Research Progress in Seed Development, Dormancy and Germination Regulated by Cytokinin
Songquan Song,1,2,*, Jun Liu,2,*, Hua Yang3, Wenhu Zhang2, Qi Zhang2, Jiadong Gao21Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 2Guangdong Provincial Key Lab for Crop Germplasm Resources Preservation and Utilization/Agro-Biological Gene Research Center, Guangdong Academy of Agricultural Sciences, Guangzhou 510640, China 3College of Bioscience and Biotechnology, Hunan Agricultural University, Changsha 410128, China
Abstract Seed germination is an important stage in the establishment, growth and propagation of plants, and plays a critical role in the life cycle of seed plants. Seed dormancy is formed during development, and reaches its peak at physiological maturity. The phytohormone regulation of seed dormancy and germination may be a highly conserved mechanism in seed plants. Cytokinin (CK) is one of the most important signal molecules in plants, and regulates many aspects of plant growth and development. The bioactive CK levels are controlled by a balance among biosynthesis, activation, deactivation, re-activation and degradation, and seed development, dormancy and germination are regulated by bioactive CK levels and signaling pathways. Here, we mainly summarize the research progresses of CK biosynthesis and catabolism, signaling, and regulation on seed development, dormancy and germination. In addition, we also propose some scientific questions that need further addressed in this field to provide some information for understanding the molecular mechanism of seed development, dormancy and germination regulated by CK. Keywords:cytokinin;metabolism;molecular mechanism;seed dormancy and germination;signaling
PDF (1221KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 宋松泉, 刘军, 杨华, 张文虎, 张琪, 高家东. 细胞分裂素调控种子发育、休眠与萌发的研究进展. 植物学报, 2021, 56(2): 218-231 doi:10.11983/CBB20141 Song Songquan, Liu Jun, Yang Hua, Zhang Wenhu, Zhang Qi, Gao Jiadong. Research Progress in Seed Development, Dormancy and Germination Regulated by Cytokinin. Bulletin of Botany, 2021, 56(2): 218-231 doi:10.11983/CBB20141
研究表明, 种子的休眠释放与萌发是由遗传和环境因素共同决定的, 其中植物激素调控可能是种子植物中一种高度保守的机制(Nonogaki, 2014, 2017)。脱落酸(abscisic acid, ABA)诱导和维持种子休眠, 抑制种子萌发; 赤霉素(gibberellin, GA)则促进种子萌发和拮抗ABA对种子萌发的抑制作用(Nonogaki, 2019; Zubo and Schaller, 2020)。除了ABA和GA外, 细胞分裂素(cytokinin, CK)、乙烯(ethylene)和生长素(auxin)也在种子的休眠释放和萌发中起重要作用(Shu et al.2016)。
细胞分裂素是一种重要的植物激素, 在调控植物生长发育的许多过程中起关键作用, 包括细胞的分裂与分化、茎和根的生长、顶端优势维持、衰老、果实和种子发育、营养信号传递以及对生物和非生物胁迫的响应(Hwang et al.2012; Jameson and Song, 2016; Wybouw and De Rybel, 2019)。近年来, CK的生物合成与代谢, 尤其是CK信号转导途径的研究取得了重要进展(Lomin et al.2018; Arkhipov et al.2019; Hallmark and Rashotte, 2019; Chen et al.2020)。本文在综述CK的生物合成与代谢及其信号转导的基础上, 重点介绍CK调控种子发育、休眠与萌发的研究进展, 并提出了在本领域需要进一步研究的科学问题, 旨在为阐明CK调控种子发育、休眠与萌发的分子机理提供参考。
1 细胞分裂素的生物合成、降解与失活
植物中天然存在的CK是腺嘌呤的衍生物, 在N6端含有1个由异戊二烯(isoprene)衍生的侧链或者1个芳香族侧链, 分别称为类异戊二烯CK (isoprenoid CK)和芳香族CK (aromatic CK) (Sakakibara, 2006)。类异戊二烯CK包括N6-异戊烯腺嘌呤(N6-isopentenyladenine, iP)、反式玉米素(trans-zeatin, tZ)、顺式玉米素(cis-zeatin, cZ)和二氢玉米素(dihydrozeatin, DZ)。其中, 主要衍生物通常是tZ和iP及其糖的结合物, 但在不同的植物物种、组织和发育阶段存在较大的差异(Sakakibara, 2006)。例如, tZ和iP型CK是拟南芥(Arabidopsis thaliana)中的主要形式, 而cZ型CK则存在于玉米、水稻和鹰嘴豆(Cicer arietinum)中。而芳香族CK, 如ortho-topolin、meta-topolin及其甲氧基衍生物(分别为ortho-methoxytopolin和meta-methoxytopolin)和6-苄基腺嘌呤(6-benzyladenine, 6-BA)仅存在于某些植物中。尽管一些人工合成的CK衍生物具有生物活性, 但还未在植物中发现。CK的活性形式是游离态(free base) CK (Hothorn et al.2011; Lomin et al.2015), 但CK也可能以非活性形式存在, 如核苷(其中1个核糖与嘌呤环的N9连接)和核苷酸(其中核糖部分包含1个磷酸基) (Kieber and Schaller, 2018)。
细胞分裂素曾被认为只能在根尖和茎尖合成, 但实际上它们能在植物体内的不同部位合成和起作用(Sakakibara, 2006)。CK通过木质部(主要以tZ-核苷的形式)从根运输到茎, 也能够通过韧皮部(主要以iP型CK的形式)从茎运输到根(Kieber and Schaller, 2018)。植物细胞中生物活性CK的水平由其生物合成、活化、失活、降解和运输调节, 从而决定细胞中CK的信号强度(Sakakibara, 2006; Jameson and Song, 2016)。植物细胞中CK及其代谢物的多样性表明, 它们之间的相互转化可能是CK作用的主要调控机制(Ho?eket al.2020)。
细胞分裂素的葡糖基结合物在生物测定中无生物活性, 这些被结合的CK不能与CK受体结合(Spichal et al.2004)。尽管O-葡糖苷和N-葡糖苷稳定性差异的生理意义至今还不清楚, 但易裂解的O-葡糖苷被认为是CK失活的稳定贮藏形式。虽然与CK O-葡糖基化有关的基因已被鉴定(Mok et al.2005), 但尚缺乏关于N-葡糖基化的分子生物学信息。在拟南芥中, UGT76C1和UGT76C2在N7或N9位置葡糖基化一系列CK碱基, 也在N7位置葡糖基化tZ, 生成tZ-O-葡糖苷(tZ-O-glucoside) (Hou et al.2004)。
2 细胞分裂素信号转导
细胞分裂素信号转导途径也被称为双组分信号转导系统(two-component signaling system), 主要包括组氨酸激酶(histidine (His) kinase, HK)、组氨酸磷酸转移蛋白(His phosphotransfer protein, HP)和反应调控因子(response regulator, RR)。其中HK作为CK的受体, HP将信号从HK传递到细胞核的RR, 从而调控靶基因转录(图1, 图2) (Hwang et al.2012; Kieber and Schaller, 2018; Lomin et al.2018; Arkhipov et al.2019; Hallmark and Rashotte, 2019; Zubo and Schaller, 2020)。双分子荧光互补(bimolecular fluorescence complementation, BiFC)实验证明, 拟南芥CK受体AHK (Arabidopsis HK) 2-4均能与AHP (Arabidopsis HP)1-3相互作用, 且无明显的偏好; CK受体能形成同源和异源二聚体(Arkhipov et al.2019)。AHK-AHK和AHK-AHP的相互作用主要定位于内质网(endoplasmic reticulum, ER); 相反, 各种磷酸转移蛋白(AHP)的同源和异源二聚体主要存在于植物的细胞核中(Arkhipov et al.2019)。
细胞分裂素信号被膜定位的组氨酸(His)激酶受体感受, 通过His磷酸转移蛋白被传递到细胞核, 从而激活细胞核中的转录因子家族。CK: 细胞分裂素; ATP: 腺苷三磷酸; ADP: 腺苷二磷酸; P: 磷酸; D: 保守的天冬氨酸; H: 保守的His; DI: 感受器模块的二聚体界面结构域; PAS和PAS-like: 感受器模块的CHASE结构域的子结构域; TM1和TM2: 跨膜结构域1和2; HisKA (DHpD): His激酶A结构域(二聚合作用和His磷酸转移结构域); H-ATPase (CAD): 腺苷三磷酸酶结构域(催化与ATP结合结构域); REC-like: 类接收器结构域; REC: 接收器结构域; HPt: 含有His的磷酸转移蛋白(磷酸传递蛋白); RR-B: B型反应调控因子(转录因子)。蛋白-蛋白相互作用(PPI)用红色虚线表示。 Figure 2Intermolecular interactions in the cytokinin signaling pathway (modified from Lomin et al.2018; Arkhipov et al.2019)
Cytokinin signaling is perceived by membrane-localized Histidine (His) kinase receptors and is transduced to nucleus through a His phosphotransfer protein to activate a family of transcription factors in the nucleus. CK: Cytokinin; ATP: Adenosine triphosphate; ADP: Adenosine diphosphate; P: Phosphate; D: Conserved Aspartate; H: Conserved His; DI: Dimerization interface domain of the sensor module; PAS and PAS-like: Subdomains of the CHASE domain of the sensor module; TM1 and TM2: Transmembrane domain 1 and 2; HisKA (DHpD): His kinase A domain (dimerization and His phosphotransfer domain); H-ATPase (CAD): Adenosine triphosphatase domain (catalytic and ATP-binding domain); REC-like: Receiver-like domain; REC: Receiver domain; HPt: His-containing phosphotransfer protein (phosphotransmitter); RR-B: Type B response regulator (transcription factor). Protein-protein interactions (PPI) are indicated by a red dotted line.
2.1 组氨酸激酶
组氨酸激酶包含1个保守的与CK结合的胞外CHASE (cyclases/His kinases-associated sensing extracellular)区域、至少2个跨膜(transmembrane, TM)结构域和1个细胞质区域(图2) (Hwang et al.2012; Lomin et al.2018; Arkhipov et al.2019)。HK主要定位于ER膜(Wulfetange et al.2011)。CHASE区域即感受器模块(sensor module, SM), 面向ER腔或者质外体(apoplast), 包括二聚界面(dimerization interface, DI)、PAS (Per-Arnt-Sim)结构域和类PAS结构域(Hothorn et al.2011; Steklov et al.2013; Romanov et al.2018)。TM区域由2-5个α-螺旋延伸组成。在马铃薯(S. tuberosum) CK受体的N端, StHK2、StHK3和StHK4的上游TM螺旋的数量分别为3、2和1个(Steklov et al.2013)。HK的细胞质部分包括催化模块(catalytic module)和接收器模块(receiver module)。催化模块由HK-二聚体(HisKA或者DHpD)和H-ATPase (CAD)结构域组成(Pekárováet al.2016); 接收器模块含有类接收器结构域(receiver-like domain, REC-like)和接收器结构域(receiver domain, REC), 它们定位于HK蛋白的C-端(图2) (Lomin et al.2018; Pekarova et al.2018; Arkhipov et al.2019)。值得注意的是, AHK4同时具有组氨酸激酶和磷酸酶活性; 在配体缺乏时, AHK4的磷酸酶活性显著高于激酶活性, 能水解其接收器结构域上的磷酸基, 从而消耗可传递的磷酸基团(M?h?nenet al.2006)。
水稻、拟南芥和马铃薯CK受体序列的比对分析表明, 马铃薯同源基因中存在已知CK受体中所有的典型基序(Lomin et al.2018)。在马铃薯催化模块中已鉴定了H、N、G1、F和G2基序, 在接收器模块中鉴定了DD-D-K基序。保守序列含有可磷酸化的His和天冬氨酸(Asp)残基。StHK2在其类接收器结构域中含有1个保守的Asp, 类似于拟南芥AHK2、番茄SlHK2及水稻OsHK3和OsHK5的同源基因。然而, 类接收器结构域中的全部类DD-D-K基序与接收器结构域中的相应序列几乎无相同点(Lomin et al.2018)。
2.2 组氨酸磷酸转移蛋白
在CK信号转导途径中, HP (也称为His磷酸传递蛋白(His phosphotransmitter, HPt))在HK受体的下游起作用, 介导磷酸基在多步骤磷酸转移中从活化的HK受体接收器结构域转移到RR接收器结构域(图1, 图2) (Hwang et al.2012; Arkhipov et al.2019)。AHP不具有催化活性, 实际上是作为RR自身磷酸化活性的高能磷酸供体。AHP是CK信号转导冗余的正调控因子(Hutchison et al.2006), 但它们也在植物其它HK下游(如CKI1 (CYTOKININ-INDEPENDENT1))起作用(Liu et al.2017)。AHP蛋白具有1个保守的半胱氨酸残基, 该残基能在体外和体内被一氧化氮(NO) S-亚硝酰化, 抑制AHP作为磷酸转移蛋白的作用, 降低AHP从HK接收磷酸基并将其转移到RR的能力(Feng et al.2013)。
磷酸传递的信号转导、蛋白互作和高阶(higher- order)功能缺失突变的分析表明, AHP在功能上冗余且可以不加选择地与受体相互作用, 磷酸基的转移是双向的(Deng et al.2010)。在6个AHP (AHP1-6)中, AHP6缺乏保守的His残基, 因此不能接受磷酸基, 被称为伪AHP (pseudo-AHP)。AHP6负调控信号转导途径活性, 很可能与AHP1-5竞争与活化受体的相互作用(Lomin et al.2018)。
2.3 反应调控因子
反应调控因子是一个转录因子家族, 包含1个保守的接受磷酸基的Asp残基(Keshishian and Rashotte, 2015)。根据其系统发生和结构域, RR家族成员可分为3种类型: A型、B型和C型(Schaller et al.2007), 但只有A型和B型RR与CK信号转导有关(Kieber and Schaller, 2018; Tuan et al.2019; Zubo and Schaller, 2020)。B型RR包括ARR (Arabidopsis RR) 1、ARR2、ARR10-14和ARR18-21, 能被HP接收器结构域中的Asp残基磷酸化激活, 作为CK反应的正调控因子起作用(Hwang et al.2012)。B型RR的特征是包含1个接收器结构域和1个含有类MYB DNA结合结构域的长C-端延伸。接收器结构域包含1个保守的Asp残基, 当多步骤磷酸传递被激活时, Asp可作为调控磷酸化的位点。类MYB DNA结合结构域与1个短的(A/G) GAT核心DNA序列结合, 该序列对B型RR的结合作用非常关键(Zubo et al.2017; Zubo and Schaller, 2020)。ChIP-seq (chromatin immunoprecipitation combined with sequencing)实验显示, B型RR在植物体内以CK依赖的方式与其靶点结合, 这种结合以类似于细菌RR的方式被接收器结构域磷酸化激活(Zubo et al.2017)。
A型ARR (如ARR3-9、ARR15-17)由1个接收器结构域和1个短的C-端延伸组成, 作为CK信号转导的负反馈调控因子起作用; 但与B型RR不同, 它们缺乏典型的转录调控输出结构域(output domain) (Kieber and Schaller, 2018)。在对CK的反应中, A型RR通过B型RR的直接激活被诱导转录(Taniguchi et al.2007), CK也以磷酸化依赖的方式稳定许多A型RR (To et al.2007)。A型ARR负调控CK信号转导的机制是与B型ARR竞争AHP的磷酸化, 从而导致多步骤磷酸传递与B型ARR转录调控因子解离, 以减少其对基因表达的影响(Kieber and Schaller, 2014; Zubo and Schaller, 2020)。
2.4 细胞分裂素信号转导通路
细胞分裂素与HK的PAS结合, 导致受体的二聚作用并被激活, CK的信号转导通过类似于原核生物的His-Asp磷酸传递途径进行(Keshishian and Rashotte, 2015; Zschiedrich et al.2016)。在激活的受体ATP结合结构域中, ATP发生水解, 释放的磷酸基团与HisKA结构域中1个保守的His残基结合, 形成磷酸组氨酸(phosphohistidine)。然后, 活化(hot)的磷酸基在分子内被转移到接收器结构域中1个保守的Asp残基上, 依次与HPt相互作用, 将活化的磷酸基转移到HPt保守的His残基上。HPt携带该活化的磷酸基并转移到细胞核和磷酸化RR蛋白保守的Asp残基。最后, 磷酸化的B型RR能够与DNA结合, 激活CK靶基因的转录(图1, 图2) (Kieber and Schaller, 2018; Pekarova et al.2018; Arkhipov et al.2019)。
3 细胞分裂素对种子发育、休眠与萌发的调控
种子成熟过程中在母体植株上获得的休眠类型称为初生休眠(primary dormancy), 种子初生休眠的诱导和维持受植物激素调控(Shu et al.2016; Nonogaki, 2017, 2019; 宋松泉等, 2020a)。研究表明, 种子中CK的水平和信号转导调控种子的发育、休眠与萌发(图1, 图3) (Kieber and Schaller, 2018; Tuan et al.2019; Nguyen et al.2020)。后熟(after-ripening)是一种释放种子休眠的方式, 也通过影响CK的水平和信号转导来调控种子的休眠与萌发(Chitnis et al.2014)。
图3
新窗口打开|下载原图ZIP|生成PPT 图3种子发育和萌发过程中与细胞分裂素(CK)代谢和信号转导组分有关的蛋白(酶)的变化模式图(改自Jameson et al.2016; Tuan et al.2019; Nguyen et al.2020)
IPT: 异戊烯基转移酶; LOG: CK核苷5′-单磷酸磷酸核糖水解酶; CKX: CK氧化酶/脱氢酶; HK: 组氨酸激酶; HP: 组氨酸磷酸转移蛋白; Type-B RRs: 类型-B反应调控因子; Type-A RRs: 类型-A反应调控因子 Figure 3The model of the change of proteins (enzymes) involved in cytokinin (CK) metabolism and signaling components during seed development and germination (modified from Jameson et al.2016; Tuan et al.2019; Nguyen et al.2020)
IPT: Isopentenyltransferase; LOG: CK nucleoside 5′-monophosphate phosphoribohydrolase; CKX: CK oxidase/ dehydrogenase; HK: Histidin (His) kinase; HP: His phosphotransfer protein; Type-B RRs: Type-B response regulators; Type-A RRs: Type-A response regulators
在小麦种子成熟过程中, TaAHK4、TaARR9和TaARR12的表达水平在AC Domain和RL4452的胚中均表现下降, 这与iP和tZ的积累下降有关。然而, 在开花后20天, AC Domain胚中TaARR12的表达水平比RL4452更低(Tuan et al.2019)。鉴于TaARR12属于B型ARR, 且作为CK反应的正调控因子起作用(Kieber and Schaller, 2018), TaARR12很可能抑制AC Domain胚中的CK反应, 从而在小麦种子成熟过程中起诱导休眠的作用。2种基因型胚乳中TaAHP2探针的表达随着种子成熟而下降。然而, 在种子成熟中期(开花后30天), TaAHK4探针在AC Domain胚乳中的表达水平比RL4452更低(Tuan et al.2019)。由于AHK是CK的受体(Keshishian and Rashotte, 2015), 因此上述结果表明, RL4452对CK的反应敏感性增加, 从而抑制种子的休眠。另一方面, 在种子成熟的早期到中期(开花后20-30天), AC Domain胚乳中TaARR9的表达水平比RL4452更高(Tuan et al.2019)。由于TaARR9属于负调控CK信号转导的A型ARR (Kieber and Schaller, 2018), 在成熟时具有深休眠的AC Domain种子胚乳对CK的反应敏感性降低, 其结果是成熟时休眠程度高。此外, AC Domain胚乳中AHK探针的表达下降也可能表明CK反应受到抑制, 从而诱导休眠。
在豌豆(Pisum sativum)种子萌发过程中, 3个PsIPT基因在吸胀后4小时的子叶中表达, 其表达丰度在种子萌发过程中升高, 在萌发后降低; PsIPT在新生的根和芽中的表达丰度最高(Jameson et al.2016)。3个LOG基因在吸胀后4小时的子叶中表达量显著低于萌发后期, 但在根和芽中均有表达(Jameson et al.2016)。PsCKX2的表达水平在吸胀后4小时的子叶中比其它4个PsCKX家族成员(PsCKX1、PsCKX3、PsCKX5和PsCKX7)更高, 但在萌发过程中变化很小。PsCKX2在发育的根和芽中维持组成型表达, 而PsCKX1、PsCKX3、PsCKX5和PsCKX7在发育的芽和根中的表达显著增高(Jameson et al.2016)。在吸胀后4小时的豌豆种子中能检测到游离态CK和CK核苷, 包括tZ和iP, 但DZ和cZ的水平很低; 未能检测到CK O-或者N-葡糖苷及其核苷酸。吸胀后2天, CK的水平显著增高, 主要是由于CK核苷酸特别是iPRMP (iP ribosyl monophosphate)的增加, cZRMP (cis-zeatin riboside monophosphate)和tZRMP增加较少。子叶和根中的CK水平在吸胀后11天达到峰值, 然后下降; 但芽中的CK水平在吸胀后5天最高, 然后下降, 主要是CK核苷酸水平的下降(Jameson et al.2016)。这些结果表明, CK的生物合成和分解代谢与种子的萌发密切相关。
后熟(干藏)通过影响CK的水平和信号转导调控种子的休眠与萌发。小麦(AC domain)种子在吸胀24小时后萌发率为0, 吸胀7天后萌发率为11%, 表明种子具有休眠特性; 经后熟10个月的种子在吸胀24小时内的萌发率为95%, 表明后熟促进小麦种子的萌发(Chitnis et al.2014)。利用基因芯片技术, Chitnis等(2014)发现, 与未后熟的种子相比, 后熟种子在吸胀过程中有1个LOG、3个GLU、1个CKX和2个ZOG基因显著上调; 在40个注释为CK信号转导基因的探针中, 只有1个编码伪反应调控因子(pseudo-response regulator)的APRR9基因在后熟种子的吸胀过程中下调2倍以上(Chitnis et al.2014)。
3.3 细胞分裂素与其它植物激素的互作及在种子休眠与萌发中的作用
3.3.1 细胞分裂素与脱落酸在种子萌发中的拮抗作用
ABA抑制种子萌发, CK能逆转ABA对莴苣(Lactuca sativa)种子萌发的抑制作用(Kham, 1968)。1 mmol·L-1 ABA对黑黄檀(Dalbergia fusca)种子萌发的抑制作用能够被0.001-0.1 mmol·L-1 6-BA所拮抗(邓志军和宋松泉, 2008)。过表达AtIPT8能降低gim1 (germination insensitive to ABA mutant 1)突变体种子萌发对ABA的敏感性; AtIPT8的功能丧失导致ipt8-1-ko和ipt8-2-ko突变体产生ABA过敏性反应; 在gim1中, AtIPT8的过表达能够通过促进CK的生物合成来增强CK信号, 从而上调CK信号转导的转录调控因子(如A型ARR4-6) (Wang et al.2011)。在野生型拟南芥Columbia-0背景下, ABA处理能够通过抑制AtIPT3和AtIPT8的表达水平减少CK的内源生物合成, 随后降低A型ARR4-6的表达水平(Wang et al.2011)。此外, 在萌发10天的gim1幼苗中, ABA处理也会抑制AtIPT3和AtIPT8的表达, 降低A型ARR4-6的表达丰度(Wang et al.2011)。值得注意的是, gim1突变体中CK水平的增高不是由于ABA生物合成的改变, 而是降低了ABA和胁迫反应基因(如ABI5(ABSCISIC ACID INSENSITIVE 5))的表达水平(Wang et al.2011)。
ABI5是一种碱性亮氨酸拉链转录因子(basic leucine zipper transcription factor), 在ABA介导的抑制种子萌发和萌发后生长的调控中起重要作用(Guan et al.2014; 宋松泉等, 2020b)。在种子萌发过程中, ABI5功能丧失能够降低ABA的敏感性, 而ABI5功能获得能增加ABA的敏感性(Liu and Stone, 2010)。研究表明, ABI5与ARR4-6发生物理相互作用(Wang et al.2011)。在arr3/arr4/arr5/arr6四突变体中, CK诱导的ABI5降低减少, 这表明A型ARR蛋白与ABI5的结合可能在调控ABI5蛋白的稳定性中起重要作用(Guan et al.2014)。由于A型ARR mRNA的稳定水平和A型ARR蛋白的积累被CK正调控(Ren et al.2009), 因此, A型ARR-ABI5复合物可能抑制ABI5蛋白与蛋白酶体降解复合物的相互作用。
Guan等(2014)研究了CK对ABA信号转导组分表达水平的影响, 包括SnRK2 (sucrose non-fermenting1-related kinase 2)基因(SnRK2.2和SnRK2.3)、ABF/AREB(ABA-responsive element binding factor/ABA-responsive element binding protein)家族基因(ABF1-4)以及ABI (ABI1-5和ABI8)基因, 发现CK对这些基因的表达无显著影响; 大多数ABA调控基因的表达水平也不受CK影响, ABA诱导的ABI5的表达仅被CK轻微降低。用CK处理72-96小时后, ABI5蛋白几乎被完全降解, 但ABI5 mRNA被适度降低(Guan et al.2014), 这表明ABI5蛋白而不是ABI5mRNA的稳定性是CK介导ABA信号转导的主要调控步骤。CK诱导的ABI5蛋白的降解被蛋白酶体降解复合物专一性抑制剂MG132逆转。这些结果表明, CK调控的ABI5积累减少可能是由26S蛋白酶体途径介导(Liu and Stone, 2010; Guan et al.2014)。
AraújoS, PaganoA, DondiD, LazzaroniS, PinelaE, MacoveiA, BalestrazziA (2019). Metabolic signatures of germination triggered by kinetin in Medicago truncatula Sci Rep9, 10466. DOI:10.1038/s41598-019-46866-6URL [本文引用: 1]
ArkhipovDV, LominSN, MyakushinaYA, SavelievaEM, OsolodkinDI, RomanovGA (2019). Modeling of protein-protein interactions in cytokinin signal transduction Int J Mol Sci20,2096. DOI:10.3390/ijms20092096URL [本文引用: 10]
BaskinCC, BaskinJM (2014). Seeds:Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd edn. Amsterdam: Academic Press. pp.5-77. [本文引用: 1]
BelmonteMF, KirkbrideRC, StoneSL, PelletierJM, BuiAQ, YeungEC, HashimotoM, FeiJ, HaradaCM, MunozMD, LeBH, DrewsGN, BradySM, GoldbergRB, HaradaJJ (2013). Comprehensive developmental profiles of gene activity in regions and subregions of the Arabidopsis seed Proc Natl Acad Sci USA110,E435- E444. DOI:10.1073/pnas.1222061110URL [本文引用: 1]
BewleyJD, BradfordKJ, HilhorstHWM, NonogakiH (2013). Physiology of Development,Germination and Dormancy, 3rd edn. New York: Springer. pp.27-83. [本文引用: 1]
CairnsJRK, EsenA (2010). β-glucosidases Cell Mol Life Sci67,3389-3405. DOI:10.1007/s00018-010-0399-2URL [本文引用: 1] β-Glucosidases (3.2.1.21) are found in all domains of living organisms, where they play essential roles in the removal of nonreducing terminal glucosyl residues from saccharides and glycosides. β-Glucosidases function in glycolipid and exogenous glycoside metabolism in animals, defense, cell wall lignification, cell wall β-glucan turnover, phytohormone activation, and release of aromatic compounds in plants, and biomass conversion in microorganisms. These functions lead to many agricultural and industrial applications. β-Glucosidases have been classified into glycoside hydrolase (GH) families GH1, GH3, GH5, GH9, and GH30, based on their amino acid sequences, while other β-glucosidases remain to be classified. The GH1, GH5, and GH30 β-glucosidases fall in GH Clan A, which consists of proteins with (β/α)<sub>8</sub>-barrel structures. In contrast, the active site of GH3 enzymes comprises two domains, while GH9 enzymes have (α/α)<sub>6</sub> barrel structures. The mechanism by which GH1 enzymes recognize and hydrolyze substrates with different specificities remains an area of intense study.
ChenL, ZhaoJQ, SongJC, JamesonPE (2020). Cytokinin dehydrogenase: a genetic target for yield improvement in wheat Plant Biotechnol J18,614-630. DOI:10.1111/pbi.13305PMID:WOS:000503784700001 [本文引用: 2] The plant hormone group, the cytokinins, is implicated in both qualitative and quantitative components of yield. Cytokinins have opposing actions in shoot and root growth-actions shown to involve cytokinin dehydrogenase (CKX), the enzyme that inactivates cytokinin. We revise and provide unambiguous names for the CKX gene family members in wheat, based on the most recently released wheat genome database, IWGSC RefSeq v1.0 & v2.0. We review expression data of CKX gene family members in wheat, revealing tissue-specific gene family member expression as well as sub-genome-specific expression. Manipulation of CKX in cereals shows clear impacts on yield, root growth and orientation, and Zn nutrition, but this also emphasizes the necessity to unlink promotive effects on grain yield from negative effects of cytokinin on root growth and uptake of mineral nutrients, particularly Zn and Fe. Wheat is the most widely grown cereal crop globally, yet is under-research compared with rice and maize. We highlight gaps in our knowledge of the involvement of CKX for wheat. We also highlight the necessity for accurate analysis of endogenous cytokinins, acknowledging why this is challenging, and provide examples where inadequate analyses of endogenous cytokinins have led to unjustified conclusions. We acknowledge that the allohexaploid nature of bread wheat poses challenges in terms of uncovering useful mutations. However, we predict TILLING followed by whole-exome sequencing will uncover informative mutations and we indicate the potential for stacking mutations within the three genomes to modify yield components. We model a wheat ideotype based on CKX manipulation.
ChitnisVR, GaoF, YaoZ, JordanMC, ParkS, AyeleBT (2014). After-ripening induced transcriptional changes of hormonal genes in wheat seeds: the cases of brassinosteroids, ethylene, cytokinin and salicylic acid PLoS One9,e87543. DOI:10.1371/journal.pone.0087543URL [本文引用: 4]
CorbineauF, XiaQ, BaillyC, EI-Maarouf-BouteauH (2014). Ethylene, a key factor in the regulation of seed dormancy Front Plant Sci5,539. [本文引用: 1]
DayRC, HerridgeRP, AmbroseBA, MacknightRC (2008). Transcriptome analysis of proliferating Arabidopsis endosperm reveals biological implications for the control of syncytial division, cytokinin signaling, and gene expression regulation Plant Physiol148,1964-1984. DOI:10.1104/pp.108.128108URL [本文引用: 1]
DengY, DongHL, MuJY, RenB, ZhengBL, JiZD, YangWC, LiangY, ZuoJR (2010). Arabidopsis histidine kinase CKI1 acts upstream of histidine phosphotransfer proteins to regulate female gametophyte development and vegetative growth Plant Cell22,1232-1248. DOI:10.1105/tpc.108.065128URL [本文引用: 1]
EastwoodD, TavenerRJA, LaidmanDL (1969). Sequential action of cytokinin and gibberellic acid in wheat aleurone tissue Nature221,1267. [本文引用: 1]
FrébortI, KowalskaM, HluskaT, FrébortováJ, GaluszkaP (2011). Evolution of cytokinin biosynthesis and degradation J Exp Bot62,2431-2452. DOI:10.1093/jxb/err004URL [本文引用: 4] Cytokinin hormones are important regulators of development and environmental responses of plants that execute their action via the molecular machinery of signal perception and transduction. The limiting step of the whole process is the availability of the hormone in suitable concentrations in the right place and at the right time to interact with the specific receptor. Hence, the hormone concentrations in individual tissues, cells, and organelles must be properly maintained by biosynthetic and metabolic enzymes. Although there are merely two active cytokinins, isopentenyladenine and its hydroxylated derivative zeatin, a variety of conjugates they may form and the number of enzymes/isozymes with varying substrate specificity involved in their biosynthesis and conversion gives the plant a variety of tools for fine tuning of the hormone level. Recent genome-wide studies revealed the existence of the respective coding genes and gene families in plants and in some bacteria. This review summarizes present knowledge on the enzymes that synthesize cytokinins, form cytokinin conjugates, and carry out irreversible elimination of the hormones, including their phylogenetic analysis and possible variations in different organisms.
GuanCM, WangXC, FengJ, HongSL, LiangY, RenB, ZuoJR (2014). Cytokinin antagonizes abscisic acid-mediated inhibition of cotyledon greening by promoting the degradation of ABSCISIC ACID INSENSITIVE 5 protein in Arabidopsis Plant Physiol164, 1515-1526. DOI:10.1104/pp.113.234740URL [本文引用: 5] In higher plants, seed germination is followed by postgerminative growth. One of the key developmental events during postgerminative growth is cotyledon greening, which enables a seedling to establish photosynthetic capacity. The plant phytohormone abscisic acid (ABA) plays a vital role by inhibiting seed germination and postgerminative growth in response to dynamically changing internal and environmental cues. It has been shown that ABSCISIC ACID INSENSITIVE5 (ABI5), a basic leucine zipper transcription factor, is an important factor in the regulation of the ABA-mediated inhibitory effect on seed germination and postgerminative growth. Conversely, the phytohormone cytokinin has been proposed to promote seed germination by antagonizing the ABA-mediated inhibitory effect. However, the underpinning molecular mechanism of cytokinin-repressed ABA signaling is largely unknown. Here, we show that cytokinin specifically antagonizes ABA-mediated inhibition of cotyledon greening with minimal effects on seed germination in Arabidopsis (Arabidopsis thaliana). We found that the cytokinin-antagonized ABA effect is dependent on a functional cytokinin signaling pathway, mainly involved in the cytokinin receptor gene CYTOKININ RESPONSE1/ARABIDOPSIS HISTIDINE KINASE4, downstream histidine phosphotransfer protein genes AHP2, AHP3, and AHP5, and a type B response regulator gene, ARR12, which genetically acts upstream of ABI5 to regulate cotyledon greening. Cytokinin has no apparent effect on the transcription of ABI5. However, cytokinin efficiently promotes the proteasomal degradation of ABI5 in a cytokinin signaling-dependentmanner. These results define a genetic pathway through which cytokinin specifically induces the degradation of ABI5 protein, thereby antagonizing ABA-mediated inhibition of postgerminative growth.
HouBK, LimEK, HigginsGS, BowlesDJ (2004). N-glucosylation of cytokinins by glycosyltransferases of Arabidopsis thaliana J Biol Chem279,47822-47832. DOI:10.1074/jbc.M409569200URL [本文引用: 1]
HutchisonCE, LiJ, ArguesoC, GonzalezM, LeeE, LewisMW, MaxwellBB, PerdueTD, SchallerGE, AlonsoJM, EckerJR, KieberJJ (2006). The Arabidopsis histidine phosphotransfer proteins are redundant positive regulators of cytokinin signaling Plant Cell18,3073-3087. PMID:17122069 [本文引用: 1] Arabidopsis thaliana histidine phosphotransfer proteins (AHPs) are similar to bacterial and yeast histidine phosphotransfer proteins (HPts), which act in multistep phosphorelay signaling pathways. A phosphorelay pathway is the current model for cytokinin signaling. To assess the role of AHPs in cytokinin signaling, we isolated T-DNA insertions in the five AHP genes that are predicted to encode functional HPts and constructed multiple insertion mutants, including an ahp1,2,3,4,5 quintuple mutant. Single ahp mutants were indistinguishable from wild-type seedlings in cytokinin response assays. However, various higher-order mutants displayed reduced sensitivity to cytokinin in diverse cytokinin assays, indicating both a positive role for AHPs in cytokinin signaling and functional overlap among the AHPs. In contrast with the other four AHPs, AHP4 may play a negative role in some cytokinin responses. The quintuple ahp mutant showed various abnormalities in growth and development, including reduced fertility, increased seed size, reduced vascular development, and a shortened primary root. These data indicate that most of the AHPs are redundant, positive regulators of cytokinin signaling and affect multiple aspects of plant development.
HwangI, SheenJ, MüllerB (2012). Cytokinin signaling networks Annu Rev Plant Biol63,353-380. URL [本文引用: 6] Despite long-standing observations on diverse cytokinin actions, the discovery path to cytokinin signaling mechanisms was tortuous. Unyielding to conventional genetic screens, experimental innovations were paramount in unraveling the core cytokinin signaling circuitry, which employs a large repertoire of genes with overlapping and specific functions. The canonical two-component transcription circuitry involves His kinases that perceive cytokinin and initiate signaling, as well as His-to-Asp phosphorelay proteins that transfer phosphoryl groups to response regulators, transcriptional activators, or repressors. Recent advances have revealed the complex physiological functions of cytokinins, including interactions with auxin and other signal transduction pathways. This review begins by outlining the historical path to cytokinin discovery and then elucidates the diverse cytokinin functions and key signaling components. Highlights focus on the integration of cytokinin signaling components into regulatory networks in specific contexts, ranging from molecular, cellular, and developmental regulations in the embryo, root apical meristem, shoot apical meristem, stem and root vasculature, and nodule organogenesis to organismal responses underlying immunity, stress tolerance, and senescence.
JamesonPE, DhandapaniP, NovakO, SongJC (2016). Cytokinins and expression of SWEET, SUT, CWINV and AAP genes increase as pea seeds germinate Int J Mol Sci17,2013. DOI:10.3390/ijms17122013URL [本文引用: 6]
KakimotoT (2001). Identification of plant cytokinin biosynthetic enzymes as dimethylallyl diphosphate: ATP/ADP isopentenyltransferases Plant Cell Physiol42,677-685. PMID:11479373 [本文引用: 1] It has been believed that the key step in cytokinin biosynthesis is the addition of a 5-carbon chain to the N(6) of AMP. To identify cytokinin biosynthesis enzymes that catalyze the formation of the isopentenyl side chain of cytokinins, the Arabidopsis genomic sequence was searched for genes that could code for isopentenyltransferases. This resulted in the identification of nine putative genes for isopentenyltransferases. One of these, AtIPT4, was subjected to detailed analysis. Overexpression of AtIPT4 caused cytokinin-independent shoot formation on calli. As shoot formation on calli normally occurs only when cytokinins are applied, it suggested that this gene product catalyzed cytokinin biosynthesis in plants. Recombinant AtIPT4 catalyzed the transfer of an isopentenyl group from dimethylallyl diphosphate to the N(6) of ATP and ADP, but not to that of AMP. AtIPT4 did not exhibit the DMAPP:tRNA isopentenyltransferase activity. These results indicate that cytokinins are, at least in part, synthesized from ATP and ADP in plants.
KasaharaH, TakeiK, UedaN, HishiyamaS, YamayaT, KamiyaY, YamaguchiS, SakakibaraH (2004). Distinct isoprenoid origins of cis- and trans-zeatin biosyntheses in Arabidopsis J Biol Chem279,14049-14054. DOI:10.1074/jbc.M314195200URL [本文引用: 2]
KhanAA (1968). Inhibition of gibberellic acid-induced germination by abscisic acid and reversal by cytokinins Plant Physiol43,1463-1465. PMID:16656940 [本文引用: 1]
KieberJJ, SchallerGE (2018). Cytokinin signaling in plant development Development145,dev149344. DOI:10.1242/dev.149344URL [本文引用: 12]
KurakawaT, UedaN, MaekawaM, KobayashiK, KojimaM, NagatoY, SakakibaraH, KyozukaJ (2007). Direct control of shoot meristem activity by a cytokinin-activating enzyme Nature445,652-655. DOI:10.1038/nature05504URL [本文引用: 2]
LiYJ, ChengHY, SongSQ (2009). Effects of temperature, after-ripening, stratification, and scarification plus hormone treatments on dormancy release and germination of Acer truncatum seeds Seed Sci Technol37,554-562. DOI:10.15258/sstURL [本文引用: 1]
LiuXD, ZhangH, ZhaoY, FengZY, LiQ, YangHQ, LuanS, LiJM, HeZH (2013). Auxin controls seed dormancy through stimulation of abscisic acid signaling by inducing ARF-mediated ABI3 activation in Arabidopsis Proc Natl Acad Sci USA110,15485-15490. DOI:10.1073/pnas.1304651110URL [本文引用: 1]
LiuZN, YuanL, SongXY, YuXL, SundaresanV (2017). AHP2, AHP3, and AHP5 act downstream of CKI1 in Arabidopsis female gametophyte development J Exp Bot68,3365-3373. DOI:10.1093/jxb/erx181URL [本文引用: 1]
LominSN, KrivosheevDM, SteklovMY, ArkhipovDV, OsolodkinDI, SchmüllingT, RomanovGA (2015). Plant membrane assays with cytokinin receptors underpin the unique role of free cytokinin bases as biologically active ligands J Exp Bot66,1851-1863. DOI:10.1093/jxb/eru522URL [本文引用: 1]
LominSN, MyakushinaYA, KolachevskayaOO, GetmanIA, ArkhipovDV, SavelievaEM, OsolodkinDI, RomanovGA (2018). Cytokinin perception in potato: new features of canonical players J Exp Bot69,3839-3853. DOI:10.1093/jxb/ery199URL [本文引用: 9]
LurHS, SetterTL (1993). Role of auxin in maize endosperm development (timing of nuclear DNA endoreduplication, zein expression, and cytokinin) Plant Physiol103,273- 280. DOI:10.1104/pp.103.1.273URL [本文引用: 1]
M?h?nenAP, HiguchiM, TormakangasK, MiyawakiK, PischkeMS, SussmanMR, HelariuttaY, KakimotoT (2006). Cytokinins regulate a bidirectional phosphorelay network in Arabidopsis Curr Biol16, 1116-1122. PMID:16753566 [本文引用: 1] The cytokinin class of plant hormones plays key roles in regulating diverse developmental and physiological processes. Arabidopsis perceives cytokinins with three related and partially redundant receptor histidine kinases (HKs): CRE1 (the same protein as WOL and AHK4), AHK2, and AHK3 (CRE-family receptors). It is suggested that binding of cytokinins induces autophosphorylation of these HKs and subsequent transfer of the phosphoryl group to a histidine phosphotransfer protein (HPt) and then to a response regulator (RR), ultimately regulating downstream signaling events. Here we demonstrate that, in vitro and in a yeast system, CRE1 is not only a kinase that phosphorylates HPts in the presence of cytokinin but is also a phosphatase that dephosphorylates HPts in the absence of cytokinin. To explore the roles of these activities in planta, we replaced CRE1 with mutant versions of the gene or with AHK2. Replacing CRE1 with CRE1(T278I), which lacks cytokinin binding activity and is locked in the phosphatase form, decreased cytokinin sensitivity. Conversely, replacing CRE1 with AHK2, which favors kinase activity, increased cytokinin sensitivity. These results indicate that in the presence of cytokinins, cytokinin receptors feed phosphate to phosphorelay-integrating HPt proteins. In the absence of cytokinins, CRE1 removes phosphate from HPt proteins, decreasing the system phosphoload.
Marín-dela Rosa N, PfeifferA, HillK, LocascioA, BhaleraoRP, MiskolcziP, GronlundAL, Wanchoo-Kohli A, ThomasSG, BennettMJ, LohmannJU, BlázquezMA, AlabadíD (2015). Genome wide binding site analysis reveals transcriptional coactivation of cytokinin- responsive genes by DELLA proteins PLoS Genet11,e1005337. DOI:10.1371/journal.pgen.1005337URL [本文引用: 1]
MiyawakiK, Matsumoto-KitanoM, KakimotoT (2004). Expression of cytokinin biosynthetic isopentenyltransferase genes in Arabidopsis: tissue specificity and regulation by auxin, cytokinin, and nitrate Plant J37,128-138. [本文引用: 1]
MokMC, MartinRC, DobrevPI, VankováR, HoPS, Yonekura-SakakibaraK, SakakibaraH, MokDWS (2005). Topolins and hydroxylated thidiazuron derivatives are substrates of cytokinin O-glucosyltransferase with position specificity related to receptor recognition Plant Physiol137,1057-1066. [本文引用: 2]
MüllerB, SheenJ (2008). Cytokinin and auxin interaction in root stem-cell specification during early embryogenesis Nature453,1094-1097. [本文引用: 2]
NguyenHN, PerryL, KisialaA, OlechowskiH, EmeryRJN (2020). Cytokinin activity during early kernel development corresponds positively with yield potential and later stage ABA accumulation in field-grown wheat ( Triticum aestivum L.) Planta252,76. [本文引用: 5]
NonogakiH (2014). Seed dormancy and germination- emerging mechanisms and new hypotheses Front Plant Sci5,233. [本文引用: 2]
NonogakiH (2017). Seed biology updates—highlights and new discoveries in seed dormancy and germination research Front Plant Sci8,524. [本文引用: 3]
NonogakiH (2019). Seed germination and dormancy: the classic story, new puzzles, and evolution J Integr Plant Biol61,541-563. [本文引用: 2]
PekárováB, SzmitkowskaA, DopitováR, DegtjarikO, ?ídekL, HejátkoJ (2016). Structural aspects of multistep phosphorelay-mediated signaling in plants Mol Plant9,71-85. [本文引用: 1]
RenB, LiangY, DengY, ChenQG, ZhangJ, YangXH, ZuoJR (2009). Genome-wide comparative analysis of type-A Arabidopsis response regulator genes by overexpression studies reveals their diverse roles and regulatory mechanisms in cytokinin signaling Cell Res19, 1178- 1190. [本文引用: 1]
RijavecT, DermastiaM (2010). Cytokinins and their function in developing seeds Acta Chim Slov57,617-629. [本文引用: 1]
RomanovGA, LominSN, SchmüllingT (2018). Cytokinin signaling: from the ER or from the PM? That is the question! New Phytol218,41-53. [本文引用: 1]
SakakibaraH (2006). Cytokinins: activity, biosynthesis, and translocation Annu Rev Plant Biol57,431-449. [本文引用: 16]
SakakibaraH, KasaharaH, UedaN, KojimaM, TakeiK, HishiyamaS, AsamiT, OkadaK, KamiyaY, YamayaT, YamaguchiS (2005). Agrobacterium tumefaciens increases cytokinin production in plastids by modifying the biosynthetic pathway in the host plant Proc Natl Acad Sci USA102,9972-9977. [本文引用: 1]
SakanoY, OkadaY, MatsunagaA, SuwamaT, KanekoT, ItoK, NoguchiH, AbeI (2004). Molecular cloning, expression, and characterization of adenylate isopentenyltransferase from hop ( Humulus lupulus L.) Phytochemistry65,2439-2446. [本文引用: 2]
SchallerGE, DoiK, HwangI, KieberJJ, KhuranaJP, KurataN, MizunoT, PareekA, ShiuSH, WuP, YipWK (2007). Nomenclature for two-component signaling elements of rice Plant Physiol143,555-557. [本文引用: 1]
ShuK, LiuXD, XieQ, HeZH (2016). Two faces of one seed: hormonal regulation of dormancy and germination Mol Plant9,34-45. [本文引用: 2]
ShuaiHW, MengYJ, LuoXF, ChenF, ZhouWG, DaiYJ, QiY, DuJB, YangF, LiuJ, YangWY, ShuK (2017). Exogenous auxin represses soybean seed germination through decreasing the gibberellin/abscisic acid (GA/ABA) ratio Sci Rep7,12620. [本文引用: 1]
SpíchalL, RakovaNY, RieflerM, MizunoT, RomanovGA, StrnadM, SchmüllingT (2004). Two cytokinin receptors of Arabidopsis thaliana, CRE1/AHK4 and AHK3, differ in their ligand specificity in a bacterial assay Plant Cell Physiol45,1299-1305. [本文引用: 1]
SteklovMY, LominSN, OsolodkinDI, RomanovGA (2013). Structural basis for cytokinin receptor signaling: an evolutionary approach Plant Cell Rep32,781-793. [本文引用: 2]
StrnadM (1997). The aromatic cytokinins Physiol Plant101,674-688. [本文引用: 1]
TakeiK, SakakibaraH, SugiyamaT (2001). Identification of genes encoding adenylate isopentenyltransferase, a cytokinin biosynthesis enzyme, in Arabidopsis thaliana J Biol Chem276,26405-26410. [本文引用: 3]
TaniguchiM, SasakiN, TsugeT, AoyamaT, OkaA (2007). ARR1 directly activates cytokinin response genes that encode proteins with diverse regulatory functions Plant Cell Physiol48,263-277. [本文引用: 1]
TarkowskaD, Dole?alK, TarkowskiP, ?stotC, HolubJ, FuksováK, SchmüllingT, SandbergG, StrnadM (2003). Identification of new aromatic cytokinins in Arabidopsis thaliana and Populus × canadensis leaves by LC- (+) ESI-MS and capillary liquid chromatography/frit- fast atom bombardment mass spectrometry Physiol Plant117,579-590. [本文引用: 1]
ToJPC, DeruèreJ, MaxwellBB, MorrisVF, HutchisonCE, FerreiraFJ, SchallerGE, KieberJJ (2007). Cytokinin regulates type-A Arabidopsis response regulator activity and protein stability via two-component phosphorelay Plant Cell19,3901-3914. [本文引用: 1]
TuanPA, YamasakiY, KannoY, SeoM, AyeleBT (2019). Transcriptomics of cytokinin and auxin metabolism and signaling genes during seed maturation in dormant and non-dormant wheat genotypes Sci Rep9,3983. [本文引用: 12]
WangYP, LiL, YeTT, ZhaoSJ, LiuZ, FengYQ, WuY (2011). Cytokinin antagonizes ABA suppression to seed germination of Arabidopsis by downregulating ABI5 expression Plant J68,249-261. [本文引用: 5]
WulfetangeK, LominSN, RomanovGA, StolzA, HeylA, SchmüllingT (2011). The cytokinin receptors of Arabidopsis are located mainly to the endoplasmic reticulum Plant Physiol156,1808-1818. [本文引用: 1]
WybouwB, DeRybel B (2019). Cytokinin—a developing story Trends Plant Sci24,177-185. [本文引用: 1]
Zalaba?kD, GaluszkaP, MrízováK, Podle?a?kova?K, GuRL, Fre?bortova?J (2014). Biochemical characterization of the maize cytokinin dehydrogenase family and cytokinin profiling in developing maize plantlets in relation to the expression of cytokinin dehydrogenase genes Plant Physiol Biochem74,283-293. [本文引用: 1]
ZdarskaM, DobisováT, GelováZ, PernisováM, DabravolskiS, HejátkoJ (2015). Illuminating light, cytokinin, and ethylene signaling crosstalk in plant development J Exp Bot66,4913-4931. [本文引用: 1]
ZschiedrichCP, KeidelV, SzurmantH (2016). Molecular mechanisms of two-component signal transduction J Mol Biol428,3752-3775. [本文引用: 1]
ZubkoE, AdamsCJ, MacháèkováI, MalbeckJ, ScollanC, MeyerP (2002). Activation tagging identifies a gene from Petunia hybrida responsible for the production of active cytokinins in plants Plant J29,797-808. [本文引用: 1]
ZuboYO, BlakleyIC, YamburenkoMV, WorthenJM, StreetIH, Franco-ZorrillaJM, ZhangWJ, HillK, RainesT, SolanoR, KieberJJ, LoraineAE, SchallerGE (2017). Cytokinin induces genome-wide binding of the type-B response regulator ARR10 to regulate growth and development in Arabidopsis Proc Natl Acad Sci USA114,E5995-E6004. [本文引用: 3]
ZuboYO, SchallerGE (2020). Role of the cytokinin-activated type-B response regulators in hormone crosstalk Plants9,166. [本文引用: 12]
Modeling of protein-protein interactions in cytokinin signal transduction 10 2019
... 细胞分裂素是一种重要的植物激素, 在调控植物生长发育的许多过程中起关键作用, 包括细胞的分裂与分化、茎和根的生长、顶端优势维持、衰老、果实和种子发育、营养信号传递以及对生物和非生物胁迫的响应(Hwang et al.2012; Jameson and Song, 2016; Wybouw and De Rybel, 2019).近年来, CK的生物合成与代谢, 尤其是CK信号转导途径的研究取得了重要进展(Lomin et al.2018; Arkhipov et al.2019; Hallmark and Rashotte, 2019; Chen et al.2020).本文在综述CK的生物合成与代谢及其信号转导的基础上, 重点介绍CK调控种子发育、休眠与萌发的研究进展, 并提出了在本领域需要进一步研究的科学问题, 旨在为阐明CK调控种子发育、休眠与萌发的分子机理提供参考. ...
... 细胞分裂素信号转导途径也被称为双组分信号转导系统(two-component signaling system), 主要包括组氨酸激酶(histidine (His) kinase, HK)、组氨酸磷酸转移蛋白(His phosphotransfer protein, HP)和反应调控因子(response regulator, RR).其中HK作为CK的受体, HP将信号从HK传递到细胞核的RR, 从而调控靶基因转录(图1, 图2) (Hwang et al.2012; Kieber and Schaller, 2018; Lomin et al.2018; Arkhipov et al.2019; Hallmark and Rashotte, 2019; Zubo and Schaller, 2020).双分子荧光互补(bimolecular fluorescence complementation, BiFC)实验证明, 拟南芥CK受体AHK (Arabidopsis HK) 2-4均能与AHP (Arabidopsis HP)1-3相互作用, 且无明显的偏好; CK受体能形成同源和异源二聚体(Arkhipov et al.2019).AHK-AHK和AHK-AHP的相互作用主要定位于内质网(endoplasmic reticulum, ER); 相反, 各种磷酸转移蛋白(AHP)的同源和异源二聚体主要存在于植物的细胞核中(Arkhipov et al.2019). ...
... 2019).AHK-AHK和AHK-AHP的相互作用主要定位于内质网(endoplasmic reticulum, ER); 相反, 各种磷酸转移蛋白(AHP)的同源和异源二聚体主要存在于植物的细胞核中(Arkhipov et al.2019). ...
... 细胞分裂素信号被膜定位的组氨酸(His)激酶受体感受, 通过His磷酸转移蛋白被传递到细胞核, 从而激活细胞核中的转录因子家族.CK: 细胞分裂素; ATP: 腺苷三磷酸; ADP: 腺苷二磷酸; P: 磷酸; D: 保守的天冬氨酸; H: 保守的His; DI: 感受器模块的二聚体界面结构域; PAS和PAS-like: 感受器模块的CHASE结构域的子结构域; TM1和TM2: 跨膜结构域1和2; HisKA (DHpD): His激酶A结构域(二聚合作用和His磷酸转移结构域); H-ATPase (CAD): 腺苷三磷酸酶结构域(催化与ATP结合结构域); REC-like: 类接收器结构域; REC: 接收器结构域; HPt: 含有His的磷酸转移蛋白(磷酸传递蛋白); RR-B: B型反应调控因子(转录因子).蛋白-蛋白相互作用(PPI)用红色虚线表示. Intermolecular interactions in the cytokinin signaling pathway (modified from Lomin <i>et al.</i> <xref ref-type="bibr" rid="b42">2018</xref>; Arkhipov <i>et al.</i> <xref ref-type="bibr" rid="b7">2019</xref>) Cytokinin signaling is perceived by membrane-localized Histidine (His) kinase receptors and is transduced to nucleus through a His phosphotransfer protein to activate a family of transcription factors in the nucleus. CK: Cytokinin; ATP: Adenosine triphosphate; ADP: Adenosine diphosphate; P: Phosphate; D: Conserved Aspartate; H: Conserved His; DI: Dimerization interface domain of the sensor module; PAS and PAS-like: Subdomains of the CHASE domain of the sensor module; TM1 and TM2: Transmembrane domain 1 and 2; HisKA (DHpD): His kinase A domain (dimerization and His phosphotransfer domain); H-ATPase (CAD): Adenosine triphosphatase domain (catalytic and ATP-binding domain); REC-like: Receiver-like domain; REC: Receiver domain; HPt: His-containing phosphotransfer protein (phosphotransmitter); RR-B: Type B response regulator (transcription factor). Protein-protein interactions (PPI) are indicated by a red dotted line. ...
... 组氨酸激酶包含1个保守的与CK结合的胞外CHASE (cyclases/His kinases-associated sensing extracellular)区域、至少2个跨膜(transmembrane, TM)结构域和1个细胞质区域(图2) (Hwang et al.2012; Lomin et al.2018; Arkhipov et al.2019).HK主要定位于ER膜(Wulfetange et al.2011).CHASE区域即感受器模块(sensor module, SM), 面向ER腔或者质外体(apoplast), 包括二聚界面(dimerization interface, DI)、PAS (Per-Arnt-Sim)结构域和类PAS结构域(Hothorn et al.2011; Steklov et al.2013; Romanov et al.2018).TM区域由2-5个α-螺旋延伸组成.在马铃薯(S. tuberosum) CK受体的N端, StHK2、StHK3和StHK4的上游TM螺旋的数量分别为3、2和1个(Steklov et al.2013).HK的细胞质部分包括催化模块(catalytic module)和接收器模块(receiver module).催化模块由HK-二聚体(HisKA或者DHpD)和H-ATPase (CAD)结构域组成(Pekárováet al.2016); 接收器模块含有类接收器结构域(receiver-like domain, REC-like)和接收器结构域(receiver domain, REC), 它们定位于HK蛋白的C-端(图2) (Lomin et al.2018; Pekarova et al.2018; Arkhipov et al.2019).值得注意的是, AHK4同时具有组氨酸激酶和磷酸酶活性; 在配体缺乏时, AHK4的磷酸酶活性显著高于激酶活性, 能水解其接收器结构域上的磷酸基, 从而消耗可传递的磷酸基团(M?h?nenet al.2006). ...
... 在CK信号转导途径中, HP (也称为His磷酸传递蛋白(His phosphotransmitter, HPt))在HK受体的下游起作用, 介导磷酸基在多步骤磷酸转移中从活化的HK受体接收器结构域转移到RR接收器结构域(图1, 图2) (Hwang et al.2012; Arkhipov et al.2019).AHP不具有催化活性, 实际上是作为RR自身磷酸化活性的高能磷酸供体.AHP是CK信号转导冗余的正调控因子(Hutchison et al.2006), 但它们也在植物其它HK下游(如CKI1 (CYTOKININ-INDEPENDENT1))起作用(Liu et al.2017).AHP蛋白具有1个保守的半胱氨酸残基, 该残基能在体外和体内被一氧化氮(NO) S-亚硝酰化, 抑制AHP作为磷酸转移蛋白的作用, 降低AHP从HK接收磷酸基并将其转移到RR的能力(Feng et al.2013). ...
... 细胞分裂素与HK的PAS结合, 导致受体的二聚作用并被激活, CK的信号转导通过类似于原核生物的His-Asp磷酸传递途径进行(Keshishian and Rashotte, 2015; Zschiedrich et al.2016).在激活的受体ATP结合结构域中, ATP发生水解, 释放的磷酸基团与HisKA结构域中1个保守的His残基结合, 形成磷酸组氨酸(phosphohistidine).然后, 活化(hot)的磷酸基在分子内被转移到接收器结构域中1个保守的Asp残基上, 依次与HPt相互作用, 将活化的磷酸基转移到HPt保守的His残基上.HPt携带该活化的磷酸基并转移到细胞核和磷酸化RR蛋白保守的Asp残基.最后, 磷酸化的B型RR能够与DNA结合, 激活CK靶基因的转录(图1, 图2) (Kieber and Schaller, 2018; Pekarova et al.2018; Arkhipov et al.2019). ...
... Day等(2008)利用激光捕获显微切割(laser capture microdissection, LCM)技术对授粉后4天的种子进行研究, 在合胞体胚乳(syncytial endosperm)中发现与CK生物合成和信号转导相关的基因高丰度表达, 尤其是AtIPT8在发育中的胚乳合点区表达丰度更高.Belmonte等(2013)研究了拟南芥种子发育过程中亚区域基因的表达, 发现在种子发育的形态发生阶段IPT4和IPT8的高丰度表达局限于合点胚乳(chalazal endosperm).这些结果表明发育中的种子是CK生物合成的场所.小麦基因芯片(GeneChip)研究及其随后所选择基因的qRT-PCR分析表明, AC domain (休眠)和RL4452 (非休眠) 2种基因型种子的成熟都与胚和胚乳组织中注释为TaIPT2-2探针(基因)的增强表达有关; 在这2种基因型的种子成熟过程中, TaCKX4-3和TaCKX7-2探针在胚中的表达水平下降, 在胚乳中维持相似的水平(Tuan et al.2019).IPT编码催化CK生物合成限速步骤的酶, 且CKX催化CK的失活(Sakakibara, 2006; Frébortet al.2011), 这些结果表明, 种子成熟与胚和胚乳中CK水平的升高有关.相反, 随着种子成熟, 胚和胚乳组织中的iP水平显著降低, 但与基因型无关; 而tZ仅在2种基因型的胚组织中被检测到, 且在AC Domain基因型中的含量比在RL4452基因型中更低(Tuan et al.2019).在2种基因型种子成熟过程中, 胚和胚乳中iP水平的下降与胚TaLOG8和胚乳TaLOG3基因的表达水平下降有关(Tuan et al.2019).这些基因编码CK激活酶(LOG), 该酶直接将失活的CK核苷酸转化为生物活性的游离态CK (Kurakawa et al.2007).此外, 种子成熟时2种基因型的胚和胚乳组织中iP水平的下降以及AC Domain胚中tZ水平的显著下降与其相应组织中注释为TaGLU4探针的表达水平下降一致(Tuan et al.2019).TaGLU4编码一种通过羟基化作用从CK结合物中释放生物活性CK的酶(Cairns and Esen, 2010). ...
Cytokinin dehydrogenase: a genetic target for yield improvement in wheat 2 2020
... 细胞分裂素是一种重要的植物激素, 在调控植物生长发育的许多过程中起关键作用, 包括细胞的分裂与分化、茎和根的生长、顶端优势维持、衰老、果实和种子发育、营养信号传递以及对生物和非生物胁迫的响应(Hwang et al.2012; Jameson and Song, 2016; Wybouw and De Rybel, 2019).近年来, CK的生物合成与代谢, 尤其是CK信号转导途径的研究取得了重要进展(Lomin et al.2018; Arkhipov et al.2019; Hallmark and Rashotte, 2019; Chen et al.2020).本文在综述CK的生物合成与代谢及其信号转导的基础上, 重点介绍CK调控种子发育、休眠与萌发的研究进展, 并提出了在本领域需要进一步研究的科学问题, 旨在为阐明CK调控种子发育、休眠与萌发的分子机理提供参考. ...
... 细胞分裂素氧化酶/脱氢酶(cytokinin oxidase/dehydrogenase, CKX)在N6侧链上不可逆地裂解游离态CK和CK核苷, 以降低生物活性CK的水平(图1) (Sakakibara, 2006; Tuan et al.2019; Chen et al.2020).研究发现, CKX的过表达导致内源CK水平降低和多种发育缺陷; tZ和iP都能被CKX裂解, 但DZ和人工合成的CK (如激动素(kinetin)和6-BA)对CKX的裂解具有抗性(Zalabáket al.2014). ...
After-ripening induced transcriptional changes of hormonal genes in wheat seeds: the cases of brassinosteroids, ethylene, cytokinin and salicylic acid 4 2014
... 种子成熟过程中在母体植株上获得的休眠类型称为初生休眠(primary dormancy), 种子初生休眠的诱导和维持受植物激素调控(Shu et al.2016; Nonogaki, 2017, 2019; 宋松泉等, 2020a).研究表明, 种子中CK的水平和信号转导调控种子的发育、休眠与萌发(图1, 图3) (Kieber and Schaller, 2018; Tuan et al.2019; Nguyen et al.2020).后熟(after-ripening)是一种释放种子休眠的方式, 也通过影响CK的水平和信号转导来调控种子的休眠与萌发(Chitnis et al.2014). ...
... 后熟(干藏)通过影响CK的水平和信号转导调控种子的休眠与萌发.小麦(AC domain)种子在吸胀24小时后萌发率为0, 吸胀7天后萌发率为11%, 表明种子具有休眠特性; 经后熟10个月的种子在吸胀24小时内的萌发率为95%, 表明后熟促进小麦种子的萌发(Chitnis et al.2014).利用基因芯片技术, Chitnis等(2014)发现, 与未后熟的种子相比, 后熟种子在吸胀过程中有1个LOG、3个GLU、1个CKX和2个ZOG基因显著上调; 在40个注释为CK信号转导基因的探针中, 只有1个编码伪反应调控因子(pseudo-response regulator)的APRR9基因在后熟种子的吸胀过程中下调2倍以上(Chitnis et al.2014). ...
Ethylene, a key factor in the regulation of seed dormancy 1 2014
... 研究表明, 种子的萌发能力与乙烯的产生有关, 外源乙烯或乙烯利(ethrel, 乙烯释放剂)能解除种子的初生和次生休眠(second dormancy) (Corbineau et al.2014).CK通过转录和转录后机制激活ACC合酶(ACC synthase, ACS), 促进乙烯的生物合成; 乙烯的产生促进被CK抑制的暗生长幼苗的下胚轴伸长以及抑制根系的生长(Zdarska et al.2015).此外, B型ARR与许多影响乙烯信号转导的基因结合.尽管乙烯受体ETR2被CK下调表达, 但CK调控大多数编码调节乙烯信号转导途径活性的元件, 而不是途径本身.调节原件包括ARGOS基因家族的ARGOS和ARL以及TRP1、ETP和EBF基因.ARGOS是在受体水平起作用的跨膜蛋白, 可使植物对乙烯去敏感; 作为乙烯信号转导的正调控因子, TRP1可能通过与受体结合和干扰与CTR1的互作起作用; ETP和EBF基因分别编码参与EIN2和EIN3降解的F-box蛋白(Zubo and Schaller, 2020). ...
Transcriptome analysis of proliferating Arabidopsis endosperm reveals biological implications for the control of syncytial division, cytokinin signaling, and gene expression regulation 1 2008
... Day等(2008)利用激光捕获显微切割(laser capture microdissection, LCM)技术对授粉后4天的种子进行研究, 在合胞体胚乳(syncytial endosperm)中发现与CK生物合成和信号转导相关的基因高丰度表达, 尤其是AtIPT8在发育中的胚乳合点区表达丰度更高.Belmonte等(2013)研究了拟南芥种子发育过程中亚区域基因的表达, 发现在种子发育的形态发生阶段IPT4和IPT8的高丰度表达局限于合点胚乳(chalazal endosperm).这些结果表明发育中的种子是CK生物合成的场所.小麦基因芯片(GeneChip)研究及其随后所选择基因的qRT-PCR分析表明, AC domain (休眠)和RL4452 (非休眠) 2种基因型种子的成熟都与胚和胚乳组织中注释为TaIPT2-2探针(基因)的增强表达有关; 在这2种基因型的种子成熟过程中, TaCKX4-3和TaCKX7-2探针在胚中的表达水平下降, 在胚乳中维持相似的水平(Tuan et al.2019).IPT编码催化CK生物合成限速步骤的酶, 且CKX催化CK的失活(Sakakibara, 2006; Frébortet al.2011), 这些结果表明, 种子成熟与胚和胚乳中CK水平的升高有关.相反, 随着种子成熟, 胚和胚乳组织中的iP水平显著降低, 但与基因型无关; 而tZ仅在2种基因型的胚组织中被检测到, 且在AC Domain基因型中的含量比在RL4452基因型中更低(Tuan et al.2019).在2种基因型种子成熟过程中, 胚和胚乳中iP水平的下降与胚TaLOG8和胚乳TaLOG3基因的表达水平下降有关(Tuan et al.2019).这些基因编码CK激活酶(LOG), 该酶直接将失活的CK核苷酸转化为生物活性的游离态CK (Kurakawa et al.2007).此外, 种子成熟时2种基因型的胚和胚乳组织中iP水平的下降以及AC Domain胚中tZ水平的显著下降与其相应组织中注释为TaGLU4探针的表达水平下降一致(Tuan et al.2019).TaGLU4编码一种通过羟基化作用从CK结合物中释放生物活性CK的酶(Cairns and Esen, 2010). ...
Arabidopsis histidine kinase CKI1 acts upstream of histidine phosphotransfer proteins to regulate female gametophyte development and vegetative growth 1 2010
... 磷酸传递的信号转导、蛋白互作和高阶(higher- order)功能缺失突变的分析表明, AHP在功能上冗余且可以不加选择地与受体相互作用, 磷酸基的转移是双向的(Deng et al.2010).在6个AHP (AHP1-6)中, AHP6缺乏保守的His残基, 因此不能接受磷酸基, 被称为伪AHP (pseudo-AHP).AHP6负调控信号转导途径活性, 很可能与AHP1-5竞争与活化受体的相互作用(Lomin et al.2018). ...
Sequential action of cytokinin and gibberellic acid in wheat aleurone tissue 1 1969
S-nitrosylation of phosphotransfer proteins represses cytokinin signaling 1 2013
... 在CK信号转导途径中, HP (也称为His磷酸传递蛋白(His phosphotransmitter, HPt))在HK受体的下游起作用, 介导磷酸基在多步骤磷酸转移中从活化的HK受体接收器结构域转移到RR接收器结构域(图1, 图2) (Hwang et al.2012; Arkhipov et al.2019).AHP不具有催化活性, 实际上是作为RR自身磷酸化活性的高能磷酸供体.AHP是CK信号转导冗余的正调控因子(Hutchison et al.2006), 但它们也在植物其它HK下游(如CKI1 (CYTOKININ-INDEPENDENT1))起作用(Liu et al.2017).AHP蛋白具有1个保守的半胱氨酸残基, 该残基能在体外和体内被一氧化氮(NO) S-亚硝酰化, 抑制AHP作为磷酸转移蛋白的作用, 降低AHP从HK接收磷酸基并将其转移到RR的能力(Feng et al.2013). ...
Evolution of cytokinin biosynthesis and degradation 4 2011
... Day等(2008)利用激光捕获显微切割(laser capture microdissection, LCM)技术对授粉后4天的种子进行研究, 在合胞体胚乳(syncytial endosperm)中发现与CK生物合成和信号转导相关的基因高丰度表达, 尤其是AtIPT8在发育中的胚乳合点区表达丰度更高.Belmonte等(2013)研究了拟南芥种子发育过程中亚区域基因的表达, 发现在种子发育的形态发生阶段IPT4和IPT8的高丰度表达局限于合点胚乳(chalazal endosperm).这些结果表明发育中的种子是CK生物合成的场所.小麦基因芯片(GeneChip)研究及其随后所选择基因的qRT-PCR分析表明, AC domain (休眠)和RL4452 (非休眠) 2种基因型种子的成熟都与胚和胚乳组织中注释为TaIPT2-2探针(基因)的增强表达有关; 在这2种基因型的种子成熟过程中, TaCKX4-3和TaCKX7-2探针在胚中的表达水平下降, 在胚乳中维持相似的水平(Tuan et al.2019).IPT编码催化CK生物合成限速步骤的酶, 且CKX催化CK的失活(Sakakibara, 2006; Frébortet al.2011), 这些结果表明, 种子成熟与胚和胚乳中CK水平的升高有关.相反, 随着种子成熟, 胚和胚乳组织中的iP水平显著降低, 但与基因型无关; 而tZ仅在2种基因型的胚组织中被检测到, 且在AC Domain基因型中的含量比在RL4452基因型中更低(Tuan et al.2019).在2种基因型种子成熟过程中, 胚和胚乳中iP水平的下降与胚TaLOG8和胚乳TaLOG3基因的表达水平下降有关(Tuan et al.2019).这些基因编码CK激活酶(LOG), 该酶直接将失活的CK核苷酸转化为生物活性的游离态CK (Kurakawa et al.2007).此外, 种子成熟时2种基因型的胚和胚乳组织中iP水平的下降以及AC Domain胚中tZ水平的显著下降与其相应组织中注释为TaGLU4探针的表达水平下降一致(Tuan et al.2019).TaGLU4编码一种通过羟基化作用从CK结合物中释放生物活性CK的酶(Cairns and Esen, 2010). ...
... 细胞分裂素对种子发育、休眠与萌发的调控通过生物活性CK水平和信号转导途径来实现(图1, 图3) (Hwang et al.2012; Nguyen et al.2020; Zubo and Schaller, 2020).尽管近年来这些领域的研究已经取得了重要进展, 但仍然有一些科学问题需要进一步探索.例如, 已知生物活性CK可被CKX失活、通过cZOG可逆失活、被UGT不可逆失活以及通过βGlc重新活化(Frébortet al.2011; Jameson and Song, 2016), 那么, 这些反应是否同时存在于同一物种的同一组织, 还是具有物种、组织和发育阶段的特异性? 在响应种子发育、休眠与萌发和环境变化时, 为了控制组织或细胞内的生物活性CK水平, 这些途径怎样被启动、运行和协调? ...
Cytokinin antagonizes abscisic acid-mediated inhibition of cotyledon greening by promoting the degradation of ABSCISIC ACID INSENSITIVE 5 protein in Arabidopsis 5 2014
... ABI5是一种碱性亮氨酸拉链转录因子(basic leucine zipper transcription factor), 在ABA介导的抑制种子萌发和萌发后生长的调控中起重要作用(Guan et al.2014; 宋松泉等, 2020b).在种子萌发过程中, ABI5功能丧失能够降低ABA的敏感性, 而ABI5功能获得能增加ABA的敏感性(Liu and Stone, 2010).研究表明, ABI5与ARR4-6发生物理相互作用(Wang et al.2011).在arr3/arr4/arr5/arr6四突变体中, CK诱导的ABI5降低减少, 这表明A型ARR蛋白与ABI5的结合可能在调控ABI5蛋白的稳定性中起重要作用(Guan et al.2014).由于A型ARR mRNA的稳定水平和A型ARR蛋白的积累被CK正调控(Ren et al.2009), 因此, A型ARR-ABI5复合物可能抑制ABI5蛋白与蛋白酶体降解复合物的相互作用. ...
... 2014).由于A型ARR mRNA的稳定水平和A型ARR蛋白的积累被CK正调控(Ren et al.2009), 因此, A型ARR-ABI5复合物可能抑制ABI5蛋白与蛋白酶体降解复合物的相互作用. ...
... Guan等(2014)研究了CK对ABA信号转导组分表达水平的影响, 包括SnRK2 (sucrose non-fermenting1-related kinase 2)基因(SnRK2.2和SnRK2.3)、ABF/AREB(ABA-responsive element binding factor/ABA-responsive element binding protein)家族基因(ABF1-4)以及ABI (ABI1-5和ABI8)基因, 发现CK对这些基因的表达无显著影响; 大多数ABA调控基因的表达水平也不受CK影响, ABA诱导的ABI5的表达仅被CK轻微降低.用CK处理72-96小时后, ABI5蛋白几乎被完全降解, 但ABI5 mRNA被适度降低(Guan et al.2014), 这表明ABI5蛋白而不是ABI5mRNA的稳定性是CK介导ABA信号转导的主要调控步骤.CK诱导的ABI5蛋白的降解被蛋白酶体降解复合物专一性抑制剂MG132逆转.这些结果表明, CK调控的ABI5积累减少可能是由26S蛋白酶体途径介导(Liu and Stone, 2010; Guan et al.2014). ...
... 2014), 这表明ABI5蛋白而不是ABI5mRNA的稳定性是CK介导ABA信号转导的主要调控步骤.CK诱导的ABI5蛋白的降解被蛋白酶体降解复合物专一性抑制剂MG132逆转.这些结果表明, CK调控的ABI5积累减少可能是由26S蛋白酶体途径介导(Liu and Stone, 2010; Guan et al.2014). ...
... 2014). ...
Review—cytokinin response factors: responding to more than cytokinin 2 2019
... 细胞分裂素是一种重要的植物激素, 在调控植物生长发育的许多过程中起关键作用, 包括细胞的分裂与分化、茎和根的生长、顶端优势维持、衰老、果实和种子发育、营养信号传递以及对生物和非生物胁迫的响应(Hwang et al.2012; Jameson and Song, 2016; Wybouw and De Rybel, 2019).近年来, CK的生物合成与代谢, 尤其是CK信号转导途径的研究取得了重要进展(Lomin et al.2018; Arkhipov et al.2019; Hallmark and Rashotte, 2019; Chen et al.2020).本文在综述CK的生物合成与代谢及其信号转导的基础上, 重点介绍CK调控种子发育、休眠与萌发的研究进展, 并提出了在本领域需要进一步研究的科学问题, 旨在为阐明CK调控种子发育、休眠与萌发的分子机理提供参考. ...
... 细胞分裂素信号转导途径也被称为双组分信号转导系统(two-component signaling system), 主要包括组氨酸激酶(histidine (His) kinase, HK)、组氨酸磷酸转移蛋白(His phosphotransfer protein, HP)和反应调控因子(response regulator, RR).其中HK作为CK的受体, HP将信号从HK传递到细胞核的RR, 从而调控靶基因转录(图1, 图2) (Hwang et al.2012; Kieber and Schaller, 2018; Lomin et al.2018; Arkhipov et al.2019; Hallmark and Rashotte, 2019; Zubo and Schaller, 2020).双分子荧光互补(bimolecular fluorescence complementation, BiFC)实验证明, 拟南芥CK受体AHK (Arabidopsis HK) 2-4均能与AHP (Arabidopsis HP)1-3相互作用, 且无明显的偏好; CK受体能形成同源和异源二聚体(Arkhipov et al.2019).AHK-AHK和AHK-AHP的相互作用主要定位于内质网(endoplasmic reticulum, ER); 相反, 各种磷酸转移蛋白(AHP)的同源和异源二聚体主要存在于植物的细胞核中(Arkhipov et al.2019). ...
Regulation of cytokinin biosynthesis, compartmentalization and translocation 1 2008
Distinct metabolism of N-glucosides of isopentenyladenine and trans-zeatin determines cytokinin metabolic spectrum in Arabidopsis 2 2020
... 细胞分裂素曾被认为只能在根尖和茎尖合成, 但实际上它们能在植物体内的不同部位合成和起作用(Sakakibara, 2006).CK通过木质部(主要以tZ-核苷的形式)从根运输到茎, 也能够通过韧皮部(主要以iP型CK的形式)从茎运输到根(Kieber and Schaller, 2018).植物细胞中生物活性CK的水平由其生物合成、活化、失活、降解和运输调节, 从而决定细胞中CK的信号强度(Sakakibara, 2006; Jameson and Song, 2016).植物细胞中CK及其代谢物的多样性表明, 它们之间的相互转化可能是CK作用的主要调控机制(Ho?eket al.2020). ...
... 目前已在拟南芥(Takei et al.2001)、矮牵牛(Petunia hybrida) (Zubko et al.2002)和啤酒花(Humulus lupulus) (Sakano et al.2004)中鉴定了IPT基因.在拟南芥中, 7个IPT基因(AtIPT1和AtIPT3-8)参与CK的生物合成(Takei et al.2001).生化研究表明, 植物IPT优先利用ADP或ATP而不是AMP作为异戊二烯基的受体, 产生iP-核苷5′-二磷酸(iP riboside 5′-diphosphate, iPRDP)或iP-核苷5′-三磷酸(iP riboside 5'-triphosphate, iPRTP) (Sakanoet al.2004; Ho?eket al.2020).AtIPT1对AMP、ADP和ATP的Km值分别为18、14.6和11.4 μmol·L -1, 对DMAPP的Km值为8.3 μmol·L -1 (Takei et al.2001).AtIPT4对ADP、ATP和DMAPP的Km值分别为9.1、3.4和11.6 μmol·L -1(Kakimoto, 2001).在过表达任一AtIPT的拟南芥转基因株系中, iP型CK显著积累, 表明所有的AtIPT对于类异戊二烯供体都具有相似的底物偏好(Sakakibara et al.2005). ...
Structural basis for cytokinin recognition by Arabidopsis thaliana histidine kinase 4 2 2011
... 植物中天然存在的CK是腺嘌呤的衍生物, 在N6端含有1个由异戊二烯(isoprene)衍生的侧链或者1个芳香族侧链, 分别称为类异戊二烯CK (isoprenoid CK)和芳香族CK (aromatic CK) (Sakakibara, 2006).类异戊二烯CK包括N6-异戊烯腺嘌呤(N6-isopentenyladenine, iP)、反式玉米素(trans-zeatin, tZ)、顺式玉米素(cis-zeatin, cZ)和二氢玉米素(dihydrozeatin, DZ).其中, 主要衍生物通常是tZ和iP及其糖的结合物, 但在不同的植物物种、组织和发育阶段存在较大的差异(Sakakibara, 2006).例如, tZ和iP型CK是拟南芥(Arabidopsis thaliana)中的主要形式, 而cZ型CK则存在于玉米、水稻和鹰嘴豆(Cicer arietinum)中.而芳香族CK, 如ortho-topolin、meta-topolin及其甲氧基衍生物(分别为ortho-methoxytopolin和meta-methoxytopolin)和6-苄基腺嘌呤(6-benzyladenine, 6-BA)仅存在于某些植物中.尽管一些人工合成的CK衍生物具有生物活性, 但还未在植物中发现.CK的活性形式是游离态(free base) CK (Hothorn et al.2011; Lomin et al.2015), 但CK也可能以非活性形式存在, 如核苷(其中1个核糖与嘌呤环的N9连接)和核苷酸(其中核糖部分包含1个磷酸基) (Kieber and Schaller, 2018). ...
... 组氨酸激酶包含1个保守的与CK结合的胞外CHASE (cyclases/His kinases-associated sensing extracellular)区域、至少2个跨膜(transmembrane, TM)结构域和1个细胞质区域(图2) (Hwang et al.2012; Lomin et al.2018; Arkhipov et al.2019).HK主要定位于ER膜(Wulfetange et al.2011).CHASE区域即感受器模块(sensor module, SM), 面向ER腔或者质外体(apoplast), 包括二聚界面(dimerization interface, DI)、PAS (Per-Arnt-Sim)结构域和类PAS结构域(Hothorn et al.2011; Steklov et al.2013; Romanov et al.2018).TM区域由2-5个α-螺旋延伸组成.在马铃薯(S. tuberosum) CK受体的N端, StHK2、StHK3和StHK4的上游TM螺旋的数量分别为3、2和1个(Steklov et al.2013).HK的细胞质部分包括催化模块(catalytic module)和接收器模块(receiver module).催化模块由HK-二聚体(HisKA或者DHpD)和H-ATPase (CAD)结构域组成(Pekárováet al.2016); 接收器模块含有类接收器结构域(receiver-like domain, REC-like)和接收器结构域(receiver domain, REC), 它们定位于HK蛋白的C-端(图2) (Lomin et al.2018; Pekarova et al.2018; Arkhipov et al.2019).值得注意的是, AHK4同时具有组氨酸激酶和磷酸酶活性; 在配体缺乏时, AHK4的磷酸酶活性显著高于激酶活性, 能水解其接收器结构域上的磷酸基, 从而消耗可传递的磷酸基团(M?h?nenet al.2006). ...
N-glucosylation of cytokinins by glycosyltransferases of Arabidopsis thaliana 1 2004
... 细胞分裂素的葡糖基结合物在生物测定中无生物活性, 这些被结合的CK不能与CK受体结合(Spichal et al.2004).尽管O-葡糖苷和N-葡糖苷稳定性差异的生理意义至今还不清楚, 但易裂解的O-葡糖苷被认为是CK失活的稳定贮藏形式.虽然与CK O-葡糖基化有关的基因已被鉴定(Mok et al.2005), 但尚缺乏关于N-葡糖基化的分子生物学信息.在拟南芥中, UGT76C1和UGT76C2在N7或N9位置葡糖基化一系列CK碱基, 也在N7位置葡糖基化tZ, 生成tZ-O-葡糖苷(tZ-O-glucoside) (Hou et al.2004). ...
The Arabidopsis histidine phosphotransfer proteins are redundant positive regulators of cytokinin signaling 1 2006
... 在CK信号转导途径中, HP (也称为His磷酸传递蛋白(His phosphotransmitter, HPt))在HK受体的下游起作用, 介导磷酸基在多步骤磷酸转移中从活化的HK受体接收器结构域转移到RR接收器结构域(图1, 图2) (Hwang et al.2012; Arkhipov et al.2019).AHP不具有催化活性, 实际上是作为RR自身磷酸化活性的高能磷酸供体.AHP是CK信号转导冗余的正调控因子(Hutchison et al.2006), 但它们也在植物其它HK下游(如CKI1 (CYTOKININ-INDEPENDENT1))起作用(Liu et al.2017).AHP蛋白具有1个保守的半胱氨酸残基, 该残基能在体外和体内被一氧化氮(NO) S-亚硝酰化, 抑制AHP作为磷酸转移蛋白的作用, 降低AHP从HK接收磷酸基并将其转移到RR的能力(Feng et al.2013). ...
Cytokinin signaling networks 6 2012
... 细胞分裂素是一种重要的植物激素, 在调控植物生长发育的许多过程中起关键作用, 包括细胞的分裂与分化、茎和根的生长、顶端优势维持、衰老、果实和种子发育、营养信号传递以及对生物和非生物胁迫的响应(Hwang et al.2012; Jameson and Song, 2016; Wybouw and De Rybel, 2019).近年来, CK的生物合成与代谢, 尤其是CK信号转导途径的研究取得了重要进展(Lomin et al.2018; Arkhipov et al.2019; Hallmark and Rashotte, 2019; Chen et al.2020).本文在综述CK的生物合成与代谢及其信号转导的基础上, 重点介绍CK调控种子发育、休眠与萌发的研究进展, 并提出了在本领域需要进一步研究的科学问题, 旨在为阐明CK调控种子发育、休眠与萌发的分子机理提供参考. ...
... 细胞分裂素信号转导途径也被称为双组分信号转导系统(two-component signaling system), 主要包括组氨酸激酶(histidine (His) kinase, HK)、组氨酸磷酸转移蛋白(His phosphotransfer protein, HP)和反应调控因子(response regulator, RR).其中HK作为CK的受体, HP将信号从HK传递到细胞核的RR, 从而调控靶基因转录(图1, 图2) (Hwang et al.2012; Kieber and Schaller, 2018; Lomin et al.2018; Arkhipov et al.2019; Hallmark and Rashotte, 2019; Zubo and Schaller, 2020).双分子荧光互补(bimolecular fluorescence complementation, BiFC)实验证明, 拟南芥CK受体AHK (Arabidopsis HK) 2-4均能与AHP (Arabidopsis HP)1-3相互作用, 且无明显的偏好; CK受体能形成同源和异源二聚体(Arkhipov et al.2019).AHK-AHK和AHK-AHP的相互作用主要定位于内质网(endoplasmic reticulum, ER); 相反, 各种磷酸转移蛋白(AHP)的同源和异源二聚体主要存在于植物的细胞核中(Arkhipov et al.2019). ...
... 组氨酸激酶包含1个保守的与CK结合的胞外CHASE (cyclases/His kinases-associated sensing extracellular)区域、至少2个跨膜(transmembrane, TM)结构域和1个细胞质区域(图2) (Hwang et al.2012; Lomin et al.2018; Arkhipov et al.2019).HK主要定位于ER膜(Wulfetange et al.2011).CHASE区域即感受器模块(sensor module, SM), 面向ER腔或者质外体(apoplast), 包括二聚界面(dimerization interface, DI)、PAS (Per-Arnt-Sim)结构域和类PAS结构域(Hothorn et al.2011; Steklov et al.2013; Romanov et al.2018).TM区域由2-5个α-螺旋延伸组成.在马铃薯(S. tuberosum) CK受体的N端, StHK2、StHK3和StHK4的上游TM螺旋的数量分别为3、2和1个(Steklov et al.2013).HK的细胞质部分包括催化模块(catalytic module)和接收器模块(receiver module).催化模块由HK-二聚体(HisKA或者DHpD)和H-ATPase (CAD)结构域组成(Pekárováet al.2016); 接收器模块含有类接收器结构域(receiver-like domain, REC-like)和接收器结构域(receiver domain, REC), 它们定位于HK蛋白的C-端(图2) (Lomin et al.2018; Pekarova et al.2018; Arkhipov et al.2019).值得注意的是, AHK4同时具有组氨酸激酶和磷酸酶活性; 在配体缺乏时, AHK4的磷酸酶活性显著高于激酶活性, 能水解其接收器结构域上的磷酸基, 从而消耗可传递的磷酸基团(M?h?nenet al.2006). ...
... 在CK信号转导途径中, HP (也称为His磷酸传递蛋白(His phosphotransmitter, HPt))在HK受体的下游起作用, 介导磷酸基在多步骤磷酸转移中从活化的HK受体接收器结构域转移到RR接收器结构域(图1, 图2) (Hwang et al.2012; Arkhipov et al.2019).AHP不具有催化活性, 实际上是作为RR自身磷酸化活性的高能磷酸供体.AHP是CK信号转导冗余的正调控因子(Hutchison et al.2006), 但它们也在植物其它HK下游(如CKI1 (CYTOKININ-INDEPENDENT1))起作用(Liu et al.2017).AHP蛋白具有1个保守的半胱氨酸残基, 该残基能在体外和体内被一氧化氮(NO) S-亚硝酰化, 抑制AHP作为磷酸转移蛋白的作用, 降低AHP从HK接收磷酸基并将其转移到RR的能力(Feng et al.2013). ...
... 反应调控因子是一个转录因子家族, 包含1个保守的接受磷酸基的Asp残基(Keshishian and Rashotte, 2015).根据其系统发生和结构域, RR家族成员可分为3种类型: A型、B型和C型(Schaller et al.2007), 但只有A型和B型RR与CK信号转导有关(Kieber and Schaller, 2018; Tuan et al.2019; Zubo and Schaller, 2020).B型RR包括ARR (Arabidopsis RR) 1、ARR2、ARR10-14和ARR18-21, 能被HP接收器结构域中的Asp残基磷酸化激活, 作为CK反应的正调控因子起作用(Hwang et al.2012).B型RR的特征是包含1个接收器结构域和1个含有类MYB DNA结合结构域的长C-端延伸.接收器结构域包含1个保守的Asp残基, 当多步骤磷酸传递被激活时, Asp可作为调控磷酸化的位点.类MYB DNA结合结构域与1个短的(A/G) GAT核心DNA序列结合, 该序列对B型RR的结合作用非常关键(Zubo et al.2017; Zubo and Schaller, 2020).ChIP-seq (chromatin immunoprecipitation combined with sequencing)实验显示, B型RR在植物体内以CK依赖的方式与其靶点结合, 这种结合以类似于细菌RR的方式被接收器结构域磷酸化激活(Zubo et al.2017). ...
... 细胞分裂素对种子发育、休眠与萌发的调控通过生物活性CK水平和信号转导途径来实现(图1, 图3) (Hwang et al.2012; Nguyen et al.2020; Zubo and Schaller, 2020).尽管近年来这些领域的研究已经取得了重要进展, 但仍然有一些科学问题需要进一步探索.例如, 已知生物活性CK可被CKX失活、通过cZOG可逆失活、被UGT不可逆失活以及通过βGlc重新活化(Frébortet al.2011; Jameson and Song, 2016), 那么, 这些反应是否同时存在于同一物种的同一组织, 还是具有物种、组织和发育阶段的特异性? 在响应种子发育、休眠与萌发和环境变化时, 为了控制组织或细胞内的生物活性CK水平, 这些途径怎样被启动、运行和协调? ...
Cytokinins and expression of SWEET, SUT, CWINV and AAP genes increase as pea seeds germinate 6 2016
... 种子成熟过程中在母体植株上获得的休眠类型称为初生休眠(primary dormancy), 种子初生休眠的诱导和维持受植物激素调控(Shu et al.2016; Nonogaki, 2017, 2019; 宋松泉等, 2020a).研究表明, 种子中CK的水平和信号转导调控种子的发育、休眠与萌发(图1, 图3) (Kieber and Schaller, 2018; Tuan et al.2019; Nguyen et al.2020).后熟(after-ripening)是一种释放种子休眠的方式, 也通过影响CK的水平和信号转导来调控种子的休眠与萌发(Chitnis et al.2014). 10.11983/CBB20141.F0003
... 细胞分裂素是一种重要的植物激素, 在调控植物生长发育的许多过程中起关键作用, 包括细胞的分裂与分化、茎和根的生长、顶端优势维持、衰老、果实和种子发育、营养信号传递以及对生物和非生物胁迫的响应(Hwang et al.2012; Jameson and Song, 2016; Wybouw and De Rybel, 2019).近年来, CK的生物合成与代谢, 尤其是CK信号转导途径的研究取得了重要进展(Lomin et al.2018; Arkhipov et al.2019; Hallmark and Rashotte, 2019; Chen et al.2020).本文在综述CK的生物合成与代谢及其信号转导的基础上, 重点介绍CK调控种子发育、休眠与萌发的研究进展, 并提出了在本领域需要进一步研究的科学问题, 旨在为阐明CK调控种子发育、休眠与萌发的分子机理提供参考. ...
... 细胞分裂素曾被认为只能在根尖和茎尖合成, 但实际上它们能在植物体内的不同部位合成和起作用(Sakakibara, 2006).CK通过木质部(主要以tZ-核苷的形式)从根运输到茎, 也能够通过韧皮部(主要以iP型CK的形式)从茎运输到根(Kieber and Schaller, 2018).植物细胞中生物活性CK的水平由其生物合成、活化、失活、降解和运输调节, 从而决定细胞中CK的信号强度(Sakakibara, 2006; Jameson and Song, 2016).植物细胞中CK及其代谢物的多样性表明, 它们之间的相互转化可能是CK作用的主要调控机制(Ho?eket al.2020). ...
... 细胞分裂素对种子发育、休眠与萌发的调控通过生物活性CK水平和信号转导途径来实现(图1, 图3) (Hwang et al.2012; Nguyen et al.2020; Zubo and Schaller, 2020).尽管近年来这些领域的研究已经取得了重要进展, 但仍然有一些科学问题需要进一步探索.例如, 已知生物活性CK可被CKX失活、通过cZOG可逆失活、被UGT不可逆失活以及通过βGlc重新活化(Frébortet al.2011; Jameson and Song, 2016), 那么, 这些反应是否同时存在于同一物种的同一组织, 还是具有物种、组织和发育阶段的特异性? 在响应种子发育、休眠与萌发和环境变化时, 为了控制组织或细胞内的生物活性CK水平, 这些途径怎样被启动、运行和协调? ...
Identification of plant cytokinin biosynthetic enzymes as dimethylallyl diphosphate: ATP/ADP isopentenyltransferases 1 2001
... 目前已在拟南芥(Takei et al.2001)、矮牵牛(Petunia hybrida) (Zubko et al.2002)和啤酒花(Humulus lupulus) (Sakano et al.2004)中鉴定了IPT基因.在拟南芥中, 7个IPT基因(AtIPT1和AtIPT3-8)参与CK的生物合成(Takei et al.2001).生化研究表明, 植物IPT优先利用ADP或ATP而不是AMP作为异戊二烯基的受体, 产生iP-核苷5′-二磷酸(iP riboside 5′-diphosphate, iPRDP)或iP-核苷5′-三磷酸(iP riboside 5'-triphosphate, iPRTP) (Sakanoet al.2004; Ho?eket al.2020).AtIPT1对AMP、ADP和ATP的Km值分别为18、14.6和11.4 μmol·L -1, 对DMAPP的Km值为8.3 μmol·L -1 (Takei et al.2001).AtIPT4对ADP、ATP和DMAPP的Km值分别为9.1、3.4和11.6 μmol·L -1(Kakimoto, 2001).在过表达任一AtIPT的拟南芥转基因株系中, iP型CK显著积累, 表明所有的AtIPT对于类异戊二烯供体都具有相似的底物偏好(Sakakibara et al.2005). ...
Distinct isoprenoid origins of cis- and trans-zeatin biosyntheses in Arabidopsis 2 2004
... 反应调控因子是一个转录因子家族, 包含1个保守的接受磷酸基的Asp残基(Keshishian and Rashotte, 2015).根据其系统发生和结构域, RR家族成员可分为3种类型: A型、B型和C型(Schaller et al.2007), 但只有A型和B型RR与CK信号转导有关(Kieber and Schaller, 2018; Tuan et al.2019; Zubo and Schaller, 2020).B型RR包括ARR (Arabidopsis RR) 1、ARR2、ARR10-14和ARR18-21, 能被HP接收器结构域中的Asp残基磷酸化激活, 作为CK反应的正调控因子起作用(Hwang et al.2012).B型RR的特征是包含1个接收器结构域和1个含有类MYB DNA结合结构域的长C-端延伸.接收器结构域包含1个保守的Asp残基, 当多步骤磷酸传递被激活时, Asp可作为调控磷酸化的位点.类MYB DNA结合结构域与1个短的(A/G) GAT核心DNA序列结合, 该序列对B型RR的结合作用非常关键(Zubo et al.2017; Zubo and Schaller, 2020).ChIP-seq (chromatin immunoprecipitation combined with sequencing)实验显示, B型RR在植物体内以CK依赖的方式与其靶点结合, 这种结合以类似于细菌RR的方式被接收器结构域磷酸化激活(Zubo et al.2017). ...
... 细胞分裂素与HK的PAS结合, 导致受体的二聚作用并被激活, CK的信号转导通过类似于原核生物的His-Asp磷酸传递途径进行(Keshishian and Rashotte, 2015; Zschiedrich et al.2016).在激活的受体ATP结合结构域中, ATP发生水解, 释放的磷酸基团与HisKA结构域中1个保守的His残基结合, 形成磷酸组氨酸(phosphohistidine).然后, 活化(hot)的磷酸基在分子内被转移到接收器结构域中1个保守的Asp残基上, 依次与HPt相互作用, 将活化的磷酸基转移到HPt保守的His残基上.HPt携带该活化的磷酸基并转移到细胞核和磷酸化RR蛋白保守的Asp残基.最后, 磷酸化的B型RR能够与DNA结合, 激活CK靶基因的转录(图1, 图2) (Kieber and Schaller, 2018; Pekarova et al.2018; Arkhipov et al.2019). ...
... 在小麦种子成熟过程中, TaAHK4、TaARR9和TaARR12的表达水平在AC Domain和RL4452的胚中均表现下降, 这与iP和tZ的积累下降有关.然而, 在开花后20天, AC Domain胚中TaARR12的表达水平比RL4452更低(Tuan et al.2019).鉴于TaARR12属于B型ARR, 且作为CK反应的正调控因子起作用(Kieber and Schaller, 2018), TaARR12很可能抑制AC Domain胚中的CK反应, 从而在小麦种子成熟过程中起诱导休眠的作用.2种基因型胚乳中TaAHP2探针的表达随着种子成熟而下降.然而, 在种子成熟中期(开花后30天), TaAHK4探针在AC Domain胚乳中的表达水平比RL4452更低(Tuan et al.2019).由于AHK是CK的受体(Keshishian and Rashotte, 2015), 因此上述结果表明, RL4452对CK的反应敏感性增加, 从而抑制种子的休眠.另一方面, 在种子成熟的早期到中期(开花后20-30天), AC Domain胚乳中TaARR9的表达水平比RL4452更高(Tuan et al.2019).由于TaARR9属于负调控CK信号转导的A型ARR (Kieber and Schaller, 2018), 在成熟时具有深休眠的AC Domain种子胚乳对CK的反应敏感性降低, 其结果是成熟时休眠程度高.此外, AC Domain胚乳中AHK探针的表达下降也可能表明CK反应受到抑制, 从而诱导休眠. ...
Inhibition of gibberellic acid-induced germination by abscisic acid and reversal by cytokinins 1 1968
... ABA抑制种子萌发, CK能逆转ABA对莴苣(Lactuca sativa)种子萌发的抑制作用(Kham, 1968).1 mmol·L-1 ABA对黑黄檀(Dalbergia fusca)种子萌发的抑制作用能够被0.001-0.1 mmol·L-1 6-BA所拮抗(邓志军和宋松泉, 2008).过表达AtIPT8能降低gim1 (germination insensitive to ABA mutant 1)突变体种子萌发对ABA的敏感性; AtIPT8的功能丧失导致ipt8-1-ko和ipt8-2-ko突变体产生ABA过敏性反应; 在gim1中, AtIPT8的过表达能够通过促进CK的生物合成来增强CK信号, 从而上调CK信号转导的转录调控因子(如A型ARR4-6) (Wang et al.2011).在野生型拟南芥Columbia-0背景下, ABA处理能够通过抑制AtIPT3和AtIPT8的表达水平减少CK的内源生物合成, 随后降低A型ARR4-6的表达水平(Wang et al.2011).此外, 在萌发10天的gim1幼苗中, ABA处理也会抑制AtIPT3和AtIPT8的表达, 降低A型ARR4-6的表达丰度(Wang et al.2011).值得注意的是, gim1突变体中CK水平的增高不是由于ABA生物合成的改变, 而是降低了ABA和胁迫反应基因(如ABI5(ABSCISIC ACID INSENSITIVE 5))的表达水平(Wang et al.2011). ...
Cytokinins 1 2014
... A型ARR (如ARR3-9、ARR15-17)由1个接收器结构域和1个短的C-端延伸组成, 作为CK信号转导的负反馈调控因子起作用; 但与B型RR不同, 它们缺乏典型的转录调控输出结构域(output domain) (Kieber and Schaller, 2018).在对CK的反应中, A型RR通过B型RR的直接激活被诱导转录(Taniguchi et al.2007), CK也以磷酸化依赖的方式稳定许多A型RR (To et al.2007).A型ARR负调控CK信号转导的机制是与B型ARR竞争AHP的磷酸化, 从而导致多步骤磷酸传递与B型ARR转录调控因子解离, 以减少其对基因表达的影响(Kieber and Schaller, 2014; Zubo and Schaller, 2020). ...
Cytokinin signaling in plant development 12 2018
... 植物中天然存在的CK是腺嘌呤的衍生物, 在N6端含有1个由异戊二烯(isoprene)衍生的侧链或者1个芳香族侧链, 分别称为类异戊二烯CK (isoprenoid CK)和芳香族CK (aromatic CK) (Sakakibara, 2006).类异戊二烯CK包括N6-异戊烯腺嘌呤(N6-isopentenyladenine, iP)、反式玉米素(trans-zeatin, tZ)、顺式玉米素(cis-zeatin, cZ)和二氢玉米素(dihydrozeatin, DZ).其中, 主要衍生物通常是tZ和iP及其糖的结合物, 但在不同的植物物种、组织和发育阶段存在较大的差异(Sakakibara, 2006).例如, tZ和iP型CK是拟南芥(Arabidopsis thaliana)中的主要形式, 而cZ型CK则存在于玉米、水稻和鹰嘴豆(Cicer arietinum)中.而芳香族CK, 如ortho-topolin、meta-topolin及其甲氧基衍生物(分别为ortho-methoxytopolin和meta-methoxytopolin)和6-苄基腺嘌呤(6-benzyladenine, 6-BA)仅存在于某些植物中.尽管一些人工合成的CK衍生物具有生物活性, 但还未在植物中发现.CK的活性形式是游离态(free base) CK (Hothorn et al.2011; Lomin et al.2015), 但CK也可能以非活性形式存在, 如核苷(其中1个核糖与嘌呤环的N9连接)和核苷酸(其中核糖部分包含1个磷酸基) (Kieber and Schaller, 2018). ...
... 细胞分裂素曾被认为只能在根尖和茎尖合成, 但实际上它们能在植物体内的不同部位合成和起作用(Sakakibara, 2006).CK通过木质部(主要以tZ-核苷的形式)从根运输到茎, 也能够通过韧皮部(主要以iP型CK的形式)从茎运输到根(Kieber and Schaller, 2018).植物细胞中生物活性CK的水平由其生物合成、活化、失活、降解和运输调节, 从而决定细胞中CK的信号强度(Sakakibara, 2006; Jameson and Song, 2016).植物细胞中CK及其代谢物的多样性表明, 它们之间的相互转化可能是CK作用的主要调控机制(Ho?eket al.2020). ...
A model of cytokinin (CK) metabolism and signaling transduction (modified from Kieber and Schaller, <xref ref-type="bibr" rid="b35">2018</xref>; Zubo and Schaller, <xref ref-type="bibr" rid="b80">2020</xref>) Ade: Adenine; ADP: Adenosine diphosphate; AMP: Adenosine monophosphate; ATP: Adenosine triphosphate; CKX: CK oxidase/dehydrogenase; CYP735A: Cytochrome P450 monooxygenase; cZOG: cis-Zeatin-O-glucosyltransferase; DMAPP: Dimethylallyl diphosphate; HK: Histidin (His) kinase; HP: His phosphotransfer protein; IPT: Isopentenyltransferase; LOG: CK nucleoside 5′-monophosphate phosphoribohydrolase; SC: Sidechain; Type-B RRs: Type-B response regulators; Type-A RRs: Type-A response regulators; UGT: UDP glycosyltransferase ...
... 细胞分裂素信号转导途径也被称为双组分信号转导系统(two-component signaling system), 主要包括组氨酸激酶(histidine (His) kinase, HK)、组氨酸磷酸转移蛋白(His phosphotransfer protein, HP)和反应调控因子(response regulator, RR).其中HK作为CK的受体, HP将信号从HK传递到细胞核的RR, 从而调控靶基因转录(图1, 图2) (Hwang et al.2012; Kieber and Schaller, 2018; Lomin et al.2018; Arkhipov et al.2019; Hallmark and Rashotte, 2019; Zubo and Schaller, 2020).双分子荧光互补(bimolecular fluorescence complementation, BiFC)实验证明, 拟南芥CK受体AHK (Arabidopsis HK) 2-4均能与AHP (Arabidopsis HP)1-3相互作用, 且无明显的偏好; CK受体能形成同源和异源二聚体(Arkhipov et al.2019).AHK-AHK和AHK-AHP的相互作用主要定位于内质网(endoplasmic reticulum, ER); 相反, 各种磷酸转移蛋白(AHP)的同源和异源二聚体主要存在于植物的细胞核中(Arkhipov et al.2019). ...
... 反应调控因子是一个转录因子家族, 包含1个保守的接受磷酸基的Asp残基(Keshishian and Rashotte, 2015).根据其系统发生和结构域, RR家族成员可分为3种类型: A型、B型和C型(Schaller et al.2007), 但只有A型和B型RR与CK信号转导有关(Kieber and Schaller, 2018; Tuan et al.2019; Zubo and Schaller, 2020).B型RR包括ARR (Arabidopsis RR) 1、ARR2、ARR10-14和ARR18-21, 能被HP接收器结构域中的Asp残基磷酸化激活, 作为CK反应的正调控因子起作用(Hwang et al.2012).B型RR的特征是包含1个接收器结构域和1个含有类MYB DNA结合结构域的长C-端延伸.接收器结构域包含1个保守的Asp残基, 当多步骤磷酸传递被激活时, Asp可作为调控磷酸化的位点.类MYB DNA结合结构域与1个短的(A/G) GAT核心DNA序列结合, 该序列对B型RR的结合作用非常关键(Zubo et al.2017; Zubo and Schaller, 2020).ChIP-seq (chromatin immunoprecipitation combined with sequencing)实验显示, B型RR在植物体内以CK依赖的方式与其靶点结合, 这种结合以类似于细菌RR的方式被接收器结构域磷酸化激活(Zubo et al.2017). ...
... A型ARR (如ARR3-9、ARR15-17)由1个接收器结构域和1个短的C-端延伸组成, 作为CK信号转导的负反馈调控因子起作用; 但与B型RR不同, 它们缺乏典型的转录调控输出结构域(output domain) (Kieber and Schaller, 2018).在对CK的反应中, A型RR通过B型RR的直接激活被诱导转录(Taniguchi et al.2007), CK也以磷酸化依赖的方式稳定许多A型RR (To et al.2007).A型ARR负调控CK信号转导的机制是与B型ARR竞争AHP的磷酸化, 从而导致多步骤磷酸传递与B型ARR转录调控因子解离, 以减少其对基因表达的影响(Kieber and Schaller, 2014; Zubo and Schaller, 2020). ...
... 细胞分裂素与HK的PAS结合, 导致受体的二聚作用并被激活, CK的信号转导通过类似于原核生物的His-Asp磷酸传递途径进行(Keshishian and Rashotte, 2015; Zschiedrich et al.2016).在激活的受体ATP结合结构域中, ATP发生水解, 释放的磷酸基团与HisKA结构域中1个保守的His残基结合, 形成磷酸组氨酸(phosphohistidine).然后, 活化(hot)的磷酸基在分子内被转移到接收器结构域中1个保守的Asp残基上, 依次与HPt相互作用, 将活化的磷酸基转移到HPt保守的His残基上.HPt携带该活化的磷酸基并转移到细胞核和磷酸化RR蛋白保守的Asp残基.最后, 磷酸化的B型RR能够与DNA结合, 激活CK靶基因的转录(图1, 图2) (Kieber and Schaller, 2018; Pekarova et al.2018; Arkhipov et al.2019). ...
... 种子成熟过程中在母体植株上获得的休眠类型称为初生休眠(primary dormancy), 种子初生休眠的诱导和维持受植物激素调控(Shu et al.2016; Nonogaki, 2017, 2019; 宋松泉等, 2020a).研究表明, 种子中CK的水平和信号转导调控种子的发育、休眠与萌发(图1, 图3) (Kieber and Schaller, 2018; Tuan et al.2019; Nguyen et al.2020).后熟(after-ripening)是一种释放种子休眠的方式, 也通过影响CK的水平和信号转导来调控种子的休眠与萌发(Chitnis et al.2014). ...
... 在小麦种子成熟过程中, TaAHK4、TaARR9和TaARR12的表达水平在AC Domain和RL4452的胚中均表现下降, 这与iP和tZ的积累下降有关.然而, 在开花后20天, AC Domain胚中TaARR12的表达水平比RL4452更低(Tuan et al.2019).鉴于TaARR12属于B型ARR, 且作为CK反应的正调控因子起作用(Kieber and Schaller, 2018), TaARR12很可能抑制AC Domain胚中的CK反应, 从而在小麦种子成熟过程中起诱导休眠的作用.2种基因型胚乳中TaAHP2探针的表达随着种子成熟而下降.然而, 在种子成熟中期(开花后30天), TaAHK4探针在AC Domain胚乳中的表达水平比RL4452更低(Tuan et al.2019).由于AHK是CK的受体(Keshishian and Rashotte, 2015), 因此上述结果表明, RL4452对CK的反应敏感性增加, 从而抑制种子的休眠.另一方面, 在种子成熟的早期到中期(开花后20-30天), AC Domain胚乳中TaARR9的表达水平比RL4452更高(Tuan et al.2019).由于TaARR9属于负调控CK信号转导的A型ARR (Kieber and Schaller, 2018), 在成熟时具有深休眠的AC Domain种子胚乳对CK的反应敏感性降低, 其结果是成熟时休眠程度高.此外, AC Domain胚乳中AHK探针的表达下降也可能表明CK反应受到抑制, 从而诱导休眠. ...
... 属于负调控CK信号转导的A型ARR (Kieber and Schaller, 2018), 在成熟时具有深休眠的AC Domain种子胚乳对CK的反应敏感性降低, 其结果是成熟时休眠程度高.此外, AC Domain胚乳中AHK探针的表达下降也可能表明CK反应受到抑制, 从而诱导休眠. ...
... Day等(2008)利用激光捕获显微切割(laser capture microdissection, LCM)技术对授粉后4天的种子进行研究, 在合胞体胚乳(syncytial endosperm)中发现与CK生物合成和信号转导相关的基因高丰度表达, 尤其是AtIPT8在发育中的胚乳合点区表达丰度更高.Belmonte等(2013)研究了拟南芥种子发育过程中亚区域基因的表达, 发现在种子发育的形态发生阶段IPT4和IPT8的高丰度表达局限于合点胚乳(chalazal endosperm).这些结果表明发育中的种子是CK生物合成的场所.小麦基因芯片(GeneChip)研究及其随后所选择基因的qRT-PCR分析表明, AC domain (休眠)和RL4452 (非休眠) 2种基因型种子的成熟都与胚和胚乳组织中注释为TaIPT2-2探针(基因)的增强表达有关; 在这2种基因型的种子成熟过程中, TaCKX4-3和TaCKX7-2探针在胚中的表达水平下降, 在胚乳中维持相似的水平(Tuan et al.2019).IPT编码催化CK生物合成限速步骤的酶, 且CKX催化CK的失活(Sakakibara, 2006; Frébortet al.2011), 这些结果表明, 种子成熟与胚和胚乳中CK水平的升高有关.相反, 随着种子成熟, 胚和胚乳组织中的iP水平显著降低, 但与基因型无关; 而tZ仅在2种基因型的胚组织中被检测到, 且在AC Domain基因型中的含量比在RL4452基因型中更低(Tuan et al.2019).在2种基因型种子成熟过程中, 胚和胚乳中iP水平的下降与胚TaLOG8和胚乳TaLOG3基因的表达水平下降有关(Tuan et al.2019).这些基因编码CK激活酶(LOG), 该酶直接将失活的CK核苷酸转化为生物活性的游离态CK (Kurakawa et al.2007).此外, 种子成熟时2种基因型的胚和胚乳组织中iP水平的下降以及AC Domain胚中tZ水平的显著下降与其相应组织中注释为TaGLU4探针的表达水平下降一致(Tuan et al.2019).TaGLU4编码一种通过羟基化作用从CK结合物中释放生物活性CK的酶(Cairns and Esen, 2010). ...
Effects of temperature, after-ripening, stratification, and scarification plus hormone treatments on dormancy release and germination of Acer truncatum seeds 1 2009
Abscisic acid increases Arabidopsis ABI5 transcription factor levels by promoting KEG E3 ligase self-ubiquitination and proteasomal degradation 2 2010
... ABI5是一种碱性亮氨酸拉链转录因子(basic leucine zipper transcription factor), 在ABA介导的抑制种子萌发和萌发后生长的调控中起重要作用(Guan et al.2014; 宋松泉等, 2020b).在种子萌发过程中, ABI5功能丧失能够降低ABA的敏感性, 而ABI5功能获得能增加ABA的敏感性(Liu and Stone, 2010).研究表明, ABI5与ARR4-6发生物理相互作用(Wang et al.2011).在arr3/arr4/arr5/arr6四突变体中, CK诱导的ABI5降低减少, 这表明A型ARR蛋白与ABI5的结合可能在调控ABI5蛋白的稳定性中起重要作用(Guan et al.2014).由于A型ARR mRNA的稳定水平和A型ARR蛋白的积累被CK正调控(Ren et al.2009), 因此, A型ARR-ABI5复合物可能抑制ABI5蛋白与蛋白酶体降解复合物的相互作用. ...
... Guan等(2014)研究了CK对ABA信号转导组分表达水平的影响, 包括SnRK2 (sucrose non-fermenting1-related kinase 2)基因(SnRK2.2和SnRK2.3)、ABF/AREB(ABA-responsive element binding factor/ABA-responsive element binding protein)家族基因(ABF1-4)以及ABI (ABI1-5和ABI8)基因, 发现CK对这些基因的表达无显著影响; 大多数ABA调控基因的表达水平也不受CK影响, ABA诱导的ABI5的表达仅被CK轻微降低.用CK处理72-96小时后, ABI5蛋白几乎被完全降解, 但ABI5 mRNA被适度降低(Guan et al.2014), 这表明ABI5蛋白而不是ABI5mRNA的稳定性是CK介导ABA信号转导的主要调控步骤.CK诱导的ABI5蛋白的降解被蛋白酶体降解复合物专一性抑制剂MG132逆转.这些结果表明, CK调控的ABI5积累减少可能是由26S蛋白酶体途径介导(Liu and Stone, 2010; Guan et al.2014). ...
Auxin controls seed dormancy through stimulation of abscisic acid signaling by inducing ARF-mediated ABI3 activation in Arabidopsis 1 2013
... 在黑暗条件下, 1 μmol·L -1外源吲哚乙酸(indole acitic acid, IAA)处理显著延缓大豆(Glycine max)种子的萌发进程, 且随着IAA浓度的增加其抑制作用增强(Shuai et al.2017).外源IAA也以剂量依赖的方式有效增强野生型拟南芥新鲜收获种子的休眠(Liu et al.2013).基因表达分析显示, 生长素诱导A型ARR7和ARR15的转录, 抑制拟南芥根发生过程中的CK反应; 相反, 在茎分生组织中, 生长素抑制A型ARR的表达, 增强这些细胞对CK的敏感性.CK抑制生长素生物合成基因IAMT1和ILL6的表达, 诱导失活基因GH3.6的表达.CK还抑制AUX1生长素输出载体的表达, 影响生长素从根尖向茎的运输, 导致根细胞增殖减少.B型ARR的结合位点与编码生长素信号转导途径组分的基因(包括2个编码生长素受体的基因、10个AUX/IAA基因和4个ARF基因)有关.CK也诱导AUX/IAA基因SHY2/IAA3的表达, 该基因作为生长素信号转导的负调控因子起作用(Zubo and Schaller, 2020). ...
AHP2, AHP3, and AHP5 act downstream of CKI1 in Arabidopsis female gametophyte development 1 2017
... 在CK信号转导途径中, HP (也称为His磷酸传递蛋白(His phosphotransmitter, HPt))在HK受体的下游起作用, 介导磷酸基在多步骤磷酸转移中从活化的HK受体接收器结构域转移到RR接收器结构域(图1, 图2) (Hwang et al.2012; Arkhipov et al.2019).AHP不具有催化活性, 实际上是作为RR自身磷酸化活性的高能磷酸供体.AHP是CK信号转导冗余的正调控因子(Hutchison et al.2006), 但它们也在植物其它HK下游(如CKI1 (CYTOKININ-INDEPENDENT1))起作用(Liu et al.2017).AHP蛋白具有1个保守的半胱氨酸残基, 该残基能在体外和体内被一氧化氮(NO) S-亚硝酰化, 抑制AHP作为磷酸转移蛋白的作用, 降低AHP从HK接收磷酸基并将其转移到RR的能力(Feng et al.2013). ...
Plant membrane assays with cytokinin receptors underpin the unique role of free cytokinin bases as biologically active ligands 1 2015
... 植物中天然存在的CK是腺嘌呤的衍生物, 在N6端含有1个由异戊二烯(isoprene)衍生的侧链或者1个芳香族侧链, 分别称为类异戊二烯CK (isoprenoid CK)和芳香族CK (aromatic CK) (Sakakibara, 2006).类异戊二烯CK包括N6-异戊烯腺嘌呤(N6-isopentenyladenine, iP)、反式玉米素(trans-zeatin, tZ)、顺式玉米素(cis-zeatin, cZ)和二氢玉米素(dihydrozeatin, DZ).其中, 主要衍生物通常是tZ和iP及其糖的结合物, 但在不同的植物物种、组织和发育阶段存在较大的差异(Sakakibara, 2006).例如, tZ和iP型CK是拟南芥(Arabidopsis thaliana)中的主要形式, 而cZ型CK则存在于玉米、水稻和鹰嘴豆(Cicer arietinum)中.而芳香族CK, 如ortho-topolin、meta-topolin及其甲氧基衍生物(分别为ortho-methoxytopolin和meta-methoxytopolin)和6-苄基腺嘌呤(6-benzyladenine, 6-BA)仅存在于某些植物中.尽管一些人工合成的CK衍生物具有生物活性, 但还未在植物中发现.CK的活性形式是游离态(free base) CK (Hothorn et al.2011; Lomin et al.2015), 但CK也可能以非活性形式存在, 如核苷(其中1个核糖与嘌呤环的N9连接)和核苷酸(其中核糖部分包含1个磷酸基) (Kieber and Schaller, 2018). ...
Cytokinin perception in potato: new features of canonical players 9 2018
... 细胞分裂素是一种重要的植物激素, 在调控植物生长发育的许多过程中起关键作用, 包括细胞的分裂与分化、茎和根的生长、顶端优势维持、衰老、果实和种子发育、营养信号传递以及对生物和非生物胁迫的响应(Hwang et al.2012; Jameson and Song, 2016; Wybouw and De Rybel, 2019).近年来, CK的生物合成与代谢, 尤其是CK信号转导途径的研究取得了重要进展(Lomin et al.2018; Arkhipov et al.2019; Hallmark and Rashotte, 2019; Chen et al.2020).本文在综述CK的生物合成与代谢及其信号转导的基础上, 重点介绍CK调控种子发育、休眠与萌发的研究进展, 并提出了在本领域需要进一步研究的科学问题, 旨在为阐明CK调控种子发育、休眠与萌发的分子机理提供参考. ...
... 细胞分裂素信号转导途径也被称为双组分信号转导系统(two-component signaling system), 主要包括组氨酸激酶(histidine (His) kinase, HK)、组氨酸磷酸转移蛋白(His phosphotransfer protein, HP)和反应调控因子(response regulator, RR).其中HK作为CK的受体, HP将信号从HK传递到细胞核的RR, 从而调控靶基因转录(图1, 图2) (Hwang et al.2012; Kieber and Schaller, 2018; Lomin et al.2018; Arkhipov et al.2019; Hallmark and Rashotte, 2019; Zubo and Schaller, 2020).双分子荧光互补(bimolecular fluorescence complementation, BiFC)实验证明, 拟南芥CK受体AHK (Arabidopsis HK) 2-4均能与AHP (Arabidopsis HP)1-3相互作用, 且无明显的偏好; CK受体能形成同源和异源二聚体(Arkhipov et al.2019).AHK-AHK和AHK-AHP的相互作用主要定位于内质网(endoplasmic reticulum, ER); 相反, 各种磷酸转移蛋白(AHP)的同源和异源二聚体主要存在于植物的细胞核中(Arkhipov et al.2019). ...
... 细胞分裂素信号被膜定位的组氨酸(His)激酶受体感受, 通过His磷酸转移蛋白被传递到细胞核, 从而激活细胞核中的转录因子家族.CK: 细胞分裂素; ATP: 腺苷三磷酸; ADP: 腺苷二磷酸; P: 磷酸; D: 保守的天冬氨酸; H: 保守的His; DI: 感受器模块的二聚体界面结构域; PAS和PAS-like: 感受器模块的CHASE结构域的子结构域; TM1和TM2: 跨膜结构域1和2; HisKA (DHpD): His激酶A结构域(二聚合作用和His磷酸转移结构域); H-ATPase (CAD): 腺苷三磷酸酶结构域(催化与ATP结合结构域); REC-like: 类接收器结构域; REC: 接收器结构域; HPt: 含有His的磷酸转移蛋白(磷酸传递蛋白); RR-B: B型反应调控因子(转录因子).蛋白-蛋白相互作用(PPI)用红色虚线表示. Intermolecular interactions in the cytokinin signaling pathway (modified from Lomin <i>et al.</i> <xref ref-type="bibr" rid="b42">2018</xref>; Arkhipov <i>et al.</i> <xref ref-type="bibr" rid="b7">2019</xref>) Cytokinin signaling is perceived by membrane-localized Histidine (His) kinase receptors and is transduced to nucleus through a His phosphotransfer protein to activate a family of transcription factors in the nucleus. CK: Cytokinin; ATP: Adenosine triphosphate; ADP: Adenosine diphosphate; P: Phosphate; D: Conserved Aspartate; H: Conserved His; DI: Dimerization interface domain of the sensor module; PAS and PAS-like: Subdomains of the CHASE domain of the sensor module; TM1 and TM2: Transmembrane domain 1 and 2; HisKA (DHpD): His kinase A domain (dimerization and His phosphotransfer domain); H-ATPase (CAD): Adenosine triphosphatase domain (catalytic and ATP-binding domain); REC-like: Receiver-like domain; REC: Receiver domain; HPt: His-containing phosphotransfer protein (phosphotransmitter); RR-B: Type B response regulator (transcription factor). Protein-protein interactions (PPI) are indicated by a red dotted line. ...
... 组氨酸激酶包含1个保守的与CK结合的胞外CHASE (cyclases/His kinases-associated sensing extracellular)区域、至少2个跨膜(transmembrane, TM)结构域和1个细胞质区域(图2) (Hwang et al.2012; Lomin et al.2018; Arkhipov et al.2019).HK主要定位于ER膜(Wulfetange et al.2011).CHASE区域即感受器模块(sensor module, SM), 面向ER腔或者质外体(apoplast), 包括二聚界面(dimerization interface, DI)、PAS (Per-Arnt-Sim)结构域和类PAS结构域(Hothorn et al.2011; Steklov et al.2013; Romanov et al.2018).TM区域由2-5个α-螺旋延伸组成.在马铃薯(S. tuberosum) CK受体的N端, StHK2、StHK3和StHK4的上游TM螺旋的数量分别为3、2和1个(Steklov et al.2013).HK的细胞质部分包括催化模块(catalytic module)和接收器模块(receiver module).催化模块由HK-二聚体(HisKA或者DHpD)和H-ATPase (CAD)结构域组成(Pekárováet al.2016); 接收器模块含有类接收器结构域(receiver-like domain, REC-like)和接收器结构域(receiver domain, REC), 它们定位于HK蛋白的C-端(图2) (Lomin et al.2018; Pekarova et al.2018; Arkhipov et al.2019).值得注意的是, AHK4同时具有组氨酸激酶和磷酸酶活性; 在配体缺乏时, AHK4的磷酸酶活性显著高于激酶活性, 能水解其接收器结构域上的磷酸基, 从而消耗可传递的磷酸基团(M?h?nenet al.2006). ...
... 2018; Pekarova et al.2018; Arkhipov et al.2019).值得注意的是, AHK4同时具有组氨酸激酶和磷酸酶活性; 在配体缺乏时, AHK4的磷酸酶活性显著高于激酶活性, 能水解其接收器结构域上的磷酸基, 从而消耗可传递的磷酸基团(M?h?nenet al.2006). ...
... 水稻、拟南芥和马铃薯CK受体序列的比对分析表明, 马铃薯同源基因中存在已知CK受体中所有的典型基序(Lomin et al.2018).在马铃薯催化模块中已鉴定了H、N、G1、F和G2基序, 在接收器模块中鉴定了DD-D-K基序.保守序列含有可磷酸化的His和天冬氨酸(Asp)残基.StHK2在其类接收器结构域中含有1个保守的Asp, 类似于拟南芥AHK2、番茄SlHK2及水稻OsHK3和OsHK5的同源基因.然而, 类接收器结构域中的全部类DD-D-K基序与接收器结构域中的相应序列几乎无相同点(Lomin et al.2018). ...
... 2018). ...
... 磷酸传递的信号转导、蛋白互作和高阶(higher- order)功能缺失突变的分析表明, AHP在功能上冗余且可以不加选择地与受体相互作用, 磷酸基的转移是双向的(Deng et al.2010).在6个AHP (AHP1-6)中, AHP6缺乏保守的His残基, 因此不能接受磷酸基, 被称为伪AHP (pseudo-AHP).AHP6负调控信号转导途径活性, 很可能与AHP1-5竞争与活化受体的相互作用(Lomin et al.2018). ...
Role of auxin in maize endosperm development (timing of nuclear DNA endoreduplication, zein expression, and cytokinin) 1 1993
Expression of cytokinin biosynthetic isopentenyltransferase genes in Arabidopsis: tissue specificity and regulation by auxin, cytokinin, and nitrate 1 2004
Topolins and hydroxylated thidiazuron derivatives are substrates of cytokinin O-glucosyltransferase with position specificity related to receptor recognition 2 2005
... 芳香族CK (如6-BA和topolin)已经在一些植物中被鉴定, 包括杨树(Populus) (Strnad, 1997)和拟南芥(Tarkowska et al.2003), 但它们是否普遍存在于植物中还不清楚.尽管它们表现出较强的CK活性, 但其生物合成和降解的途径尚不明确.芳香族CK与类异戊二烯CK所涉及的酶和受体都能识别这2种类型的CK成员, 因此二者的糖基化及其与细胞信号转导系统相互作用的机制可能相同(Mok et al.2005).在番茄(Solanum lycopersicum)果皮组织中, 6-BA的修饰被一种P450酶的有效抑制剂美替拉酮(metyrapone)强烈抑制(Sakakibara, 2006).虽然具体作用方式尚不清楚, 但CYP735A或者其它的P450可能参与topolin的生物合成(Sakakibara, 2006). ...
... 细胞分裂素的葡糖基结合物在生物测定中无生物活性, 这些被结合的CK不能与CK受体结合(Spichal et al.2004).尽管O-葡糖苷和N-葡糖苷稳定性差异的生理意义至今还不清楚, 但易裂解的O-葡糖苷被认为是CK失活的稳定贮藏形式.虽然与CK O-葡糖基化有关的基因已被鉴定(Mok et al.2005), 但尚缺乏关于N-葡糖基化的分子生物学信息.在拟南芥中, UGT76C1和UGT76C2在N7或N9位置葡糖基化一系列CK碱基, 也在N7位置葡糖基化tZ, 生成tZ-O-葡糖苷(tZ-O-glucoside) (Hou et al.2004). ...
Cytokinin and auxin interaction in root stem-cell specification during early embryogenesis 2 2008
... 基因的表达.上述研究为胚胎早期发生过程中可能的CK信号转导提供了最初的证据(Müller and Sheen,2008). ...
Cytokinin activity during early kernel development corresponds positively with yield potential and later stage ABA accumulation in field-grown wheat ( Triticum aestivum L.) 5 2020
... 种子成熟过程中在母体植株上获得的休眠类型称为初生休眠(primary dormancy), 种子初生休眠的诱导和维持受植物激素调控(Shu et al.2016; Nonogaki, 2017, 2019; 宋松泉等, 2020a).研究表明, 种子中CK的水平和信号转导调控种子的发育、休眠与萌发(图1, 图3) (Kieber and Schaller, 2018; Tuan et al.2019; Nguyen et al.2020).后熟(after-ripening)是一种释放种子休眠的方式, 也通过影响CK的水平和信号转导来调控种子的休眠与萌发(Chitnis et al.2014). ...
... 种子成熟过程中在母体植株上获得的休眠类型称为初生休眠(primary dormancy), 种子初生休眠的诱导和维持受植物激素调控(Shu et al.2016; Nonogaki, 2017, 2019; 宋松泉等, 2020a).研究表明, 种子中CK的水平和信号转导调控种子的发育、休眠与萌发(图1, 图3) (Kieber and Schaller, 2018; Tuan et al.2019; Nguyen et al.2020).后熟(after-ripening)是一种释放种子休眠的方式, 也通过影响CK的水平和信号转导来调控种子的休眠与萌发(Chitnis et al.2014). ...
Structural aspects of multistep phosphorelay-mediated signaling in plants 1 2016
... 组氨酸激酶包含1个保守的与CK结合的胞外CHASE (cyclases/His kinases-associated sensing extracellular)区域、至少2个跨膜(transmembrane, TM)结构域和1个细胞质区域(图2) (Hwang et al.2012; Lomin et al.2018; Arkhipov et al.2019).HK主要定位于ER膜(Wulfetange et al.2011).CHASE区域即感受器模块(sensor module, SM), 面向ER腔或者质外体(apoplast), 包括二聚界面(dimerization interface, DI)、PAS (Per-Arnt-Sim)结构域和类PAS结构域(Hothorn et al.2011; Steklov et al.2013; Romanov et al.2018).TM区域由2-5个α-螺旋延伸组成.在马铃薯(S. tuberosum) CK受体的N端, StHK2、StHK3和StHK4的上游TM螺旋的数量分别为3、2和1个(Steklov et al.2013).HK的细胞质部分包括催化模块(catalytic module)和接收器模块(receiver module).催化模块由HK-二聚体(HisKA或者DHpD)和H-ATPase (CAD)结构域组成(Pekárováet al.2016); 接收器模块含有类接收器结构域(receiver-like domain, REC-like)和接收器结构域(receiver domain, REC), 它们定位于HK蛋白的C-端(图2) (Lomin et al.2018; Pekarova et al.2018; Arkhipov et al.2019).值得注意的是, AHK4同时具有组氨酸激酶和磷酸酶活性; 在配体缺乏时, AHK4的磷酸酶活性显著高于激酶活性, 能水解其接收器结构域上的磷酸基, 从而消耗可传递的磷酸基团(M?h?nenet al.2006). ...
Cytokinin and ethylene signaling 2 2018
... 组氨酸激酶包含1个保守的与CK结合的胞外CHASE (cyclases/His kinases-associated sensing extracellular)区域、至少2个跨膜(transmembrane, TM)结构域和1个细胞质区域(图2) (Hwang et al.2012; Lomin et al.2018; Arkhipov et al.2019).HK主要定位于ER膜(Wulfetange et al.2011).CHASE区域即感受器模块(sensor module, SM), 面向ER腔或者质外体(apoplast), 包括二聚界面(dimerization interface, DI)、PAS (Per-Arnt-Sim)结构域和类PAS结构域(Hothorn et al.2011; Steklov et al.2013; Romanov et al.2018).TM区域由2-5个α-螺旋延伸组成.在马铃薯(S. tuberosum) CK受体的N端, StHK2、StHK3和StHK4的上游TM螺旋的数量分别为3、2和1个(Steklov et al.2013).HK的细胞质部分包括催化模块(catalytic module)和接收器模块(receiver module).催化模块由HK-二聚体(HisKA或者DHpD)和H-ATPase (CAD)结构域组成(Pekárováet al.2016); 接收器模块含有类接收器结构域(receiver-like domain, REC-like)和接收器结构域(receiver domain, REC), 它们定位于HK蛋白的C-端(图2) (Lomin et al.2018; Pekarova et al.2018; Arkhipov et al.2019).值得注意的是, AHK4同时具有组氨酸激酶和磷酸酶活性; 在配体缺乏时, AHK4的磷酸酶活性显著高于激酶活性, 能水解其接收器结构域上的磷酸基, 从而消耗可传递的磷酸基团(M?h?nenet al.2006). ...
... 细胞分裂素与HK的PAS结合, 导致受体的二聚作用并被激活, CK的信号转导通过类似于原核生物的His-Asp磷酸传递途径进行(Keshishian and Rashotte, 2015; Zschiedrich et al.2016).在激活的受体ATP结合结构域中, ATP发生水解, 释放的磷酸基团与HisKA结构域中1个保守的His残基结合, 形成磷酸组氨酸(phosphohistidine).然后, 活化(hot)的磷酸基在分子内被转移到接收器结构域中1个保守的Asp残基上, 依次与HPt相互作用, 将活化的磷酸基转移到HPt保守的His残基上.HPt携带该活化的磷酸基并转移到细胞核和磷酸化RR蛋白保守的Asp残基.最后, 磷酸化的B型RR能够与DNA结合, 激活CK靶基因的转录(图1, 图2) (Kieber and Schaller, 2018; Pekarova et al.2018; Arkhipov et al.2019). ...
Genome-wide comparative analysis of type-A Arabidopsis response regulator genes by overexpression studies reveals their diverse roles and regulatory mechanisms in cytokinin signaling 1 2009
... ABI5是一种碱性亮氨酸拉链转录因子(basic leucine zipper transcription factor), 在ABA介导的抑制种子萌发和萌发后生长的调控中起重要作用(Guan et al.2014; 宋松泉等, 2020b).在种子萌发过程中, ABI5功能丧失能够降低ABA的敏感性, 而ABI5功能获得能增加ABA的敏感性(Liu and Stone, 2010).研究表明, ABI5与ARR4-6发生物理相互作用(Wang et al.2011).在arr3/arr4/arr5/arr6四突变体中, CK诱导的ABI5降低减少, 这表明A型ARR蛋白与ABI5的结合可能在调控ABI5蛋白的稳定性中起重要作用(Guan et al.2014).由于A型ARR mRNA的稳定水平和A型ARR蛋白的积累被CK正调控(Ren et al.2009), 因此, A型ARR-ABI5复合物可能抑制ABI5蛋白与蛋白酶体降解复合物的相互作用. ...
Cytokinins and their function in developing seeds 1 2010
... Day等(2008)利用激光捕获显微切割(laser capture microdissection, LCM)技术对授粉后4天的种子进行研究, 在合胞体胚乳(syncytial endosperm)中发现与CK生物合成和信号转导相关的基因高丰度表达, 尤其是AtIPT8在发育中的胚乳合点区表达丰度更高.Belmonte等(2013)研究了拟南芥种子发育过程中亚区域基因的表达, 发现在种子发育的形态发生阶段IPT4和IPT8的高丰度表达局限于合点胚乳(chalazal endosperm).这些结果表明发育中的种子是CK生物合成的场所.小麦基因芯片(GeneChip)研究及其随后所选择基因的qRT-PCR分析表明, AC domain (休眠)和RL4452 (非休眠) 2种基因型种子的成熟都与胚和胚乳组织中注释为TaIPT2-2探针(基因)的增强表达有关; 在这2种基因型的种子成熟过程中, TaCKX4-3和TaCKX7-2探针在胚中的表达水平下降, 在胚乳中维持相似的水平(Tuan et al.2019).IPT编码催化CK生物合成限速步骤的酶, 且CKX催化CK的失活(Sakakibara, 2006; Frébortet al.2011), 这些结果表明, 种子成熟与胚和胚乳中CK水平的升高有关.相反, 随着种子成熟, 胚和胚乳组织中的iP水平显著降低, 但与基因型无关; 而tZ仅在2种基因型的胚组织中被检测到, 且在AC Domain基因型中的含量比在RL4452基因型中更低(Tuan et al.2019).在2种基因型种子成熟过程中, 胚和胚乳中iP水平的下降与胚TaLOG8和胚乳TaLOG3基因的表达水平下降有关(Tuan et al.2019).这些基因编码CK激活酶(LOG), 该酶直接将失活的CK核苷酸转化为生物活性的游离态CK (Kurakawa et al.2007).此外, 种子成熟时2种基因型的胚和胚乳组织中iP水平的下降以及AC Domain胚中tZ水平的显著下降与其相应组织中注释为TaGLU4探针的表达水平下降一致(Tuan et al.2019).TaGLU4编码一种通过羟基化作用从CK结合物中释放生物活性CK的酶(Cairns and Esen, 2010). ...
Agrobacterium tumefaciens increases cytokinin production in plastids by modifying the biosynthetic pathway in the host plant 1 2005
... 目前已在拟南芥(Takei et al.2001)、矮牵牛(Petunia hybrida) (Zubko et al.2002)和啤酒花(Humulus lupulus) (Sakano et al.2004)中鉴定了IPT基因.在拟南芥中, 7个IPT基因(AtIPT1和AtIPT3-8)参与CK的生物合成(Takei et al.2001).生化研究表明, 植物IPT优先利用ADP或ATP而不是AMP作为异戊二烯基的受体, 产生iP-核苷5′-二磷酸(iP riboside 5′-diphosphate, iPRDP)或iP-核苷5′-三磷酸(iP riboside 5'-triphosphate, iPRTP) (Sakanoet al.2004; Ho?eket al.2020).AtIPT1对AMP、ADP和ATP的Km值分别为18、14.6和11.4 μmol·L -1, 对DMAPP的Km值为8.3 μmol·L -1 (Takei et al.2001).AtIPT4对ADP、ATP和DMAPP的Km值分别为9.1、3.4和11.6 μmol·L -1(Kakimoto, 2001).在过表达任一AtIPT的拟南芥转基因株系中, iP型CK显著积累, 表明所有的AtIPT对于类异戊二烯供体都具有相似的底物偏好(Sakakibara et al.2005). ...
Molecular cloning, expression, and characterization of adenylate isopentenyltransferase from hop ( Humulus lupulus L.) 2 2004
... 目前已在拟南芥(Takei et al.2001)、矮牵牛(Petunia hybrida) (Zubko et al.2002)和啤酒花(Humulus lupulus) (Sakano et al.2004)中鉴定了IPT基因.在拟南芥中, 7个IPT基因(AtIPT1和AtIPT3-8)参与CK的生物合成(Takei et al.2001).生化研究表明, 植物IPT优先利用ADP或ATP而不是AMP作为异戊二烯基的受体, 产生iP-核苷5′-二磷酸(iP riboside 5′-diphosphate, iPRDP)或iP-核苷5′-三磷酸(iP riboside 5'-triphosphate, iPRTP) (Sakanoet al.2004; Ho?eket al.2020).AtIPT1对AMP、ADP和ATP的Km值分别为18、14.6和11.4 μmol·L -1, 对DMAPP的Km值为8.3 μmol·L -1 (Takei et al.2001).AtIPT4对ADP、ATP和DMAPP的Km值分别为9.1、3.4和11.6 μmol·L -1(Kakimoto, 2001).在过表达任一AtIPT的拟南芥转基因株系中, iP型CK显著积累, 表明所有的AtIPT对于类异戊二烯供体都具有相似的底物偏好(Sakakibara et al.2005). ...
... 2004; Ho?eket al.2020).AtIPT1对AMP、ADP和ATP的Km值分别为18、14.6和11.4 μmol·L -1, 对DMAPP的Km值为8.3 μmol·L -1 (Takei et al.2001).AtIPT4对ADP、ATP和DMAPP的Km值分别为9.1、3.4和11.6 μmol·L -1(Kakimoto, 2001).在过表达任一AtIPT的拟南芥转基因株系中, iP型CK显著积累, 表明所有的AtIPT对于类异戊二烯供体都具有相似的底物偏好(Sakakibara et al.2005). ...
Nomenclature for two-component signaling elements of rice 1 2007
... 反应调控因子是一个转录因子家族, 包含1个保守的接受磷酸基的Asp残基(Keshishian and Rashotte, 2015).根据其系统发生和结构域, RR家族成员可分为3种类型: A型、B型和C型(Schaller et al.2007), 但只有A型和B型RR与CK信号转导有关(Kieber and Schaller, 2018; Tuan et al.2019; Zubo and Schaller, 2020).B型RR包括ARR (Arabidopsis RR) 1、ARR2、ARR10-14和ARR18-21, 能被HP接收器结构域中的Asp残基磷酸化激活, 作为CK反应的正调控因子起作用(Hwang et al.2012).B型RR的特征是包含1个接收器结构域和1个含有类MYB DNA结合结构域的长C-端延伸.接收器结构域包含1个保守的Asp残基, 当多步骤磷酸传递被激活时, Asp可作为调控磷酸化的位点.类MYB DNA结合结构域与1个短的(A/G) GAT核心DNA序列结合, 该序列对B型RR的结合作用非常关键(Zubo et al.2017; Zubo and Schaller, 2020).ChIP-seq (chromatin immunoprecipitation combined with sequencing)实验显示, B型RR在植物体内以CK依赖的方式与其靶点结合, 这种结合以类似于细菌RR的方式被接收器结构域磷酸化激活(Zubo et al.2017). ...
Two faces of one seed: hormonal regulation of dormancy and germination 2 2016
... 2001).AtIPT4对ADP、ATP和DMAPP的Km值分别为9.1、3.4和11.6 μmol·L -1(Kakimoto, 2001).在过表达任一AtIPT的拟南芥转基因株系中, iP型CK显著积累, 表明所有的AtIPT对于类异戊二烯供体都具有相似的底物偏好(Sakakibara et al.2005). ...
ARR1 directly activates cytokinin response genes that encode proteins with diverse regulatory functions 1 2007
... A型ARR (如ARR3-9、ARR15-17)由1个接收器结构域和1个短的C-端延伸组成, 作为CK信号转导的负反馈调控因子起作用; 但与B型RR不同, 它们缺乏典型的转录调控输出结构域(output domain) (Kieber and Schaller, 2018).在对CK的反应中, A型RR通过B型RR的直接激活被诱导转录(Taniguchi et al.2007), CK也以磷酸化依赖的方式稳定许多A型RR (To et al.2007).A型ARR负调控CK信号转导的机制是与B型ARR竞争AHP的磷酸化, 从而导致多步骤磷酸传递与B型ARR转录调控因子解离, 以减少其对基因表达的影响(Kieber and Schaller, 2014; Zubo and Schaller, 2020). ...
Identification of new aromatic cytokinins in Arabidopsis thaliana and Populus × canadensis leaves by LC- (+) ESI-MS and capillary liquid chromatography/frit- fast atom bombardment mass spectrometry 1 2003
... 芳香族CK (如6-BA和topolin)已经在一些植物中被鉴定, 包括杨树(Populus) (Strnad, 1997)和拟南芥(Tarkowska et al.2003), 但它们是否普遍存在于植物中还不清楚.尽管它们表现出较强的CK活性, 但其生物合成和降解的途径尚不明确.芳香族CK与类异戊二烯CK所涉及的酶和受体都能识别这2种类型的CK成员, 因此二者的糖基化及其与细胞信号转导系统相互作用的机制可能相同(Mok et al.2005).在番茄(Solanum lycopersicum)果皮组织中, 6-BA的修饰被一种P450酶的有效抑制剂美替拉酮(metyrapone)强烈抑制(Sakakibara, 2006).虽然具体作用方式尚不清楚, 但CYP735A或者其它的P450可能参与topolin的生物合成(Sakakibara, 2006). ...
Cytokinin regulates type-A Arabidopsis response regulator activity and protein stability via two-component phosphorelay 1 2007
... A型ARR (如ARR3-9、ARR15-17)由1个接收器结构域和1个短的C-端延伸组成, 作为CK信号转导的负反馈调控因子起作用; 但与B型RR不同, 它们缺乏典型的转录调控输出结构域(output domain) (Kieber and Schaller, 2018).在对CK的反应中, A型RR通过B型RR的直接激活被诱导转录(Taniguchi et al.2007), CK也以磷酸化依赖的方式稳定许多A型RR (To et al.2007).A型ARR负调控CK信号转导的机制是与B型ARR竞争AHP的磷酸化, 从而导致多步骤磷酸传递与B型ARR转录调控因子解离, 以减少其对基因表达的影响(Kieber and Schaller, 2014; Zubo and Schaller, 2020). ...
Transcriptomics of cytokinin and auxin metabolism and signaling genes during seed maturation in dormant and non-dormant wheat genotypes 12 2019
... 细胞分裂素氧化酶/脱氢酶(cytokinin oxidase/dehydrogenase, CKX)在N6侧链上不可逆地裂解游离态CK和CK核苷, 以降低生物活性CK的水平(图1) (Sakakibara, 2006; Tuan et al.2019; Chen et al.2020).研究发现, CKX的过表达导致内源CK水平降低和多种发育缺陷; tZ和iP都能被CKX裂解, 但DZ和人工合成的CK (如激动素(kinetin)和6-BA)对CKX的裂解具有抗性(Zalabáket al.2014). ...
... 反应调控因子是一个转录因子家族, 包含1个保守的接受磷酸基的Asp残基(Keshishian and Rashotte, 2015).根据其系统发生和结构域, RR家族成员可分为3种类型: A型、B型和C型(Schaller et al.2007), 但只有A型和B型RR与CK信号转导有关(Kieber and Schaller, 2018; Tuan et al.2019; Zubo and Schaller, 2020).B型RR包括ARR (Arabidopsis RR) 1、ARR2、ARR10-14和ARR18-21, 能被HP接收器结构域中的Asp残基磷酸化激活, 作为CK反应的正调控因子起作用(Hwang et al.2012).B型RR的特征是包含1个接收器结构域和1个含有类MYB DNA结合结构域的长C-端延伸.接收器结构域包含1个保守的Asp残基, 当多步骤磷酸传递被激活时, Asp可作为调控磷酸化的位点.类MYB DNA结合结构域与1个短的(A/G) GAT核心DNA序列结合, 该序列对B型RR的结合作用非常关键(Zubo et al.2017; Zubo and Schaller, 2020).ChIP-seq (chromatin immunoprecipitation combined with sequencing)实验显示, B型RR在植物体内以CK依赖的方式与其靶点结合, 这种结合以类似于细菌RR的方式被接收器结构域磷酸化激活(Zubo et al.2017). ...
... 种子成熟过程中在母体植株上获得的休眠类型称为初生休眠(primary dormancy), 种子初生休眠的诱导和维持受植物激素调控(Shu et al.2016; Nonogaki, 2017, 2019; 宋松泉等, 2020a).研究表明, 种子中CK的水平和信号转导调控种子的发育、休眠与萌发(图1, 图3) (Kieber and Schaller, 2018; Tuan et al.2019; Nguyen et al.2020).后熟(after-ripening)是一种释放种子休眠的方式, 也通过影响CK的水平和信号转导来调控种子的休眠与萌发(Chitnis et al.2014). ...
... IPT: 异戊烯基转移酶; LOG: CK核苷5′-单磷酸磷酸核糖水解酶; CKX: CK氧化酶/脱氢酶; HK: 组氨酸激酶; HP: 组氨酸磷酸转移蛋白; Type-B RRs: 类型-B反应调控因子; Type-A RRs: 类型-A反应调控因子 The model of the change of proteins (enzymes) involved in cytokinin (CK) metabolism and signaling components during seed development and germination (modified from Jameson <i>et al.</i> <xref ref-type="bibr" rid="b28">2016</xref>; Tuan <i>et al.</i> <xref ref-type="bibr" rid="b71">2019</xref>; Nguyen <i>et al.</i> <xref ref-type="bibr" rid="b49">2020</xref>) IPT: Isopentenyltransferase; LOG: CK nucleoside 5′-monophosphate phosphoribohydrolase; CKX: CK oxidase/ dehydrogenase; HK: Histidin (His) kinase; HP: His phosphotransfer protein; Type-B RRs: Type-B response regulators; Type-A RRs: Type-A response regulators ...
... Day等(2008)利用激光捕获显微切割(laser capture microdissection, LCM)技术对授粉后4天的种子进行研究, 在合胞体胚乳(syncytial endosperm)中发现与CK生物合成和信号转导相关的基因高丰度表达, 尤其是AtIPT8在发育中的胚乳合点区表达丰度更高.Belmonte等(2013)研究了拟南芥种子发育过程中亚区域基因的表达, 发现在种子发育的形态发生阶段IPT4和IPT8的高丰度表达局限于合点胚乳(chalazal endosperm).这些结果表明发育中的种子是CK生物合成的场所.小麦基因芯片(GeneChip)研究及其随后所选择基因的qRT-PCR分析表明, AC domain (休眠)和RL4452 (非休眠) 2种基因型种子的成熟都与胚和胚乳组织中注释为TaIPT2-2探针(基因)的增强表达有关; 在这2种基因型的种子成熟过程中, TaCKX4-3和TaCKX7-2探针在胚中的表达水平下降, 在胚乳中维持相似的水平(Tuan et al.2019).IPT编码催化CK生物合成限速步骤的酶, 且CKX催化CK的失活(Sakakibara, 2006; Frébortet al.2011), 这些结果表明, 种子成熟与胚和胚乳中CK水平的升高有关.相反, 随着种子成熟, 胚和胚乳组织中的iP水平显著降低, 但与基因型无关; 而tZ仅在2种基因型的胚组织中被检测到, 且在AC Domain基因型中的含量比在RL4452基因型中更低(Tuan et al.2019).在2种基因型种子成熟过程中, 胚和胚乳中iP水平的下降与胚TaLOG8和胚乳TaLOG3基因的表达水平下降有关(Tuan et al.2019).这些基因编码CK激活酶(LOG), 该酶直接将失活的CK核苷酸转化为生物活性的游离态CK (Kurakawa et al.2007).此外, 种子成熟时2种基因型的胚和胚乳组织中iP水平的下降以及AC Domain胚中tZ水平的显著下降与其相应组织中注释为TaGLU4探针的表达水平下降一致(Tuan et al.2019).TaGLU4编码一种通过羟基化作用从CK结合物中释放生物活性CK的酶(Cairns and Esen, 2010). ...
... 2019).在2种基因型种子成熟过程中, 胚和胚乳中iP水平的下降与胚TaLOG8和胚乳TaLOG3基因的表达水平下降有关(Tuan et al.2019).这些基因编码CK激活酶(LOG), 该酶直接将失活的CK核苷酸转化为生物活性的游离态CK (Kurakawa et al.2007).此外, 种子成熟时2种基因型的胚和胚乳组织中iP水平的下降以及AC Domain胚中tZ水平的显著下降与其相应组织中注释为TaGLU4探针的表达水平下降一致(Tuan et al.2019).TaGLU4编码一种通过羟基化作用从CK结合物中释放生物活性CK的酶(Cairns and Esen, 2010). ...
... 2019).这些基因编码CK激活酶(LOG), 该酶直接将失活的CK核苷酸转化为生物活性的游离态CK (Kurakawa et al.2007).此外, 种子成熟时2种基因型的胚和胚乳组织中iP水平的下降以及AC Domain胚中tZ水平的显著下降与其相应组织中注释为TaGLU4探针的表达水平下降一致(Tuan et al.2019).TaGLU4编码一种通过羟基化作用从CK结合物中释放生物活性CK的酶(Cairns and Esen, 2010). ...
... 2019).TaGLU4编码一种通过羟基化作用从CK结合物中释放生物活性CK的酶(Cairns and Esen, 2010). ...
... 在小麦种子成熟过程中, TaAHK4、TaARR9和TaARR12的表达水平在AC Domain和RL4452的胚中均表现下降, 这与iP和tZ的积累下降有关.然而, 在开花后20天, AC Domain胚中TaARR12的表达水平比RL4452更低(Tuan et al.2019).鉴于TaARR12属于B型ARR, 且作为CK反应的正调控因子起作用(Kieber and Schaller, 2018), TaARR12很可能抑制AC Domain胚中的CK反应, 从而在小麦种子成熟过程中起诱导休眠的作用.2种基因型胚乳中TaAHP2探针的表达随着种子成熟而下降.然而, 在种子成熟中期(开花后30天), TaAHK4探针在AC Domain胚乳中的表达水平比RL4452更低(Tuan et al.2019).由于AHK是CK的受体(Keshishian and Rashotte, 2015), 因此上述结果表明, RL4452对CK的反应敏感性增加, 从而抑制种子的休眠.另一方面, 在种子成熟的早期到中期(开花后20-30天), AC Domain胚乳中TaARR9的表达水平比RL4452更高(Tuan et al.2019).由于TaARR9属于负调控CK信号转导的A型ARR (Kieber and Schaller, 2018), 在成熟时具有深休眠的AC Domain种子胚乳对CK的反应敏感性降低, 其结果是成熟时休眠程度高.此外, AC Domain胚乳中AHK探针的表达下降也可能表明CK反应受到抑制, 从而诱导休眠. ...
... 2019).由于AHK是CK的受体(Keshishian and Rashotte, 2015), 因此上述结果表明, RL4452对CK的反应敏感性增加, 从而抑制种子的休眠.另一方面, 在种子成熟的早期到中期(开花后20-30天), AC Domain胚乳中TaARR9的表达水平比RL4452更高(Tuan et al.2019).由于TaARR9属于负调控CK信号转导的A型ARR (Kieber and Schaller, 2018), 在成熟时具有深休眠的AC Domain种子胚乳对CK的反应敏感性降低, 其结果是成熟时休眠程度高.此外, AC Domain胚乳中AHK探针的表达下降也可能表明CK反应受到抑制, 从而诱导休眠. ...
... 2019).由于TaARR9属于负调控CK信号转导的A型ARR (Kieber and Schaller, 2018), 在成熟时具有深休眠的AC Domain种子胚乳对CK的反应敏感性降低, 其结果是成熟时休眠程度高.此外, AC Domain胚乳中AHK探针的表达下降也可能表明CK反应受到抑制, 从而诱导休眠. ...
Cytokinin antagonizes ABA suppression to seed germination of Arabidopsis by downregulating ABI5 expression 5 2011
... ABA抑制种子萌发, CK能逆转ABA对莴苣(Lactuca sativa)种子萌发的抑制作用(Kham, 1968).1 mmol·L-1 ABA对黑黄檀(Dalbergia fusca)种子萌发的抑制作用能够被0.001-0.1 mmol·L-1 6-BA所拮抗(邓志军和宋松泉, 2008).过表达AtIPT8能降低gim1 (germination insensitive to ABA mutant 1)突变体种子萌发对ABA的敏感性; AtIPT8的功能丧失导致ipt8-1-ko和ipt8-2-ko突变体产生ABA过敏性反应; 在gim1中, AtIPT8的过表达能够通过促进CK的生物合成来增强CK信号, 从而上调CK信号转导的转录调控因子(如A型ARR4-6) (Wang et al.2011).在野生型拟南芥Columbia-0背景下, ABA处理能够通过抑制AtIPT3和AtIPT8的表达水平减少CK的内源生物合成, 随后降低A型ARR4-6的表达水平(Wang et al.2011).此外, 在萌发10天的gim1幼苗中, ABA处理也会抑制AtIPT3和AtIPT8的表达, 降低A型ARR4-6的表达丰度(Wang et al.2011).值得注意的是, gim1突变体中CK水平的增高不是由于ABA生物合成的改变, 而是降低了ABA和胁迫反应基因(如ABI5(ABSCISIC ACID INSENSITIVE 5))的表达水平(Wang et al.2011). ...
... 2011).此外, 在萌发10天的gim1幼苗中, ABA处理也会抑制AtIPT3和AtIPT8的表达, 降低A型ARR4-6的表达丰度(Wang et al.2011).值得注意的是, gim1突变体中CK水平的增高不是由于ABA生物合成的改变, 而是降低了ABA和胁迫反应基因(如ABI5(ABSCISIC ACID INSENSITIVE 5))的表达水平(Wang et al.2011). ...
... 2011).值得注意的是, gim1突变体中CK水平的增高不是由于ABA生物合成的改变, 而是降低了ABA和胁迫反应基因(如ABI5(ABSCISIC ACID INSENSITIVE 5))的表达水平(Wang et al.2011). ...
... 2011). ...
... ABI5是一种碱性亮氨酸拉链转录因子(basic leucine zipper transcription factor), 在ABA介导的抑制种子萌发和萌发后生长的调控中起重要作用(Guan et al.2014; 宋松泉等, 2020b).在种子萌发过程中, ABI5功能丧失能够降低ABA的敏感性, 而ABI5功能获得能增加ABA的敏感性(Liu and Stone, 2010).研究表明, ABI5与ARR4-6发生物理相互作用(Wang et al.2011).在arr3/arr4/arr5/arr6四突变体中, CK诱导的ABI5降低减少, 这表明A型ARR蛋白与ABI5的结合可能在调控ABI5蛋白的稳定性中起重要作用(Guan et al.2014).由于A型ARR mRNA的稳定水平和A型ARR蛋白的积累被CK正调控(Ren et al.2009), 因此, A型ARR-ABI5复合物可能抑制ABI5蛋白与蛋白酶体降解复合物的相互作用. ...
The cytokinin receptors of Arabidopsis are located mainly to the endoplasmic reticulum 1 2011
... 组氨酸激酶包含1个保守的与CK结合的胞外CHASE (cyclases/His kinases-associated sensing extracellular)区域、至少2个跨膜(transmembrane, TM)结构域和1个细胞质区域(图2) (Hwang et al.2012; Lomin et al.2018; Arkhipov et al.2019).HK主要定位于ER膜(Wulfetange et al.2011).CHASE区域即感受器模块(sensor module, SM), 面向ER腔或者质外体(apoplast), 包括二聚界面(dimerization interface, DI)、PAS (Per-Arnt-Sim)结构域和类PAS结构域(Hothorn et al.2011; Steklov et al.2013; Romanov et al.2018).TM区域由2-5个α-螺旋延伸组成.在马铃薯(S. tuberosum) CK受体的N端, StHK2、StHK3和StHK4的上游TM螺旋的数量分别为3、2和1个(Steklov et al.2013).HK的细胞质部分包括催化模块(catalytic module)和接收器模块(receiver module).催化模块由HK-二聚体(HisKA或者DHpD)和H-ATPase (CAD)结构域组成(Pekárováet al.2016); 接收器模块含有类接收器结构域(receiver-like domain, REC-like)和接收器结构域(receiver domain, REC), 它们定位于HK蛋白的C-端(图2) (Lomin et al.2018; Pekarova et al.2018; Arkhipov et al.2019).值得注意的是, AHK4同时具有组氨酸激酶和磷酸酶活性; 在配体缺乏时, AHK4的磷酸酶活性显著高于激酶活性, 能水解其接收器结构域上的磷酸基, 从而消耗可传递的磷酸基团(M?h?nenet al.2006). ...
Cytokinin—a developing story 1 2019
... 细胞分裂素是一种重要的植物激素, 在调控植物生长发育的许多过程中起关键作用, 包括细胞的分裂与分化、茎和根的生长、顶端优势维持、衰老、果实和种子发育、营养信号传递以及对生物和非生物胁迫的响应(Hwang et al.2012; Jameson and Song, 2016; Wybouw and De Rybel, 2019).近年来, CK的生物合成与代谢, 尤其是CK信号转导途径的研究取得了重要进展(Lomin et al.2018; Arkhipov et al.2019; Hallmark and Rashotte, 2019; Chen et al.2020).本文在综述CK的生物合成与代谢及其信号转导的基础上, 重点介绍CK调控种子发育、休眠与萌发的研究进展, 并提出了在本领域需要进一步研究的科学问题, 旨在为阐明CK调控种子发育、休眠与萌发的分子机理提供参考. ...
Biochemical characterization of the maize cytokinin dehydrogenase family and cytokinin profiling in developing maize plantlets in relation to the expression of cytokinin dehydrogenase genes 1 2014
... 细胞分裂素氧化酶/脱氢酶(cytokinin oxidase/dehydrogenase, CKX)在N6侧链上不可逆地裂解游离态CK和CK核苷, 以降低生物活性CK的水平(图1) (Sakakibara, 2006; Tuan et al.2019; Chen et al.2020).研究发现, CKX的过表达导致内源CK水平降低和多种发育缺陷; tZ和iP都能被CKX裂解, 但DZ和人工合成的CK (如激动素(kinetin)和6-BA)对CKX的裂解具有抗性(Zalabáket al.2014). ...
Illuminating light, cytokinin, and ethylene signaling crosstalk in plant development 1 2015
... 研究表明, 种子的萌发能力与乙烯的产生有关, 外源乙烯或乙烯利(ethrel, 乙烯释放剂)能解除种子的初生和次生休眠(second dormancy) (Corbineau et al.2014).CK通过转录和转录后机制激活ACC合酶(ACC synthase, ACS), 促进乙烯的生物合成; 乙烯的产生促进被CK抑制的暗生长幼苗的下胚轴伸长以及抑制根系的生长(Zdarska et al.2015).此外, B型ARR与许多影响乙烯信号转导的基因结合.尽管乙烯受体ETR2被CK下调表达, 但CK调控大多数编码调节乙烯信号转导途径活性的元件, 而不是途径本身.调节原件包括ARGOS基因家族的ARGOS和ARL以及TRP1、ETP和EBF基因.ARGOS是在受体水平起作用的跨膜蛋白, 可使植物对乙烯去敏感; 作为乙烯信号转导的正调控因子, TRP1可能通过与受体结合和干扰与CTR1的互作起作用; ETP和EBF基因分别编码参与EIN2和EIN3降解的F-box蛋白(Zubo and Schaller, 2020). ...
Molecular mechanisms of two-component signal transduction 1 2016
... 细胞分裂素与HK的PAS结合, 导致受体的二聚作用并被激活, CK的信号转导通过类似于原核生物的His-Asp磷酸传递途径进行(Keshishian and Rashotte, 2015; Zschiedrich et al.2016).在激活的受体ATP结合结构域中, ATP发生水解, 释放的磷酸基团与HisKA结构域中1个保守的His残基结合, 形成磷酸组氨酸(phosphohistidine).然后, 活化(hot)的磷酸基在分子内被转移到接收器结构域中1个保守的Asp残基上, 依次与HPt相互作用, 将活化的磷酸基转移到HPt保守的His残基上.HPt携带该活化的磷酸基并转移到细胞核和磷酸化RR蛋白保守的Asp残基.最后, 磷酸化的B型RR能够与DNA结合, 激活CK靶基因的转录(图1, 图2) (Kieber and Schaller, 2018; Pekarova et al.2018; Arkhipov et al.2019). ...
Activation tagging identifies a gene from Petunia hybrida responsible for the production of active cytokinins in plants 1 2002
... 目前已在拟南芥(Takei et al.2001)、矮牵牛(Petunia hybrida) (Zubko et al.2002)和啤酒花(Humulus lupulus) (Sakano et al.2004)中鉴定了IPT基因.在拟南芥中, 7个IPT基因(AtIPT1和AtIPT3-8)参与CK的生物合成(Takei et al.2001).生化研究表明, 植物IPT优先利用ADP或ATP而不是AMP作为异戊二烯基的受体, 产生iP-核苷5′-二磷酸(iP riboside 5′-diphosphate, iPRDP)或iP-核苷5′-三磷酸(iP riboside 5'-triphosphate, iPRTP) (Sakanoet al.2004; Ho?eket al.2020).AtIPT1对AMP、ADP和ATP的Km值分别为18、14.6和11.4 μmol·L -1, 对DMAPP的Km值为8.3 μmol·L -1 (Takei et al.2001).AtIPT4对ADP、ATP和DMAPP的Km值分别为9.1、3.4和11.6 μmol·L -1(Kakimoto, 2001).在过表达任一AtIPT的拟南芥转基因株系中, iP型CK显著积累, 表明所有的AtIPT对于类异戊二烯供体都具有相似的底物偏好(Sakakibara et al.2005). ...
Cytokinin induces genome-wide binding of the type-B response regulator ARR10 to regulate growth and development in Arabidopsis 3 2017
... 反应调控因子是一个转录因子家族, 包含1个保守的接受磷酸基的Asp残基(Keshishian and Rashotte, 2015).根据其系统发生和结构域, RR家族成员可分为3种类型: A型、B型和C型(Schaller et al.2007), 但只有A型和B型RR与CK信号转导有关(Kieber and Schaller, 2018; Tuan et al.2019; Zubo and Schaller, 2020).B型RR包括ARR (Arabidopsis RR) 1、ARR2、ARR10-14和ARR18-21, 能被HP接收器结构域中的Asp残基磷酸化激活, 作为CK反应的正调控因子起作用(Hwang et al.2012).B型RR的特征是包含1个接收器结构域和1个含有类MYB DNA结合结构域的长C-端延伸.接收器结构域包含1个保守的Asp残基, 当多步骤磷酸传递被激活时, Asp可作为调控磷酸化的位点.类MYB DNA结合结构域与1个短的(A/G) GAT核心DNA序列结合, 该序列对B型RR的结合作用非常关键(Zubo et al.2017; Zubo and Schaller, 2020).ChIP-seq (chromatin immunoprecipitation combined with sequencing)实验显示, B型RR在植物体内以CK依赖的方式与其靶点结合, 这种结合以类似于细菌RR的方式被接收器结构域磷酸化激活(Zubo et al.2017). ...
... 2017). ...
... 此外, CK也与油菜素甾醇(brassinosteroid)、茉莉酸(jasmonic acid)和水杨酸(salicylic acid)产生广泛互作(crosstalk), B型ARR的结合位点与这些植物激素的生物合成、降解/失活、运输和信号转导的基因有关(Zubo et al.2017; Zubo and Schaller, 2020). ...
Role of the cytokinin-activated type-B response regulators in hormone crosstalk 12 2020
A model of cytokinin (CK) metabolism and signaling transduction (modified from Kieber and Schaller, <xref ref-type="bibr" rid="b35">2018</xref>; Zubo and Schaller, <xref ref-type="bibr" rid="b80">2020</xref>) Ade: Adenine; ADP: Adenosine diphosphate; AMP: Adenosine monophosphate; ATP: Adenosine triphosphate; CKX: CK oxidase/dehydrogenase; CYP735A: Cytochrome P450 monooxygenase; cZOG: cis-Zeatin-O-glucosyltransferase; DMAPP: Dimethylallyl diphosphate; HK: Histidin (His) kinase; HP: His phosphotransfer protein; IPT: Isopentenyltransferase; LOG: CK nucleoside 5′-monophosphate phosphoribohydrolase; SC: Sidechain; Type-B RRs: Type-B response regulators; Type-A RRs: Type-A response regulators; UGT: UDP glycosyltransferase ...
... 细胞分裂素信号转导途径也被称为双组分信号转导系统(two-component signaling system), 主要包括组氨酸激酶(histidine (His) kinase, HK)、组氨酸磷酸转移蛋白(His phosphotransfer protein, HP)和反应调控因子(response regulator, RR).其中HK作为CK的受体, HP将信号从HK传递到细胞核的RR, 从而调控靶基因转录(图1, 图2) (Hwang et al.2012; Kieber and Schaller, 2018; Lomin et al.2018; Arkhipov et al.2019; Hallmark and Rashotte, 2019; Zubo and Schaller, 2020).双分子荧光互补(bimolecular fluorescence complementation, BiFC)实验证明, 拟南芥CK受体AHK (Arabidopsis HK) 2-4均能与AHP (Arabidopsis HP)1-3相互作用, 且无明显的偏好; CK受体能形成同源和异源二聚体(Arkhipov et al.2019).AHK-AHK和AHK-AHP的相互作用主要定位于内质网(endoplasmic reticulum, ER); 相反, 各种磷酸转移蛋白(AHP)的同源和异源二聚体主要存在于植物的细胞核中(Arkhipov et al.2019). ...
... 反应调控因子是一个转录因子家族, 包含1个保守的接受磷酸基的Asp残基(Keshishian and Rashotte, 2015).根据其系统发生和结构域, RR家族成员可分为3种类型: A型、B型和C型(Schaller et al.2007), 但只有A型和B型RR与CK信号转导有关(Kieber and Schaller, 2018; Tuan et al.2019; Zubo and Schaller, 2020).B型RR包括ARR (Arabidopsis RR) 1、ARR2、ARR10-14和ARR18-21, 能被HP接收器结构域中的Asp残基磷酸化激活, 作为CK反应的正调控因子起作用(Hwang et al.2012).B型RR的特征是包含1个接收器结构域和1个含有类MYB DNA结合结构域的长C-端延伸.接收器结构域包含1个保守的Asp残基, 当多步骤磷酸传递被激活时, Asp可作为调控磷酸化的位点.类MYB DNA结合结构域与1个短的(A/G) GAT核心DNA序列结合, 该序列对B型RR的结合作用非常关键(Zubo et al.2017; Zubo and Schaller, 2020).ChIP-seq (chromatin immunoprecipitation combined with sequencing)实验显示, B型RR在植物体内以CK依赖的方式与其靶点结合, 这种结合以类似于细菌RR的方式被接收器结构域磷酸化激活(Zubo et al.2017). ...
... ; Zubo and Schaller, 2020).ChIP-seq (chromatin immunoprecipitation combined with sequencing)实验显示, B型RR在植物体内以CK依赖的方式与其靶点结合, 这种结合以类似于细菌RR的方式被接收器结构域磷酸化激活(Zubo et al.2017). ...
... A型ARR (如ARR3-9、ARR15-17)由1个接收器结构域和1个短的C-端延伸组成, 作为CK信号转导的负反馈调控因子起作用; 但与B型RR不同, 它们缺乏典型的转录调控输出结构域(output domain) (Kieber and Schaller, 2018).在对CK的反应中, A型RR通过B型RR的直接激活被诱导转录(Taniguchi et al.2007), CK也以磷酸化依赖的方式稳定许多A型RR (To et al.2007).A型ARR负调控CK信号转导的机制是与B型ARR竞争AHP的磷酸化, 从而导致多步骤磷酸传递与B型ARR转录调控因子解离, 以减少其对基因表达的影响(Kieber and Schaller, 2014; Zubo and Schaller, 2020). ...
... 在黑暗条件下, 1 μmol·L -1外源吲哚乙酸(indole acitic acid, IAA)处理显著延缓大豆(Glycine max)种子的萌发进程, 且随着IAA浓度的增加其抑制作用增强(Shuai et al.2017).外源IAA也以剂量依赖的方式有效增强野生型拟南芥新鲜收获种子的休眠(Liu et al.2013).基因表达分析显示, 生长素诱导A型ARR7和ARR15的转录, 抑制拟南芥根发生过程中的CK反应; 相反, 在茎分生组织中, 生长素抑制A型ARR的表达, 增强这些细胞对CK的敏感性.CK抑制生长素生物合成基因IAMT1和ILL6的表达, 诱导失活基因GH3.6的表达.CK还抑制AUX1生长素输出载体的表达, 影响生长素从根尖向茎的运输, 导致根细胞增殖减少.B型ARR的结合位点与编码生长素信号转导途径组分的基因(包括2个编码生长素受体的基因、10个AUX/IAA基因和4个ARF基因)有关.CK也诱导AUX/IAA基因SHY2/IAA3的表达, 该基因作为生长素信号转导的负调控因子起作用(Zubo and Schaller, 2020). ...
... 研究表明, 种子的萌发能力与乙烯的产生有关, 外源乙烯或乙烯利(ethrel, 乙烯释放剂)能解除种子的初生和次生休眠(second dormancy) (Corbineau et al.2014).CK通过转录和转录后机制激活ACC合酶(ACC synthase, ACS), 促进乙烯的生物合成; 乙烯的产生促进被CK抑制的暗生长幼苗的下胚轴伸长以及抑制根系的生长(Zdarska et al.2015).此外, B型ARR与许多影响乙烯信号转导的基因结合.尽管乙烯受体ETR2被CK下调表达, 但CK调控大多数编码调节乙烯信号转导途径活性的元件, 而不是途径本身.调节原件包括ARGOS基因家族的ARGOS和ARL以及TRP1、ETP和EBF基因.ARGOS是在受体水平起作用的跨膜蛋白, 可使植物对乙烯去敏感; 作为乙烯信号转导的正调控因子, TRP1可能通过与受体结合和干扰与CTR1的互作起作用; ETP和EBF基因分别编码参与EIN2和EIN3降解的F-box蛋白(Zubo and Schaller, 2020). ...
... 此外, CK也与油菜素甾醇(brassinosteroid)、茉莉酸(jasmonic acid)和水杨酸(salicylic acid)产生广泛互作(crosstalk), B型ARR的结合位点与这些植物激素的生物合成、降解/失活、运输和信号转导的基因有关(Zubo et al.2017; Zubo and Schaller, 2020). ...
... 细胞分裂素对种子发育、休眠与萌发的调控通过生物活性CK水平和信号转导途径来实现(图1, 图3) (Hwang et al.2012; Nguyen et al.2020; Zubo and Schaller, 2020).尽管近年来这些领域的研究已经取得了重要进展, 但仍然有一些科学问题需要进一步探索.例如, 已知生物活性CK可被CKX失活、通过cZOG可逆失活、被UGT不可逆失活以及通过βGlc重新活化(Frébortet al.2011; Jameson and Song, 2016), 那么, 这些反应是否同时存在于同一物种的同一组织, 还是具有物种、组织和发育阶段的特异性? 在响应种子发育、休眠与萌发和环境变化时, 为了控制组织或细胞内的生物活性CK水平, 这些途径怎样被启动、运行和协调? ...
 ,1,2,*, 刘军
,1,2,*, 刘军
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT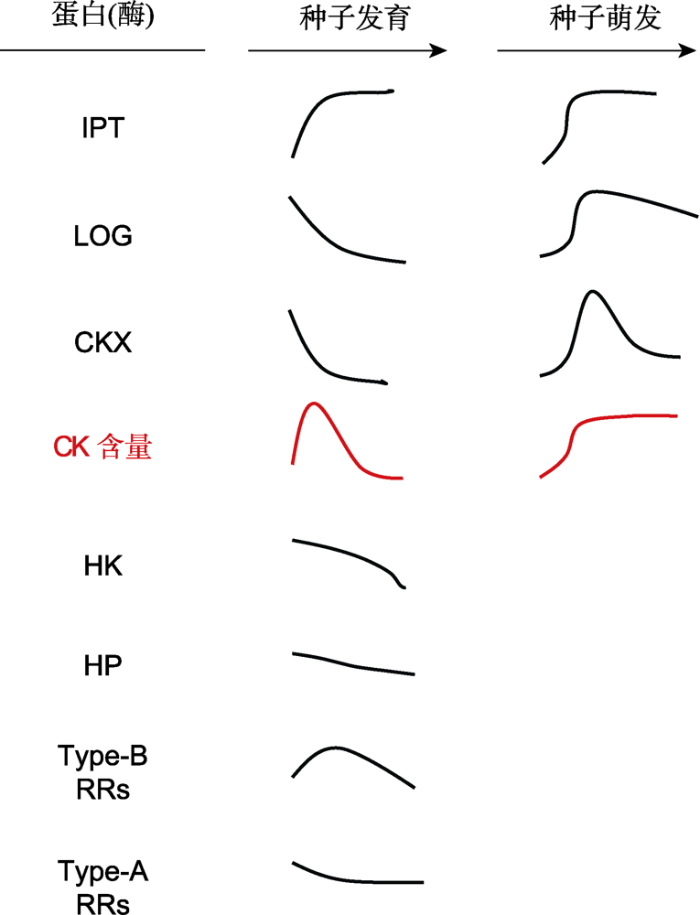 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}