 ,*, 上官周平,*西北农林科技大学, 黄土高原土壤侵蚀与旱地农业国家重点实验室, 杨凌 712100
,*, 上官周平,*西北农林科技大学, 黄土高原土壤侵蚀与旱地农业国家重点实验室, 杨凌 712100Research Advances in the Main Ecological Functions of Root Exudates
Jiajia Li, Miaochun Fan,*, Zhouping Shangguan,*State Key Laboratory of Soil Erosion and Dryland Farming on the Loess Plateau, Northwest A&F University, Yangling 712100, China通讯作者: *E-mail:fanmc16@126.com;shangguan@ms.iswc.ac.cn
责任编辑: 白羽红
收稿日期:2020-03-5接受日期:2020-09-8网络出版日期:2020-11-01
| 基金资助: |
Corresponding authors: *E-mail:fanmc16@126.comshangguan@ms.iswc.ac.cn
Received:2020-03-5Accepted:2020-09-8Online:2020-11-01

摘要
根系分泌物在植物根系-土壤-微生物互作过程及其生态反馈机制中发挥重要作用。在植物根际复杂网络互作过程中, 根系分泌物被认为是“根际对话”的媒介, 其在调控植物适应微生境、缓解根际养分竞争及构建根际微生物群落结构方面意义重大。该文结合国内外该领域主要研究成果, 综述了根系分泌物对植物生长、土壤微生物特性及土壤养分循环的影响, 并展望了未来根系分泌物的研究方向。
关键词:
Abstract
Root exudates play an important role in root-soil-microorganism interactions and ecological feedback mechanisms. Root exudates are considered as the medium of “rhizosphere dialogue” in the process of complex plant rhizosphere network interaction, with great impact in regulating plant adaptation to microhabitats, thus alleviating rhizosphere nutrient competition and constructing rhizosphere microbial community structure. Here, we review the recent advances on the effects of root exudates on plant growth, soil microbial characteristics such as soil enzymes, microbial biomass, and microbial community, and soil nutrient circulation. We also propose the important directions and contents of future research on root exudates.
Keywords:
PDF (1141KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
李佳佳, 樊妙春, 上官周平. 植物根系分泌物主要生态功能研究进展. 植物学报, 2020, 55(6): 788-796 doi:10.11983/CBB20036
Li Jiajia, Fan Miaochun, Shangguan Zhouping.

植物在生长过程中通过根系主动或被动向根际释放的化合物被称为植物根系分泌物(root exudate)。根系分泌物的作用早在18世纪已被发现, 随后人们发现其可以促进或抑制周围植物生长(洪常青和聂艳丽, 2003)。近年来, 随着新技术和方法不断出现, 根系分泌物研究得以丰富和发展, 在选择作物种植方式、构建间种套作配置模式、生物入侵防控、生态恢复重建以及污染土壤的生物修复方面发挥重要作用。
根系分泌物可影响微生物生长, 调控土壤有机质分解和土壤氮(N)循环(Meier et al., 2017)。植物根系则可通过根系分泌物浓度的变化发出环境变化反馈信号, 调控根系生长以及碳(C)分配(Canarini et al., 2019)。植物-土壤互作调控根际过程释放的CO2约占陆地生态系统CO2总释放量的1/2 (Schimel, 1995; Cheng et al., 2013; Hopkins et al., 2013)。深入了解根系分泌物介导的植物-土壤互作过程, 有助于准确评估陆地生态系统碳储存和模型构建; 特别是在气候变化大背景下, 了解植物根系分泌物如何响应环境变化、评估根系分泌物对陆地生态系统地下碳循环的贡献是未来地下生态过程研究的重点和难点。因此, 根系分泌物及其生态功能的研究成为当今植物生态学、土壤生态学、植物营养学与全球变化生态学关注的热点。本文论述了根系分泌物输入土壤后对植物生长、土壤微生物特性以及土壤养分循环的影响, 以期厘清植物根系分泌物在植物根系-土壤-微生物互作过程中的作用机制, 从而明确未来植物根系分泌物的研究方向。
1 根系分泌物研究方法
根系分泌物是植物通过生物代谢(包括初生代谢和次生代谢, 其代谢产物分别为初生代谢物质和次生代谢物质)和非生物代谢产生的一些代谢产物。根系分泌物主要包括低分子化合物(脂肪酸和多糖等)、高分子黏胶物质(氨基酸、酚酸类化合物和有机酸类等)、无机离子(阴离子和阳离子, 如K+、Na+、NH4+、NO3-)及其它物质(磷酸酶和生长类物质等)。1.1 根系分泌物的收集
根系分泌物收集方法的研究起步较早, 不同的分离方法各有利弊。收集方法有水培收集、基质收集、吸附剂薄膜收集法、连续性根分泌收集系统法和根系分泌物收集培养箱等(王树起等, 2007)。水培法是将植物培养在水中, 待植物长大, 从水中提取根系分泌物, 该方法操作简单, 但是水培环境与自然条件差别较大。基质培养法是将植物培养在特定的固定基质中, 之后取出基质反复冲洗, 然后过滤基质收集根系分泌物, 该方法的不足之处是收集比较困难, 且难以保持根系分泌物的稳定性。吸附剂薄膜收集法和连续性根系分泌收集系统法都具有即时性。根系分泌物收集培养箱是将植物种植在特定培养箱中, 培养箱下设置一个根系分泌物收集装置, 该方法操作简便、即时, 但是取样空间小、重复性差且容易损伤根系(王占义等, 2010)。原位收集装置近年来被用于收集森林根系分泌物, 可以缩短抽滤时间和减少根系机械损伤, 操作简单, 无需移植植物(刘庆等, 2012; 王小平等, 2018)。目前, 在根系分泌物收集方法的时效性、稳定性和便利性等方面已有很大改进, 但不同根系形态和土壤特性的空间分布异质性很难保证在不损害根系的情况下获得正常的根系分泌物, 野外无菌条件更是很难保证, 不同分离方法对根系扰动和微生物干扰程度也不尽相同, 这给根系分泌物的收集带来不同程度的困难。
1.2 根系分泌物的提取和鉴定
除样品收集之外, 根系分泌物的提取也是根系分泌物研究的重要环节。通常情况下, 收集到的分泌物浓度很低且混有各种杂质, 必须进一步浓缩提取才可用于组分检测。常用的根系分泌物提取方法有离子交换法(ion exchange method)、萃取法(extraction method)、薄层层析法(thin layer chromatography)和超声浸提法(ultrasonic extraction)等。根系分泌物因植物种类不同而异, 适用方法也存在差异。石鑫(2012)通过超声波浸提、三性萃取、萃取硅烷化和萃取甲酯化4种方法分别提取刺槐(Robinia pseudoacacia)根系分泌物, 结果发现超声浸提法不能全面检测到刺槐根系分泌物组分, 其它方法则较为适合。核磁共振技术(nuclear magnetic resonance, NMR)、气相色谱-质谱(gas chromatography-mass spectrometry, GC/MS)、高效液相色谱(high performance liquid chromatography, HPLC)、离子色谱(ion chromatography, IC)和紫外可见光谱(ultraviolet- visible spectroscopy, UV-vis)等仪器已应用于植物及土壤代谢相关研究, 为代谢物组分系统性研究提供便利。张文明等(2015)采用液相色谱质谱联用仪(liquid chromatography mass spectrometry, LC/MS)对连作马铃薯(Solanum tuberosum)苗期和现蕾期根系分泌物进行鉴定, 发现苗期和现蕾期根系分泌物含有棕榈酸和邻苯二甲酸二丁酯。杨瑞秀等(2014)采用HPLC法对甜瓜(Cucumis melo)根系分泌物进行分离鉴定, 发现其含有没食子酸、邻苯二甲酸、丁香酸、水杨酸、苯甲酸和肉桂酸等酚酸类物质。Kumar等(2009)利用GC/MS在红树木榄(Bruguiera gymnorhiza)和海漆(Excoecaria agallocha)的根系分泌物中检出氨基嘧啶、棕榈酸及硬脂酸等。
尽管GC/MS和LC/MS技术已广泛应用于根系分泌物研究, 但也有不足之处。其忽略了含量低但非常重要的特异组分; 收集过程对根系生长影响比较大且在分离时容易被其它因素干扰, 目前尚无法实现对根系分泌物的动态监测(尹华军等, 2018)。因此开发原位、无菌收集方式是获得根系分泌物的前提。今后有望结合多组学及先进设备(如蛋白质组学、代谢组学、生物传感器和比率荧光化学传感器)建立高效完善的现代分析方法, 进而实现森林根系分泌物的定性定量分析(Fuhrer and Zamboni, 2015; Martinière et al., 2018)。
2 根系分泌物的生态功能
植物-植物、土壤-微生物、微生物-植物、微生物-微生物在植物根际形成复杂的网络关系。根系分泌物是其“交流”的媒介(吴林坤等, 2014), 在植物生长发育、提高植物抗性和适应能力方面发挥不可忽视的作用(图1)。图1
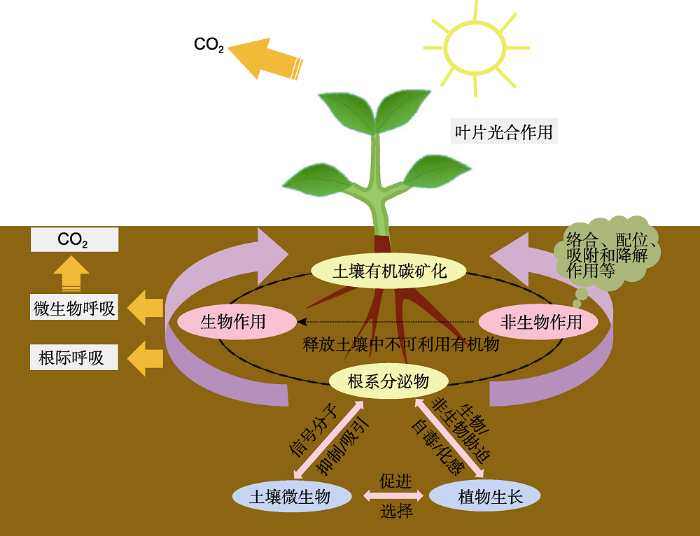 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1根系分泌物的主要生态功能
Figure 1Ecological functions of root exudates
2.1 根系分泌物对植物生长的影响
在根际周围, 许多生物因子和非生物因子交互作用,共同影响植物的生长发育。根系分泌物是调控根际生物和非生物互作的重要因子。根系分泌物作为化学信号或者化感物质, 在根际层面做出反应, 进而影响植物生长、微生物侵染以及根际微环境, 促使植物种间关系正向或负向发展(王树起等, 2007; 王建花等, 2013; 夏志超, 2017; 章芳芳, 2018)。首先, 植物可以通过化感/自毒作用(allelopathy/ autotoxicity)直接影响植物的共生关系或通过改变土壤生物特性来影响植物的生长(王建花等, 2013)。刘苹等(2010)证实花生(Arachis hypogaea)根系分泌物中存在自毒物质, 其对花生胚根生长具有抑制作用。杨树(Populus × euramericana)根系分泌的8种酚酸(苯甲酸、肉桂酸、阿魏酸、香草酸、香草醛、对羧基苯甲酸、异香草醛和香豆素)对土壤脲酶、蔗糖酶、碱性磷酸酶以及土壤微生物均有明显的自毒效应(陆茜, 2016)。白三叶(Trifolium repens)根系分泌的酚类化合物对各受体(黑麦草(Lolium perenne)、紫羊茅(Festuca rubra)、高羊茅(F. arundinacea)、草地早熟禾(Poa pratensis)和葡萄翦股颖(Agrostis stolonifera))幼苗根生长具有明显的抑制作用(余婷等, 2013)。以上研究表明, 植物根系分泌的一些化合物在植物种间产生负作用。
此外, 根系分泌物还可以通过直接或间接调控土壤中的生物和非生物因子促进植物的生长发育。短花针茅(Stipa breviflora)、无芒隐子草(Cleistogenes songorica)和冷蒿(Artemisia frigida) 3种荒漠草原优势植物根系分泌物的共同组分(1,2-二甲苯)在显著提高土壤速效氮磷和有机碳含量的同时, 能提高植物对土壤微环境的适应性(高雪峰, 2017)。拟南芥(Arabidopsis thaliana)根系分泌物影响根际微生物群落, 并通过产生IAA调节开花时间, 通过调控氮素利用来影响营养生长; 反之IAA也可以进一步促进植物生长和根系分泌物的释放(Lu et al., 2018)。研究表明, 植物根系分泌物可在根际招募有益菌群来抑制根际病原微生物, 从而进一步提高自身防御能力(周宝利等, 2011; 张文明等, 2015; 刘艳霞等, 2016)。小麦(Triticum aestvum)和玉米(Zea mays)根系释放苯并恶嗪酮, 该物质可促进茉莉酸信号物质释放而提高自身和子代防御能力(Hu et al., 2018)。感病拟南芥可通过改变根系分泌物组分调控根际微生物群落, 增强自身抗病能力(Yuan et al., 2018a)。
综上所述, 阐明根系分泌物对植物生长的影响机理可丰富植物种间化学通讯理论, 有助于解决植物持续多代种植过程中出现的品种退化和地力衰弱问题, 为林木可持续发展和多代经营提供科学依据。
2.2 根系分泌物对土壤微生物特性的影响
土壤微生物在土壤物质循环过程和生态系统服务中扮演着重要角色。根系分泌物和土壤微生物是根际微生态领域研究的重要方向, 开展根系分泌物与土壤微生物研究可推动植物根系与土壤互作调控机理的研究。目前, 根系分泌物对土壤微生物特性的影响主要为微生物生物量、微生物群落结构及土壤酶活性等方面。2.2.1 根系分泌物对土壤微生物生物量的影响
土壤微生物生物量是土壤环境变化的指示因子(马志良等, 2019), 是土壤有机碳组分中最活跃的部分, 对土壤养分转化和物质循环具有重要作用(Khan et al., 2016)。粗枝云杉(Picea asperata)根系分泌物对根际微生物生物量和根际微生物群组影响呈正效应(李娇等, 2014)。研究表明, 根系分泌物的分泌速率和微生物碳氮变化具有明显的季节动态, 两者呈正相关, 根际微生物活性和根际微生物生长代谢调控有赖于根系分泌物的释放(王小平等, 2018)。高浓度根系分泌物可通过刺激微生物生长(李杨等, 2019)和提高土壤微生物对底物碳源的利用能力(黄玉茜等, 2015)来提高土壤微生物生物量碳含量。
Du等(2020)发现, 玉米根系分泌物可增加邻苯二甲酯的生物利用性以及微生物碳氮含量, 且在不同品种间差异显著。根系分泌物与土壤微生物互作可能受多方面因素的影响。根系分泌物量(Eisenhauer et al., 2017)、土壤化学性质、土壤细菌和真菌自身的微生物生物量等都可能影响土壤微生物生物量对根系分泌物的响应(梁儒彪等, 2015)。关于根系分泌物对微生物生物量的影响目前尚缺乏系统性研究。
2.2.2 根系分泌物对微生物群落结构的影响
植物健康生长取决于自然宏观有机体和微观有机体互作(Sasse et al., 2018)。在根-土壤界面, 根系与根际微生物互作促进植物更好地适应环境(王孝林和王二涛, 2019)。例如, 有些植物可以通过根系分泌的特异信号分子独脚金内酯, 诱导丛枝菌根真菌分支向宿主靠近, 进而形成菌根共生体, 提高宿主对水分和磷元素的吸收(尚赏等, 2011)。
植物根系分泌有益于微生物生长的物质而吸引其在根际聚集(史刚荣, 2004), 而根系分泌物对土壤微生物群落影响具有选择性。短花针茅、无芒隐子草和冷蒿分泌的乙酰柠檬酸三丁酯、1,2-二甲苯和芥酸酰胺可以显著改变土壤微生物群落, 尤其是细菌群落组成(高雪峰, 2017)。玉米根系分泌的黏液也对根际细菌群落具有显著影响(Benizri et al., 2007)。研究表明, 土壤微生物对根系分泌物的偏好可驱动根际微生物群落构建(Zhalnina et al., 2018)。例如, 革兰氏阴性菌和真菌能快速同化根际碳(Paterson et al., 2007); 黑麦草根际真菌是根际碳利用的主要成员(Ku?lien? et al., 2014)。
根系分泌物作为植物和微生物协同进化的媒介(Morgan et al., 2005), 在调控根际微环境、促进植物应对各种生物或非生物胁迫中发挥重要作用。植物通过根系分泌物选择根际微生物群落(Broeckling et al., 2008); 反之, 根际微生物群落影响植物根系分泌物的释放(刘子雄等, 2005)。研究表明, 微生物种群结构的短暂变化, 由根系分泌物和环境选择压力驱动, 而选择压力来源于微生物种群的种内交换和种间迁移(Yuan et al., 2018b)。贾甜华(2019)通过分析根系分泌物与土壤微生物的相关性, 提出根系分泌物对土壤微生物的作用可能不是直接的, 可能由微生物之间的食物网决定, 但不排除植物选择的结果。
根系分泌物在介导植物-微生物互作过程中发挥重要作用, 但对其具体机制还知之甚少, 未来有望结合多组学或生物探针等方法进一步揭示植物根际的奥秘。
2.2.3 根系分泌物对土壤酶活性的影响
土壤酶参与土壤各种代谢过程和能量转化, 是土壤生态系统中物质循环和能量转化的生物催化剂。植物根系分泌物是土壤酶的主要来源之一(张丽莉等, 2004), 也可被微生物分解利用(孙悦等, 2014), 因此根系分泌物的输入可能会改变土壤酶的功能属性以及作用强度和方向。
关于植物根系分泌物对土壤酶活性的影响已有相关报道。小麦、油菜(Brassica napus)和白菜(B. chinensis)根系分泌物可缓解砷污染对土壤酶(脲酶、过氧化氢酶、酸性磷酸酶、脱氢酶和蔗糖酶)活性的抑制作用(李美璇, 2018); 甜菜(Beta vulgaris)根系分泌物可降低磷酸酶活性, 但提高土壤蔗糖酶活性; 玉米根系分泌物对磷酸酶活性的提高作用弱于对蔗糖酶(耿贵, 2011); 棉花(Gossypium hirsutum)根系分泌物可通过促进细菌和真菌的繁殖来提高土壤转化酶和磷酸酶活性(赵小亮等, 2009)。由此可见, 根系分泌物输入后会改变土壤酶活性, 进而影响土壤养分物质循环。此过程依赖于微生物代谢需求和土壤基质养分有效性调控(Caldwell, 2005)。Tian等(2019)发现土壤条件、根系分泌物以及枯落物共同影响土壤酶功能和枯落物分解过程, 这为探索根系分泌物介导土壤碳氮循环过程提供了新思路。
2.3 根系分泌物对土壤养分循环的影响
根系分泌物除了调节土壤微生物群落和土壤酶活性等生物活性外, 其在养分有效性(Mommer et al., 2016)及养分循环(Keiluweit et al., 2015)等方面也发挥重要作用。根系分泌物通过生物/非生物作用激发土壤-碳养分循环过程或根际激发效应(Cheng et al., 2014; 孙悦等, 2014; Zhu et al., 2014; 尹华军等, 2018)。我们通过以下几个理论来揭示根系分泌物介导的土壤-碳养分循环过程。微生物共代谢理论, 即根系分泌物输入为微生物生长发育提供了可利用能源, 从而使微生物区系发生变化(微生物活性和胞外酶增加), 进而促进土壤有机质的分解和养分循环(Wutzler and Reichstein, 2008; Zhu et al., 2014; Finzi et al., 2015)。调控闸门假设阐述了土壤中生物不可利用的有机物质可以经非生物调控(根系分泌物络合作用、溶解和有机质吸附等)转化为生物可利用的有机物质, 再经过微生物矿化分解(Kemmitt et al., 2008)。该假说体现了根系分泌物的生物和非生物作用。
根际碳矿化和养分循环过程中微生物和胞外酶活性受到根际氮有效性调控(Dijkstra et al., 2008; Drake et al., 2013; Sullivan and Hart, 2013)。微生物氮矿化假说和化学计量分解假说揭示了氮有效性对土壤有机质分解的影响(Craine et al., 2007)。微生物氮矿化假说认为氮缺乏表现正效应, 氮富余则相反(孙悦等, 2014)。化学计量分解假说认为只有基质碳氮满足微生物需求并与微生物碳氮比匹配时, 微生物活性和分解速率才最高(Hessen et al., 2004)。
以上假说均把根系分泌物作为一种外源碳, 微生物利用根际碳源促进有机质分解和根际无机碳氮释放。Shen等(2020)认为植物根系分泌物增加可能是对养分需求的响应, 研究发现草地退化使根系生物量和植物枯落物减少, 却使根系的输出率增加。由于退化草地根系周转对土壤碳输入贡献降低, 而根系分泌物对土壤碳输入的相对贡献增加, 因此, 草地退化过程中根系分泌物相对贡献的变化会影响草地土壤有机碳的形成和稳定。根系分泌物、根系周转和土壤微生物等是地下碳输入和输出过程的重要组成部分(肖春旺等, 2017)。量化根系分泌物输入对地下碳输入的贡献及其生物调控机制, 特别是在叠加气候变化、CO2升高和氮沉降等环境干扰因素下, 有助于进一步了解根系分泌物长期影响根周转率和土壤有机碳存储的作用机制。这将为精确探讨地下碳氮养分循环及其模型构建奠定理论基础。
3 研究展望
以往根际研究的特点是以生产实践问题为导向, 根系分泌物在调控根际环境中发挥的主要功能也集中在农业生态系统中。目前有关根系分泌物在森林生态系统中的重要作用与调控机制的研究较少, 一定程度上限制了对林木根系地下生态过程及其对生境变化响应的认识。因此, 强化森林根系分泌物-土壤微生物互作过程及其生态作用的研究, 已经成为全球气候变化下植物生态学与土壤生态学的重要内容。未来土壤生态学的研究方向将围绕以下几方面。(1) 森林根系分泌物-土壤-微生物系统协同反馈作用的潜在机制。植物根系分泌物的种类和数量变化是植物根系长期进化和对环境适应的结果, 并在根际区域形成一系列特殊根系和微生物功能性状, 具有重要的生态学功能和进化学意义。相应地, 在森林生态系统中了解森林根系分泌物演替变化和根际功能属性变化, 不仅对植被恢复有重要意义, 还可最大限度地发挥该系统的功能及合理定向调控; 并为揭示地下根际功能属性在种群分布和生物多样性中的作用提供新的研究思路。
(2) 季节变化对森林根系分泌物-土壤-微生物系统的影响。关于根系分泌物-土壤-微生物系统的研究主要集中在生长季节, 而关于根系分泌物的季节变化动态相关研究较为缺乏。例如, 不同生长季植物根系分泌物输入速率以及输入通量, 不同生长季节地上地下互作关系。随着新技术的发展, 通过多组学和多学科交叉等方法展开研究将有助于了解气候变化对植物根系分泌物以及根际微环境的影响, 以应对气候变化对植物生长的影响。
(3) 植物根系分泌物输入对土壤碳动态变化的调控机制。根系分泌物通过调控微生物活性和合成胞外酶的能力来调控土壤碳养分循环, 但是关于根系分泌物输入对碳代谢相关功能基因以及相关酶的调控还缺乏足够的了解。特别是土壤碳组分对根系分泌物C/N比值变化的敏感性以及根系分泌物输入对土壤碳的贡献, 未来可以通过宏基因组学和代谢组学等技术手段促进该领域研究。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1128/AEM.02188-07URLPMID:18083870 [本文引用: 1]

Plants are in constant contact with a community of soil biota that contains fungi ranging from pathogenic to symbiotic. A few studies have demonstrated a critical role of chemical communication in establishing highly specialized relationships, but the general role for root exudates in structuring the soil fungal community is poorly described. This study demonstrates that two model plant species (Arabidopsis thaliana and Medicago truncatula) are able to maintain resident soil fungal populations but unable to maintain nonresident soil fungal populations. This is mediated largely through root exudates: the effects of adding in vitro-generated root exudates to the soil fungal community were qualitatively and quantitatively similar to the results observed for plants grown in those same soils. This effect is observed for total fungal biomass, phylotype diversity, and overall community similarity to the starting community. Nonresident plants and root exudates influenced the fungal community by both positively and negatively impacting the relative abundance of individual phylotypes. A net increase in fungal biomass was observed when nonresident root exudates were added to resident plant treatments, suggesting that increases in specific carbon substrates and/or signaling compounds support an increased soil fungal population load. This study establishes root exudates as a mechanism through which a plant is able to regulate soil fungal community composition.
[本文引用: 1]
DOI:10.3389/fpls.2019.00157URLPMID:30881364 [本文引用: 1]
Root exudation is an important process determining plant interactions with the soil environment. Many studies have linked this process to soil nutrient mobilization. Yet, it remains unresolved how exudation is controlled and how exactly and under what circumstances plants benefit from exudation. The majority of root exudates including primary metabolites (sugars, amino acids, and organic acids) are believed to be passively lost from the root and used by rhizosphere-dwelling microbes. In this review, we synthetize recent advances in ecology and plant biology to explain and propose mechanisms by which root exudation of primary metabolites is controlled, and what role their exudation plays in plant nutrient acquisition strategies. Specifically, we propose a novel conceptual framework for root exudates. This framework is built upon two main concepts: (1) root exudation of primary metabolites is driven by diffusion, with plants and microbes both modulating concentration gradients and therefore diffusion rates to soil depending on their nutritional status; (2) exuded metabolite concentrations can be sensed at the root tip and signals are translated to modify root architecture. The flux of primary metabolites through root exudation is mostly located at the root tip, where the lack of cell differentiation favors diffusion of metabolites to the soil. We show examples of how the root tip senses concentration changes of exuded metabolites and translates that into signals to modify root growth. Plants can modify the concentration of metabolites either by controlling source/sink processes or by expressing and regulating efflux carriers, therefore challenging the idea of root exudation as a purely unregulated passive process. Through root exudate flux, plants can locally enhance concentrations of many common metabolites, which can serve as sensors and integrators of the plant nutritional status and of the nutrient availability in the surrounding environment. Plant-associated micro-organisms also constitute a strong sink for plant carbon, thereby increasing concentration gradients of metabolites and affecting root exudation. Understanding the mechanisms of and the effects that environmental stimuli have on the magnitude and type of root exudation will ultimately improve our knowledge of processes determining soil CO2 emissions, ecosystem functioning, and how to improve the sustainability of agricultural production.
URLPMID:23952258 [本文引用: 2]
URLPMID:17824441 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.envres.2020.109611URLPMID:32668551 [本文引用: 1]
Phthalic acid esters (PAEs) is a class of prevalent pollutants in agricultural soil, threating food safety through crop uptake and accumulation of PAEs. Accumulation of PAEs varies largely among crop species and cultivars. Nevertheless, how root exudates affect PAE bioavailability, dissipation, uptake and accumulation is still not well understood. In the present study, desorption and pot experiments were designed to investigate how root exudates from high-(Peizataifeng) and low-(Fengyousimiao) PAE accumulating rice cultivars affect soil PAE bioavailability, dissipation, and accumulation variation. Rice root exudates including low molecular weight organic acids (LMWOAs) of Peizataifeng and Fengyousimiao could enhance desorption of two typical PAE compounds, di-n-butyl phthalate (DBP) and di-(2-ethylhexyl) phthalate (DEHP), from aged soil to their available fractions by increasing soil dissolved organic carbon (DOC), thus improving their bioavailability in soil. Peizataifeng produced twice higher amounts of oxalic acid, critic acid and malonic acid in root exudates, and exhibited stronger effects on enhancing desorption and bioavailability of DBP and DEHP than Fengyousimiao. Higher (by about 50%) total organic carbon contents of root exudates from Peizataifeng led to higher (by 10-30%) soil microbial biomass carbon and nitrogen than Fengyousimiao, and thus promoted more PAE dissipation from soil than Fengyousimiao. Nevertheless, higher (by 20-50%) soil DOC and significantly higher PAE bioavailability in the soils planted Peizataifeng resulted in greater (by 53-93%) PAE accumulation in roots and shoots of Peizataifeng than Fengyousimiao, confirming by higher (by 1.82-3.48 folds) shoot and root bioconcentration factors of Peizataifeng than Fengyousimiao. This study reveals that the difference in root exudate extent and LMWOAs between Peizataifeng and Fengyousimiao differentiates PAE accumulation.
URLPMID:28374800 [本文引用: 1]
[本文引用: 1]
URLPMID:25197792 [本文引用: 1]
[本文引用: 1]
URLPMID:23943914 [本文引用: 1]
URLPMID:30013066 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.geoderma.2016.02.019URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1186/s40168-018-0615-0URLPMID:30587246 [本文引用: 1]
BACKGROUND: Plant phenology has crucial biological, physical, and chemical effects on the biosphere. Phenological drivers have largely been studied, but the role of plant microbiota, particularly rhizosphere microbiota, has not been considered. RESULTS: We discovered that rhizosphere microbial communities could modulate the timing of flowering of Arabidopsis thaliana. Rhizosphere microorganisms that increased and prolonged N bioavailability by nitrification delayed flowering by converting tryptophan to the phytohormone indole acetic acid (IAA), thus downregulating genes that trigger flowering, and stimulating further plant growth. The addition of IAA to hydroponic cultures confirmed this metabolic network. CONCLUSIONS: We document a novel metabolic network in which soil microbiota influenced plant flowering time, thus shedding light on the key role of soil microbiota on plant functioning. This opens up multiple opportunities for application, from helping to mitigate some of the effects of climate change and environmental stress on plants (e.g. abnormal temperature variation, drought, salinity) to manipulating plant characteristics using microbial inocula to increase crop potential.
DOI:10.1073/pnas.1721769115URLPMID:29866831 [本文引用: 1]
Building a proton gradient across a biological membrane and between different tissues is a matter of great importance for plant development and nutrition. To gain a better understanding of proton distribution in the plant root apoplast as well as across the plasma membrane, we generated Arabidopsis plants expressing stable membrane-anchored ratiometric fluorescent sensors based on pHluorin. These sensors enabled noninvasive pH-specific measurements in mature root cells from the medium-epidermis interface up to the inner cell layers that lie beyond the Casparian strip. The membrane-associated apoplastic pH was much more alkaline than the overall apoplastic space pH. Proton concentration associated with the plasma membrane was very stable, even when the growth medium pH was altered. This is in apparent contradiction with the direct connection between root intercellular space and the external medium. The plasma membrane-associated pH in the stele was the most preserved and displayed the lowest apoplastic pH (6.0 to 6.1) and the highest transmembrane delta pH (1.5 to 2.2). Both pH values also correlated well with optimal activities of channels and transporters involved in ion uptake and redistribution from the root to the aerial part. In growth medium where ionic content is minimized, the root plasma membrane-associated pH was more affected by environmental proton changes, especially for the most external cell layers. Calcium concentration appears to play a major role in apoplastic pH under these restrictive conditions, supporting a role for the cell wall in pH homeostasis of the unstirred surface layer of plasma membrane in mature roots.
[本文引用: 1]
URLPMID:26832947 [本文引用: 1]
[本文引用: 1]
DOI:10.1111/j.1469-8137.2006.01929.xURLPMID:17096787 [本文引用: 1]
DOI:10.1016/j.tplants.2017.09.003URL [本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.soilbio.2020.107817URL [本文引用: 1]
DOI:10.1016/j.soilbio.2012.12.007URL [本文引用: 1]
Soil organic matter (SUM) decomposition has the potential to radically affect carbon dioxide concentrations in the atmosphere. Priming, the increased decomposition of SUM after the addition of a labile carbon (C) source, may be an important regulator of SUM dynamics, yet little is known about the mechanisms of the priming effect. Two hypotheses generated in the last decade have suggested that priming is caused by either the nutrient conditions in soil or the response of the microbial community to labile C addition. We used a three million year substrate age gradient, with associated changes in nutrient availability and microbial communities, to test these two hypotheses. We added C-13 labeled glucose to soil in quantities similar to increases in root exudation that can be expected in a high carbon dioxide environment, and traced the effect of C addition on soil C, the microbial community, and soil nutrient pools and fluxes. We observed positive priming, negative priming, and no net priming depending on substrate age. Priming was most positive at the youngest sites with the smallest nitrogen (N) pools, and most negative at the site with the most available N. In contrast, we found no significant relationships between priming and phosphorus availability. Though components of the microbial community size and structure (measured by phospholipid fatty acid analysis) changed as a result of C addition, soil N availability was a better explanatory mechanism of priming effects than microbial community dynamics. Our results suggest close linkages between C and N cycles regulate the magnitude and direction of priming. If general, the stability of SUM in temperate ecosystems is likely to be governed by soil N status. (C) 2013 Elsevier Ltd.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1186/s40168-018-0537-xURLPMID:30208962 [本文引用: 1]
6) and lower exudation levels for sugars, alcohols, and short-chain organic acids (SCOAs) (C
DOI:10.1038/s41564-018-0129-3URLPMID:29556109 [本文引用: 1]
Like all higher organisms, plants have evolved in the context of a microbial world, shaping both their evolution and their contemporary ecology. Interactions between plant roots and soil microorganisms are critical for plant fitness in natural environments. Given this co-evolution and the pivotal importance of plant-microbial interactions, it has been hypothesized, and a growing body of literature suggests, that plants may regulate the composition of their rhizosphere to promote the growth of microorganisms that improve plant fitness in a given ecosystem. Here, using a combination of comparative genomics and exometabolomics, we show that pre-programmed developmental processes in plants (Avena barbata) result in consistent patterns in the chemical composition of root exudates. This chemical succession in the rhizosphere interacts with microbial metabolite substrate preferences that are predictable from genome sequences. Specifically, we observed a preference by rhizosphere bacteria for consumption of aromatic organic acids exuded by plants (nicotinic, shikimic, salicylic, cinnamic and indole-3-acetic). The combination of these plant exudation traits and microbial substrate uptake traits interact to yield the patterns of microbial community assembly observed in the rhizosphere of an annual grass. This discovery provides a mechanistic underpinning for the process of rhizosphere microbial community assembly and provides an attractive direction for the manipulation of the rhizosphere microbiome for beneficial outcomes.
DOI:10.1016/j.soilbio.2014.04.033URL [本文引用: 2]
短花针茅荒漠草原优势植物根系分泌物及其主要组分对土壤微生物的影响
2
2017
... 此外, 根系分泌物还可以通过直接或间接调控土壤中的生物和非生物因子促进植物的生长发育.短花针茅(Stipa breviflora)、无芒隐子草(Cleistogenes songorica)和冷蒿(Artemisia frigida) 3种荒漠草原优势植物根系分泌物的共同组分(1,2-二甲苯)在显著提高土壤速效氮磷和有机碳含量的同时, 能提高植物对土壤微环境的适应性(
... 植物根系分泌有益于微生物生长的物质而吸引其在根际聚集(
作物根系分泌物对土壤碳、氮含量、微生物数量和酶活性的影响
1
2011
... 关于植物根系分泌物对土壤酶活性的影响已有相关报道.小麦、油菜(Brassica napus)和白菜(B. chinensis)根系分泌物可缓解砷污染对土壤酶(脲酶、过氧化氢酶、酸性磷酸酶、脱氢酶和蔗糖酶)活性的抑制作用(
根系分泌物及其在植物营养中的作用
1
2003
... 植物在生长过程中通过根系主动或被动向根际释放的化合物被称为植物根系分泌物(root exudate).根系分泌物的作用早在18世纪已被发现, 随后人们发现其可以促进或抑制周围植物生长(
花生根系分泌物对土壤微生物学特性及群落功能多样性的影响
1
2015
... 土壤微生物生物量是土壤环境变化的指示因子(
三江源区退化高寒草地毒杂草植物根际微生物与根系分泌物的变化及相关性分析
1
2019
... 根系分泌物作为植物和微生物协同进化的媒介(
不同林龄云杉人工林的根系分泌物与土壤微生物
1
2014
... 土壤微生物生物量是土壤环境变化的指示因子(
根系分泌物对砷污染土壤中砷酸还原菌存活效应的影响
1
2018
... 关于植物根系分泌物对土壤酶活性的影响已有相关报道.小麦、油菜(Brassica napus)和白菜(B. chinensis)根系分泌物可缓解砷污染对土壤酶(脲酶、过氧化氢酶、酸性磷酸酶、脱氢酶和蔗糖酶)活性的抑制作用(
不同浓度和多样性的根系分泌物对土壤团聚体稳定性的影响
1
2019
... 土壤微生物生物量是土壤环境变化的指示因子(
模拟根系分泌物C:N化学计量特征对川西亚高山森林土壤碳动态和微生物群落结构的影响
1
2015
...
花生根系分泌物自毒作用研究
1
2010
... 首先, 植物可以通过化感/自毒作用(allelopathy/ autotoxicity)直接影响植物的共生关系或通过改变土壤生物特性来影响植物的生长(
一种根系分泌物原位收集装置及系统
1
2012
... 根系分泌物收集方法的研究起步较早, 不同的分离方法各有利弊.收集方法有水培收集、基质收集、吸附剂薄膜收集法、连续性根分泌收集系统法和根系分泌物收集培养箱等(
烟草根系分泌物酚酸类物质的鉴定及其对根际微生物的影响
1
2016
... 此外, 根系分泌物还可以通过直接或间接调控土壤中的生物和非生物因子促进植物的生长发育.短花针茅(Stipa breviflora)、无芒隐子草(Cleistogenes songorica)和冷蒿(Artemisia frigida) 3种荒漠草原优势植物根系分泌物的共同组分(1,2-二甲苯)在显著提高土壤速效氮磷和有机碳含量的同时, 能提高植物对土壤微环境的适应性(
林木根系分泌物与根际微生物研究进展
1
2005
... 根系分泌物作为植物和微生物协同进化的媒介(
杨树连栽对根际土壤环境演变的影响及其自毒效应研究
1
2016
... 首先, 植物可以通过化感/自毒作用(allelopathy/ autotoxicity)直接影响植物的共生关系或通过改变土壤生物特性来影响植物的生长(
增温对高寒灌丛根际和非根际土壤微生物生物量碳氮的影响
1
2019
... 土壤微生物生物量是土壤环境变化的指示因子(
丛枝菌根形成过程及其信号转导途径
1
2011
... 植物健康生长取决于自然宏观有机体和微观有机体互作(
植物根系分泌物的生态效应
1
2004
... 植物根系分泌有益于微生物生长的物质而吸引其在根际聚集(
刺槐根系分泌物研究
1
2012
... 除样品收集之外, 根系分泌物的提取也是根系分泌物研究的重要环节.通常情况下, 收集到的分泌物浓度很低且混有各种杂质, 必须进一步浓缩提取才可用于组分检测.常用的根系分泌物提取方法有离子交换法(ion exchange method)、萃取法(extraction method)、薄层层析法(thin layer chromatography)和超声浸提法(ultrasonic extraction)等.根系分泌物因植物种类不同而异, 适用方法也存在差异.
根际激发效应的发生机制及其生态重要性
3
2014
... 土壤酶参与土壤各种代谢过程和能量转化, 是土壤生态系统中物质循环和能量转化的生物催化剂.植物根系分泌物是土壤酶的主要来源之一(
... 根系分泌物除了调节土壤微生物群落和土壤酶活性等生物活性外, 其在养分有效性(
... 根际碳矿化和养分循环过程中微生物和胞外酶活性受到根际氮有效性调控(
植物化感作用类型及其在农业中的应用
2
2013
... 在根际周围, 许多生物因子和非生物因子交互作用,共同影响植物的生长发育.根系分泌物是调控根际生物和非生物互作的重要因子.根系分泌物作为化学信号或者化感物质, 在根际层面做出反应, 进而影响植物生长、微生物侵染以及根际微环境, 促使植物种间关系正向或负向发展(
... 首先, 植物可以通过化感/自毒作用(allelopathy/ autotoxicity)直接影响植物的共生关系或通过改变土壤生物特性来影响植物的生长(
根系分泌物的化感作用及其对土壤微生物的影响
2
2007
... 根系分泌物收集方法的研究起步较早, 不同的分离方法各有利弊.收集方法有水培收集、基质收集、吸附剂薄膜收集法、连续性根分泌收集系统法和根系分泌物收集培养箱等(
... 在根际周围, 许多生物因子和非生物因子交互作用,共同影响植物的生长发育.根系分泌物是调控根际生物和非生物互作的重要因子.根系分泌物作为化学信号或者化感物质, 在根际层面做出反应, 进而影响植物生长、微生物侵染以及根际微环境, 促使植物种间关系正向或负向发展(
根际微生物促进水稻氮利用的机制
1
2019
... 植物健康生长取决于自然宏观有机体和微观有机体互作(
连香树人工林根系分泌物输入季节性变化及其驱动的根际微生物特性研究
2
2018
... 根系分泌物收集方法的研究起步较早, 不同的分离方法各有利弊.收集方法有水培收集、基质收集、吸附剂薄膜收集法、连续性根分泌收集系统法和根系分泌物收集培养箱等(
... 土壤微生物生物量是土壤环境变化的指示因子(
一种新型根系分泌物收集装置与收集方法的介绍
1
2010
... 根系分泌物收集方法的研究起步较早, 不同的分离方法各有利弊.收集方法有水培收集、基质收集、吸附剂薄膜收集法、连续性根分泌收集系统法和根系分泌物收集培养箱等(
根系分泌物介导下植物-土壤-微生物互作关系研究进展与展望
1
2014
... 植物-植物、土壤-微生物、微生物-植物、微生物-微生物在植物根际形成复杂的网络关系.根系分泌物是其“交流”的媒介(
根系分泌物介导的植物种间地下化学作用
1
2017
... 在根际周围, 许多生物因子和非生物因子交互作用,共同影响植物的生长发育.根系分泌物是调控根际生物和非生物互作的重要因子.根系分泌物作为化学信号或者化感物质, 在根际层面做出反应, 进而影响植物生长、微生物侵染以及根际微环境, 促使植物种间关系正向或负向发展(
陆地生态系统地下碳输入与输出过程研究进展
1
2017
... 以上假说均把根系分泌物作为一种外源碳, 微生物利用根际碳源促进有机质分解和根际无机碳氮释放.
甜瓜根系分泌物中酚酸物质对尖孢镰孢菌的化感效应
1
2014
... 核磁共振技术(nuclear magnetic resonance, NMR)、气相色谱-质谱(gas chromatography-mass spectrometry, GC/MS)、高效液相色谱(high performance liquid chromatography, HPLC)、离子色谱(ion chromatography, IC)和紫外可见光谱(ultraviolet- visible spectroscopy, UV-vis)等仪器已应用于植物及土壤代谢相关研究, 为代谢物组分系统性研究提供便利.
森林根系分泌物生态学研究: 问题与展望
2
2018
... 尽管GC/MS和LC/MS技术已广泛应用于根系分泌物研究, 但也有不足之处.其忽略了含量低但非常重要的特异组分; 收集过程对根系生长影响比较大且在分离时容易被其它因素干扰, 目前尚无法实现对根系分泌物的动态监测(
... 根系分泌物除了调节土壤微生物群落和土壤酶活性等生物活性外, 其在养分有效性(
白三叶根系分泌物对5种草坪草的化感作用
1
2013
... 首先, 植物可以通过化感/自毒作用(allelopathy/ autotoxicity)直接影响植物的共生关系或通过改变土壤生物特性来影响植物的生长(
根系分泌物对间作体系种间根系相互作用的调控及其关键成分研究
1
2018
... 在根际周围, 许多生物因子和非生物因子交互作用,共同影响植物的生长发育.根系分泌物是调控根际生物和非生物互作的重要因子.根系分泌物作为化学信号或者化感物质, 在根际层面做出反应, 进而影响植物生长、微生物侵染以及根际微环境, 促使植物种间关系正向或负向发展(
稻-麦轮作系统土壤糖酶活性对开放式CO2浓度增高的响应
1
2004
... 土壤酶参与土壤各种代谢过程和能量转化, 是土壤生态系统中物质循环和能量转化的生物催化剂.植物根系分泌物是土壤酶的主要来源之一(
马铃薯根系分泌物成分鉴别及其对立枯丝核菌的影响
2
2015
... 核磁共振技术(nuclear magnetic resonance, NMR)、气相色谱-质谱(gas chromatography-mass spectrometry, GC/MS)、高效液相色谱(high performance liquid chromatography, HPLC)、离子色谱(ion chromatography, IC)和紫外可见光谱(ultraviolet- visible spectroscopy, UV-vis)等仪器已应用于植物及土壤代谢相关研究, 为代谢物组分系统性研究提供便利.
... 此外, 根系分泌物还可以通过直接或间接调控土壤中的生物和非生物因子促进植物的生长发育.短花针茅(Stipa breviflora)、无芒隐子草(Cleistogenes songorica)和冷蒿(Artemisia frigida) 3种荒漠草原优势植物根系分泌物的共同组分(1,2-二甲苯)在显著提高土壤速效氮磷和有机碳含量的同时, 能提高植物对土壤微环境的适应性(
棉花根系分泌物对土壤速效养分和酶活性及微生物数量的影响
1
2009
... 关于植物根系分泌物对土壤酶活性的影响已有相关报道.小麦、油菜(Brassica napus)和白菜(B. chinensis)根系分泌物可缓解砷污染对土壤酶(脲酶、过氧化氢酶、酸性磷酸酶、脱氢酶和蔗糖酶)活性的抑制作用(
嫁接茄子根系分泌物变化及其对黄萎菌的影响
1
2011
... 此外, 根系分泌物还可以通过直接或间接调控土壤中的生物和非生物因子促进植物的生长发育.短花针茅(Stipa breviflora)、无芒隐子草(Cleistogenes songorica)和冷蒿(Artemisia frigida) 3种荒漠草原优势植物根系分泌物的共同组分(1,2-二甲苯)在显著提高土壤速效氮磷和有机碳含量的同时, 能提高植物对土壤微环境的适应性(
Additions of maize root mucilage to soil changed the structure of the bacterial community
1
2007
... 植物根系分泌有益于微生物生长的物质而吸引其在根际聚集(
Root exudates regulate soil fungal community composition and diversity
1
2008
... 根系分泌物作为植物和微生物协同进化的媒介(
Enzyme activities as a component of soil biodiversity: a review
1
2005
... 关于植物根系分泌物对土壤酶活性的影响已有相关报道.小麦、油菜(Brassica napus)和白菜(B. chinensis)根系分泌物可缓解砷污染对土壤酶(脲酶、过氧化氢酶、酸性磷酸酶、脱氢酶和蔗糖酶)活性的抑制作用(
Root exudation of primary metabolites: mechanisms and their roles in plant responses to environmental stimuli
1
2019
... 根系分泌物可影响微生物生长, 调控土壤有机质分解和土壤氮(N)循环(
Synthesis and modeling perspectives of rhizosphere priming
2
2014
... 根系分泌物可影响微生物生长, 调控土壤有机质分解和土壤氮(N)循环(
... 根系分泌物除了调节土壤微生物群落和土壤酶活性等生物活性外, 其在养分有效性(
Microbial nitrogen limitation increases decomposition
1
2007
... 根际碳矿化和养分循环过程中微生物和胞外酶活性受到根际氮有效性调控(
Long-term enhancement of N availability and plant growth under elevated CO2 in a semi-arid grassland
1
2008
... 根际碳矿化和养分循环过程中微生物和胞外酶活性受到根际氮有效性调控(
Stoichiometry constrains microbial response to root exudation-insights from a model and a field experiment in a temperate forest
1
2013
... 根际碳矿化和养分循环过程中微生物和胞外酶活性受到根际氮有效性调控(
Rice root exudates enhance desorption and bioavailability of phthalic acid esters (PAEs) in soil associating with cultivar variation in PAE accumulation
1
2020
...
Root biomass and exudates link plant diversity with soil bacterial and fungal biomass
1
2017
...
Rhizosphere processes are quantitatively important components of terrestrial carbon and nutrient cycles
1
2015
... 微生物共代谢理论, 即根系分泌物输入为微生物生长发育提供了可利用能源, 从而使微生物区系发生变化(微生物活性和胞外酶增加), 进而促进土壤有机质的分解和养分循环(
High-throughput discovery metabolomics
1
2015
... 尽管GC/MS和LC/MS技术已广泛应用于根系分泌物研究, 但也有不足之处.其忽略了含量低但非常重要的特异组分; 收集过程对根系生长影响比较大且在分离时容易被其它因素干扰, 目前尚无法实现对根系分泌物的动态监测(
Carbon sequestration in ecosystems: the role of stoichiometry
1
2004
... 根际碳矿化和养分循环过程中微生物和胞外酶活性受到根际氮有效性调控(
Ecosystem- level controls on root-rhizosphere respiration
1
2013
... 根系分泌物可影响微生物生长, 调控土壤有机质分解和土壤氮(N)循环(
Root exudate metabolites drive plant-soil feedbacks on growth and defense by shaping the rhizosphere microbiota
1
2018
... 此外, 根系分泌物还可以通过直接或间接调控土壤中的生物和非生物因子促进植物的生长发育.短花针茅(Stipa breviflora)、无芒隐子草(Cleistogenes songorica)和冷蒿(Artemisia frigida) 3种荒漠草原优势植物根系分泌物的共同组分(1,2-二甲苯)在显著提高土壤速效氮磷和有机碳含量的同时, 能提高植物对土壤微环境的适应性(
Mineral protection of soil carbon counteracted by root exudates
1
2015
... 根系分泌物除了调节土壤微生物群落和土壤酶活性等生物活性外, 其在养分有效性(
Mineralization of native soil organic matter is not regulated by the size, activity or composition of the soil microbial biomass—a new perspective
1
2008
... 微生物共代谢理论, 即根系分泌物输入为微生物生长发育提供了可利用能源, 从而使微生物区系发生变化(微生物活性和胞外酶增加), 进而促进土壤有机质的分解和养分循环(
Microbial biomass, fungal and bacterial residues, and their relationships to the soil organic matter C/N/P/S ratios
1
2016
... 土壤微生物生物量是土壤环境变化的指示因子(
Preliminary GC-MS analysis of compounds present in the root exudates of three mangrove species
1
2009
... 核磁共振技术(nuclear magnetic resonance, NMR)、气相色谱-质谱(gas chromatography-mass spectrometry, GC/MS)、高效液相色谱(high performance liquid chromatography, HPLC)、离子色谱(ion chromatography, IC)和紫外可见光谱(ultraviolet- visible spectroscopy, UV-vis)等仪器已应用于植物及土壤代谢相关研究, 为代谢物组分系统性研究提供便利.
Medium-term response of microbial community to rhizodeposits of white clover and ryegrass and tracing of active processes induced by 13C and 15N labelled exudates
1
2014
... 植物根系分泌有益于微生物生长的物质而吸引其在根际聚集(
Rhizosphere microorganisms can influence the timing of plant flowering
1
2018
... 此外, 根系分泌物还可以通过直接或间接调控土壤中的生物和非生物因子促进植物的生长发育.短花针茅(Stipa breviflora)、无芒隐子草(Cleistogenes songorica)和冷蒿(Artemisia frigida) 3种荒漠草原优势植物根系分泌物的共同组分(1,2-二甲苯)在显著提高土壤速效氮磷和有机碳含量的同时, 能提高植物对土壤微环境的适应性(
Uncovering pH at both sides of the root plasma membrane interface using noninvasive imaging
1
2018
... 尽管GC/MS和LC/MS技术已广泛应用于根系分泌物研究, 但也有不足之处.其忽略了含量低但非常重要的特异组分; 收集过程对根系生长影响比较大且在分离时容易被其它因素干扰, 目前尚无法实现对根系分泌物的动态监测(
Root exudates increase N availability by stimulating microbial turnover of fast-cycling N pools
1
2017
... 根系分泌物可影响微生物生长, 调控土壤有机质分解和土壤氮(N)循环(
Root-root interactions: towards a rhizosphere framework
1
2016
... 根系分泌物除了调节土壤微生物群落和土壤酶活性等生物活性外, 其在养分有效性(
Biological costs and benefits to plant-microbe interactions in the rhizosphere
1
2005
... 根系分泌物作为植物和微生物协同进化的媒介(
Rhizodeposition shapes rhizosphere microbial community structure in organic soil
1
2007
... 植物根系分泌有益于微生物生长的物质而吸引其在根际聚集(
Feed your friends: do plant exudates shape the root microbiome?
1
2018
... 植物健康生长取决于自然宏观有机体和微观有机体互作(
Terrestrial ecosystems and the carbon cycle
1
1995
... 根系分泌物可影响微生物生长, 调控土壤有机质分解和土壤氮(N)循环(
Increased contribution of root exudates to soil carbon input during grassland degradation
1
2020
... 以上假说均把根系分泌物作为一种外源碳, 微生物利用根际碳源促进有机质分解和根际无机碳氮释放.
Evaluation of mechanisms controlling the priming of soil carbon along a substrate age gradient
1
2013
... 根际碳矿化和养分循环过程中微生物和胞外酶活性受到根际氮有效性调控(
Priming effect of litter mineralization: the role of root exudate depends on its interactions with litter quality and soil condition
1
2019
... 关于植物根系分泌物对土壤酶活性的影响已有相关报道.小麦、油菜(Brassica napus)和白菜(B. chinensis)根系分泌物可缓解砷污染对土壤酶(脲酶、过氧化氢酶、酸性磷酸酶、脱氢酶和蔗糖酶)活性的抑制作用(
Priming and substrate quality interactions in soil organic matter models
1
2013
... 微生物共代谢理论, 即根系分泌物输入为微生物生长发育提供了可利用能源, 从而使微生物区系发生变化(微生物活性和胞外酶增加), 进而促进土壤有机质的分解和养分循环(
Root exudates dominate the colonization of pathogen and plant growth-promoting rhizobacteria
1
2018a
... 此外, 根系分泌物还可以通过直接或间接调控土壤中的生物和非生物因子促进植物的生长发育.短花针茅(Stipa breviflora)、无芒隐子草(Cleistogenes songorica)和冷蒿(Artemisia frigida) 3种荒漠草原优势植物根系分泌物的共同组分(1,2-二甲苯)在显著提高土壤速效氮磷和有机碳含量的同时, 能提高植物对土壤微环境的适应性(
Root exudates drive the soil-borne legacy of aboveground pathogen infection
1
2018b
... 根系分泌物作为植物和微生物协同进化的媒介(
Dynamic root exudate chemistry and microbial substrate preferences drive patterns in rhizosphere microbial community assembly
1
2018
... 植物根系分泌有益于微生物生长的物质而吸引其在根际聚集(
Rhizosphere priming effects on soil carbon and nitrogen mineralization
2
2014
... 根系分泌物除了调节土壤微生物群落和土壤酶活性等生物活性外, 其在养分有效性(
... 微生物共代谢理论, 即根系分泌物输入为微生物生长发育提供了可利用能源, 从而使微生物区系发生变化(微生物活性和胞外酶增加), 进而促进土壤有机质的分解和养分循环(

{kind=link}
{kind=link}