Effects of Epigenetic Mechanisms on C4Phosphoenolpyruvate Carboxylase Transgenic Rice (Oryza sativa) Seed Germination Under Drought Stress
Ningxi Song1,2, Yingfeng Xie2, Xia Li1,2,3,*通讯作者: *E-mail:jspplx@jaas.ac.cn
责任编辑: 朱亚娜
收稿日期:2020-03-21接受日期:2020-08-26网络出版日期:2020-11-01
| 基金资助: |
Corresponding authors: *E-mail:jspplx@jaas.ac.cn
Editor:
Received:2020-03-21Accepted:2020-08-26Online:2020-11-01

摘要
为探究干旱胁迫下表观遗传机制对高表达玉米(Zea mays) C4型PEPC转基因水稻(Oryza sativa)种子萌发的影响, 以转C4型PEPC水稻(PC)和野生型水稻Kitaake (WT)为试材, 采用10% (m/v)聚乙二醇6000 (PEG6000)模拟干旱条件, 通过单独和联合施用PEG6000、DNA甲基化抑制剂5-氮杂胞苷(5azaC)和可变剪接抑制剂大环内酯类(PB)进行种子发芽实验, 测定种子活力、萌发过程中可溶性糖和可溶性蛋白含量、α-淀粉酶活性以及PEPC、糖信号相关基因和部分剪接因子基因的表达。结果表明, 0.25 μmol·L-1PB处理对2种供试水稻在干旱条件下种子萌发均表现出显著抑制作用, 使干旱条件下种子萌发过程中可溶性总糖、蔗糖、葡萄糖和果糖含量以及可溶性蛋白含量均有所下降, PB也抑制糖信号-蔗糖非发酵1 (SNF1)相关蛋白激酶(SnRKs)家族和剪接因子丝氨酸/精氨酸富集蛋白家族(SR proteins)相关基因的表达以及α-淀粉酶的活性, 但对PC的抑制作用小于WT。5 μmol·L-15azaC处理对干旱条件下种子萌发的效果与可变剪接抑制剂相反。5 μmol·L -1 5azaC联合PEG6000干旱处理部分减缓了干旱对水稻种子发芽率的抑制作用, 使供试材料发芽率升高, 表明DNA甲基化和可变剪接机制参与了水稻芽期干旱耐性, 其中对PC的作用更大。
关键词:
Abstract
In order to reveal the effect of epigenetic mechanism under drought stress toward seed germination of transgenic rice within high maize C4-type PEPC gene expressing, C4-PEPC transgenic rice (PC) and wild type rice Kitaake (WT) were used in this study. By introducing DNA methylation inhibitor (5-azacytidine, 5azaC) and alternative splicing inhibitors (macrolides pladienolide B, PB), the drought simulation treatments with 10% (m/v) polyethylene glycol-6000 (PEG6000) alone or combining with the inhibitors were used for seed germination experiments. Seed vigor, soluble sugar and soluble protein content, α-amylase activity and the expression levels of related genes, PEPC-related genes, sugar signal-related genes, and some splicing factor genes during germination were measured. In the results, when treated with 0.25 μmol·L-1, PB had showed a significant inhibitory effect on the seed germination of the two tested rice lines under drought conditions. The content of total soluble sugar, sucrose, glucose, fructose and soluble protein during seed germination after PB addition was reduced to a certain extent under drought conditions. PB treatment also inhibited the gene expression of sucrose nonfermenting-1 (SNF1)-related protein kinase SnRKs family and the splicing factor arginine/serine-rich proteins (SR proteins), and the activity of α-amylase as well, but the inhibitory effect on PC is less than those on WT. 5 μmol·L -1 5azaC treatment had an opposite effect with alternative splicing inhibitors. The combination treatment with 5azaC and PEG6000 partially alleviated the inhibitory effect of drought stress on rice seed germination, and the germination rate of the tested materials increased. It can be seen that the mechanism of DNA methylation and alternative splicing are involved in drought tolerance at the bud stage of rice lines, with a larger effect on PC.
Keywords:
PDF (5046KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
宋凝曦, 谢寅峰, 李霞. 干旱胁迫下表观遗传机制对转C4型PEPC基因水稻种子萌发的影响. 植物学报, 2020, 55(6): 677-692 doi:10.11983/CBB20048
Song Ningxi, Xie Yingfeng, Li Xia.

近年来, 在全球气候变化背景下, 干旱频繁发生且日趋严峻, 严重影响作物的生长, 甚至造成减产(Prasad et al., 2011)。在高光强、高温和干旱等逆境条件下, C4植物的光合效率和水分利用率高于C3植物, 进而获得较高的生物学产量(Lebouteiller et al., 2007)。磷酸烯醇式丙酮酸羧化酶(phosphoenolpyruvate carboxylase, PEPC)是重要的胞质代谢酶, 也是C4植物光合作用的关键酶, 在种子发芽和生长发育中发挥重要作用(O’Leary et al., 2011)。Ku等(1999)首次运用农杆菌介导法, 将完整的玉米(Zea mays) C4-PEPC基因(含自身启动子)成功导入水稻(Oryza sativa), 获得高表达玉米C4-PEPC的转基因水稻植株(简称PC), 该材料不仅光合效率显著提高, 而且籽粒产量显著增加。进一步研究发现, PC水稻在逆境如高光强(焦德茂等, 2001)、高温(焦德茂等, 2003)、干旱(Qian et al., 2015; Zhang et al., 2017)和低氮(Tang et al., 2018)条件下比原种表现出更大的优势。深入研究表明, 磷脂酸(Li et al., 2011)、H2O2 (Ren et al., 2014)、NO (Chen et al., 2014)和钙离子(Qian et al., 2015; 刘小龙等, 2015)等第二信使分子均参与了PC水稻C4-PEPC基因的表达和酶活性的发挥, 这些调节过程与其内源糖水平的差异密切相关(Li and Wang, 2013), 而且在干旱条件下PC内源糖代谢加强(Zhang et al., 2017)。张金飞等(2018)研究表明, 糖信号-蔗糖非发酵1 (sucrose nonfermen- ting-1, SNF1)相关蛋白激酶(sucrose nonfermen- ting-1-related protein kinase, SnRKs)家族中, SnRK3s家族基因可与钙调B类磷酸酶(calcineu- rin-B-like, CBL)作用, 诱导NO参与叶片气孔调节, 从而增强保水能力, 保持其光合能力稳定, 最终表现为耐旱。He等(2020)研究发现, SnRK2也参与PC的干旱响应。Liu等(2017)研究表明, PC水稻在响应干旱逆境时C4-PEPC会发生去甲基化现象, 有利于C4-PEPC的高表达, 同时也伴随着PEPC的磷酸化以及相关基因的表达, 表明表观遗传机制可能也参与该材料对干旱的响应, 但尚缺乏系统研究。
可变剪接(alternative splicing, AS)是一种生物体基因转录后调控的方式, 植物中的许多基因存在AS, 其在植物生长发育中具有重要作用(曲瑞莲等, 2014)。已有研究表明, AS广泛参与高等植物对非生物胁迫的转录调控, 为植物提供了一种迅速应对环境变化的有效方案(Syed et al., 2012), 在水稻中也已有相关研究(Cao and Ma, 2019)。PEPC的磷酸化在调控高等植物中枢代谢中起重要作用, 磷酸烯醇丙酮酸羧化酶激酶(phosphoenolpyruvate carboxylase kinases, PPCK)在PEPC磷酸化过程中起关键作用。已有研究在番茄(Lycopersicon esculentum)中鉴定了2个PPCK基因(LePPCK1和LePPCK2)。其中, LePPCK2还含有一种新的第二内含子, 具有可选择的剪接作用。在果实成熟期间, LePPCK2的表达显著增加, 而在成熟后子房和种子中仅包含正确剪接的LePPCK2转录物。此外, 在马铃薯(Solanum tuberosum)、茄子(S. melonge)和烟草(Nicotiana tabacum)中均存在这种组织特异性调节, 而且剪接的内含子序列在所报告的4个物种间高度保守(Marsh et al., 2003)。Ling等(2017)研究表明, 大环内酯类(pladienolide B, PB)抑制拟南芥(Arabidopsis thaliana)中的AS; 转录组测序数据显示, 采用PB模拟非生物胁迫信号(包括盐、干旱和脱落酸(abscisic acid, ABA))可激活拟南芥中非生物胁迫和ABA响应基因(RD29A和MAPK18), 并可模拟ABA对气孔孔径??的影响。Lev Maor等(2015)研究表明, DNA甲基化也调节可变剪接, 然而高表达玉米C4-PEPC转基因水稻植株在干旱响应过程中发生的DNA甲基化差异, 是否会诱导可变剪接的变化值得深入探究。
本研究以PC及非转基因野生型水稻(WT)为试材, 在芽期用PEG6000模拟干旱处理, 加入PB和DNA甲基化抑制剂5-氮杂胞苷(5-azacytidine, 5azaC), 进行不同程度的联合处理, 然后观察其对PC和WT种子发芽的影响。通过测定不同萌发时间内源可溶性糖和可溶性蛋白含量、α-淀粉酶活性以及相关基因、PEPC、糖信号以及部分剪接因子相关基因的表达, 分析不同处理下PC和WT种子萌发过程中主要相关酶活性、基因表达差异以及对PEPC基因表达的影响, 旨在揭示干旱条件下表观遗传途径对PC水稻种子萌发的影响, 为阐明PC在芽期的耐旱机制提供新线索。
1 材料与方法
1.1 供试材料
本研究所用的转基因水稻(Oryza sativa L.)是由Ku等(1999)以日本粳稻Kitaake为受体, 将完整的玉米C4-PEPC基因导入, 产生的高表达玉米C4-PEPC转基因水稻(PC), 以野生型水稻Kitaake (WT)为对照。选择同年收获的大小一致、颗粒饱满的供试种子, 先用75%乙醇消毒15分钟, 然后用无菌水冲洗干净。将种子置于含40 mL无菌水的培养皿中, 然后放入黑暗条件下的30°C培养箱中, 1天后对供试材料进行处理。1.2 干旱、DNA甲基化抑制剂以及可变剪接抑制剂处理
对照(CK)用无菌水处理, 再用10% (w/v) PEG6000进行模拟干旱处理。将浸泡后的水稻种子分别移至无菌水、10% PEG6000溶液、10% PEG6000+0.25 μmol·L-1 PB溶液、10% PEG6000+5 μmol·L-1 5azaC溶液以及10% PEG6000+0.25 μmol·L-1 PB+ 5 μmol·L-1 5azaC溶液中。取不同处理材料进行发芽实验, 待测样品按不同处理天数取样, 经液氮速冻后, 置于-80°C储存备用, 生理指标统一测定。测定基因表达水平的水稻种子处理方法: 均匀涂抹抑制剂后在黑暗中预处理10小时, 再转入上述溶液中处理2小时。1.3 种子活力测定
参照标准发芽试验法(陈蕾太等, 2016)测定种子活力。取不同浓度处理材料, 于处理第4天按照如下公式计算发芽势, 于第8天计算发芽率。取正常生长的幼苗经105°C杀青30分钟, 80°C烘干至恒量后, 称量苗干重和根干重。以标准发芽试验所测定的种子活力相关指标为对照。发芽势=(4天内正常幼苗数/供试种子数)×100%; 发芽率=(8天内正常幼苗数/供试种子数)×100%; 发芽指数(GI)=ΣGt/Dt (Gt为时间t的发芽数, Dt为相应的发芽天数); 活力指数=GI×S (S为单株幼苗干重质量(g))。
1.4 可溶性总糖测定
参考蒽酮比色法(Somani et al., 1987)测定可溶性总糖含量。将种子在10°C烘箱中烘干15分钟后调至70°C过夜。将种子磨碎后称取50 mg样品, 转入10 mL刻度离心管内, 加入4 mL 80%乙醇。水浴搅拌40分钟后离心收上清, 残渣用80%乙醇重复提取2次, 合并上清。在上清液中加入10 mg活性炭, 80°C脱色30分钟, 然后用80%乙醇定容至10 mL, 过滤后取滤液进行测定。吸取1 mL上述糖提取液, 转入干净的试管中, 加入5 mL蒽酮试剂混合, 沸水浴加热10分钟, 取出冷却, 然后采用分光光度计测定(波长为625 nm)吸光值(OD值)。根据标准曲线计算得出滤液中的糖含量, 并进一步计算样品中的含糖量。1.5 糖组分含量测定
蔗糖、葡萄糖和果糖含量参照文献(Ambavaram et al., 2014)进行测定。将蔗糖标准液用80%乙醇进行梯度(0、10、20、40、60、80和100 μg·mL-1)稀释。分别取0.4 mL溶液加200 μL 2 mol·L-1 NaOH, 100°C煮沸5分钟, 冷却后加2.8 mL 30%盐酸和0.8 mL 0.1%间苯二酚, 摇匀后于80°C水浴10分钟, 冷却后在480 nm处测定OD值, 然后计算样品中的蔗糖含量。将葡萄糖标准液用80%乙醇进行梯度(0、15、30、50、75、100、150和200 μg·mL-1)稀释。取8支试管分别加入4 mL酶制剂, 置于30°C水浴, 待管中酶液温度上升达到平衡后, 分别加入2 mL梯度稀释后的葡萄糖溶液, 摇匀后保温5分钟, 再各加入8 mL 10 mol·L-1硫酸终止反应。当酶液温度与室温平衡后, 测定460 nm处OD值, 然后计算样品中的葡萄糖含量。将果糖溶液用80%乙醇进行梯度(0、15、30、50、75、100、150和200 μg·mL-1)稀释。取8支试管, 分别加入1 mL梯度稀释后的果糖溶液, 然后分别加入2 mL 30%盐酸、1 mL 0.1%间苯二酚及1 mL H2O, 摇匀。在80°C水浴中反应10分钟, 然后冷却至室温, 480 nm处测OD值, 然后计算样品中的果糖含量。1.6 可溶性蛋白含量测定
采用考马斯亮蓝G-250法(Bradford, 1976)测定可溶性蛋白含量。以标准牛血清蛋白浓度作标准曲线, 分别加入2 mL染料试剂立即混匀, 用分光光度计测定波长595 nm处OD值, 以2 mL蒸馏水加2 mL染料试剂作为比色空白对照。通过标准曲线计算得出蛋白质质量浓度。1.7 α-淀粉酶活性测定
参考3,5-二硝基水杨酸(DNS)法(李合生, 2000)测定α-淀粉酶活性。取0.1 g萌发种子研磨至匀浆, 离心20分钟后取上清液备用。在对照和测定管中各加1 mL酶提取液, 70°C水浴15分钟后迅速冷却。在试管中各加入1 mL柠檬酸缓冲液, 对照管中各加入4 mL 0.4 mol·L-1 NaOH钝化酶活, 然后测定560 nm处OD值。1.8 总RNA提取和qRT-PCR
采用TaKaRa Mini BEST Plant RNA Extraction Kit (TaKaRa, Cat No.9769)试剂盒提取总RNA。采用TaKaRa Prime Script RT Master Mix Perfect Real Time (TaKaRa, Cat No.RR036A)试剂盒进行反转录。Real Time PCR采用TB Green Premix Ex Taq Tli RNaseH Plus (TaKaRa, Cat No.RR82LR)试剂盒进行。SR基因引物序列来自Dong等(2018)的研究, 其它引物采用Primer premier 5.0进行设计(表1)。Table 1
表1
表1引物序列
Table 1
| Gene | Forward primer (5'-3') | Reverse primer (5'-3') |
|---|---|---|
| Actin | CCCTCAAACATCGGTATGGA | TTGATCTTCATGCTGCTTGG |
| OsK1a | AACCAGAGGTAACAGGCAGG | AACCAGAGGTAACAGGCAGG |
| OsK24 | CGTGTTGGCTTCAGTGAAT | CCTTCTCTATCTAAGGGCCG |
| OsK35 | TTGTGTTGGCTTCAGTGAAA | CCTTCGCTGTCTAAGGACTG |
| C4-PEPC | CCCACTATCCTTCGCAAGAC | CTAGCCAGTGTTCTGCATGCCGG |
| Osppc2a | CTGGTTGAGATGGTTTTCGC | GGTGTGAATTCAGGCACTTC |
| SAPK8 | ATAGATGATAATGTCCAGCG | GTTCCTACAGTGGATTTTGG |
| SAPK9 | CACAGCAACGCCGTCTCC | CACACTTCCACCGCTACCAA |
| SAPK10 | TGCTGATGTGTGGTCGTGTG | TGCTGGTATGGTCGCCTCT |
| SR40 | CAATCTGGGGACTGCTTTC | TCCTGCTTGGGCTTTTACT |
| SR33 | ATATTGCCTGCTACCCGAAAG | CAGAGCAGCACCCAGTTTATTAC |
| OsAmy1A | TTTCGGTCCTCATCGTCCTCC | TCCACGACTCCCAGTTGAATC |
| OsAmy1C | TGGTATCGATCAGAAACCGGC | GTCCGACCTTCGTGATGACC |
| OsAmy3C | AAGCATTCCACCACAATGAGC | AGGAAGTTGTACCACCCACC |
| OsAmy3E | TCACCCTGTGTTGTGTCGTT | AAAGTTGTACCACCCGCCTT |
新窗口打开|下载CSV
1.9 数据处理
采用SPSS 25.0软件(SPSS, 芝加哥, IL)进行数据统计分析, 采用Microsoft Excel 2016软件作图。使用2-ΔΔCt方法(Livak and Schmittgen, 2001)分析qRT-PCR数据, 采用SPSS 25.0软件进行单因素方差分析和最小显著性差异多重比较检验(P<0.05)。2 结果与讨论
2.1 不同处理对PC和WT种子活力的影响
PEG处理可降低PC和WT的发芽势(图1A)、发芽率(图1B)、活力指数(图1C)和发芽指数(图1D)。图1E显示不同处理天数下各处理的种子发芽情况。PEG+PB处理可显著抑制种子发芽; PEG+5azaC处理可减弱干旱对PC和WT发芽势的抑制作用, 其中对PC的作用更为显著; PEG+PB+5azaC处理可减弱干旱对供试材料种子发芽率的抑制, 比PEG+PB处理对发芽的恢复作用更大。PB对PC的抑制作用小于WT, DNA甲基化抑制剂对PC发芽恢复作用更大, PC的发芽势、发芽指数和活力指数表现与其发芽率类似。图1
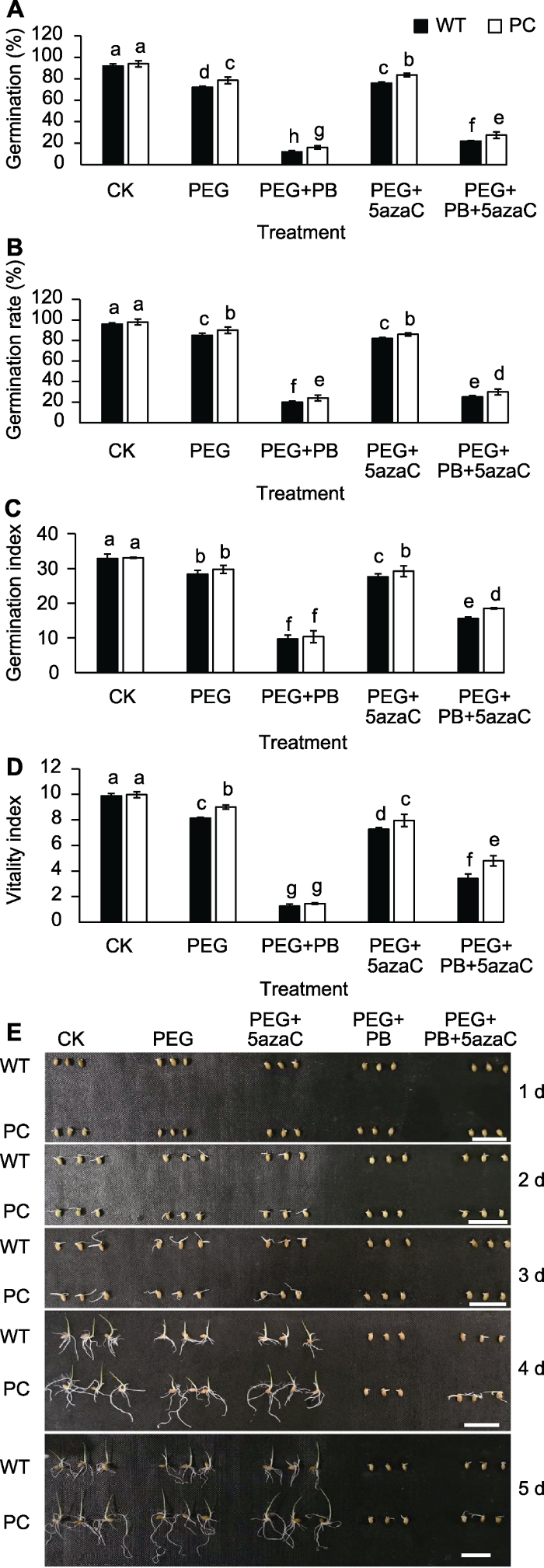 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1不同处理对转基因水稻(PC)和野生型水稻(WT)种子活力的影响
(A) 发芽势; (B) 发芽率; (C) 发芽指数; (D) 活力指数; (E) 种子发芽图片。不同小写字母表示差异显著(P<0.05)。Bars=0.5 cm
Figure 1Effects of different treatments on seed vigor of transgenic rice (PC) and wild type rice (WT)
(A) Germination; (B) Germination rate; (C) Germination index; (D) Vigor index; (E) Images of germinating seeds at different time. Different lowercase letters indicate significant differences (P<0.05). Bars=0.5 cm
2.2 不同处理对PC和WT种子根长和芽长的影响
不同处理下WT和PC种子根长(图2A)、芽长(图2B)和根芽比(图2C)的变化差异显著。与PC CK相比, PC PEG单独处理使PC根长增加, WT根长变短, 抑制了PC和WT芽的生长, 且两材料的根芽比更大。PC PEG+PB和WT PEG+PB处理可明显抑制PC和WT的根长、芽长和根芽比, 且与WT相比, 对PC的抑制作用较小。PC PEG+5azaC处理对PC生根影响不大, 但抑制WT生根, 均抑制PC和WT发芽, 但对PC的抑制作用小于WT, 且与PEG单独处理相比, 该处理对PC芽生长有缓解作用, PC和WT的根芽比更大。PC PEG+PB+5azaC和WT PEG+PB+5azaC处理抑制PC和WT的根长和芽长, 但对PC的抑制作用较小, 该处理与干旱联合PB处理相比, 可促进PC芽生长, 降低PC和WT的根芽比, 根在处理第4天之后才明显生长。上述结果表明, 可变剪接抑制剂显著抑制了供试材料根和芽长, 而甲基化抑制剂则发挥促进作用。从根芽比看, 处理后期根的伸长部分恢复, 但是两者并没有缓解可变剪接抑制剂对其根和芽生长的影响。图2
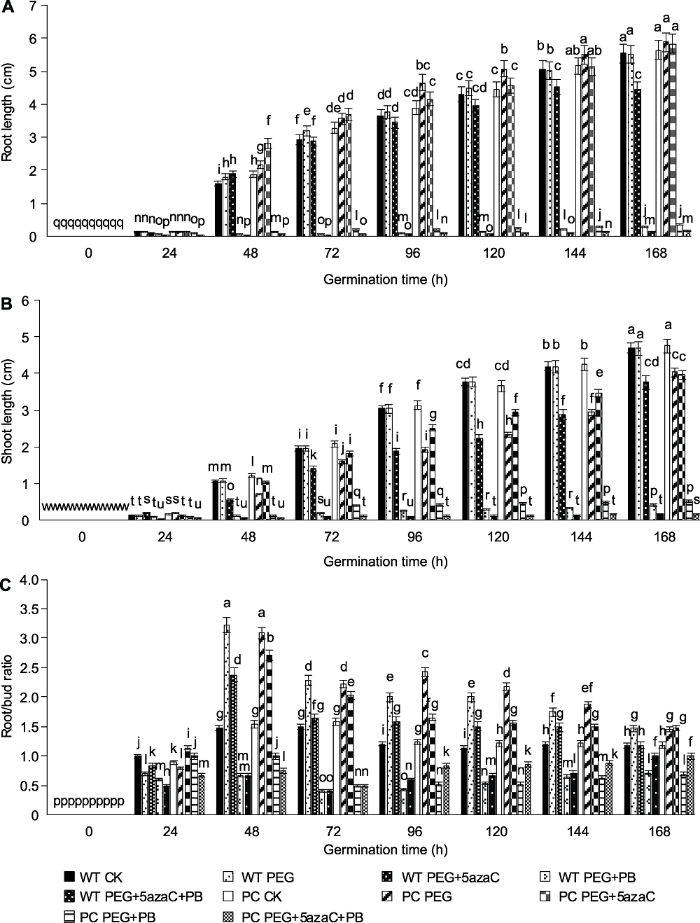 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同处理下转基因水稻(PC)和野生型水稻(WT)萌发种子根长(A)、芽长(B)和根芽比(C)的变化
不同小写字母表示差异显著(P<0.05)。
Figure 2Effects of different treatments on root length (A), shoot length (B) and root/bud ratio (C) of transgenic rice (PC) and wild type rice (WT) germinating seeds
Different lowercase letters indicate significant differences (P<0.05).
2.3 不同处理对PC和WT种子萌发过程中可溶性总糖和可溶性蛋白含量的影响
渗透调节物质(主要包括可溶性糖和可溶性蛋白)的积累是植物抵御逆境的有效方式。PEG、PEG+PB和PEG+PB+5azaC处理降低了种子萌发过程中的可溶性总糖含量(图3A); 而PEG+5azaC处理使PC种子萌发过程中可溶性总糖含量增加, 表明PEG+PB处理显著降低了可溶性总糖的含量, PEG+5azaC处理则提高了种子内可溶性总糖的含量。值得关注的是, DNA甲基化抑制剂和可变剪接抑制剂联合干旱处理均加大了两材料可溶性糖含量的差异, PC可溶性糖含量高于WT。PEG处理延缓了PC和WT可溶性蛋白含量的降低(图3B); PEG+PB处理加速PC与WT可溶性蛋白含量的下降; PEG+5azaC处理延缓了PC和WT可溶性蛋白降低的过程。表明PEG+PB处理显著降低了可溶性蛋白含量, PEG+5azaC处理则提高了种子可溶性蛋白含量, 可变剪接抑制剂联合干旱处理加大了转基因和野生型水稻可溶性蛋白含量的差异, 种子萌发过程中可溶性蛋白含量PC比WT下降缓慢。图3
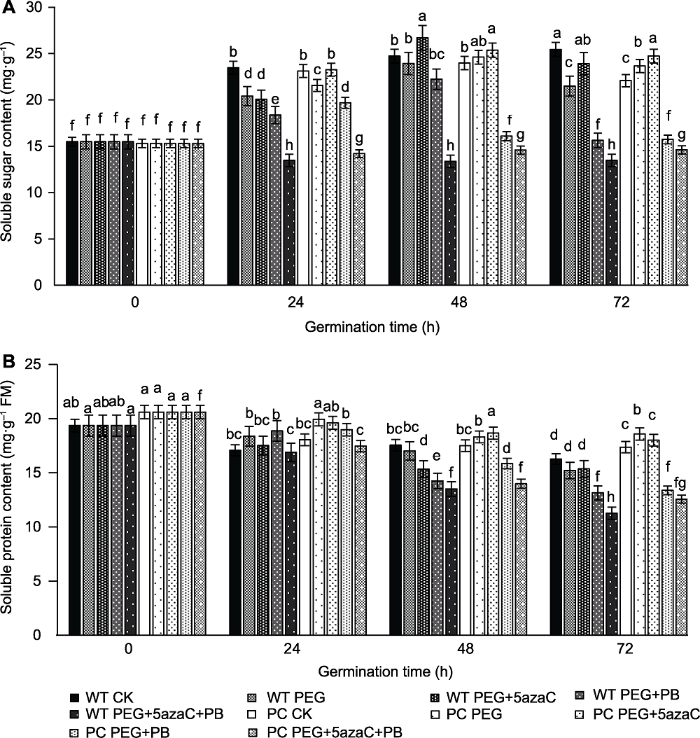 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同处理对转基因水稻(PC)和野生型水稻(WT)种子萌发过程中可溶性总糖含量(A)和可溶性蛋白含量(B)的影响
不同小写字母表示差异显著(P<0.05)。
Figure 3Effects of different treatments on total soluble sugar content (A) and soluble protein content (B) of transgenic rice (PC) and wild type rice (WT) germinating seeds
Different lowercase letters indicate significant differences (P<0.05).
2.4 不同处理对PC和WT种子萌发过程中糖组分含量的影响
可溶性总糖在萌发24和48小时出现显著差异, 我们进一步分析了主要糖组分在同期的变化。图4A-C显示PC和WT种子在不同处理24小时糖组分含量的差异。其中, PEG处理使PC和WT种子蔗糖和果糖含量降低; PEG+PB处理使两材料蔗糖、葡萄糖和果糖含量降低; PEG+5azaC处理使两材料中蔗糖、葡萄糖与果糖的含量增加; PEG+PB+5azaC处理使两材料蔗糖、葡萄糖和果糖含量降低。图4D-F显示PC和WT种子在不同处理48小时糖组分含量的差异, 其中PEG处理使PC和WT种子蔗糖和葡萄糖的含量降低; PEG+PB处理使两材料蔗糖、葡萄糖和果糖的含量降低; PEG+5azaC处理使两材料中蔗糖、葡萄糖与果糖的含量增加。此外, 用可变剪接抑制剂处理24小时后, PC的蔗糖和葡萄糖含量高于WT; 处理48小时PC的蔗糖、葡萄糖和果糖含量高于WT。图4
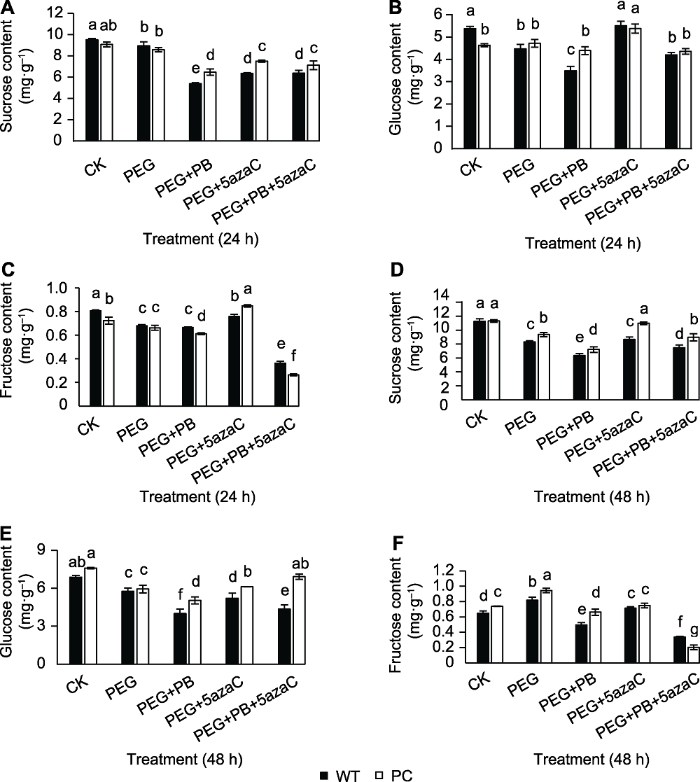 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同处理对转基因水稻(PC)和野生型水稻(WT)种子萌发过程中糖组分含量的影响
(A), (D) 分别为发芽24和48小时的蔗糖含量; (B), (E) 分别为发芽24和48小时的葡萄糖含量; (C), (F) 分别为发芽24和48小时的果糖含量。不同小写字母表示差异显著(P<0.05)。
Figure 4Effects of different treatments on sugar content of transgenic rice (PC) and wild type rice (WT) germinating seeds
(A), (D) Sucrose content at 24 h and 48 h after germination, respectively; (B), (E) Glucose content at 24 h and 48 h after germination, respectively; (C), (F) Fructose content at 24 h and 48 h after germination, respectively. Different lowercase letters indicate significant differences (P<0.05).
2.5 不同处理对PC和WT种子萌发过程中SnRK家族相关基因表达的影响
在不同处理条件下, PC和WT糖组分的差异反映糖和能量水平的变化。为验证这种变化是否与糖信号SnRK家族相关基因有关, 我们选取与能量水平密切相关的3个SnRK1家族基因(OsK1a、OsK24和OsK35)以及与逆境相关的ABA依赖的3个SnRK2家族基因(SAPK8、SAPK9和SAPK10), 分析其基因表达响应干旱的变化。图5A-C显示, PEG+PB处理抑制供试材料SnRK1相关基因的表达, PEG+5azaC处理显著提高SnRK1相关基因的表达水平, 表明两种表观遗传机制的确影响了SnRK1基因的表达, 而且与蔗糖的含量呈现同步。图5D-F显示, PEG+PB处理下调SnRK2相关基因的表达, 而且与葡萄糖的含量同步; PEG+5azaC处理使SnRK2相关基因的表达量增加。值得关注的是, PC可溶性糖的降幅始终小于WT, 因此, PC的糖类相关基因表达仍高于WT。图5
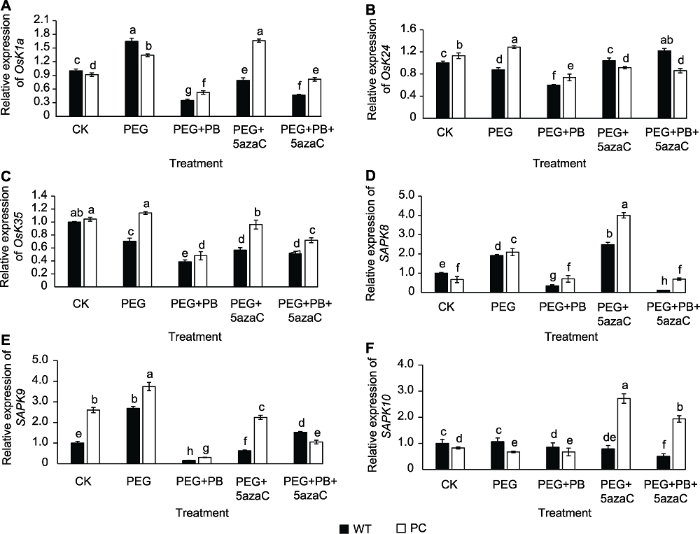 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5不同处理对转基因水稻(PC)和野生型水稻(WT)种子萌发过程中SnRK相关基因表达的影响
不同小写字母表示差异显著(P<0.05)。
Figure 5Effects of different treatments on the expression of SnRK related genes in transgenic rice (PC) and wild type rice (WT) germinating seeds
Different lowercase letters indicate significant differences (P<0.05).
2.6 不同处理对PC和WT种子萌发过程中剪接因子相关基因表达的影响
丝氨酸/精氨酸丰富蛋白家族(arginine/serine-rich proteins, SR proteins)是真核生物中的一类剪接因子, 在前体mRNA的组成性和选择性剪接中起作用(Reddy and Shad, 2011)。其中SR为植物所特有, 在植物响应环境胁迫中起重要作用(Staiger and Brown, 2013)。拟南芥SR40的突变体比野生型对脱落酸更敏感(Cruz et al., 2014), 在矿质元素缺乏条件下SR33具有调节水稻中磷元素分布的作用(Dong et al., 2018)。PEG处理上调PC和WT中SR33基因的表达; PEG+PB和PEG+5azaC处理均下调SR40和SR33基因的表达, 其中PC中SR40和SR33的表达比WT更高(图6A, B)。以上结果表明, 可变剪接抑制剂对不同剪接因子基因表达的影响与其糖信号的表现类似。图6
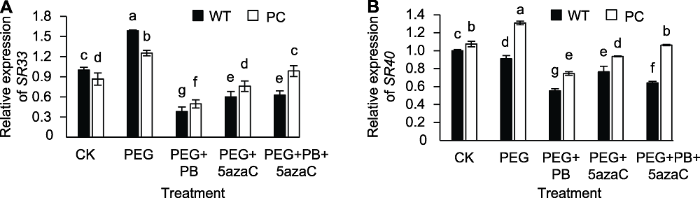 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6不同处理对转基因水稻(PC)和野生型水稻(WT)种子萌发过程中剪接因子相关基因表达的影响
不同小写字母表示差异显著(P<0.05)。
Figure 6Effects of different treatments on expression of splicing factor related genes in transgenic rice (PC) and wild type rice (WT) germinating seeds
Different lowercase letters indicate significant differences (P<0.05).
2.7 不同处理对PC和WT种子萌发过程中PEPC相关基因表达的影响
剪接因子基因的变化是否改变了PC的PEPC基因的变化值得关注。PC不仅包括内源C3型PEPC基因(Osppc2a), 还包括外源导入的玉米C4-PEPC基因(C4-PEPC)。本研究检测了C4-PEPC和Osppc2a基因表达水平。结果表明, PEG单独处理可上调PC中C4-PEPC的表达; PEG+PB处理则下调PC中C4-PEPC的表达; PEG+PB+5azaC处理可上调PC中C4-PEPC的表达(图7A)。图7B显示, PEG处理均上调供试材料Osppc2a基因的表达; PEG+PB处理下调了Osppc2a基因的表达; PEG+5azaC处理也上调两供试材料Osppc2a基因的表达; PEG+PB+5azaC处理上调两供试材料Osppc2a基因表达。以上结果表明,两种表观遗传抑制剂确实影响了C4-PEPC和Osppc2a的表达, 但并未完全抑制, 暗示可能还存在其它机制参与其对干旱的响应。图7
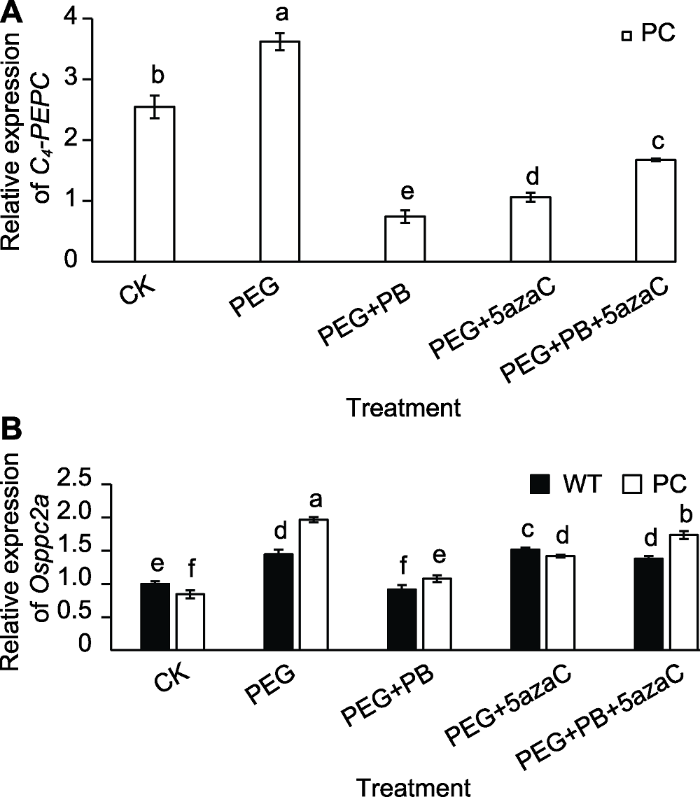 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7不同处理对转基因水稻(PC)和野生型水稻(WT)种子萌发过程中PEPC相关基因表达的影响
不同小写字母表示差异显著(P<0.05)。
Figure 7Effects of different treatments on expression of PEPC-related genes in transgenic rice (PC) and wild type rice (WT) germinating seeds
Different lowercase letters indicate significant differences (P<0.05).
2.8 不同处理对PC和WT种子萌发过程中α-淀粉酶相关基因表达的影响
图8A?D显示, PEG单独处理上调了PC和WT OsAmy3C基因的表达; PEG+PB处理促进了PC种子OsAmy1A、OsAmy1C和OsAmy3C的表达, 对其中OsAmy3C的影响与单独干旱处理类似, 表现为诱导增加, 而对OsAmy3E则显著抑制; PEG+5azaC处理均下调了OsAmy1A和OsAmy1C的表达, 上调了OsAmy3E的表达; PEG+PB+5azaC处理均下调OsAmy1A、OsAmy1C和OsAmy3E的表达。其中, 两种表观遗传抑制剂处理下PC的OsAmy1A和OsAmy3C的表达均显著高于WT, 与PC的发芽率与发芽势的表现一致。图8
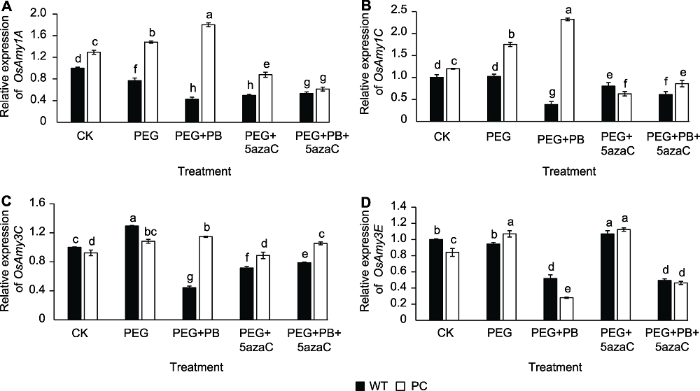 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8不同处理对转基因水稻(PC)和野生型水稻(WT)种子萌发过程中α-淀粉酶相关基因表达的影响
不同小写字母表示差异显著(P<0.05)。
Figure 8Effects of different treatments on expression of α-amylase related genes in transgenic rice (PC) and type wild rice (WT) germinating seeds
Different lowercase letters indicate significant differences (P<0.05).
2.9 不同处理对PC和WT种子萌发过程中α-淀粉酶活性的影响
图9显示在萌发24小时后, PEG单独处理下, PC和WT的α-淀粉酶活性均高于对照, 且PC高于WT; PEG+ PB处理下WT和PC表现不同, PC的表现高于其对照CK, 而WT则表现低于其对照; PEG+5azaC和PEG+ PB+5azaC处理下, PC和WT均低于对照; 在萌发后48-72小时, 各处理条件下PC和WT的α-淀粉酶活性均逐渐下降至低于对照, 但PC下降程度仍小于WT。图9
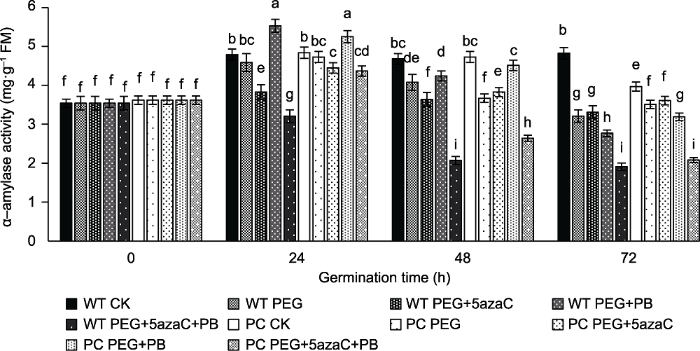 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9不同处理对转基因水稻(PC)和野生型水稻(WT)种子萌发过程中α-淀粉酶活性的影响
不同小写字母表示差异显著(P<0.05)。
Figure 9Effects of different treatments on α-amylase activity in transgenic rice (PC) and wild type rice (WT) germinating seeds
Different lowercase letters indicate significant differences (P<0.05).
2.10 表观遗传抑制剂处理下PC和WT种子萌发过程各指标相关性分析
相关性分析显示, 本研究中PC发芽势分别与SAPK10和C4-PEPC基因表达显著相关(P<0.05); 发芽率分别与α-淀粉酶活性、OsK1a、OsAmy1A以及SR40基因表达显著相关; 发芽指数分别与种子中蔗糖和葡萄糖含量显著相关(P<0.05); 可溶性蛋白与OsAmy1A基因表达量相关, α-淀粉酶活性与OsAmy1A基因表达水平呈极显著相关(P<0.01)。OsK1a表达分别与SR40、C4-PEPC基因表达显著相关; SAPK8表达分别与OsAmy1A、SR40基因表达显著相关(P<0.05)。OsAmy1A表达与SR33基因表达呈极显著相关(P<0.01); SR40表达与C4-PEPC基因表达相关; WT发芽势与OsAmy1A基因表达显著相关(P<0.05); 发芽率与SAPK8表达显著相关(P<0.05); 可溶性糖含量分别与OsK35、Osppc2a基因表达显著相关(P<0.05); 葡萄糖含量与SAPK8基因表达呈极显著相关(P<0.01); α-淀粉酶活性与OsAmy1A基因表达相关; OsK35表达与Osppc2a基因表达显著相关(P<0.05); SAPK9表达与SAPK10、OsAmy3E基因表达相关(附表1)。2.11 讨论
C4植物的磷酸烯醇式丙酮酸羧化酶可将二氧化碳输送至核酮糖-1,5-二磷酸羧化酶(Ribulose-1,5-bisphosphate carboxylase, Rubisco), 在高光强和高温下也能更快地进行光合作用, 因此C4植物光合作用效率高, 并具有更好的水分利用率(Lata et al., 2013; Muthamilarasan et al., 2014)。高表达转玉米C4型PEPC基因水稻的耐逆表现为丰富植物耐旱机理提供了很好的研究材料。本研究从供试种子的发芽情况、萌发过程中可溶性糖及可溶性蛋白含量、相关基因表达以及α-淀粉酶活性的变化进行分析, 结果表明, 表观遗传机制-DNA甲基化和可变剪接参与了供试材料芽期的干旱响应, 其中对PC的作用更明显, 与其相关的糖信号SnRK1s和SnRK2s基因的表达有关。有效的种子发芽使幼苗健壮地生长, 并具有抵御生物或非生物胁迫的能力, 从而最终发展成为高产作物(Foolad et al., 2007)。种子发芽需要有利的环境条件, 包括水、氧气和温度, 以及多种细胞过程如合成、转运和信号转导的结合(Miransari and Simith, 2014)。干种子在吸水后需通过DNA修复、mRNA的转录及翻译迅速恢复代谢活性(Dinkova et al., 2011)。水稻种子萌发过程中, α-淀粉酶起主导作用(Damaris et al., 2019)。种子田间萌发出苗过程中常遇干旱胁迫, 因此研究不同基因型水稻种子逆境萌发过程中主要相关酶活性和基因表达差异, 对进一步认识种子活力的生化和分子基础具有重要参考价值(李美玲等, 2017)。α-淀粉酶基因Amy1A、Amy1C、Amy3C和Amy3E是种子萌发过程中的重要基因(Nanjo et al., 2004; Hakata et al., 2012; Hu et al., 2016; Nakata et al., 2017)。本研究表明, 在PB联合PEG处理后, PC的α-淀粉酶活性高于WT, PC的OsAmy1A、OsAmy1C和OsAmy3C表达水平更高, 可能与PC耐旱有关; 5azaC联合PEG处理后, PC的α-淀粉酶活性高于WT, PC的OsAmy1A和OsAmy3C表达量也较高, 表明DNA甲基化和可变剪接这2种表观遗传途径参与水稻种子萌发过程中的干旱响应。
为了能够在胁迫条件下生存, 植物进化出复杂的机制来感知外部信号, 从而对环境变化做出最佳反应, 其中表观遗传机制调控植物应答外界胁迫是近年来发现的重要机制之一, DNA甲基化和可变剪接均是其中的重要途径(杜康兮等, 2018)。已有研究表明, 水稻干旱响应基因的甲基化水平显著增高(Yan et al., 2010)。在植物中包含内含子的基因有可变剪接现象, 其中拟南芥和水稻中分别有61%和33%含有内含子的基因存在可变剪接(Wang and Brendel, 2006; Marquez et al., 2012)。AS在植物发育和环境应答中的作用已受到越来越多研究者的重视, 已报道在烟草、小麦(Triticum aestivum)、水稻和拟南芥中与生物和非生物胁迫相关的基因包括HsfA2、INH2、sad1/lsm5、Wdreb2、RPS4、SR (SR32和SR33a)、TNL和CNL等(Dinesh and Baker, 2000; Egawa et al., 2006; Zhang and Gassmann, 2007; Gassmann, 2008; Brummell et al., 2011; Golisz et al., 2013; Liu et al., 2013; Zhang et al., 2013; 宋凝曦等, 2020)。研究还发现, 包括水稻在内的许多作物在响应非生物胁迫中发生了AS, 受干旱程度影响会导致可变剪接的数目上升(Filichkin et al., 2014)。PB作为植物中AS的抑制剂, 以浓度依赖性方式对植物生长和发育表现出显著的抑制作用(Ling et al., 2017)。本研究也发现, 0.25 μmol·L-1 PB可显著抑制供试材料的种子萌发能力, 加入5 μmol·L-1 5azaC并未缓解PB对种子萌发的抑制, 从供试水稻发芽率抑制程度看, AS起的作用大于DNA甲基化的作用。
丝氨酸/苏氨酸蛋白激酶是一个相对较大的蛋白激酶家族, 其中AMPK (AMP-activated kinase)/ SnRKs是高度保守的丝氨酸/苏氨酸蛋白激酶家族的重要成员。SnRKs可以细分成3个亚家族(SnRK1、SnRK2和SnRK3)。SnRK1主要在调节植株生长发育、维持能量平衡以及响应生物逆境方面发挥重要作用; SnRK2则在响应干旱以及盐碱等非生物胁迫中发挥重要作用; SnRK3更多地参与胞内离子浓度的平衡, 通过调节离子浓度迅速启动植株响应非生物胁迫(张金飞等, 2017)。本研究中, PB联合干旱处理后抑制了蔗糖相关的SnRK1亚家族基因和葡萄糖相关的依赖ABA的SnRK2亚家族基因表达, 而加入5azaC联合干旱处理则对SnRK1亚家族基因OsK1a、OsK24及SnRK2亚家族基因SAPK9有上调作用。研究表明, 5% PEG6000模拟干旱处理1小时后, 表达PC的C4-PEPC启动子-1 095至-416区域中出现去甲基化现象(Liu et al., 2017)。本研究进一步表明, PEG联合DNA甲基化抑制剂处理可上调PC的SnRK1和SnRK2相关基因表达, 可能有利于其C4-PEPC基因的表达从而响应干旱。丝氨酸/精氨酸丰富蛋白家族(arginine/serine-rich proteins, SR proteins)是在真核生物前体mRNA选择性剪接中起作用的一类剪接因子, 在植物发育以及对环境的响应中具有关键作用(Reddy and Shad, 2011)。本研究发现在PB联合干旱处理后, 供试材料种子中剪接因子相关基因SR33和SR40的表达下降, 证实PB通过调节剪切子相关基因的表达而下调AS, 从而抑制种子萌发, 调控其干旱耐受性。
本研究阐明了表观遗传机制对干旱条件下PC种子萌发的效应。结果表明, 0.25 μmol·L-1 PB联合干旱处理对供试水稻种子萌发均表现出显著的抑制作用。添加PB后, 种子萌发过程中可溶性总糖、蔗糖、葡萄糖、果糖含量以及可溶性蛋白含量下降, 该处理还抑制蔗糖相关的SnRK1亚家族基因和葡萄糖相关的SnRK2亚家族基因表达, α-淀粉酶相关基因以及剪接因子SR相关基因表达也被抑制, 但对PC的抑制作用小于WT。5 μmol·L-1 5azaC联合干旱处理则与可变剪接抑制剂的效果相反。用5azaC联合PB处理可部分缓解干旱对供试水稻种子萌发的抑制效果。
综上, 水稻种子萌发时如遇到干旱胁迫, 会通过DNA去甲基化或者增加可变剪接来响应干旱胁迫, 而且两供试材料抑制程度的差异与其糖信号相关基因的差异表达有关。本研究阐明了DNA甲基化和可变剪接途径对供试水稻芽期干旱的影响, 但是两种途径如何互作尚未探讨, 糖如何诱导PC的表观遗传途径还需后续深入研究。
附表1 表观遗传抑制剂处理下转基因水稻(PC)和野生型水稻(WT)种子萌发过程各指标相关性分析
Appendix table 1 Correlation of parameters of transgenic rice (PC) and wild type rice (WT) seeds germination process under epigenetic inhibitor treatmenthttp://www.chinbullbotany.com/fileup/1674-3466/PDF/t20-048.pdf
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
DOI:10.11983/CBB17143URL [本文引用: 1]

植物的生长发育容易受到外界环境变化的影响。非生物胁迫发生时, 表观遗传机制对胁迫应答基因的表达调控发挥了十分重要的作用。近年来, 调控植物非生物胁迫应答的表观遗传机制研究取得了一系列重要进展, 为进一步深入解析植物响应非生物胁迫的分子机制奠定了基础。该文对DNA甲基化修饰、组蛋白修饰、染色质重塑和非编码RNA等主要表观遗传调控方式在植物响应非生物胁迫中的作用进行了简要综述。
[本文引用: 1]
URL [本文引用: 1]
PEPC,PPDK,NADP-ME,PEPC+PPDK基因水稻及原种为材料,比较C4途径有关光合酶活性、叶绿素荧光参数、CO2交换等生理指标,重点研究了转PEPC基因水稻的生理特性, 结果如下:(1)转PEPC基因水稻PEPC活性比原种高20倍,光饱和光合速率比原种高55%,羧化效率提高50%,CO2补偿点降低27%;(2)在高光强(3 h)或光氧化剂甲基紫精(MV)处理后, 与原种相比,转PEPC基因水稻的PSⅡ光化学效率(Fv /Fm)、光化学猝灭(qP)下降较少,证明其耐光抑制和耐光氧化能力增强;(3)在高光强条件下,转PEPC基因水稻中RuBPCase活性变化不明显,但碳酸酐酶(CA)诱导活性增加1.8倍.这些结果为揭示高光合效率机理和高光合效率育种提供了有益的启示.]]>
[本文引用: 1]
[本文引用: 1]
DOI:10.3724/SP.J.1259.2015.00206URL [本文引用: 1]
4植物光合作用酶。为了揭示高光效转C4PEPC基因水稻(Oryza sativa)对干旱胁迫的适应机理, 以高表达转C4PEPC水稻(PC)和野生型水稻Kitaake (WT)为供试材料, 在植株的4–5叶期, 使用不同浓度外源CaCl2溶液处理, 测定在15%聚乙二醇6000 (polyethylene glycol-6000, PEG-6000)胁迫下叶片相对含水量、光合参数、内源钙总含量、叶片总蛋白激酶活性、PEPC酶活性以及相关基因表达和蛋白质含量。结果表明, 0.5 mmol·L–1 CaCl2明显提高PC叶片相对含水量(P<0.05), 2 mmol·L–1和10 mmol·L–1 CaCl2则作用不显著, 对WT则影响不显著。不同浓度钙处理对PEG处理PC的净光合速率影响不显著, 而通过维持气孔导度减少水分胁迫。内源总钙浓度的数据显示, 在PEG6000处理下, PC具有维持稳定内源Ca2+浓度的能力, 过高浓度(10 mmol·L–1 CaCl2)钙处理反而降低了PEPC酶活性、PEPC基因表达和可溶性蛋白的含量。]]>
[本文引用: 1]
[本文引用: 1]
DOI:10.3724/SP.J.1006.2018.00082URL [本文引用: 1]
2+、NO含量、己糖激酶活性、B类钙调磷酸酶(calcineurin B-like, CBL)与蔗糖非发酵1 (sucrose nonfermenting-1, SNF1)相关蛋白激酶(SNF1-related protein kinase 3s, SnRK3s)基因表达的变化。结果表明,在盆栽试验中,分蘖期外施3%葡萄糖联合干旱处理对水稻的产量及其构成因子影响不显著,而在孕穗期处理,PC的株高、穗数、每穗实粒重和单株产量均显著高于WT。在水培试验中,外施1%葡萄糖和12% (m/v)聚乙二醇6000 (polyethylene glycol-6000, PEG-6000)模拟干旱处理,均显著提高了PC功能叶片的净光合速率(Pn)、气孔导度(Gs)和羧化效率(CE),叶片内蔗糖和果糖的含量也均显著高于WT。值得关注的是,PC叶片己糖激酶(hexokinase, HXK)活性、CBL1/SnRK3.1/SnRK3.4/ SnRK3.21基因的相对表达量在外施1%葡萄糖联合12% PEG模拟干旱处理下均显著低于12%PEG处理,而NO含量则显著上升。相关性分析也表明,PC中Pn、胞间CO2浓度(Ci)和Gs分别与葡萄糖含量、HXK活性和SnRK3.16基因表达显著相关。外施葡萄糖处理可上调PC糖水平,下调其CBL和SnRK3s基因表达,诱导NO参与叶片气孔调节,从而增强保水能力,保持光合能力稳定,最终表现为耐旱。]]>
DOI:10.11983/CBB16095URL [本文引用: 1]
蔗糖非发酵1 (SNF1)相关蛋白激酶家族(SnRKs)是植物胁迫响应过程中的一类关键蛋白激酶。在响应生物胁迫时, SnRKs可通过参与活性氧和水杨酸介导的信号转导途径, 增强植物对生物侵害的耐受性。在响应非生物胁迫时, SnRKs通过脱落酸(ABA)介导的信号通路, 增强植株对干旱、盐碱和高温等的耐受性; 且通过独立于ABA的信号通路, SnRKs可调控胞内能量状态, 维持离子平衡。该文总结了SnRKs蛋白激酶作为胁迫信号通路中的主要调节因子的最新研究进展, 并展望了未来的研究方向。
DOI:10.1038/ncomms6302URLPMID:25358745 [本文引用: 1]
Plants capture solar energy and atmospheric carbon dioxide (CO2) through photosynthesis, which is the primary component of crop yield, and needs to be increased considerably to meet the growing global demand for food. Environmental stresses, which are increasing with climate change, adversely affect photosynthetic carbon metabolism (PCM) and limit yield of cereals such as rice (Oryza sativa) that feeds half the world. To study the regulation of photosynthesis, we developed a rice gene regulatory network and identified a transcription factor HYR (HIGHER YIELD RICE) associated with PCM, which on expression in rice enhances photosynthesis under multiple environmental conditions, determining a morpho-physiological programme leading to higher grain yield under normal, drought and high-temperature stress conditions. We show HYR is a master regulator, directly activating photosynthesis genes, cascades of transcription factors and other downstream genes involved in PCM and yield stability under drought and high-temperature environmental stress conditions.
URLPMID:942051 [本文引用: 1]
DOI:10.1093/jxb/err043URLPMID:21393382 [本文引用: 1]
Cold storage of tubers of potato (Solanum tuberosum L.) compromises tuber quality in many cultivars by the accumulation of hexose sugars in a process called cold-induced sweetening. This is caused by the breakdown of starch to sucrose, which is cleaved to glucose and fructose by vacuolar acid invertase. During processing of affected tubers, the high temperatures involved in baking and frying cause the Maillard reaction between reducing sugars and free amino acids, resulting in the accumulation of acrylamide. cDNA clones with deduced proteins homologous to known invertase inhibitors were isolated and the two most abundant forms, termed INH1 and INH2, were shown to possess apoplastic and vacuolar localization, respectively. The INH2 gene showed developmentally regulated alternative splicing, so, in addition to the INH2alpha transcript encoding the full-length protein, two hybrid mRNAs (INH2beta*A and INH2beta*B) that encoded deduced vacuolar invertase inhibitors with divergent C-termini were detected, the result of mRNA splicing of an upstream region of INH2 to a downstream region of INH1. Hybrid RNAs are common in animals, where they may add to the diversity of the proteome, but are rarely described in plants. During cold storage, INH2alpha and the hybrid INH2beta mRNAs accumulated to higher abundance in cultivars resistant to cold-induced sweetening than in susceptible cultivars. Increased amounts of invertase inhibitor may contribute to the suppression of acid invertase activity and prevent cleavage of sucrose. Evidence for increased RNA splicing activity was detected in several resistant lines, a mechanism that in some circumstances may generate a range of proteins with additional functional capacity to aid adaptability.
DOI:10.3389/fpls.2019.01222URLPMID:31632433 [本文引用: 1]
Gene expression in eukaryotes is controlled at multiple levels, including transcriptional and post-transcriptional levels. The transcriptional regulation of gene expression is complex and includes the regulation of the initiation and elongation phases of transcription. Meanwhile, the post-transcriptional regulation of gene expression includes precursor messenger RNA (pre-mRNA) splicing, 5' capping, and 3' polyadenylation. Among these events, pre-mRNA splicing, conducted by the spliceosome, plays a key role in the regulation of gene expression, and the efficiency and precision of pre-mRNA splicing are critical for gene function. Ski-interacting protein (SKIP) is an evolutionarily conserved protein from yeast to humans. In plants, SKIP is a bifunctional regulator that works as a splicing factor as part of the spliceosome and as a transcriptional regulator via interactions with the transcriptional regulatory complex. Here, we review how the functions of SKIP as a splicing factor and a transcriptional regulator affect environmental fitness and development in plants.
[本文引用: 1]
DOI:10.3390/ijms151017541URLPMID:25268622 [本文引用: 1]
Serine/arginine-rich (SR) proteins are major modulators of alternative splicing, a key generator of proteomic diversity and flexible means of regulating gene expression likely to be crucial in plant environmental responses. Indeed, mounting evidence implicates splicing factors in signal transduction of the abscisic acid (ABA) phytohormone, which plays pivotal roles in the response to various abiotic stresses. Using real-time RT-qPCR, we analyzed total steady-state transcript levels of the 18 SR and two SR-like genes from Arabidopsis thaliana in seedlings treated with ABA and in genetic backgrounds with altered expression of the ABA-biosynthesis ABA2 and the ABA-signaling ABI1 and ABI4 genes. We also searched for ABA-responsive cis elements in the upstream regions of the 20 genes. We found that members of the plant-specific SC35-Like (SCL) Arabidopsis SR protein subfamily are distinctively responsive to exogenous ABA, while the expression of seven SR and SR-related genes is affected by alterations in key components of the ABA pathway. Finally, despite pervasiveness of established ABA-responsive promoter elements in Arabidopsis SR and SR-like genes, their expression is likely governed by additional, yet unidentified cis-acting elements. Overall, this study pinpoints SR34, SR34b, SCL30a, SCL28, SCL33, RS40, SR45 and SR45a as promising candidates for involvement in ABA-mediated stress responses.
[本文引用: 1]
DOI:10.1073/pnas.020367497URLPMID:10660679 [本文引用: 1]
The N gene, a member of the Toll-IL-1 homology region-nucleotide binding site-leucine-rich repeat region (LRR) class of plant resistance genes, encodes two transcripts, N(S) and N(L), via alternative splicing of the alternative exon present in the intron III. The N(S) transcript, predicted to encode the full-length N protein containing the Toll-IL-1 homology region, nucleotide binding site, and LRR, is more prevalent before and for 3 hr after tobacco mosaic virus (TMV) infection. The N(L) transcript, predicted to encode a truncated N protein (N(tr)) lacking 13 of the 14 repeats of the LRR, is more prevalent 4-8 hr after TMV infection. Plants harboring a cDNA-N(S) transgene, capable of encoding an N protein but not an N(tr) protein, fail to exhibit complete resistance to TMV. Transgenic plants containing a cDNA-N(S)-bearing intron III and containing 3' N-genomic sequences, encoding both N(S) and N(L) transcripts, exhibit complete resistance to TMV. These results suggest that both N transcripts and presumably their encoded protein products are necessary to confer complete resistance to TMV.
[本文引用: 1]
DOI:10.1105/tpc.18.00051URLPMID:30254029 [本文引用: 2]
Alternative splicing (AS) of pre-mRNAs promotes transcriptome and proteome diversity and plays important roles in a wide range of biological processes. However, the role of AS in maintaining mineral nutrient homeostasis in plants is largely unknown. To clarify this role, we obtained whole transcriptome RNA sequencing data from rice (Oryza sativa) roots grown in the presence or absence of several mineral nutrients (Fe, Zn, Cu, Mn, and P). Our systematic analysis revealed 13,291 alternatively spliced genes, representing approximately 53.3% of the multiexon genes in the rice genome. As the overlap between differentially expressed genes and differentially alternatively spliced genes is small, a molecular understanding of the plant's response to mineral deficiency is limited by analyzing differentially expressed genes alone. We found that the targets of AS are highly nutrient-specific. To verify the role of AS in mineral nutrition, we characterized mutants in genes encoding Ser/Arg (SR) proteins that function in AS. We identified several SR proteins as critical regulators of Zn, Mn, and P nutrition and showed that three SR protein-encoding genes regulate P uptake and remobilization between leaves and shoots of rice, demonstrating that AS has a key role in regulating mineral nutrient homeostasis in rice.
DOI:10.1266/ggs.81.77URLPMID:16755132 [本文引用: 1]
A number of cold responsive (Cor)/late embryogenesis abundant (Lea) genes are induced by both low temperature (LT) and dehydration. To understand the molecular basis of cold acclimation and its relationship with drought stress response in wheat seedlings, we isolated a DREB2 homolog Wdreb2, which is the candidate gene for a transcription factor of the Cor/Lea genes. The Wdreb2 expression was activated by cold, drought, salt and exogenous ABA treatment. Detailed expression studies of Wdreb2 indicated the involvement of two distinct pathways of its activation, a drought and salt stress-responsive pathway and a cold-responsive pathway. The transient expression analysis showed that the Wrab19 expression was directly activated by the WDREB2 transcription factor in wheat cells. Three transcript forms of Wdreb2 (Wdreb2alpha, Wdreb2beta and Wdreb2gamma) were produced through alternative splicing. Under drought and salt stress conditions, the amount of the Wdreb2beta form remained fairly constant during 24-hour treatment, while those of the Wdreb2alpha and Wdreb2gamma forms showed transient increases. On the other hand, the LT treatment resulted in increased transcript levels of all three forms of Wdreb2. Thus, under the LT and drought/salt stress conditions the amount of the WDREB2 transcription factors in wheat is differentially controlled by the level of transcription and alternative splicing.
DOI:10.1016/j.molp.2014.10.011URLPMID:25680774 [本文引用: 1]
Environmental stresses profoundly altered accumulation of nonsense mRNAs including intron-retaining (IR) transcripts in Arabidopsis. Temporal patterns of stress-induced IR mRNAs were dissected using both oscillating and non-oscillating transcripts. Broad-range thermal cycles triggered a sharp increase in the long IR CCA1 isoforms and altered their phasing to different times of day. Both abiotic and biotic stresses such as drought or Pseudomonas syringae infection induced a similar increase. Thermal stress induced a time delay in accumulation of CCA1 I4Rb transcripts, whereas functional mRNA showed steady oscillations. Our data favor a hypothesis that stress-induced instabilities of the central oscillator can be in part compensated through fluctuations in abundance and out-of-phase oscillations of CCA1 IR transcripts. Taken together, our results support a concept that mRNA abundance can be modulated through altering ratios between functional and nonsense/IR transcripts. SR45 protein specifically bound to the retained CCA1 intron in vitro, suggesting that this splicing factor could be involved in regulation of intron retention. Transcriptomes of nonsense-mediated mRNA decay (NMD)-impaired and heat-stressed plants shared a set of retained introns associated with stress- and defense-inducible transcripts. Constitutive activation of certain stress response networks in an NMD mutant could be linked to disequilibrium between functional and nonsense mRNAs.
URLPMID:18317505 [本文引用: 1]
[本文引用: 1]
DOI:10.1093/nar/gkt296URLPMID:23620288 [本文引用: 1]
Sm-like (Lsm) proteins have been identified in all organisms and are related to RNA metabolism. Here, we report that Arabidopsis nuclear AtLSM8 protein, as well as AtLSM5, which localizes to both the cytoplasm and nucleus, function in pre-mRNA splicing, while AtLSM5 and the exclusively cytoplasmic AtLSM1 contribute to 5'-3' mRNA decay. In lsm8 and sad1/lsm5 mutants, U6 small nuclear RNA (snRNA) was reduced and unspliced mRNA precursors accumulated, whereas mRNA stability was mainly affected in plants lacking AtLSM1 and AtLSM5. Some of the mRNAs affected in lsm1a lsm1b and sad1/lsm5 plants were also substrates of the cytoplasmic 5'-3' exonuclease AtXRN4 and of the decapping enzyme AtDCP2. Surprisingly, a subset of substrates was also stabilized in the mutant lacking AtLSM8, which supports the notion that plant mRNAs are actively degraded in the nucleus. Localization of LSM components, purification of LSM-interacting proteins as well as functional analyses strongly suggest that at least two LSM complexes with conserved activities in RNA metabolism, AtLSM1-7 and AtLSM2-8, exist also in plants.
DOI:10.1111/j.1467-7652.2012.00741.xURLPMID:22967050 [本文引用: 1]
High temperature impairs rice (Oryza sativa) grain filling by inhibiting the deposition of storage materials such as starch, resulting in mature grains with a chalky appearance, currently a major problem for rice farming in Asian countries. Such deterioration of grain quality is accompanied by the altered expression of starch metabolism-related genes. Here we report the involvement of a starch-hydrolyzing enzyme, alpha-amylase, in high temperature-triggered grain chalkiness. In developing seeds, high temperature induced the expression of alpha-amylase genes, namely Amy1A, Amy1C, Amy3A, Amy3D and Amy3E, as well as alpha-amylase activity, while it decreased an alpha-amylase-repressing plant hormone, ABA, suggesting starch to be degraded by alpha-amylase in developing grains under elevated temperature. Furthermore, RNAi-mediated suppression of alpha-amylase genes in ripening seeds resulted in fewer chalky grains under high-temperature conditions. As the extent of the decrease in chalky grains was highly correlated to decreases in the expression of Amy1A, Amy1C, Amy3A and Amy3B, these genes would be involved in the chalkiness through degradation of starch accumulating in the developing grains. The results show that activation of alpha-amylase by high temperature is a crucial trigger for grain chalkiness and that its suppression is a potential strategy for ameliorating grain damage from global warming.
[本文引用: 1]
[本文引用: 1]
DOI:10.1038/5256URLPMID:9920274 [本文引用: 2]
Using an Agrobacterium-mediated transformation system, we have introduced the intact gene of maize phosphoenolpyruvate carboxylase (PEPC), which catalyzes the initial fixation of atmospheric CO2 in C4 plants into the C3 crop rice. Most transgenic rice plants showed high-level expression of the maize gene; the activities of PEPC in leaves of some transgenic plants were two- to threefold higher than those in maize, and the enzyme accounted for up to 12% of the total leaf soluble protein. RNA gel blot and Southern blot analyses showed that the level of expression of the maize PEPC in transgenic rice plants correlated with the amount of transcript and the copy number of the inserted maize gene. Physiologically, the transgenic plants exhibited reduced O2 inhibition of photosynthesis and photosynthetic rates comparable to those of untransformed plants. The results demonstrate a successful strategy for installing the key biochemical component of the C4 pathway of photosynthesis in C3 plants.
DOI:10.3109/07388551.2012.716809URLPMID:22985089 [本文引用: 1]
Foxtail millet is one of the oldest domesticated diploid C4 Panicoid crops having a comparatively small genome size of approximately 515 Mb, short life cycle, and inbreeding nature. Its two species, Setaria italica (domesticated) and Setaria viridis (wild progenitor), have characteristics that classify them as excellent model systems to examine several aspects of architectural, evolutionary, and physiological importance in Panicoid grasses especially the biofuel crops such as switchgrass and napiergrass. Foxtail millet is a staple crop used extensively for food and fodder in parts of Asia and Africa. In its long history of cultivation, it has been adapted to arid and semi-arid areas of Asia, North Africa, South and North America. Foxtail millet has one of the largest collections of cultivated as well as wild-type germplasm rich with phenotypic variations and hence provides prospects for association mapping and allele-mining of elite and novel variants to be incorporated in crop improvement programs. Most of the foxtail millet accessions can be primarily abiotic stress tolerant particularly to drought and salinity, and therefore exploiting these agronomic traits can enhance its efficacy in marker-aided breeding as well as in genetic engineering for abiotic stress tolerance. In addition, the release of draft genome sequence of foxtail millet would be useful to the researchers worldwide in not only discerning the molecular basis of biomass production in biofuel crops and the methods to improve it, but also for the introgression of beneficial agronomically important characteristics in foxtail millet as well as in related Panicoid bioenergy grasses.
[本文引用: 1]
DOI:10.1016/j.tig.2015.03.002URLPMID:25837375 [本文引用: 1]
Although DNA methylation was originally thought to only affect transcription, emerging evidence shows that it also regulates alternative splicing. Exons, and especially splice sites, have higher levels of DNA methylation than flanking introns, and the splicing of about 22% of alternative exons is regulated by DNA methylation. Two different mechanisms convey DNA methylation information into the regulation of alternative splicing. The first involves modulation of the elongation rate of RNA polymerase II (Pol II) by CCCTC-binding factor (CTCF) and methyl-CpG binding protein 2 (MeCP2); the second involves the formation of a protein bridge by heterochromatin protein 1 (HP1) that recruits splicing factors onto transcribed alternative exons. These two mechanisms, however, regulate only a fraction of such events, implying that more underlying mechanisms remain to be found.
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/tpj.13383URLPMID:27664942 [本文引用: 2]
Alternative splicing (AS) of precursor RNAs enhances transcriptome plasticity and proteome diversity in response to diverse growth and stress cues. Recent work has shown that AS is pervasive across plant species, with more than 60% of intron-containing genes producing different isoforms. Mammalian cell-based assays have discovered various inhibitors of AS. Here, we show that the macrolide pladienolide B (PB) inhibits constitutive splicing and AS in plants. Also, our RNA sequencing (RNA-seq) data revealed that PB mimics abiotic stress signals including salt, drought and abscisic acid (ABA). PB activates the abiotic stress- and ABA-responsive reporters RD29A::LUC and MAPKKK18::uidA in Arabidopsis thaliana and mimics the effects of ABA on stomatal aperture. Genome-wide analysis of AS by RNA-seq revealed that PB perturbs the splicing machinery and leads to a striking increase in intron retention and a reduction in other forms of AS. Interestingly, PB treatment activates the ABA signaling pathway by inhibiting the splicing of clade A PP2C phosphatases while still maintaining to some extent the splicing of ABA-activated SnRK2 kinases. Taken together, our data establish PB as an inhibitor and modulator of splicing and a mimic of abiotic stress signals in plants. Thus, PB reveals the molecular underpinnings of the interplay between stress responses, ABA signaling and post-transcriptional regulation in plants.
URLPMID:23503691 [本文引用: 1]
URLPMID:27592839 [本文引用: 2]
DOI:10.1006/meth.2001.1262URLPMID:11846609 [本文引用: 1]
The two most commonly used methods to analyze data from real-time, quantitative PCR experiments are absolute quantification and relative quantification. Absolute quantification determines the input copy number, usually by relating the PCR signal to a standard curve. Relative quantification relates the PCR signal of the target transcript in a treatment group to that of another sample such as an untreated control. The 2(-Delta Delta C(T)) method is a convenient way to analyze the relative changes in gene expression from real-time quantitative PCR experiments. The purpose of this report is to present the derivation, assumptions, and applications of the 2(-Delta Delta C(T)) method. In addition, we present the derivation and applications of two variations of the 2(-Delta Delta C(T)) method that may be useful in the analysis of real-time, quantitative PCR data.
DOI:10.1101/gr.134106.111URLPMID:22391557 [本文引用: 1]
Alternative splicing (AS) is a key regulatory mechanism that contributes to transcriptome and proteome diversity. As very few genome-wide studies analyzing AS in plants are available, we have performed high-throughput sequencing of a normalized cDNA library which resulted in a high coverage transcriptome map of Arabidopsis. We detect approximately 150,000 splice junctions derived mostly from typical plant introns, including an eightfold increase in the number of U12 introns (2069). Around 61% of multiexonic genes are alternatively spliced under normal growth conditions. Moreover, we provide experimental validation of 540 AS transcripts (from 256 genes coding for important regulatory factors) using high-resolution RT-PCR and Sanger sequencing. Intron retention (IR) is the most frequent AS event ( approximately 40%), but many IRs have relatively low read coverage and are less well-represented in assembled transcripts. Additionally, approximately 51% of Arabidopsis genes produce AS transcripts which do not involve IR. Therefore, the significance of IR in generating transcript diversity was generally overestimated in previous assessments. IR analysis allowed the identification of a large set of cryptic introns inside annotated coding exons. Importantly, a significant fraction of these cryptic introns are spliced out in frame, indicating a role in protein diversity. Furthermore, we show extensive AS coupled to nonsense-mediated decay in AFC2, encoding a highly conserved LAMMER kinase which phosphorylates splicing factors, thus establishing a complex loop in AS regulation. We provide the most comprehensive analysis of AS to date which will serve as a valuable resource for the plant community to study transcriptome complexity and gene regulation.
URLPMID:14630963 [本文引用: 1]
[本文引用: 1]
DOI:10.1371/journal.pone.0109920URLPMID:25279462 [本文引用: 1]
MYB proteins represent one of the largest transcription factor families in plants, playing important roles in diverse developmental and stress-responsive processes. Considering its significance, several genome-wide analyses have been conducted in almost all land plants except foxtail millet. Foxtail millet (Setaria italica L.) is a model crop for investigating systems biology of millets and bioenergy grasses. Further, the crop is also known for its potential abiotic stress-tolerance. In this context, a comprehensive genome-wide survey was conducted and 209 MYB protein-encoding genes were identified in foxtail millet. All 209 S. italica MYB (SiMYB) genes were physically mapped onto nine chromosomes of foxtail millet. Gene duplication study showed that segmental- and tandem-duplication have occurred in genome resulting in expansion of this gene family. The protein domain investigation classified SiMYB proteins into three classes according to number of MYB repeats present. The phylogenetic analysis categorized SiMYBs into ten groups (I-X). SiMYB-based comparative mapping revealed a maximum orthology between foxtail millet and sorghum, followed by maize, rice and Brachypodium. Heat map analysis showed tissue-specific expression pattern of predominant SiMYB genes. Expression profiling of candidate MYB genes against abiotic stresses and hormone treatments using qRT-PCR revealed specific and/or overlapping expression patterns of SiMYBs. Taken together, the present study provides a foundation for evolutionary and functional characterization of MYB TFs in foxtail millet to dissect their functions in response to environmental stimuli.
DOI:10.3389/fpls.2017.02089URLPMID:29270189 [本文引用: 1]
Global warming impairs grain filling in rice and reduces starch accumulation in the endosperm, leading to chalky-appearing grains, which damages their market value. We found previously that high temperature-induced expression of starch-lytic alpha-amylases during ripening is crucial for grain chalkiness. Because the rice genome carries at least eight functional alpha-amylase genes, identification of the alpha-amylase(s) that contribute most strongly to the production of chalky grains could accelerate efficient breeding. To identify alpha-amylase genes responsible for the production of chalky grains, we characterized the histological expression pattern of eight alpha-amylase genes and the influences of their overexpression on grain appearance and carbohydrate components through a series of experiments with transgenic rice plants. The promoter activity of most alpha-amylase genes was elevated to various extents at high temperature. Among them, the expression of Amy1A and Amy3C was induced in the internal, especially basal to dorsal, region of developing endosperm, whereas that of Amy3D was confined near the ventral aleurone. These regions coincided with the site of occurrence of chalkiness, which was in clear contrast to conventionally known expression patterns of the enzyme in the scutellum and aleurone during seed germination. Furthermore, overexpression of alpha-amylase genes, except for Amy3E, in developing endosperm produced various degrees of chalky grains without heat exposure, whereas that of Amy3E yielded normal translucent grains, as was the case in the vector control, even though Amy3E-overexpressing grains contained enhanced alpha-amylase activities. The weight of the chalky grains was decreased due to reduced amounts of starch, and microscopic observation of the chalky part of these grains revealed that their endosperm consisted of loosely packed round starch granules that had numerous pits on their surface, confirming the hydrolysis of the starch reserve by alpha-amylases. Moreover, the chalky grains contained increased amounts of soluble sugars including maltooligosaccharides at the expense of starch. The integrated analyses proposed that expression of Amy1A, Amy3C, and Amy3D at the specific regions of the developing endosperm could generate the chalkiness. This finding provides the fundamental knowledge to narrow down the targets for the development of high temperature-tolerant premium rice.
DOI:10.1271/bbb.68.112URLPMID:14745172 [本文引用: 1]
We isolated and identified 10 alpha-amylase isoforms by using beta-cyclodextrin Sepharose affinity column chromatography and two-dimensional polyacrylamide gel electrophoresis from germinating rice (Oryza sativa L.) seeds. Immunoblots with anti-alpha-amylase I-1 and II-4 antibodies indicated that 8 isoforms in 10 are distinguishable from alpha-amylase I-1 and II-4. Peptide mass fingerprinting analysis showed that there exist novel isoforms encoded by RAmy3B and RAmy3C genes. The optimum temperature for enzyme reaction of the RAmy3B and RAmy3C coding isoforms resembled that of alpha-amylase isoform II-4 (RAmy3D). Furthermore, complex protein polymorphism resulted from a single alpha-amylase gene was found to occur not only in RAmy3D, but also in RAmy3B.
URLPMID:21524275 [本文引用: 1]
[本文引用: 1]
DOI:10.1111/jipb.12283URLPMID:25231250 [本文引用: 2]
To understand the molecular responses of PC (Overexpressing the maize C4-pepc gene, which encodes phosphoenolpyruvate carboxylase (PEPC)), to drought stress at cell level, we analyzed changes in the levels of signaling molecules (hydrogen peroxide (H2O2), calcium ion (Ca(2+)), and nitric oxide (NO)) in suspension-cultured PC and wild-type (WT) rice (Oryza sativa L.) cell under drought stress induced by 20% polyethylene glycol 6000 (PEG-6000). Results demonstrated that PC improved drought tolerance by enhancing antioxidant defense, retaining higher relative water content, survival percentages, and dry weight of cells. In addition, PEPC activity in PC under PEG treatment was strengthened by addition of H2O2 inhibitor, dimethylthiourea (DMTU) and NO synthesis inhibitor, 2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide (cPTIO), respectively, while that in PC was weakened by addition of free calcium chelator, ethylene glycol-bis(b-aminoethylether)-N,N,N',N'-tetraacetic acid (EGTA) + calcium channel outflow inhibitor, ruthenium red (RR) + plasma membrane channel blocker La(NO3)3, but EGTA + RR did not. Results also showed that NO and Ca(2+) was lying downstream of H2O2 in drought-induced signaling. Calcium ion was also involved in the expression of C4-pepc in PC. These results suggested that PC could improve oxidative tolerance in suspension-cultured cells and the acquisition of this tolerance required downregulation of H2O2 and the entry of extracellular Ca(2+) into cells across the plasma membrane for regulation of PEPC activity and C4-pepc expression.
DOI:10.1002/wrna.98URLPMID:21766458 [本文引用: 2]
Global analyses of splicing of precursor messenger RNAs (pre-mRNAs) have revealed that alternative splicing (AS) is highly pervasive in plants. Despite the widespread occurrence of AS in plants, the mechanisms that control splicing and the roles of splice variants generated from a gene are poorly understood. Studies on plant serine/arginine-rich (SR) proteins, a family of highly conserved proteins, suggest their role in both constitutive splicing and AS of pre-mRNAs. SR proteins have a characteristic domain structure consisting of one or two RNA recognition motifs at the N-terminus and a C-terminal RS domain rich in arginine/serine dipeptides. Plants have many more SR proteins compared to animals including several plant-specific subfamilies. Pre-mRNAs of plant SR proteins are extensively alternatively spliced to increase the transcript complexity by about six-fold. Some of this AS is controlled in a tissue- and development-specific manner. Furthermore, AS of SR pre-mRNAs is altered by various stresses, raising the possibility of rapid reprogramming of the whole transcriptome by external signals through regulation of the splicing of these master regulators of splicing. Most SR splice variants contain a premature termination codon and are degraded by up-frameshift 3 (UPF3)-mediated nonsense-mediated decay (NMD), suggesting a link between NMD and regulation of expression of the functional transcripts of SR proteins. Limited functional studies with plant SRs suggest key roles in growth and development and plant responses to the environment. Here, we discuss the current status of research on plant SRs and some promising approaches to address many unanswered questions about plant SRs.
DOI:10.1016/j.plaphy.2013.11.011URLPMID:24316011 [本文引用: 1]
In this study, we investigated the photosynthetic physiological basis in 'PC' transgenic rice (Oryza sativa L.), showing high-level expression of the gene encoding C4 phosphoenolpyruvate carboxylase (pepc), by hydrogen peroxide (H2O2). The C4-PEPC gene (pepc) from maize in the transgenic rice plants was checked by PCR. Comparison of yield components and photosynthetic indices between PC and untransformed wild-type (WT) plants indicated that increased yield in PC was associated with higher net photosynthetic rate and higher activities of phosphoenolpyruvate carboxylase (PEPC). Both PC and WT plants were treated with 1 mmol L(-1) abscisic acid (ABA), 0.04% 1-butanol (BA), 2 mmol L(-1) neomycin (NS), or 2 mmol L(-1) diphenyleneiodonium chloride (DPI) to investigate the relationship between photosynthesis and levels of H2O2 and phosphatidic acid. In both PC and WT, ABA induced H2O2 generation and simultaneous decrease in stomatal conductance (g(s)). PC plants treated with BA showed decreased H2O2 content and strongly increased g(s) within 2 h of treatment. Similar results were observed in response to DPI treatment in PC. However, WT did not observe the decrease of H2O2 during the treatments of BA and DPI. The reduced H2O2 content in PC caused by BA treatment differed to that induced by DPI because BA did not inhibit NADPH oxidase activities. While BA induced a larger PEPC activity in PC, and higher catalase activity as well. These results indicated that the regulation of endogenous H2O2 metabolism of PC could be helpful for enhancing photosynthetic capability.
DOI:10.1016/0003-2697(87)90172-2URLPMID:3442328 [本文引用: 1]
Two milliliters of a reagent consisting of anthrone and tryptophan each at a 0.01% concentration in 75% sulfuric acid, when added to 0.9 ml of solution containing D-fructose, produced on heating (55 degrees C, 90 min) a pink color (lambdamax 520 nm) with an absorbance of 0.009 A/nmol. The absorbance is about three times higher than that of the standard anthrone-sulfuric acid reagent. Glucose yields a color only about 1% as intense as that yielded by fructose. The method is useful for the estimation of fructose in the presence of proteins. Synthetic Amadori compounds and glycosylated proteins containing ketoamine-linked fructose, however, were unreactive.
DOI:10.1105/tpc.113.113803URLPMID:24179132 [本文引用: 1]
High-throughput sequencing for transcript profiling in plants has revealed that alternative splicing (AS) affects a much higher proportion of the transcriptome than was previously assumed. AS is involved in most plant processes and is particularly prevalent in plants exposed to environmental stress. The identification of mutations in predicted splicing factors and spliceosomal proteins that affect cell fate, the circadian clock, plant defense, and tolerance/sensitivity to abiotic stress all point to a fundamental role of splicing/AS in plant growth, development, and responses to external cues. Splicing factors affect the AS of multiple downstream target genes, thereby transferring signals to alter gene expression via splicing factor/AS networks. The last two to three years have seen an ever-increasing number of examples of functional AS. At a time when the identification of AS in individual genes and at a global level is exploding, this review aims to bring together such examples to illustrate the extent and importance of AS, which are not always obvious from individual publications. It also aims to ensure that plant scientists are aware that AS is likely to occur in the genes that they study and that dynamic changes in AS and its consequences need to be considered routinely.
DOI:10.1016/j.tplants.2012.06.001URLPMID:22743067 [本文引用: 1]
More than 60% of intron-containing genes undergo alternative splicing (AS) in plants. This number will increase when AS in different tissues, developmental stages, and environmental conditions are explored. Although the functional impact of AS on protein complexity is still understudied in plants, recent examples demonstrate its importance in regulating plant processes. AS also regulates transcript levels and the link with nonsense-mediated decay and generation of unproductive mRNAs illustrate the need for both transcriptional and AS data in gene expression analyses. AS has influenced the evolution of the complex networks of regulation of gene expression and variation in AS contributed to adaptation of plants to their environment and therefore will impact strategies for improving plant and crop phenotypes.
DOI:10.1016/j.plaphy.2018.08.013URLPMID:30114676 [本文引用: 1]
The objective of this study was to reveal the physiological and molecular mechanisms of low-nitrogen (N) tolerance in transgenic plant lines containing C4 phosphoenolpyruvate carboxylase (C4-PEPC) gene. The transgenic rice lines only over-expressing the maize C4-PEPC) (PC) and their untransformed wild type, Kitaake (WT), were used in this study. At different N levels, the dry weight, total N content, carbon and N levels, photorespiration-related enzymatic activities, gene expression levels and photorespiration-related product accumulations were measured, as were the transgenic lines' agronomic traits. The PC line, having lower total N and higher soluble sugar contents, was more tolerant to low-N stress than WT, which was consistent with its higher PEPC and lower N-assimilation-related enzyme activity levels. The photosynthetic parameters, enzymatic activity levels, transcripts and products related to photorespiration in PC were also greater than in WT under low-N conditions. This study showed that increased carbon levels in transgenic rice lines overexpressing C4-PEPC could help regulate the photorespiratory pathway under low-N conditions, conferring low-N tolerance and a higher grain yield per plant.
DOI:10.1073/pnas.0602039103URLPMID:16632598 [本文引用: 1]
Alternative splicing (AS) has been extensively studied in mammalian systems but much less in plants. Here we report AS events deduced from EST/cDNA analysis in two model plants: Arabidopsis and rice. In Arabidopsis, 4,707 (21.8%) of the genes with EST/cDNA evidence show 8,264 AS events. Approximately 56% of these events are intron retention (IntronR), and only 8% are exon skipping. In rice, 6,568 (21.2%) of the expressed genes display 14,542 AS events, of which 53.5% are IntronR and 13.8% are exon skipping. The consistent high frequency of IntronR suggests prevalence of splice site recognition by intron definition in plants. Different AS events within a given gene occur, for the most part, independently. In total, 36-43% of the AS events produce transcripts that would be targets of the non-sense-mediated decay pathway, if that pathway were to operate in plants as in humans. Forty percent of Arabidopsis AS genes are alternatively spliced also in rice, with some examples strongly suggesting a role of the AS event as an evolutionary conserved mechanism of posttranscriptional regulation. We created a comprehensive web-interfaced database to compile and visualize the evidence for alternative splicing in plants (Alternative Splicing in Plants, available at www.plantgdb.org/ASIP).
DOI:10.1111/j.1365-313X.2010.04246.xURLPMID:20487381 [本文引用: 1]
We conducted genome-wide mapping of cytosine methylation using methylcytosine immunoprecipitation combined with Illumina sequencing. The chromosomal distribution pattern of methylated DNA is similar to the heterochromatin distribution pattern on rice chromosomes. The DNA methylation patterns of rice genes are similar to those in Arabidopsis thaliana, including distinct methylation patterns asssociated with gene bodies and promoters. The DNA sequences in the core domains of rice Cen4, Cen5 and Cen8 showed elevated methylation levels compared with sequences in the pericentromeric regions. In addition, elevated methylation levels were associated with the DNA sequences in the CENH3-binding subdomains, compared with the sequences in the flanking H3 subdomains. In contrast, the centromeric domain of Cen11, which is composed exclusively of centromeric satellite DNA, is hypomethylated compared with the pericentromeric domains. Thus, the DNA sequences associated with functional centromeres can be either hypomethylated or hypermethylated. The methylation patterns of centromeric DNA appear to be correlated with the composition of the associated DNA sequences. We propose that both hypomethylation and hypermethylation of CENH3-associated DNA sequences can serve as epigenetic marks to distinguish where CENH3 deposition will occur within the surrounding H3 chromatin.
URLPMID:28415033 [本文引用: 2]
DOI:10.1016/S2095-3119(13)60260-9URL [本文引用: 1]
DOI:10.1104/pp.107.108720URLPMID:17951452 [本文引用: 1]
The Arabidopsis (Arabidopsis thaliana) disease resistance protein RESISTANCE TO PSEUDOMONAS SYRINGAE4 (RPS4) activates defenses in response to bacterial pathogens expressing avrRps4 in a gene-for-gene specific manner. The RPS4 gene produces multiple transcripts via alternative splicing of two regular introns flanking exon 3 and a cryptic intron within exon 3. We showed previously that RPS4-mediated resistance requires the combined presence of transcripts encoding both full-length and truncated open reading frames. Here, we demonstrate that alternative splicing of RPS4 undergoes dynamic changes specifically during the resistance response. Furthermore, RPS4 expression was induced by the presence of AvrRps4 in an EDS1-dependent manner. Interestingly, inducible alternative splicing was not limited to the avrRps4-RPS4 interaction, indicating that regulation of alternative splicing may be a general response to prime the plant stress response system. Intron-deficient transgenes lacking only one intron were previously shown to be nonfunctional. Here, we establish quantitatively that the absence of one intron had no effect on the splicing frequency of remaining introns. Given the lack of functionality of single intron-deficient transgenes, this suggests that the products of individual transcripts have distinct functions during RPS4-triggered resistance. Transient expression of truncated RPS4 proteins in Nicotiana benthamiana induced hypersensitive response-like cell death in the absence of AvrRps4. Interestingly, different truncated proteins had markedly differing stability. In summary, RPS4 function is regulated at multiple levels, including gene expression, alternative splicing, and protein stability, presumably to fine-tune activity and limit damage inflicted by activated RPS4 protein.
基于小麦种子发芽逆境抗逆指数的种子活力评价
1
2016
... 参照标准发芽试验法(
表观遗传调控植物响应非生物胁迫的研究进展
1
2018
... 为了能够在胁迫条件下生存, 植物进化出复杂的机制来感知外部信号, 从而对环境变化做出最佳反应, 其中表观遗传机制调控植物应答外界胁迫是近年来发现的重要机制之一, DNA甲基化和可变剪接均是其中的重要途径(
转PEPC基因水稻具有初级CO2浓缩机制的生理特点
1
2003
... 近年来, 在全球气候变化背景下, 干旱频繁发生且日趋严峻, 严重影响作物的生长, 甚至造成减产(
转PEPC基因水稻的光合CO2同化和叶绿素荧光特性
1
2001
... 近年来, 在全球气候变化背景下, 干旱频繁发生且日趋严峻, 严重影响作物的生长, 甚至造成减产(
1
2000
... 参考3,5-二硝基水杨酸(DNS)法(
小麦干热风抗性鉴定及热胁迫相关基因TaHSPs的表达分析
1
2017
... 有效的种子发芽使幼苗健壮地生长, 并具有抵御生物或非生物胁迫的能力, 从而最终发展成为高产作物(
外源Ca2+对PEG处理下转C4型PEPC基因水稻光合生理的调节
1
2015
... 近年来, 在全球气候变化背景下, 干旱频繁发生且日趋严峻, 严重影响作物的生长, 甚至造成减产(
mRNA选择性剪切在植物发育中的作用
1
2014
... 可变剪接(alternative splicing, AS)是一种生物体基因转录后调控的方式, 植物中的许多基因存在AS, 其在植物生长发育中具有重要作用(
可变剪接在植物响应胁迫中的作用
1
2020
... 为了能够在胁迫条件下生存, 植物进化出复杂的机制来感知外部信号, 从而对环境变化做出最佳反应, 其中表观遗传机制调控植物应答外界胁迫是近年来发现的重要机制之一, DNA甲基化和可变剪接均是其中的重要途径(
外源葡萄糖增强高表达转玉米C4型PEPC水稻耐旱性的生理机制
1
2018
... 近年来, 在全球气候变化背景下, 干旱频繁发生且日趋严峻, 严重影响作物的生长, 甚至造成减产(
植物SnRKs家族在胁迫信号通路中的调节作用
1
2017
... 丝氨酸/苏氨酸蛋白激酶是一个相对较大的蛋白激酶家族, 其中AMPK (AMP-activated kinase)/ SnRKs是高度保守的丝氨酸/苏氨酸蛋白激酶家族的重要成员.SnRKs可以细分成3个亚家族(SnRK1、SnRK2和SnRK3).SnRK1主要在调节植株生长发育、维持能量平衡以及响应生物逆境方面发挥重要作用; SnRK2则在响应干旱以及盐碱等非生物胁迫中发挥重要作用; SnRK3更多地参与胞内离子浓度的平衡, 通过调节离子浓度迅速启动植株响应非生物胁迫(
Coordinated regulation of photosynthesis in rice increases yield and tolerance to environmental stress
1
2014
... 蔗糖、葡萄糖和果糖含量参照文献(
A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding
1
1976
... 采用考马斯亮蓝G-250法(
Induction of vacuolar invertase inhibitor mRNA in potato tubers contributes to cold-induced sweetening resistance and includes spliced hybrid mRNA variants
1
2011
... 为了能够在胁迫条件下生存, 植物进化出复杂的机制来感知外部信号, 从而对环境变化做出最佳反应, 其中表观遗传机制调控植物应答外界胁迫是近年来发现的重要机制之一, DNA甲基化和可变剪接均是其中的重要途径(
To splice or to transcribe: SKIP- mediated environmental fitness and development in plants
1
2019
... 可变剪接(alternative splicing, AS)是一种生物体基因转录后调控的方式, 植物中的许多基因存在AS, 其在植物生长发育中具有重要作用(
Promotion of photosynthesis in transgenic rice over-expressing of maize C4 phosphoenolpyruvate carboxylase gene by nitric oxide donors
1
2014
... 近年来, 在全球气候变化背景下, 干旱频繁发生且日趋严峻, 严重影响作物的生长, 甚至造成减产(
Abscisic acid (ABA) regulation of Arabidopsis SR protein gene expression
1
2014
... 丝氨酸/精氨酸丰富蛋白家族(arginine/serine-rich proteins, SR proteins)是真核生物中的一类剪接因子, 在前体mRNA的组成性和选择性剪接中起作用(
The rice α-amylase, conserved regulator of seed maturation and germination
1
2019
... 有效的种子发芽使幼苗健壮地生长, 并具有抵御生物或非生物胁迫的能力, 从而最终发展成为高产作物(
Alternatively spliced N resistance gene transcripts: their possible role in tobacco mosaic virus resistance
1
2000
... 为了能够在胁迫条件下生存, 植物进化出复杂的机制来感知外部信号, 从而对环境变化做出最佳反应, 其中表观遗传机制调控植物应答外界胁迫是近年来发现的重要机制之一, DNA甲基化和可变剪接均是其中的重要途径(
Tight translational control by the initiation factors eIF4E and eIF(iso)4E is required for maize seed germination
1
2011
... 有效的种子发芽使幼苗健壮地生长, 并具有抵御生物或非生物胁迫的能力, 从而最终发展成为高产作物(
Alternative splicing plays a critical role in maintaining mineral nutrient homeostasis in rice (Oryza sativa)
2
2018
... 采用TaKaRa Mini BEST Plant RNA Extraction Kit (TaKaRa, Cat No.9769)试剂盒提取总RNA.采用TaKaRa Prime Script RT Master Mix Perfect Real Time (TaKaRa, Cat No.RR036A)试剂盒进行反转录.Real Time PCR采用TB Green Premix Ex Taq Tli RNaseH Plus (TaKaRa, Cat No.RR82LR)试剂盒进行.SR基因引物序列来自
... 丝氨酸/精氨酸丰富蛋白家族(arginine/serine-rich proteins, SR proteins)是真核生物中的一类剪接因子, 在前体mRNA的组成性和选择性剪接中起作用(
Differential regulation of transcript accumulation and alternative splicing of a DREB2 homolog under abiotic stress conditions in common wheat
1
2006
... 为了能够在胁迫条件下生存, 植物进化出复杂的机制来感知外部信号, 从而对环境变化做出最佳反应, 其中表观遗传机制调控植物应答外界胁迫是近年来发现的重要机制之一, DNA甲基化和可变剪接均是其中的重要途径(
Environmental stresses modulate abundance and timing of alternatively spliced circadian transcripts in Arabidopsis
1
2015
... 为了能够在胁迫条件下生存, 植物进化出复杂的机制来感知外部信号, 从而对环境变化做出最佳反应, 其中表观遗传机制调控植物应答外界胁迫是近年来发现的重要机制之一, DNA甲基化和可变剪接均是其中的重要途径(
Common QTL affect the rate of tomato seed germination under different stress and nonstress conditions
1
2007
... 有效的种子发芽使幼苗健壮地生长, 并具有抵御生物或非生物胁迫的能力, 从而最终发展成为高产作物(
Alternative splicing in plant defense
1
2008
... 为了能够在胁迫条件下生存, 植物进化出复杂的机制来感知外部信号, 从而对环境变化做出最佳反应, 其中表观遗传机制调控植物应答外界胁迫是近年来发现的重要机制之一, DNA甲基化和可变剪接均是其中的重要途径(
Arabidopsis thaliana LSM proteins function in mRNA splicing and degradation
1
2013
... 为了能够在胁迫条件下生存, 植物进化出复杂的机制来感知外部信号, 从而对环境变化做出最佳反应, 其中表观遗传机制调控植物应答外界胁迫是近年来发现的重要机制之一, DNA甲基化和可变剪接均是其中的重要途径(
Suppression of α-amylase genes improves quality of rice grain ripened under high temperature
1
2012
... 有效的种子发芽使幼苗健壮地生长, 并具有抵御生物或非生物胁迫的能力, 从而最终发展成为高产作物(
Drought tolerance of transgenic rice overexpressing maize C4-PEPC gene related to increased anthocyanin synthesis regulated by sucrose and calcium
1
2020
... 近年来, 在全球气候变化背景下, 干旱频繁发生且日趋严峻, 严重影响作物的生长, 甚至造成减产(
Inhibitory effect of chemical combinations on seed germination and pre-harvest sprouting in hybrid rice
1
2016
... 有效的种子发芽使幼苗健壮地生长, 并具有抵御生物或非生物胁迫的能力, 从而最终发展成为高产作物(
High-level expression of maize phosphoenolpyruvate carboxylase in transgenic rice plants
2
1999
... 近年来, 在全球气候变化背景下, 干旱频繁发生且日趋严峻, 严重影响作物的生长, 甚至造成减产(
... 本研究所用的转基因水稻(Oryza sativa L.)是由
Foxtail millet: a model crop for genetic and genomic studies in bioenergy grasses
1
2013
... C4植物的磷酸烯醇式丙酮酸羧化酶可将二氧化碳输送至核酮糖-1,5-二磷酸羧化酶(Ribulose-1,5-bisphosphate carboxylase, Rubisco), 在高光强和高温下也能更快地进行光合作用, 因此C4植物光合作用效率高, 并具有更好的水分利用率(
Physiological impacts of modulating phosphoenolpyruvate carboxylase levels in leaves and seeds of Arabidopsis thaliana
1
2007
... 近年来, 在全球气候变化背景下, 干旱频繁发生且日趋严峻, 严重影响作物的生长, 甚至造成减产(
The alternative role of DNA methylation in splicing regulation
1
2015
... 可变剪接(alternative splicing, AS)是一种生物体基因转录后调控的方式, 植物中的许多基因存在AS, 其在植物生长发育中具有重要作用(
Physiological and metabolic enzymes activity changes in transgenic rice plants with increased phosphoenolpyruvate carboxylase activity during the flowering stage
1
2013
... 近年来, 在全球气候变化背景下, 干旱频繁发生且日趋严峻, 严重影响作物的生长, 甚至造成减产(
Effects of 1-butanol, neomycin and calcium on the photosynthetic characteristics of pepc transgenic rice
1
2011
... 近年来, 在全球气候变化背景下, 干旱频繁发生且日趋严峻, 严重影响作物的生长, 甚至造成减产(
Pre-mRNA splicing repression triggers abiotic stress signaling in plants
2
2017
... 可变剪接(alternative splicing, AS)是一种生物体基因转录后调控的方式, 植物中的许多基因存在AS, 其在植物生长发育中具有重要作用(
... 为了能够在胁迫条件下生存, 植物进化出复杂的机制来感知外部信号, 从而对环境变化做出最佳反应, 其中表观遗传机制调控植物应答外界胁迫是近年来发现的重要机制之一, DNA甲基化和可变剪接均是其中的重要途径(
An autoregulatory loop controlling Arabidopsis HsfA2 expression: role of heat shock-induced alternative splicing
1
2013
... 为了能够在胁迫条件下生存, 植物进化出复杂的机制来感知外部信号, 从而对环境变化做出最佳反应, 其中表观遗传机制调控植物应答外界胁迫是近年来发现的重要机制之一, DNA甲基化和可变剪接均是其中的重要途径(
Phosphoenolpyruvate carboxylase regulation in C4-PEPC-expressing transgenic rice during early responses to drought stress
2
2017
... 近年来, 在全球气候变化背景下, 干旱频繁发生且日趋严峻, 严重影响作物的生长, 甚至造成减产(
... 丝氨酸/苏氨酸蛋白激酶是一个相对较大的蛋白激酶家族, 其中AMPK (AMP-activated kinase)/ SnRKs是高度保守的丝氨酸/苏氨酸蛋白激酶家族的重要成员.SnRKs可以细分成3个亚家族(SnRK1、SnRK2和SnRK3).SnRK1主要在调节植株生长发育、维持能量平衡以及响应生物逆境方面发挥重要作用; SnRK2则在响应干旱以及盐碱等非生物胁迫中发挥重要作用; SnRK3更多地参与胞内离子浓度的平衡, 通过调节离子浓度迅速启动植株响应非生物胁迫(
Analysis of relative gene expression data using reatime quantitative PCR and the 2-ΔΔCt method
1
2001
... 采用SPSS 25.0软件(SPSS, 芝加哥, IL)进行数据统计分析, 采用Microsoft Excel 2016软件作图.使用2-ΔΔCt方法(
Transcriptome survey reveals increased comp- lexity of the alternative splicing landscape in Arabidopsis
1
2012
... 为了能够在胁迫条件下生存, 植物进化出复杂的机制来感知外部信号, 从而对环境变化做出最佳反应, 其中表观遗传机制调控植物应答外界胁迫是近年来发现的重要机制之一, DNA甲基化和可变剪接均是其中的重要途径(
Structure and expression of phospho enolpyruvate carboxylase kinase genes in solanaceae. A novel gene exhibits alternative splicing
1
2003
... 可变剪接(alternative splicing, AS)是一种生物体基因转录后调控的方式, 植物中的许多基因存在AS, 其在植物生长发育中具有重要作用(
Plant hormones and seed germination
1
2014
... 有效的种子发芽使幼苗健壮地生长, 并具有抵御生物或非生物胁迫的能力, 从而最终发展成为高产作物(
Identification and molecular characterization of MYB transcription factor superfamily in C4 model plant foxtail millet ( Setaria italica L.)
1
2014
... C4植物的磷酸烯醇式丙酮酸羧化酶可将二氧化碳输送至核酮糖-1,5-二磷酸羧化酶(Ribulose-1,5-bisphosphate carboxylase, Rubisco), 在高光强和高温下也能更快地进行光合作用, 因此C4植物光合作用效率高, 并具有更好的水分利用率(
High temperature-induced expression of rice α-amylases in developing endosperm produces chalky grains
1
2017
... 有效的种子发芽使幼苗健壮地生长, 并具有抵御生物或非生物胁迫的能力, 从而最终发展成为高产作物(
Proteomic identification of α-amylase isoforms encoded by RAmy3B/3C from germinating rice seeds
1
2004
... 有效的种子发芽使幼苗健壮地生长, 并具有抵御生物或非生物胁迫的能力, 从而最终发展成为高产作物(
The remarkable diversity of plant PEPC (phosphoenolpyruvate carboxylase): recent insights into the physiological functions and post-translational controls of non-photosynthetic PEPCs
1
2011
... 近年来, 在全球气候变化背景下, 干旱频繁发生且日趋严峻, 严重影响作物的生长, 甚至造成减产(
Independent and combined effects of high temperature and drought stress during grain filling on plant yield and chloroplast EF-Tu expression in spring wheat
1
2011
... 近年来, 在全球气候变化背景下, 干旱频繁发生且日趋严峻, 严重影响作物的生长, 甚至造成减产(
Improved oxidative tolerance in suspension-cultured cells of C4-pepc trans- genic rice by H2O2 and Ca2+ under PEG-6000
2
2015
... 近年来, 在全球气候变化背景下, 干旱频繁发生且日趋严峻, 严重影响作物的生长, 甚至造成减产(
... )和钙离子(
Plant serine/arginine-rich proteins: roles in precursor messenger RNA splicing, plant development, and stress responses
2
2011
... 丝氨酸/精氨酸丰富蛋白家族(arginine/serine-rich proteins, SR proteins)是真核生物中的一类剪接因子, 在前体mRNA的组成性和选择性剪接中起作用(
... 丝氨酸/苏氨酸蛋白激酶是一个相对较大的蛋白激酶家族, 其中AMPK (AMP-activated kinase)/ SnRKs是高度保守的丝氨酸/苏氨酸蛋白激酶家族的重要成员.SnRKs可以细分成3个亚家族(SnRK1、SnRK2和SnRK3).SnRK1主要在调节植株生长发育、维持能量平衡以及响应生物逆境方面发挥重要作用; SnRK2则在响应干旱以及盐碱等非生物胁迫中发挥重要作用; SnRK3更多地参与胞内离子浓度的平衡, 通过调节离子浓度迅速启动植株响应非生物胁迫(
Hydrogen peroxide regulated photosynthesis in C4- pepc transgenic rice
1
2014
... 近年来, 在全球气候变化背景下, 干旱频繁发生且日趋严峻, 严重影响作物的生长, 甚至造成减产(
A modified anthrone-sulfuric acid method for the determination of fructose in the presence of certain proteins
1
1987
... 参考蒽酮比色法(
Alternative splicing at the intersection of biological timing, development, and stress responses
1
2013
... 丝氨酸/精氨酸丰富蛋白家族(arginine/serine-rich proteins, SR proteins)是真核生物中的一类剪接因子, 在前体mRNA的组成性和选择性剪接中起作用(
Alternative splicing in plants-coming of age
1
2012
... 可变剪接(alternative splicing, AS)是一种生物体基因转录后调控的方式, 植物中的许多基因存在AS, 其在植物生长发育中具有重要作用(
Transgenic rice over-expressing maize C4 phosphoenolpyruvate carboxylase gene contributes to alleviating low nitrogen stress
1
2018
... 近年来, 在全球气候变化背景下, 干旱频繁发生且日趋严峻, 严重影响作物的生长, 甚至造成减产(
Genomewide comparative analysis of alternative splicing in plants
1
2006
... 为了能够在胁迫条件下生存, 植物进化出复杂的机制来感知外部信号, 从而对环境变化做出最佳反应, 其中表观遗传机制调控植物应答外界胁迫是近年来发现的重要机制之一, DNA甲基化和可变剪接均是其中的重要途径(
Genome-wide mapping of cytosine methylation revealed dynamic DNA methylation patterns associated with genes and centromeres in rice
1
2010
... 为了能够在胁迫条件下生存, 植物进化出复杂的机制来感知外部信号, 从而对环境变化做出最佳反应, 其中表观遗传机制调控植物应答外界胁迫是近年来发现的重要机制之一, DNA甲基化和可变剪接均是其中的重要途径(
Physiological investigation of C4-phosphoenolpyruvate- carboxylase-introduced rice line shows that sucrose metabolism is involved in the improved drought tolerance
2
2017
... 近年来, 在全球气候变化背景下, 干旱频繁发生且日趋严峻, 严重影响作物的生长, 甚至造成减产(
... ), 而且在干旱条件下PC内源糖代谢加强(
Alterations of alternative splicing patterns of Ser/Arg-rich (SR) genes in response to hormones and stresses treatments in different ecotypes of rice (Oryza sativa)
1
2013
... 为了能够在胁迫条件下生存, 植物进化出复杂的机制来感知外部信号, 从而对环境变化做出最佳反应, 其中表观遗传机制调控植物应答外界胁迫是近年来发现的重要机制之一, DNA甲基化和可变剪接均是其中的重要途径(
Alternative splicing and mRNA levels of the disease resistance gene RPS4 are induced during defense responses
1
2007
... 为了能够在胁迫条件下生存, 植物进化出复杂的机制来感知外部信号, 从而对环境变化做出最佳反应, 其中表观遗传机制调控植物应答外界胁迫是近年来发现的重要机制之一, DNA甲基化和可变剪接均是其中的重要途径(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}