insights into wall polysaccharide
1
2018
... XyG的乙酰化主要发生在XXGG型XyG主链的葡萄糖残基及XXXG型XyG侧链的半乳糖残基上(
Downregulation of RWA genes in hybrid aspen affects xylan acetylation and wood saccharification
1
2017
... XyG的乙酰化主要发生在XXGG型XyG主链的葡萄糖残基及XXXG型XyG侧链的半乳糖残基上(
A galacturonic acid-containing xyloglucan is involved in Arabi-dopsis root hair tip growth
1
2012
... 大多数XyG木糖残基在GT47家族的一些酶作用下被其它糖基进一步取代, 其中最常见的是D-半乳糖.UDP-半乳糖在β-半乳糖基转移酶(β-galactosyltransferases, β-GAL)的作用下掺入XyG.例如, MUR3专一性负责XXXG第3位木糖残基的半乳糖化, 将XXXG变成XXLG (
Arabidopsis and Brachypodium distachyon transgenic plants expressing Aspergillus nidu- lans acetylesterases have decreased degree of polysacc- haride acetylation and increased resistance to pathogens
1
2013
... 有研究表明, XyG乙酰化程度与植物的抗逆性相关.例如, 将真菌的鼠李糖半乳糖醛酸乙酰酯酶转入拟南芥后, 植株XyG的乙酰化程度显著降低, 转基因植株对灰葡萄孢菌(Botrytis cinerea)等真菌的抗性增强, 但不能增强对病原细菌的抗性(
The Arabidopsis TCH4 xyloglucan endotransglycosylase: substrate specificity, pH optimum, and cold tolerance
1
1997
... 早在20多年前, 拟南芥XyG特异糖基化基因TCH4 (即AtXTH22)就已被证明非常耐(12-18)°C低温(
Brassinosteroid mediated cell wall remodeling in grasses under abiotic stress
1
2017
... 早在20多年前, 拟南芥XyG特异糖基化基因TCH4 (即AtXTH22)就已被证明非常耐(12-18)°C低温(
The XTH family of enzymes involved in xyloglucan endotransglucosylation and endohydrolysis: current perspectives and a new unifying nomenclature
1
2002
... 植物XyG代谢相关基因与其抗盐性的研究多见于XTH基因家族.例如, 转辣椒(Capsicum annuum) CaXTH3的拟南芥耐盐性增强, 推测其通过调节气孔关闭来防止过度失水, 从而提高耐盐性(
Functional analysis of cellulose and xyloglucan in the walls of stomatal guard cells of Arabidopsis
1
2016
... 研究表明, 植物XyG降解相关基因参与抗旱/涝过程.
AtBGAL10 is the main xyloglucan β-galac-tosidase in Arabidopsis, and its absence results in unusual xyloglucan subunits and growth defects
1
2012
... 参与XyG支链降解的糖苷酶主要包括α-木糖苷酶(如AtXYL1/AXY3) (
Soluble and membrane-bound β-glucosidases are involved in trimming the xyloglucan backbone
0
2017
Hemicelluloses
2
2010
... 木葡聚糖(XyG)是双子叶植物初生壁中含量最为丰富的半纤维素成分, 约占细胞壁重量的20%-25%, 而禾本科植物中一般仅为2%-10% (
... XyG是一种由D-吡喃葡萄糖残基以β-1→4键相连构成主链的杂多糖, 葡萄糖残基在O-6位上可被D-吡喃木糖残基所取代(
The identification of two arabinosyltransferases from tomato reveals functional equivalency of xyloglucan side chain substituents
1
2013
... 大多数XyG木糖残基在GT47家族的一些酶作用下被其它糖基进一步取代, 其中最常见的是D-半乳糖.UDP-半乳糖在β-半乳糖基转移酶(β-galactosyltransferases, β-GAL)的作用下掺入XyG.例如, MUR3专一性负责XXXG第3位木糖残基的半乳糖化, 将XXXG变成XXLG (
The role of the plant-specific ALTERED XYLOGLU-CAN 9 protein in Arabidopsis cell wall polysaccharide O-acetylation
1
2015
... XyG的乙酰化主要发生在XXGG型XyG主链的葡萄糖残基及XXXG型XyG侧链的半乳糖残基上(
Xyloglucan endotransglycosylase/hydrolase genes from a susceptible and resistant jute species show opposite expression pattern following Macrophomina phaseolina infection
1
2012
... 质外体组织是植物抵御病原菌的关键战场.病原菌分泌质外体XEGs降解植物XyG的主链, 相应地, 植物通过诱导表达其抑制蛋白(xyloglucan-speci?c endo- 1,4-glucanase inhibitor protein, XEGIP)的方式进行抵抗.这在苹果(Malus x domestica)和大豆等多种植物中得到证实.植物XEGIP通过与病原菌XEG结合从而使其失活, 进而抑制病原菌对植物细胞壁XyG的降解(
α-xylosidase plays essential roles in xyloglucan remodelling, maintenance of cell wall integrity, and seed germination in Arabidopsis thaliana
1
2016
... 参与XyG支链降解的糖苷酶主要包括α-木糖苷酶(如AtXYL1/AXY3) (
The plant cell-wall enzyme AtXTH3 catalyses covalent cross-linking between cellulose and cello-oligosaccharide
1
2017
... 植物XyG内转糖苷酶/水解酶(endotransglycosylase/ hydrolases, XTHs)基因属于CAZy糖基水解酶16 (glycoside hydrolase 16, GH16)家族基因, 编码的蛋白可能具有2种不同的催化活性, 对XyG的影响不同.其中XyG特异内水解酶(XyG: endohydrolase, XEH)活性在形式上是XyG特异的内切葡聚糖酶(xyloglucan-specific endo-1,4-glucanases, XEGs), 降解XyG主链骨架(
Characterization of the XTH gene family: new insight to the roles in soybean flooding tolerance
1
2018
... 研究表明, 植物XyG降解相关基因参与抗旱/涝过程.
Inhibition of tobacco necrosis virus infection by xyloglucan fragments
1
1994
... 早期人们发现, XyG降解产生的寡糖片段XGO可抑制烟草坏死病毒, 具有诱导抗性的功能(
Both cold and sub-zero acclimation induce cell wall modification and changes in the extracellular proteome in Arabidopsis thaliana
1
2019
... 早在20多年前, 拟南芥XyG特异糖基化基因TCH4 (即AtXTH22)就已被证明非常耐(12-18)°C低温(
The SENSITIVE TO FREEZING 2 gene, required for freezing tolerance in Arabidopsis thaliana, encodes a β-glucosidase
1
2004
... 早在20多年前, 拟南芥XyG特异糖基化基因TCH4 (即AtXTH22)就已被证明非常耐(12-18)°C低温(
Cell wall integrity maintenance during plant development and interaction with the environment
1
2019
... 植物细胞壁是围绕在植物原生质体外的一种细胞结构, 由初生细胞壁和次生细胞壁组成.初生细胞壁位于外层, 是由纤维素、半纤维素和果胶等多糖及结构蛋白组成的一种复杂网络结构.次生细胞壁的结构与组成均与初生细胞壁有所不同, 除了纤维素和半纤维素外, 还含有木质素.植物细胞壁除了作为细胞的主要结构成分, 承担机械支撑、维持和决定细胞形态等功能外, 还在植物的生长发育(
The mur2 mutant of Arabidopsis thaliana lacks fucosylated xyloglucan because of a lesion in fucosyltransferase AtFUT1
1
2002
... XXLG型XyG的半乳糖残基通常会被岩藻糖基进一步取代, 形成XXFG.XyG特异岩藻糖基转移酶(XyG: fucosyltransferases 1, FUT1)负责将GDP-岩藻糖添加到XXLG或XLLG的L上(
Mutation in xyloglucan 6-xylosytransferase results in abnormal root hair development in Oryza sativa
1
2014
... 所有类型的XyG葡聚糖主链都含有木糖基取代基.XyG特异木糖基转移酶(XyG: xylosyltransferase, XXT)负责将UDP-木糖添加到主链葡萄糖残基O-6位上(
Genome-wide identification and expression profiling analysis of the xyloglucan endotransglucosylase/hydrolase gene family in tobacco ( Nicotiana tabacum L.)
1
2018
... 近年来, 越来越多的研究表明, XyG代谢相关基因能够响应多种逆境或增强植物对多种逆境的抗性.例如, PeXTH基因同时具有提高胡杨耐盐及重金属的功能(
Expression and distribution of extensins and AGPs in susceptible and resistant banana cultivars in response to wounding and Fusarium oxysporum
1
2017
... 植物在整个生命周期中经常遭受各种生物和非生物逆境胁迫.相应地, 植物采取多种防御策略来应对逆境, 包括改变细胞壁结构与组成、表达包括与木葡聚糖代谢相关基因在内的抗逆基因(
Immunohistochemical analysis of cell wall hydroxyproline-rich glycoproteins in the roots of resistant and susceptible wax gourd cultivars in response to Fusarium oxysporum f. sp
1
2011
... 植物在整个生命周期中经常遭受各种生物和非生物逆境胁迫.相应地, 植物采取多种防御策略来应对逆境, 包括改变细胞壁结构与组成、表达包括与木葡聚糖代谢相关基因在内的抗逆基因(
Genome-wide analysis of Sorghum GT47 family reveals functional divergences of MUR3-like genes
1
2018
... 大多数XyG木糖残基在GT47家族的一些酶作用下被其它糖基进一步取代, 其中最常见的是D-半乳糖.UDP-半乳糖在β-半乳糖基转移酶(β-galactosyltransferases, β-GAL)的作用下掺入XyG.例如, MUR3专一性负责XXXG第3位木糖残基的半乳糖化, 将XXXG变成XXLG (
Xyloglucan endotransglucosylase-hydrolase 30 negatively affects salt tolerance in Arabidopsis
1
2019
... 植物XyG代谢相关基因与其抗盐性的研究多见于XTH基因家族.例如, 转辣椒(Capsicum annuum) CaXTH3的拟南芥耐盐性增强, 推测其通过调节气孔关闭来防止过度失水, 从而提高耐盐性(
Variable content and distribution of arabinogalactan proteins in banana ( Musa spp.) under low temperature stress
1
2015
... 植物在整个生命周期中经常遭受各种生物和非生物逆境胁迫.相应地, 植物采取多种防御策略来应对逆境, 包括改变细胞壁结构与组成、表达包括与木葡聚糖代谢相关基因在内的抗逆基因(
BcXYG1, a secreted xyloglucanase from Botrytis cinerea, triggers both cell death and plant immune responses
1
2017
... 研究表明, 铝毒是由于Al3+与植物的细胞壁结合从而影响根系的伸长, 而XyG被认为是Al主要的结合位点(
XTH31, encoding an in vitro XEH/XET-active enzyme, regulates aluminum sensitivity by modulating in vivo XET action, cell wall xyloglucan content, and aluminum binding capacity in Arabidopsis
1
2012
... 研究表明, 铝毒是由于Al3+与植物的细胞壁结合从而影响根系的伸长, 而XyG被认为是Al主要的结合位点(

{kind=link}
{kind=link}